Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
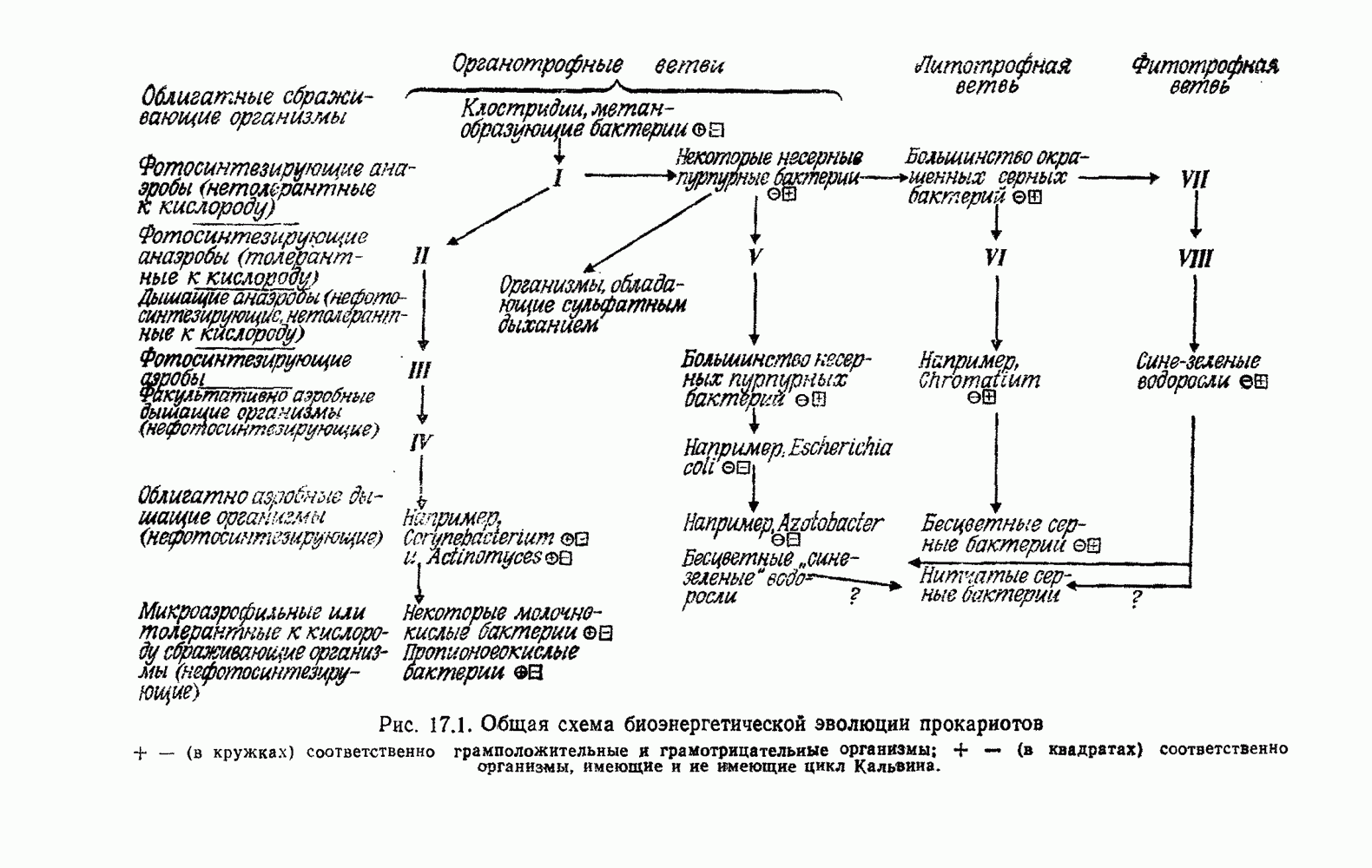
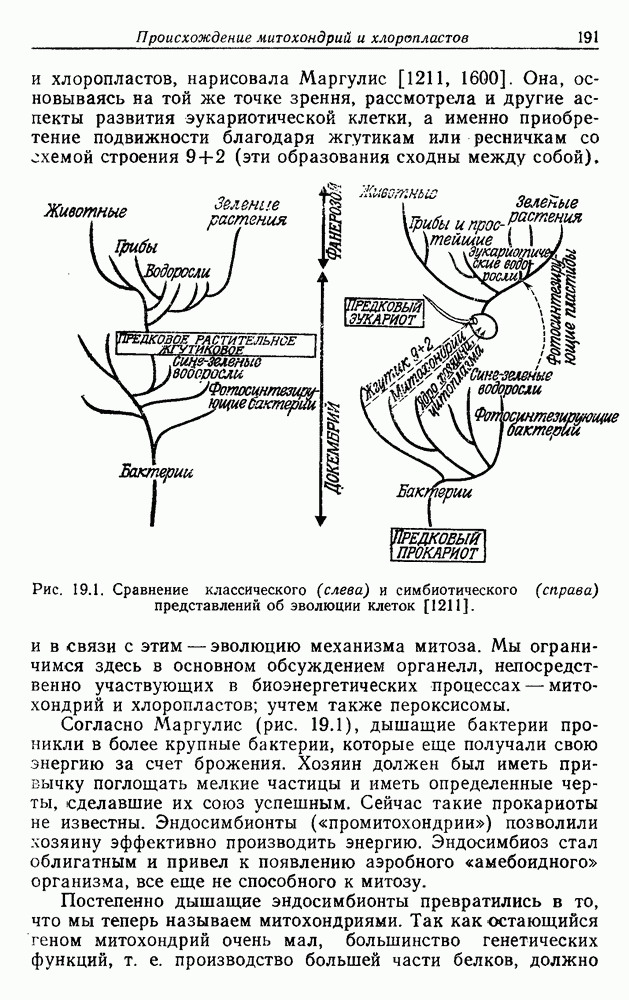
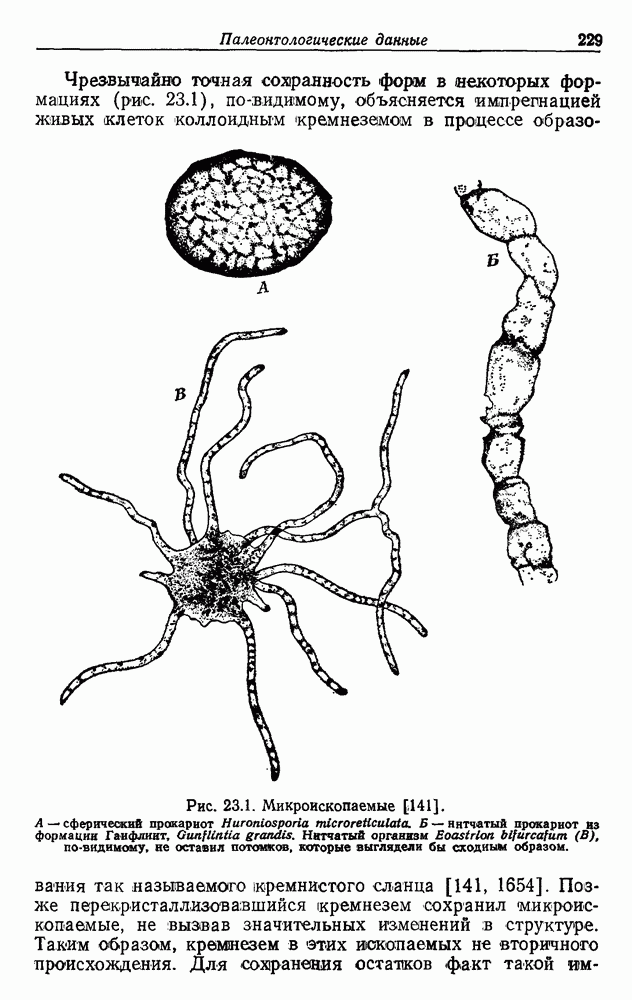
Е. ПРОИСХОЖДЕНИЕ АЭРОБОВОсновываясь на гипотезе конверсии, мы можем считать, что факультативные аэробы среди фотоорганотрофов, в частности несерные пурпурные бактерии Дрюс создал терминологию для обозначения структур клетки, которые, независимо от функции, можно употреблять при изучении родства между клеточной мембраной (КМ) и внутриклеточной мембранной системой (ВКМ). Эта последняя может занимать большую часть объема клетки. По-видимому, она возникла путем инвагинации клеточной мембраны. Фотосинтетическая система пурпурных бактерий в основном, но не исключительно, находится в ВКМ, дыхательная система — в КМ или близ нее. Следовательно, топографическое разделение двух систем, синтезирующих АТФ, неполно. Некоторые авторы склонны считать, что две системы более или менее отделены друг от друга [1749, 1750, 1513]. Другие предполагают существование сложных циклов, служащих и для фотосинтеза, и для дыхания [878, 1331, 1924]. Как бы там ни было, совершенно очевидно, что обе системы находятся в тесном взаимодействии [401, 504, 656, 663, 1147, 1148, 1245, 1344, 1856, 1910]. Для той и другой систем характерны явления индукции и репрессии, в значительной степени! обратимые. Интересно, что были получены мутанты несерной пурпурной бактерии Взаимное влияние фотосинтеза и дыхания наблюдалось также у сине-зеленых водорослей [292, 606, 1423, 1425]. Более слабая связь между этими процессами у эукариотических растений, несомненно, является следствием разделения митохондрий и хлоропластов, в результате которого и те и другие имеют четко определенные функции. В эукариотических растениях могут одновременно протекать процессы фотосинтеза и дыхания с максимальной интенсивностью, причем в обоих случаях будет синтезироваться АТФ. Позже многие из фотосинтезирующих бактерий, ставшие аэробными, утратили способность к фотосинтезу, превратившись в чисто дышащие организмы, т. е. в строгих аэробов среди бактерий. Утрата способности к фотосинтезу хорошо известна также среди сине-зеленых водорослей, от которых произошли бесцветные «скользящие бактерии», энергетика которых основана на процессах дыхания По аналогии с этим дышащие бесцветные серные бактерии (15) могут происходить от фотосинтезирующих окрашенных серобактерий (11). Остается открытым вопрос о том, в каком временном порядке появились облигатные и факультативные дышащие организмы после превращения фотосинтезирующих в дышащие. Фотосинтезирующие бактерии, способные к росту за счет одного брожения, сейчас представляют собой редкое исключение
|
 |
Оглавление
|













































































































































































































































 например
например  должны напоминать бактерий, бывших промежуточным звеном между фотосинтезирующими и дышащими. Таким образом, эти организмы могут служить моделями для понимания перехода от фотосинтеза к дыханию. И в самом деле различные авторы уделили большое внимание структурному и функциональному родству фотосинтезирующей и дыхательной систем у фотоорганотрофов.
должны напоминать бактерий, бывших промежуточным звеном между фотосинтезирующими и дышащими. Таким образом, эти организмы могут служить моделями для понимания перехода от фотосинтеза к дыханию. И в самом деле различные авторы уделили большое внимание структурному и функциональному родству фотосинтезирующей и дыхательной систем у фотоорганотрофов.
 которые в аэробных условиях в темноте вырабатывают фотосинтезирующие пигменты [1095]. Некоторые из этих мутантов тем не менее хорошо растут в аэробных условиях. С другой стороны, не способный к дыханию мутант другого вида Rhodopseudomonas является ценным экспериментальным объектом при изучении механизма подавления образования хлорофилла кислородом [1217]. В литературе указывалось, что несерные пурпурные бактерии, например
которые в аэробных условиях в темноте вырабатывают фотосинтезирующие пигменты [1095]. Некоторые из этих мутантов тем не менее хорошо растут в аэробных условиях. С другой стороны, не способный к дыханию мутант другого вида Rhodopseudomonas является ценным экспериментальным объектом при изучении механизма подавления образования хлорофилла кислородом [1217]. В литературе указывалось, что несерные пурпурные бактерии, например  способные к фотосинтетическому и окислительному фосфорилированию, представляют собой уникальные объекты для изучения энергетики роста бактериофага [1642].
способные к фотосинтетическому и окислительному фосфорилированию, представляют собой уникальные объекты для изучения энергетики роста бактериофага [1642].
 .
.
 Тем не менее возможно, что от таких организмов произошли факультативно дышащие. А быть может, облигатно дышащие организмы произошли от строго фотосинтезирующих, а позднее у них вновь развилась и заняла важное место сохранившаяся уже навсегда, но часто скрытая способность к брожению. Молочнокислые бактерии
Тем не менее возможно, что от таких организмов произошли факультативно дышащие. А быть может, облигатно дышащие организмы произошли от строго фотосинтезирующих, а позднее у них вновь развилась и заняла важное место сохранившаяся уже навсегда, но часто скрытая способность к брожению. Молочнокислые бактерии  уникальны среди анаэробов (это скорее вопрос терминологии) - в том смысле, что они вполне толерантны к кислороду. Более того, у некоторых из них обнаружены развитые до некоторой степени дыхательные системы и гемопротеиды. Они микроаэрофильны. На этом и на других основаниях было выдвинуто предположение, что молочнокислые бактерии регрессировали из аэробного состояния [301, 1979, 1980]. Пропионовокислые бактерии, которые тоже могли регрессировать, имеют больше гемопротеидов, чем молочнокислые, но они лишь микроаэротолерантны [458, 1675, 1784]. Возможно, что когда-то существовали «первичные» толерантные к кислороду бактерии, которые не регрессировали от бактерий, способных к использованию кислорода.
уникальны среди анаэробов (это скорее вопрос терминологии) - в том смысле, что они вполне толерантны к кислороду. Более того, у некоторых из них обнаружены развитые до некоторой степени дыхательные системы и гемопротеиды. Они микроаэрофильны. На этом и на других основаниях было выдвинуто предположение, что молочнокислые бактерии регрессировали из аэробного состояния [301, 1979, 1980]. Пропионовокислые бактерии, которые тоже могли регрессировать, имеют больше гемопротеидов, чем молочнокислые, но они лишь микроаэротолерантны [458, 1675, 1784]. Возможно, что когда-то существовали «первичные» толерантные к кислороду бактерии, которые не регрессировали от бактерий, способных к использованию кислорода.