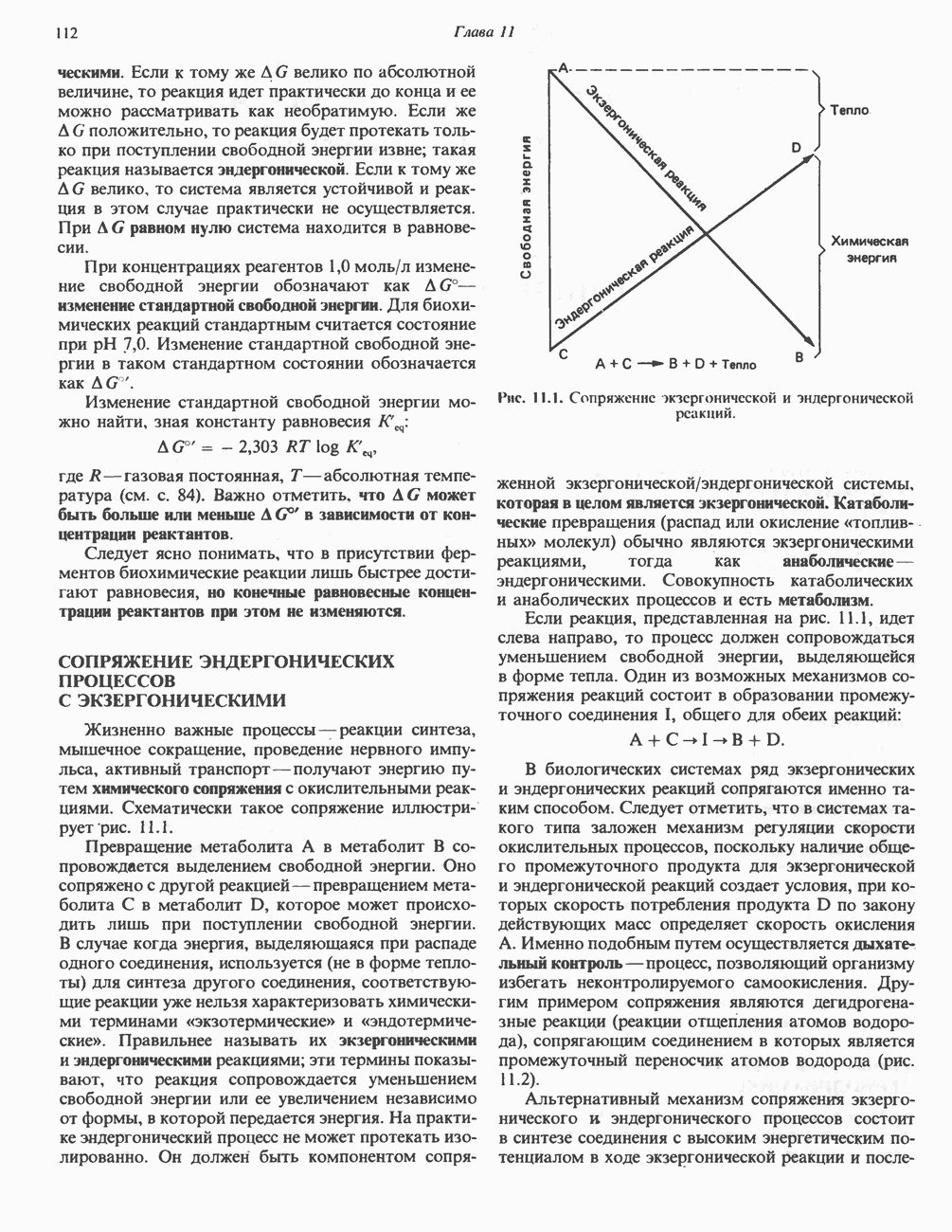
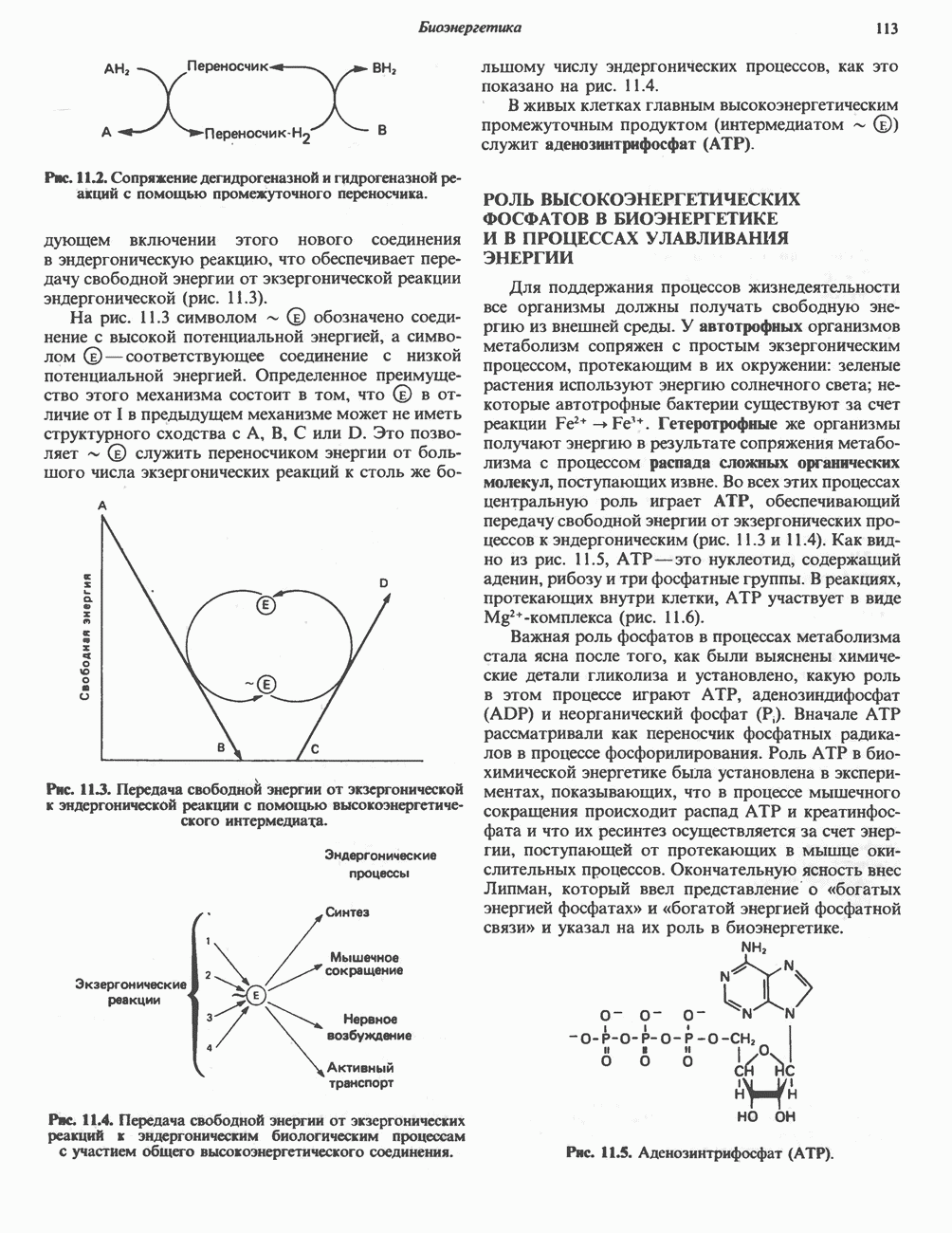
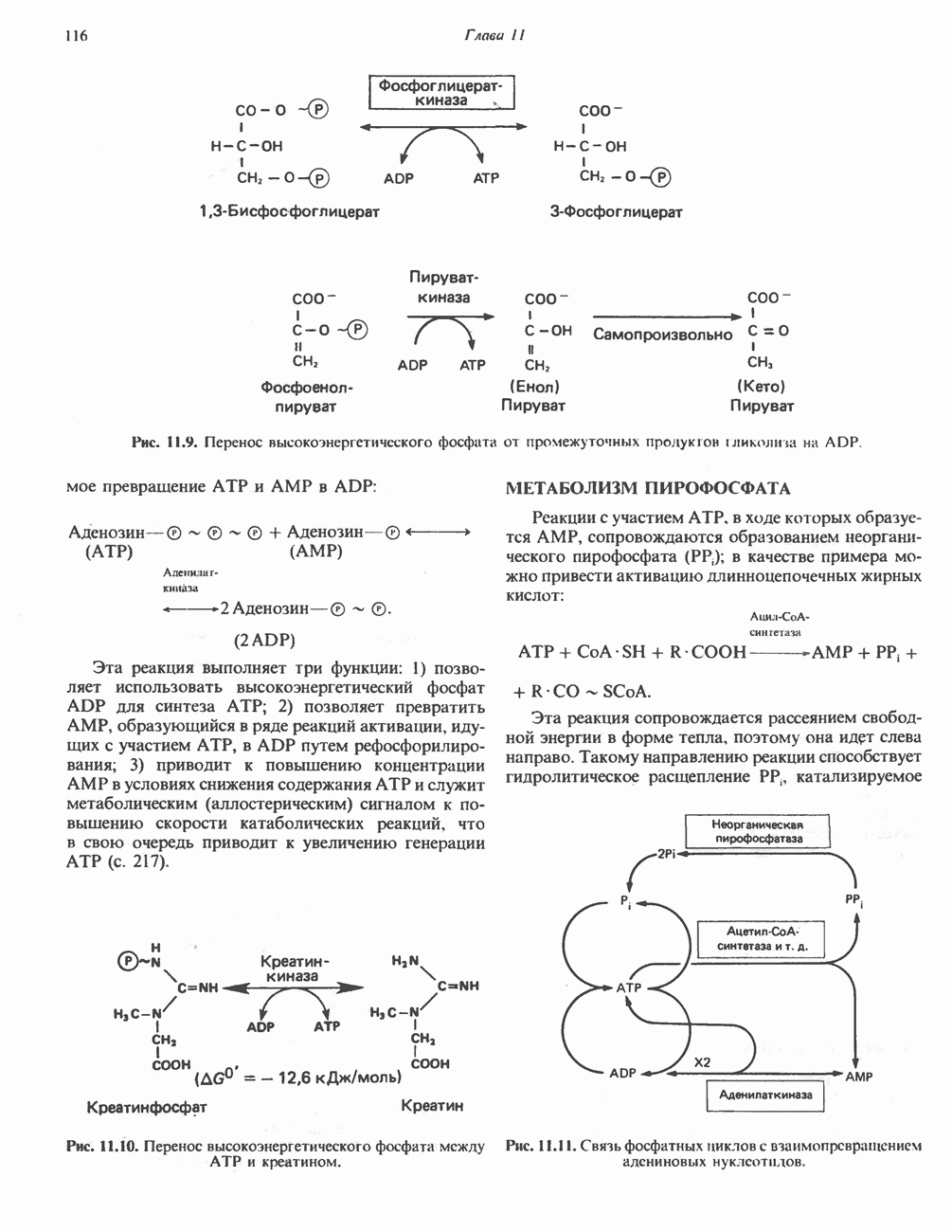
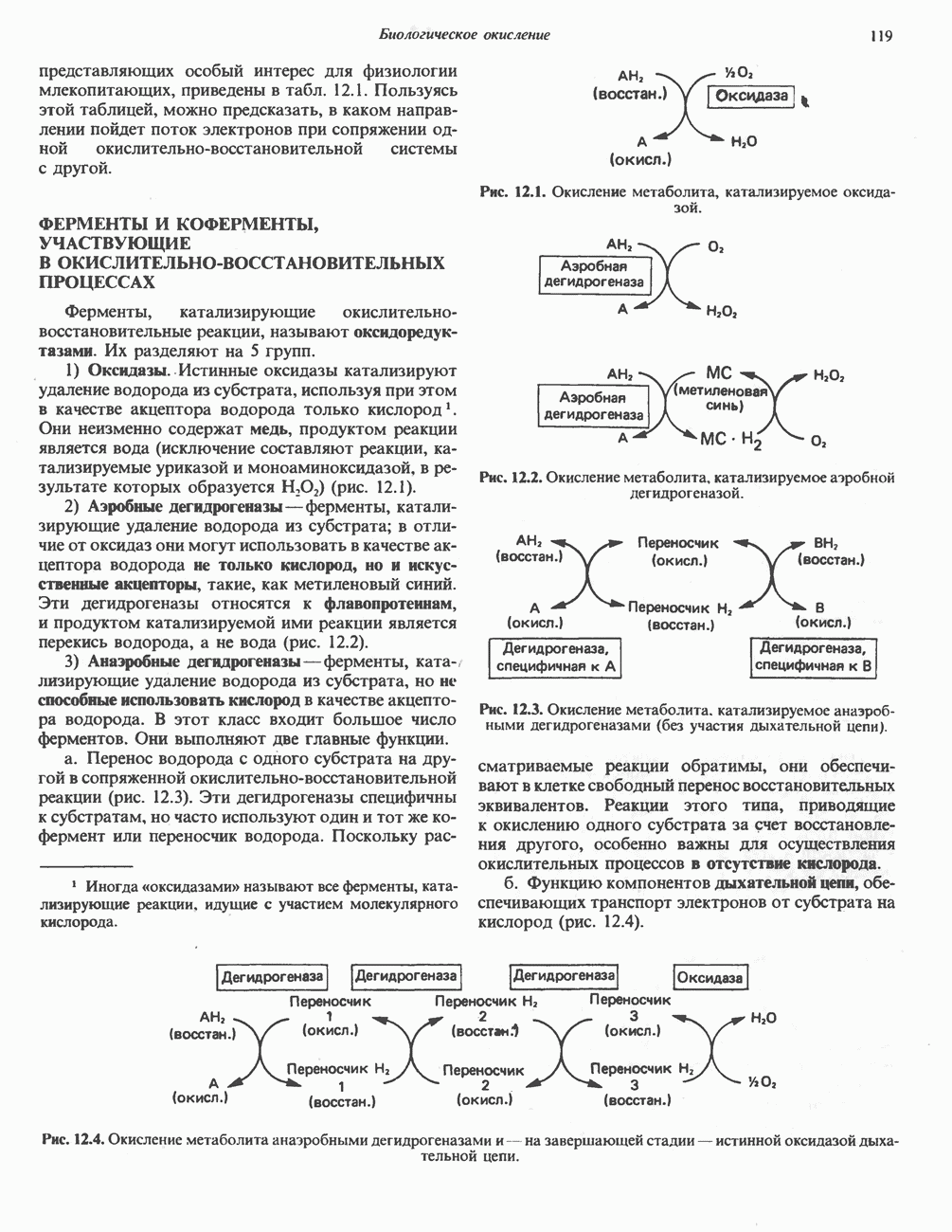
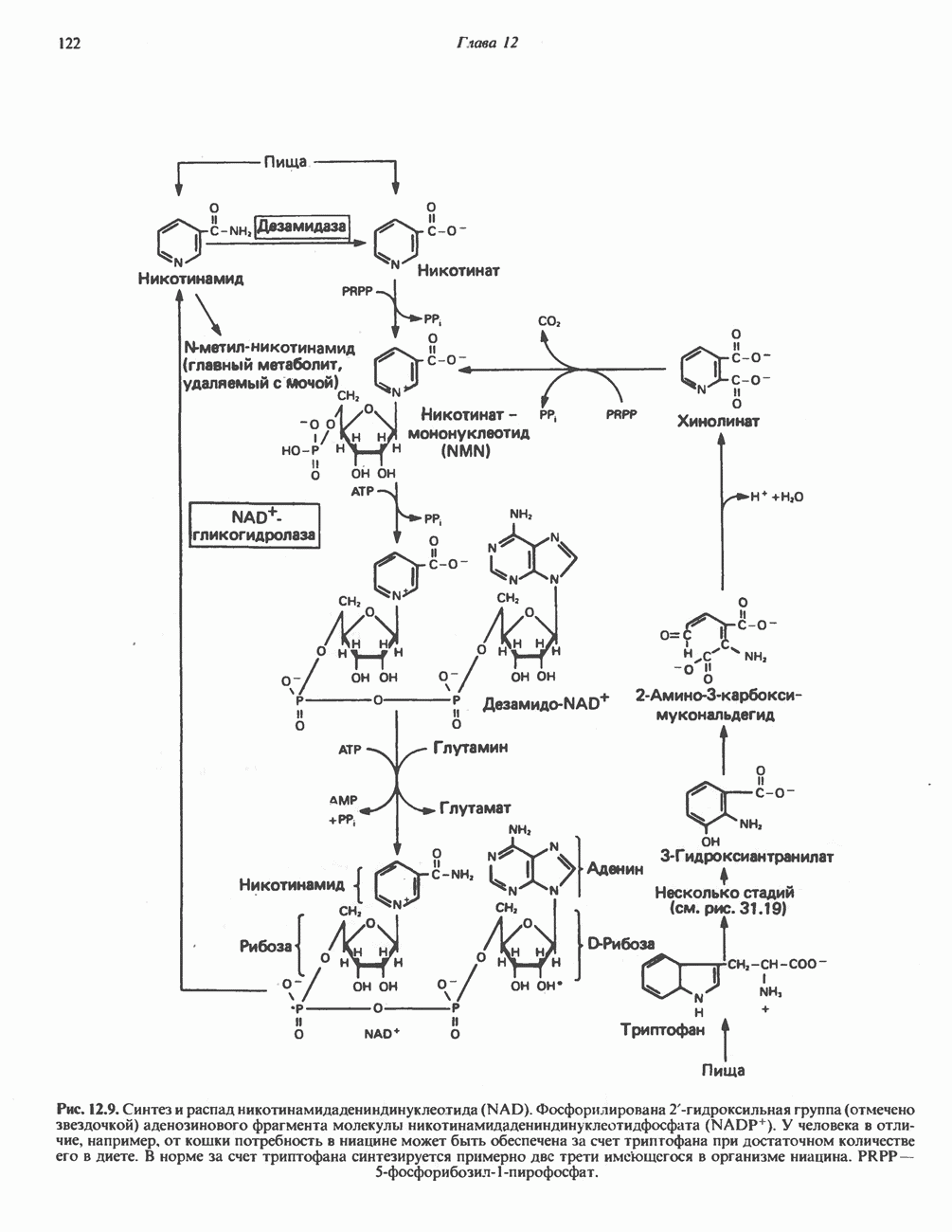
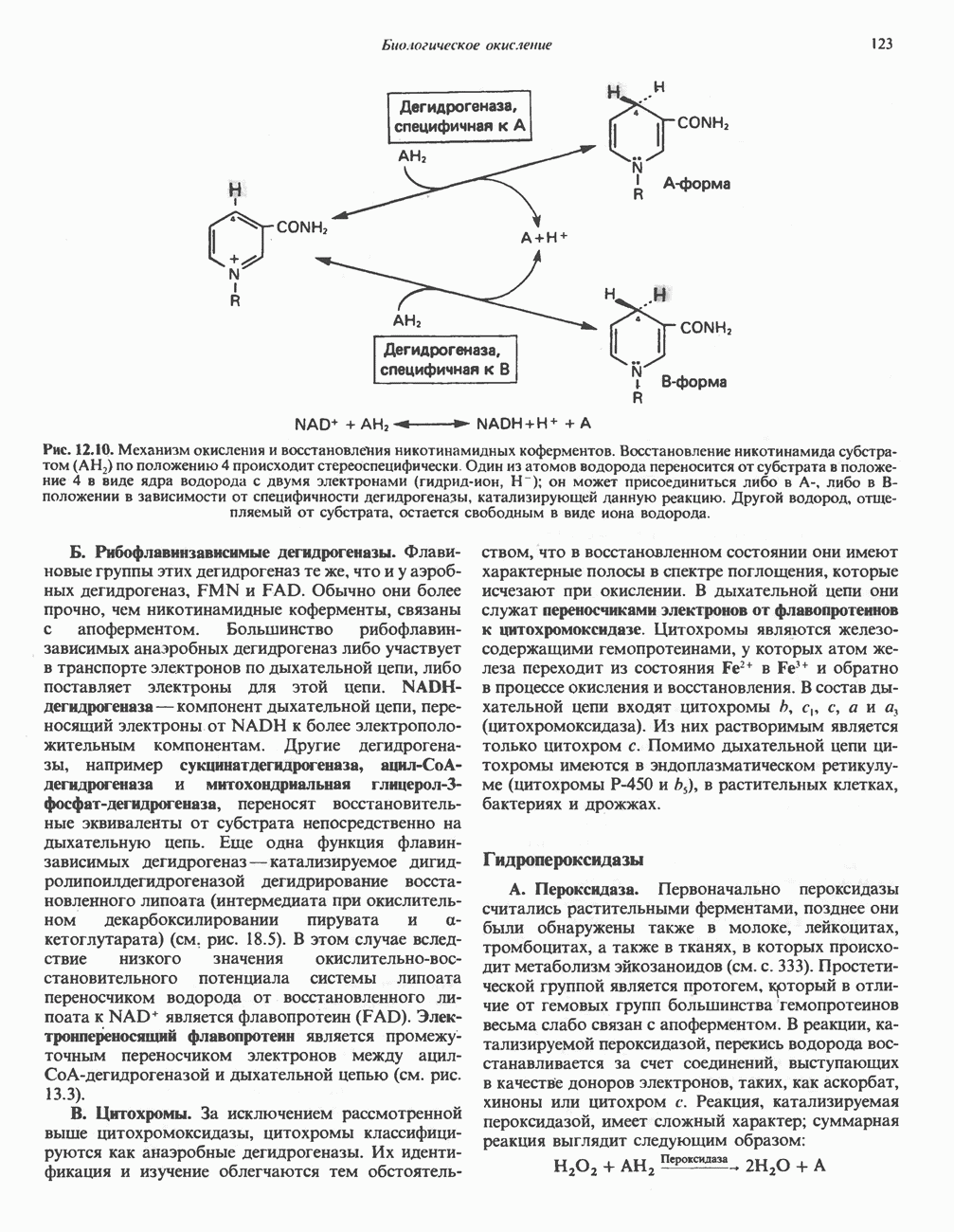
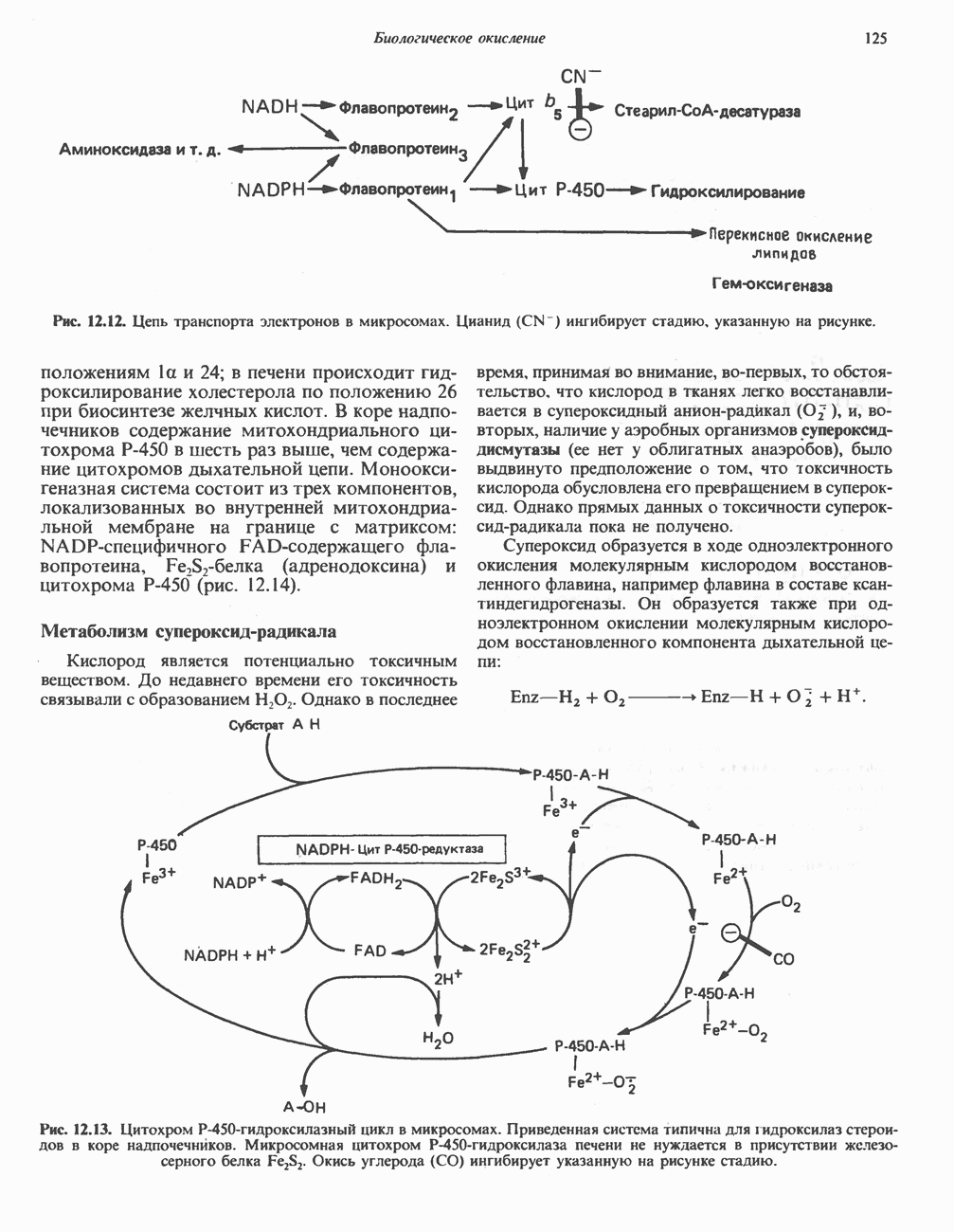
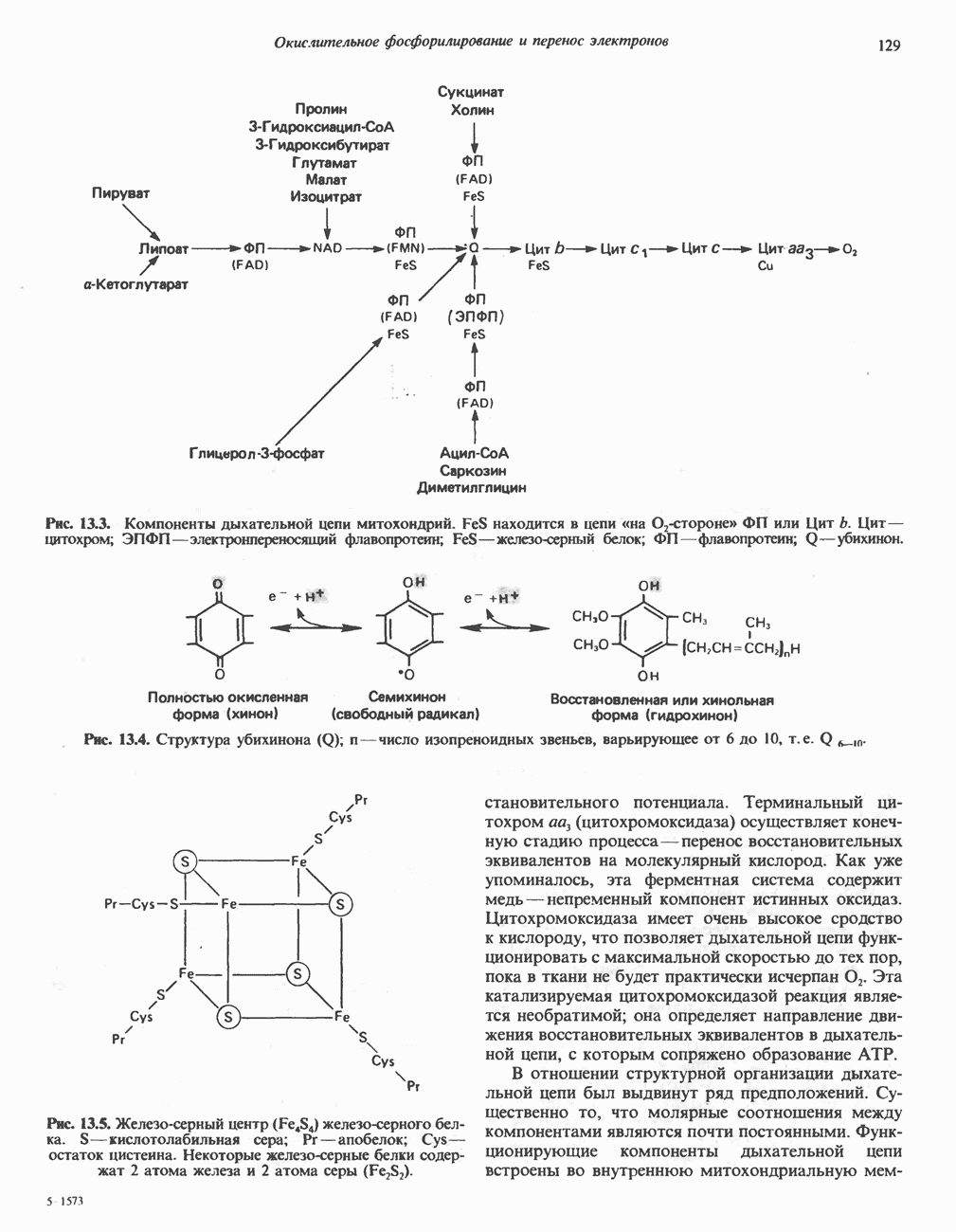
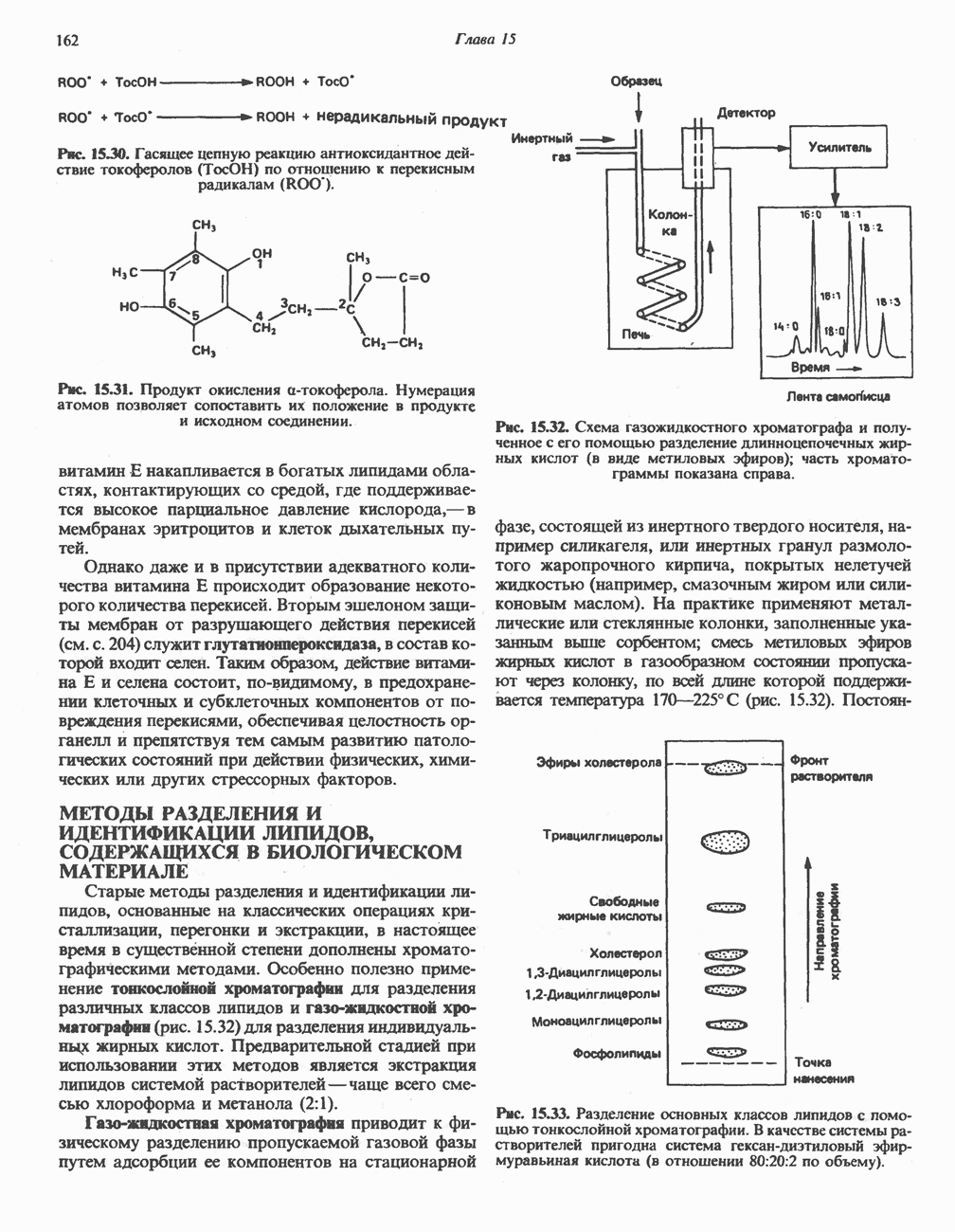
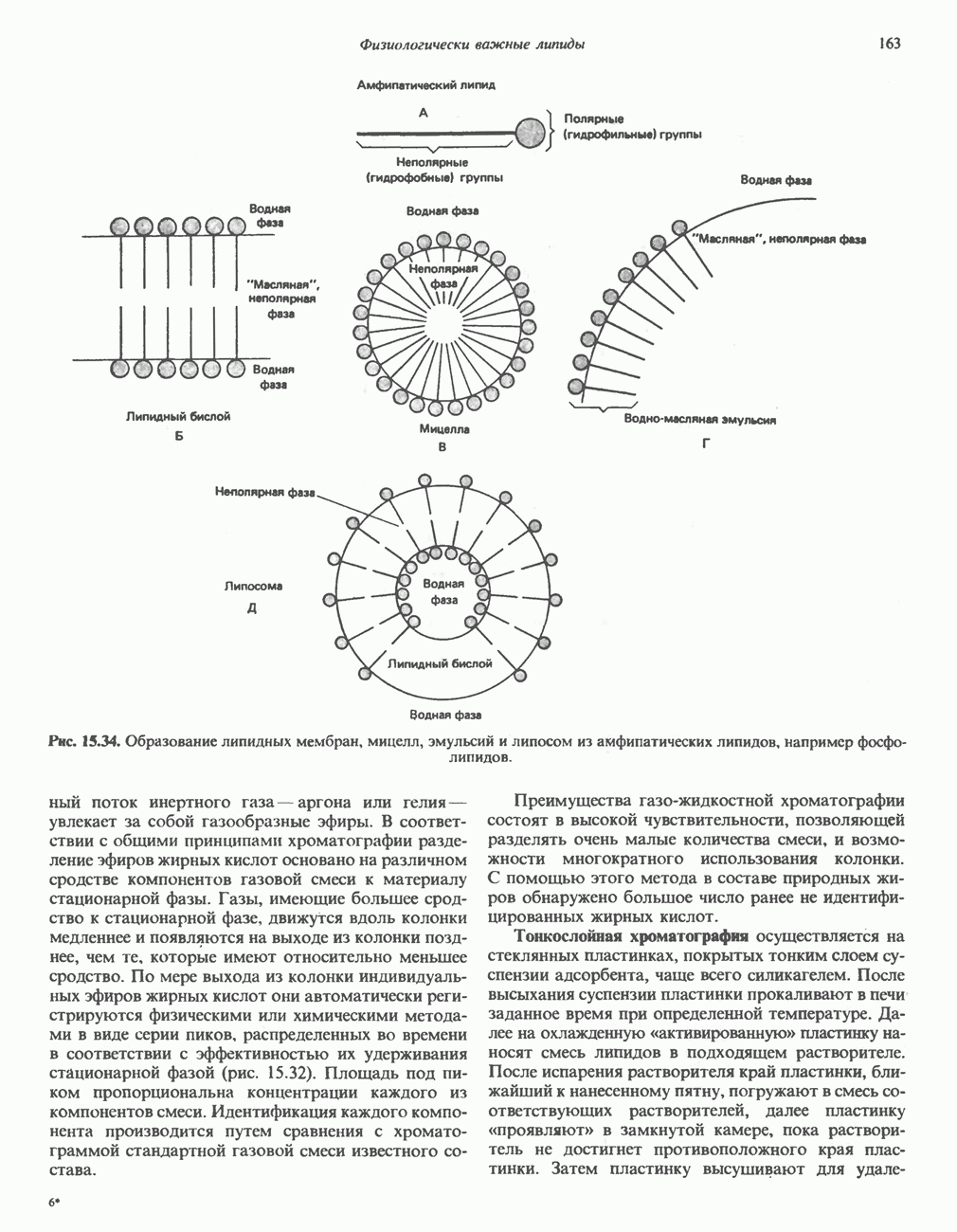
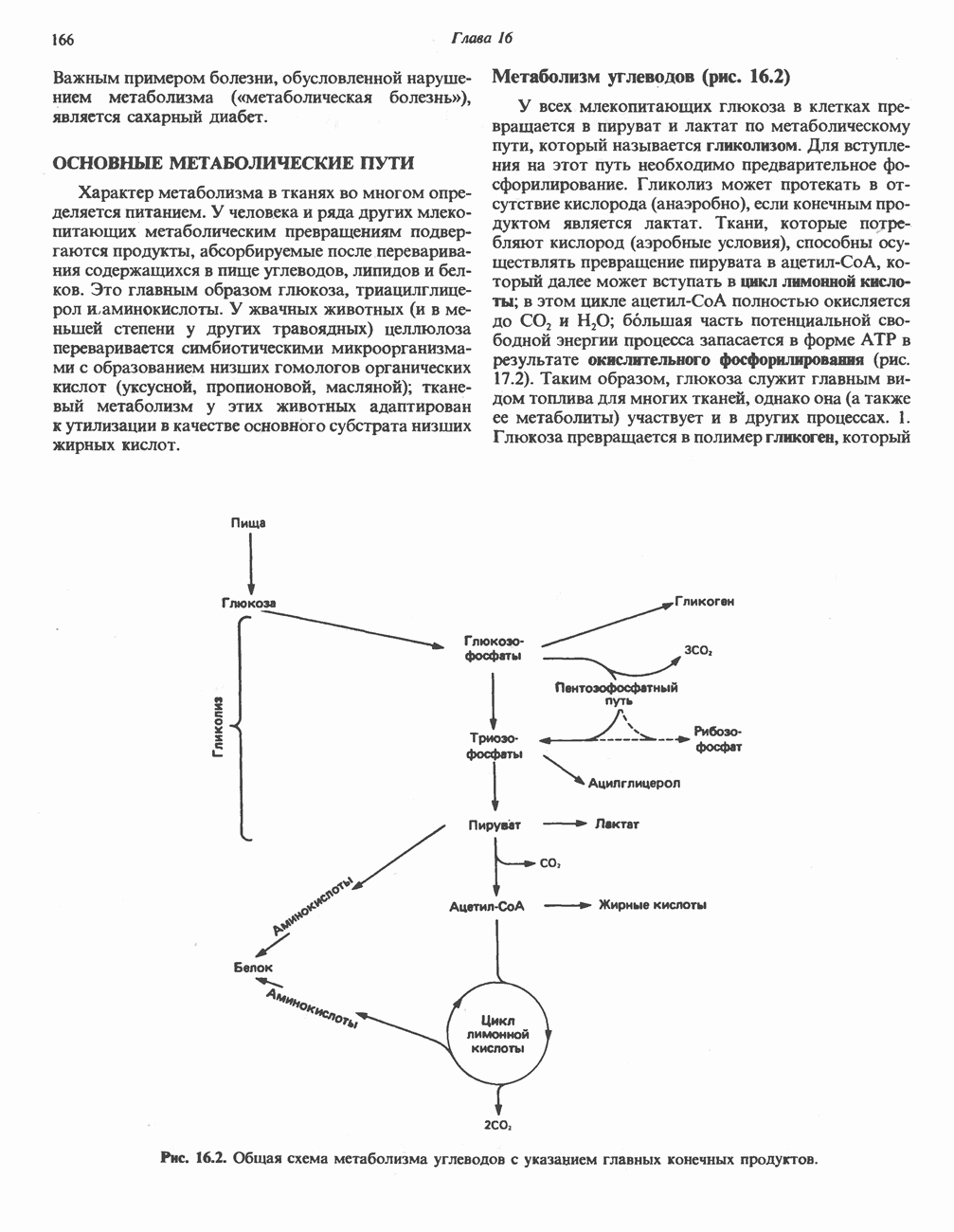
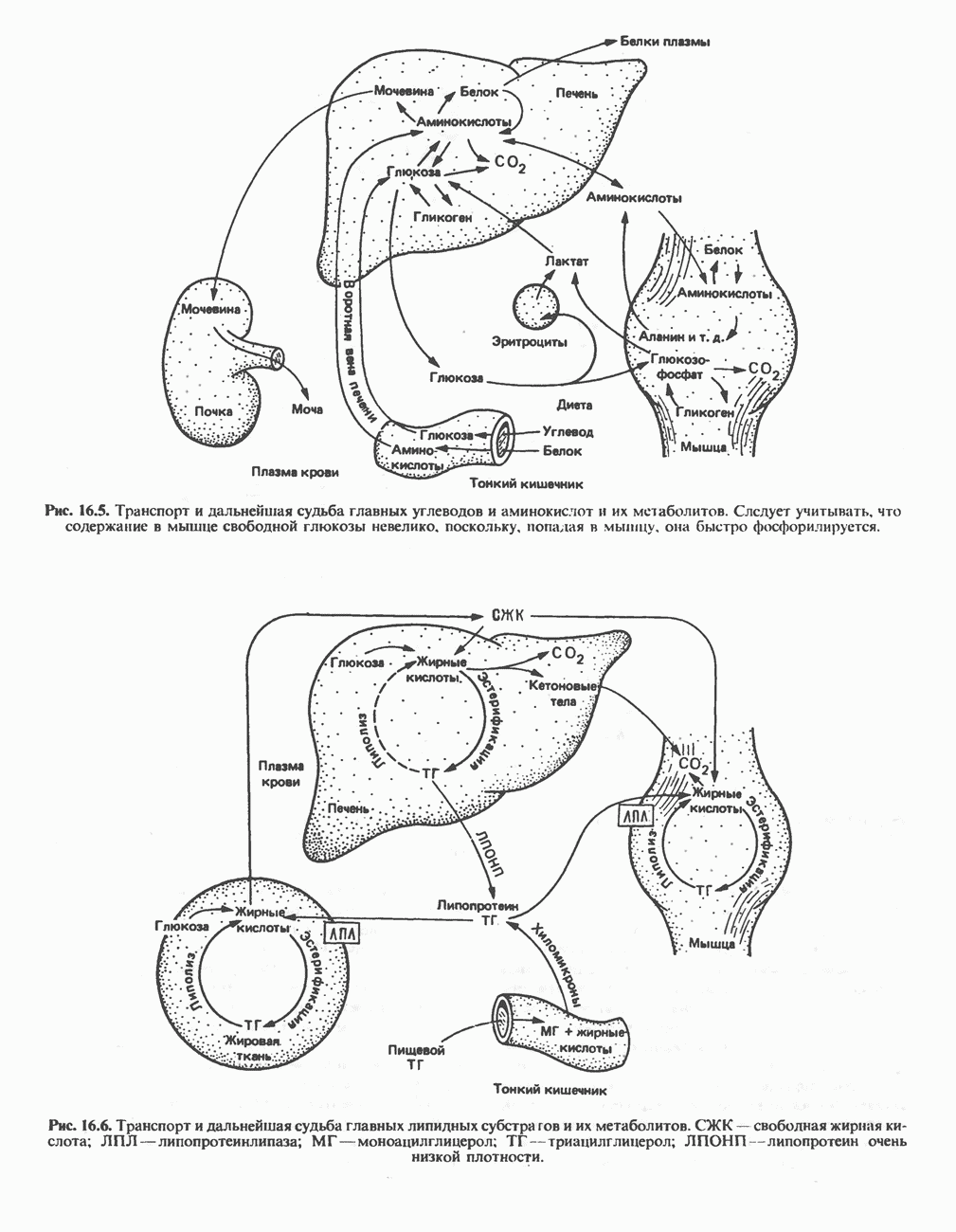
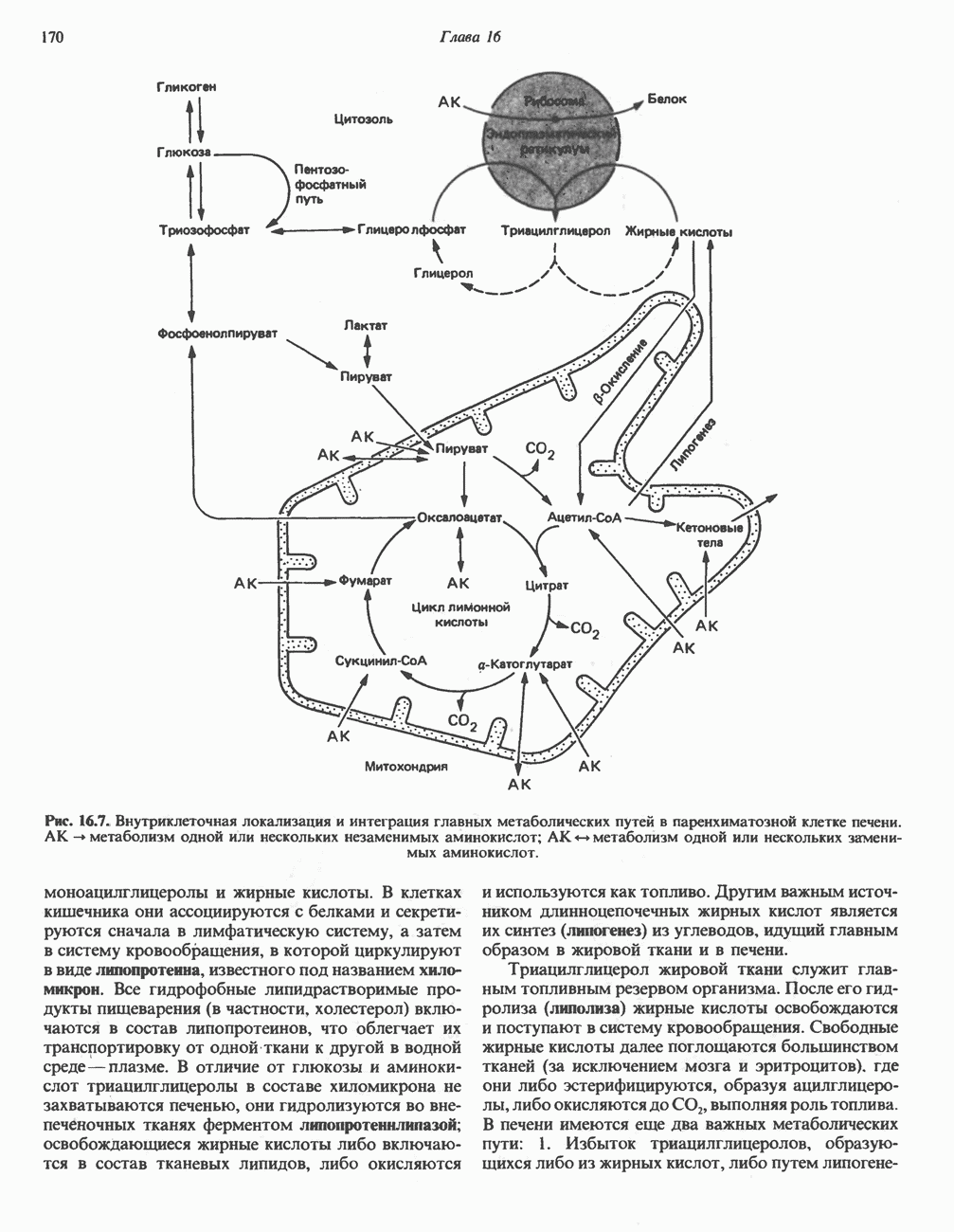
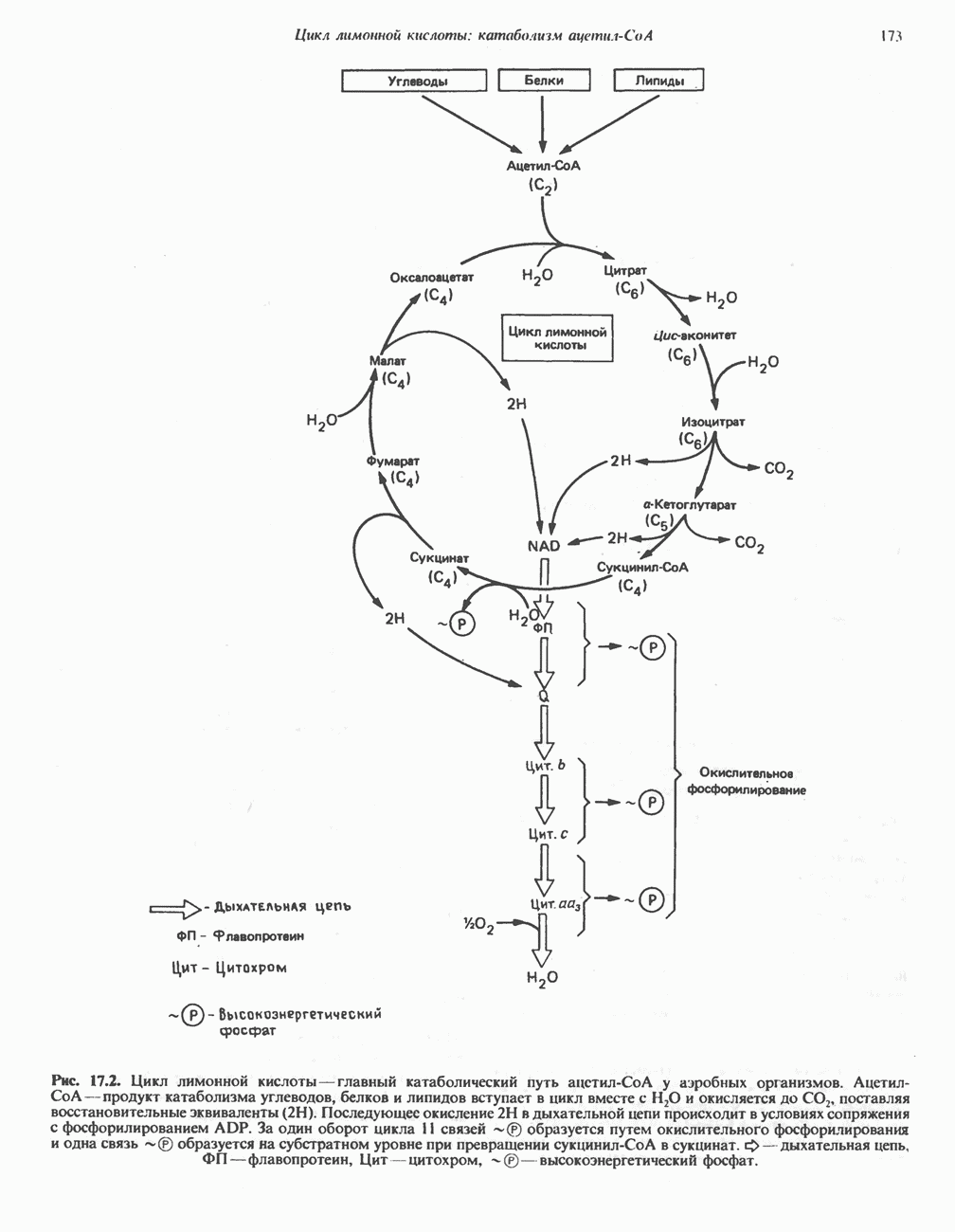
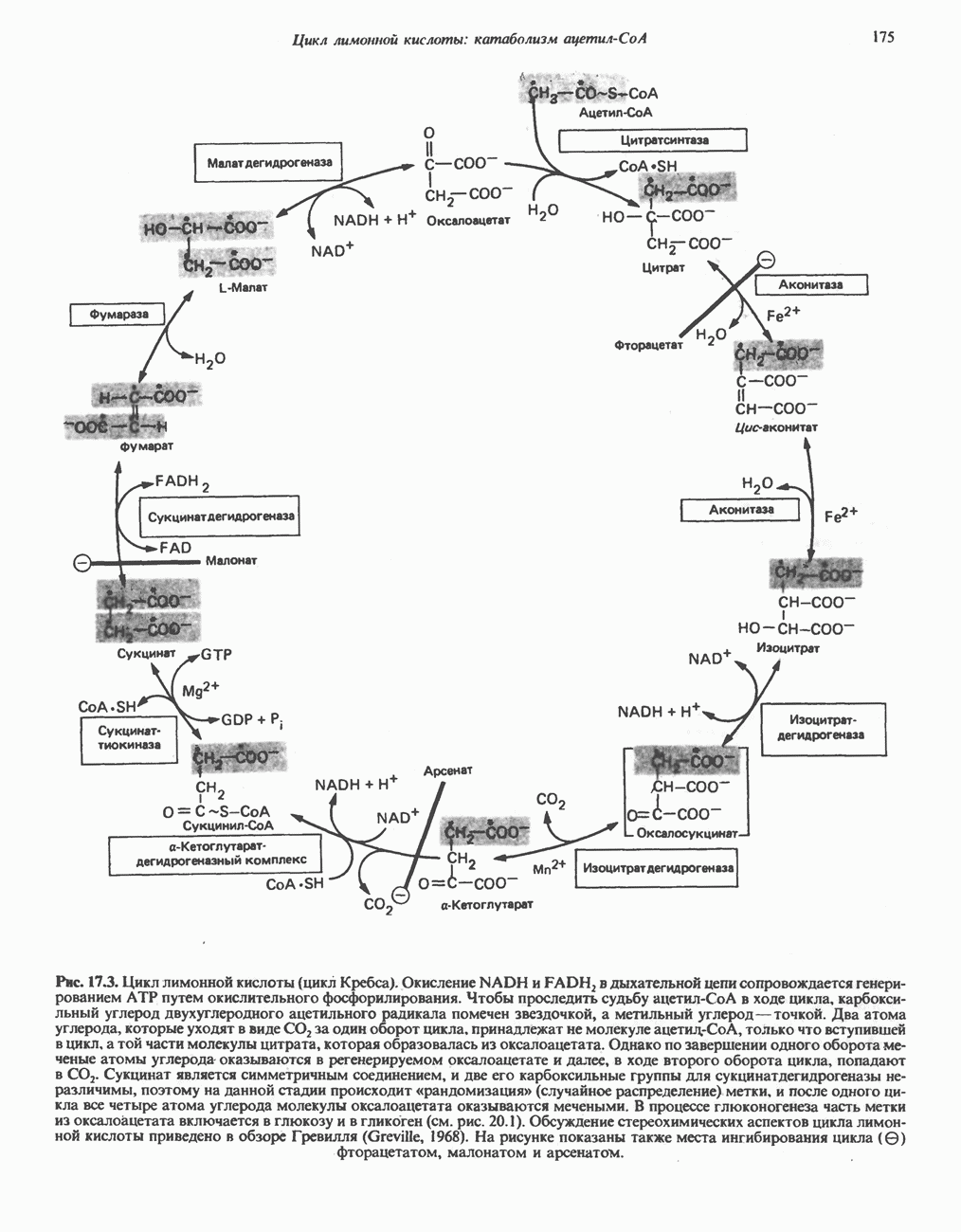
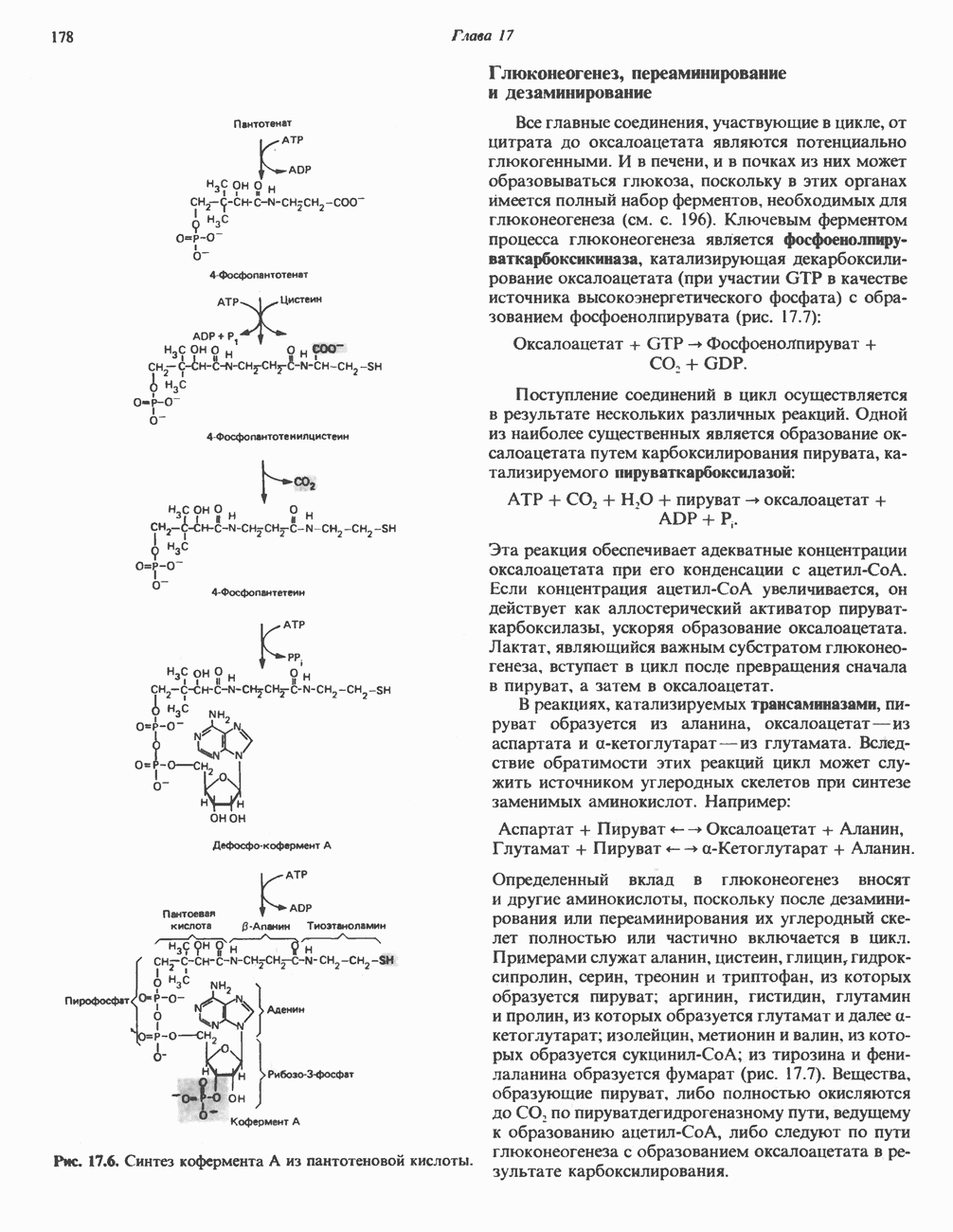
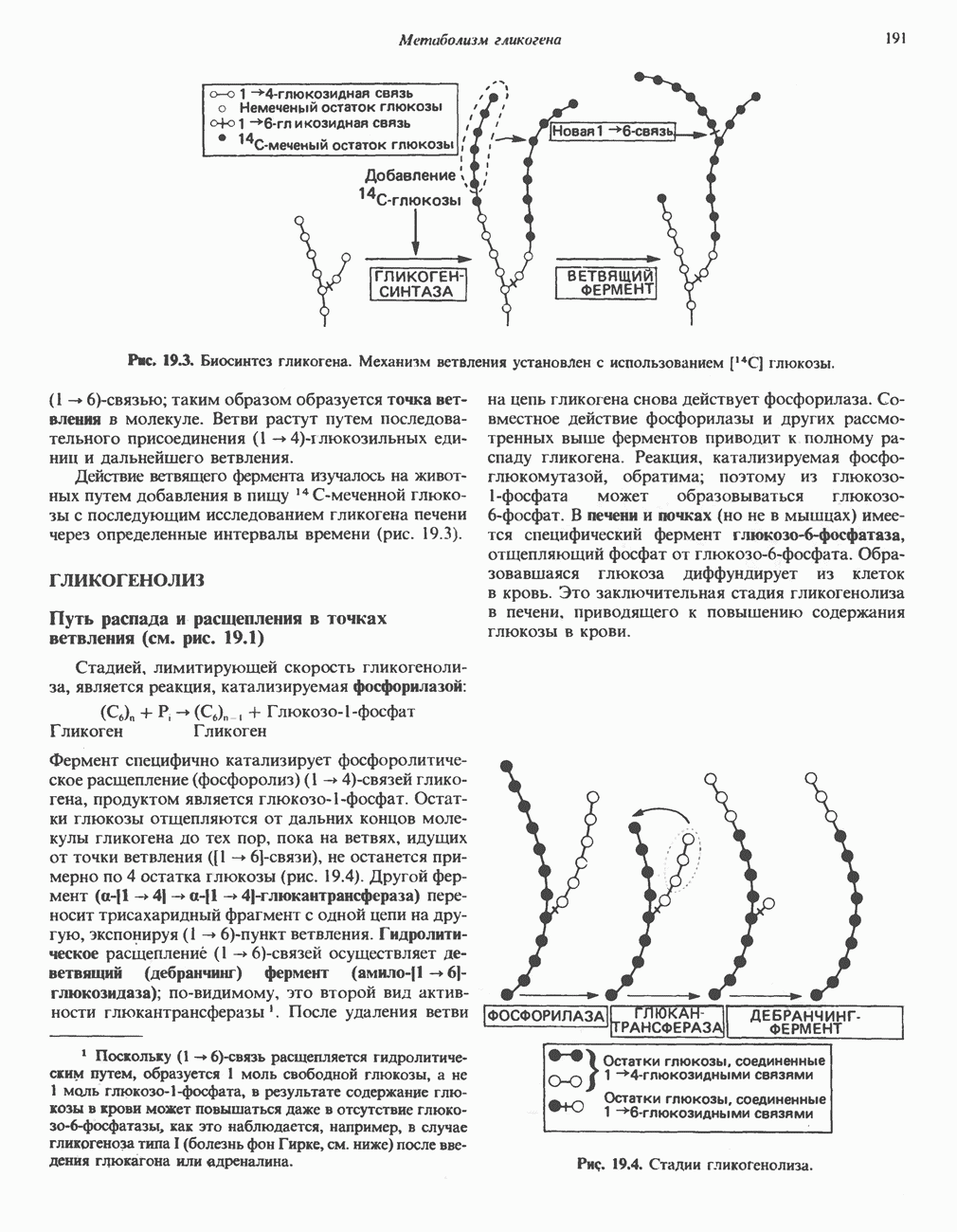
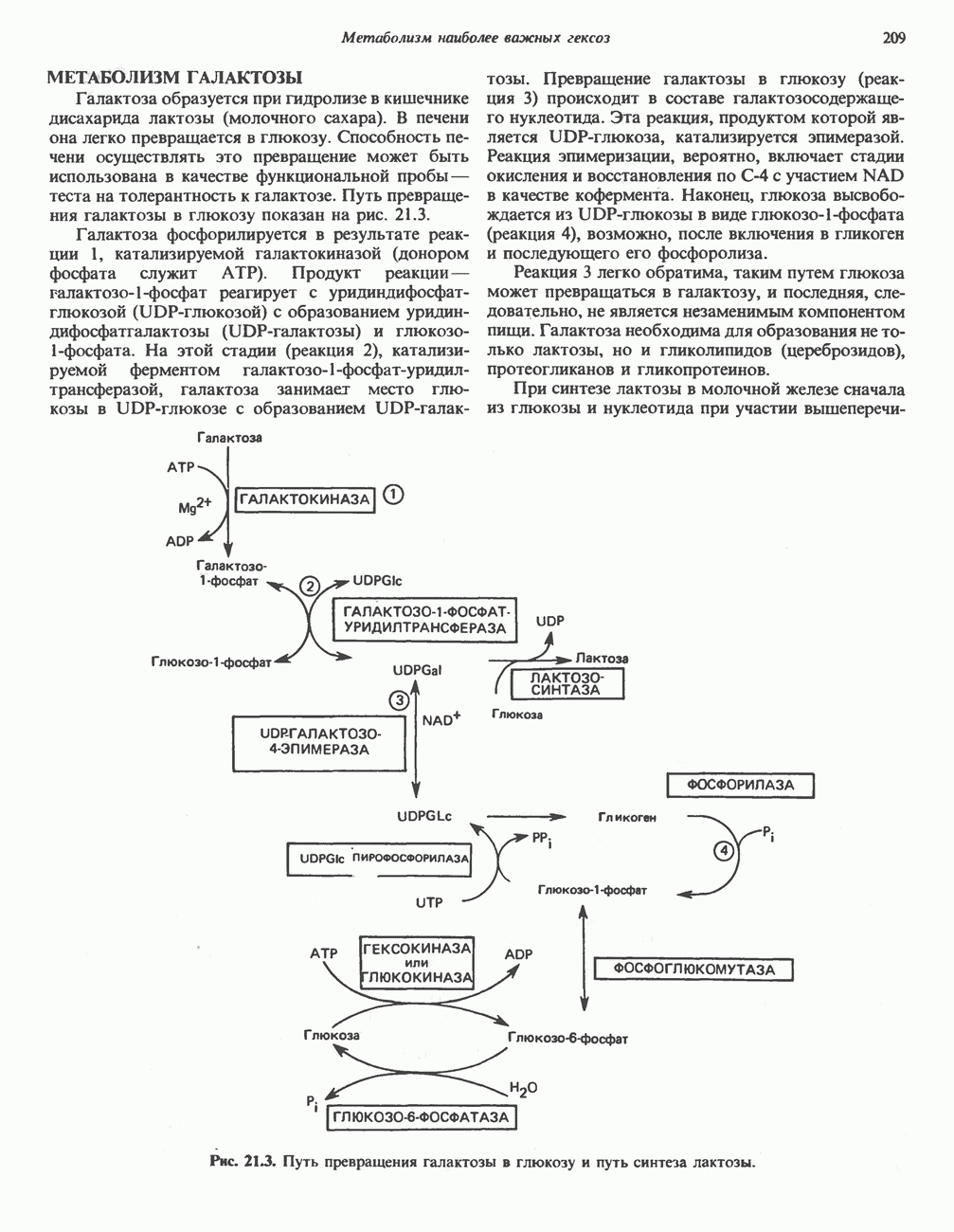
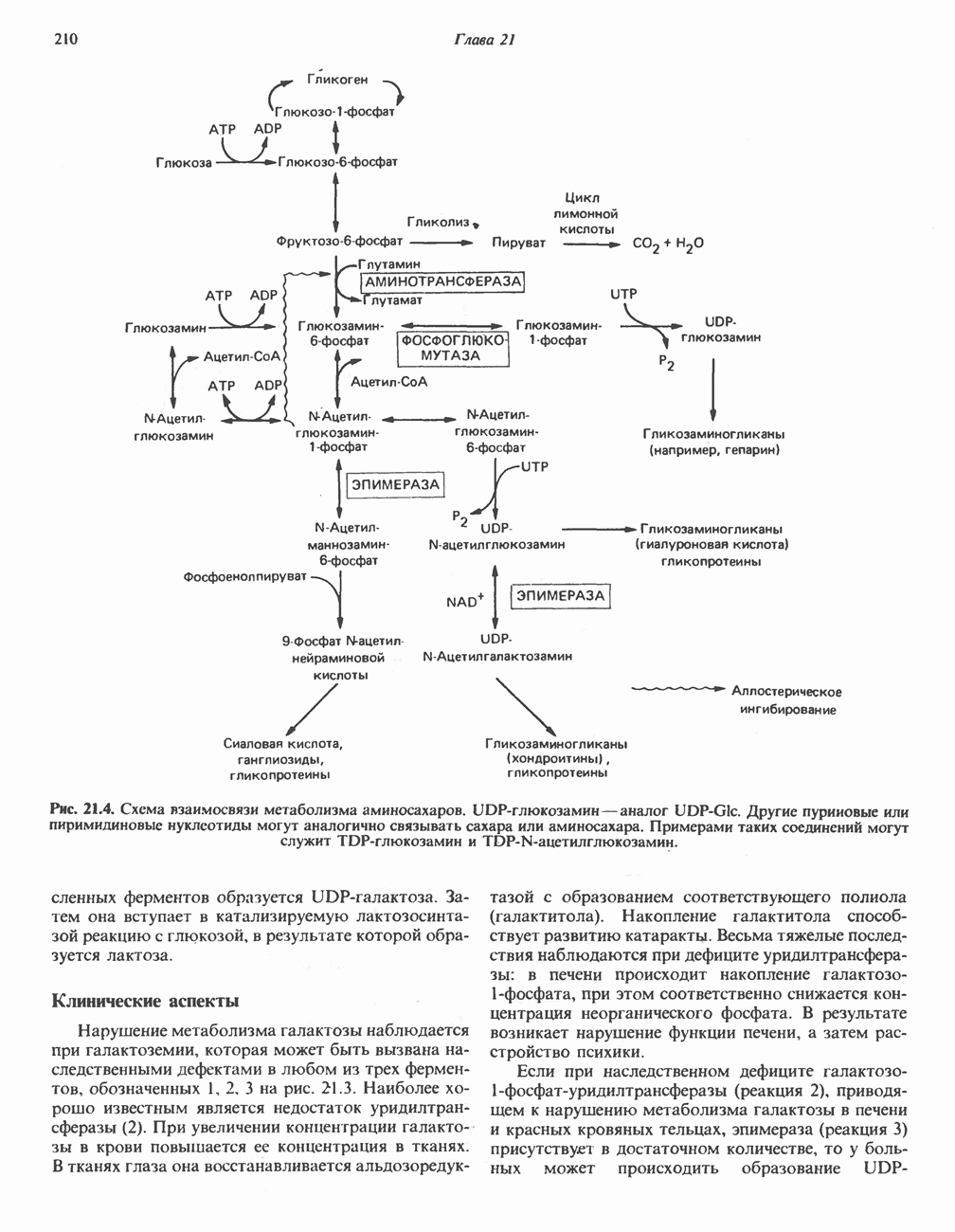
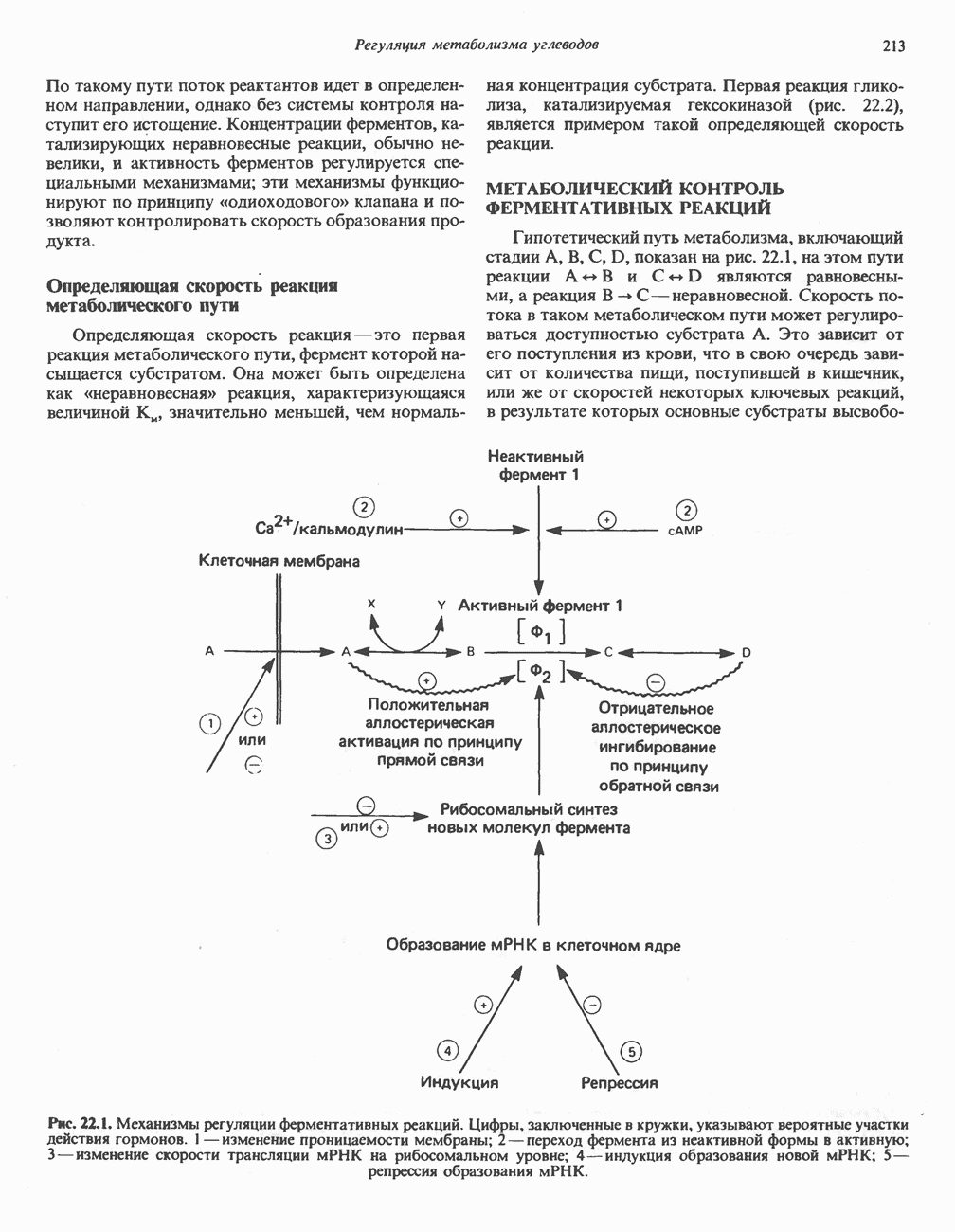
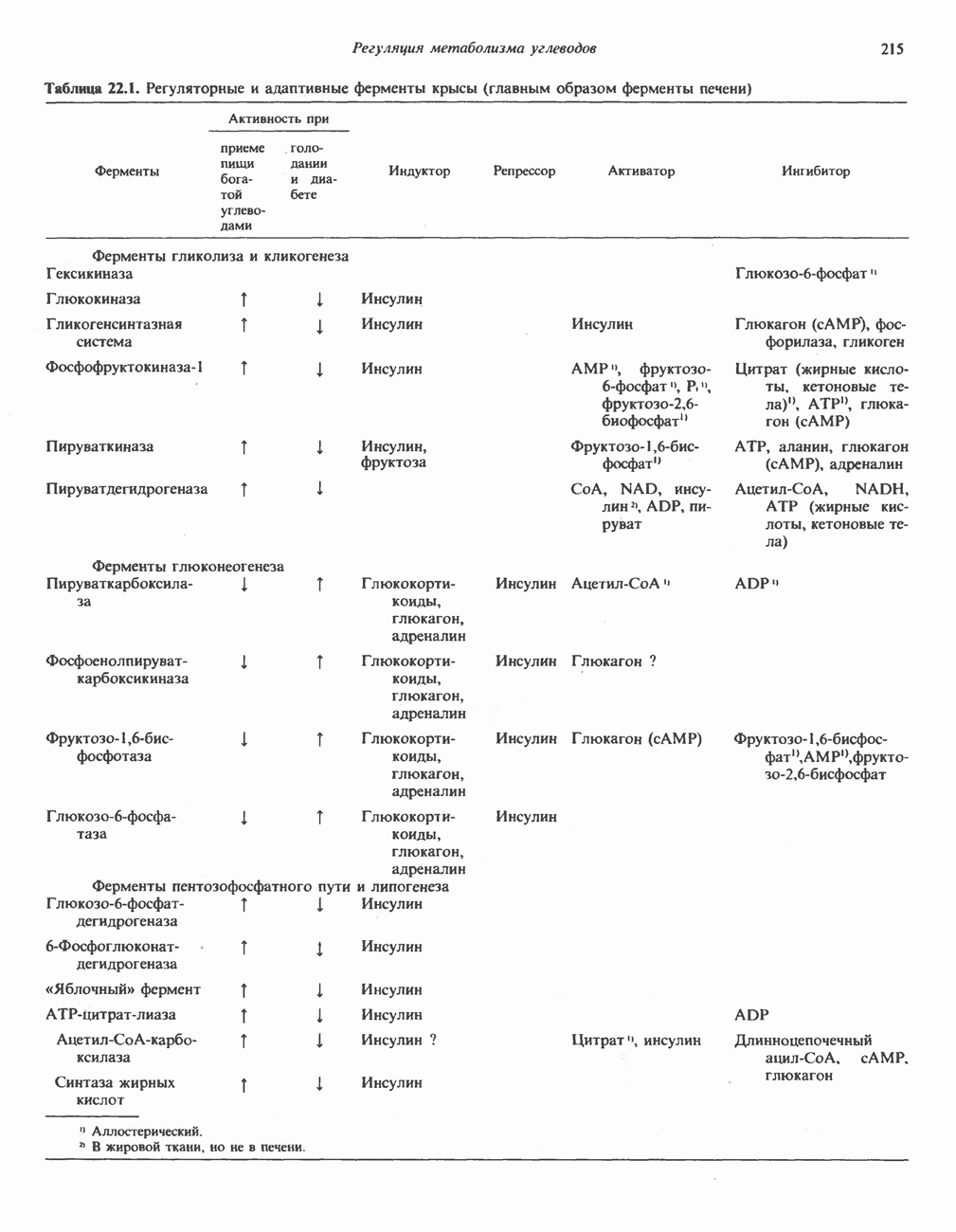
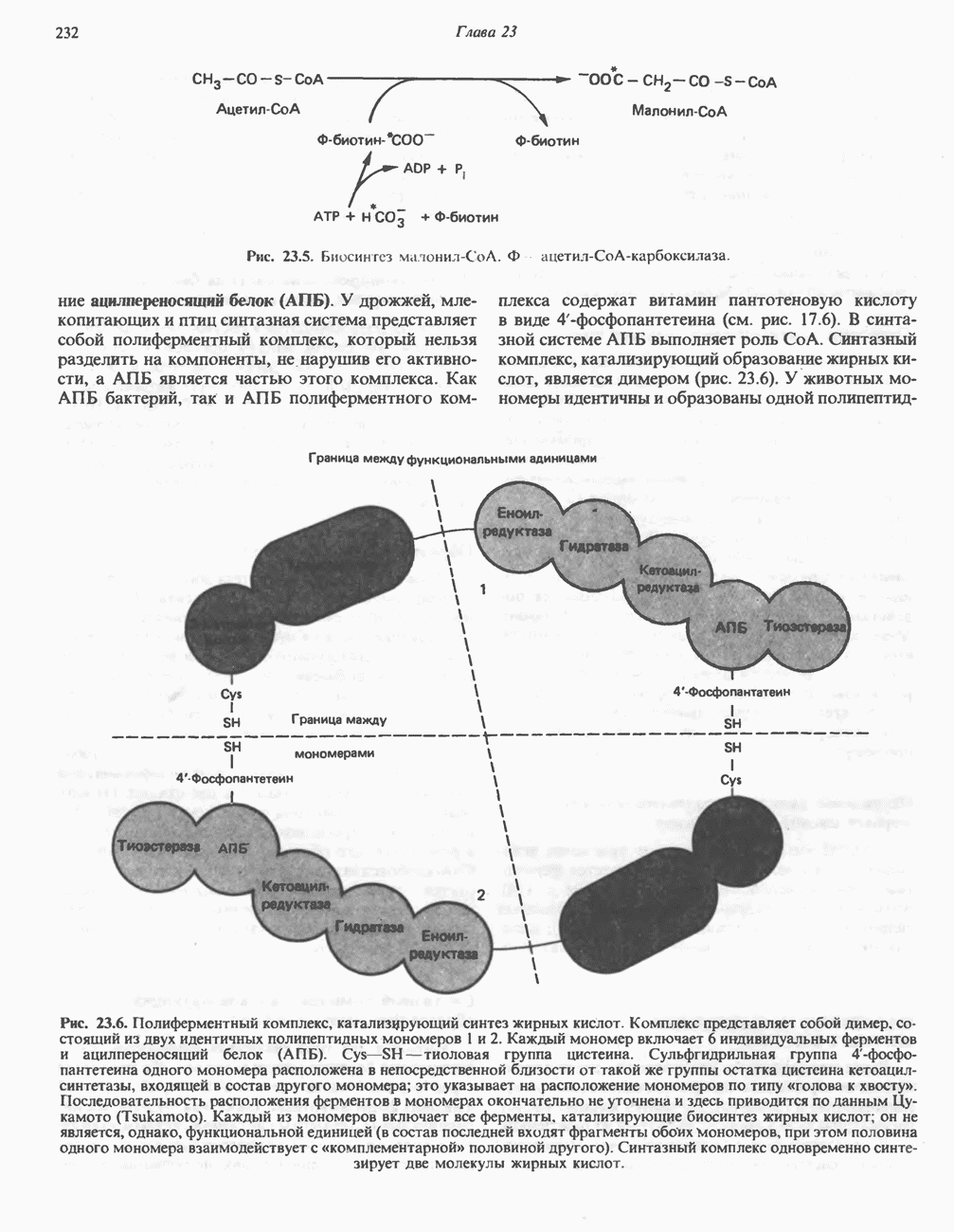
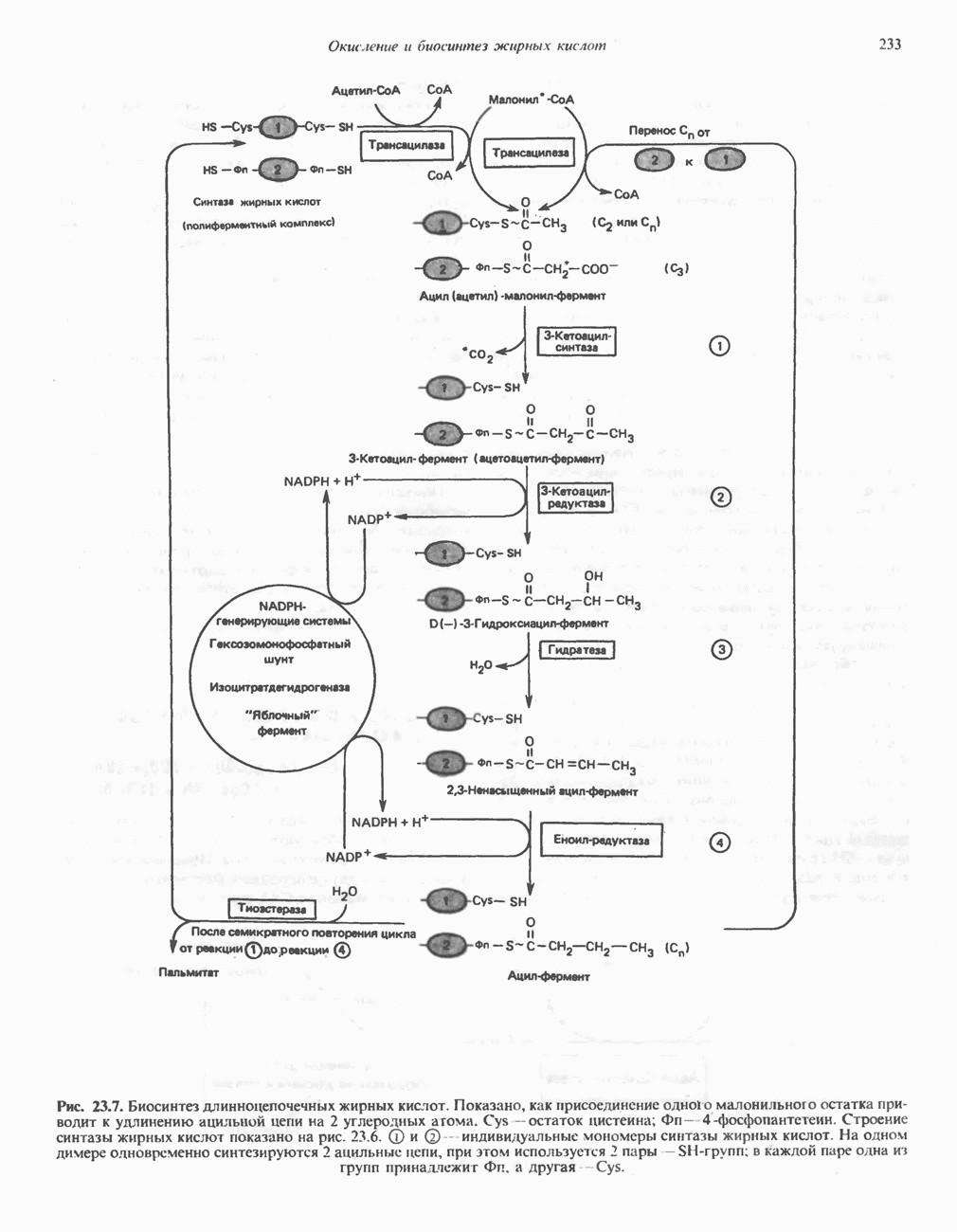
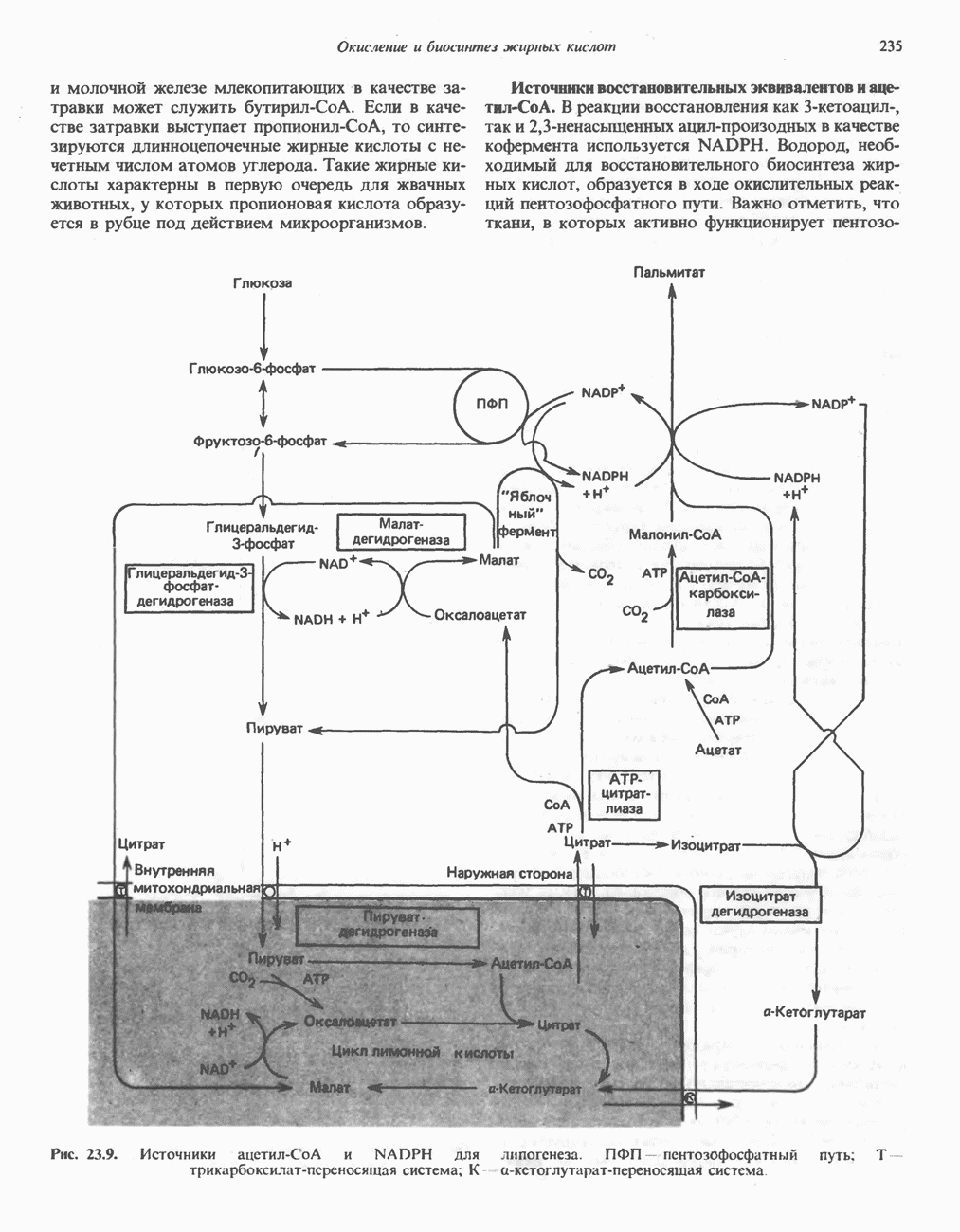
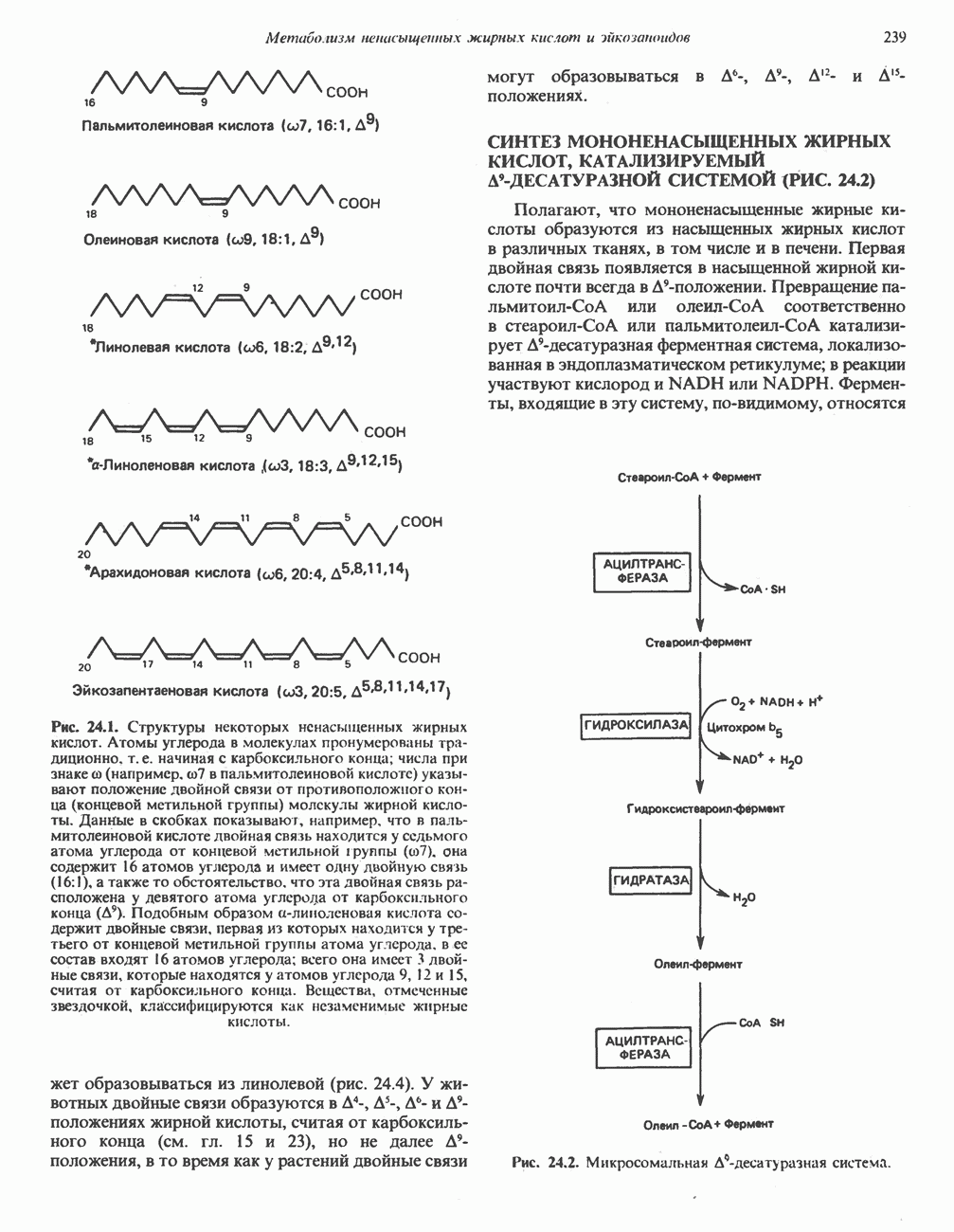
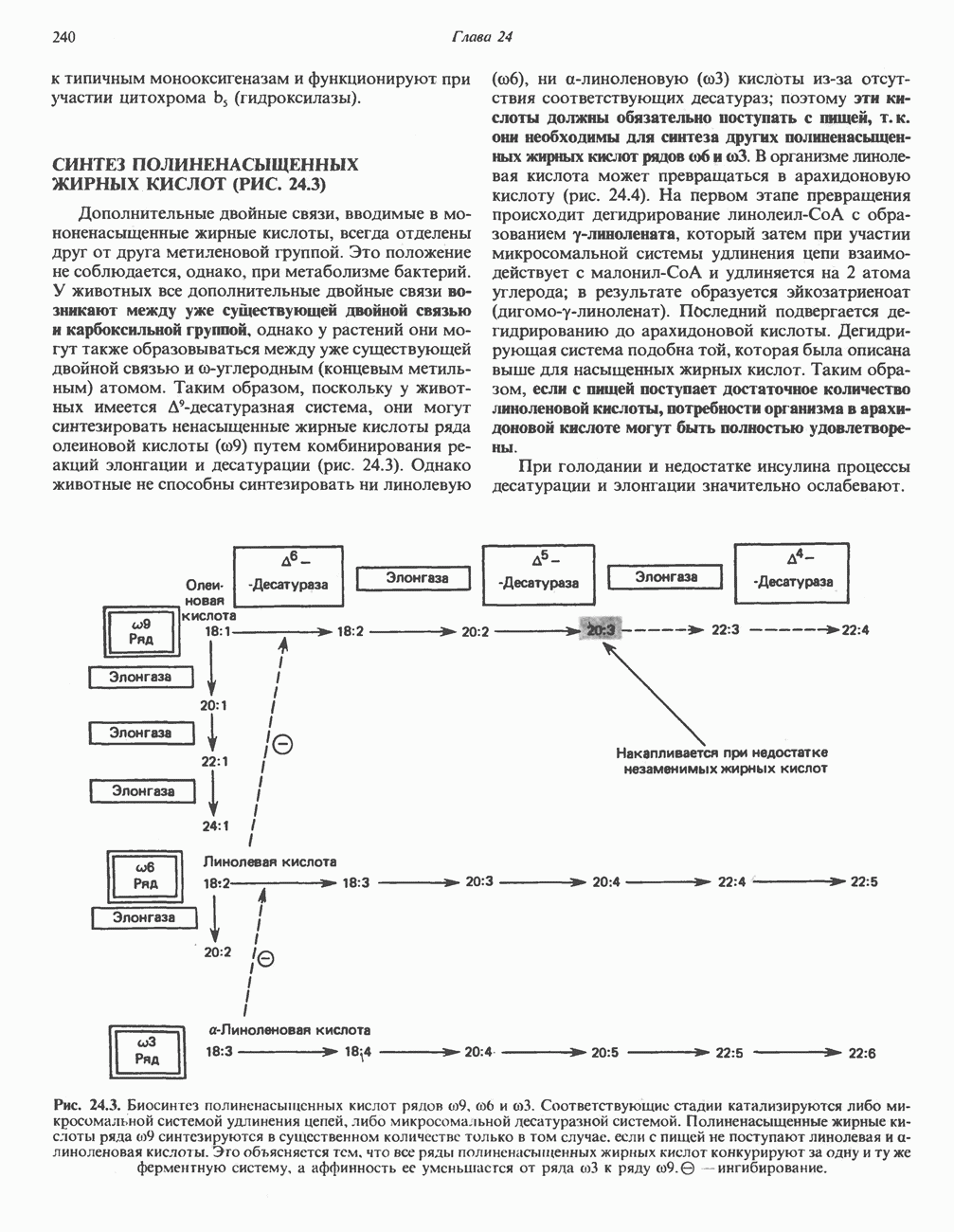
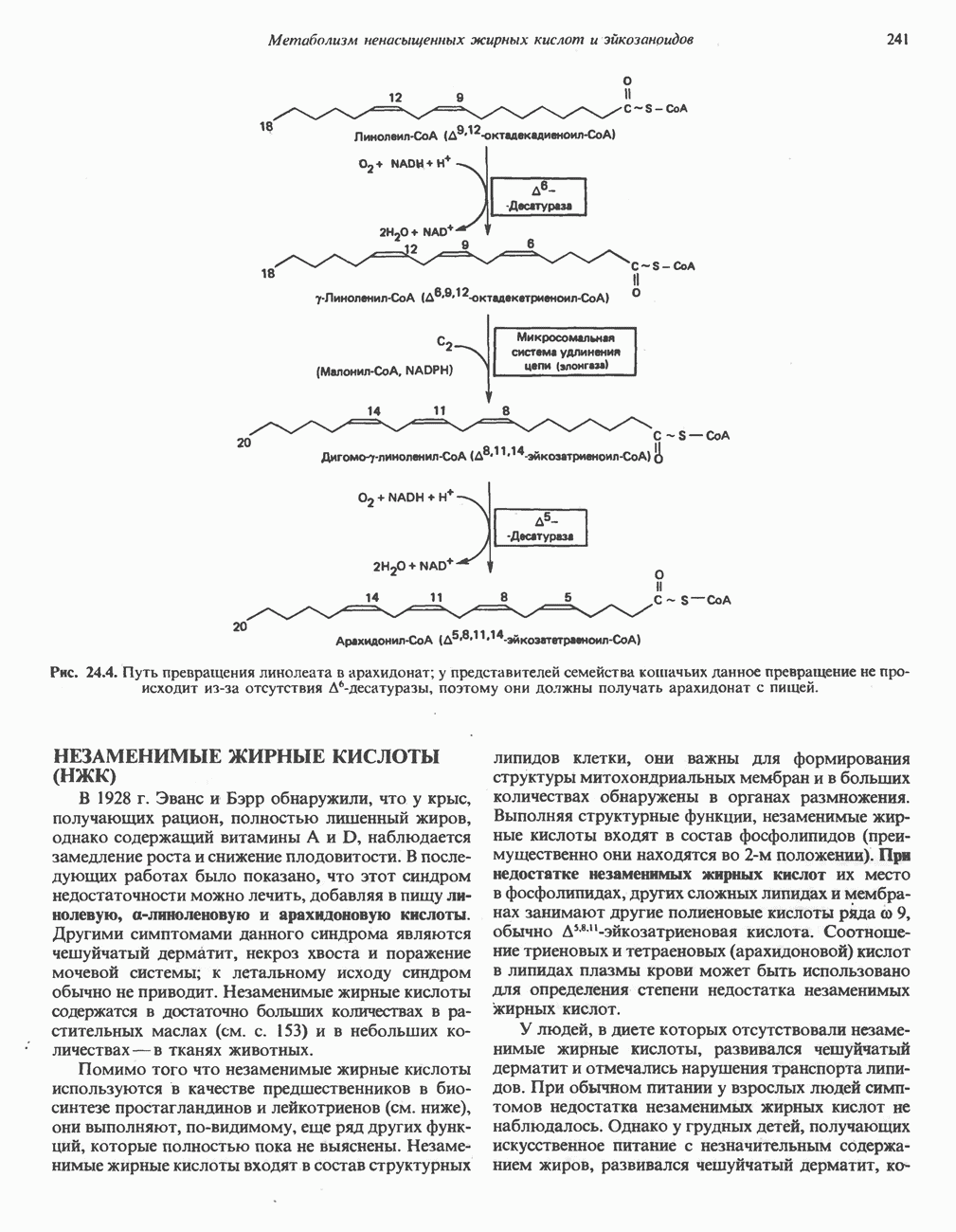
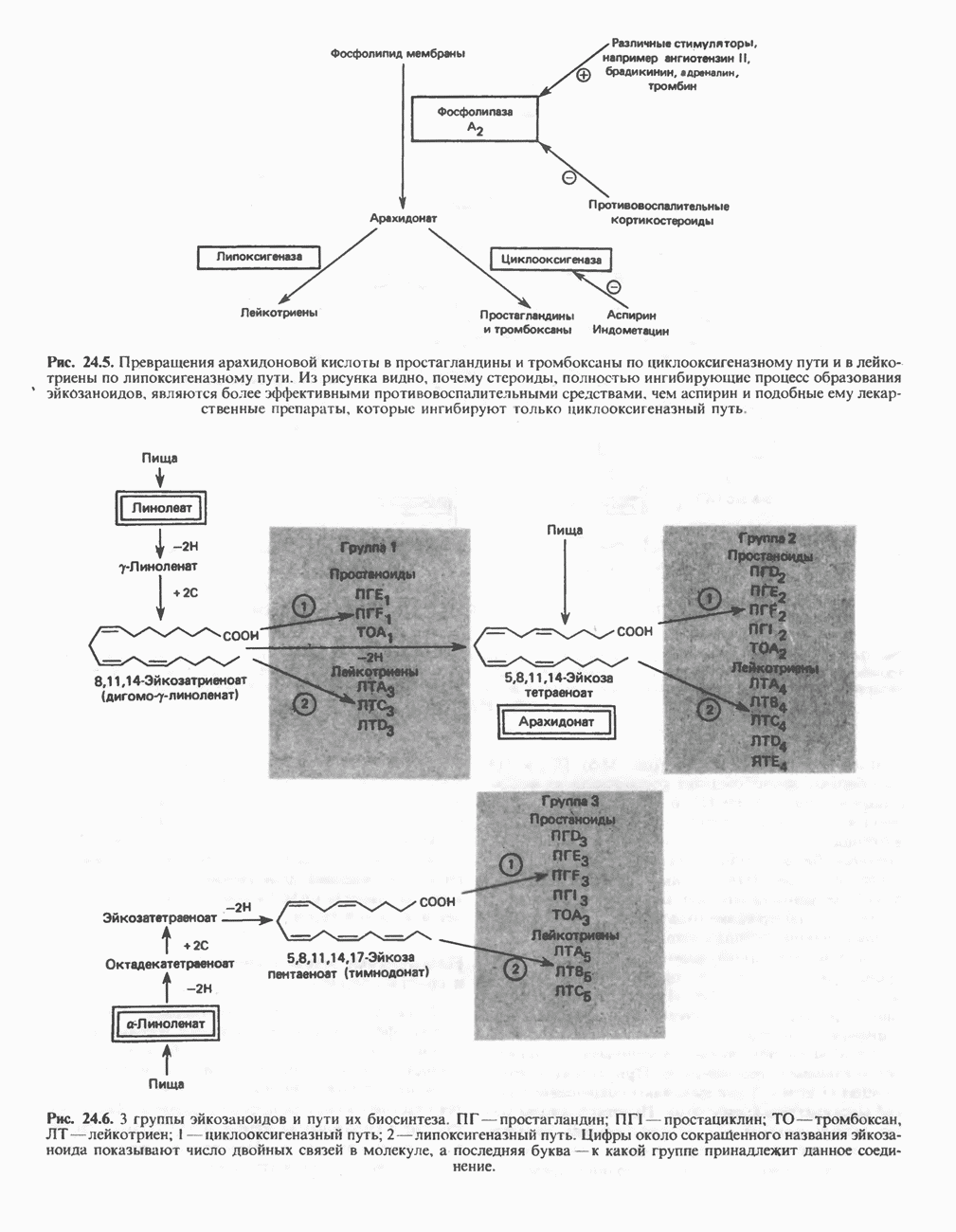
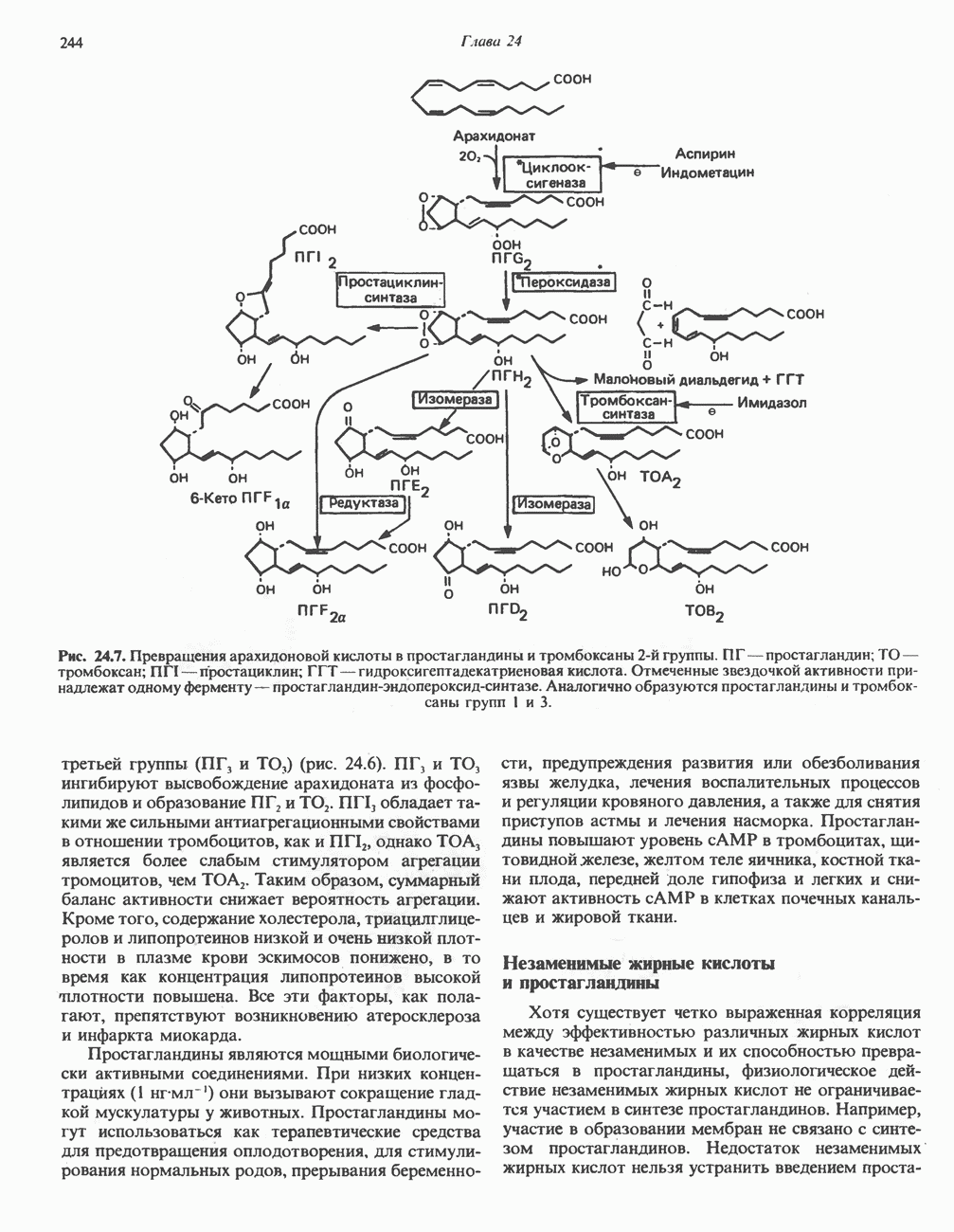
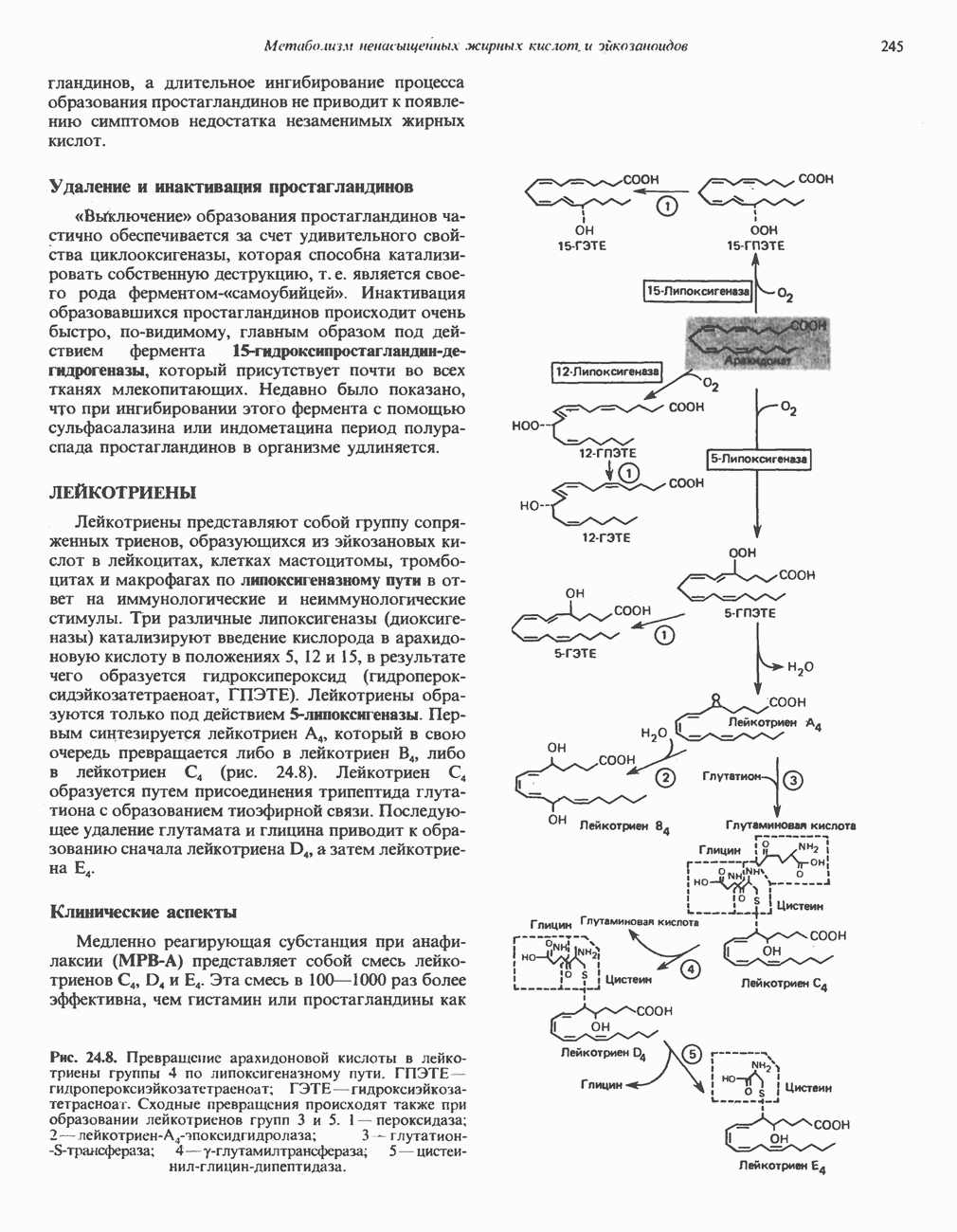
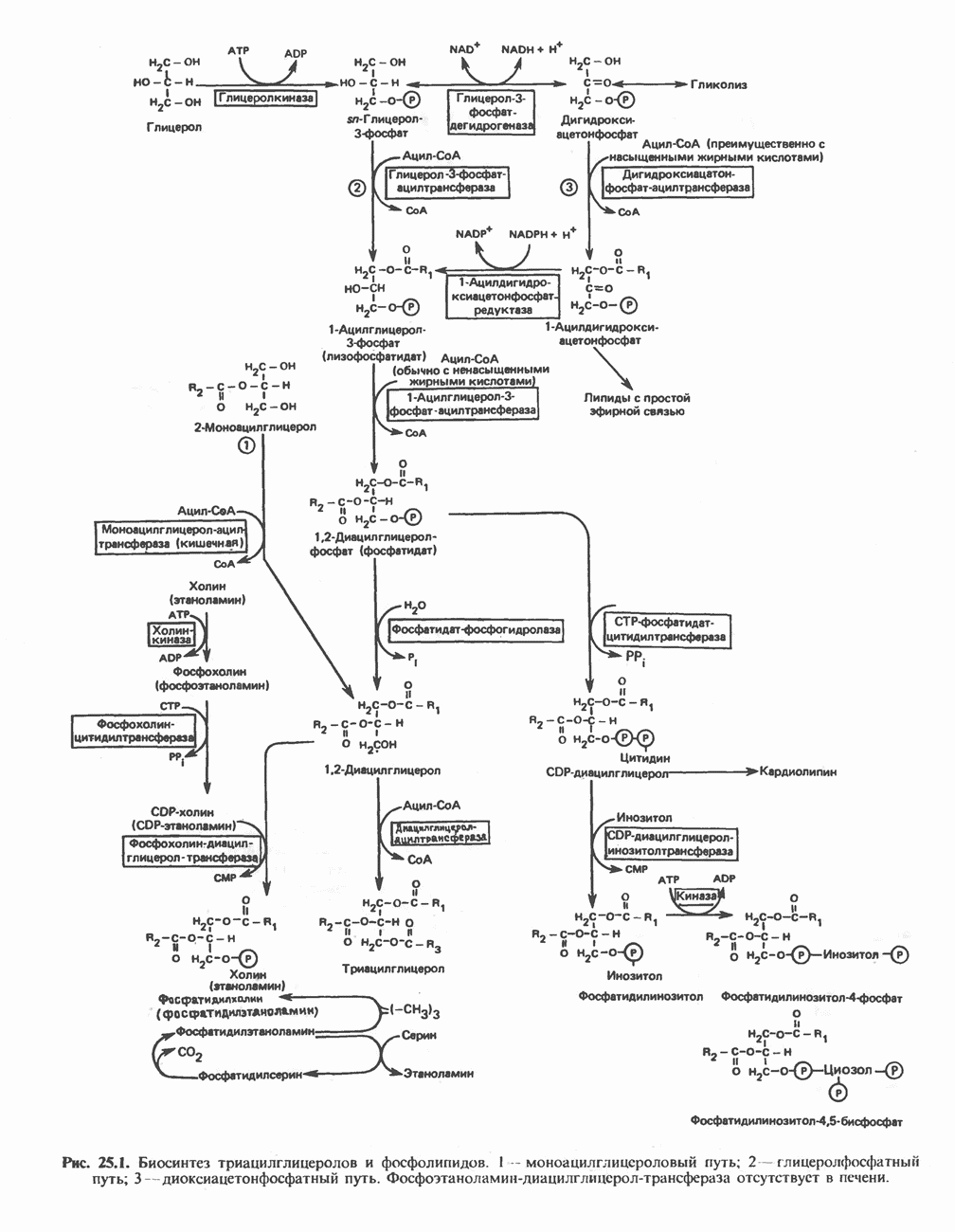
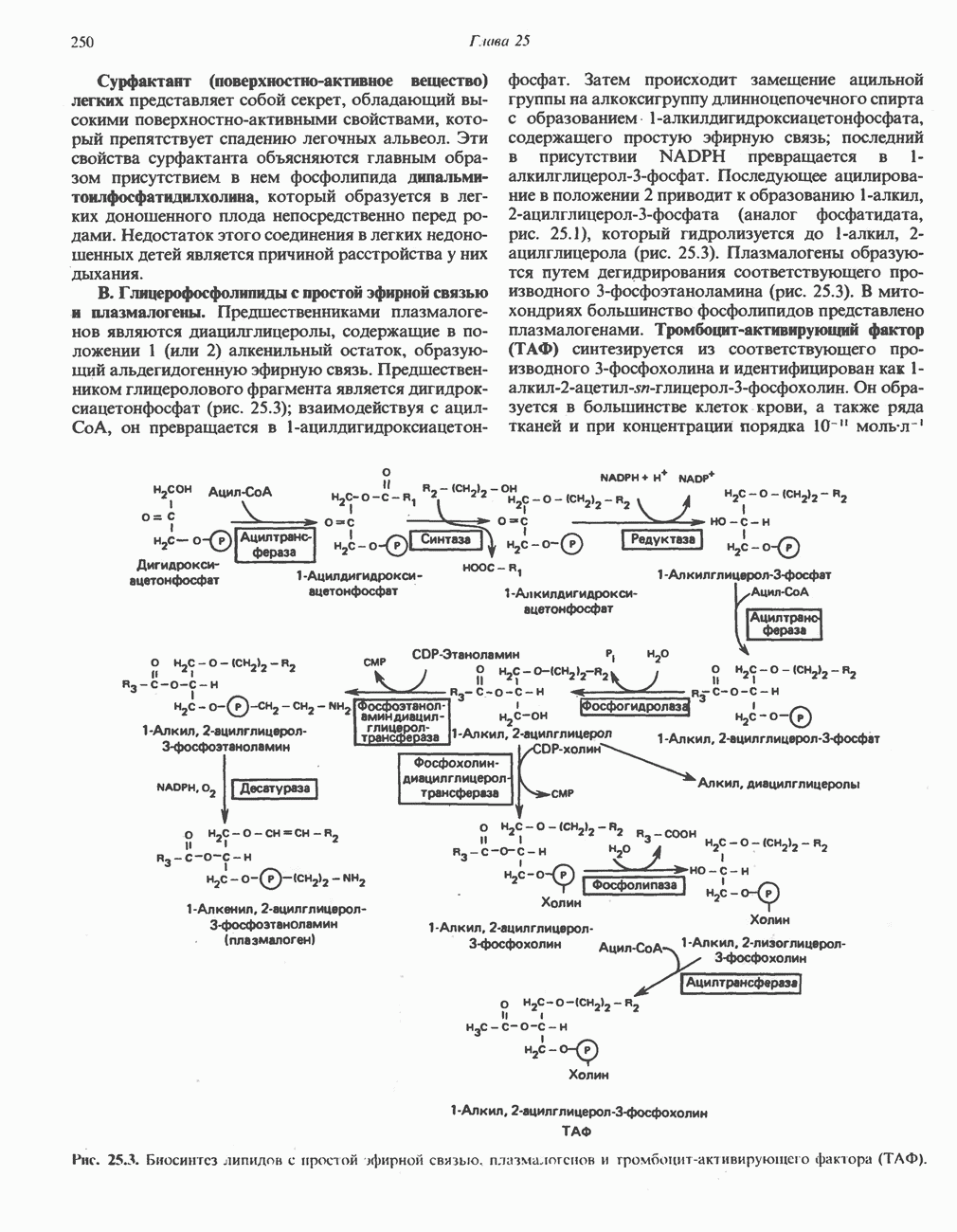
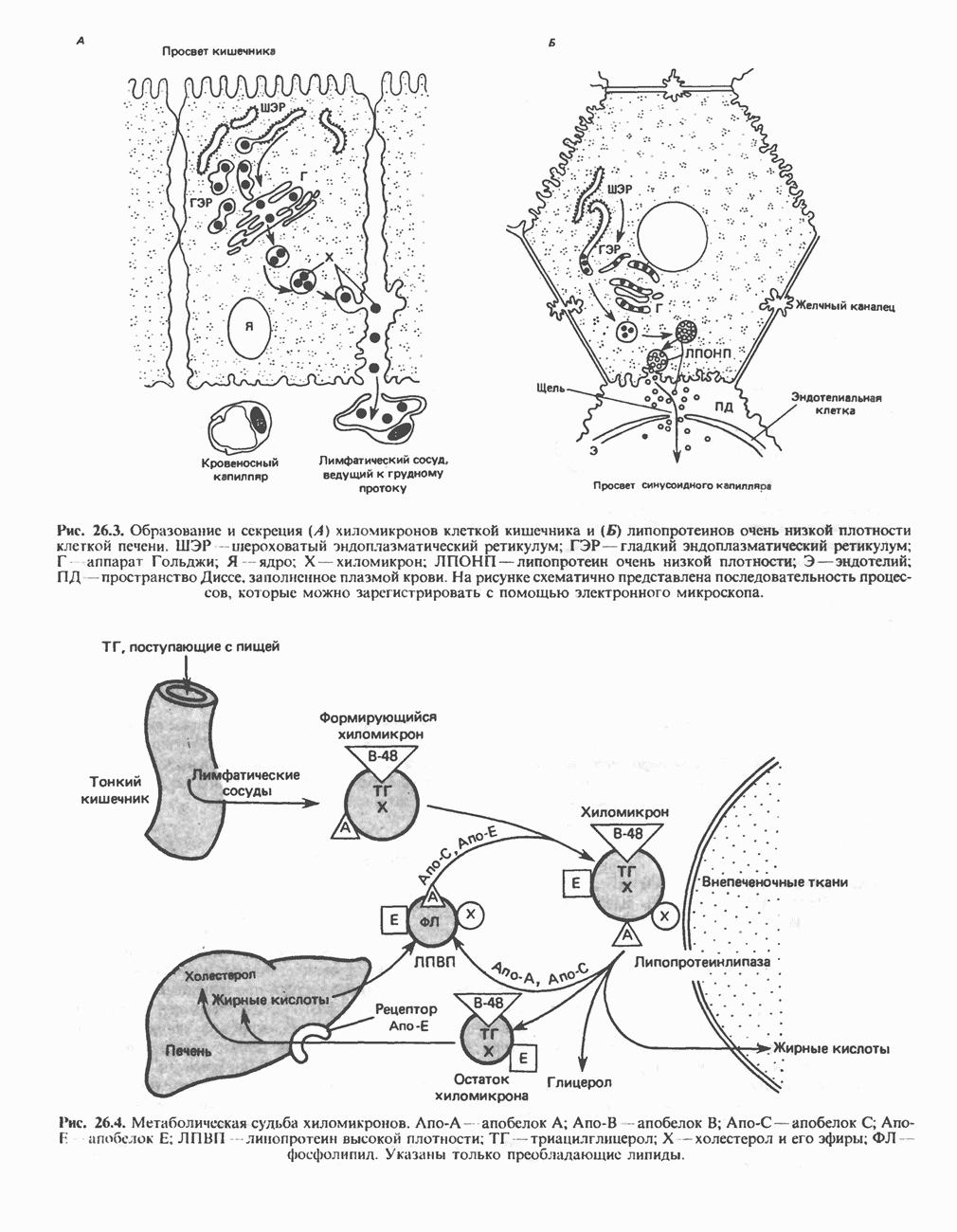
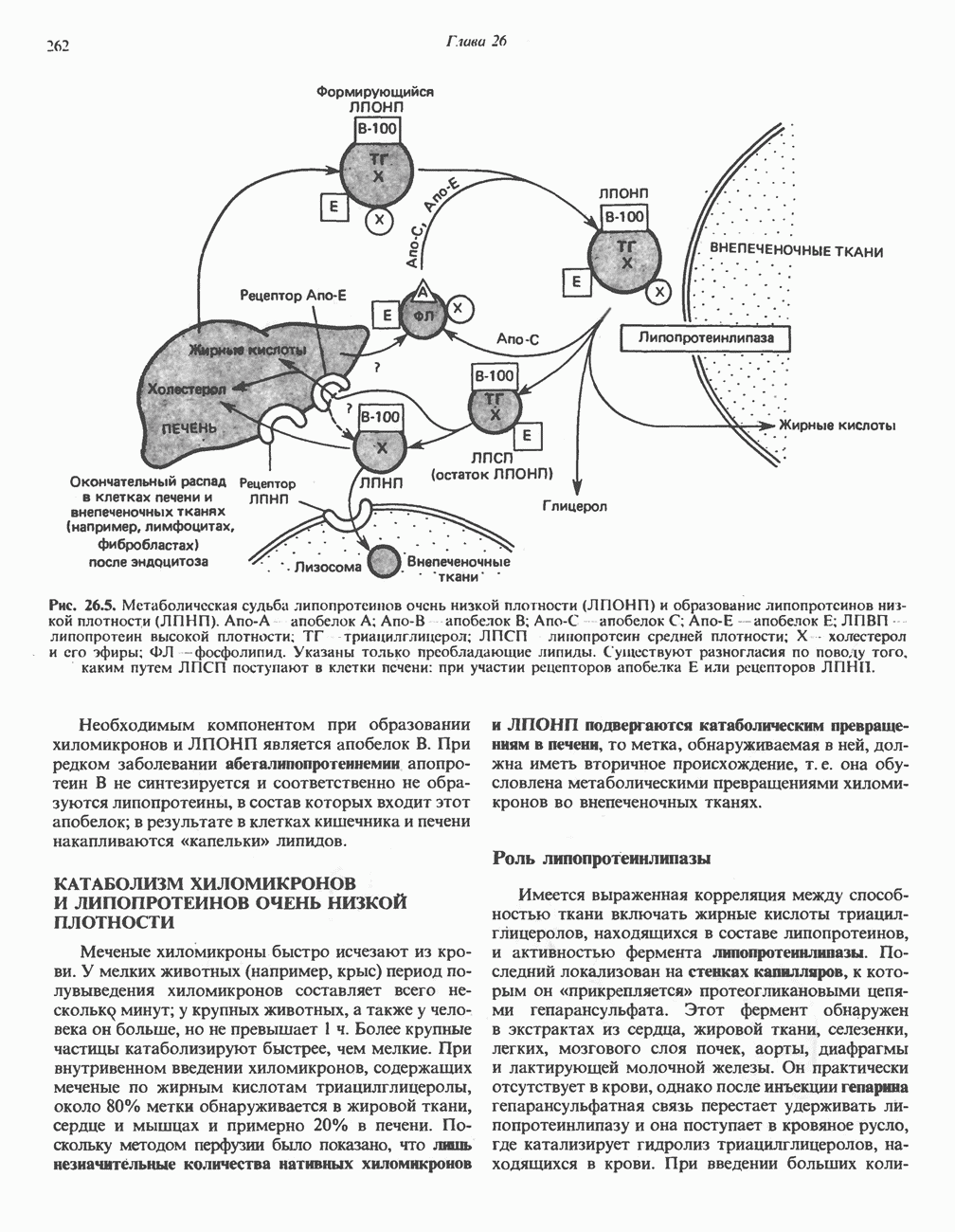
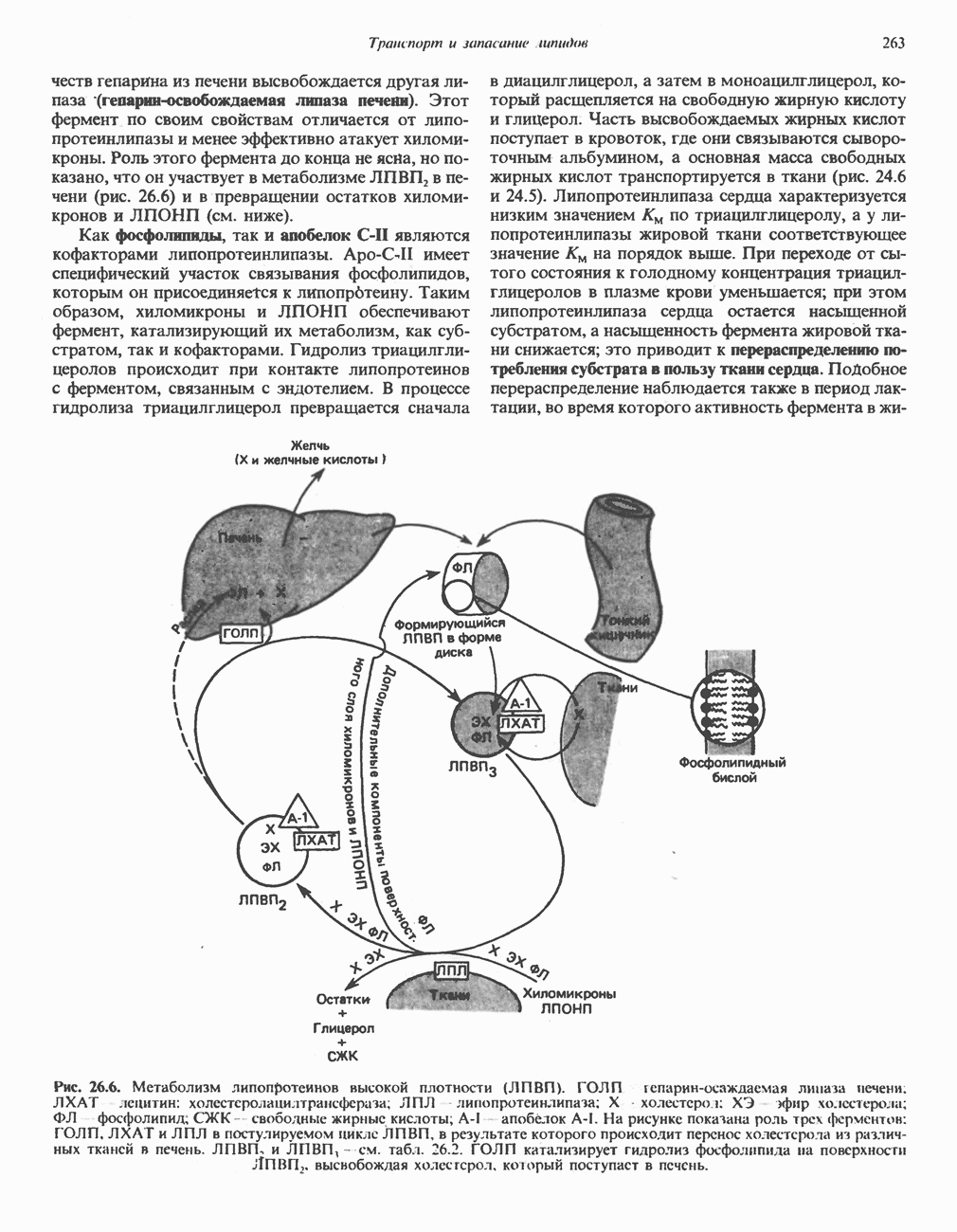
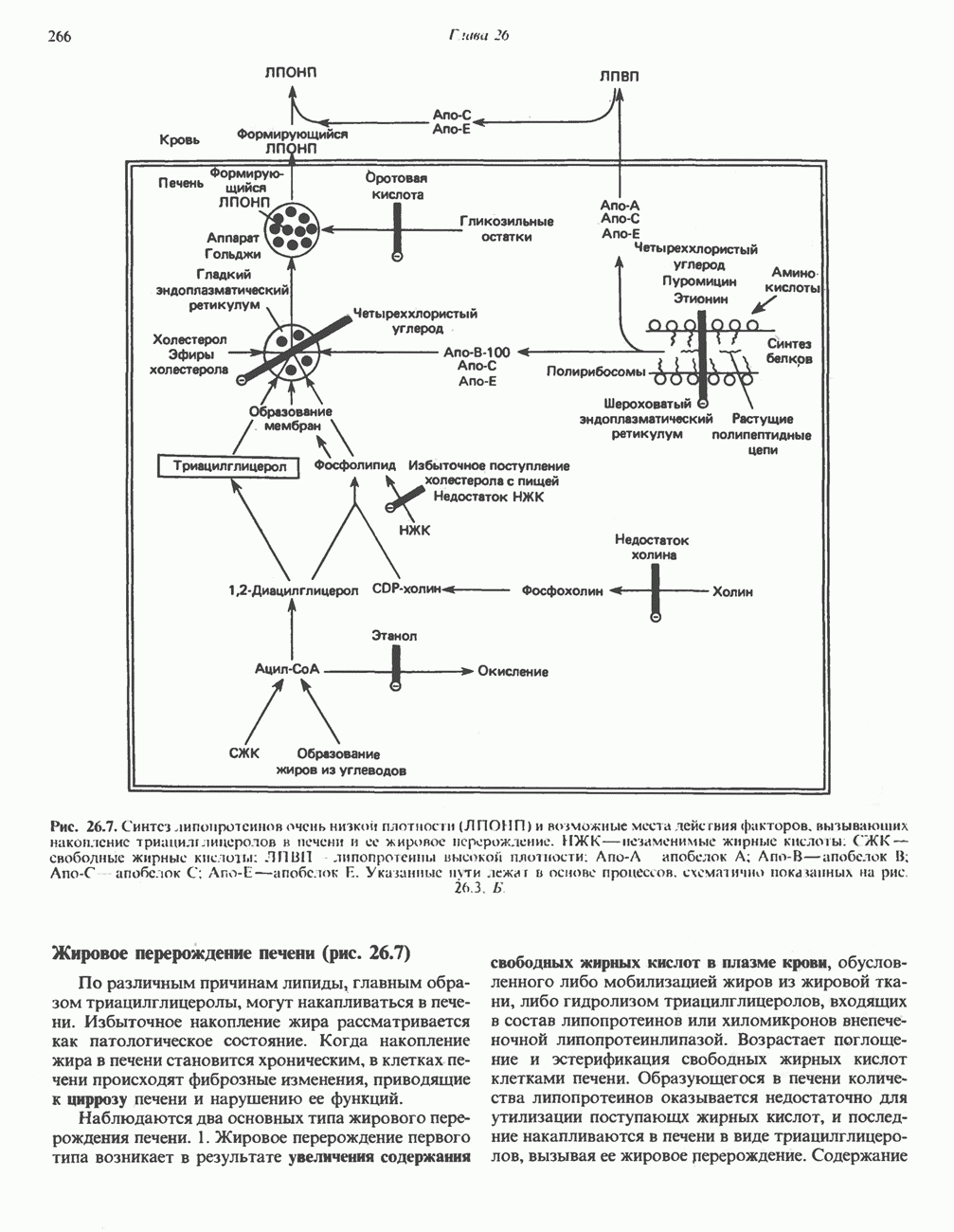
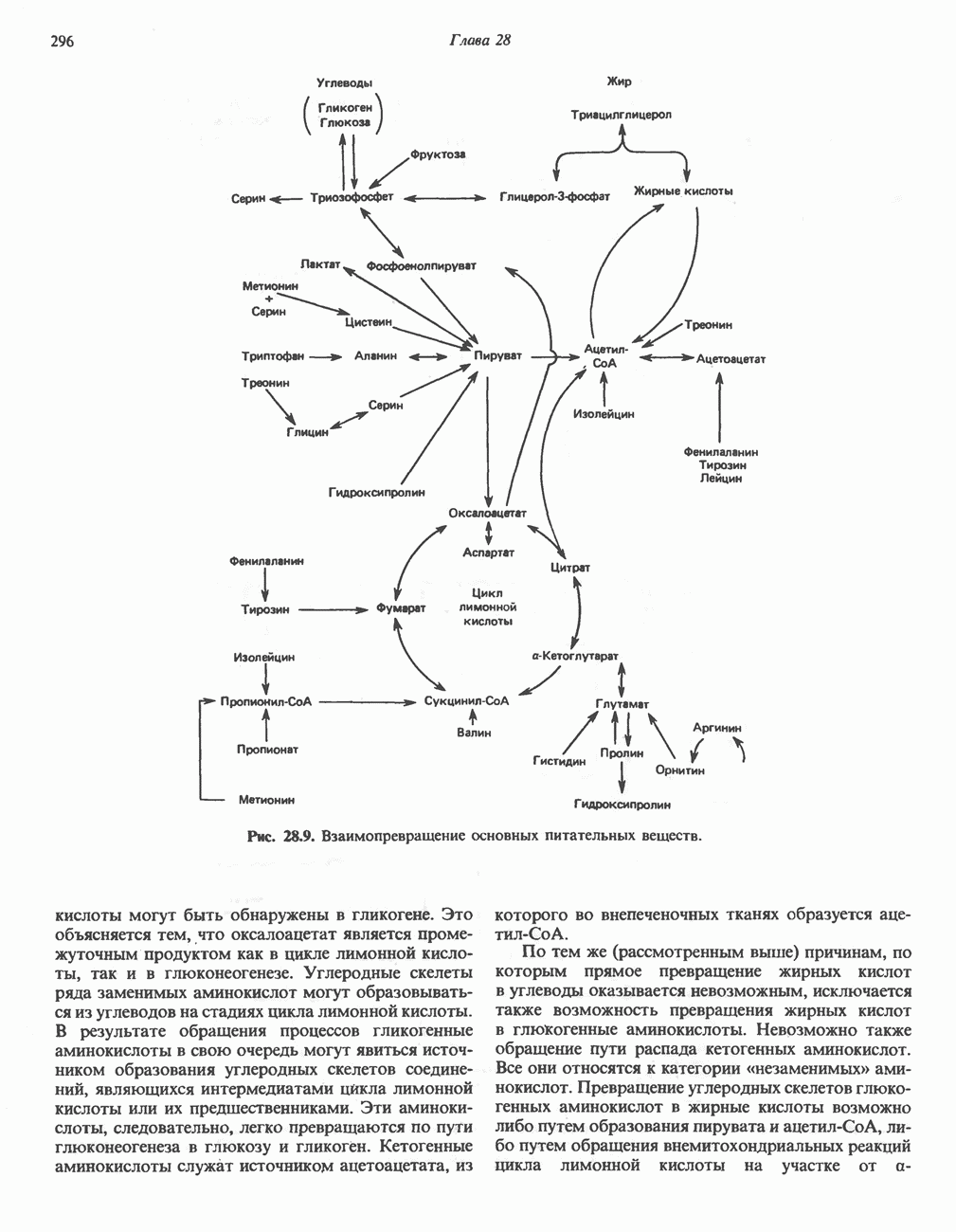
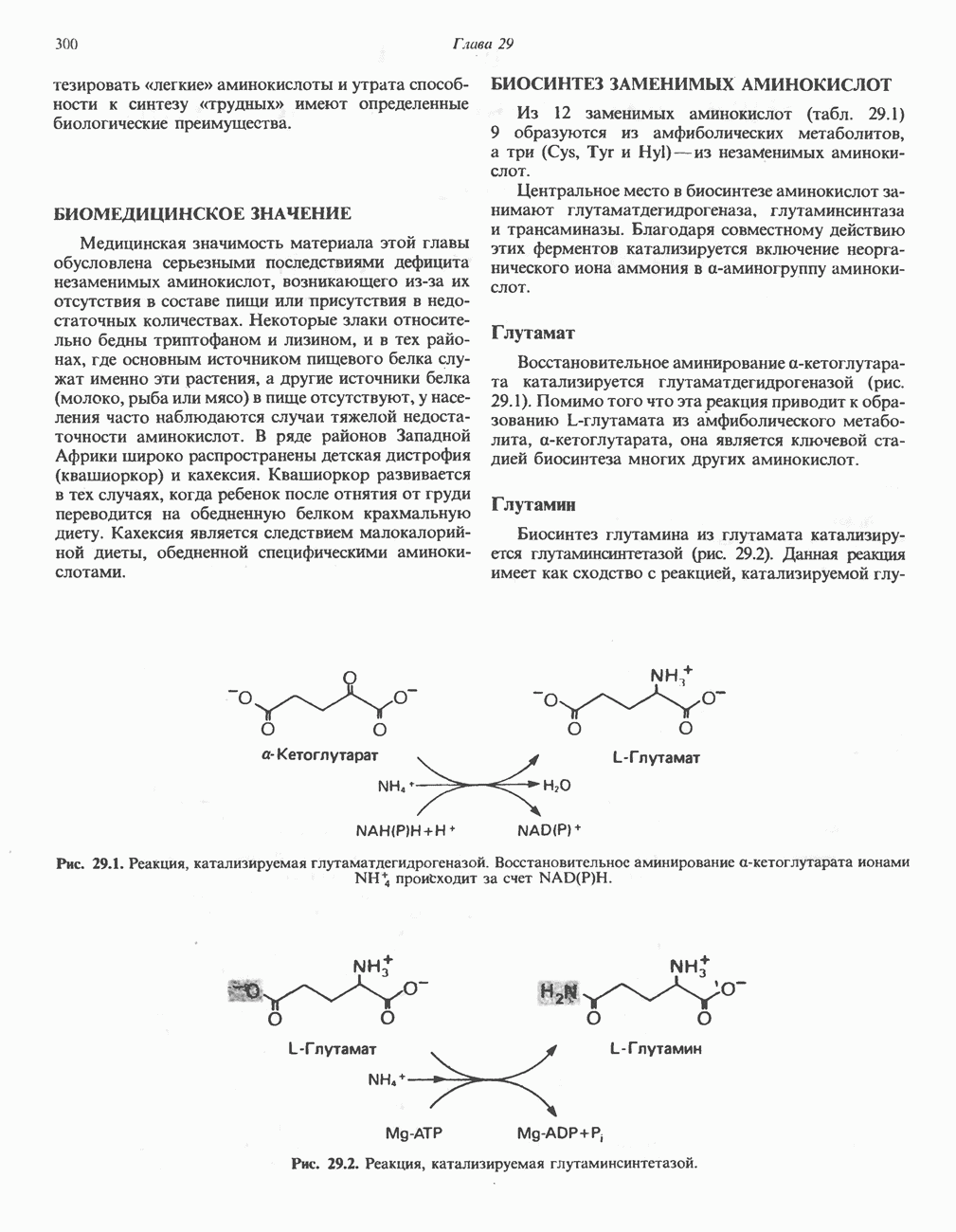
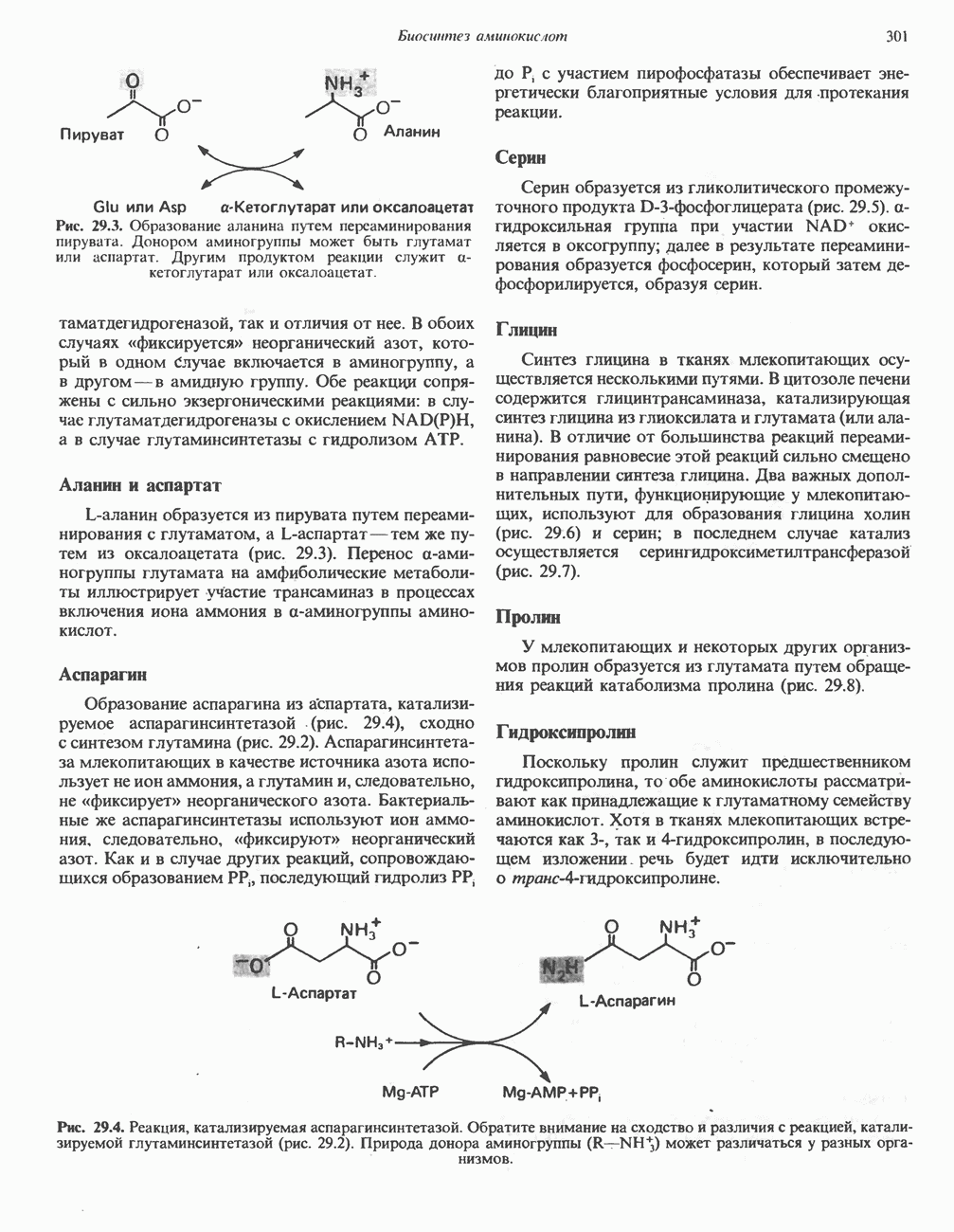
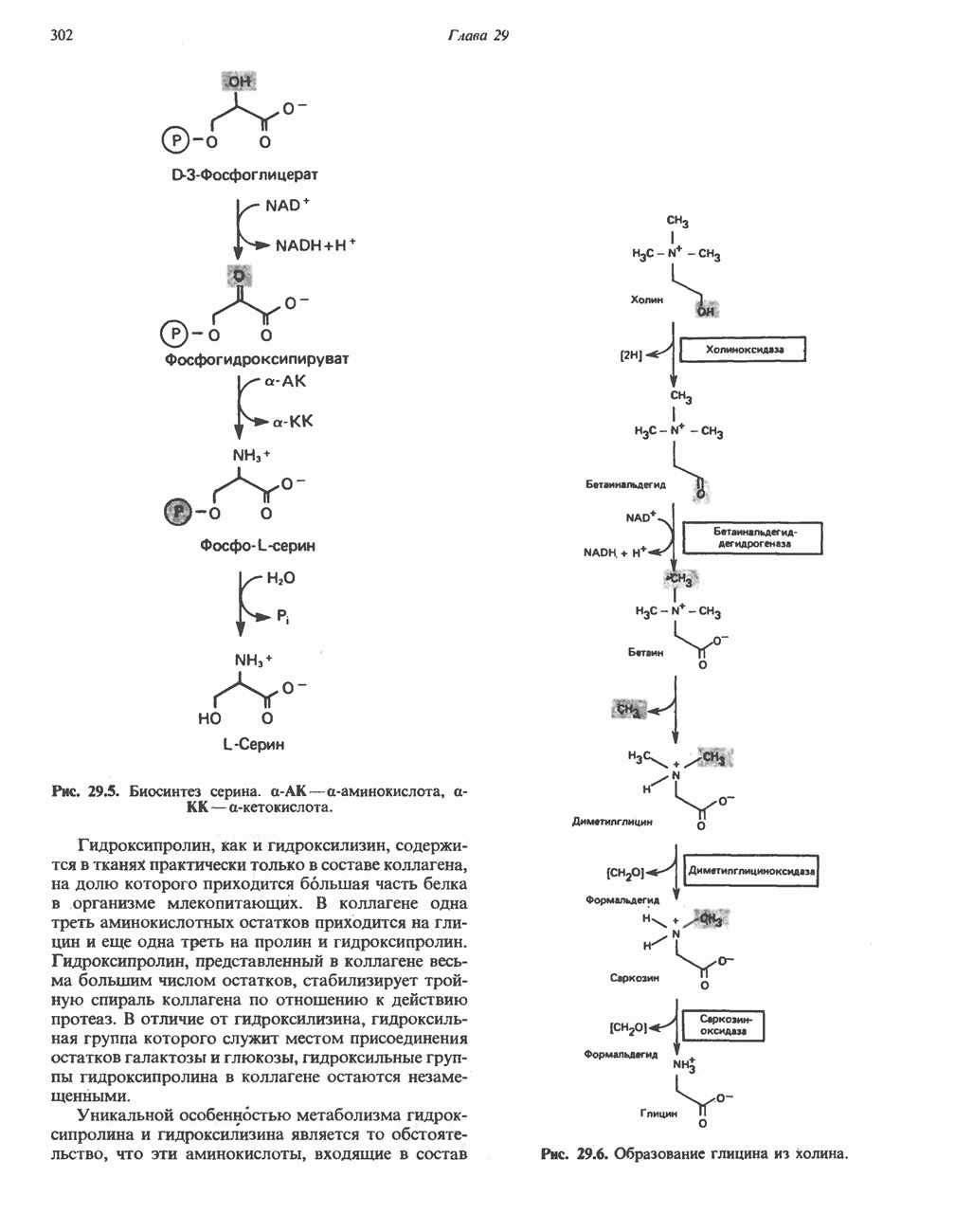
МЕХАНИЗМЫ КОНТРОЛЯ ГЛИКОГЕНОЛИЗА И ГЛИКОГЕНЕЗА
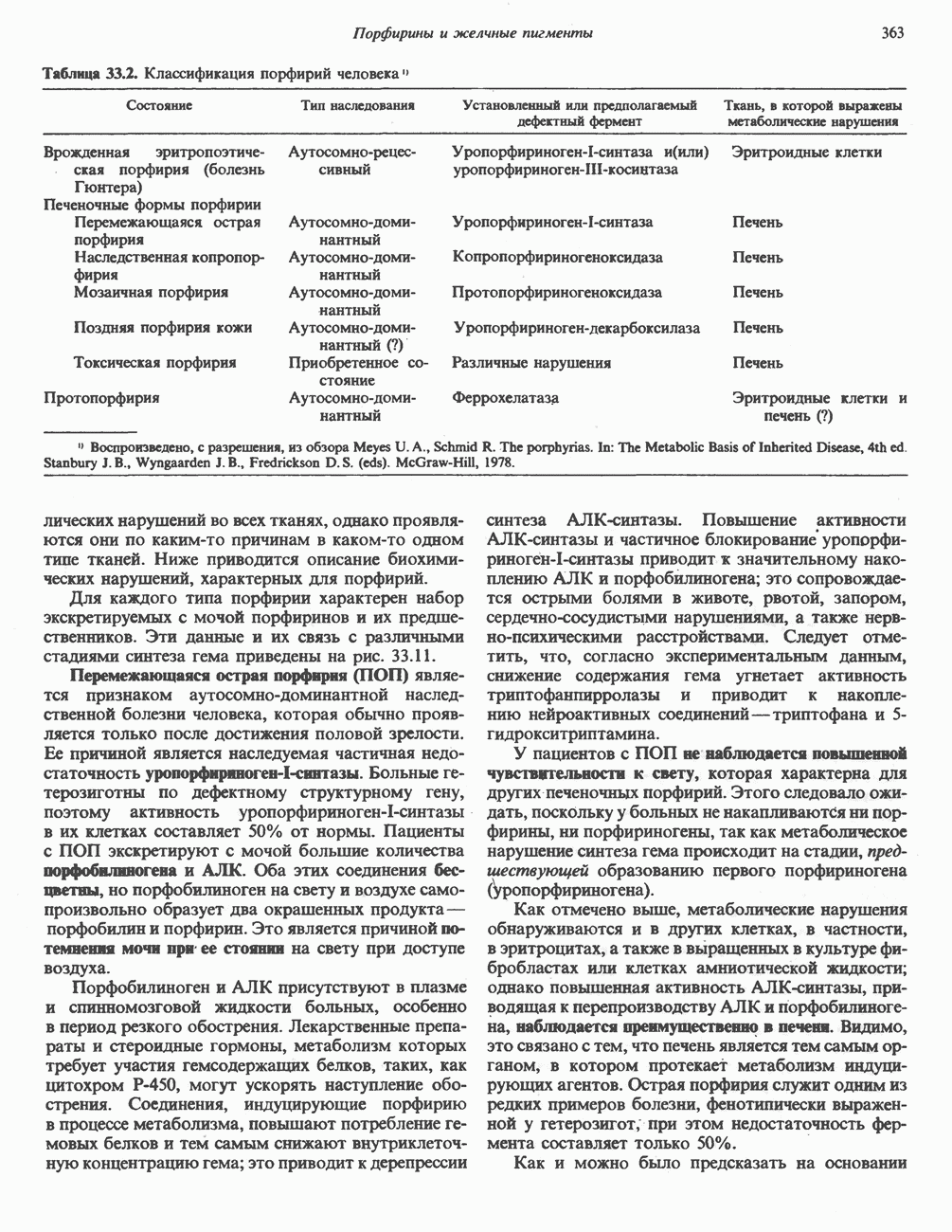
Главные ферменты, контролирующие метаболизм гликогена, — гликогенфосфорилаза и гликоген-синтаза — регулируются сложной серией реакций, в которых используются как аллостерические механизмы (см. с. 104), так и ковалентная модификация путем фосфорилирования и дефосфорилирования фермента (см. с. 108).
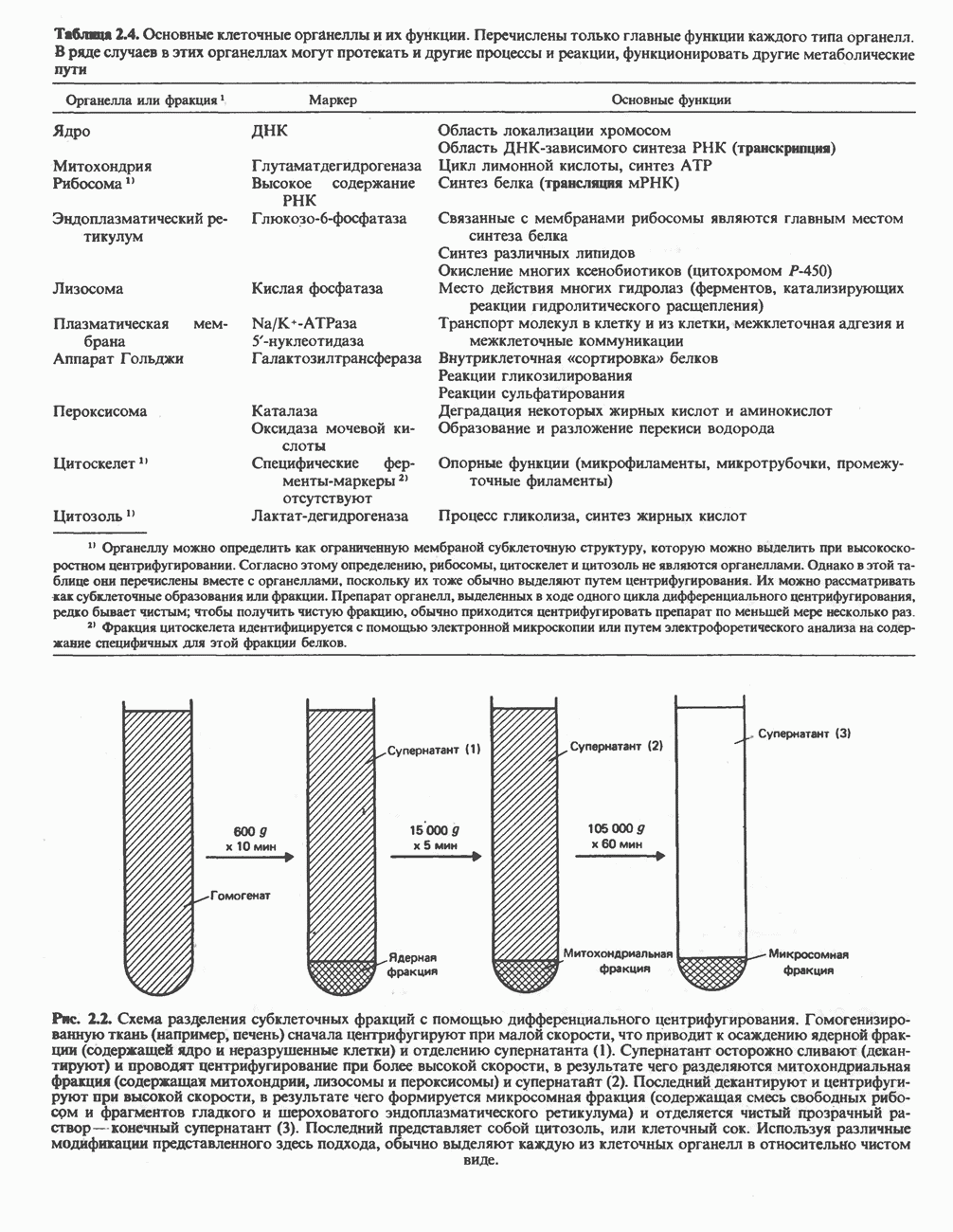
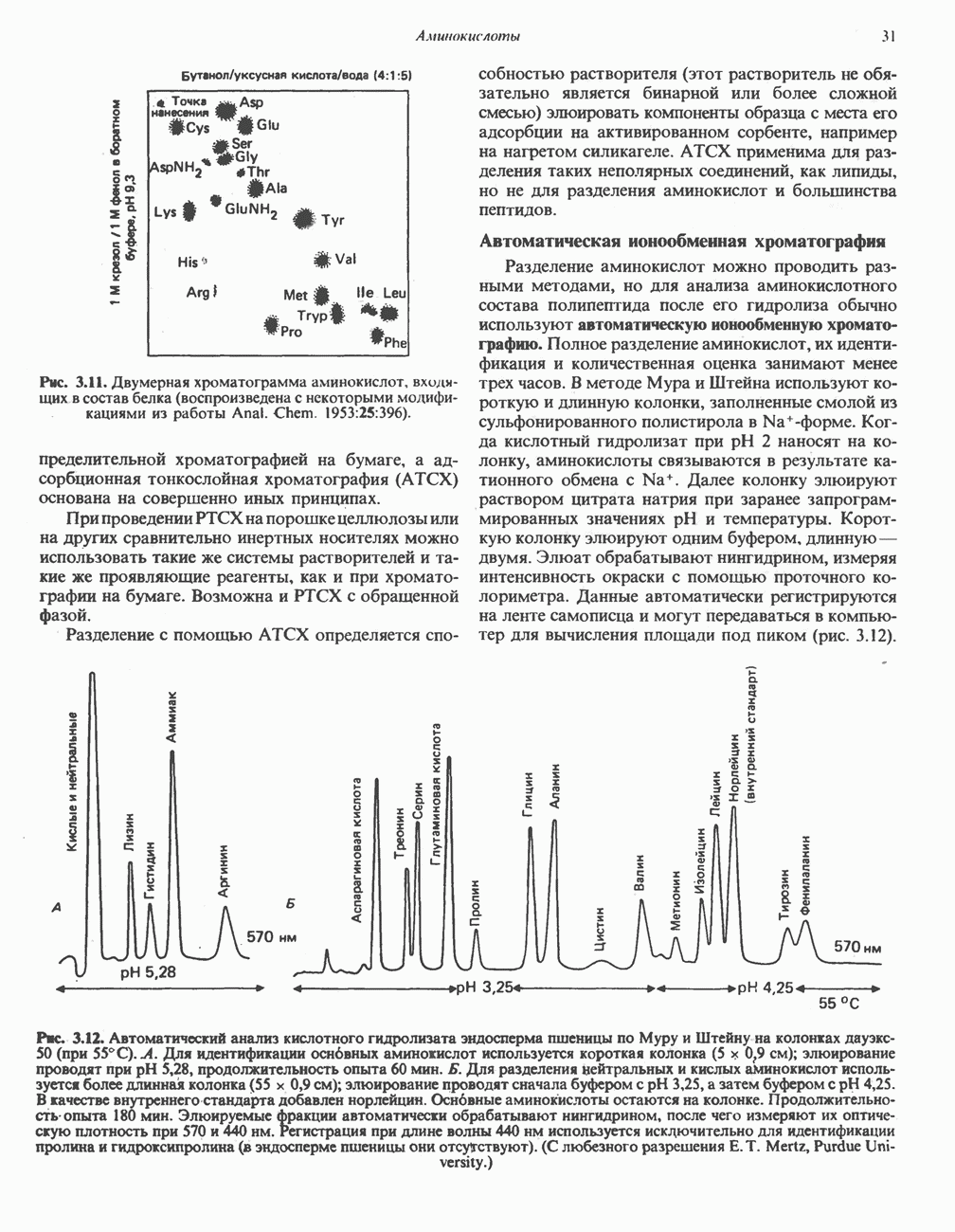
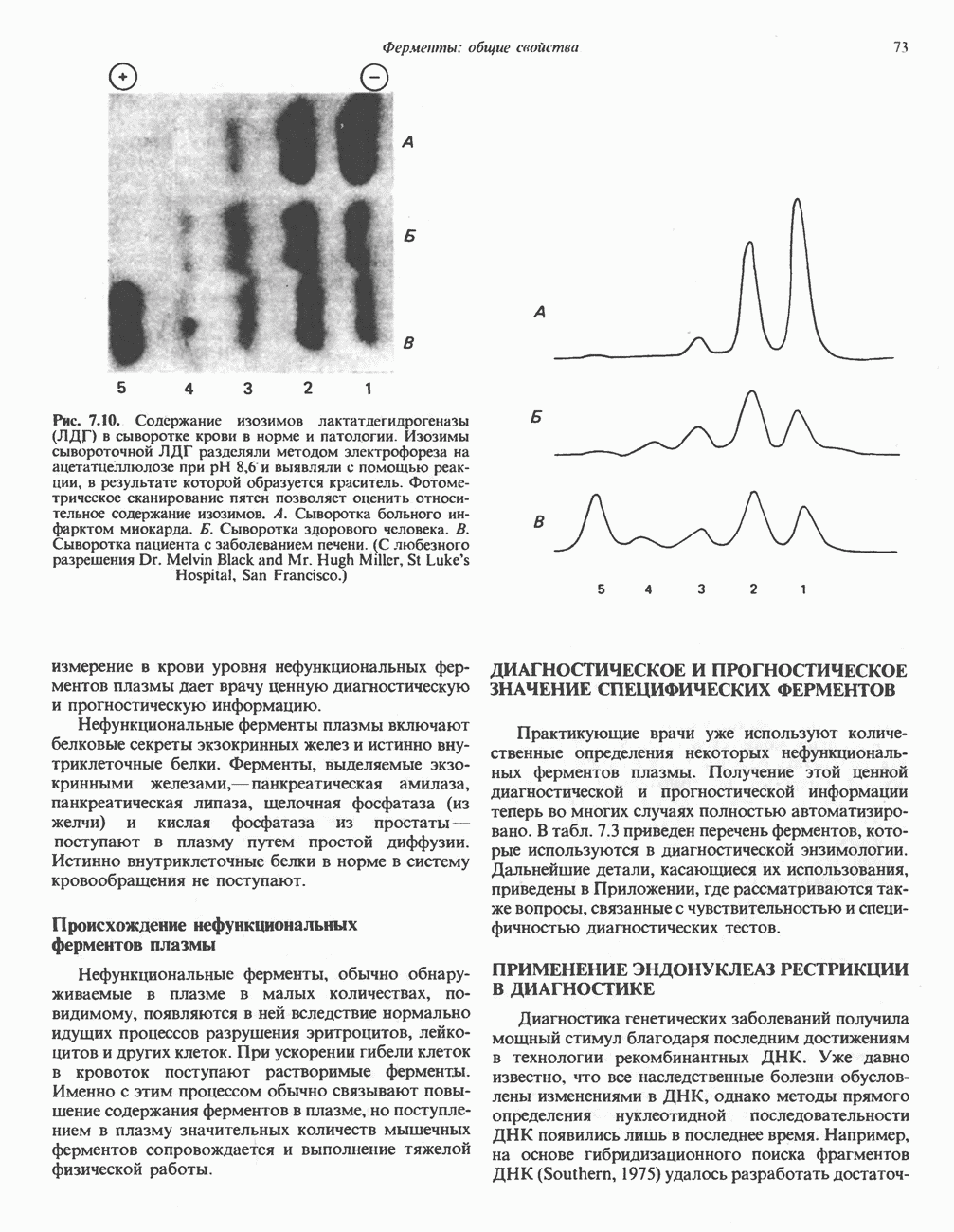
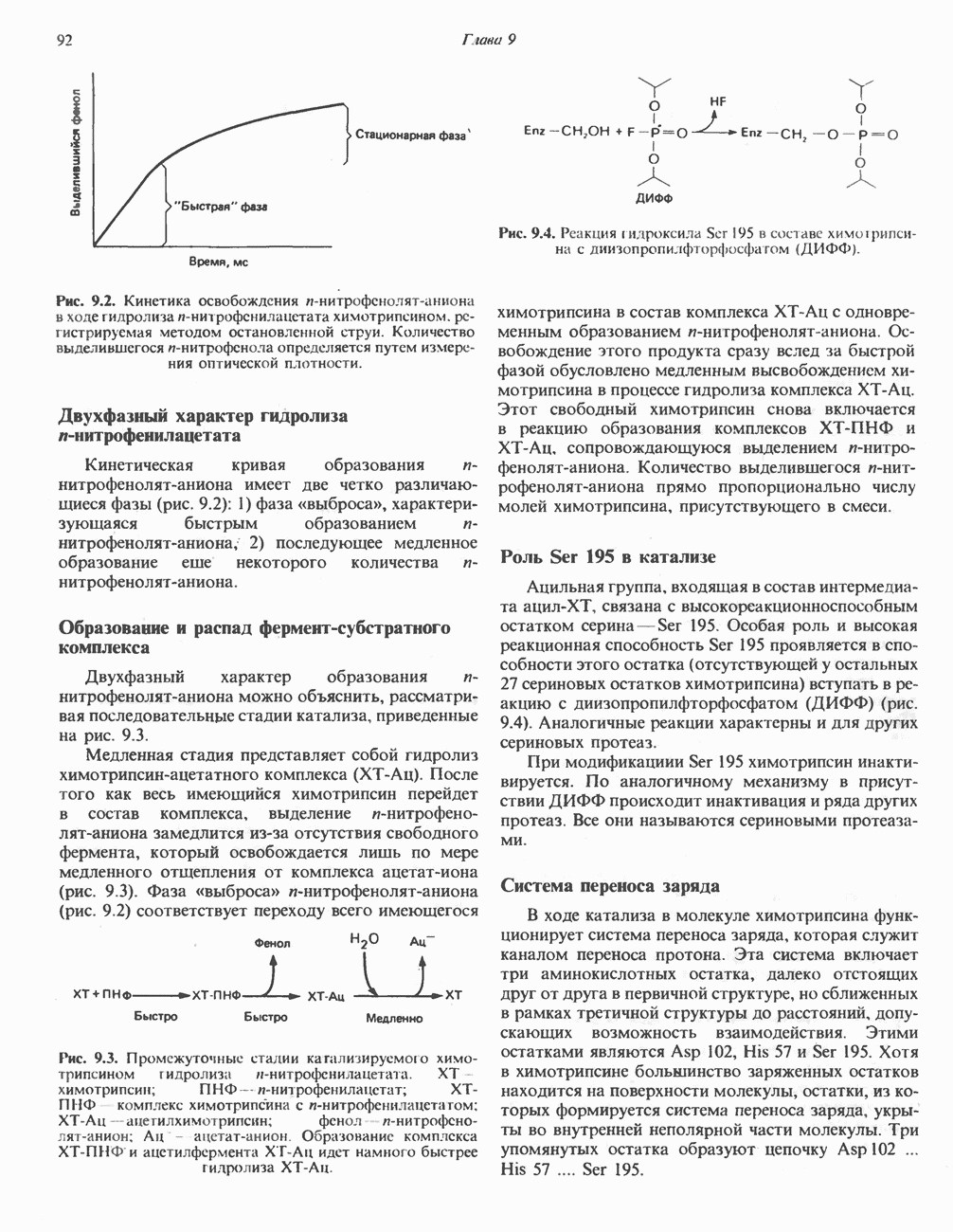
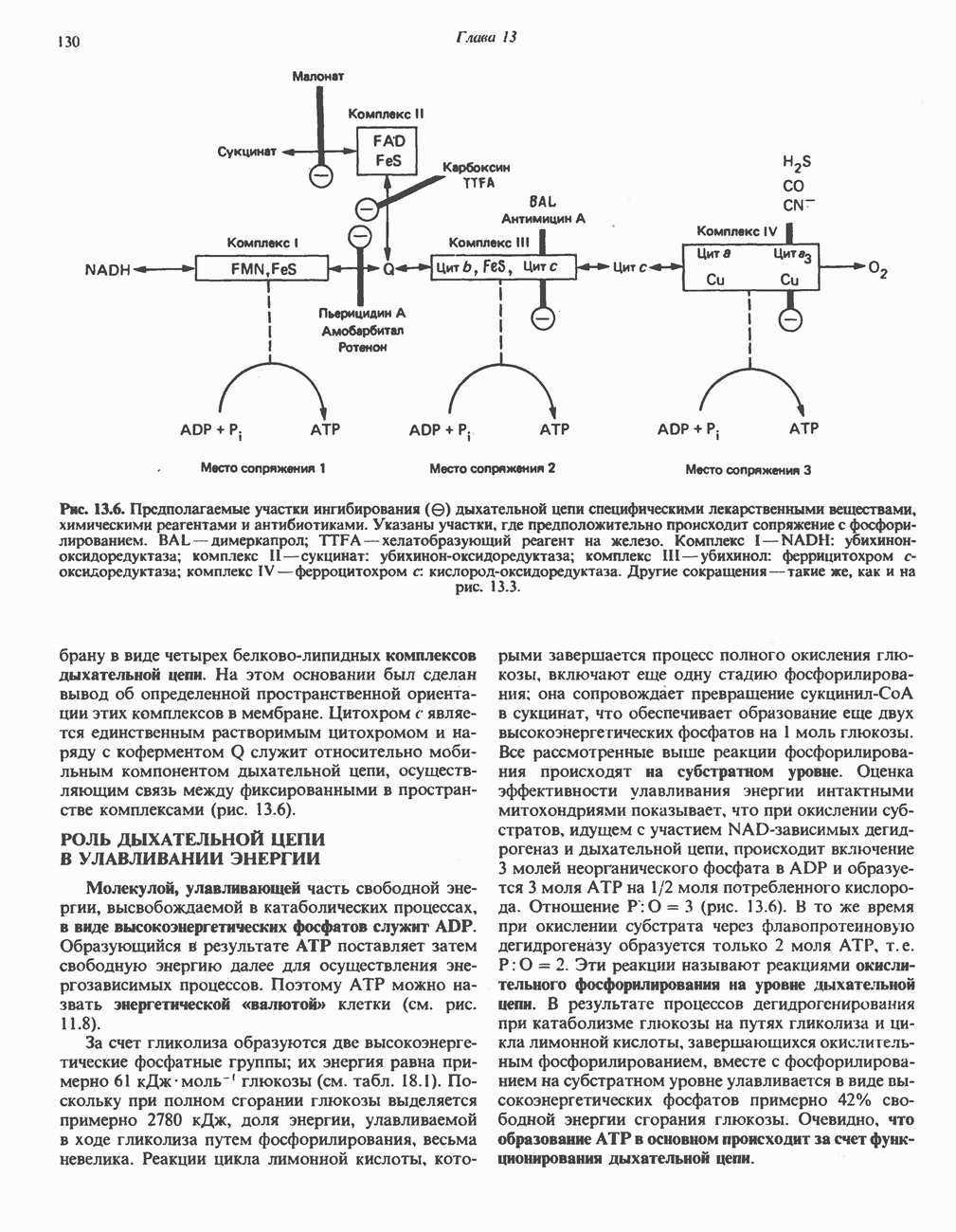
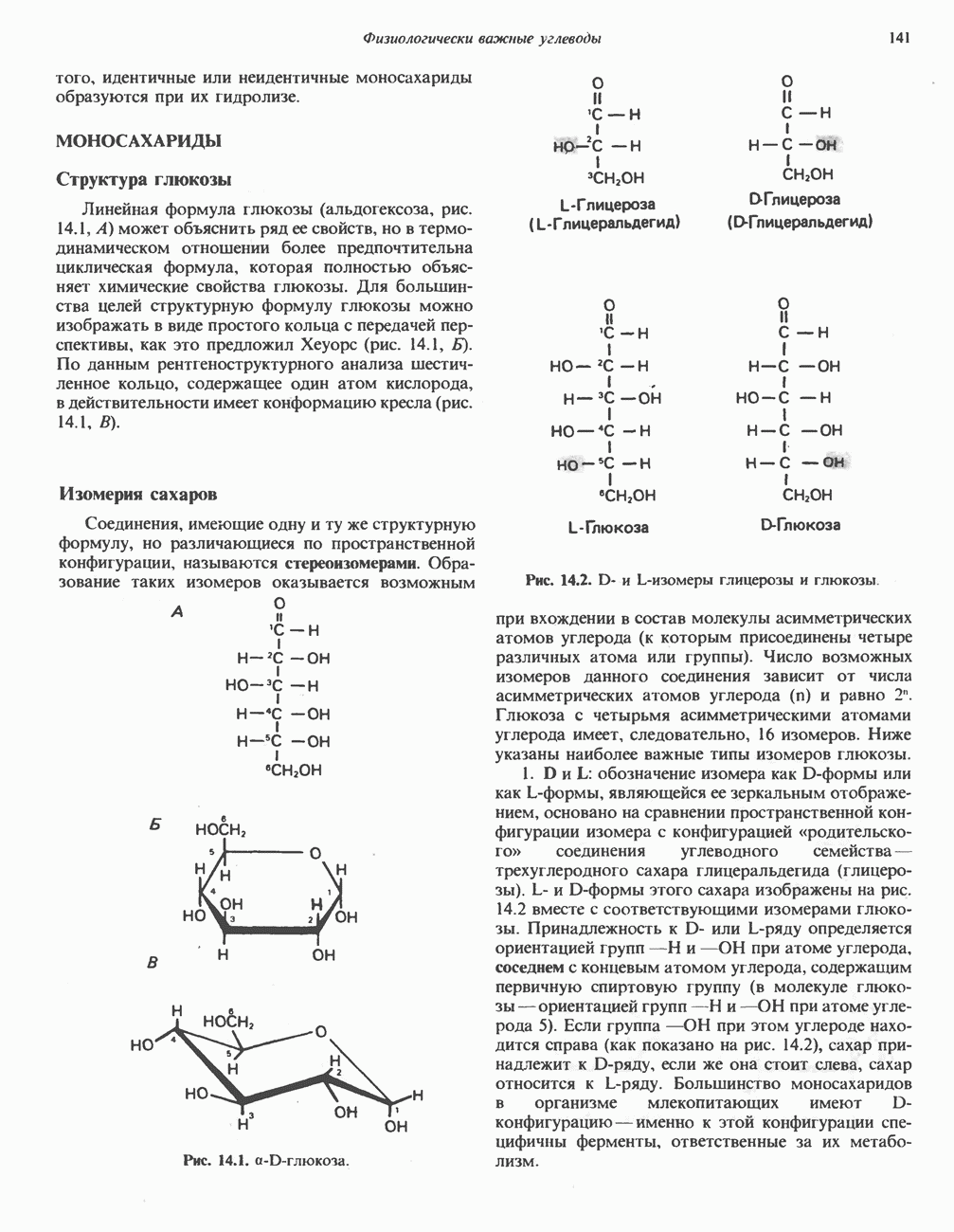
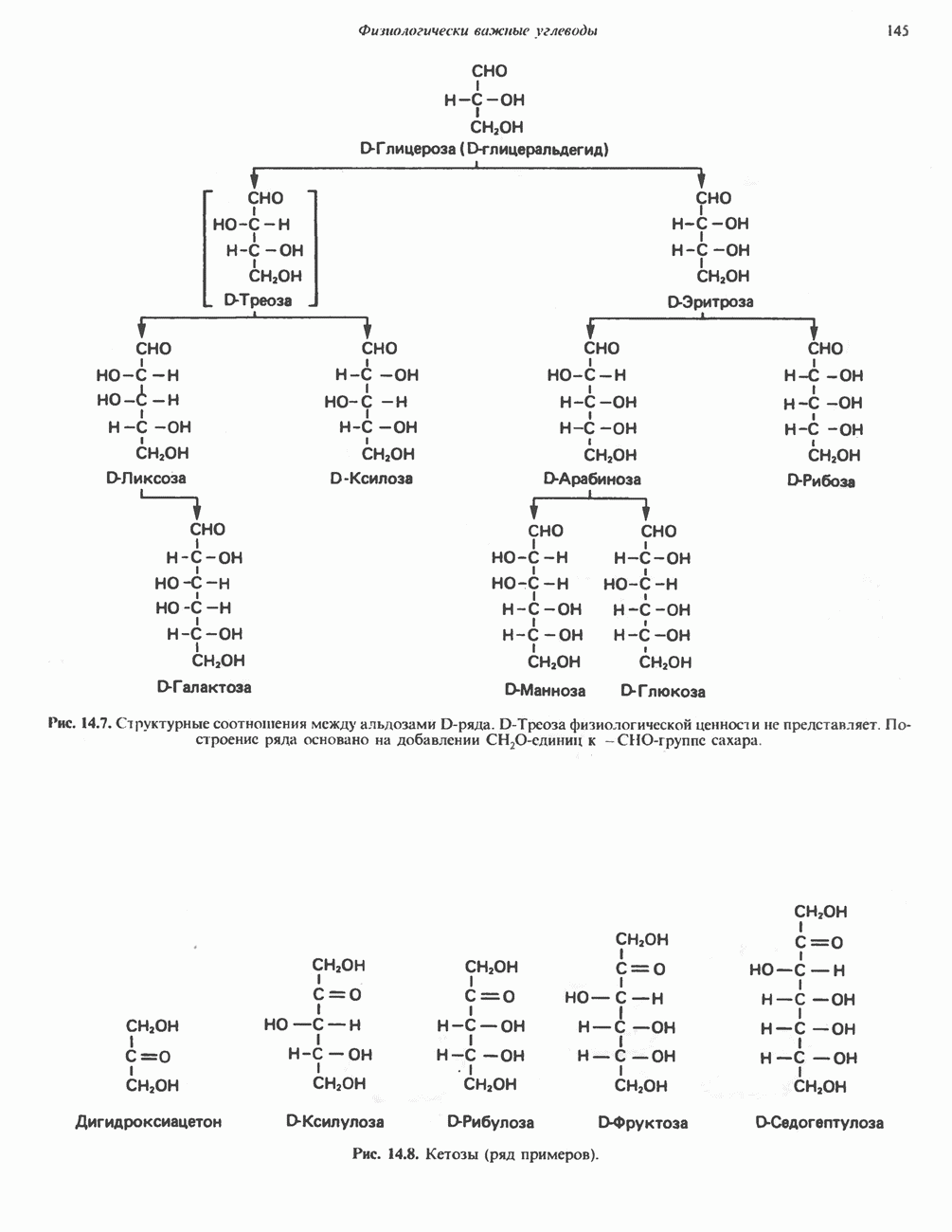
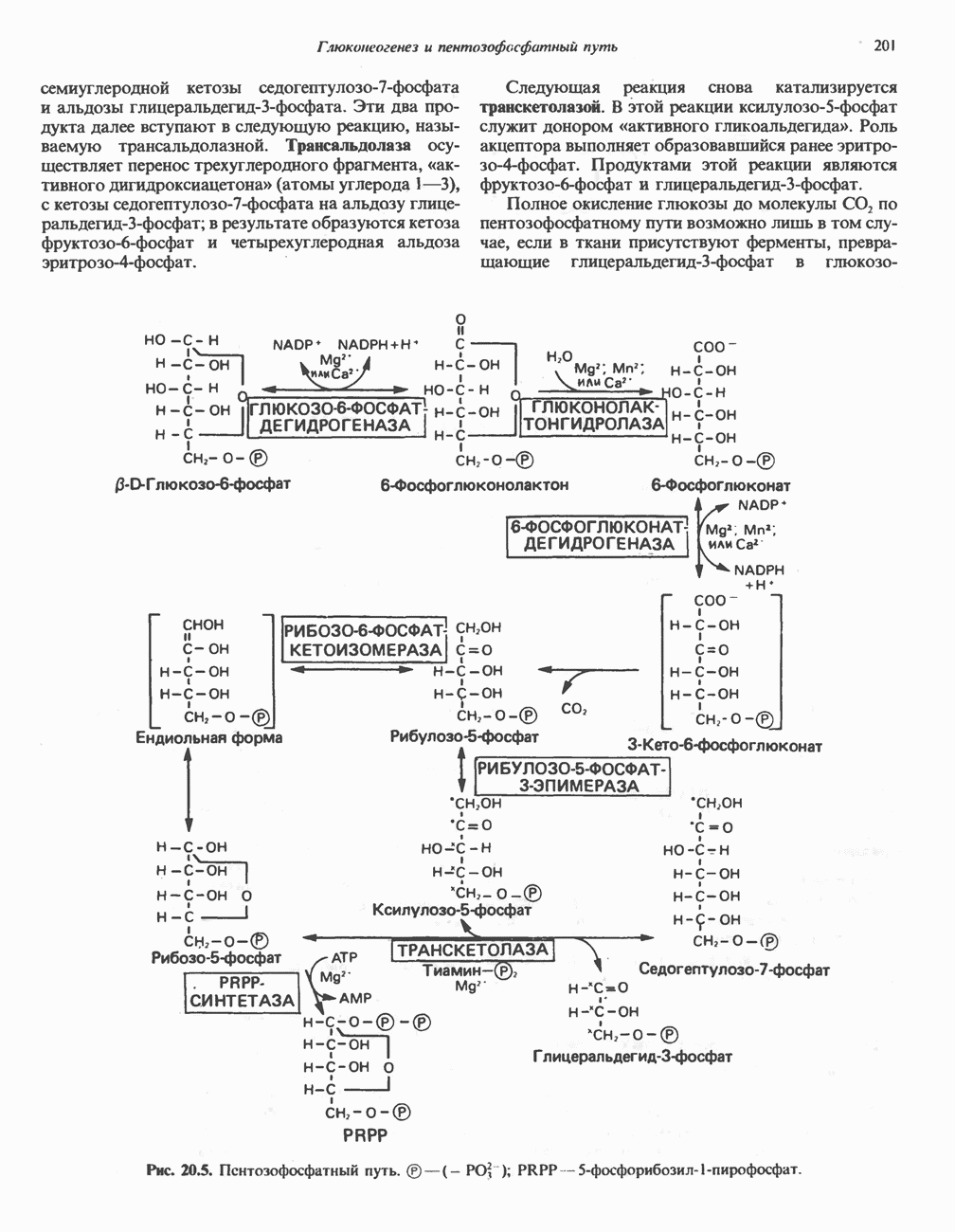
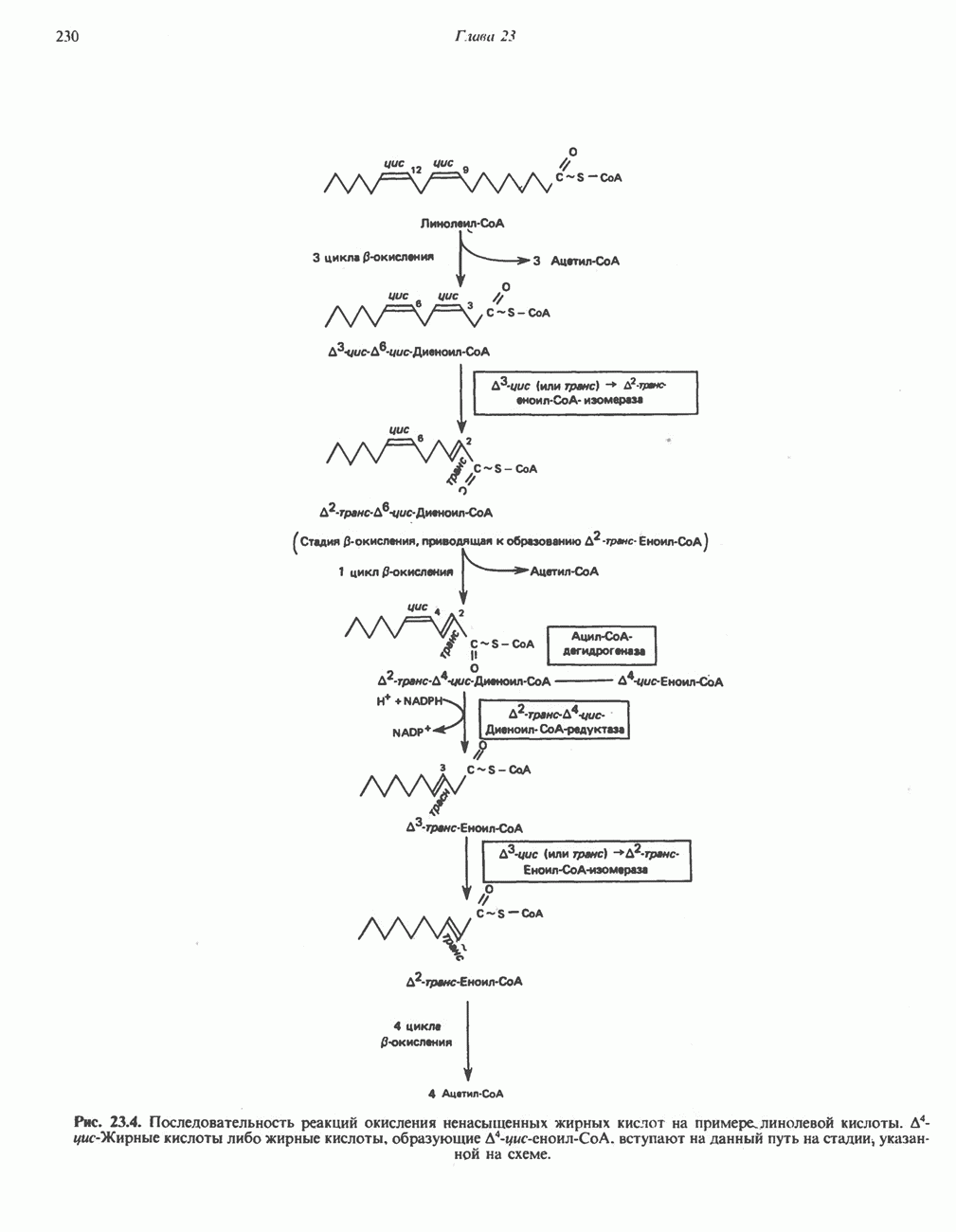
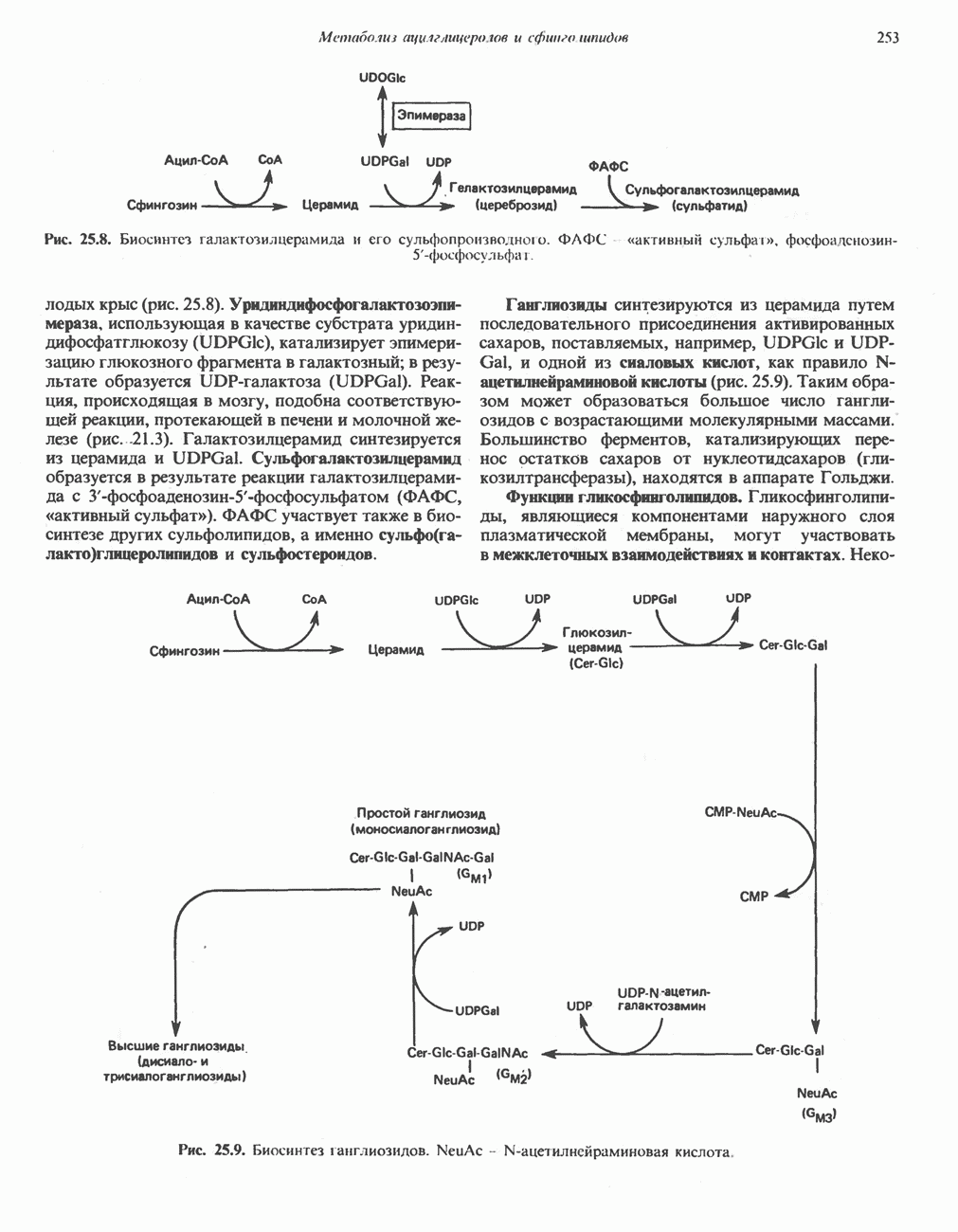
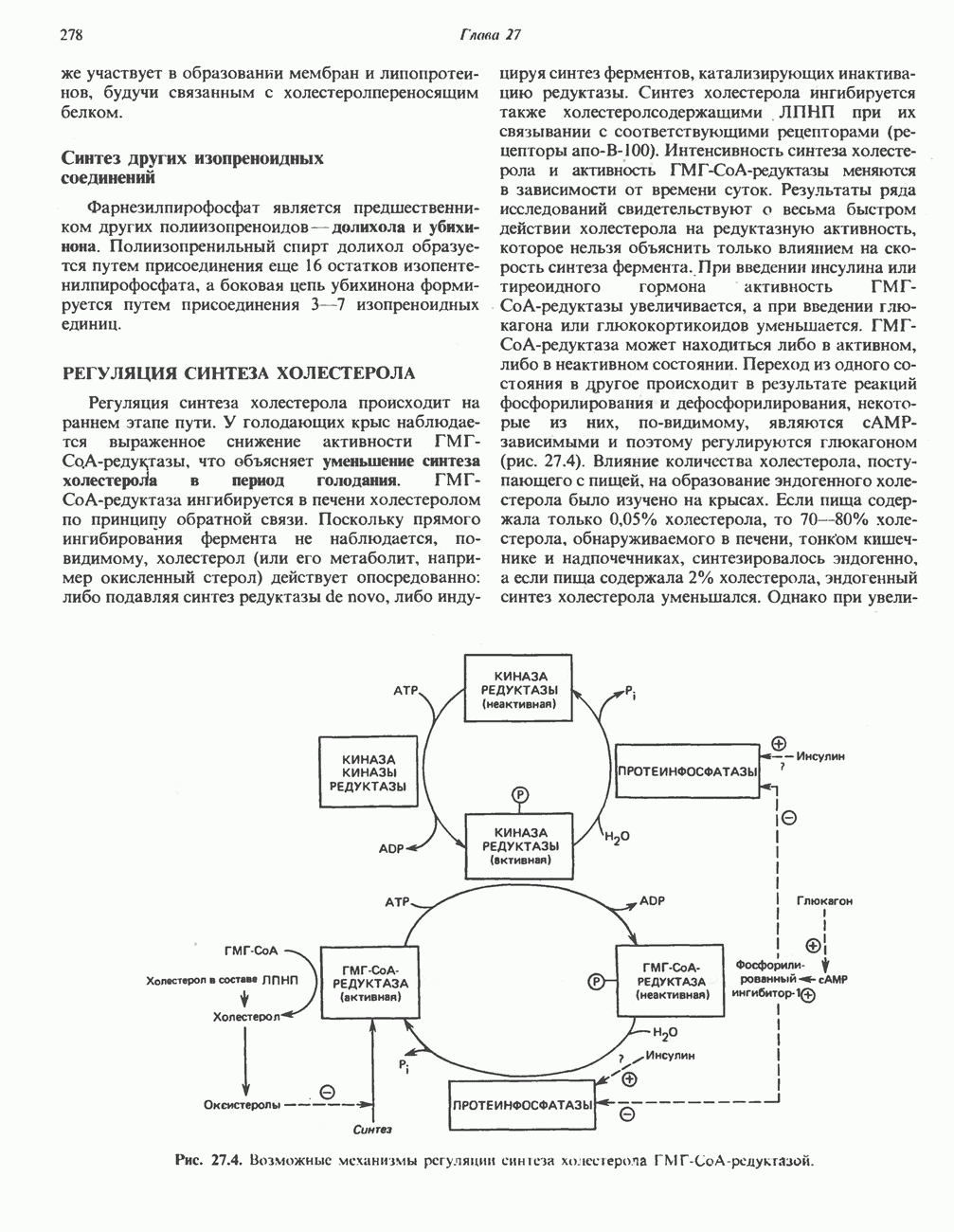
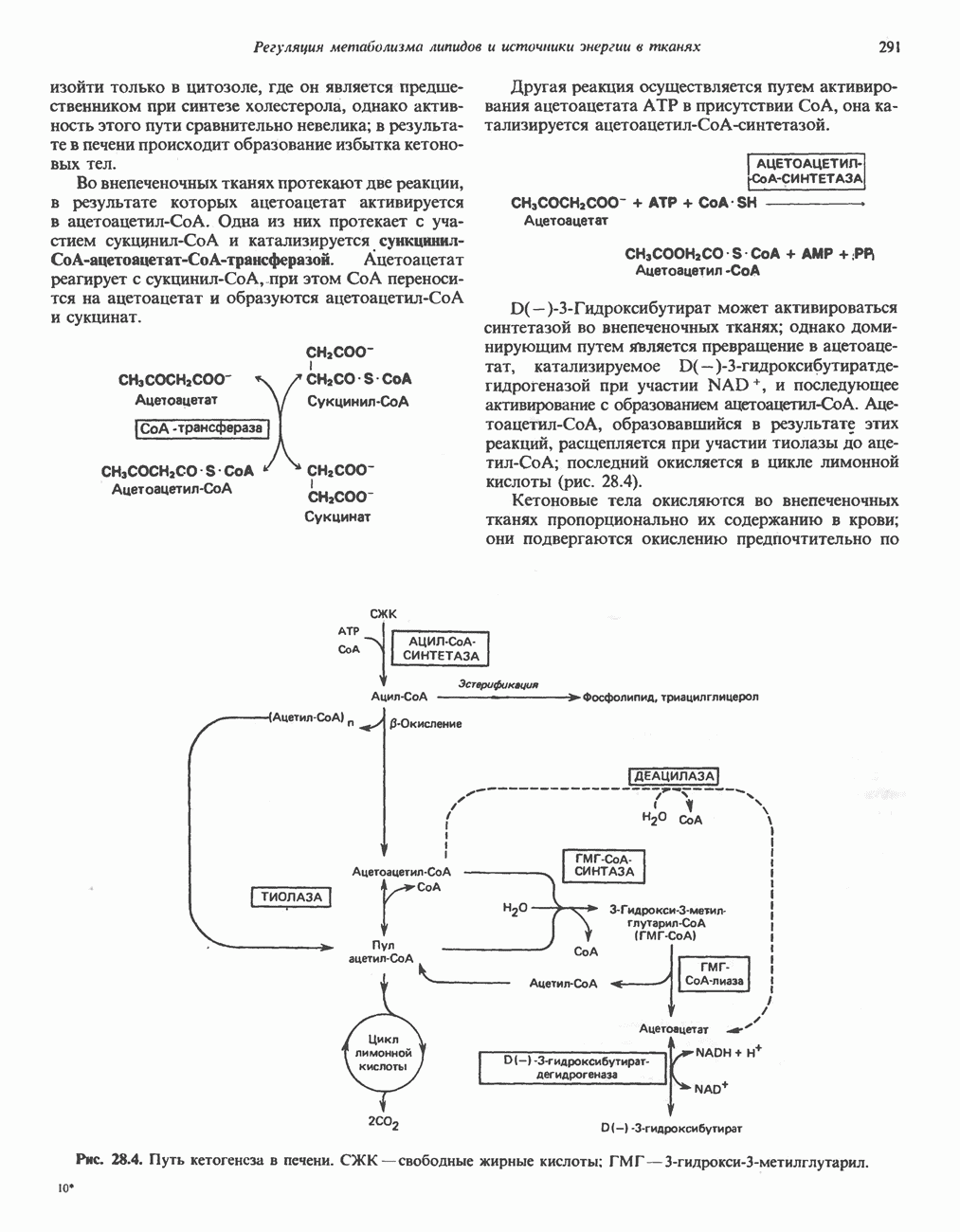
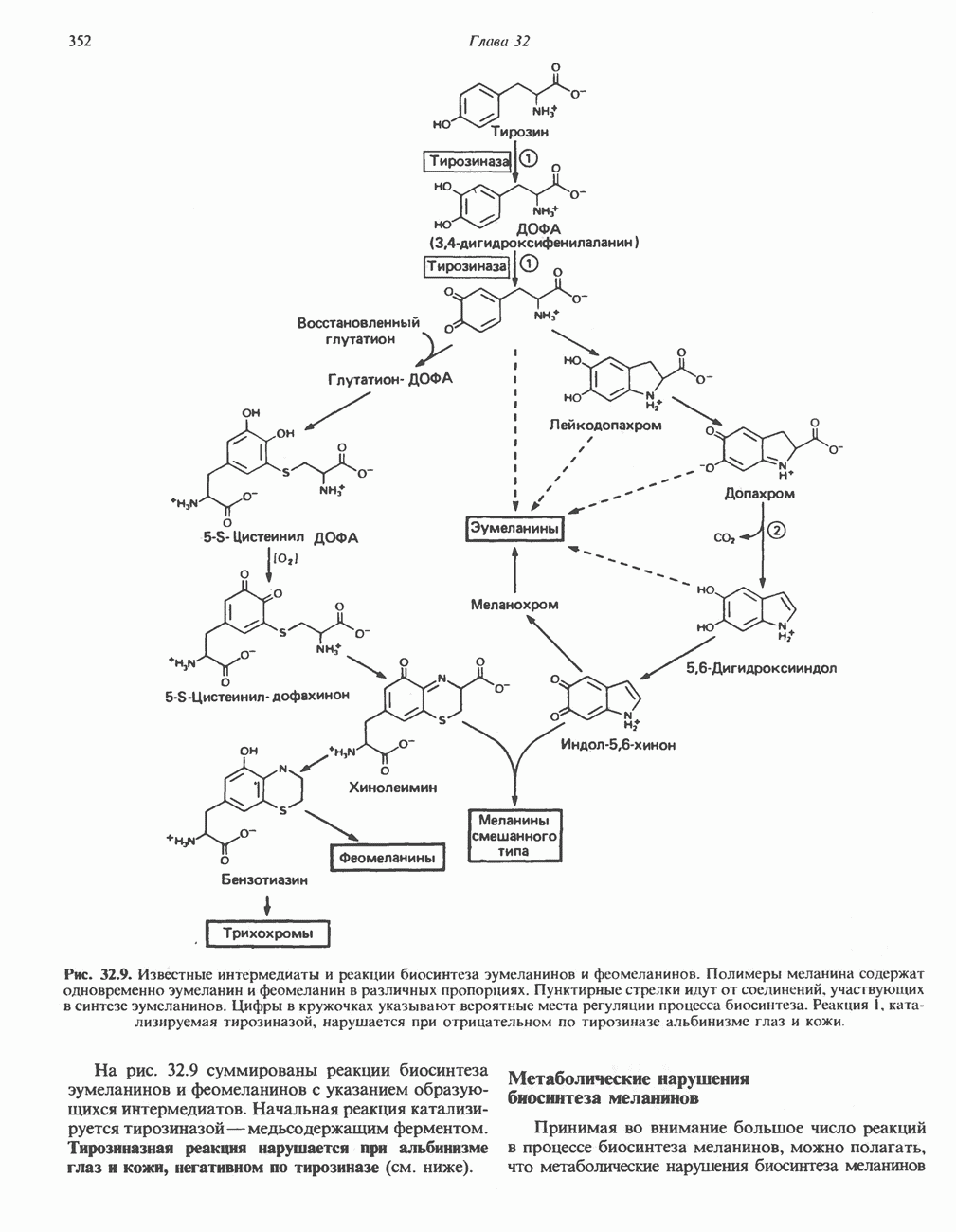
Активация и инактивация фосфорилазы (рис. 19.5)
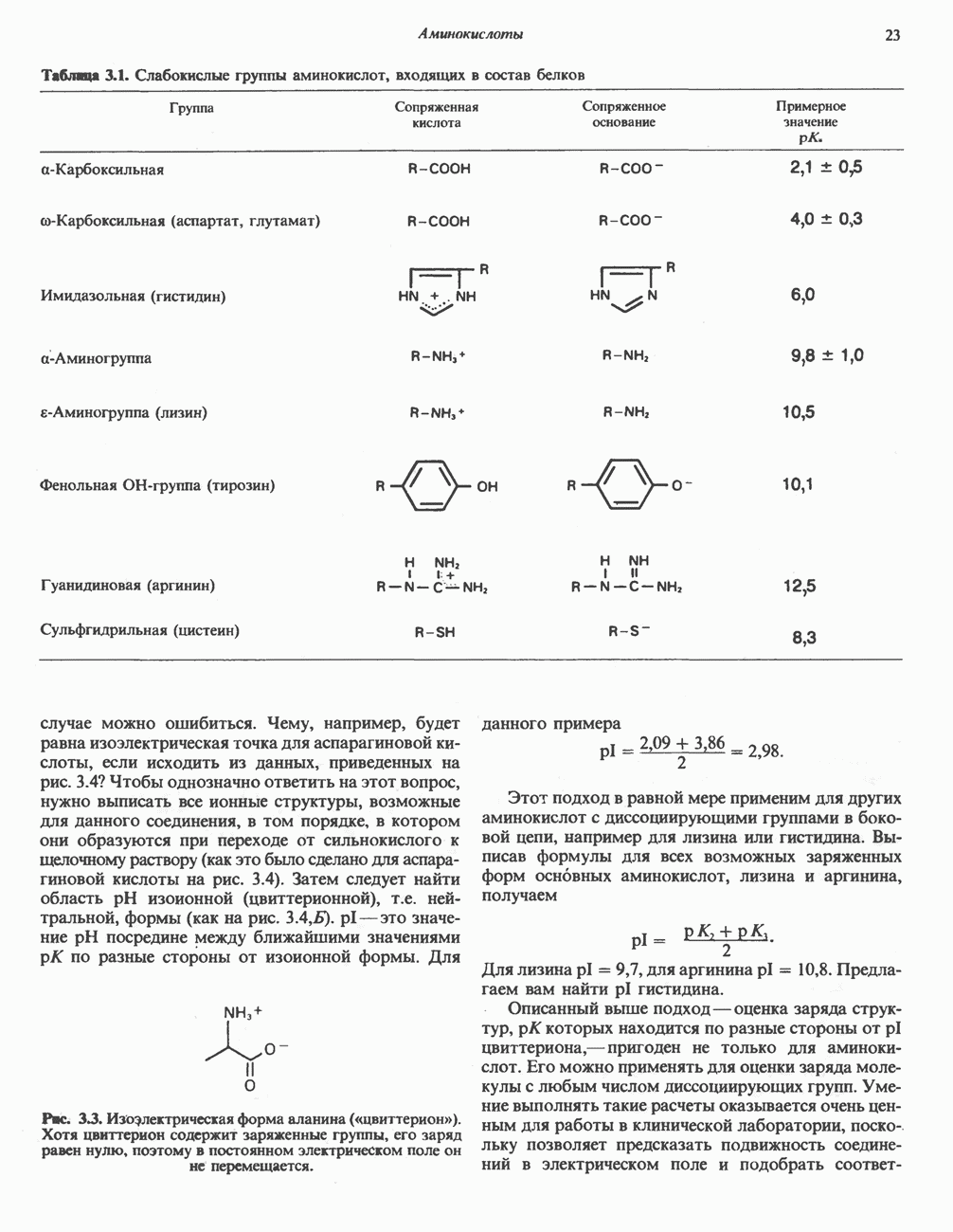
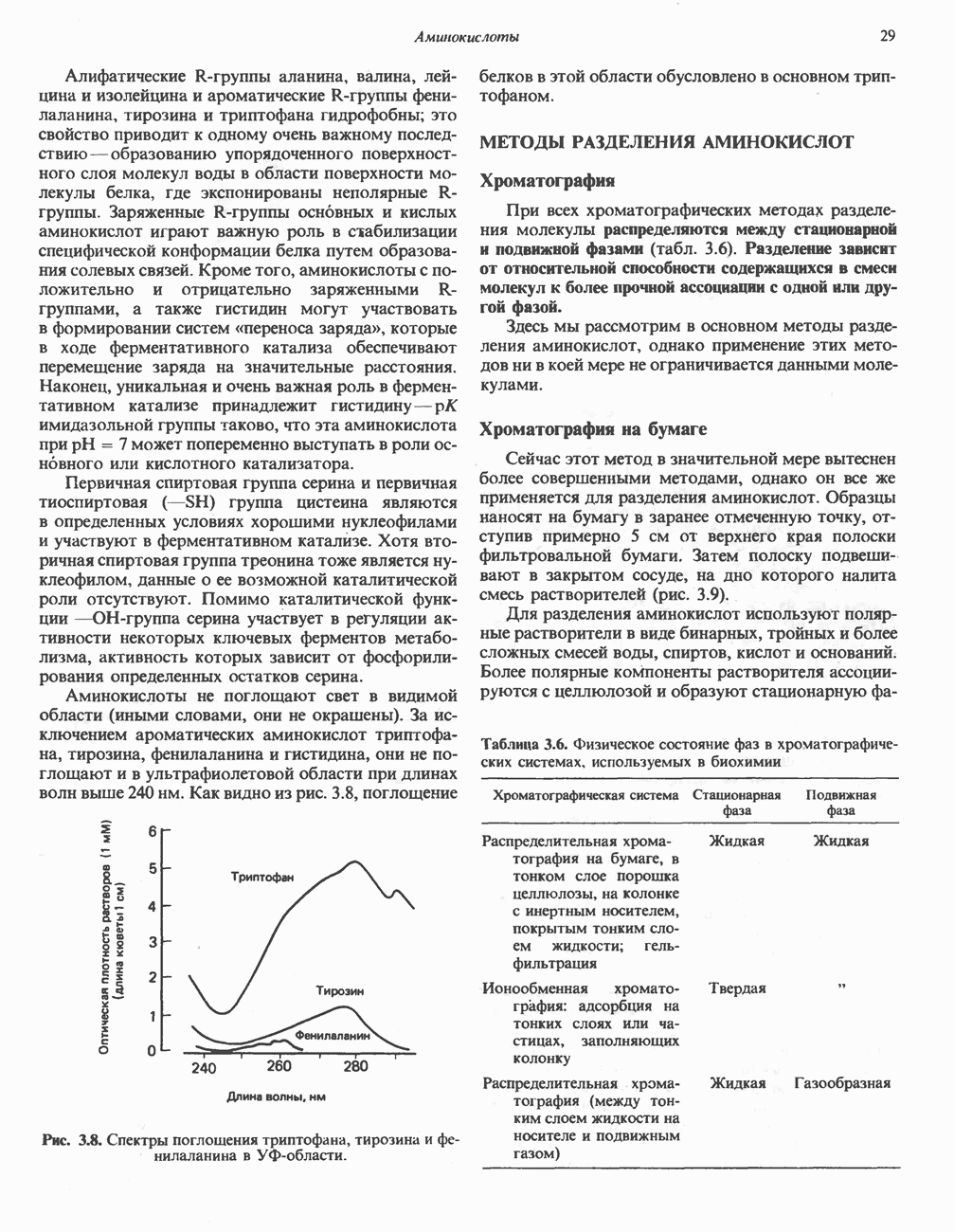
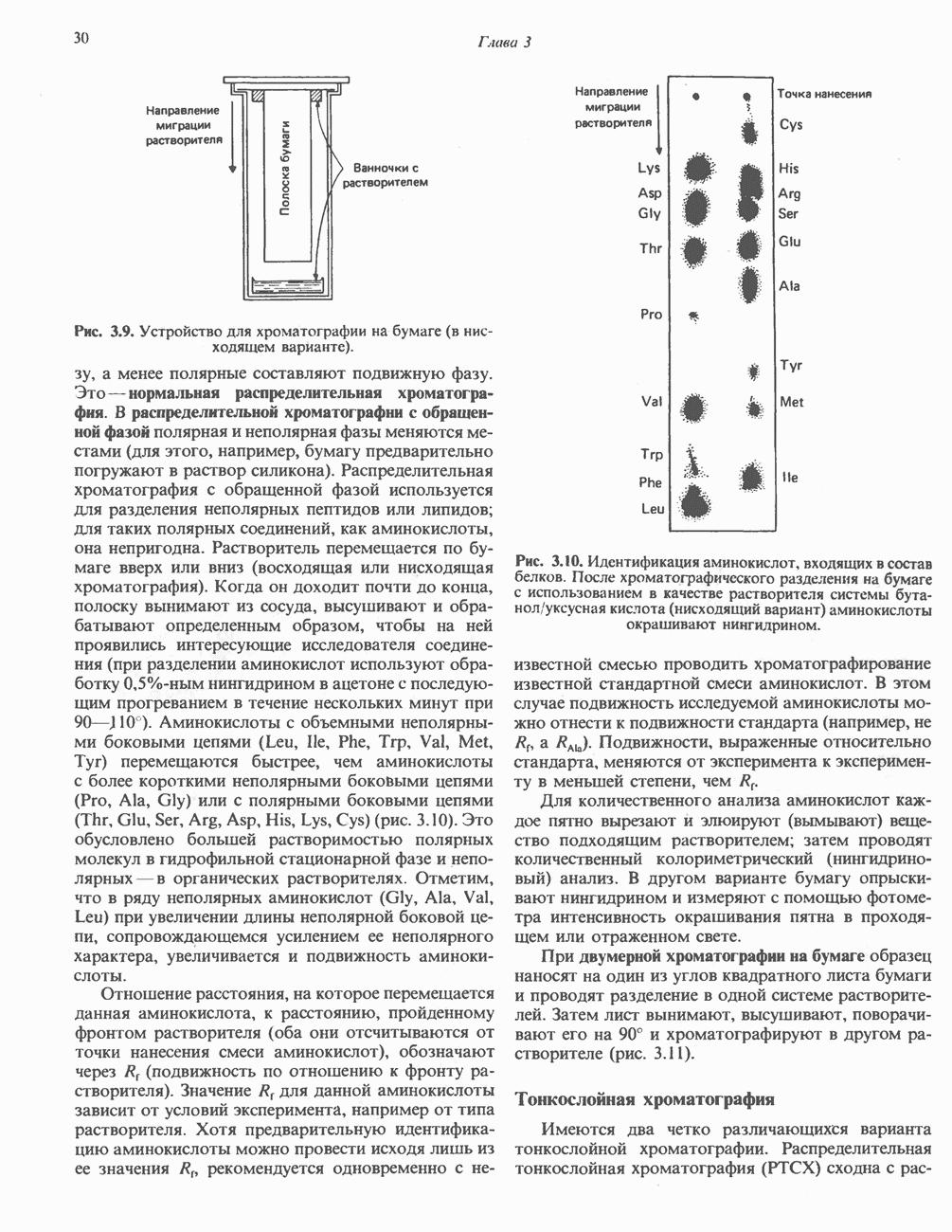
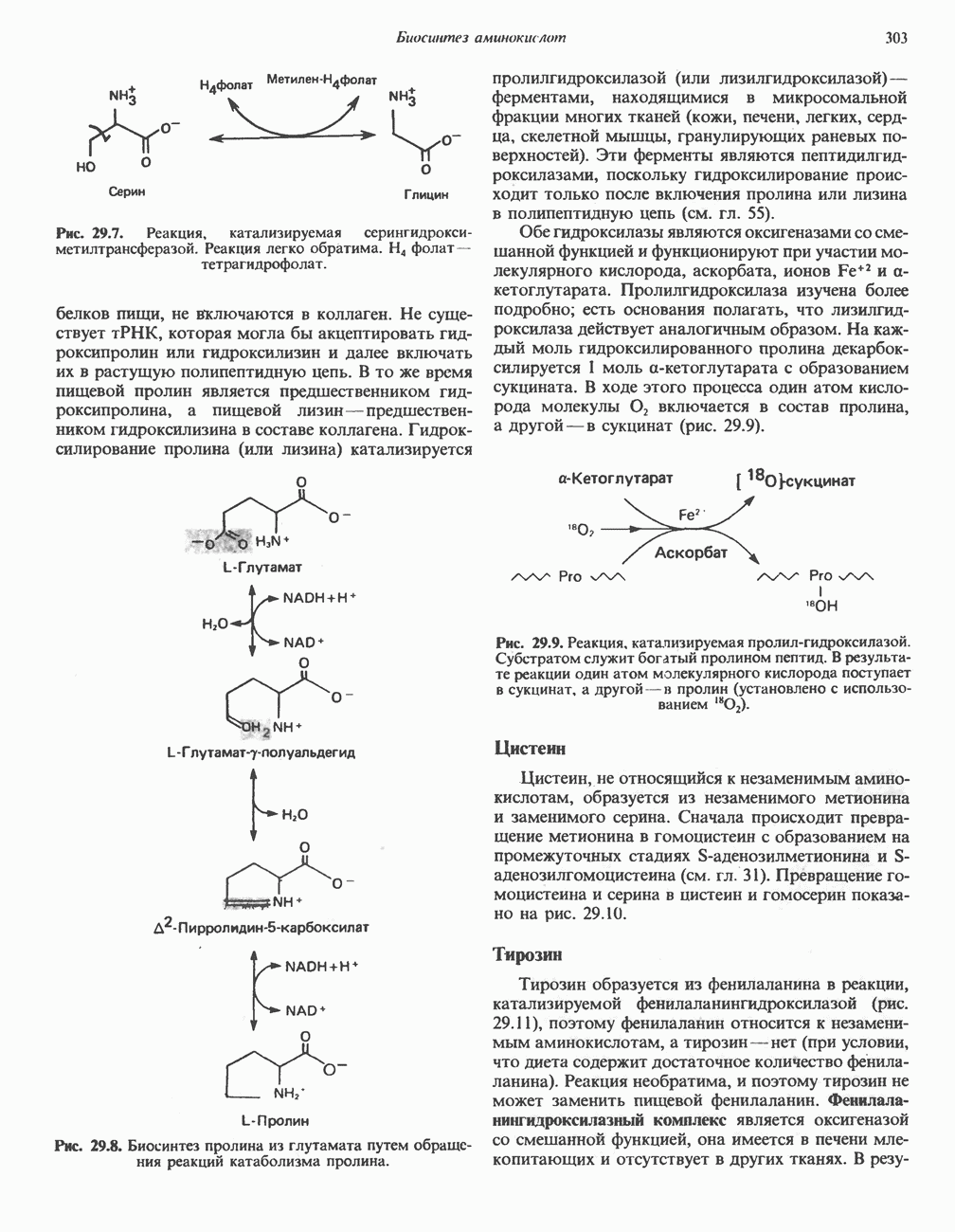
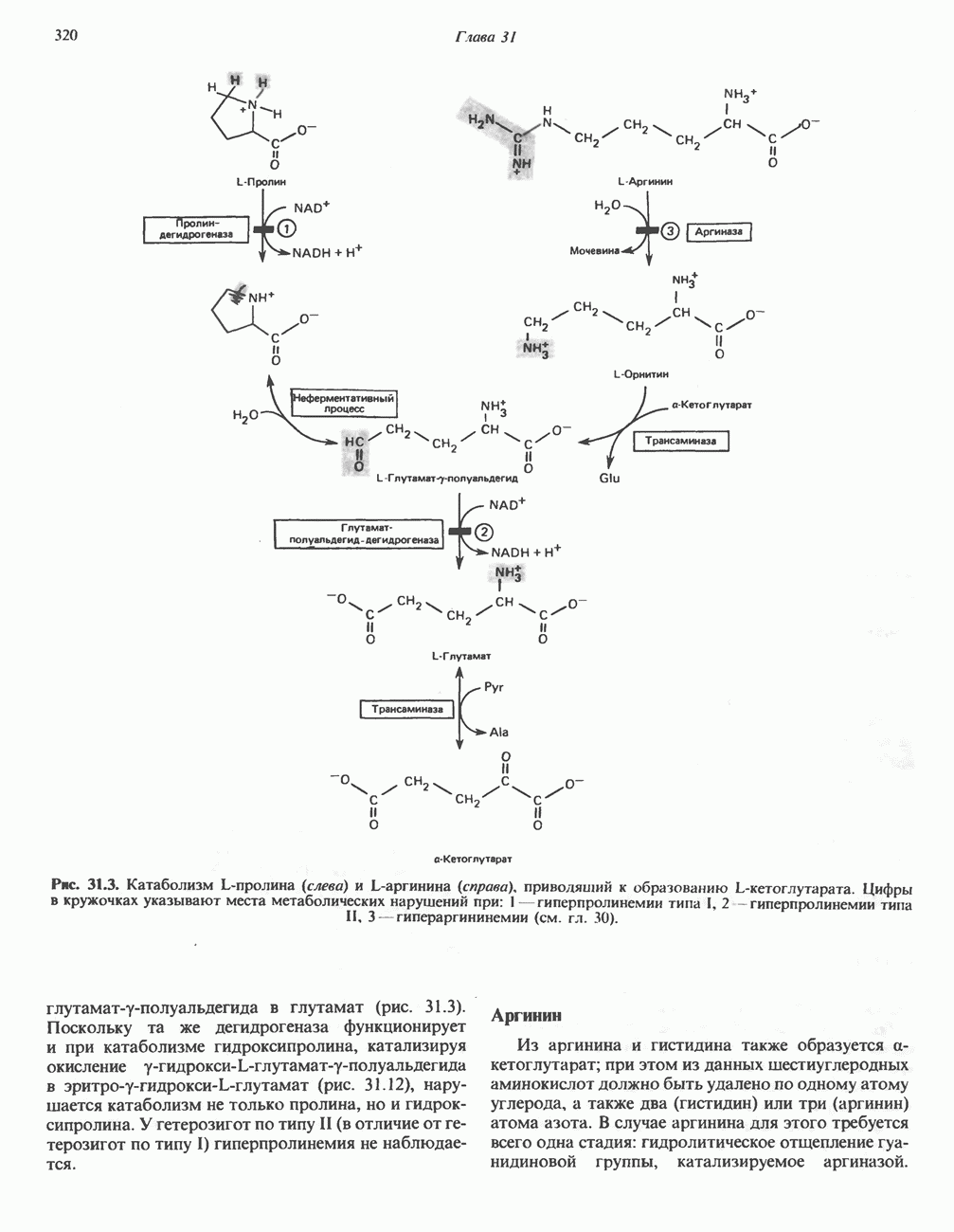
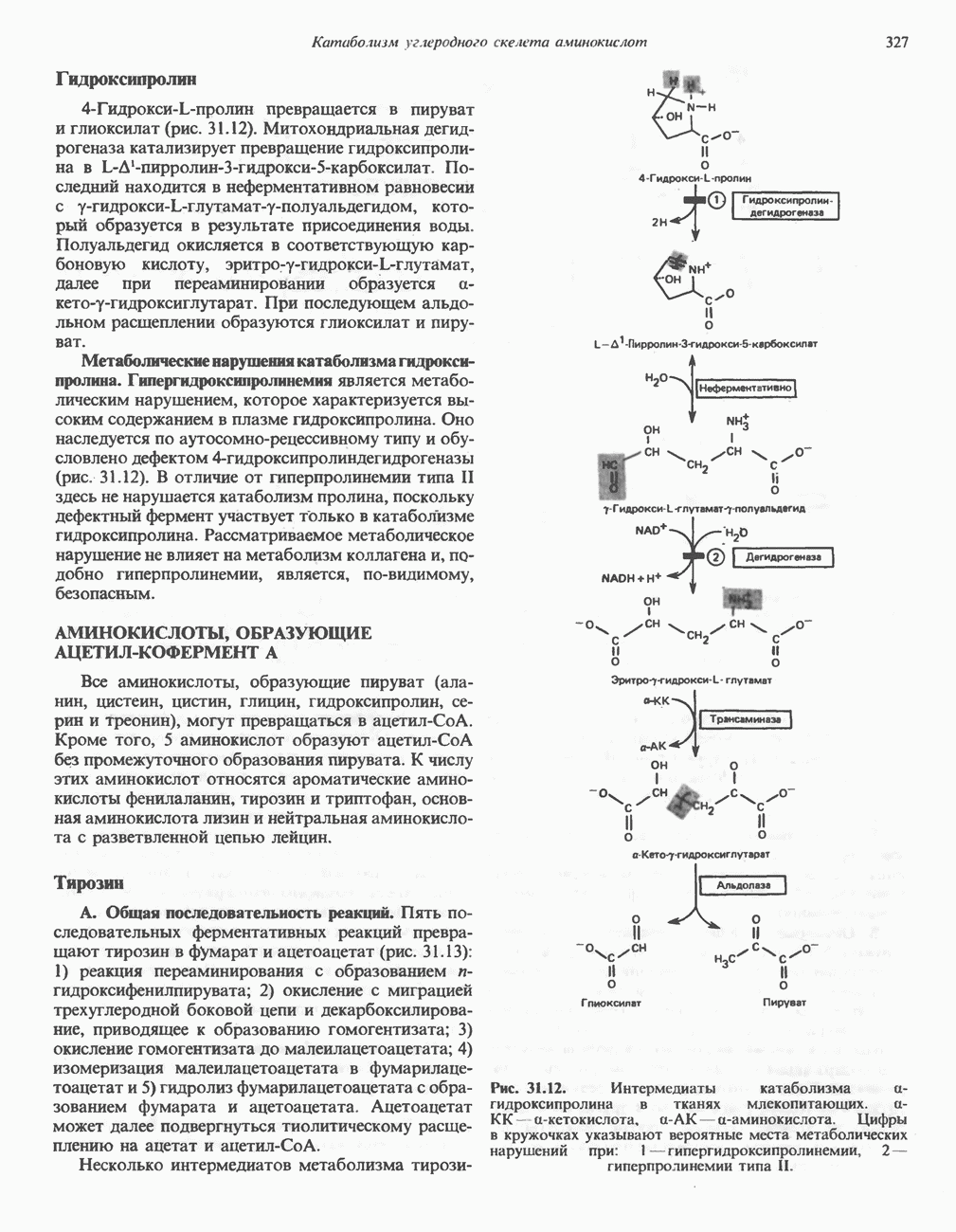
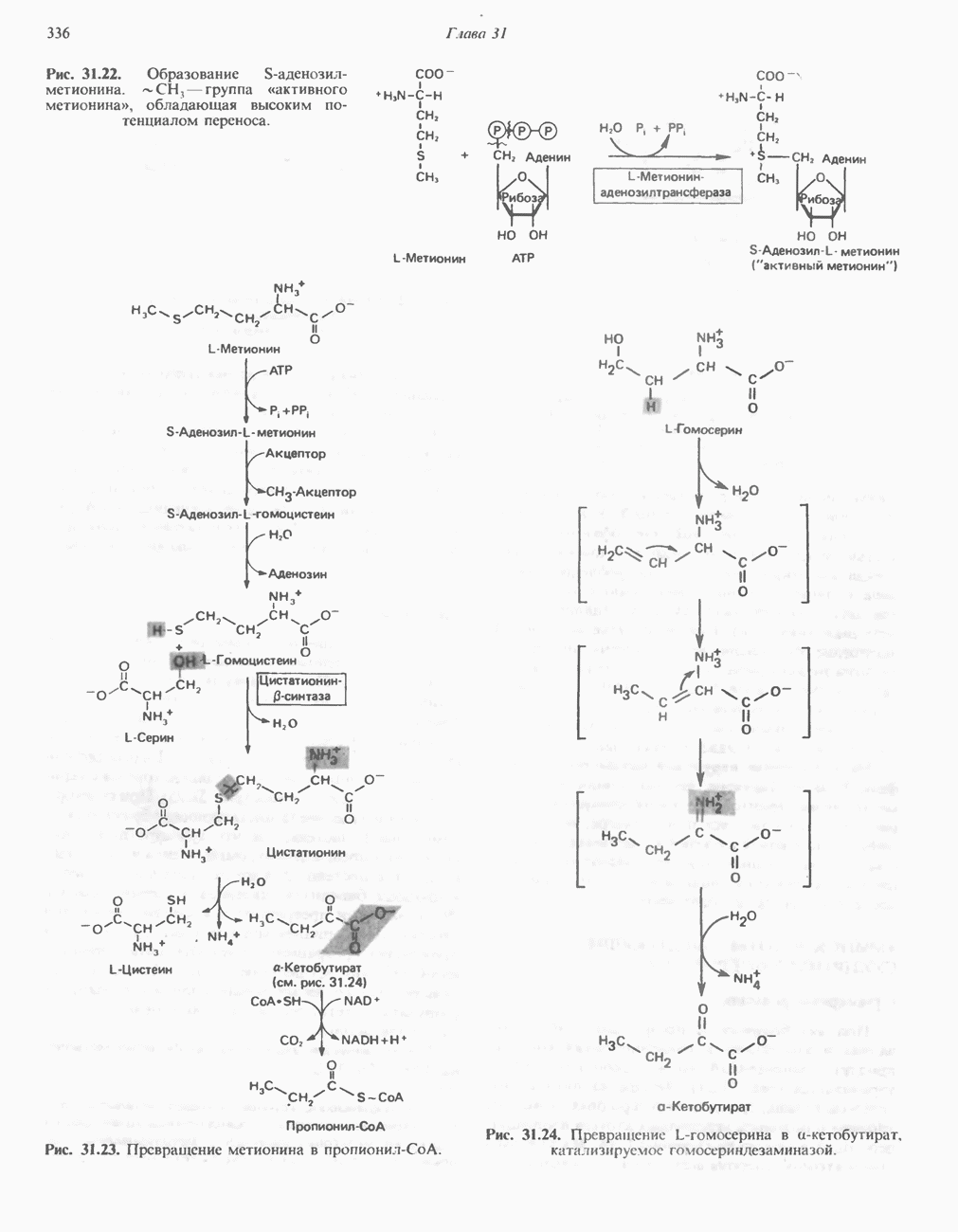
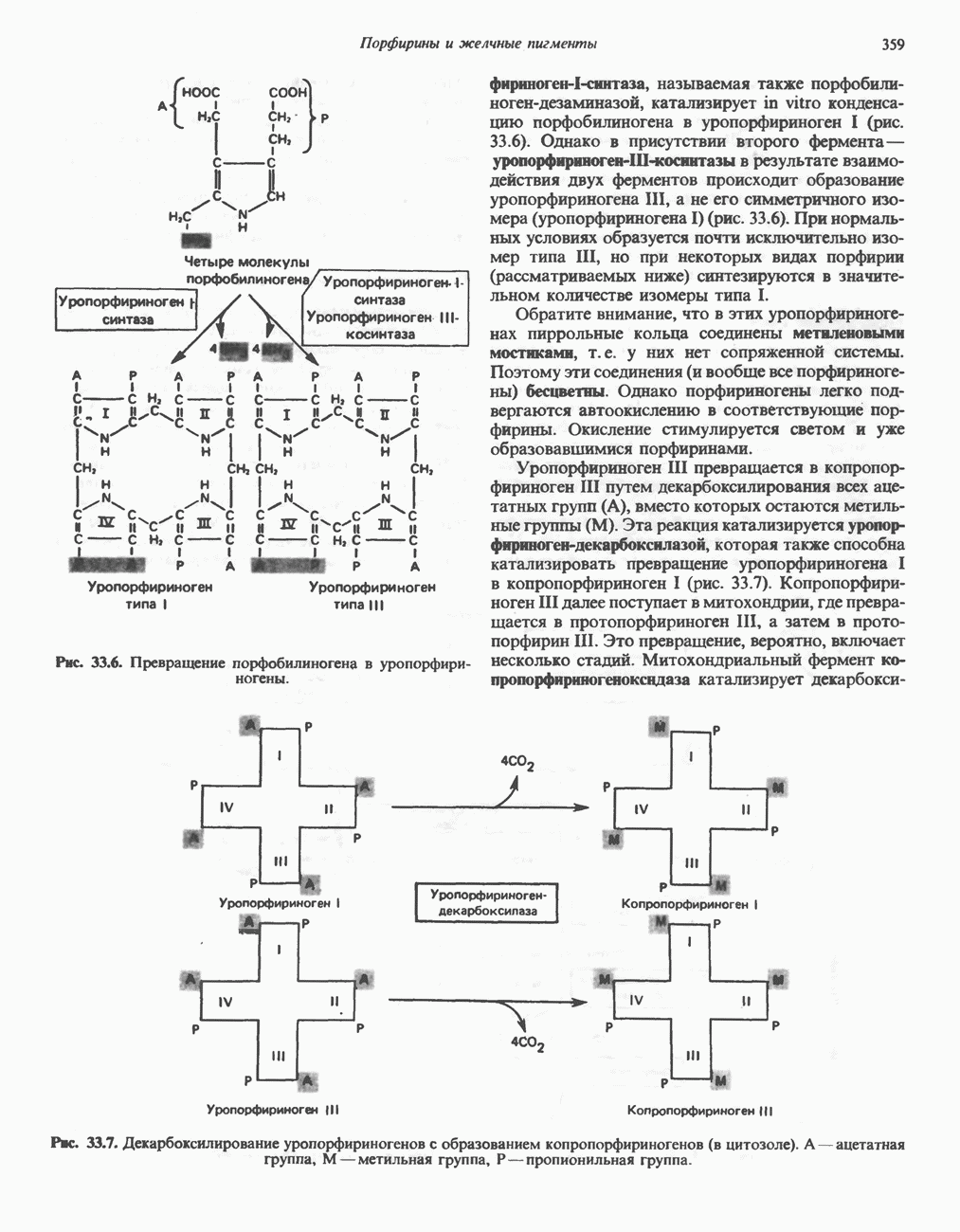
В печени фосфорилаза находится как в активной, так и в неактивной форме. В активной фосфорилазе (фосфорилазе а) гидроксильная группа одного из остатков серина фосфорилирована. Под действием специфической фосфатазы (протеинфосфатазы-1) фермент превращается в неактивную фосфорилазу b в результате гидролитического отщепления фосфата от остатка серина. Реактивация происходит путем рефосфорилирования за счет АТР при действии специфического фермента киназы фосфорилазы.
Мышечная фосфорилаза иммунологически и генетически отличается от соответствующего фермента печени. Она может находиться в двух формах: в форме фосфорилазы а—фосфорилированного фермента, активного как в присутствии, так и в отсутствие АМР (ее аллостерического модулятора), и в форме фосфорилазы b, дефосфорилированной и активной только в присутствии АМР. Фосфорилаза а является нормальной физиологически активной формой фермента. Она представляет собой димер, каждый мономер которого содержит одну молекулу пиридоксальфосфата.
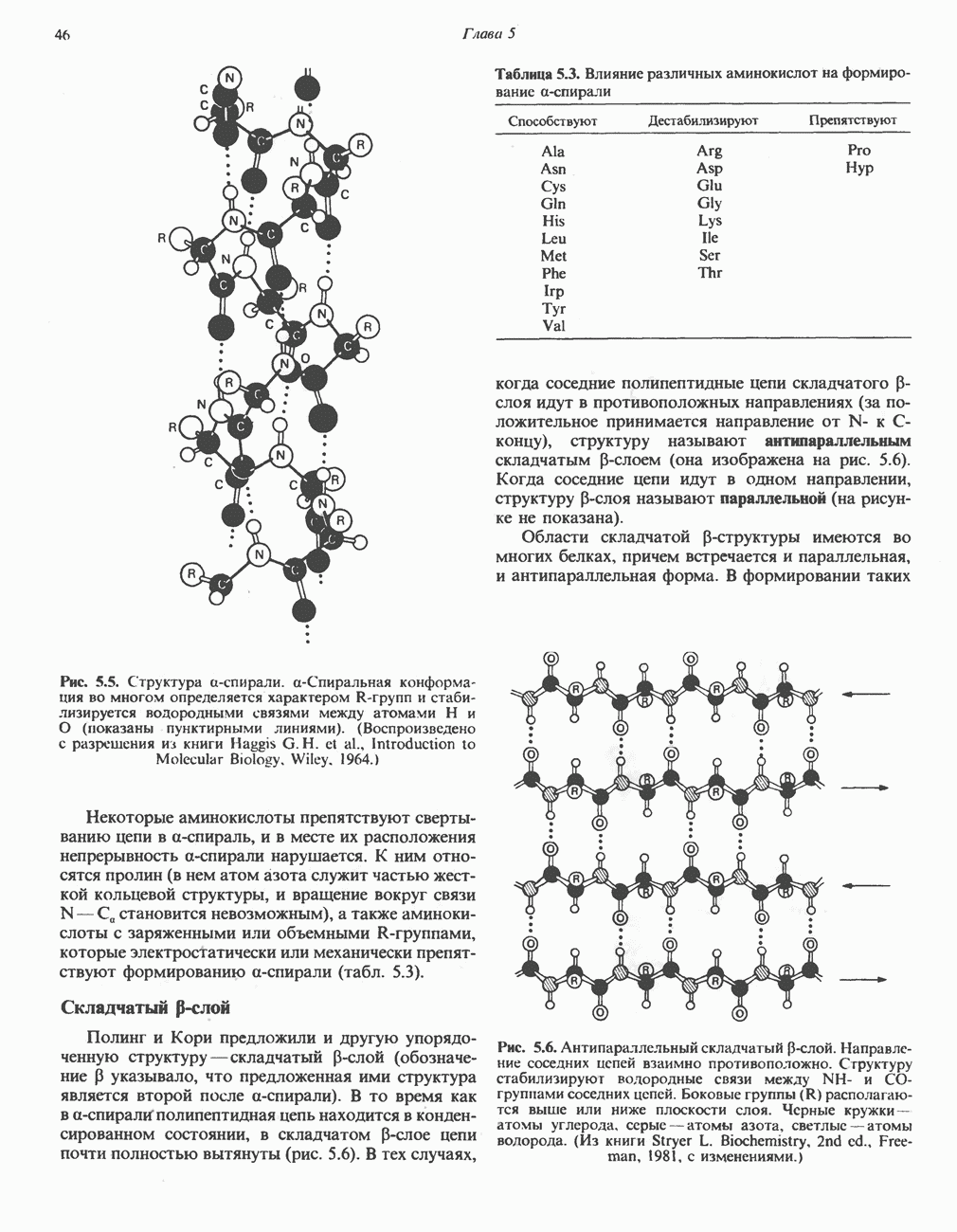
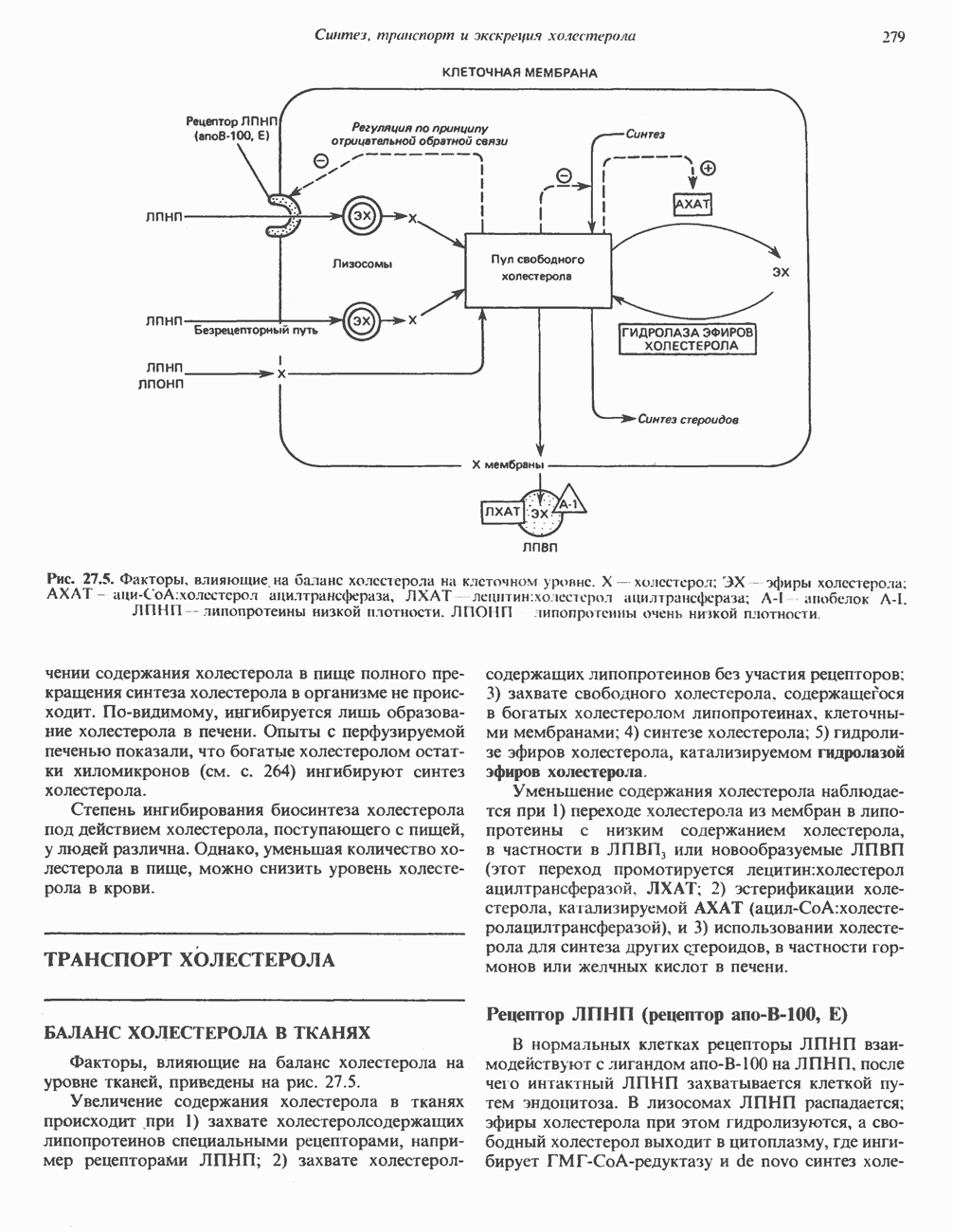
Активация с участием сАМР
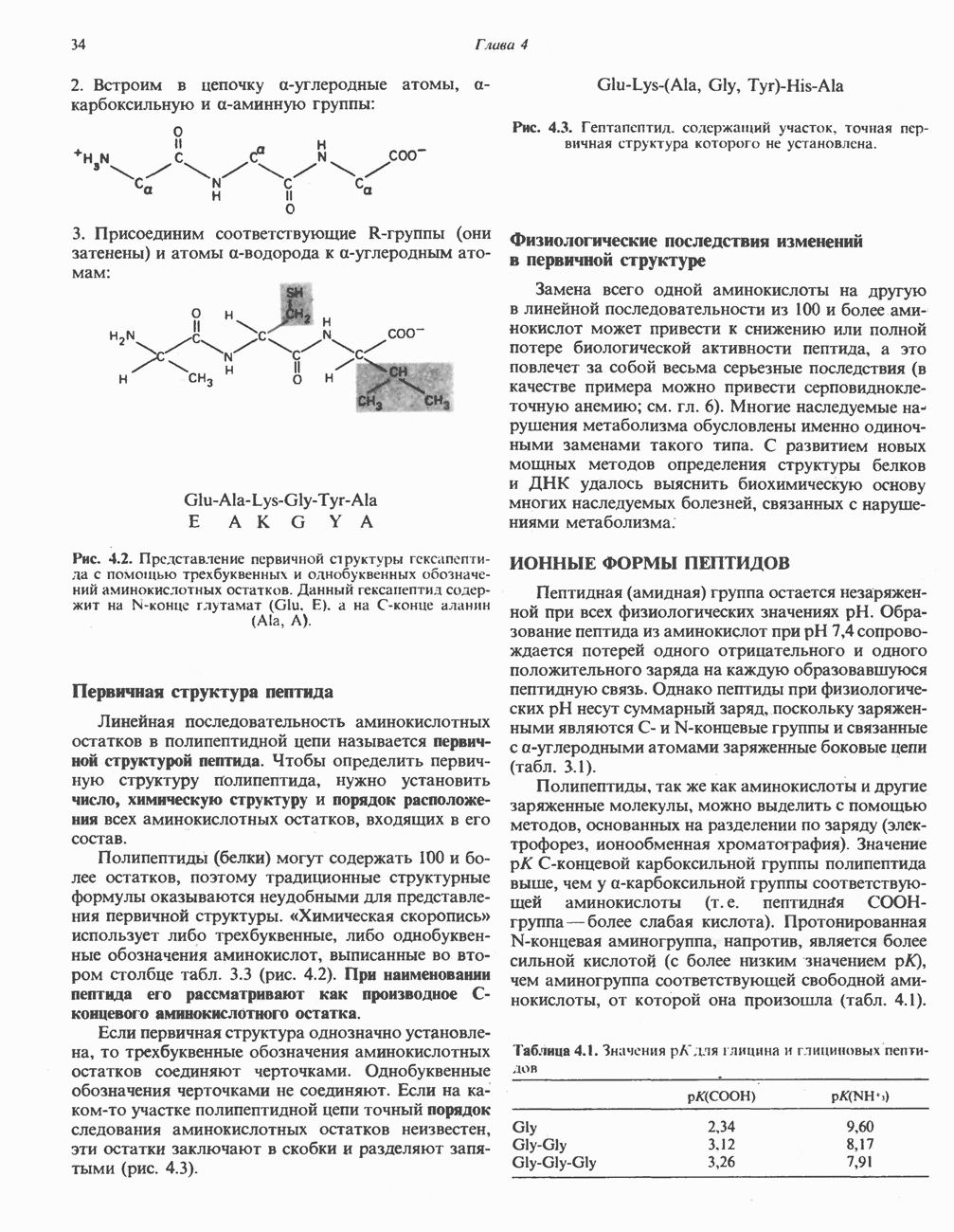
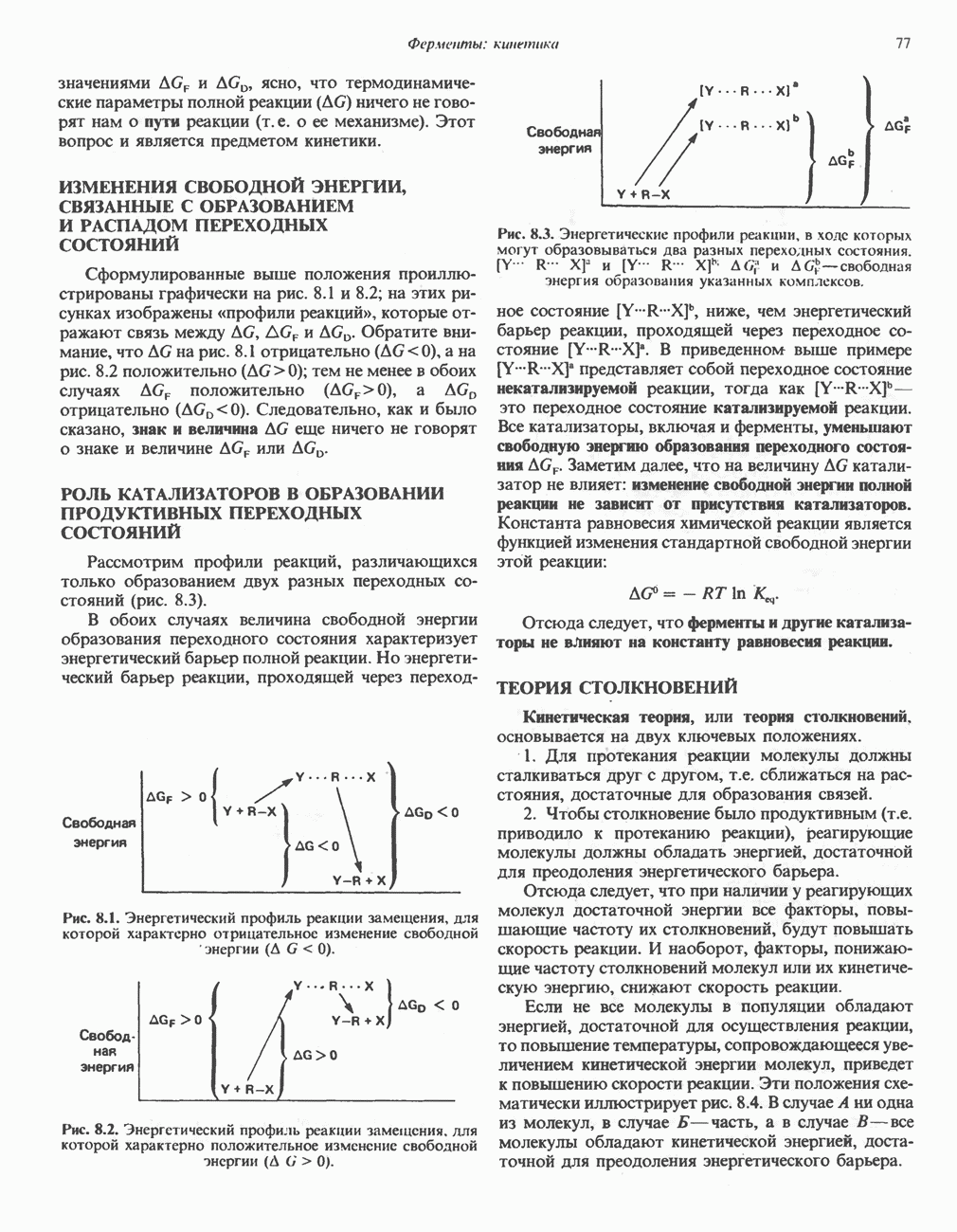
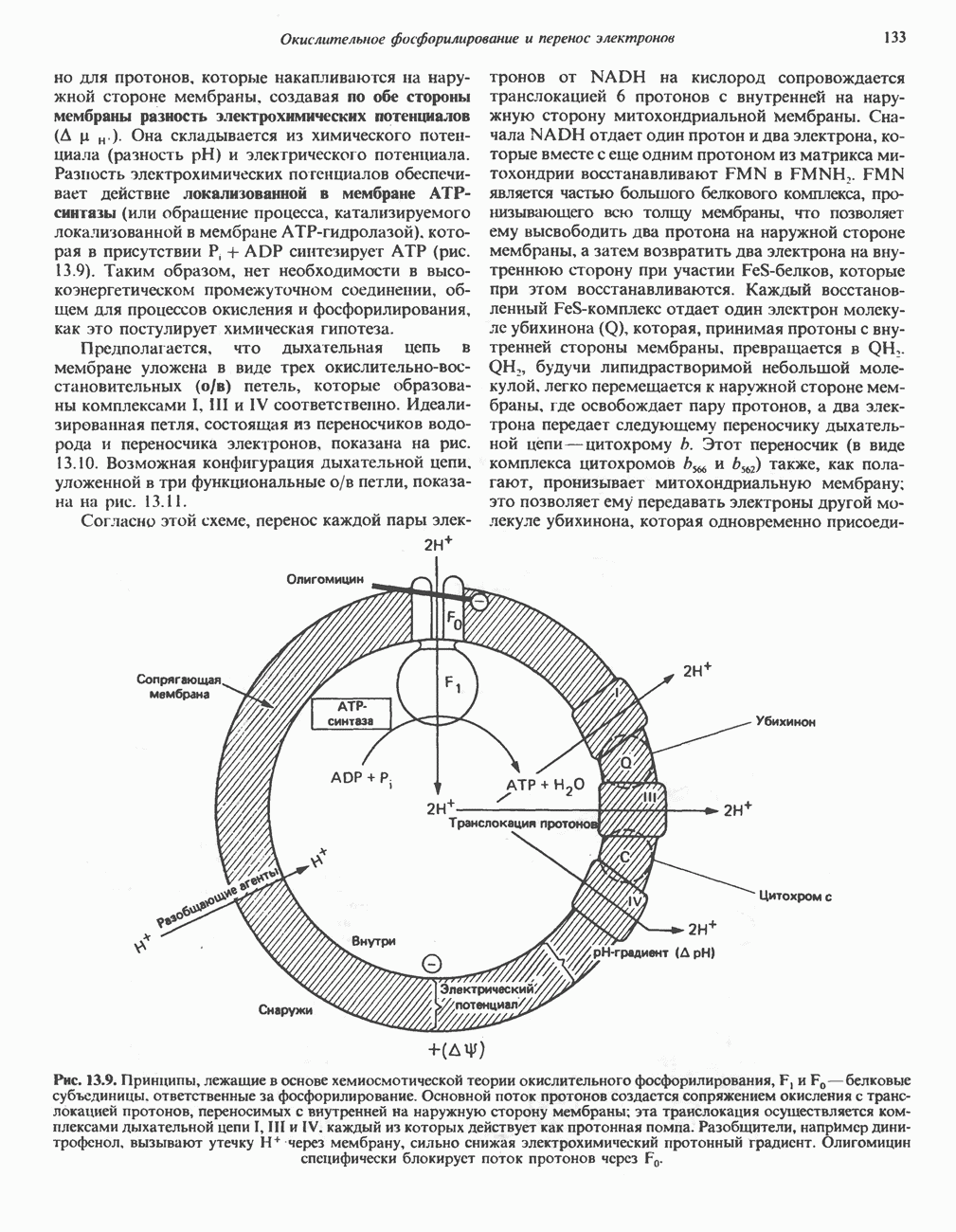
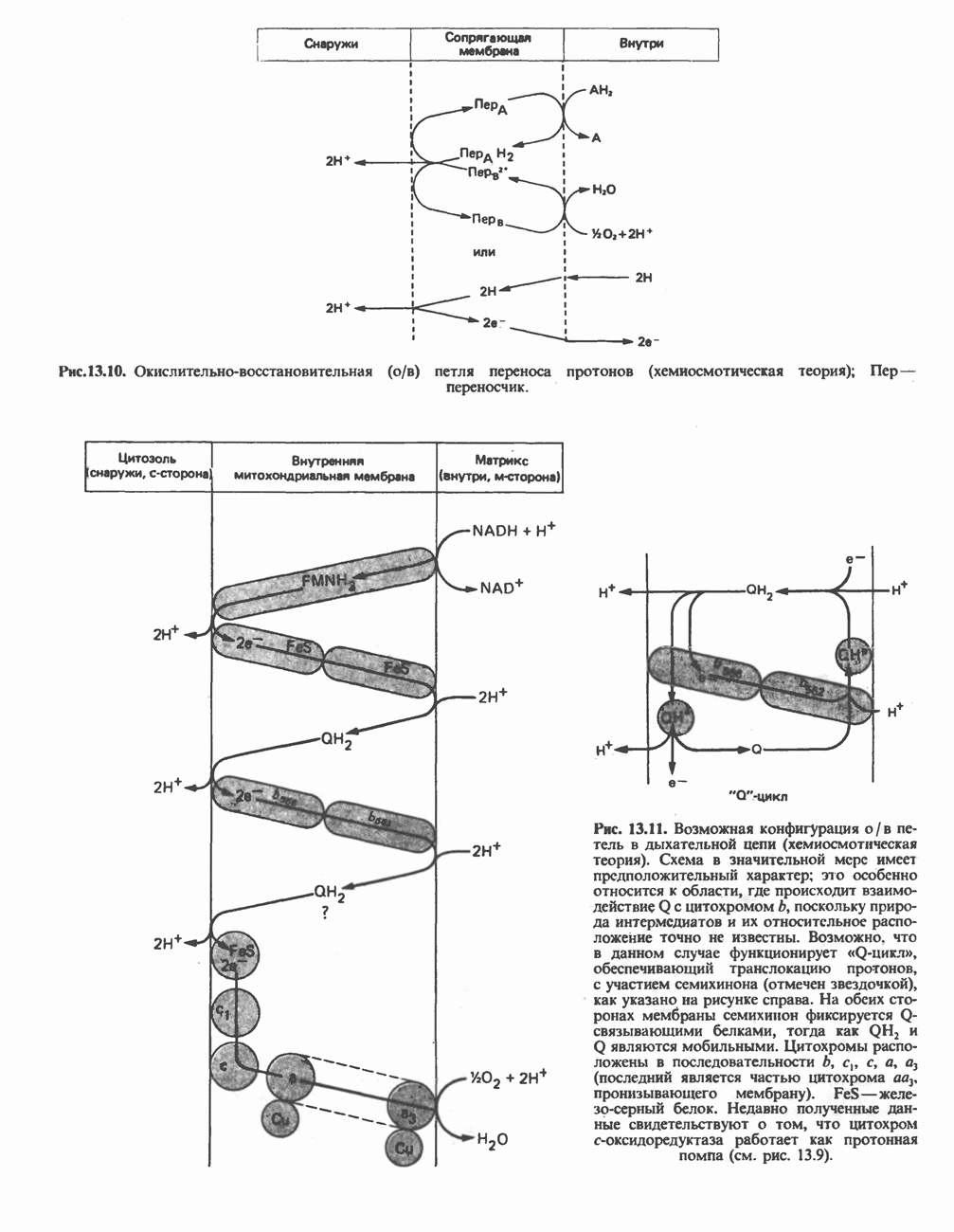
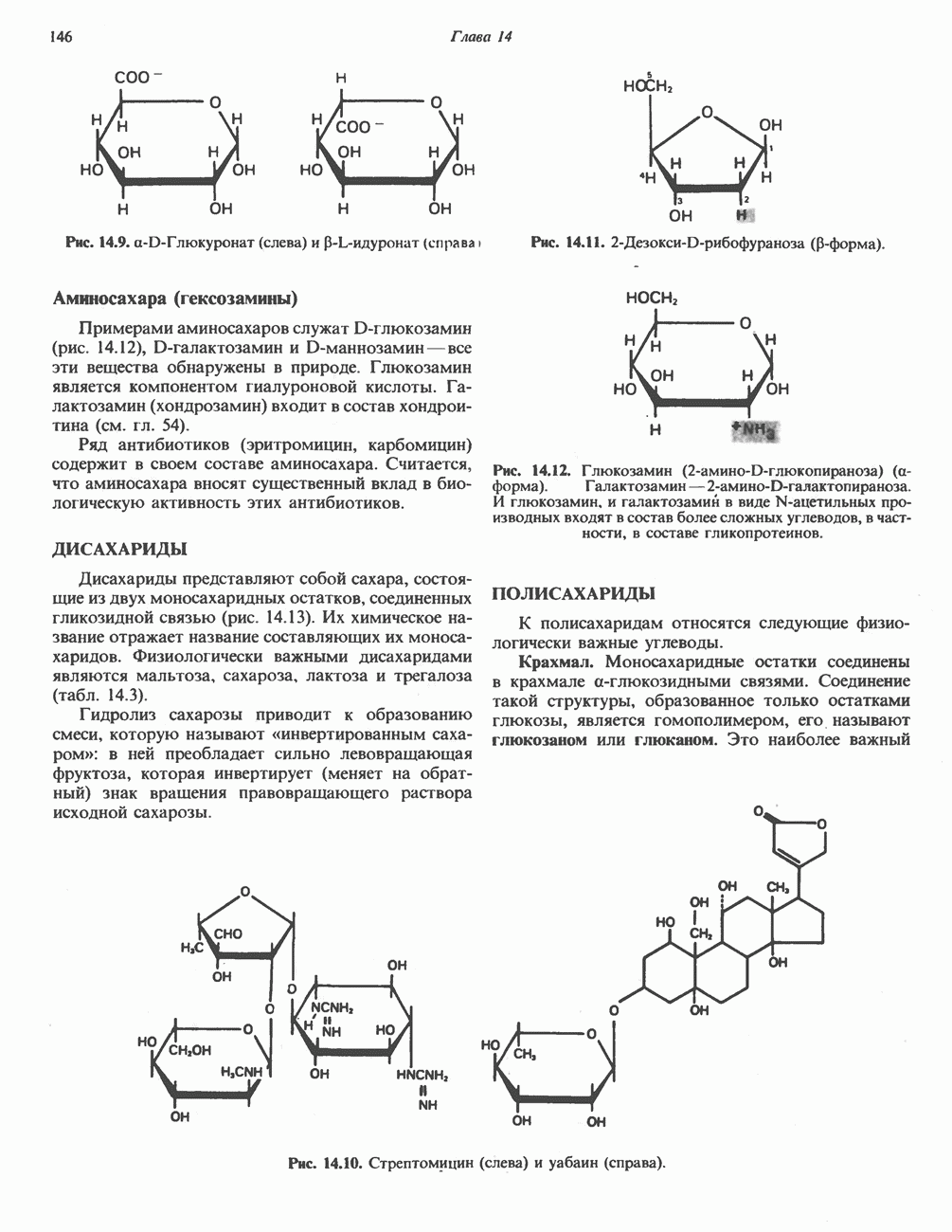
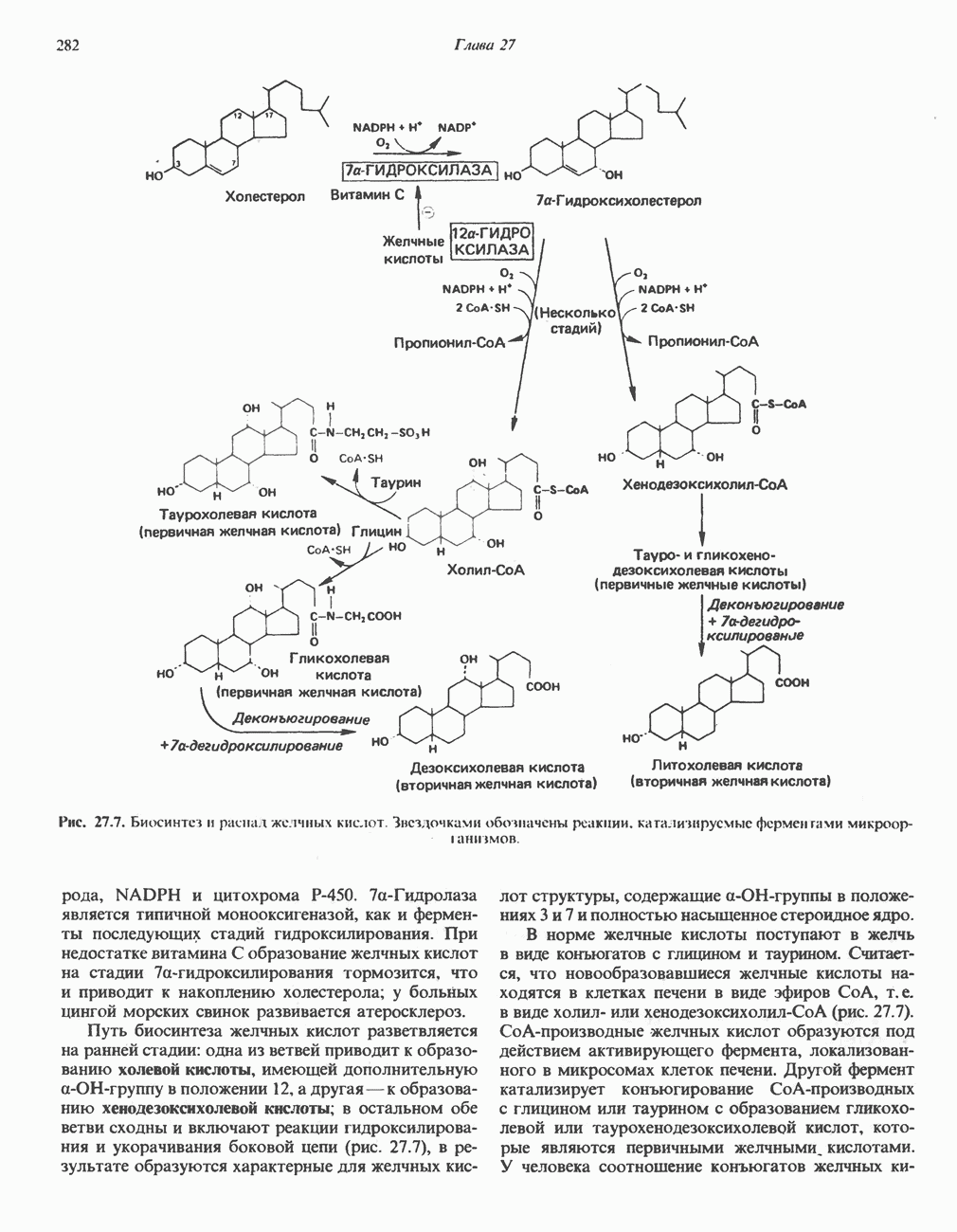
В мышце фосфорилаза активируется адреналином (рис. 19.5). Однако он оказывает не прямой эффект, а действует опосредованно через сАМР (3,5-циклоадениловую кислоту; циклический АМР) (рис. 19.6 и гл. 44). сАМР представляет собой внутриклеточный интермедиат, выступающий в роли второго посредника при действии ряда гормонов. Он образуется из АТР при действии фермента аденилат-циклазы, находящейся на внутренней поверхности клеточной мембраны. Аденилатциклаза активируется (опосредованно) гормонами адреналином и норадреналином—лигандами Р-адренергических рецепторов, локализованных в клеточной мембране; в печени она активируется глюкагоном, действующим

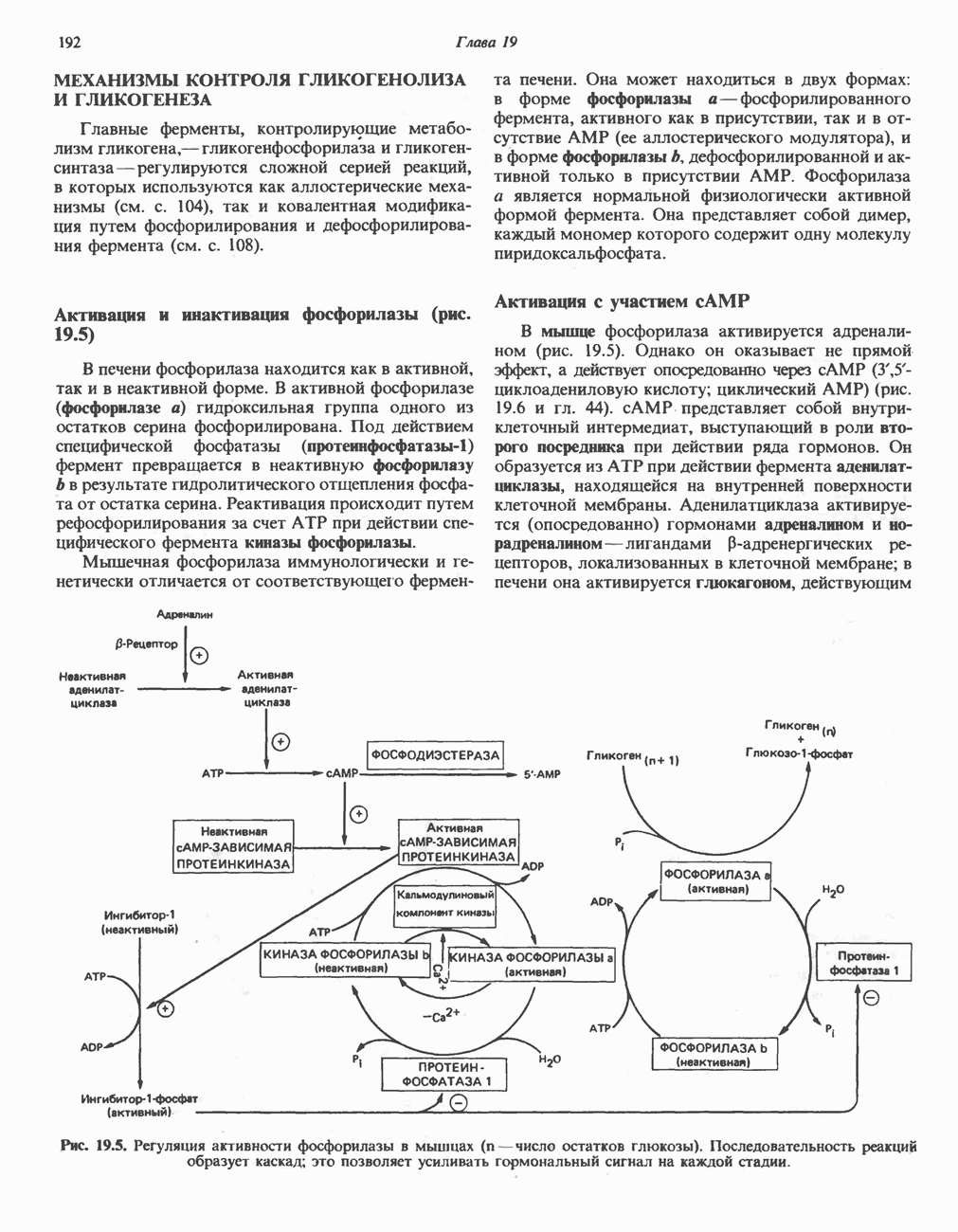
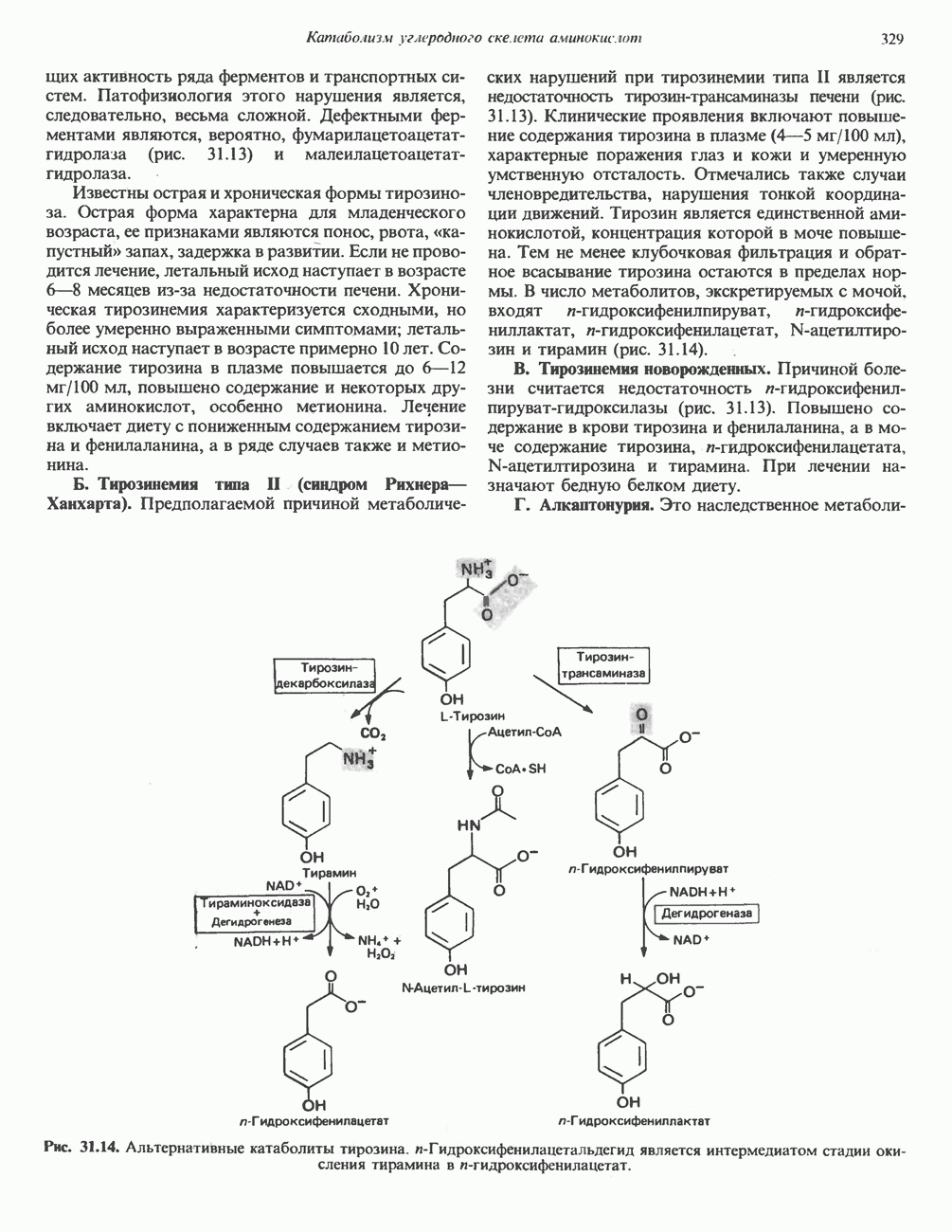
Рис. 19.5. Регуляция активности фосфорилазы в мышцах (п—число остатков глюкозы). Последовательность реакций образует каскад; это позволяет усиливать гормональный сигнал на каждой стадии.

Рис. 19.6. 3,5-Адениловая кислота (циклический АМР. сАМР).
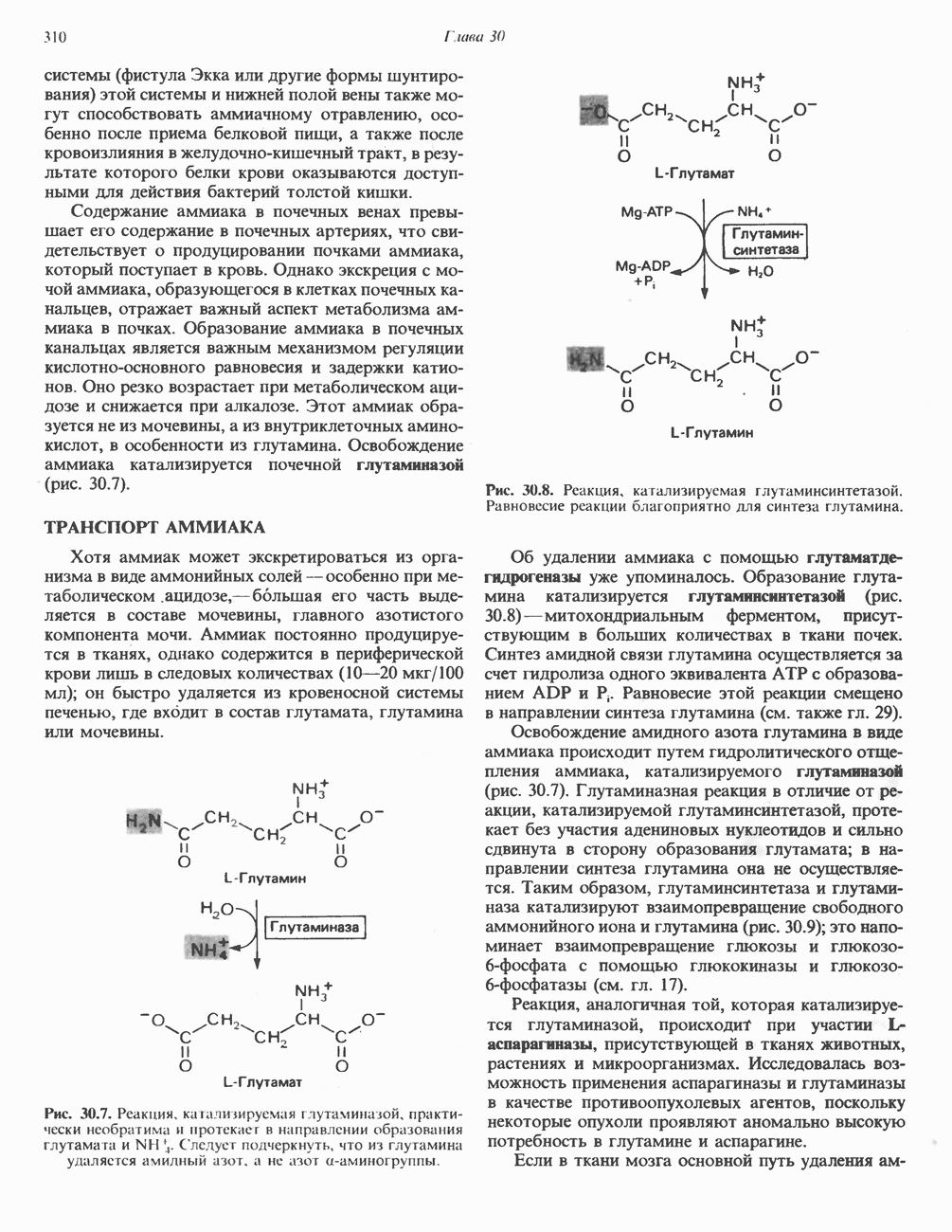
при участии специального глюкагонового рецептора. сАМР разрушается при действии фосфодиэстеразы; именно этот фермент поддерживает в норме концентрацию сАМР на низком уровне. Имеются данные о том, что инсулин повышает активность фосфодиэстеразы в печени; это приводит к понижению концентрации сАМР.
Повышение концентрации сАМР активирует фермент, обладающий весьма широкой специфичностью — сАМР-зависимую протеинкиназу. Эта киназа катализирует фосфорилирование при участии АТР неактивной киназы фосфорилазы с образованием активной киназы фосфорилазы, которая в свою очередь, путем фосфорилирования, активирует фосфорилазу b, переходящую в фосфорилазу а (рис. 19.5).
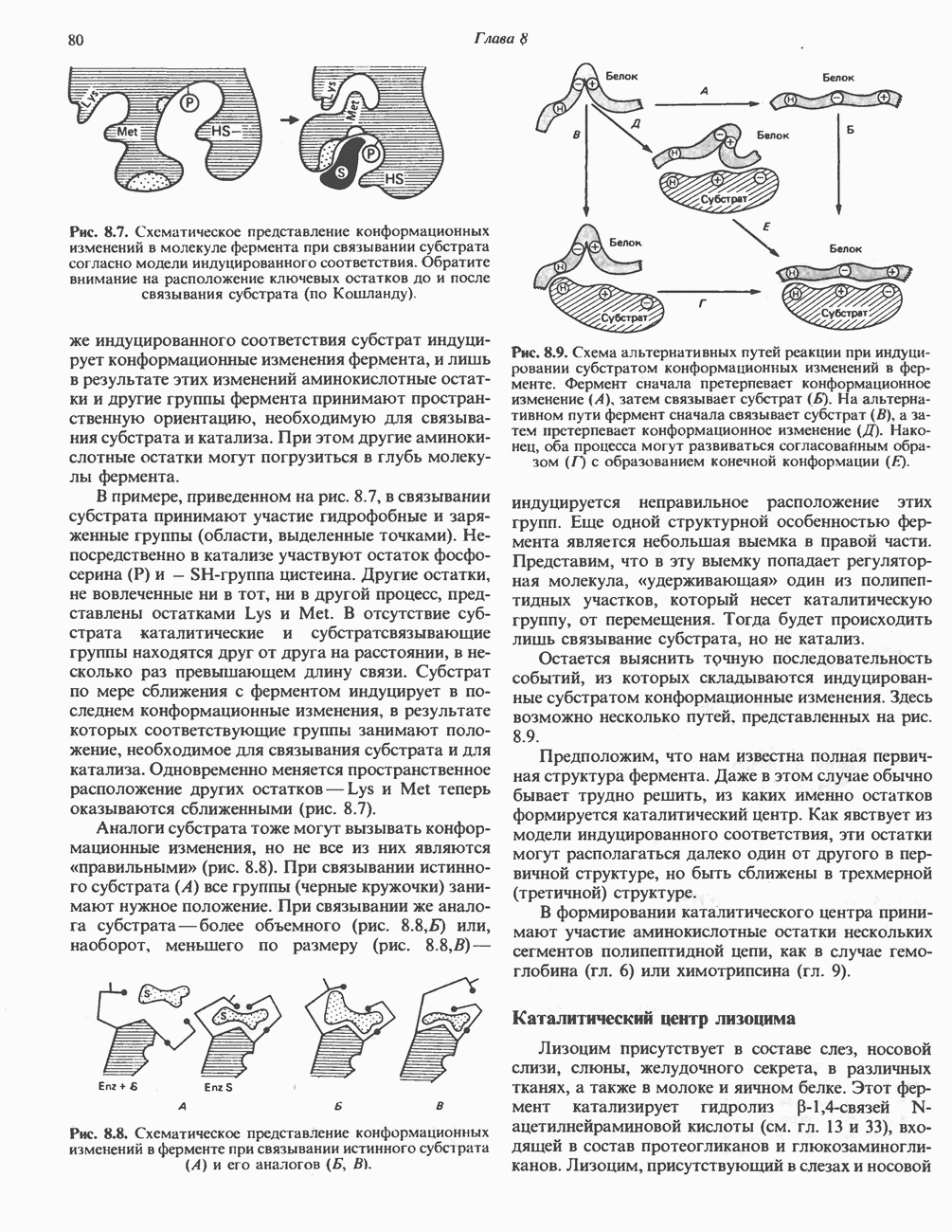
Неактивная сАМР - зависимая протеинкиназа состоит из двух пар субъединиц; в каждую пару входят регуляторная субъединица (R), способная связывать две молекулы сАМР, и каталитическая субъединица (С), структура которой включает активный центр. Связывание сАМР с комплексом  вызывает диссоциацию последнего, приводящую к освобождению активных С-мономеров (см. гл. 44):
вызывает диссоциацию последнего, приводящую к освобождению активных С-мономеров (см. гл. 44):

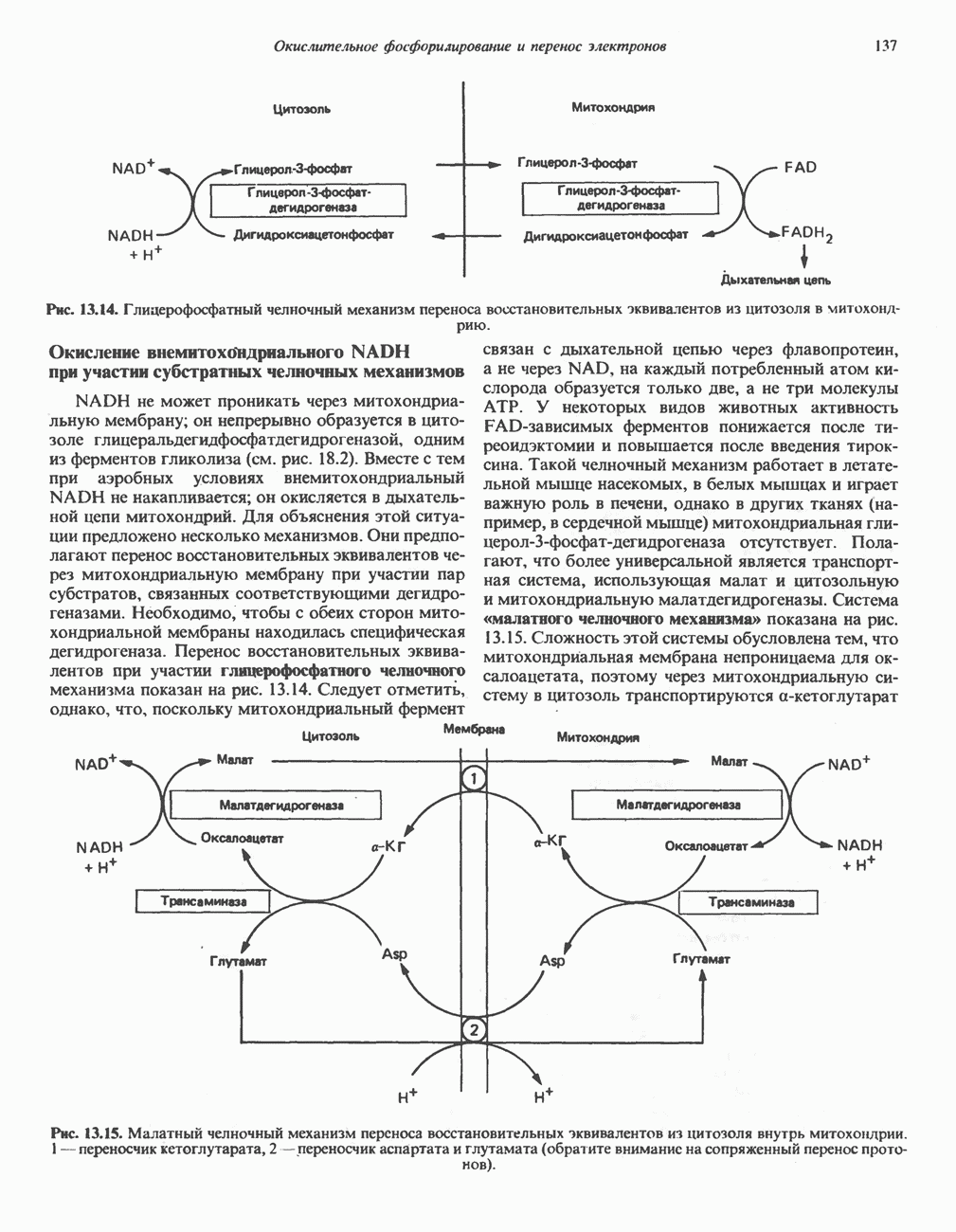
Активация ионами и синхронизация с мышечным сокращением
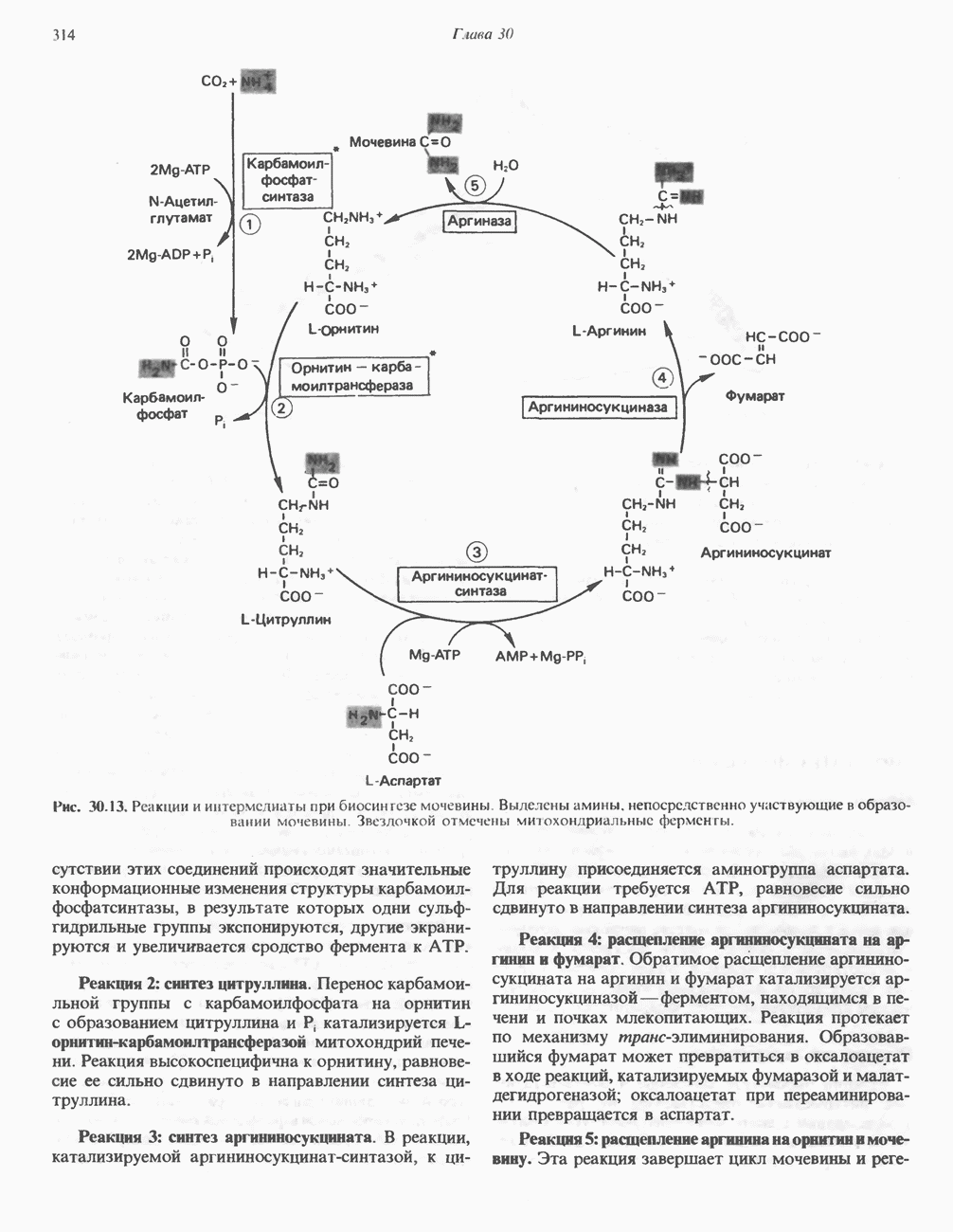
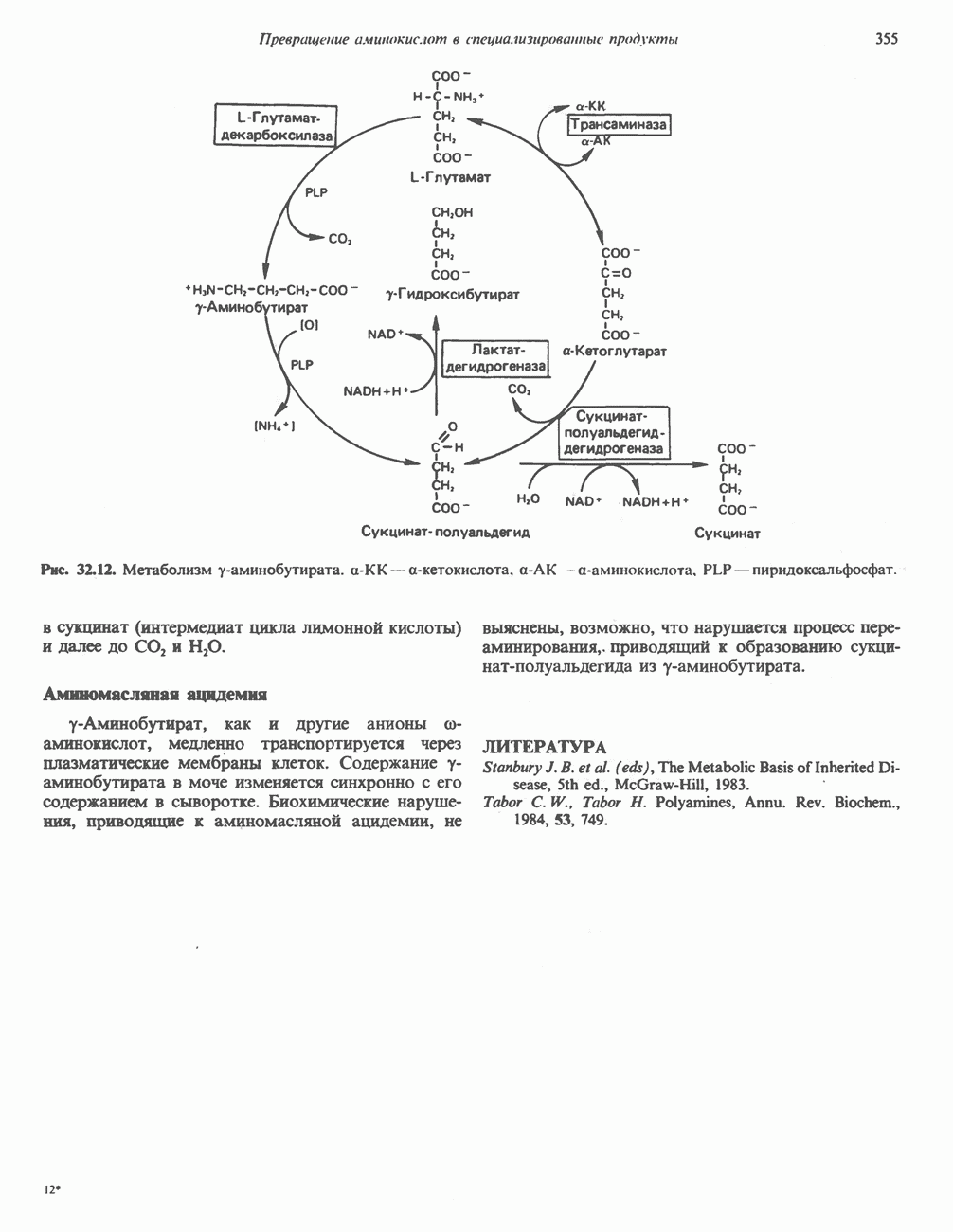
Сразу после начала сокращения мышцы гликогенолиз возрастает в несколько сотен раз. Процесс включает быструю активацию фосфорилазы благодаря активации киназы фосфорилазы ионами  тем же сигналом, который инициирует сокращение. Мышечная киназа фосфорилазы состоит из субъединиц четырех типов:
тем же сигналом, который инициирует сокращение. Мышечная киназа фосфорилазы состоит из субъединиц четырех типов:  . Ее структура —
. Ее структура —  Субъединицы а и
Субъединицы а и  содержат остатки серина, фосфорилируемые
содержат остатки серина, фосфорилируемые  -зависимой протеинкиназой. Р-Субъединица связывает четыре иона
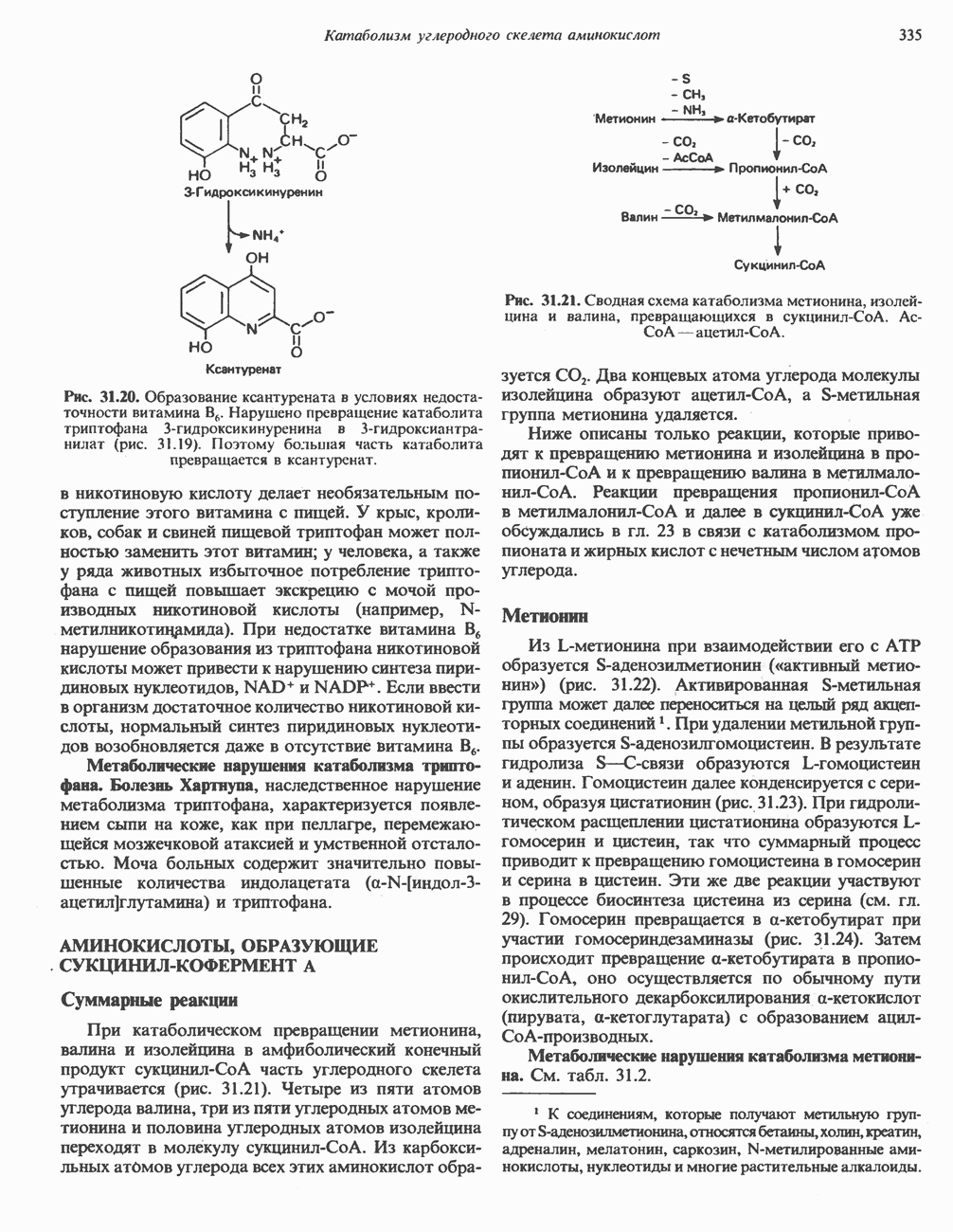
-зависимой протеинкиназой. Р-Субъединица связывает четыре иона  она идентична
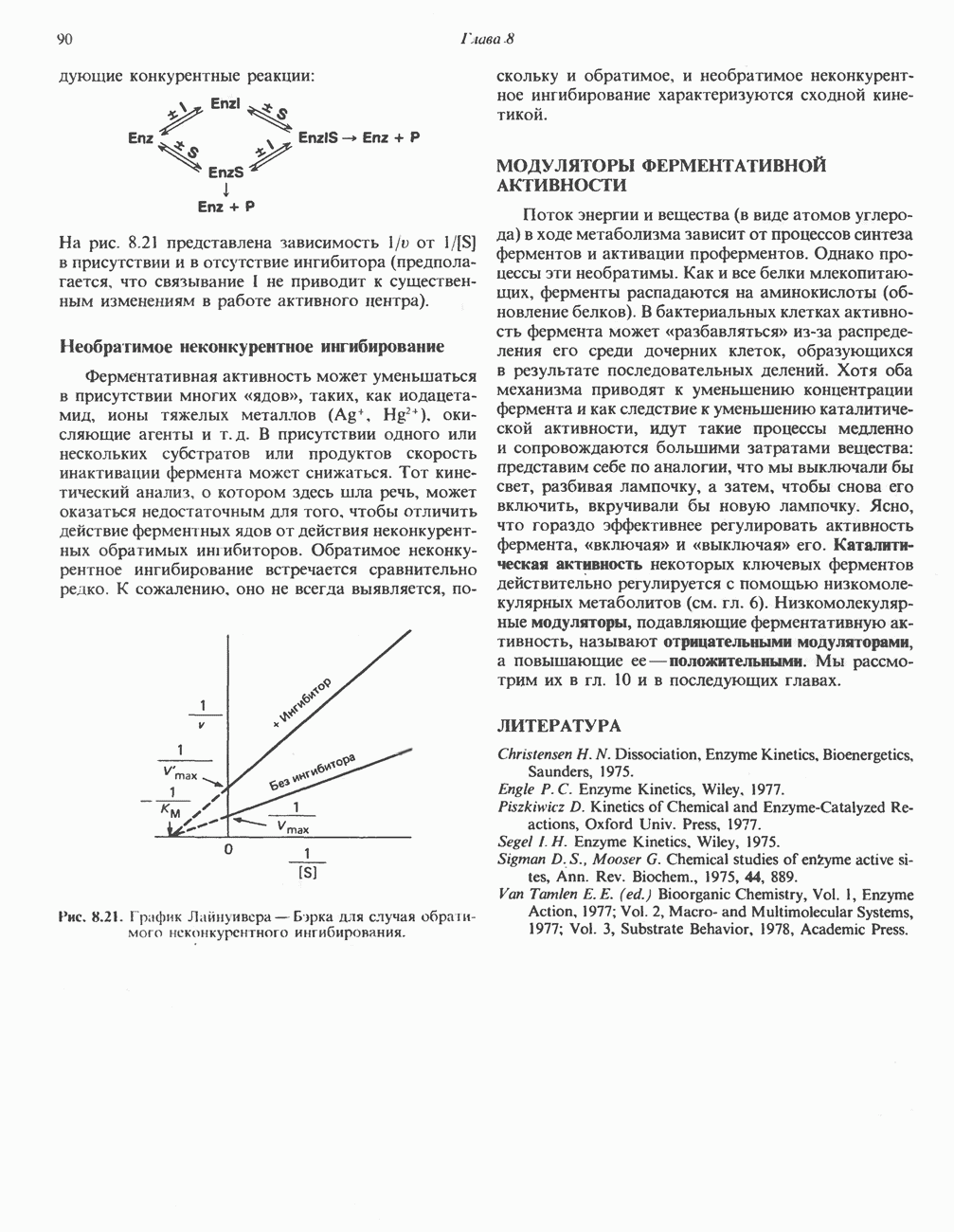
она идентична  -связывающему белку кальмодулину. Связывание ионов
-связывающему белку кальмодулину. Связывание ионов  активирует каталитический центр у-субъединицы, хотя молекула остается в дефосфорилированной конфигурации. В то же время полную активность фосфорилированная а-форма приобретает только в присутствии ионов
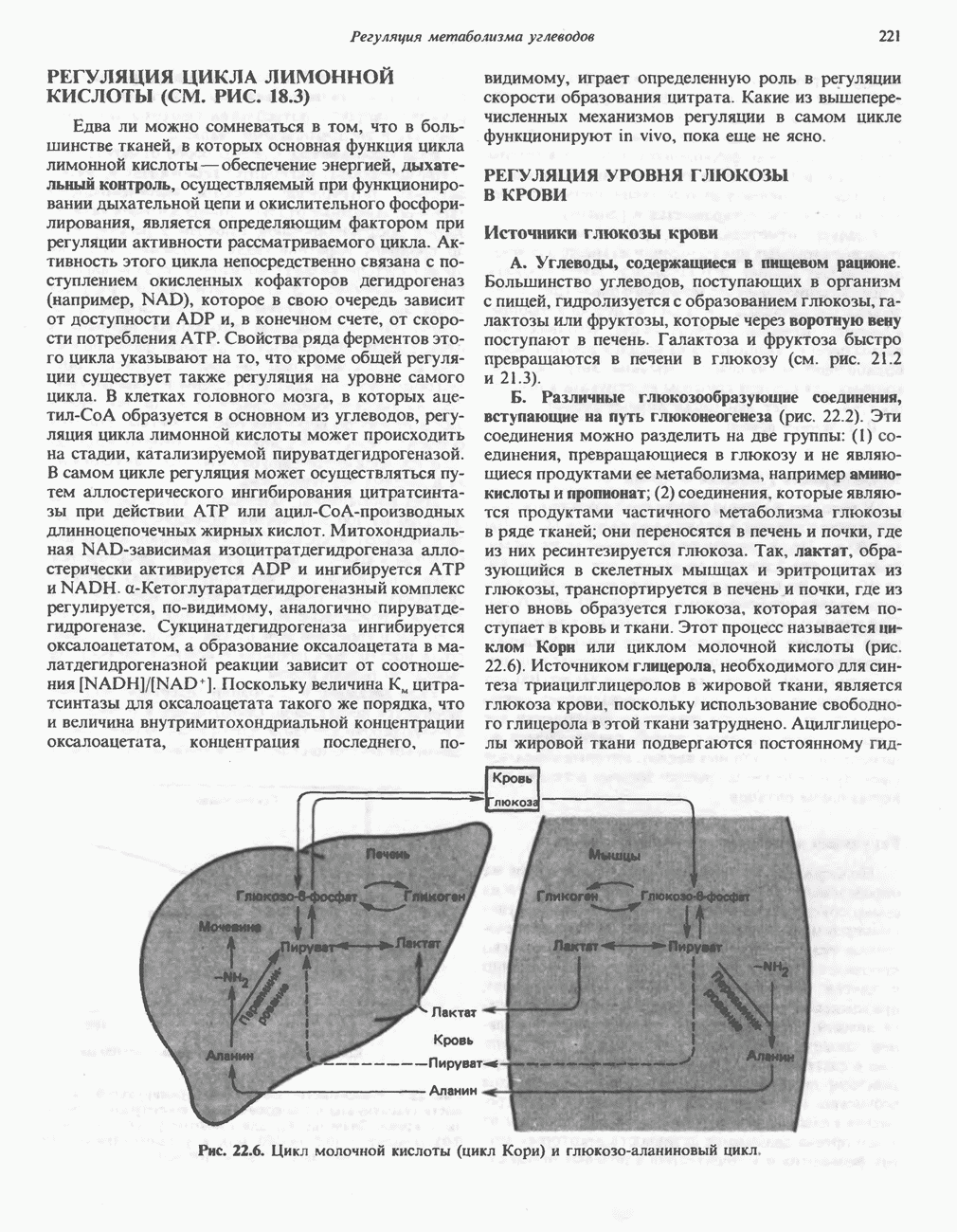
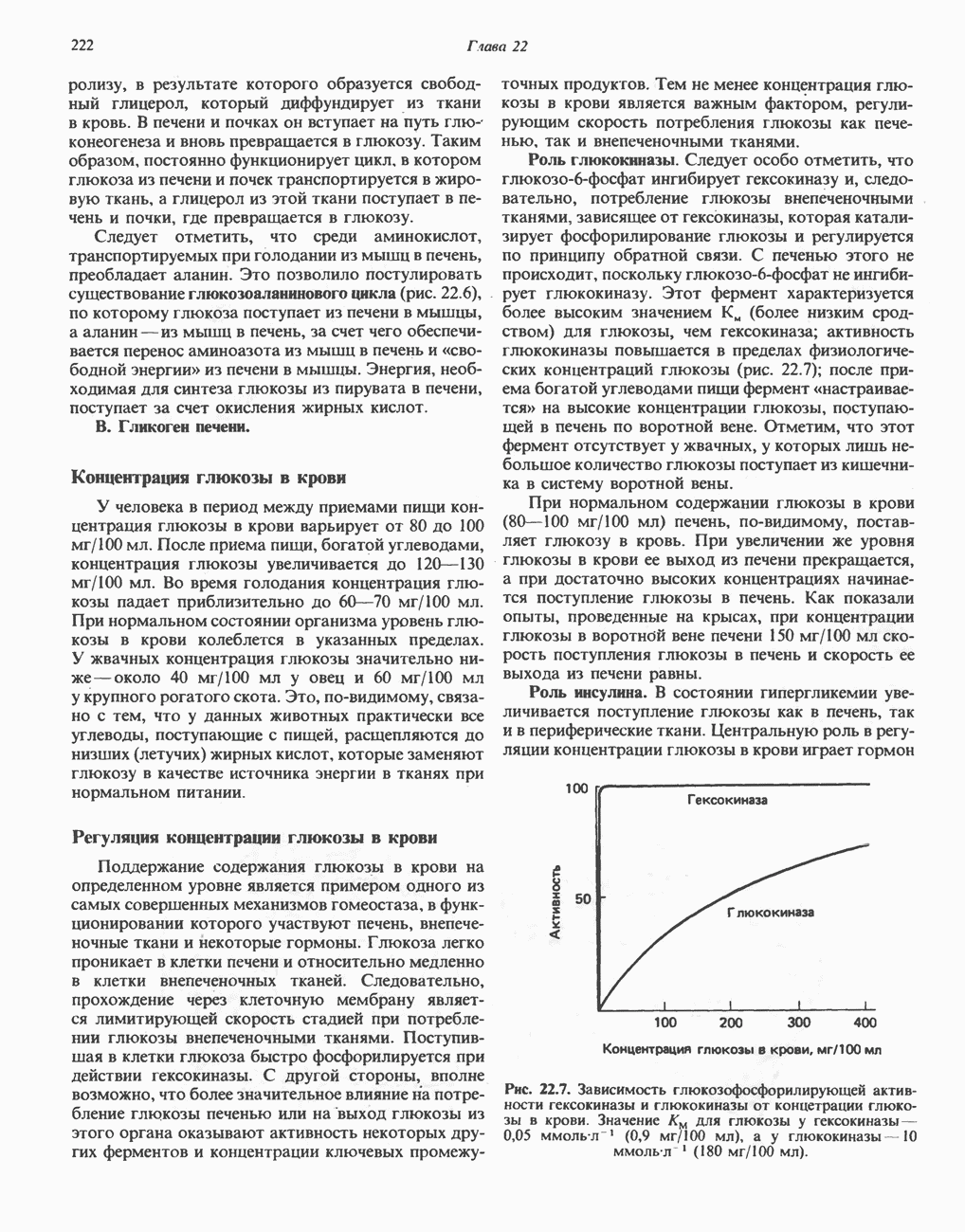
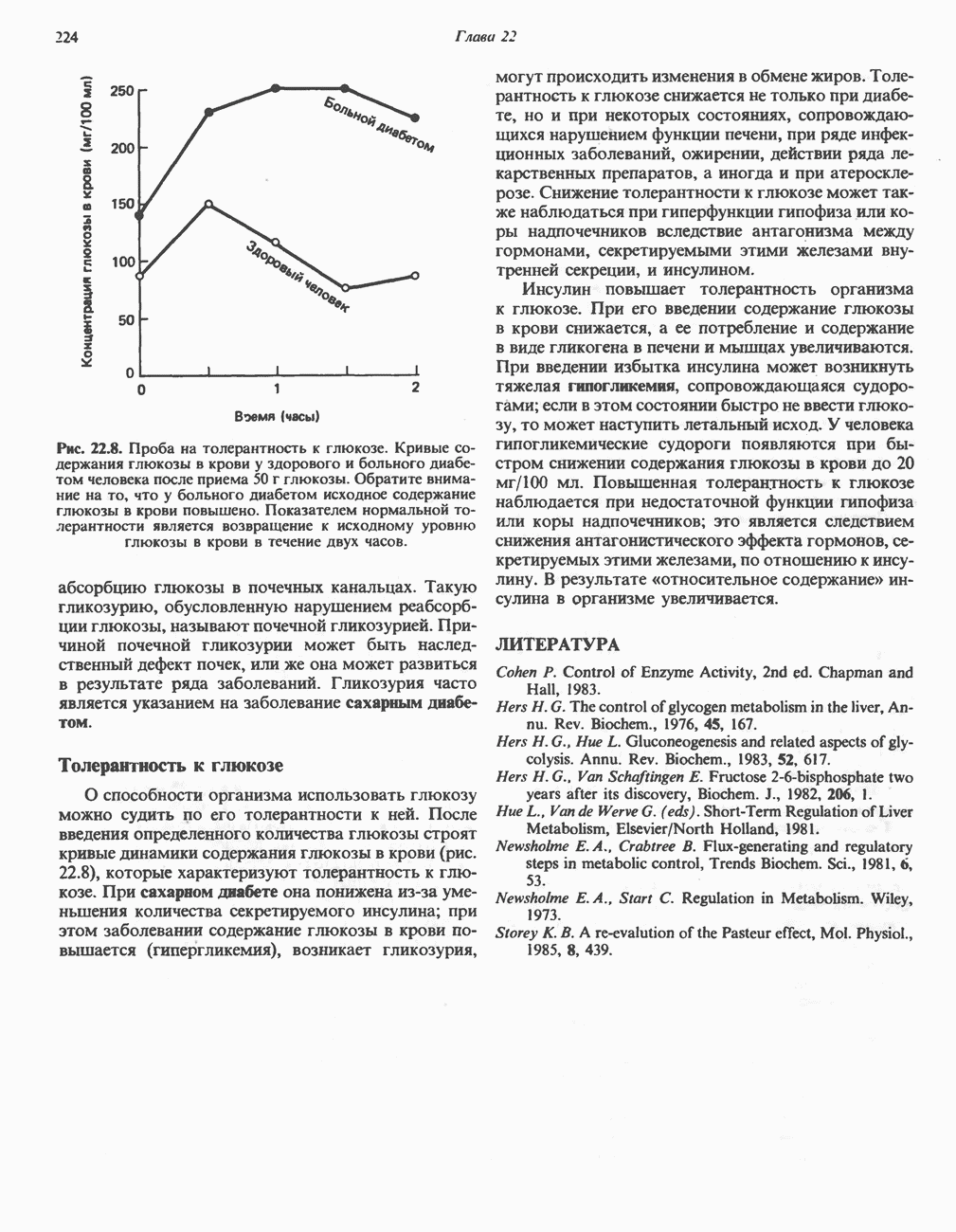
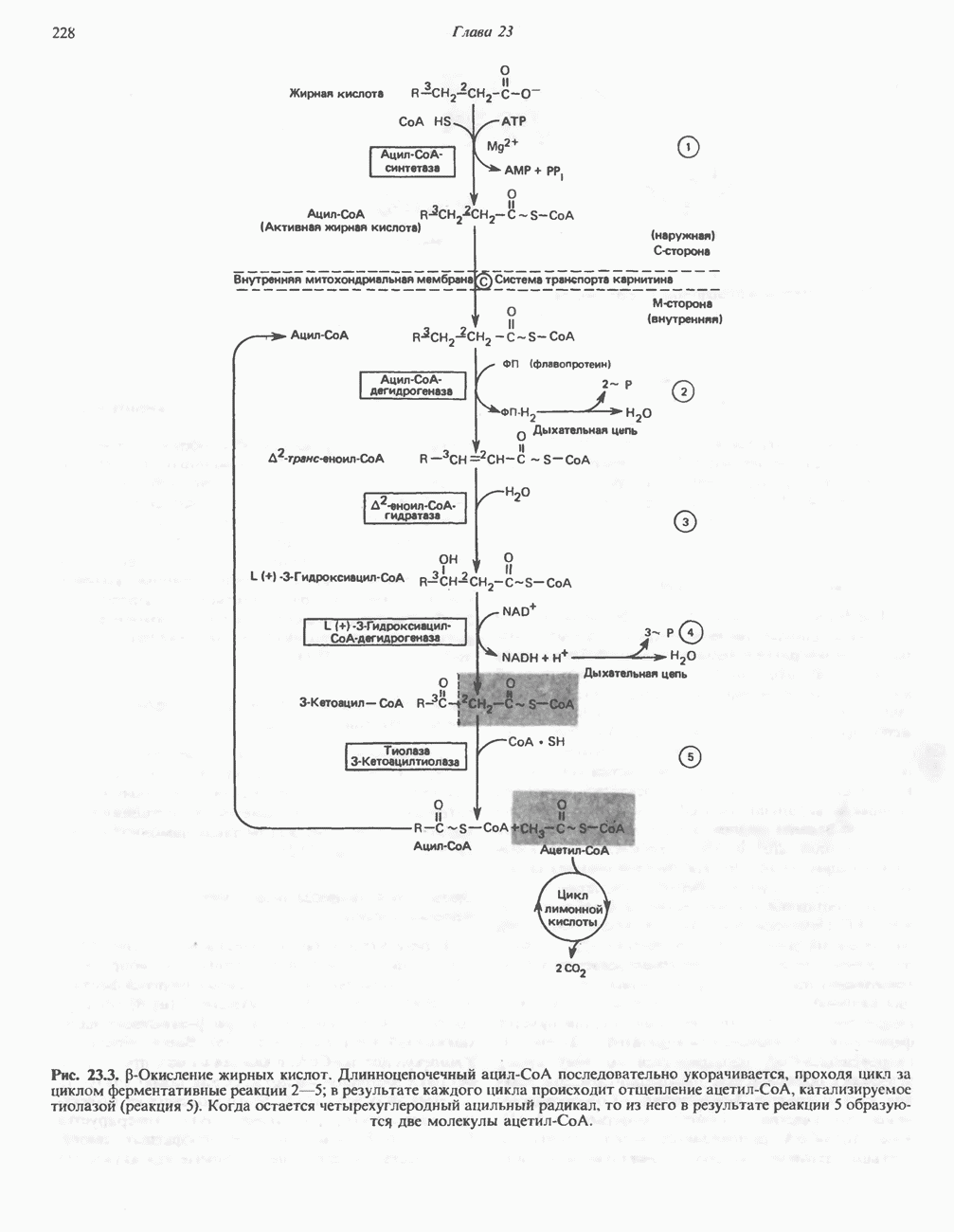
активирует каталитический центр у-субъединицы, хотя молекула остается в дефосфорилированной конфигурации. В то же время полную активность фосфорилированная а-форма приобретает только в присутствии ионов  Важно отметить, что кальмодулин имеет структурное сходство с мышечным
Важно отметить, что кальмодулин имеет структурное сходство с мышечным  - связывающим белком тропонином
- связывающим белком тропонином  . С киназой фосфорилазы может взаимодействовать вторая молекула кальмодулина или
. С киназой фосфорилазы может взаимодействовать вторая молекула кальмодулина или  вызывая дополнительную активацию фермента. Таким образом, активация мышечного сокращения и гликогенолиз осуществляются одним и тем же
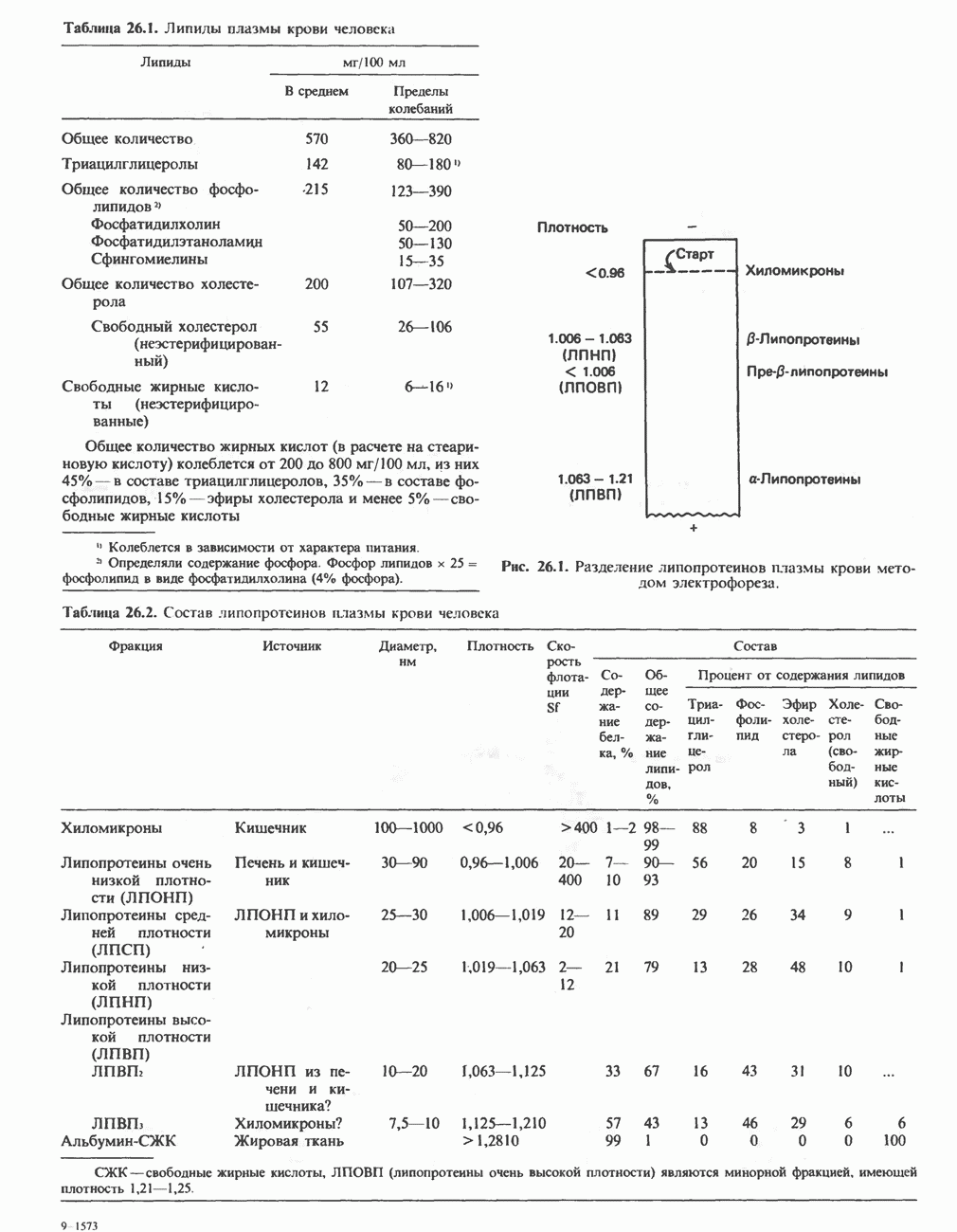
вызывая дополнительную активацию фермента. Таким образом, активация мышечного сокращения и гликогенолиз осуществляются одним и тем же  -связывающим белком. Кальмодулин — белок, который участвует во многих видах воздействия кальция на клетку (см. гл. 44).
-связывающим белком. Кальмодулин — белок, который участвует во многих видах воздействия кальция на клетку (см. гл. 44).
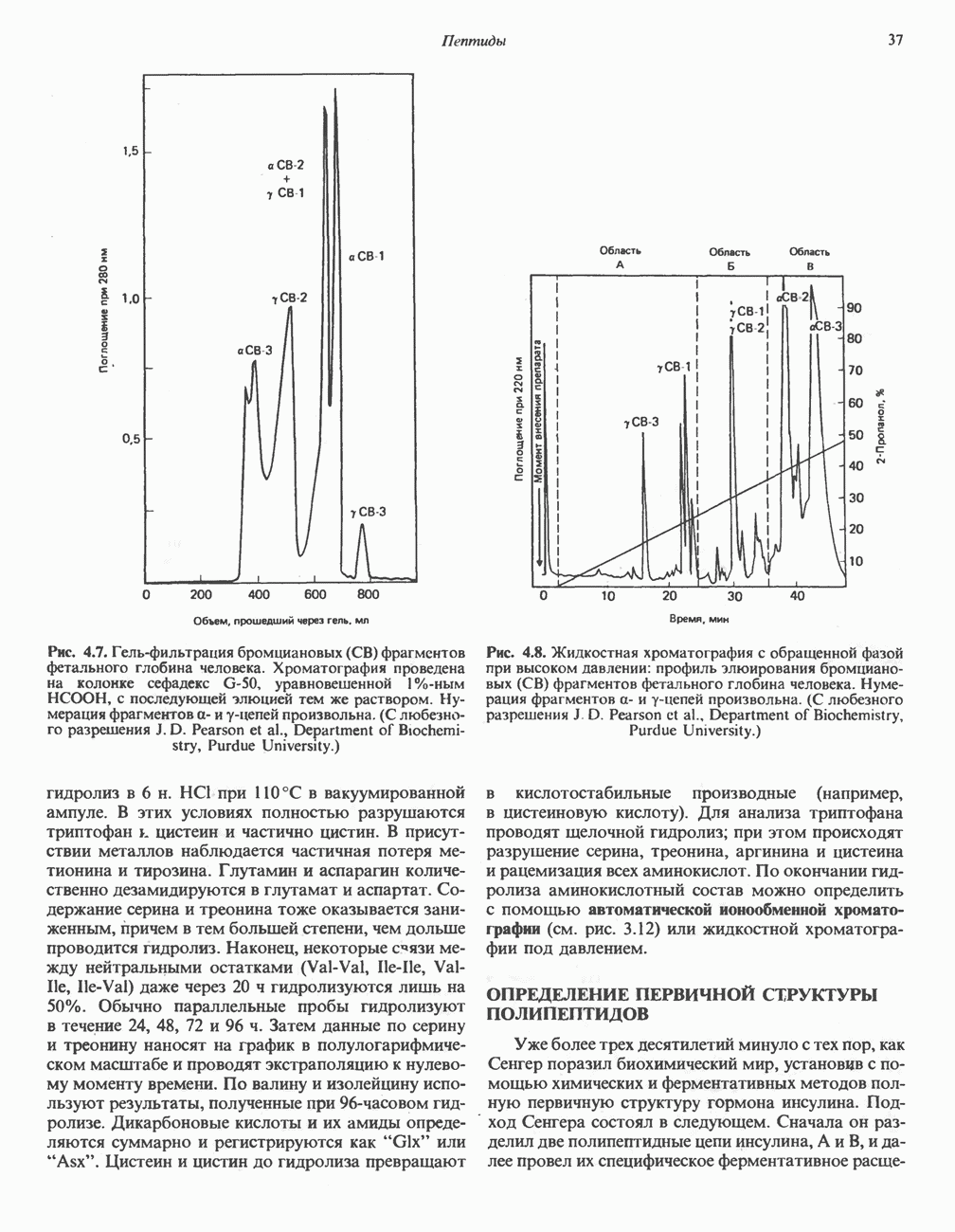
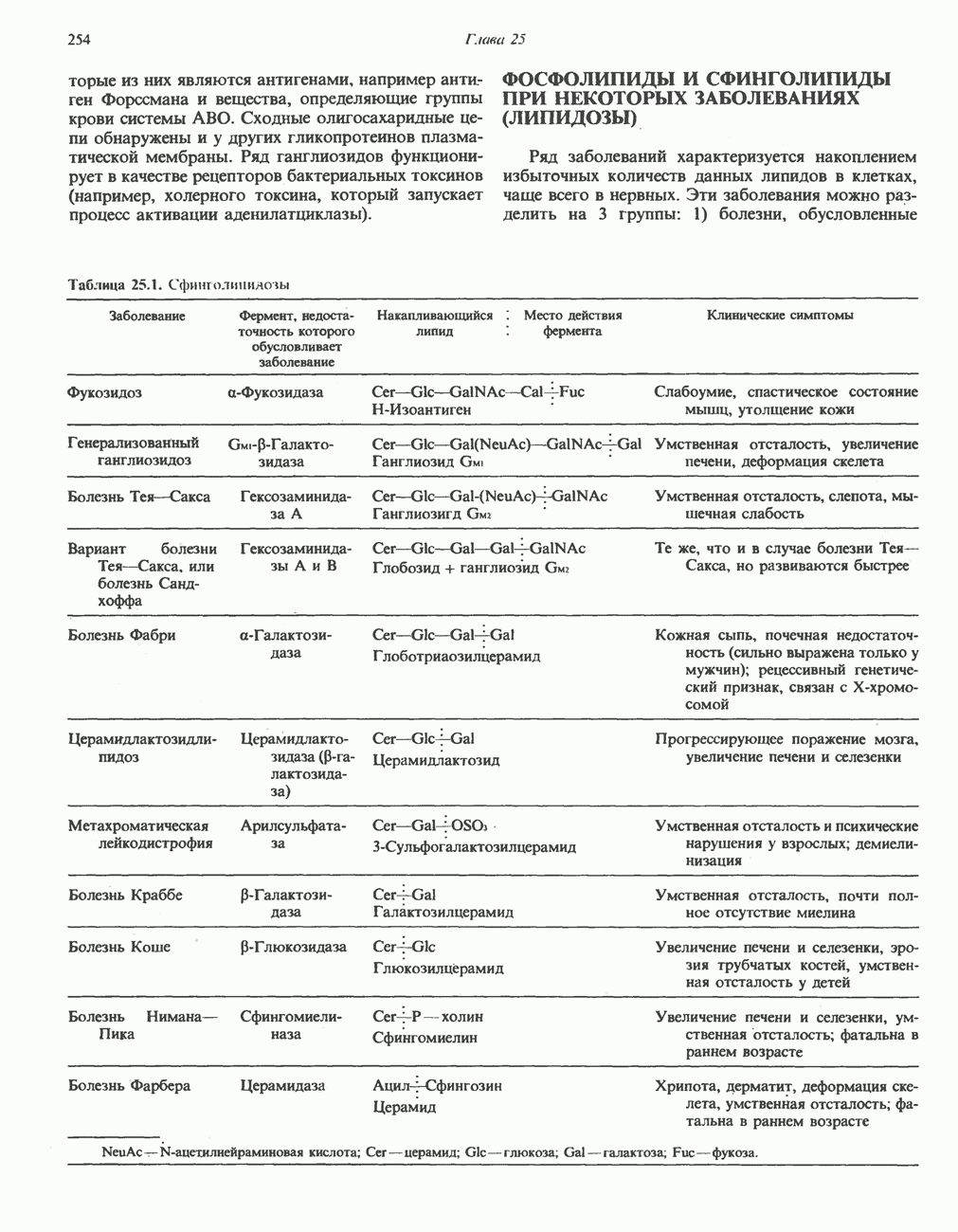
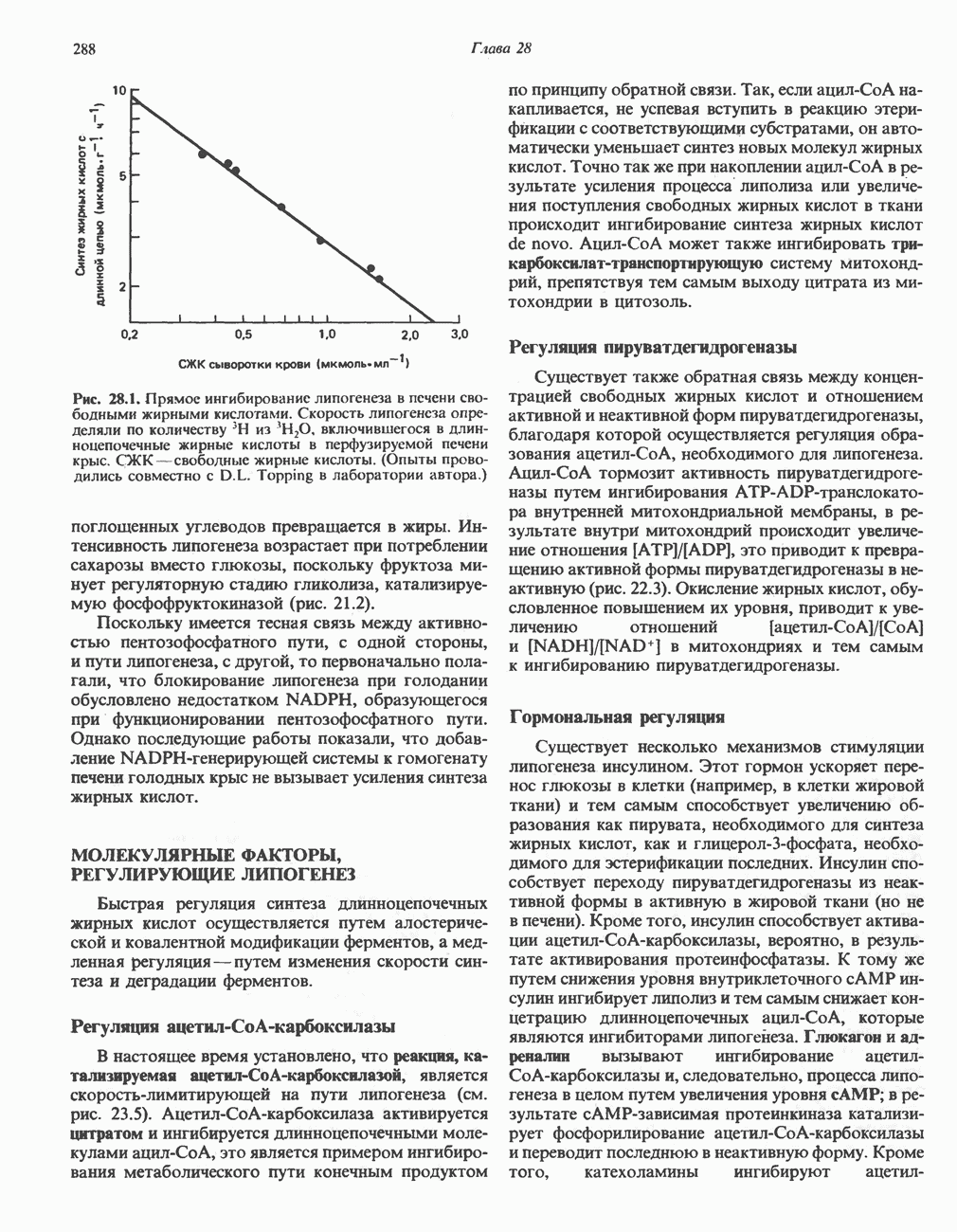
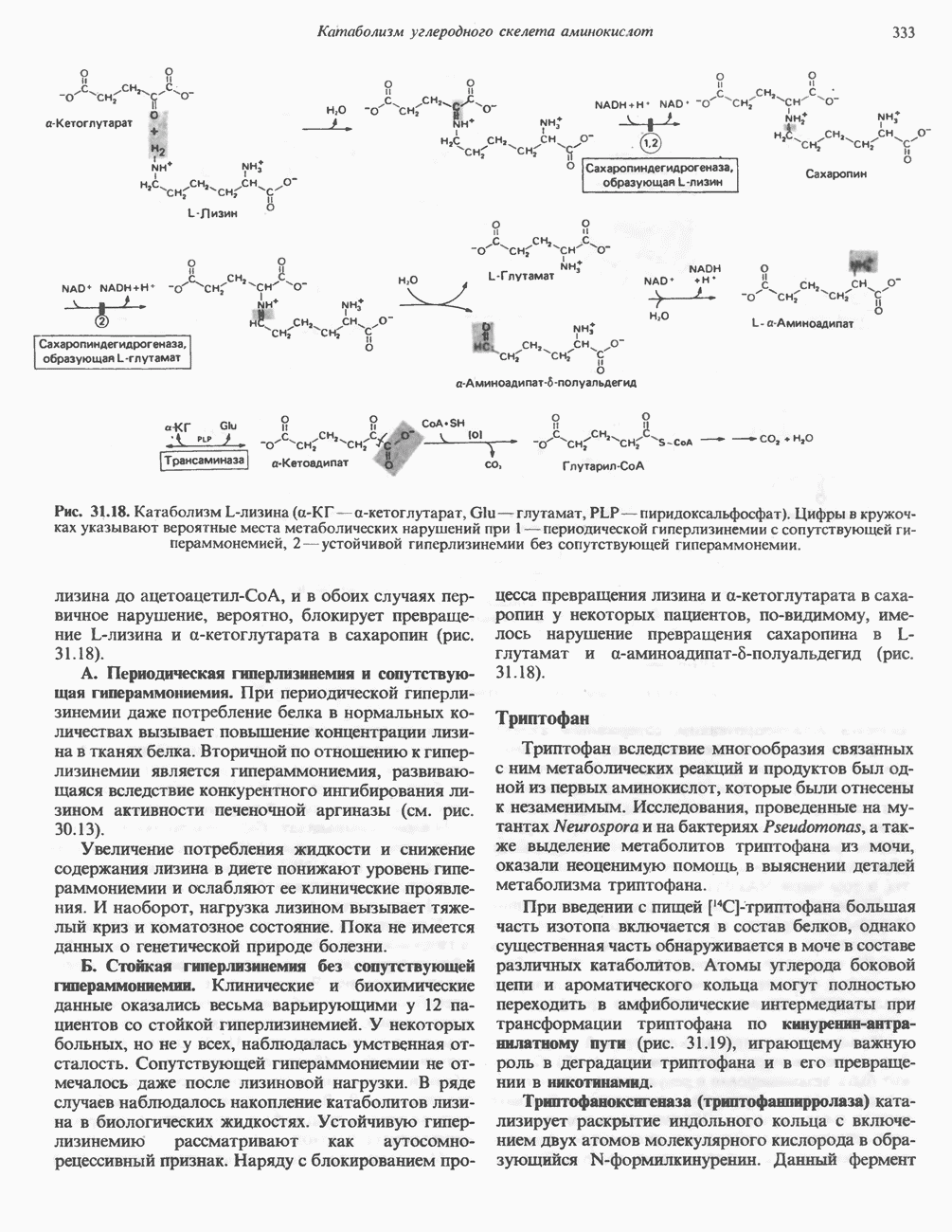
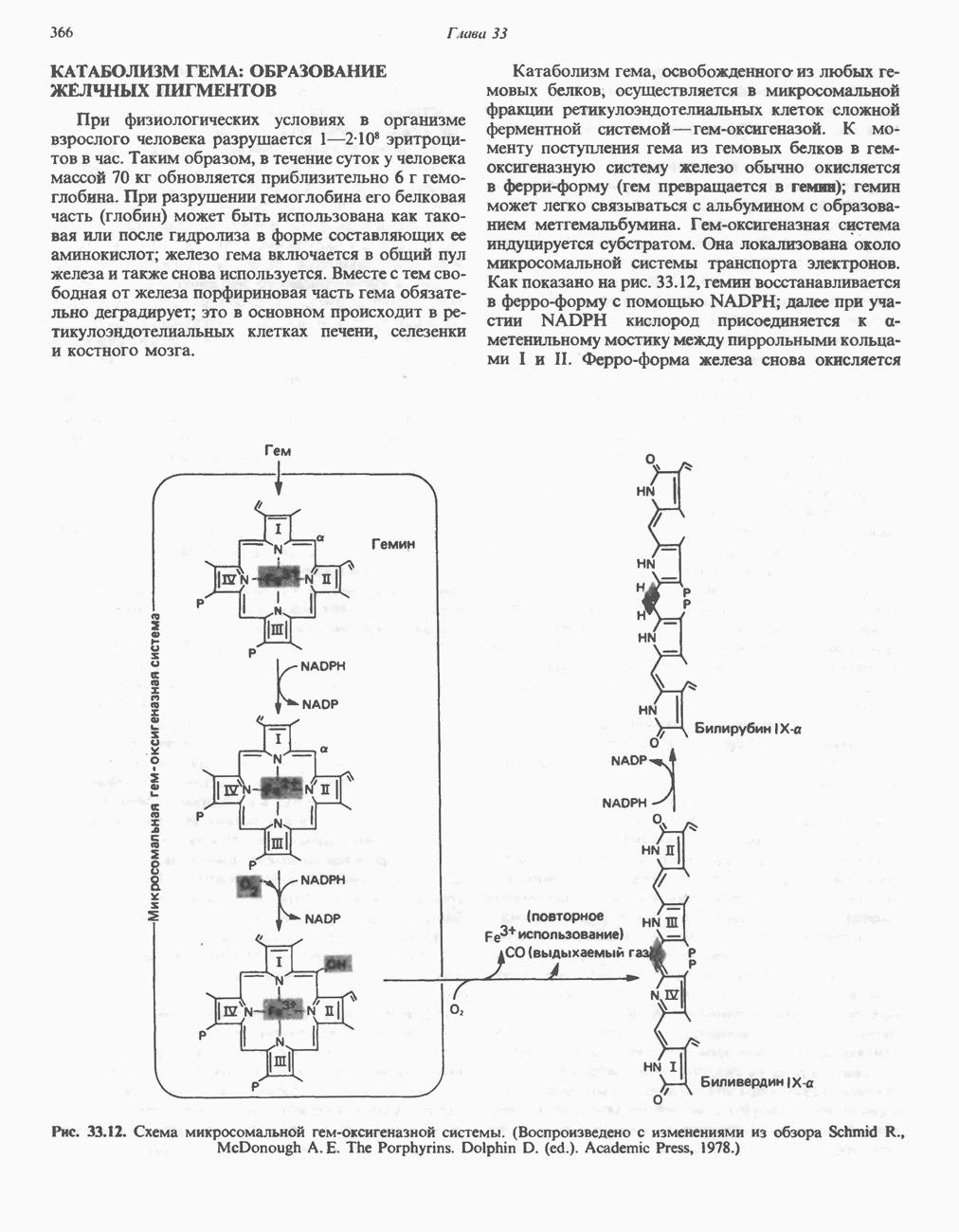
Гликогенолиз в печени
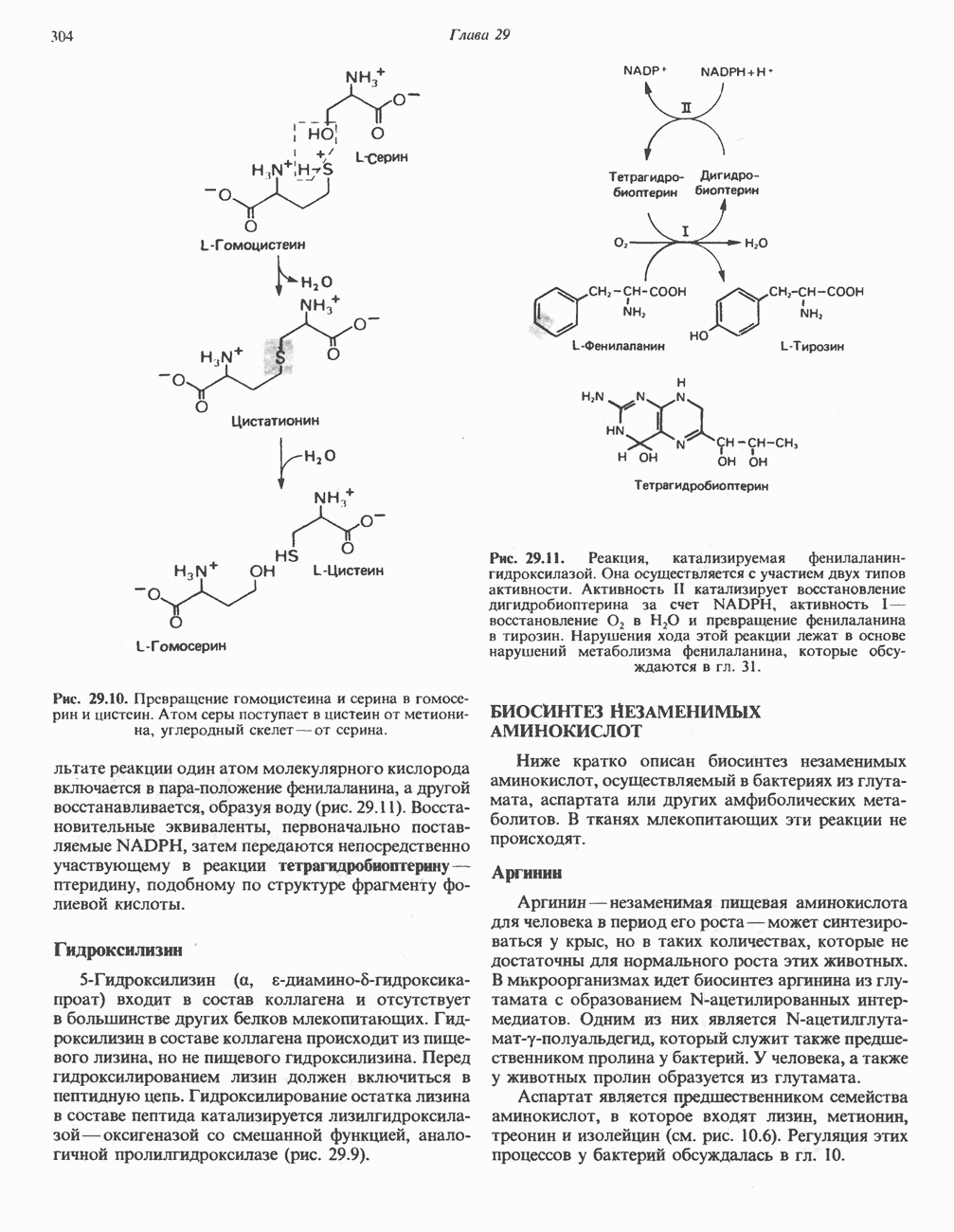
Установлено, что при стимуляции гликогенолиза катехоламинами в печени в качестве главных посредников выступают  -рецепторы. При этом происходит
-рецепторы. При этом происходит  -независимая мобилизация ионов
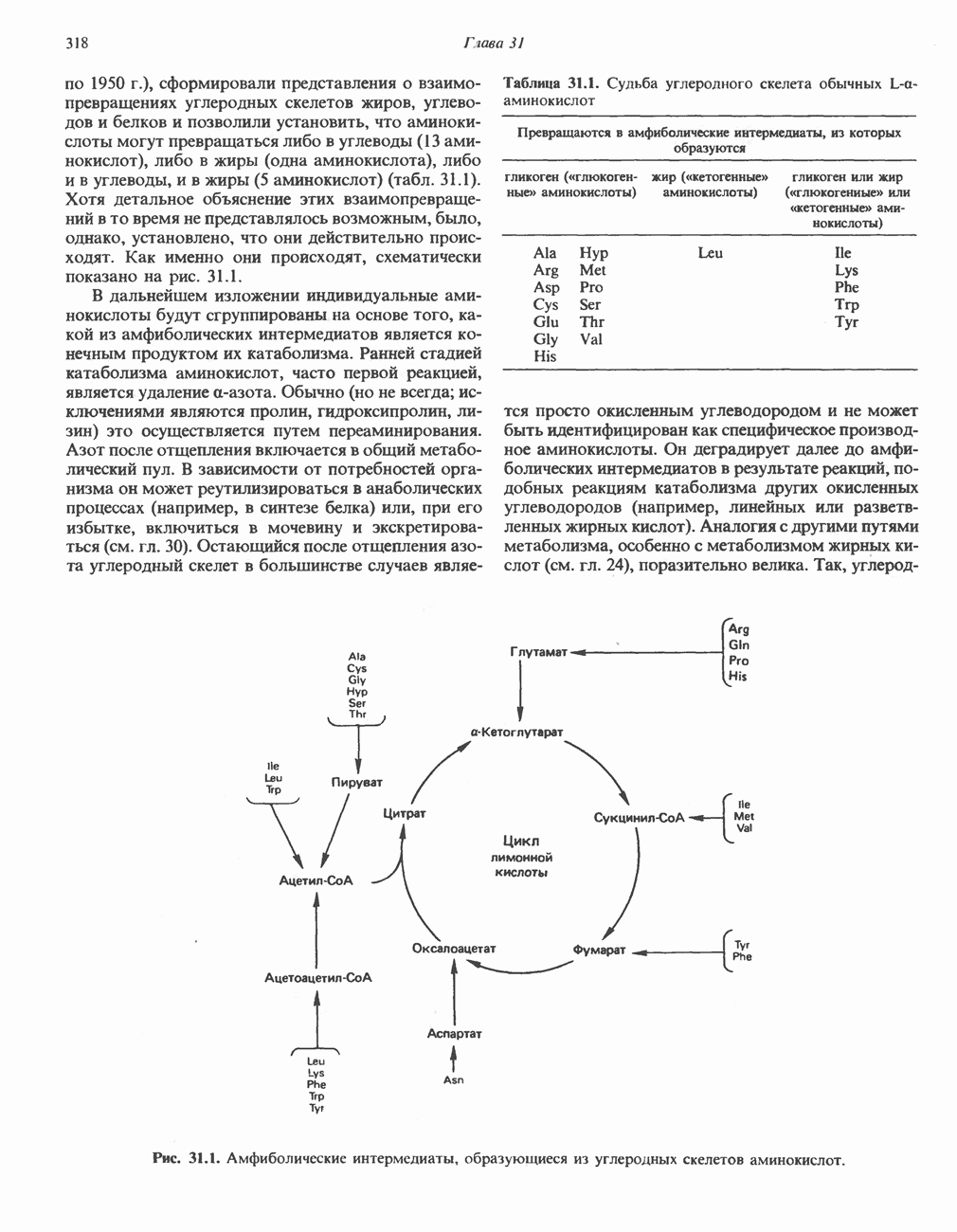
-независимая мобилизация ионов  и переход их из митохондрий в цитозоль, где они стимулируют
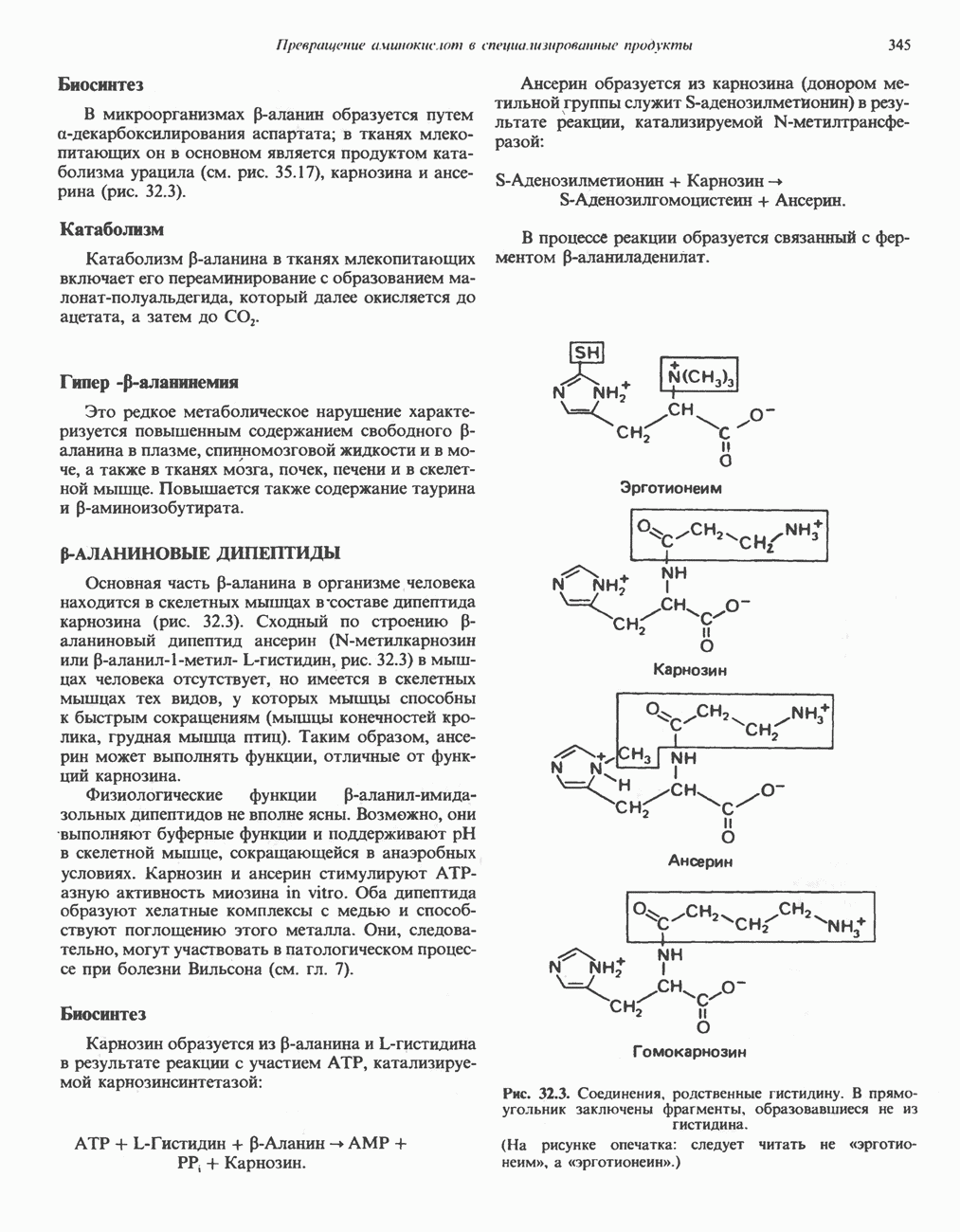
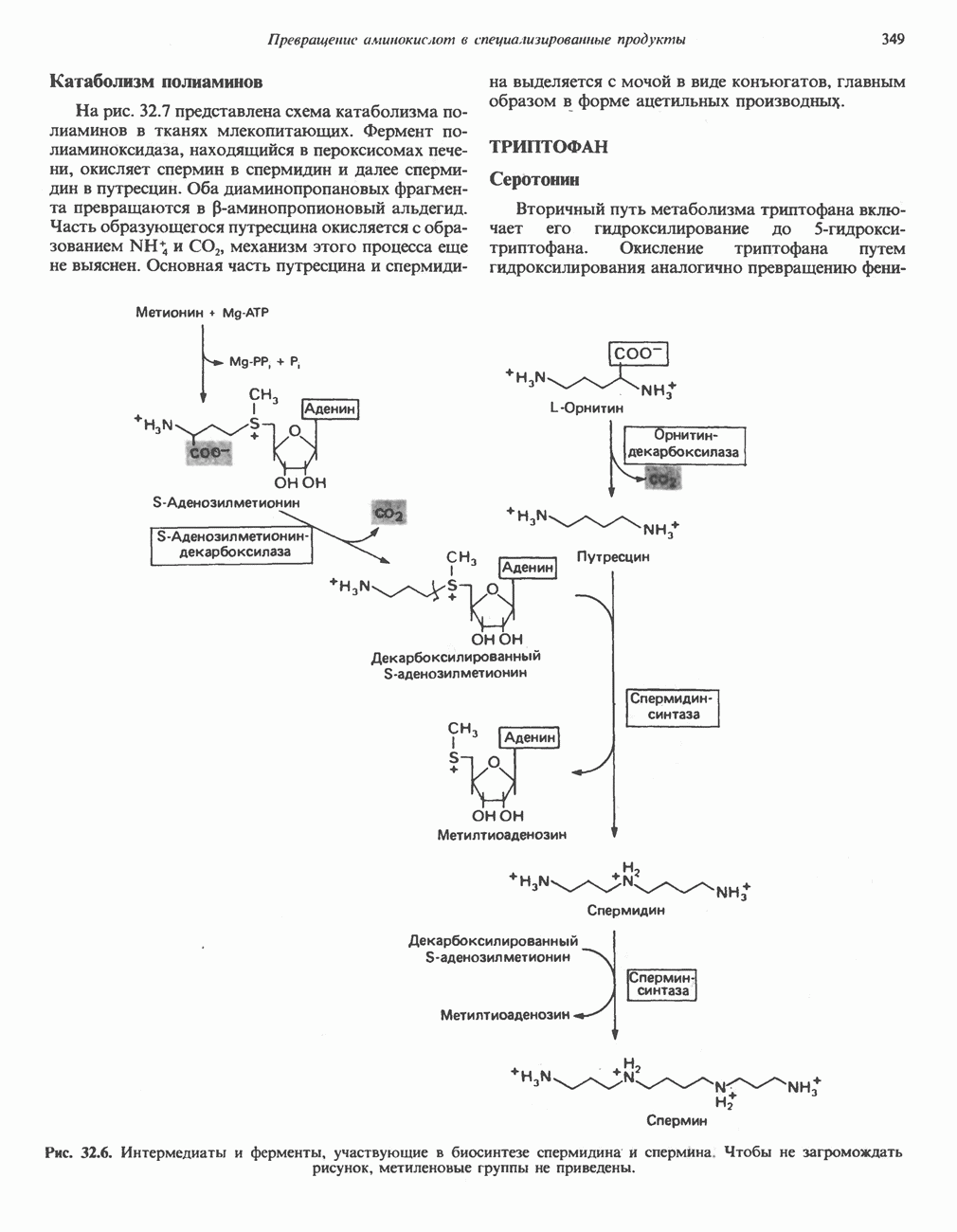
и переход их из митохондрий в цитозоль, где они стимулируют  кальмодулин чувствительную киназу фосфорилазы. Фосфорилаза скелетных мышц в отличие от фосфорилазы печени не активируется глюкагоном. Отметим, что фосфорилаза сердечной мышцы активируется этим гормоном. Другим важным отличием является ингибирование печеночной протеинфосфатазы-1 активной формой фосфорилазы.
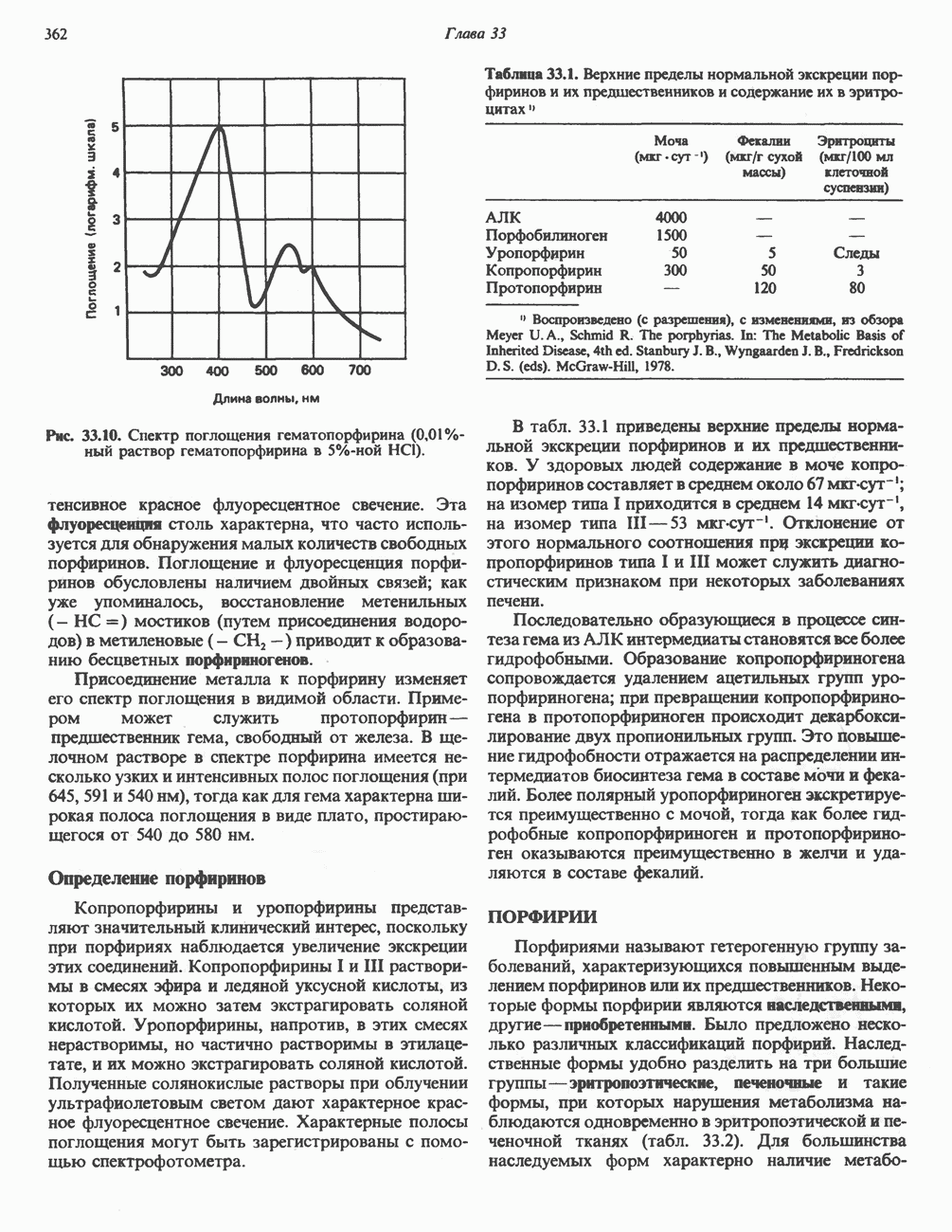
кальмодулин чувствительную киназу фосфорилазы. Фосфорилаза скелетных мышц в отличие от фосфорилазы печени не активируется глюкагоном. Отметим, что фосфорилаза сердечной мышцы активируется этим гормоном. Другим важным отличием является ингибирование печеночной протеинфосфатазы-1 активной формой фосфорилазы.
Инактивация фосфорилазы
Фосфорилаза а и киназа фосфорилазы а дефосфорилируются и инактивируются протеинфосфатазой-1. Ингибитором протеинфосфатазы-1 является белок, который называют ингибитором-1; последний становится активным только после фосфорилирования  -зависимой протеинкиназой. Таким образом,
-зависимой протеинкиназой. Таким образом,  контролирует как активацию, так и инактивацию фосфорилазы (рис. 19.5).
контролирует как активацию, так и инактивацию фосфорилазы (рис. 19.5).
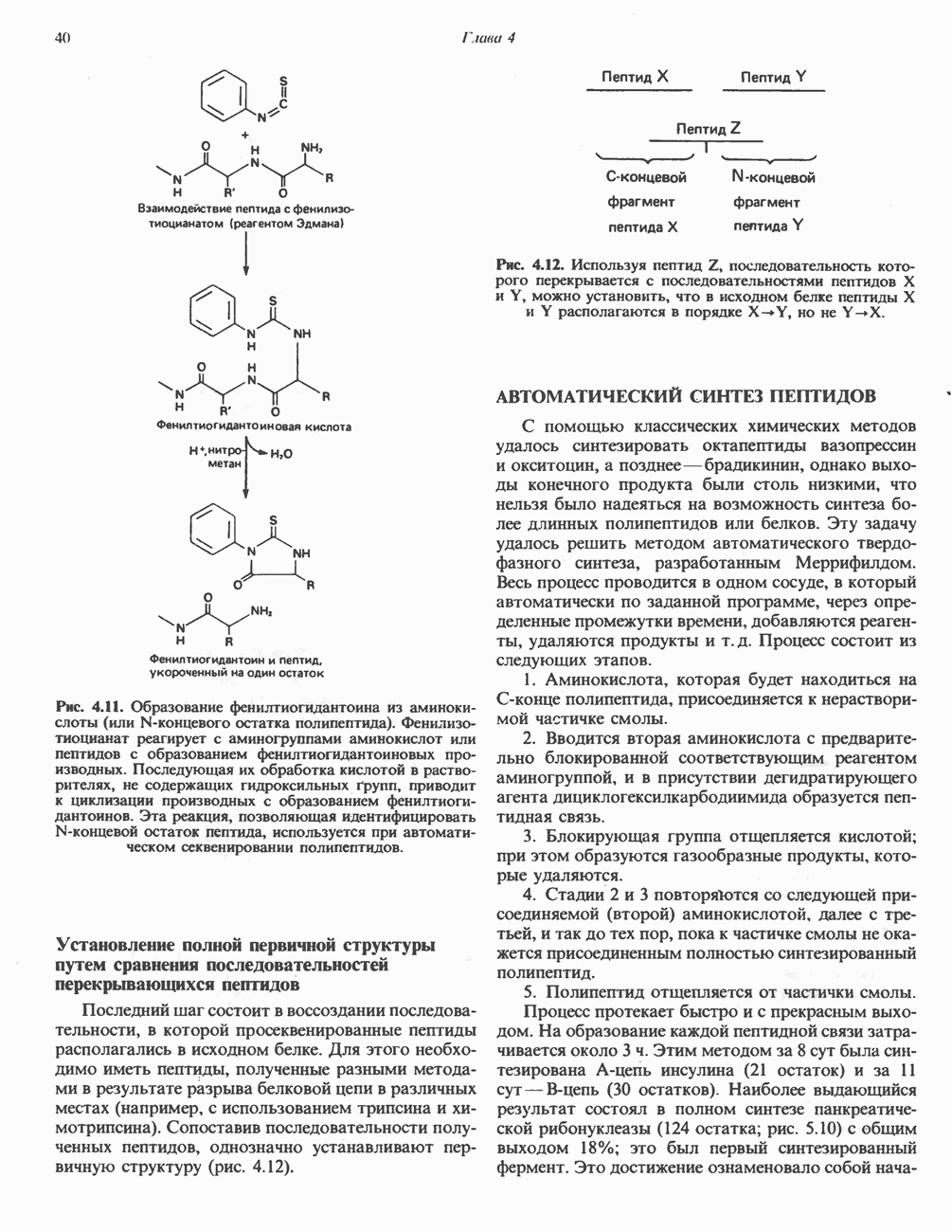
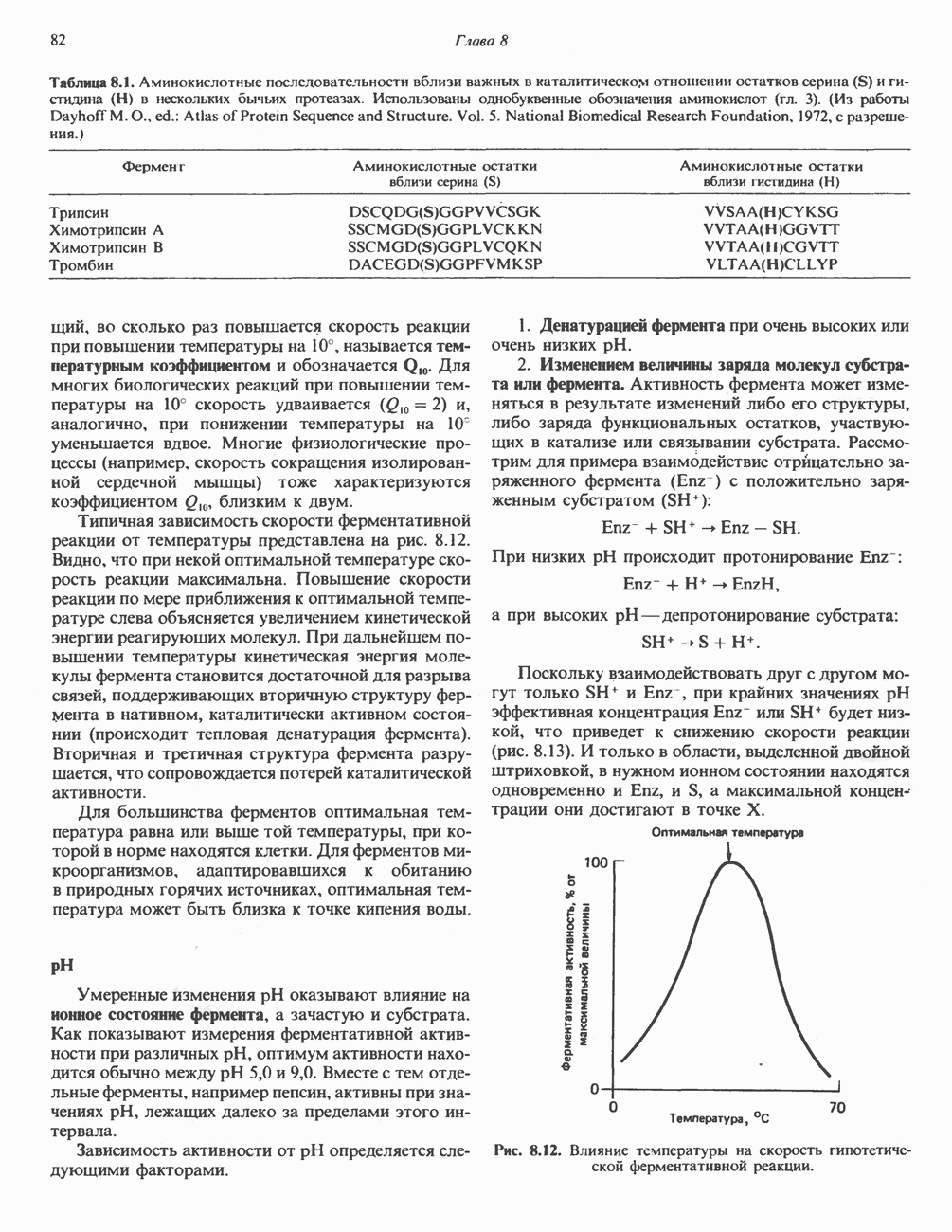
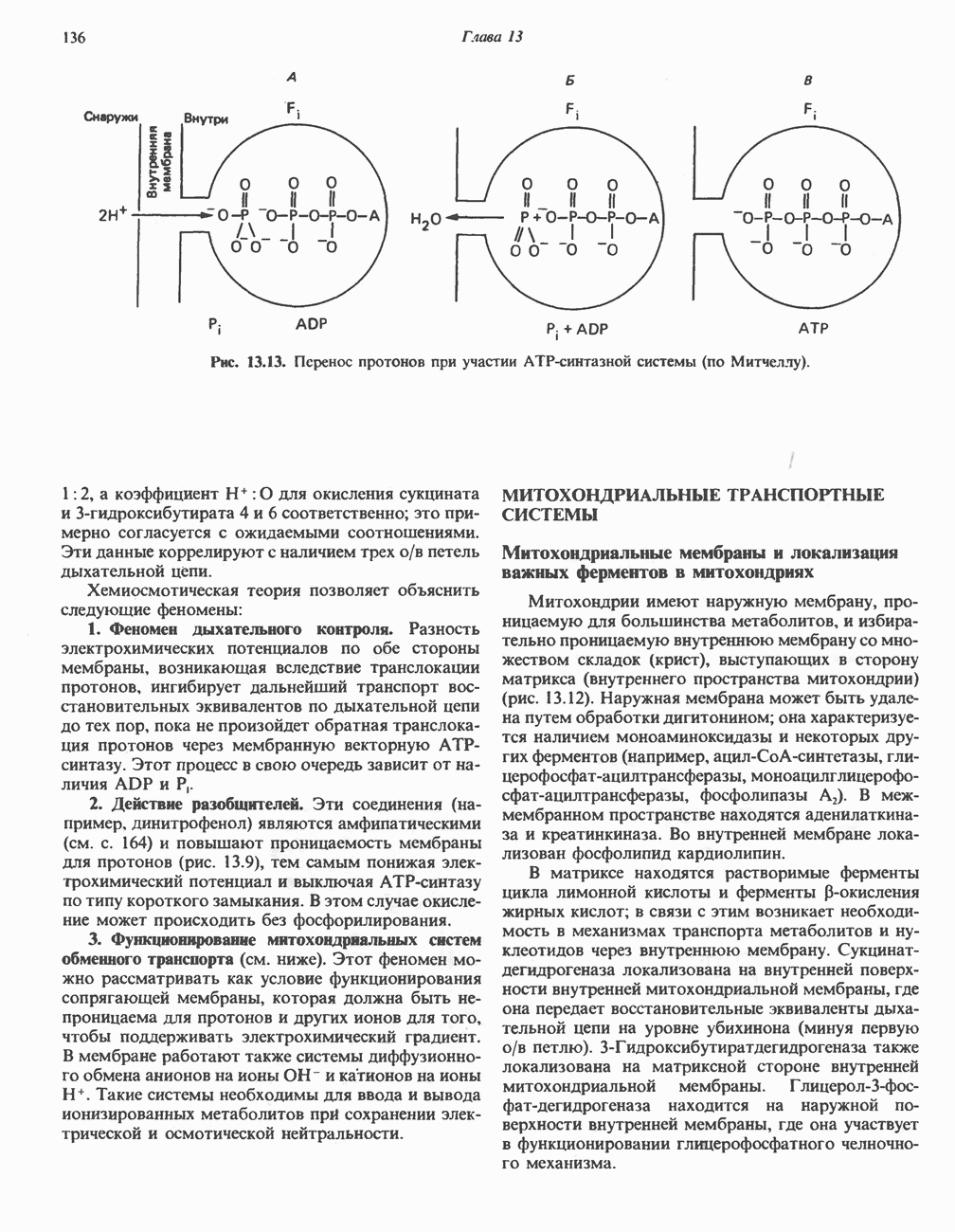
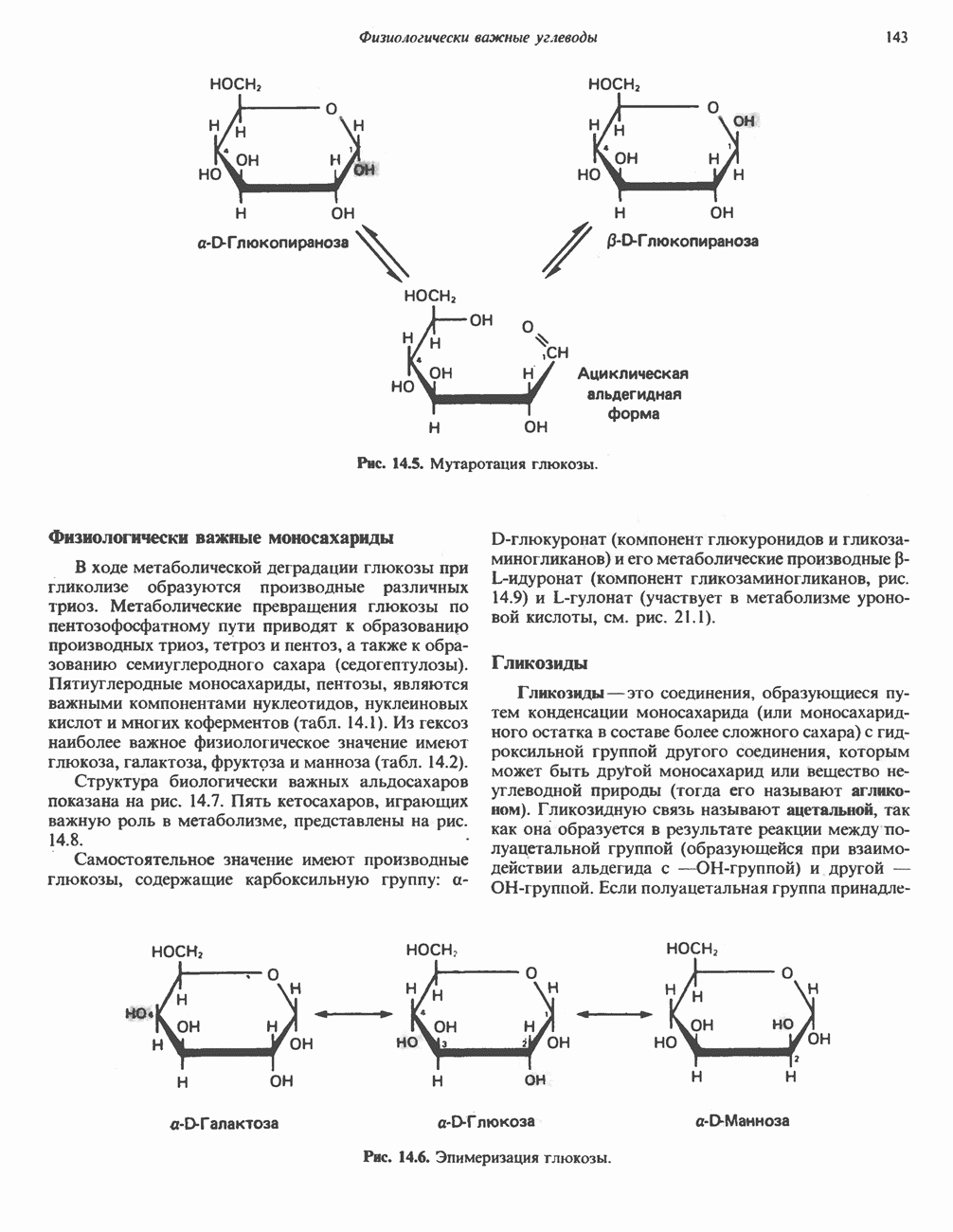
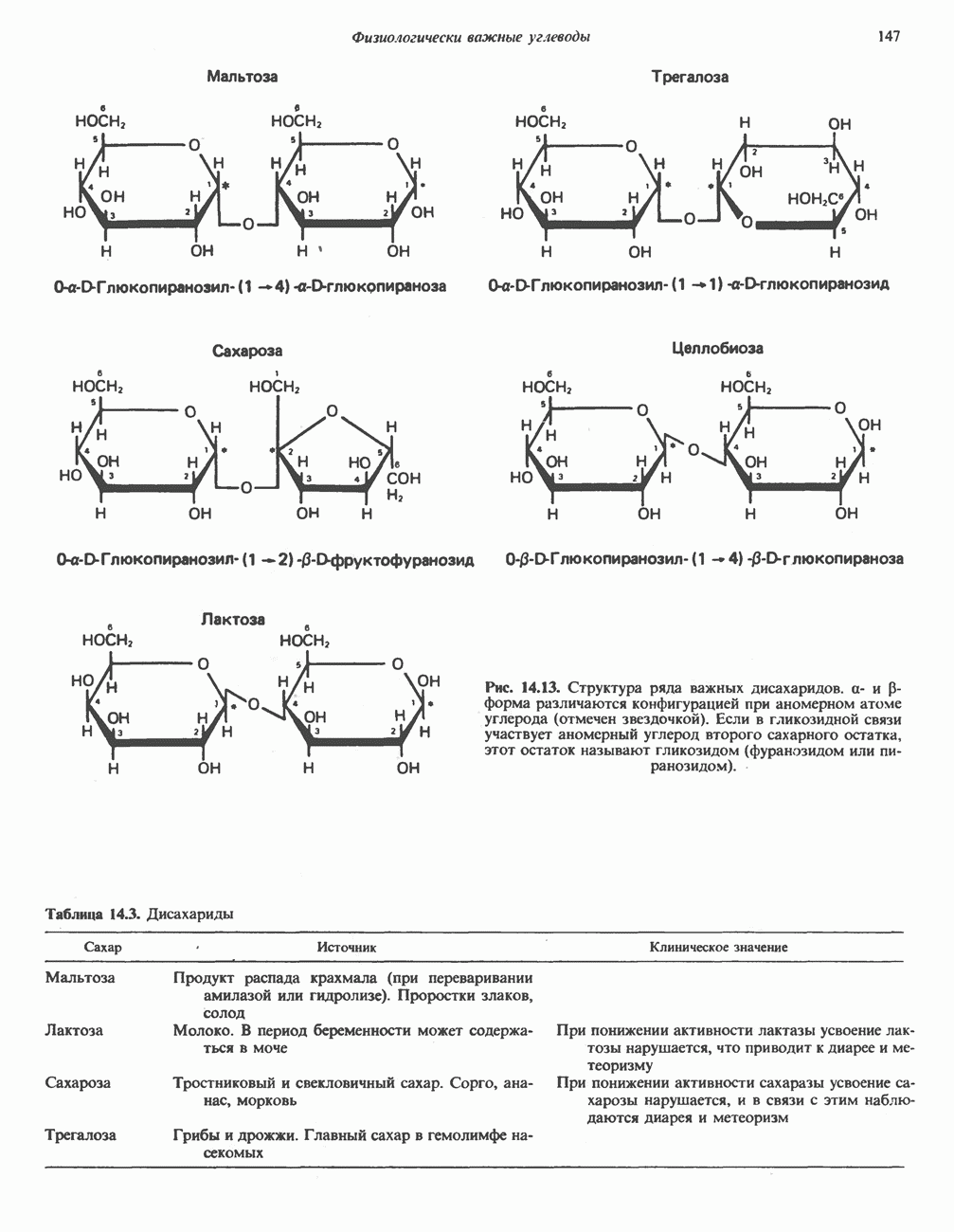
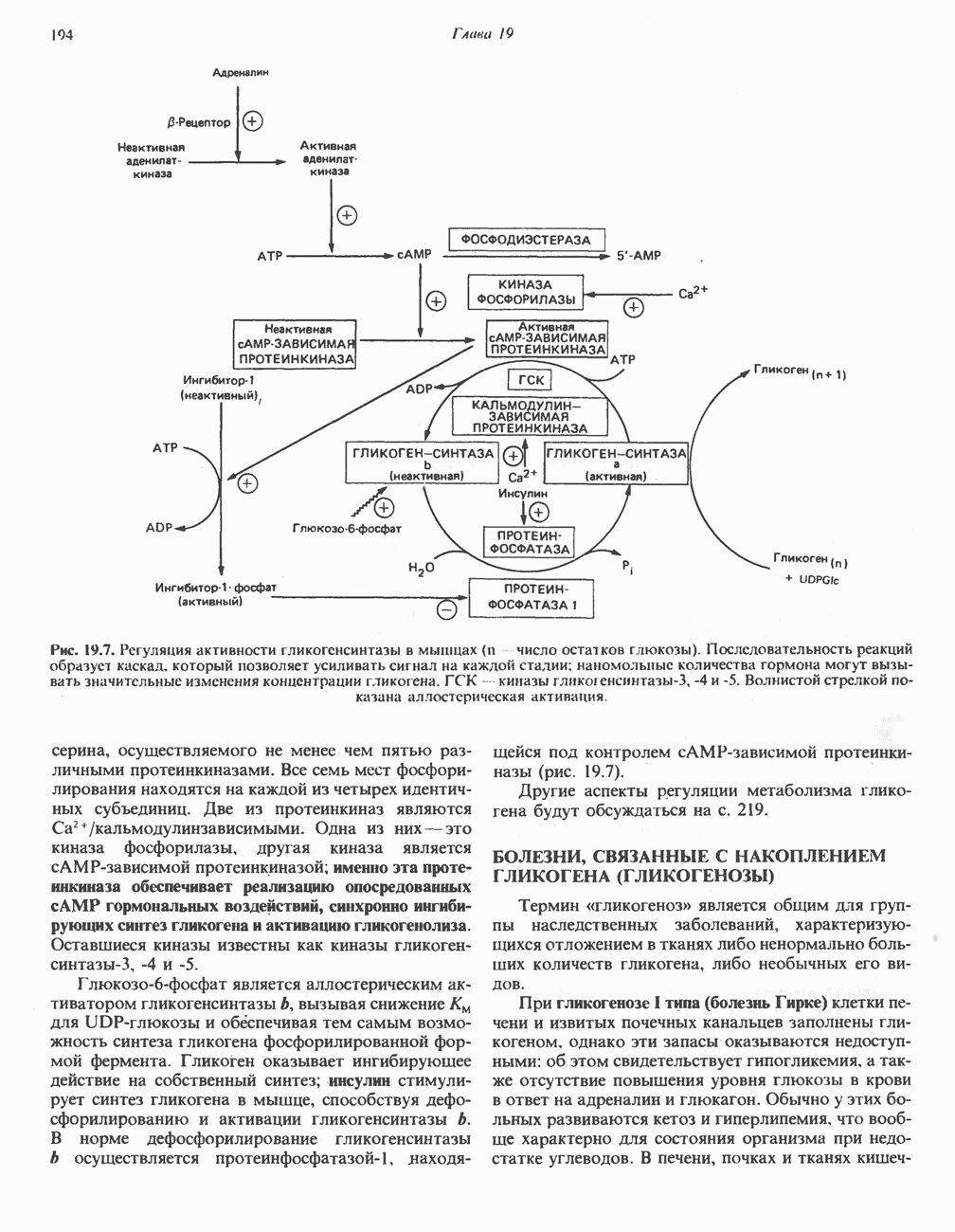
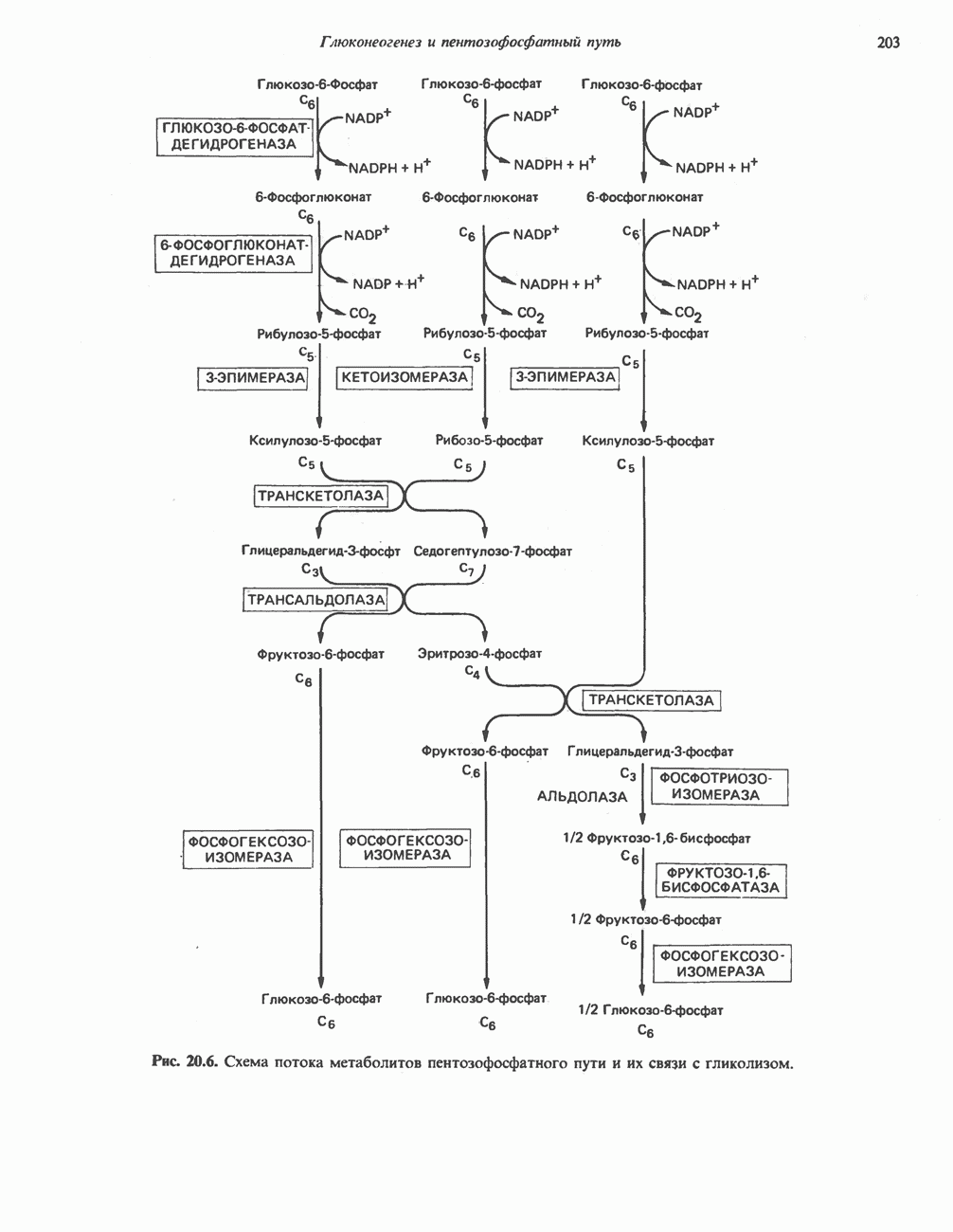
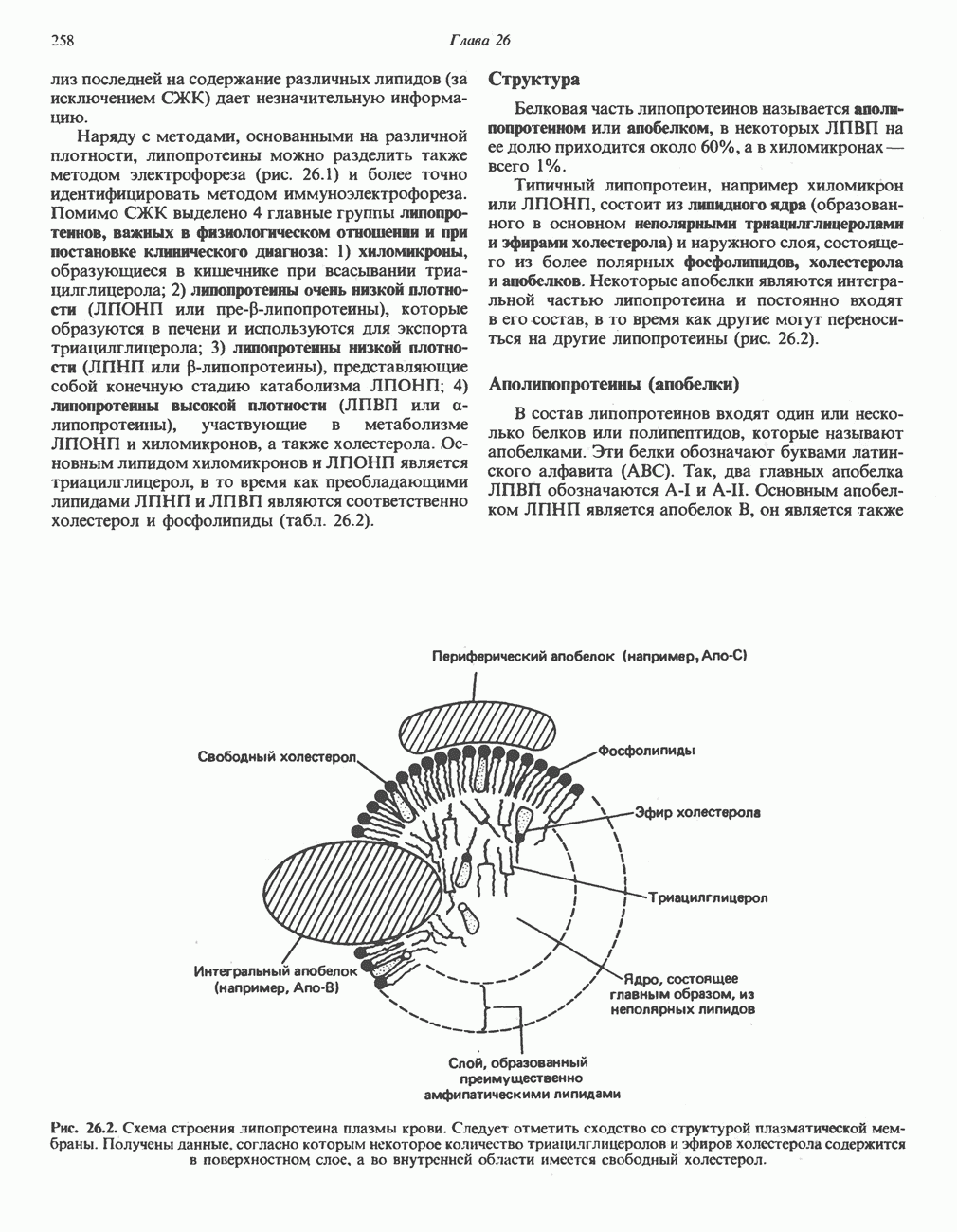
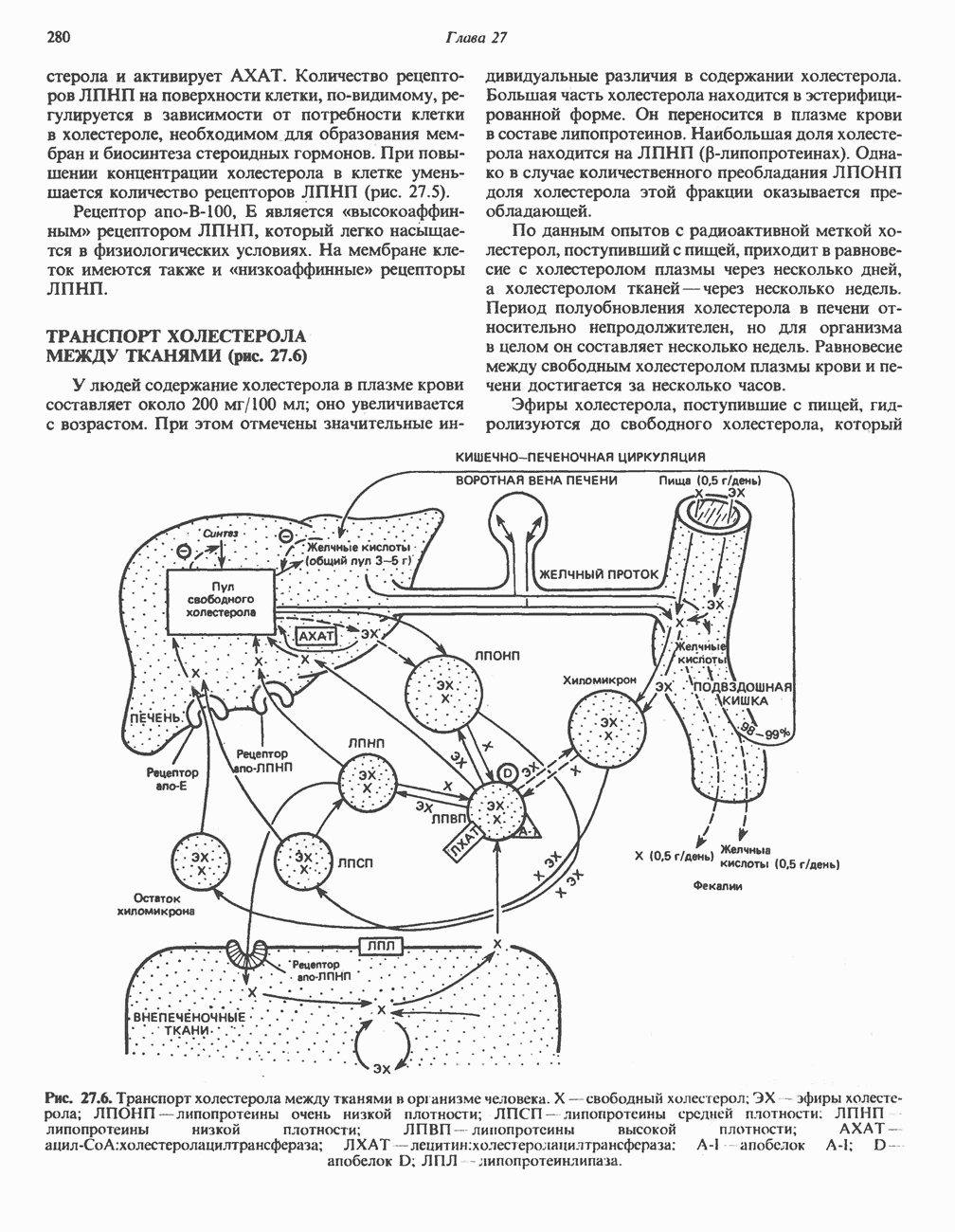
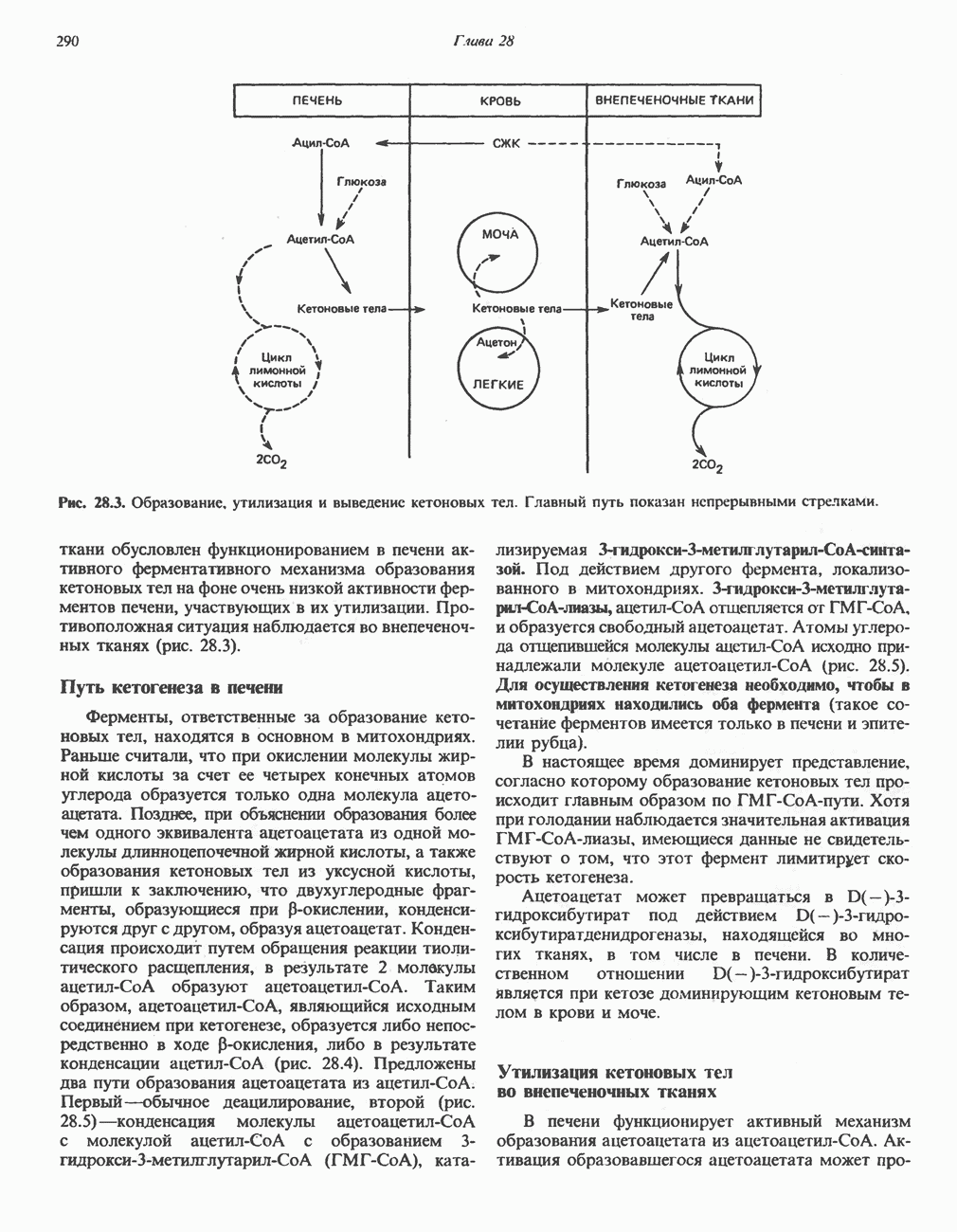
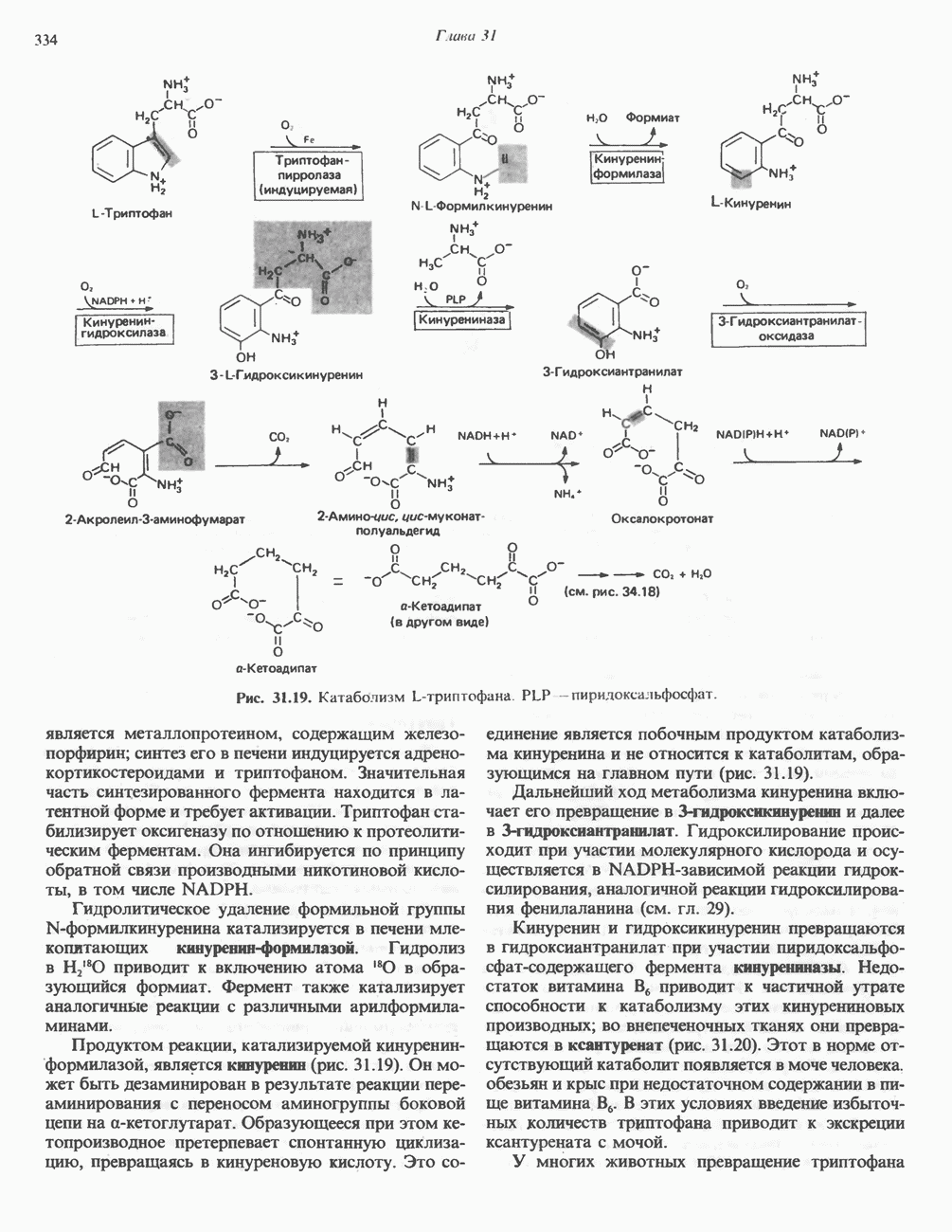
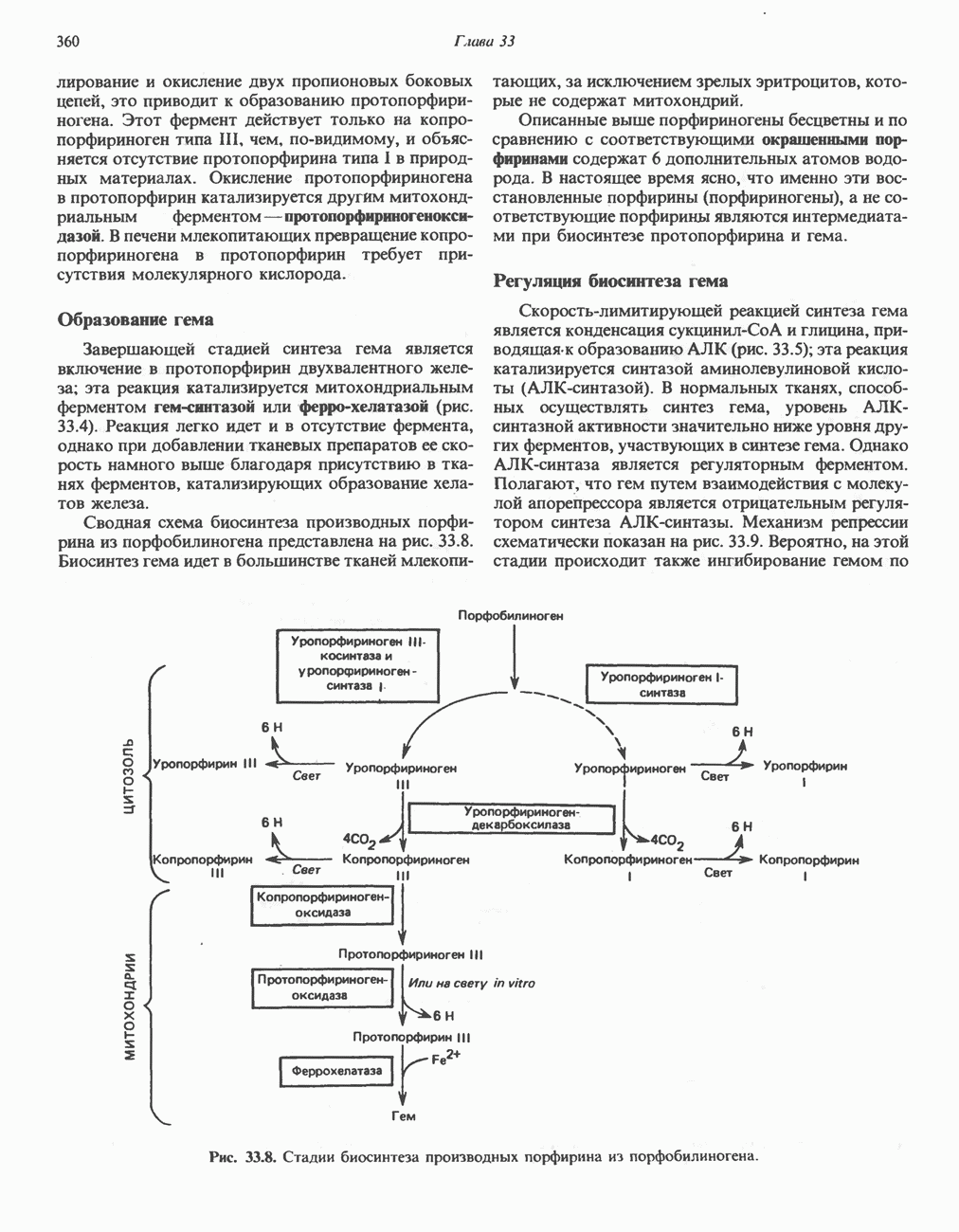
Активация и инактивация гликогенсинтазы (рис. 19.7)
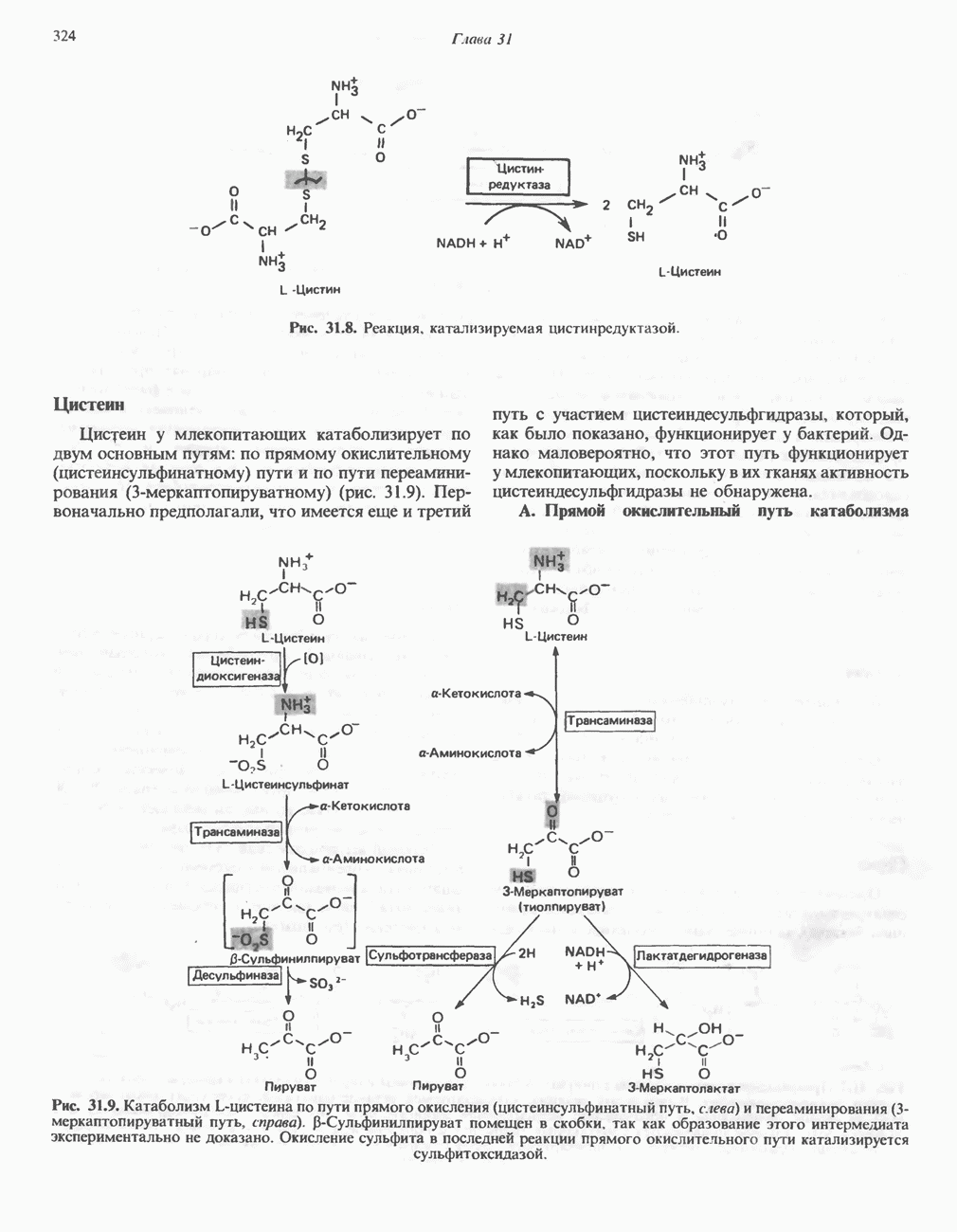
Подобно фосфорилазе, гликогенсинтаза может находиться либо в фосфорилированном, либо в нефосфорилированном состоянии. Однако в отличие от фосфорилазы в этом случае активна дефосфорилированная форма (гликогенсинтаза а), которая может быть инактивирована с образованием гликогенсинтазы b путем фосфорилирования семи остатков

Рис. 19.7. Регуляция активности гликогенсинтазы в мышцах ( число остатков глюкозы). Последовательность реакций образует каскад, который позволяет усиливать сигнал на каждой стадии; наномольные количества гормона могут вызывать значительные изменения концентрации гликогена. ГСК — киназы глнкогенсинтазы-3, -4 и -5. Волнистой стрелкой показана аллостерическая активация.
число остатков глюкозы). Последовательность реакций образует каскад, который позволяет усиливать сигнал на каждой стадии; наномольные количества гормона могут вызывать значительные изменения концентрации гликогена. ГСК — киназы глнкогенсинтазы-3, -4 и -5. Волнистой стрелкой показана аллостерическая активация.
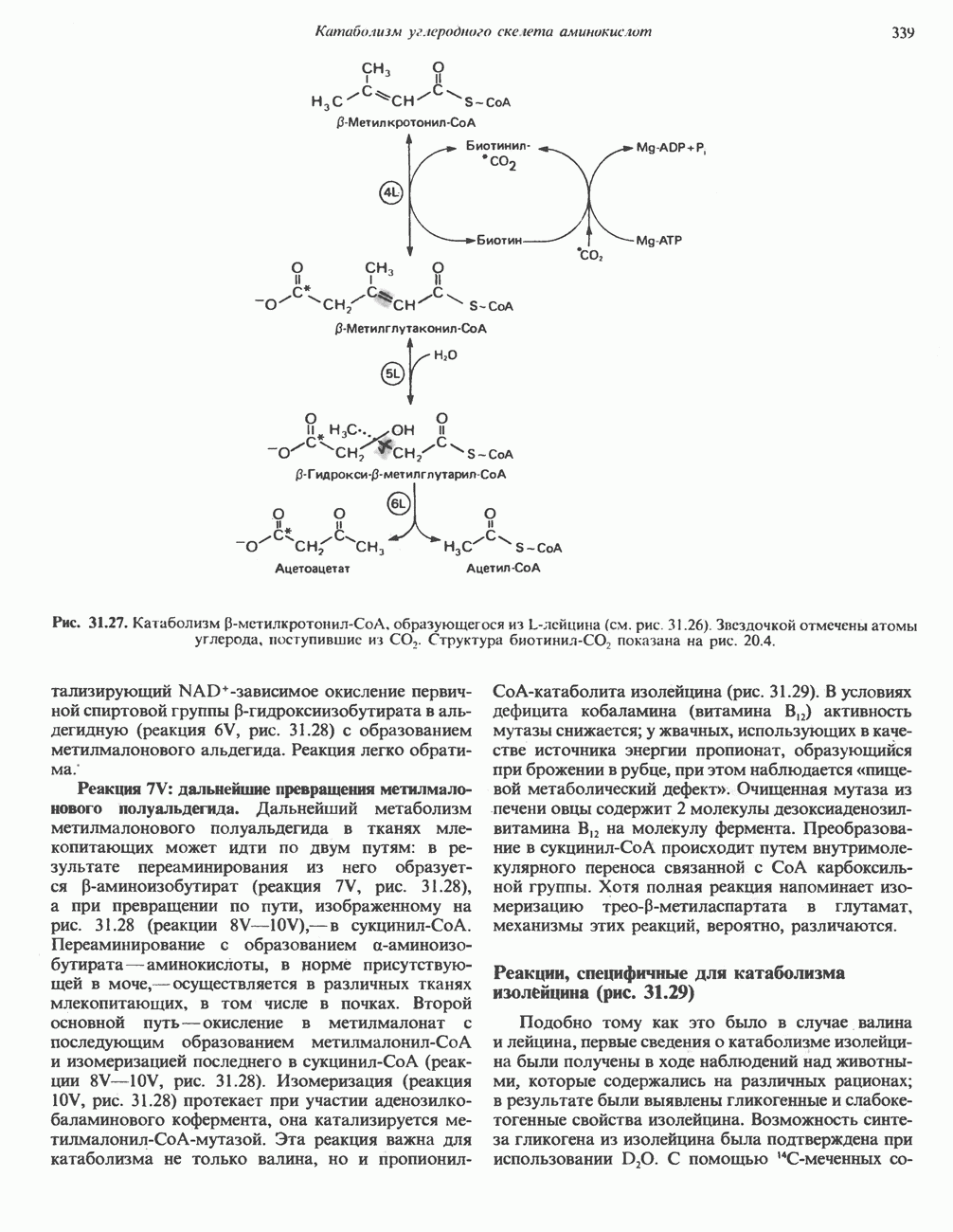
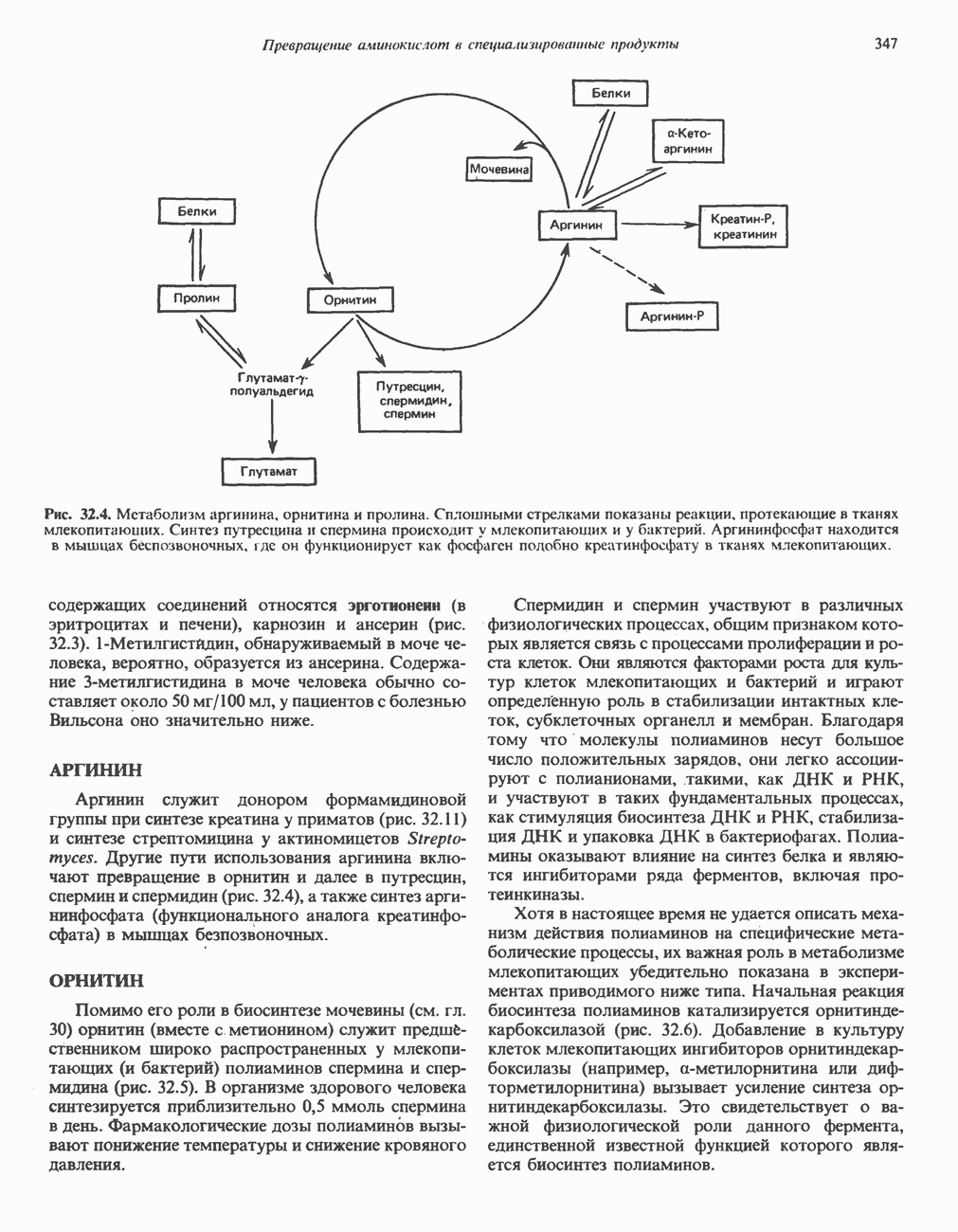
серина, осуществляемого не менее чем пятью различными протеинкиназами. Все семь мест фосфорилирования находятся на каждой из четырех идентичных субъединиц. Две из протеинкиназ являются  кальмодулинзависимыми. Одна из них — это киназа фосфорилазы, другая киназа является сАМР-зависимой протеинкиназой; именно эта протеинкиназа обеспечивает реализацию опосредованных сАМР гормональных воздействий, синхронно ингибирующих синтез гликогена и активацию гликогенолиза. Оставшиеся киназы известны как киназы гликоген-синтазы-3, -4 и -5.
кальмодулинзависимыми. Одна из них — это киназа фосфорилазы, другая киназа является сАМР-зависимой протеинкиназой; именно эта протеинкиназа обеспечивает реализацию опосредованных сАМР гормональных воздействий, синхронно ингибирующих синтез гликогена и активацию гликогенолиза. Оставшиеся киназы известны как киназы гликоген-синтазы-3, -4 и -5.
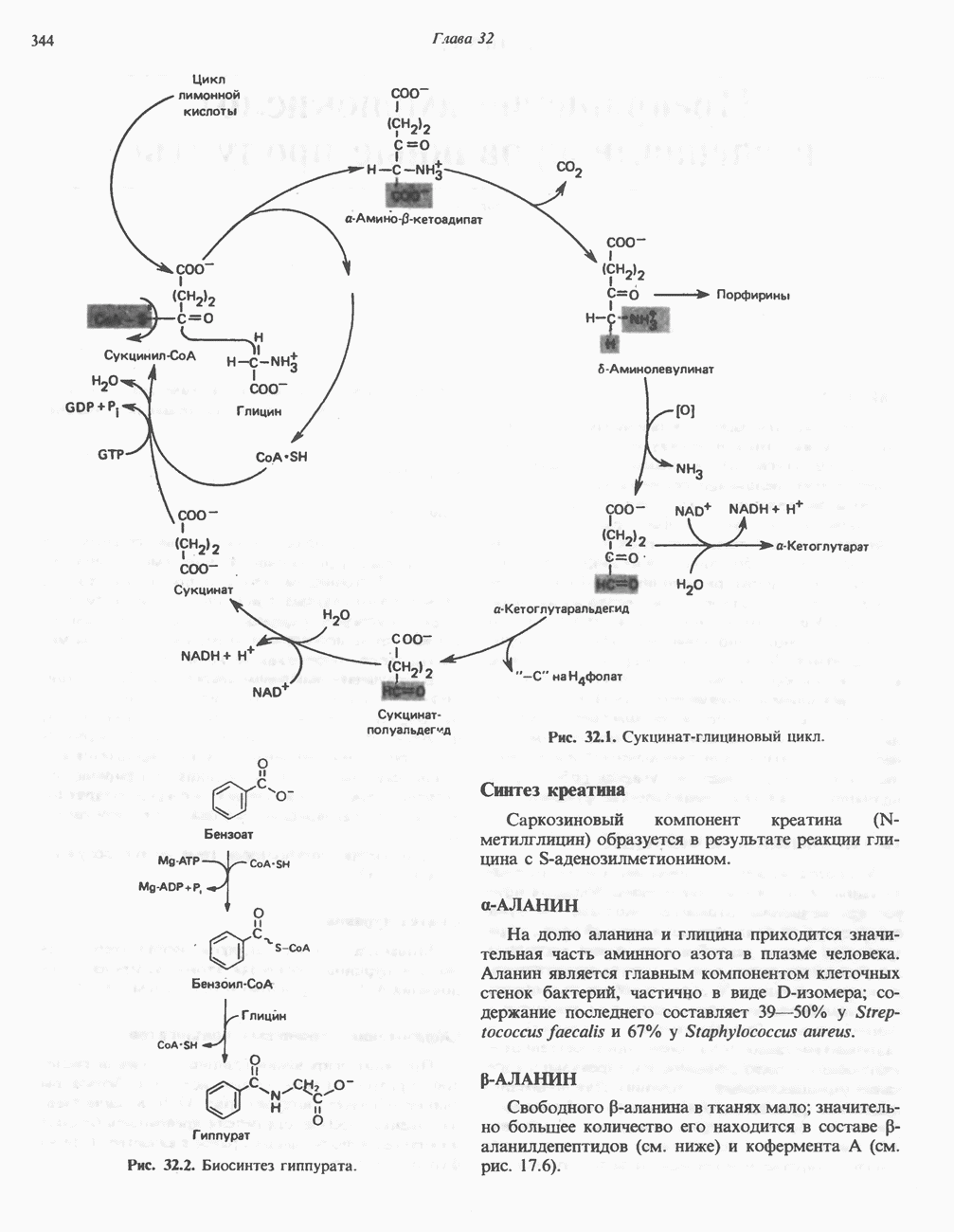
Глкжозо-6-фосфат является аллостерическим активатором гликогенсинтазы b, вызывая снижение  для UDP-глюкозы и обеспечивая тем самым возможность синтеза гликогена фосфорилированной формой фермента. Гликоген оказывает ингибирующее действие на собственный синтез; инсулин стимулирует синтез гликогена в мышце, способствуя дефосфорилированию и активации гликогенсинтазы b. В норме дефосфорилирование гликогенсинтазы b осуществляется протеинфосфатазой-1, яаходящейся под контролем сАМР-зависимой протеинкиназы (рис. 19.7).
для UDP-глюкозы и обеспечивая тем самым возможность синтеза гликогена фосфорилированной формой фермента. Гликоген оказывает ингибирующее действие на собственный синтез; инсулин стимулирует синтез гликогена в мышце, способствуя дефосфорилированию и активации гликогенсинтазы b. В норме дефосфорилирование гликогенсинтазы b осуществляется протеинфосфатазой-1, яаходящейся под контролем сАМР-зависимой протеинкиназы (рис. 19.7).
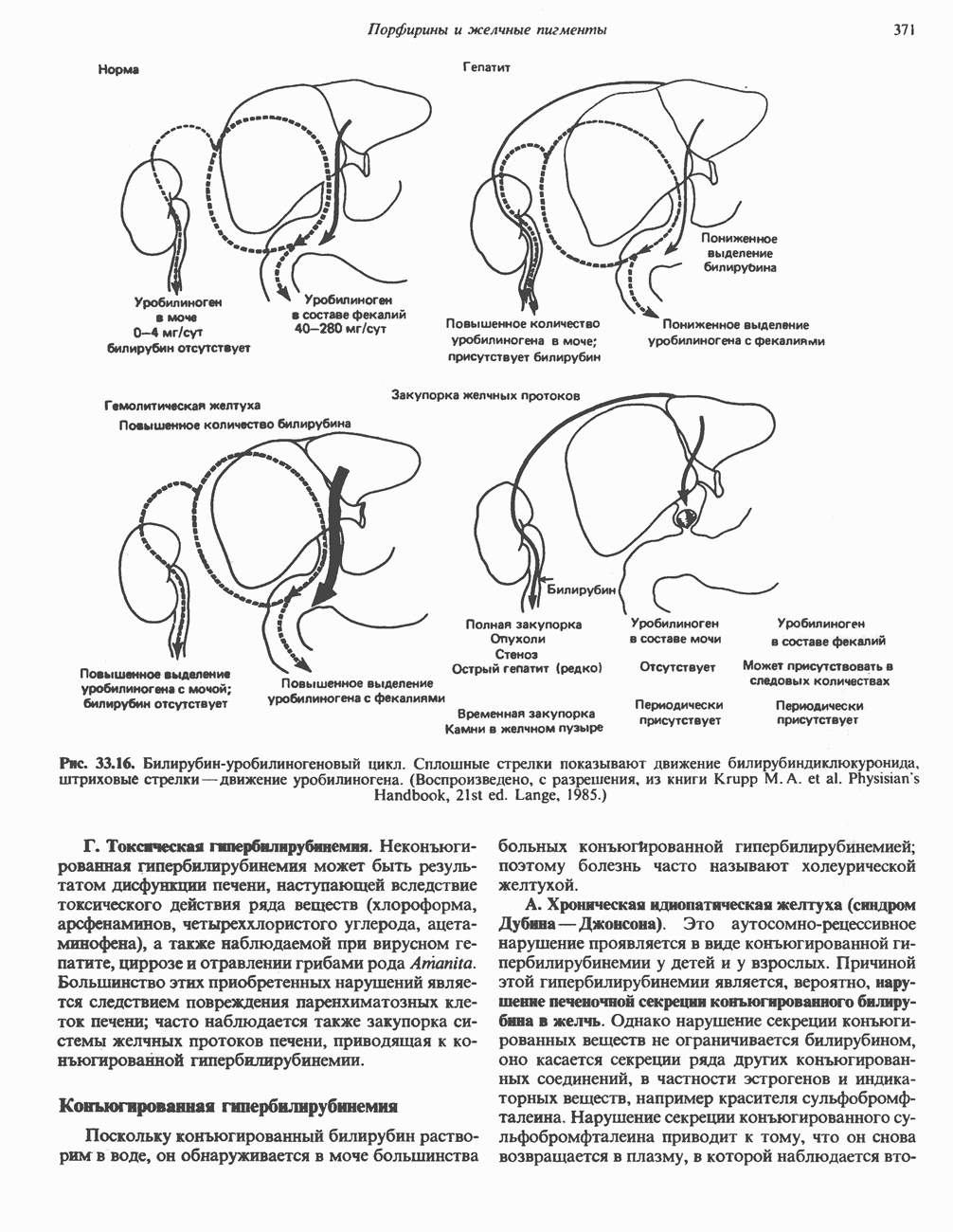
Другие аспекты регуляции метаболизма гликогена будут обсуждаться на с. 219.