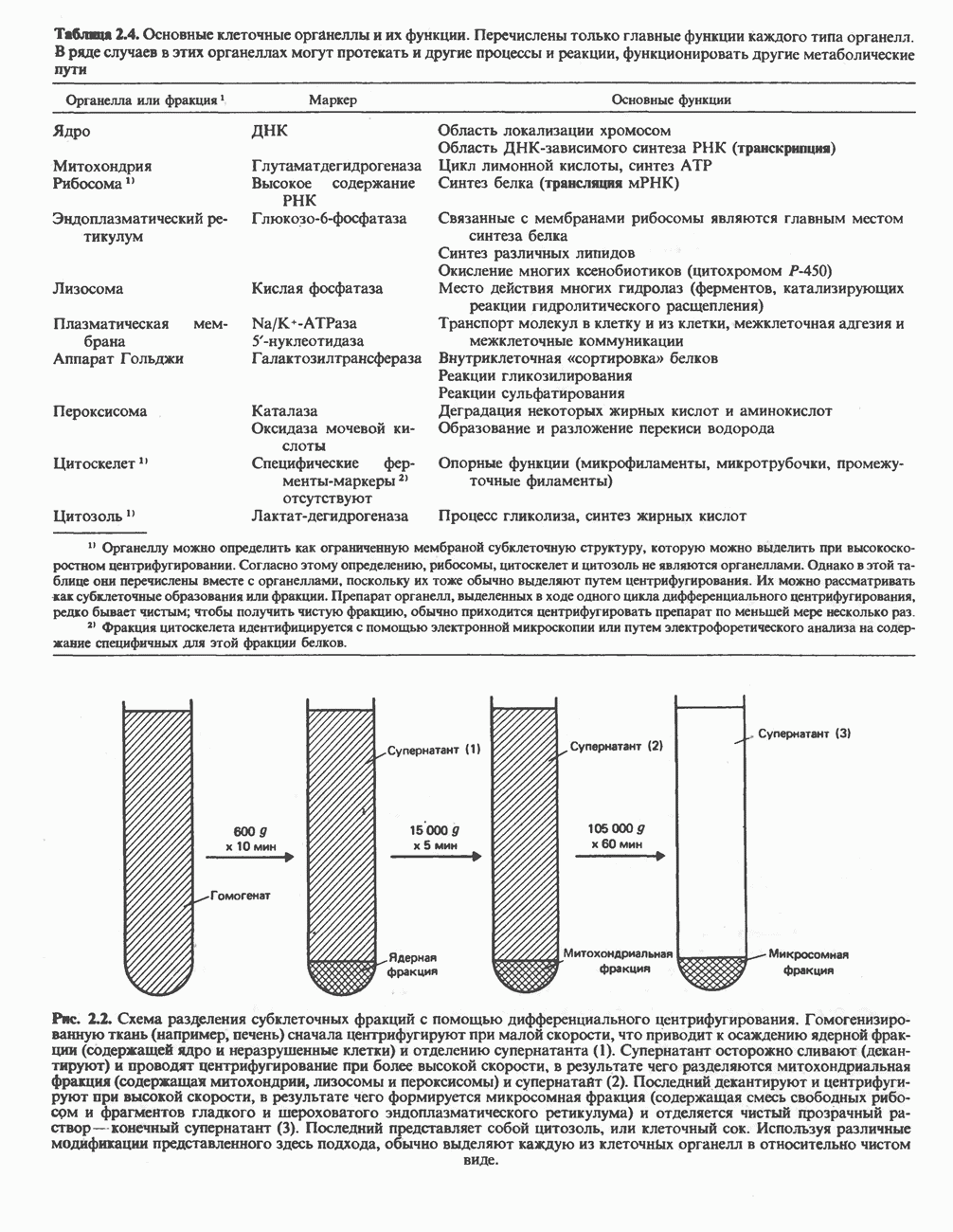
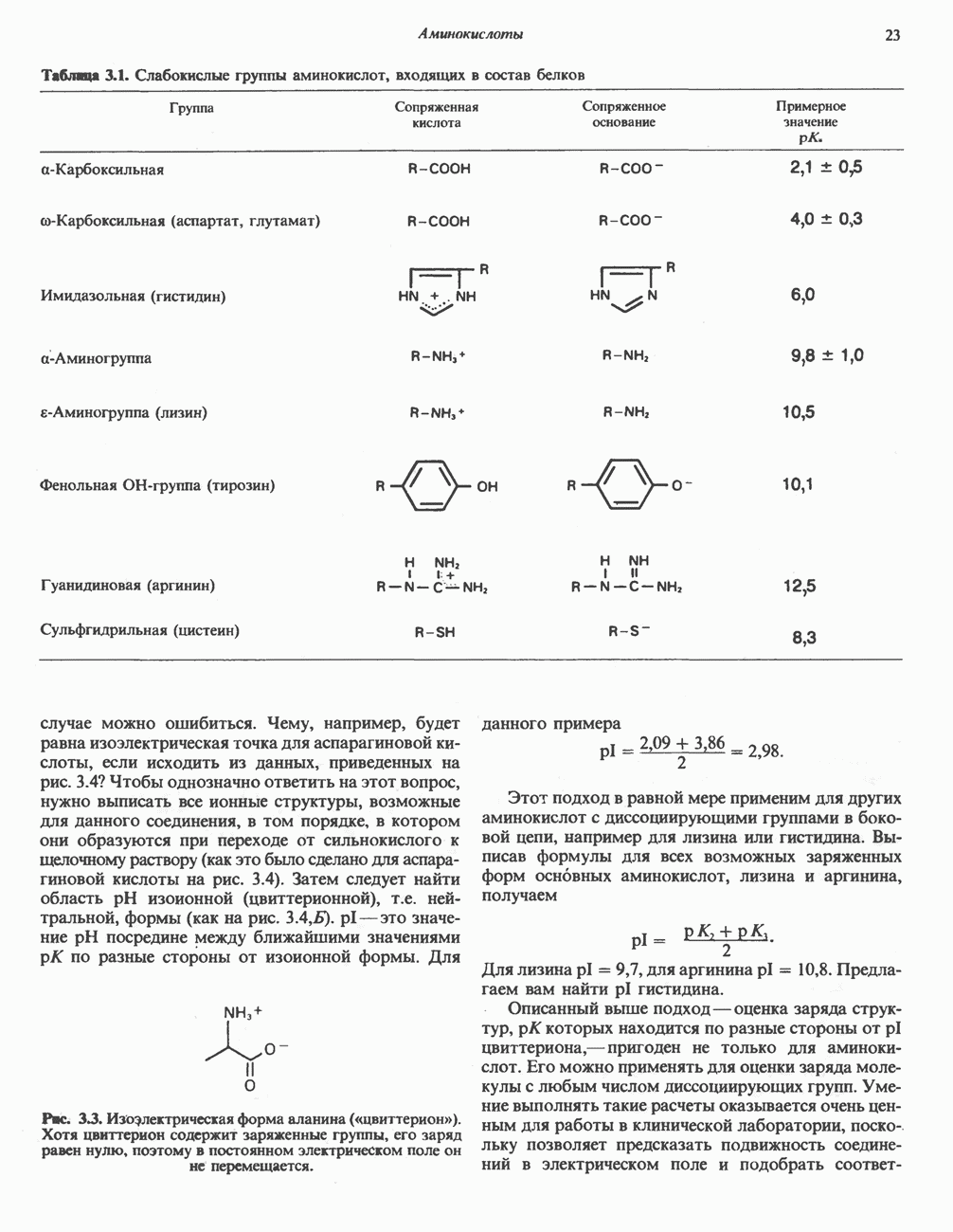
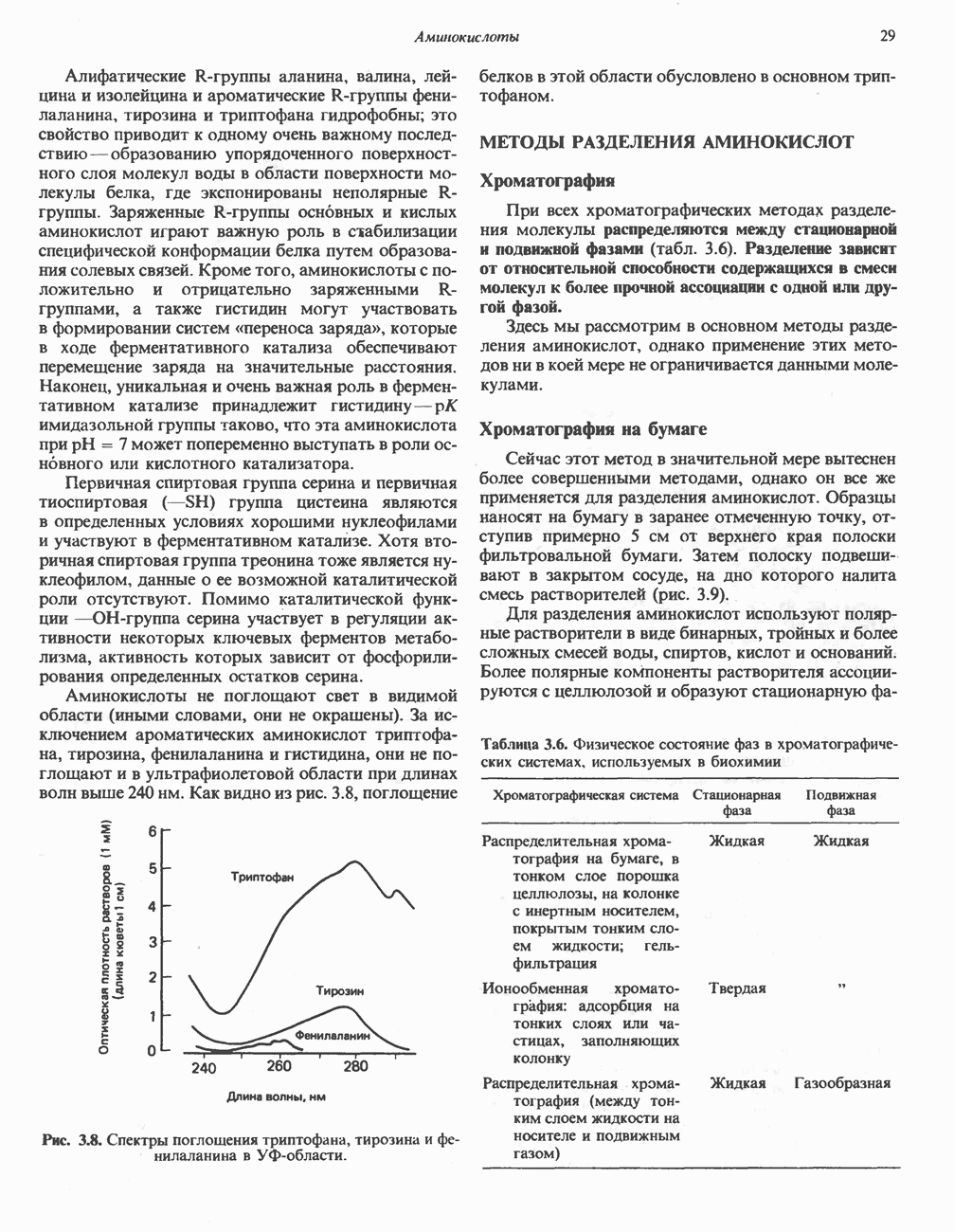
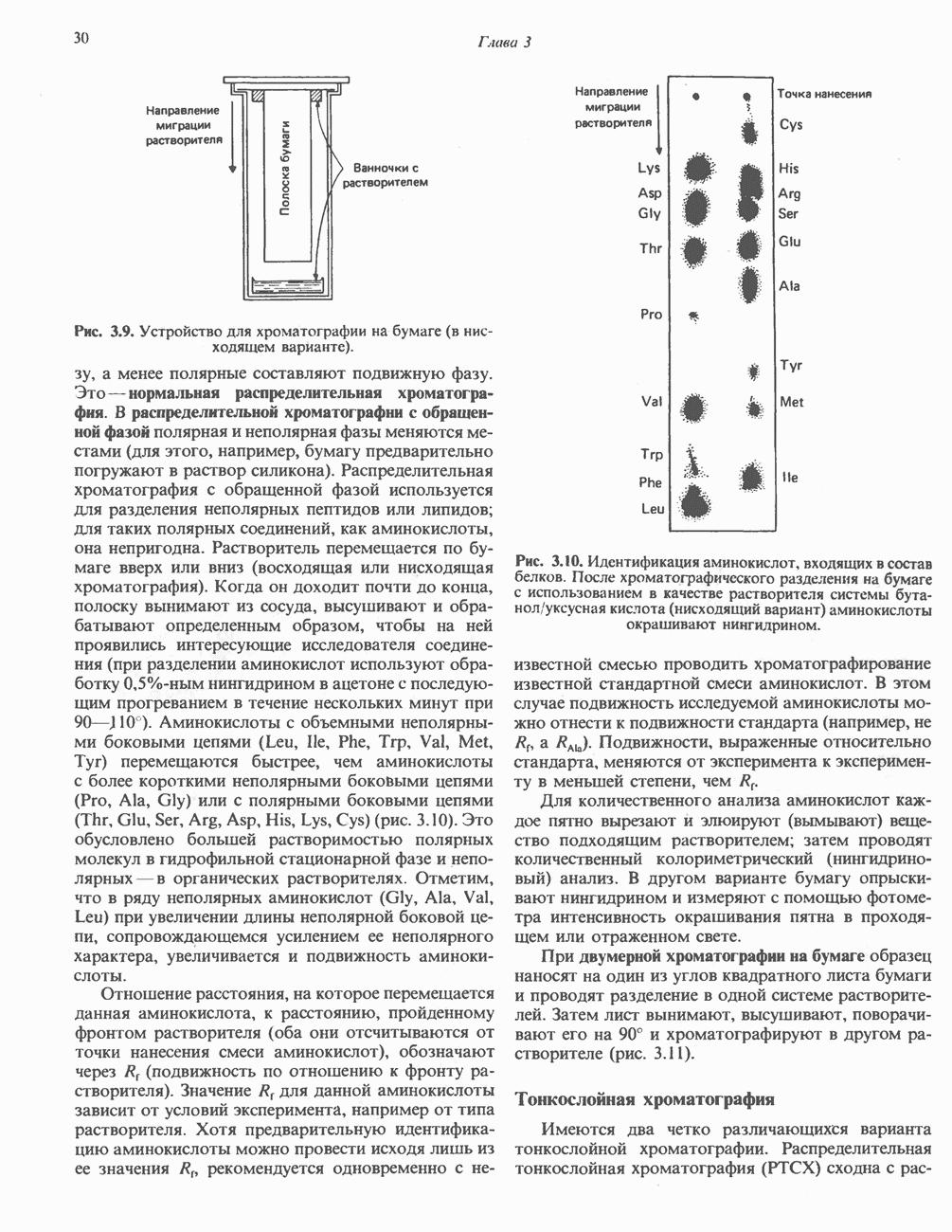
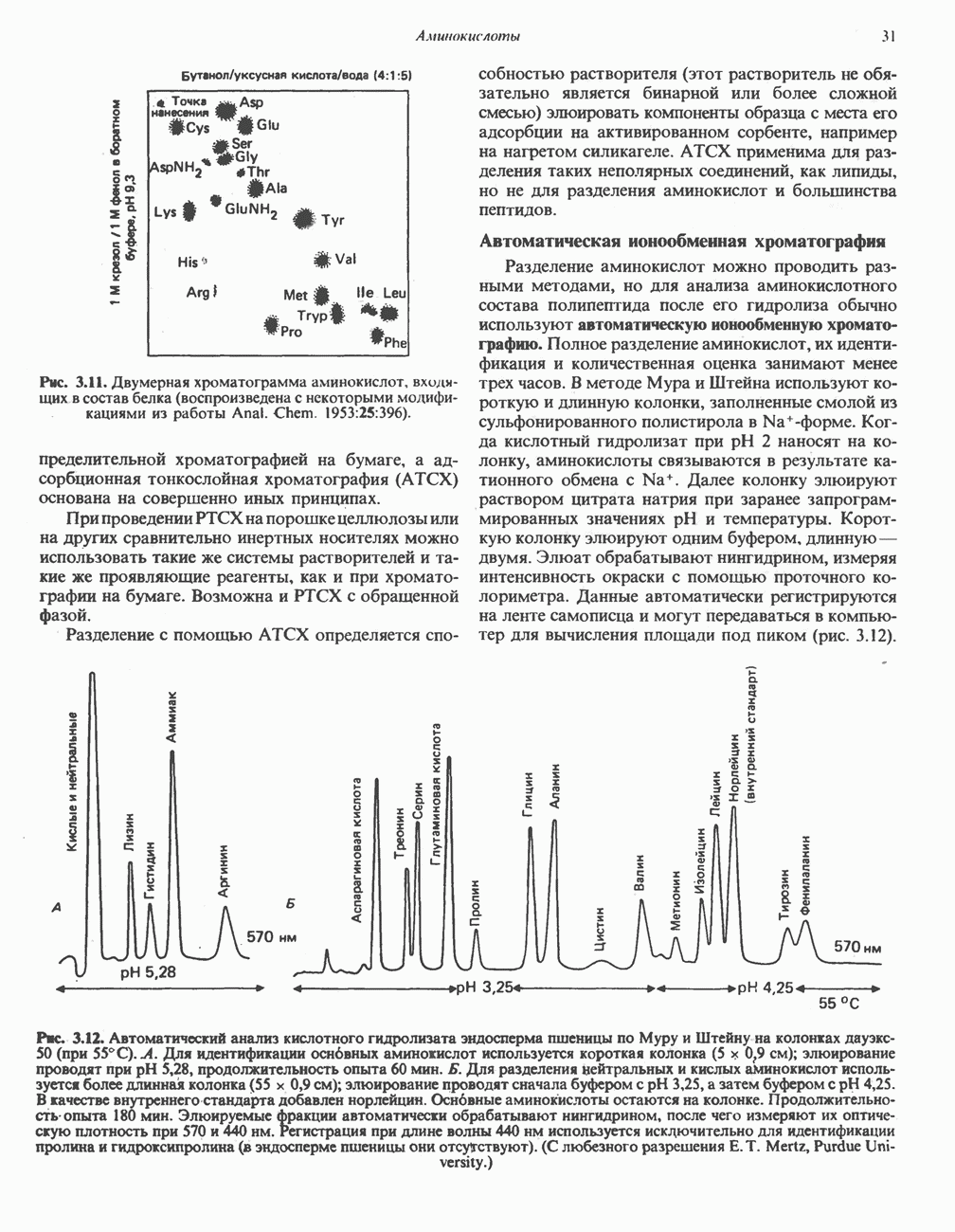
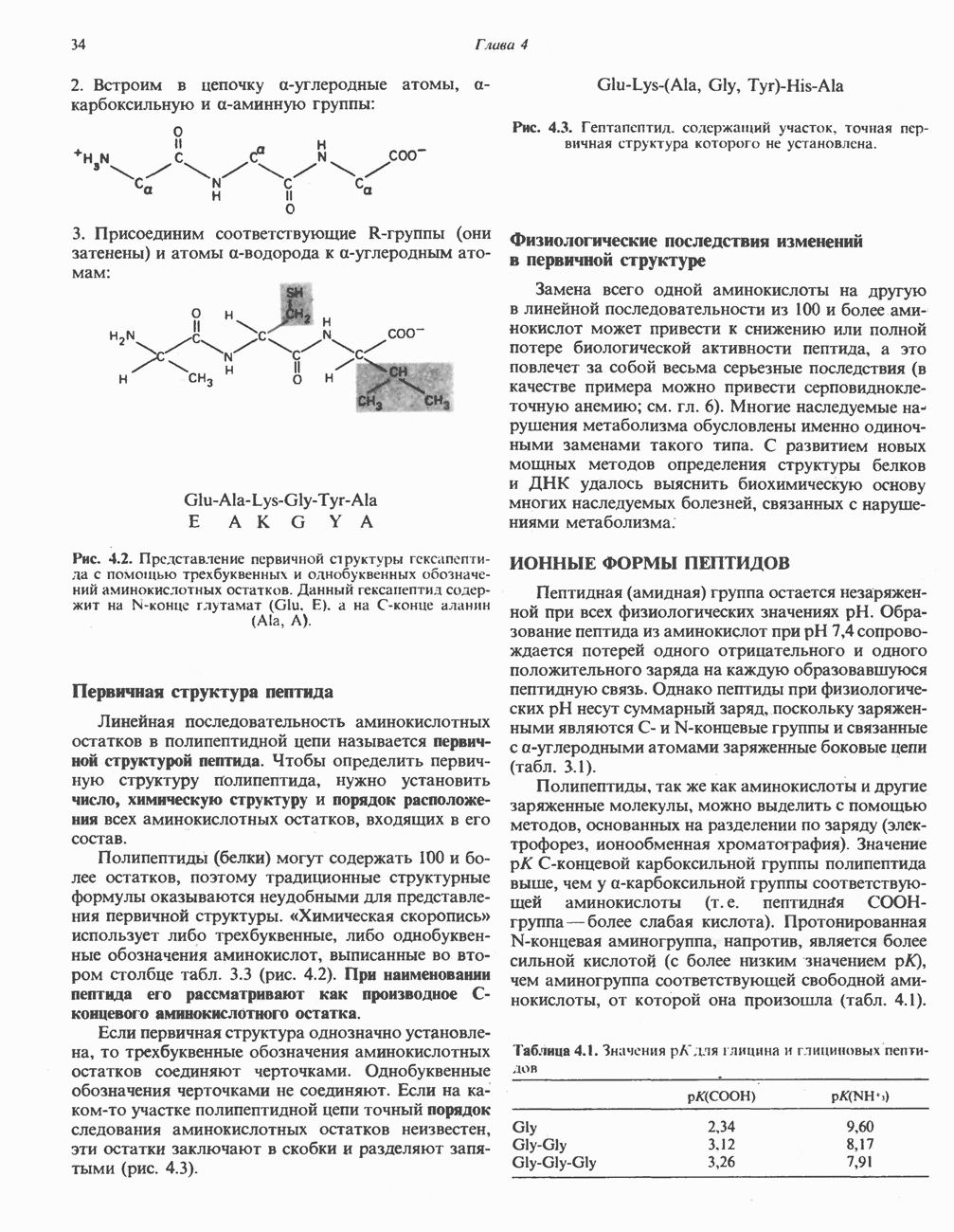
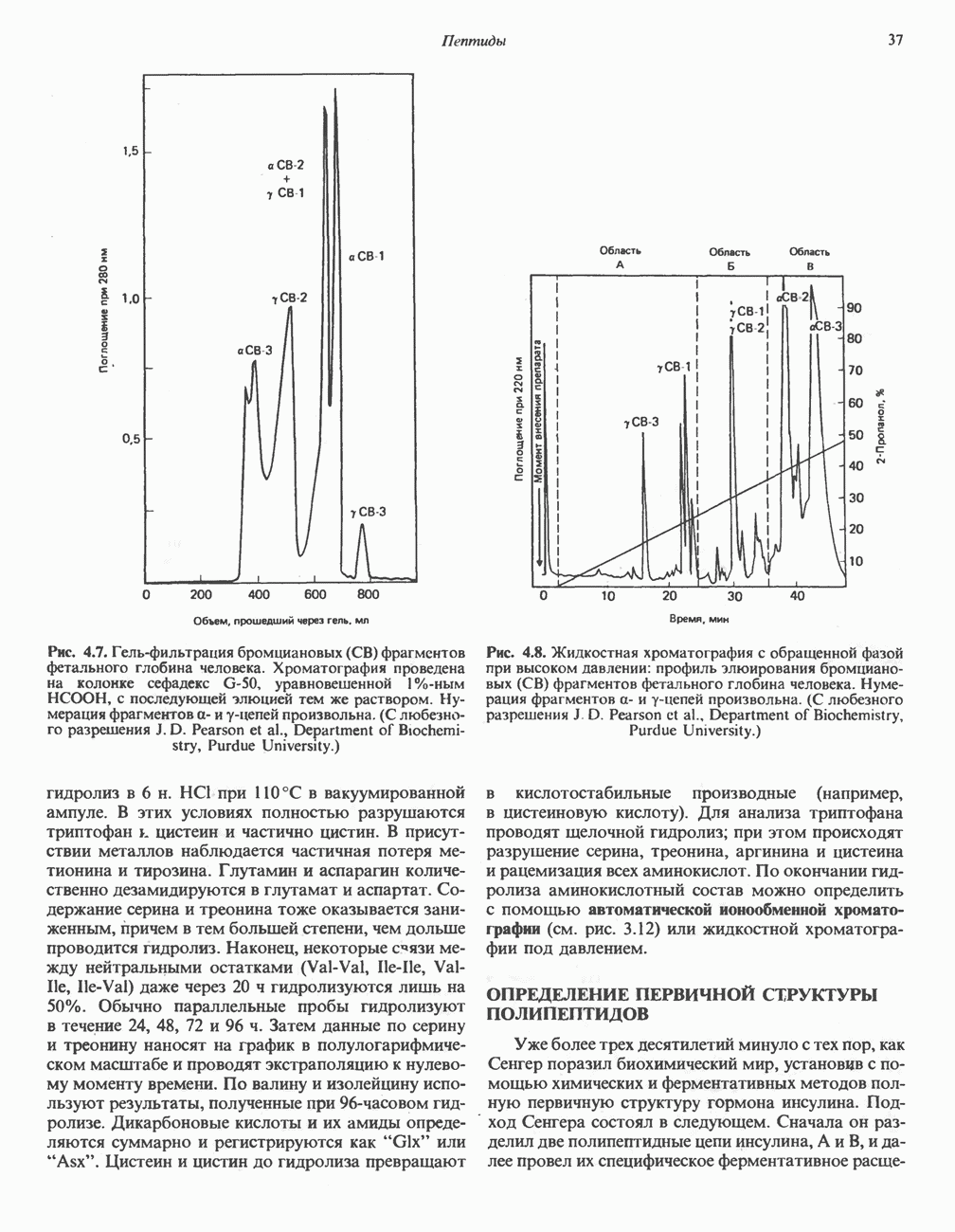
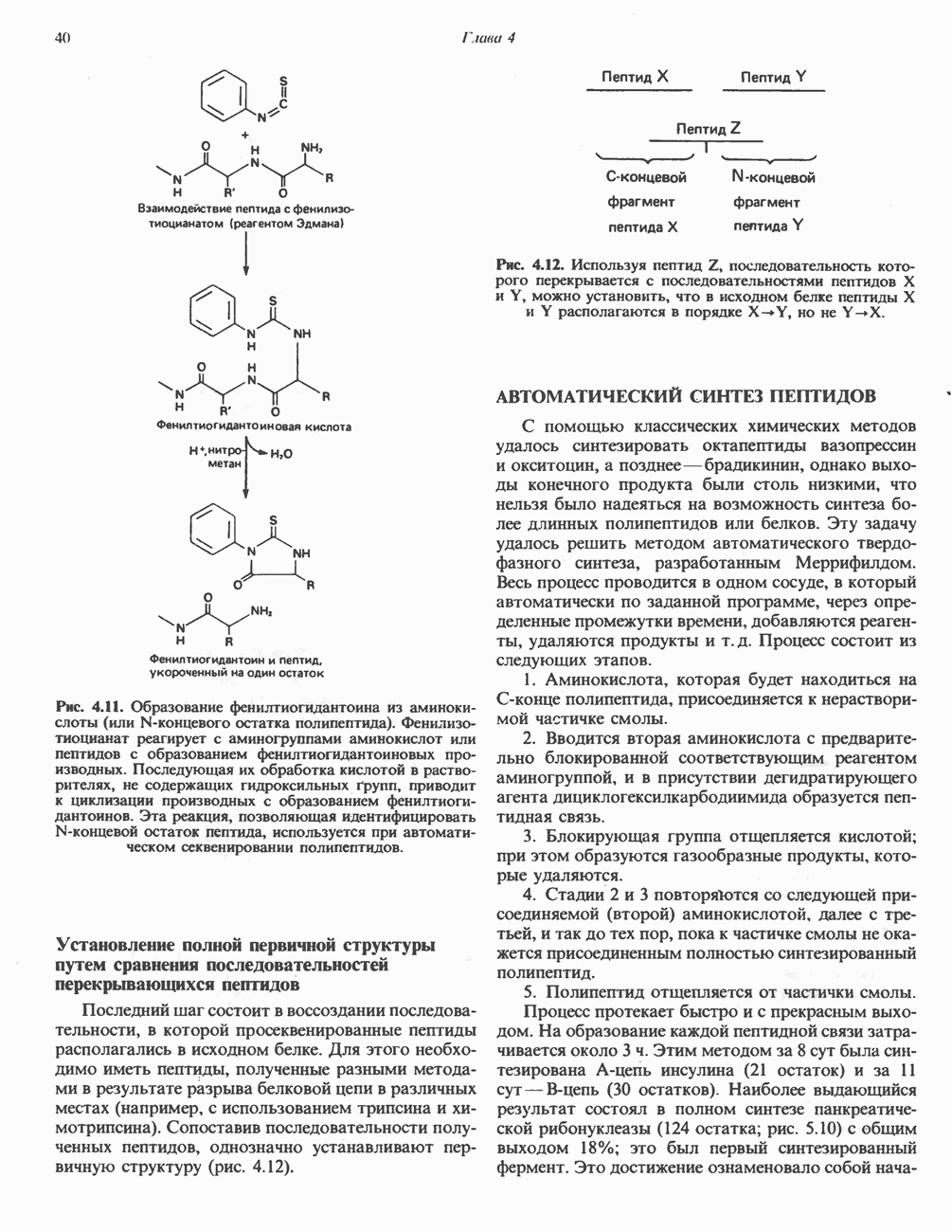
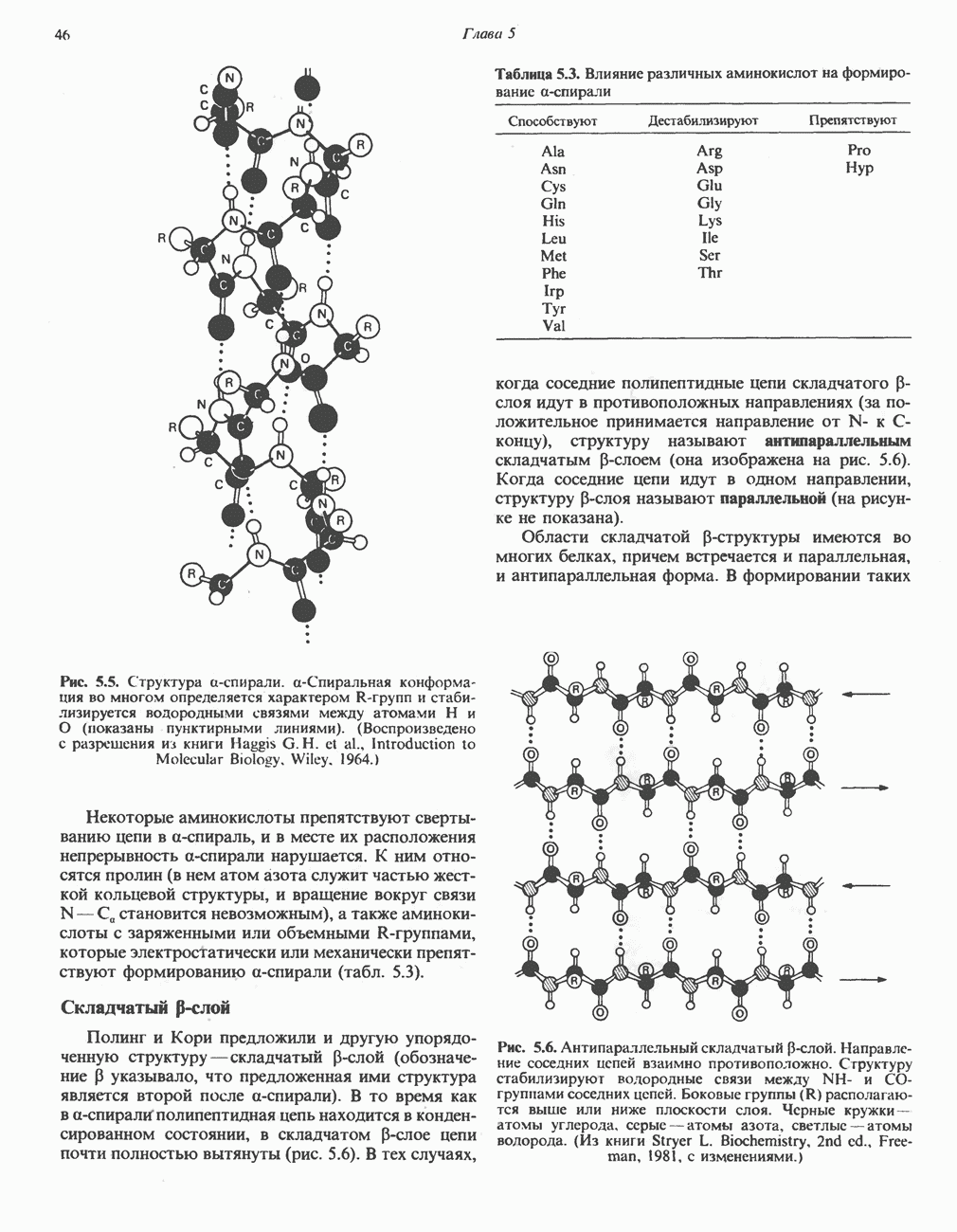
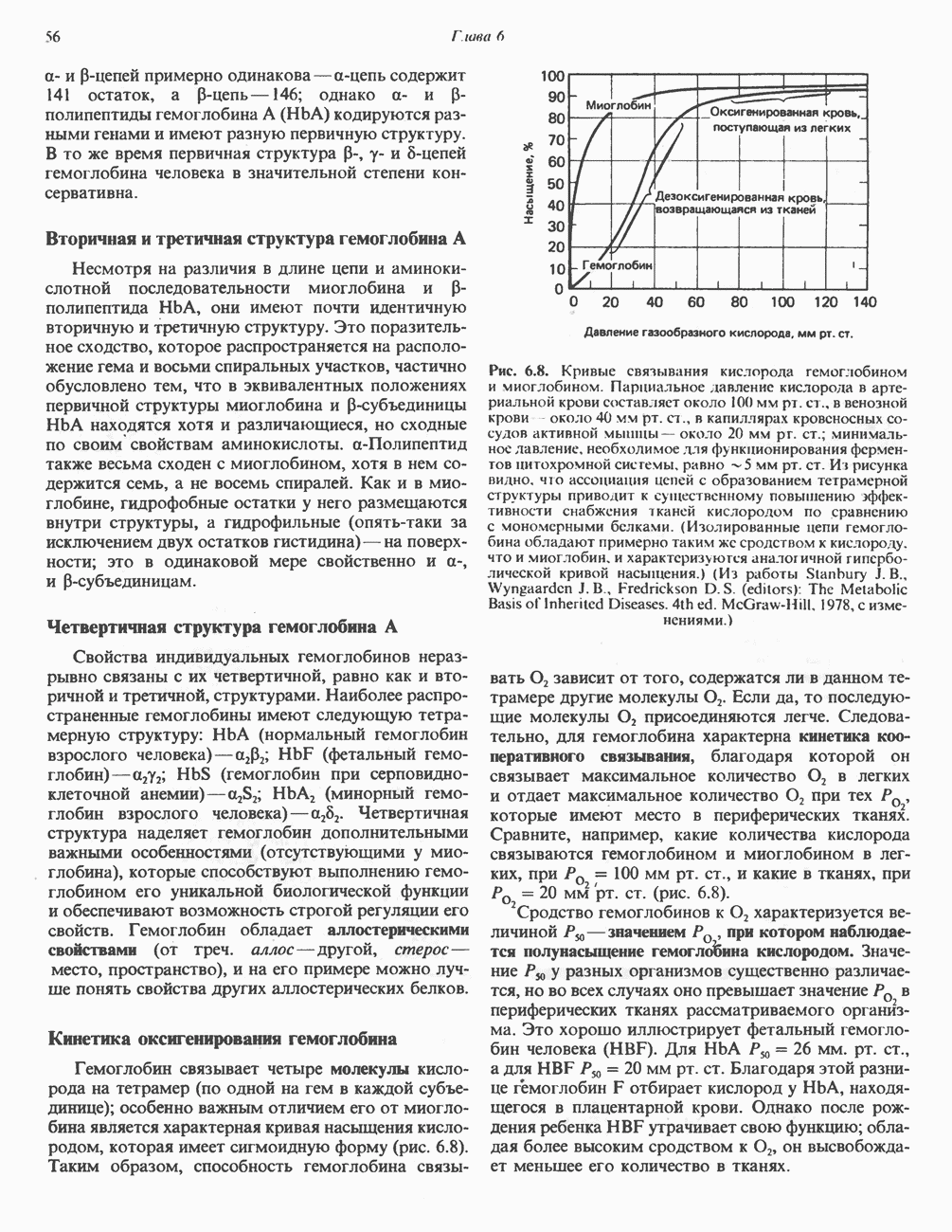
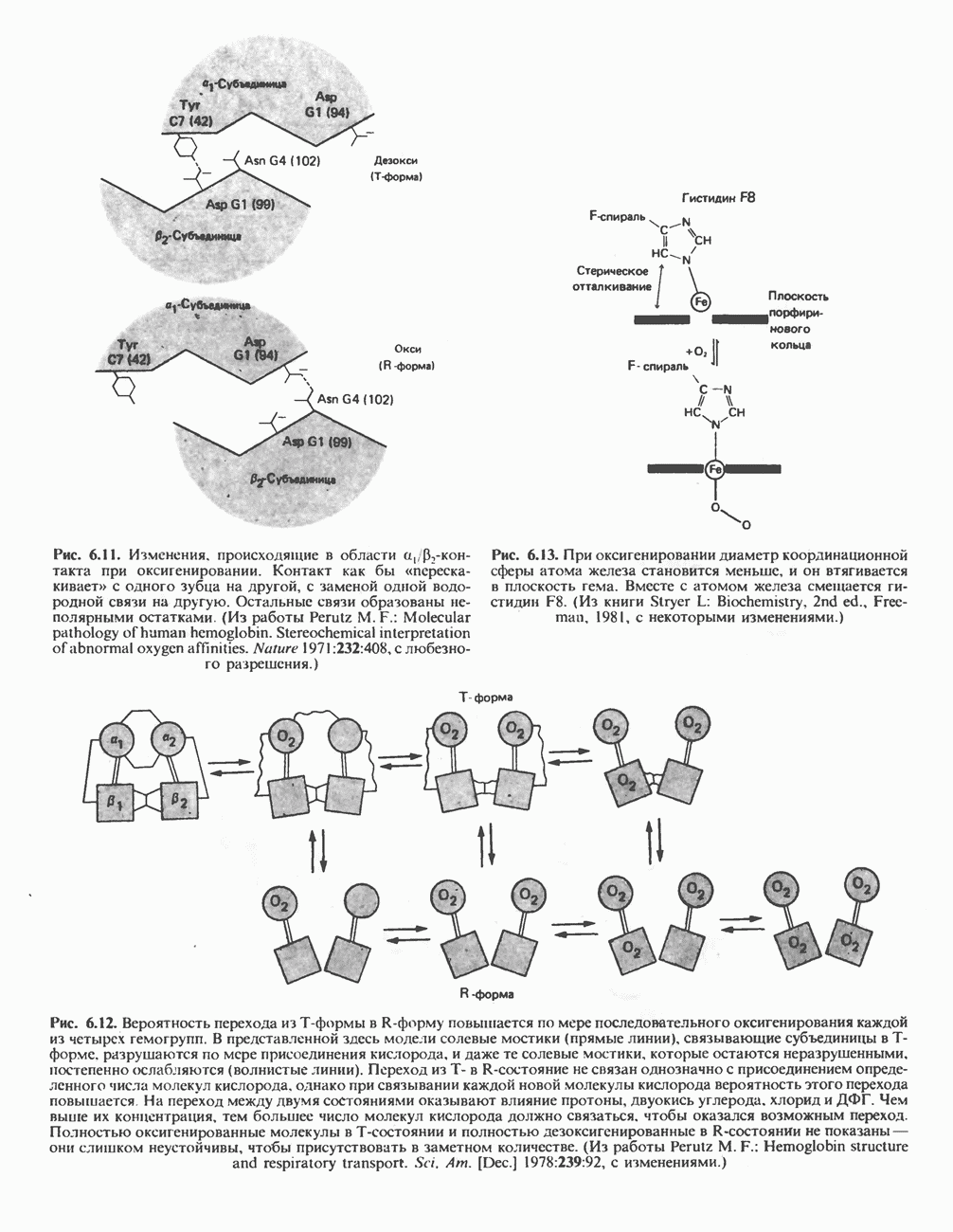
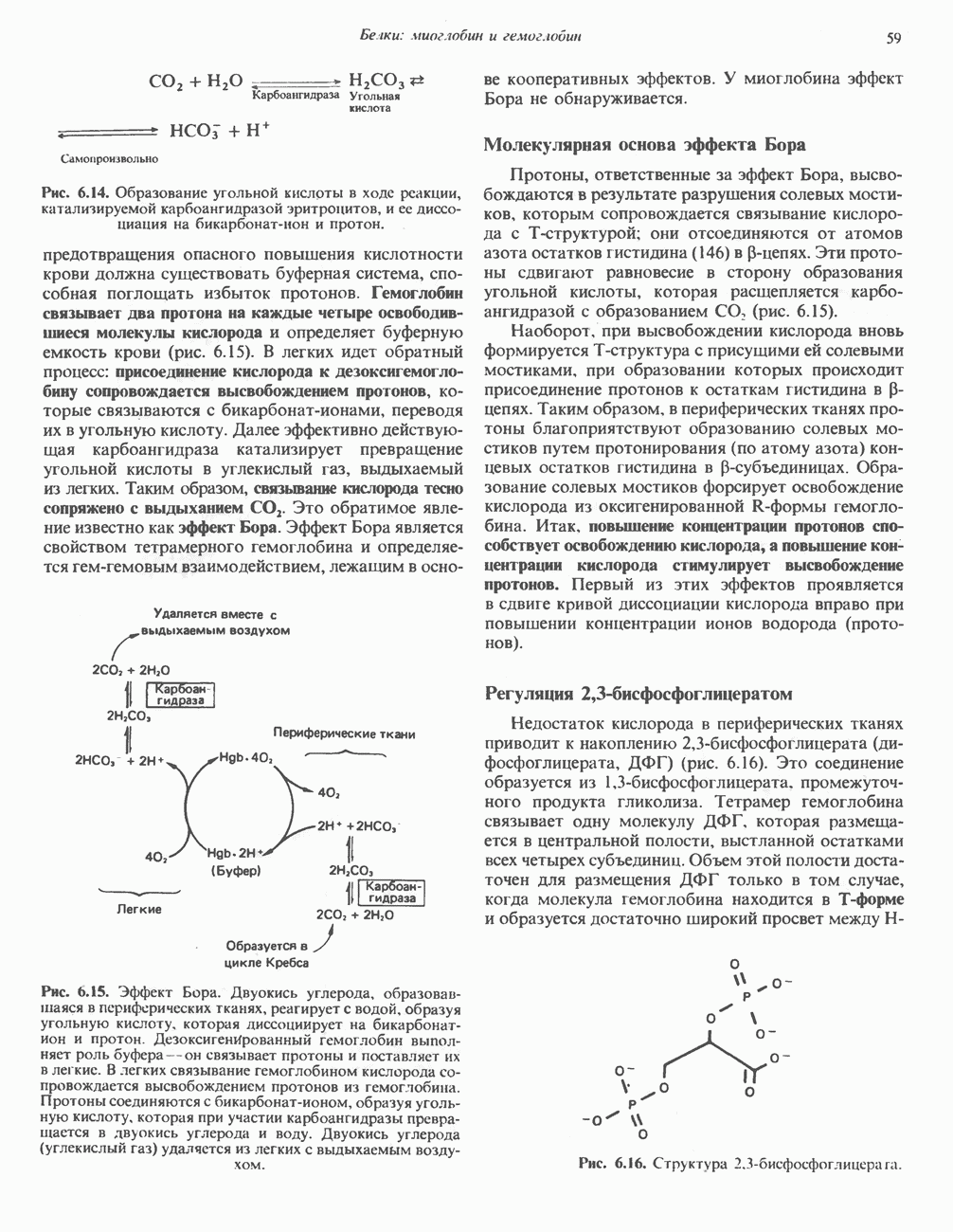
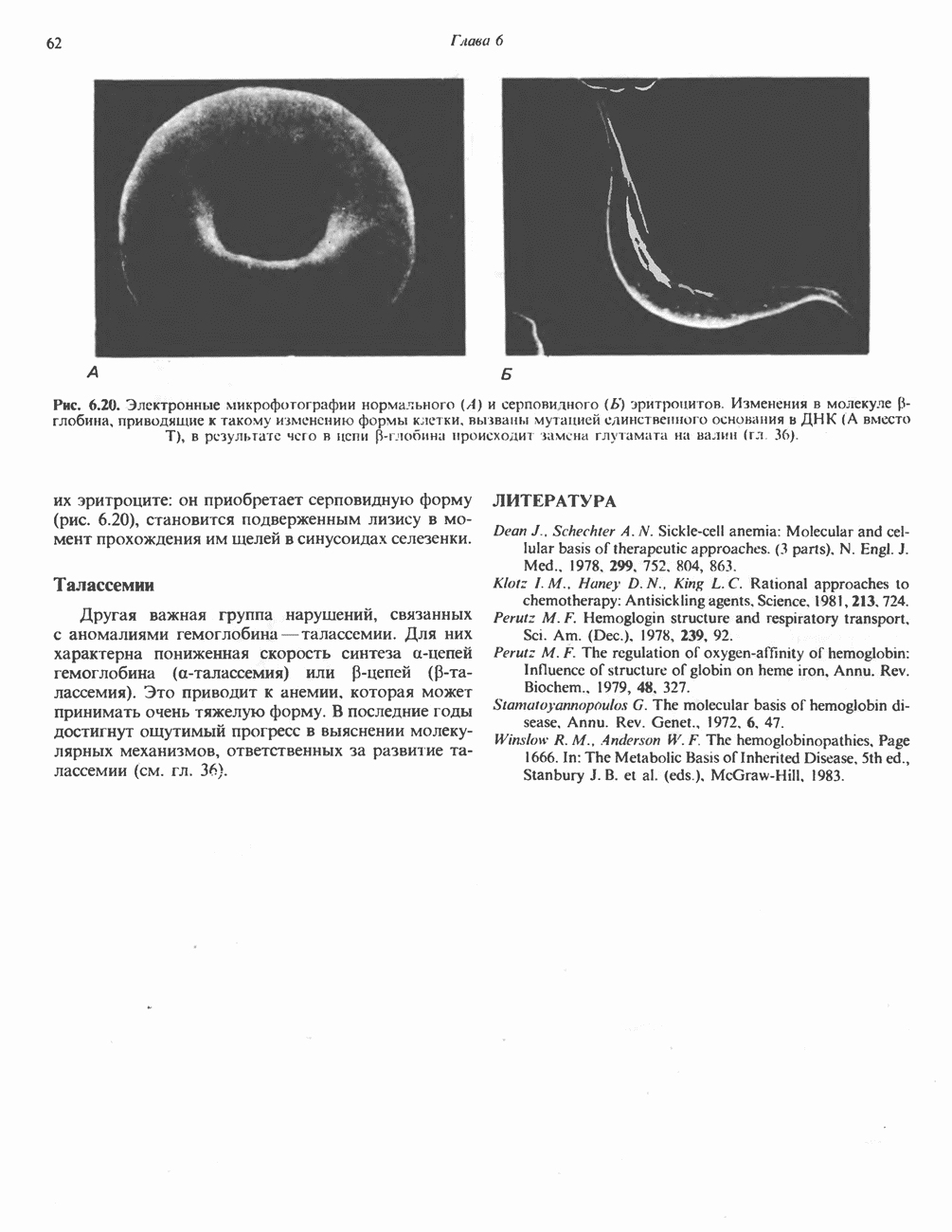
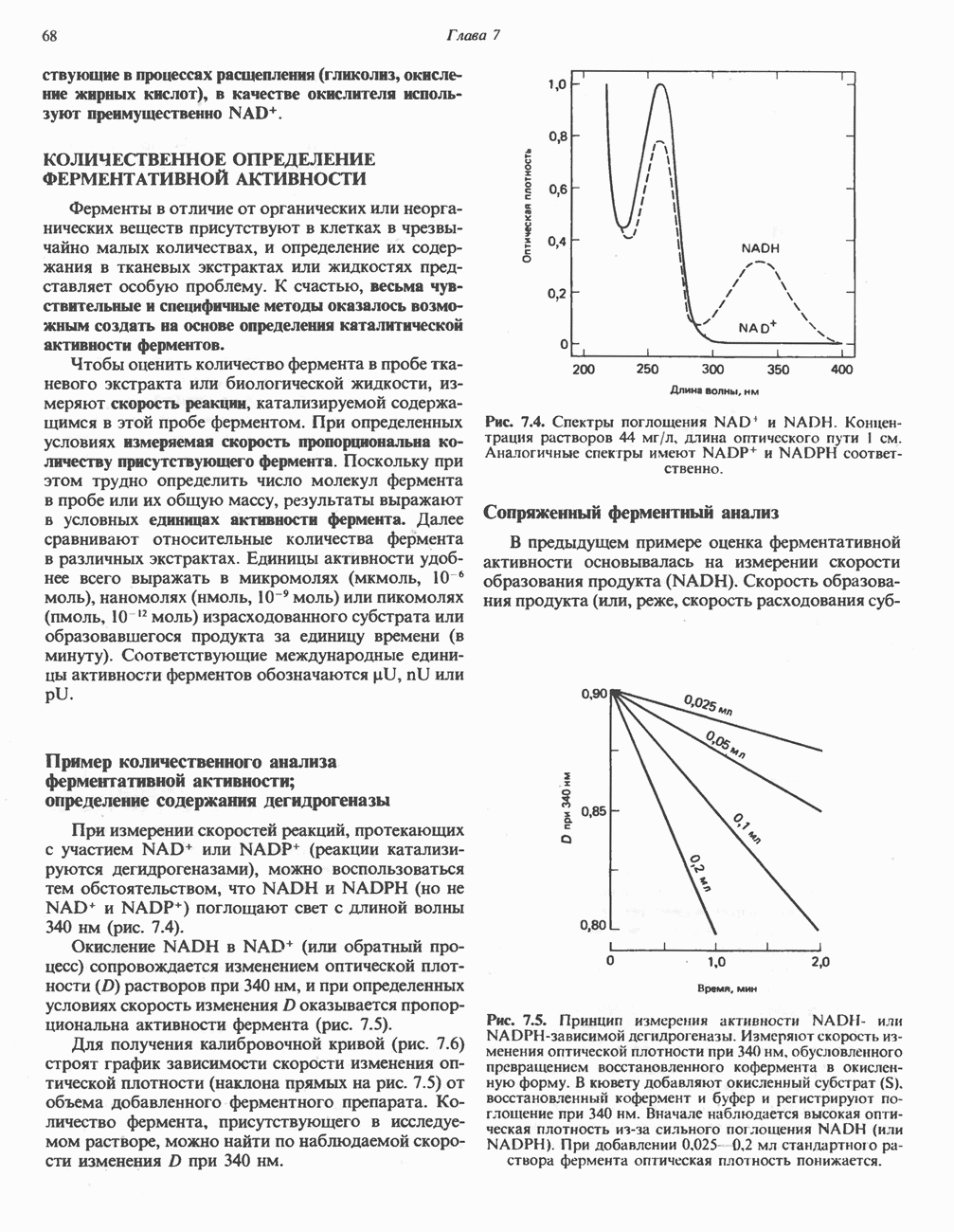
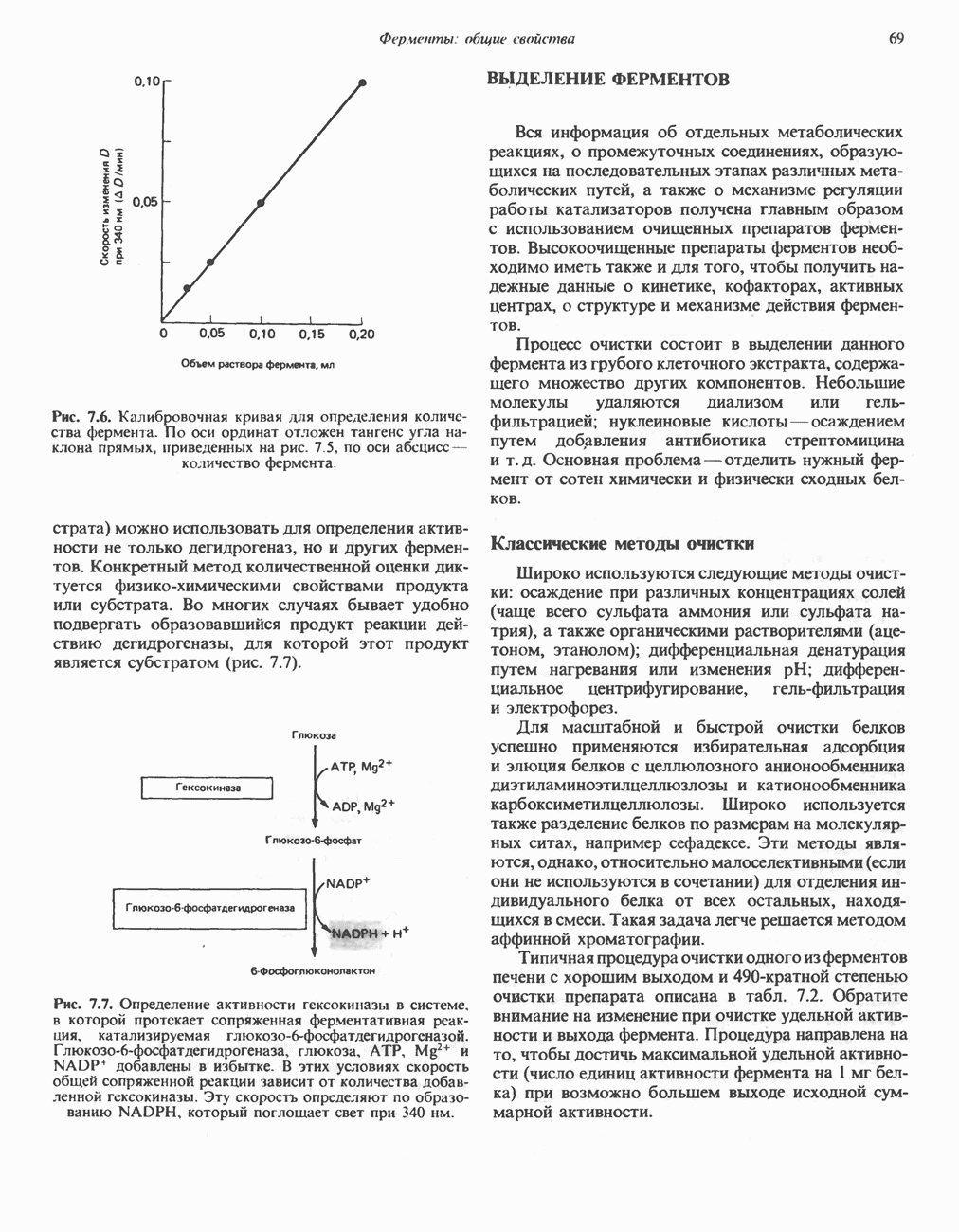
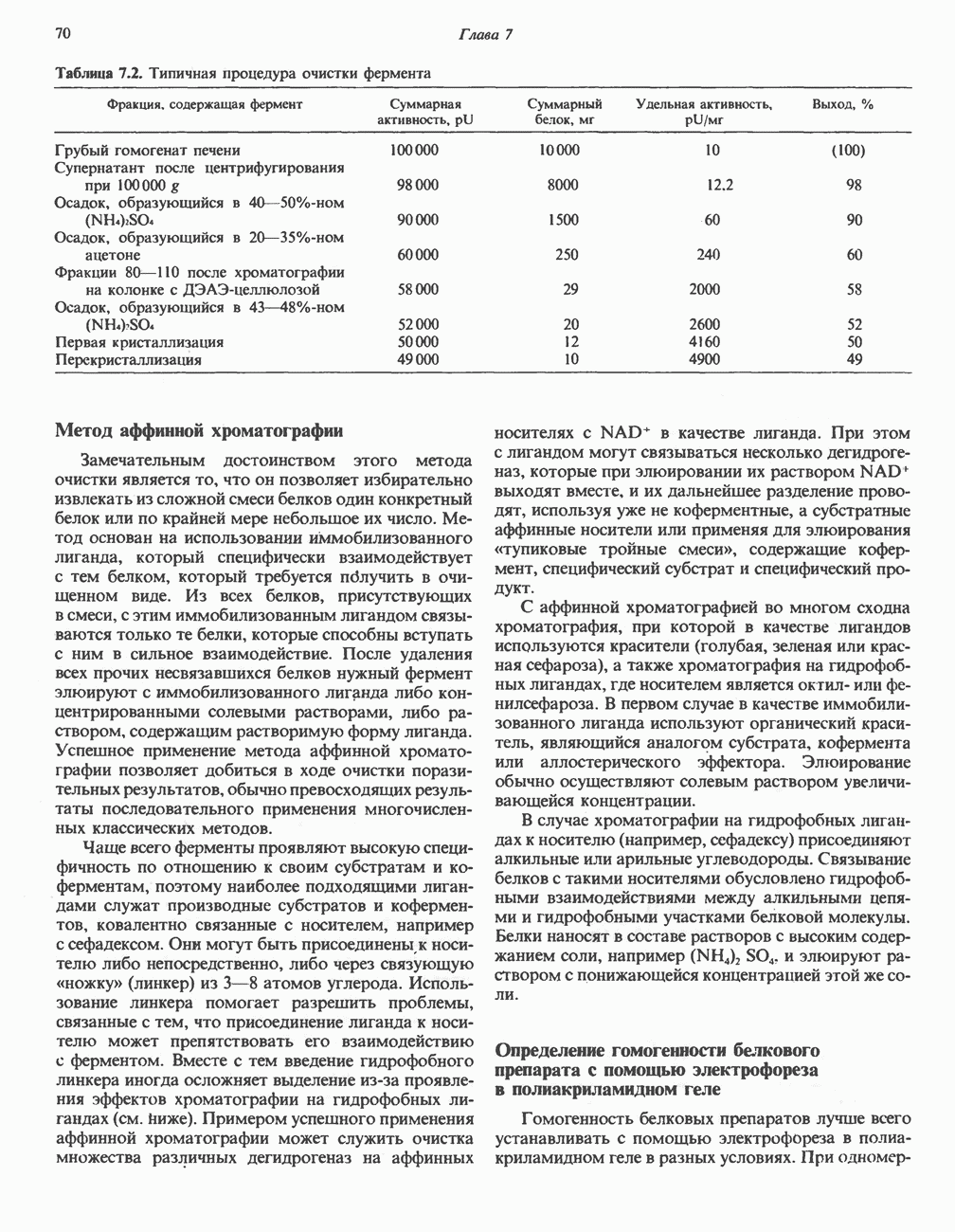
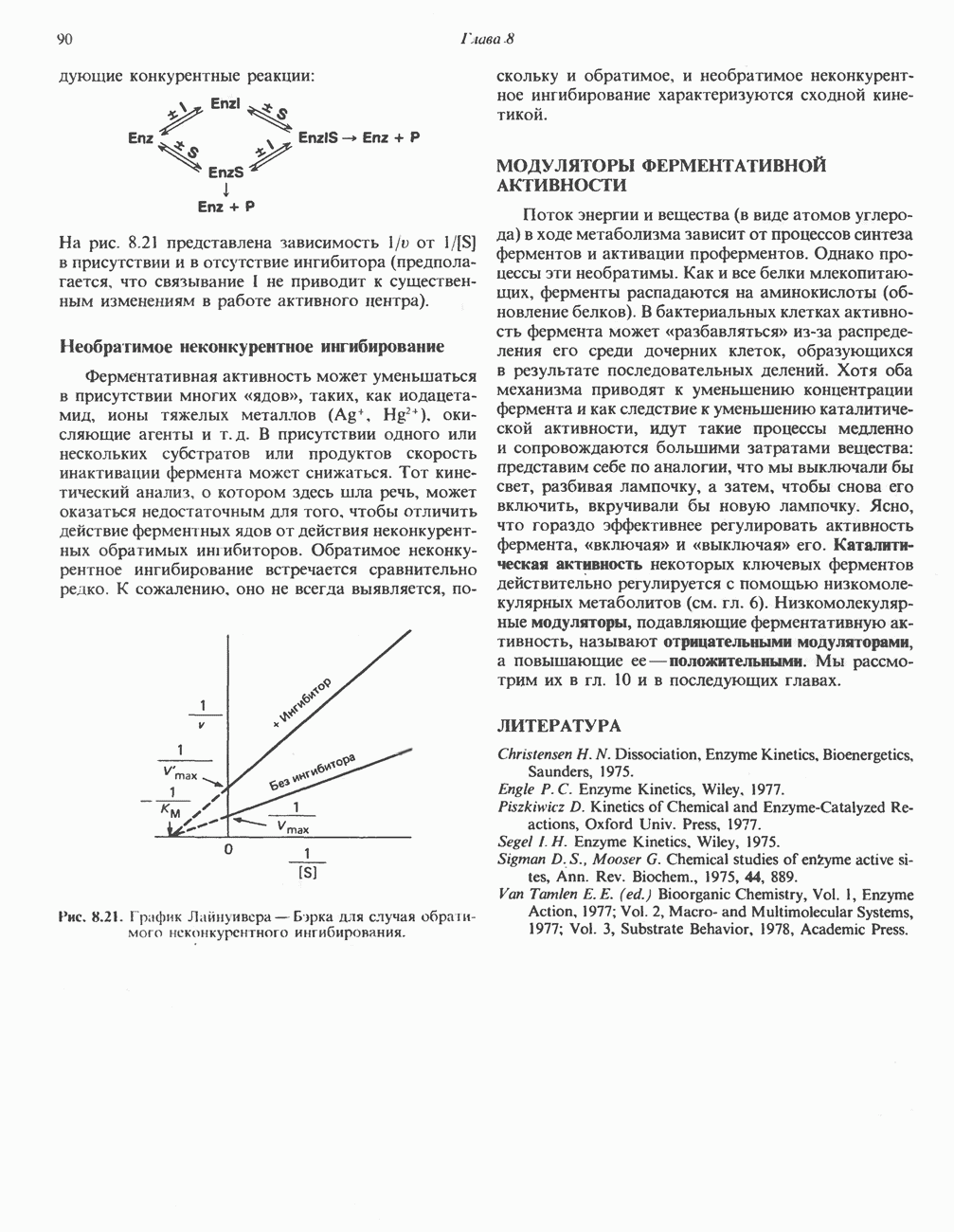
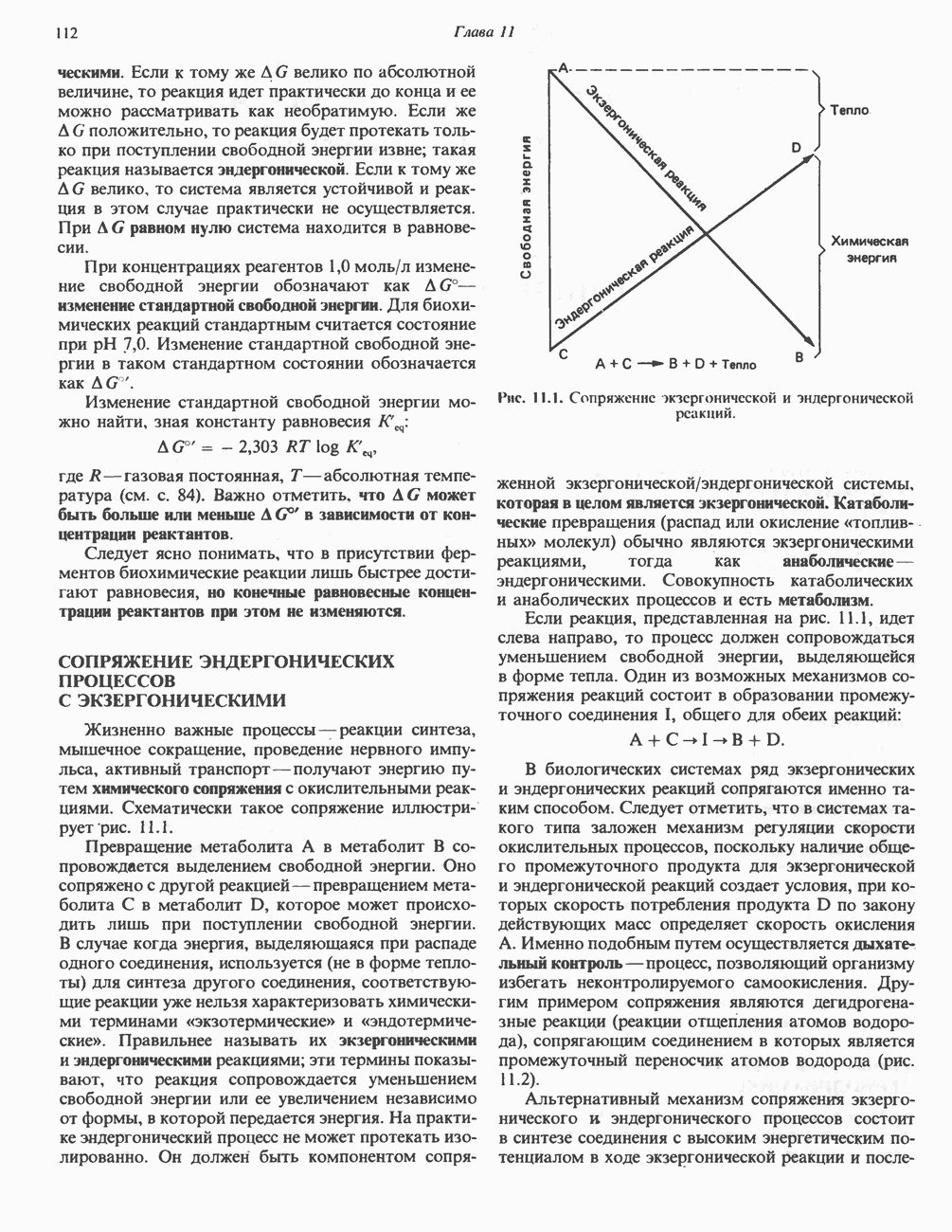
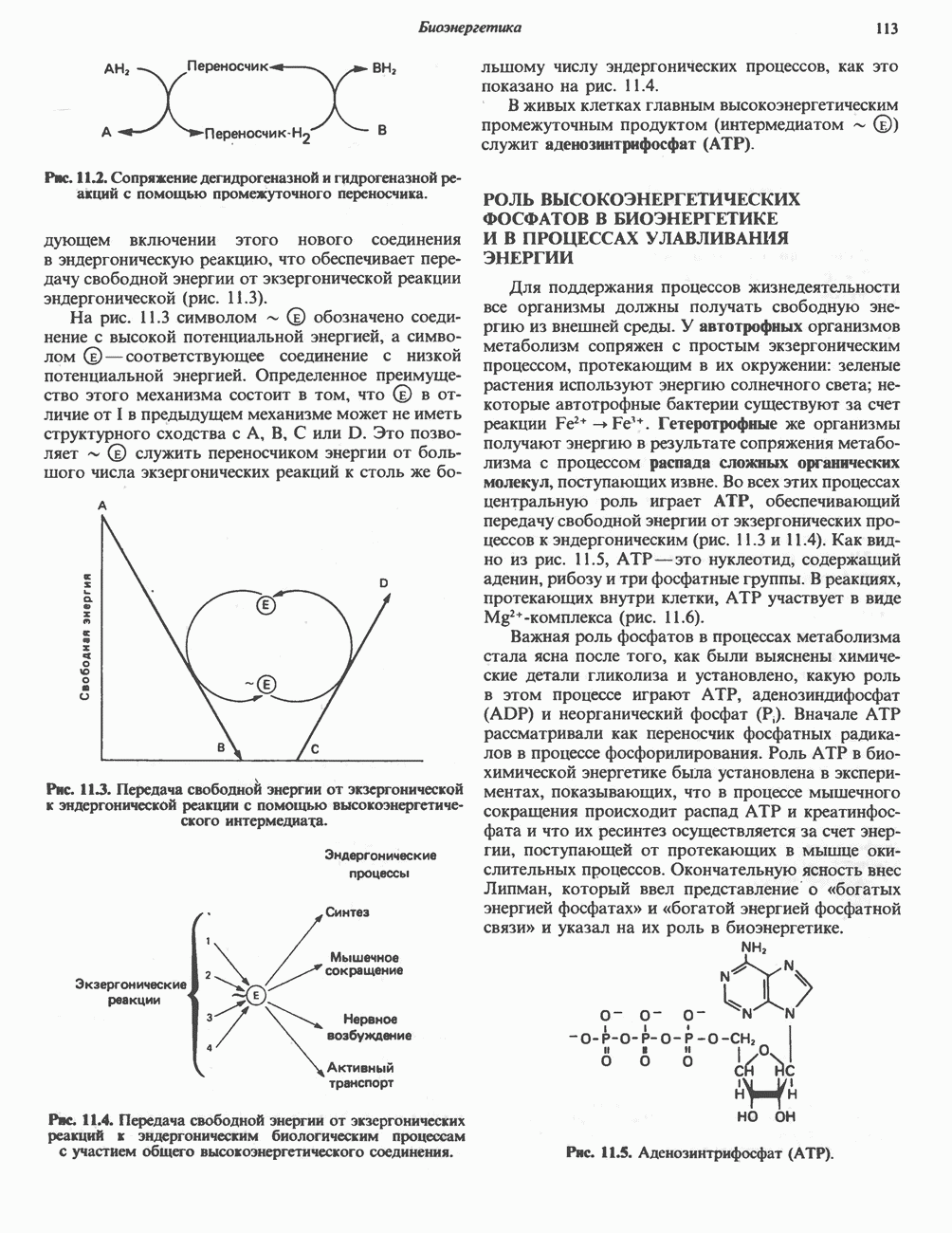
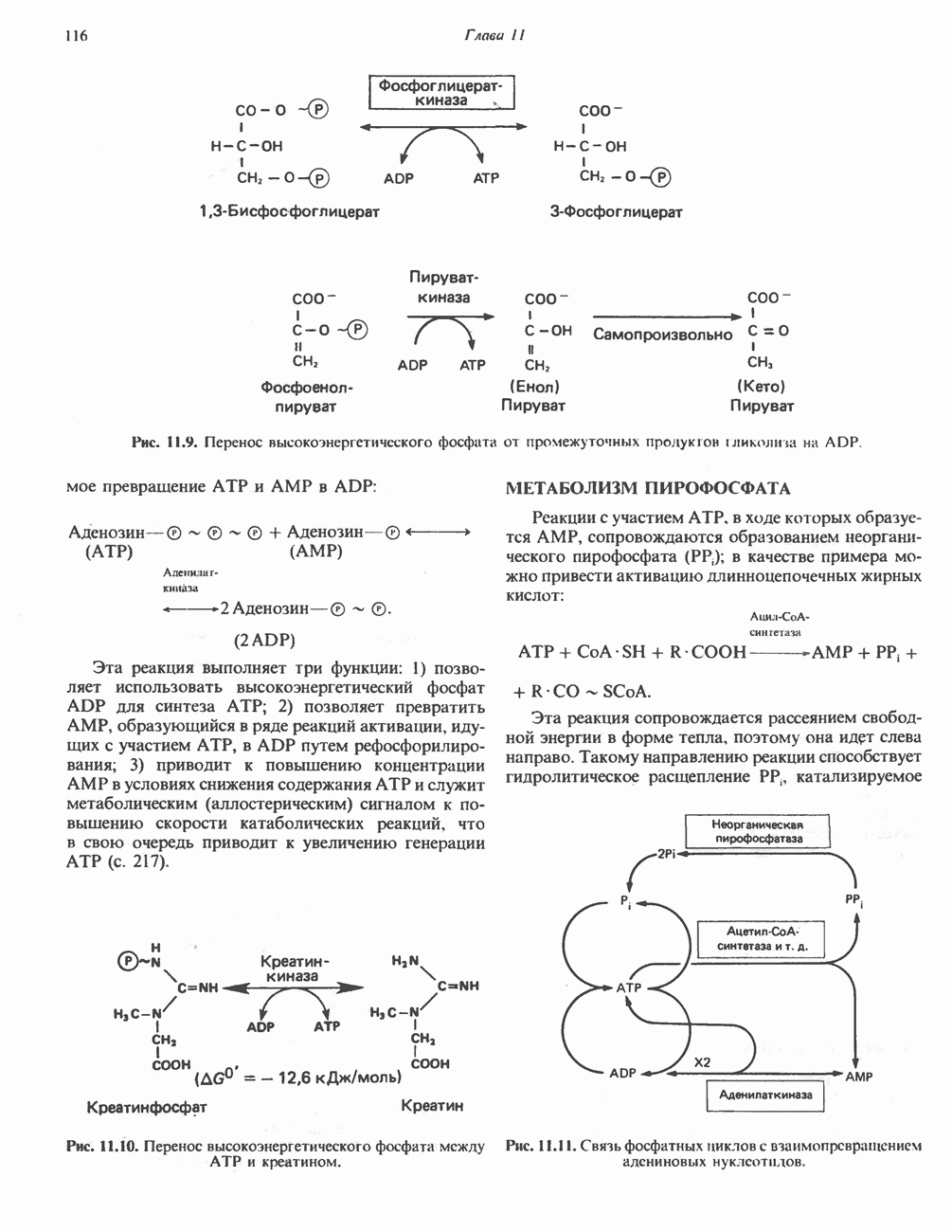
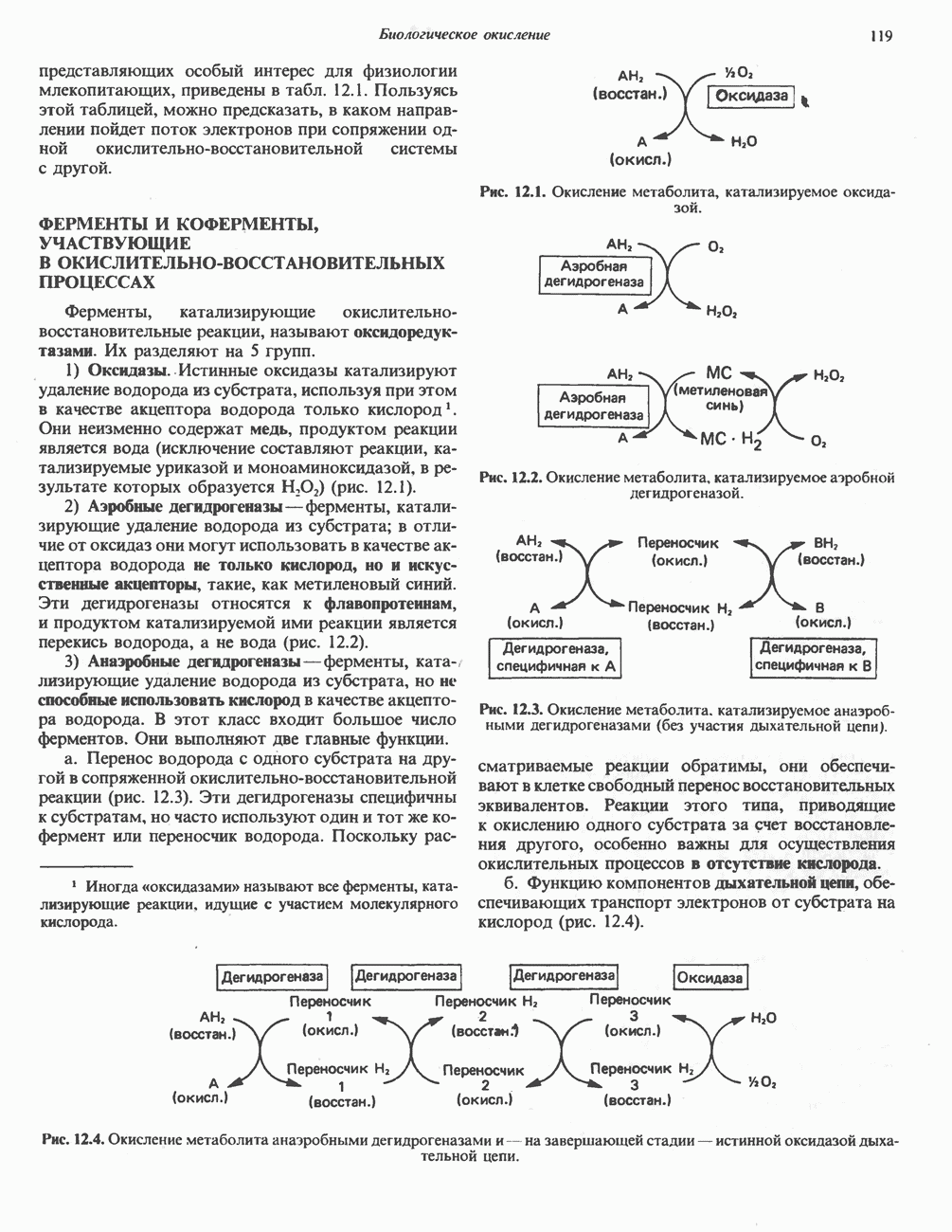
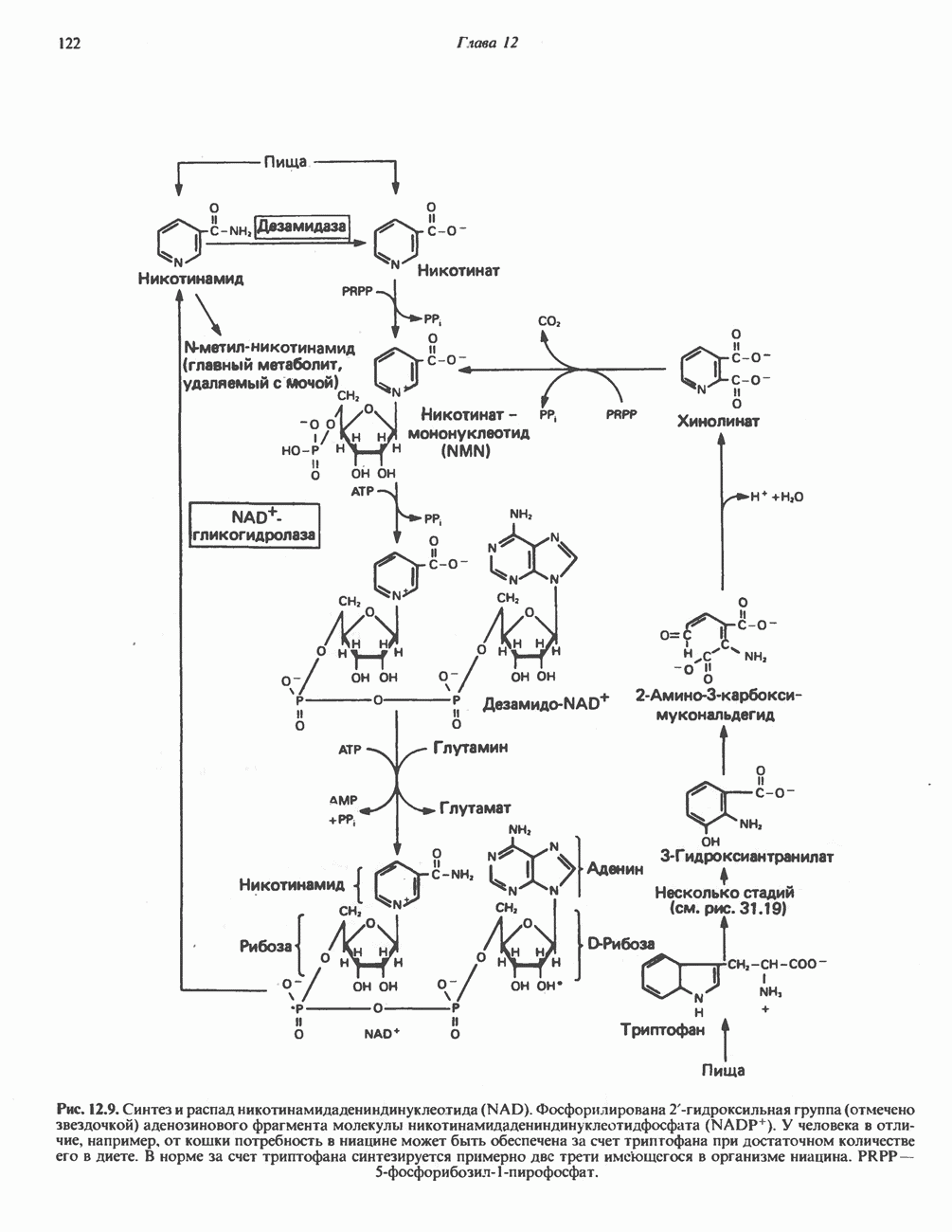
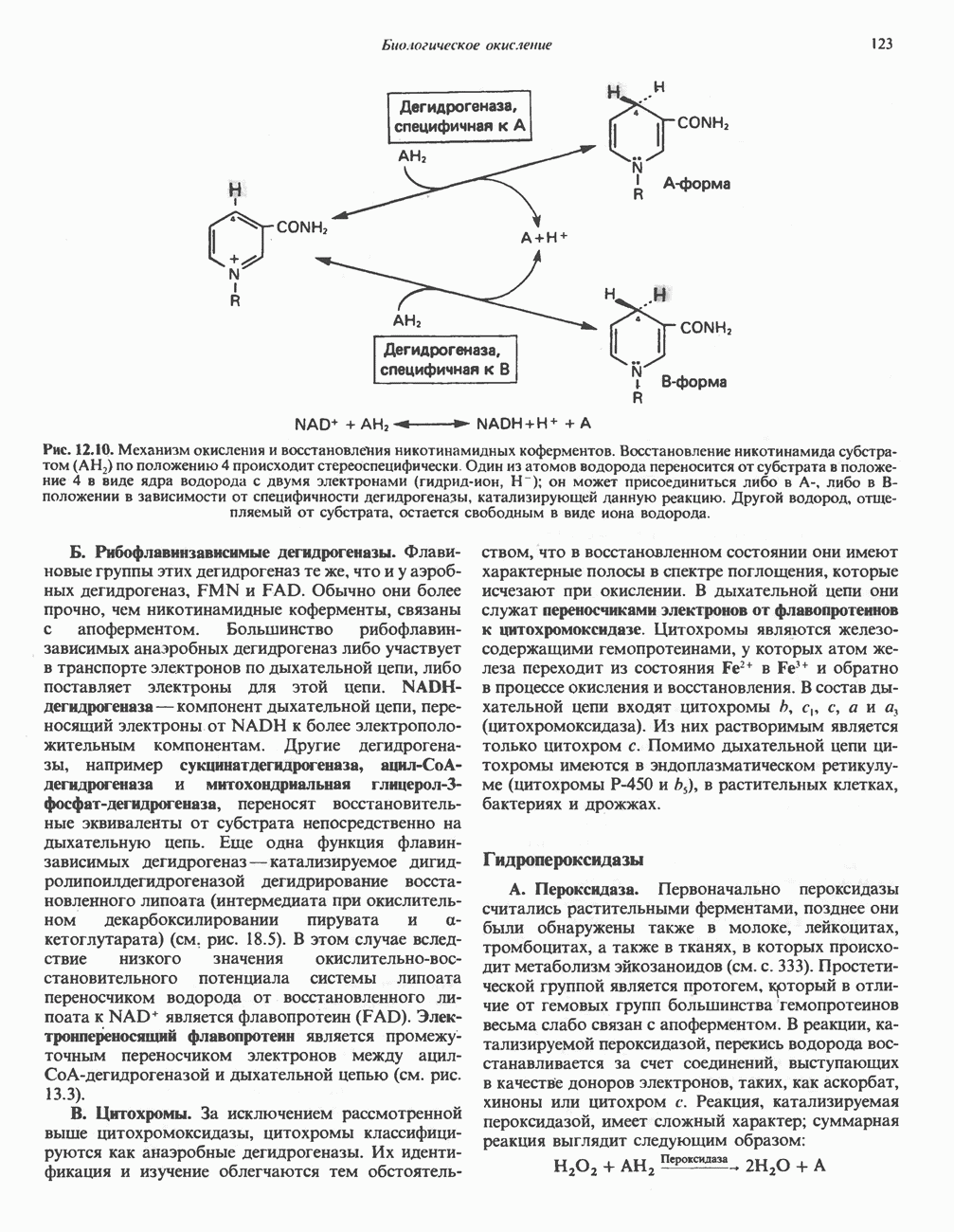
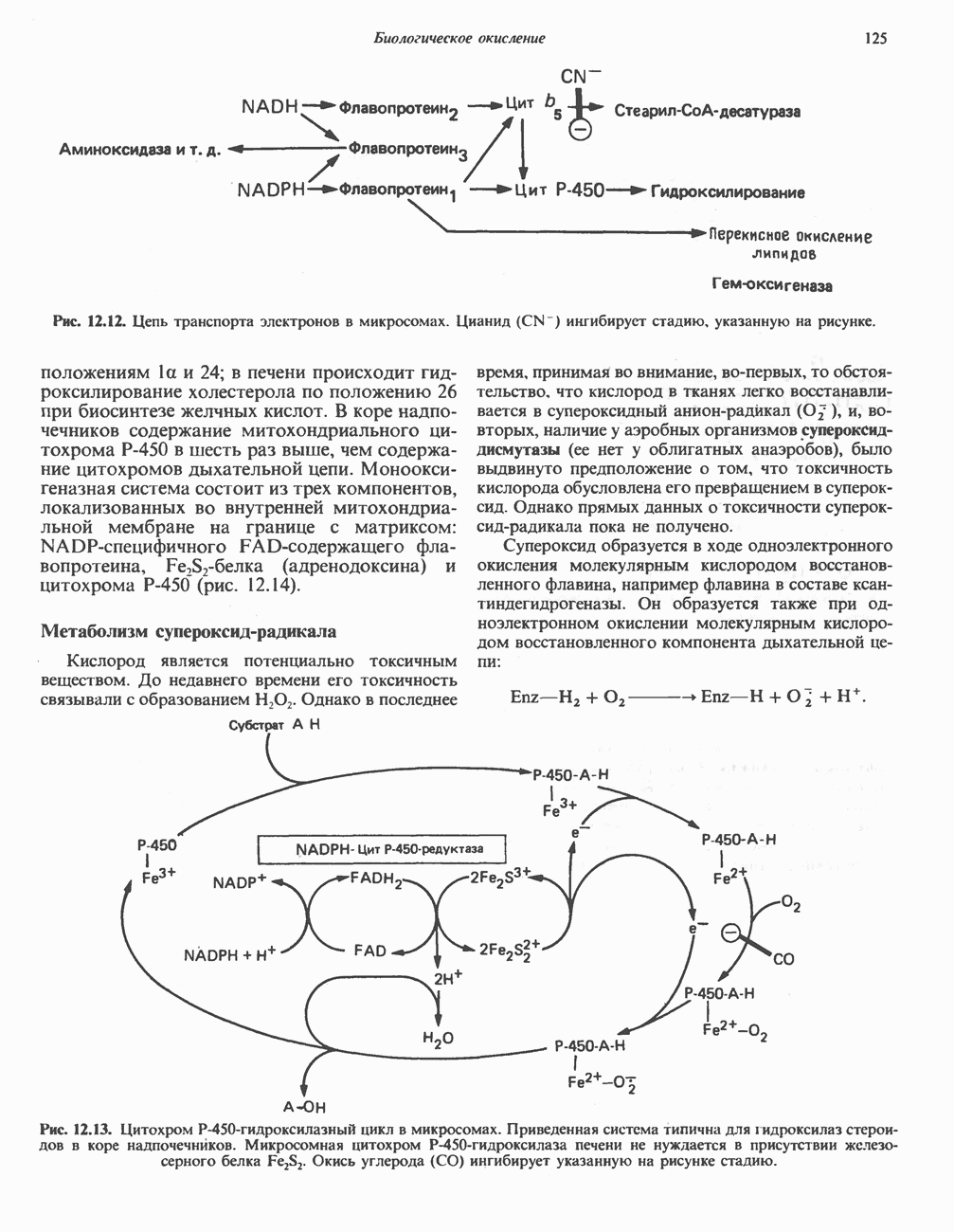
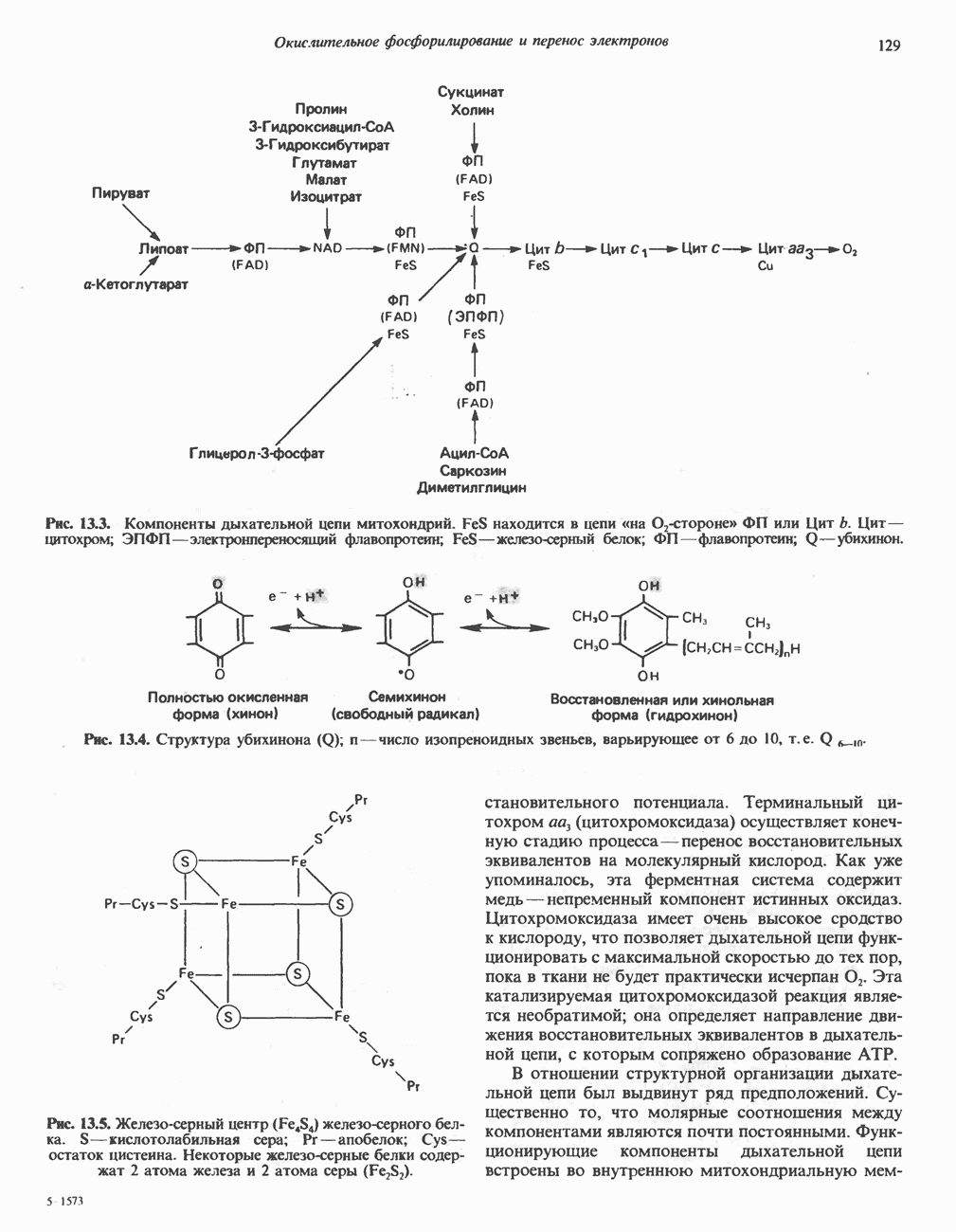
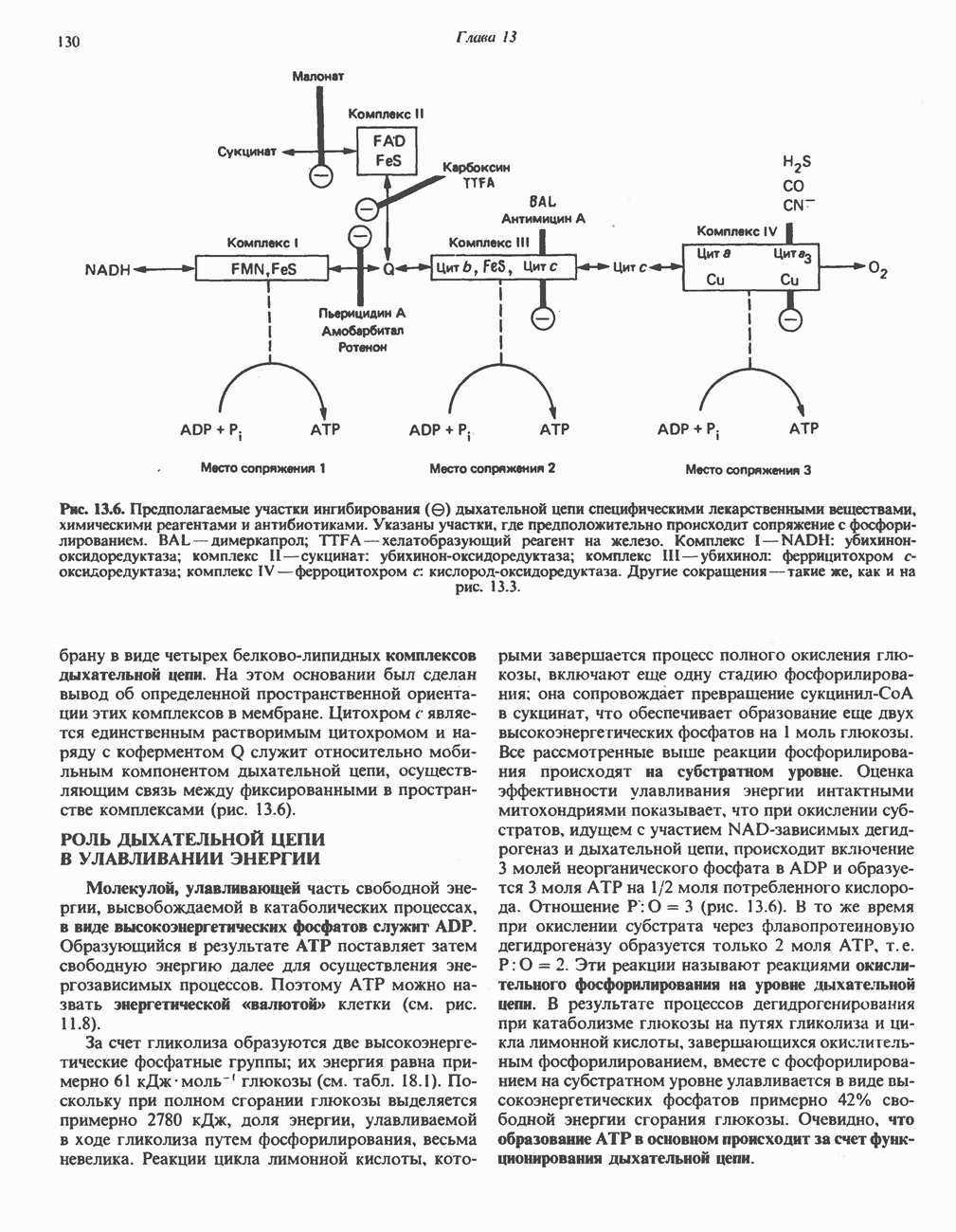
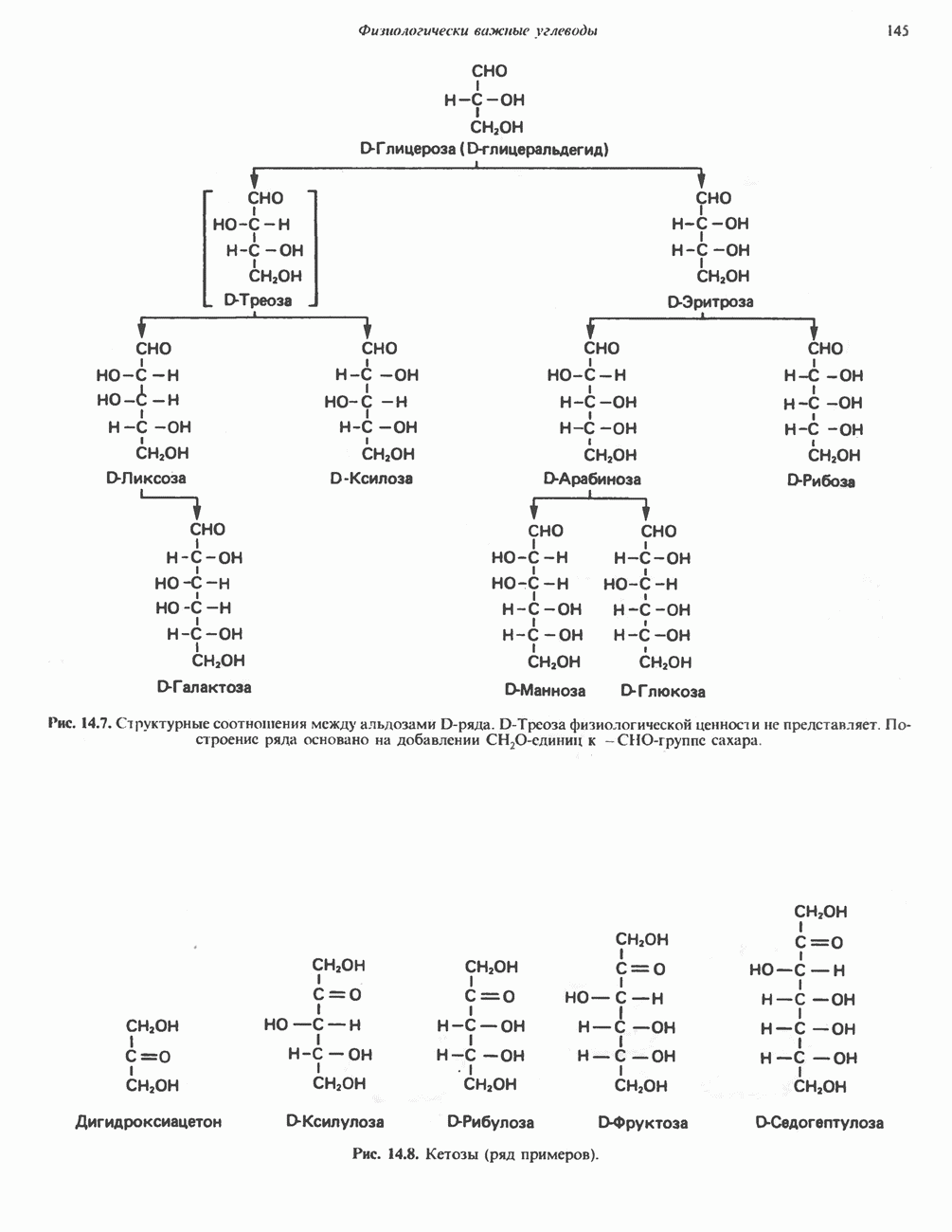
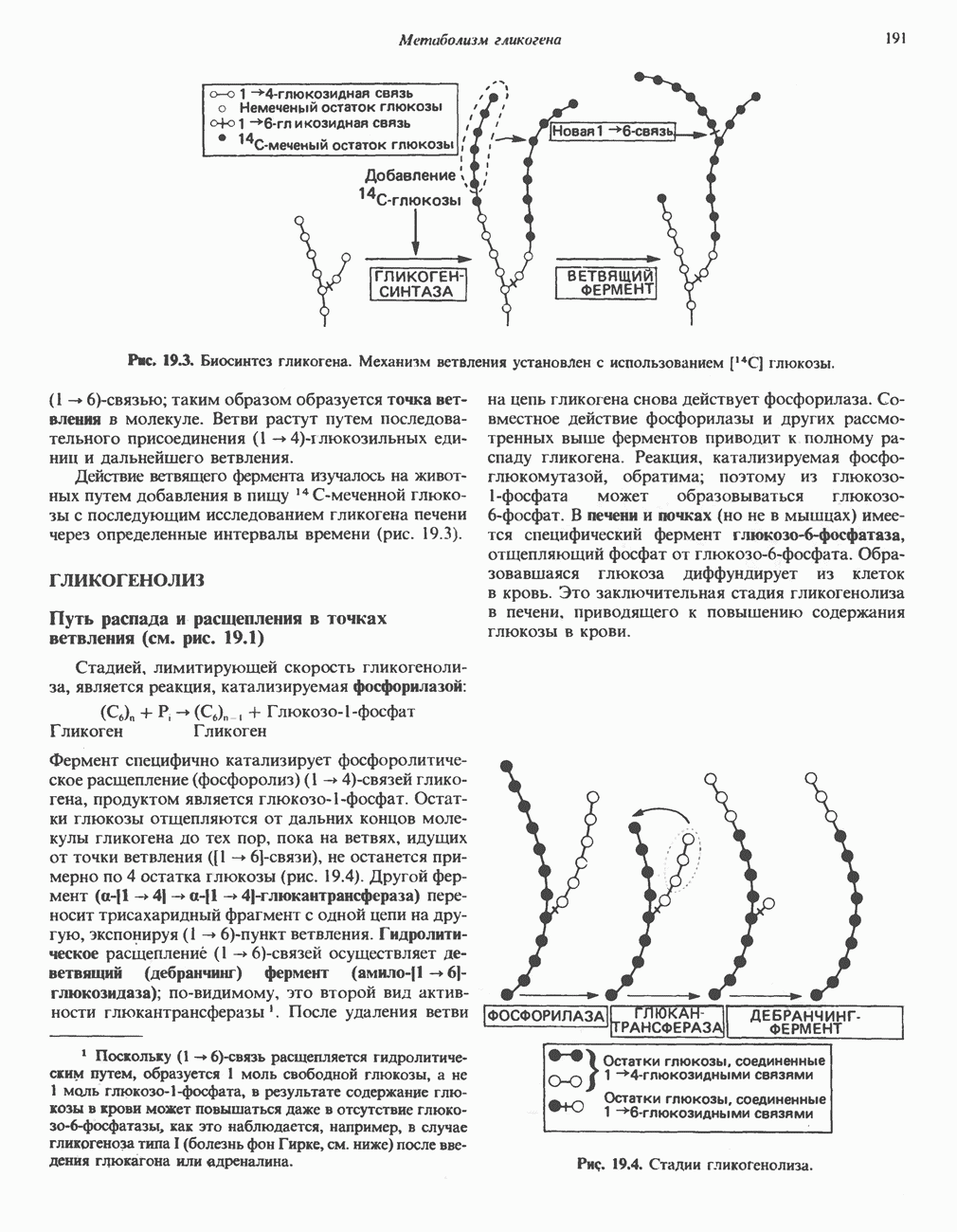
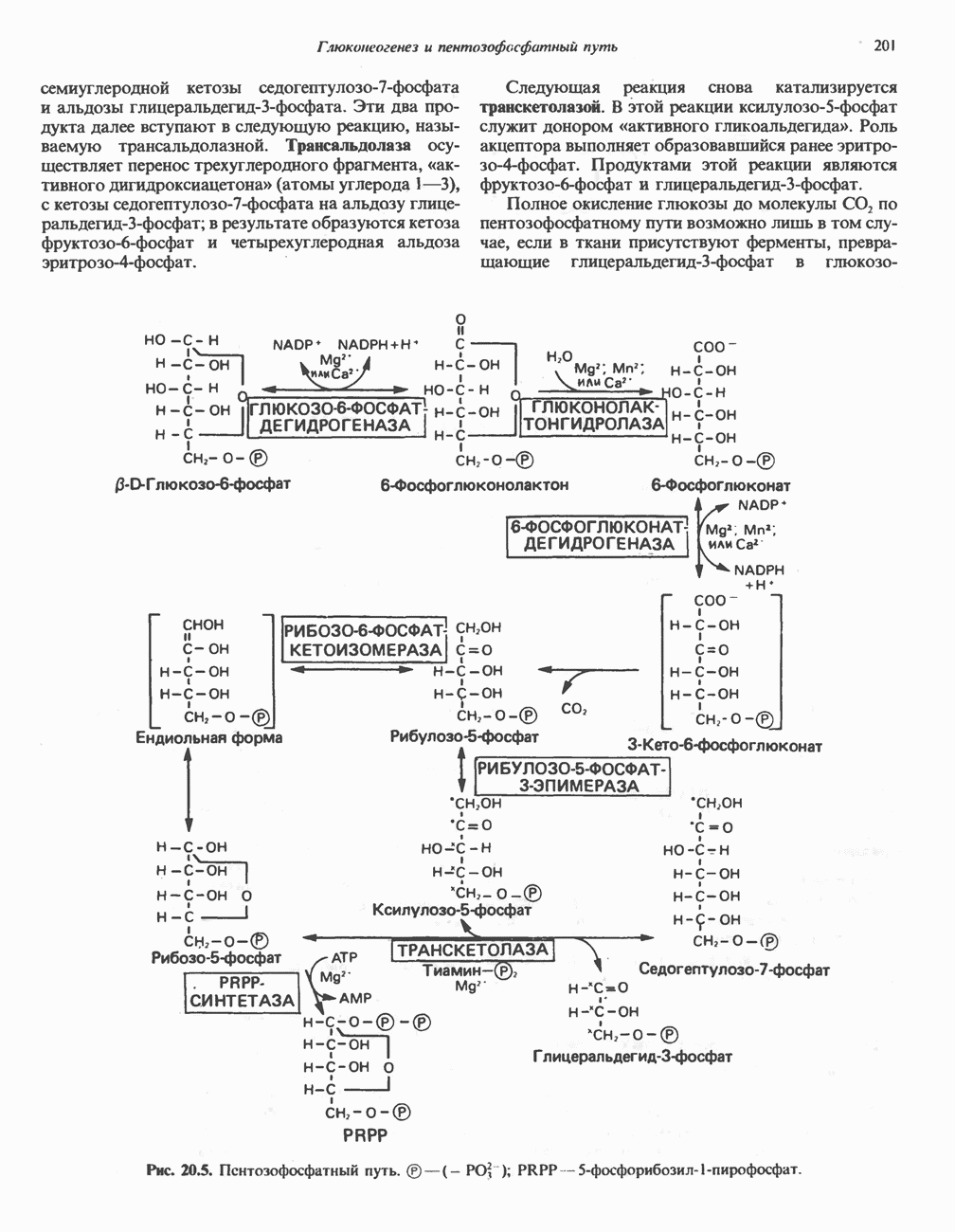
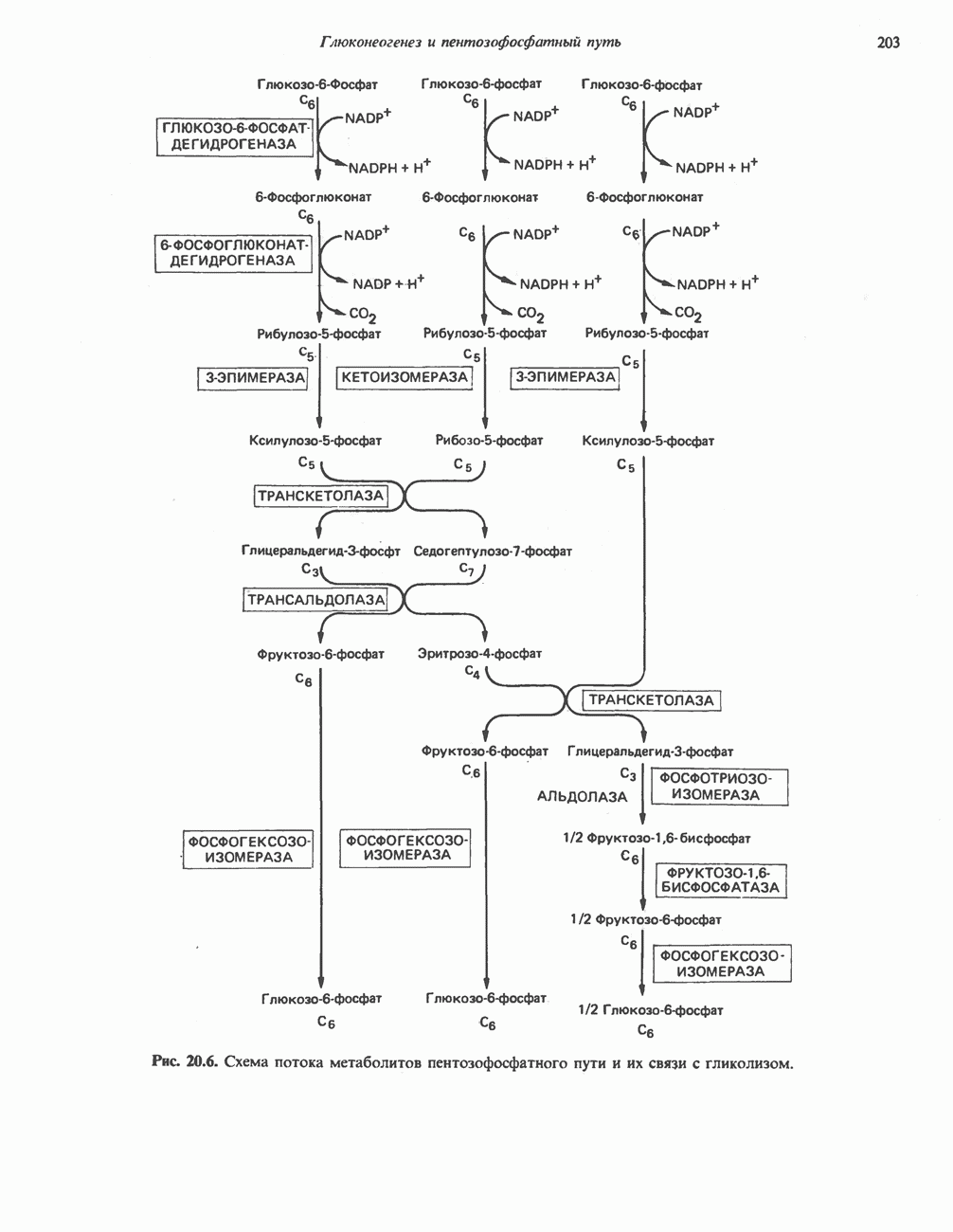
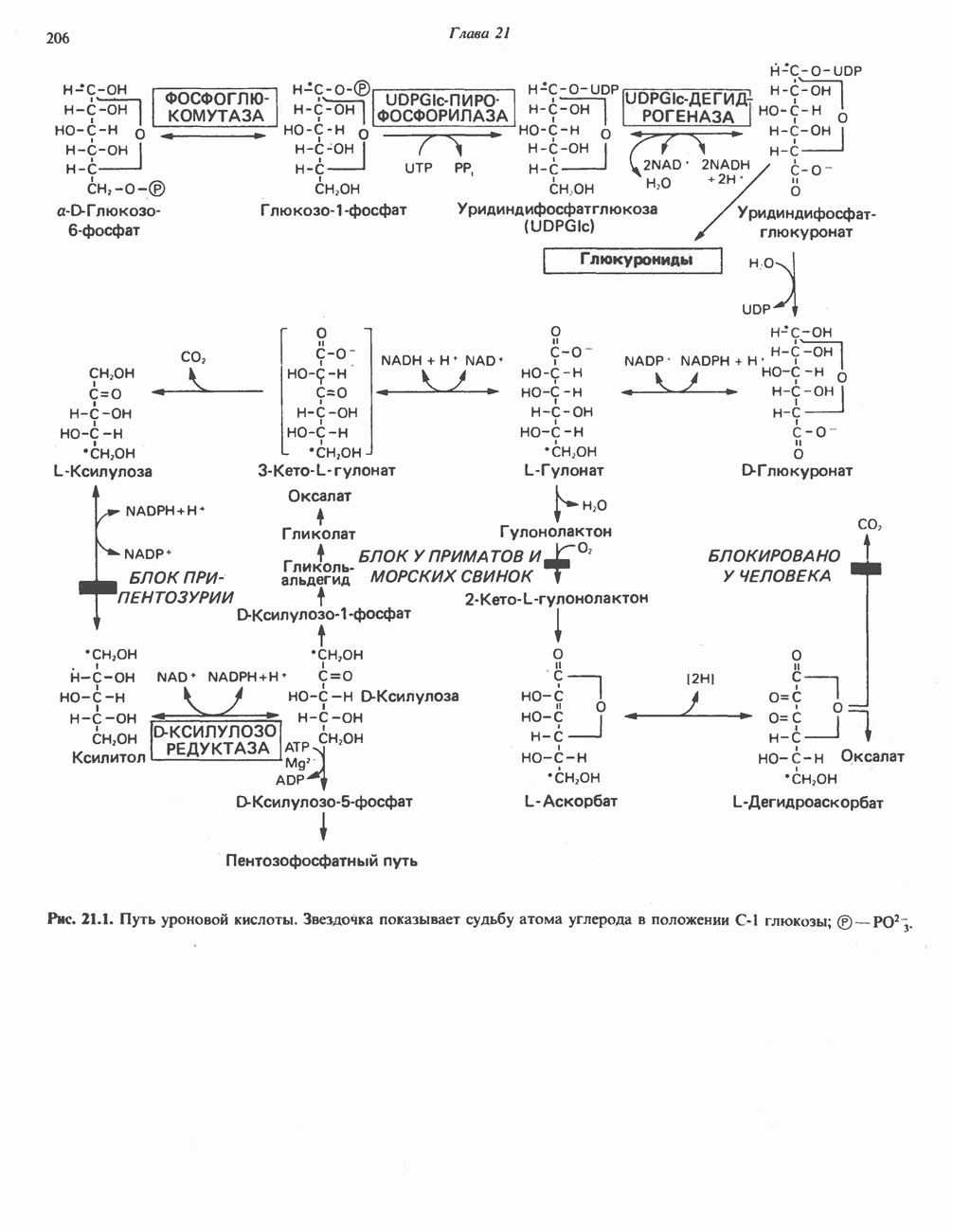
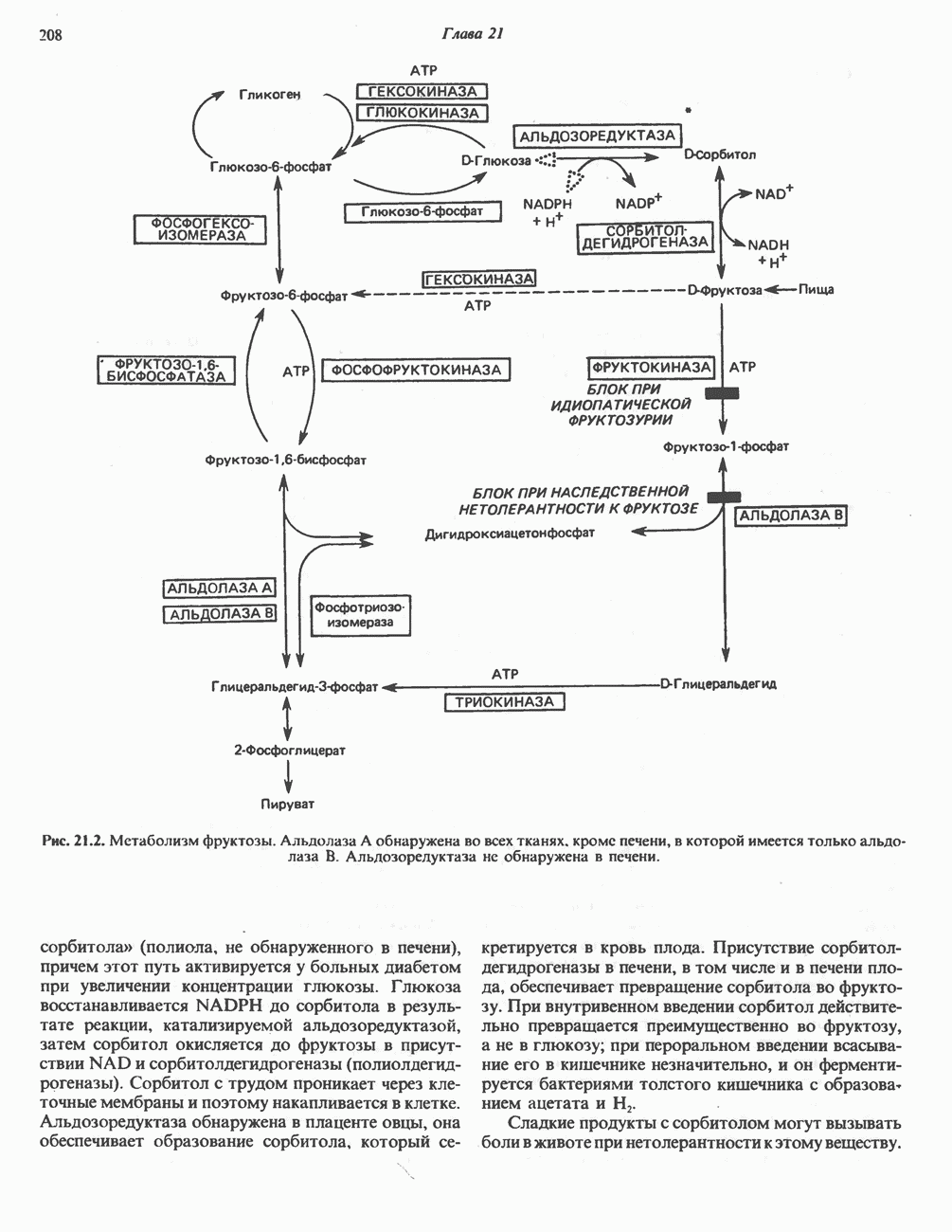
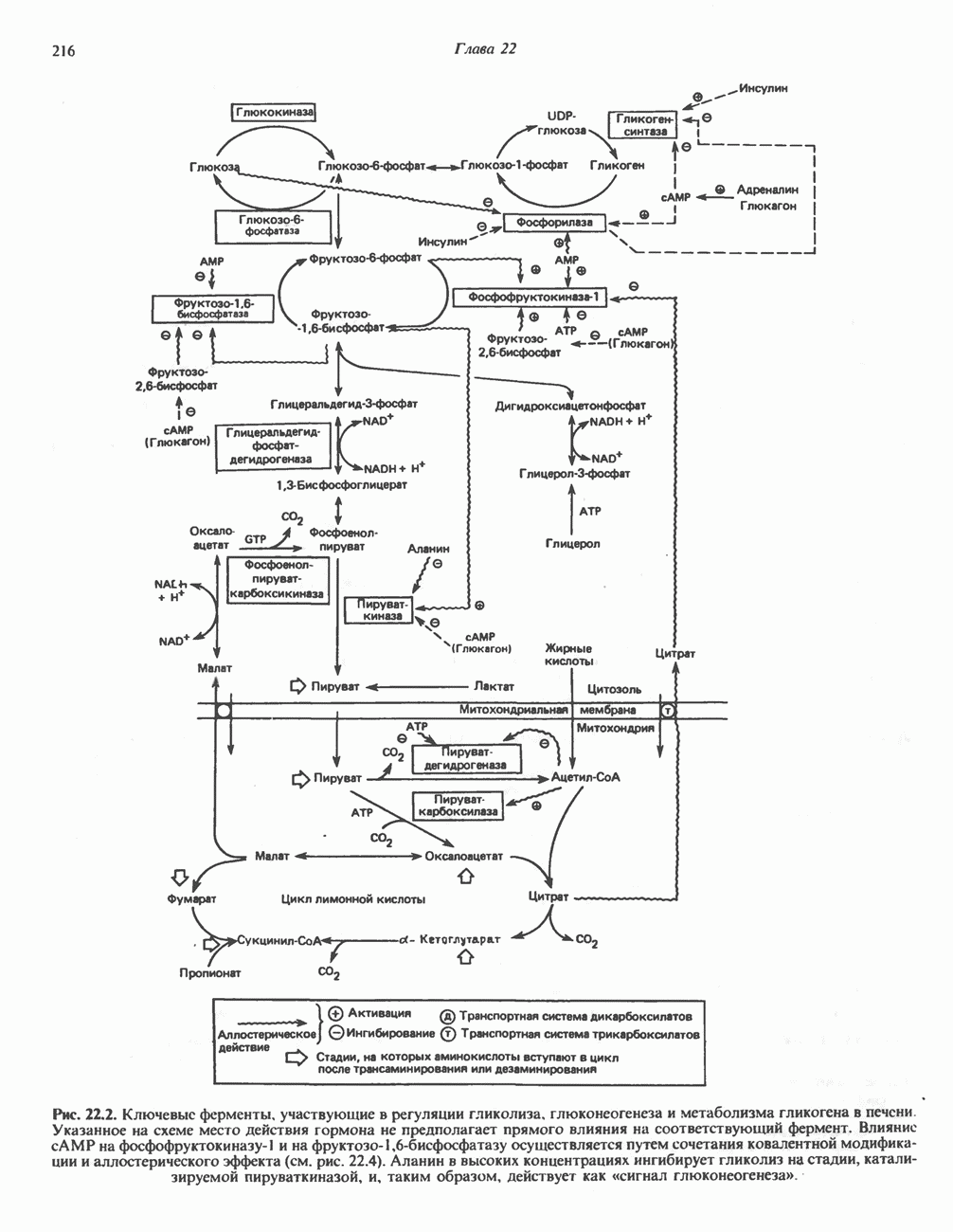
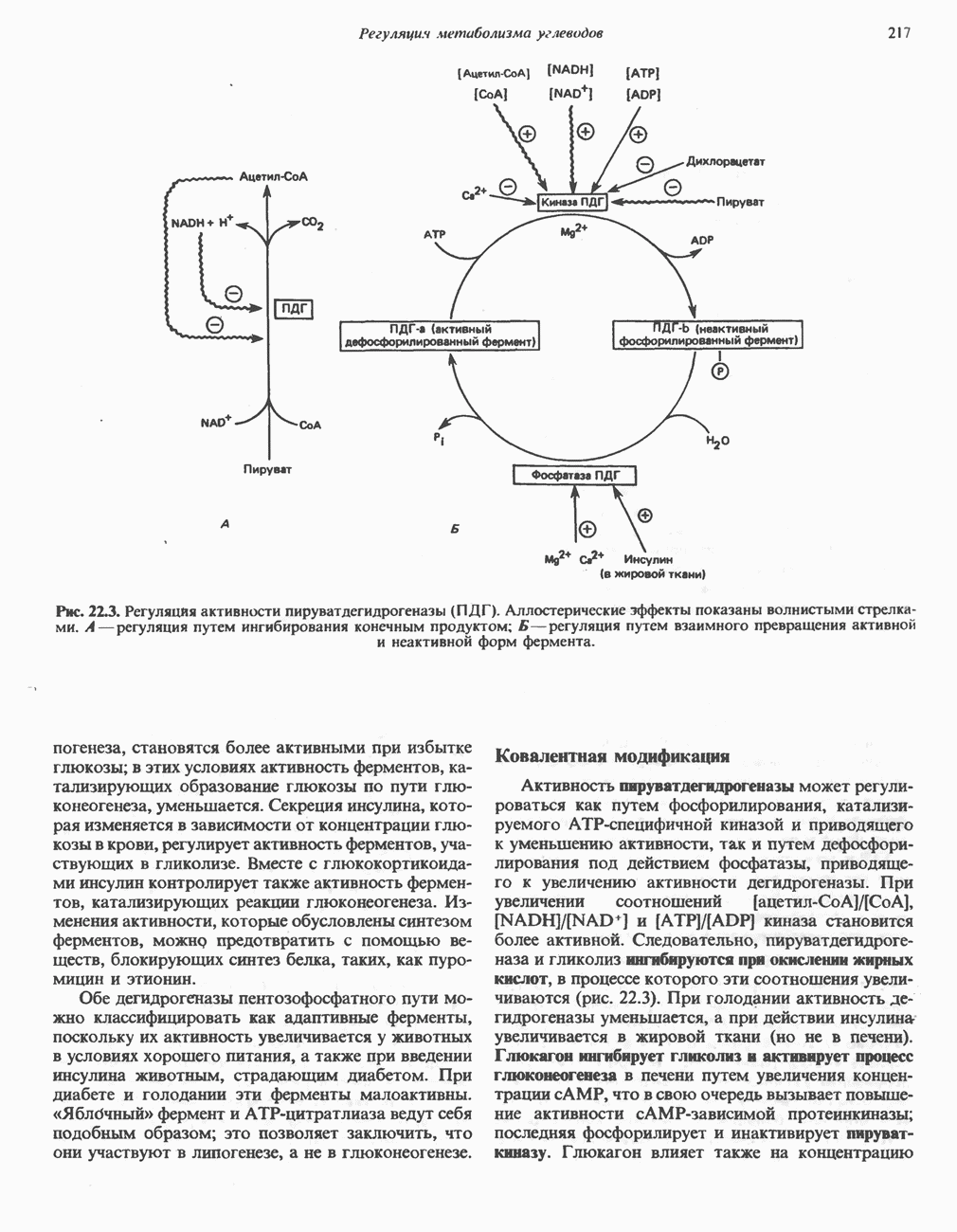
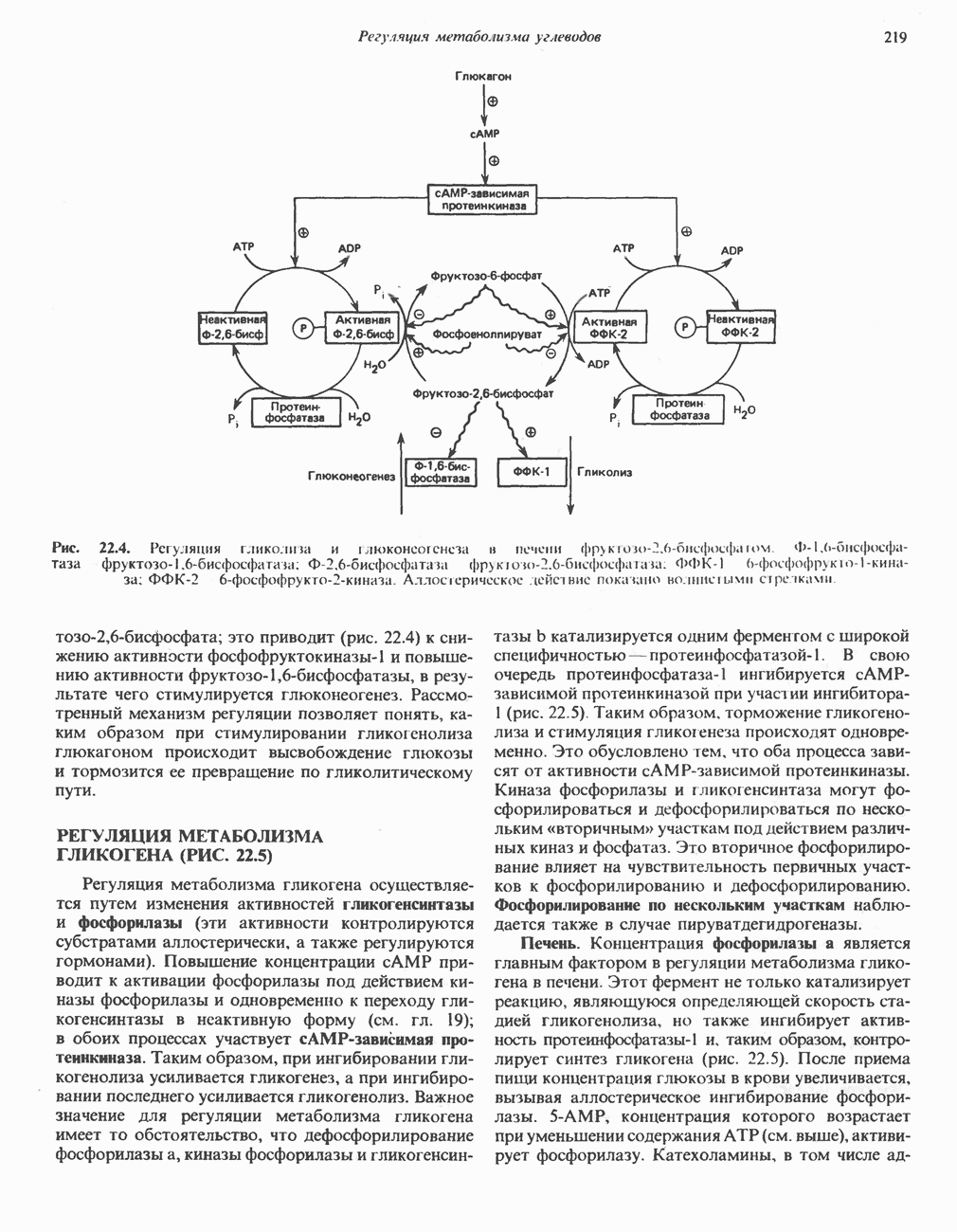
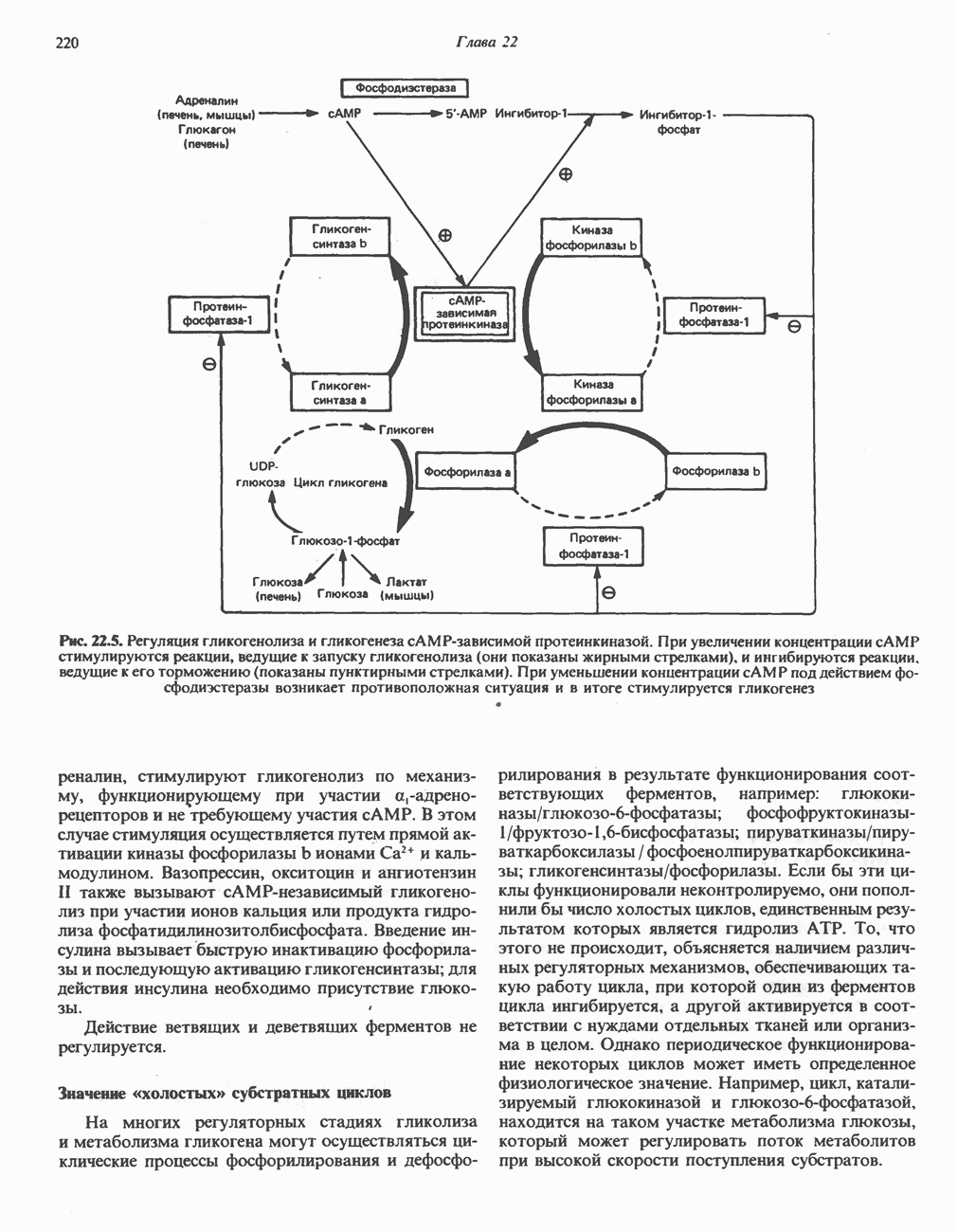
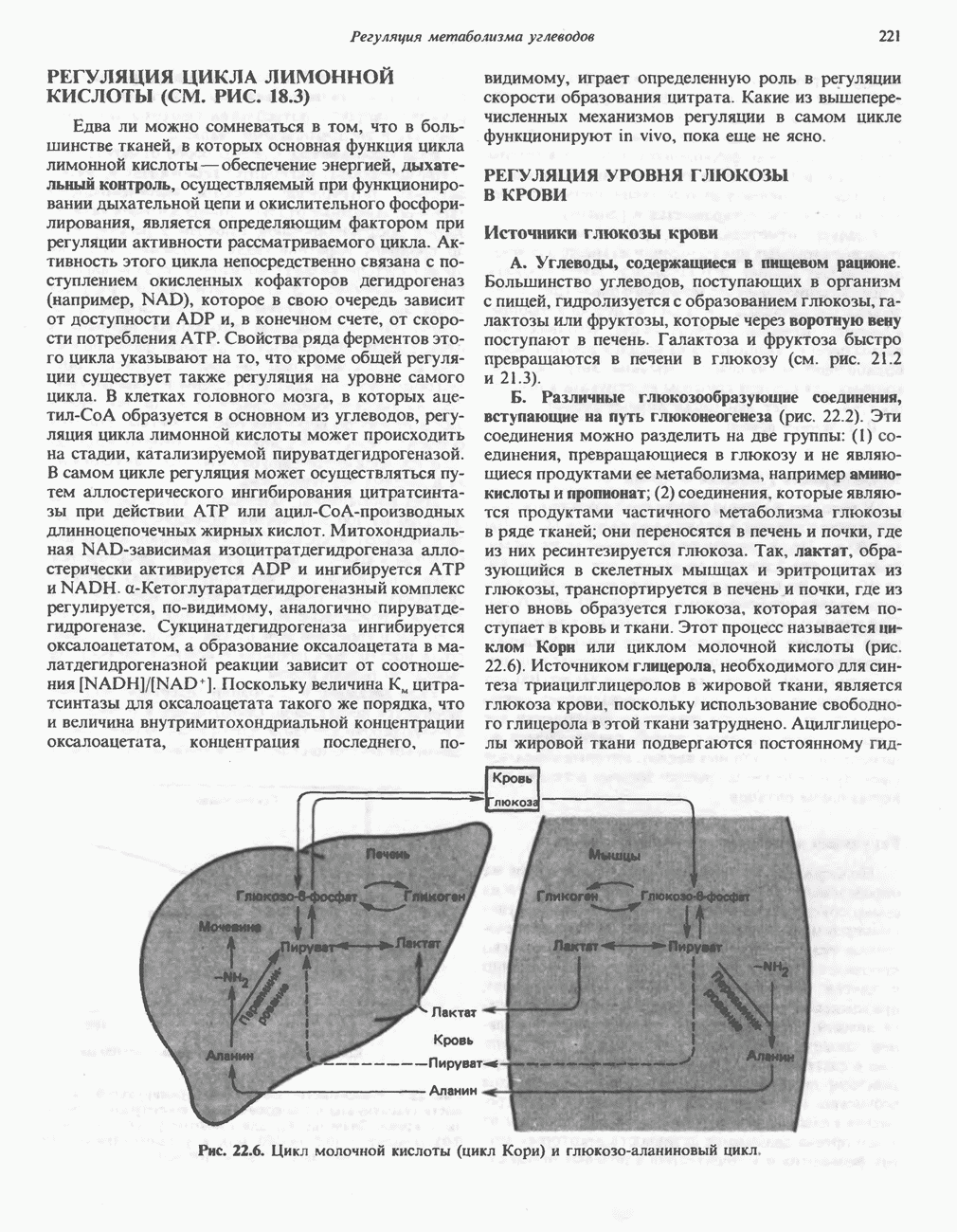
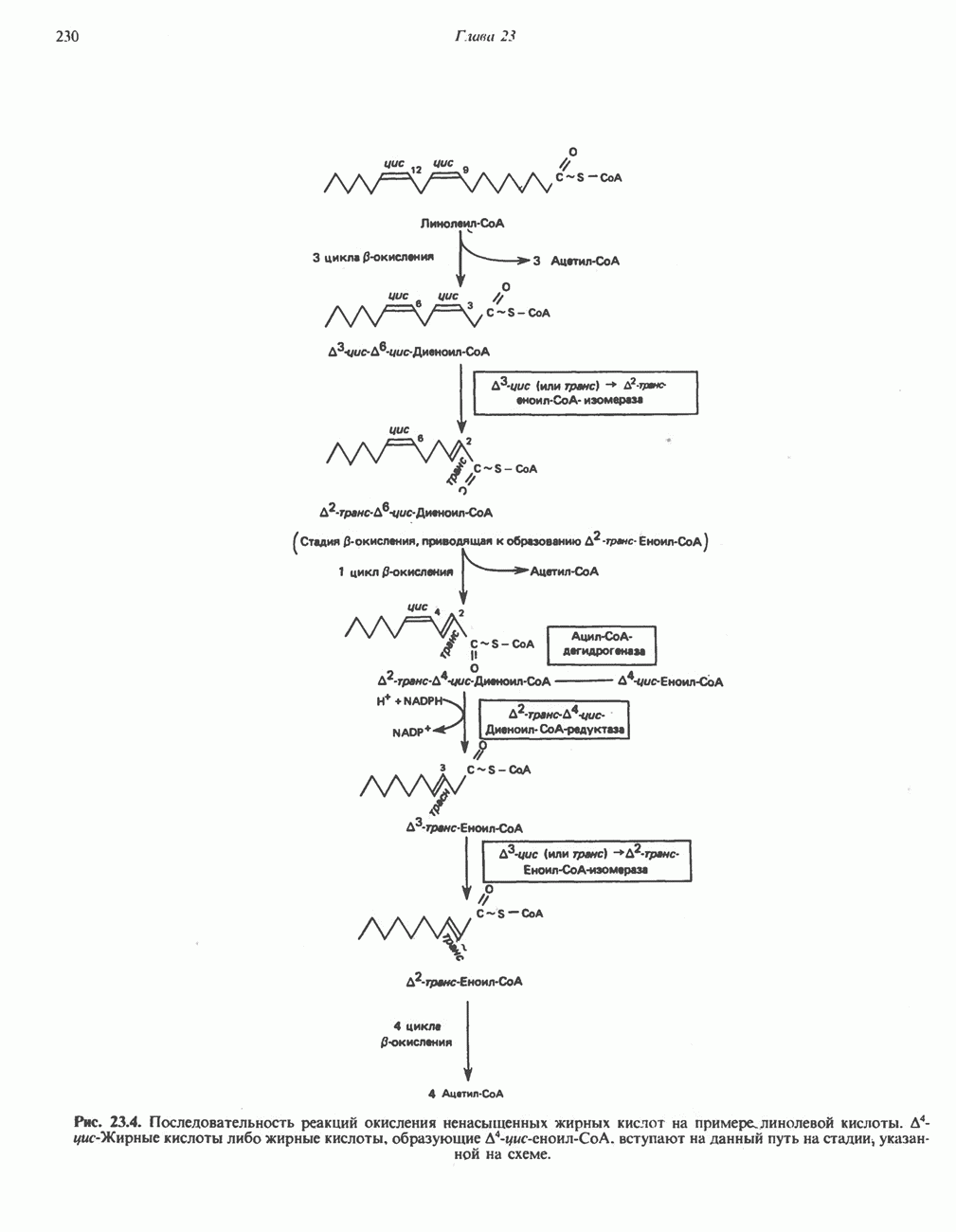
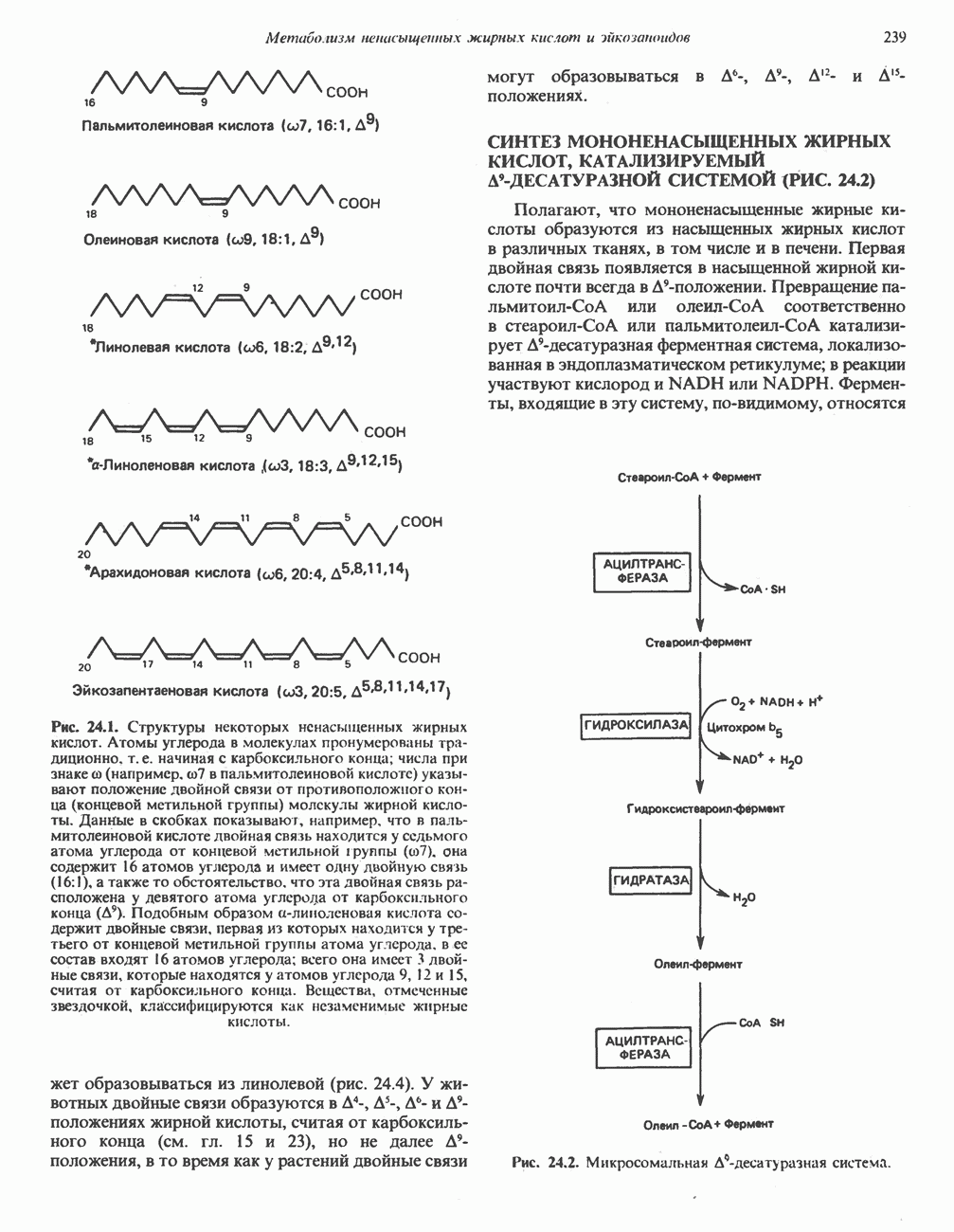
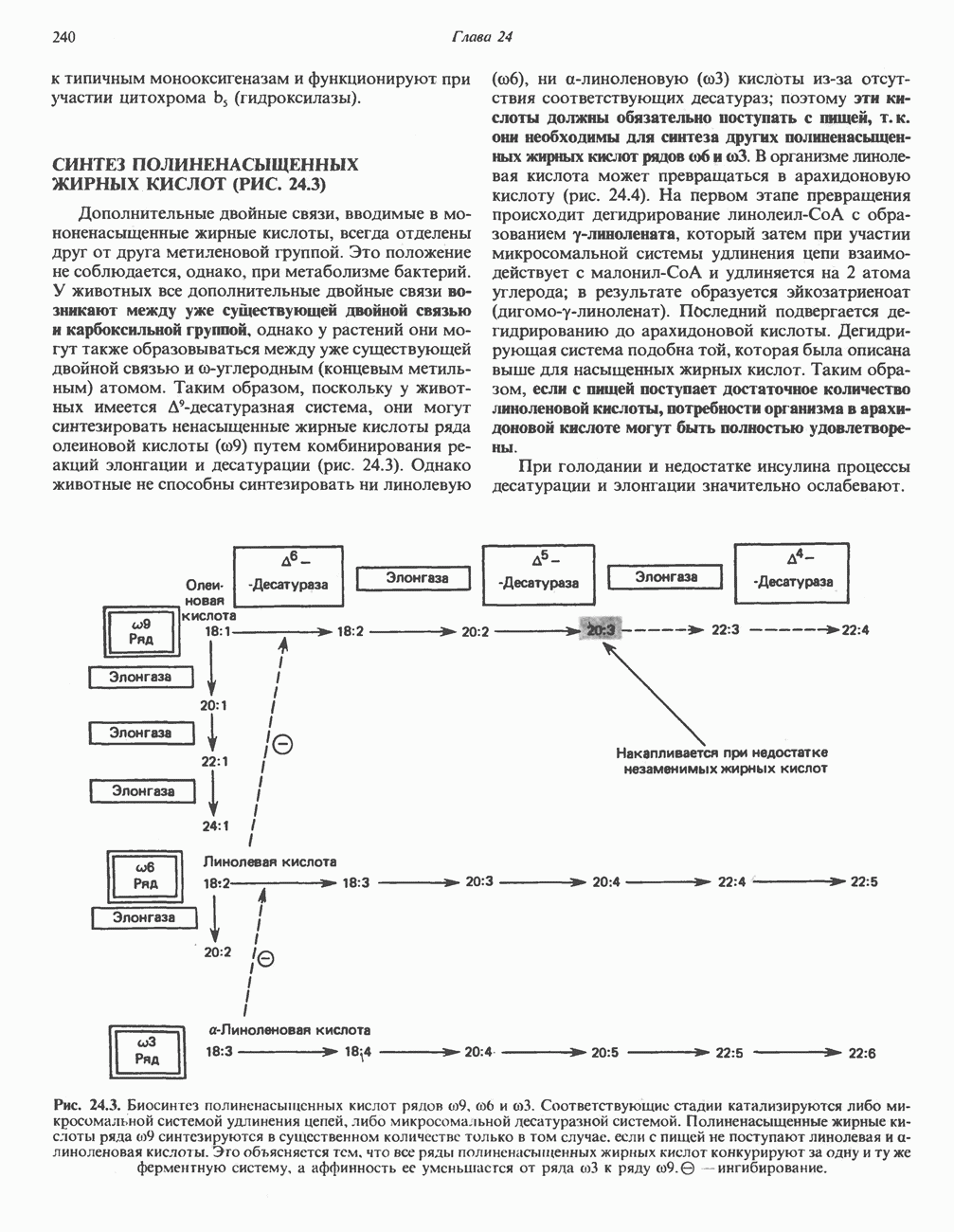
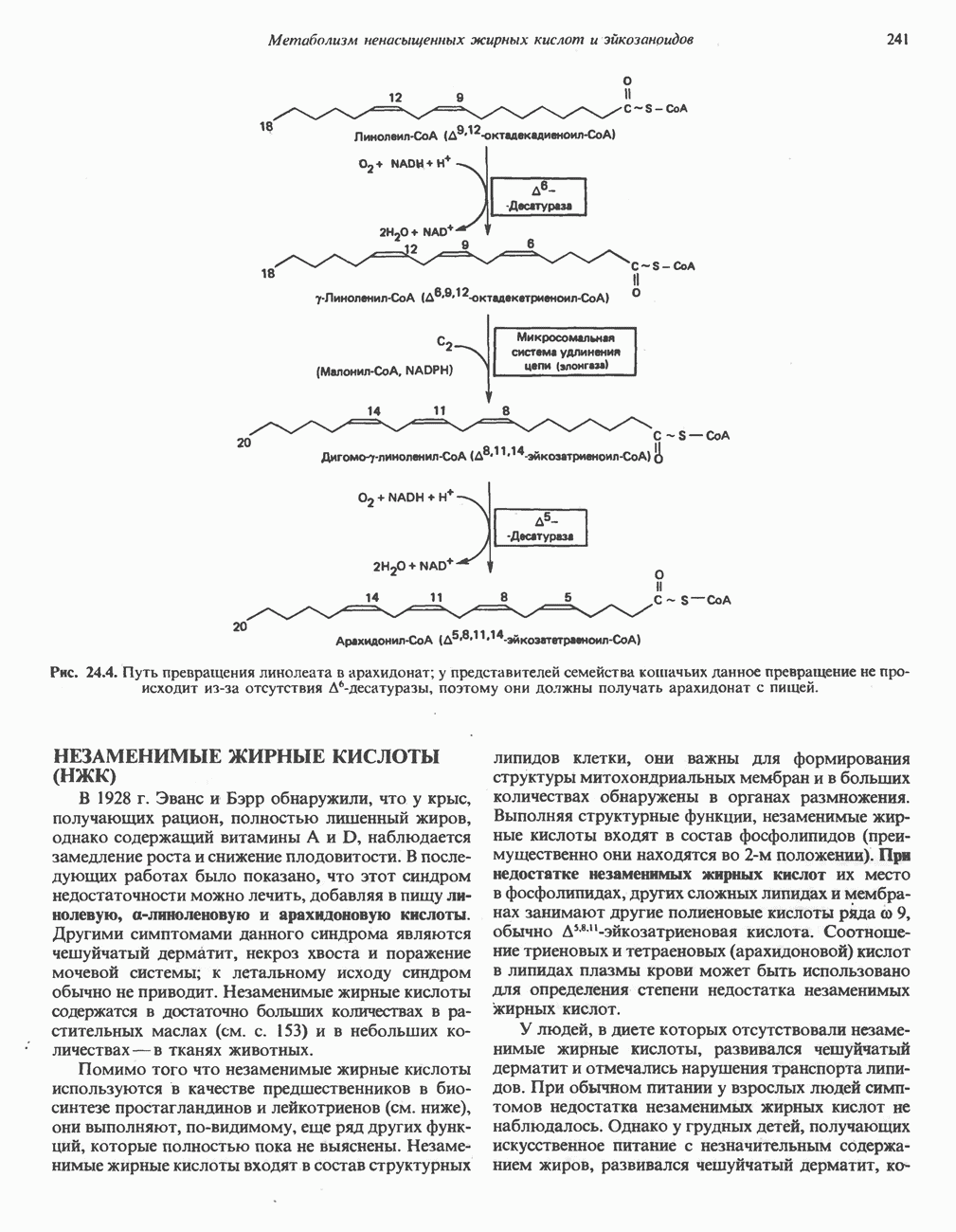
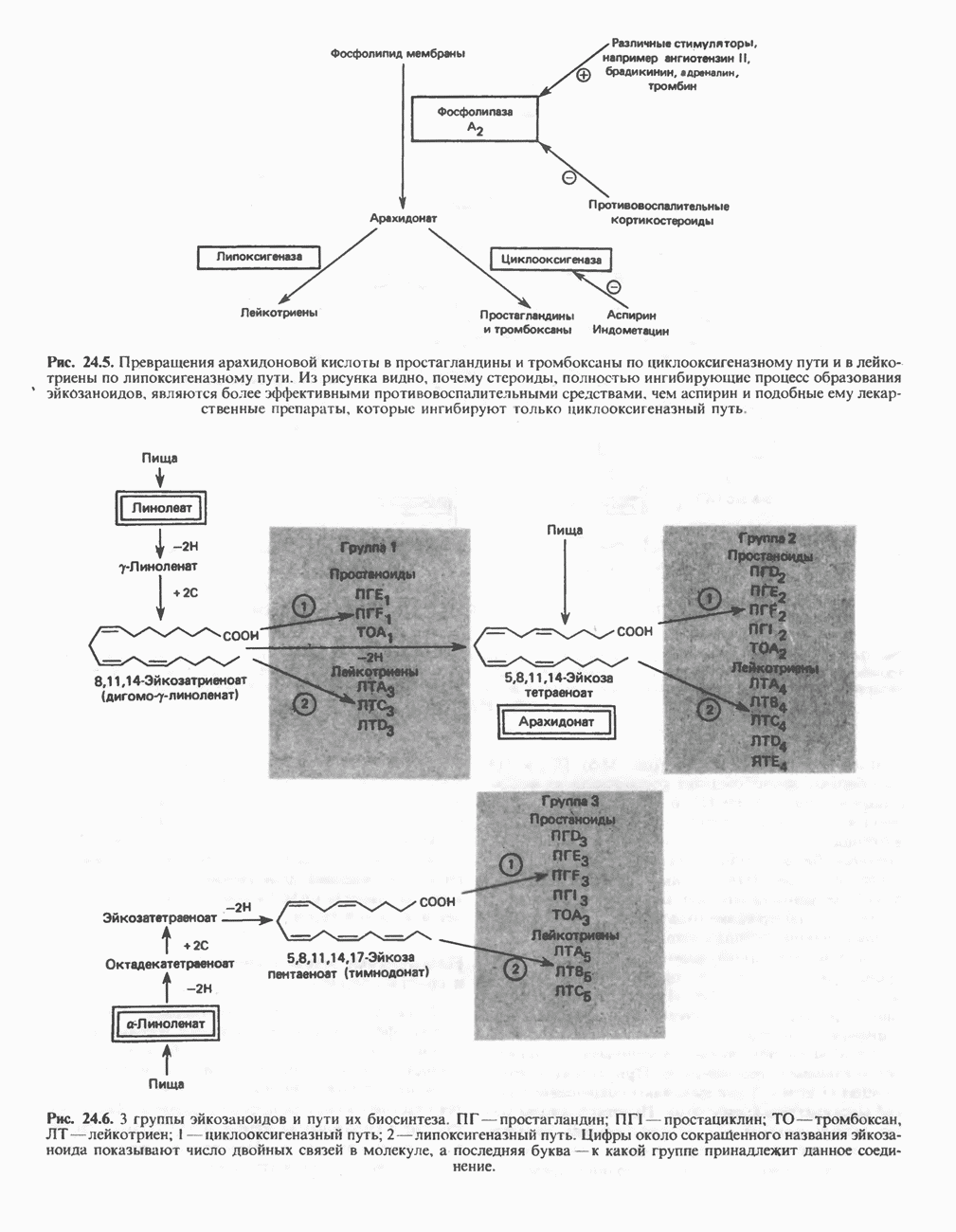
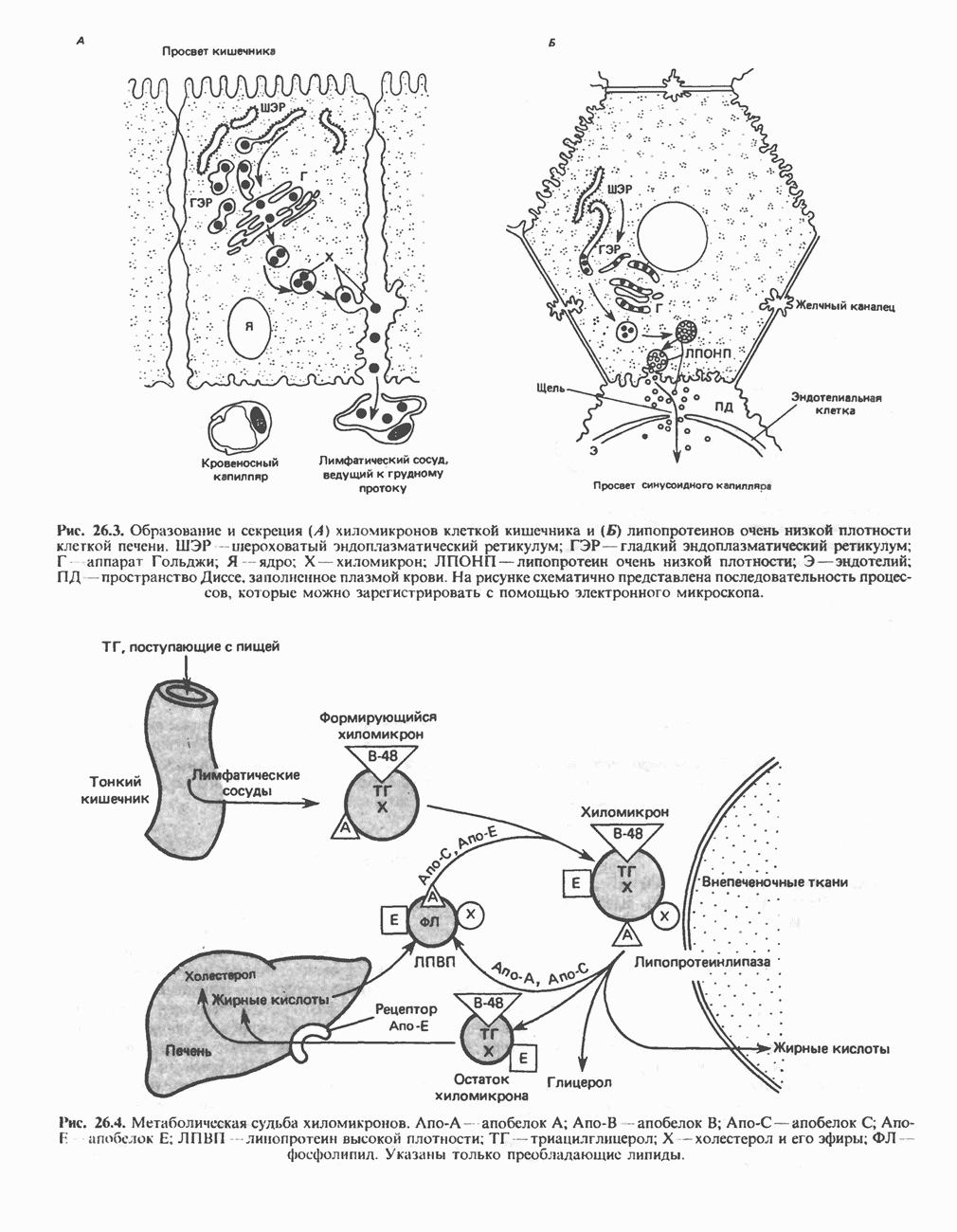
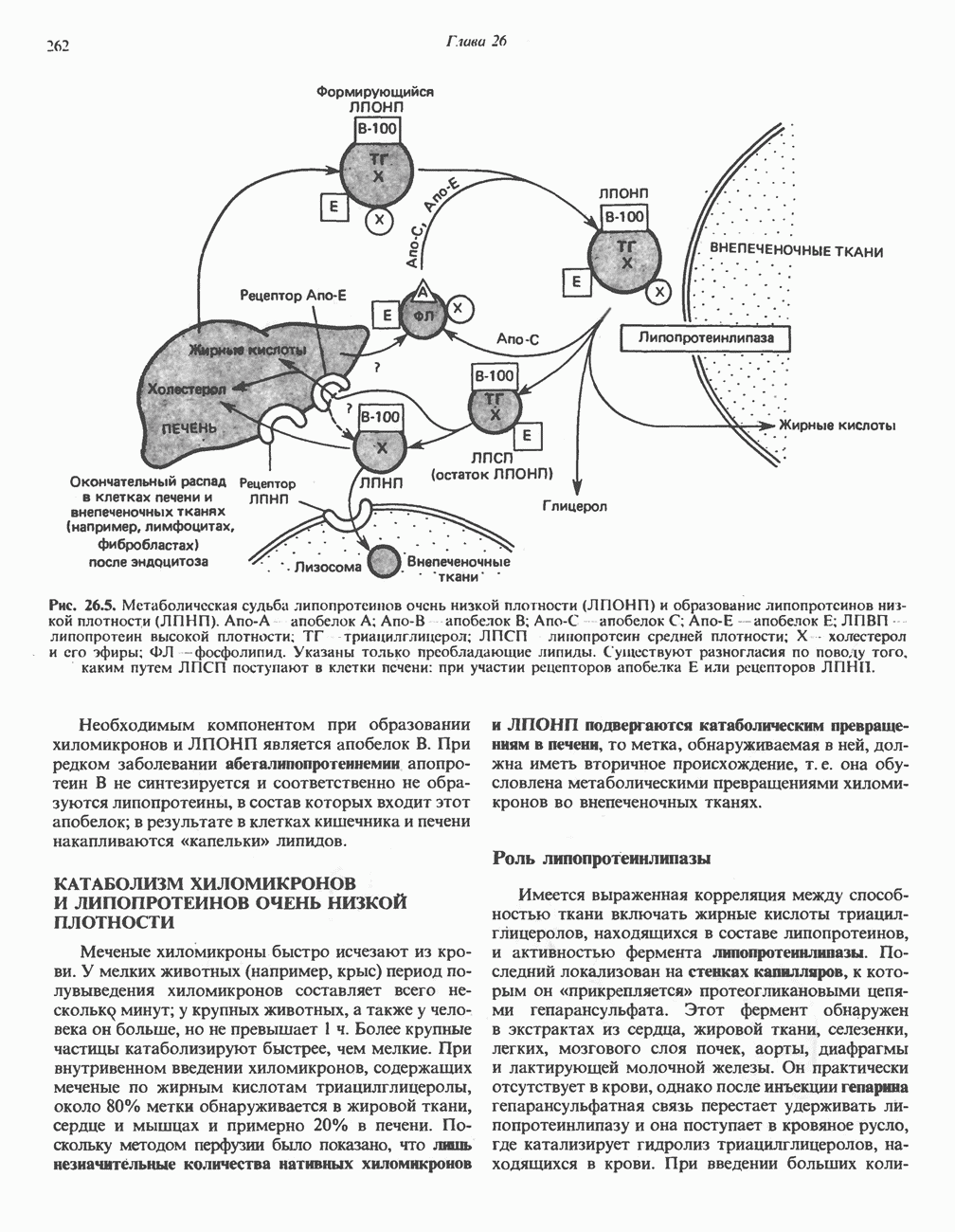
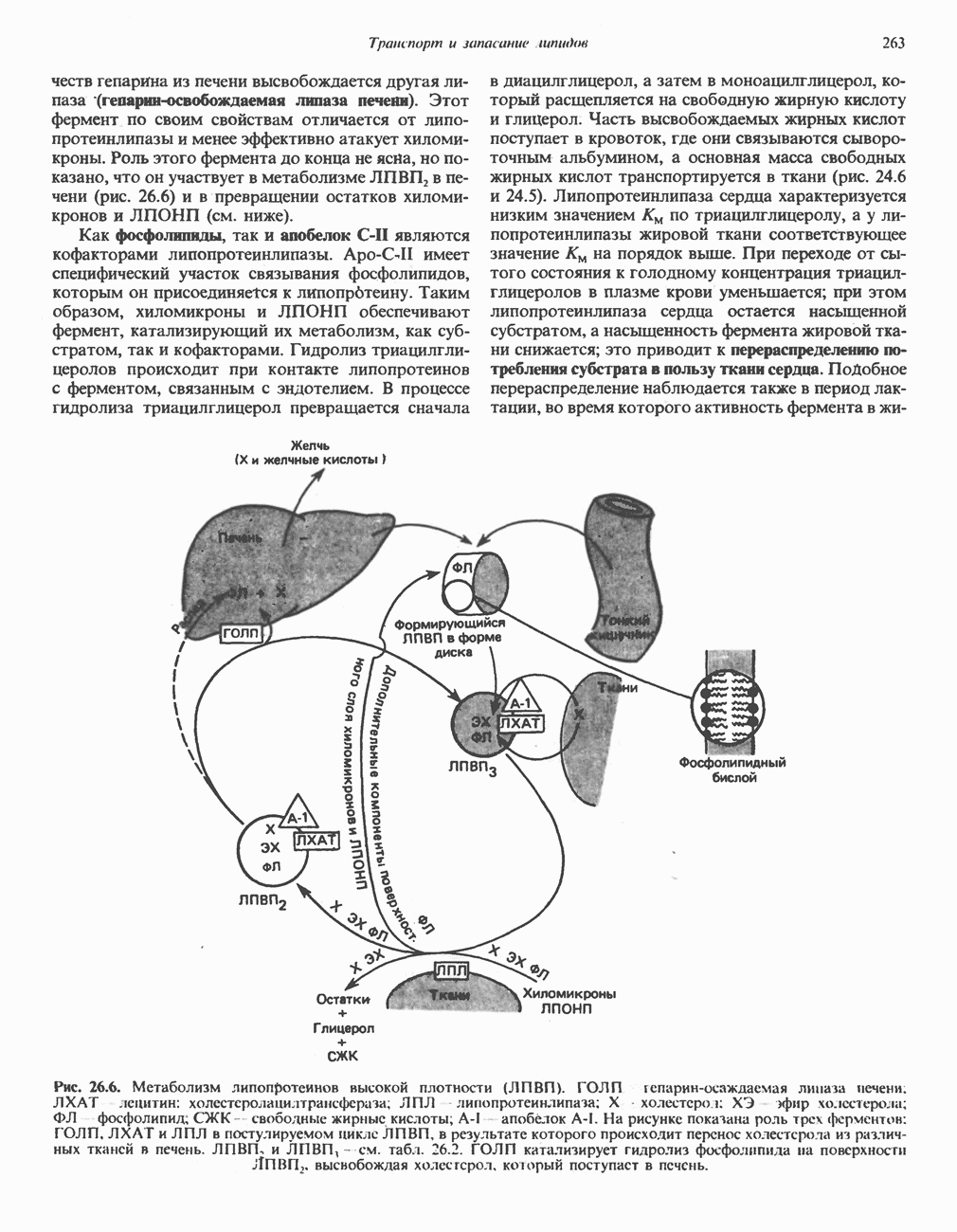
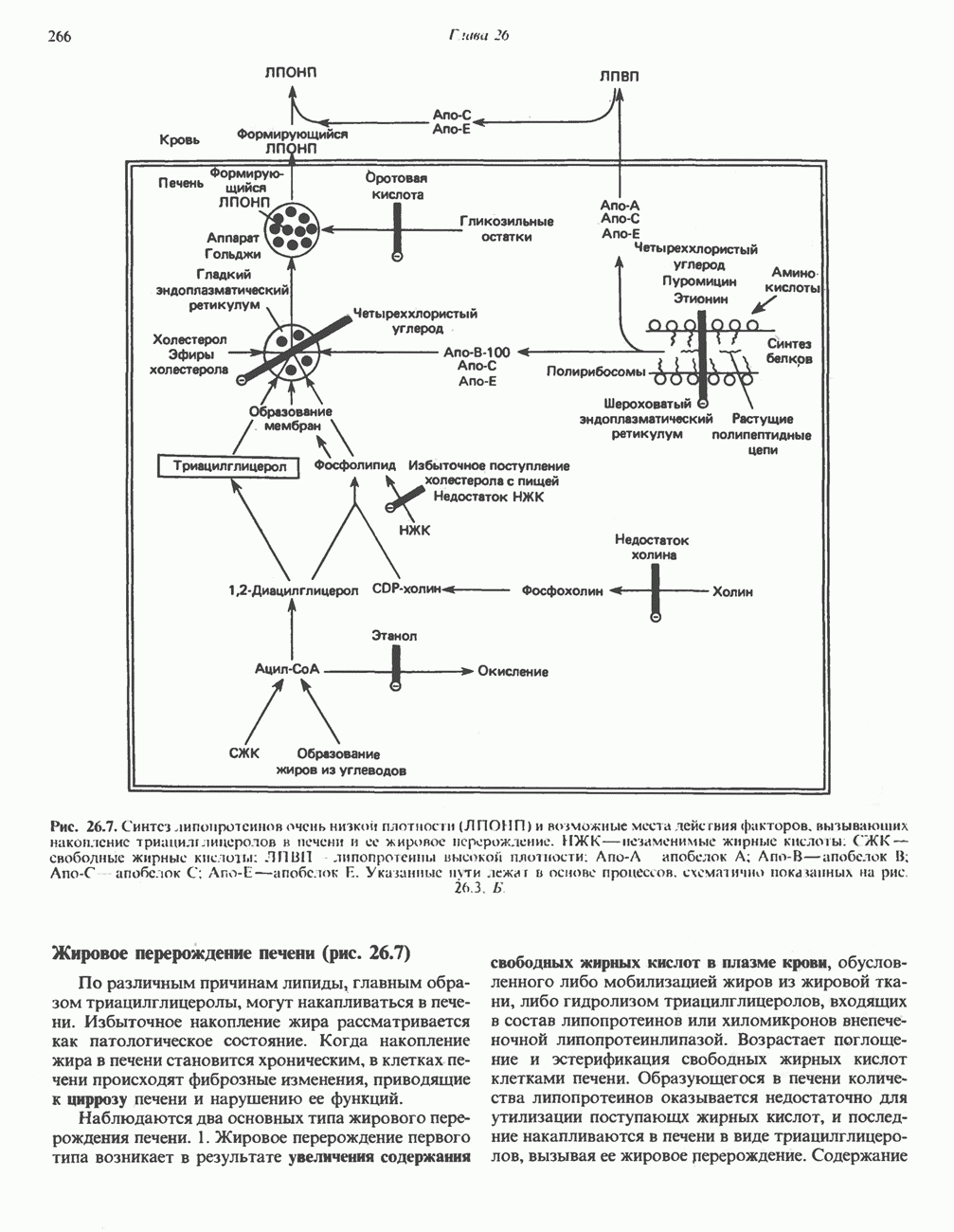
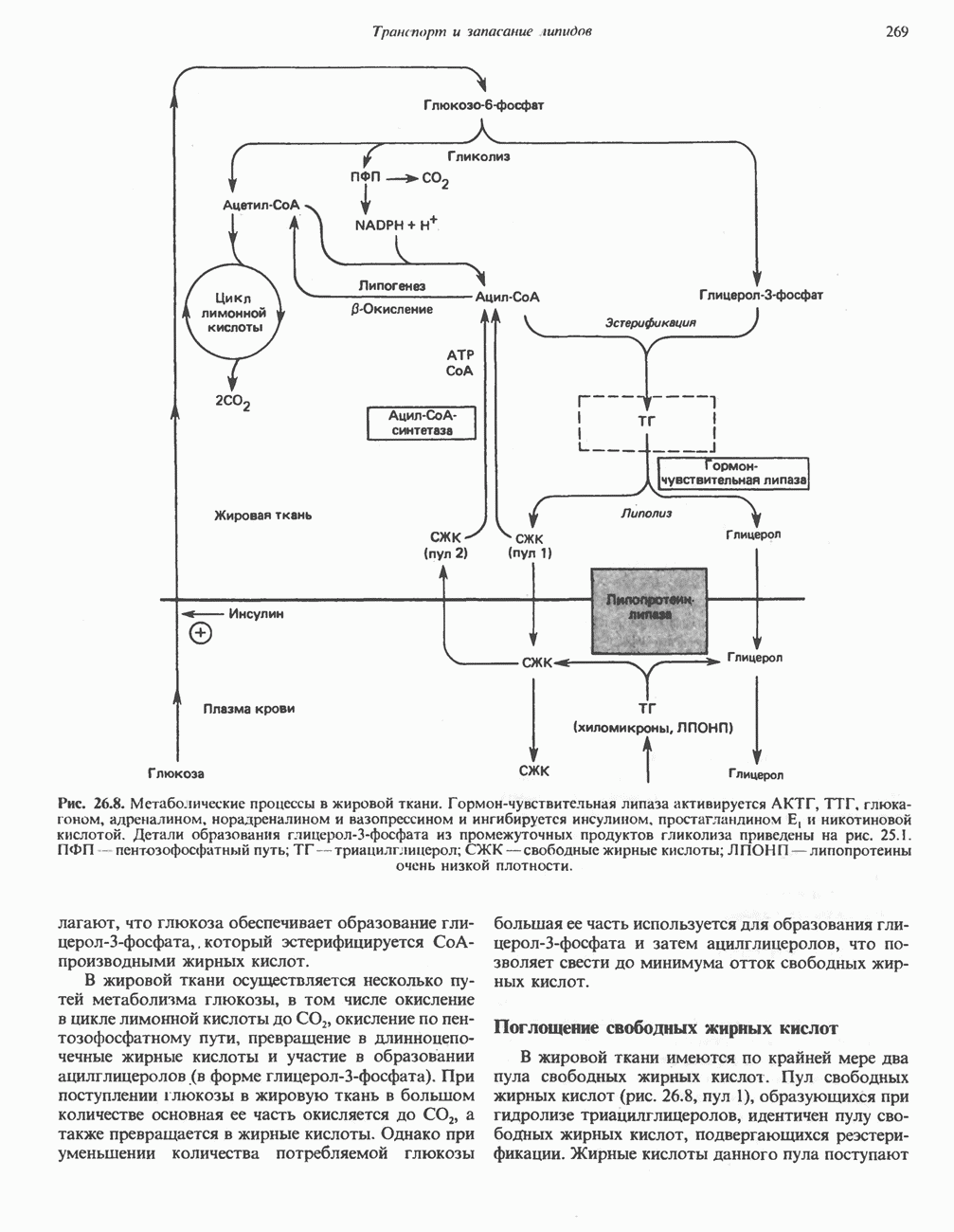
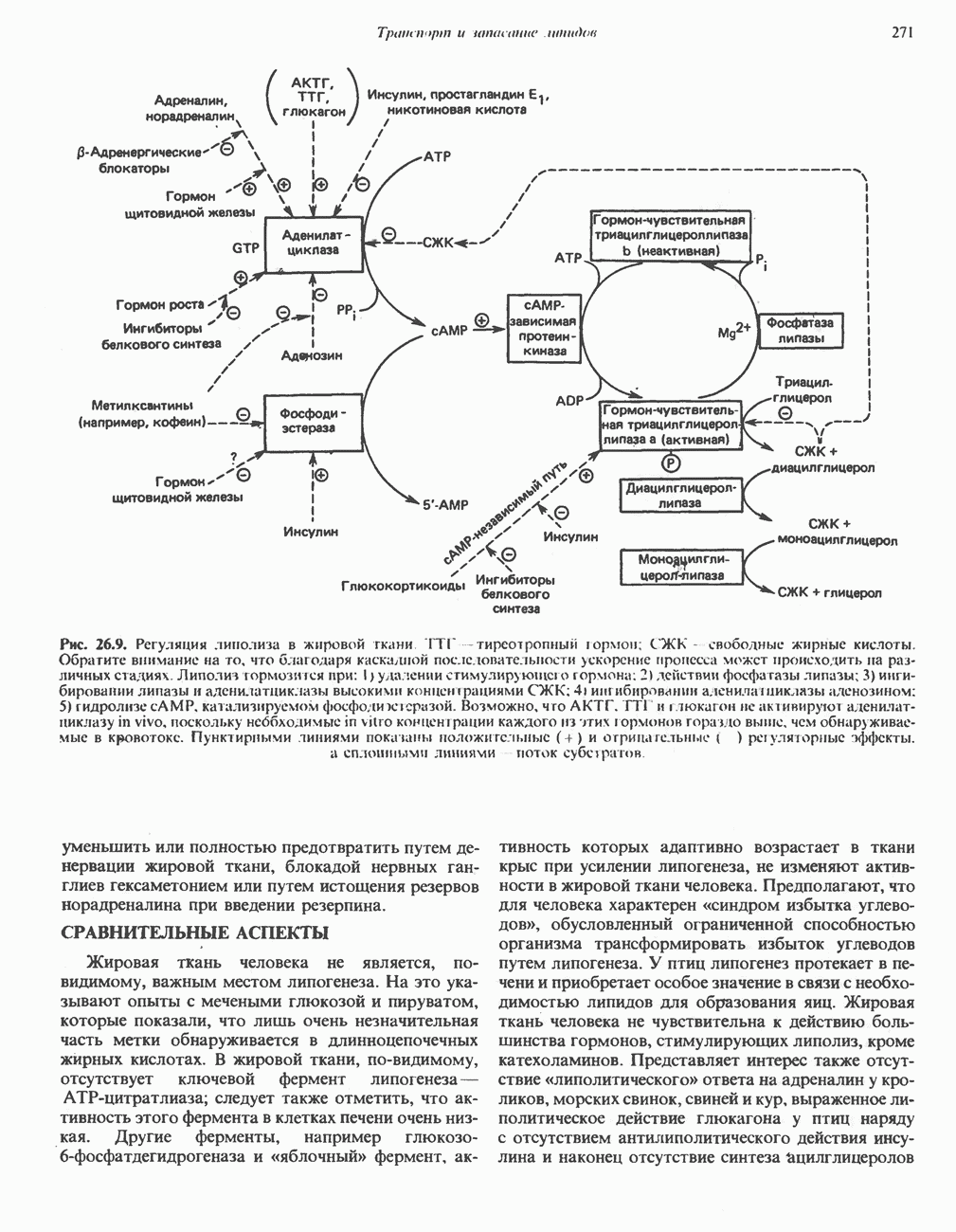
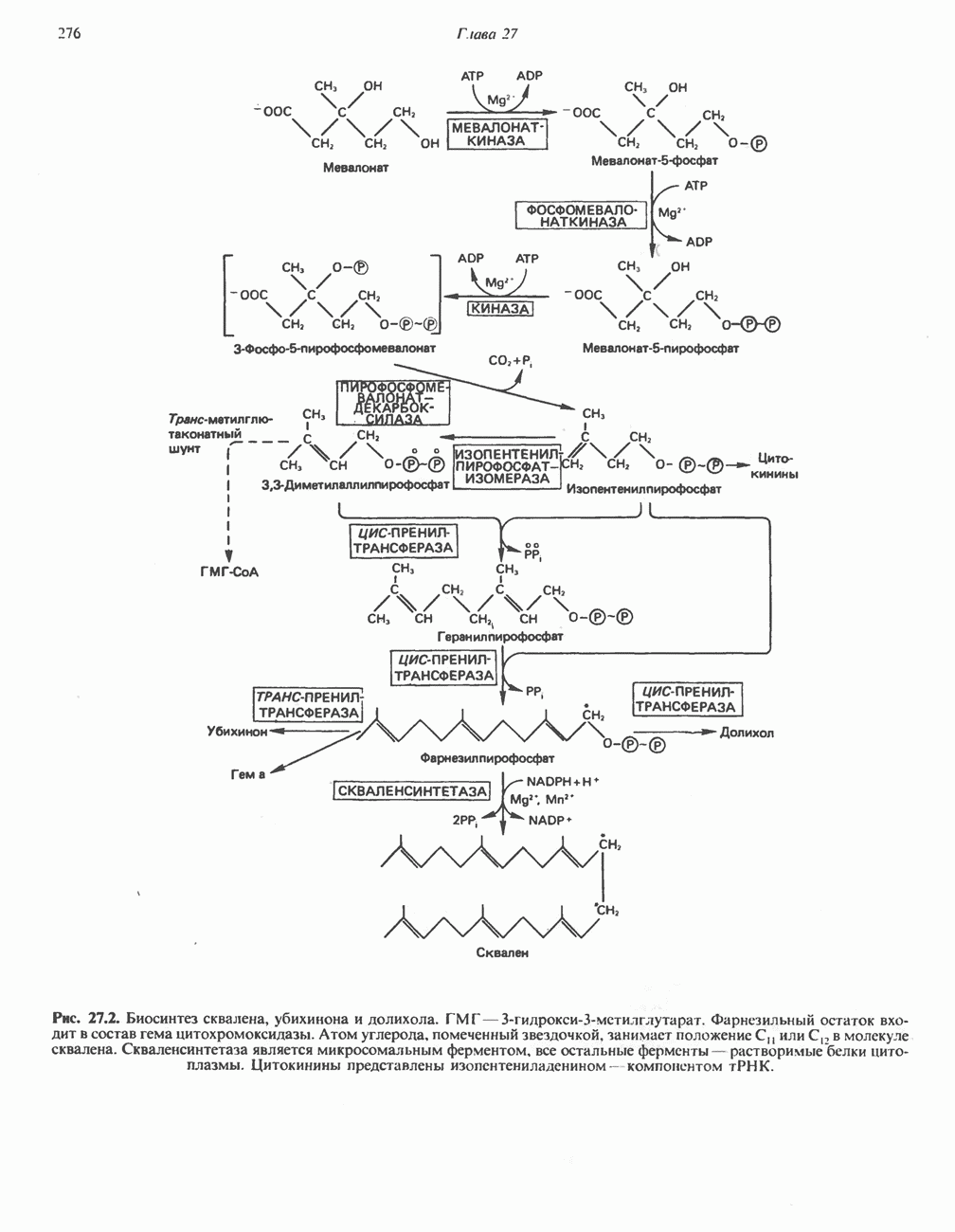
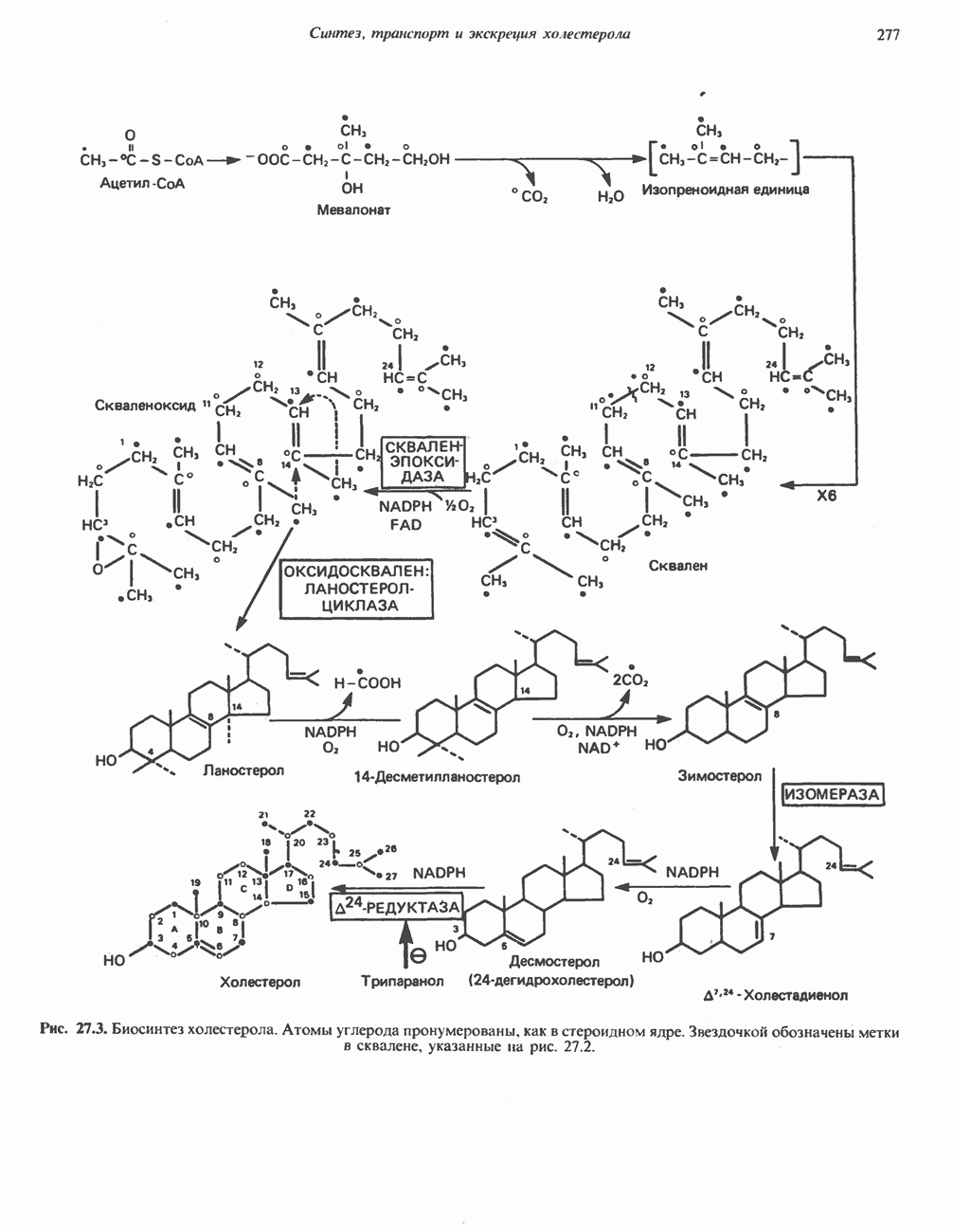
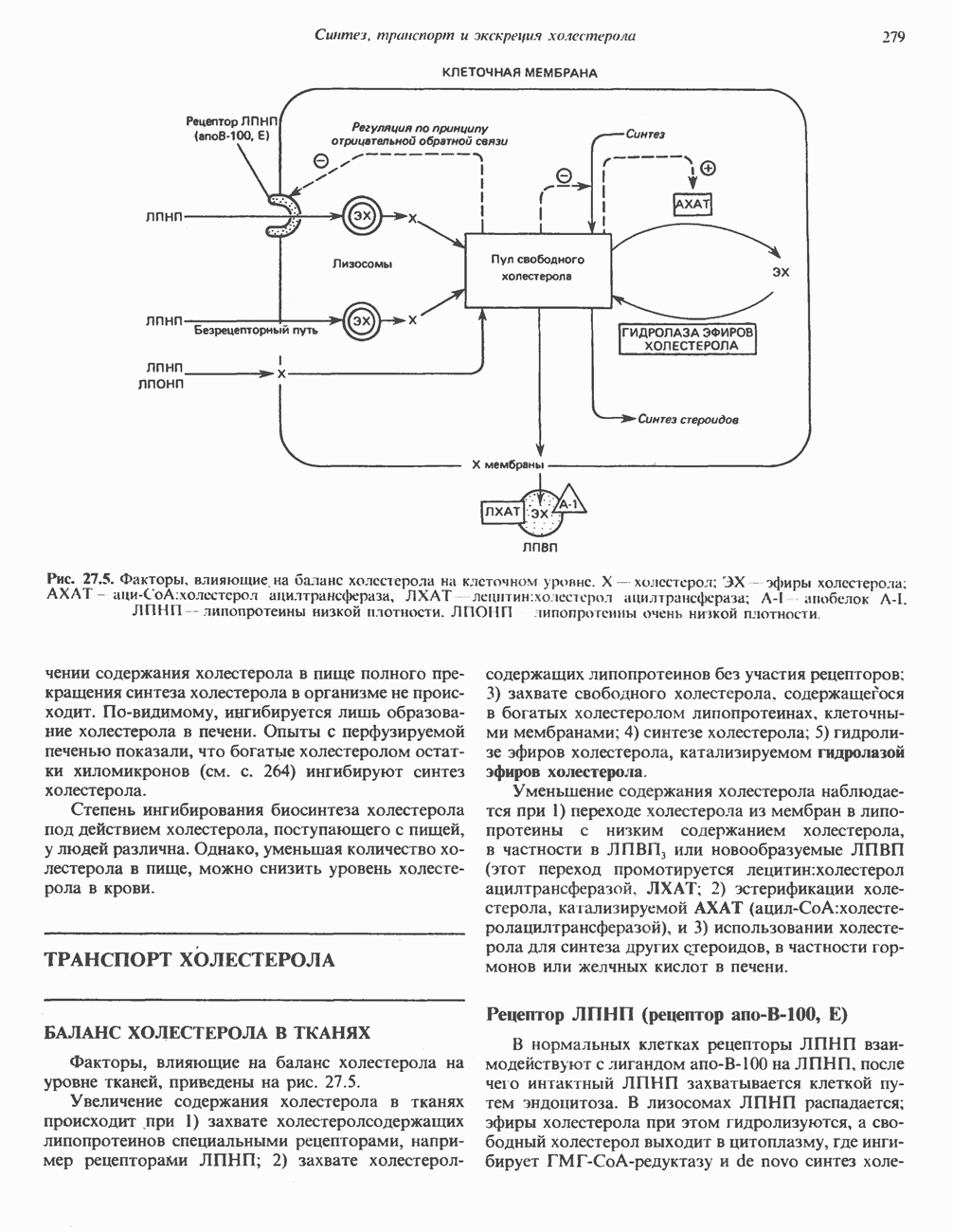
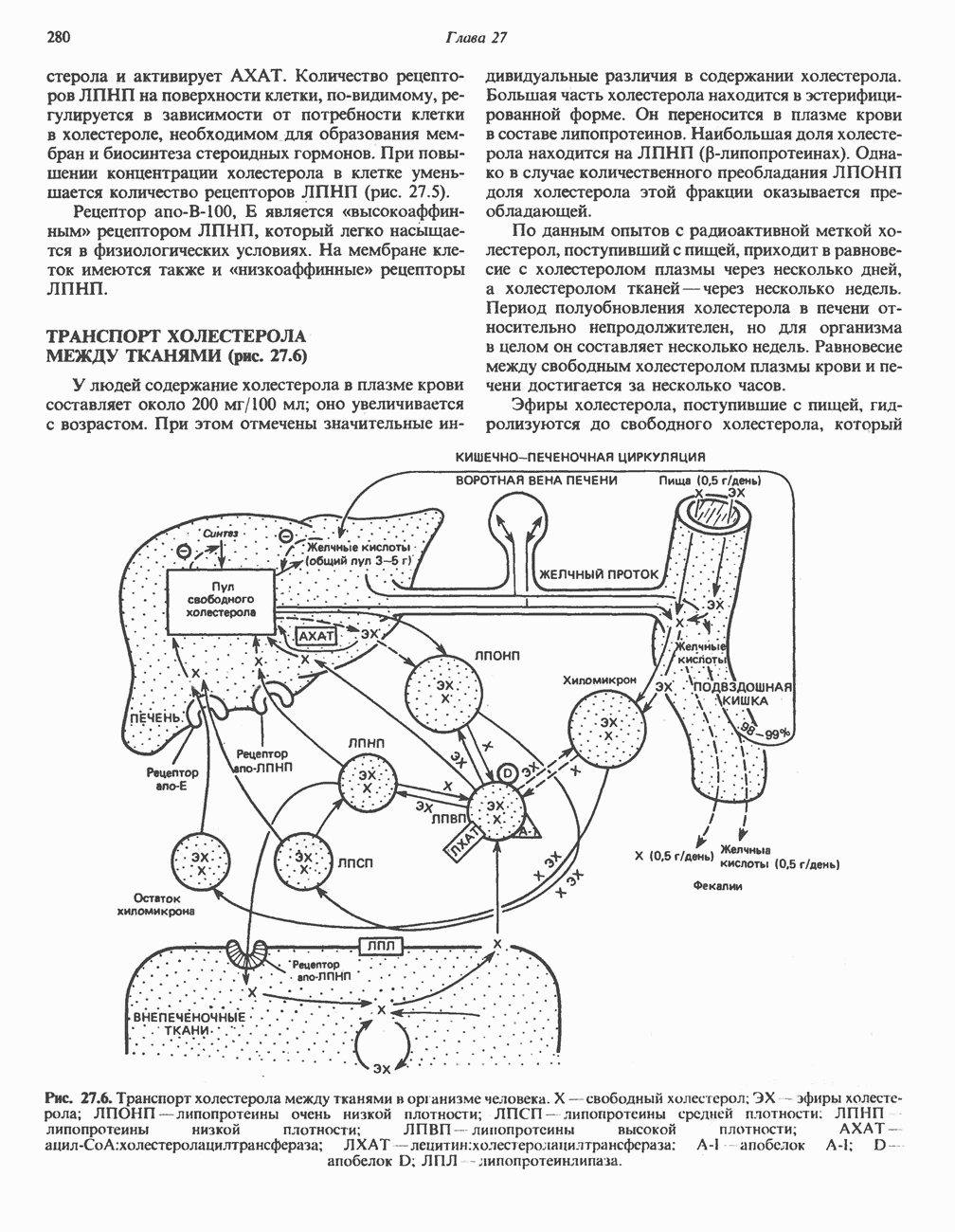
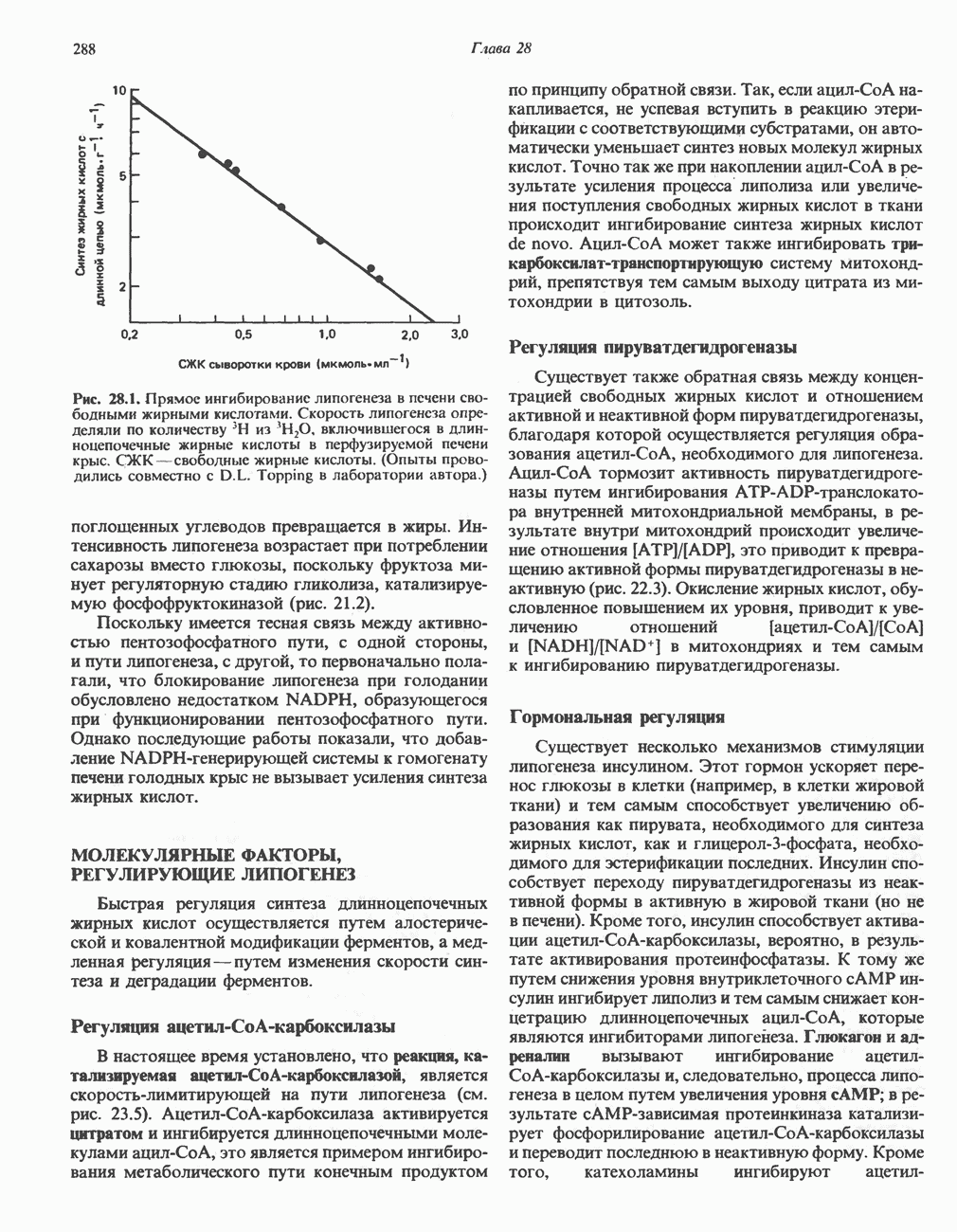
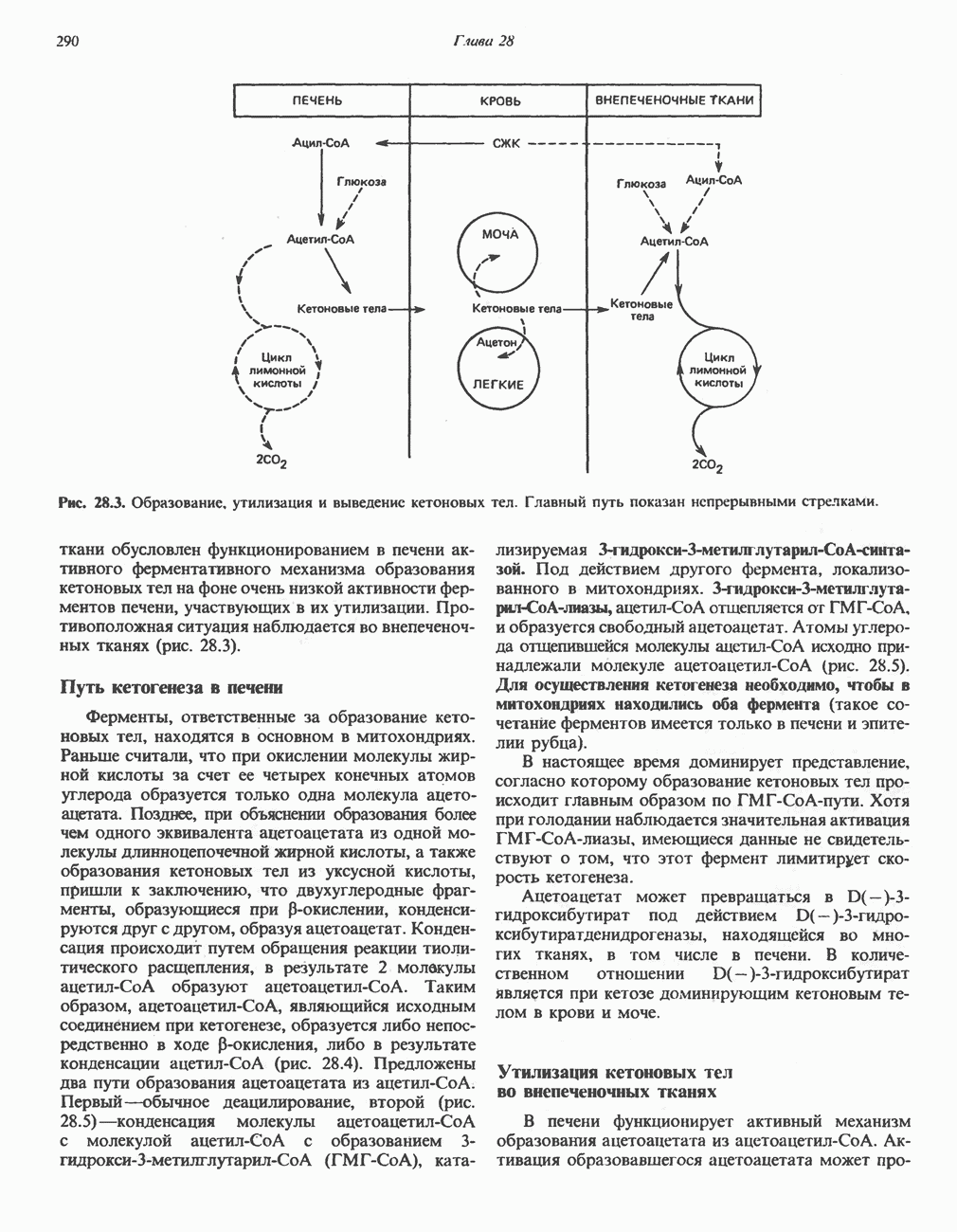
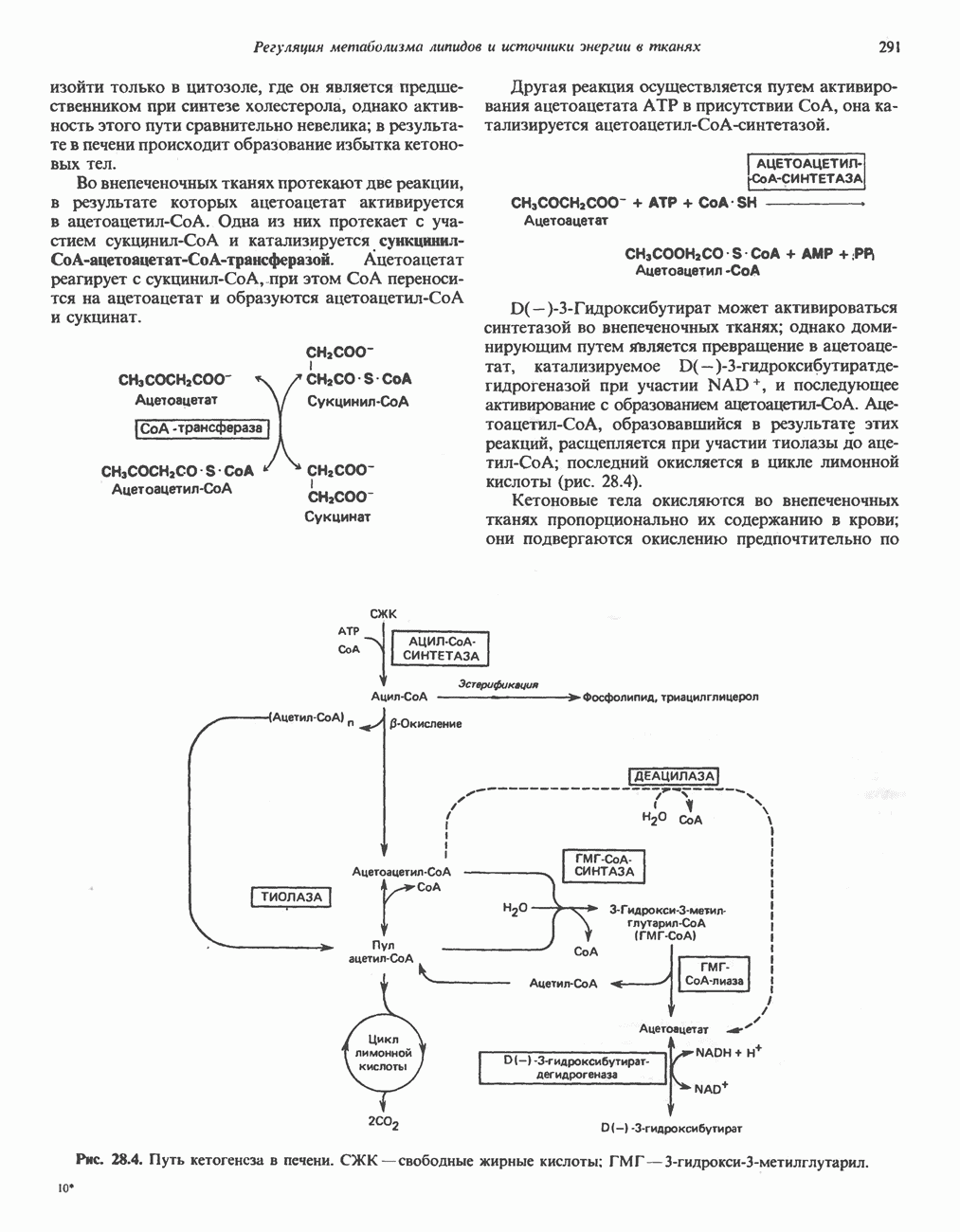
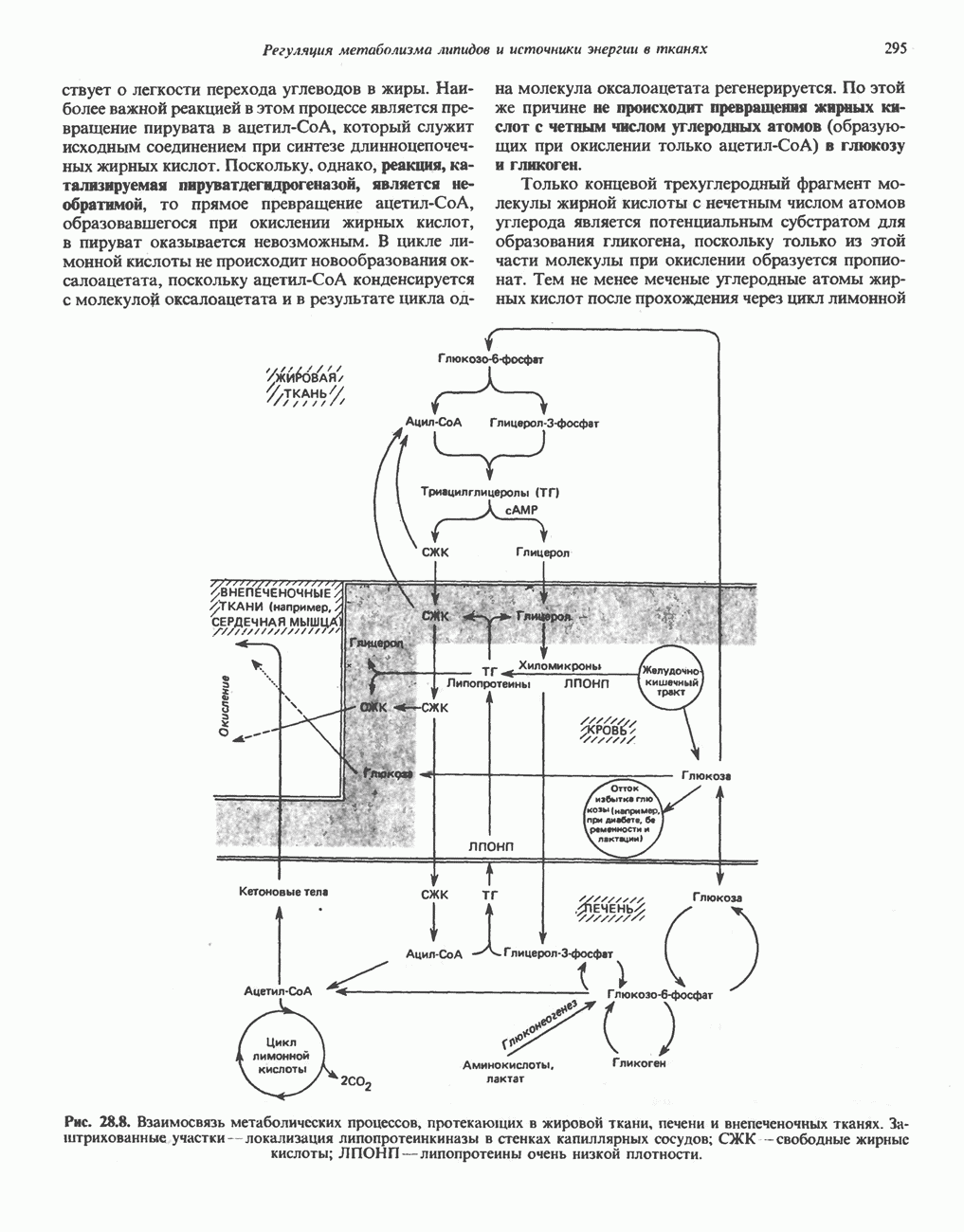
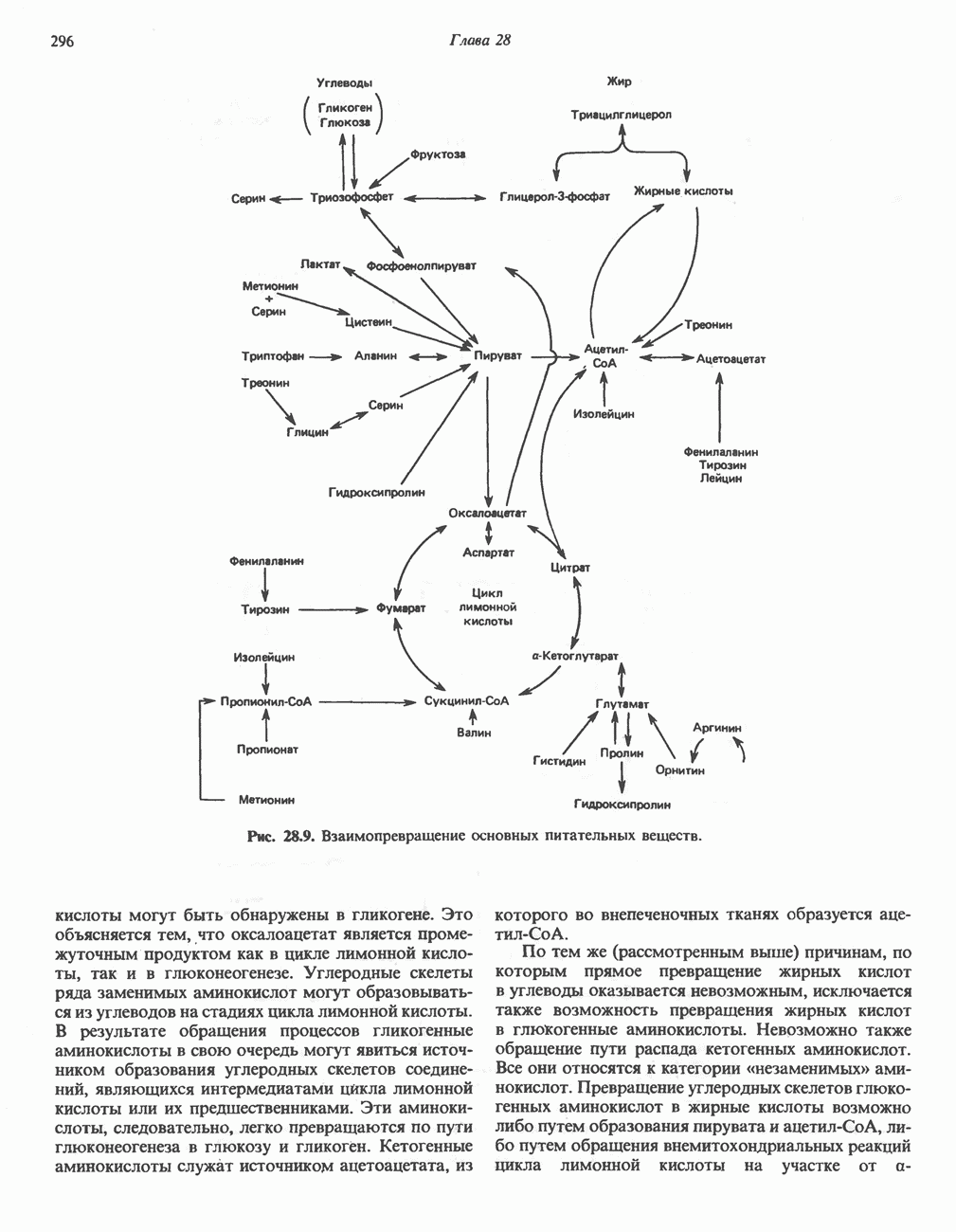
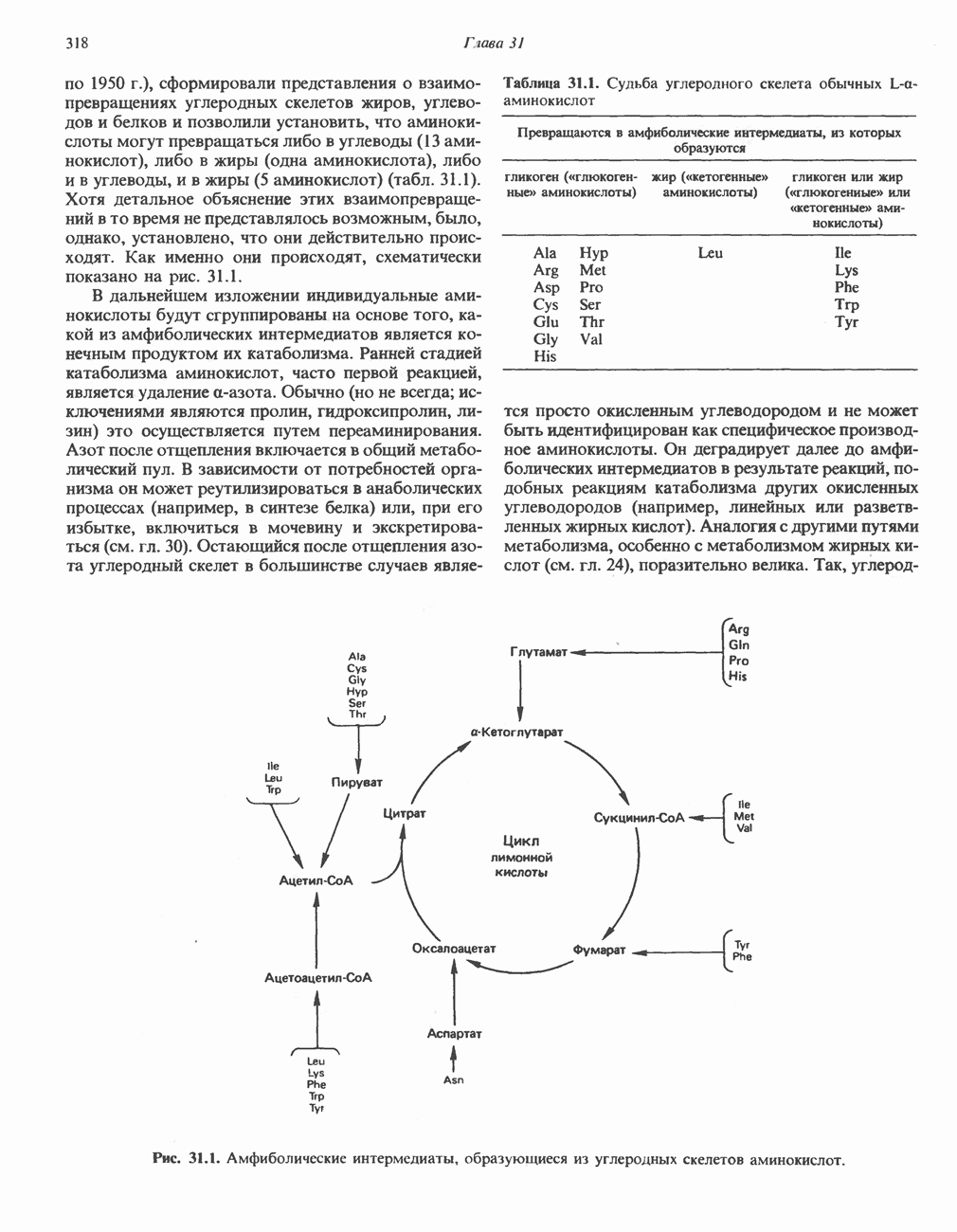
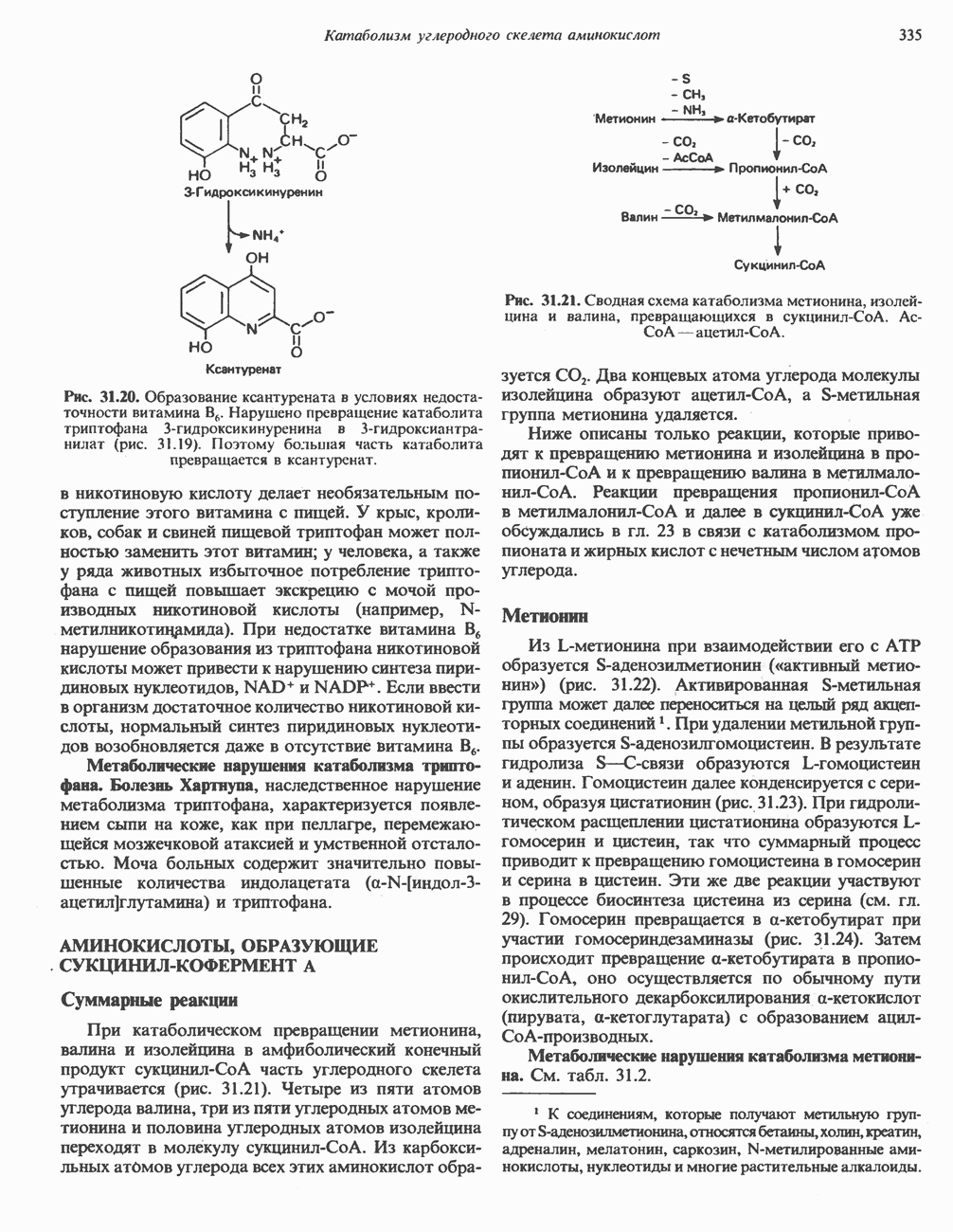
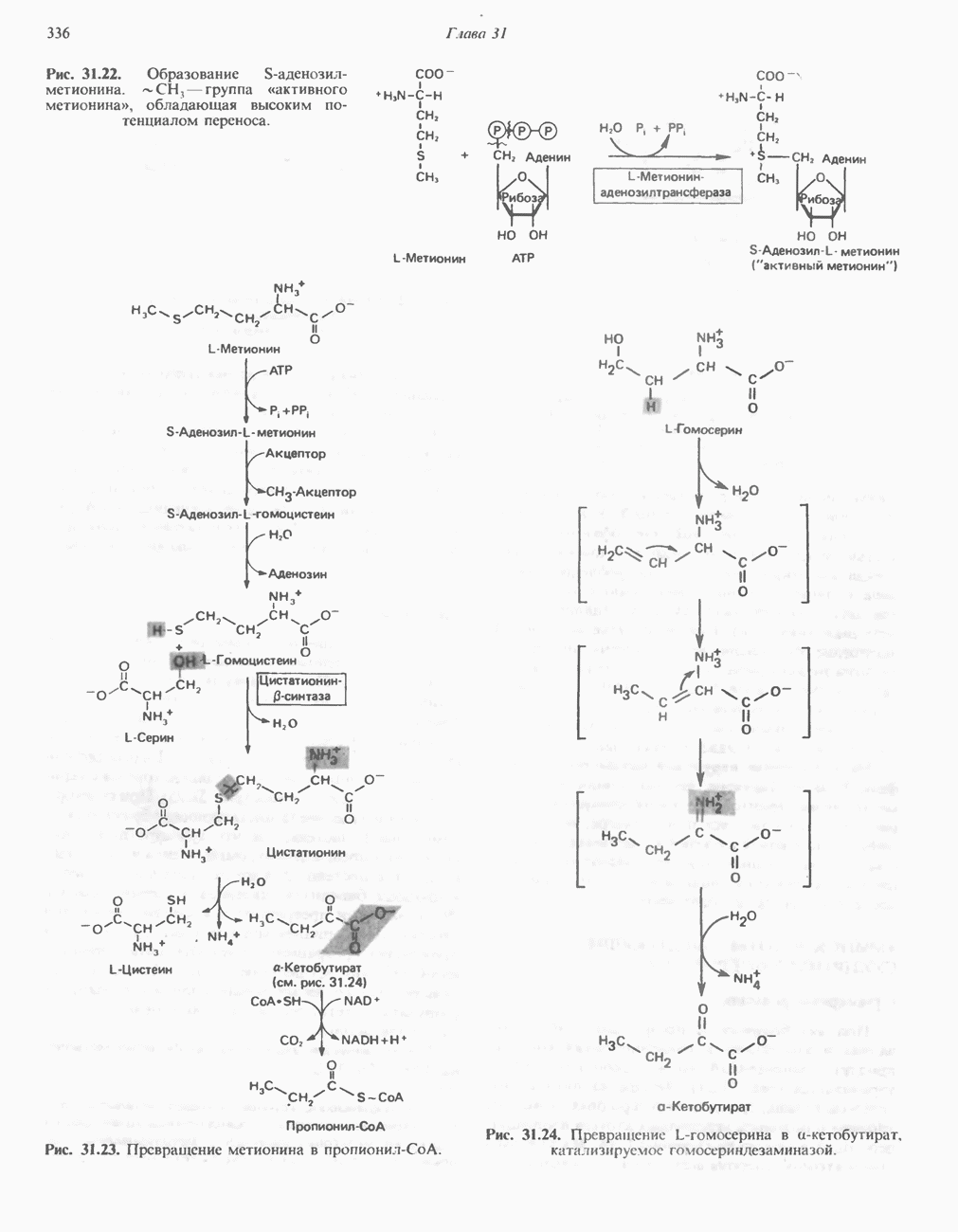
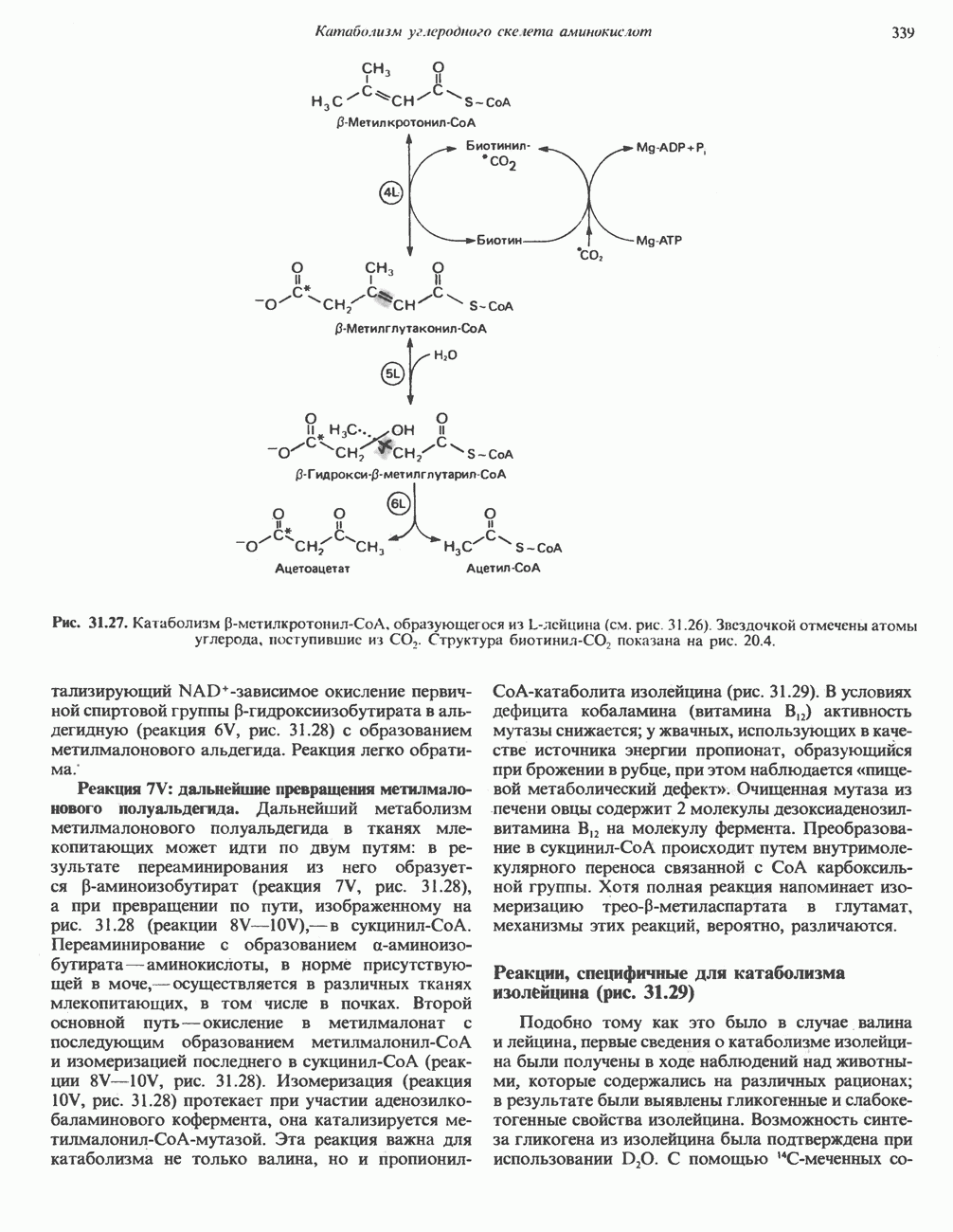
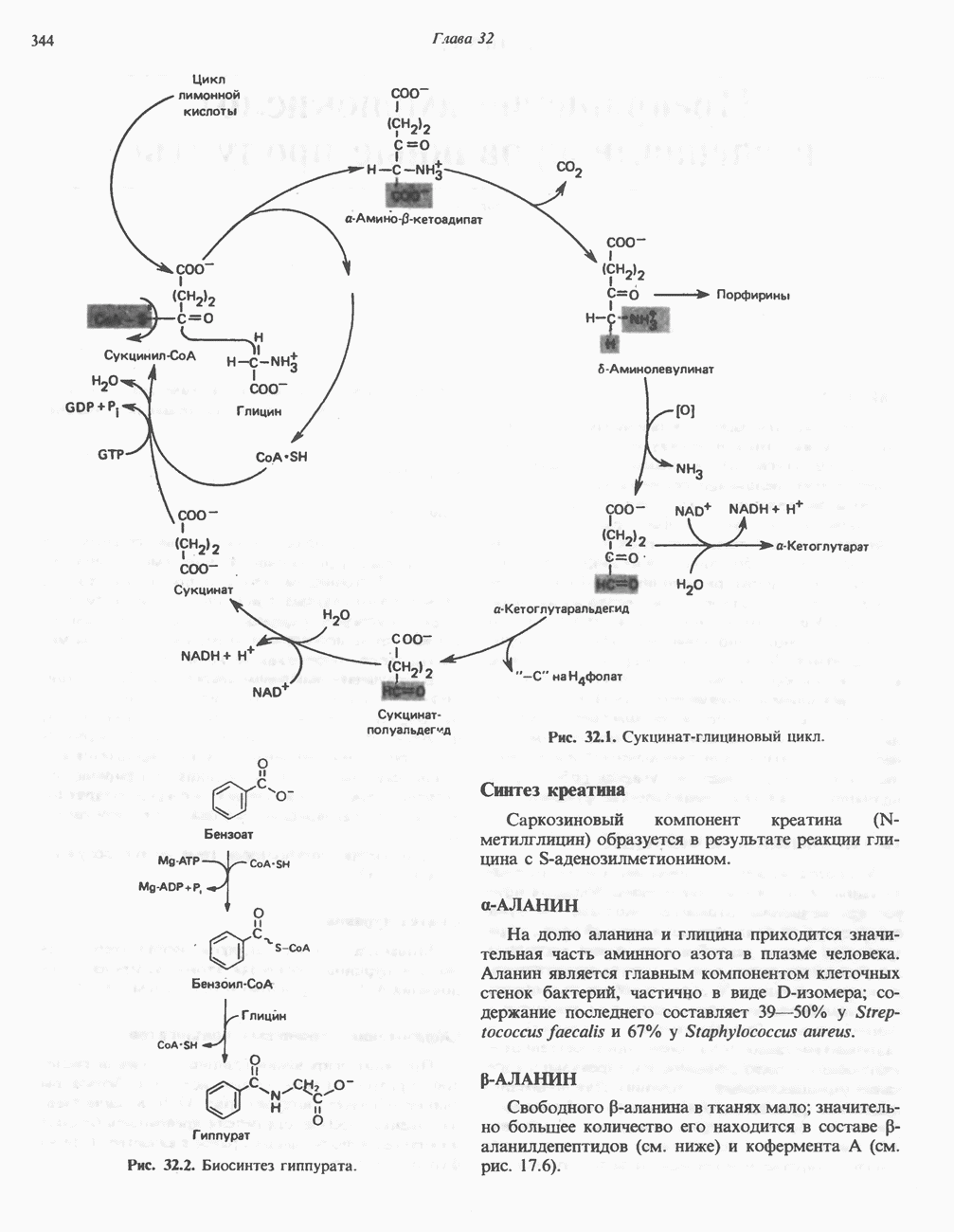
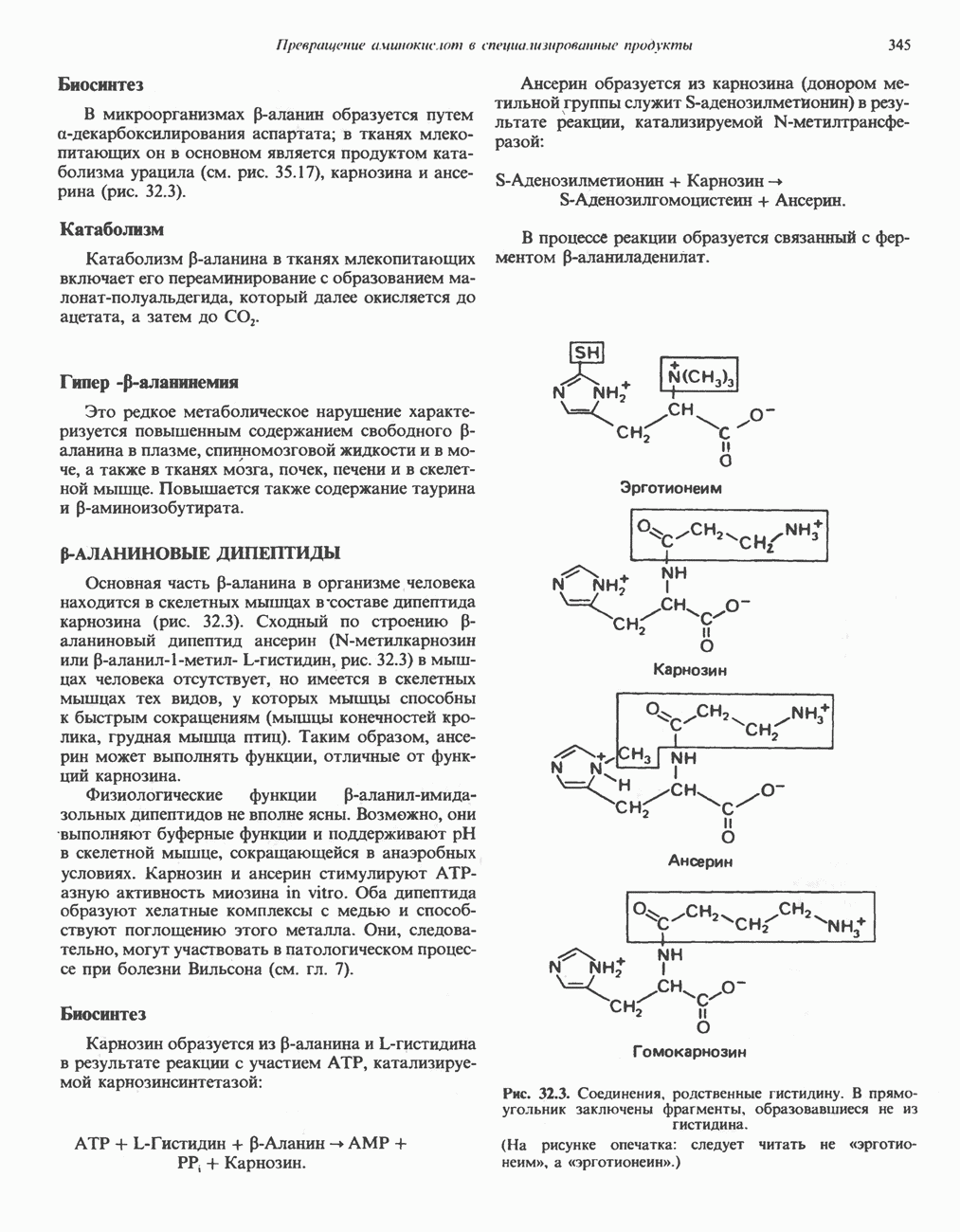
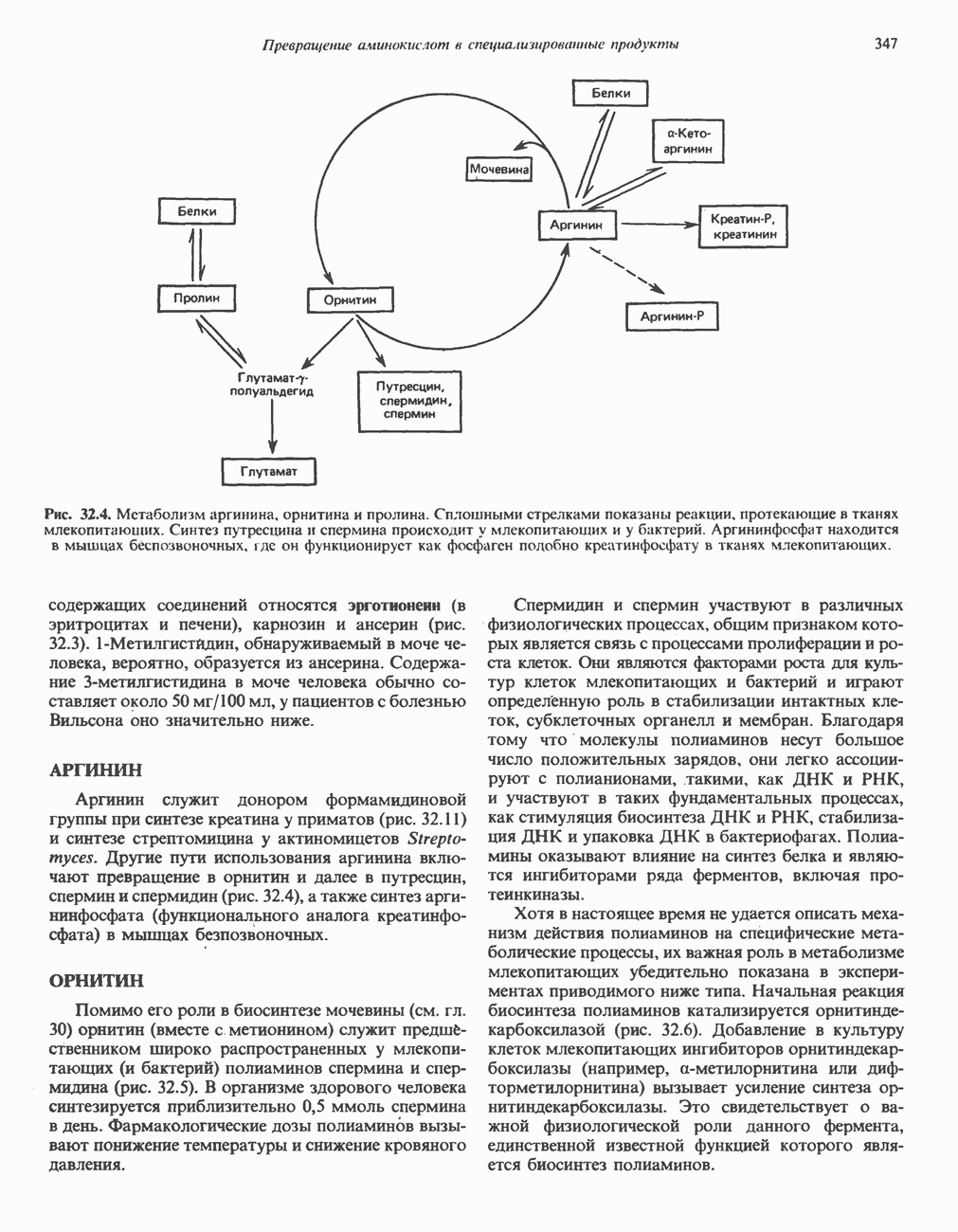
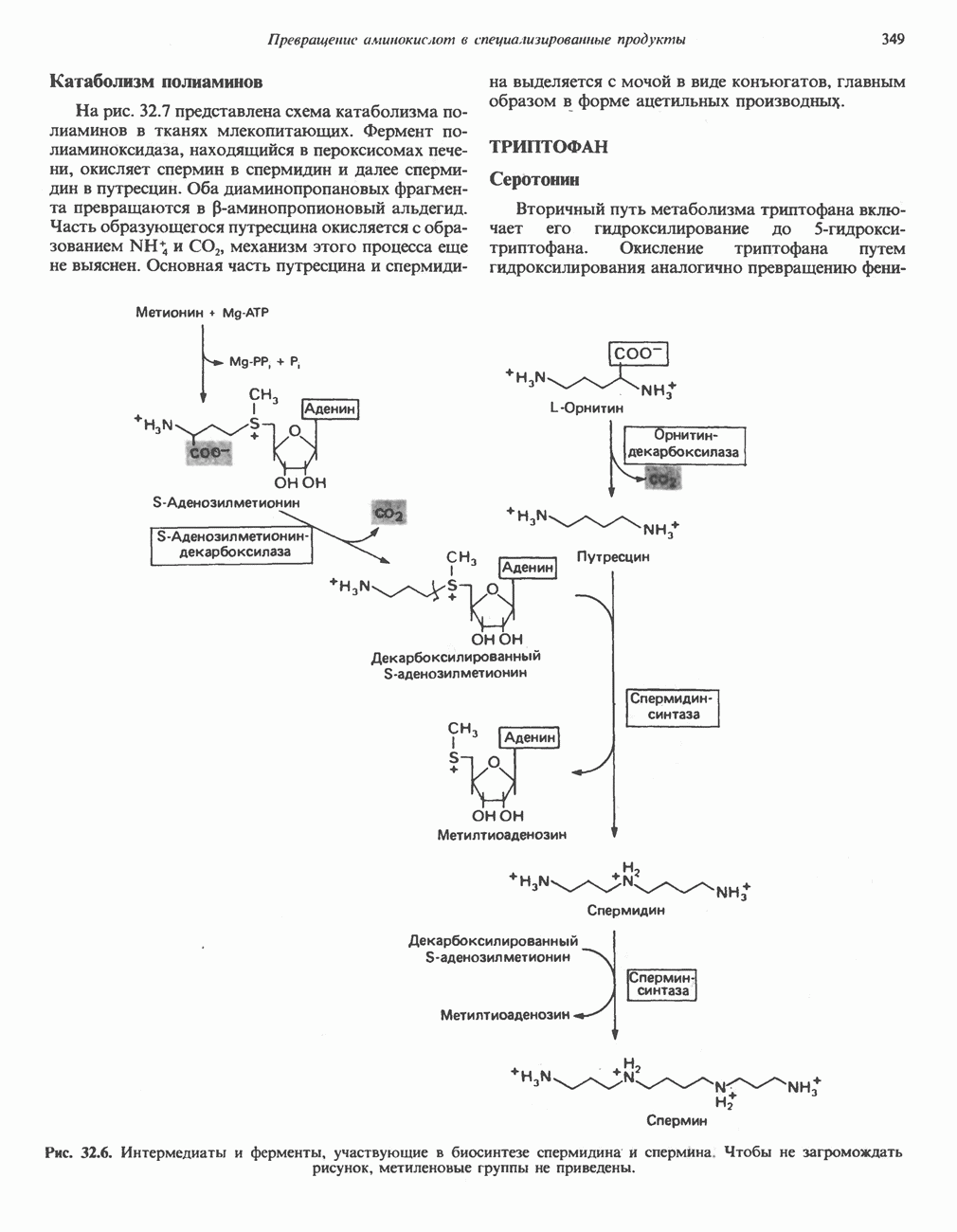
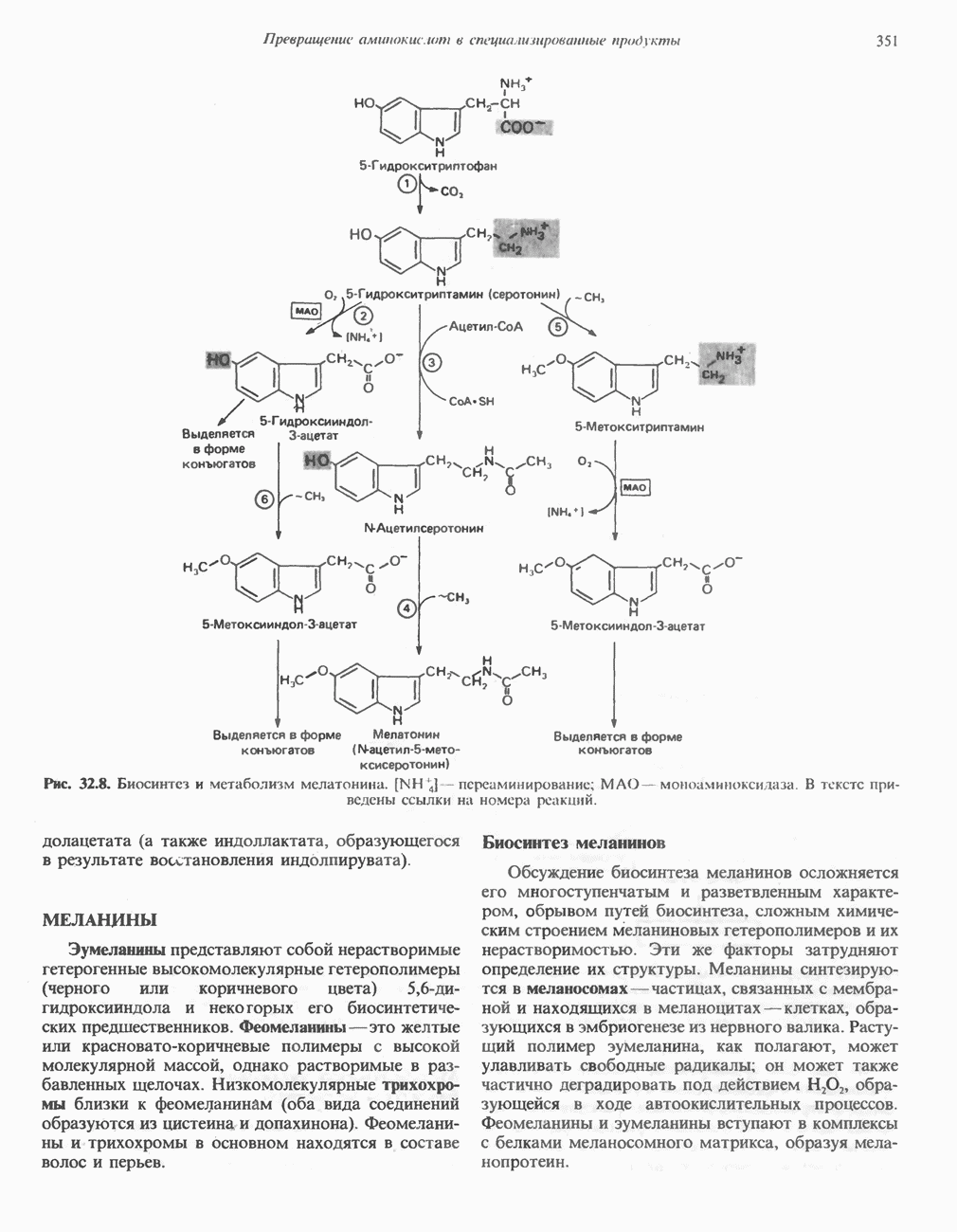
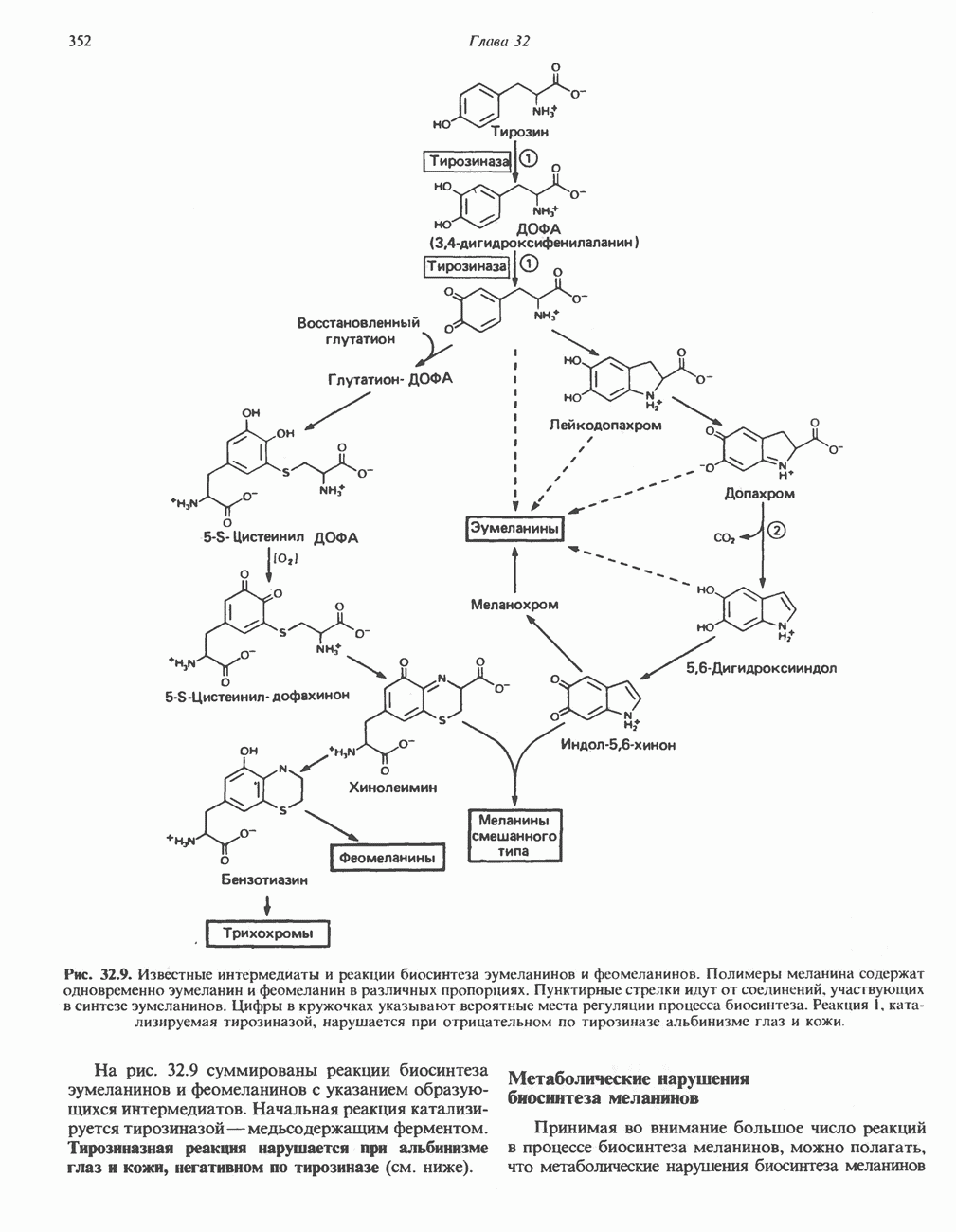
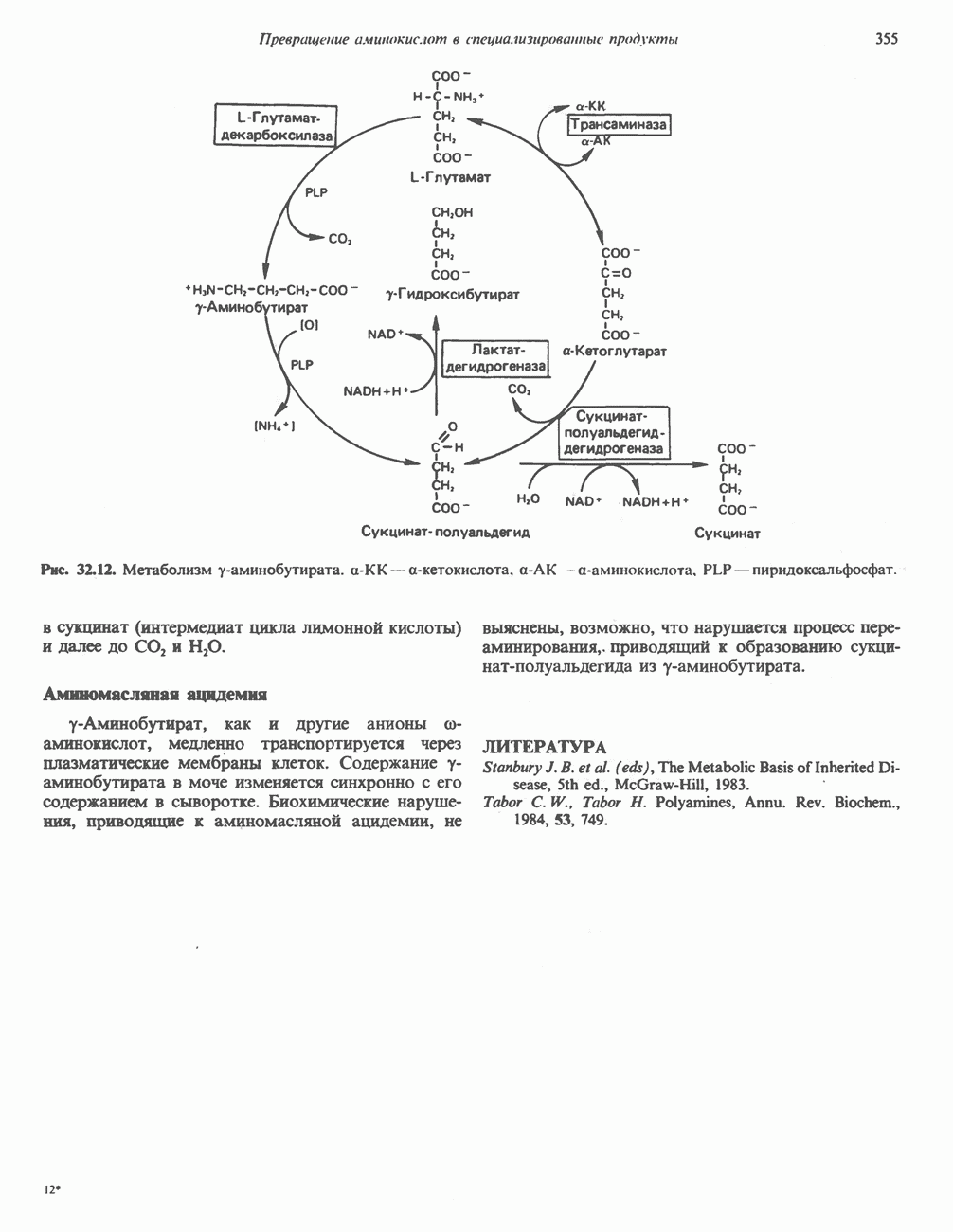
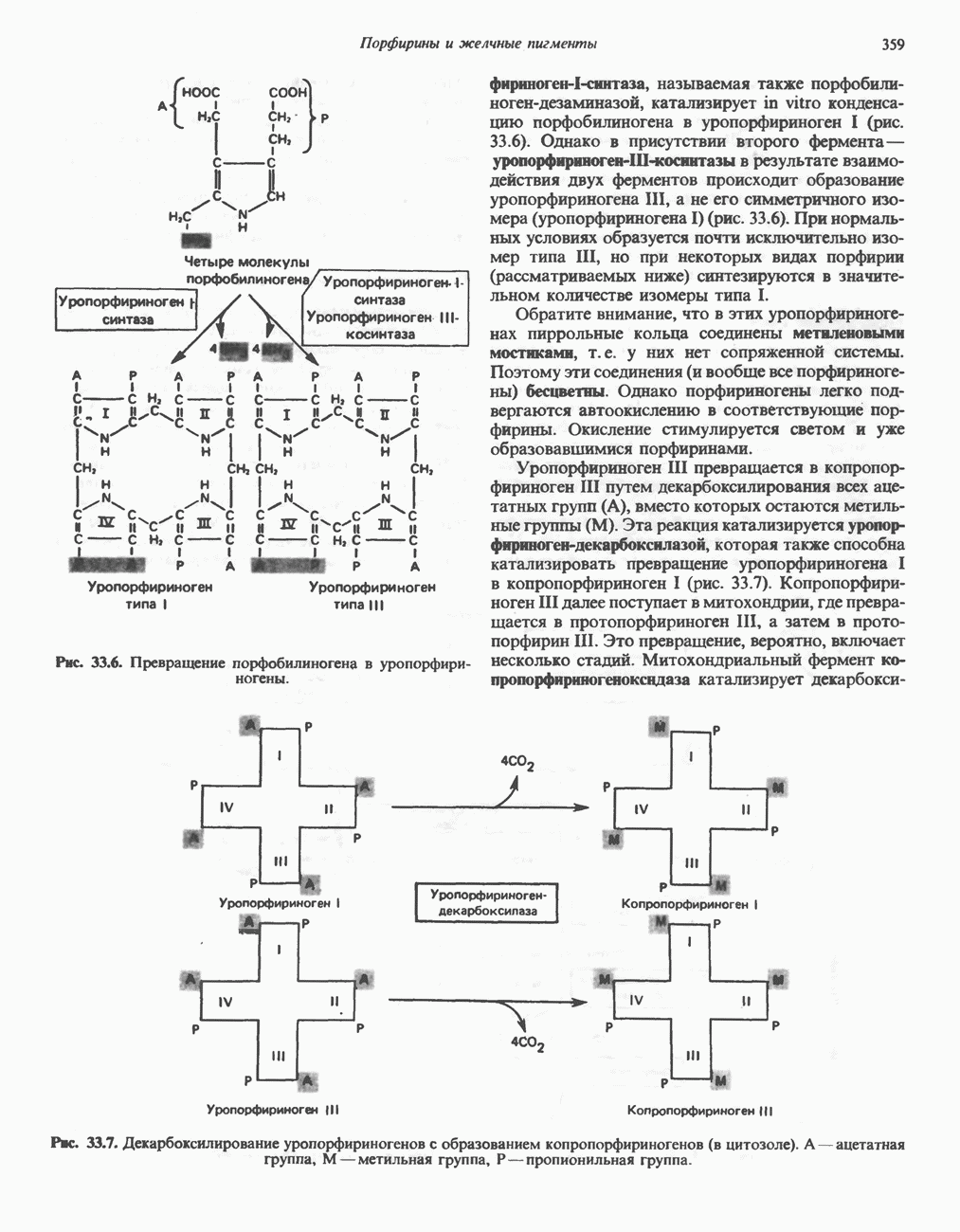
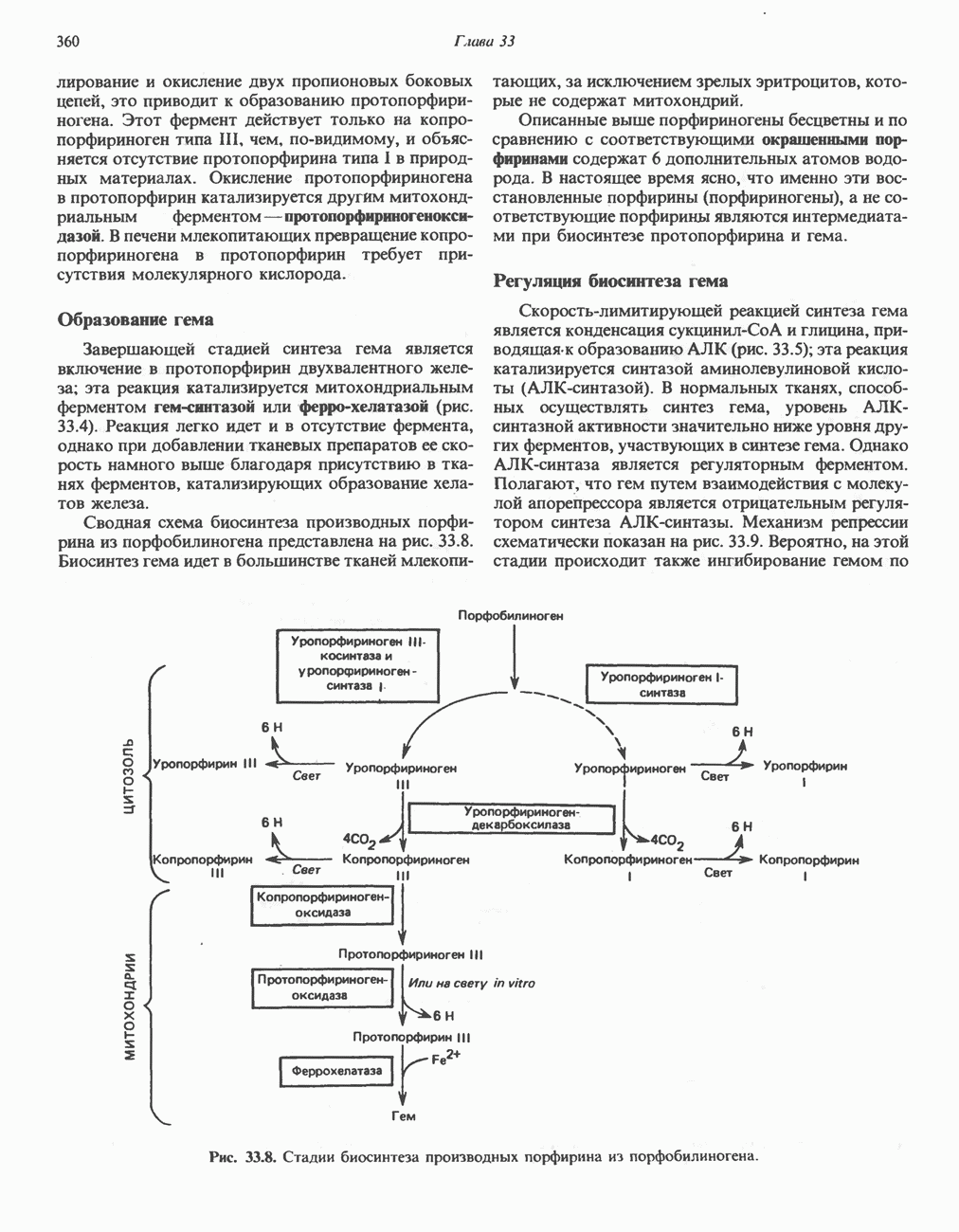
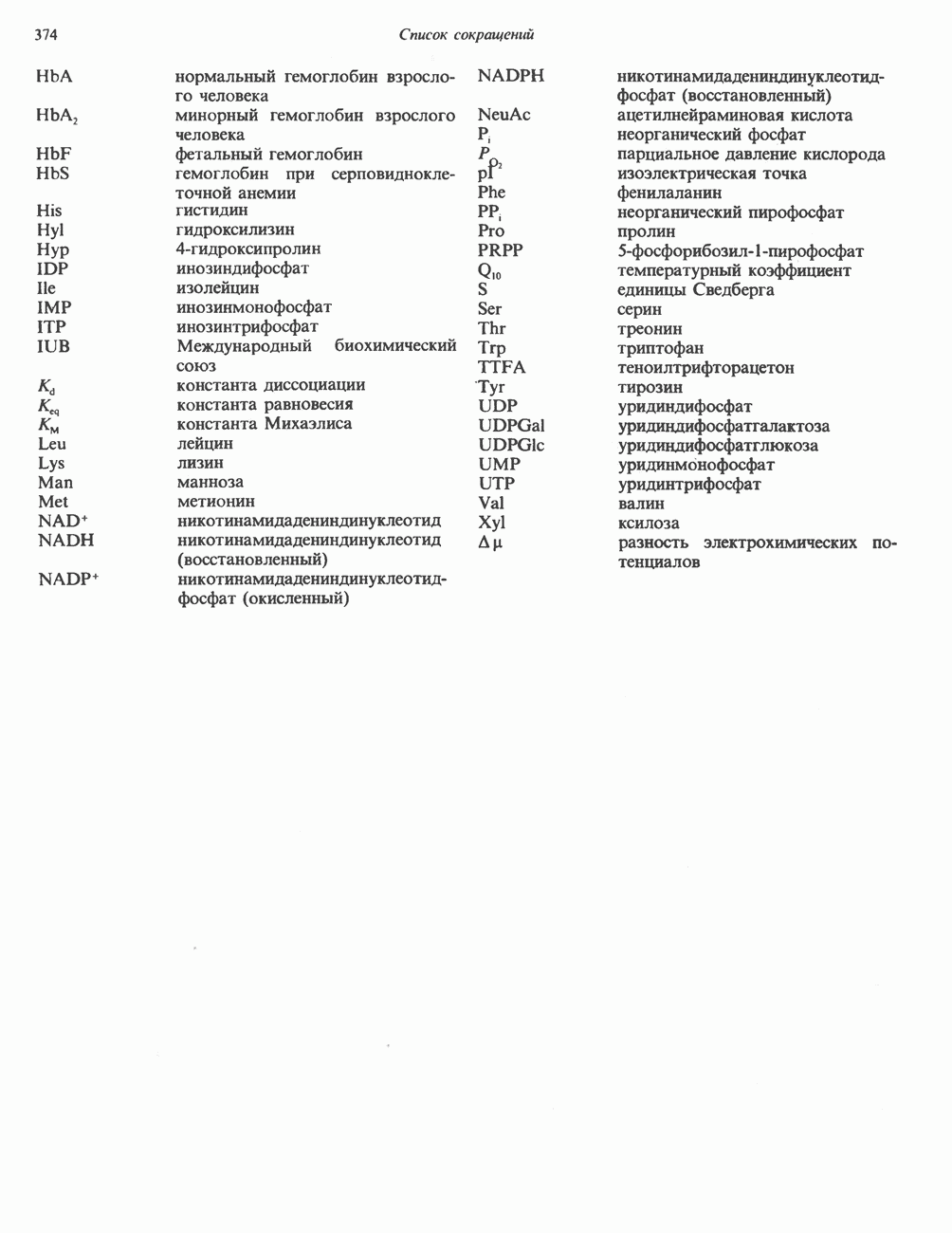
ВЫВЕДЕНИЕ ХОЛЕСТЕРОЛА И ОБРАЗОВАНИЕ СОЛЕЙ ЖЕЛЧНЫХ КИСЛОТ
Из организма человека ежедневно выводится около 1 г холестерола. Приблизительно половина этого количества экскретнруется с фекалиями после превращения в желчные кислоты. Оставшаяся часть выводится в виде нейтральных стероидов. Большая часть холестерола, поступившего в желчь, реабсорбируется; считается, что по крайней мере часть холестерола, являющегося предшественником фекальных стеролов, поступает из слизистой оболочки кишечника. Основным фекальным стеролом является копростанол, который образуется из холестерола в нижнем отделе кишечника под действием присутствующей в нем микрофлоры. Значительная доля солей желчных кислот, поступающих с желчью, всасывается в кишечнике и через воротную вену возвращается в печень, где снова поступает в желчь. Этот путь транспорта солей желчных кислот получил название кишечно-печеночной циркуляции. Оставшаяся часть солей желчных кислот, а также их производные выводятся с фекалиями. Под действием кишечных бактерий первичные желчные кислоты превращаются во вторичные.
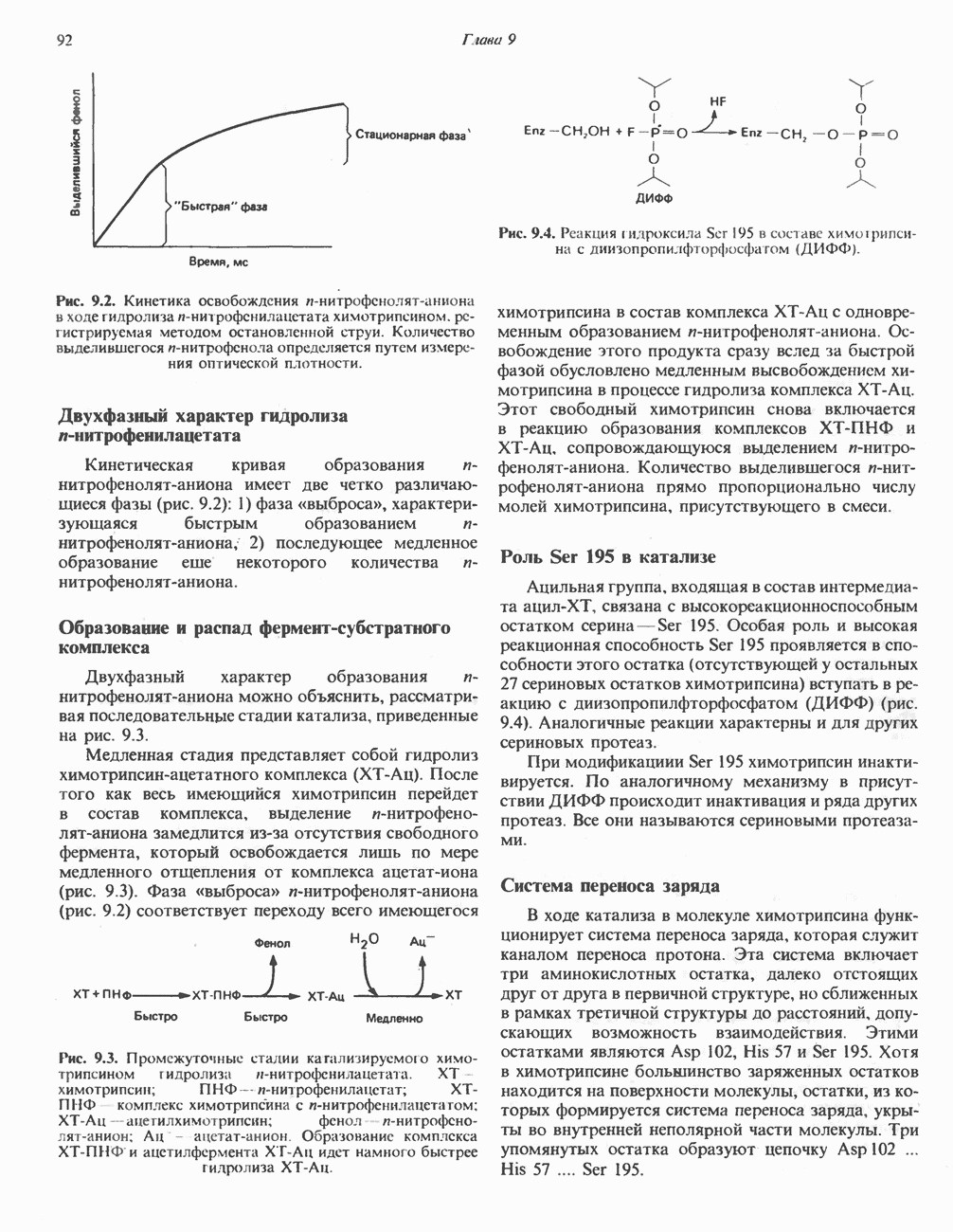
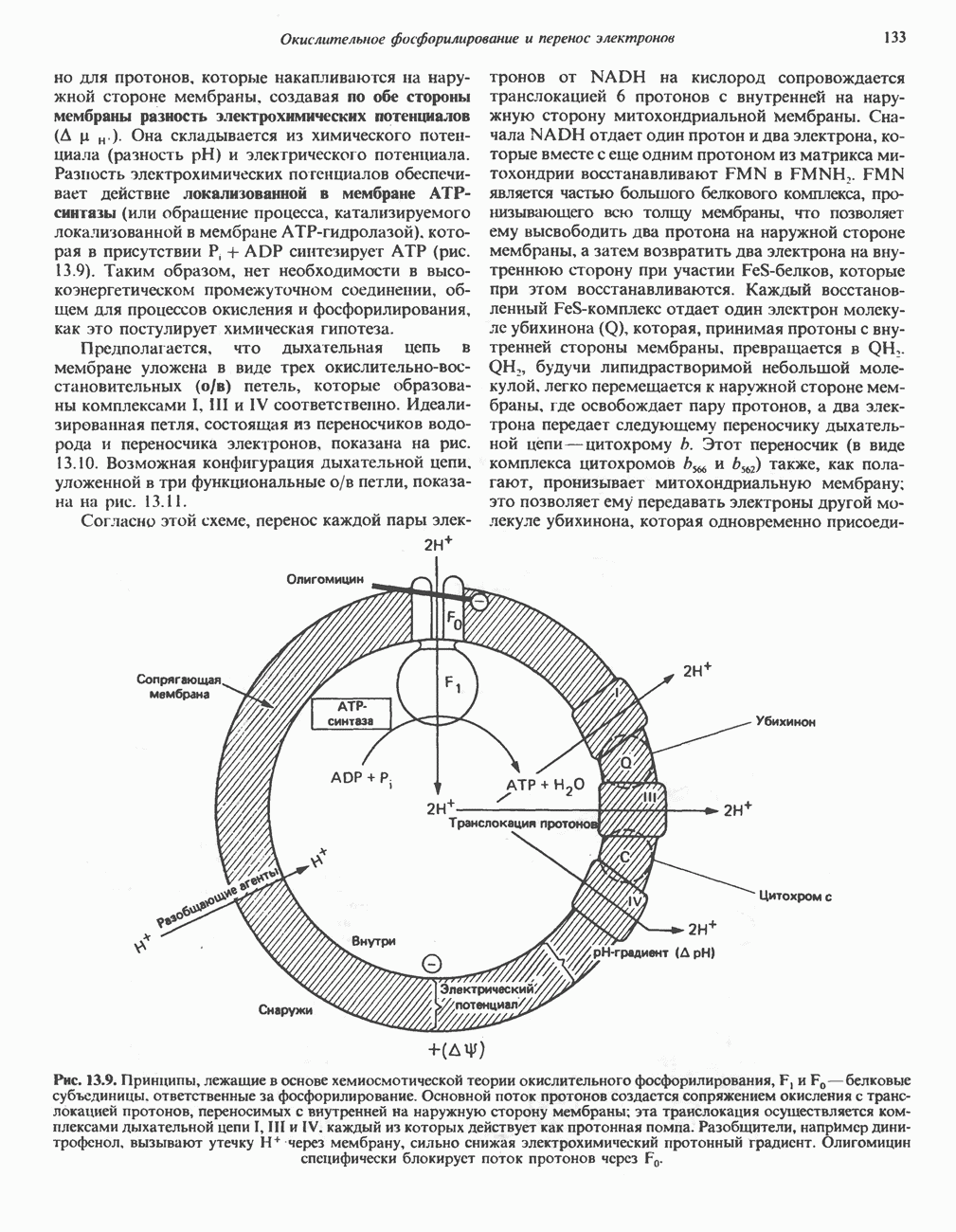
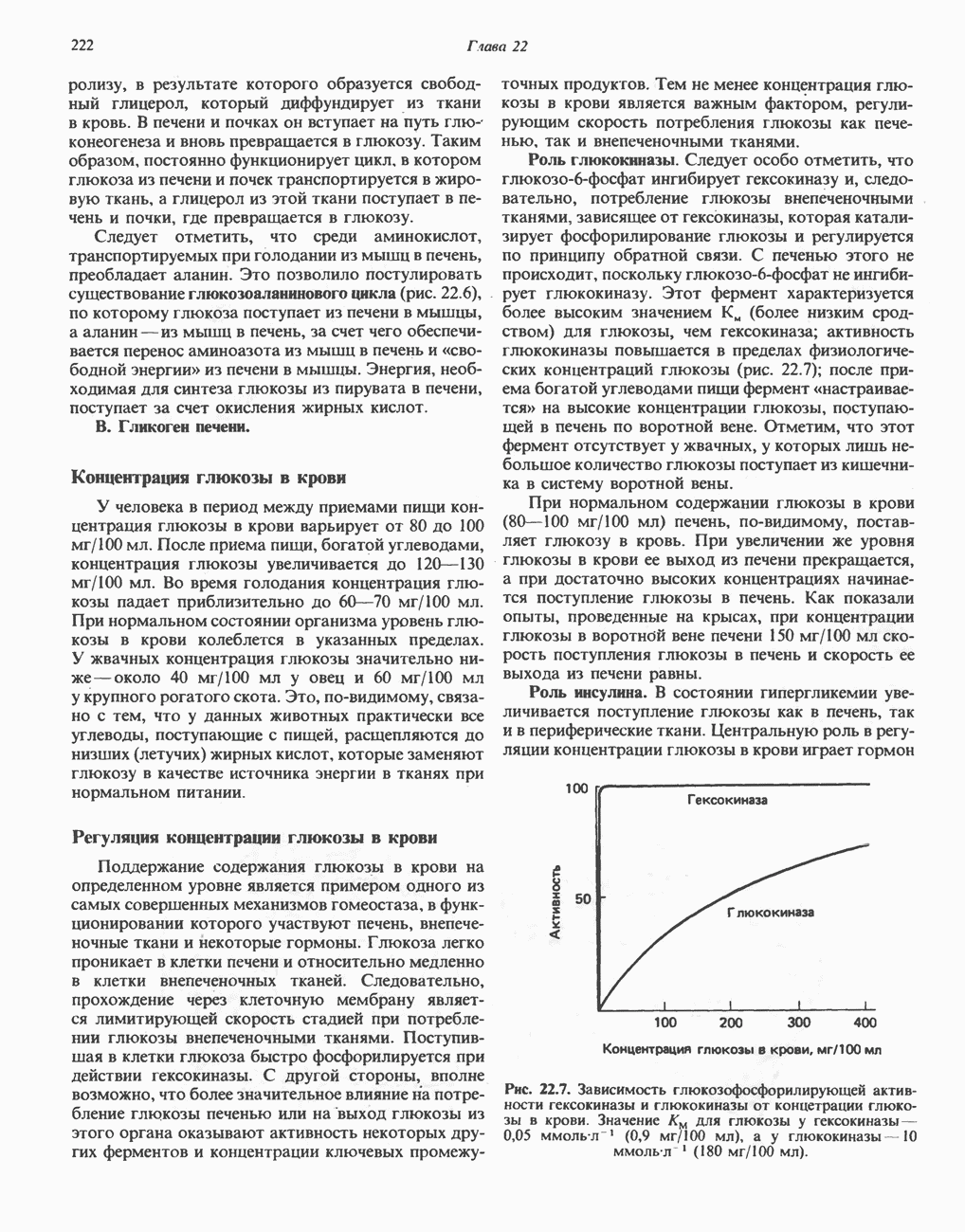
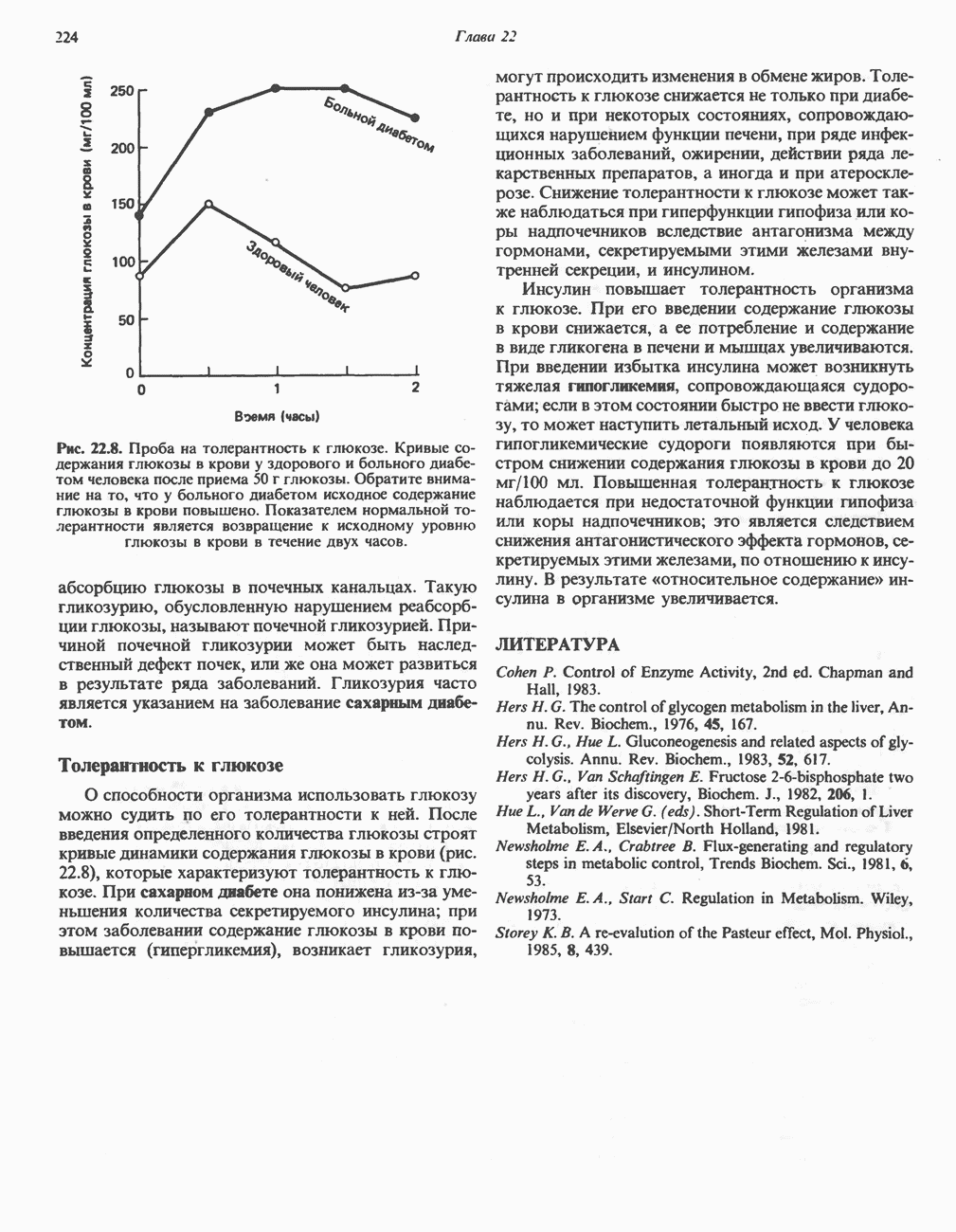
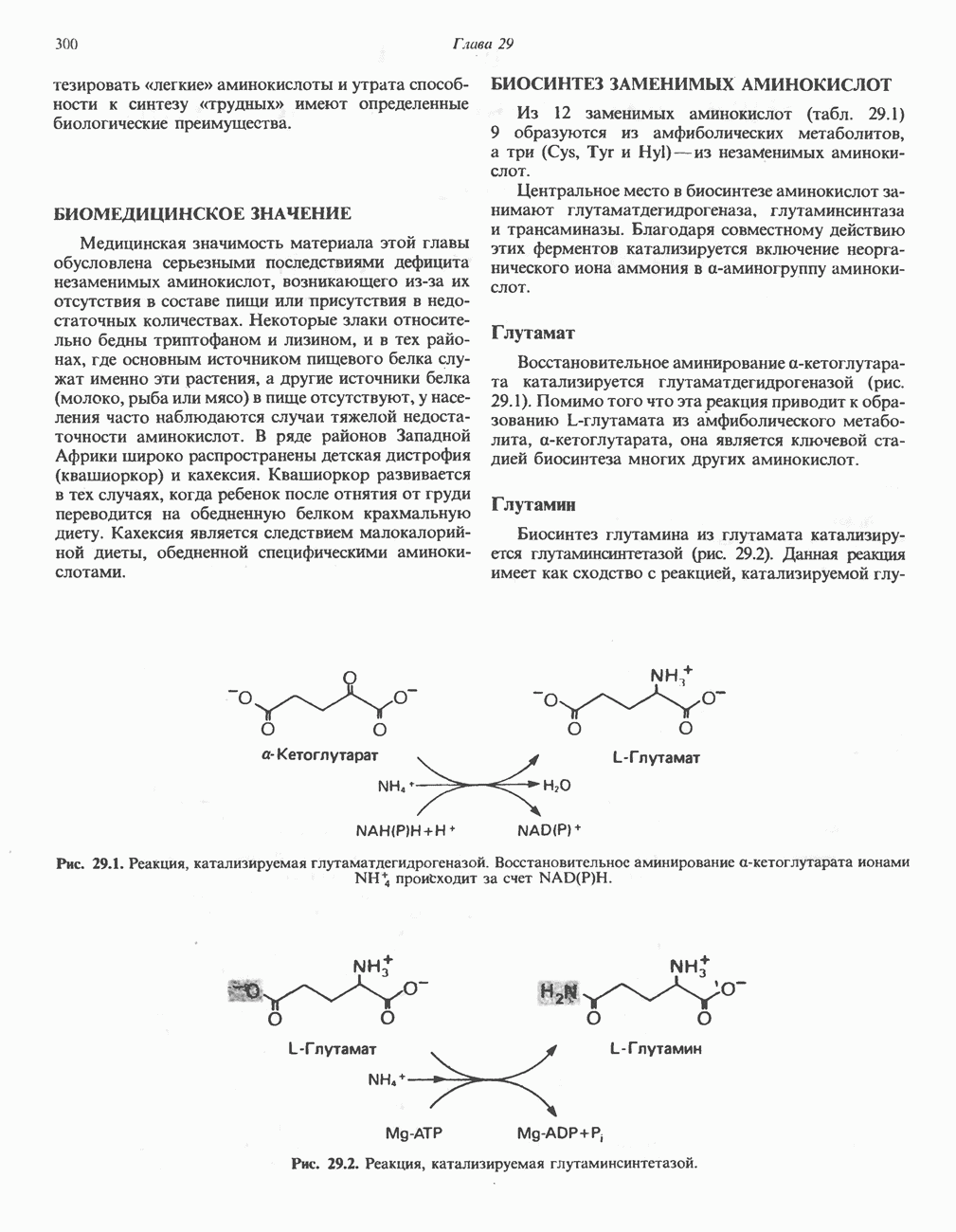
БИОСИНТЕЗ ЖЕЛЧНЫХ КИСЛОТ
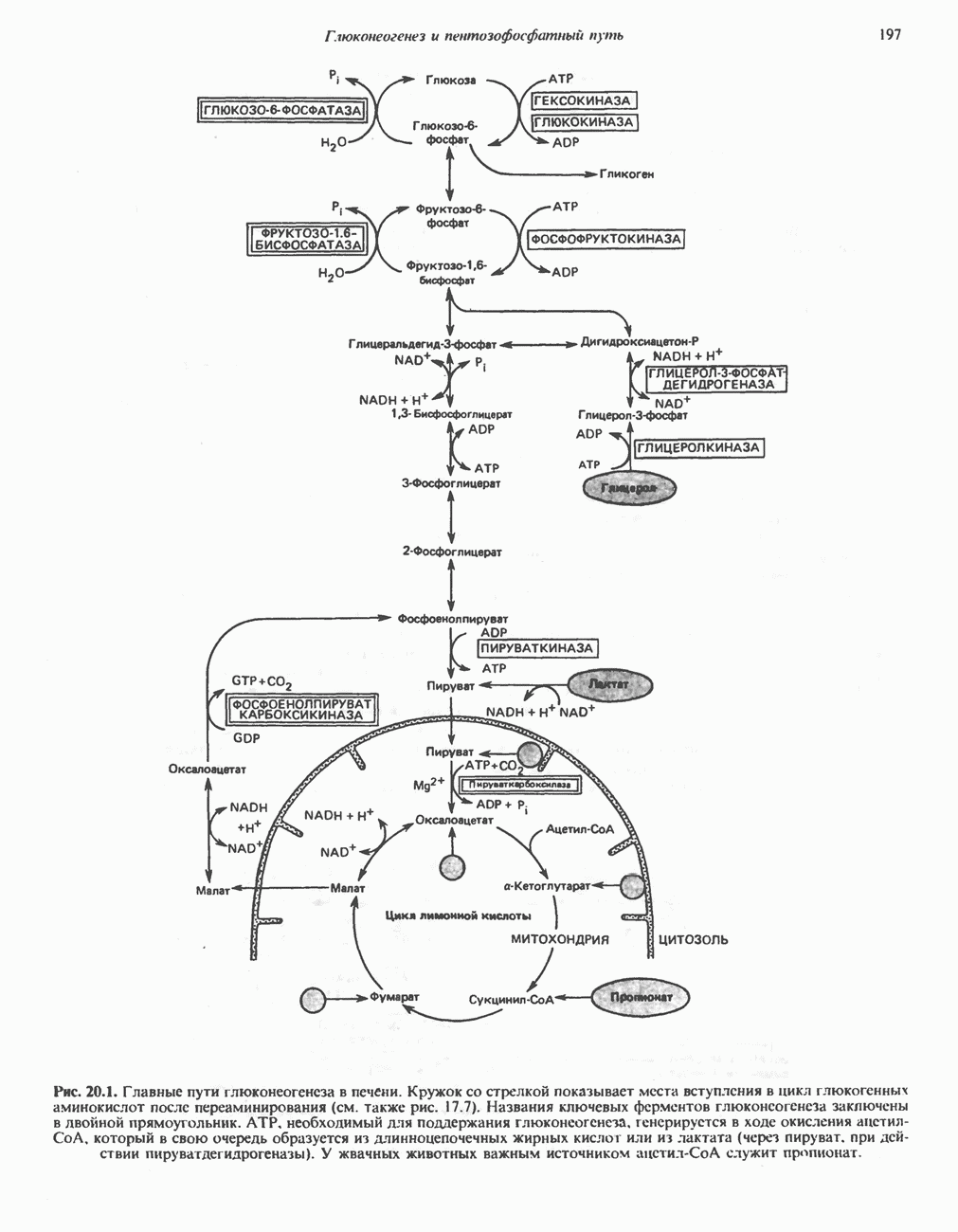
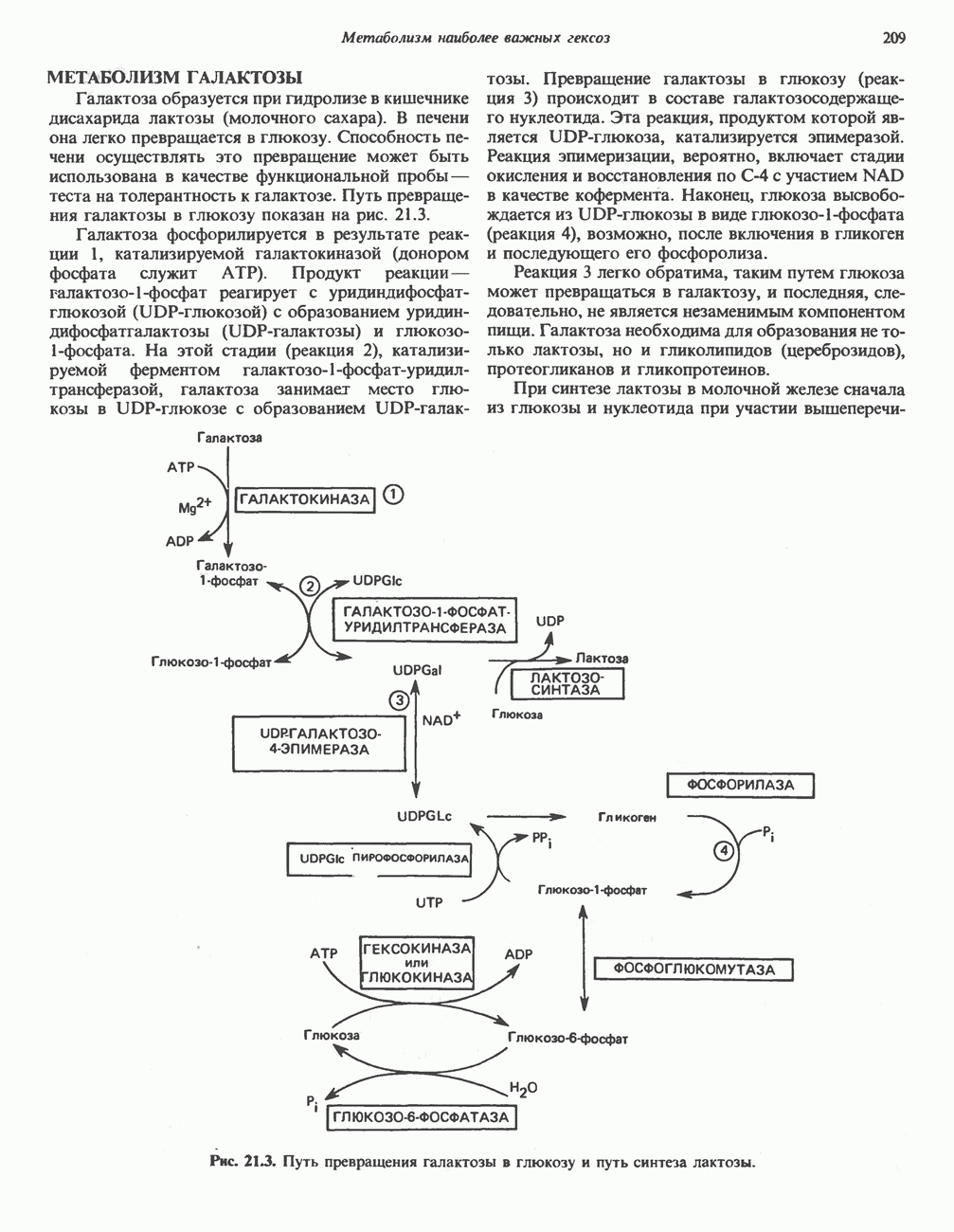
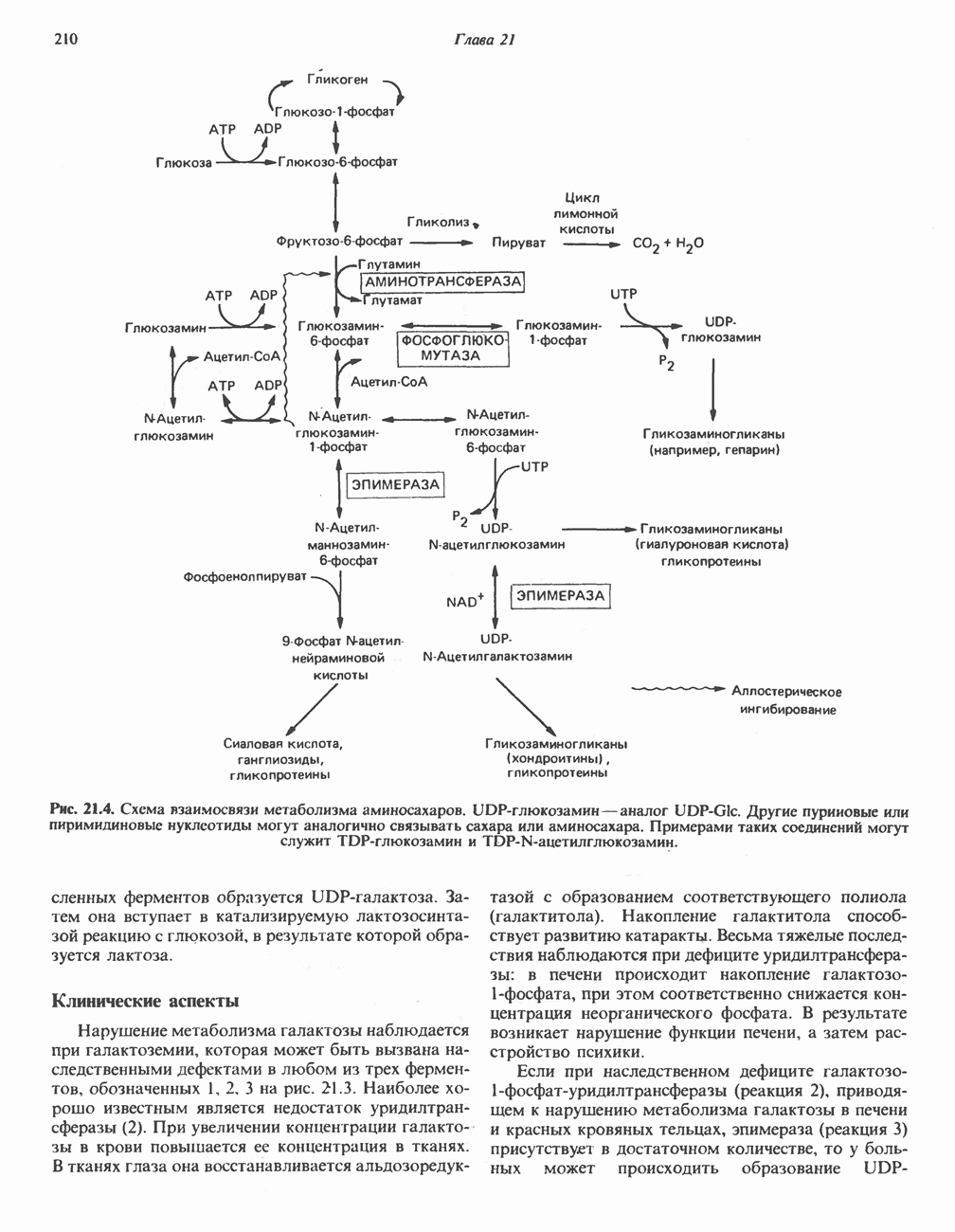
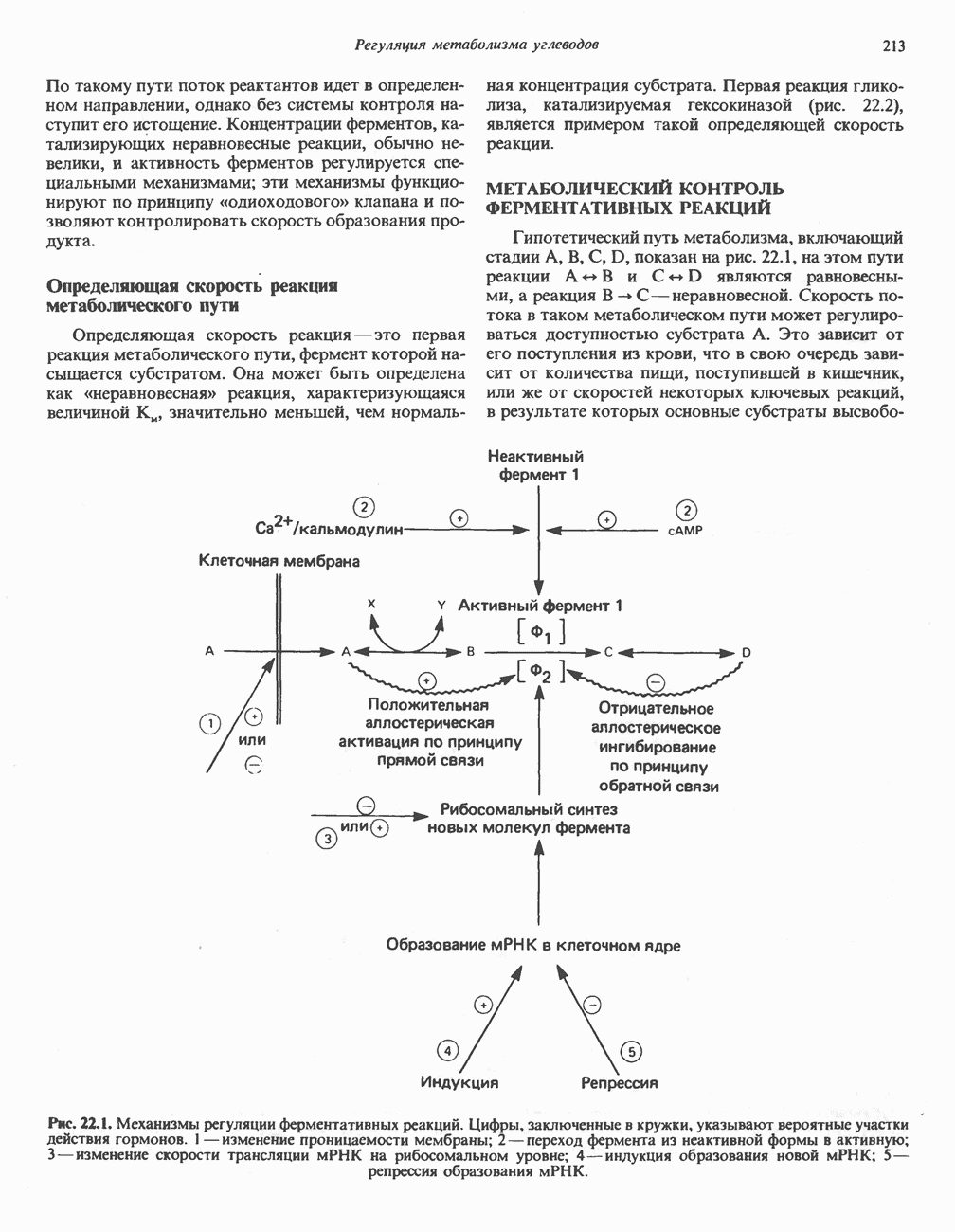
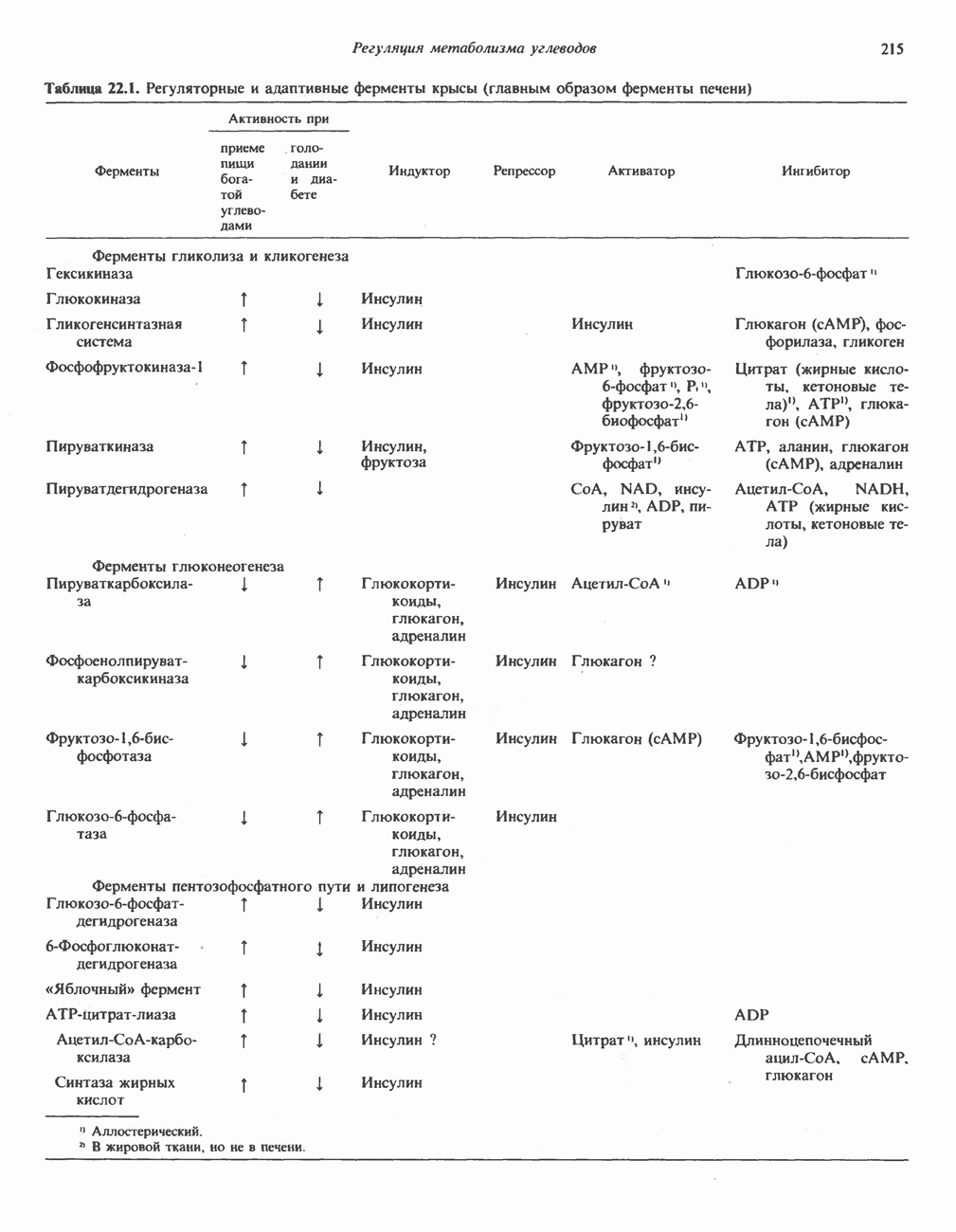
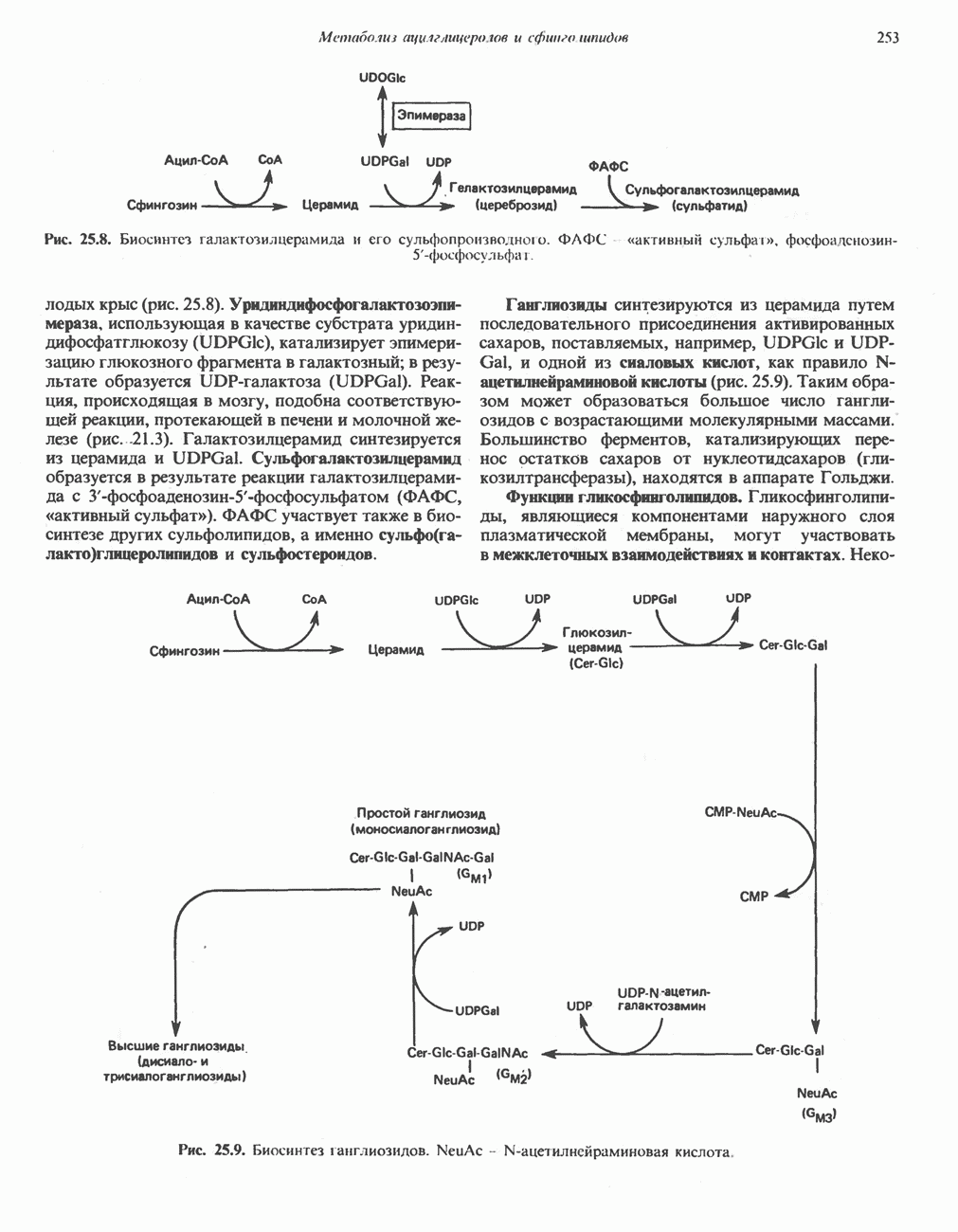
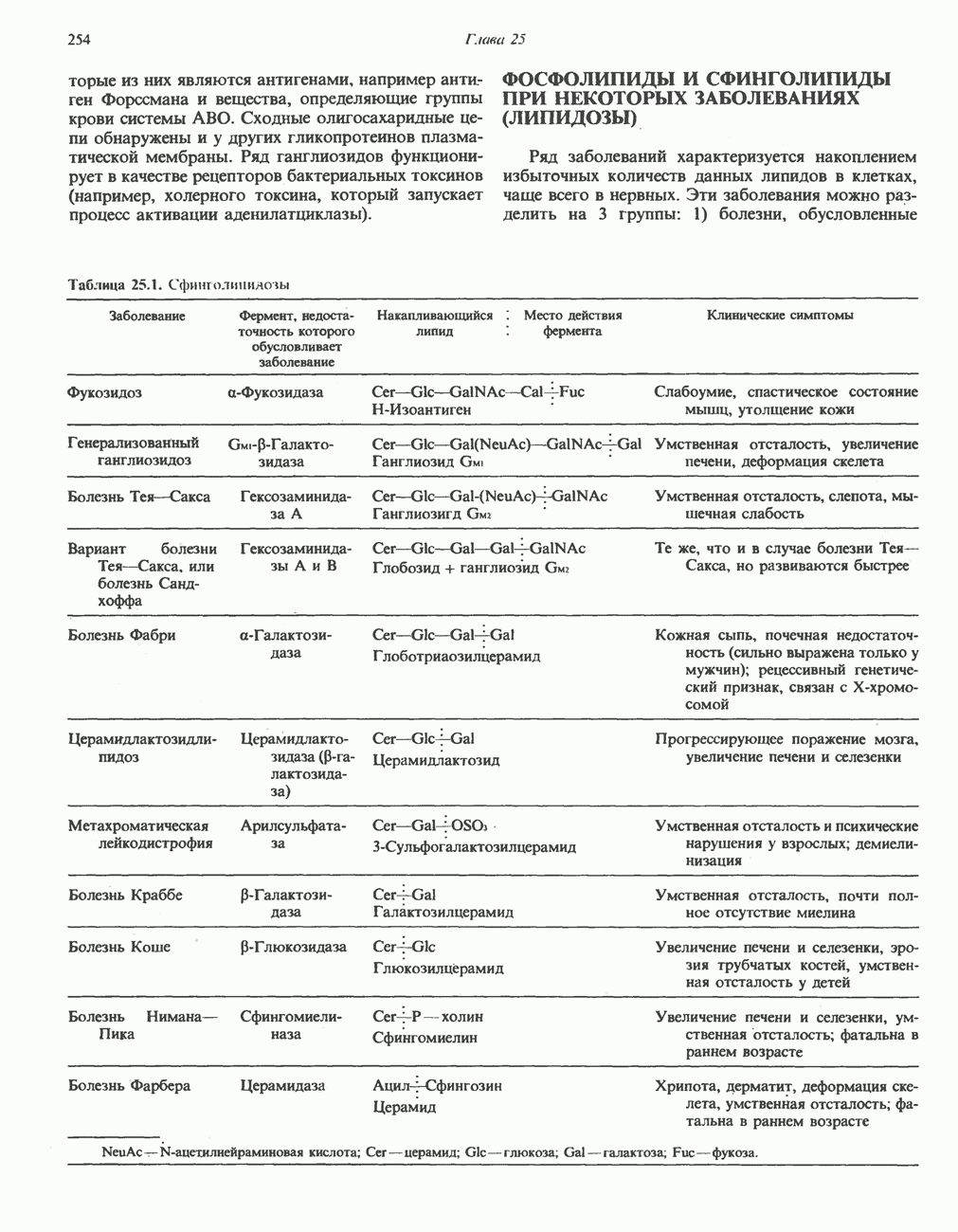
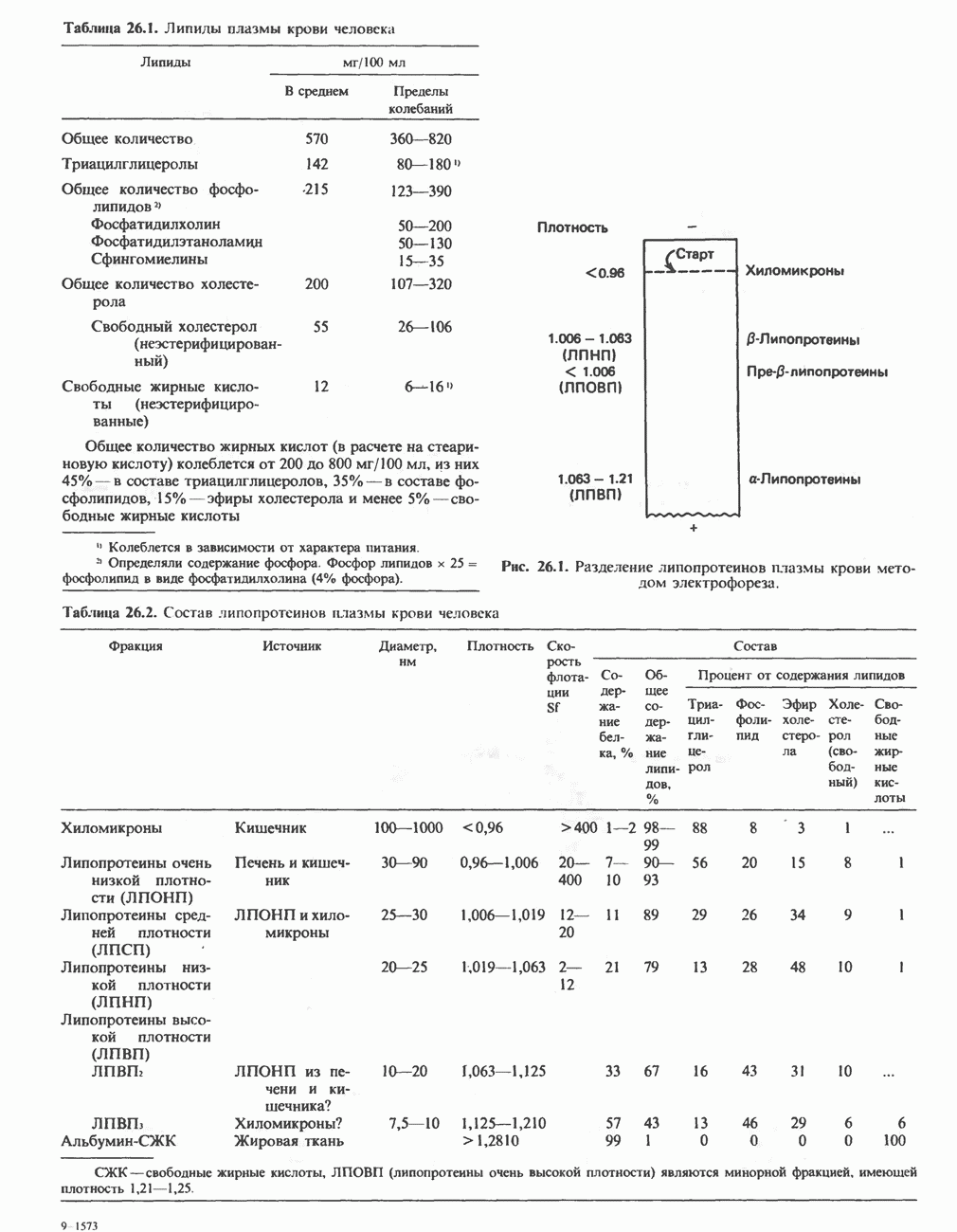
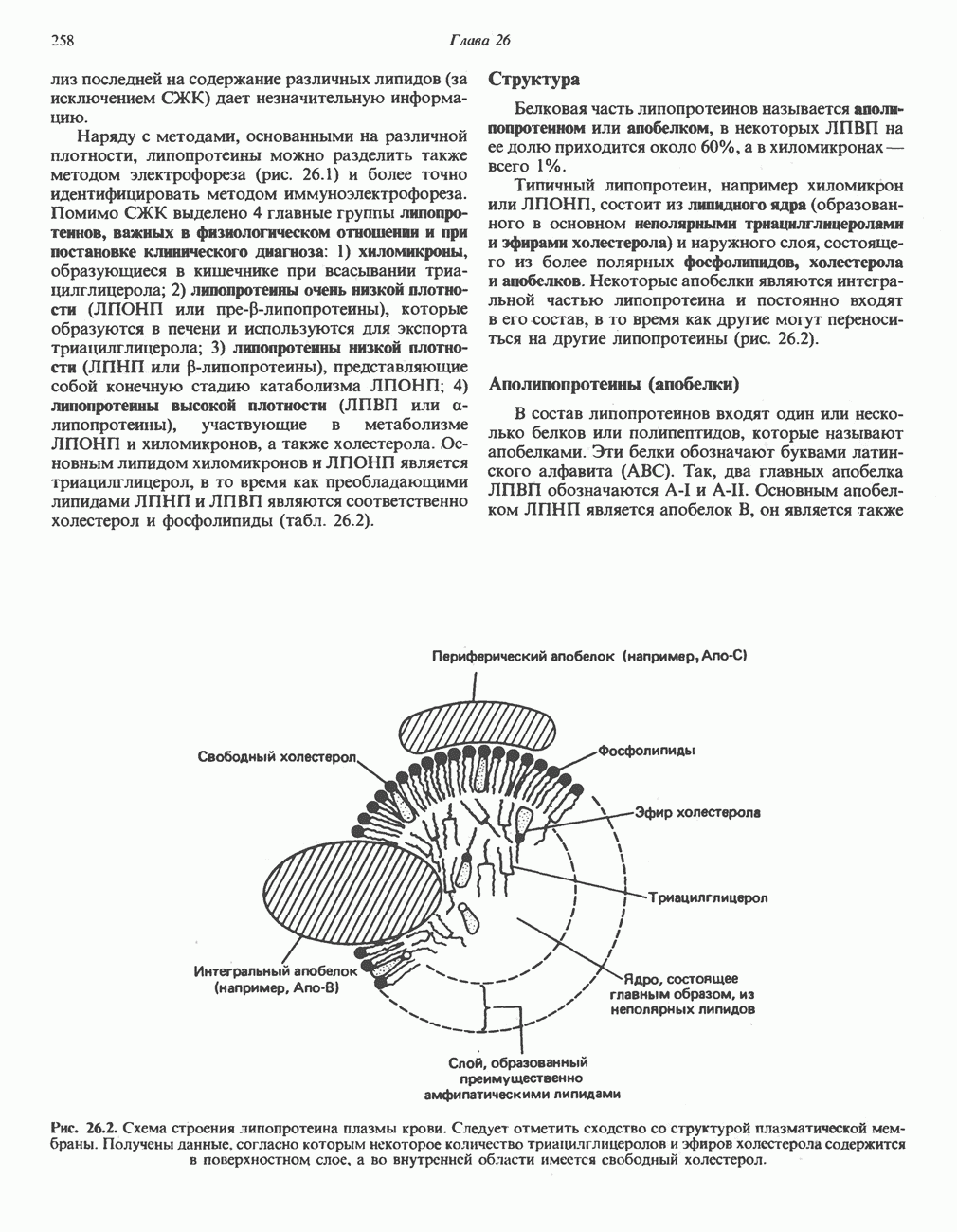
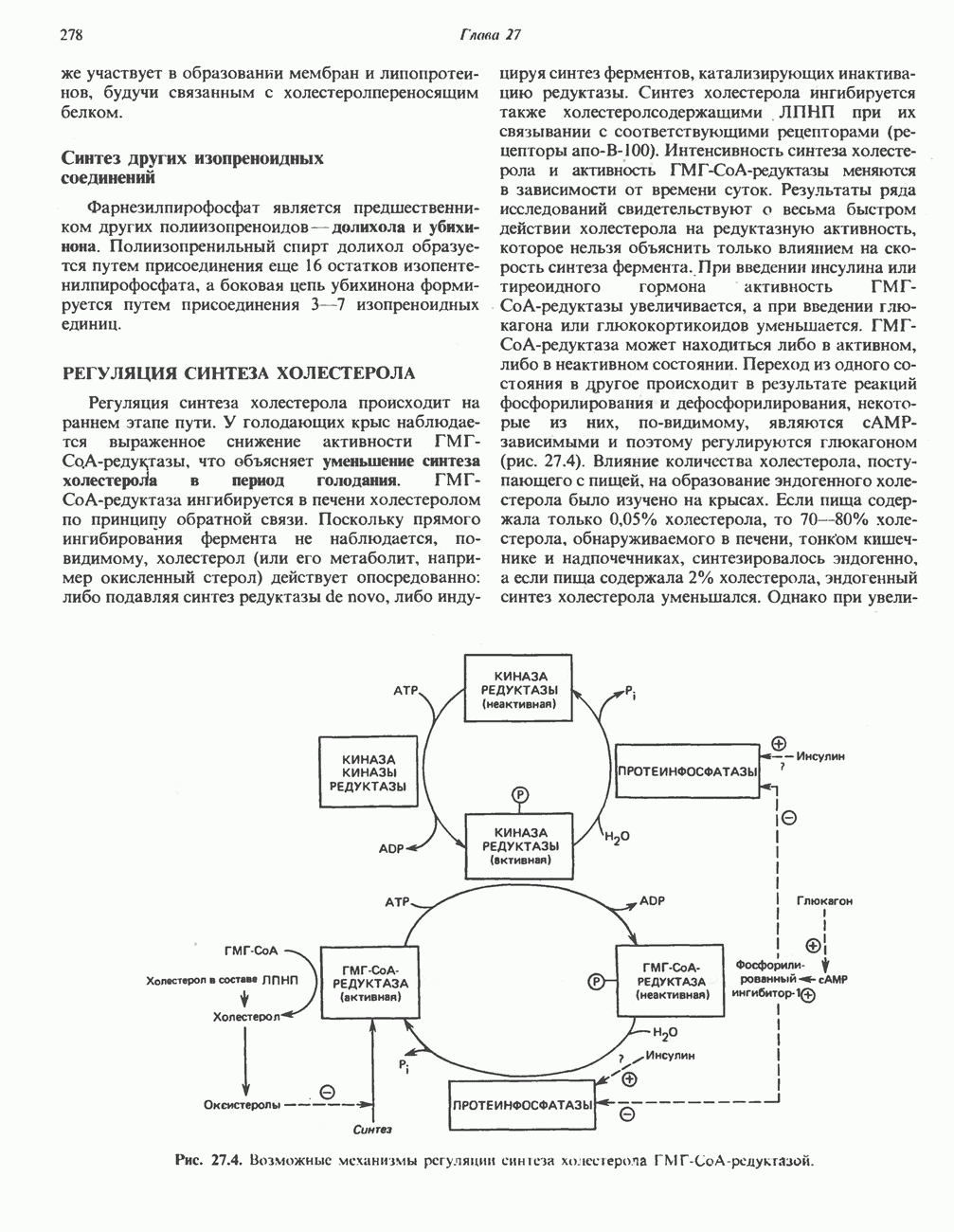
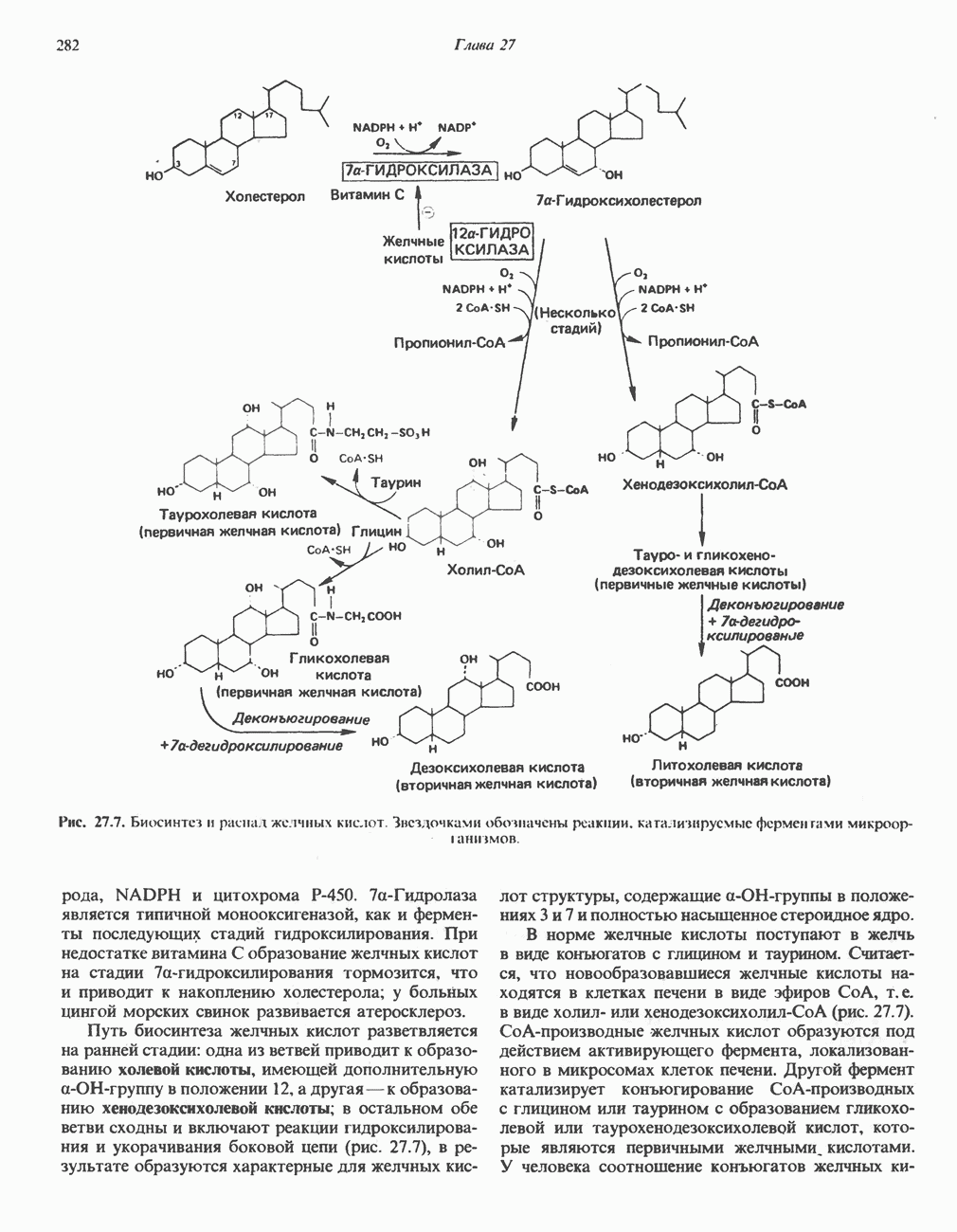
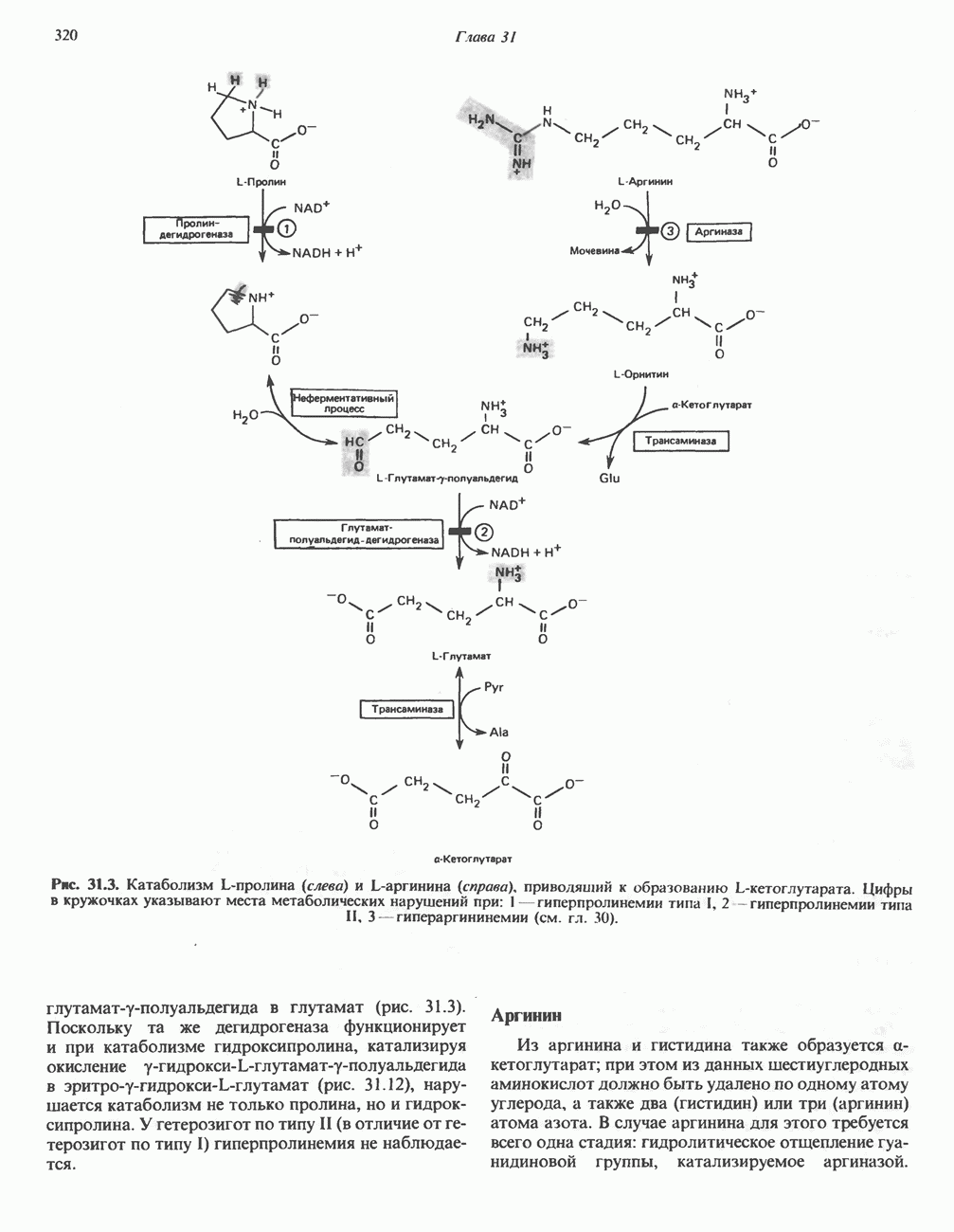
Первичные желчные кислоты синтезируются в печени из холестерола через ряд промежуточных соединений. Среди желчных кислот доминирует в количественном отношении холевая кислота. Холевая и хенодезоксихолевая кислоты имеют общего предшественника, образующегося из холестерола (рис. 27.7).
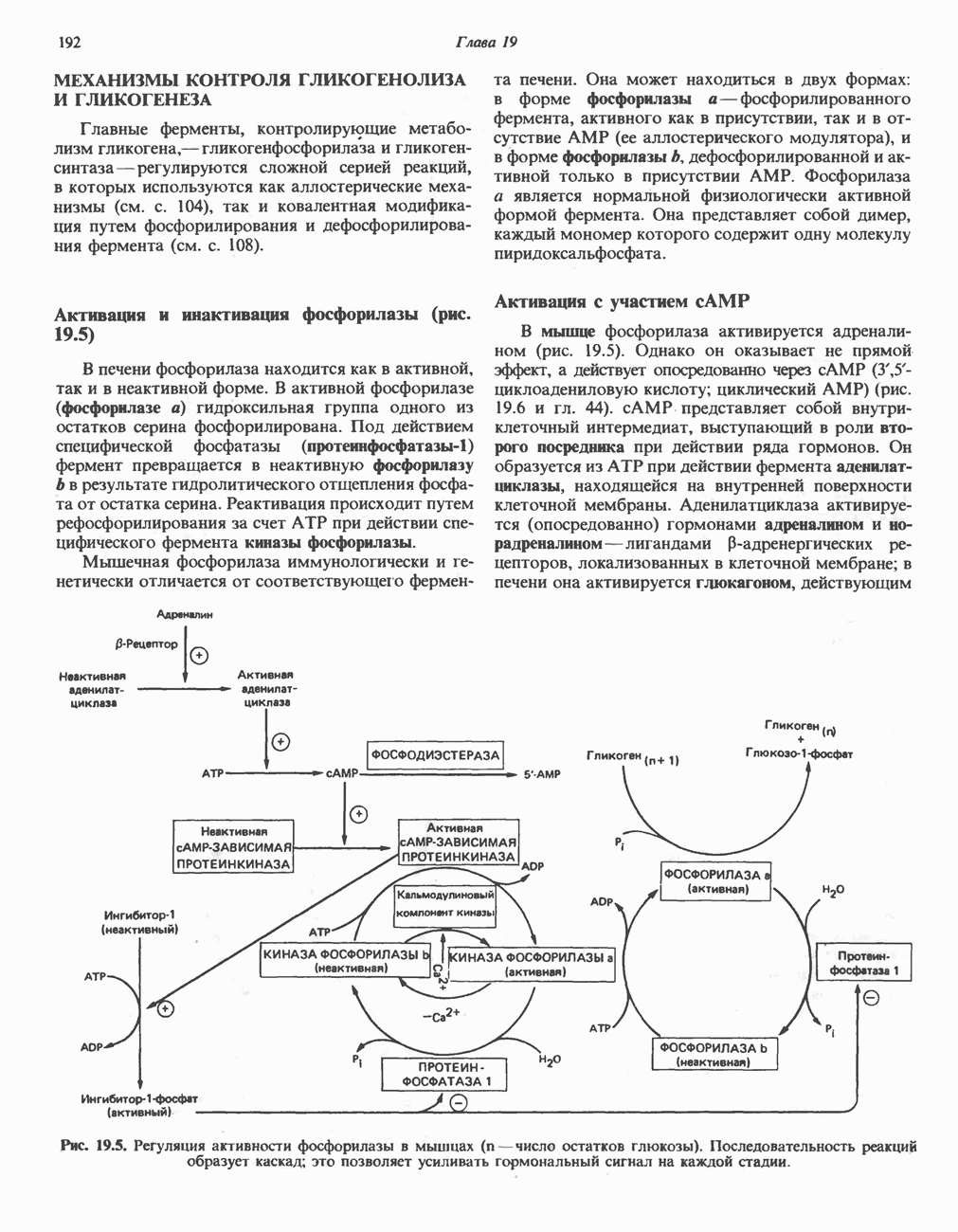
Первым этапом биосинтеза желчных кислот является  -гидроксилирование холестерола, и именно эта реакция является скорость - определяющей в процессе биосинтеза желчных кислот. Она катализируется микросомальным ферментом
-гидроксилирование холестерола, и именно эта реакция является скорость - определяющей в процессе биосинтеза желчных кислот. Она катализируется микросомальным ферментом  -гидроксилазой и протекает с участием кислорода
-гидроксилазой и протекает с участием кислорода
NADPH и цитохрома  является типичной монооксигеназой, как и ферменты последующих стадий гидроксилирования. При недостатке витамина С образование желчных кислот на стадии
является типичной монооксигеназой, как и ферменты последующих стадий гидроксилирования. При недостатке витамина С образование желчных кислот на стадии  тормозится, что и приводит к накоплению холестерола; у больных цингой морских свинок развивается атеросклероз.
тормозится, что и приводит к накоплению холестерола; у больных цингой морских свинок развивается атеросклероз.
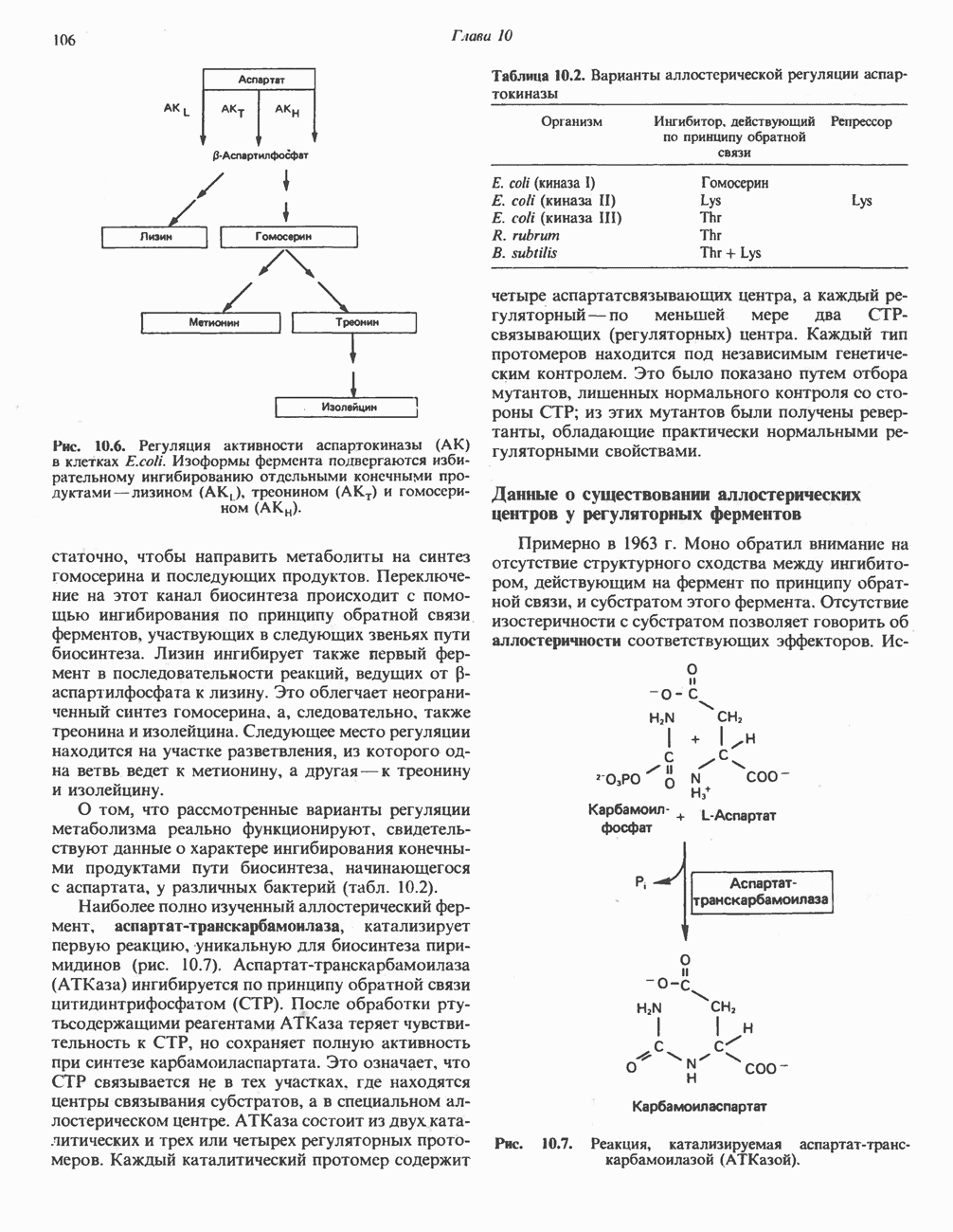
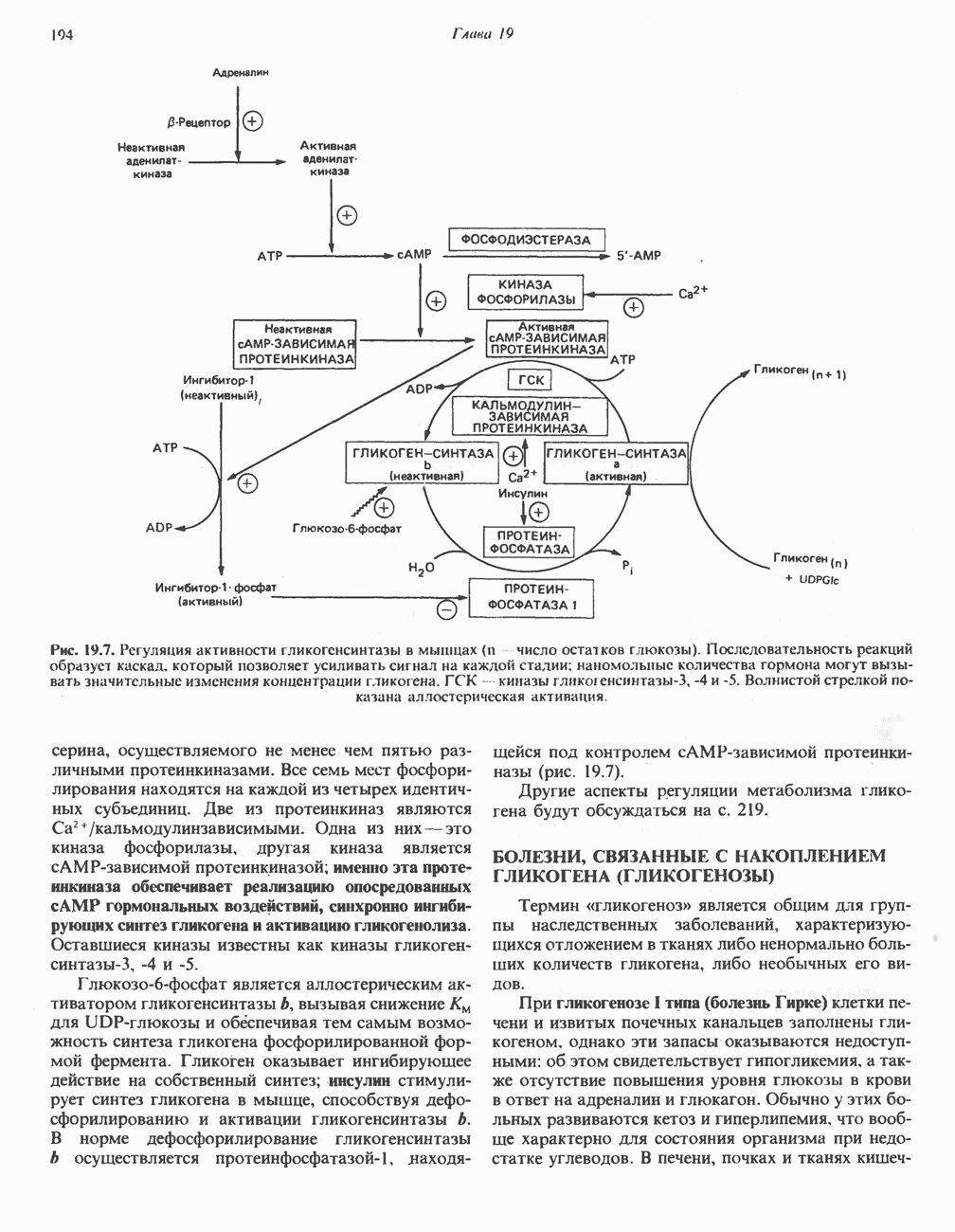
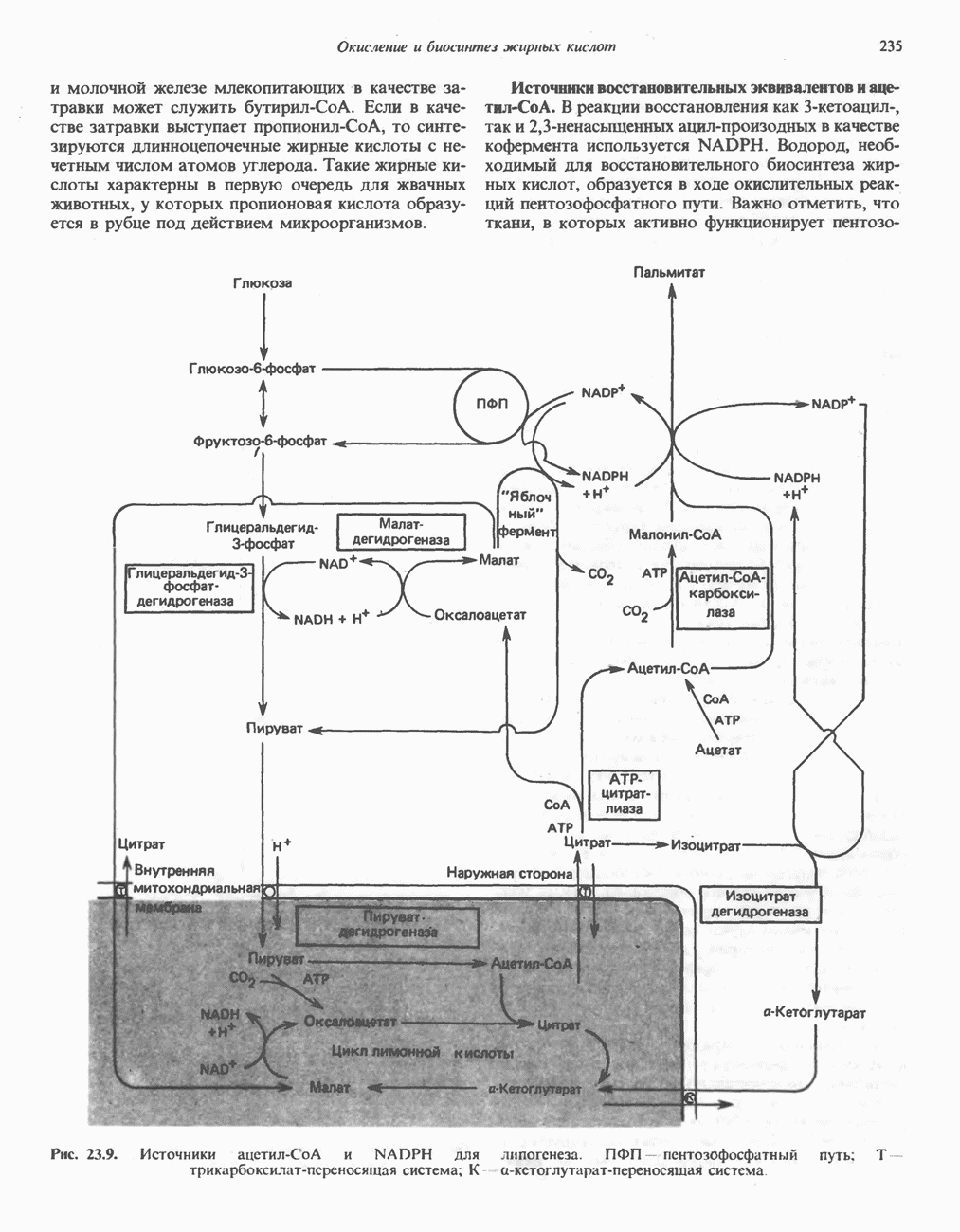
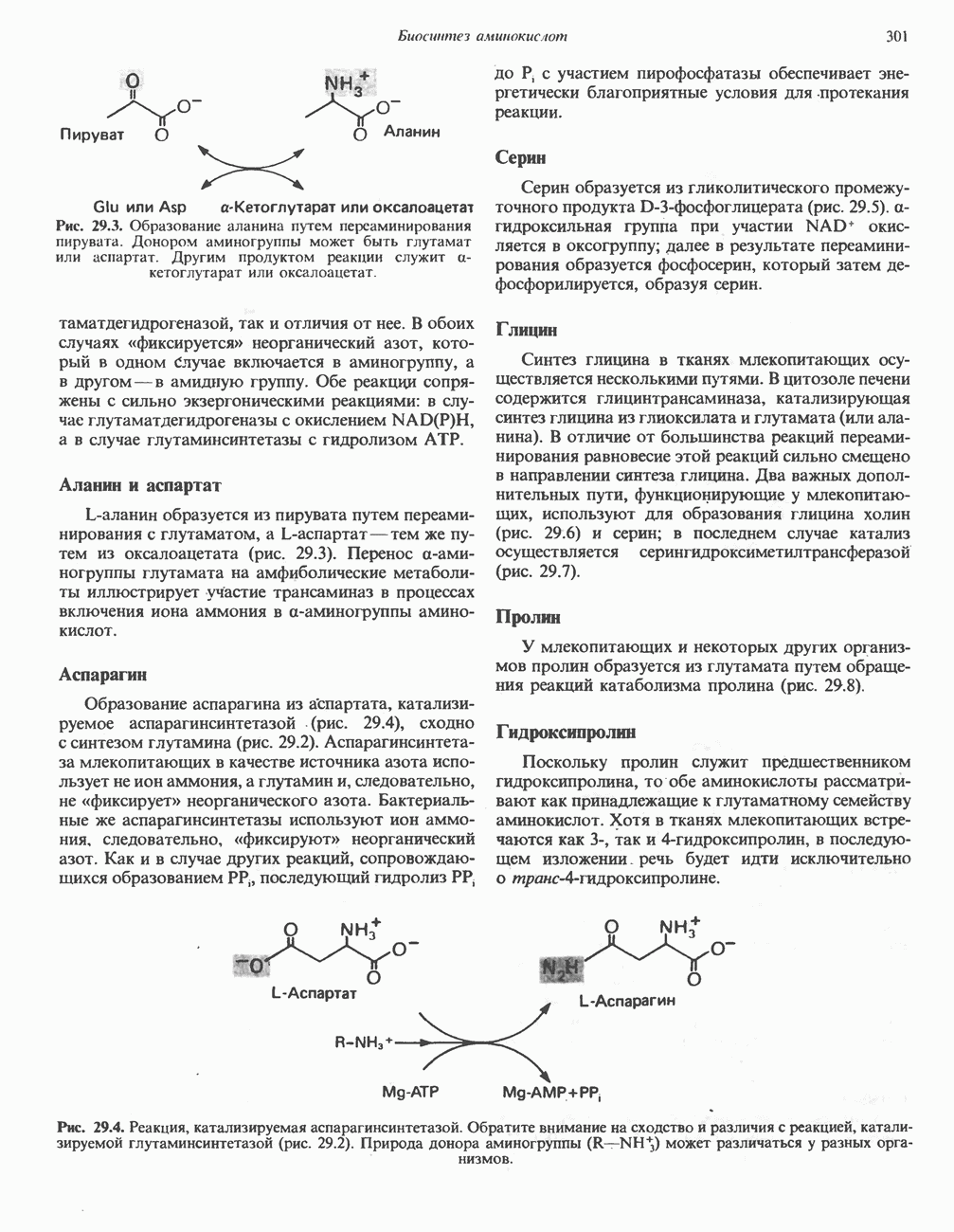
Путь биосинтеза желчных кислот разветвляется на ранней стадии: одна из ветвей приводит к образованию холевой кислоты, имеющей дополнительную а-ОН-группу в положении 12, а другая — к образованию хенодезоксихолевой кислоты; в остальном обе ветви сходны и включают реакции гидроксилирования и укорачивания боковой цепи (рис. 27.7), в результате образуются характерные для желчных кислот структуры, содержащие а-ОН-группы в положениях 3 и 7 и полностью насыщенное стероидное ядро.
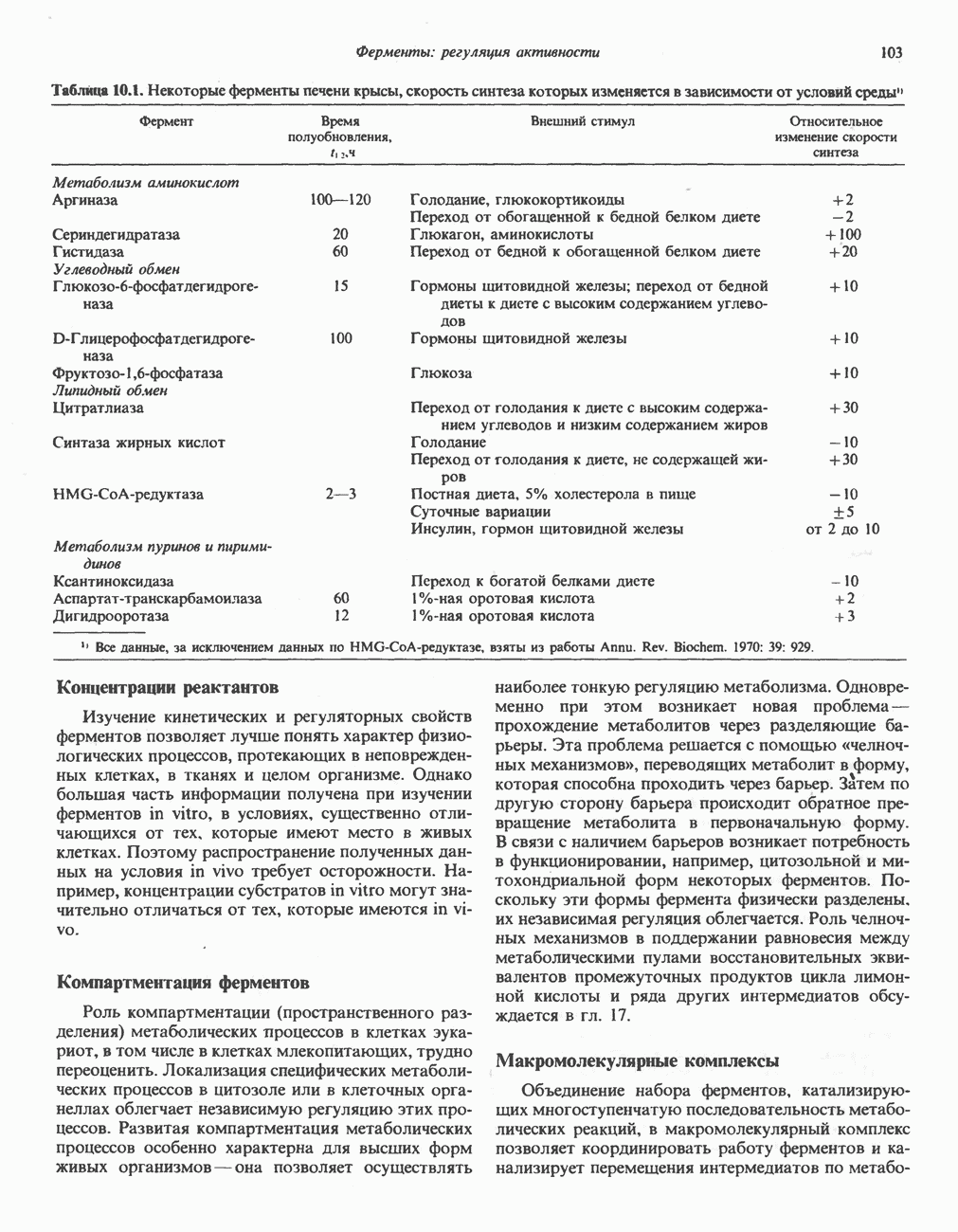
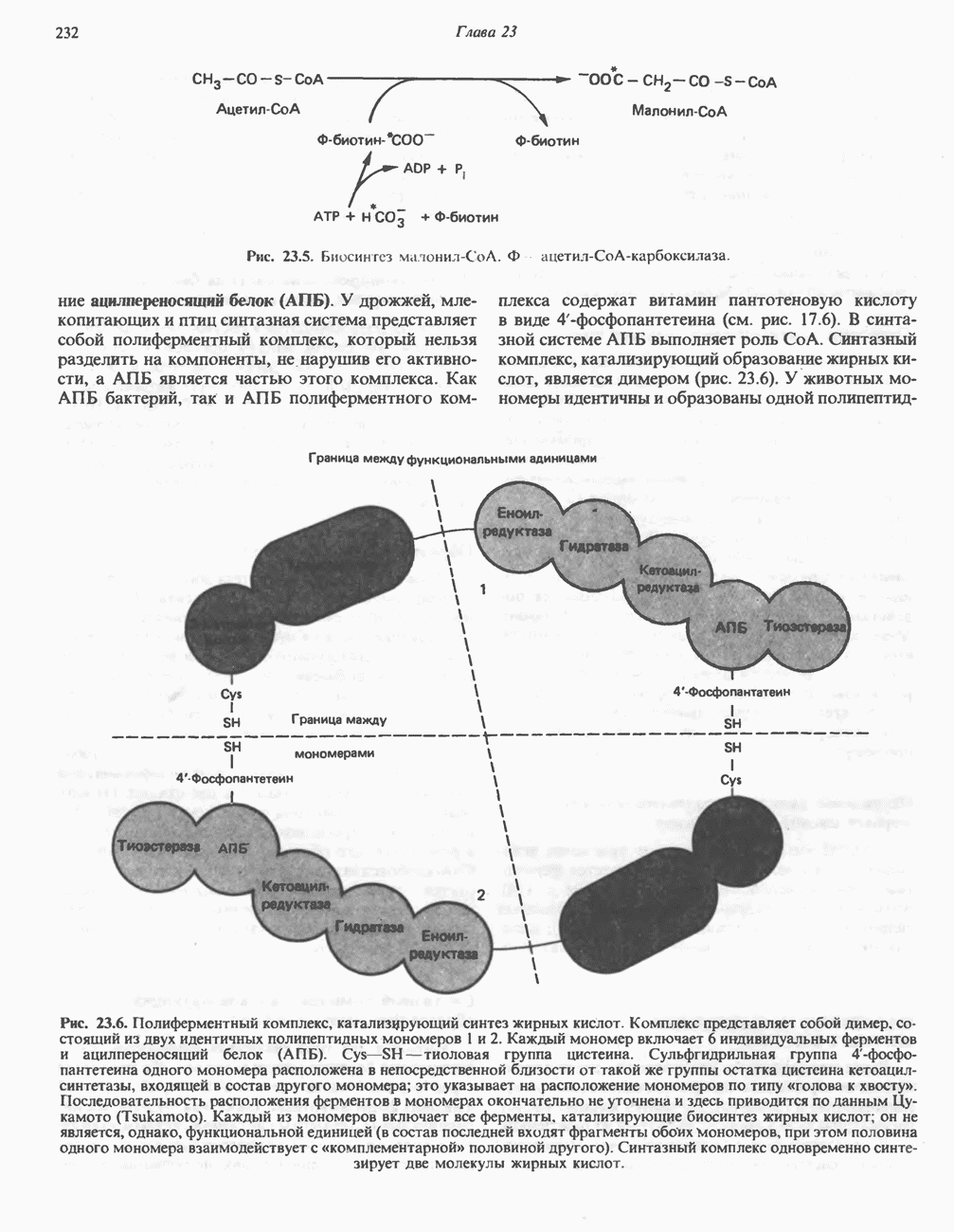
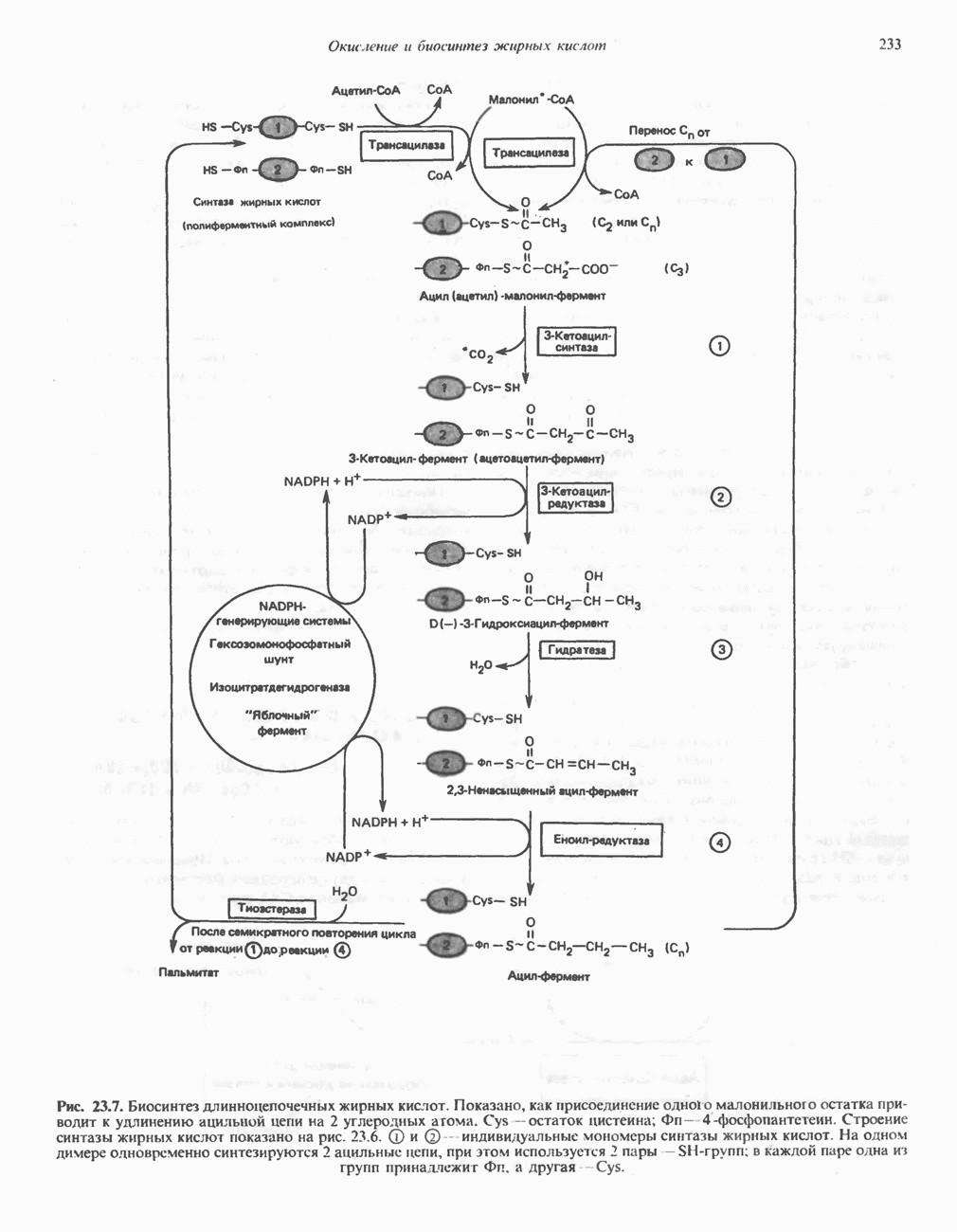
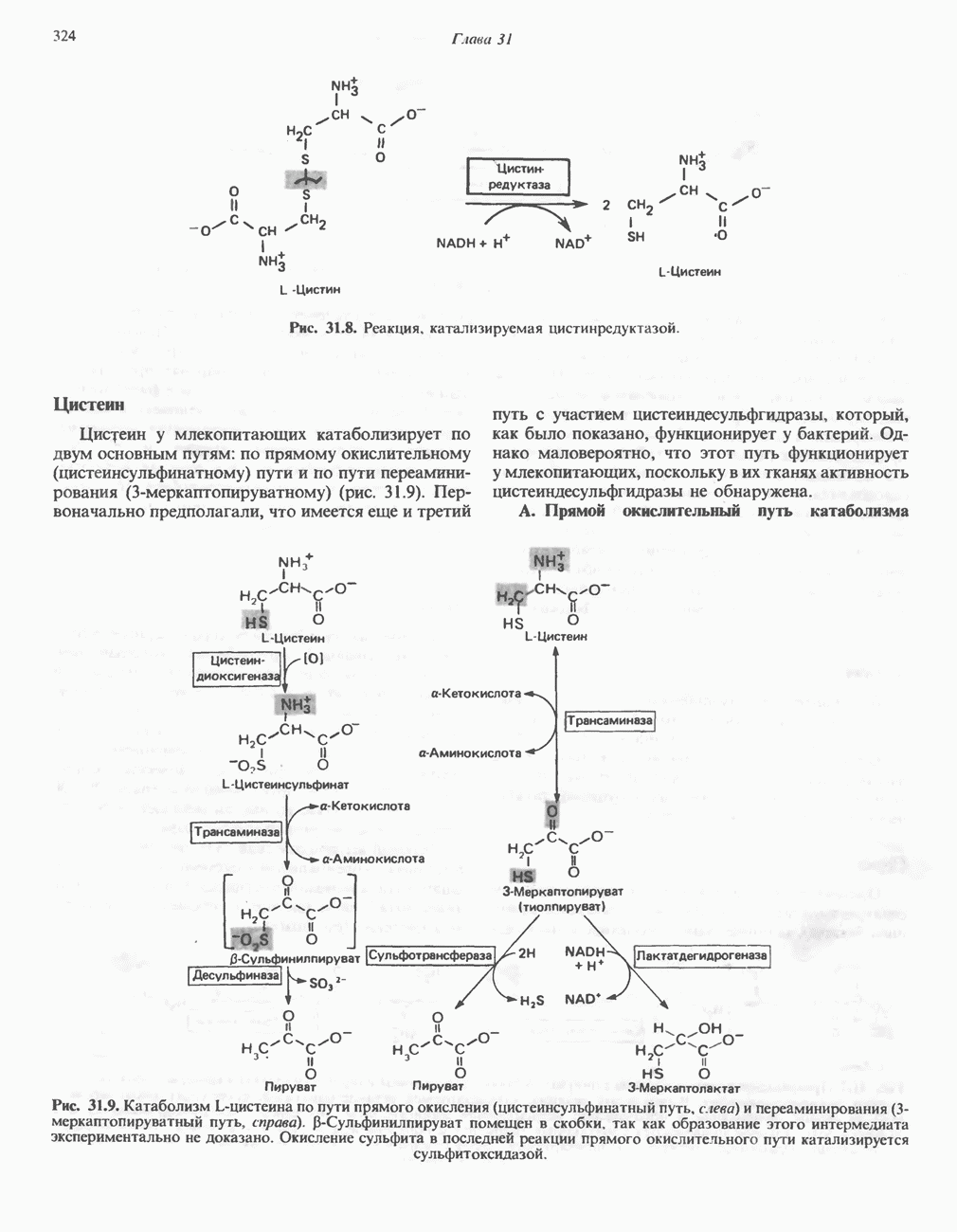
В норме желчные кислоты поступают в желчь в виде конъюгатов с глицином и таурином. Считается, что новообразовавшиеся желчные кислоты находятся в клетках печени в виде эфиров СоА, т.е. в виде холил- или хенодезоксихолил-СоА (рис. 27.7). СоА-производные желчных кислот образуются под действием активирующего фермента, локализованного в микросомах клеток печени. Другой фермент катализирует конъюгирование СоА-производных с глицином или таурином с образованием гликохолевой или таурохенодезоксихолевой кислот, которые являются первичными желчными, кислотами. У человека соотношение конъюгатов желчных
(см. скан)
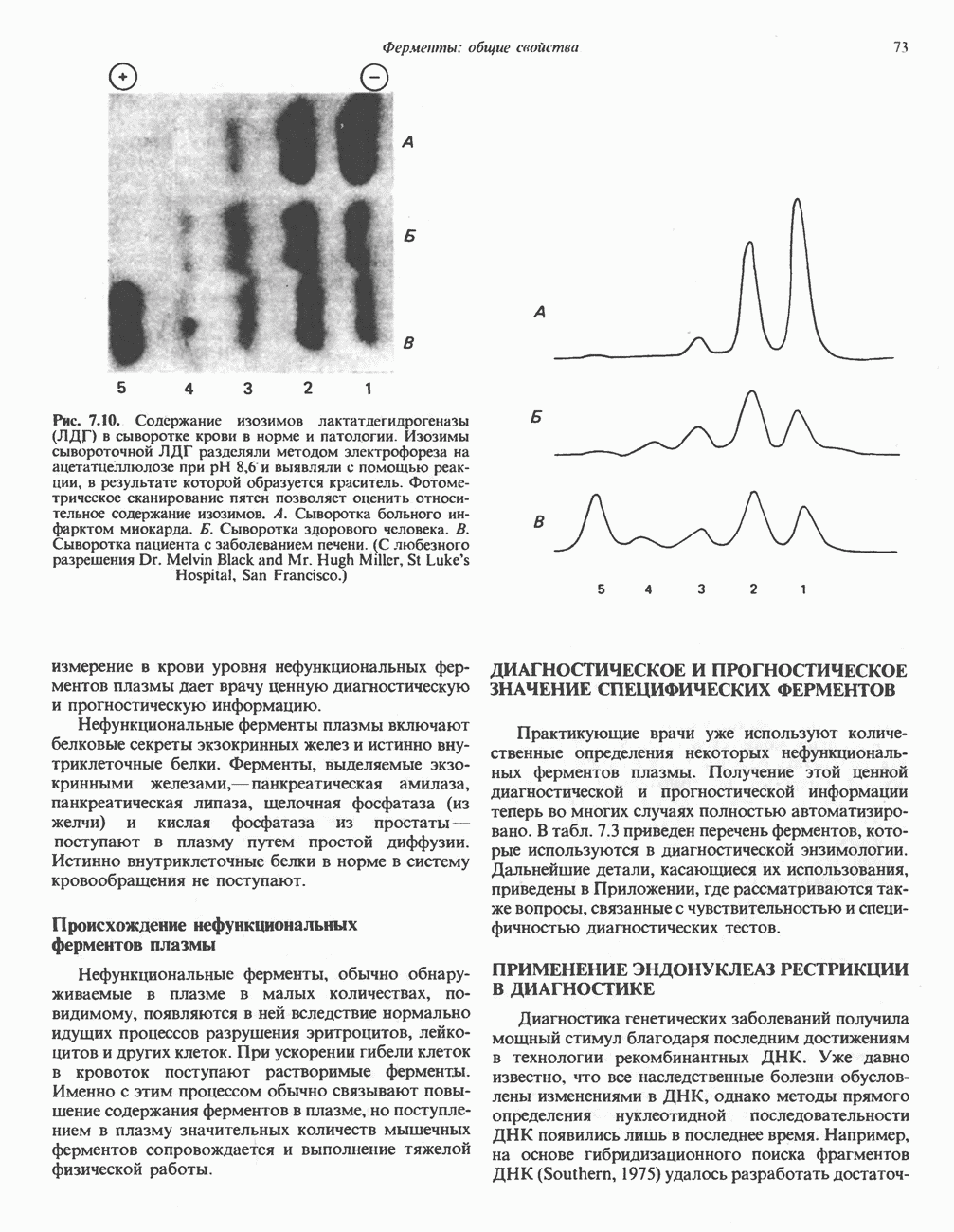
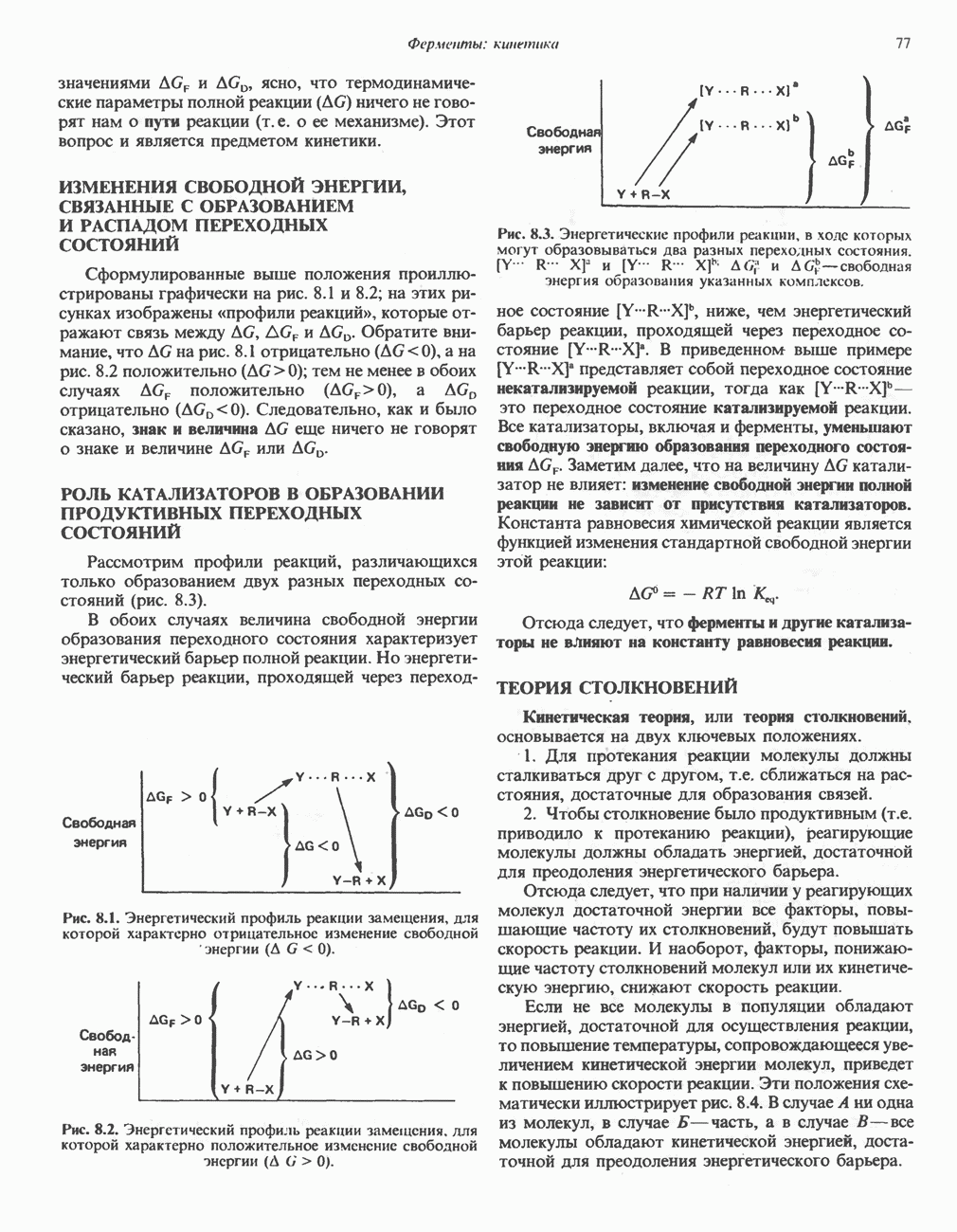
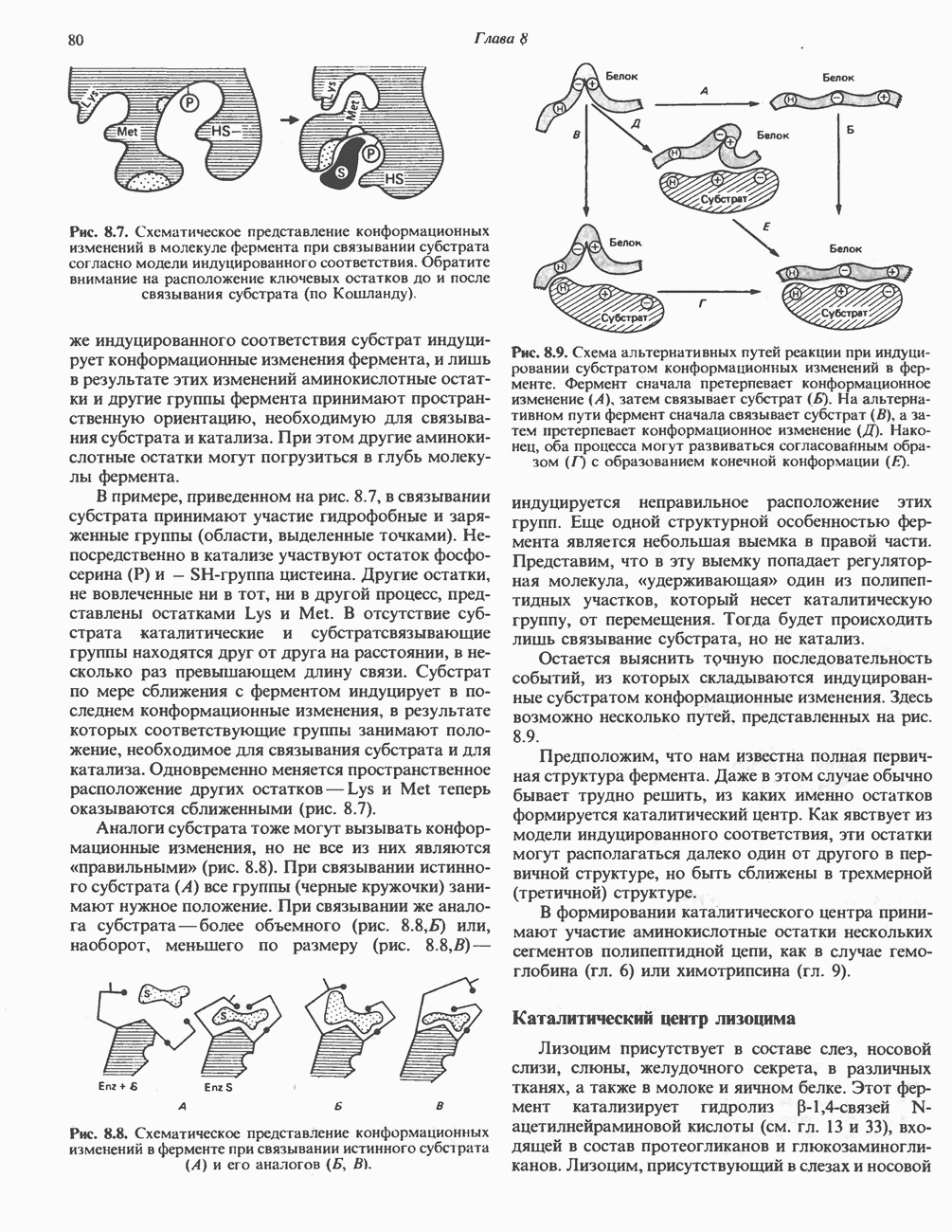
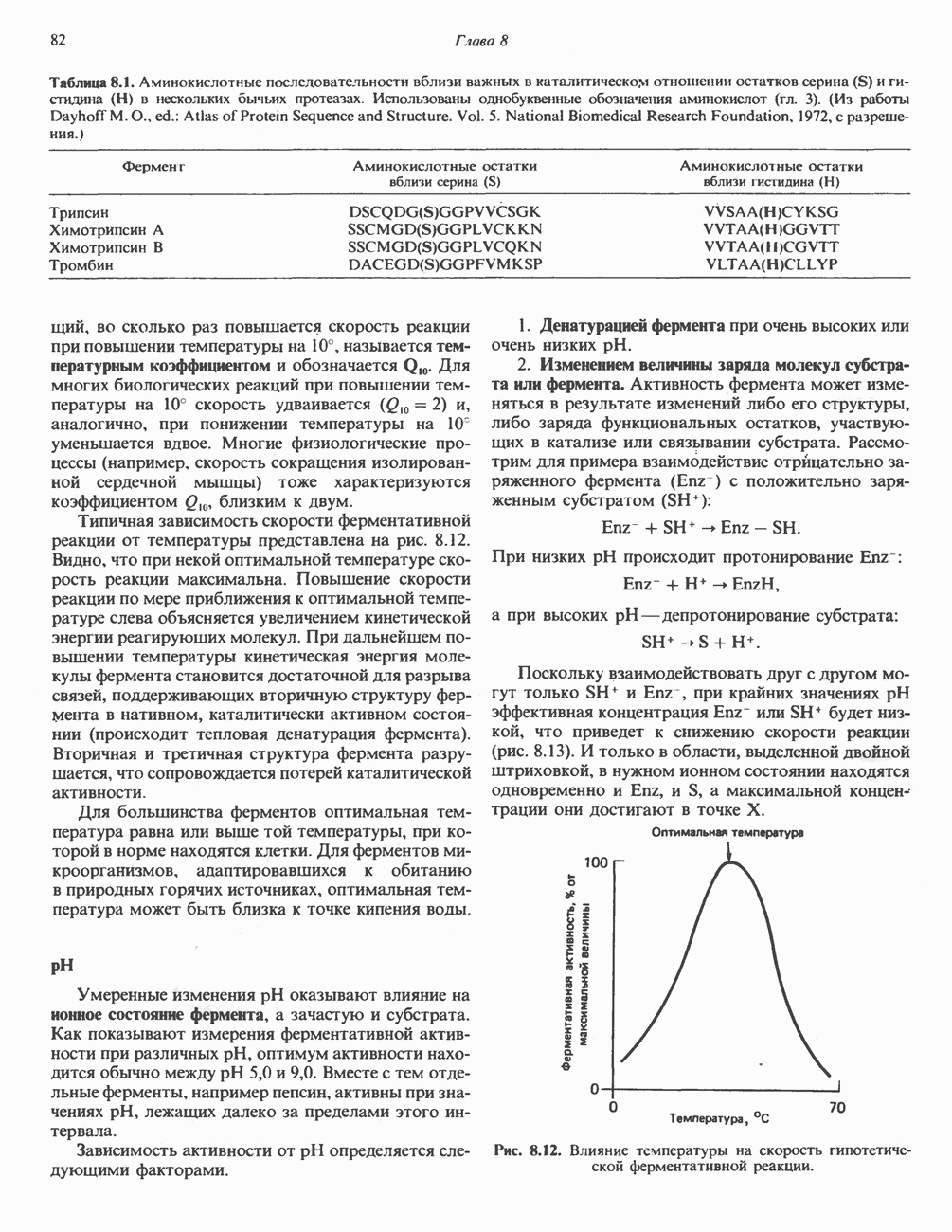
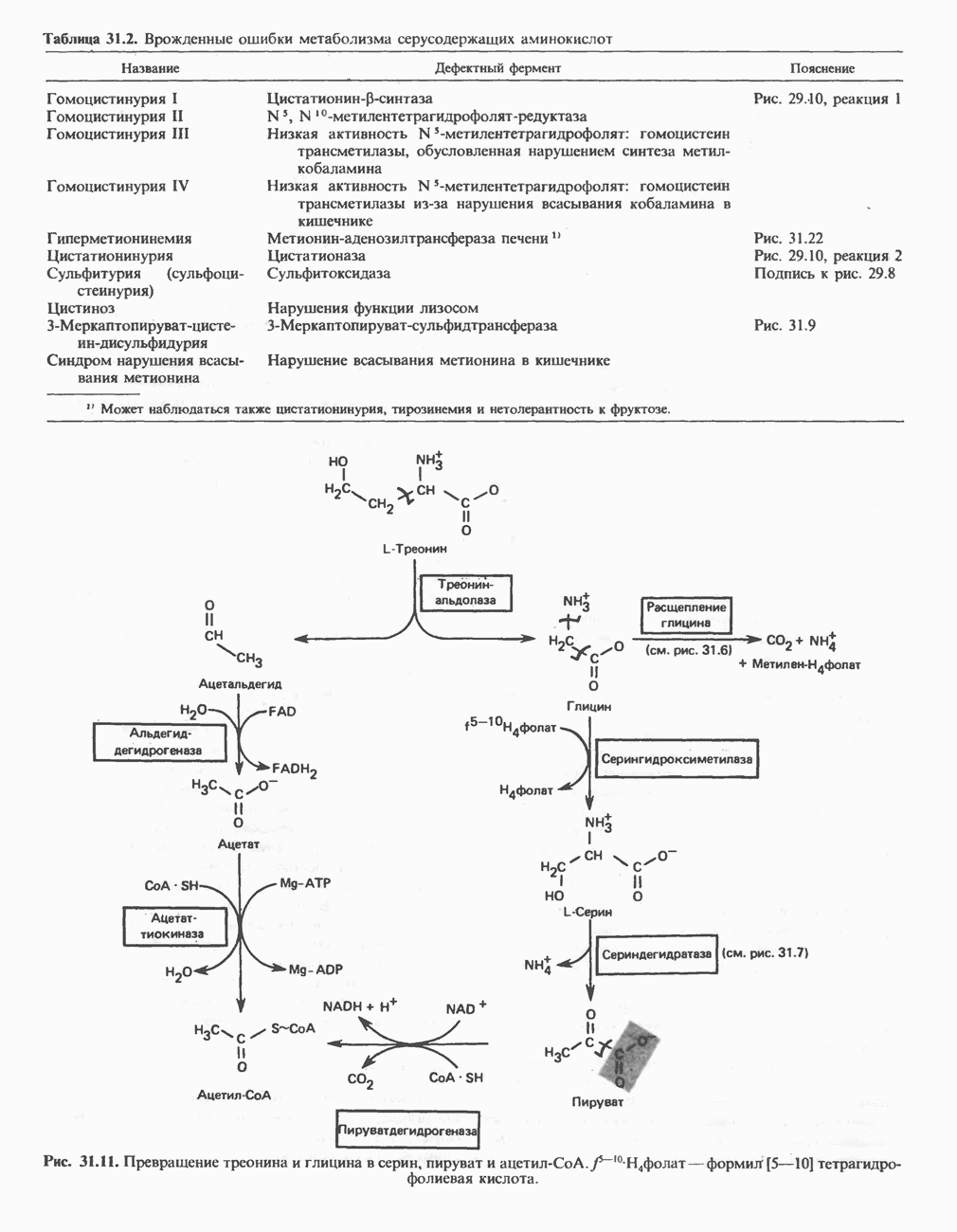
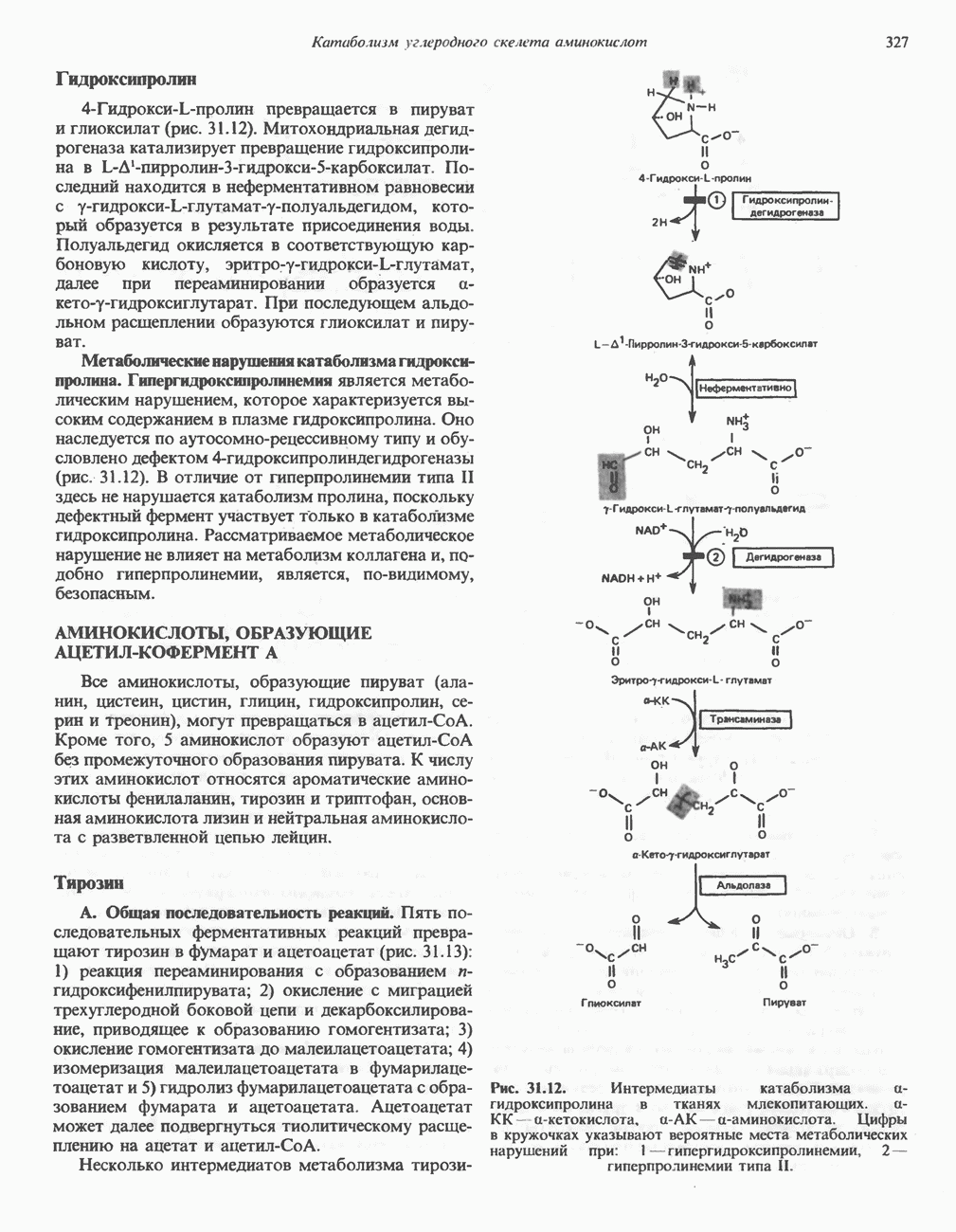
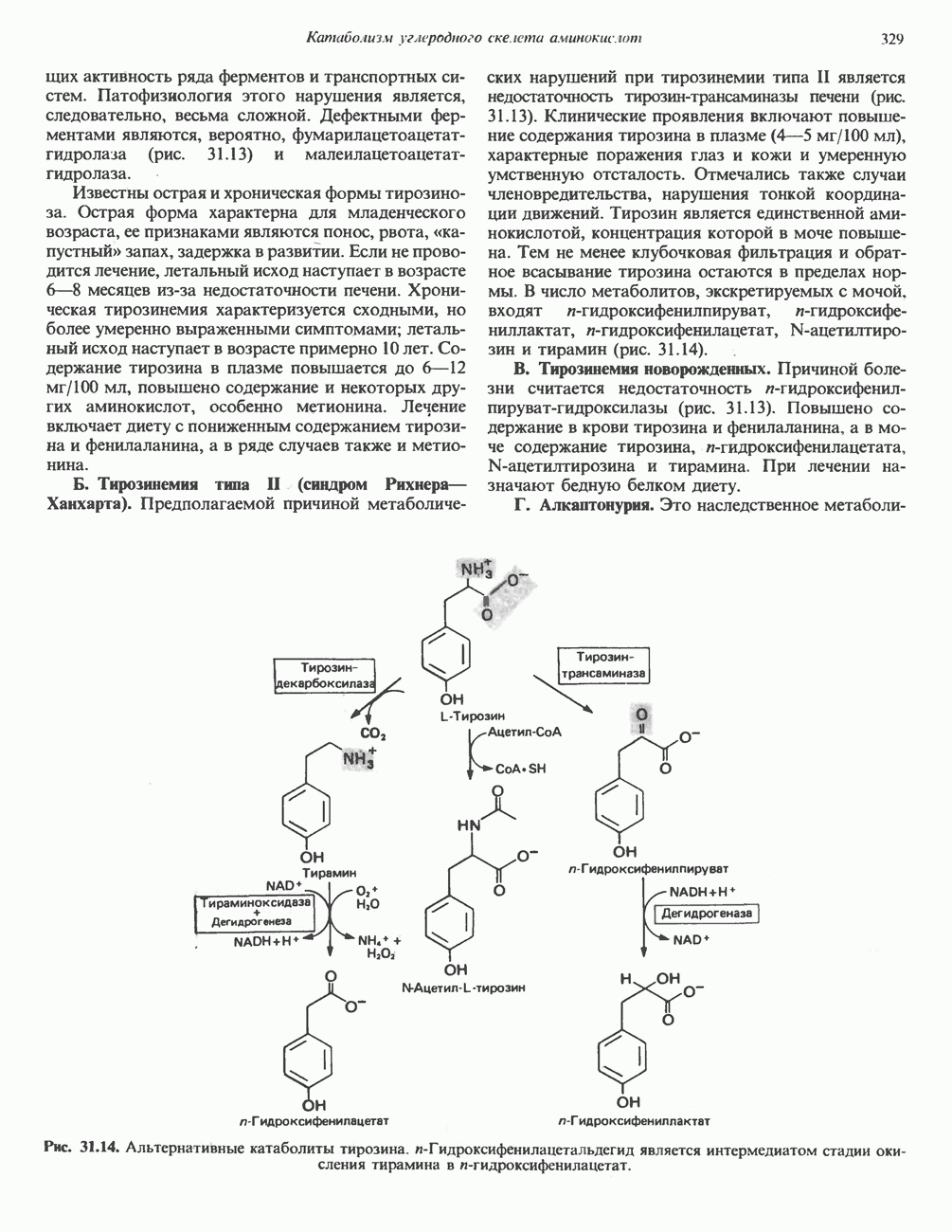
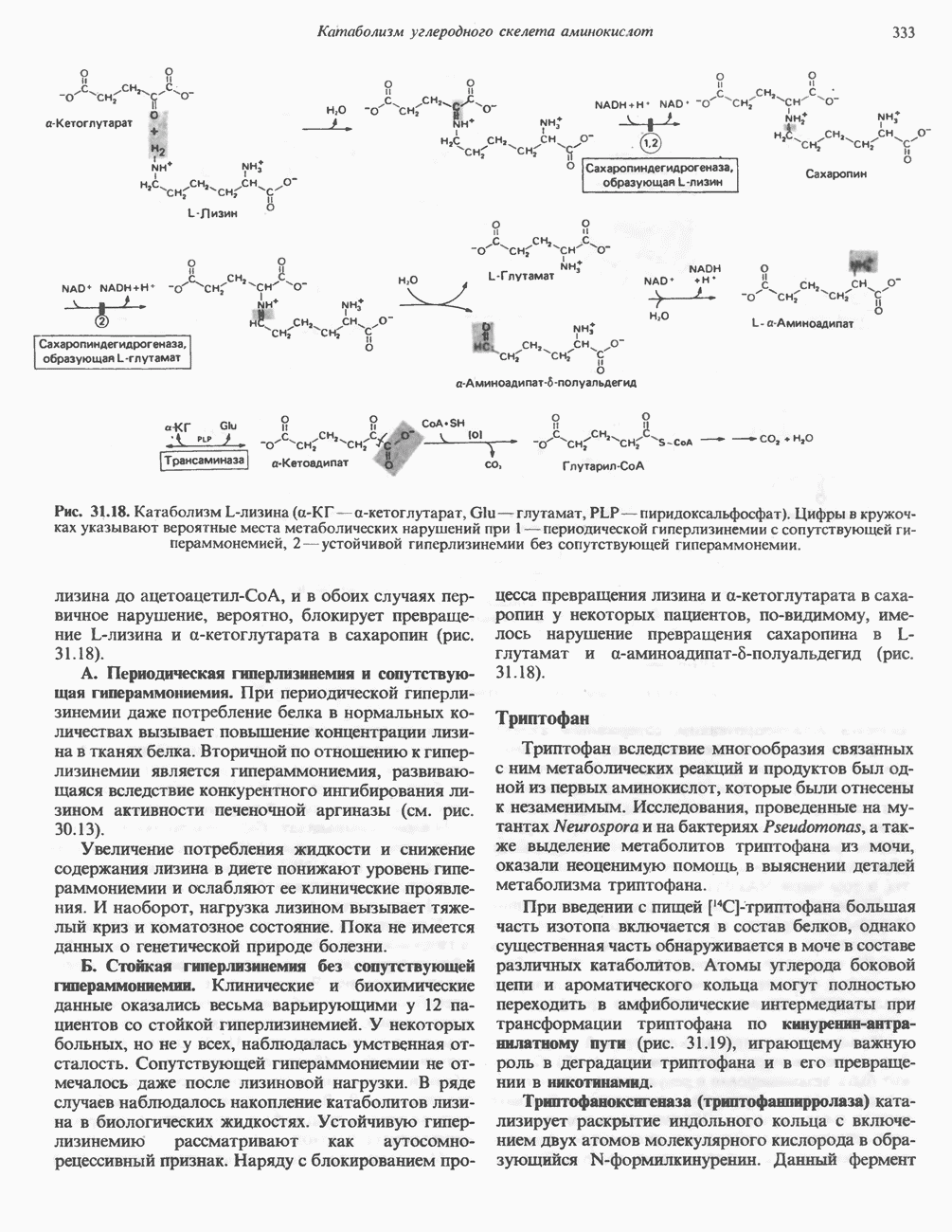
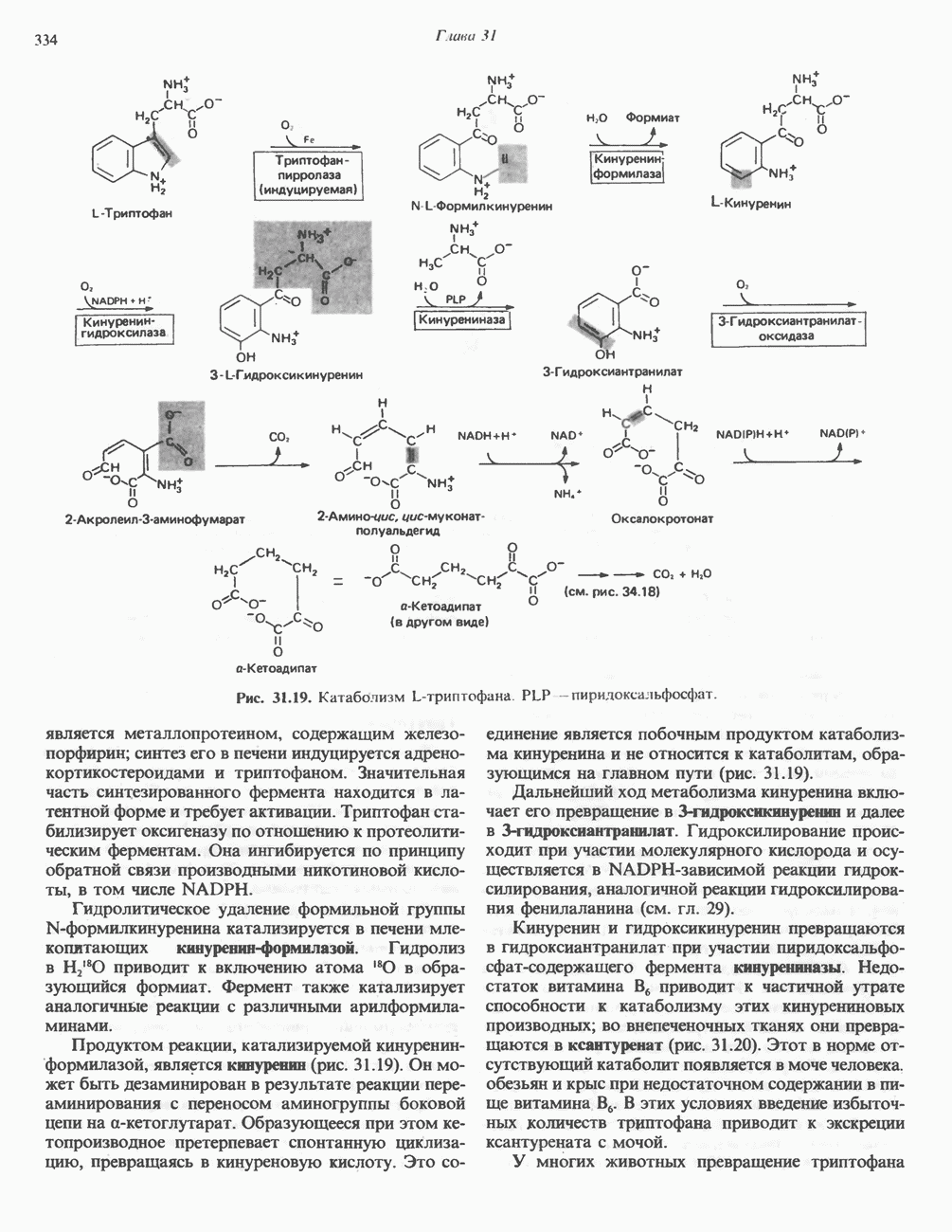
Рис. 27.7. Биосинтез и распад желчных кислот. Звездочками обозначены реакции, катализируемые ферментами микроорганизмов.
кислот с глицином и таурином составляет в норме 3:1.
Поскольку желчь содержит значительное количество ионов натрия и калия и имеет щелочную реакцию, то принято рассматривать желчные кислоты и их конъюгаты в форме солей, откуда и возник термин «желчные соли».
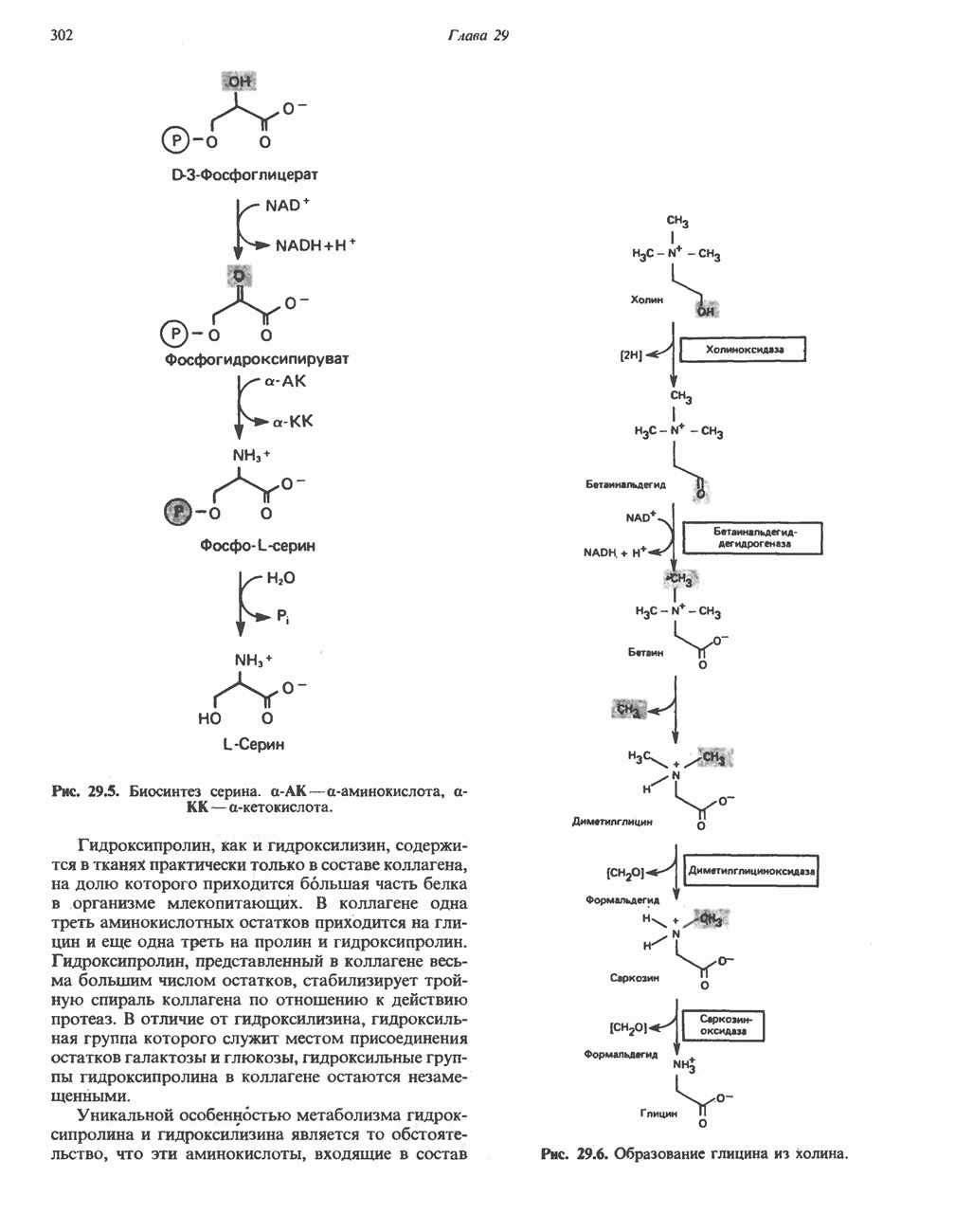
В кишечнике часть первичных желчных кислот под действием кишечных бактерий подвергается деконъюгированию и  -дегидроксилированию, в результате чего образуются вторичные желчные кислоты: из холевой кислоты—дезоксихолевая кислота и из хенодезоксихолевой кислоты — литохолевая кислота (рис. 27.7).
-дегидроксилированию, в результате чего образуются вторичные желчные кислоты: из холевой кислоты—дезоксихолевая кислота и из хенодезоксихолевой кислоты — литохолевая кислота (рис. 27.7).