Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
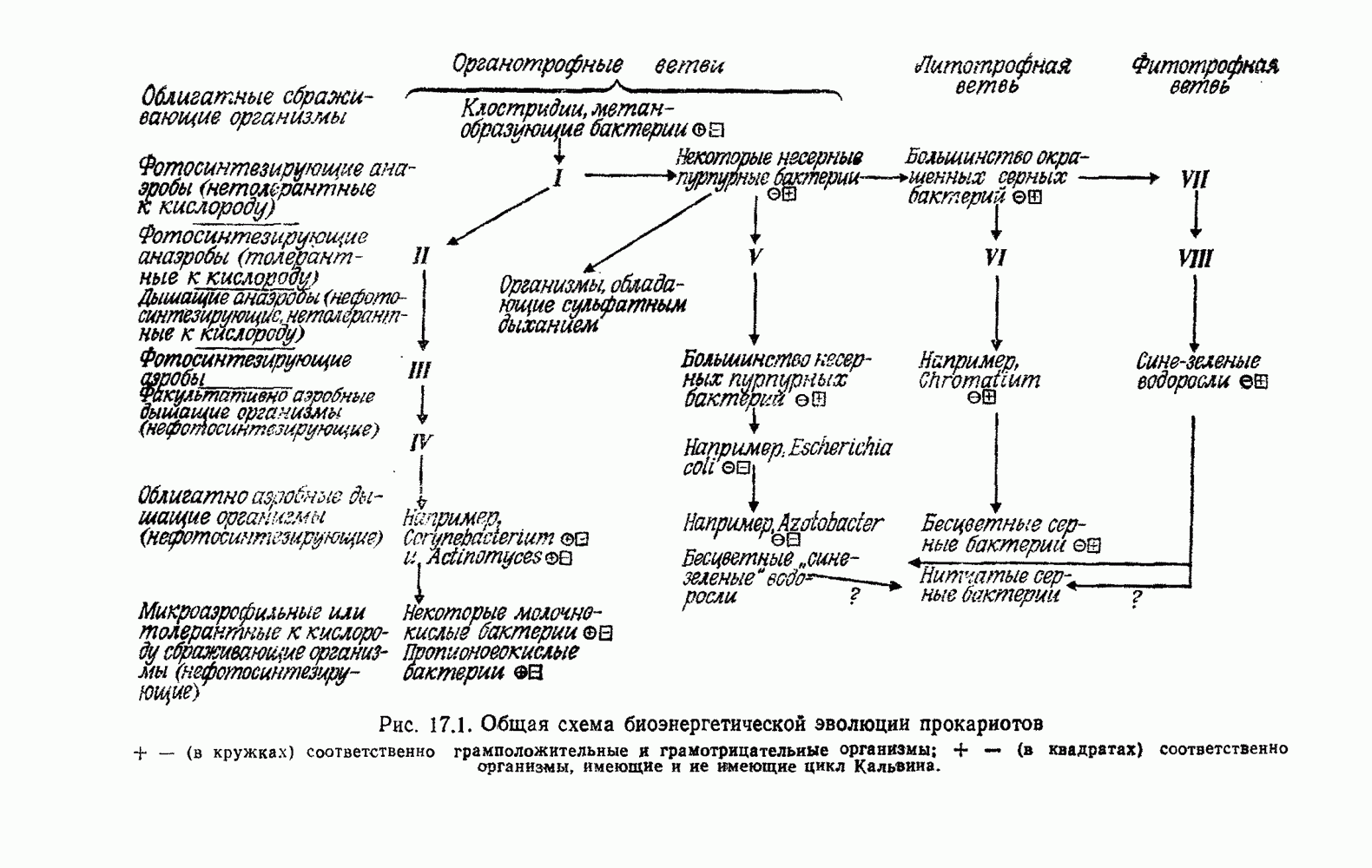
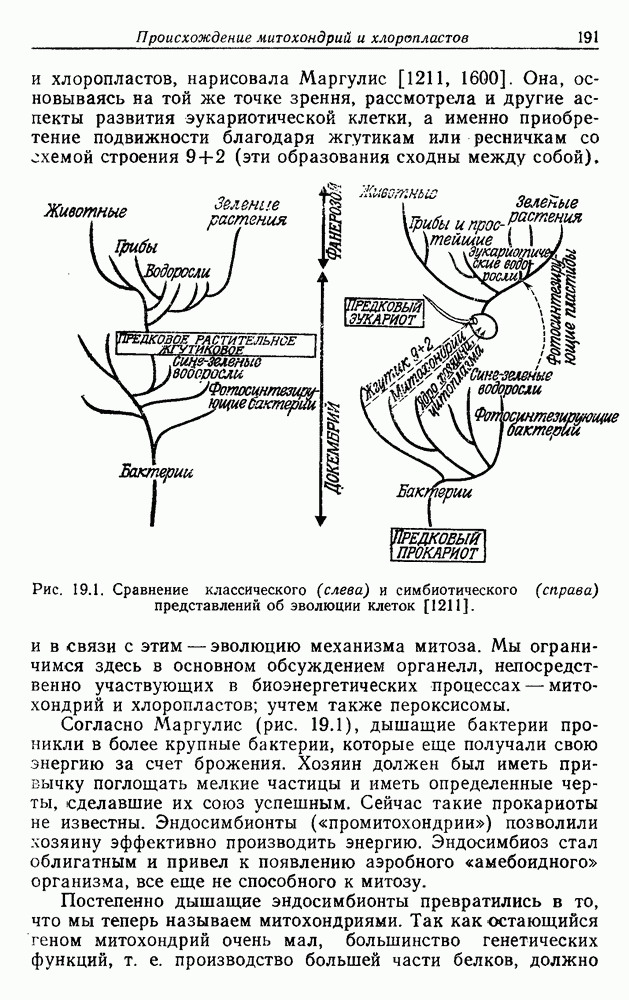
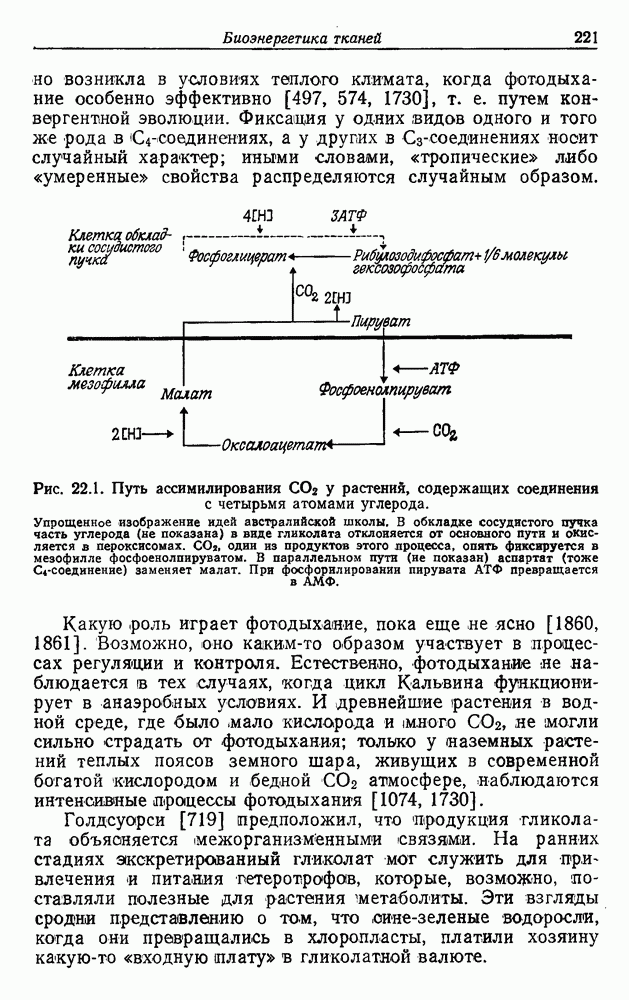
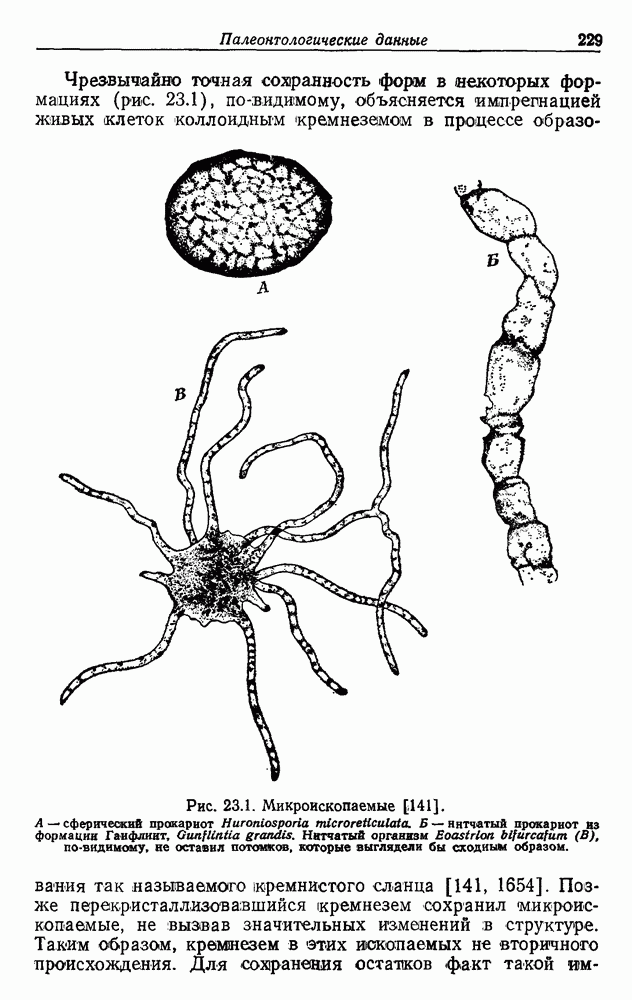
Д. ГИПОТЕЗА КОНВЕРСИИОтветом на все эти вопросы может быть положение о происхождении всех дышащих бактерий от фотосинтезирующих. Часто указывалось на общее сходство цепей переноса электронов в фотосинтезе и дыхании. Как в основе механизма дыхания, так и в основе механизма фотосинтеза всегда лежат упорядоченные ансамбли, связанные с мембранами. Более того, фотосинтетическая цепь потока электронов включает, как и дыхательная цепь, флавопротеиды, цитохромы, хиноны и негеминовые белки, содержащие серу. Не удивительно, что, по мнению многих авторов [514, 660, 1354], фотосинтетическая цепь, уже существовавшая, была изменена, приспособлена и перенята для дыхания, после того как появилась необходимость в нем. Далее было высказано предположение [284, 285], что каждый существующий вид дышащих бактерий произошел от того или иного вида фотосинтезирующих бактерий. Эту гипотезу можно назвать «гипотезой облигатной конверсии». Не слишком трудно себе представить, что такой переход мог происходить неоднократно и параллельно от многих фотосинтезирующих ко многим дышащим видам, раз при этом требовалось всего лишь переключить существующие механизмы с фотосинтеза на дыхание. Эта гипотеза объясняет удивительное разнообразие отдельных деталей в устройстве дыхательной цепи у разных бактерий. Она объясняет также гибкость дыхания. Один и тот же организм может в разных условиях использовать для бактериального дыхания разные пути, что, по-видимому, справедливо и в отношении бактериального фотосинтеза Мы не видим причин, по которым и у сине-зеленых водорослей, и у бактерий дыхание не могло бы возникать повторно несколько раз, и понятно, что механизмы при этом также должны быть разными. К сожалению, как уже указывалось, дыхательные цепи у сине-зеленых водорослей изучены мало. Сообщалось, что флавиновые системы могут использовать кислород Как известно, вне фотосинтетических мембран в клетке, помимо цитохромов, содержатся и другие гемопротеиды. Так, в аэробных условиях гемопротеиды стали полезными для избавления от ядовитой Но основная трудность, с которой мы сталкиваемся при создании любой гипотезы, пытающейся объяснить происхождение полного дыхания в процессах, основанных на участии флавина, состоит в том, что элементы связанной с мембраной дыхательной цепи и окислительного фосфорилирования должны были бы возникать de novo много раз у разных бактерий, так что, по-видимому, лучше полагать, что связанные с мембраной фотосинтетические цепи переноса электронов превратились в дыхательные. Все гемопротеиды, функционирующие вне фотосинтетических и дыхательных цепей, вполне могли произойти от цитохромов, входящих в состав этих цепей.
|
 |
Оглавление
|













































































































































































































































 . И здесь разнообразие, по всей вероятности, имеет адаптивную ценность
. И здесь разнообразие, по всей вероятности, имеет адаптивную ценность  .
.
 Однако при этом не синтезируется АТФ, поэтому Долин [487] рассматривает это бесцитохромное «дыхание» посредством флавинов (часто идущее с большой интенсивностью) как раннюю и в основном неудачную попытку справиться с
Однако при этом не синтезируется АТФ, поэтому Долин [487] рассматривает это бесцитохромное «дыхание» посредством флавинов (часто идущее с большой интенсивностью) как раннюю и в основном неудачную попытку справиться с  Можно представить, что цитохромы, как только они появились в связи с фотосинтезом
Можно представить, что цитохромы, как только они появились в связи с фотосинтезом  дополнили собой короткую, основанную на флавине «дыхательную» цепь [1093, 1715] в кислородной атмосфере, которая образовалась постепенно вследствие фотосинтеза. Между прочим, ввиду отсутствия окислительного фосфорилирования их лучше было бы называть окислительно-восстановительными, а не дыхательными цепями.
дополнили собой короткую, основанную на флавине «дыхательную» цепь [1093, 1715] в кислородной атмосфере, которая образовалась постепенно вследствие фотосинтеза. Между прочим, ввиду отсутствия окислительного фосфорилирования их лучше было бы называть окислительно-восстановительными, а не дыхательными цепями.
 продукта функционирования оксигеназ — каталазы и пероксидазы. На какой-то стадии (или стадиях) у высших организмов появились гемоглобины
продукта функционирования оксигеназ — каталазы и пероксидазы. На какой-то стадии (или стадиях) у высших организмов появились гемоглобины  . В микросомной фракции печени позвоночных, т. е. тоже у эукариотов, найдены цитохромы
. В микросомной фракции печени позвоночных, т. е. тоже у эукариотов, найдены цитохромы 