Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
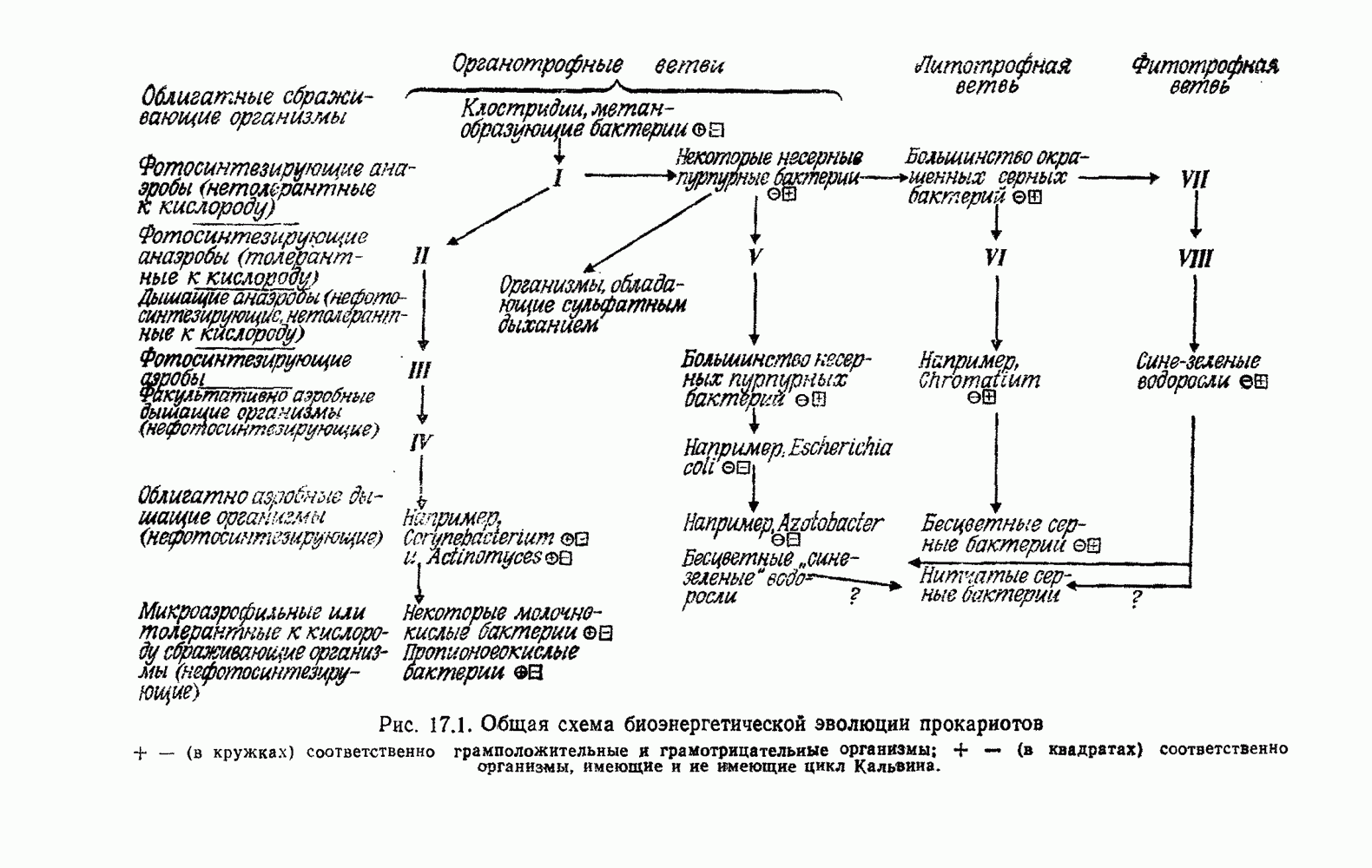
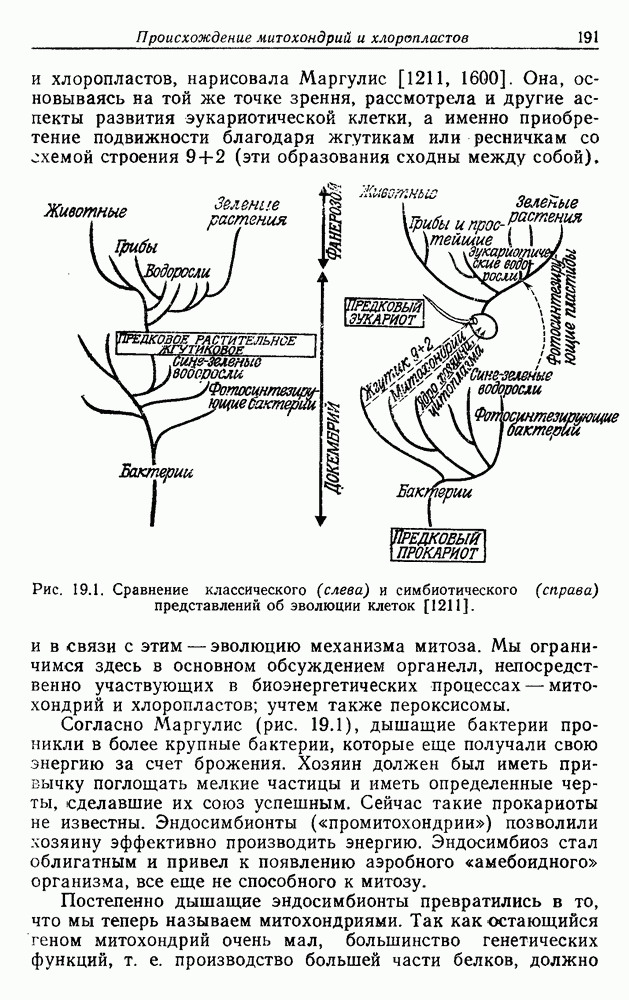
Д. БИОЭНЕРГЕТИКА И КЛАССИФИКАЦИЯВ классификации живых объектов биологи традиционно основывались главным образом на макростроении организмов и их наружных органов (морфологии) и на макрофизиологических процессах, в основном размножении и передвижении. Этот подход весьма плодотворно используется для классификации высших растений, грибов и животных, но для классификации примитивных организмов требуются дополнительные критерии, основанные на микрофизиологических и биохимических данных. Клод Бернар, А. Дорн, П. Уинстон, Г. Молиш и Гоулэнд Хопкинс были среди первых, кто подчеркнул важность этих критериев [601, 1185, 1285]. Среди активных сторонников физиологического подхода был также Львов [1185]: «Шизомицеты (бактерии, Э.Б.) ввиду разнообразия их метаболизма... должны представлять собой замечательный материал для изучения физиологической эволюции. Сравнение биохимических процессов в связи с их эволюцией должно дать результаты первостепенной важности... Морфологические признаки бактерий не позволяют удовлетворительно разделить их на отряды и семейства. Единственные критерии, которыми располагает морфолог, это форма бактериальной клетки, наличие жгутиков и особенности их прикрепления, присутствие или отсутствие спор. Ни один из этих признаков не обладает достаточной стабильностью». Сравнительные достоинства «морфологии» и «физиологии» для классификации вкратце обсуждались много раз [1203, 1305, 1754]. В общем и целом, как показал Львов, физиологические данные тем полезнее, чем примитивнее организмы. Однако проводить классификацию высших организмов на основе их физиологических особенностей явно трудно или даже практически невозможно, поскольку здесь известно огромное число морфологически различающихся видов. Пригодные для классификации метаболические различия невелики и, как правило, неизвестны. Но не следует забывать, что в конечном счете морфология, в том числе микроморфология и физиология, взаимозависимы. Много лет назад Клюйвер и Ван-Ниль [1015] сделали вывод, что для классификации бактерий «морфология остается первым и самым надежным признаком». Этот вывод позже повторили в своих работах Стэниер и Ван-Ниль [1785], а также Ван-Ниль [1912, 1916]. Впрочем, уже в то время эти авторы подчеркивали важность для классификации физиологических, конкретнее, катаболических процессов. Эти процессы в большинстве своем обеспечивают клетки энергией. Значительно позднее Стэниер пришел к выводу, что из функциональных признаков для таксономических целей наиболее удобно использовать метаболические процессы. В недавней работе Стэниер и др. [1782] еще более подчеркнули значение физиологического подхода к эубактериям (истинным бактериям): «Их разнообразие лишь в небольшой степени выражается в различии клеточных структур, так как структурная простота этих организмов значительно ограничивает число различных категорий, которые можно установить, пользуясь только структурными признаками. Разнообразие эубактерий отражено главным образом в невиданном у других организмов диапазоне специализированных физиологических и биохимических типов, существующих среди них... Большинство бактерий можно правильно описать, лишь указывая, что они делают, а не просто как они выглядят...» В качестве иллюстрации существующих трудностей авторы отмечают специфический случай фотосинтезирующих пурпурных бактерий (9,11). Их объединяет особый метаболизм, служащий для получения энергии, и в связи с этим наличие у них специфической группы пигментов. Но среди этих бактерий существуют три разных структурных типа: палочки и спирали, размножающиеся простым делением, и один овальный организм, размножающийся почкованием. Для каждого из этих трех типов известны морфологические аналоги среди нефотосинтезирующих эубактерий. Вероятно, в этом случае может помочь гипотеза конверсии В конечном итоге должны доминировать те критерии, которые позволяют ближе всего подойти к естественной (филогенетической, т. е. эволюционной) классификации организмов, в противоположность фенетической классификации, основанной только на признаках существующих ныне организмов. Цель построения полностью филогенетической классификации, конечно, так далека, что до сих пор нет единого мнения, стоит ли вообще к ней стремиться. Различные взгляды на этот вопрос можно найти в сравнительно недавних работах [216, 330, 467, 835, 837, 1754]. Современная форма фенетической классификации — «нумерическая таксономия» [1756, 1757], в которой подсчитываются признаки и затем подсчетами определяется степень родства организмов. Взвешивания признаков не производится, хотя никто не оспаривает огромного внутреннего интереса филогенетических взаимоотношений, а для их определения, видимо, требуется взвешивание. Таким образом, задача этого подхода состоит в том, чтобы классифицировать по критериям, которые считаются объективными; полагают, что любой хорошо информированный наблюдатель в конце концов придет к тем же выводам. Однако, как отмечают некоторые исследователи, уже в выборе признаков, которые будут использоваться, неизбежно проявляется субъективность суждения (см., например, [732]). При выяснении филогенетических связей может ввести в заблуждение морфологическая, физиологическая или биохимическая конвергенция. Замечательный пример конвергентной эволюции, насколько нам известно, — это путь разложения и использования бензоата и родственных соединений [344]. И у Acinetobacter, и у псевдомонад — групп, как полагают, очень удаленных друг от друга, — наблюдаются одни и те же промежуточные продукты (около дюжины), но разные механизмы регуляции. По-видимому, в данном случае химические свойства субстрата заставили организмы выработать одинаковый путь разложения; очевидно, никакой другой путь химически невозможен. Но не исключено, что здесь имел место горизонтальный перенос целого комплекса генов Оргель сравнил родство с плагиатом [1366]. Ситуация, когда два изобретателя подают заявки на одно и то же изобретение, причем тексты этих заявок тоже в основном идентичны, т. е. когда по крайней мере один из заявителей скопировал идею другого, соответствует ситуации, в которой не только продукты метаболизма, но и ведущие к ним пути идентичны. В этом случае вероятно родство. Напротив, фундаментальное различие путей указывает на конвергенцию. Но, как показывает пример с бензоатом, этот критерий не всегда справедлив. Общеизвестна, например, конвергенция в случае плавников (рыбы, дельфины, пингвины) или крыльев (птицы, рукокрылые, вымершие рептилии). Возможно, конвергенция в поведении оценена меньше. Например, кто вспоминает, что лисы и гуси, которые известны как образцовые семьянины и заботливые родители, филогенетически связаны только через явно менее добродетельных рептилий?
|
 |
Оглавление
|













































































































































































































































 . В общем можно сказать, что любое кажущееся противоречие между результатами морфологического и физиолого-биохимического исследований, включая биоэнергетику, следует рассматривать как вызов, как приглашение к более углубленному анализу.
. В общем можно сказать, что любое кажущееся противоречие между результатами морфологического и физиолого-биохимического исследований, включая биоэнергетику, следует рассматривать как вызов, как приглашение к более углубленному анализу.
 а механизмы регуляции изменились позже.
а механизмы регуляции изменились позже.