Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
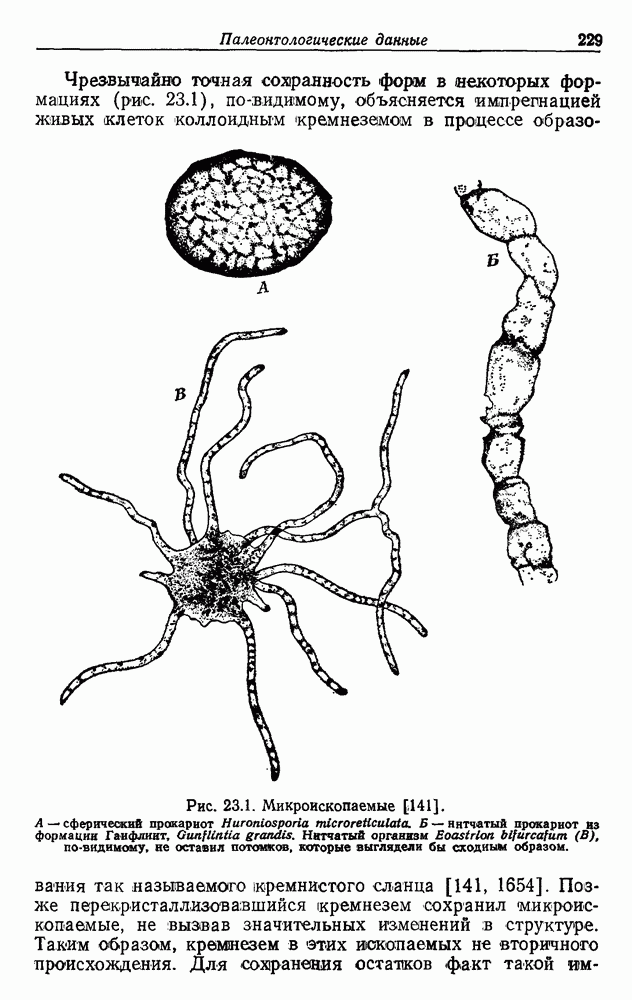
Г. ПРОЦЕССЫ И ВЕЩЕСТВАКак говорил Гоулэнд Хопкинс, жизнь — это такая штука, которая происходит. Существенная черта жизни — это ее химические процессы в их интегрированной целостности. Они описываются «динамической биохимией». Биомолекулы (нук-леотиды, белки, углеводы, липиды, светочувствительные вещества, минеральные вещества и т. д.) можно рассматривать как опоры, на которых держится сложное переплетение процессов, и это сложное переплетение связывает опоры и поддерживает их. Характер процессов зависит от участвующих в них веществ, а образующиеся продукты в свою очередь зависят от характера процессов. Из этого следует, что биоэнергетическую эволюцию можно описывать, говоря не о процессах, а о веществах. При достаточно глубоком знании оба описания должны быть эквивалентны. Но если деятельность определенной клетки (конкретнее говоря, биоэнергетические процессы) часто может быть хорошо описана в рамках динамической биохимии, то достаточно подробное описание на языке биомолекул и их пространственного расположения еще долгое время будет для нас практически невозможным. И все же эволюционная динамическая биохимия нуждается в сотрудничестве с «хемотаксономией» [39, 40, 825, 1563, 1826—1831], т. е. со сравнительным изучением химического состава тканей и органов у разных видов. Хемотаксономия еще до возникновения молекулярной биологии привела к тому, что позже назвали «молекулярным подходом к филогении» [596, 597, 600]. В тех случаях, когда имеется информация о тонкой структуре биомолекул и о их надмолекулярных агрегатах, возможность вклада в эволюционную теорию огромна. Среди специфических продуктов клеток, т. е. биохимических маркеров [1], можно упомянуть пигменты (каротиноиды [725, 1831]) и компоненты клеточной оболочки 1831]; последние обусловливают также реакцию по Граму Основная схема синтеза биомолекул заложена в конечном счете в ДНК Чаргафф [369] заметил, что молярное содержание гуанина и цитозина, с одной стороны, и аденина и тимина — с другой в любом данном препарате природной ДНК одинаковы. Это и понятно, и даже необходимо, в свете предложенной позже, в 1953 г., двойной спирали Крика и Уотсона. Аденин спаривается с тимином, а гуанин — с цитозином. Поэтому нуклеотидный состав ДНК любого вида можно выразить одним показателем. Этот показатель был определен для многих групп организмов; у бактерий он варьирует между 23 и 74% ГЦ (см., например, [469, 470]). Разумеется, что в таком случае показатель для АТ-пар будет составлять 77— 26%. Предполагается, что количества ГЦ у родственных групп сравнительно близки. (Однако следует быть осторожным с такими выводами [1710]. Оказалось, что содержание ГЦ-пар положительно коррелирует со степенью облучения экосистемы, в которой в норме живут определенные виды бактерий, ультрафиолетовыми лучами. Возможно, это связано с хорошо известной большей чувствительностью тимина к фотохимической димеризации. Поэтому не исключено, что по содержанию ГЦ не всегда с достаточным основанием можно говорить о степени родства.) Более точную информацию о сходствах и различиях между молекулами ДНК разных видов («гомологичности ДНК») дают эксперименты по гибридизации (спариванию оснований) ДНК. В многочисленных работах описано много различных методик для определения гомологии и приведено много результатов [493, 470, 887, 1203, 1215, 1521, 1628, 1755, 1831]. Максимум информации можно было бы получить, установив полностью последовательность оснований в ДНК. К сожалению, из-за методических трудностей [1204, 1602] полная последовательность оснований молекулы ДНК пока не определена. Но участки таких молекул транслируются с помощью мРНК на язык белков, определяя таким образом последовательность аминокислот в молекуле белка. Белок содержит меньше информации, чем соответствующая часть молекулы ДНК, поскольку одна и та же аминокислота может кодироваться несколькими разными триплетами нуклеотидов До сих пор еще не пришли к единому мнению по вопросу о том, были ли медленные эволюционные изменения состава белка в результате отбора биологически полезными, как считают обычно, или нейтральными [589, 987—990, 1203, 1207, 1540, 2060]. Обсуждение этого важного вопроса с точки зрения скорее организменной, чем молекулярной биологии можно найти в работах Симпсона [1708] и Айалы [107]. Для выяснения последовательности аминокислот чаще всего используют цитохром с [477, 478, 588, 589, 1205— 1207, 1334]. Кроме того, для подобных целей использовали и другие белки, в том числе фибринопептид, гистон IV и гемоглобин [447, 477]. Прочитана также последовательность аминокислот и в некоторых белках из высших растений [254—258, 1956]. К сожалению, большинство работ по цитохрому с проведено на высших организмах. Бактериями, особенно интересными с точки зрения биоэнергетики, в основном пренебрегают. По-видимому, частично это происходит потому, что не всегда ясно, какой цитохром определенной бактерии соответствует цитохрому с высших организмов. Другие упомянутые выше белки даже не встречаются у бактерий. Только недавно метод выяснения последовательности аминокислот в цитохроме с был применен к фотосинтезирующей бактерии Rhodospirillum rubrum, а выяснение последовательности нуклеотидов тРНК было проведено на других бактериях [1237]. Возможно, самым важным объектом анализа последовательности для филогении окажется древний белок ферредоксин (Кстати, Яманака [2036, 2037] попытался использовать избирательное сродство цитохрома с к цитохромоксидазе и нитритредуктазе от разных организмов для построения филогенетического древа; результаты оказались чрезвычайно интересными!) Результаты расшифровки последовательностей используются с помощью сложных математических методов [446, 447] для построения отдельных частей филогенетических древес. Более того, если использовать палеонтологические данные о времени, прошедшем с момента определенных ответвлений для калибровки, то можно рассчитать средние скорости эволюционных изменений; они различны для разных информационных макромолекул. На основании знания этих скоростей определили, что позвоночные отделились от беспозвоночных примерно 0,6 гигалет (Гл) 1 назад. Высшие организмы (эукариоты (18), т. е. растения, кроме сине-зеленых водорослей, животные и грибы) отделились от бактерий (прокариотов, 18) в 2,7 раза раньше, чем произошло разделение эукариотов на разные группы [1237]. Ниже будут приведены данные об эволюции биологического превращения энергии, полученные исходя из структуры белка В литературе имеются указания, что в случае бактерий горизонтальный перенос генов может быть источником ошибок в филогенетических выводах, сделанных на основании анализа последовательностей аминокислот [823].
|
 |
Оглавление
|













































































































































































































































 .
.
 . Следовательно, данные об эволюционном родстве можно, по-видимому, искать, сравнивая «содержание ГЦ».
. Следовательно, данные об эволюционном родстве можно, по-видимому, искать, сравнивая «содержание ГЦ».
 . И все же для выяснения филогенетических взаимоотношений изучение последовательности с помощью белков имеет чрезвычайное значение [69, 422, 452, 2032, 2061 — 2063]. Определение полных аминокислотных последовательностей технически возможно уже много лет [448, 589, 933, 1208].
. И все же для выяснения филогенетических взаимоотношений изучение последовательности с помощью белков имеет чрезвычайное значение [69, 422, 452, 2032, 2061 — 2063]. Определение полных аминокислотных последовательностей технически возможно уже много лет [448, 589, 933, 1208].
 который, видимо, есть у всех организмов [341, 449, 450, 527, 773—777, 1158, 1988].
который, видимо, есть у всех организмов [341, 449, 450, 527, 773—777, 1158, 1988].
