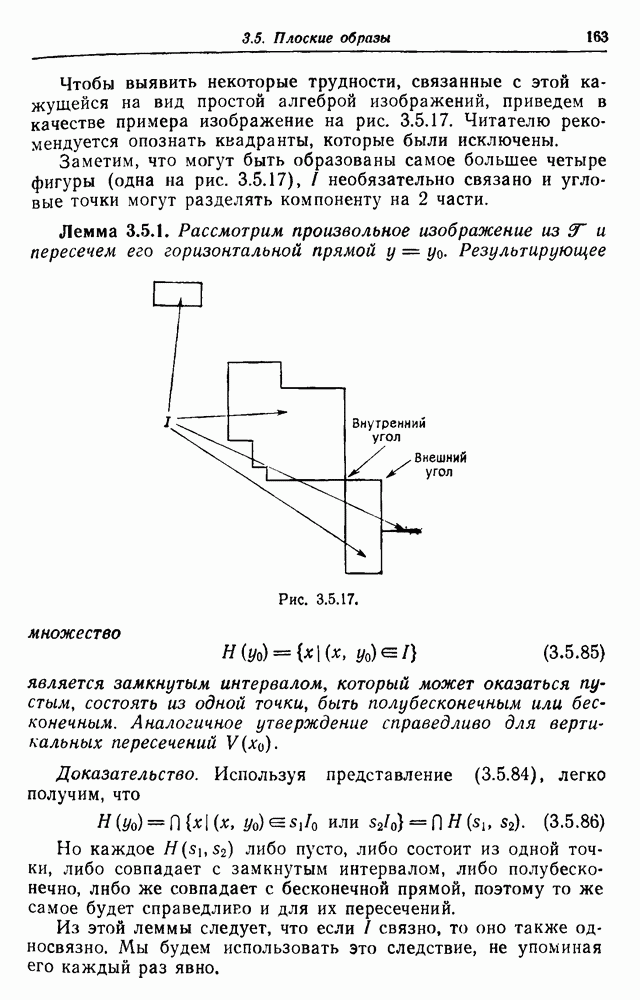
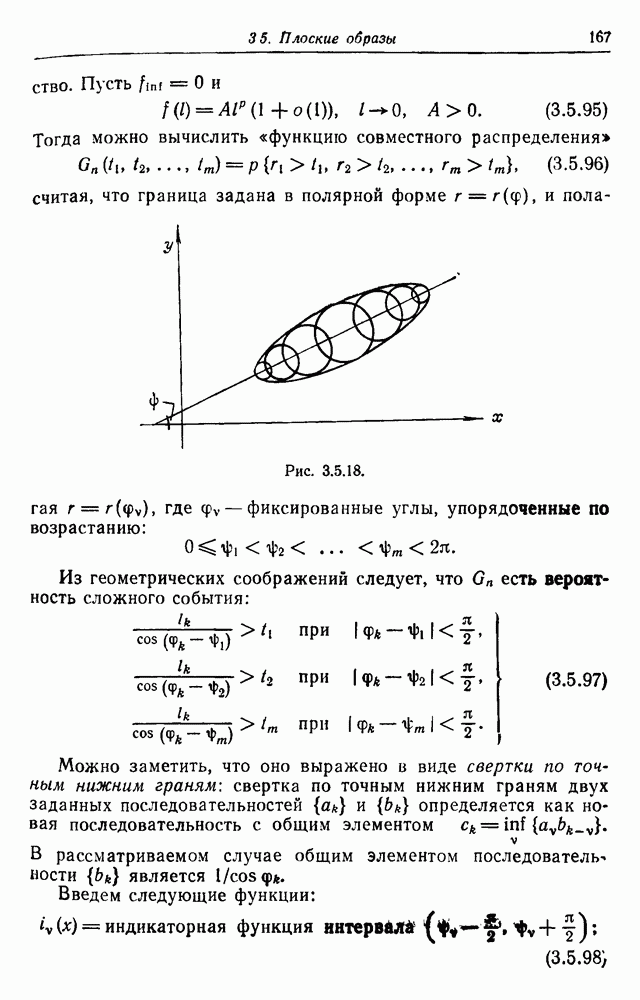
Пред.
След.
Макеты страниц
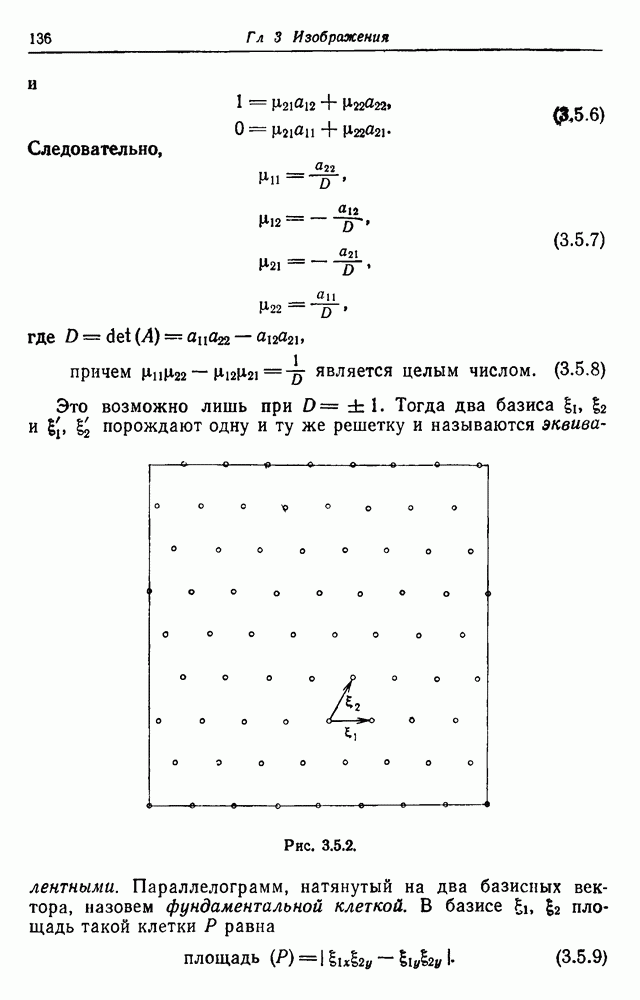
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
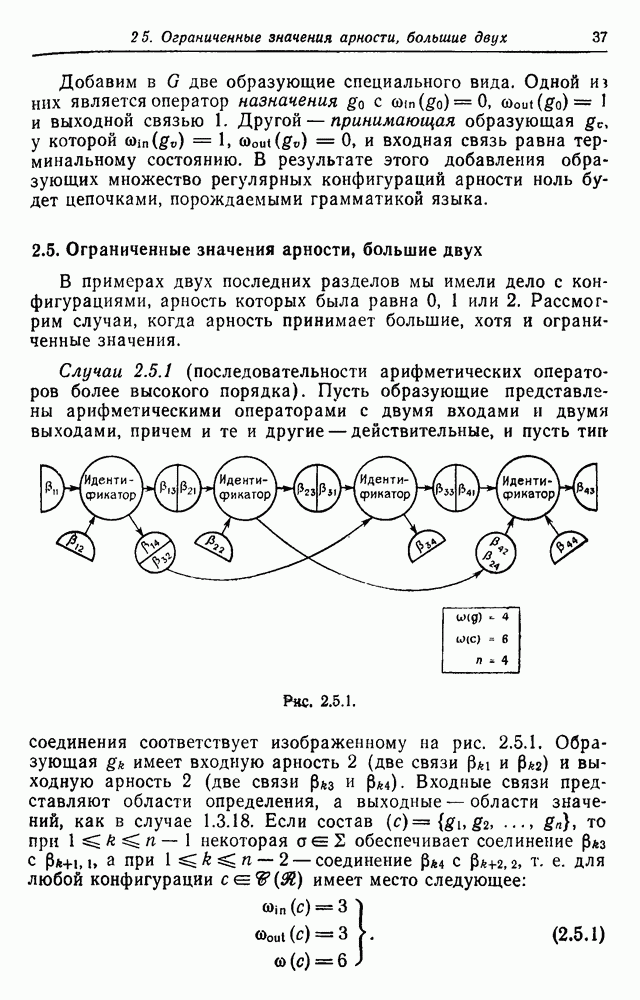
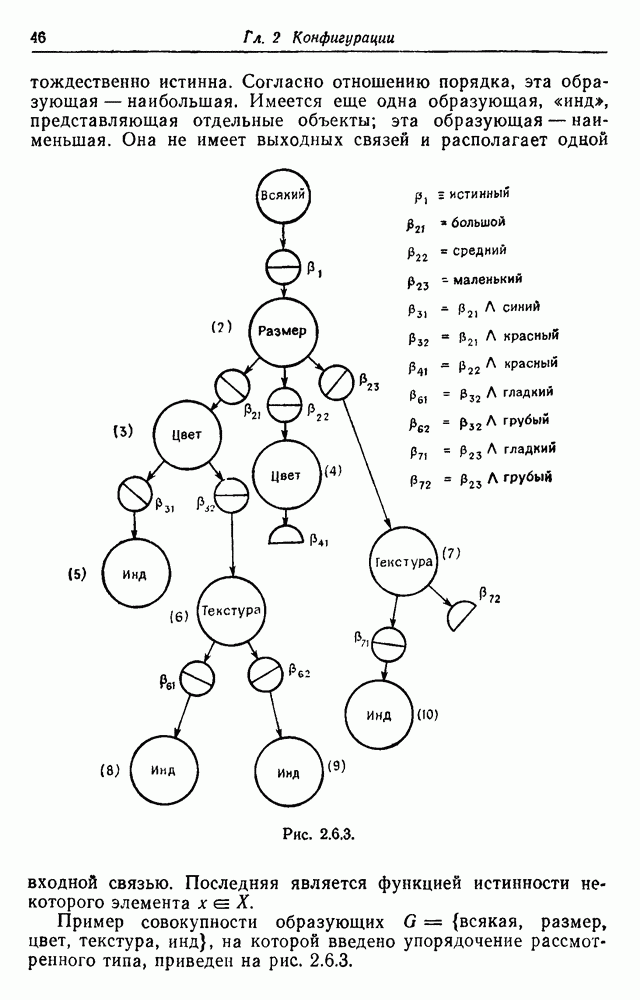
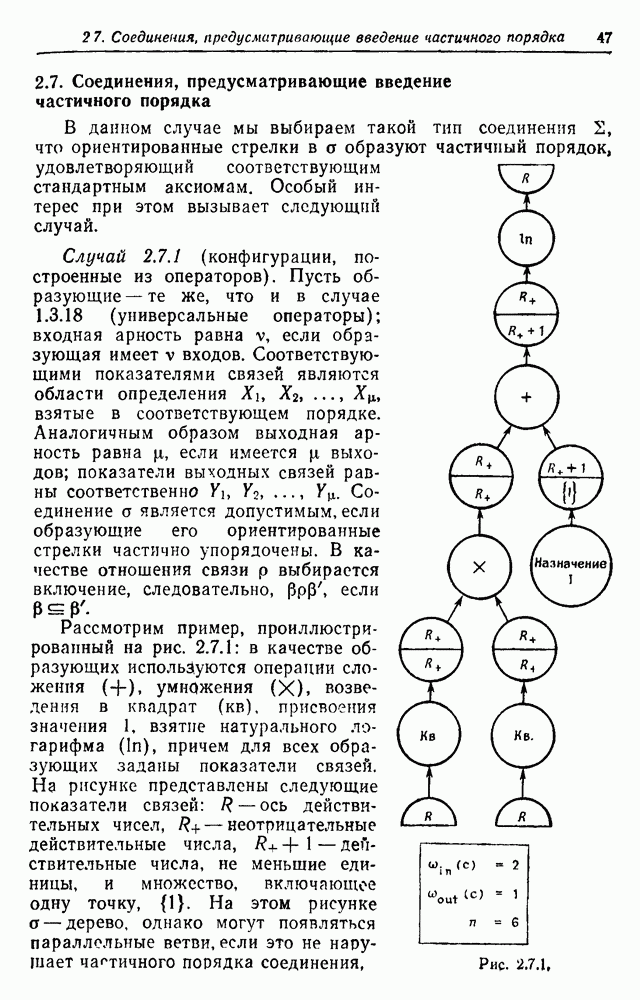
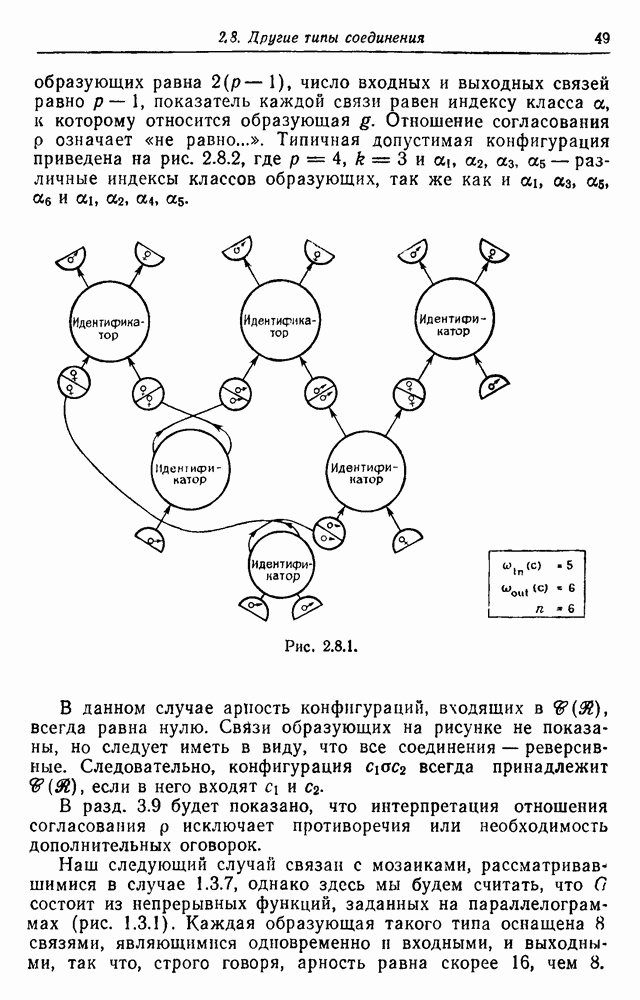
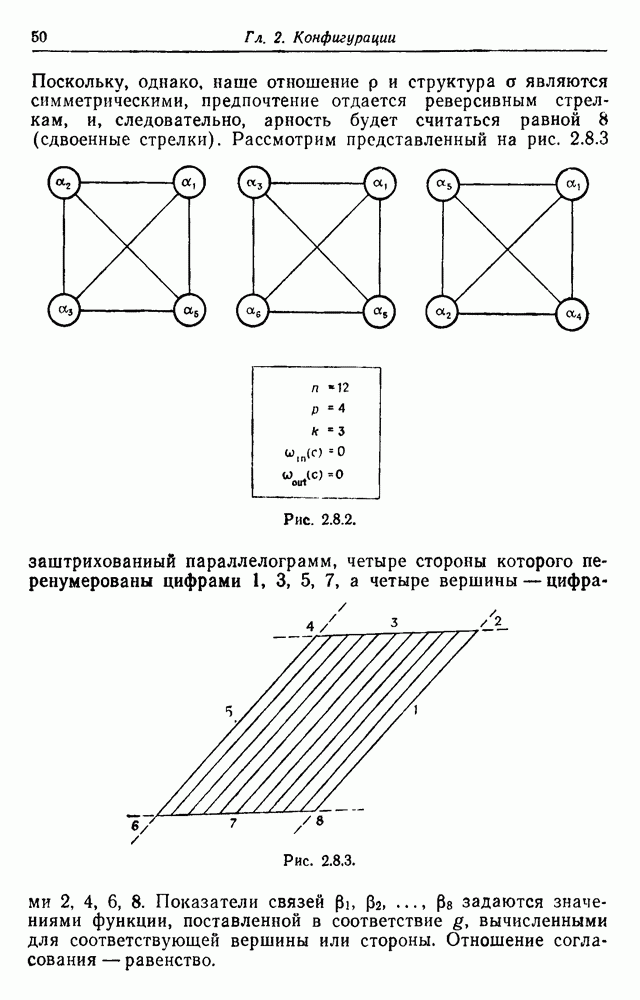
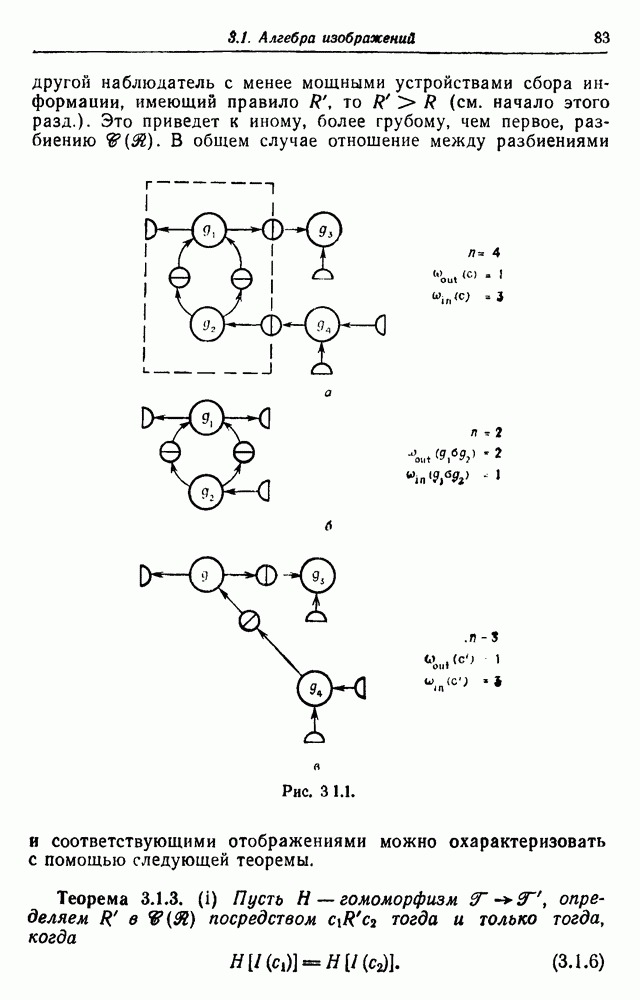
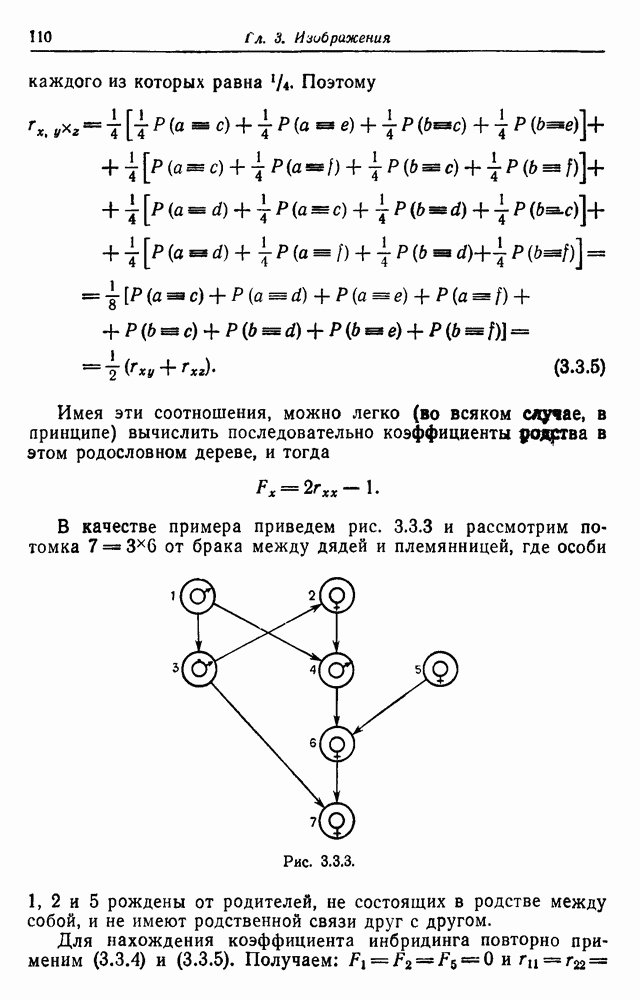
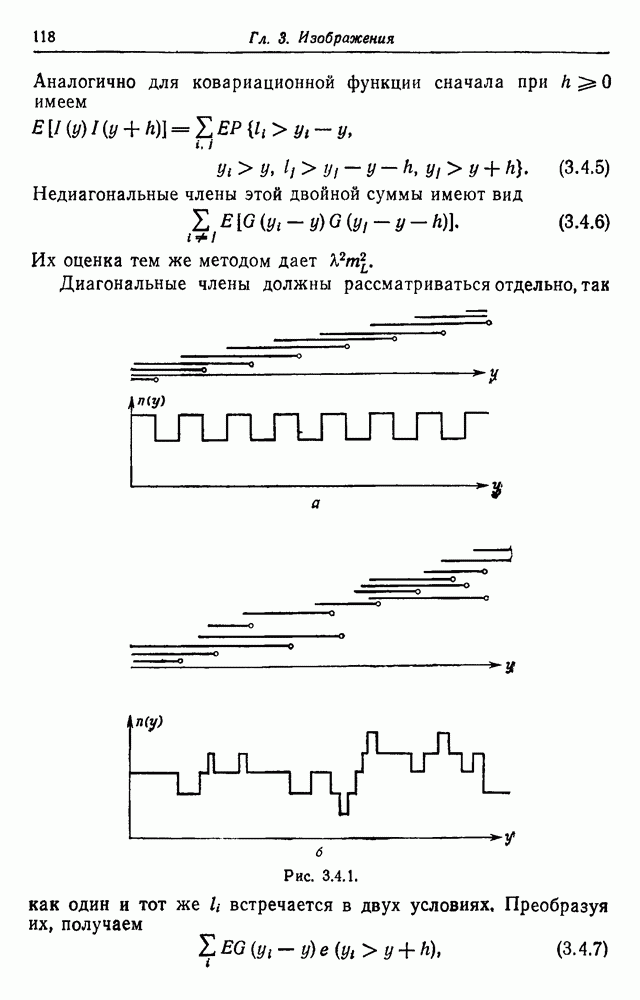
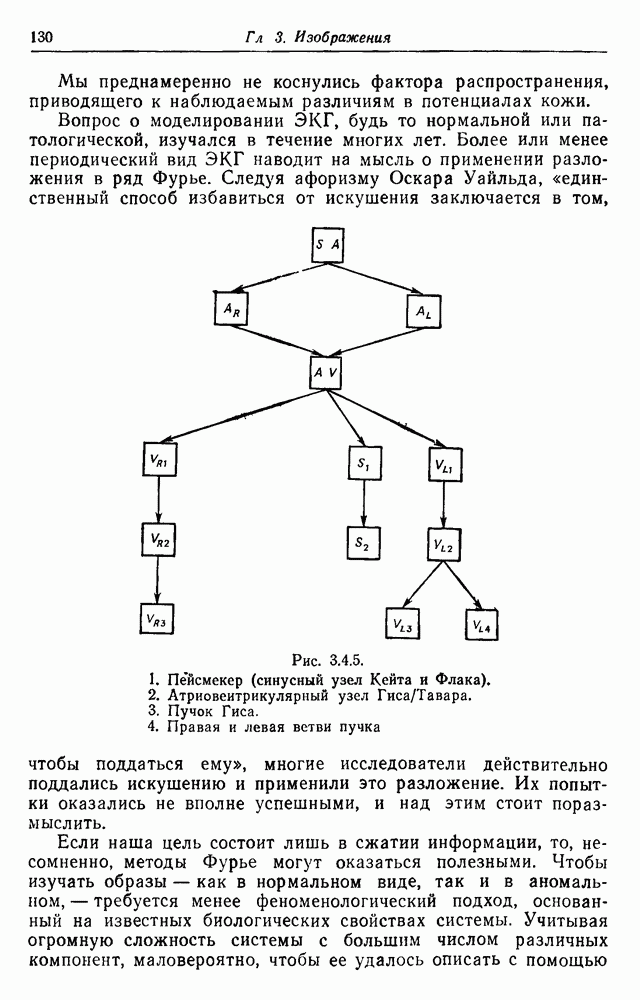
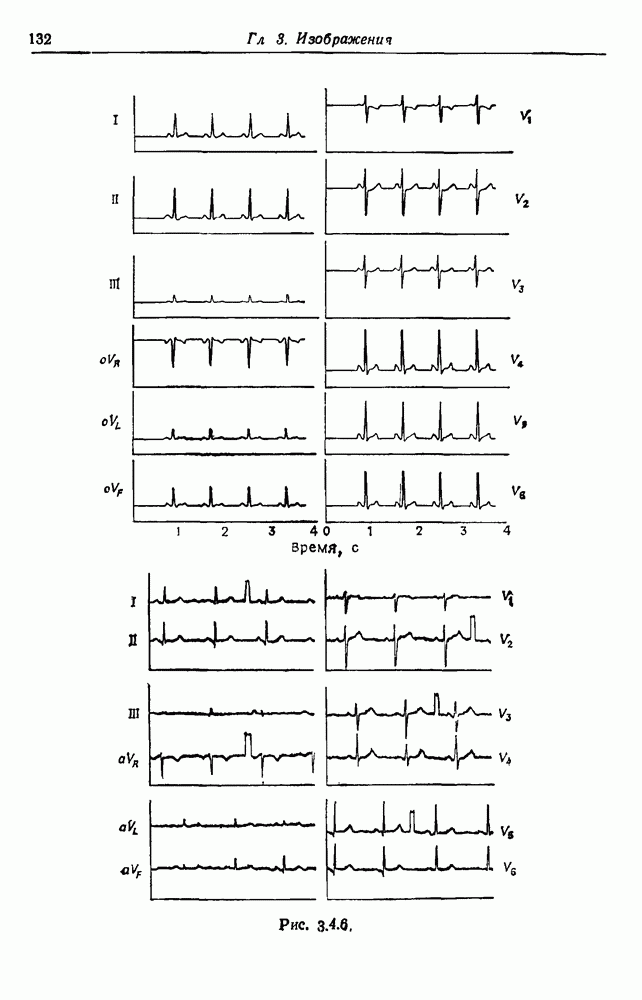
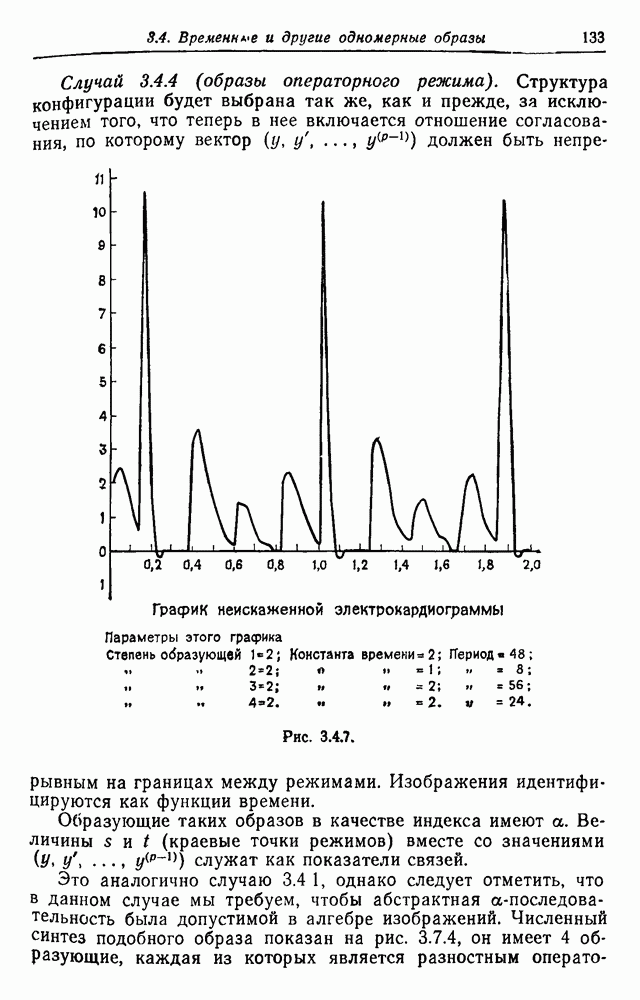
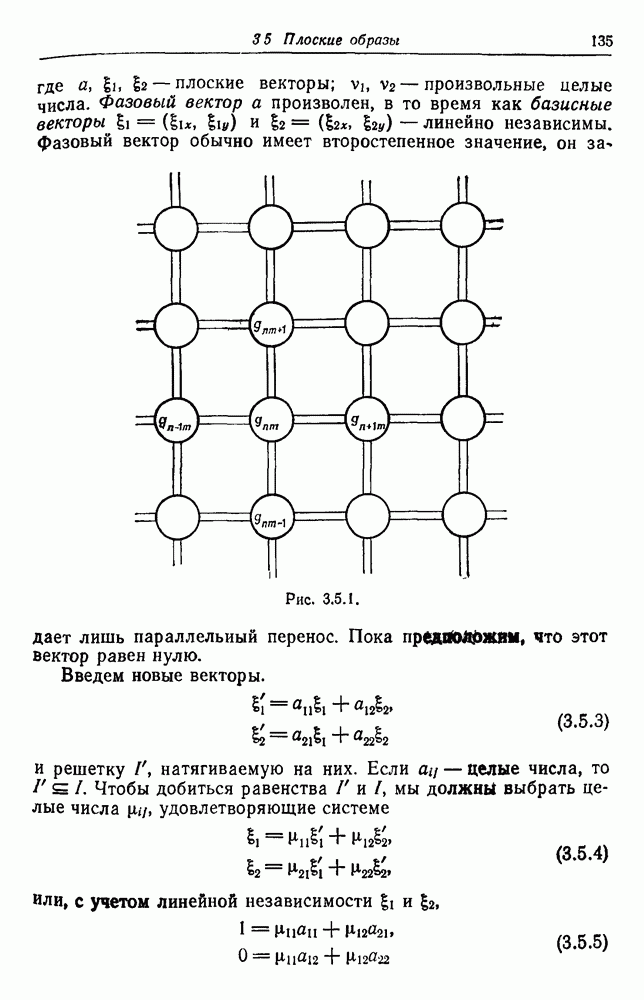
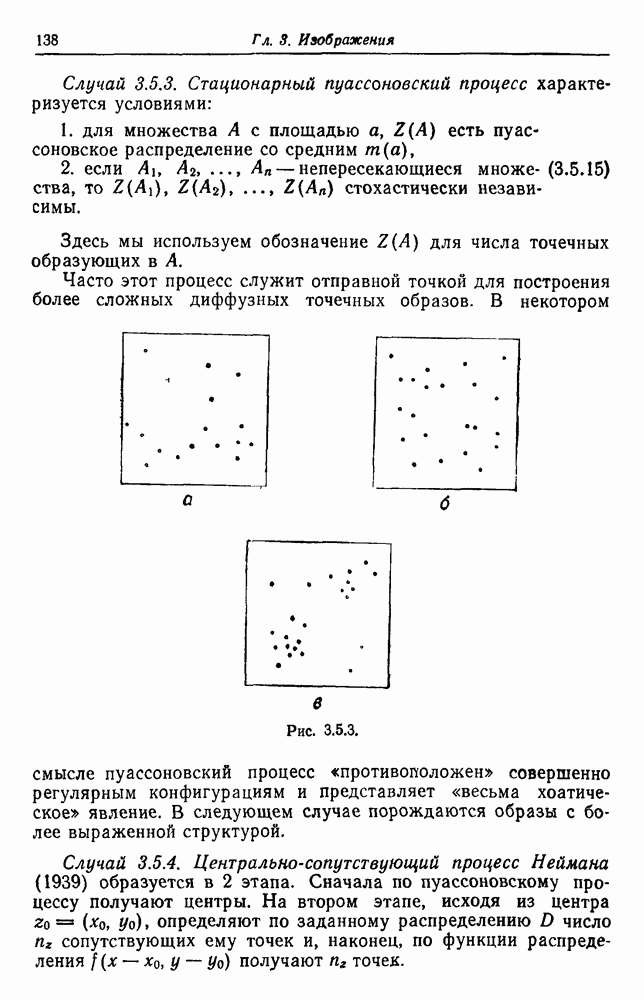
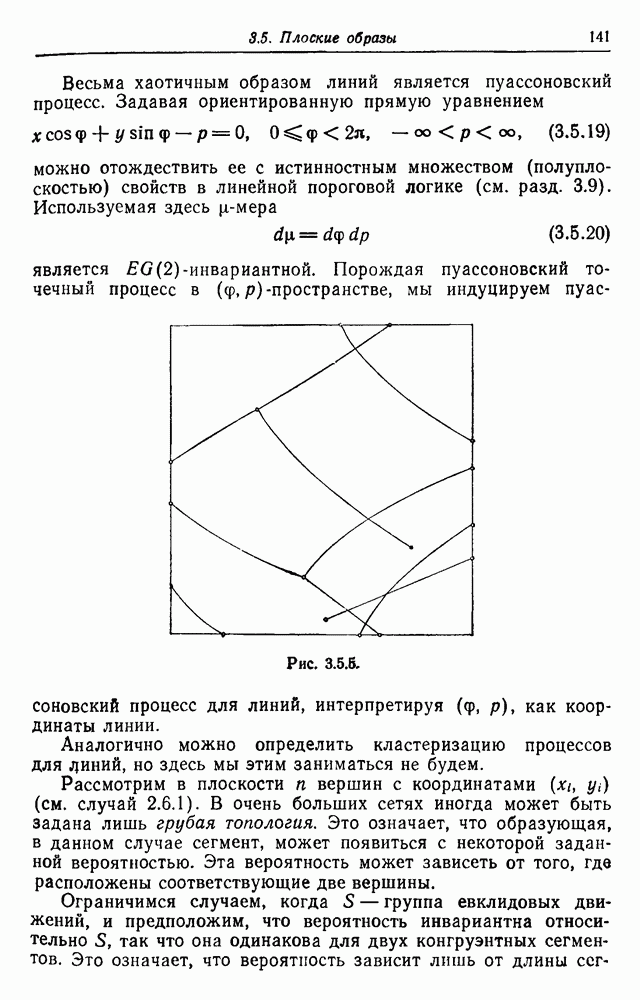
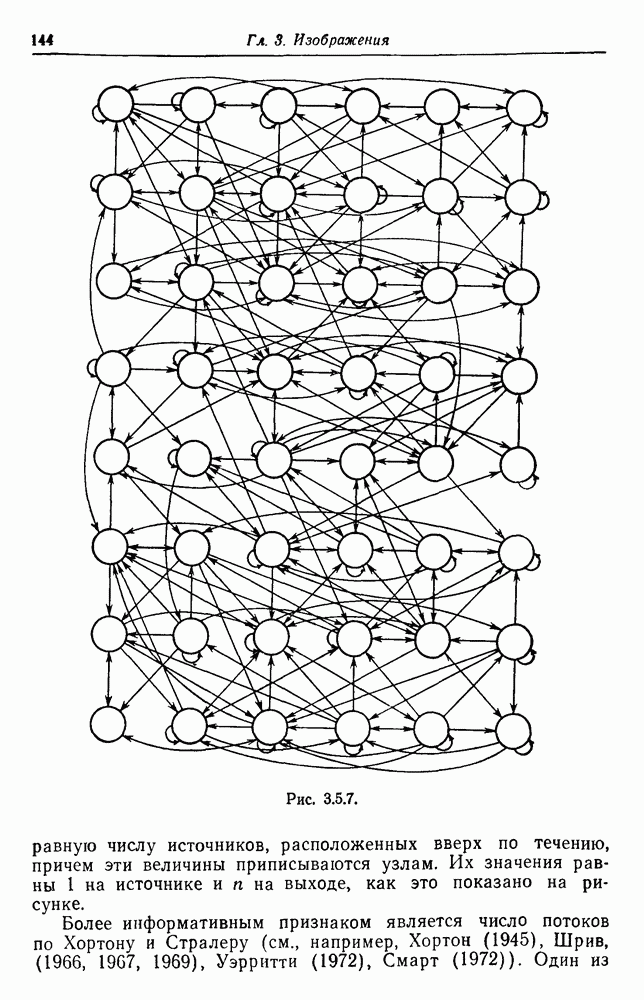
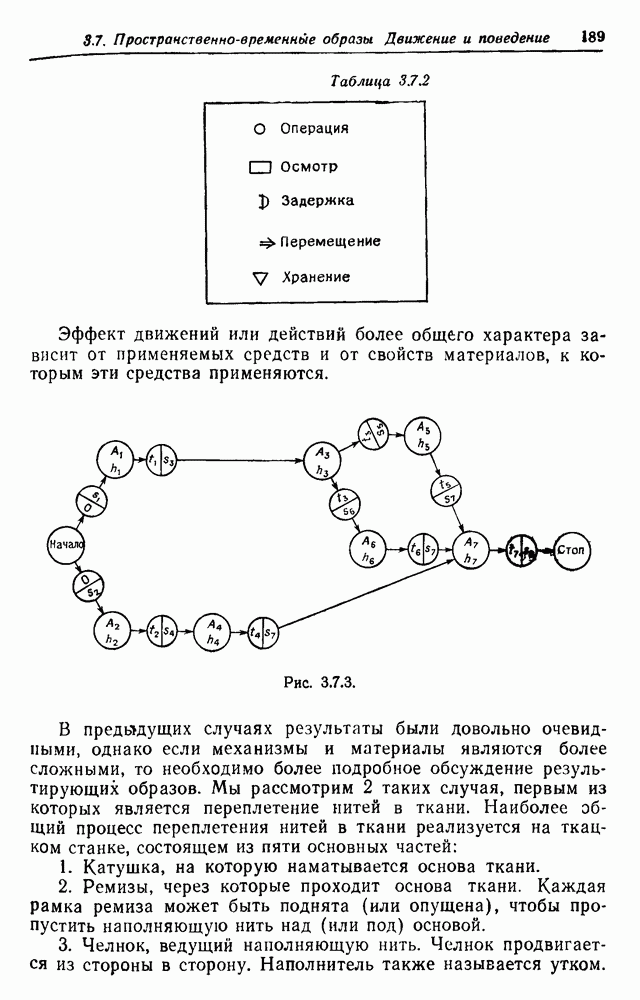
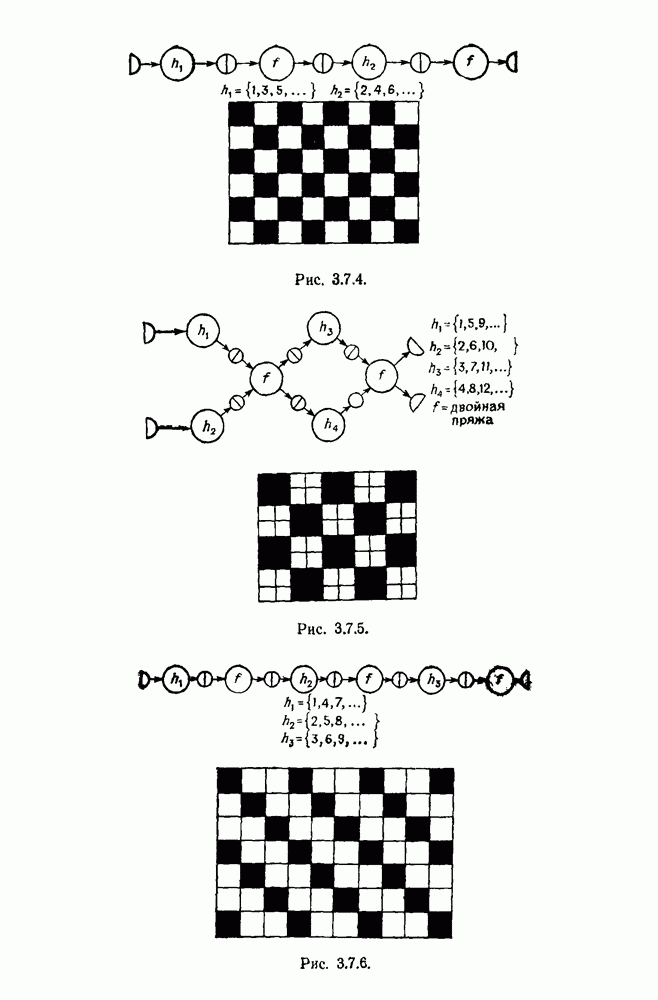
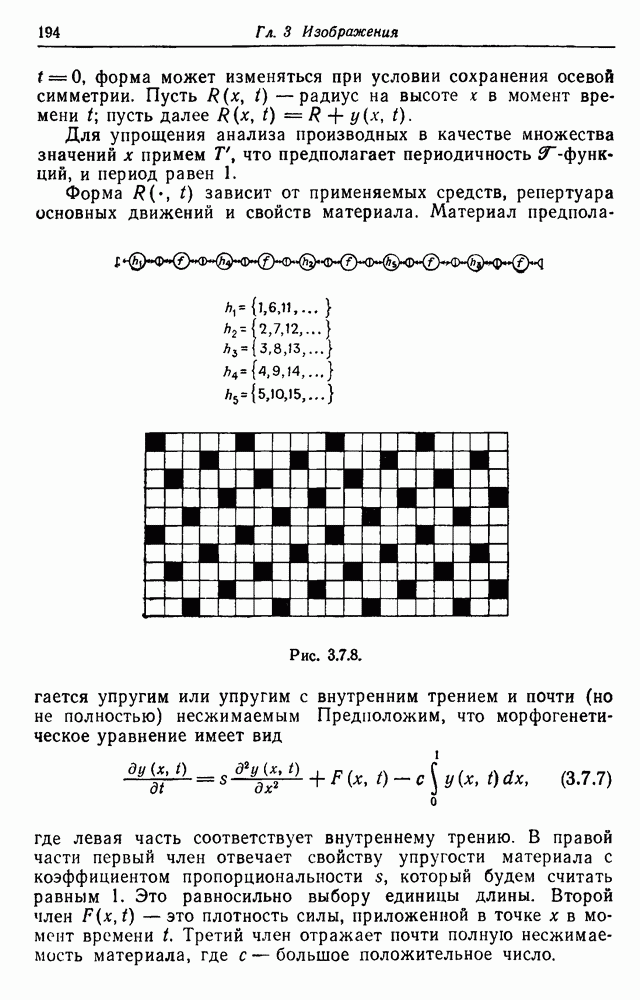
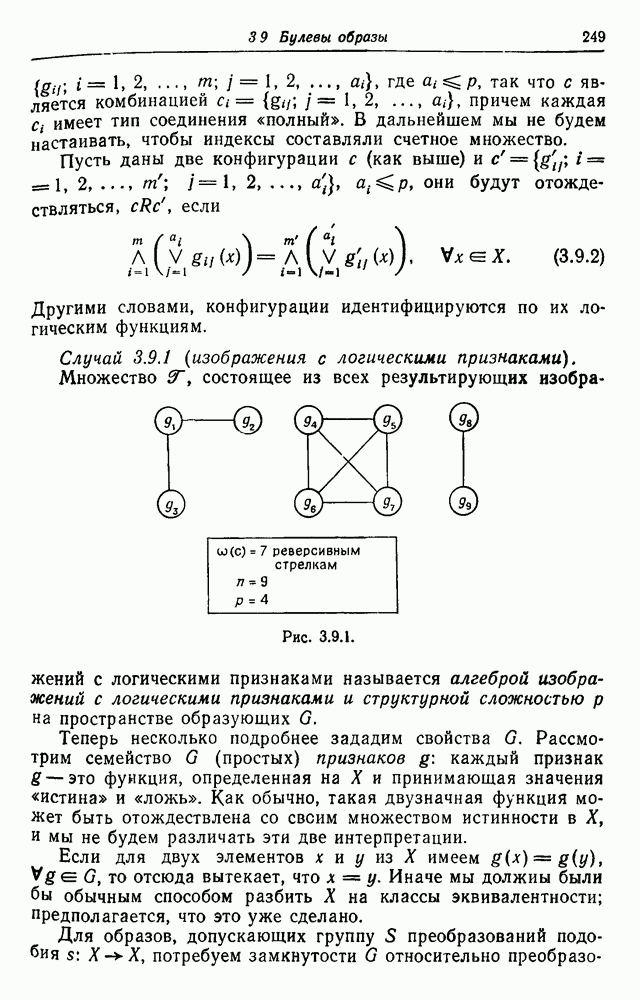
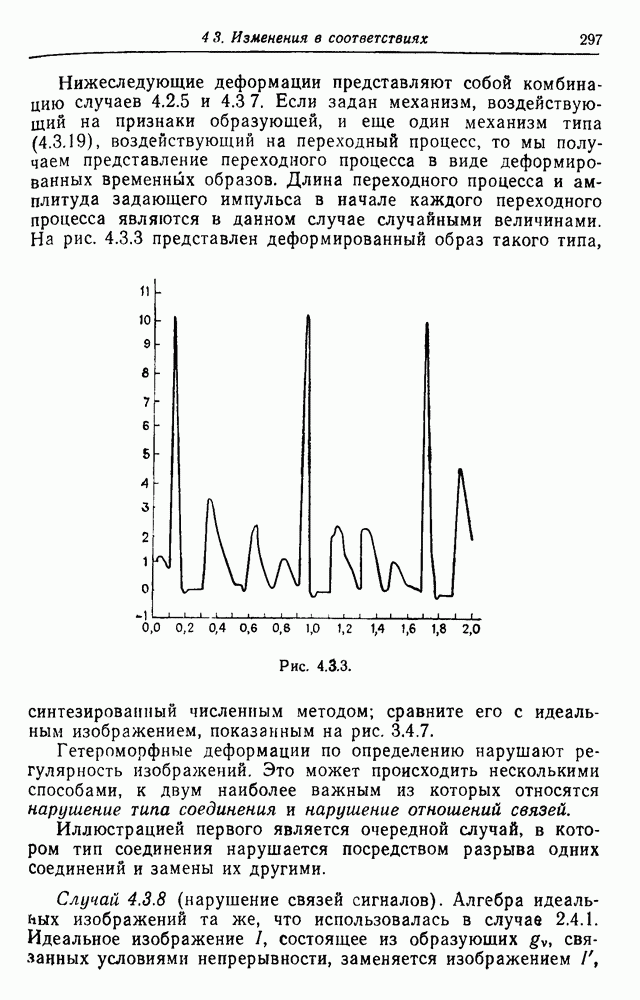
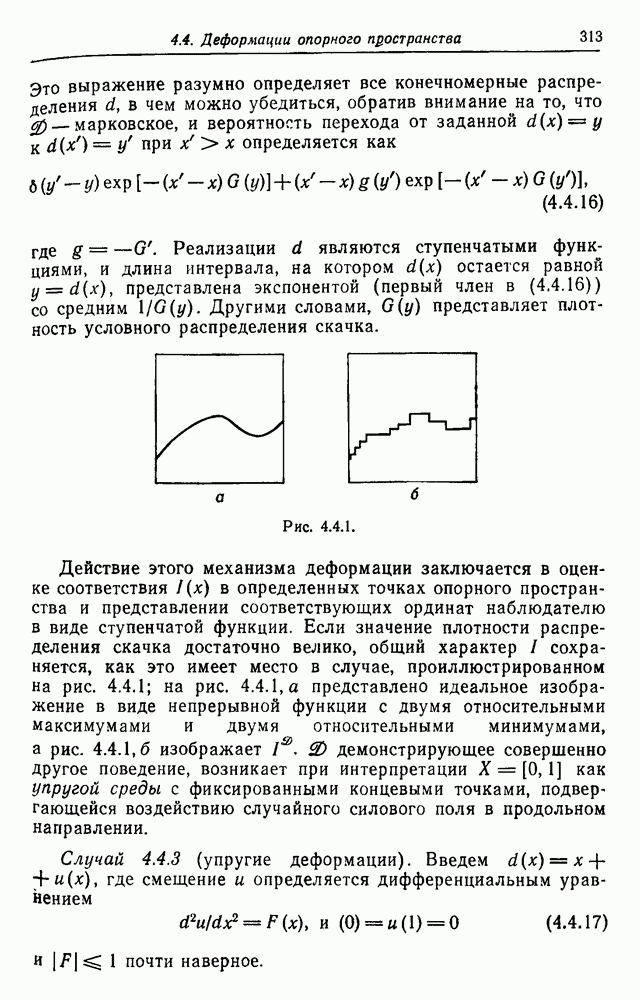
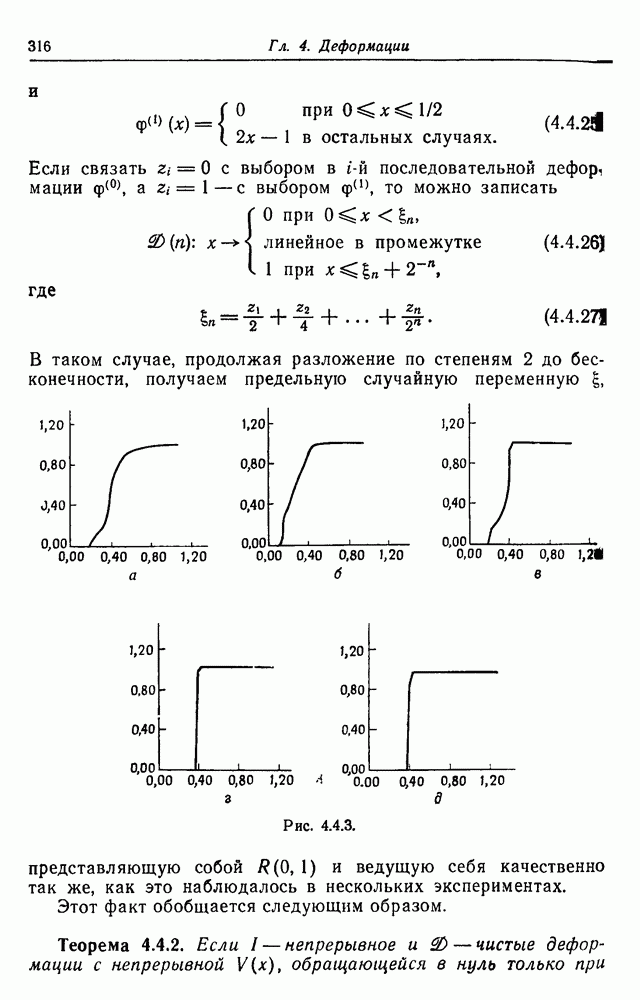
3.3. Другие абстрактные изображенияПусть образующие по-прежнему являются абстрактными символами, смысл которых выражается непосредственно без обращения к среде-носителю информации или опорному пространству. Теперь рассмотрим несколько случаев, когда тип соединения 2 не является ни линейным, ни древовидным. Случай 3.3.1. Раскрашенные графы. Пусть образующие перечисляются так: (i) n = m, (ii) существует перестановка Можно считать, что Очень сложные изображения встречаются при изучении родственных связей, которые столь дороги сердцам антропологов и специалистов по генетике популяций. В частности, будут чены два взаимодополняющих друг друга аспекта, инбридинг и запреты на кровосмешения. Рассмотрим родословное дерево, приведенное на рис. 3.3.2, указывающее некоторых прародителей известного быка Комета. В данном случае имеем лишь два признака — самцов и самок:
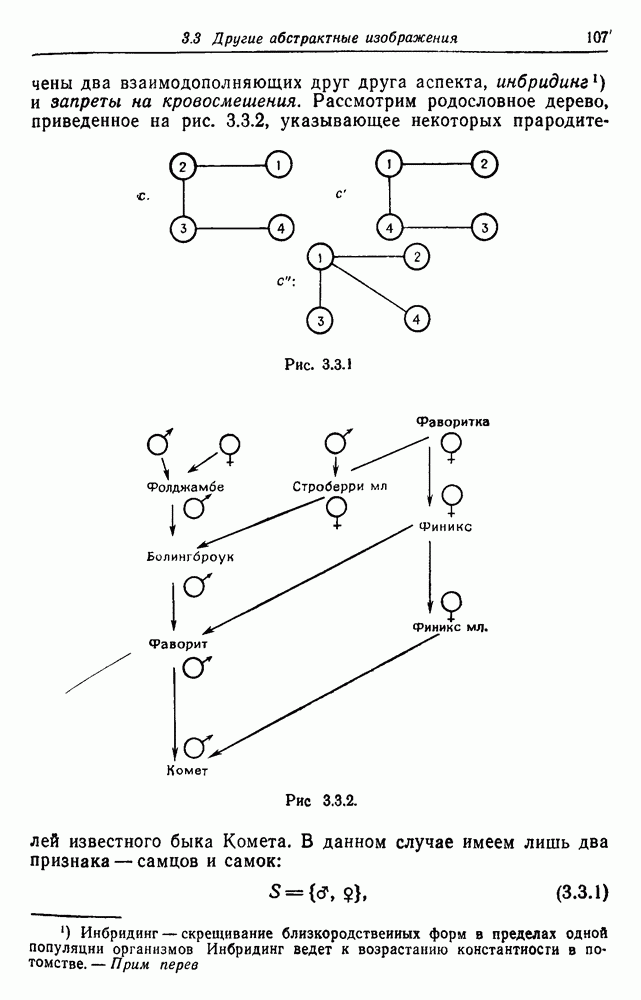
Рис. 3.3.1 (см. скан) и их различные представители обозначаются именами. Это небольшое изображение не обладает заметной регулярностью, однако благодаря инбридингу существуют некоторые кратные родственные отношения. Комет — внук Финикс как по материнской линии через Финикс мл., так и по отцовской линии через Фаворита. Он также является собственным кузеном и племянником своей матери. Выбор типа соединения 2 зависит от системы спаривания, которую мы обсудим позднее. Во всяком случае, ясно, что петли всегда исключаются. Ясно также, что родословное дерево такого типа имеет не древовидную структуру, так как отдельная особь может происходить от двух особей с общим прародителем. Чтобы задать регулярную конфигурацию, мы должны определить два фактора: правила спаривания и степень полноты графа. В сообществах людей браки обычно ограничиваются запретами, исключающими некоторые родственные отношения. Они широко варьируются, и мы должны соответствующим образом выбирать эти ограничения в каждом конкретном случае. Могут также существовать правила, определяющие предпочтительные браки в группах. С другой стороны, полнота должна означать, что никакие родственные отношения не исключаются из графа. Однако для любой конечной конфигурации всегда будут существовать элементы, в которых присутствует не более одного родителя, и такие элементы, в которых все дети принадлежат конфигурации. В качестве минимальных ограничений примем следующие. 1. С каждым элементом с связан не более чем один родитель мужского пола и не более чем один родитель женского пола. 2. Для элементов с допускаются все отношения между родителями и детьми. Случайные источники допустимых конфигураций представляют интерес как в генетике, так и в антропологии, и проще всего определять их рекурсивно. Для заданного множества образующих необходима следующая детализация. 1. Случайный механизм для спаривания образующих в соответствии с ограничениями 52. 2. Случайный механизм для определения числа и пола появившихся потомков. 3. Механизм, определяющий, как именно поколения следуют друг за другом, например путем задания распределения продолжительности жизни и той ее части, когда возможно размножение. Раз мы имеем конфигурацию, возникает вопрос о ее функциональном описании. Для того чтобы охарактеризовать инбридинг и родство в родословных образах, Райтом (1941) были выдвинуты следующие соображения, уточненные впоследствии Малеко (1948). Рассмотрим два гена популяции, С другой стороны, может случиться, что они идентичны, но не происходят от одной и той же хромосомы какого-либо общего предка. Тогда Коэффициент инбридинга
Для двух особей
Между
С другой стороны, рассмотрим потомка
так что (см. уравнение (3.3.2))
Более общее соотношение получается следующим образом. Рассмотрим особей каждого из которых равна
Имея эти соотношения, можно легко (во всяком случае, в принципе) вычислить последовательно коэффициенты родрва в этом родословном дереве, и тогда
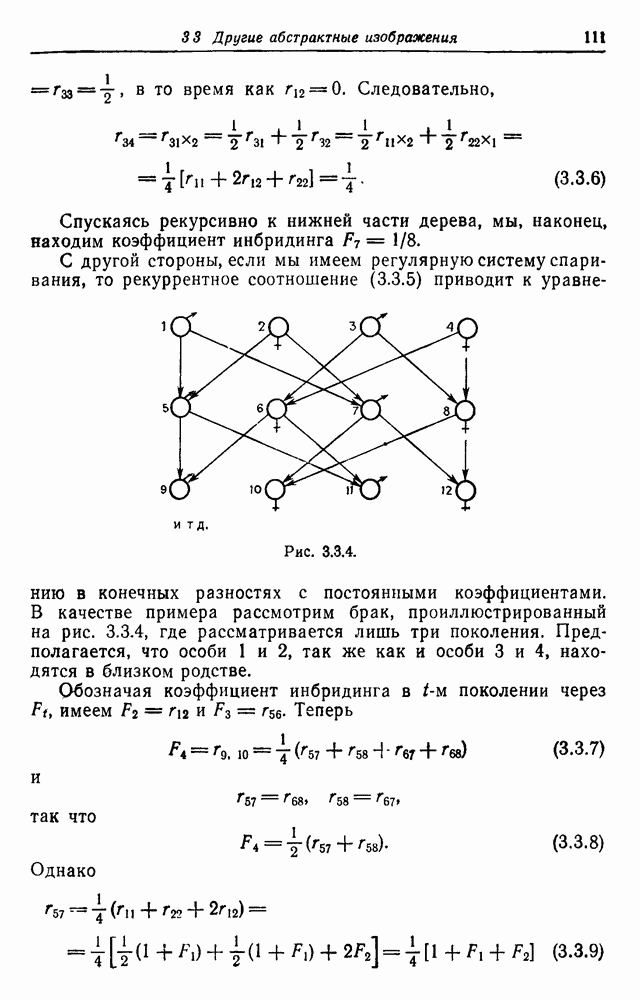
В качестве примера приведем рис. 3.3.3 и рассмотрим потомка
Рис. 3.3.3. Для нахождения коэффициента инбридинга повторно применим (3.3.4) и (3.3.5). Получаем:
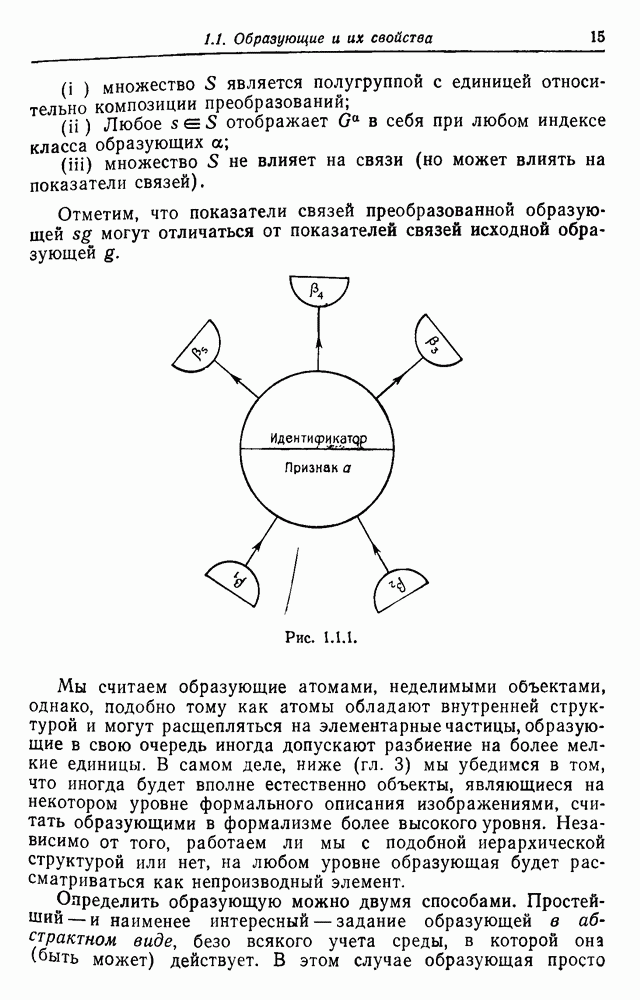
Спускаясь рекурсивно к нижней части дерева, мы, наконец, находим коэффициент инбридинга
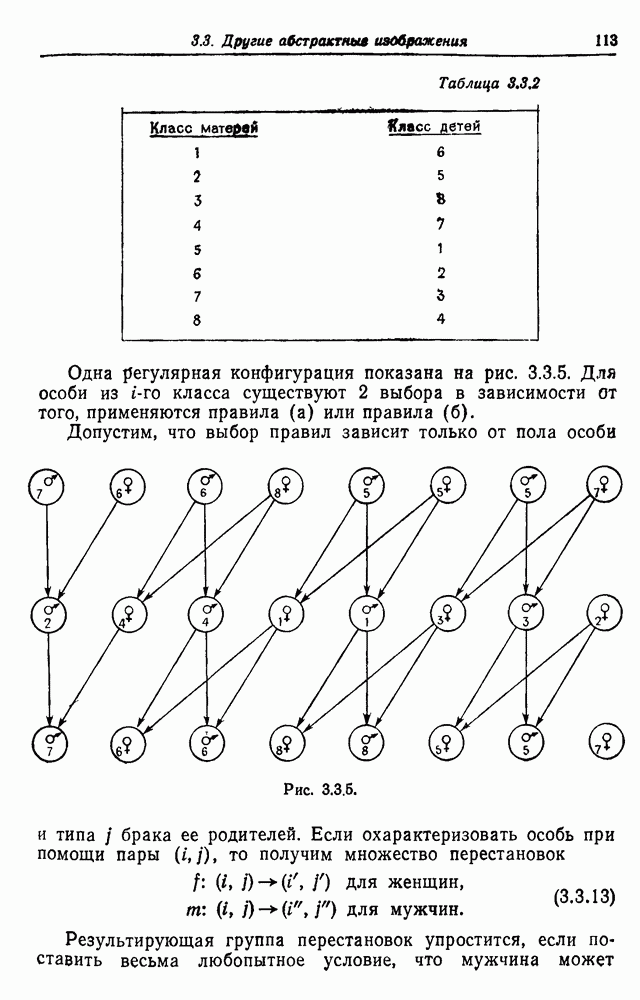
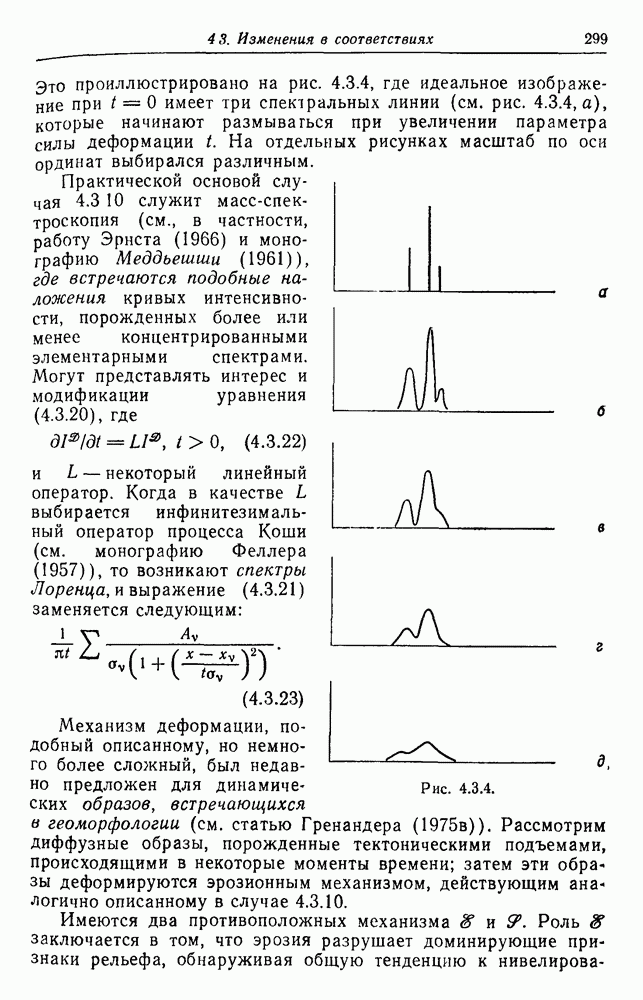
Рис. 3.3.4. С другой стороны, если мы имеем регулярную систему спаривания, то рекуррентное соотношение (3.3.5) приводит к уравнению в конечных разностях с постоянными коэффициентами. В качестве примера рассмотрим брак, проиллюстрированный на рис. 3.3.4, где рассматривается лишь три поколения. Предполагается, что особи 1 и 2, так же как и особи 3 и 4, находятся в близком родстве. Обозначая коэффициент инбридинга в
и
так что
Однако
и
Аналогичным путем для произвольного поколения
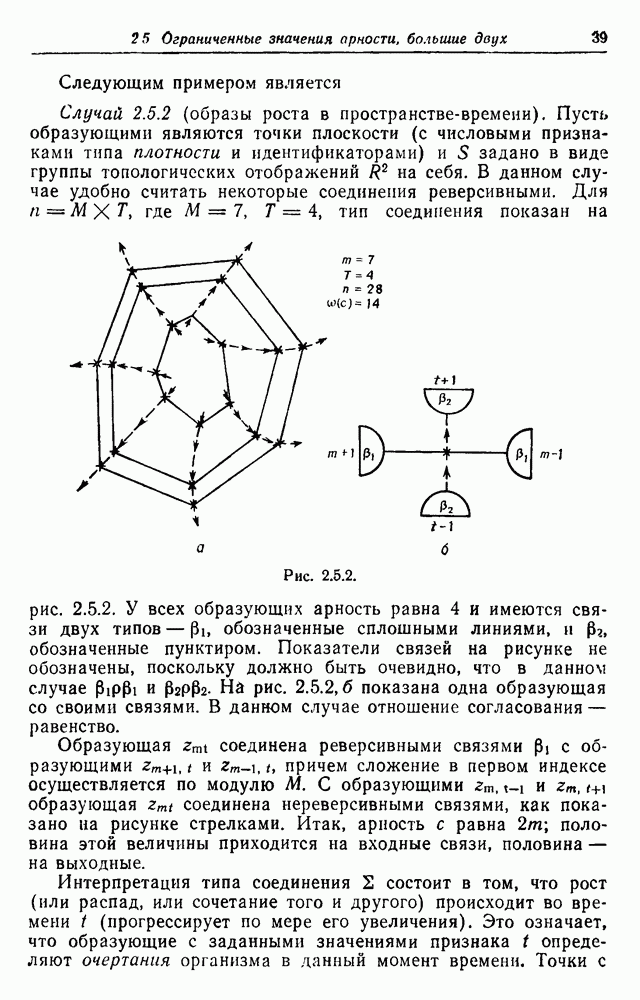
которое можно решить стандартным методом. Таким образом, мы можем описать особей с точки зрения инбридинга, и, накапливая статистику, говорить о среднем инбридинге и т. д. Рассматривая с другой точки зрения конфигурацию того же типа, можно поставить вопрос: какими структурными свойствами должны обладать результирующие конфигурации, если заданы ограничения на бракосочетания. Известным примером такого рода является система Мернгина (см. замечания). Популяция разбивается на восемь групп:
Возможные брачные отношения приведены в табл. 3.3.1, состоящей из двух множеств правил: а и б. Мужчина из первой группы может жениться на женщине из третьей группы (см. (а)) или из четвертой группы (см. (б)), мужчина из второй группы может жениться на женщине из четвертой группы (см. (а)) или из третьей группы и т. д. В нашем примере существует 16 брачных типов. Присвоим им номера Таблица 3.3.1а (см. скан) Таблица 3.3.1б (см. скан) Таблица 3.3.2 (см. скан) Одна регулярная конфигурация показана на рис. 3.3.5. Для особи из
Рис. 3.3.5 Допустим, что выбор правил зависит только от пола особи и типа
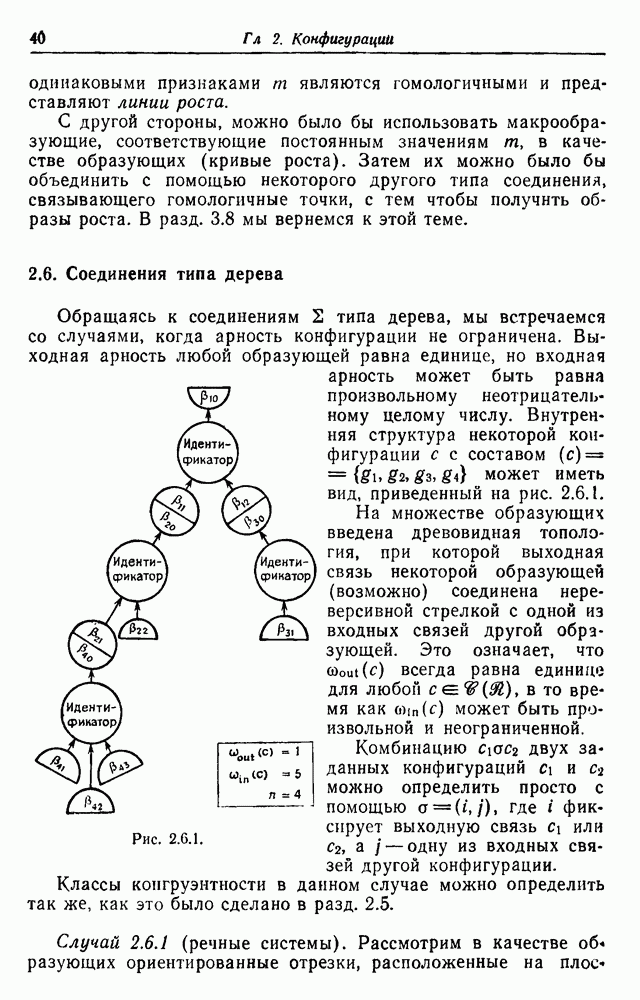
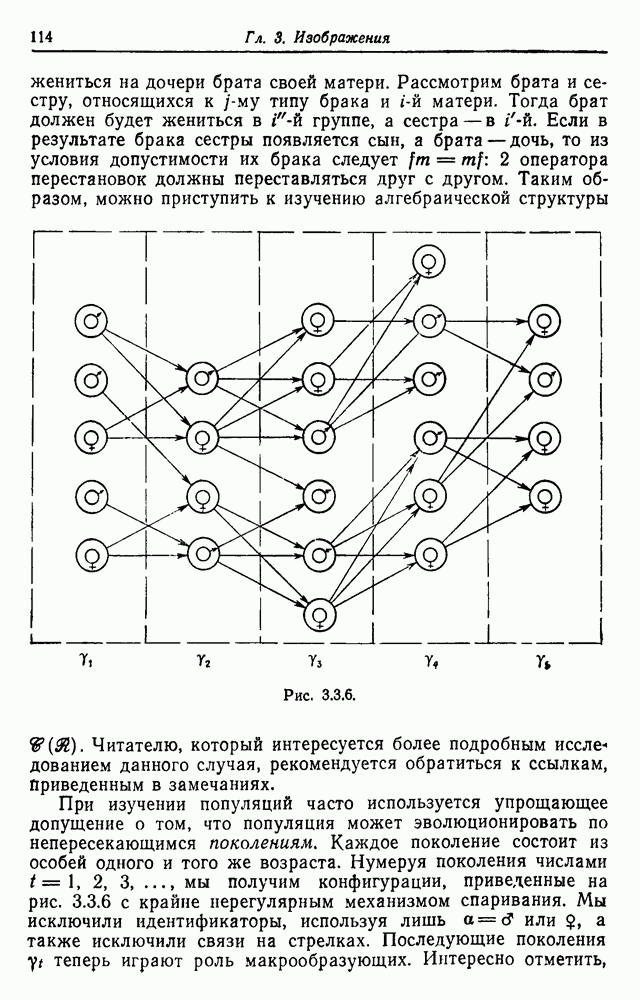
Результирующая группа перестановок упростится, если поставить весьма любопытное условие, что мужчина может жениться на дочери брата своей матери. Рассмотрим брата и сестру, относящихся к
Рис. 3.3.6. Читателю, который интересуется более подробным исследованием данного случая, рекомендуется обратиться к ссылкам, Приведенным в замечаниях. При изучении популяций часто используется упрощающее допущение о том, что популяция может эволюционировать по непересекающимся поколениям. Каждое поколение состоит из особей одного и того же возраста. Нумеруя поколения числами что В этом разделе образующие были абстрактными. Изучение аналогичных конфигураций в опорном пространстве будет отложено до разд. 5.5 и части IV.
|
 |
Оглавление
|













































































































































































































































































































 и имеют произвольные соединения в конфигурациях,
и имеют произвольные соединения в конфигурациях,  «истинный». Мы идентифицируем две конфигурации
«истинный». Мы идентифицируем две конфигурации  если
если
 первых
первых  натуральных чисел, такая, что если
натуральных чисел, такая, что если  то
то  соединяются с помощью структуры
соединяются с помощью структуры  тогда и только тогда, когда
тогда и только тогда, когда  соединяются с помощью структуры
соединяются с помощью структуры  . Такое правило
. Такое правило  называется идентификацией при помощи перестановки.
называется идентификацией при помощи перестановки.
 соответствуют краскам, в которые окрашиваются вершины графа связей. На рис. 3.3.1
соответствуют краскам, в которые окрашиваются вершины графа связей. На рис. 3.3.1  идентифицируются как конфигурации, принадлежащие к одному и тому же изображению
идентифицируются как конфигурации, принадлежащие к одному и тому же изображению  (используйте перестановку
(используйте перестановку  однако
однако  не является эквивалентом ни одной из них).
не является эквивалентом ни одной из них).


 Может случиться, что они совпадают как результат их возникновения от одного и того же гена, встречающегося в предках по обеим линиям: тогда мы скажем, что
Может случиться, что они совпадают как результат их возникновения от одного и того же гена, встречающегося в предках по обеим линиям: тогда мы скажем, что  идентичны по происхождению, и запишем:
идентичны по происхождению, и запишем: 
 называются идентичными по состоянию.
называются идентичными по состоянию.
 особи
особи  определяется как вероятность того, что два гена особи
определяется как вероятность того, что два гена особи  в конкретном локусе идентичны по происхождению, так что, если
в конкретном локусе идентичны по происхождению, так что, если  имеет генотип
имеет генотип  то
то

 коэффициент родства
коэффициент родства  определяется как вероятность того, что случайный ген
определяется как вероятность того, что случайный ген  (в конкретном локусе) идентичен по происхождению со случайным геном у (в том же локусе), так что, если
(в конкретном локусе) идентичен по происхождению со случайным геном у (в том же локусе), так что, если  имеет генотип
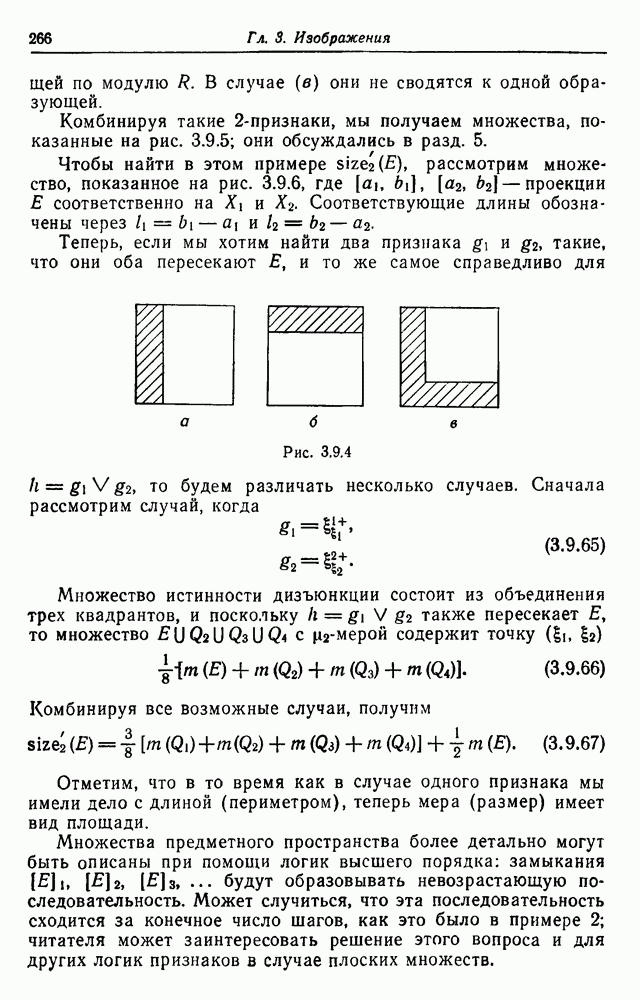
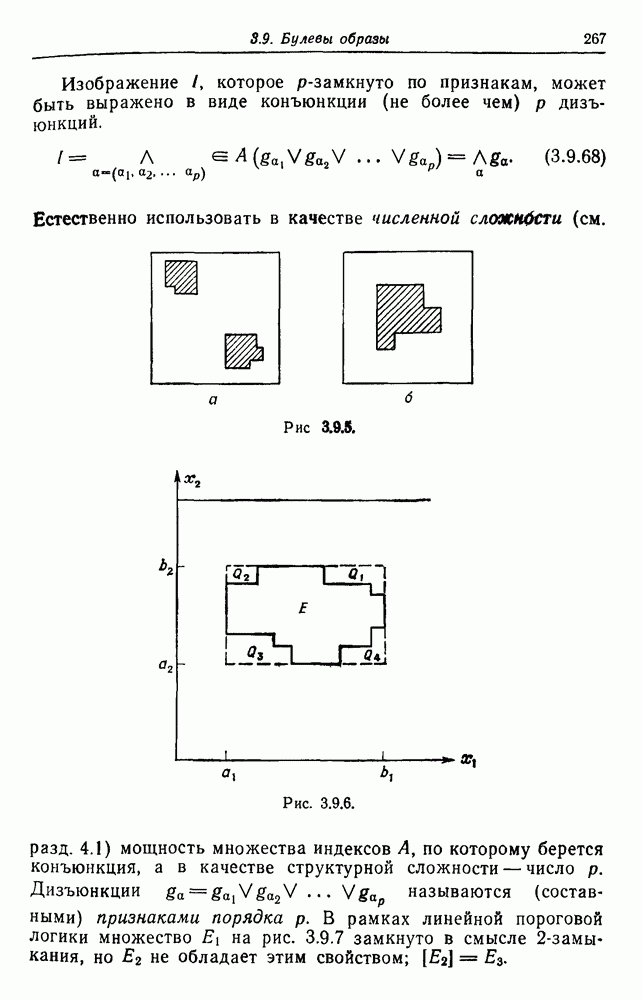
имеет генотип  а у — генотип
а у — генотип  то
то

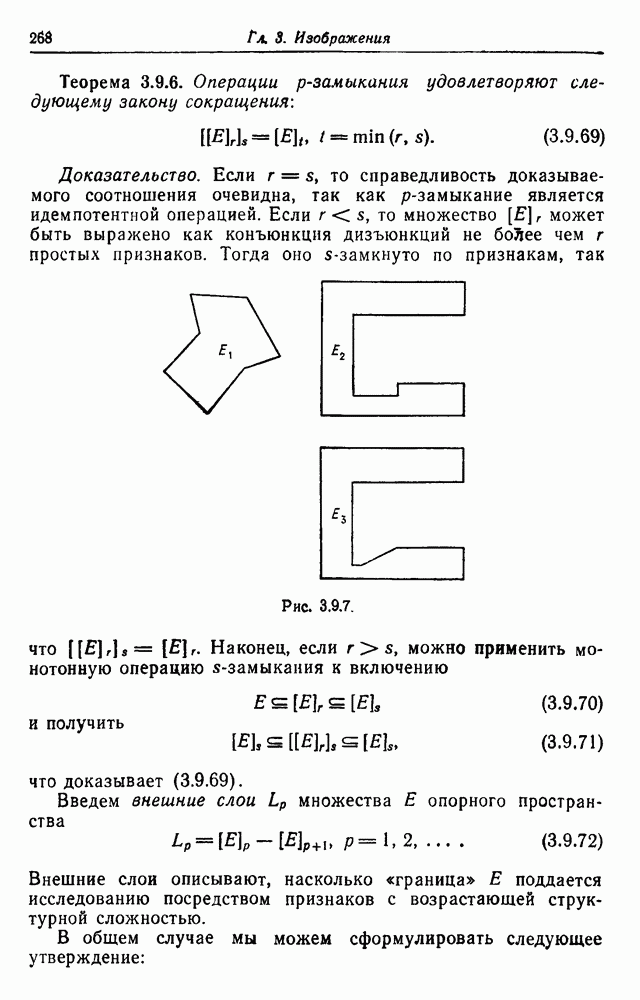
 существуют некоторые простые отношения. Во-первых, если
существуют некоторые простые отношения. Во-первых, если  имеет (в некотором локусе) генотип
имеет (в некотором локусе) генотип  то его коэффициент родства с самим собой задается так:
то его коэффициент родства с самим собой задается так:

 рожденного при скрещивании
рожденного при скрещивании  Если
Если  имеет генотип
имеет генотип  а у — генотип
а у — генотип  то
то


 с генотипами
с генотипами  Тогда потомок
Тогда потомок  будет иметь один из генотипов
будет иметь один из генотипов  вероятность
вероятность
 Поэтому
Поэтому


 от брака между дядей и племянницей, где особи 1, 2 и 5 рождены от родителей, не состоящих в родстве между собой, и не имеют родственной связи друг с другом.
от брака между дядей и племянницей, где особи 1, 2 и 5 рождены от родителей, не состоящих в родстве между собой, и не имеют родственной связи друг с другом.


 в то время как
в то время как  Следовательно,
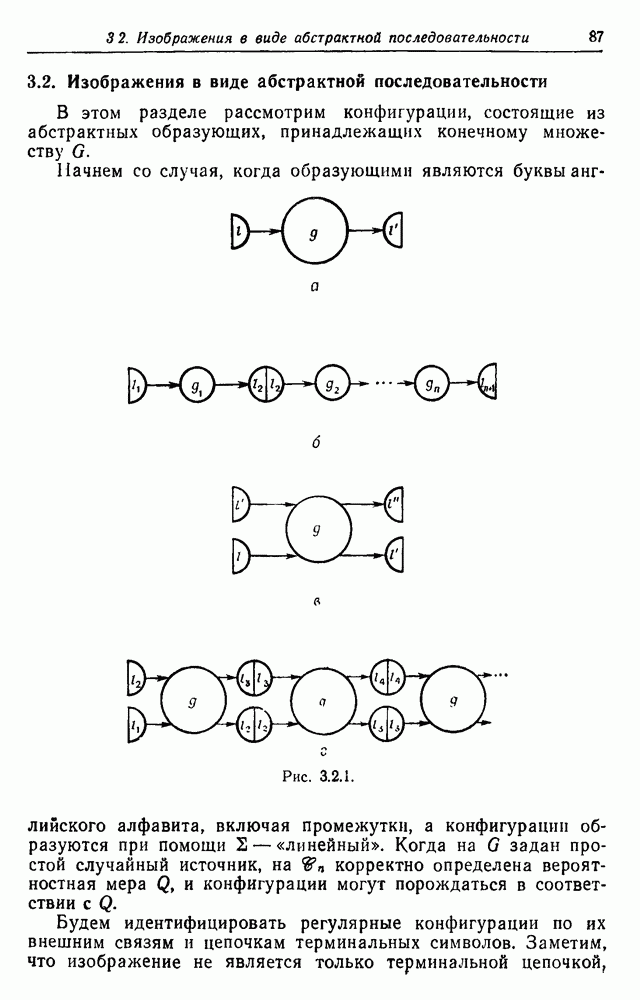
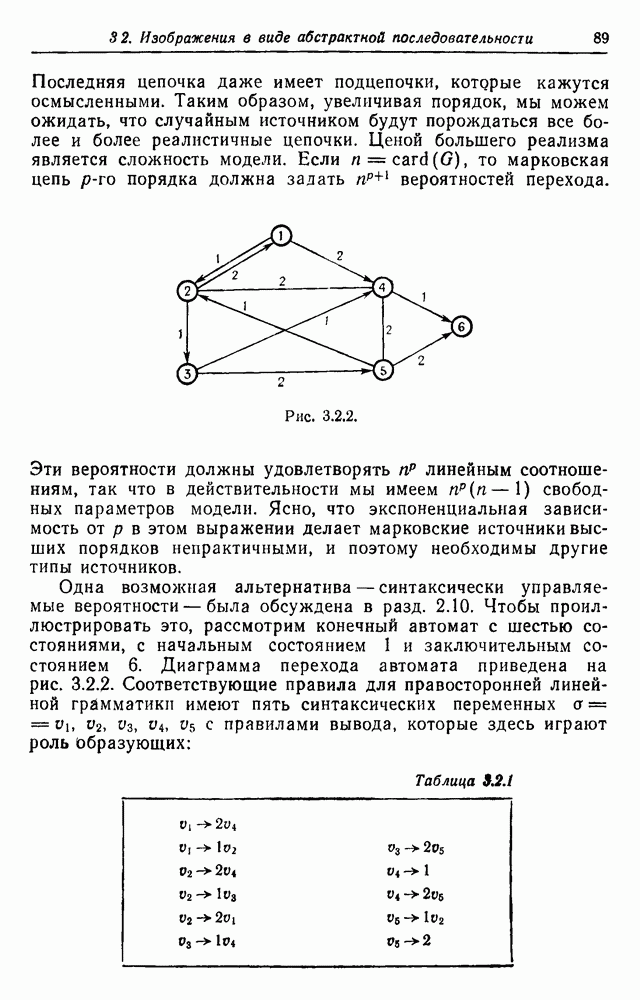
Следовательно,

 .
.

 поколении через
поколении через  имеем
имеем  Теперь
Теперь




 , так что
, так что

 получим линейное уравнение в конечных разностях третьего порядка с постоянными коэффициентами
получим линейное уравнение в конечных разностях третьего порядка с постоянными коэффициентами

 каждая из которых содержит представителей обоих полов, так что всего мы имеем 16 типов:
каждая из которых содержит представителей обоих полов, так что всего мы имеем 16 типов:

 . Группа потомка любой пары зависит лишь от того, к какой группе относится его мать, что и указано в табл. 3.3.2.
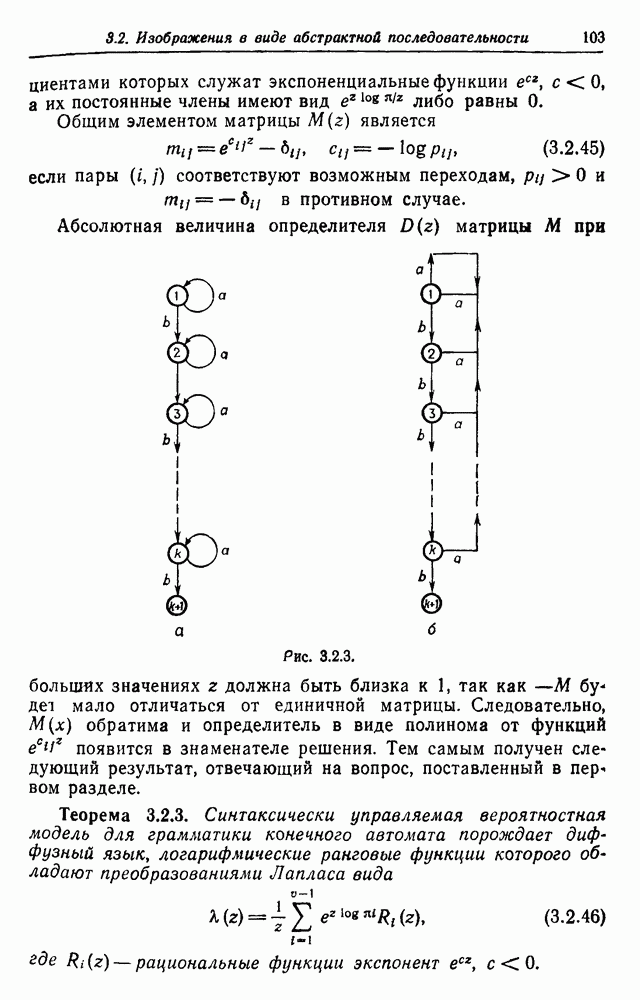
. Группа потомка любой пары зависит лишь от того, к какой группе относится его мать, что и указано в табл. 3.3.2.
 класса существуют 2 выбора в зависимости от того, применяются правила
класса существуют 2 выбора в зависимости от того, применяются правила  или правила
или правила 

 брака ее родителей. Если охарактеризовать особь при помощи пары
брака ее родителей. Если охарактеризовать особь при помощи пары  то получим множество перестановок
то получим множество перестановок

 типу брака и
типу брака и  матери. Тогда брат должен будет жениться в
матери. Тогда брат должен будет жениться в  группе, а сестра
группе, а сестра  Если в результате брака сестры появляется сын, а брата — дочь, то из условия допустимости их брака следует
Если в результате брака сестры появляется сын, а брата — дочь, то из условия допустимости их брака следует  оператора перестановок должны переставляться друг с другом. Таким образом, можно приступить к изучению алгебраической структуры
оператора перестановок должны переставляться друг с другом. Таким образом, можно приступить к изучению алгебраической структуры

 мы получим конфигурации, приведенные на рис. 3.3.6 с крайне нерегулярным механизмом спаривания. Мы исключили идентификаторы, используя лишь
мы получим конфигурации, приведенные на рис. 3.3.6 с крайне нерегулярным механизмом спаривания. Мы исключили идентификаторы, используя лишь  или
или  также исключили связи на стрелках. Последующие поколения
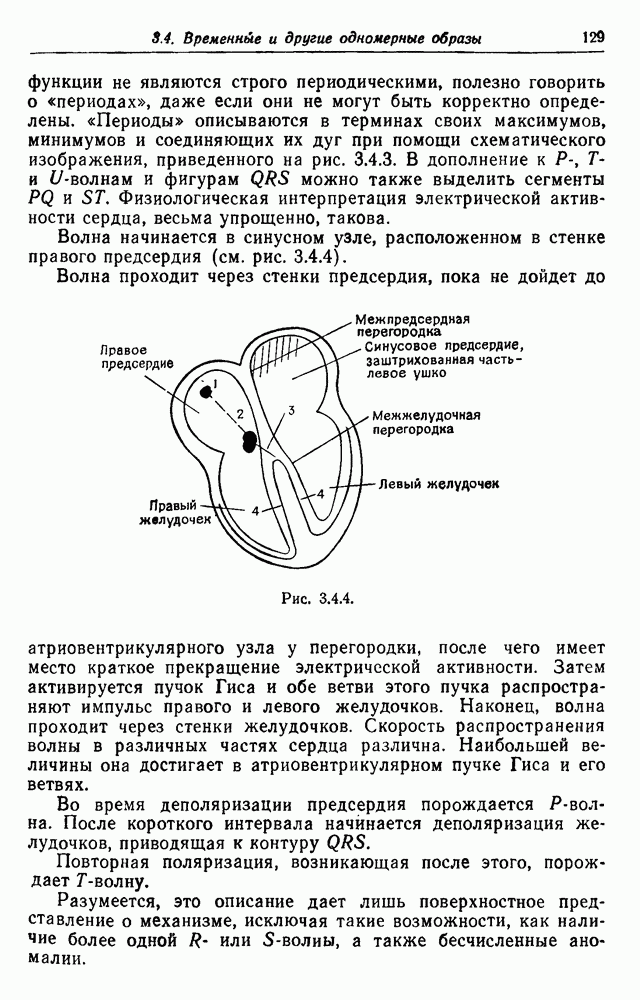
также исключили связи на стрелках. Последующие поколения  теперь играют роль макрообразующих. Интересно отметить,
теперь играют роль макрообразующих. Интересно отметить,
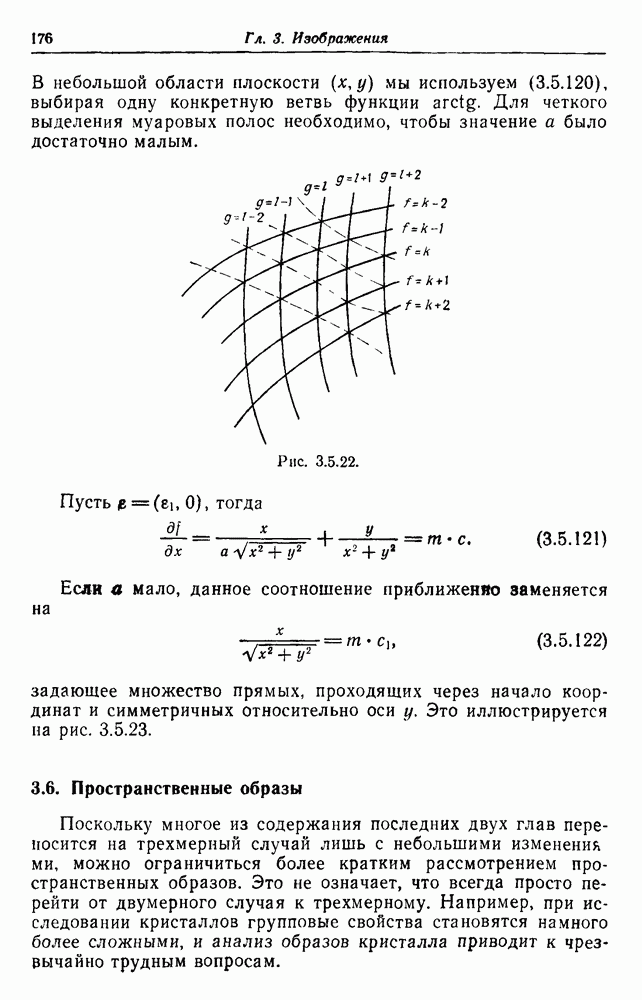
 образует марковский процесс, если спаривание случайно и число детей у каждой пары имеет распределение, не зависящее от происхождения пары.
образует марковский процесс, если спаривание случайно и число детей у каждой пары имеет распределение, не зависящее от происхождения пары.