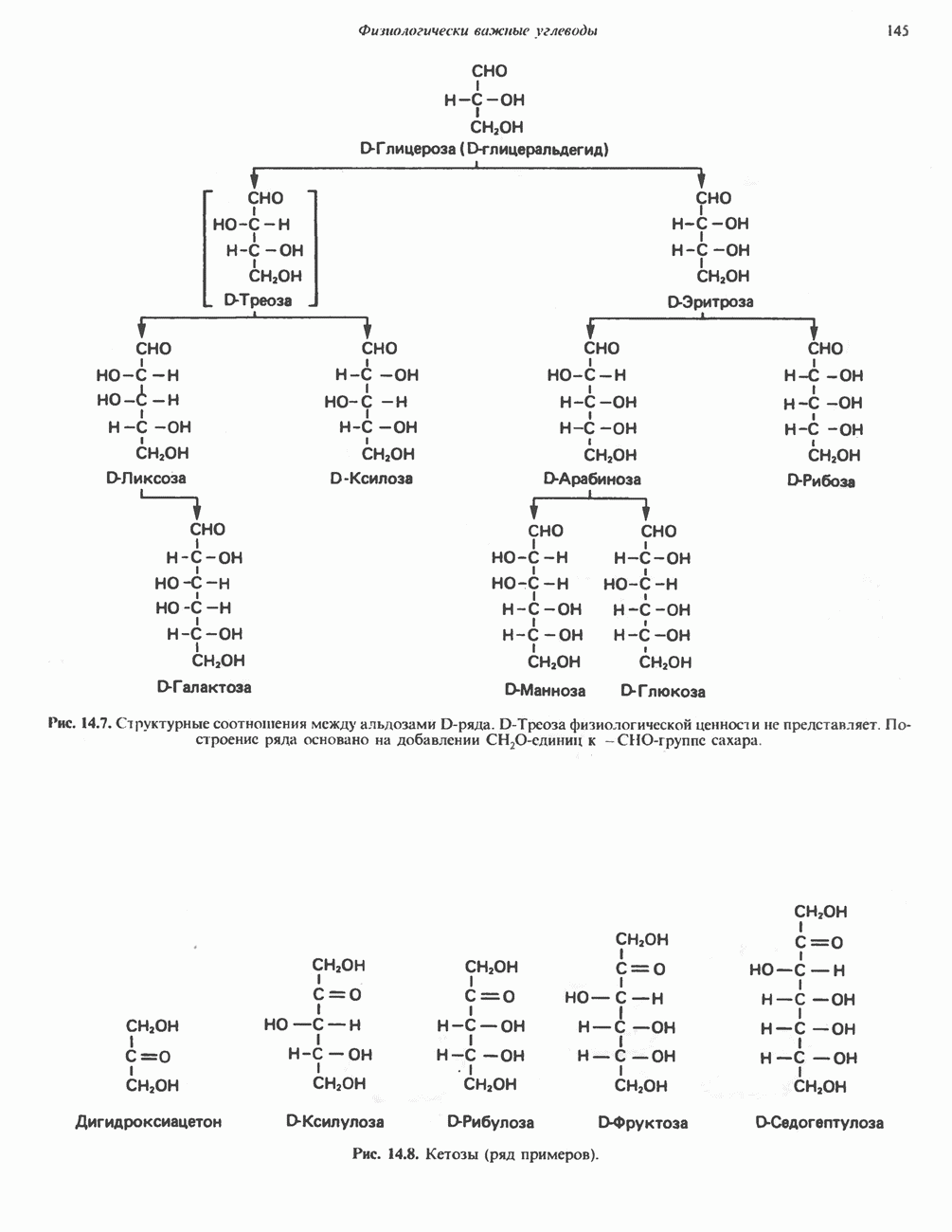
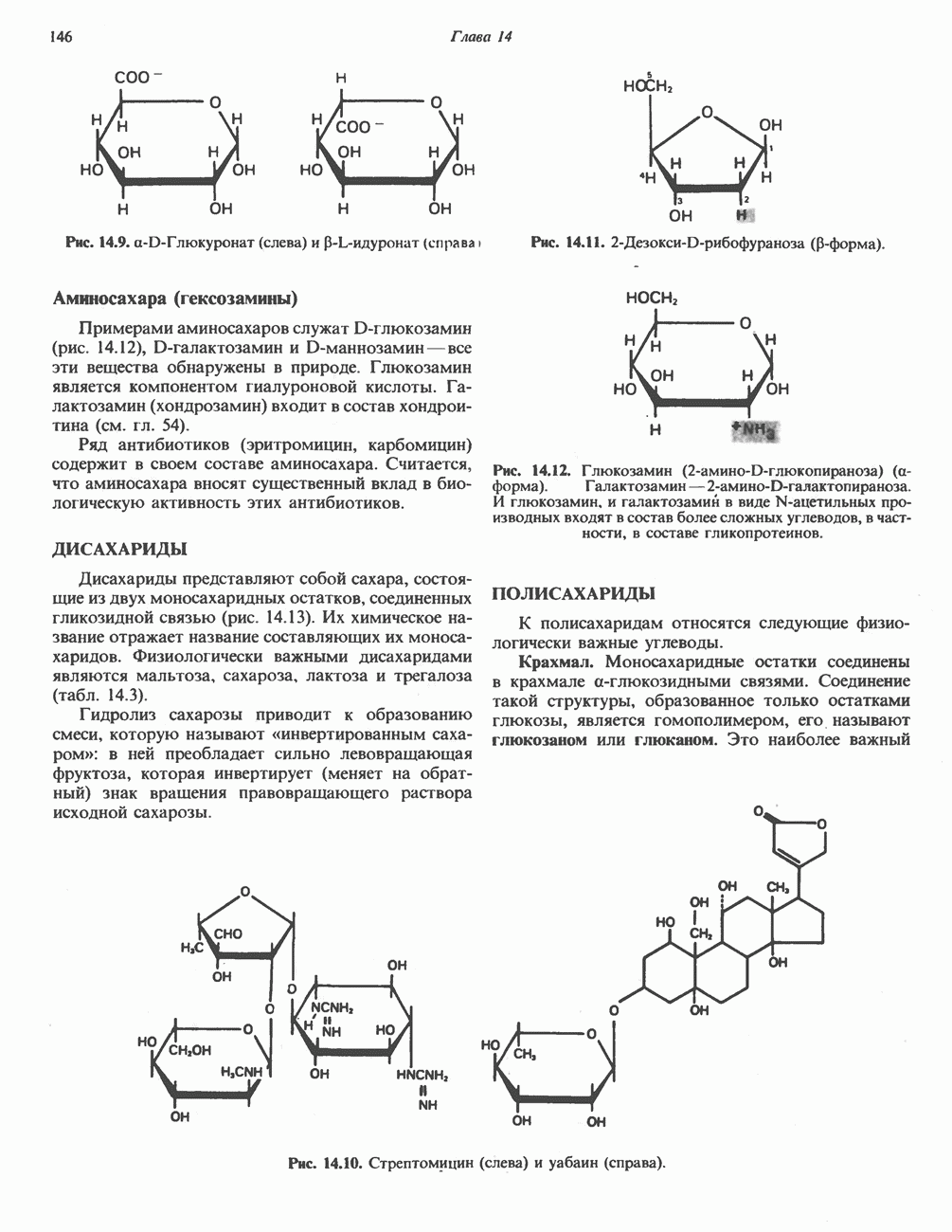
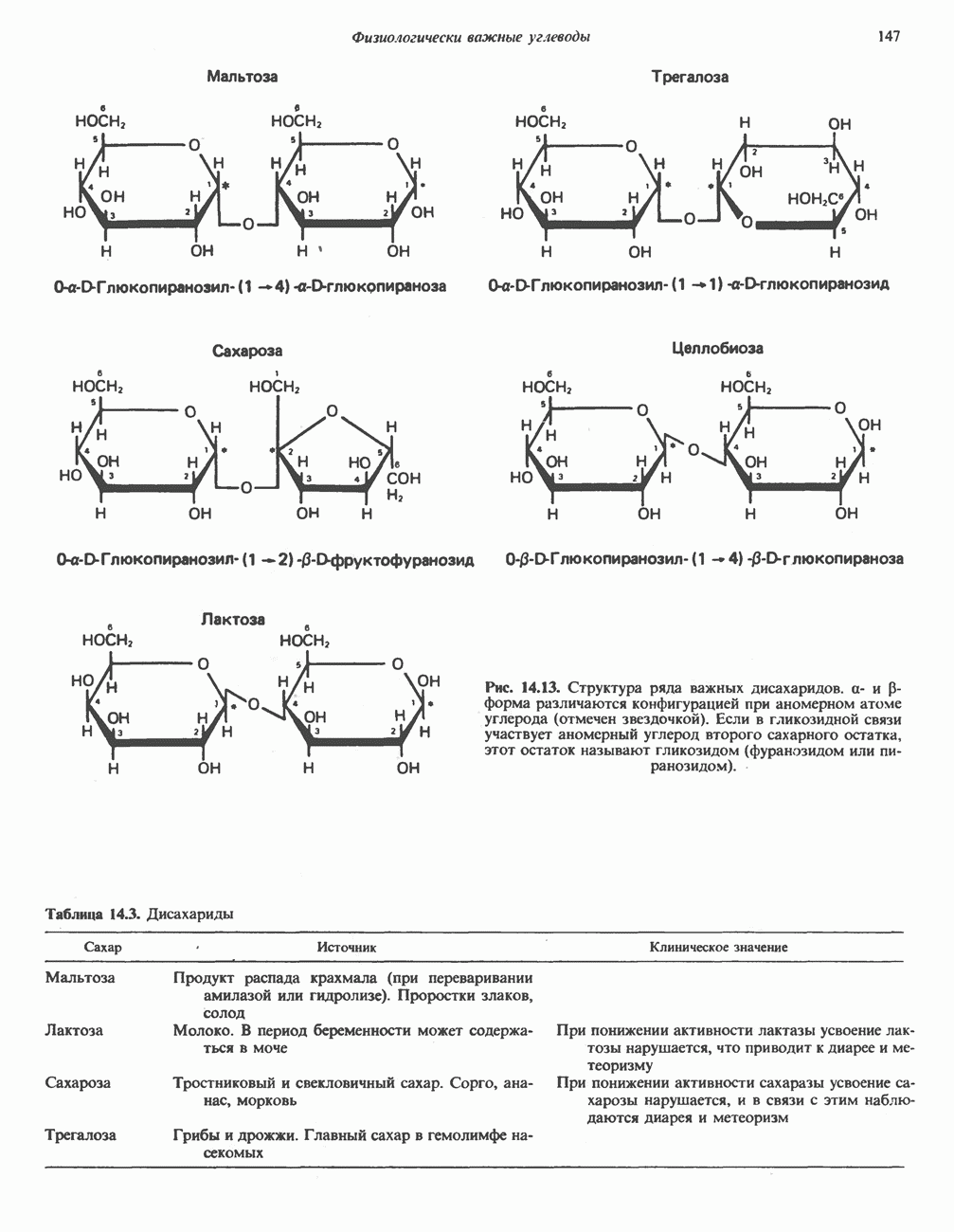
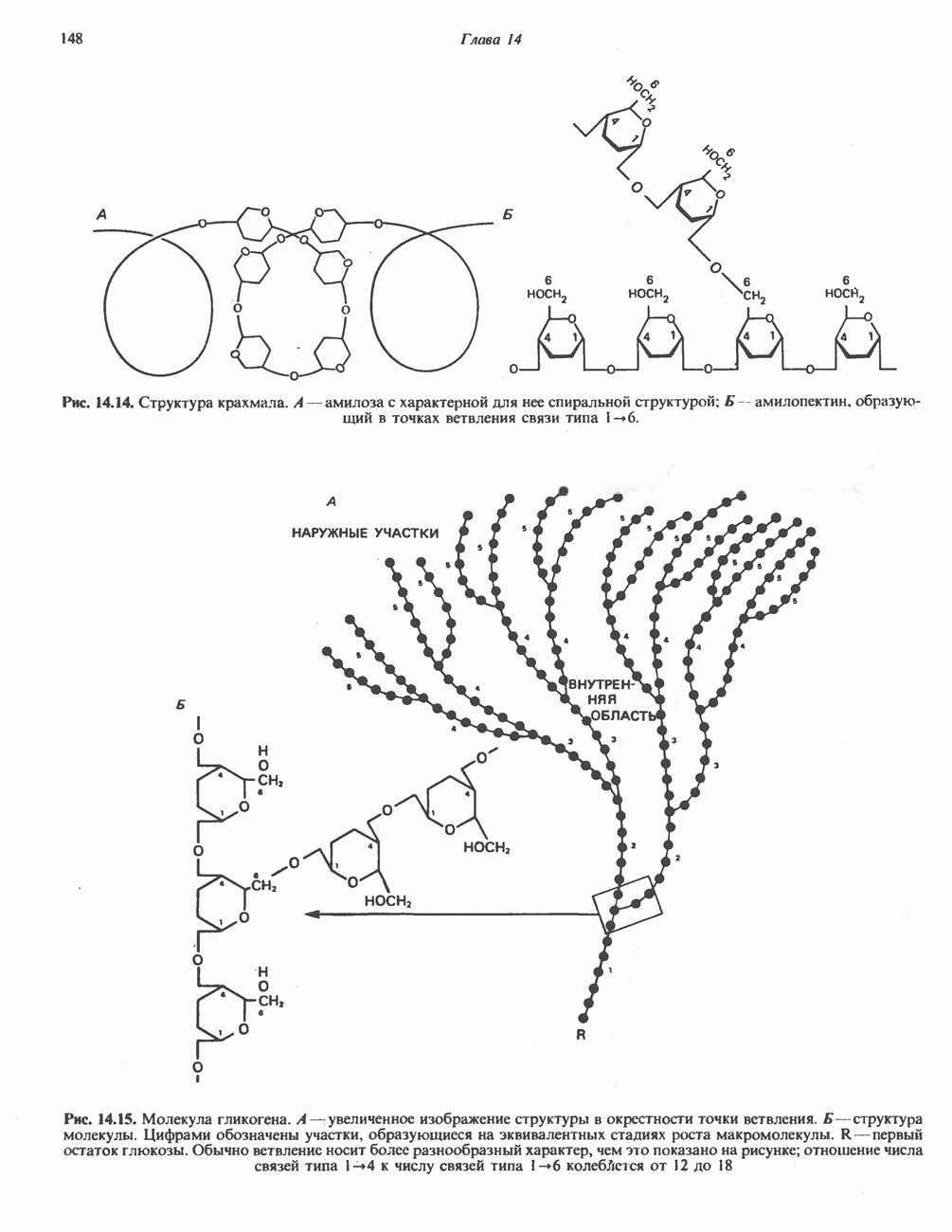
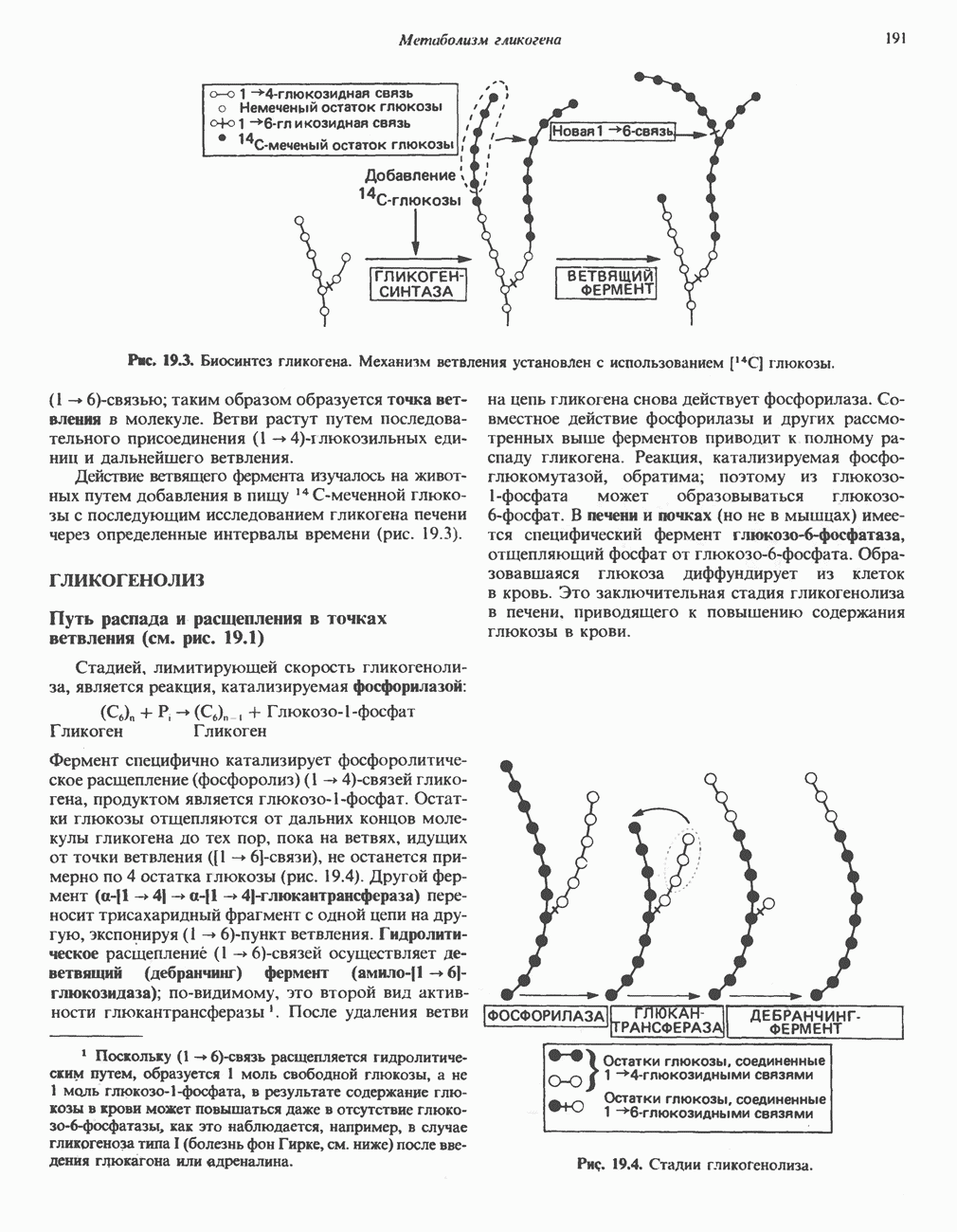
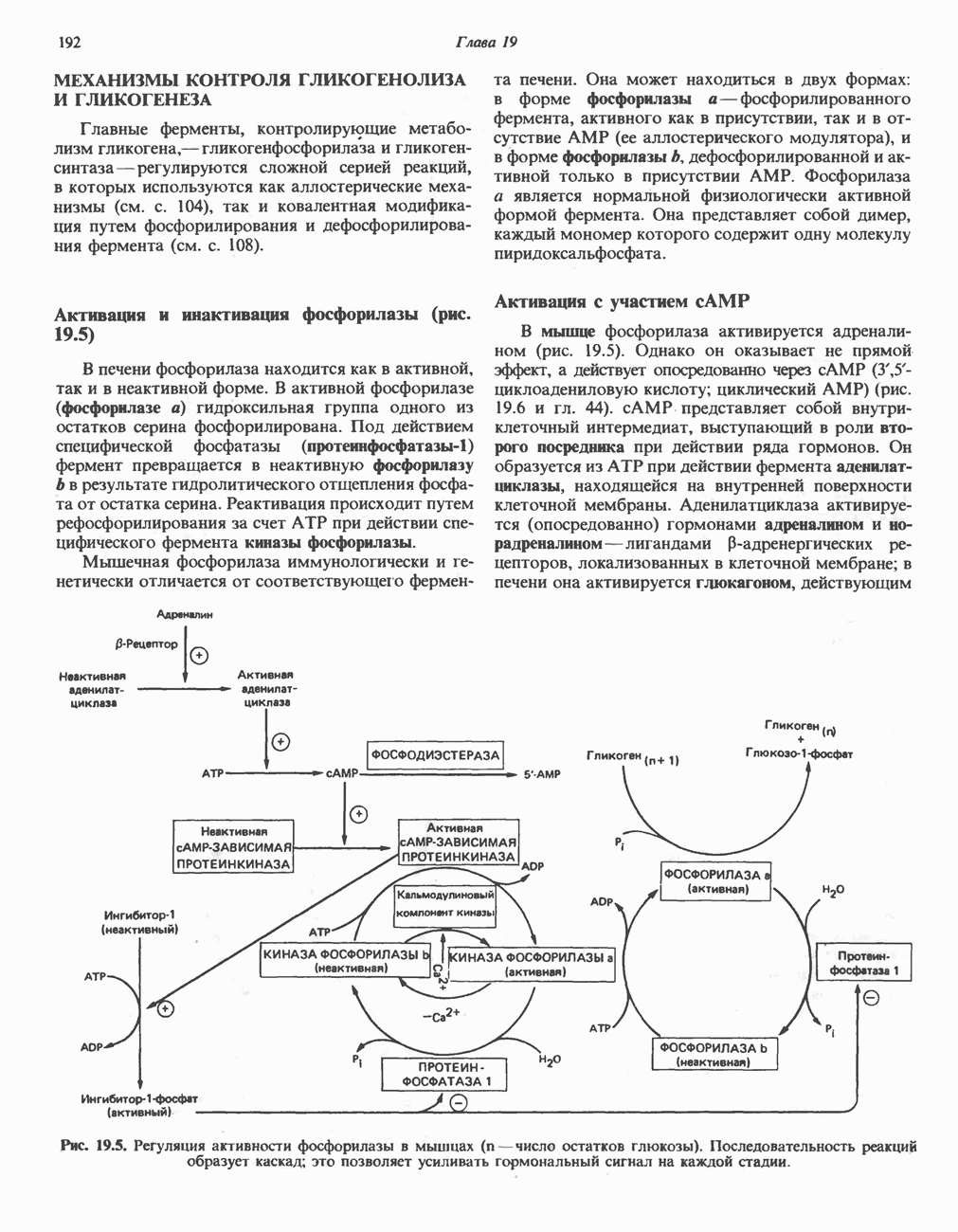
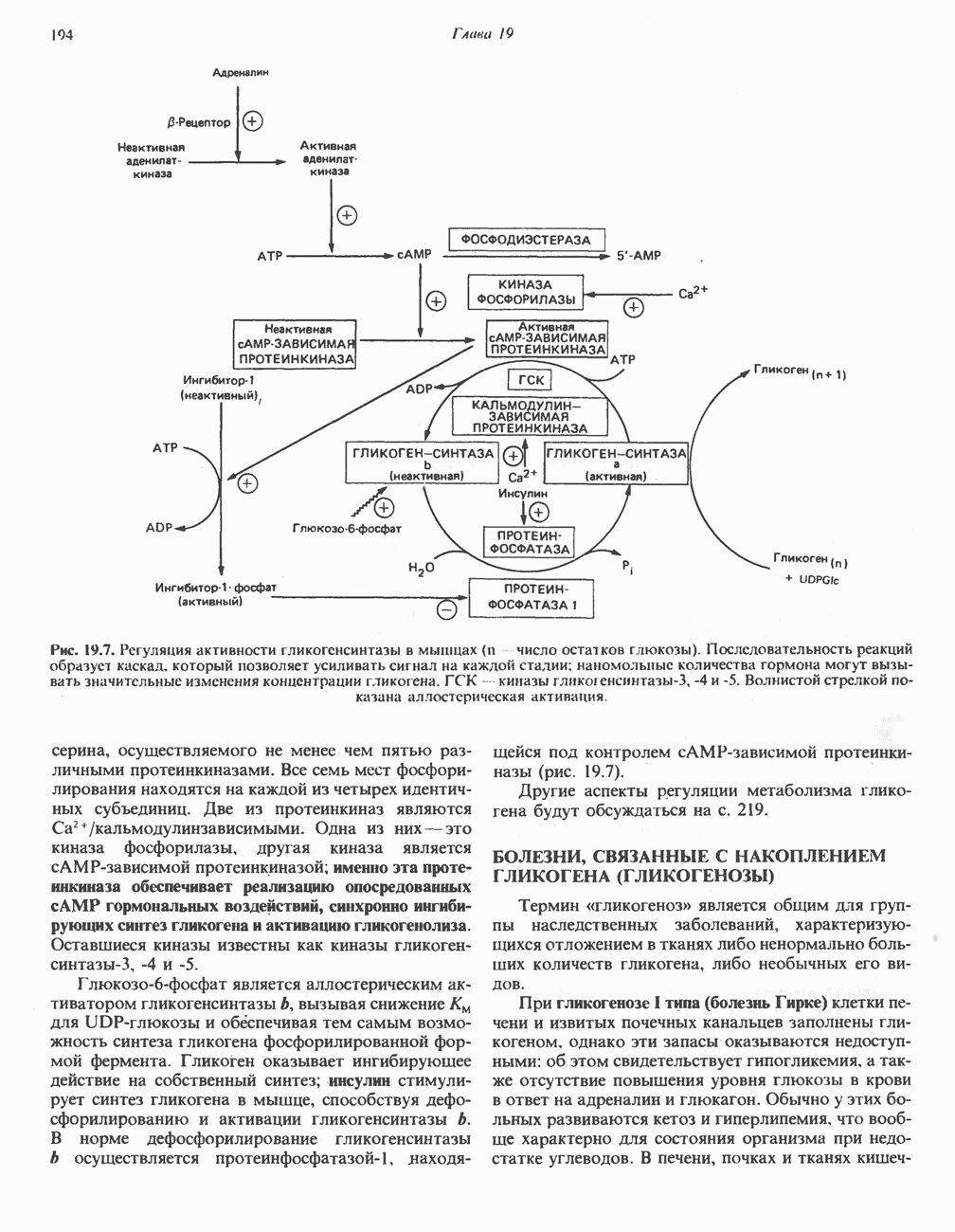
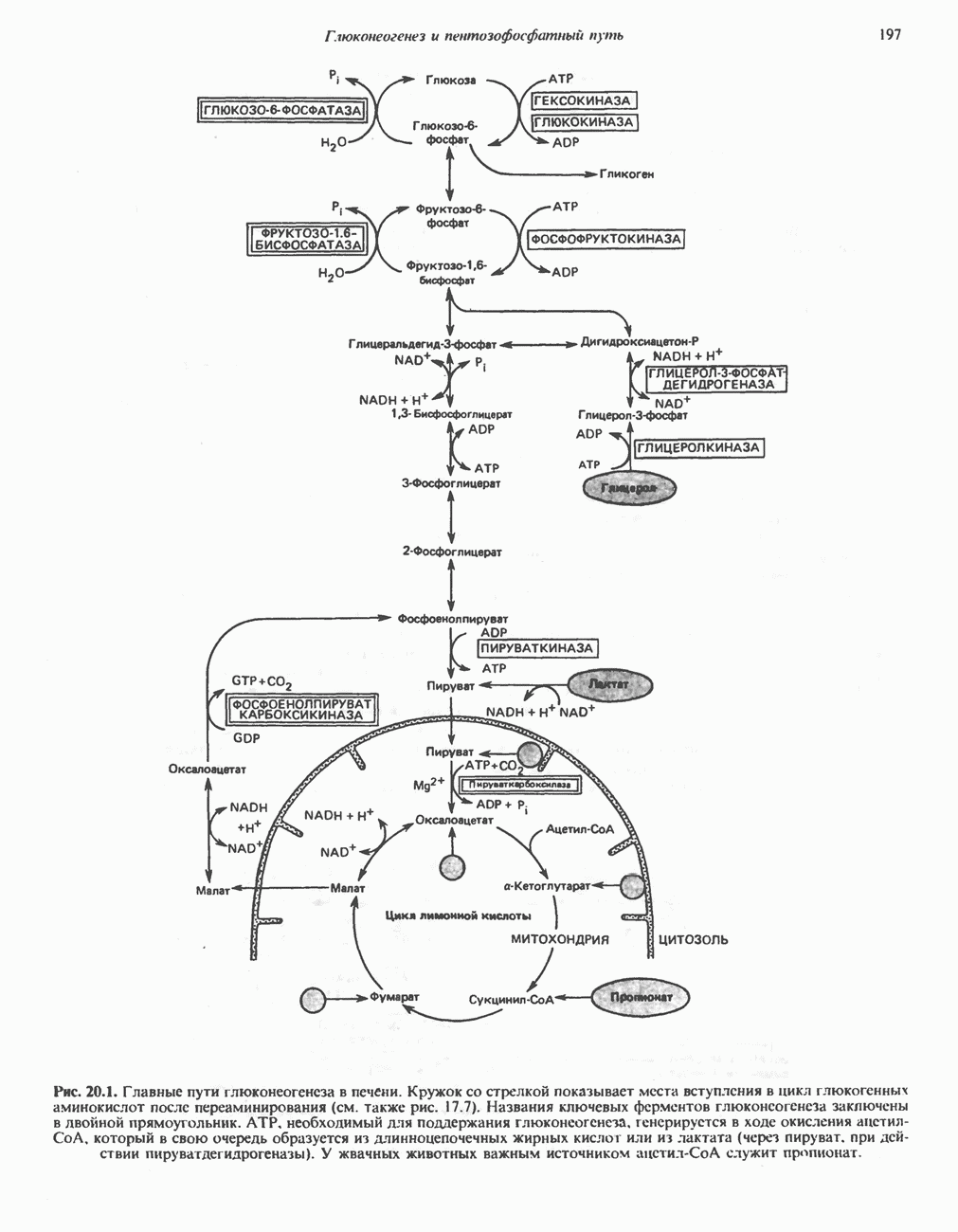
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
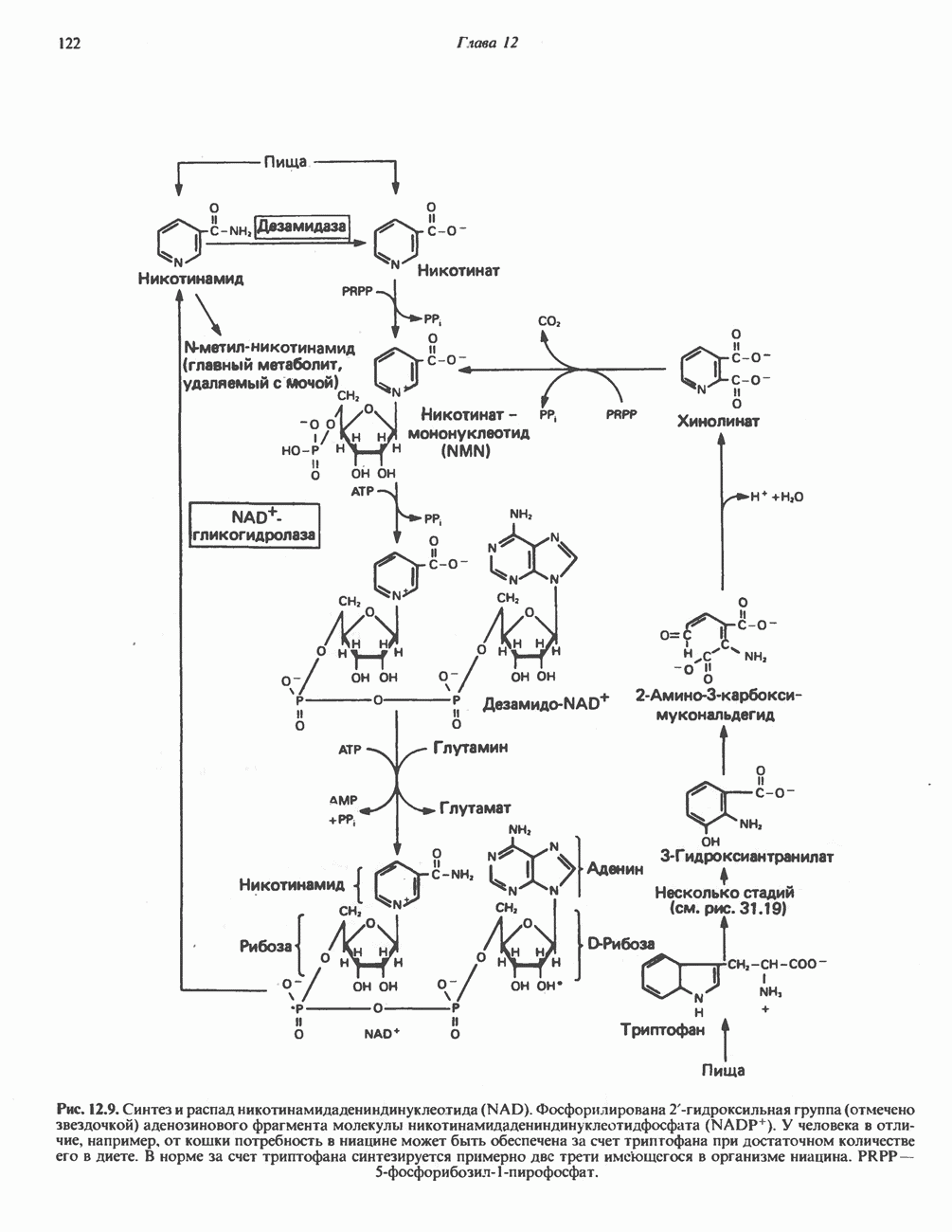
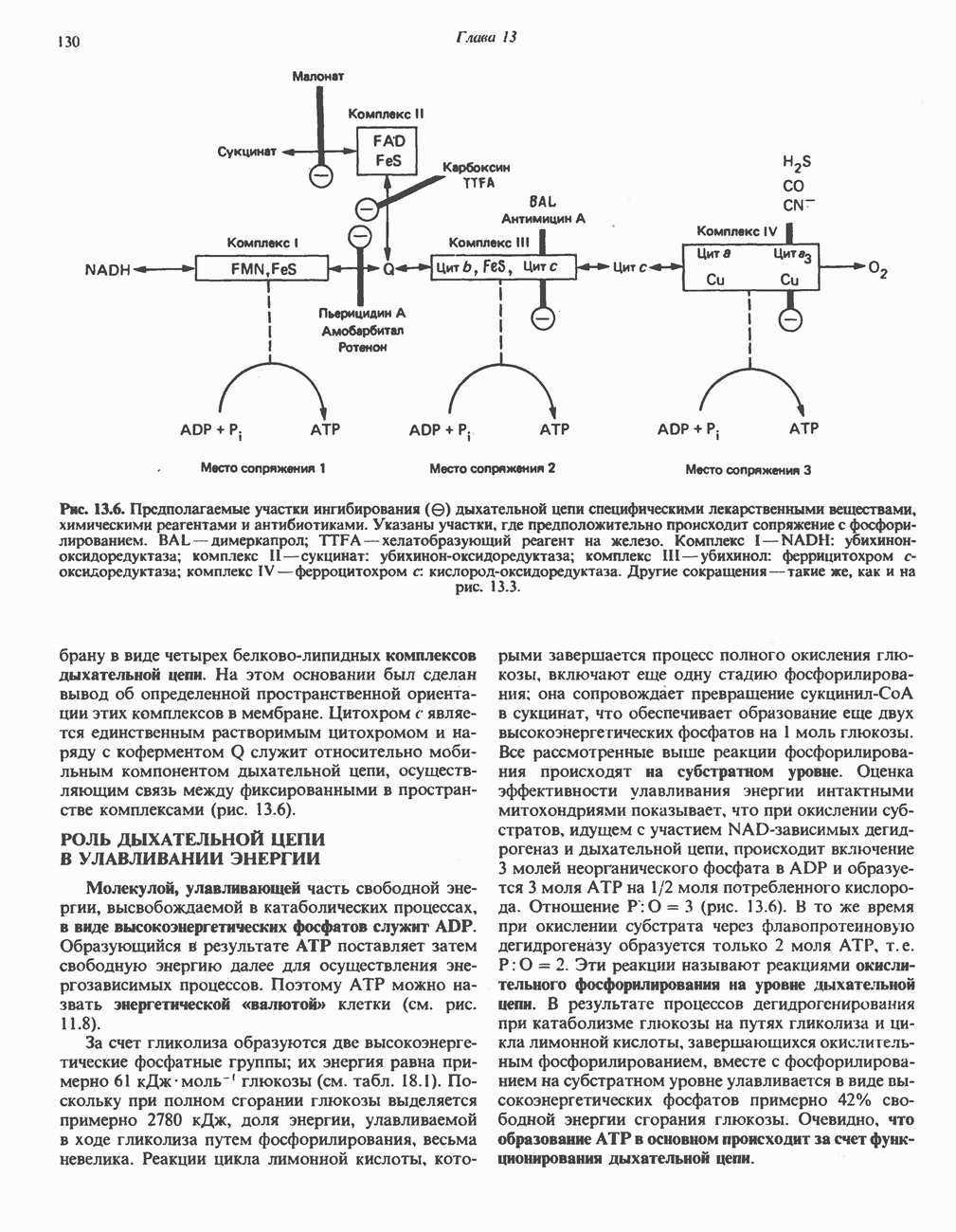
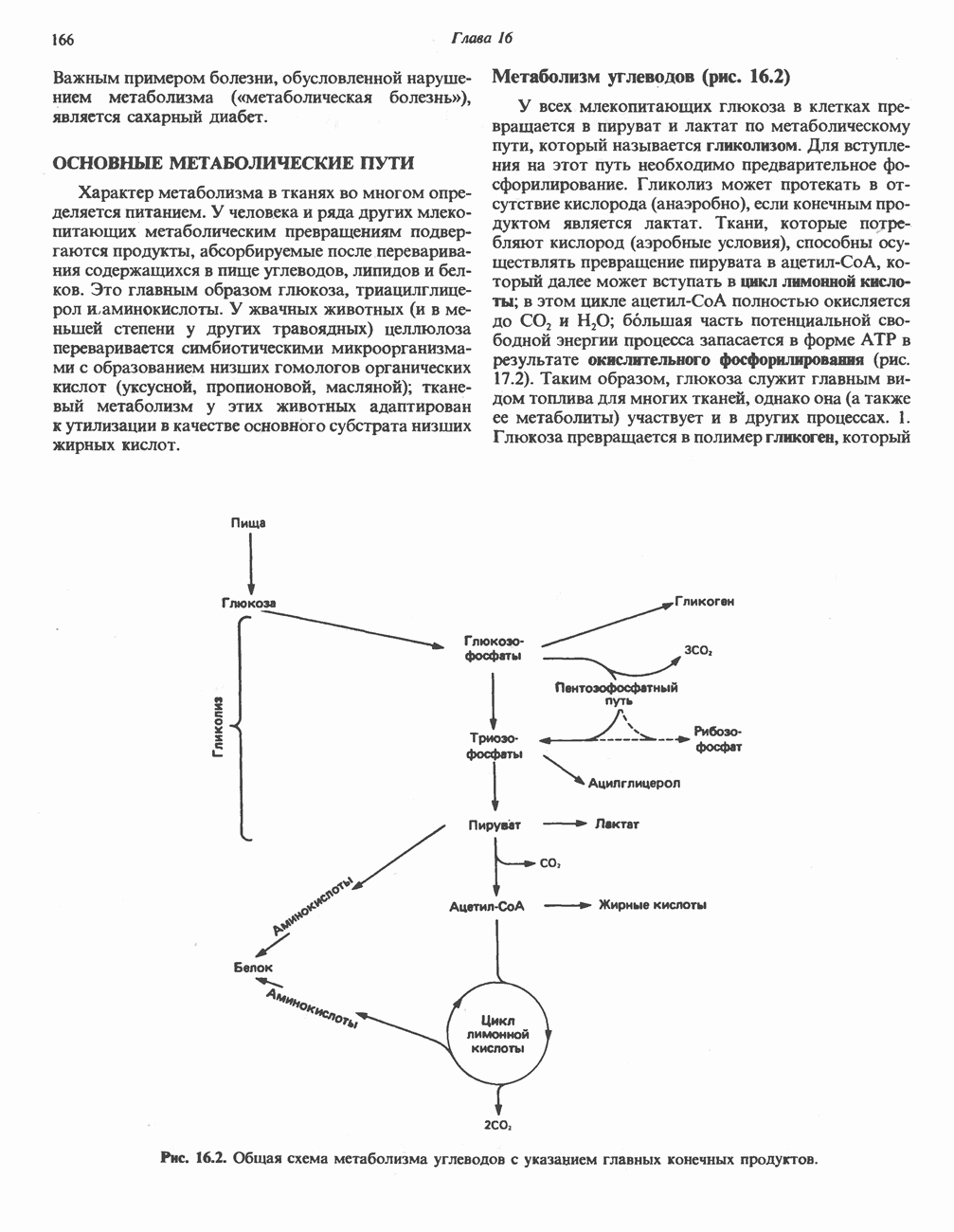
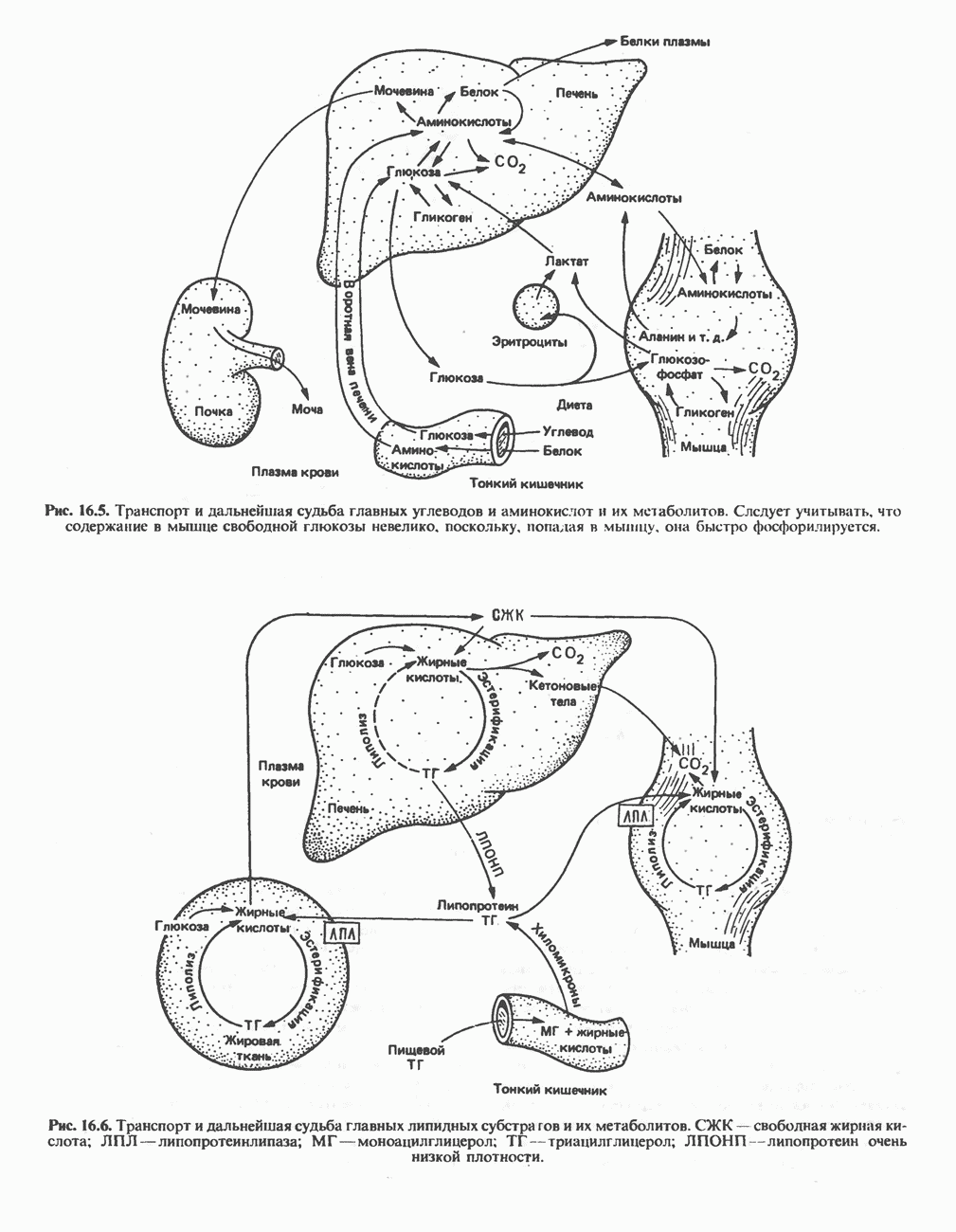
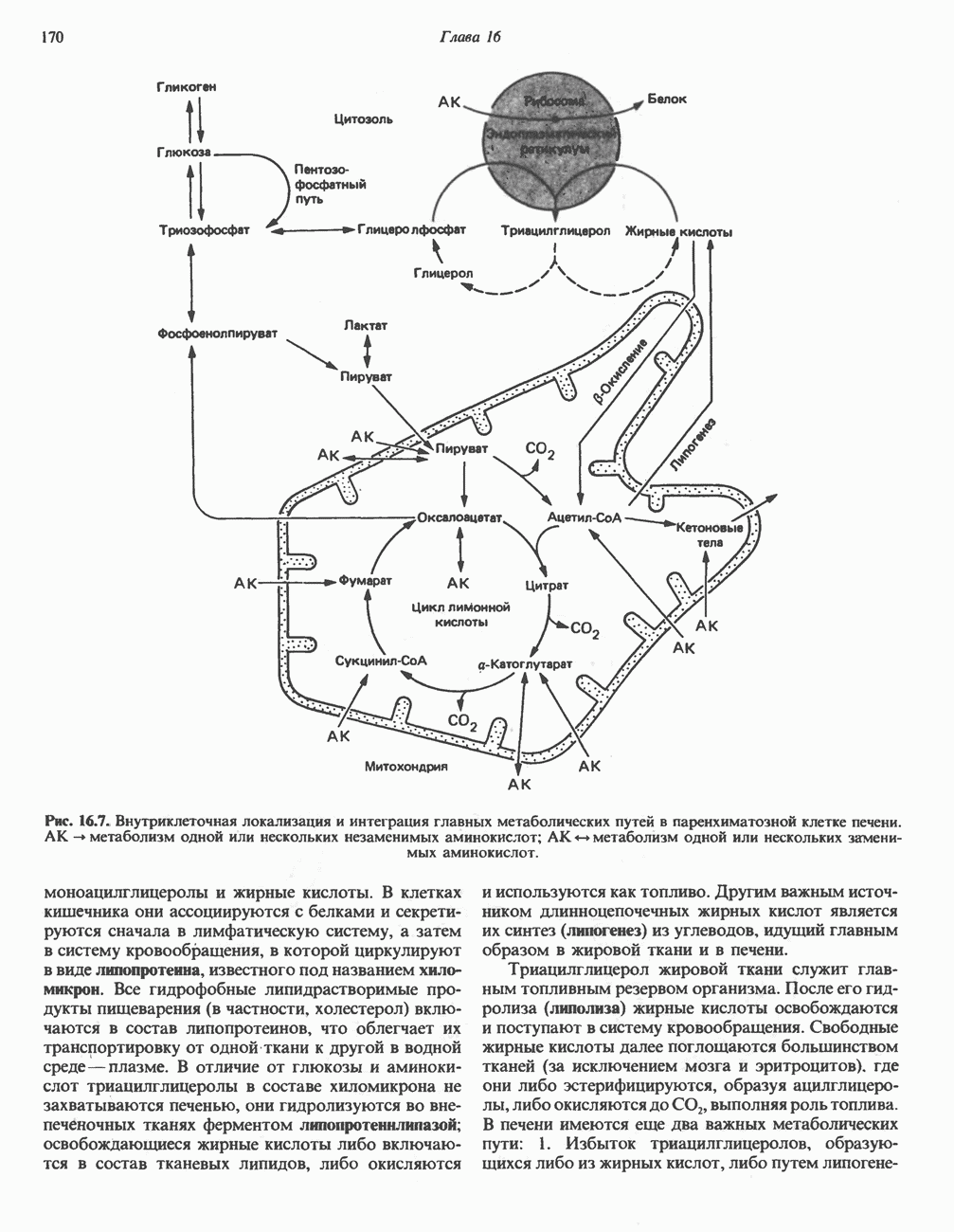
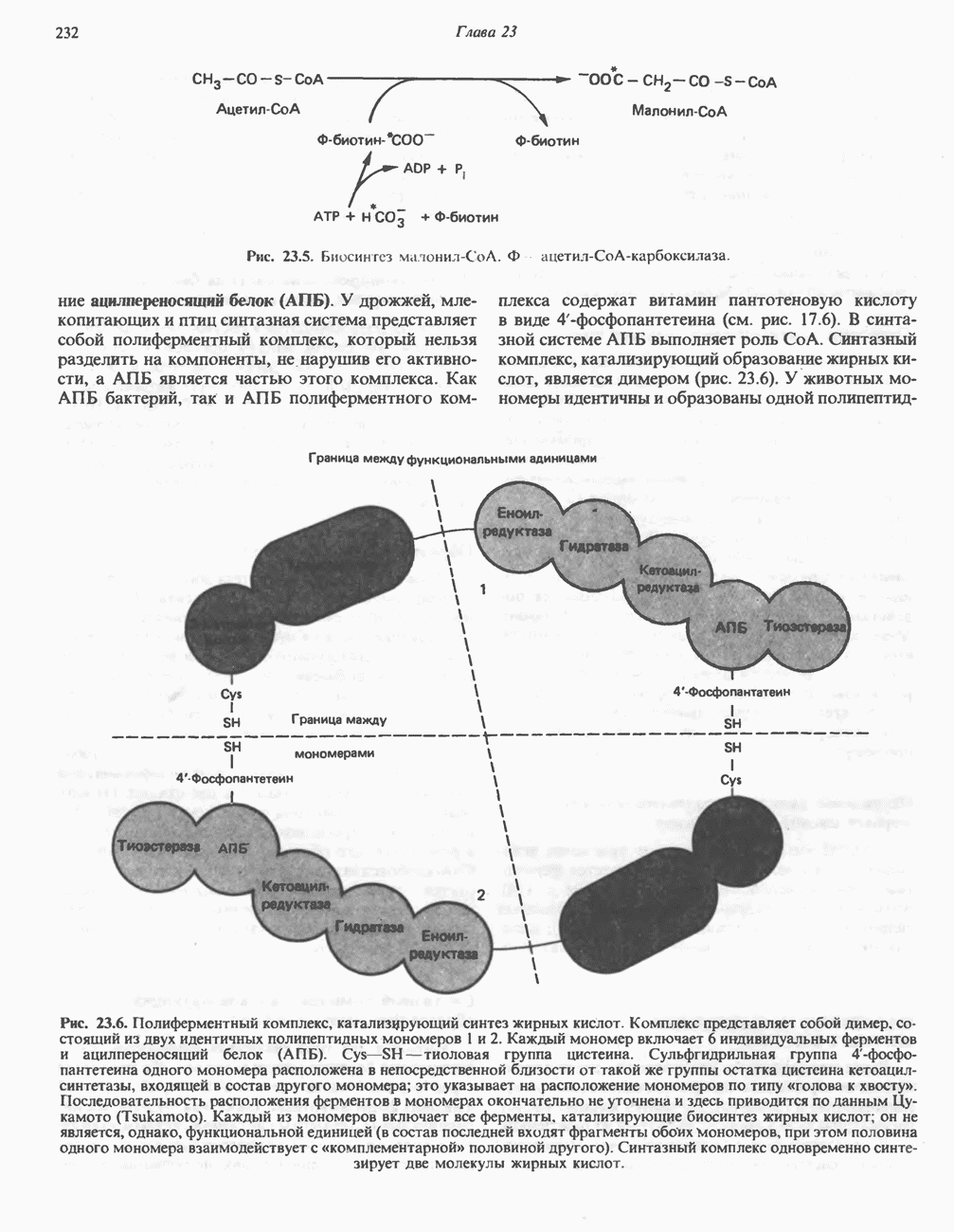
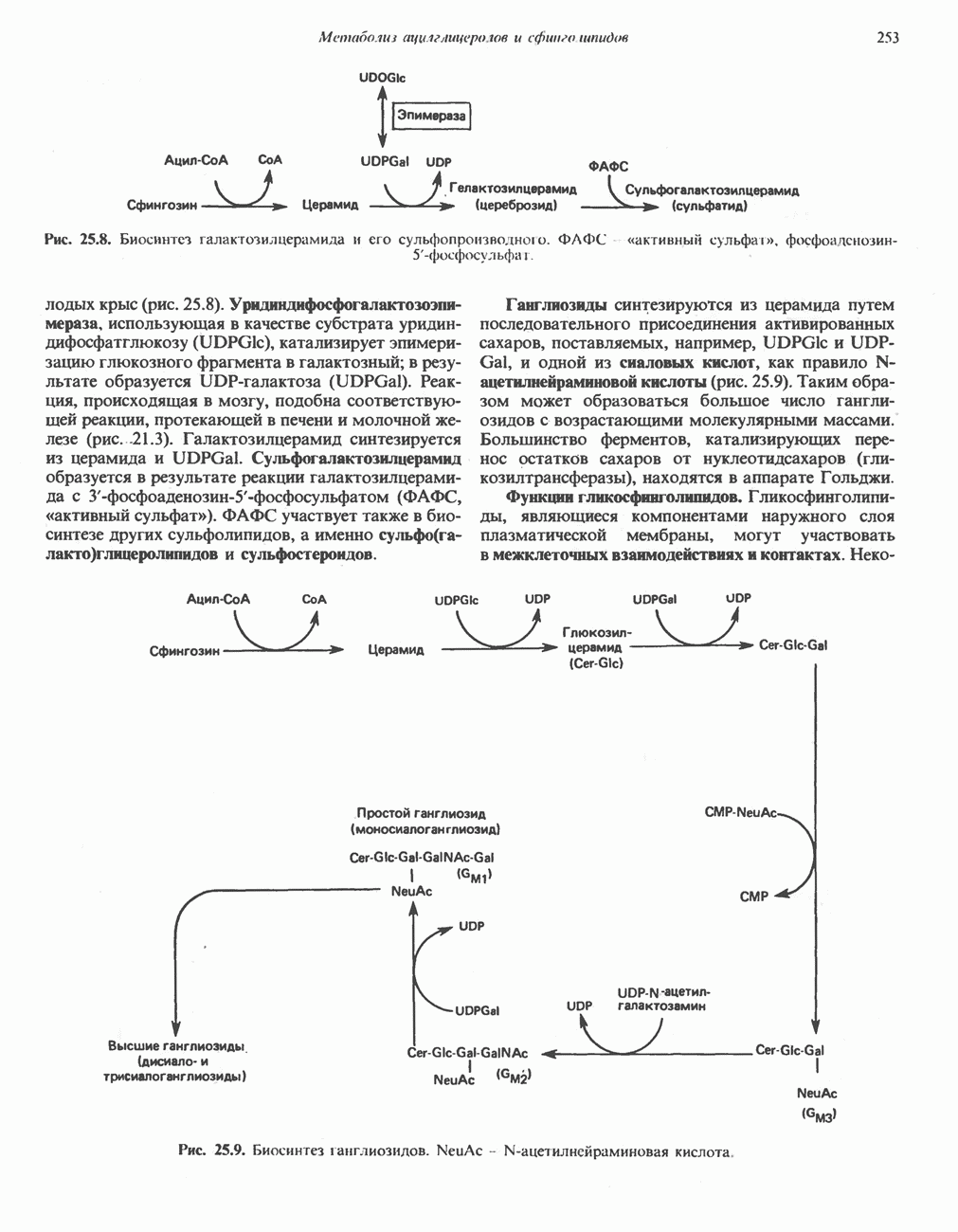
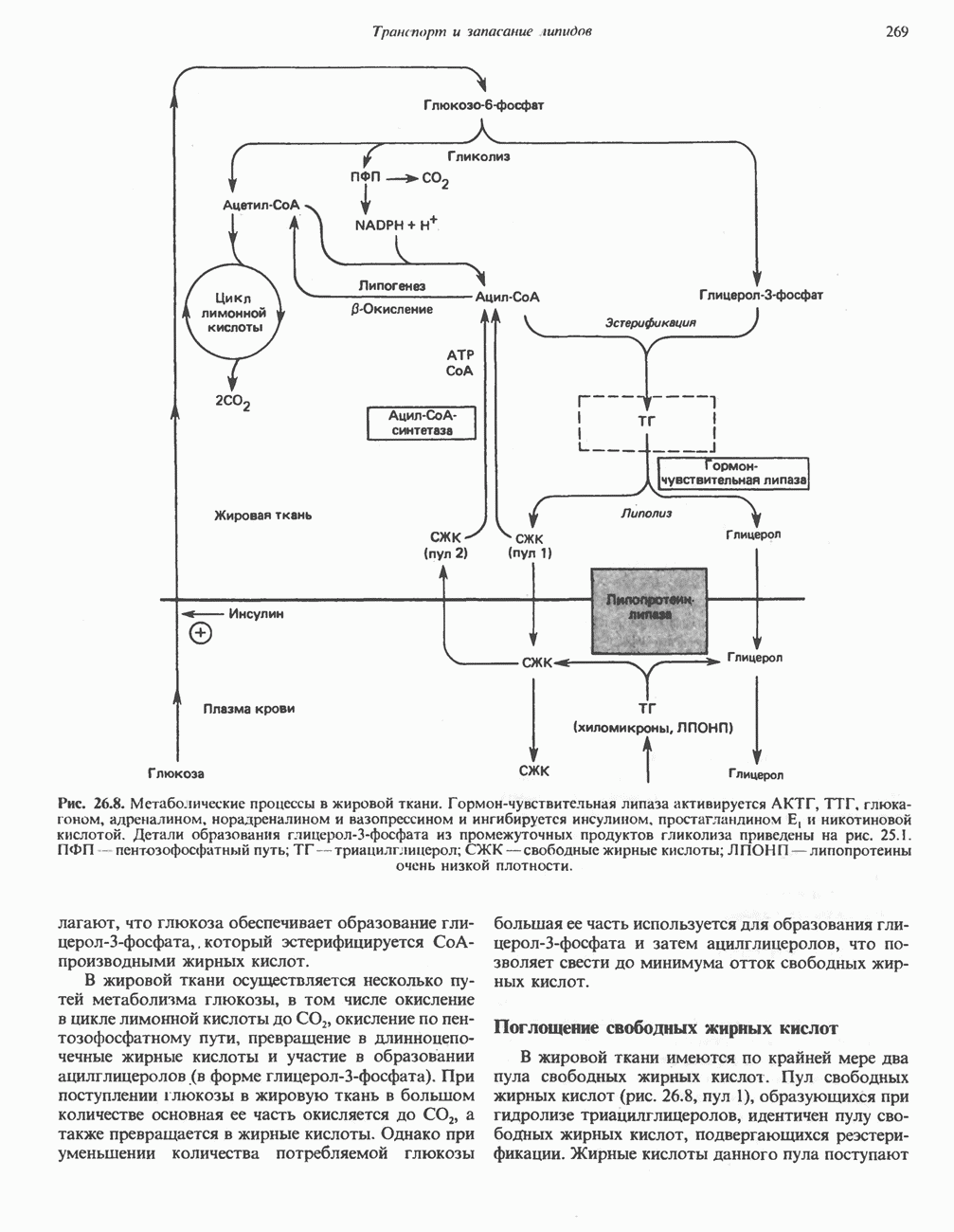
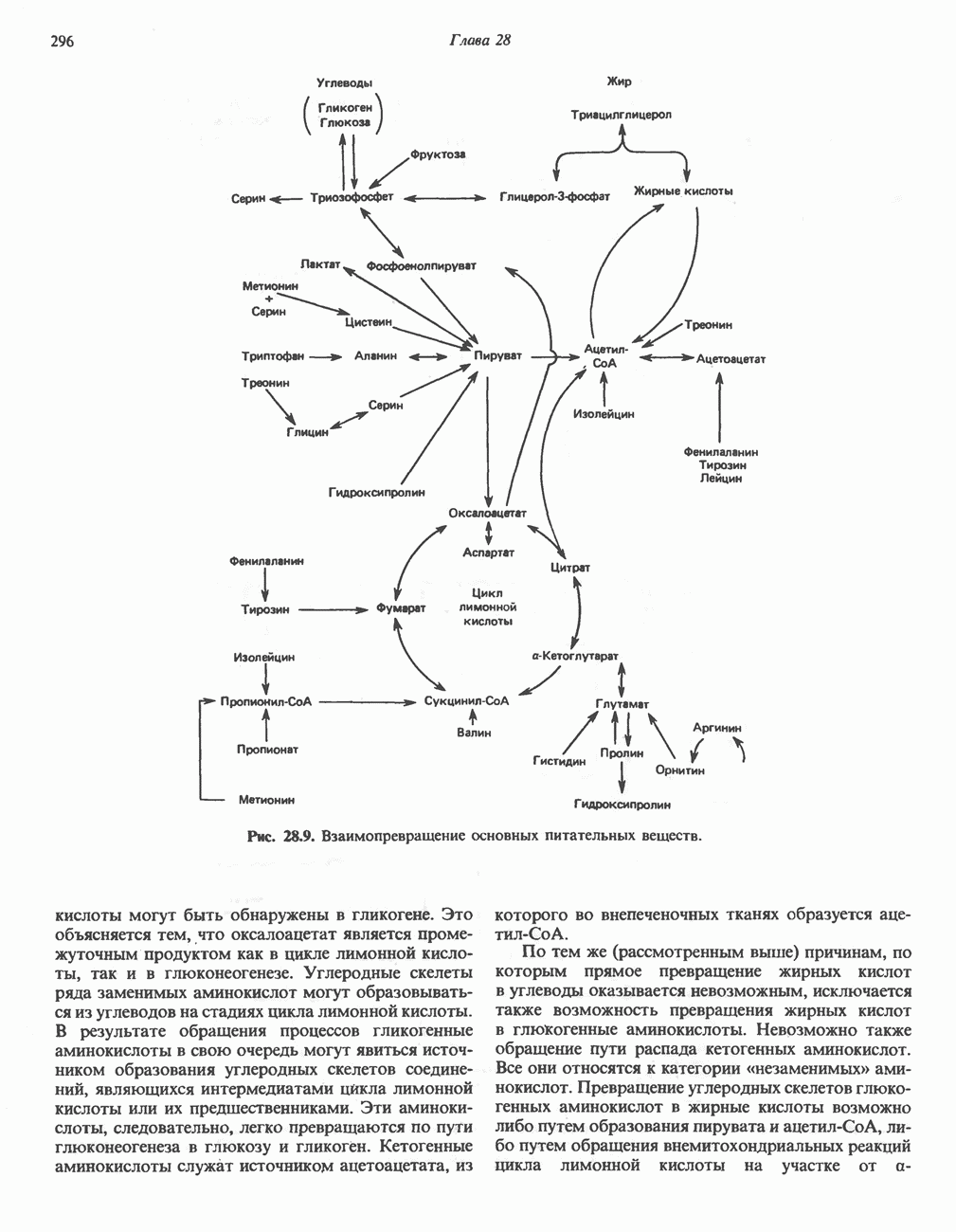
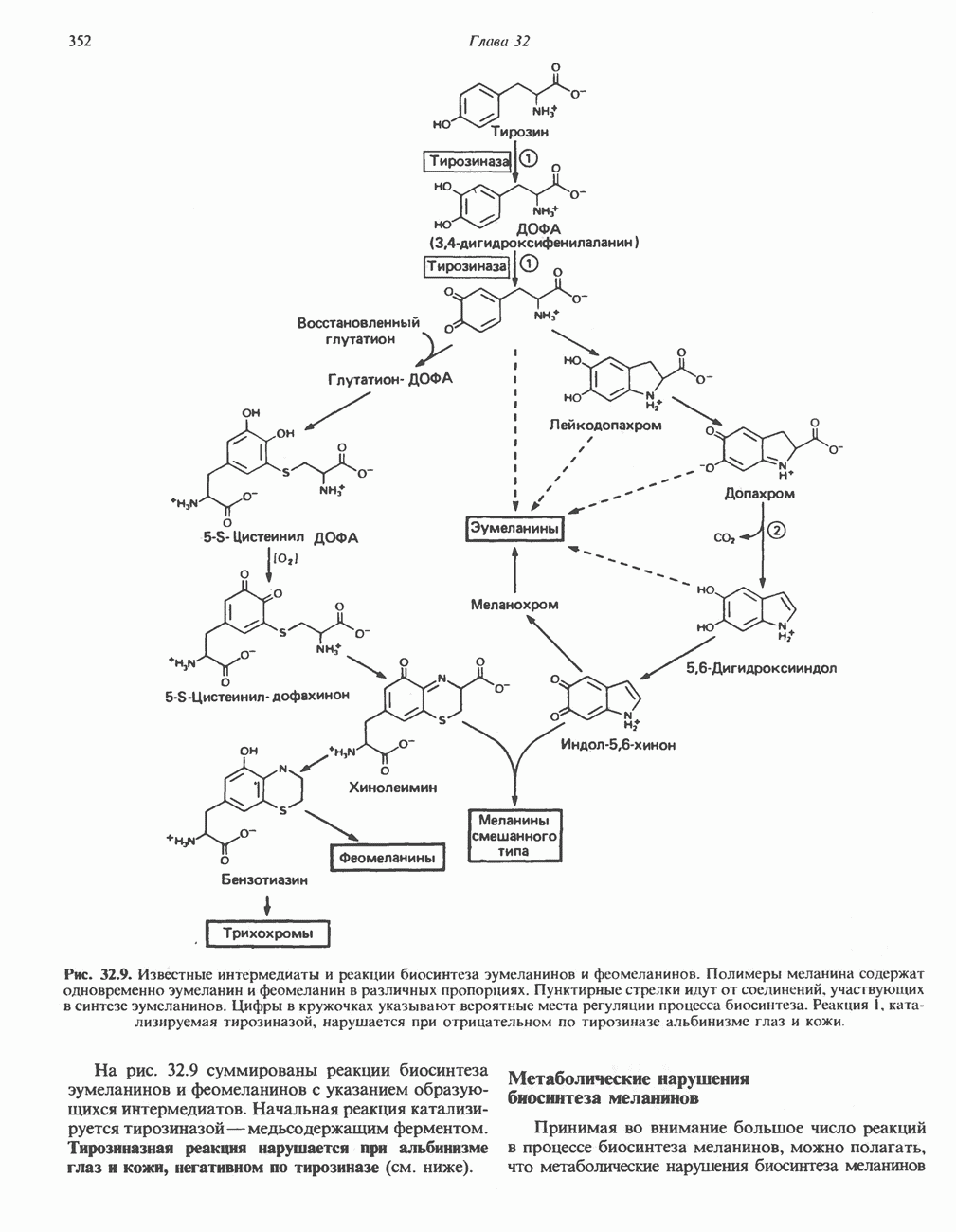
ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТСвободные жирные кислоты«Свободными жирными кислотами» (СЖК) называют жирные кислоты, находящиеся в неэстерифицированной форме; иногда их называют неэстерифицированными жирными кислотами (НЖК). В плазме крови длинноцепочечные СЖК образуют комплекс с альбумином, а в клетке — с белком, связывающим жирные кислоты, который называют Z-белком; фактически они никогда не бывают свободными. Короткоцепочечные жирные кислоты лучше растворяются в воде и находятся либо в виде неионизированной кислоты, либо в виде аниона жирной кислоты. Активация жирных кислотТак же как и в случае метаболизма глюкозы, жирная кислота прежде всего должна превратиться в активное производное в результате реакции, протекающей с участием АТР, и только после этого она способна взаимодействовать с ферментами, катализирующими дальнейшее превращение. В процессе окисления жирных кислот эта стадия является единственной, требующей энергии в виде АТР. В присутствии АТР и кофермента А фермент ацил-СоА-синтетаза (тиокиназа) катализирует превращение свободной жирной кислоты в «активную жирную кислоту» или ацил-СоА, которое осуществляется за счет расщепления одной богатой энергией фосфатной связи.
Присутствие неорганической пирофосфатазы, которая расщепляет богатую энергией фосфатную связь в пирофосфате, обеспечивает полноту протекания процесса активации. Таким образом, для активации одной молекулы жирной кислоты в итоге расходуются две богатые энергией фосфатные связи.
Ацил-СоА-синтетазы находятся в эндоплазмати-ческом ретикулуме, а также внутри митохондрий и на их наружной мембране. В литературе описан ряд ацил-СоА-синтетаз; они специфичны к жирным кислотам с определенной длиной цепи. Роль карнитина в окислении жирных кислотКарнитин
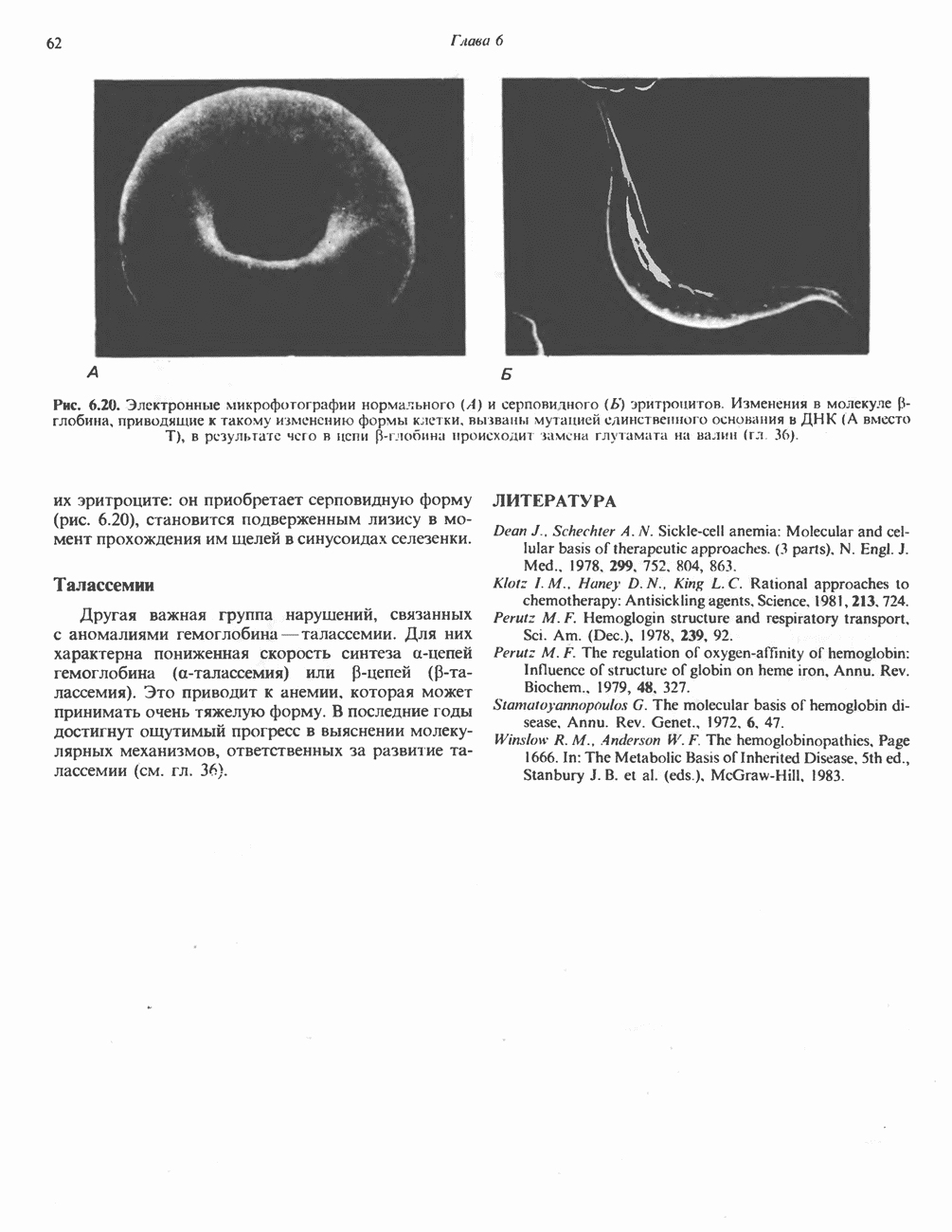
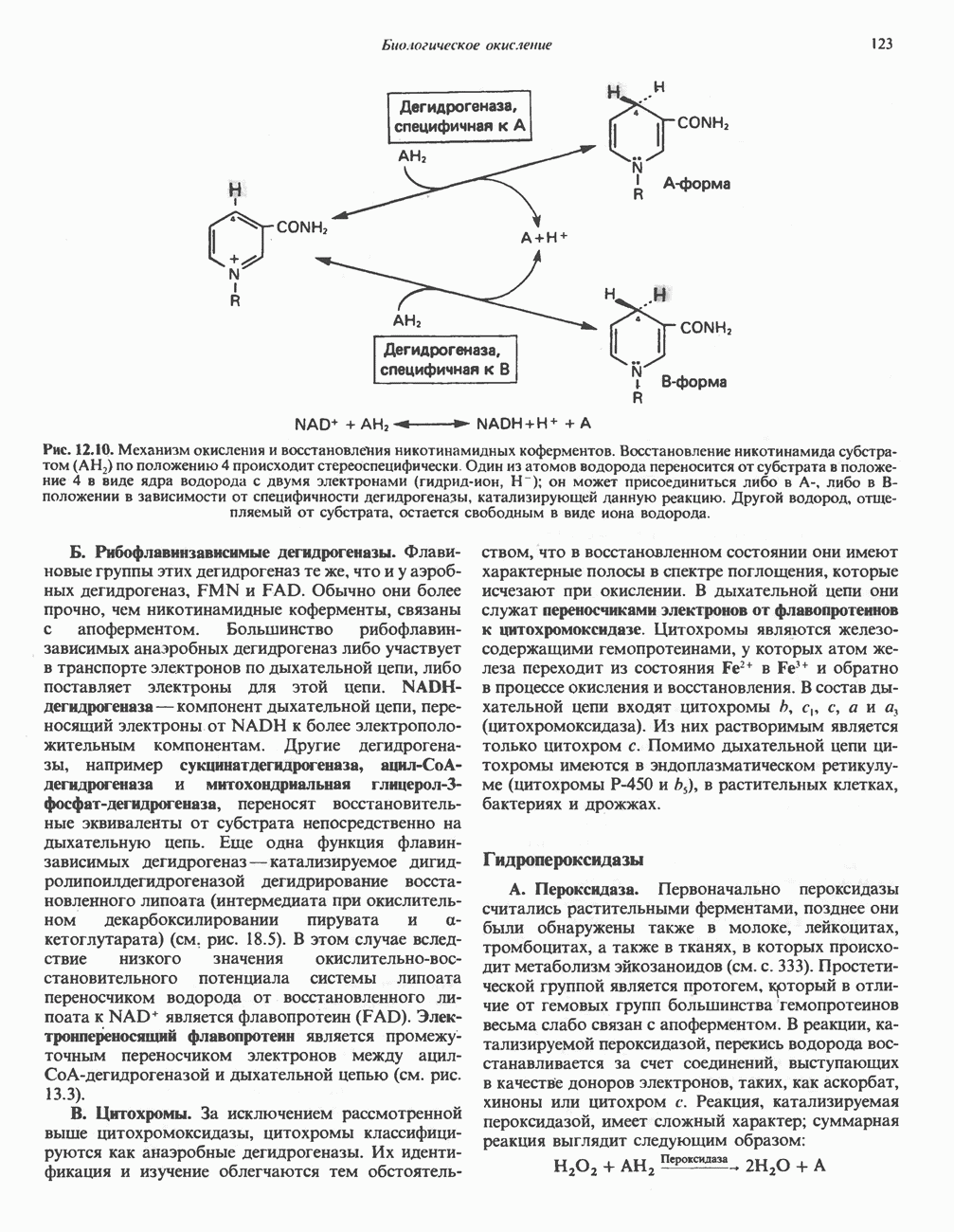
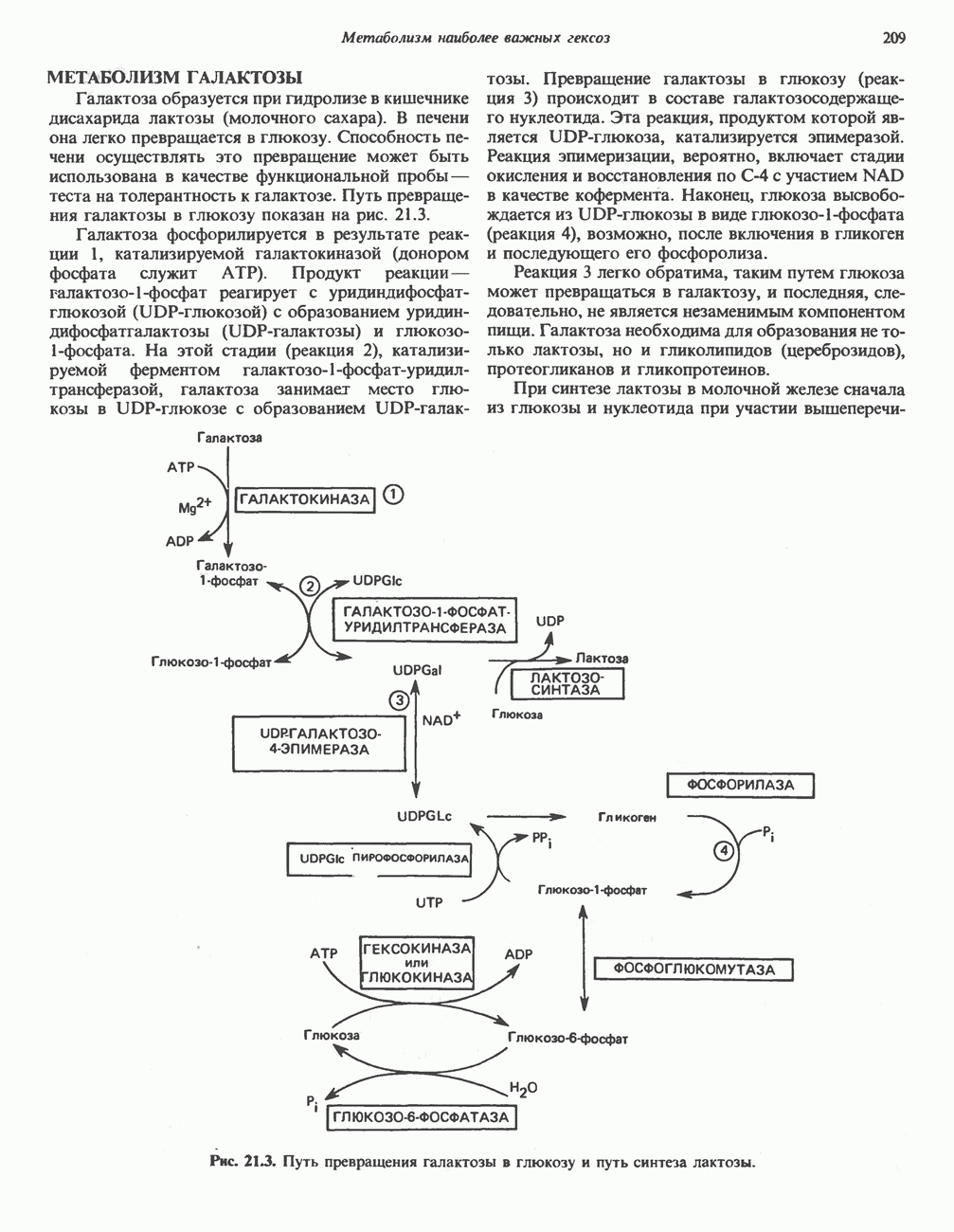
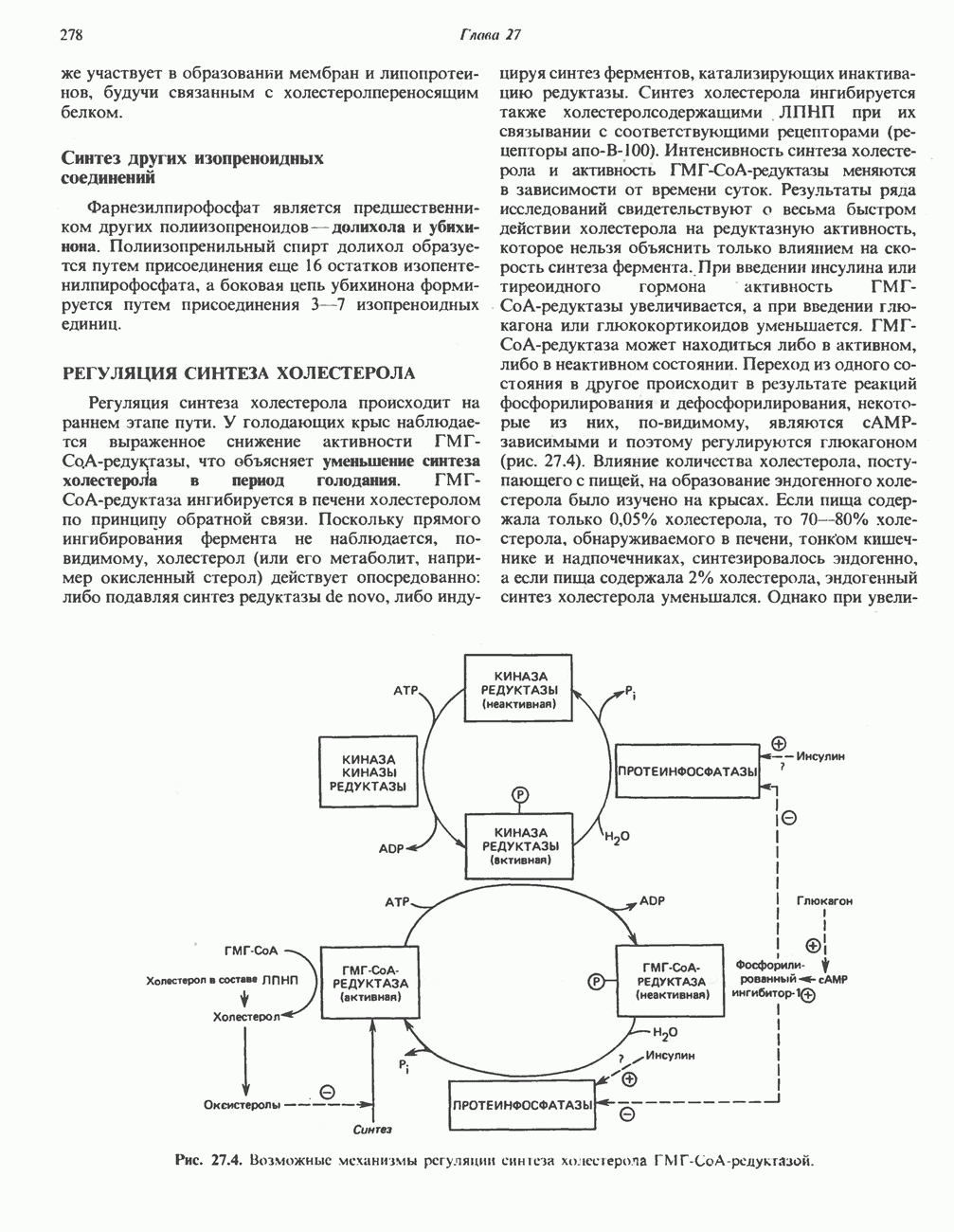
особенно много его в мышцах. Он образуется из лизина и метионина в печени и почках. Активация низших жирных кислот и их окисление могут происходить в митохондриях независимо от карнитина, однако длинноцепочечные ацил-СоА-производные (или СЖК) не могут проникать в митохондрии и окисляться, если предварительно не образуют ацилкарнитин-производных. На наружной стороне внутренней мембраны митохондрий имеется фермент карнитин-пальмитоилтрансфераза I, который переносит длинноцепочечные ацильные группы на карнитин с образованием ацилкарнитина; последний способен проникать в митохондрии, где находятся ферменты, катализирующие процесс ( Возможный механизм, объясняющий участие карнитина в окислении жирных кислот в митохондриях, приведен на рис. 23.1. Кроме того, в митохондриях находится другой фермент — карнитин-ацетилтрансфераза, который катализирует перенос короткоцепочечных ацильных групп между СоА и карнитином. Функция этого фермента пока не ясна.
Рис. 23.1. Роль карнитина в переносе длинноцепочечных жирных кислот через внутреннюю мембрану митохондрий. Длиннопепочечный ацил-СоА не способен проходить через внутреннюю мембрану митохондрий, в то время как такой способностью обладает ацилкарнитин, образую цийся при Действии карнитин-пальмитонлтрансферазы I. Карнитин-ацилкарнитин-фанслоказа является транспортной системой. осуществляющей перенос молекулы ацилкарнитина через внутреннюю мембрану митохондрии, сопряженный с выходом мопскулы свободного карнитина. Затем при действии карнитин-пальмитоилтрансферазы 11, локализованной на внутренней поверхности внутренней мембраны митохондрии, ацилкарнитин взаимодействует с СоА. В результате в митохондриальном матриксе вновь образуется ацил-СоА. а карнитин высвобождается. Возможно,
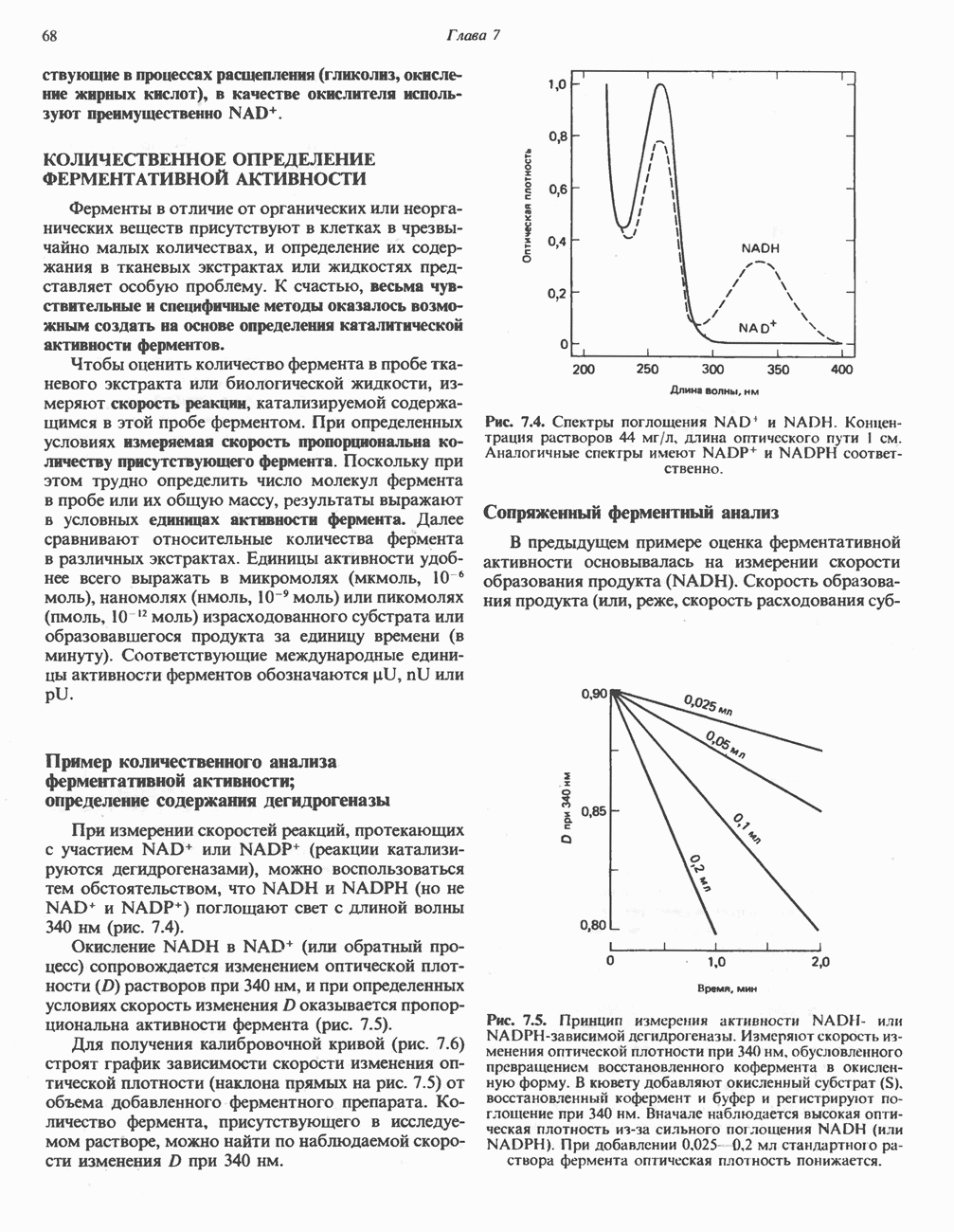
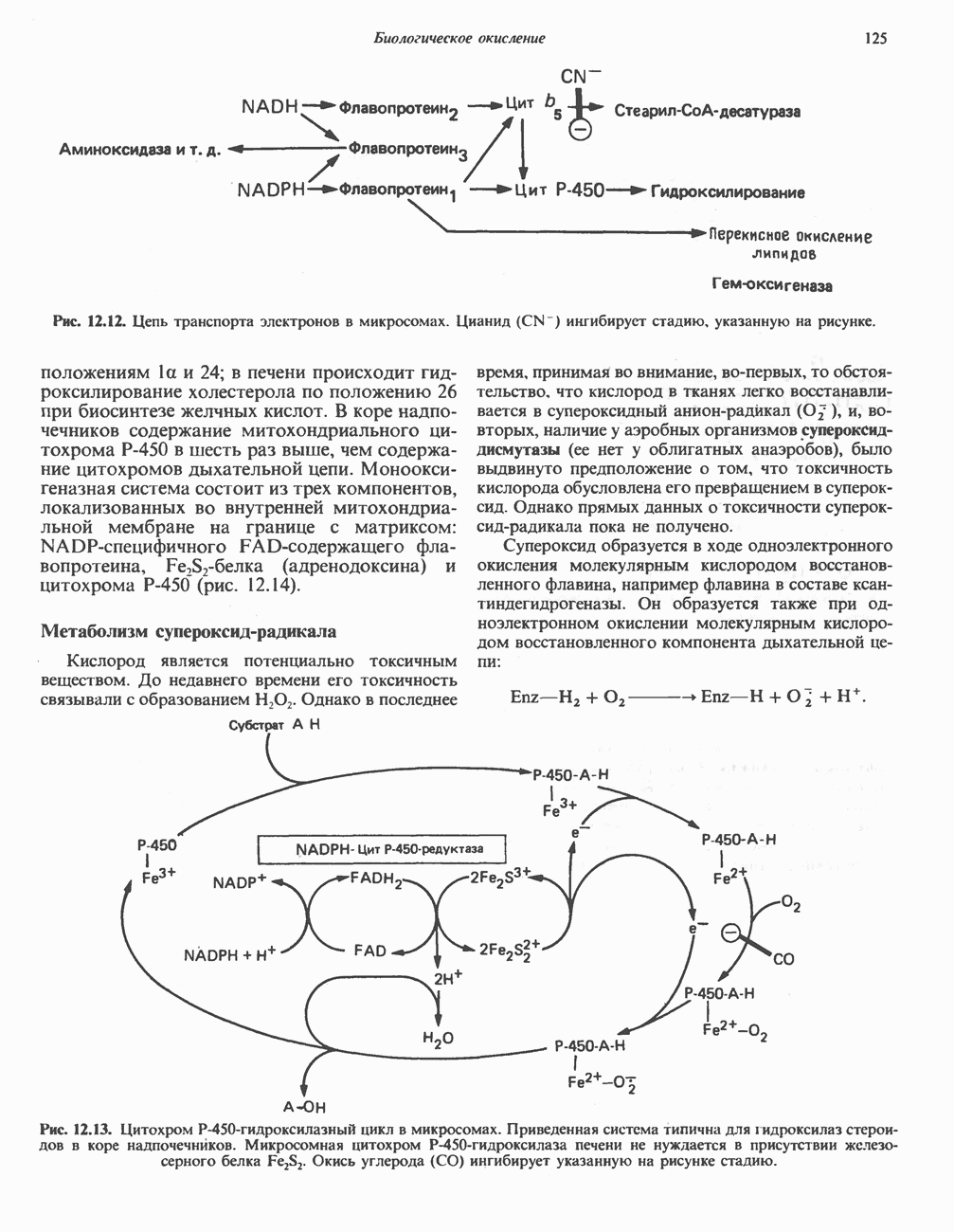
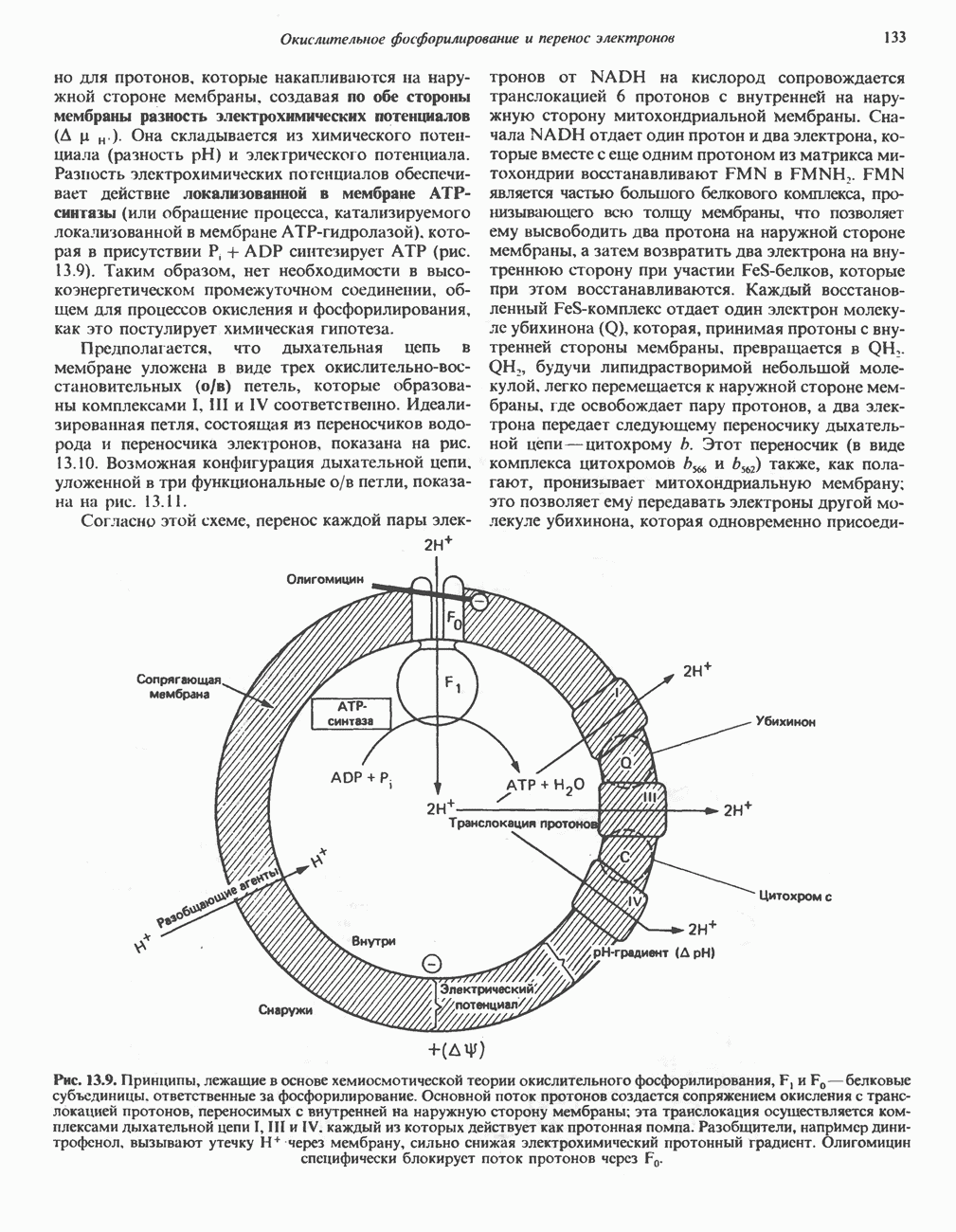
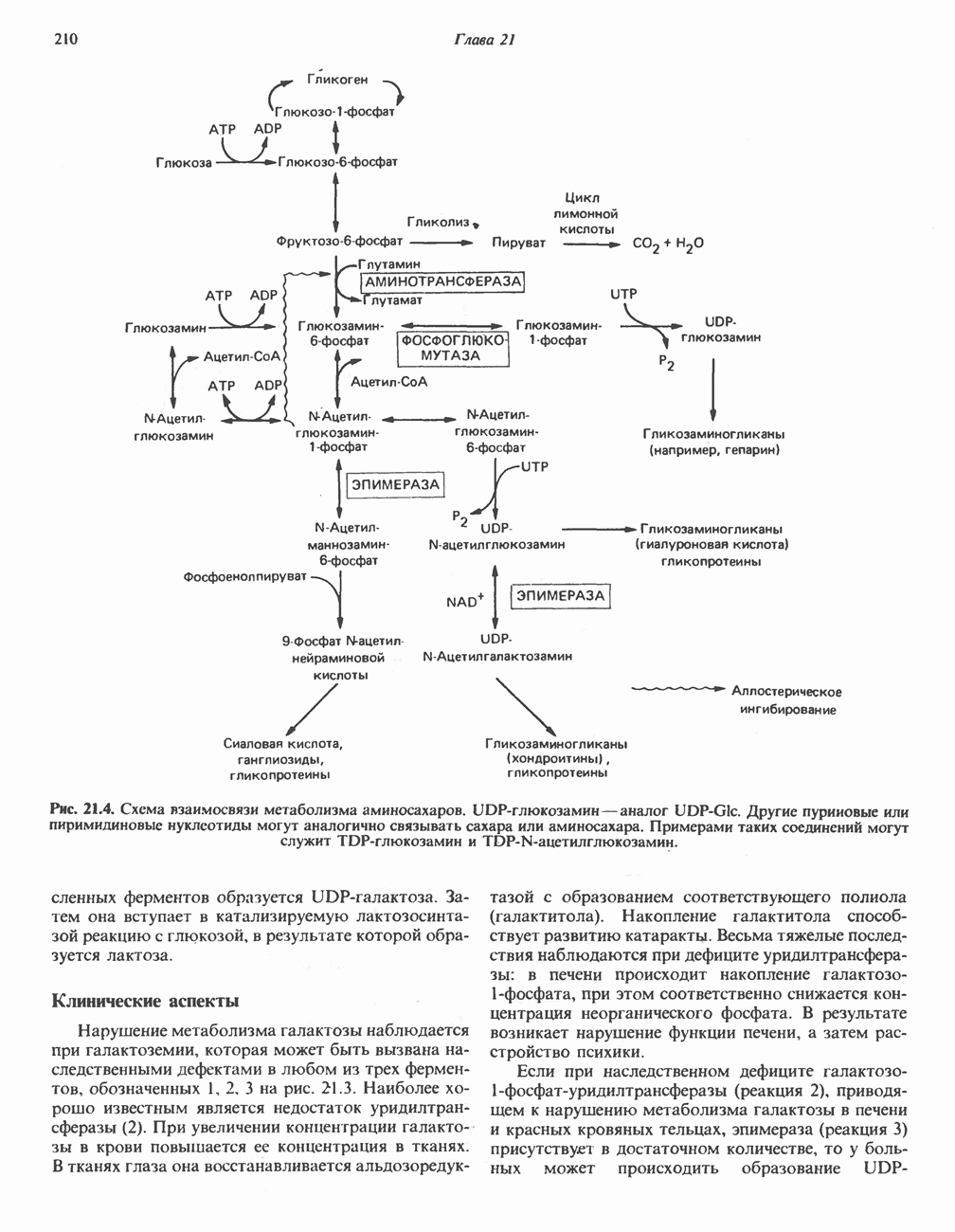
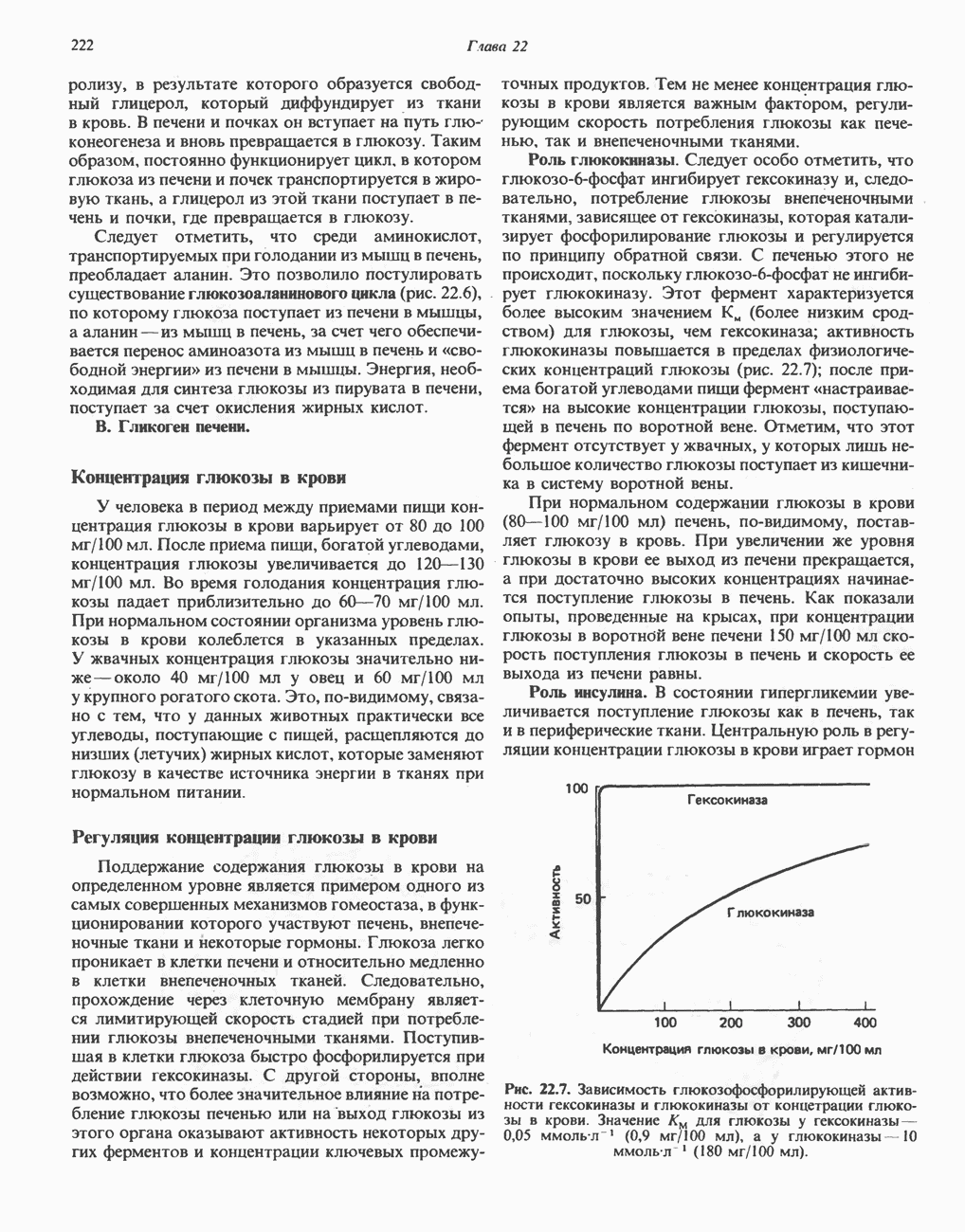
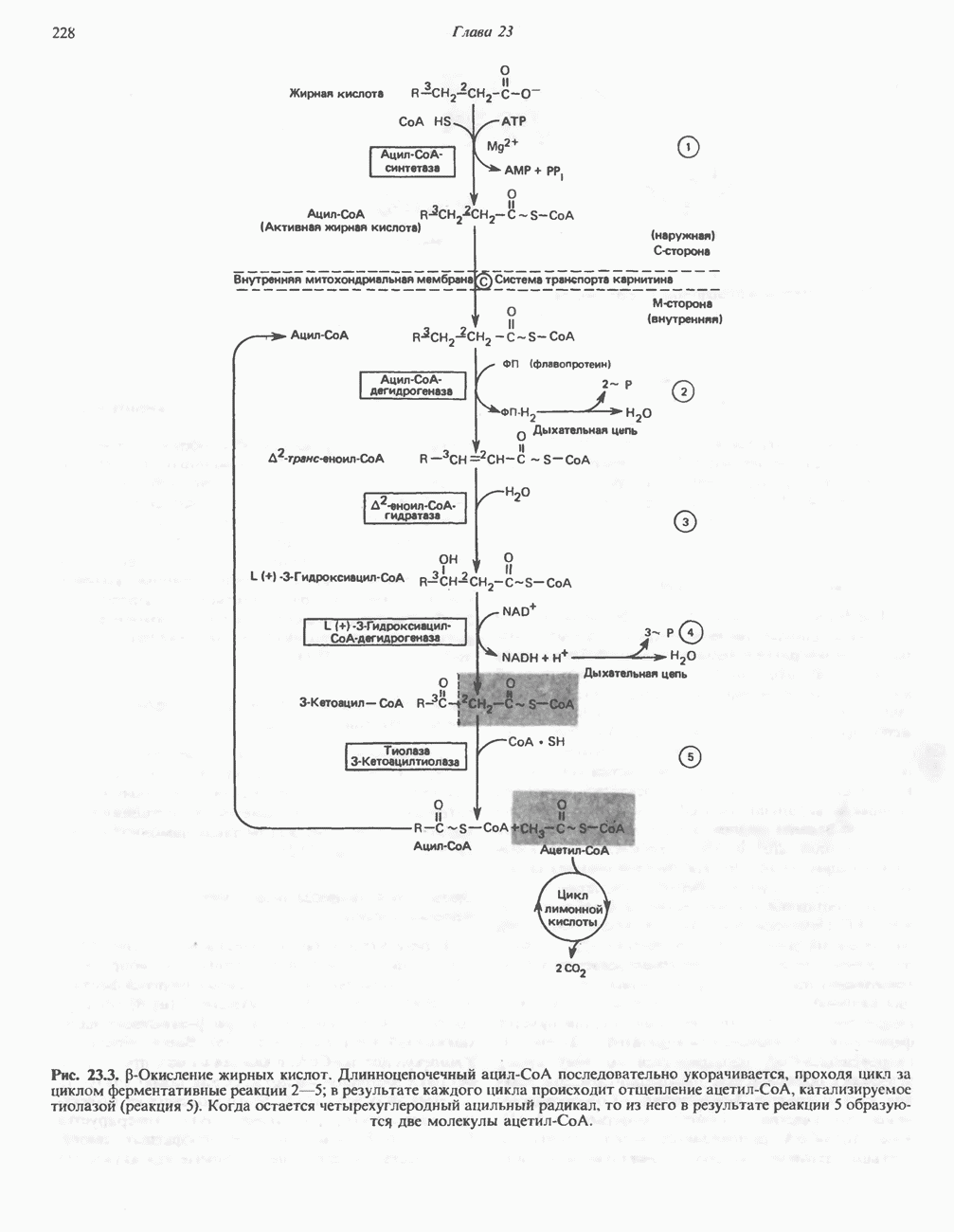
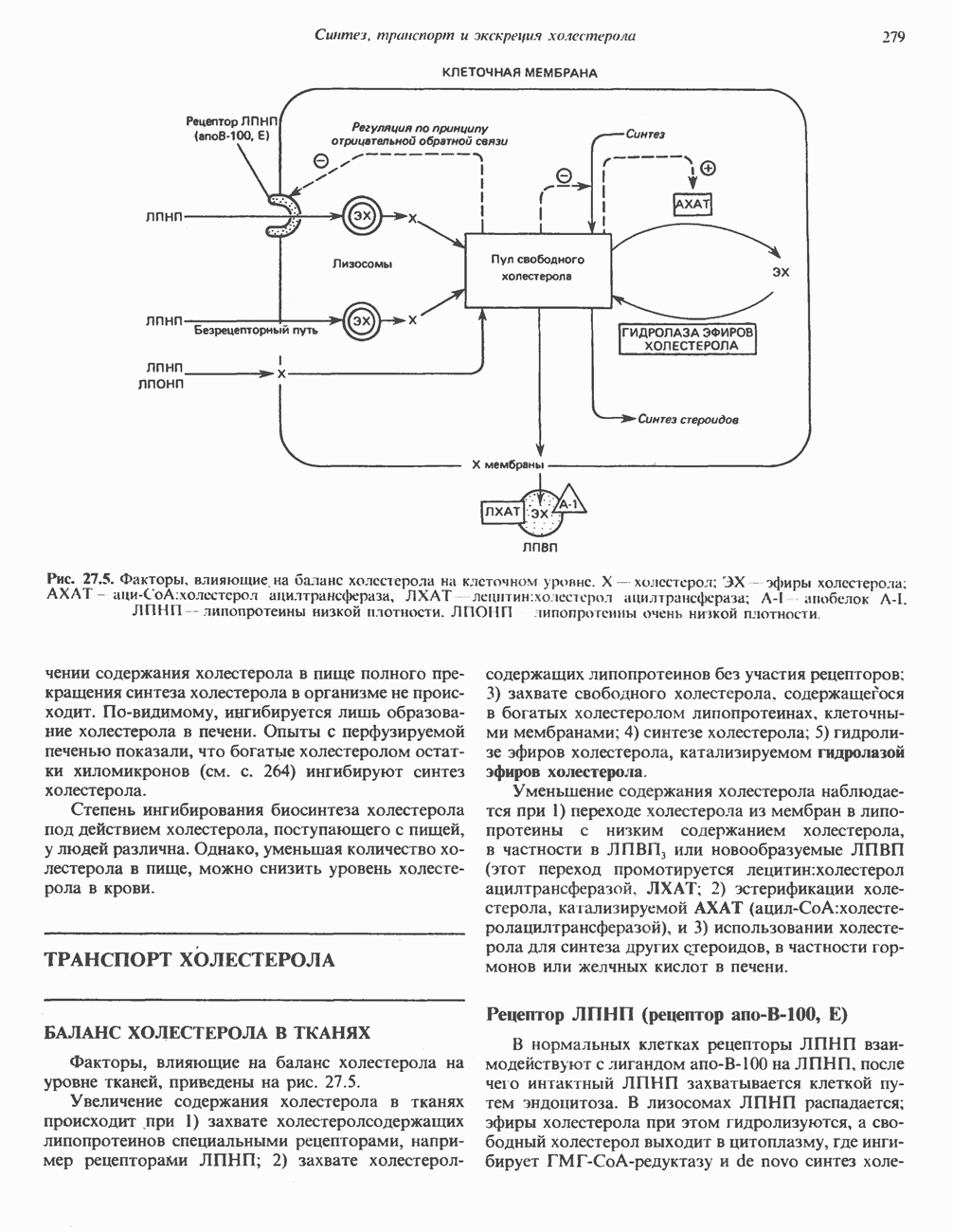
он облегчает транспорт ацетильных групп через мембрану митохондрий. b-Окисление жирных кислотОбщее представление дает рис. 23.2. При 13-окислении жирных кислот 2 атома углерода одновременно отщепляются от карбоксильного конца молекулы ацил-СоА. Углеродная цепь разрывается
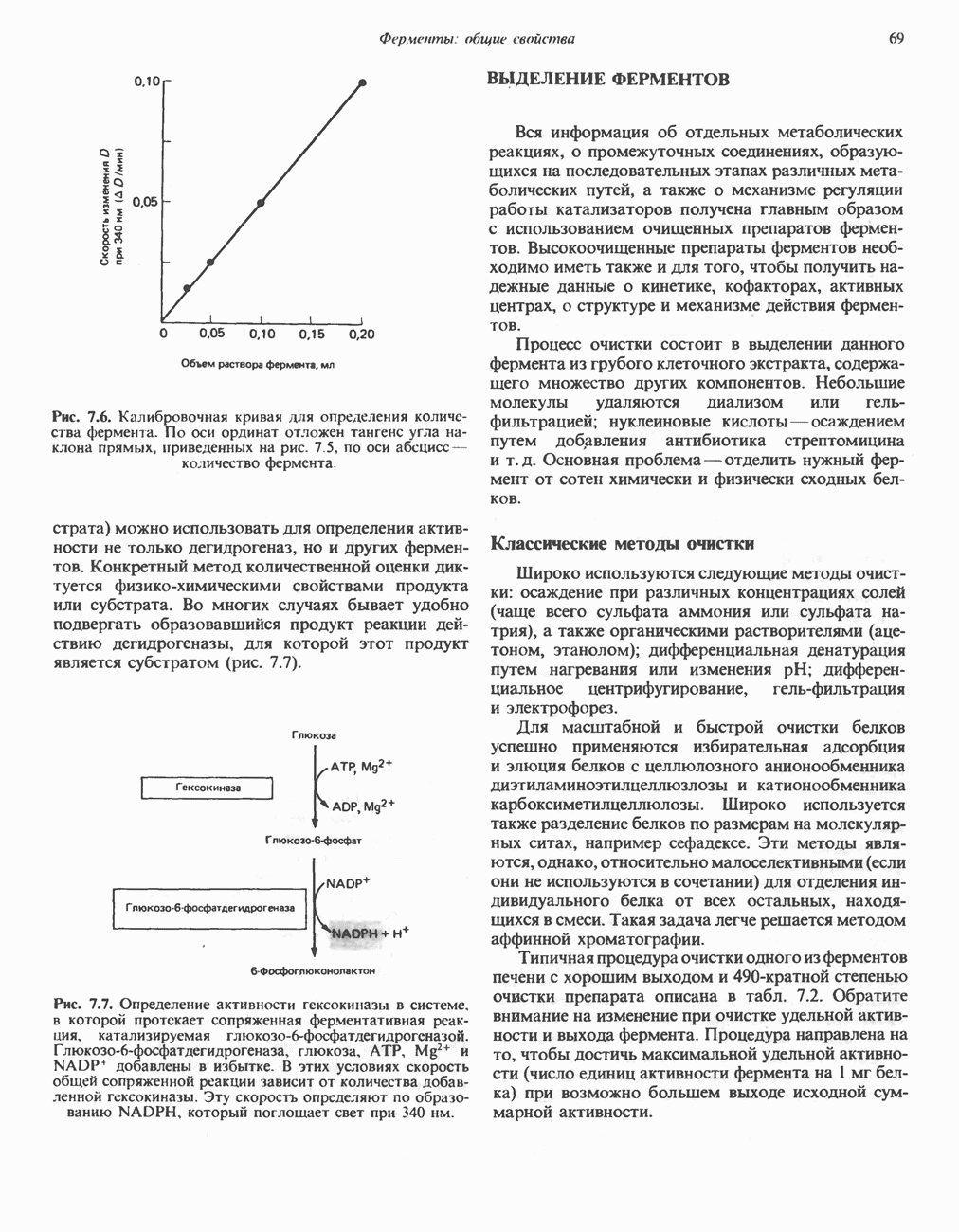
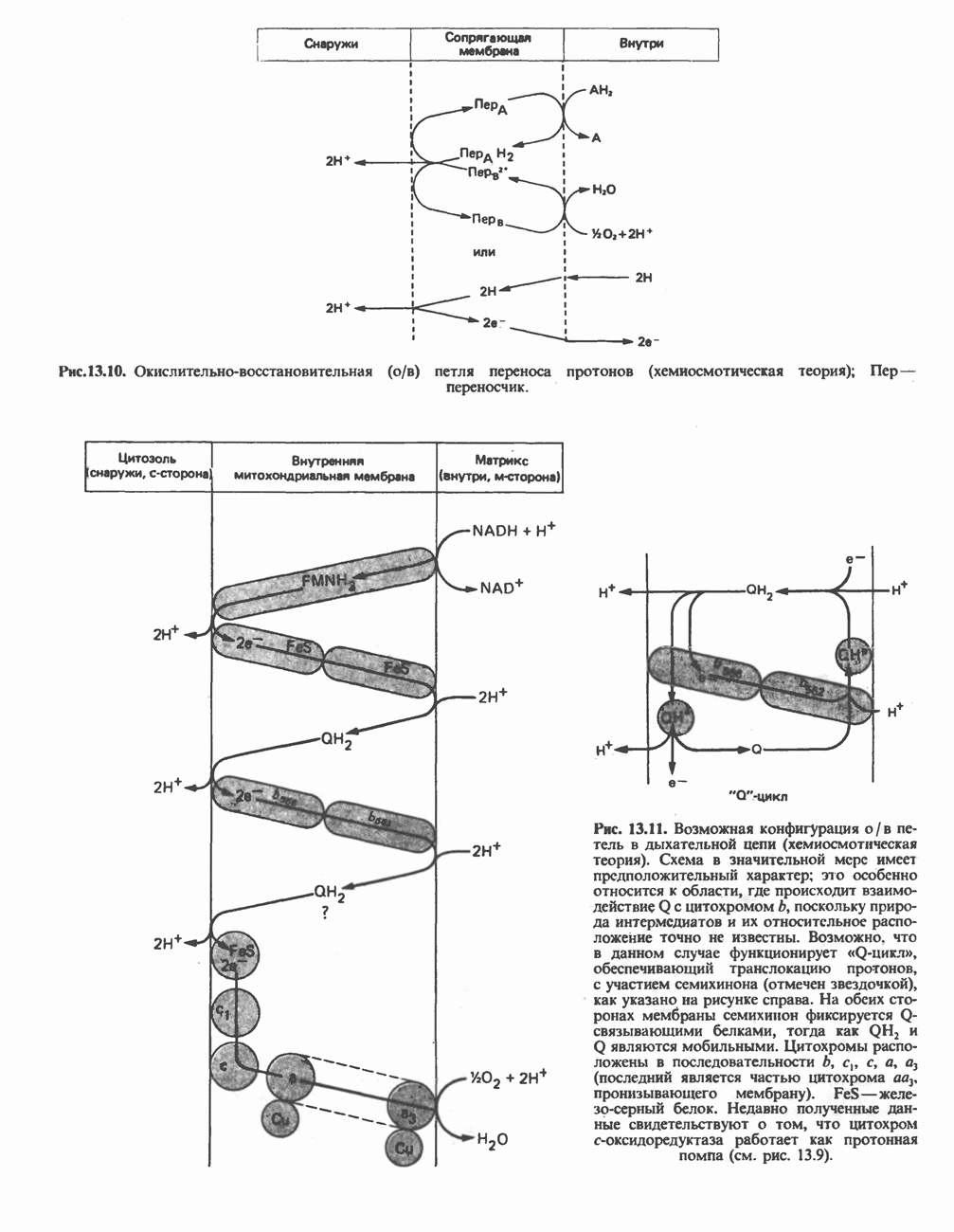
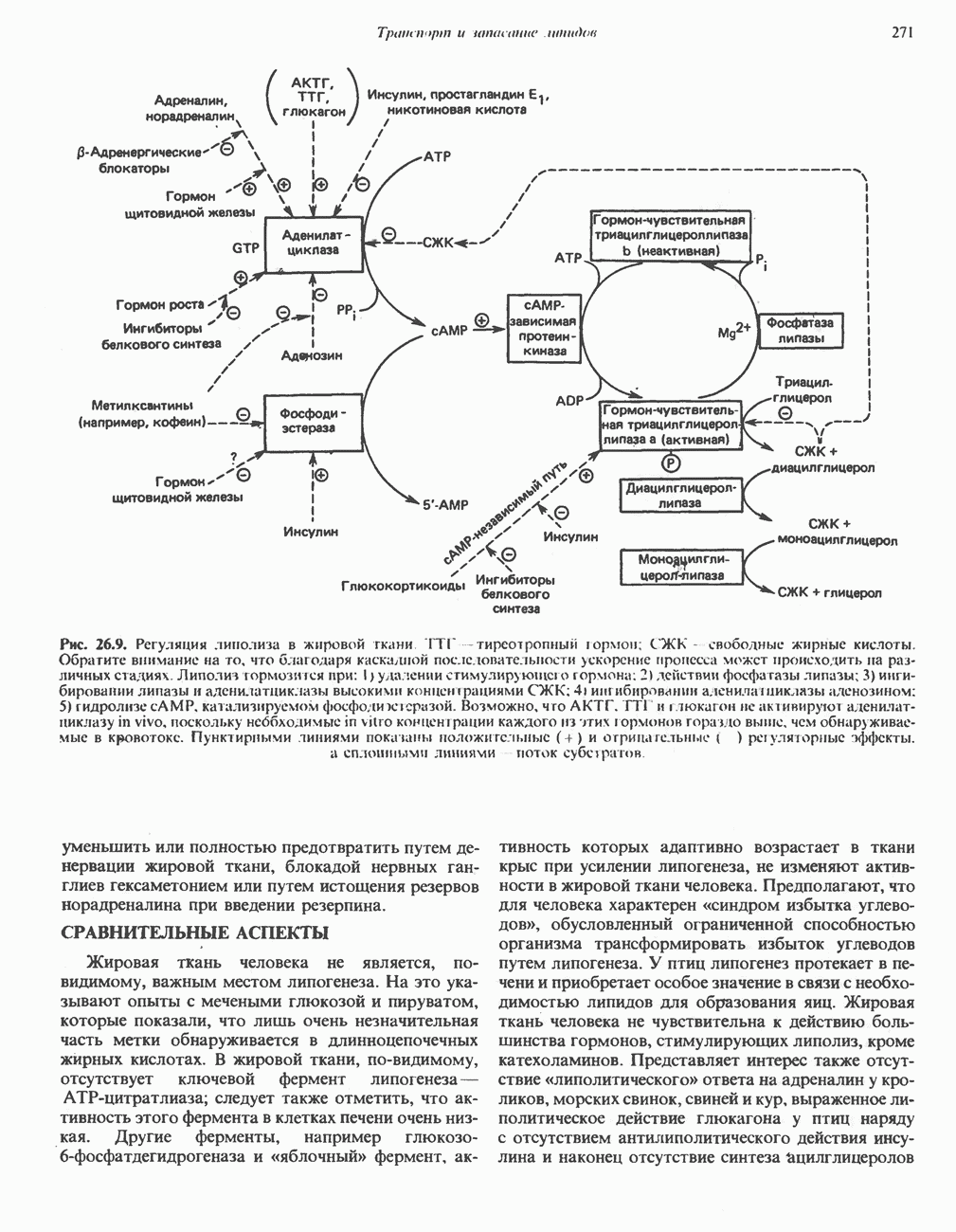
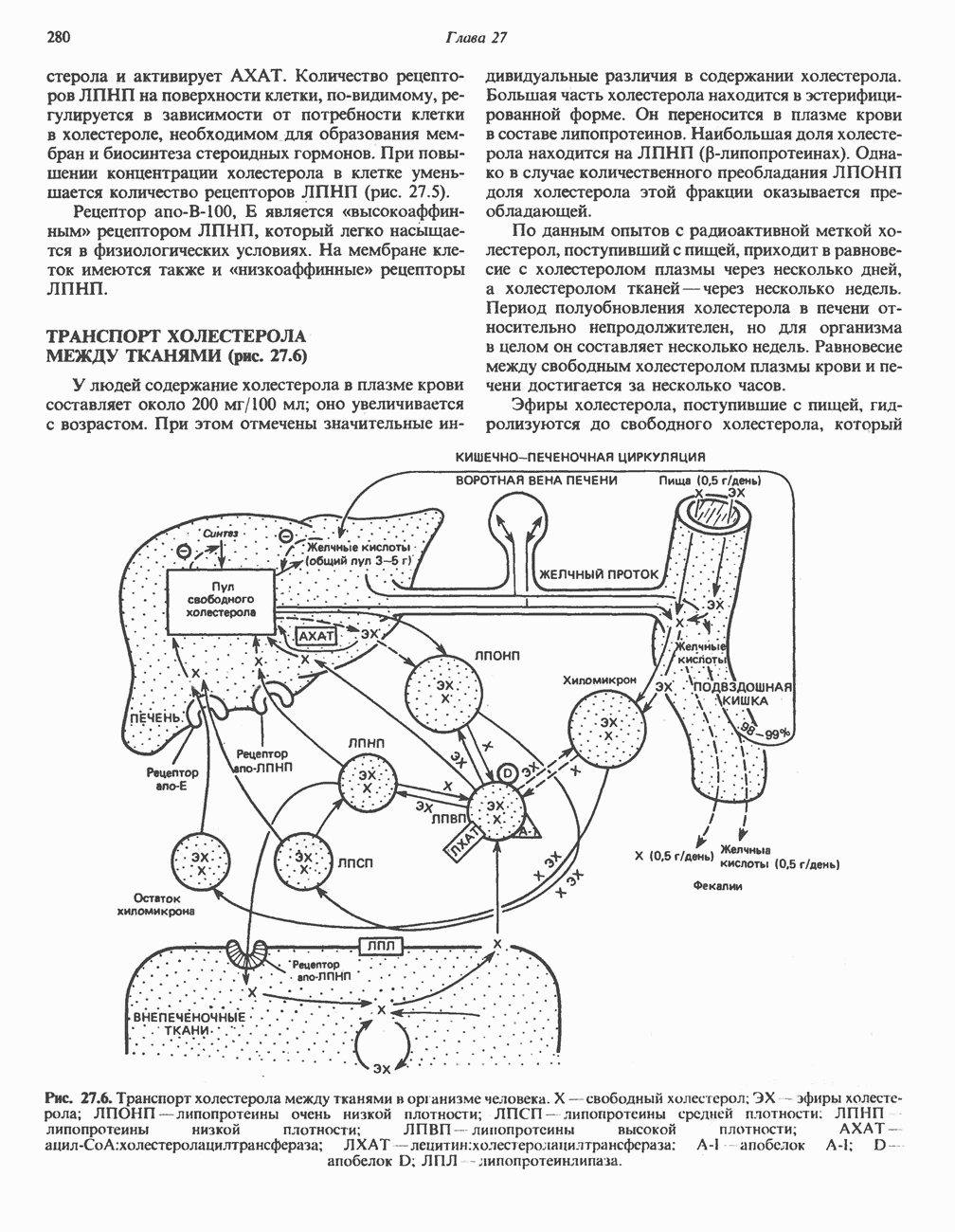
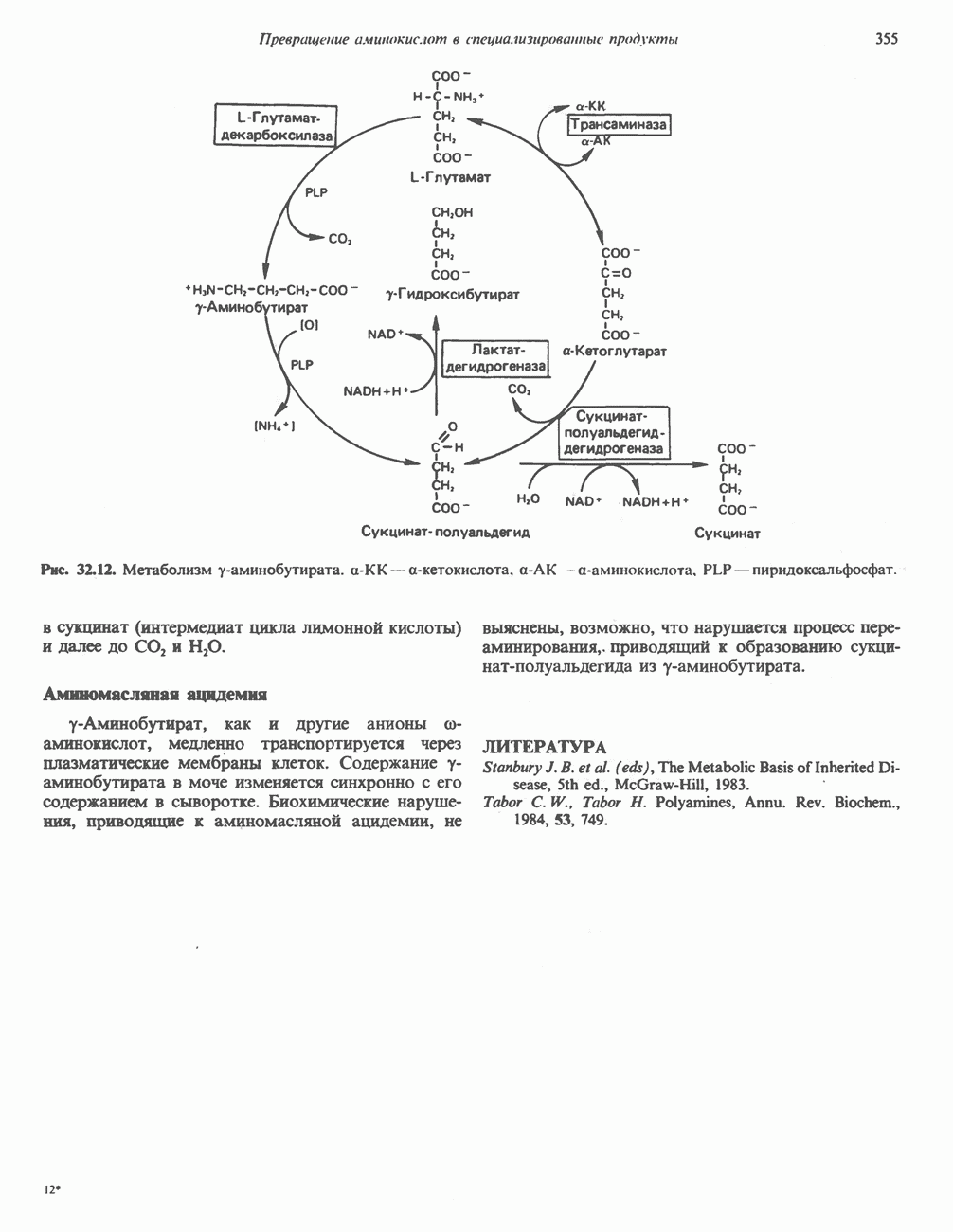
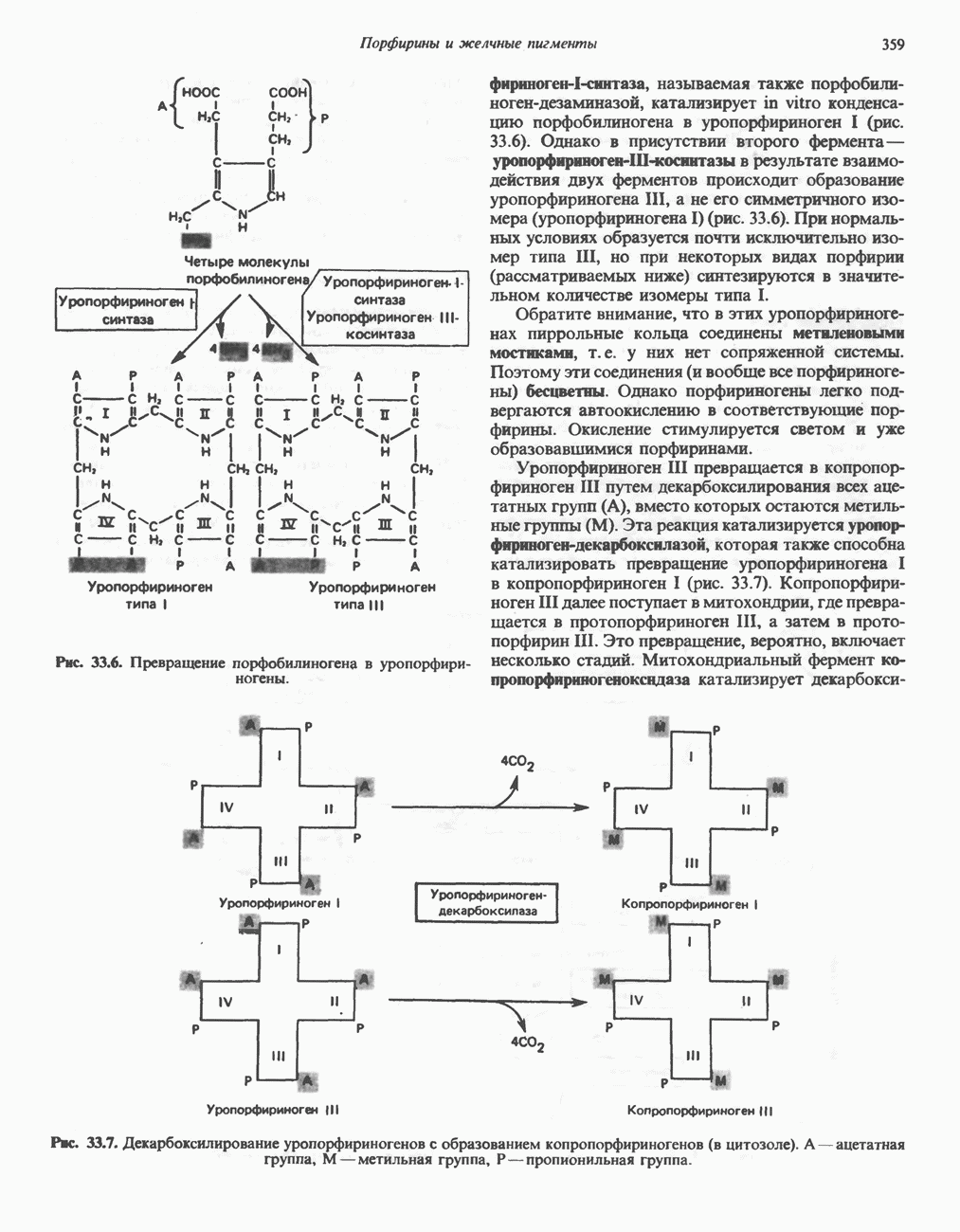
Рис. 23.2. Схема между атомами углерода в положениях Последовательность реакцийРяд ферментов, известных под общим названием «оксидазы жирных кислот», находятся в митохондриальном матриксе в непосредственной близости от дыхательной цепи, локализованной во внутренней мембране митохондрий. Эта система катализирует окисление ацил-СоА до ацетил-СоА, которое сопряжено с фосфорилированием ADP до АТР (рис. 23.3). После проникновения ацильного фрагмента через мембрану митохондрий при участии карнитиновой транспортной системы и переноса ацильной группы от карнитина на Окисление жирных кислот с нечетным числом атомов углеродаb-Окисление жирных кислот с нечетным числом атомов углерода заканчивается на стадии образования трехуглеродного фрагмента - пропионил-СоА, который затем превращается в Энергетика процесса окисления жирных кислотВ результате переноса электронов по дыхательной цепи от восстановленного флавопротеина и NAD синтезируется по 5 богатых энергией фосфатных связей (см. гл. 13) на каждые 7 (из 8) молекул ацетил-СоА, образующихся при b-окислении пальмитиновой кислоты (см. скан) Рис. 23.3. Р Окисление жирных кислот. Длинноцепочечный ацит СоА последовательно укорачивается, проходя цикт за циклом ферментативные реакции 2—5; в результате каждого цикла происходит отщепление ацетил-СоА, катализируемое тиолазой (реакция 5). Когда остается четырехуглеродный ацильный радикал, то из него в результате реакции 5 образуются две молекулы ацетил-СоА. жирной кислоты, то в общей сложности получим 129 богатых энергией связей на 1 моль или Окисление жирных кислот в пероксисомахВ пероксисомах а- и b-Окисление жирных кислот
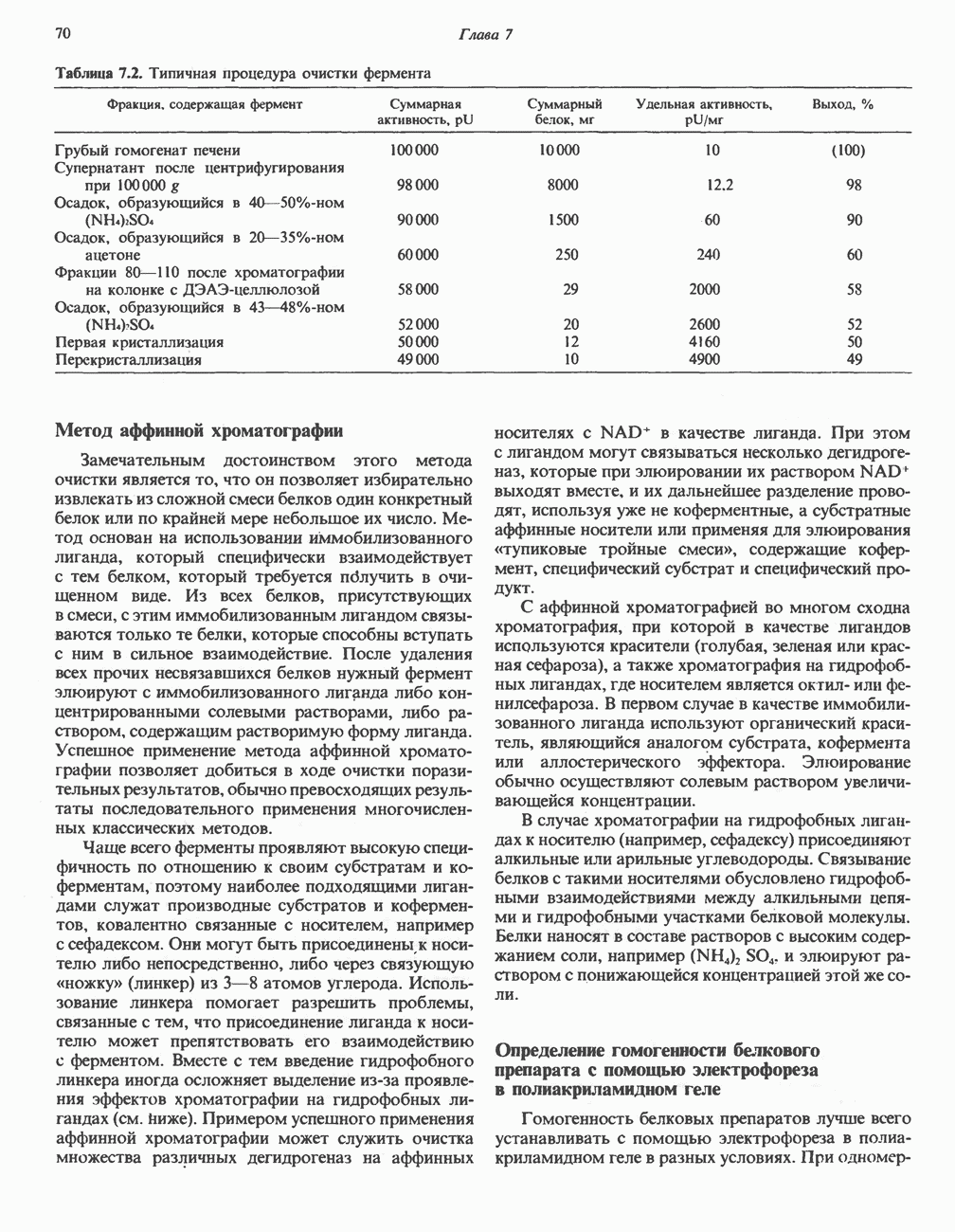
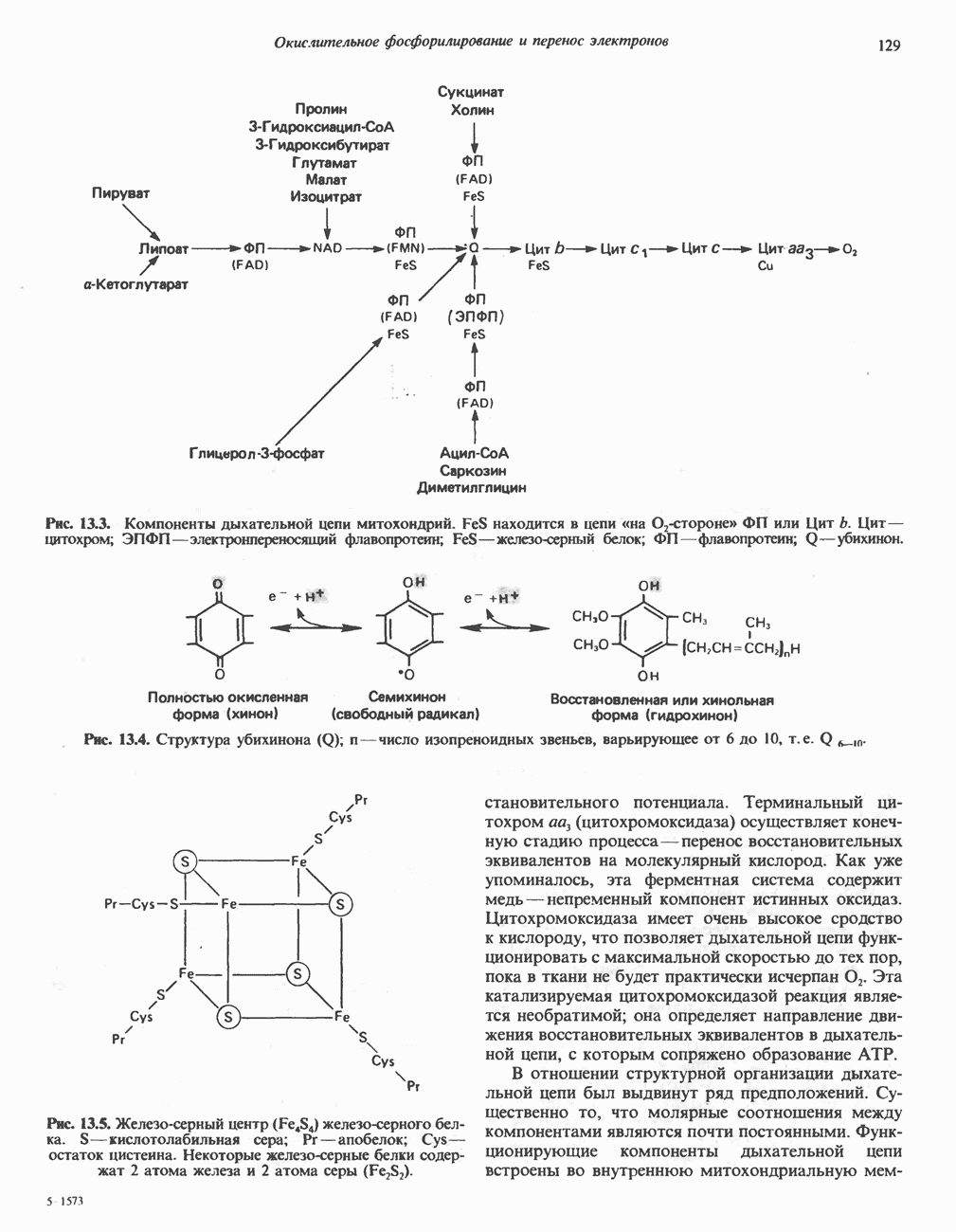
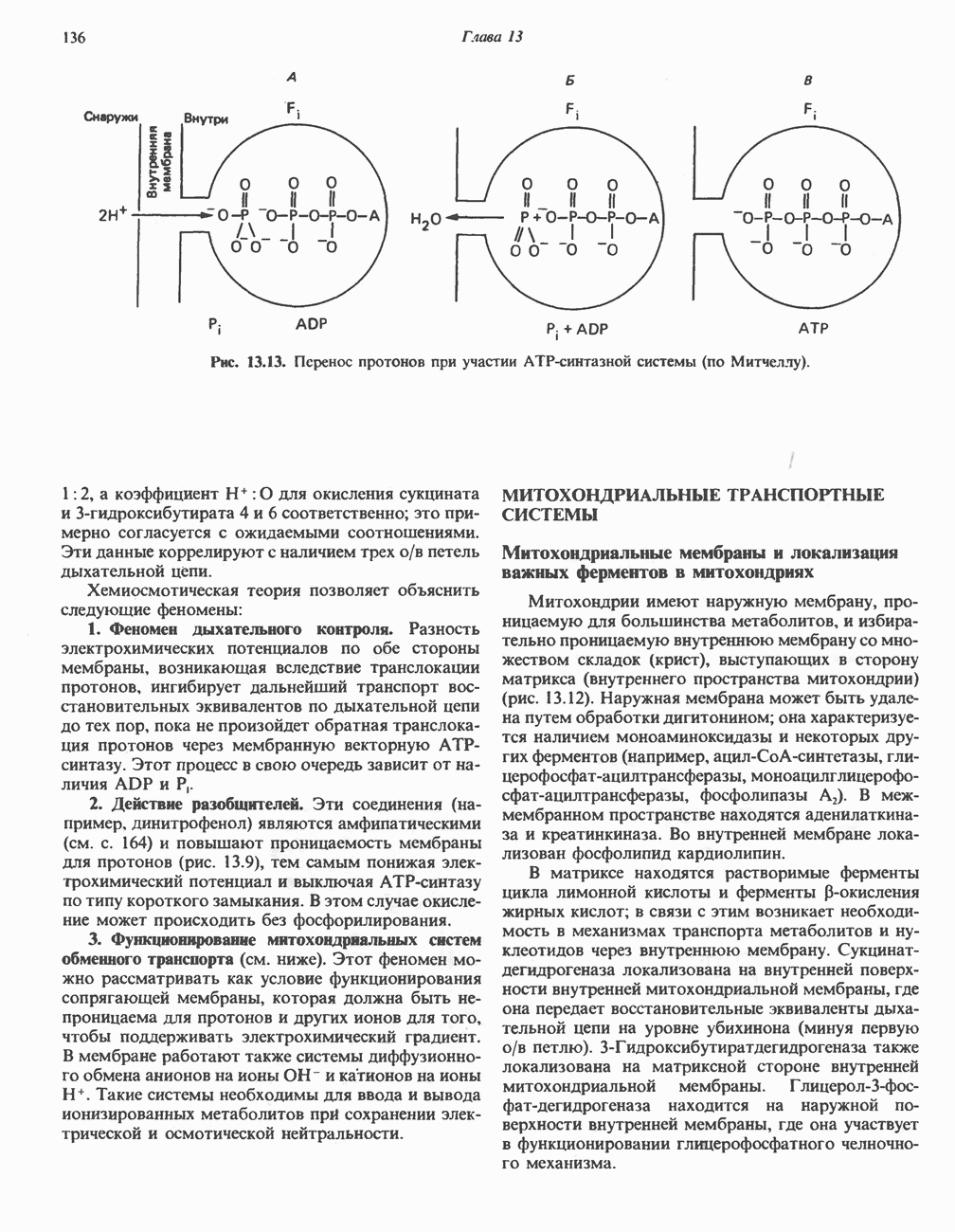
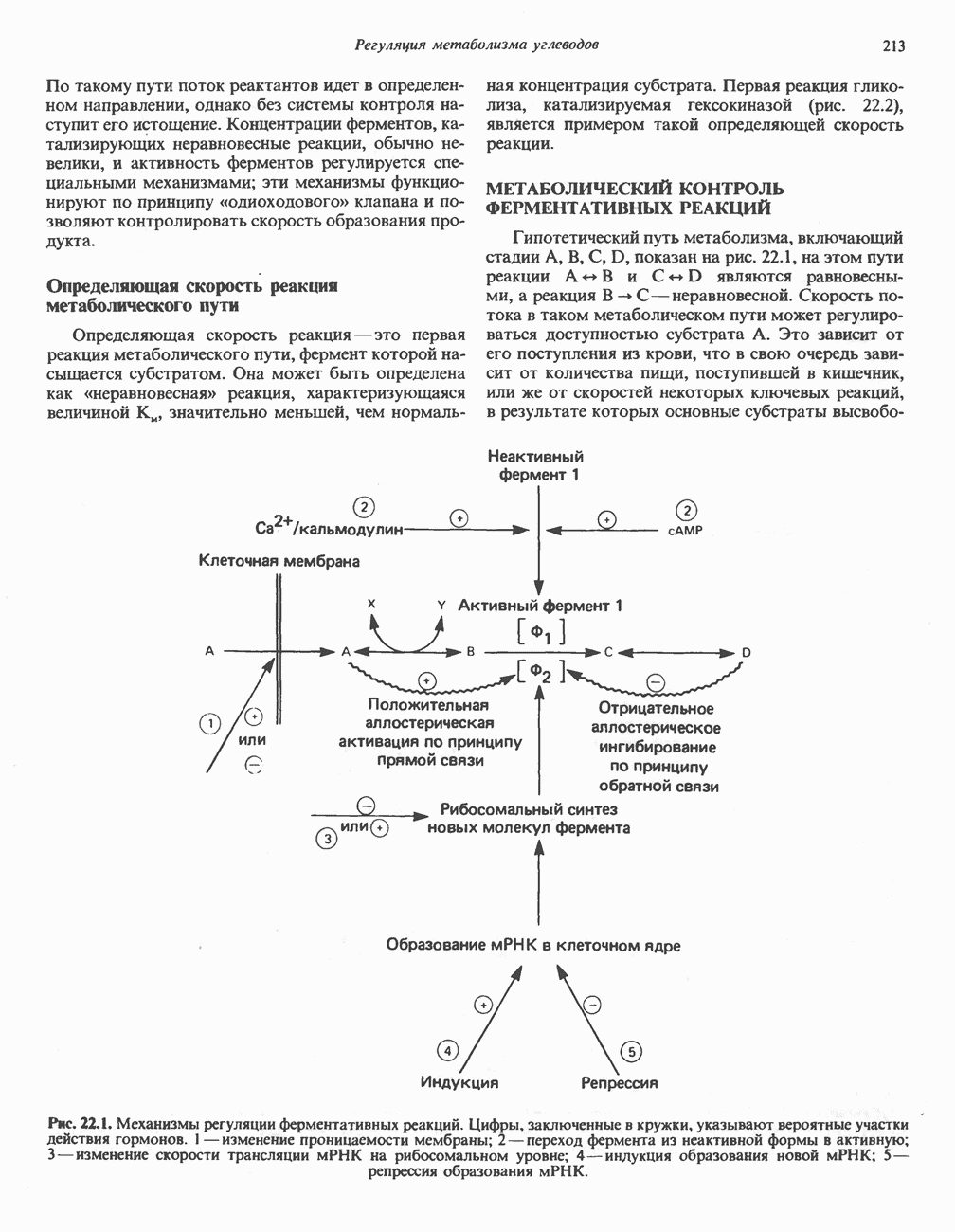
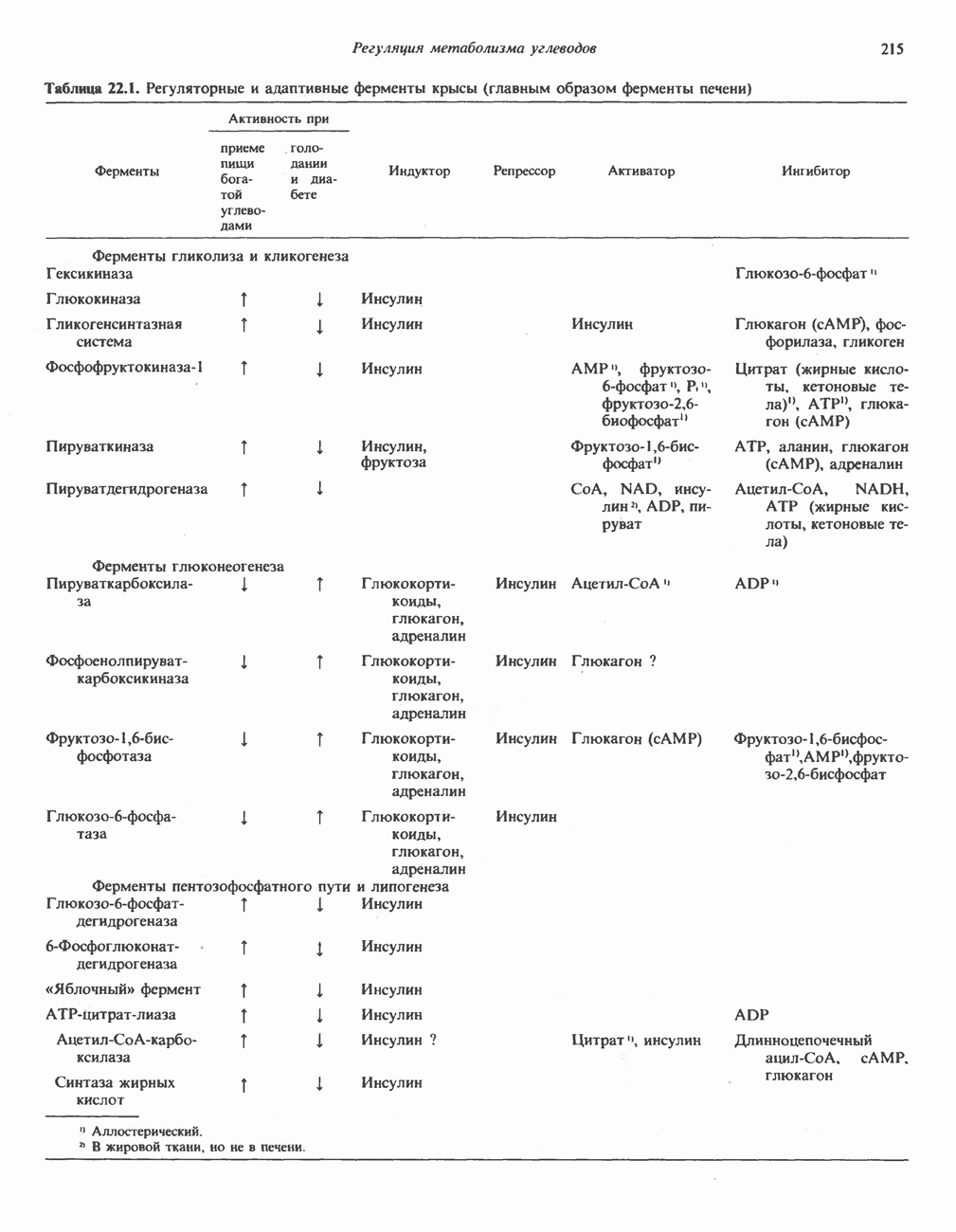
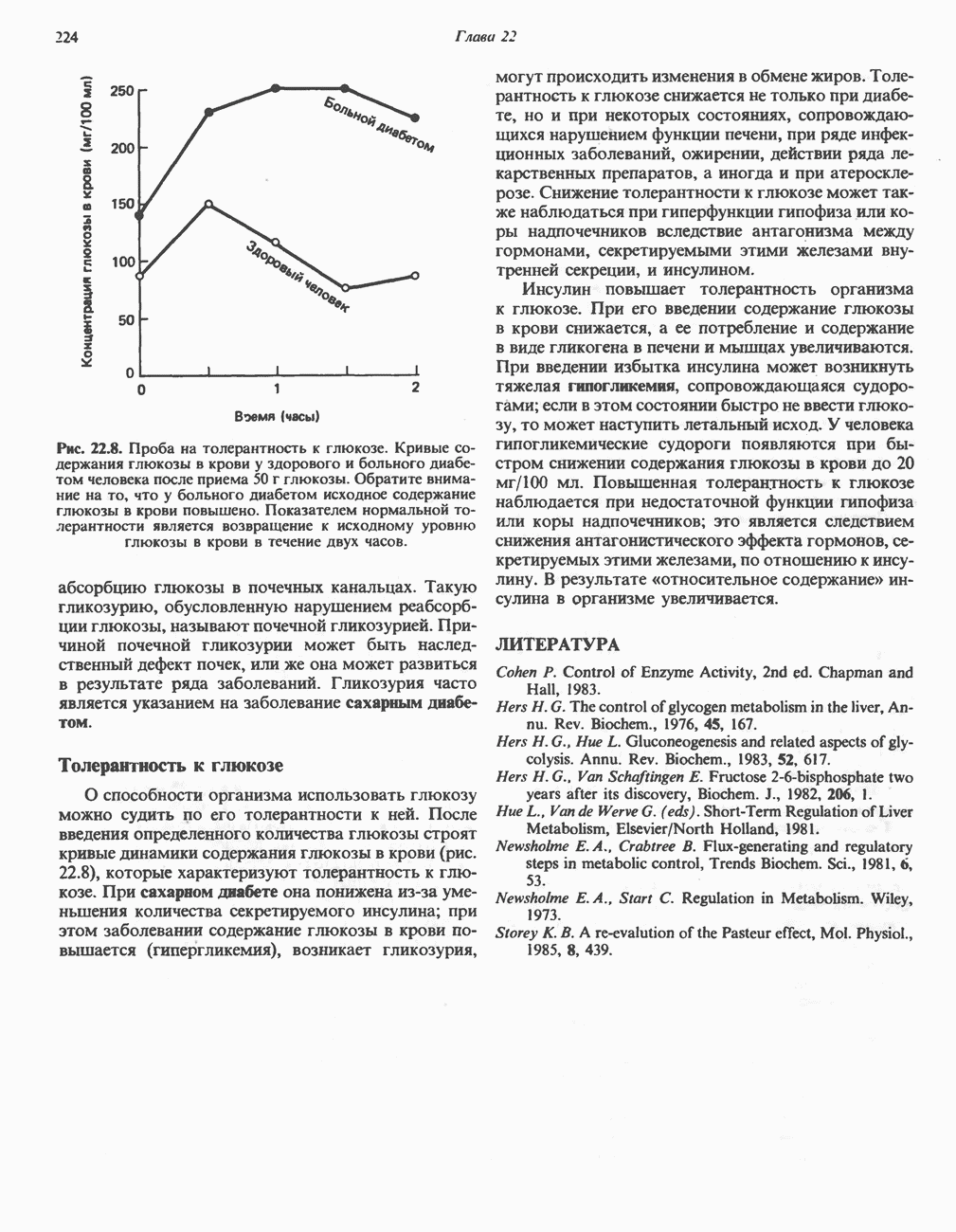
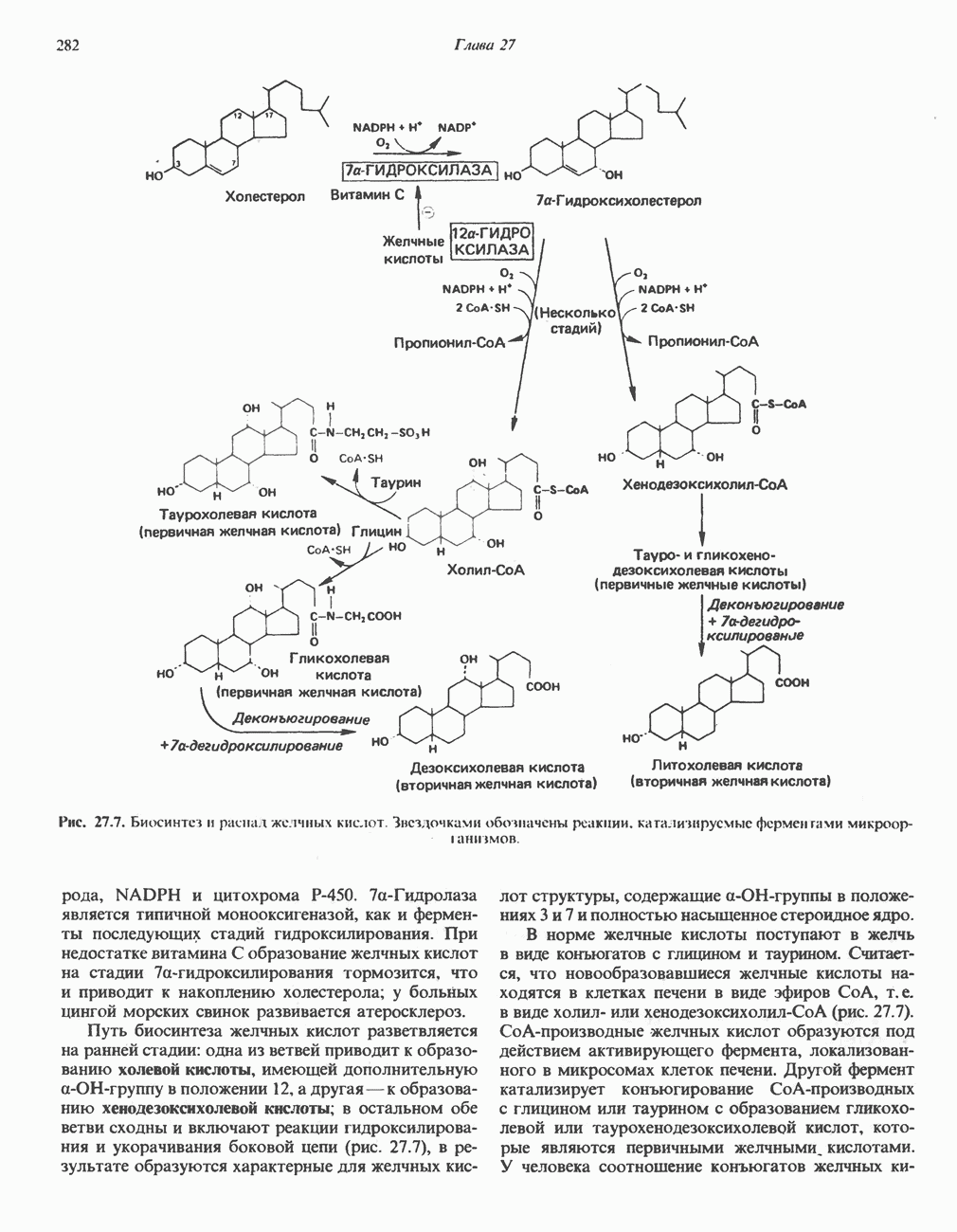
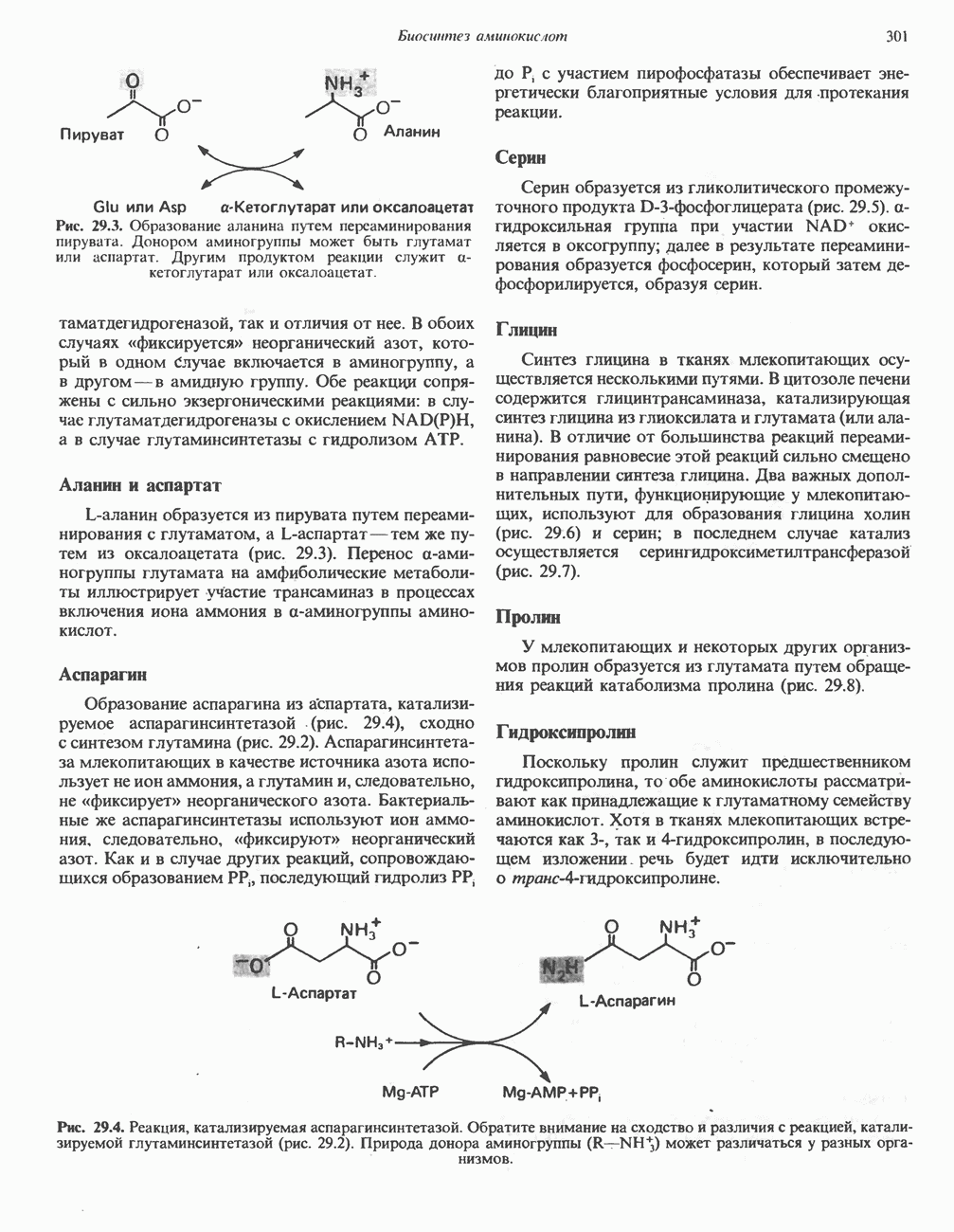
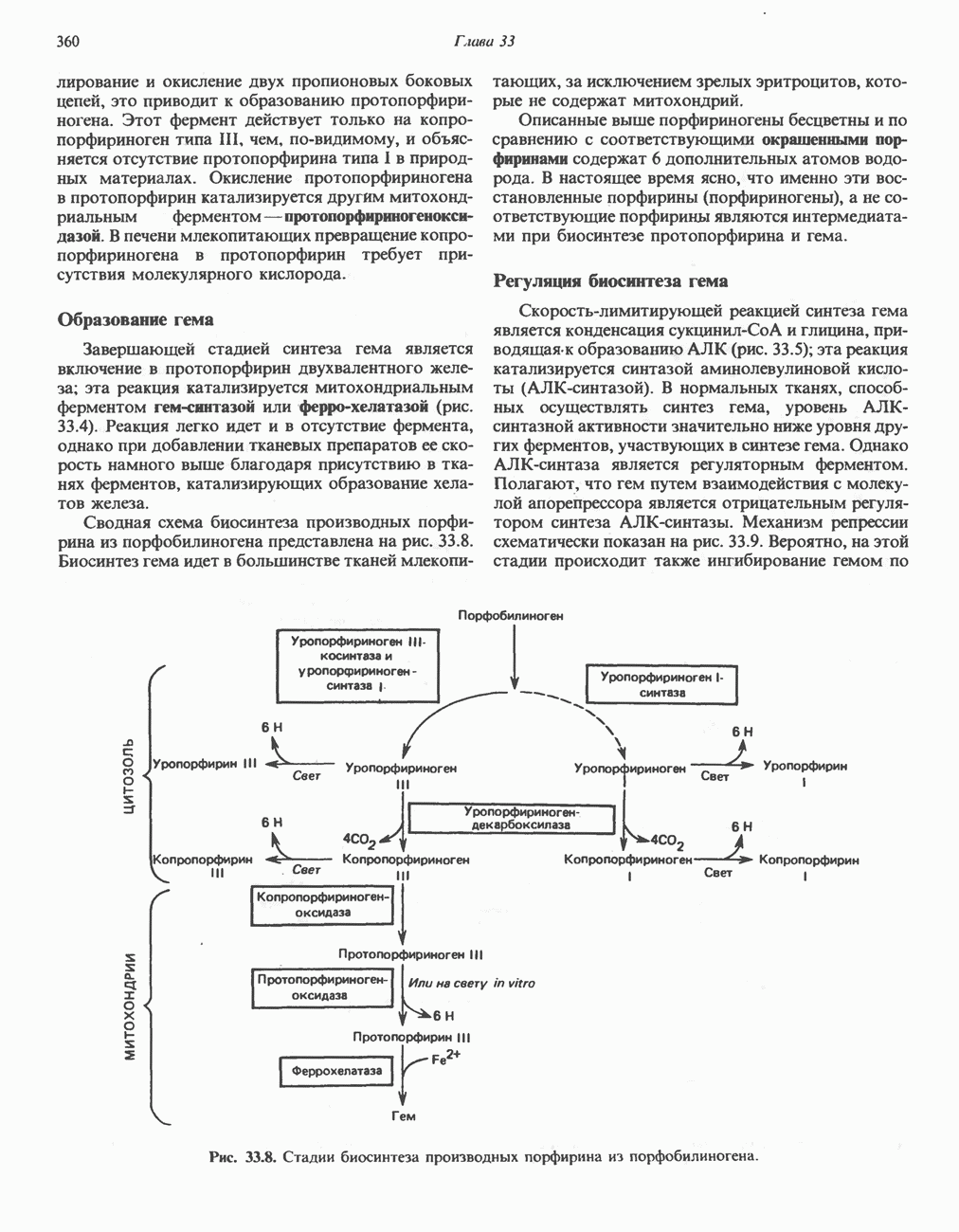
Клинические аспектыКетоз развивается при высокой скорости окисления жирных кислот в печени, особенно в тех случаях, когда оно происходит на фоне недостатка углеводов (см. с. 292). Подобное состояние возникает при приеме пищи, богатой жирами, голодании, сахарном диабете, кетозе у лактирующих коров и токсикозе беременности (кетозе) у овец. Ниже приводятся причины, вызывающие нарушение процесса окисления жирных кислот. Недостаток карнитина встречается у новорожденных, чаще всего недоношенных детей; он обусловлен либо нарушением биосинтеза карнитина; либо его «утечкой» в почках. Потери карнитина могут происходить при гемодиализе; больные, страдающие органической ацидурией, теряют большое количество карнитина, который экскретируется из организма в форме конъюгатов с органическими кислотами. Для восполнения потерь этого соединения некоторые пациенты нуждаются в особой диете, включающей продукты, содержащие карнитин. Признаками и симптомами недостатка карнитина являются приступы гипогликемии, возникающие из-за снижения глюконеогенеза в результате нарушения процесса - окисления жирных кислот, уменьшение образования кетоновых тел, сопровождающееся повышением содержания СЖК в плазме крови, мышечная слабость (миастения), а также накопление липидов. При лечении внутрь принимают препарат карнитина. Симптомы недостатка карнитина очень сходны с симптомами синдрома Рейе (Reye), при котором, однако, содержание карнитина является нормальным. Причина синдрома Рейе пока неизвестна. Снижение активности карнитинпальмитоилтрансферазы печени приводит к гипогликемии и понижению содержания кетоновых тел в плазме крови, а снижение активности карнитин-пальмитоилтраисферазы мышц — к нарушению процесса окисления жирных кислот, в результате чего периодически возникает мышечная слабость и развивается миоглобинурия. Ямайская рвотная болезнь возникает у людей после употребления в пищу незрелых плодов аки (Blig-hia sapida), которые содержат токсин гипоглицнн, инактивирующий ацил-СоА-дегидрогеназу, в результате чего ингибируется процесс При дикарбоновой ацидурии происходит экскреция Болезнь Рефсума является редким неврологическим заболеванием, которое вызывается накоплением в тканях фитановой кислоты, образующейся из фитола; последний входит в состав хлорофилла, поступающего в организм с продуктами растительного происхождения. Фитановая кислота содержит метальную группу у третьего атома углерода, это блокирует ее (см. скан) Рис. 23.4. Последовательность реакций окисления ненасыщенных жирных кислот на примере, линолевой кислоты. удаляется при а-окислении, но у людей, страдающих болезнью Рефсума, имеется врожденное нарушение системы а-окисления, что приводит к накоплению фитановой кислоты в тканях. Синдром Цельвегера (Zellweger) или цереброгепаторенальный синдром является редким наследственным заболеванием, при котором во всех тканях отсутствуют пероксисомы. У больных, страдающих синдромом Цельвегера, в мозгу накапливаются Окисление ненасыщенных жирных кислотСоА-производные ненасыщенных жирных кислот атакуются ферментами, катализирующими Р-окисление до стадии, на которой в зависимости от локализации двойных связей образуется либо Перекисное окисление полиненасыщенных жирных кислот в микросомахNADPH-зависимое перекисное окисление ненасыщенных жирных кислот катализируется ферментами, локализованными в микросомах (см. с. 124). Антиоксиданты, например БГТ (бутилированный гидрокситолуол) и а-токоферол (витамин Е), ингибируют перекисное окисление липидов в микросомах.
|
 |
Оглавление
|


































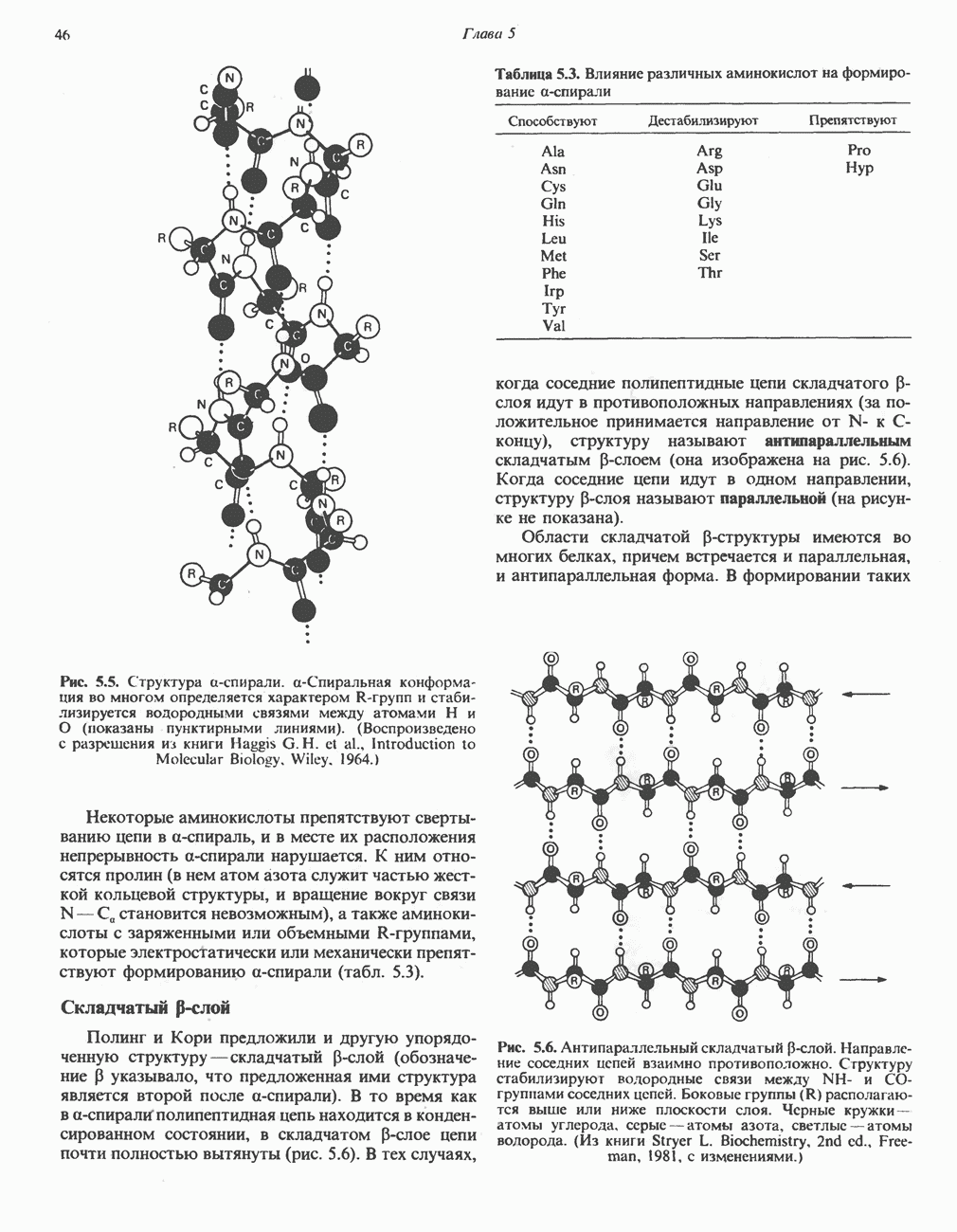
















































































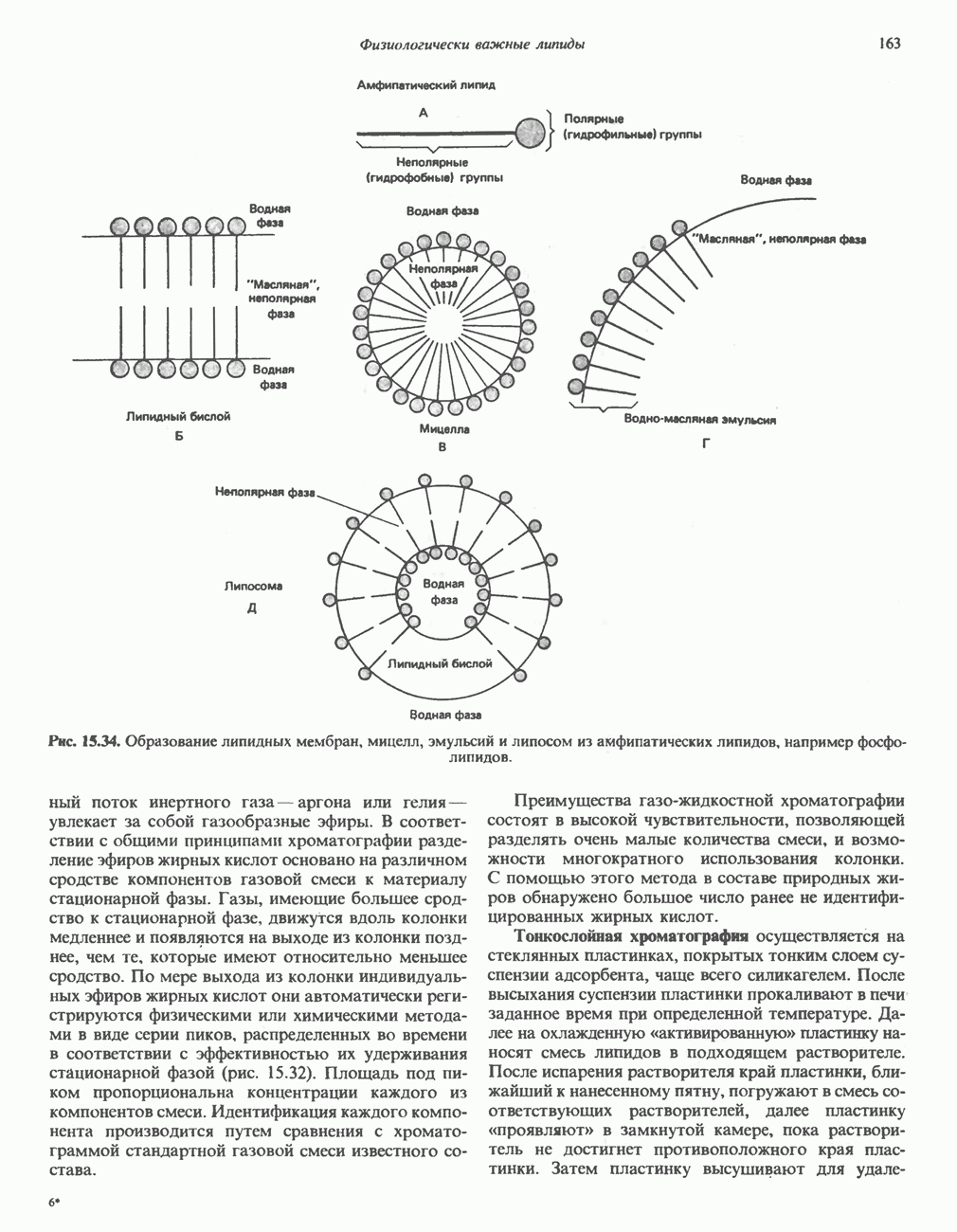






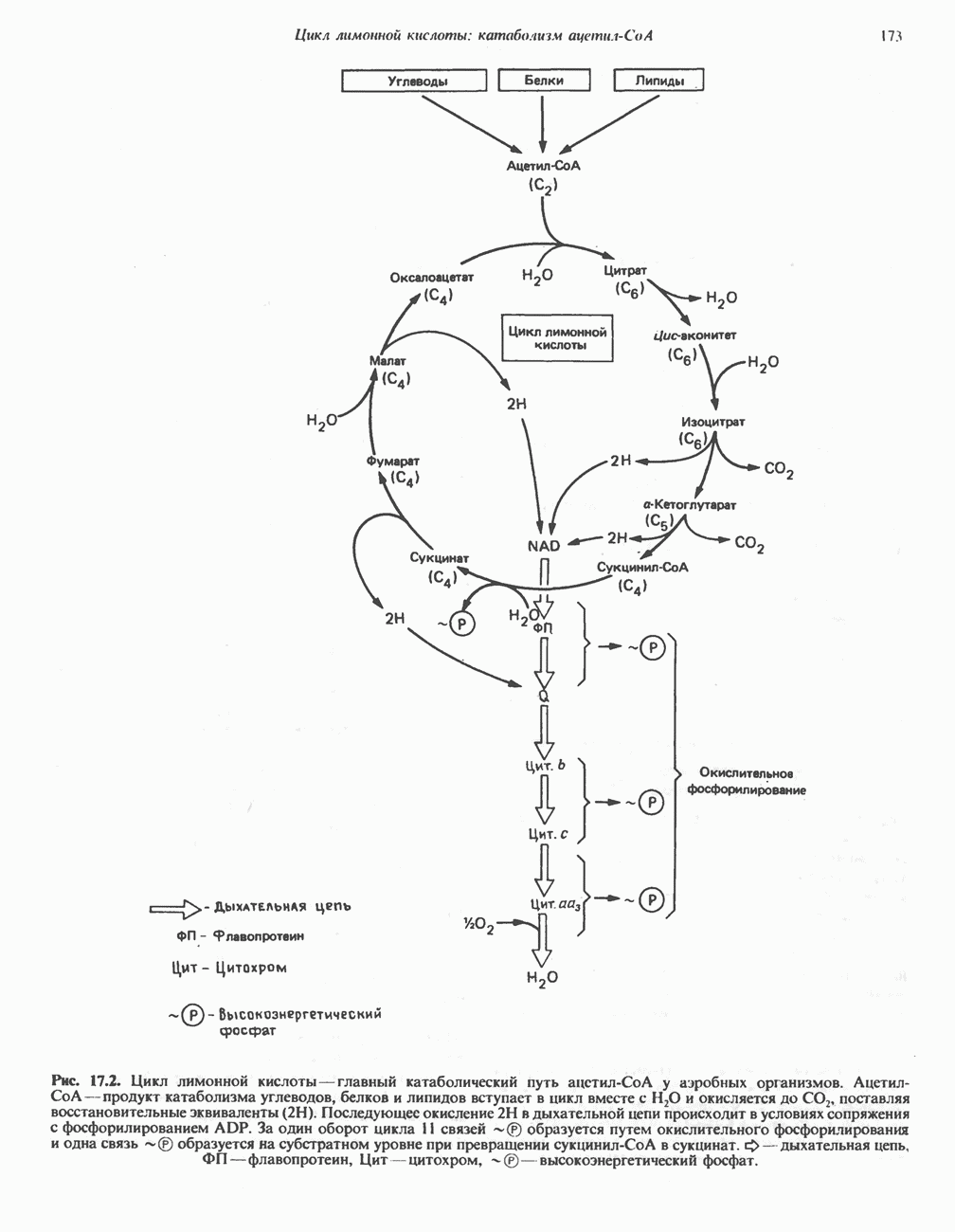

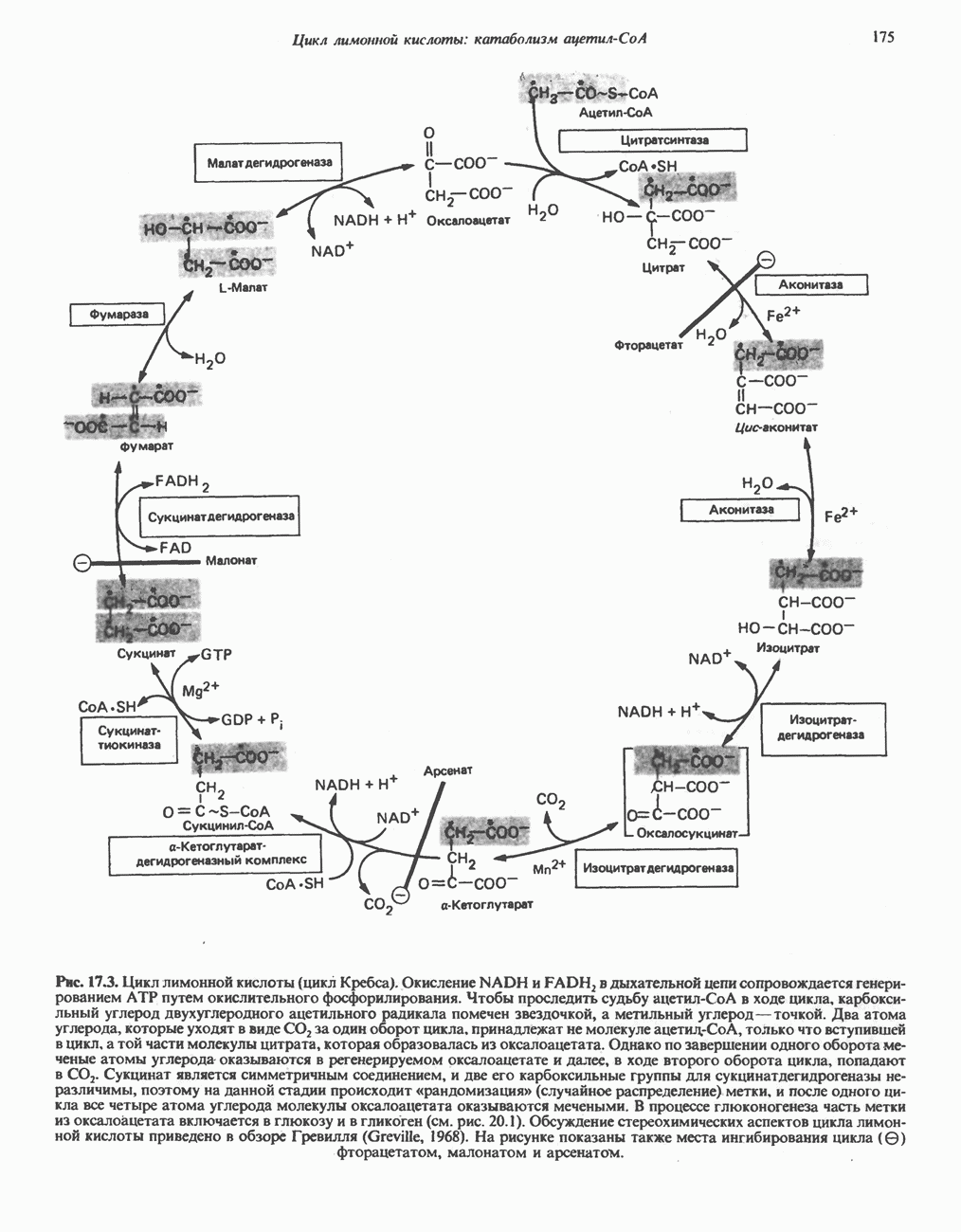























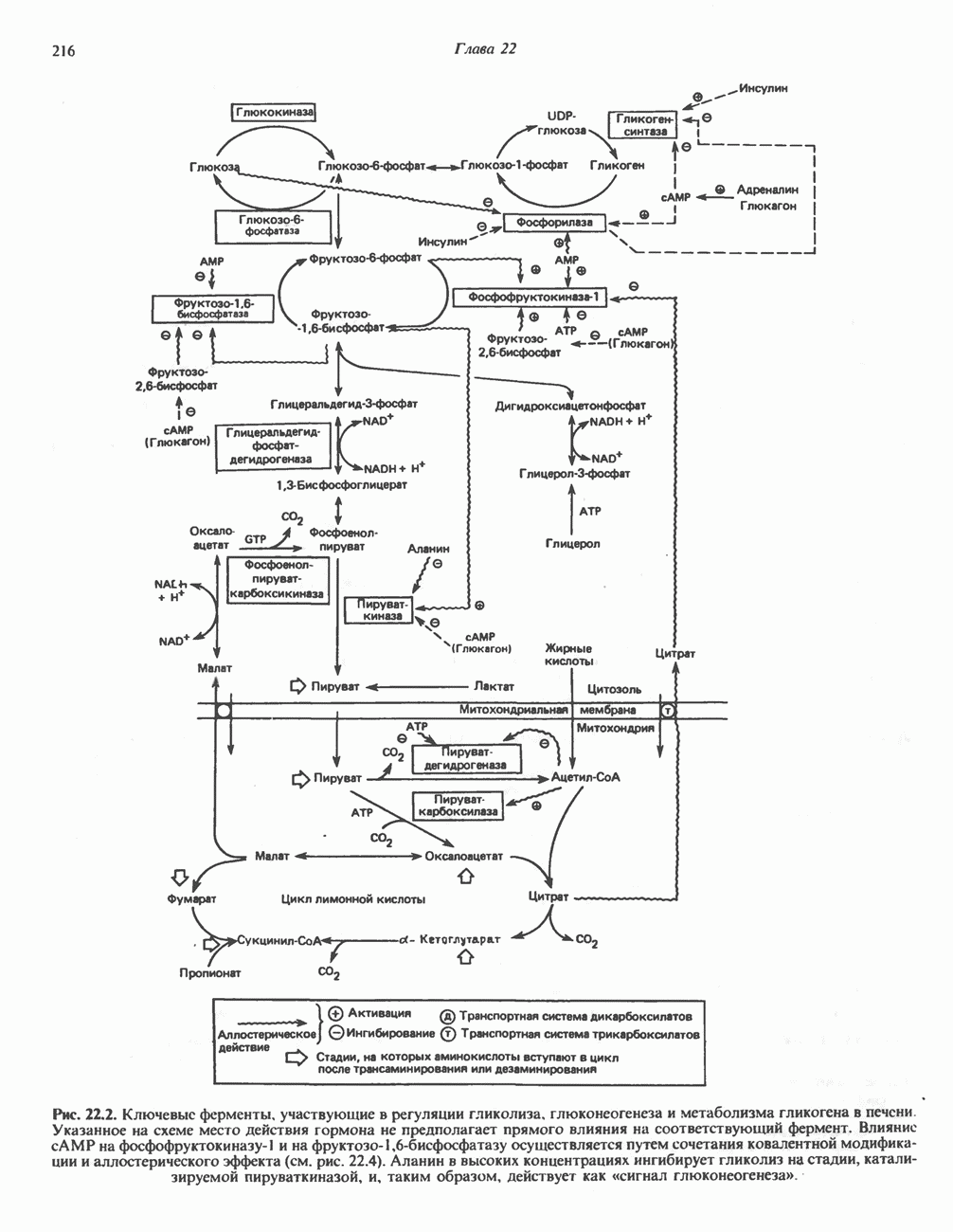
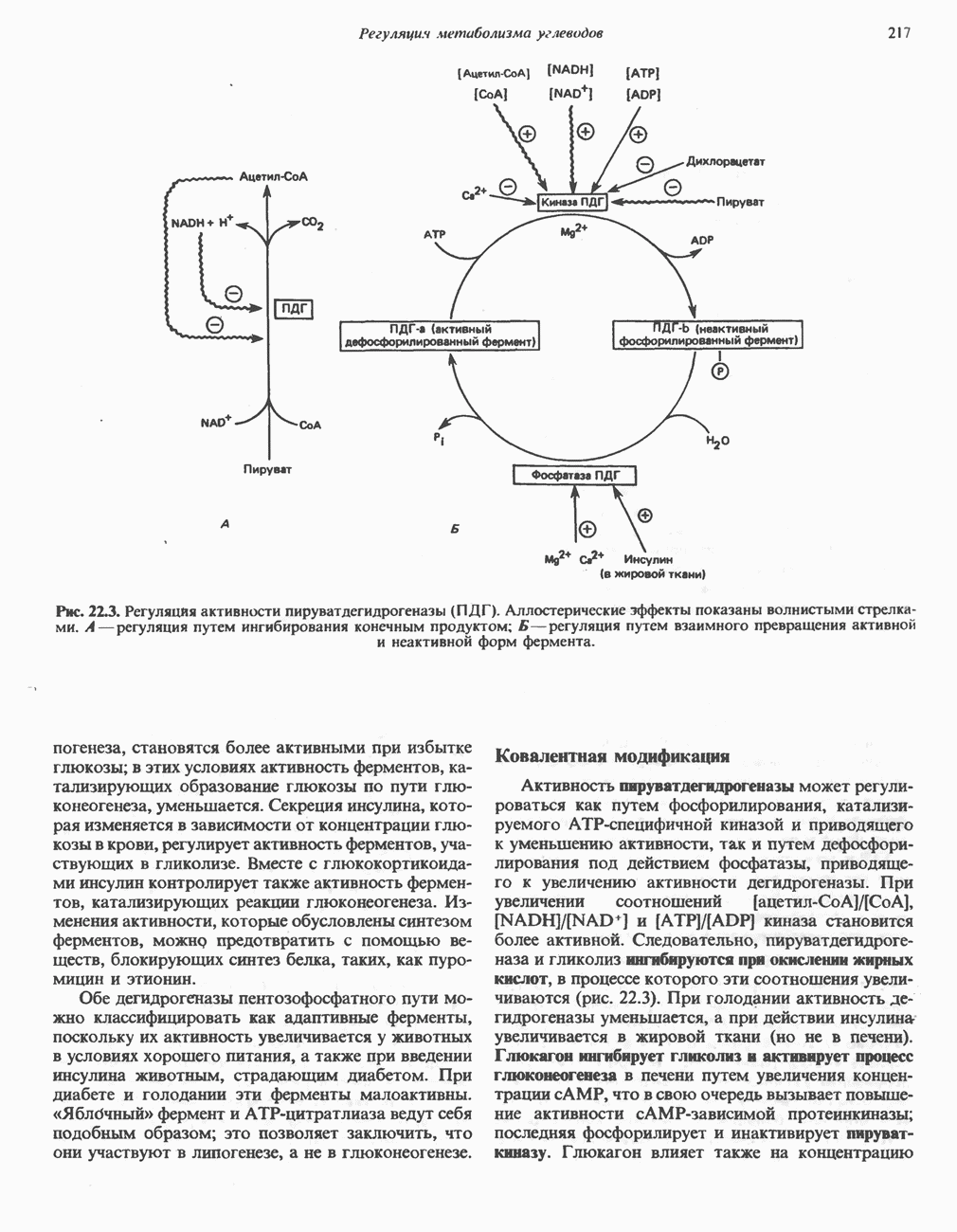

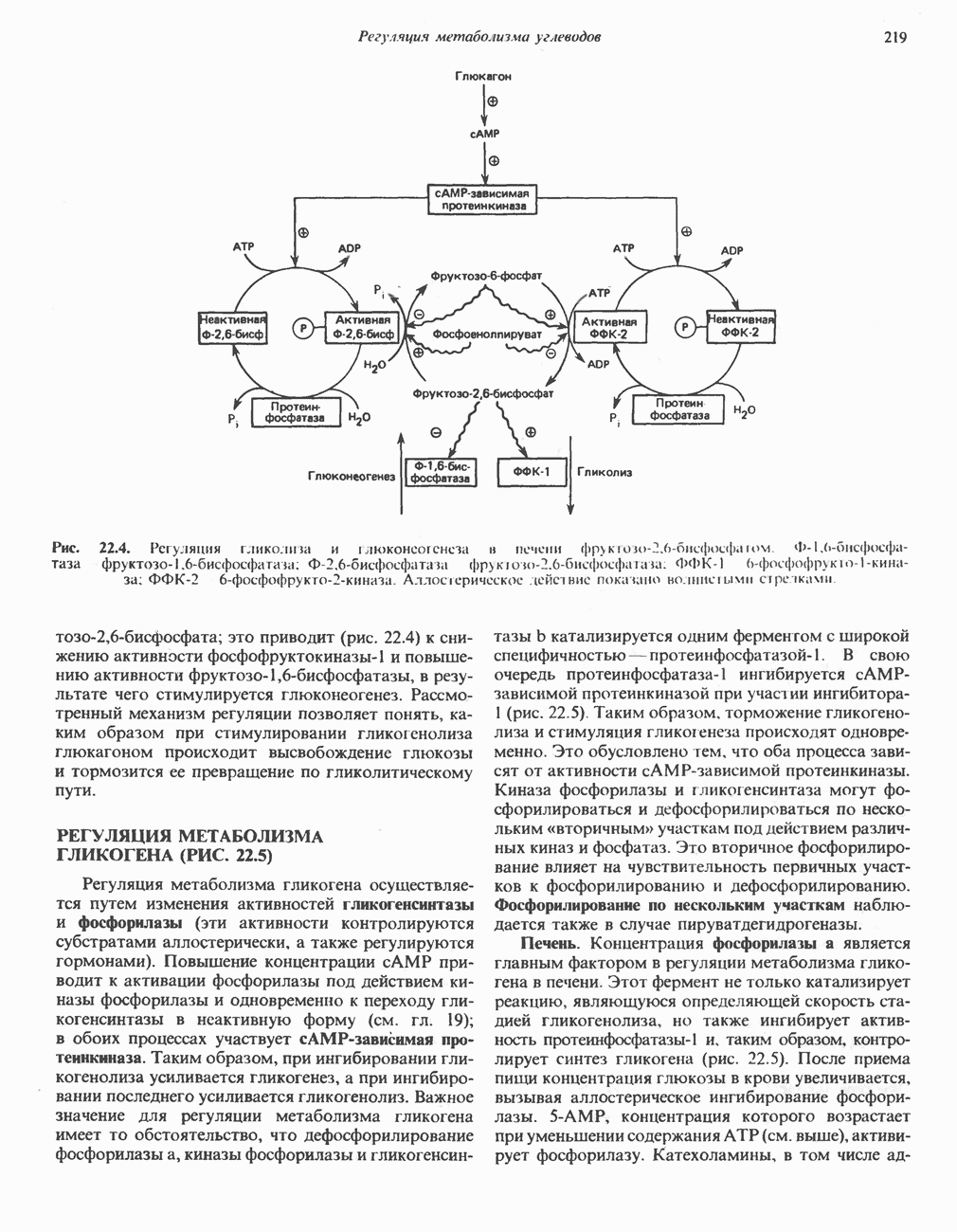
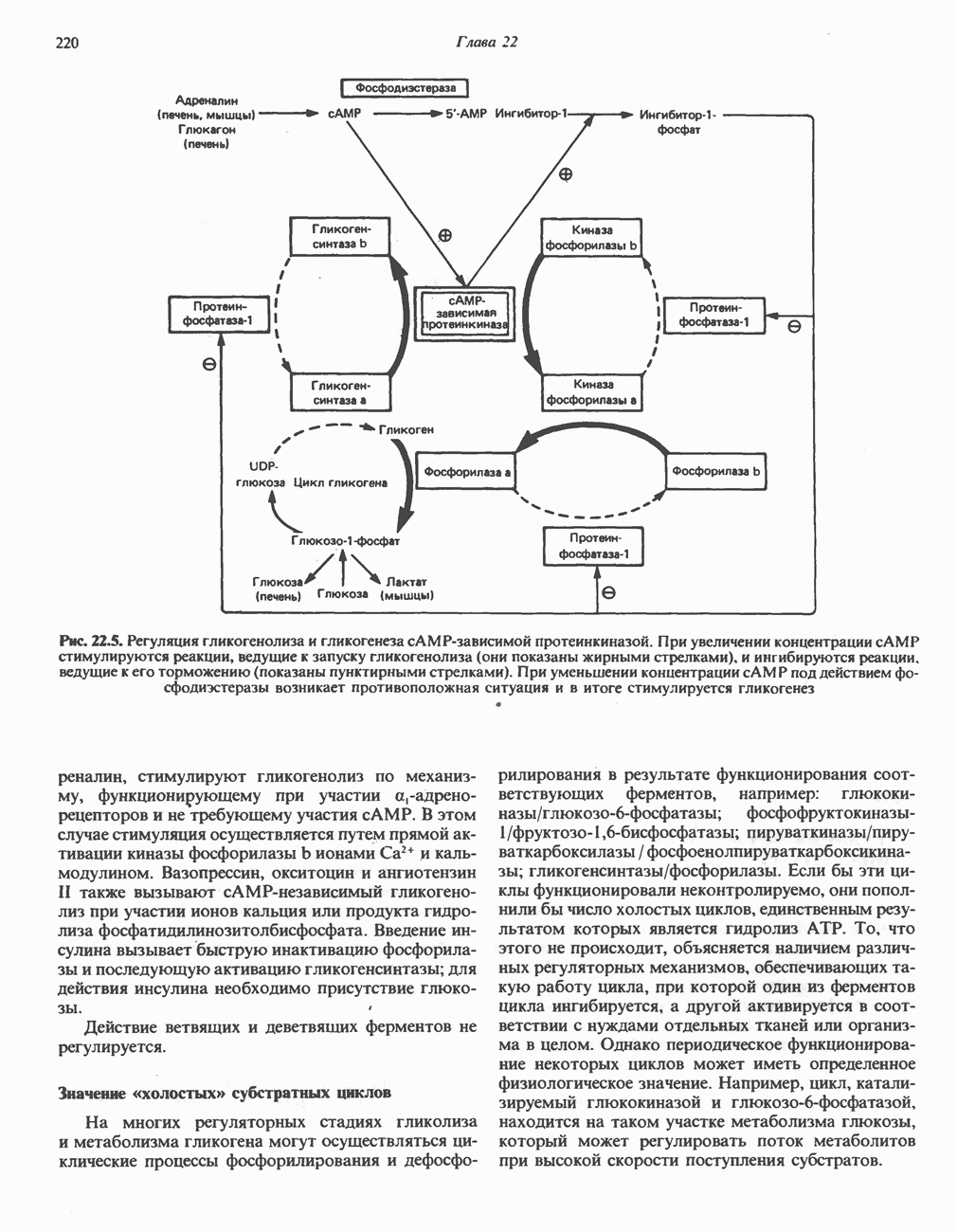






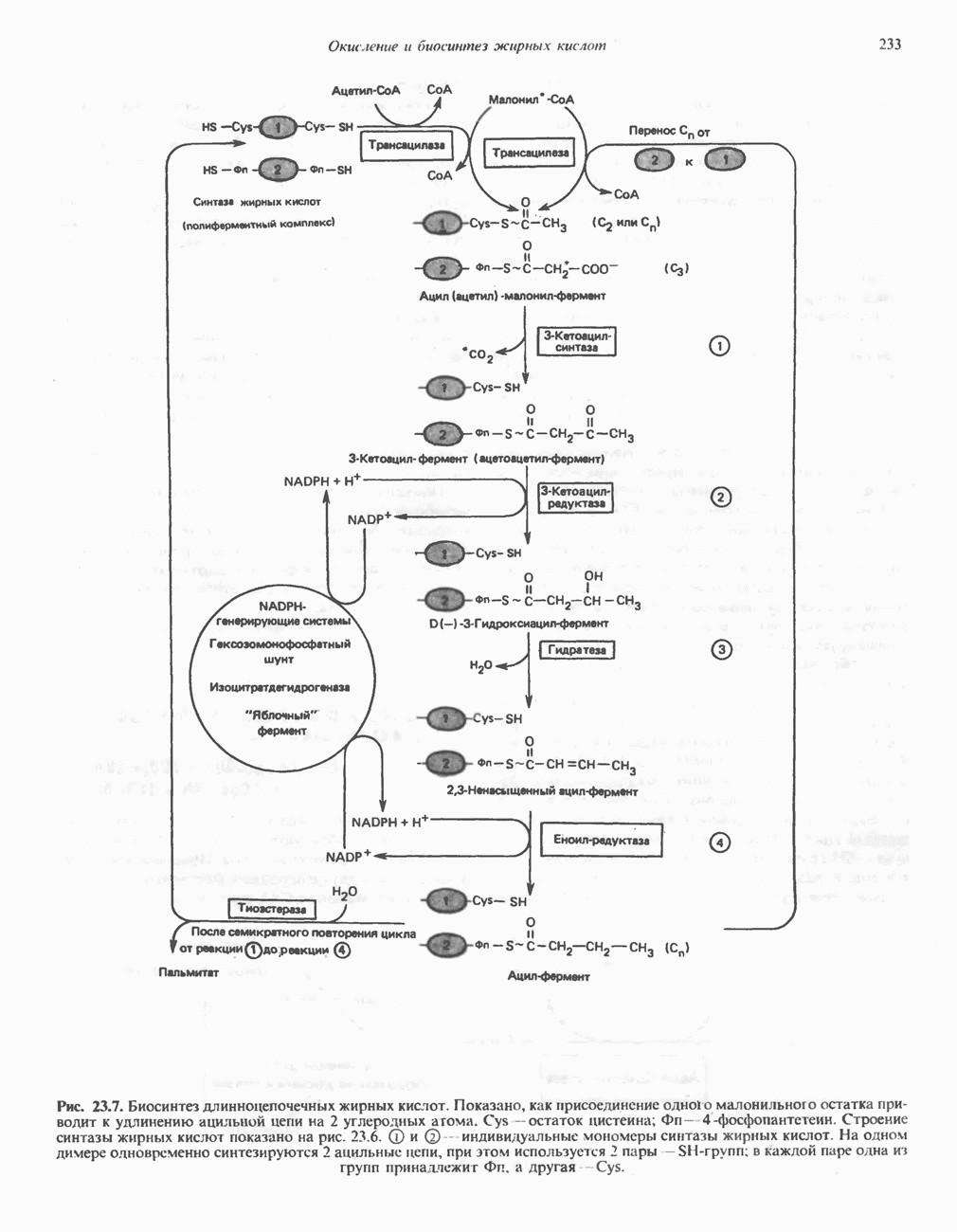

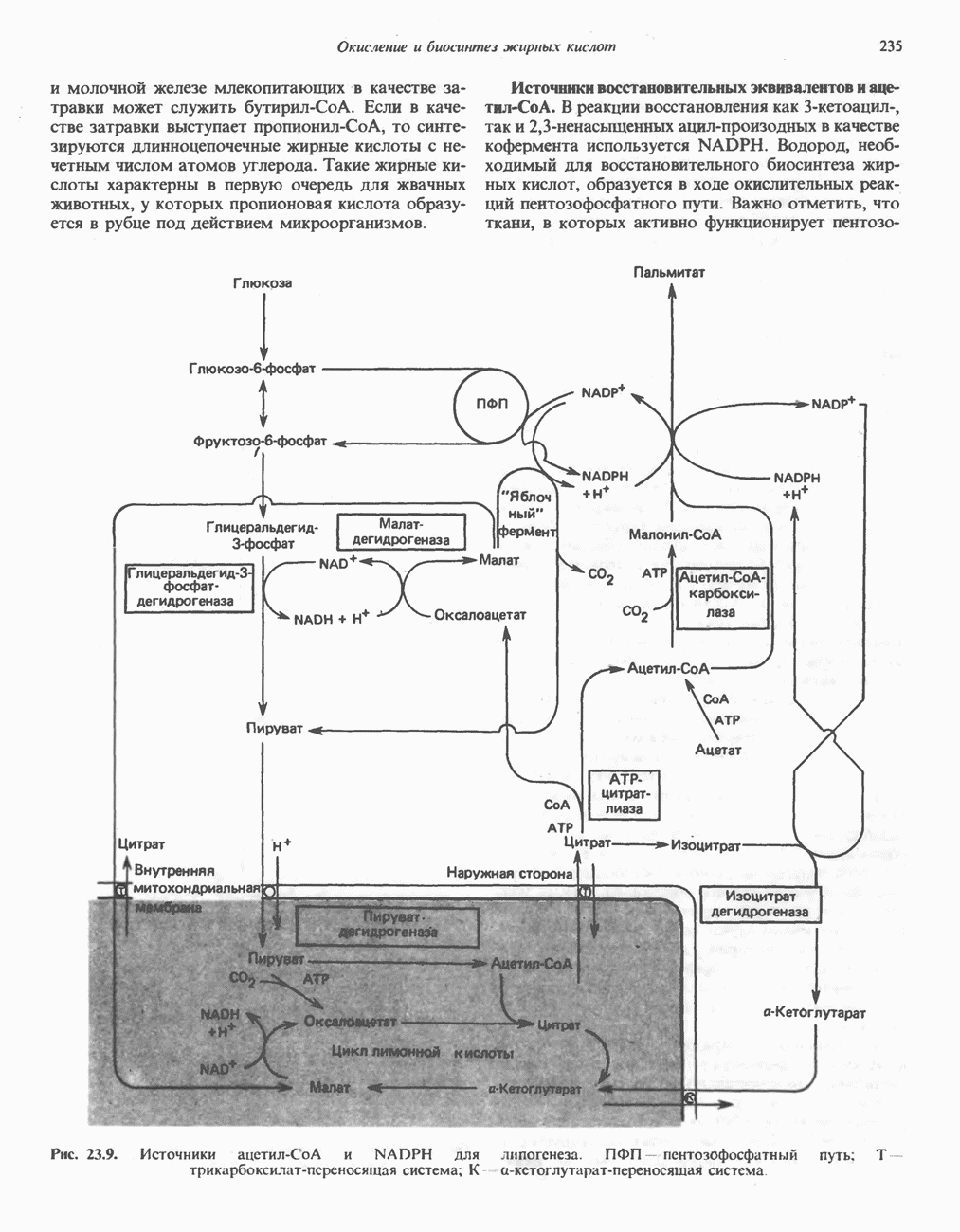



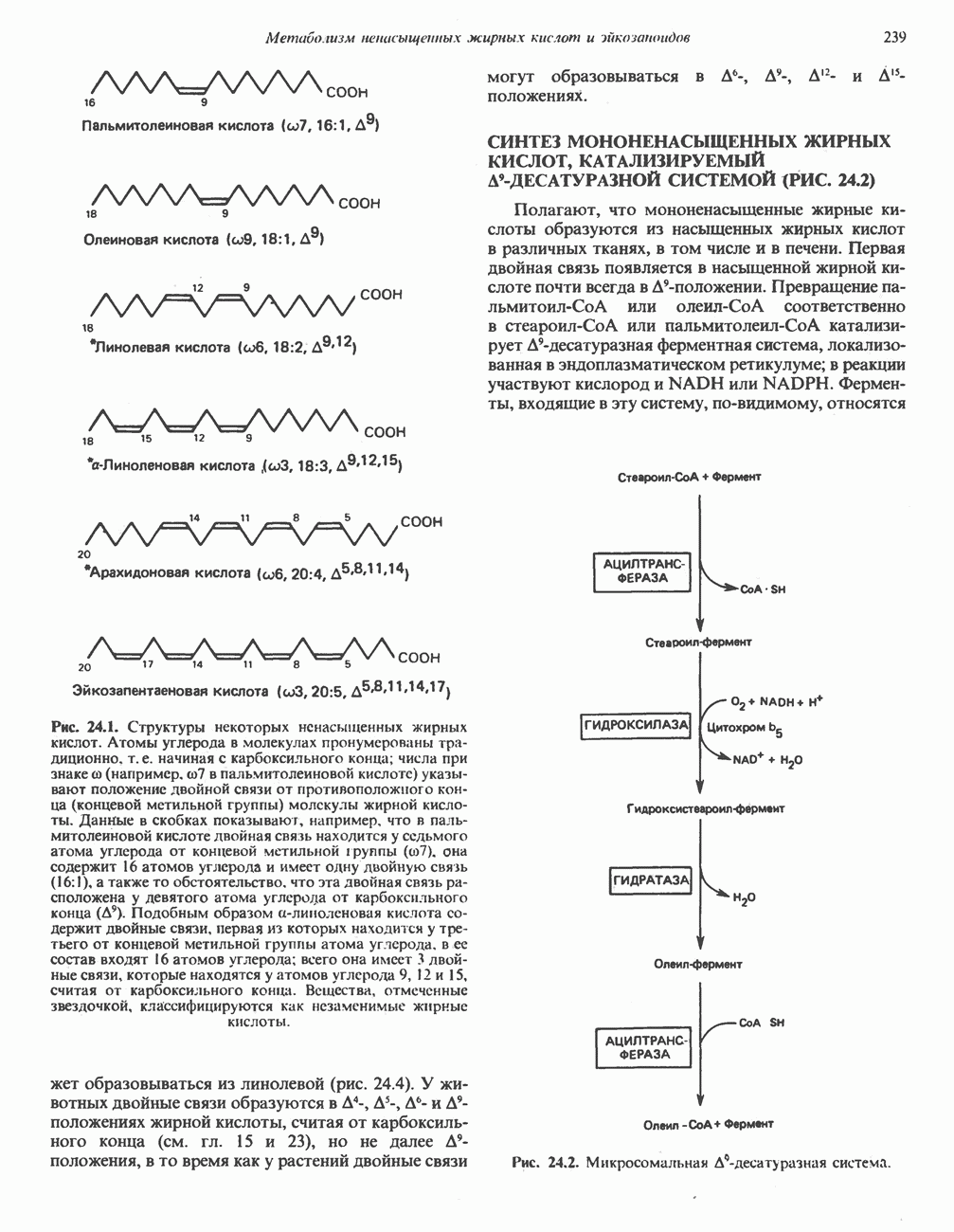
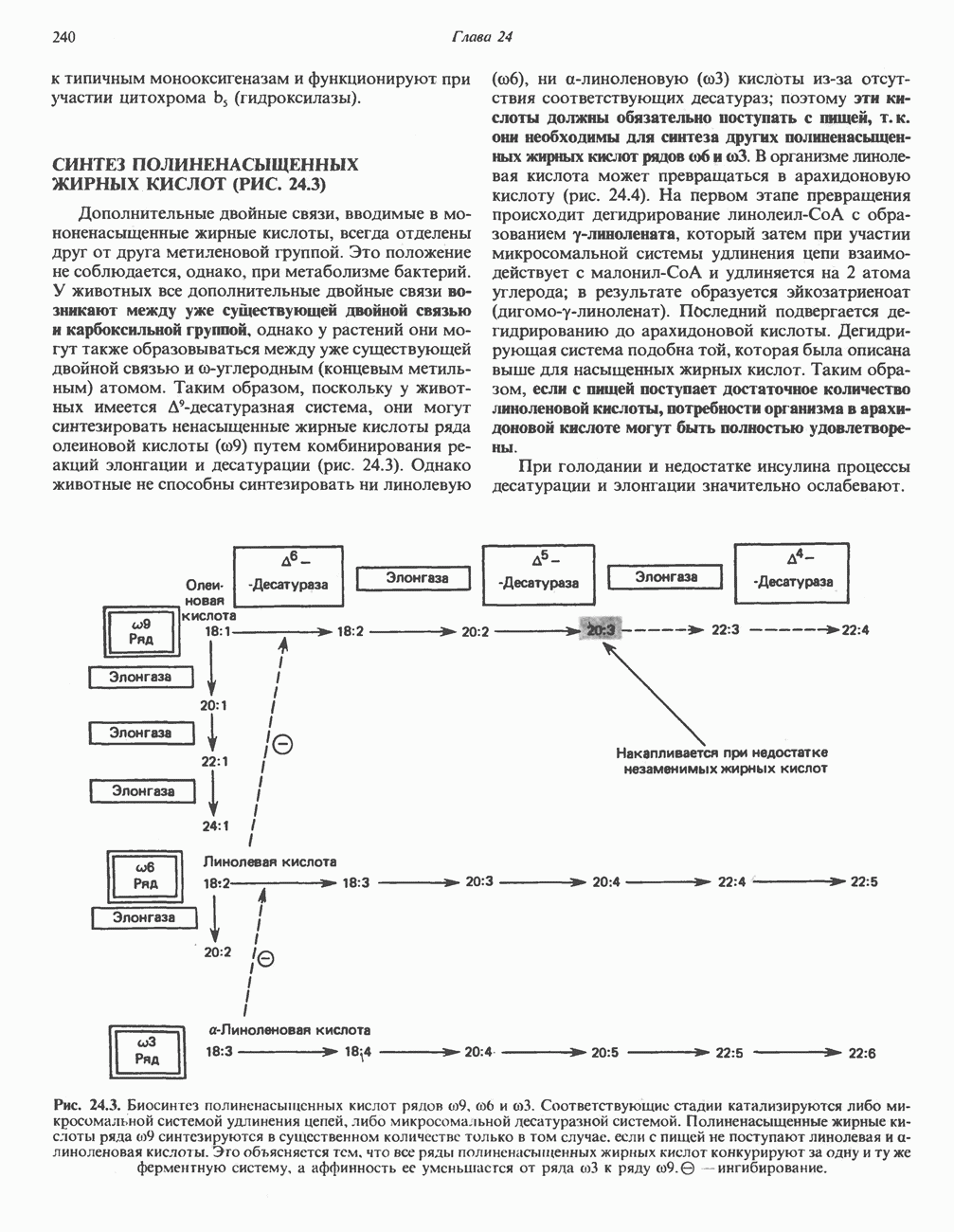
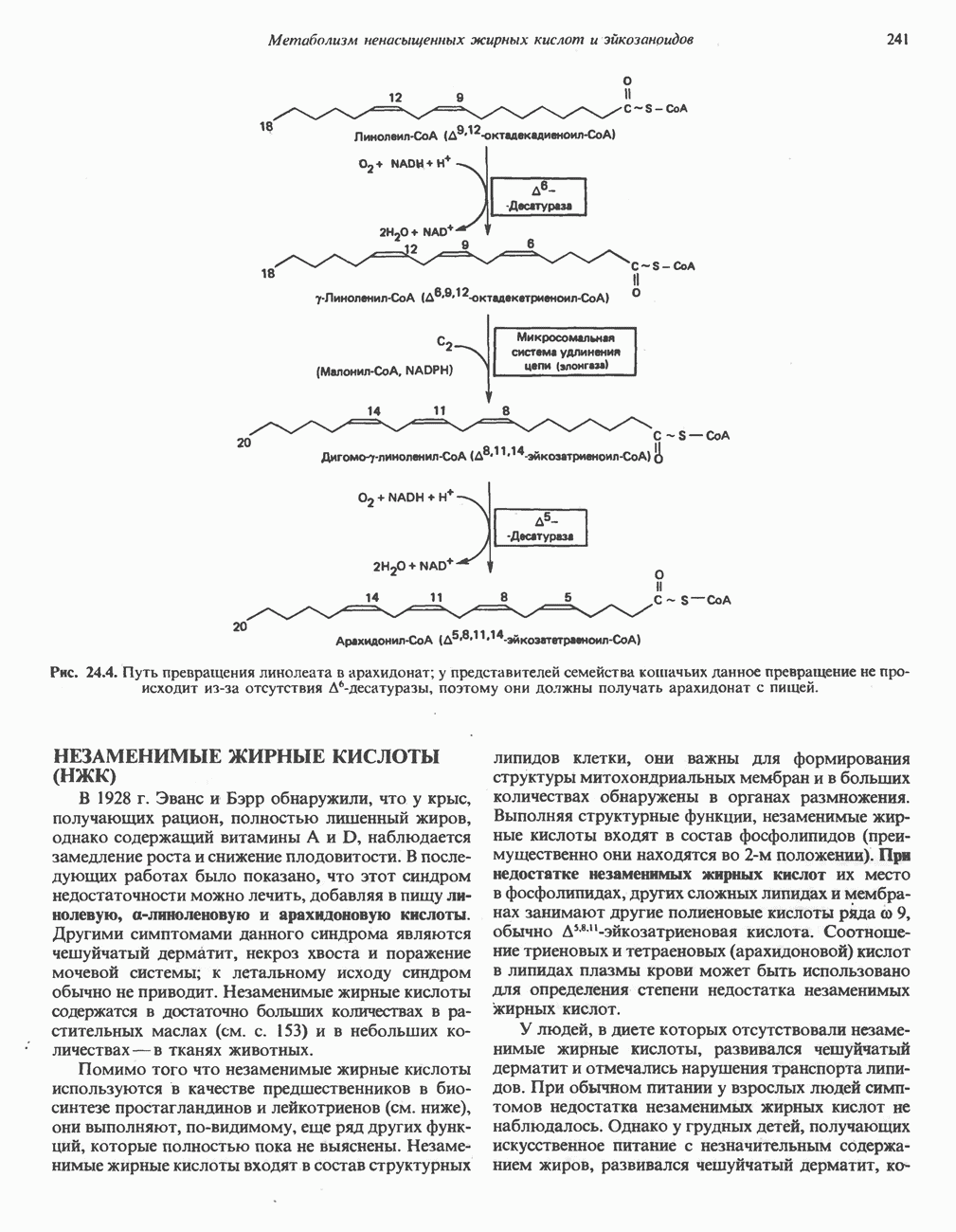

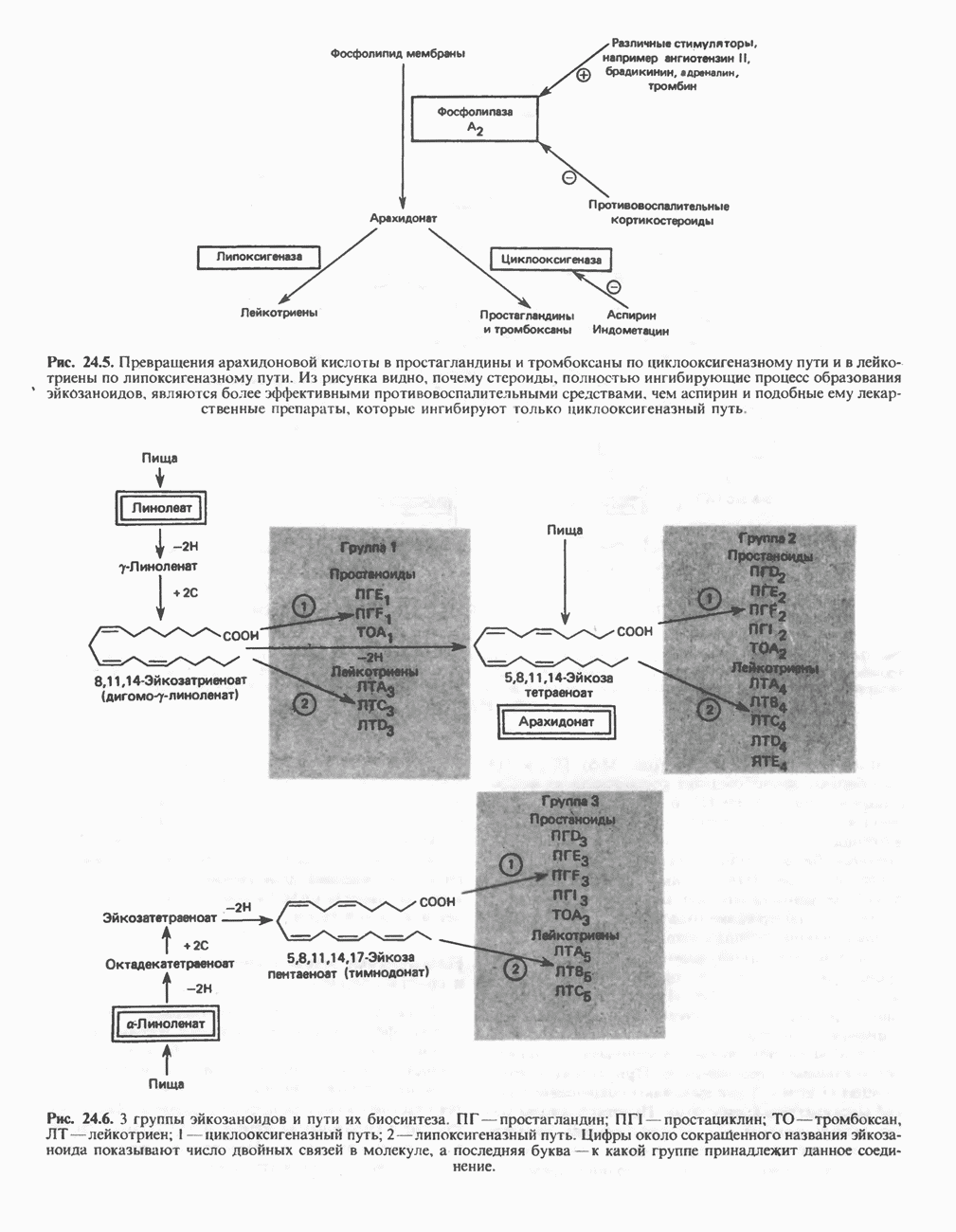







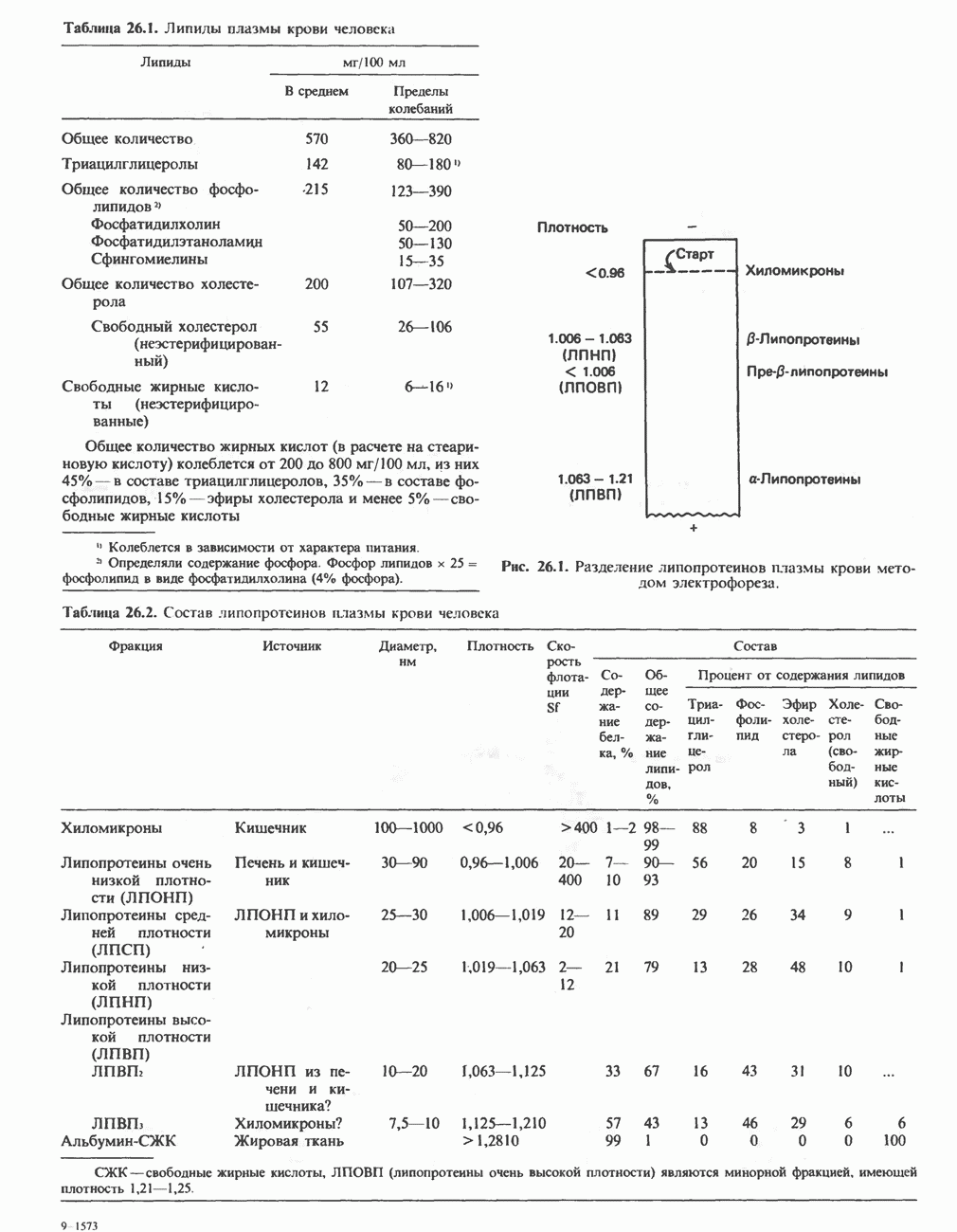





































































 является широко распространенным соединением,
является широко распространенным соединением,

 -окисления.
-окисления.



 -окисления жирных кислот.
-окисления жирных кислот.
 , откуда и возникло название
, откуда и возникло название  -окисление. Образующиеся двухуглеродные фрагменты представляют собой ацетил-СоА. Так, в случае пальмитоил-СоА образуется 8 молекул ацетил-СоА.
-окисление. Образующиеся двухуглеродные фрагменты представляют собой ацетил-СоА. Так, в случае пальмитоил-СоА образуется 8 молекул ацетил-СоА.
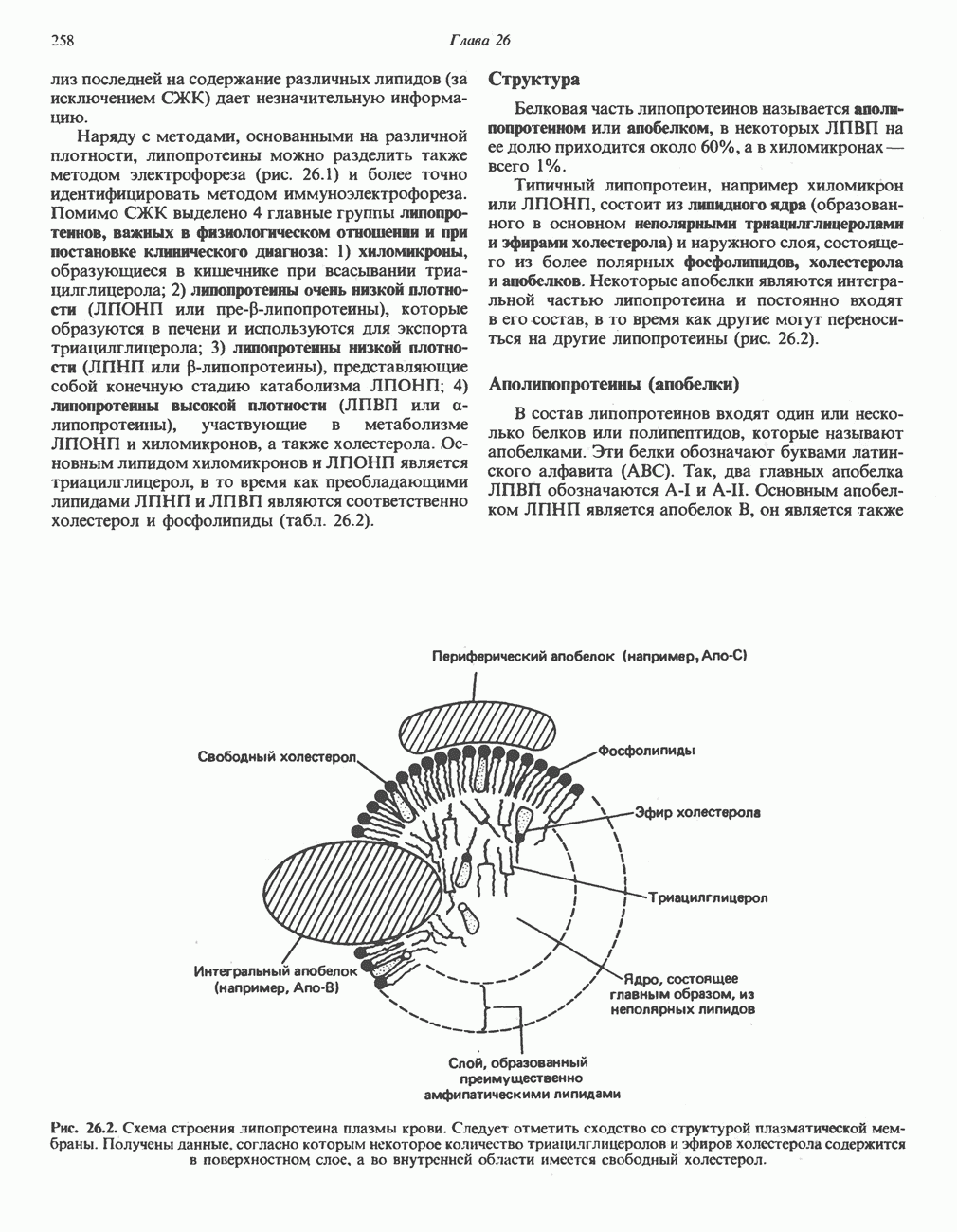
 происходит отщепление двух атомов водорода от углеродных атомов в положениях
происходит отщепление двух атомов водорода от углеродных атомов в положениях  катализируемое ацил-СоА-дегидрогеназои. Продуктом этой реакции является
катализируемое ацил-СоА-дегидрогеназои. Продуктом этой реакции является  . Фермент представляет собой флавопротеин, его простетической группой служит FAD. Окисление последнего в дыхательной цепи митохондрий происходит при участии другого флавопротеина. названного электронпереносящим флавопротеином [см. с. 123). Далее происходит гидратация двойной связи, в результате чего образуется 3-гидроксиацил-СоА. Эта реакция катализируется ферментом А2-еноил-СоА-гидратазой. Затем 3-гидроксиацил-ОоА дегидрируется по 3-му атому углерода с образованием 3-кетоацил-СоА; эта реакция катализируется 3-гидроксиацил-СоА-дегидрогеназой при,участии в качестве кофермента NAD. 3-Кетоацил-СоА расщепляется между вторым и третьим атомами углерода 3-кетотиолазой или ацетил-СоА-ацнлтрансферазой с образованием ацетил-СоА- и ацил-СоА-производного, которое на 2 атома углерода короче исходной молекулы ацил-СоА. Это тиолитическое расщепление требует участия еще одной молекулы
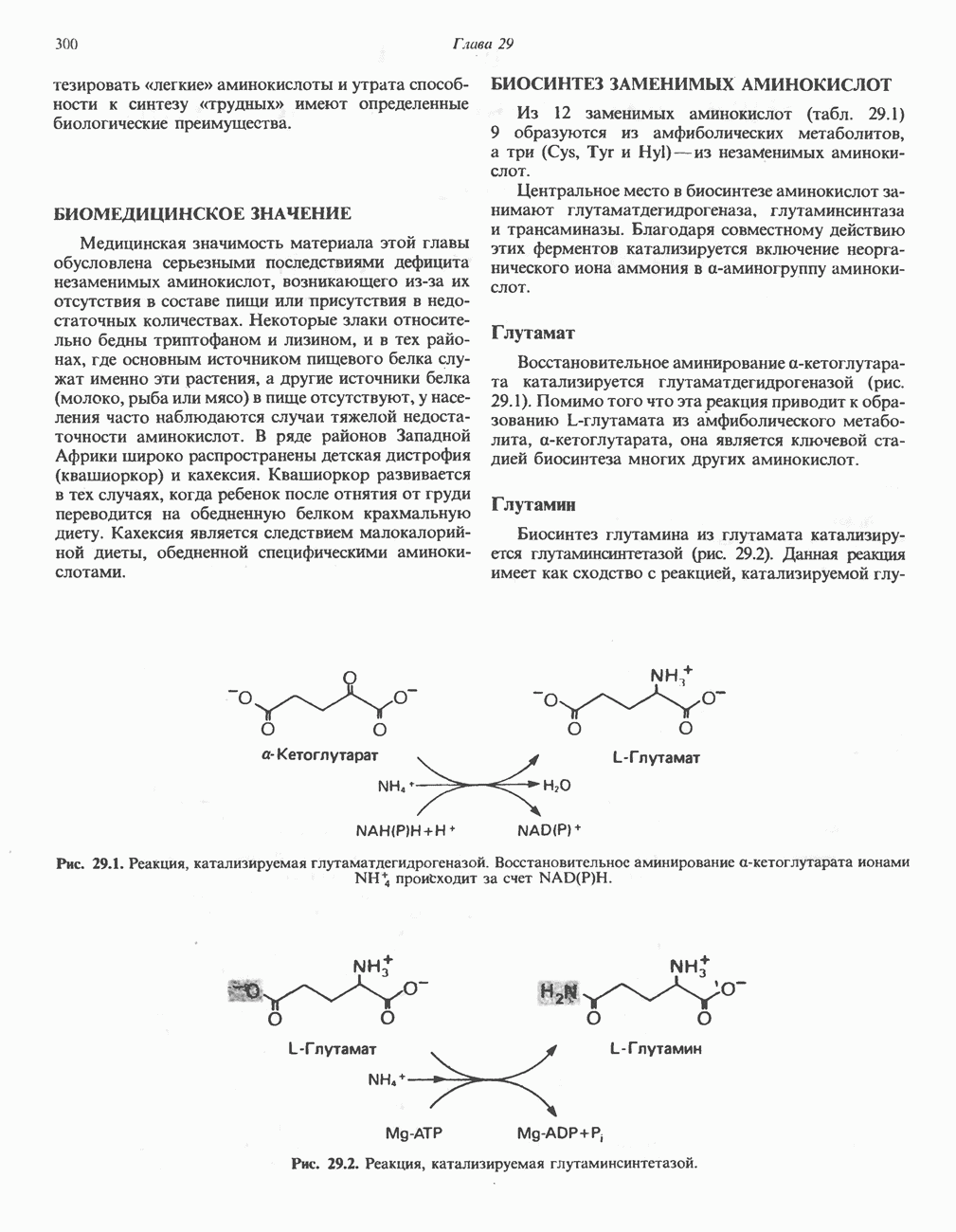
. Фермент представляет собой флавопротеин, его простетической группой служит FAD. Окисление последнего в дыхательной цепи митохондрий происходит при участии другого флавопротеина. названного электронпереносящим флавопротеином [см. с. 123). Далее происходит гидратация двойной связи, в результате чего образуется 3-гидроксиацил-СоА. Эта реакция катализируется ферментом А2-еноил-СоА-гидратазой. Затем 3-гидроксиацил-ОоА дегидрируется по 3-му атому углерода с образованием 3-кетоацил-СоА; эта реакция катализируется 3-гидроксиацил-СоА-дегидрогеназой при,участии в качестве кофермента NAD. 3-Кетоацил-СоА расщепляется между вторым и третьим атомами углерода 3-кетотиолазой или ацетил-СоА-ацнлтрансферазой с образованием ацетил-СоА- и ацил-СоА-производного, которое на 2 атома углерода короче исходной молекулы ацил-СоА. Это тиолитическое расщепление требует участия еще одной молекулы  Образующийся укороченный ацил-СоА вновь вступает в цикл Р-окисления, начиная с реакции 2 (рис. 23.3). Таким путем длинноцепочечные жирные кислоты могут полностью расщепляться до ацетил-СоА (С2-фрагментов); последние в цикле лимонной кислоты, который протекает в митохондриях, окисляются до
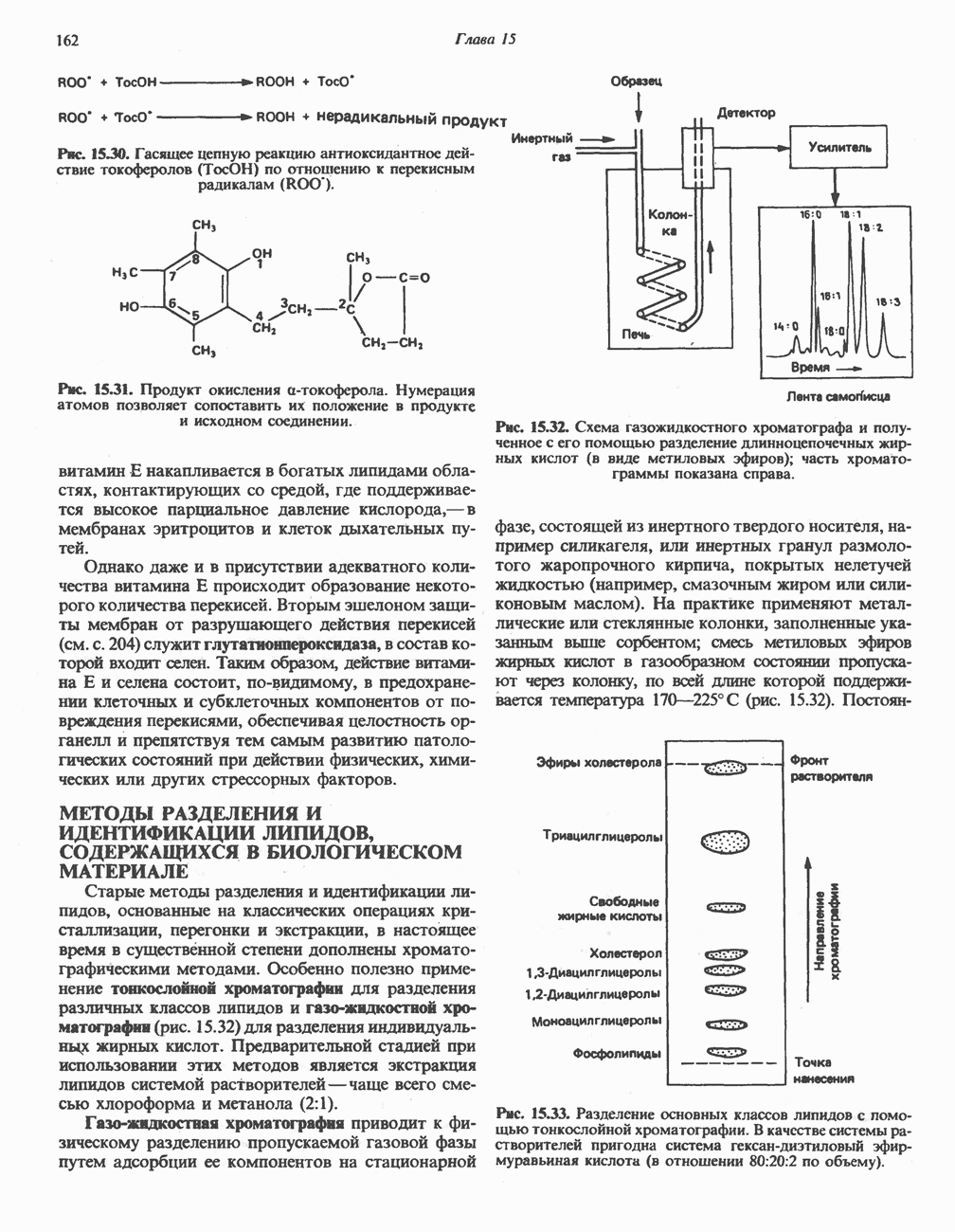
Образующийся укороченный ацил-СоА вновь вступает в цикл Р-окисления, начиная с реакции 2 (рис. 23.3). Таким путем длинноцепочечные жирные кислоты могут полностью расщепляться до ацетил-СоА (С2-фрагментов); последние в цикле лимонной кислоты, который протекает в митохондриях, окисляются до 
 являющийся интермедиатом цикла лимонной кислоты (см. также рис. 20.2).
являющийся интермедиатом цикла лимонной кислоты (см. также рис. 20.2).
 Всего образуется 8 молекул ацетил-СоА, и каждая из них, проходя через цикл лимонной кислоты, обеспечивает синтез 12 богатых энергией связей. Всего в расчете на молекулу пальмитата по этому пути генерируется 8 х 12 = 96 богатых энергией фосфатных связей. Если учесть две связи, необходимые для активации
Всего образуется 8 молекул ацетил-СоА, и каждая из них, проходя через цикл лимонной кислоты, обеспечивает синтез 12 богатых энергией связей. Всего в расчете на молекулу пальмитата по этому пути генерируется 8 х 12 = 96 богатых энергией фосфатных связей. Если учесть две связи, необходимые для активации
 кДж. Поскольку свободная энергия сгорания пальмитиновой кислоты составляет
кДж. Поскольку свободная энергия сгорания пальмитиновой кислоты составляет  то на долю энергии, запасаемой в виде фосфатных связей при окислении жирной кислоты, приходится около 40%.
то на долю энергии, запасаемой в виде фосфатных связей при окислении жирной кислоты, приходится около 40%.
 -окисление жирных кислот происходит в модифицированном виде. Продуктами окисления в данном случае являются ацетил-СоА и
-окисление жирных кислот происходит в модифицированном виде. Продуктами окисления в данном случае являются ацетил-СоА и  , последняя образуется на стадии, катализируемой связанной с флавопротеином дегидрогеназой. Этот путь окисления непосредственно не сопряжен с фосфорилированием и образованием АТР, но он обеспечивает расщепление жирных кислот с очень длинной цепью (например,
, последняя образуется на стадии, катализируемой связанной с флавопротеином дегидрогеназой. Этот путь окисления непосредственно не сопряжен с фосфорилированием и образованием АТР, но он обеспечивает расщепление жирных кислот с очень длинной цепью (например,  ); он включается при диете, богатой жирами, или приеме гиполипидемических лекарственных препаратов, таких, как клофибрат. Ферменты пероксисом не атакуют жирные кислоты с короткими цепями, и процесс Р-окисления останавливается при образовании октаноил-СоА. Октаноильные и ацетильные группы удаляются затем из пероксисом в виде октаноилкарнитина и ацетилкарнитина и окисляются в митохондриях.
); он включается при диете, богатой жирами, или приеме гиполипидемических лекарственных препаратов, таких, как клофибрат. Ферменты пероксисом не атакуют жирные кислоты с короткими цепями, и процесс Р-окисления останавливается при образовании октаноил-СоА. Октаноильные и ацетильные группы удаляются затем из пероксисом в виде октаноилкарнитина и ацетилкарнитина и окисляются в митохондриях.
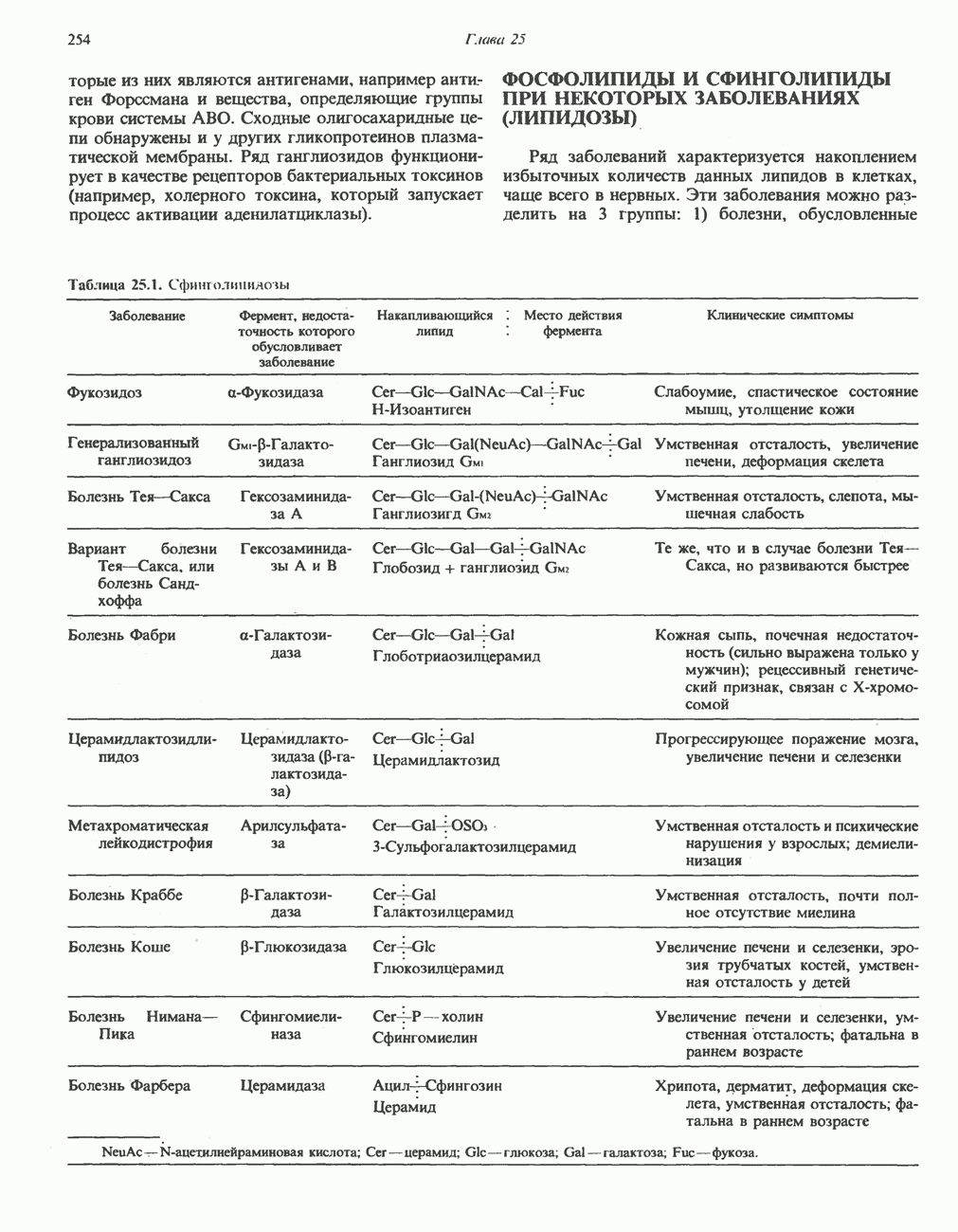
 -Окисление является основным путем катаболизма жирных кислот. Однако недавно было обнаружено, что в тканях мозга происходит
-Окисление является основным путем катаболизма жирных кислот. Однако недавно было обнаружено, что в тканях мозга происходит  -окисление жирных кислот, т. е. последовательное отщепление одноуглеродных фрагментов от карбоксильного конца молекулы. В этом процессе участвуют интермедиаты, содержащие
-окисление жирных кислот, т. е. последовательное отщепление одноуглеродных фрагментов от карбоксильного конца молекулы. В этом процессе участвуют интермедиаты, содержащие  он не сопровождается образованием богатых энергией фосфатных связей.
он не сопровождается образованием богатых энергией фосфатных связей.
 -Окисление жирных кислот в норме весьма незначительно. Этот тип окисления, катализируемый гидроксилазами при участии цитохрома
-Окисление жирных кислот в норме весьма незначительно. Этот тип окисления, катализируемый гидроксилазами при участии цитохрома  с. 123), протекает в эндоплазматическом
с. 123), протекает в эндоплазматическом  -Группа превращается в —
-Группа превращается в — -группу, которая затем окисляется до —СООН; в результате образуется дикарбоновая кислота. Последняя расщепляется путем Р-окисления обычно до адипиновой
-группу, которая затем окисляется до —СООН; в результате образуется дикарбоновая кислота. Последняя расщепляется путем Р-окисления обычно до адипиновой  и субериновой
и субериновой  кислот, которые затем удаляются с мочой.
кислот, которые затем удаляются с мочой.
 -окисления.
-окисления.
 кислот и развивается гипогликемия, не связанная с повышением содержания кетоновых тел. Причиной данного заболевания является отсутствие в митохондриях ацил-СоА-дегидрогеназы среднецепочечных жирных кислот. При этом нарушается
кислот и развивается гипогликемия, не связанная с повышением содержания кетоновых тел. Причиной данного заболевания является отсутствие в митохондриях ацил-СоА-дегидрогеназы среднецепочечных жирных кислот. При этом нарушается  -окисление и усиливается
-окисление и усиливается  -окисление длинноцепочечных жирных кислот, которые укорачиваются до среднецепочечных дикарбоновых кислот, выводимых из организма.
-окисление длинноцепочечных жирных кислот, которые укорачиваются до среднецепочечных дикарбоновых кислот, выводимых из организма.
 -окисление. В норме эта метильная группа
-окисление. В норме эта метильная группа
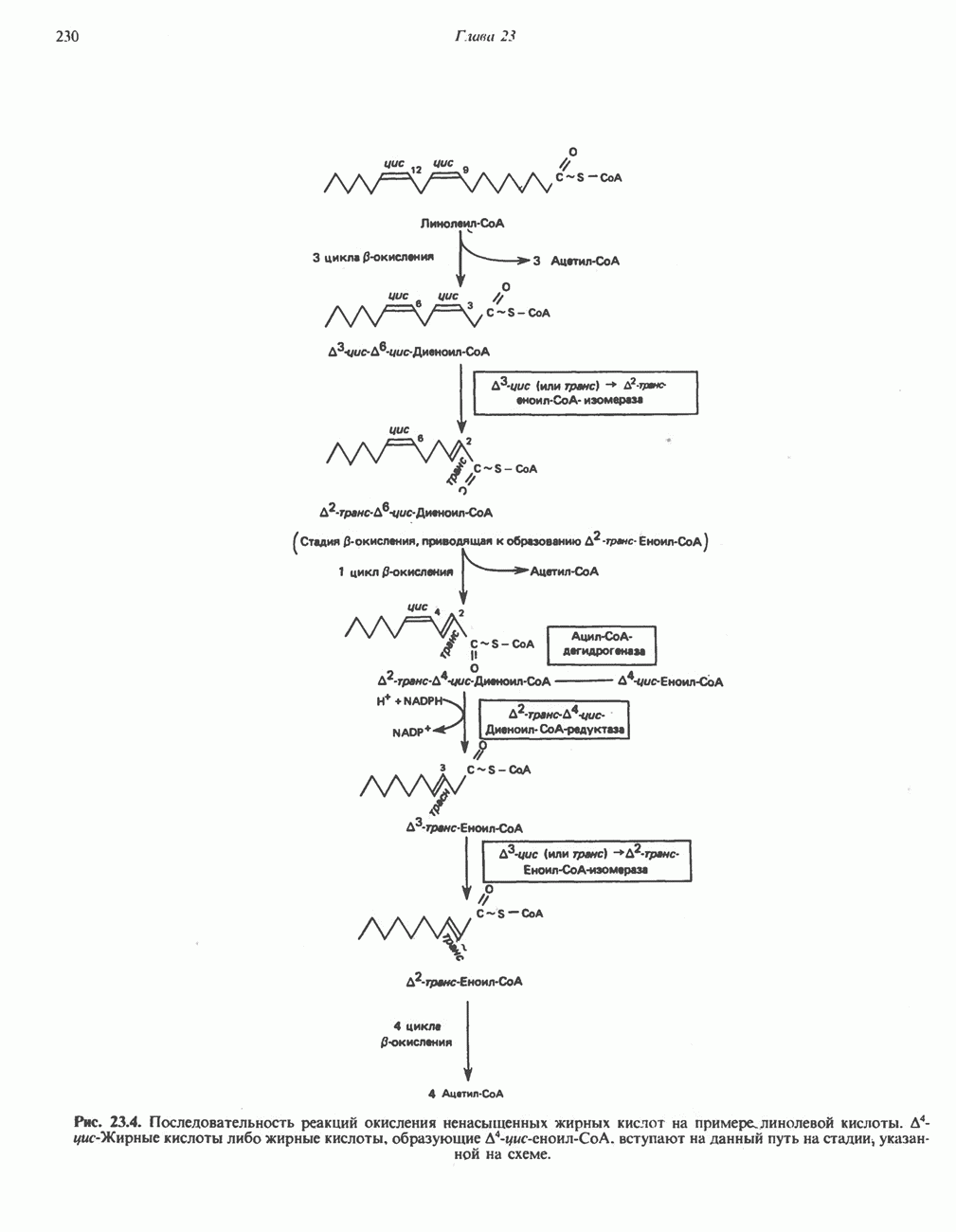
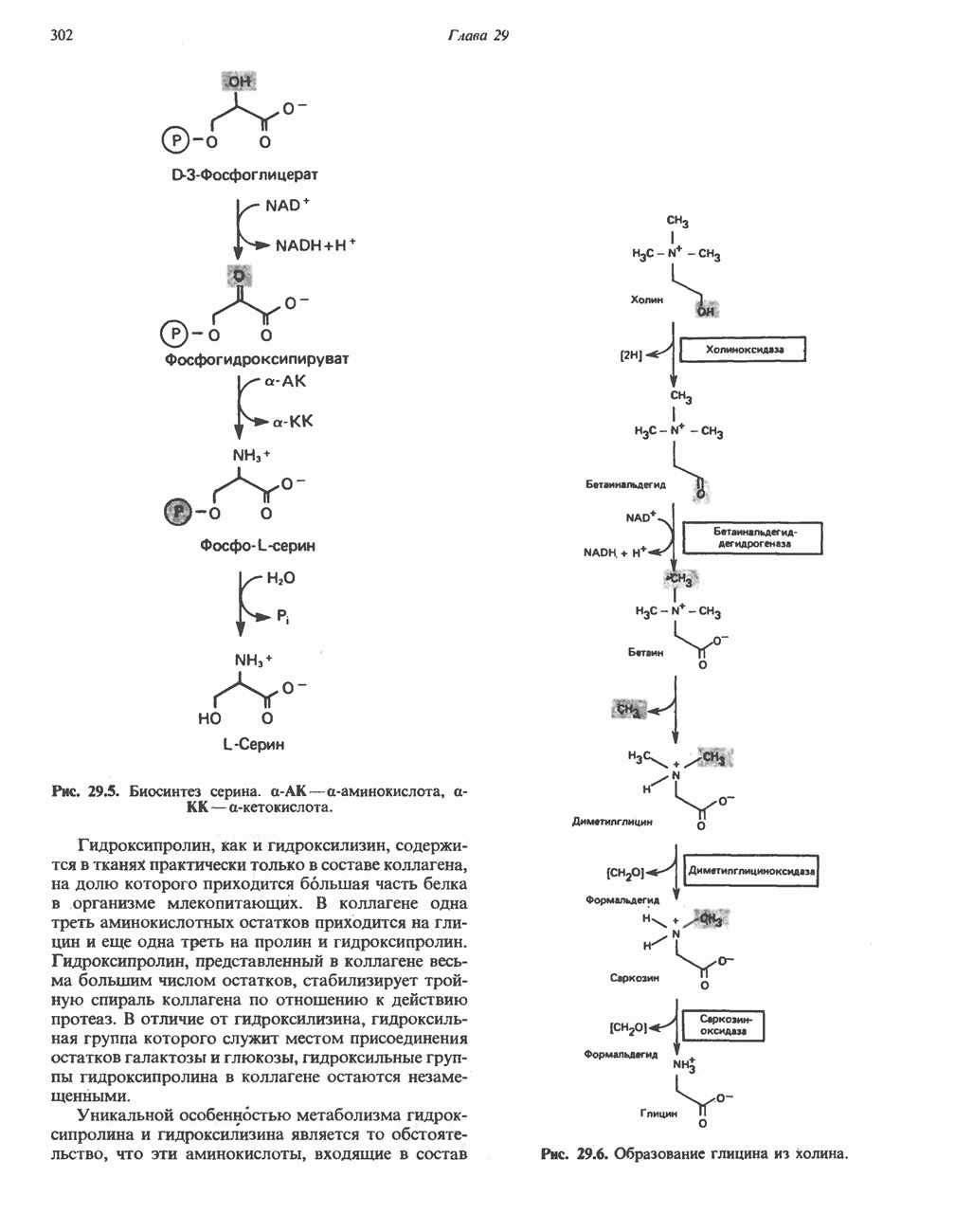
 -Жирные кислоты либо жирные кислоты, образующие
-Жирные кислоты либо жирные кислоты, образующие  вступают на данный путь на стадии указанной на схеме.
вступают на данный путь на стадии указанной на схеме.
 кислоты, поскольку из-за отсутствия пероксисом у них не происходит процесс окисления длинноцепочечных жирных кислот.
кислоты, поскольку из-за отсутствия пероксисом у них не происходит процесс окисления длинноцепочечных жирных кислот.
 либо
либо  затем
затем  изомеризуется под действием
изомеризуется под действием  с образованием
с образованием  Последний вступает далее в обычную последовательность реакций Р-окисления. Любое
Последний вступает далее в обычную последовательность реакций Р-окисления. Любое  либо образовавшееся в ходе окисления из линолевой кислоты (рис. 23.4), либо вступившее в цикл на этой стадии, превращается под действием ацил-Соа-дегидрогеназы в
либо образовавшееся в ходе окисления из линолевой кислоты (рис. 23.4), либо вступившее в цикл на этой стадии, превращается под действием ацил-Соа-дегидрогеназы в  последний затем превращается в
последний затем превращается в  под действием
под действием  -зависимого фермента
-зависимого фермента  Далее
Далее  атакует А
атакует А  связь; в результате образуется
связь; в результате образуется  являющийся промежуточным соединением при
являющийся промежуточным соединением при  -окислении.
-окислении.