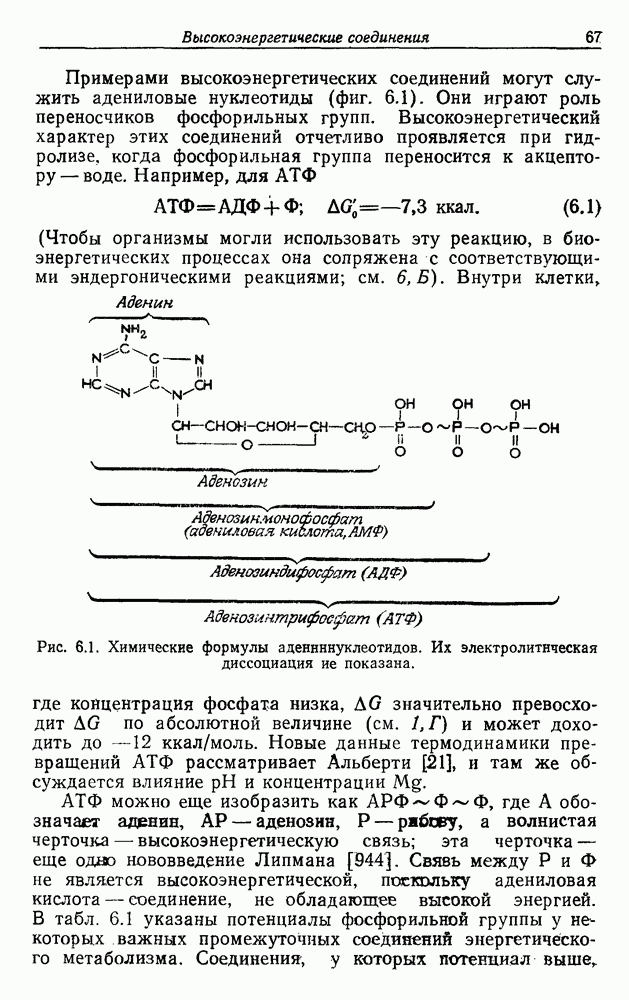
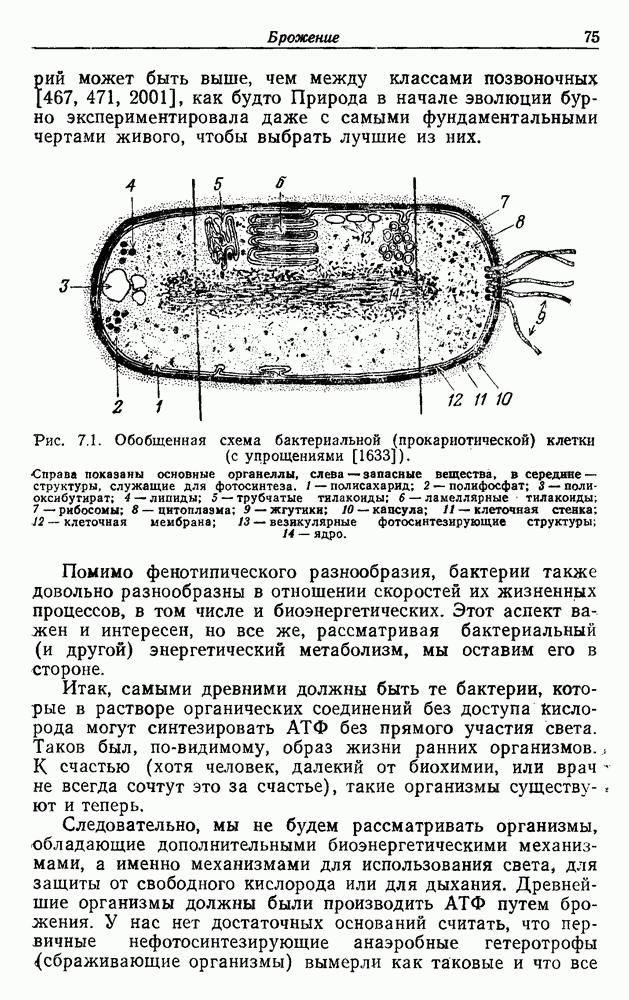
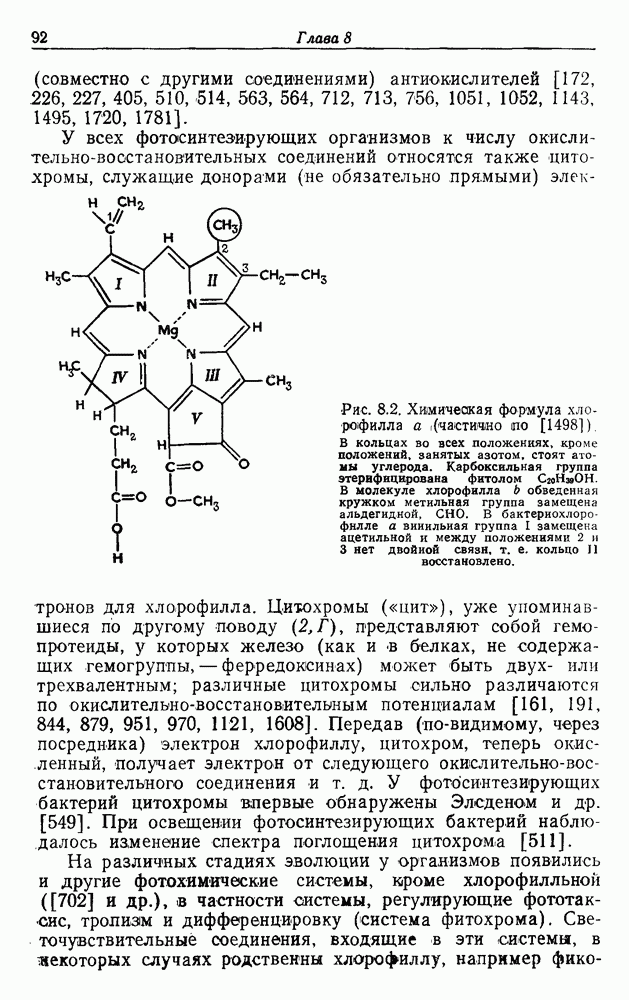
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
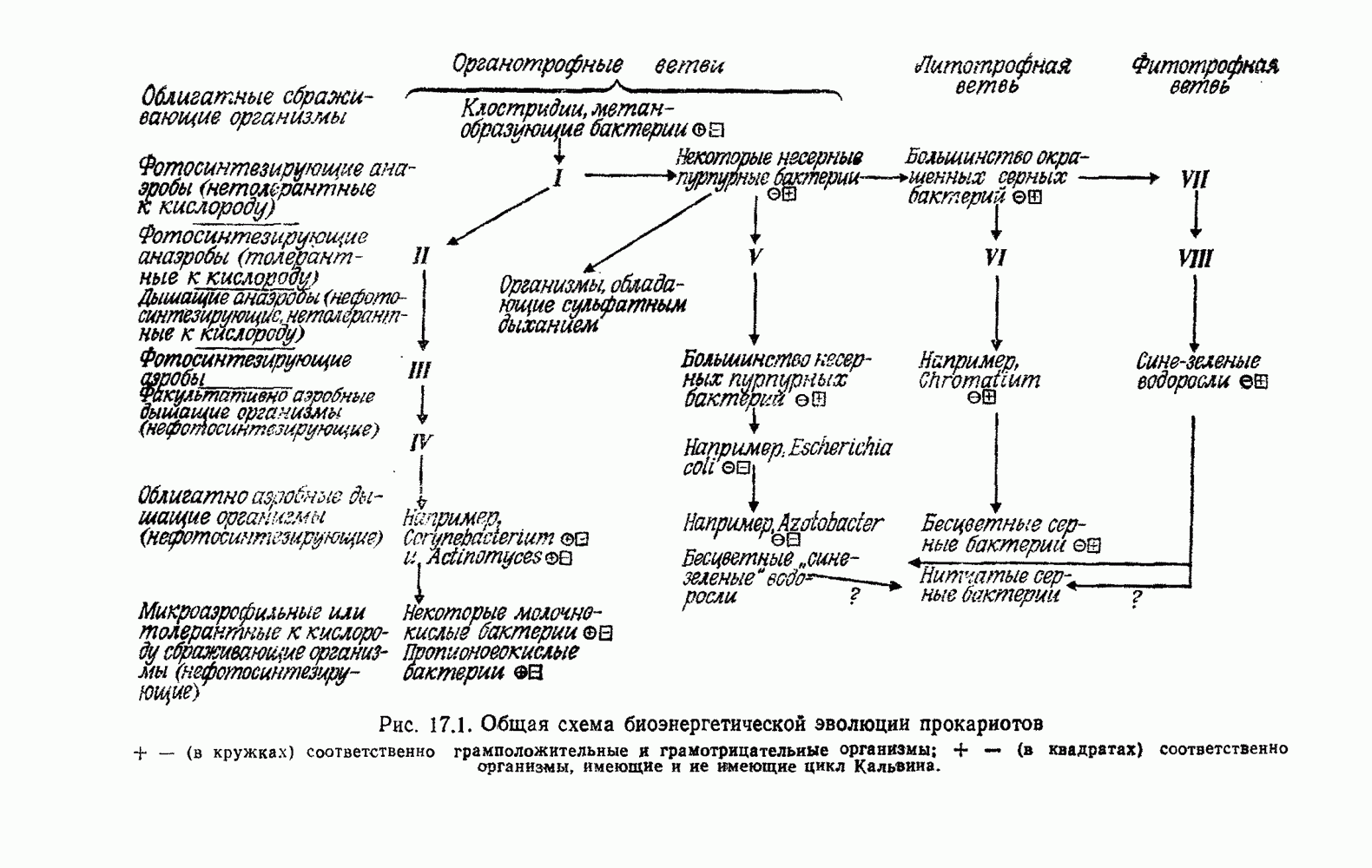
ГЛАВА 15. ХЕМОЛИТОТРОФИЯА. ОБЩИЕ ОСОБЕННОСТИКак и дышащие органотрофы (хемооргано синтезирующие литотрофы (хемолитотрофы), открытые Виноградским [2009, 2010], используют для синтеза АТФ окисление на воздухе при участии неорганических восстановителей [23, 548, 640, 673, 696, 1113, 1114, 1419]. У хемолитотрофов обычно обнаруживают внутренние мембранные системы, напоминающие мембранные системы фотолитотрофов [979, 1945]. Хемолитотрофы, подобно фотолитотрофам, грамотрицательны. Как правило, они аэробны и используют в качестве конечного акцептора электронов свободный кислород. Иногда кислород заменяется нитрат-ионом. Важные группы хемолитотрофов — тиобациллы, нитрификаторы и водородные бактерии. Они окисляют соответственно соединения серы, азота или свободный Во многих случаях для производства энергии могут также в какой-то степени использоваться экзогенные органические субстраты. В других случаях экзогенные органические вещества используются для биосинтеза, но не для производства энергии, что можно доказать с помощью изотопов. Было много споров о степени облигатности литотрофии или даже автотрофии некоторых из этих организмов [979, 1552, 1571]. Литотрофы, которые не могут использовать экзогенные органические субстраты, явно утратили способность к этому. Таким образом, в настоящее время эта проблема для хемотрофов имеет не большую важность, чем для фототрофов (8, В; 11, А). Риттенберг [1553] пишет: «Остаются некоторые виды бактерий, которых не удалось выращивать в культуре в отсутствие неорганического источника энергии или света. Такие формы называют соответственно облигатными хемолитотрофами или облигатными фототрофами. Имеющиеся данные... позволяют предположить, что нет таких бактерий, которые одновременно являлись бы облигатными автотрофами. С экологической и эволюционной точек зрения полная зависимость от Учитывая эволюционные соображения, кажется разумным отнести хемолитотрофов вместе с аэробными хемоорганотрофами к дышащим организмам. Это верно, хотя при хемолитотрофии
Рис. 15.1. Одна из возможных схем энергетически сопряженного транспорта электронов у хемолитотрофов. Первичный восстановитель Существенная черта этих организмов — производство энергии путем окисления Отнесение хемолитотрофов к дышащим организмам оправдано тем, что они синтезируют АТФ путем окислительного фосфорилирования. Это обнаружено также и в бесклеточных экстрактах, например в экстрактах из нитрифицирующих бактерий [31] или из тиобацилл [830]. Как всегда, механизмы; потока электронов и окислительного фосфорилирования связаны с мембранами и нерастворимы. (Аберрантные результаты были получены Колом и Элимом [410] на Thiobacillus navellus.) Определенное в эксперименте отношение Восстановители переносят электроны к какому-либо из звеньев цепи потока электронов [23, 28, 979, 985, 1419, 1874]. Из-за недостаточно больших отрицательных окислительно-восстановительных потенциалов восстановители (за исключением Хемолитотрофы получают углерод для биосинтеза, ассимилируя
|
 |
Оглавление
|








































































































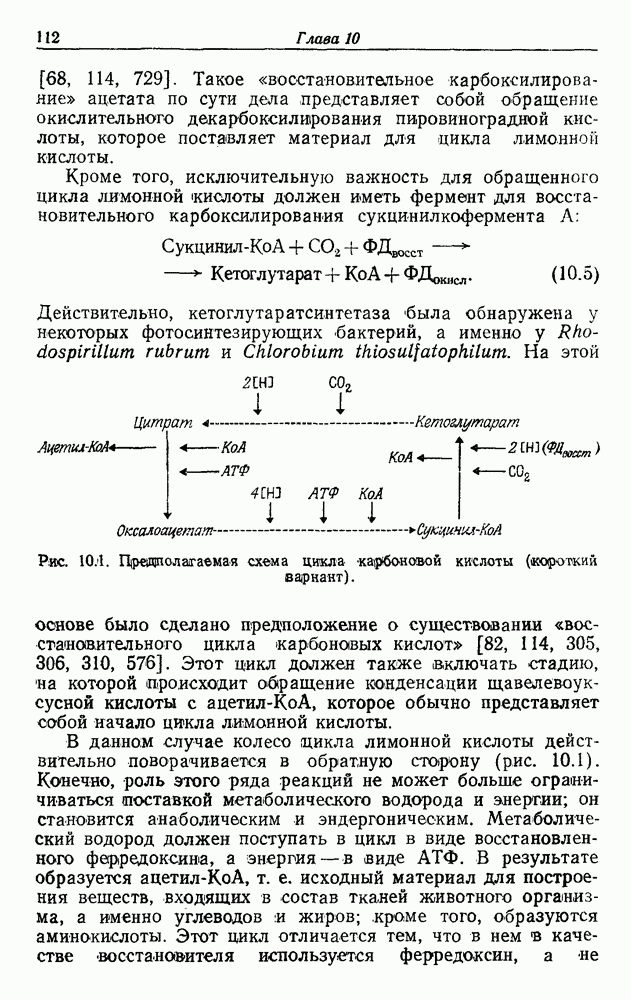










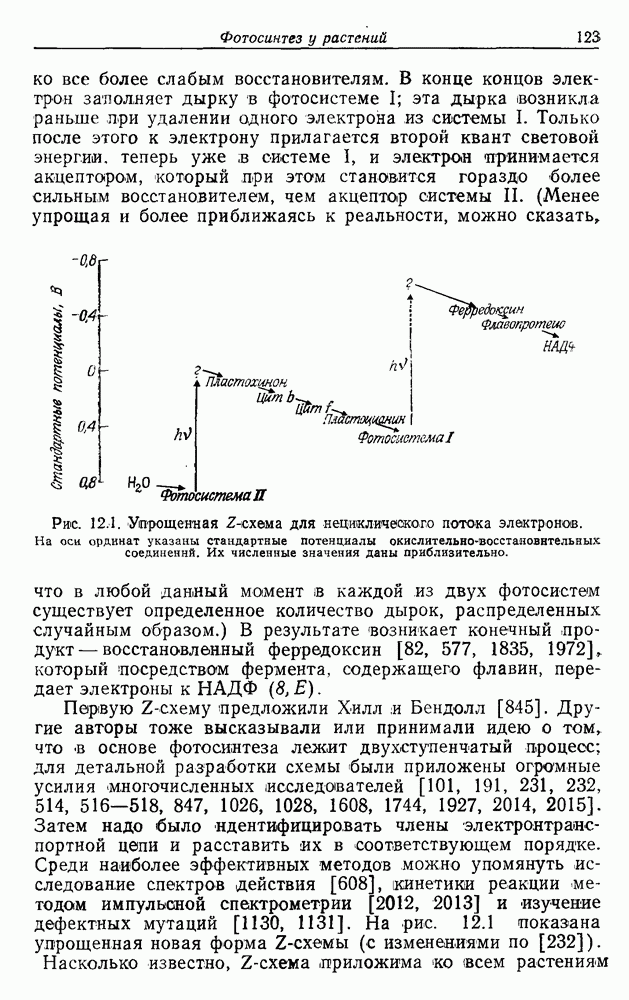
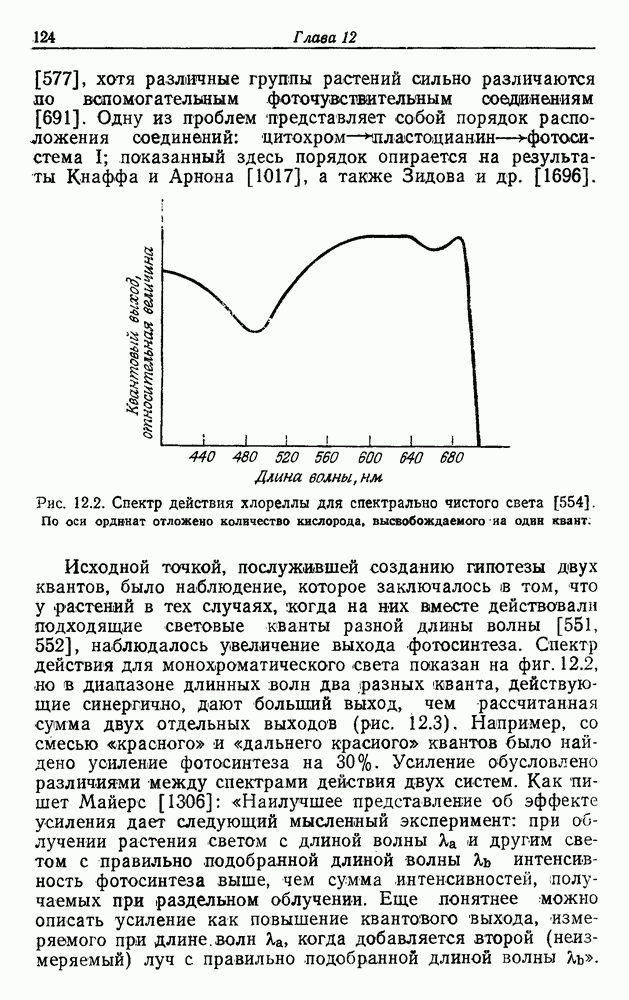
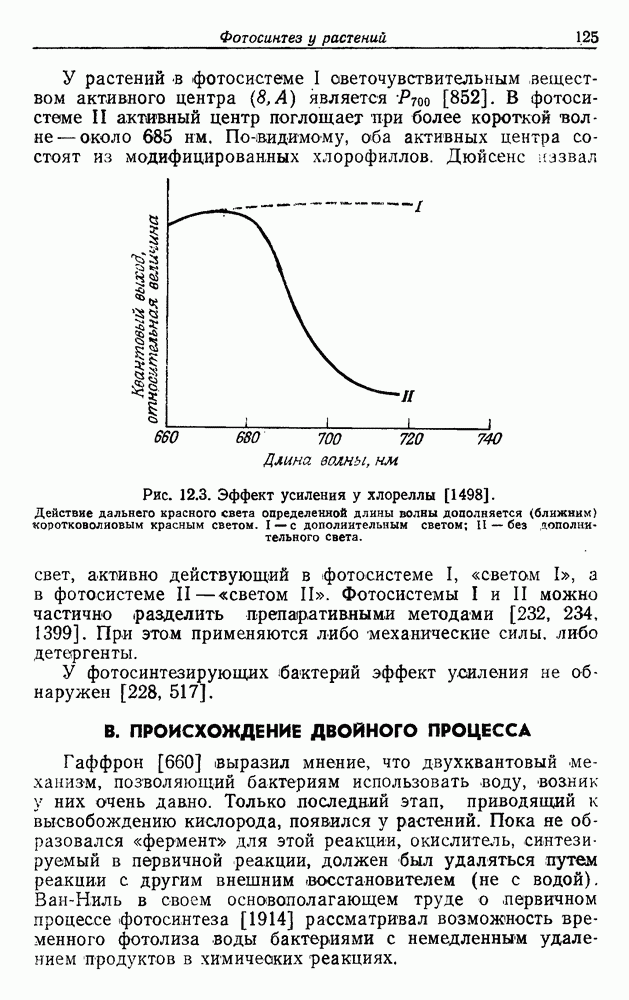



























































































































 . В отсутствие экзогенного субстрата на дыхание расходуется органическое запасное вещество [236, 985]. Очевидно, что хемолитотрофы сохранили, по крайней мере, некоторые из механизмов, необходимых для органотрофии.
. В отсутствие экзогенного субстрата на дыхание расходуется органическое запасное вещество [236, 985]. Очевидно, что хемолитотрофы сохранили, по крайней мере, некоторые из механизмов, необходимых для органотрофии.
 как источника углерода имеет мало смысла и бактерии с такими требованиями были бы анахронизмом на современной Земле».
как источника углерода имеет мало смысла и бактерии с такими требованиями были бы анахронизмом на современной Земле».
 не образуется, а, напротив, поглощается как источник углерода.
не образуется, а, напротив, поглощается как источник углерода.

 пиридиннуклеотид
пиридиннуклеотид  и конечный акцептор электрона (органическое вещество, обозначенное здесь
и конечный акцептор электрона (органическое вещество, обозначенное здесь  показаны в восстановленных состояниях.
показаны в восстановленных состояниях.
 или нитратом.
или нитратом.
 и локализация фосфорилирования значительно варьируют у разных родов и видов [29].
и локализация фосфорилирования значительно варьируют у разных родов и видов [29].
 не могут непосредственно передавать электроны к НАД (см. табл. 11.1). Например, при окислении тиосульфата бактерией
не могут непосредственно передавать электроны к НАД (см. табл. 11.1). Например, при окислении тиосульфата бактерией  сначала восстанавливается цитохром с [1616]. НАД-Н, необходимый для ассимиляции
сначала восстанавливается цитохром с [1616]. НАД-Н, необходимый для ассимиляции  затем, как правило, поставляется (рис. 15.1) обращенным потоком электронов
затем, как правило, поставляется (рис. 15.1) обращенным потоком электронов  к
к  приводимым в движение АТФ [24—27, 30, 610, 985, 1419, 1684]. Однако многие вопросы, касающиеся состава цепей переноса электронов и точек вхождения электронов в цепи, остаются открытыми [28, 985, 1945].
приводимым в движение АТФ [24—27, 30, 610, 985, 1419, 1684]. Однако многие вопросы, касающиеся состава цепей переноса электронов и точек вхождения электронов в цепи, остаются открытыми [28, 985, 1945].
 . С этой целью, как и у фотолитотрофов
. С этой целью, как и у фотолитотрофов  используется в основном восстановительный пентозофосфатный цикл [1419]. При гетеротрофном росте активность ферментов этого цикла обычно подавляется, что тоже напоминает фотолитотрофов [979, 1552]. Запасные вещества
используется в основном восстановительный пентозофосфатный цикл [1419]. При гетеротрофном росте активность ферментов этого цикла обычно подавляется, что тоже напоминает фотолитотрофов [979, 1552]. Запасные вещества  обнаруженные у хемолитотрофов, представляют собой
обнаруженные у хемолитотрофов, представляют собой  углеводы и у некоторых водородных бактерий жиры [505]. Обнаружены отдельные ферменты цикла лимонной кислоты [978, 1422], частично используемые в биосинтетических
углеводы и у некоторых водородных бактерий жиры [505]. Обнаружены отдельные ферменты цикла лимонной кислоты [978, 1422], частично используемые в биосинтетических  т. е. в анаболических, процессах [727, 979].
т. е. в анаболических, процессах [727, 979].