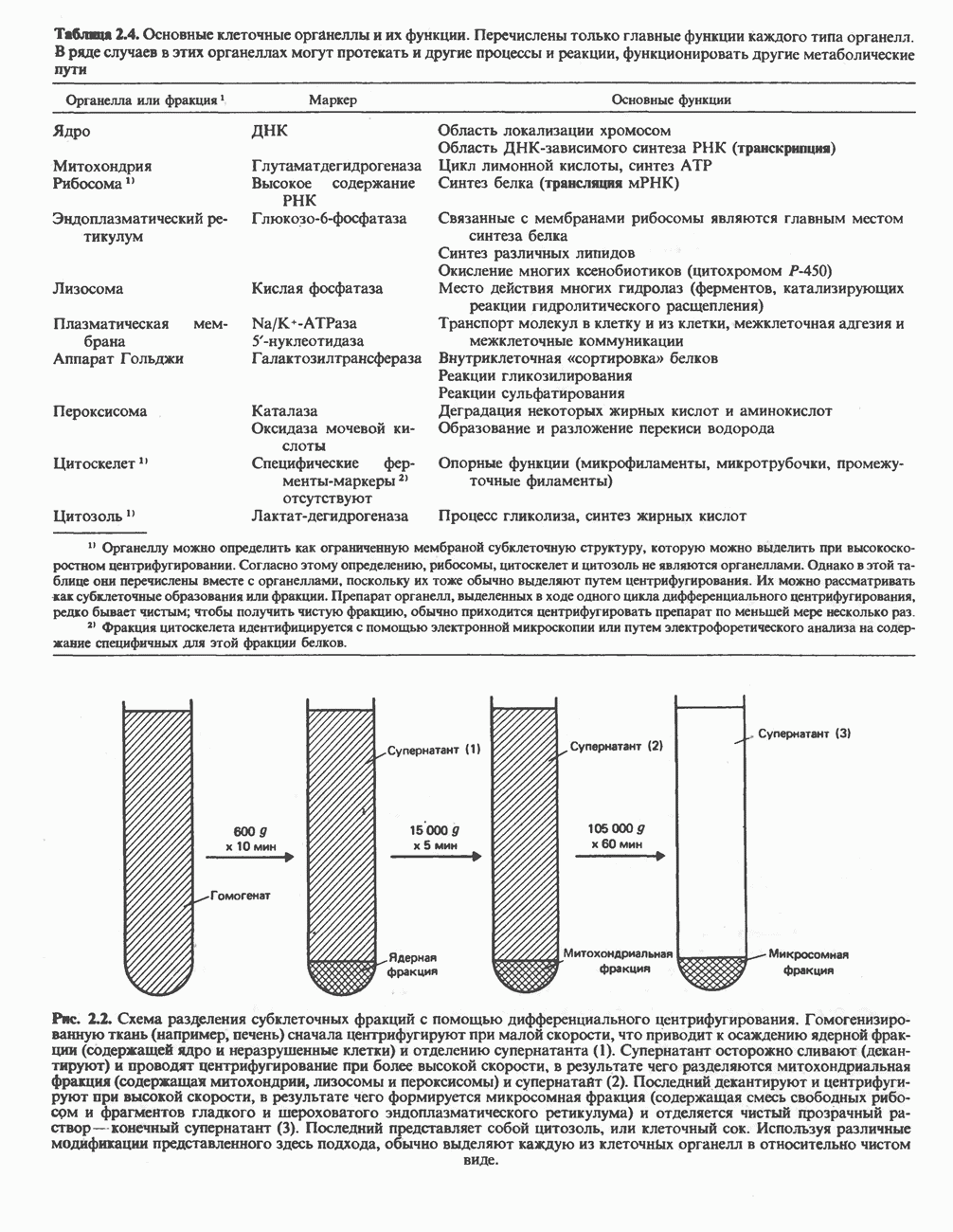
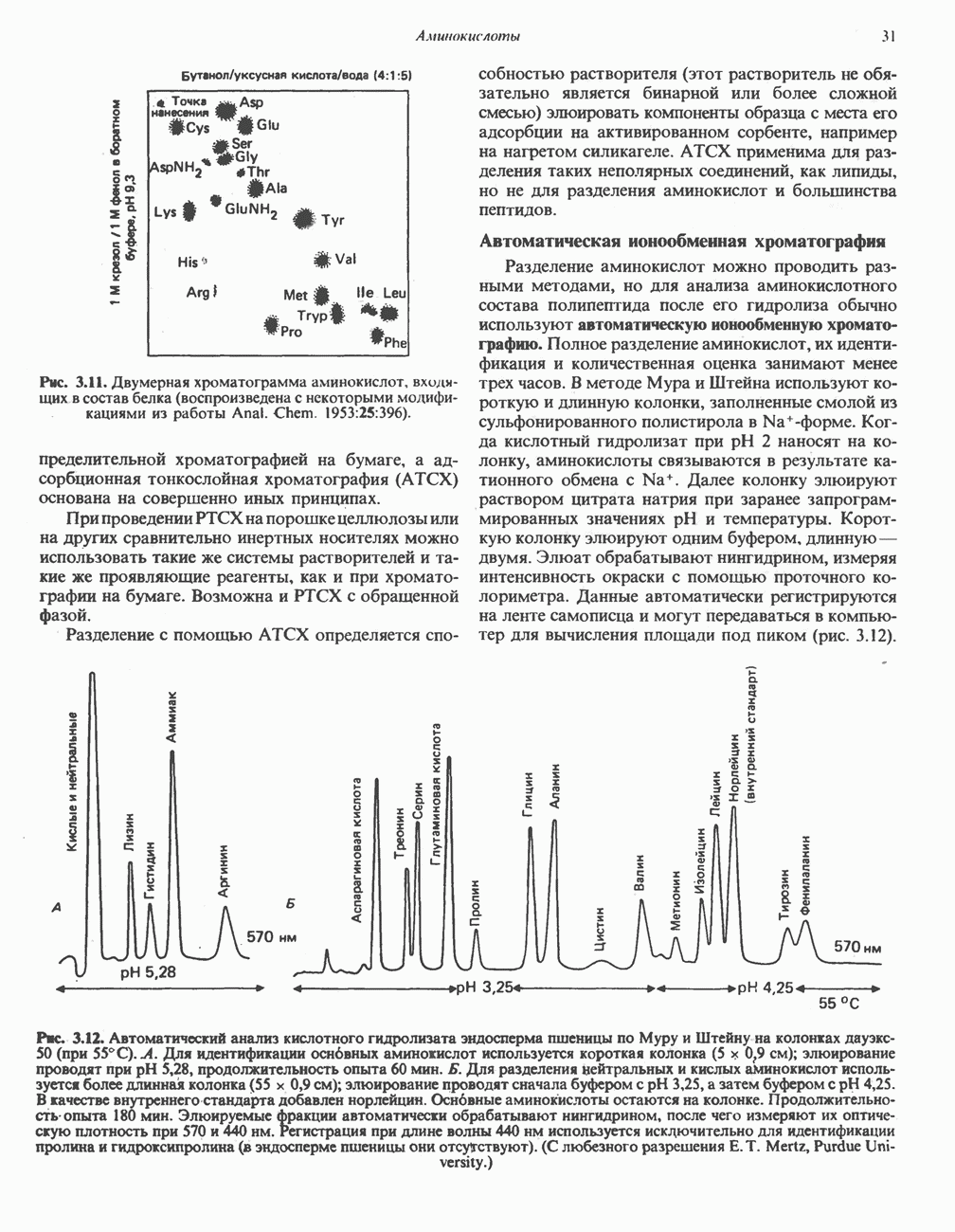
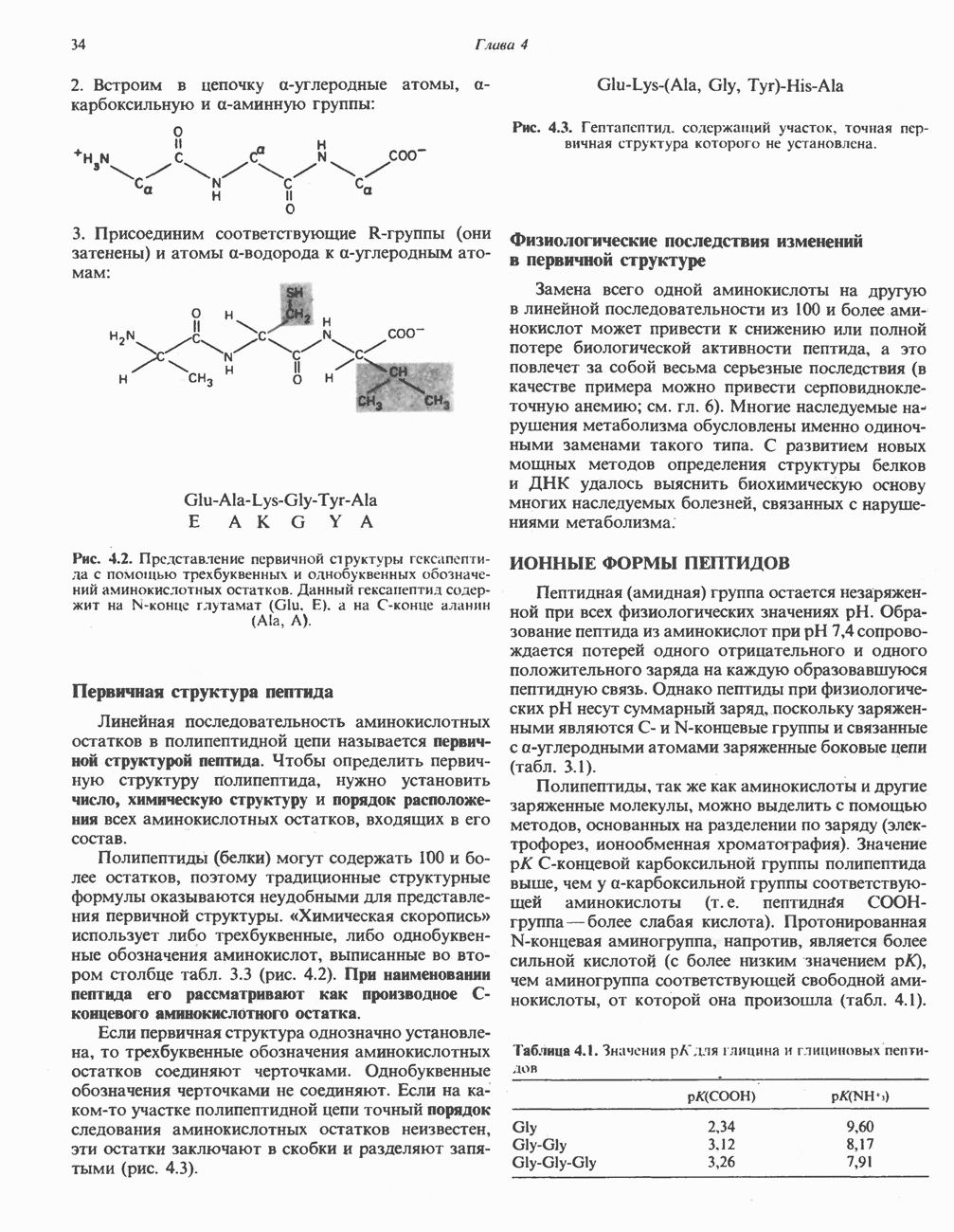
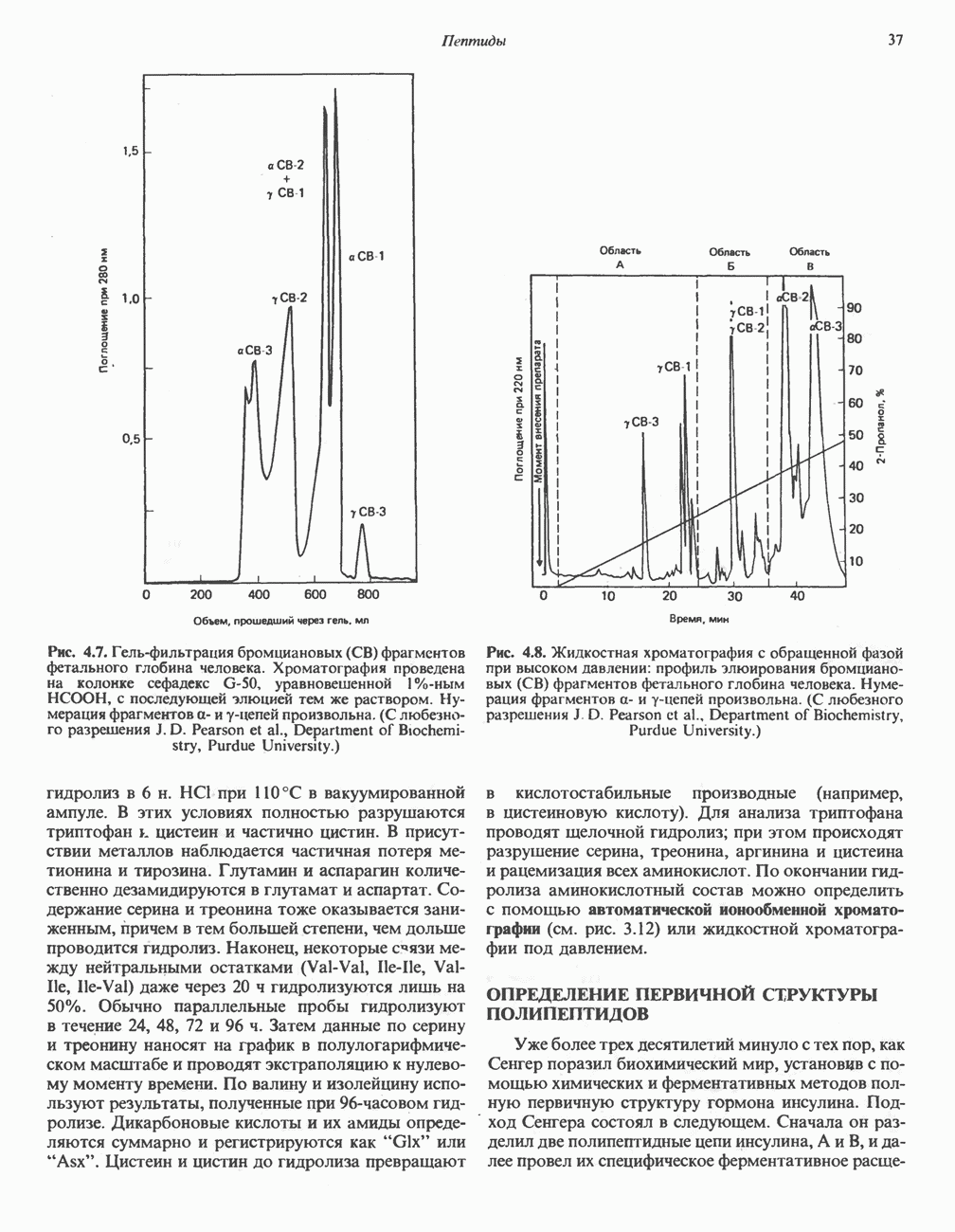
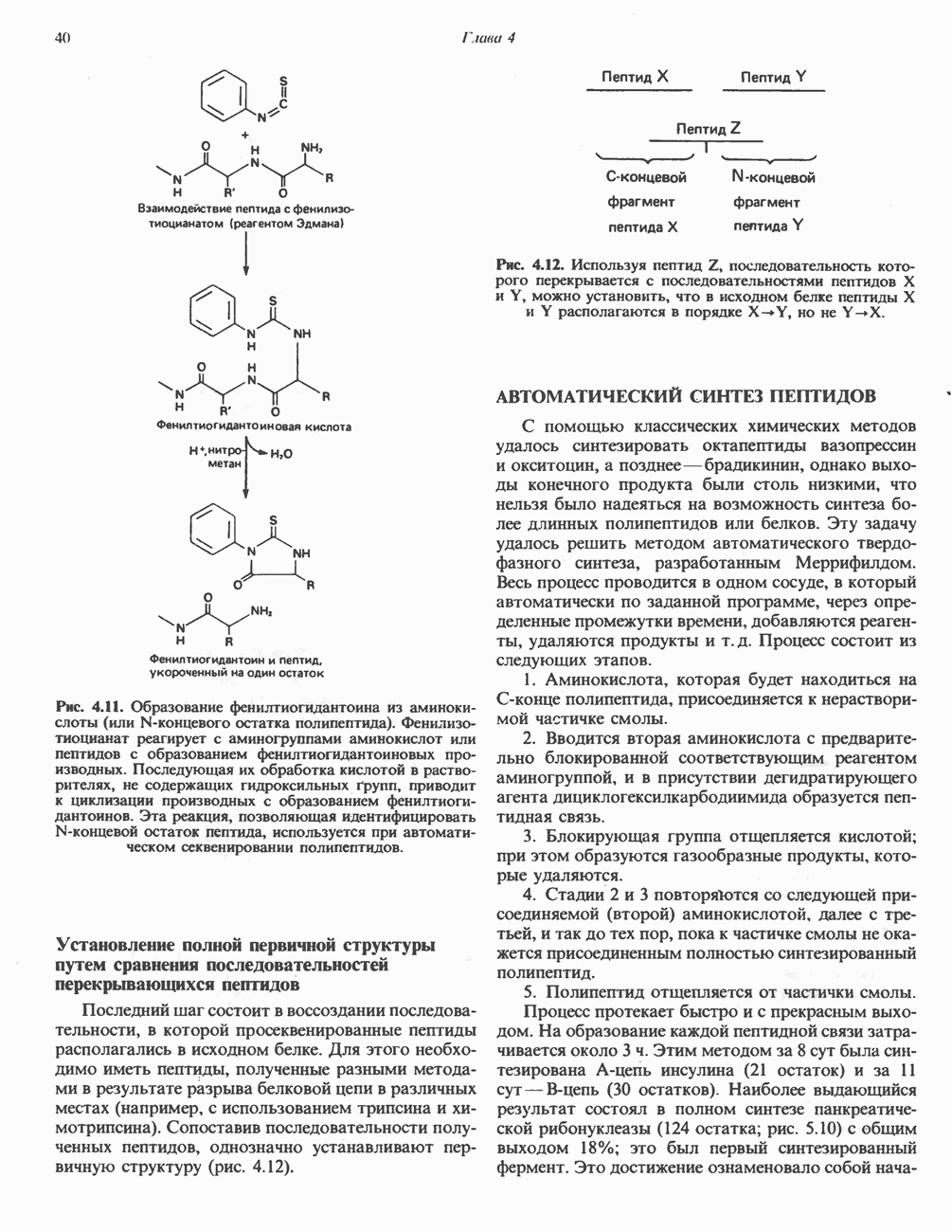
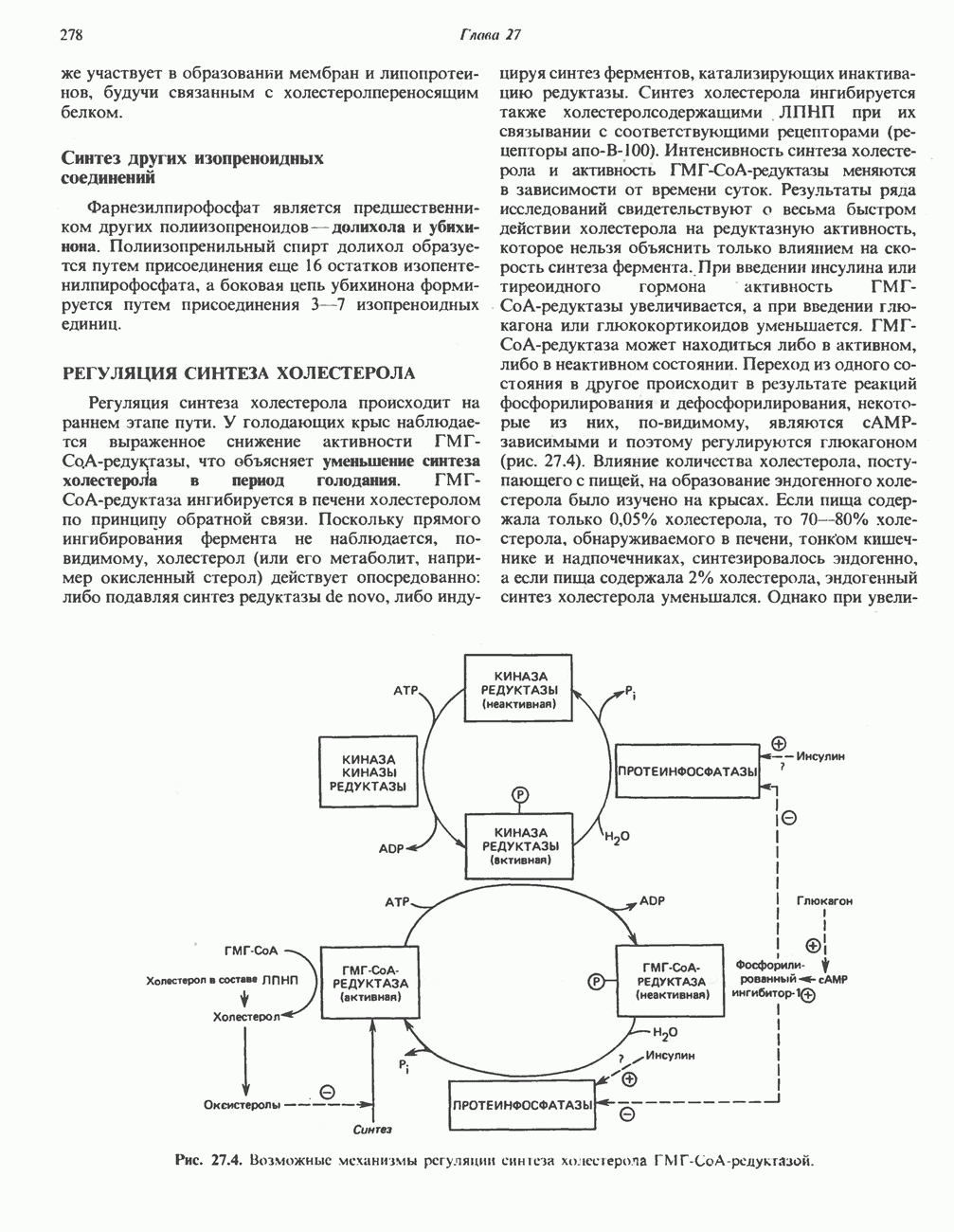
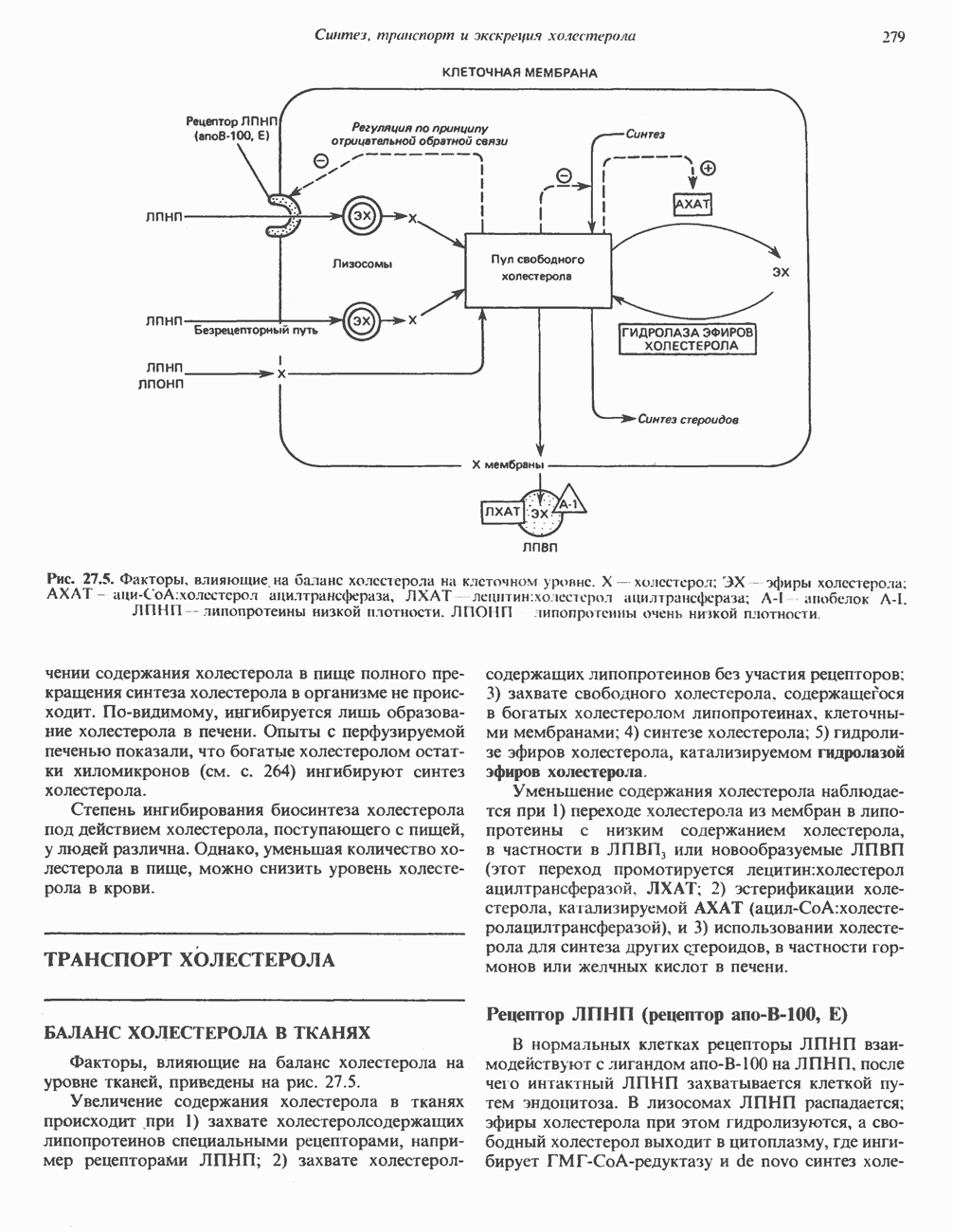
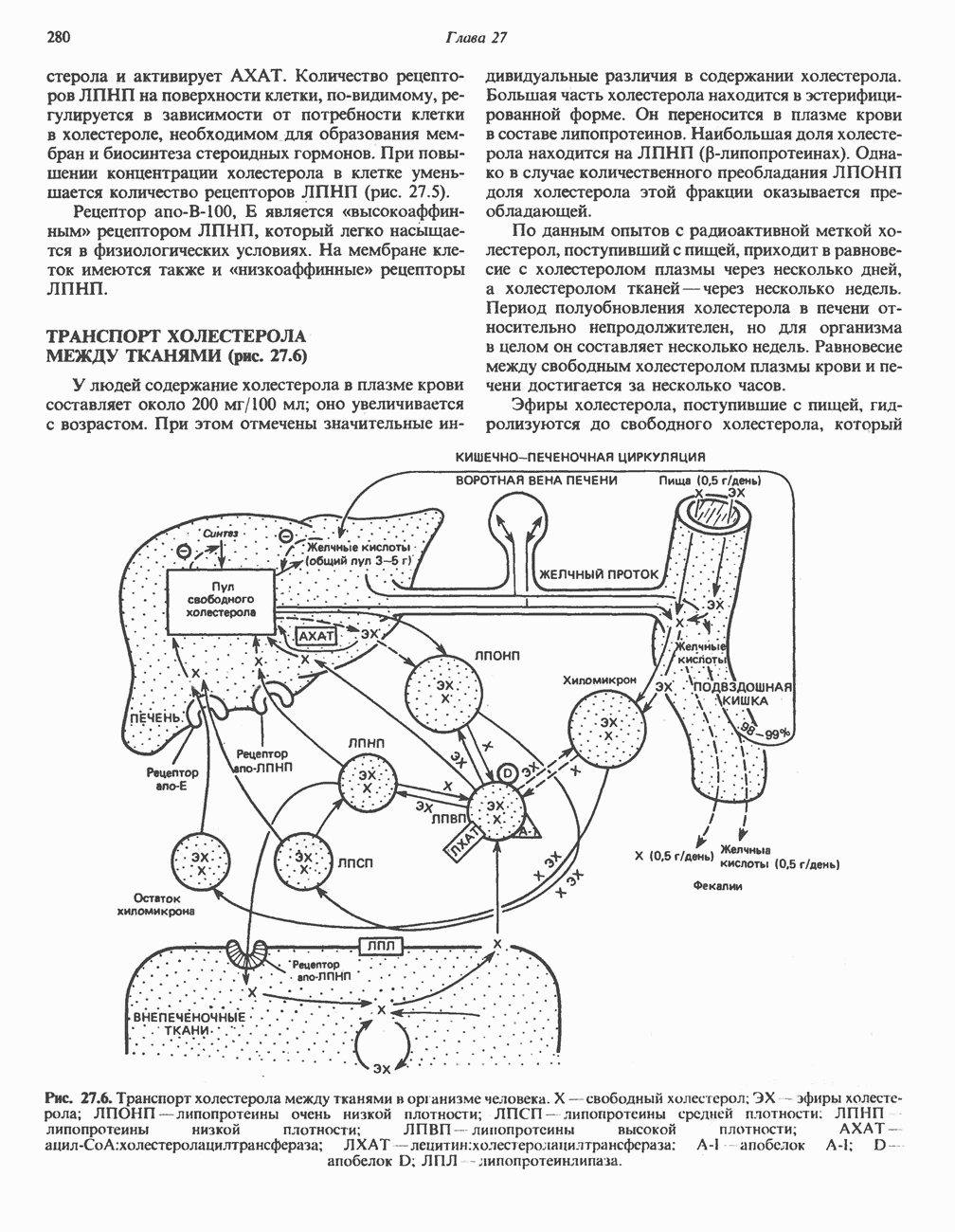
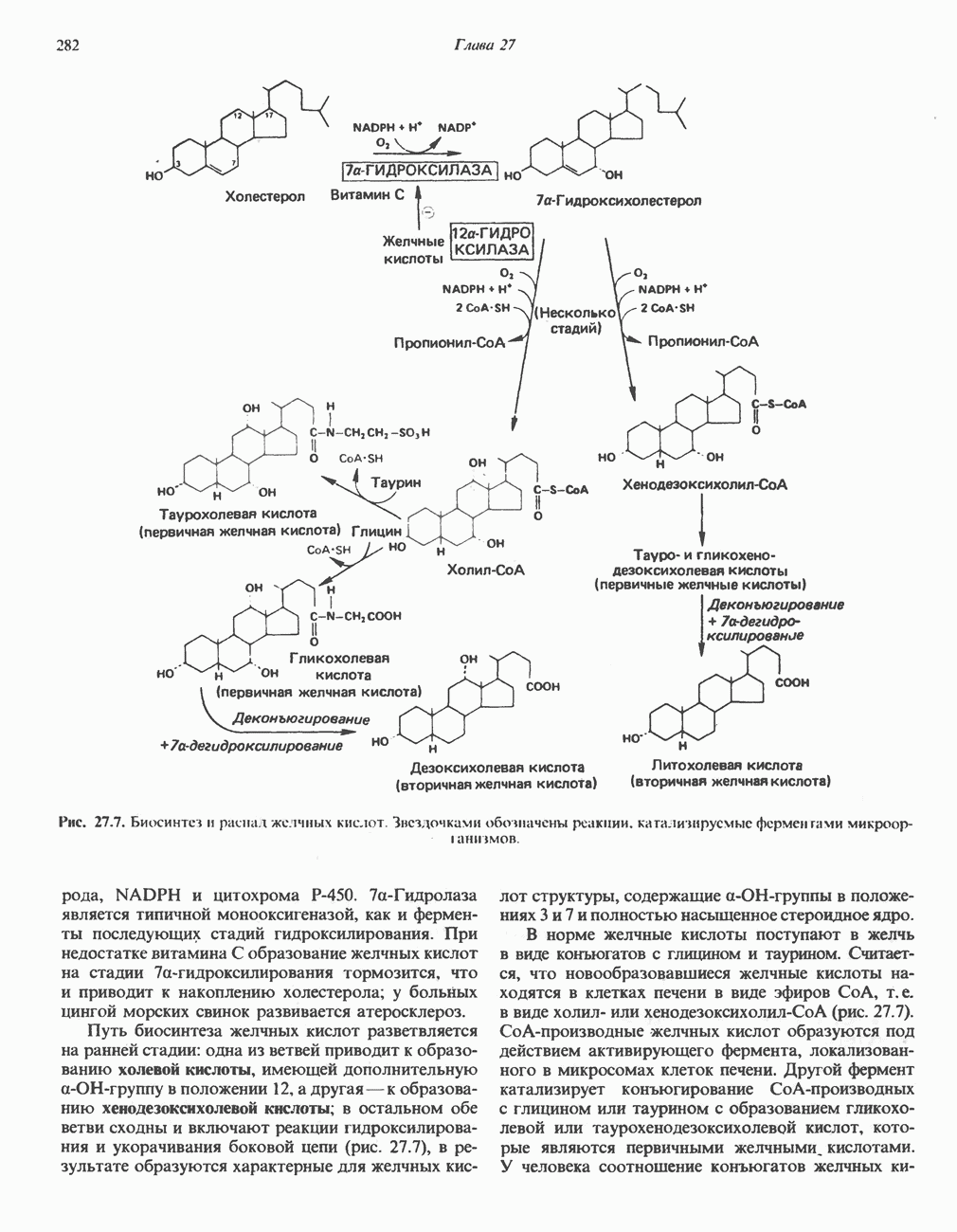
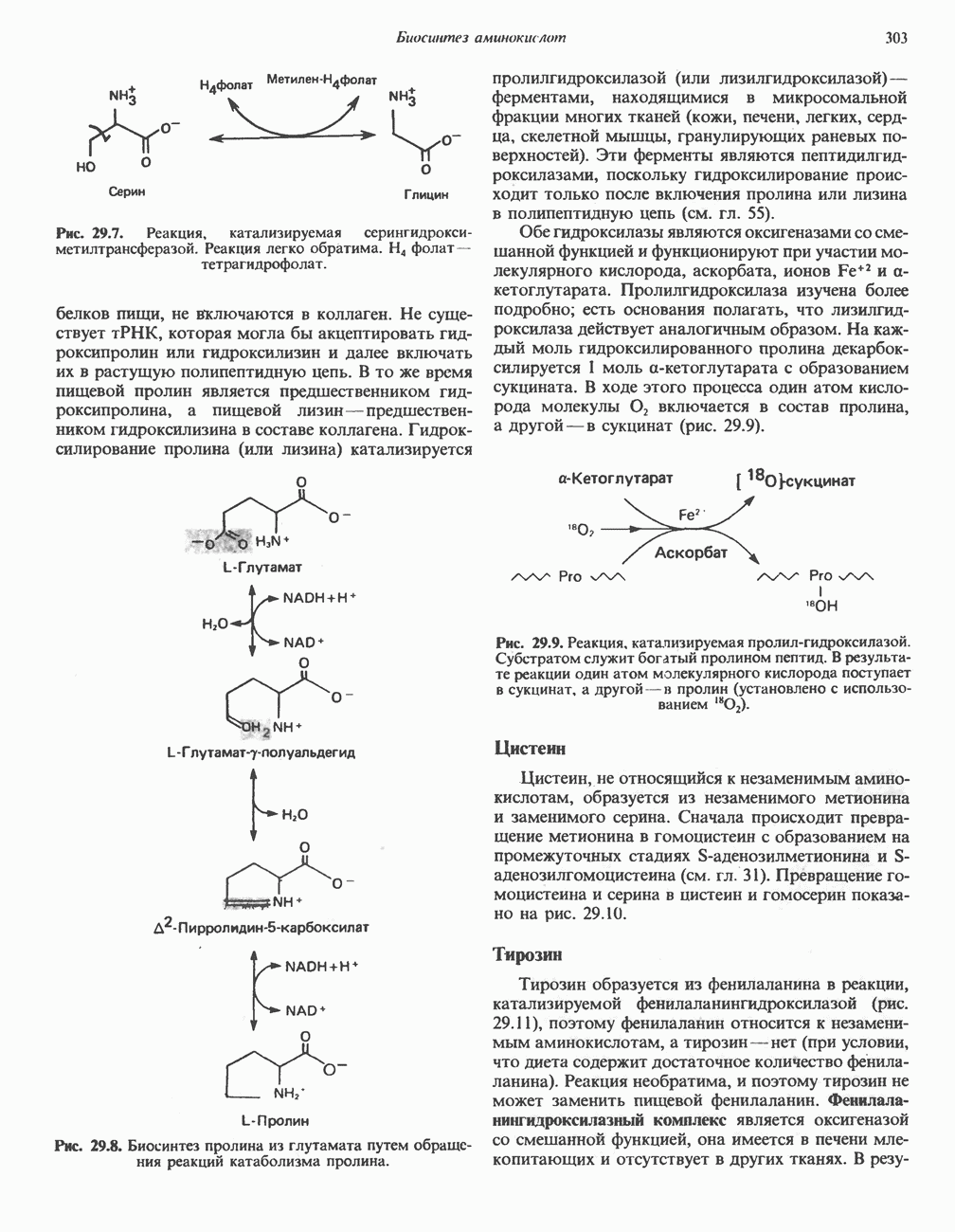
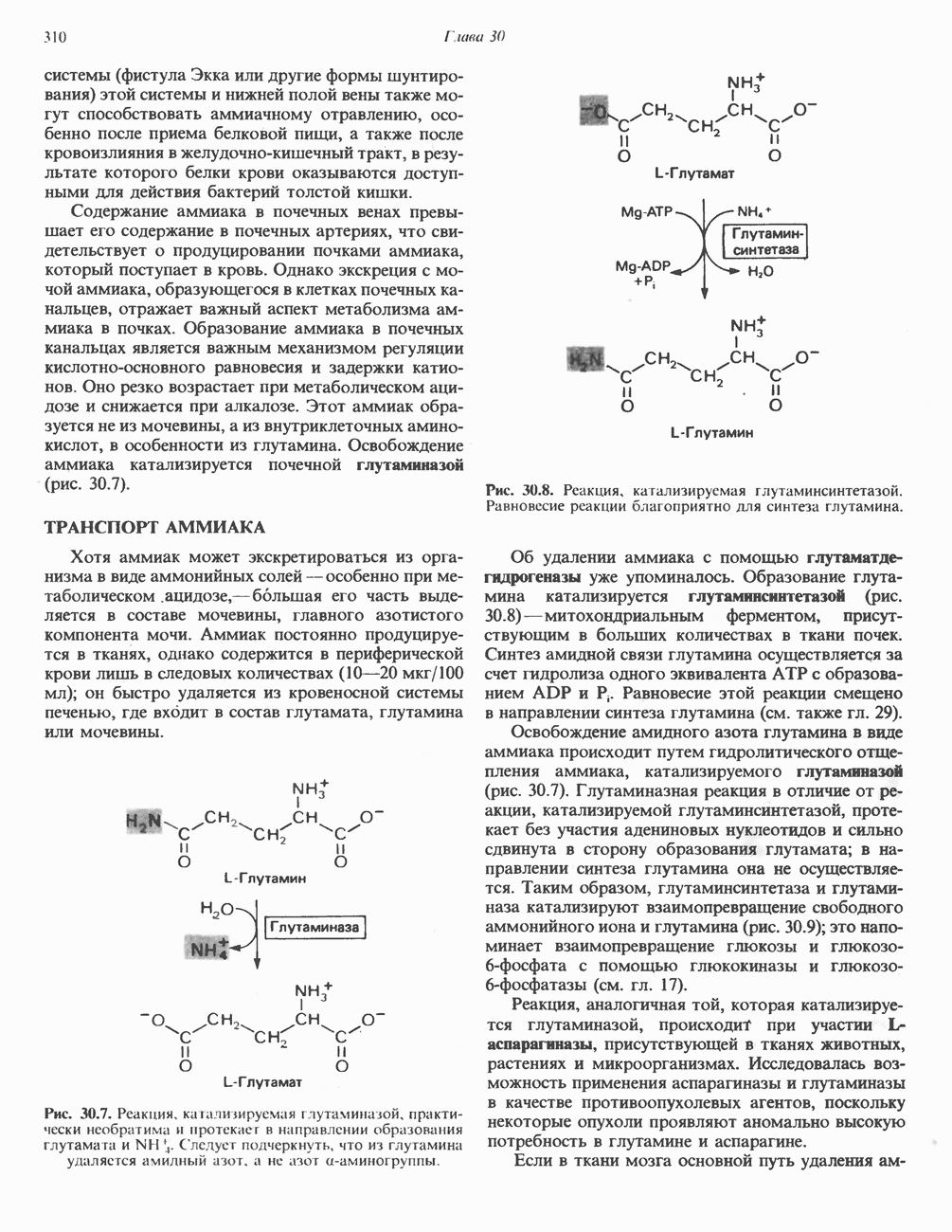
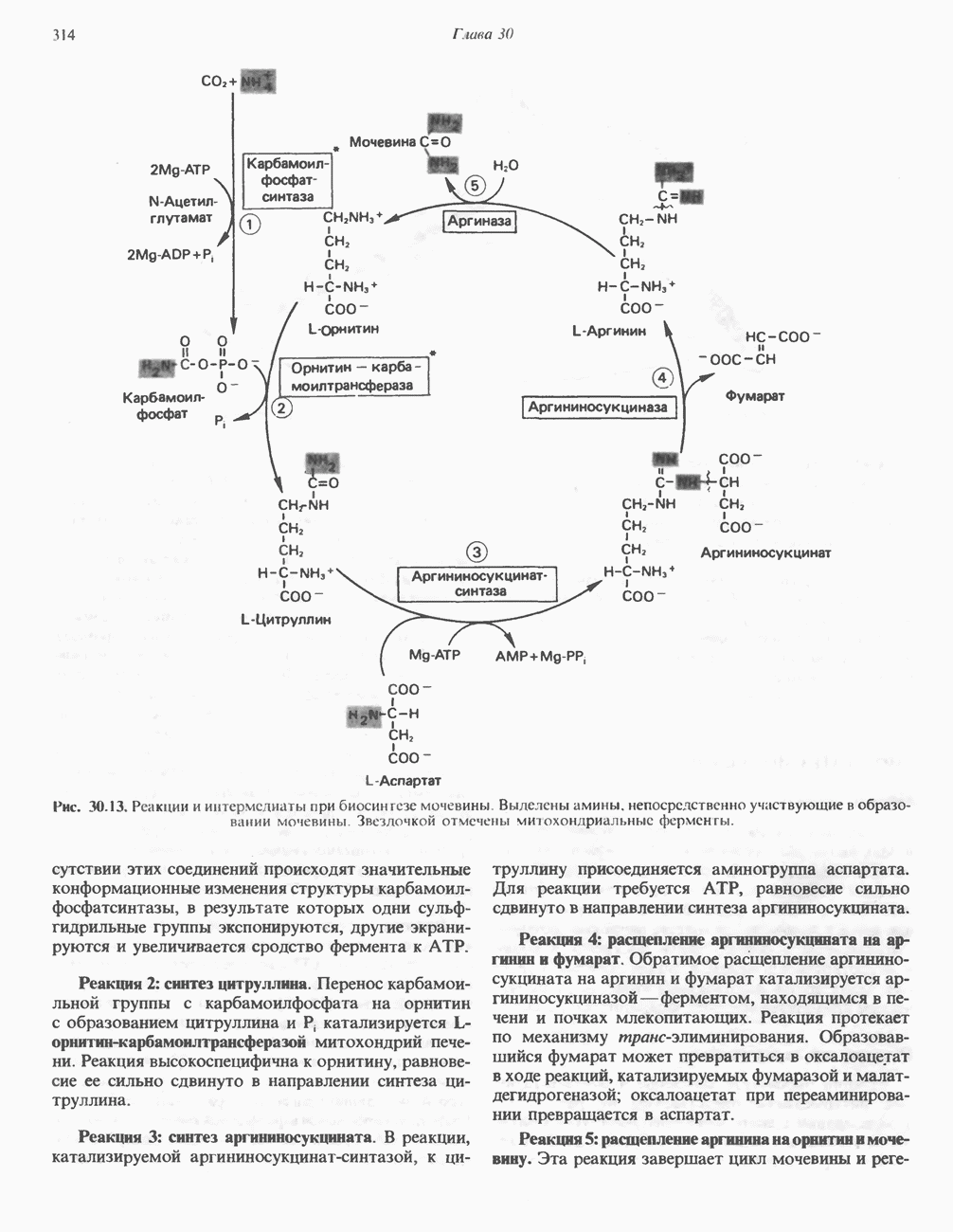
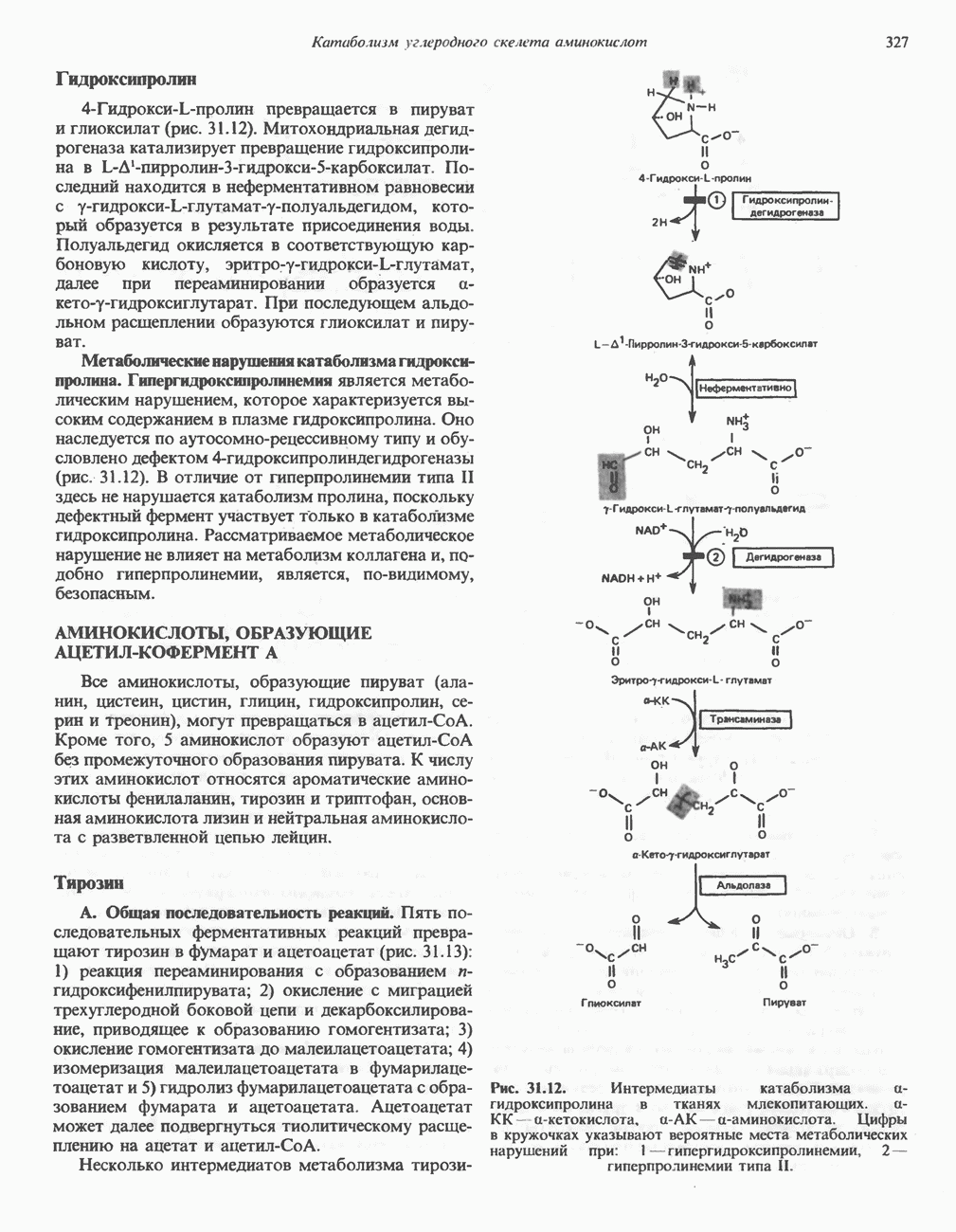
Пред.
След.
Макеты страниц
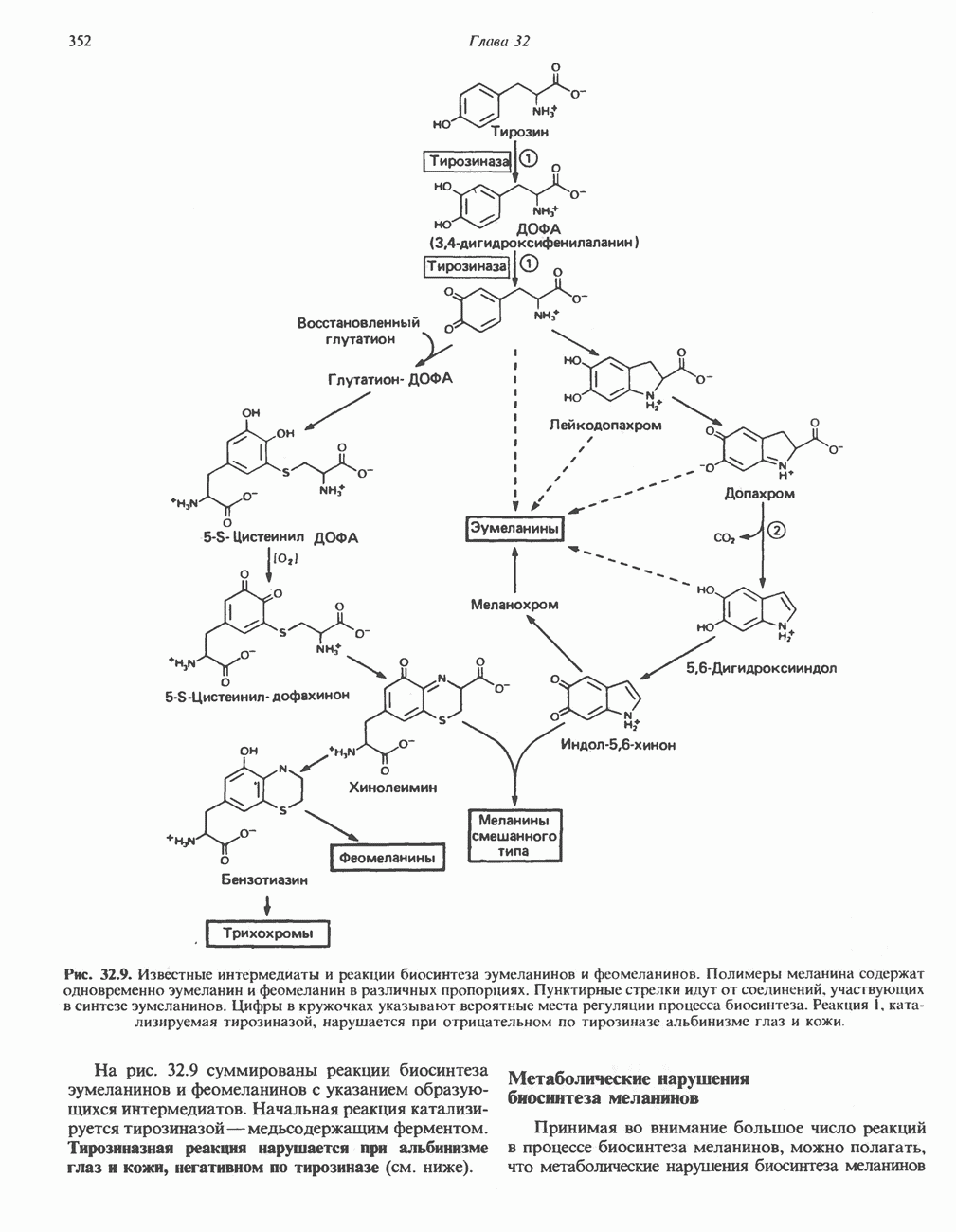
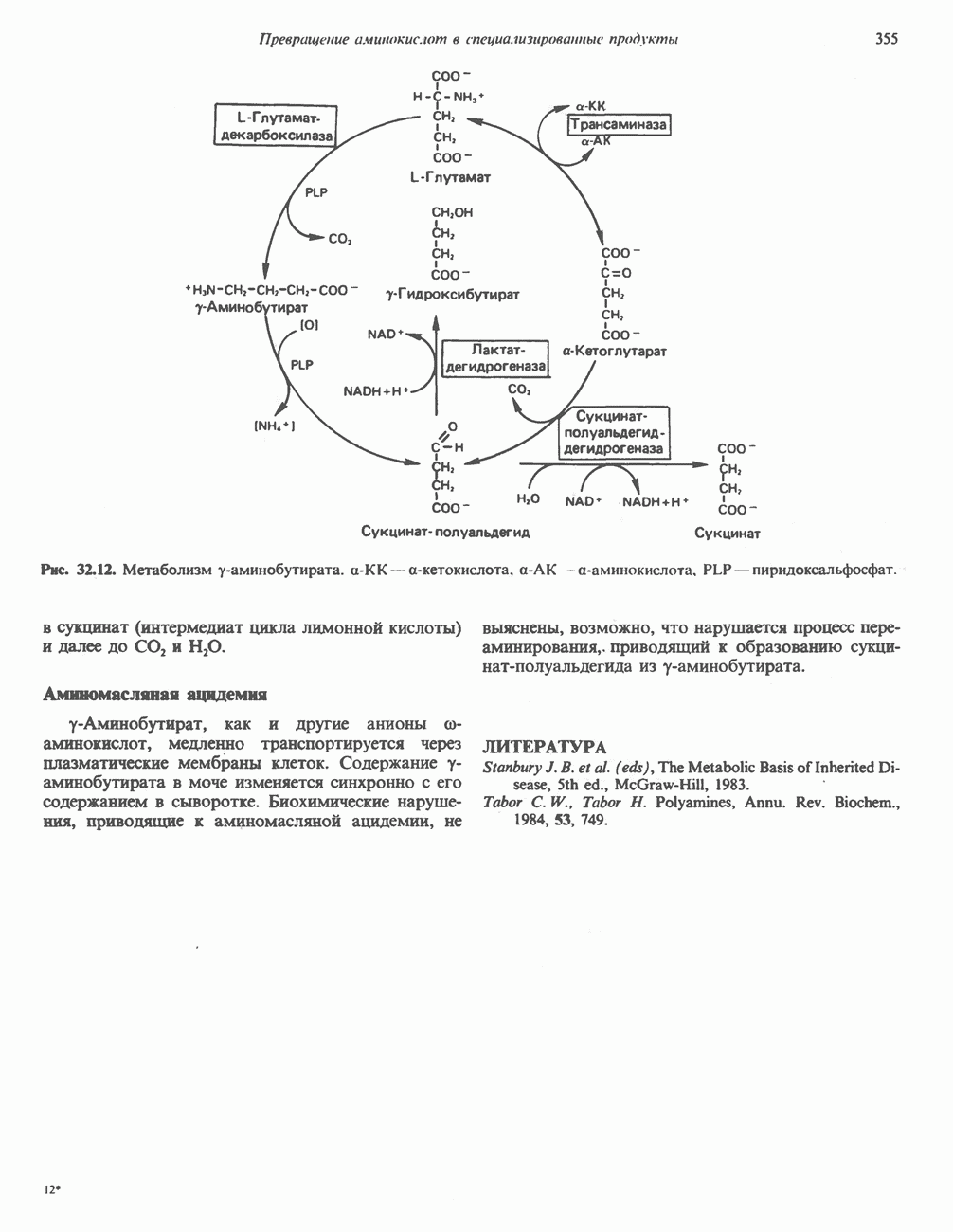
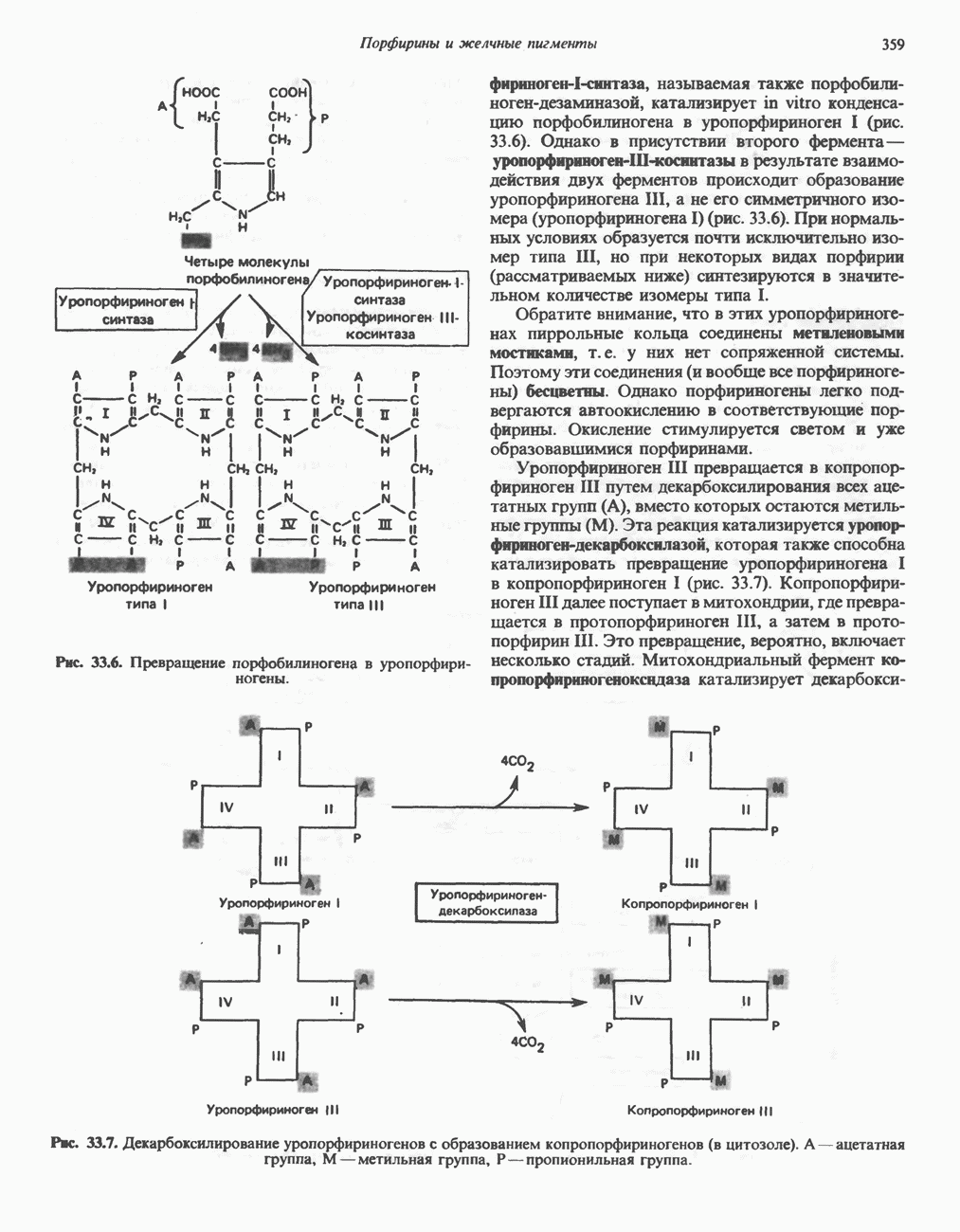
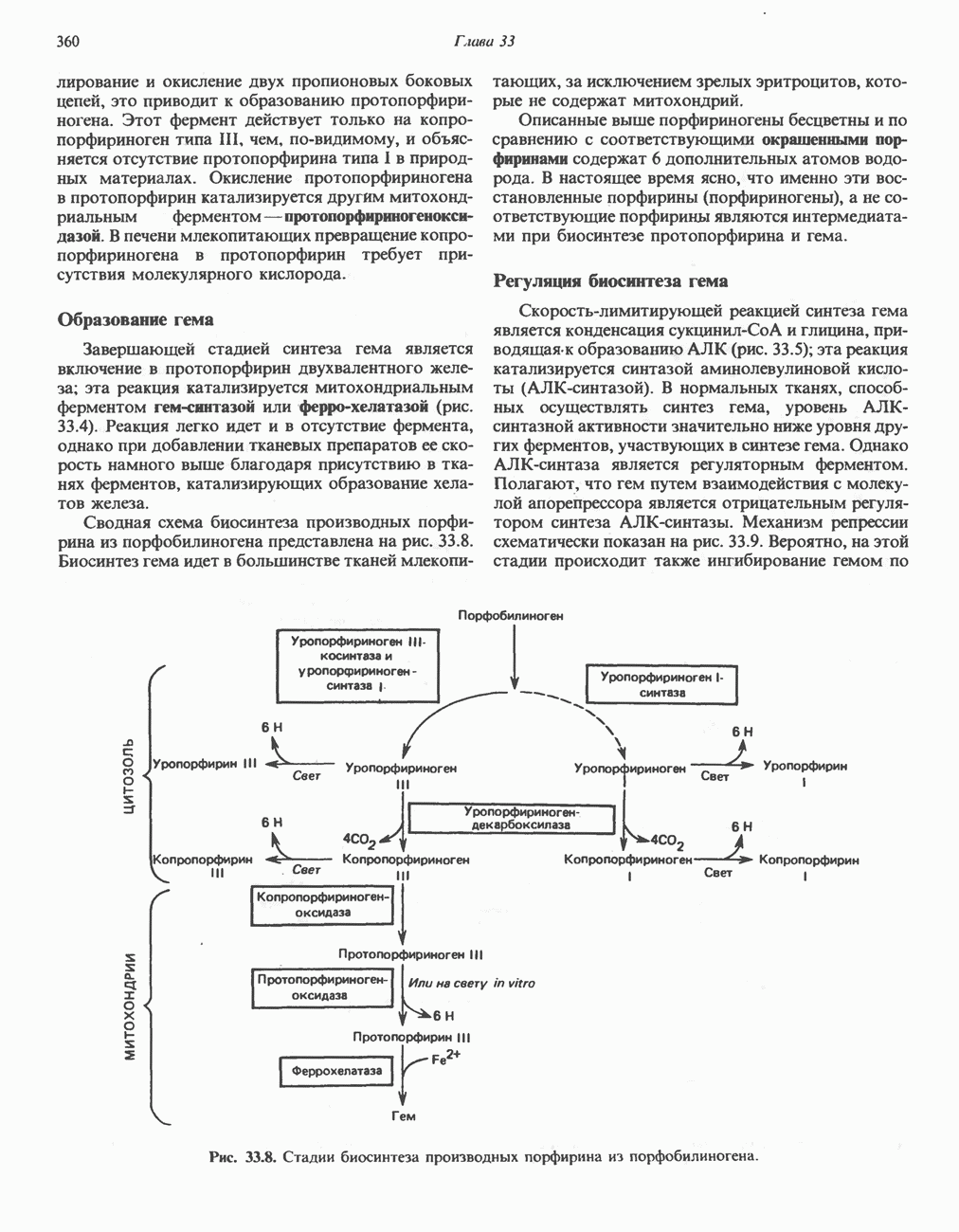
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
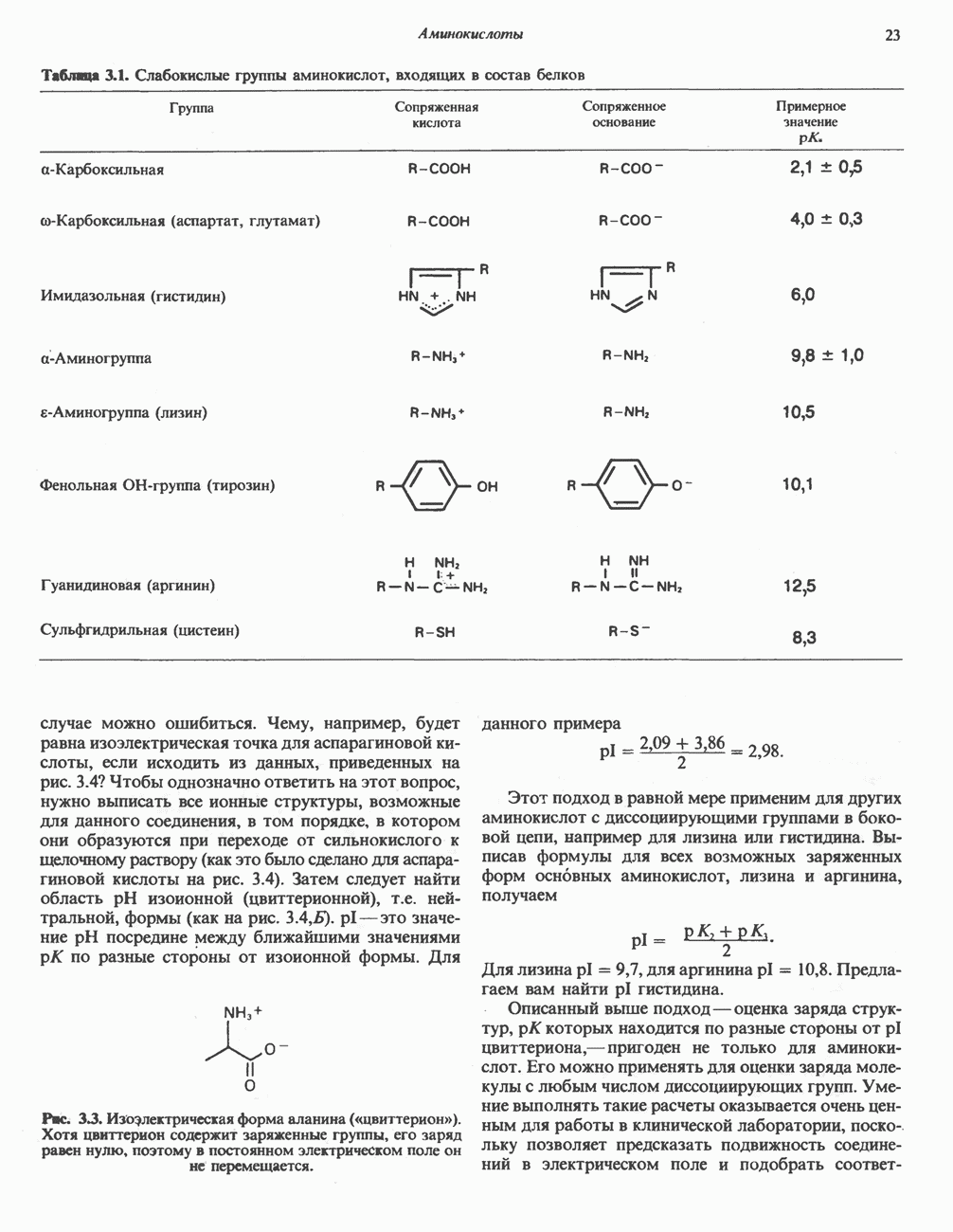
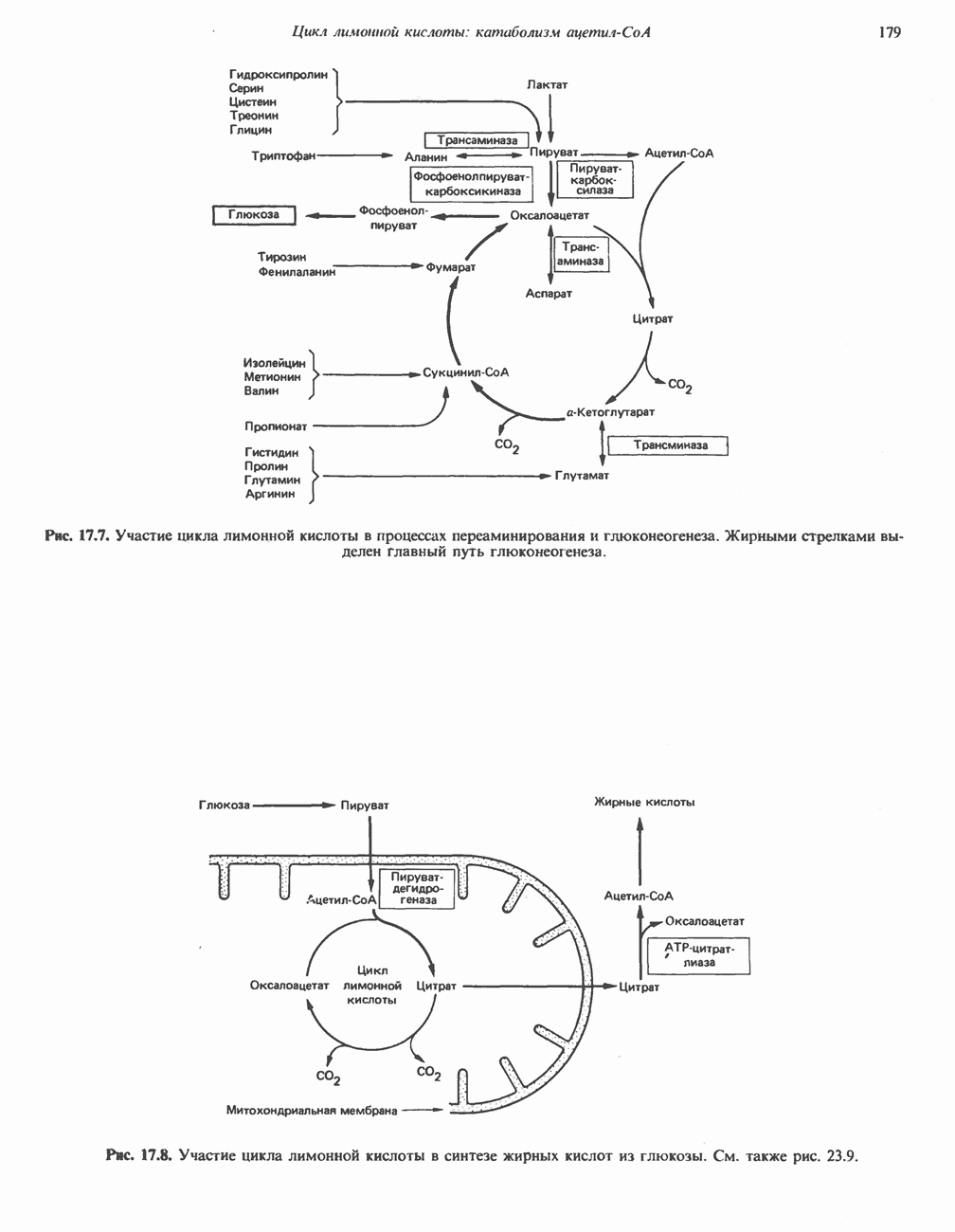
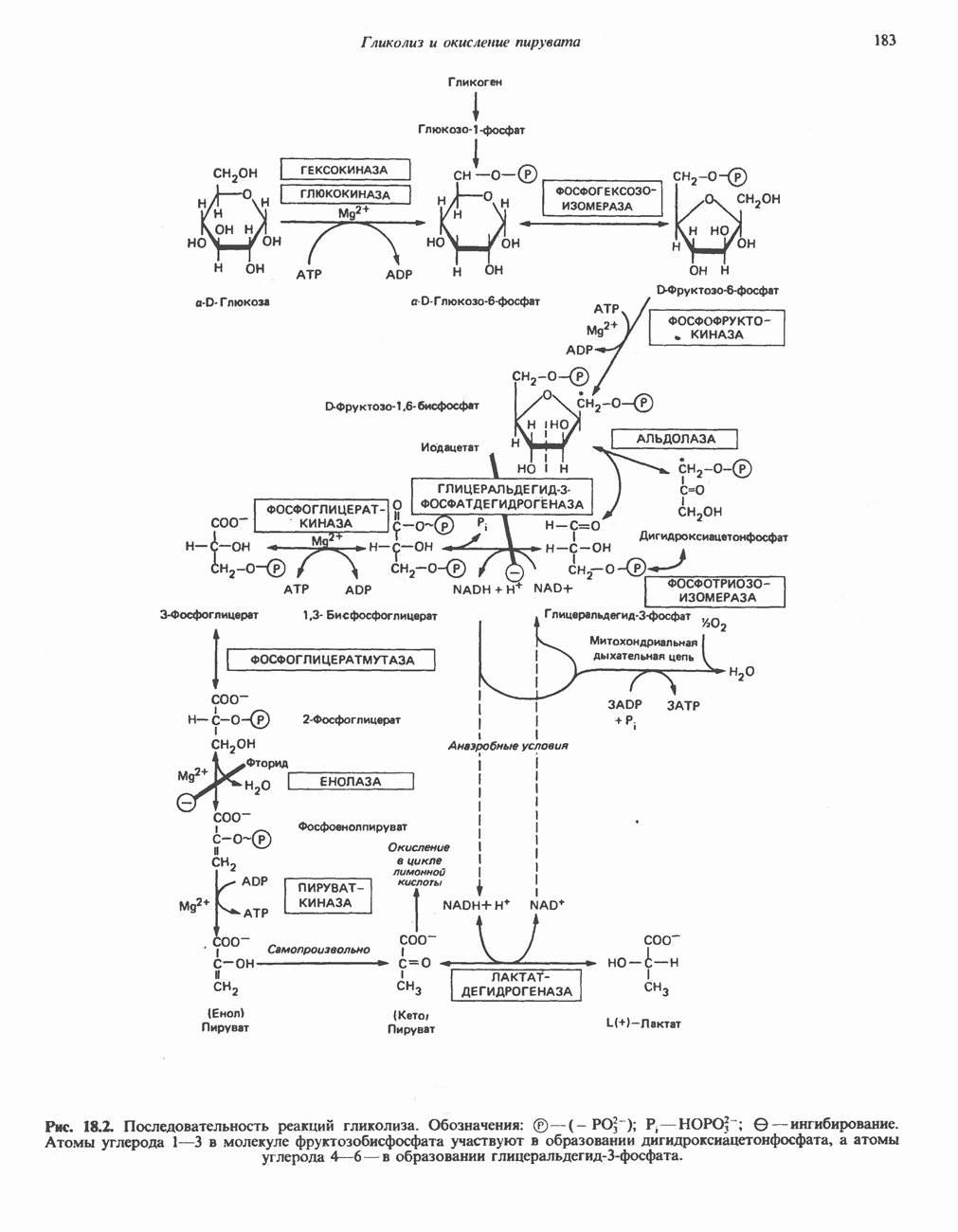
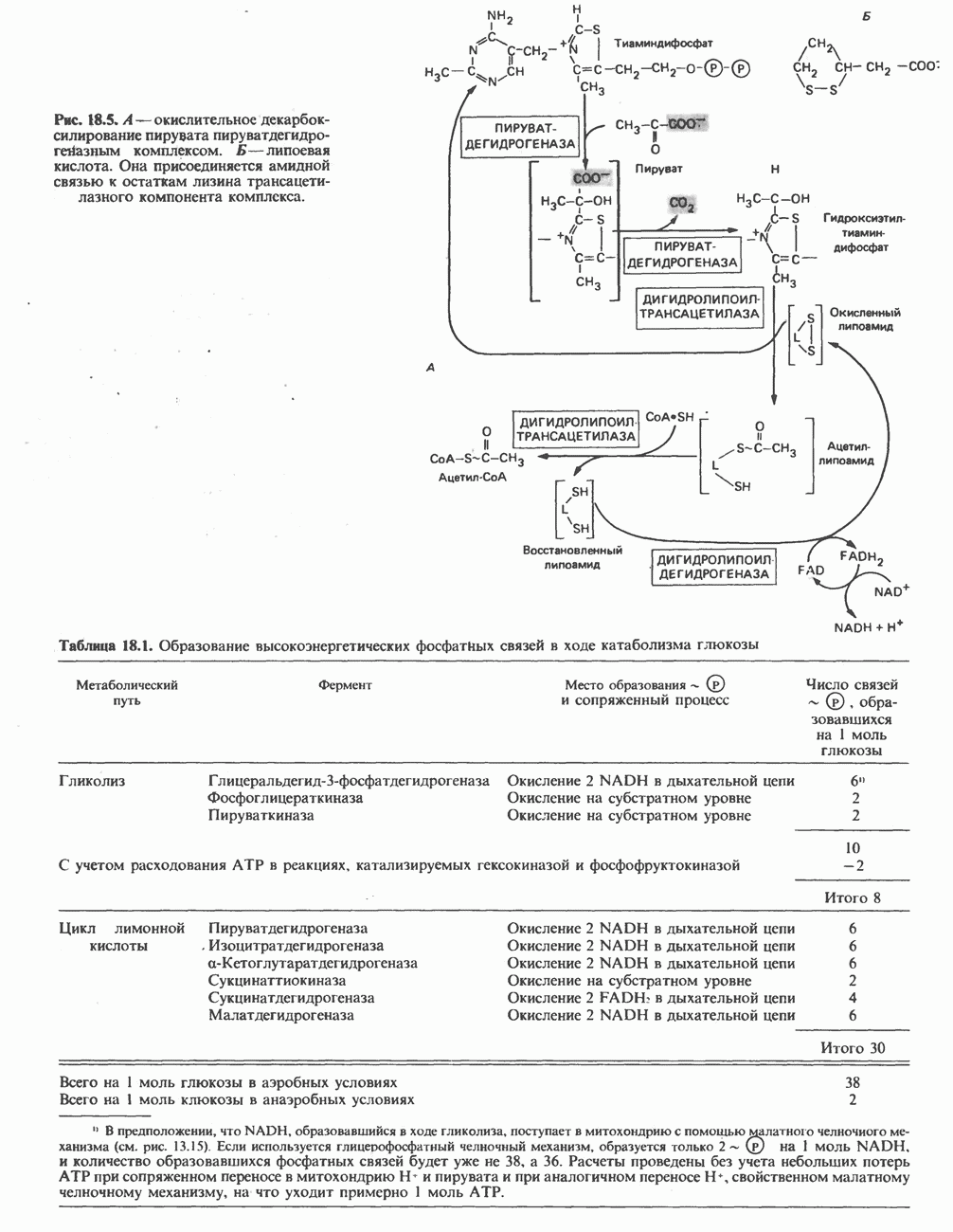
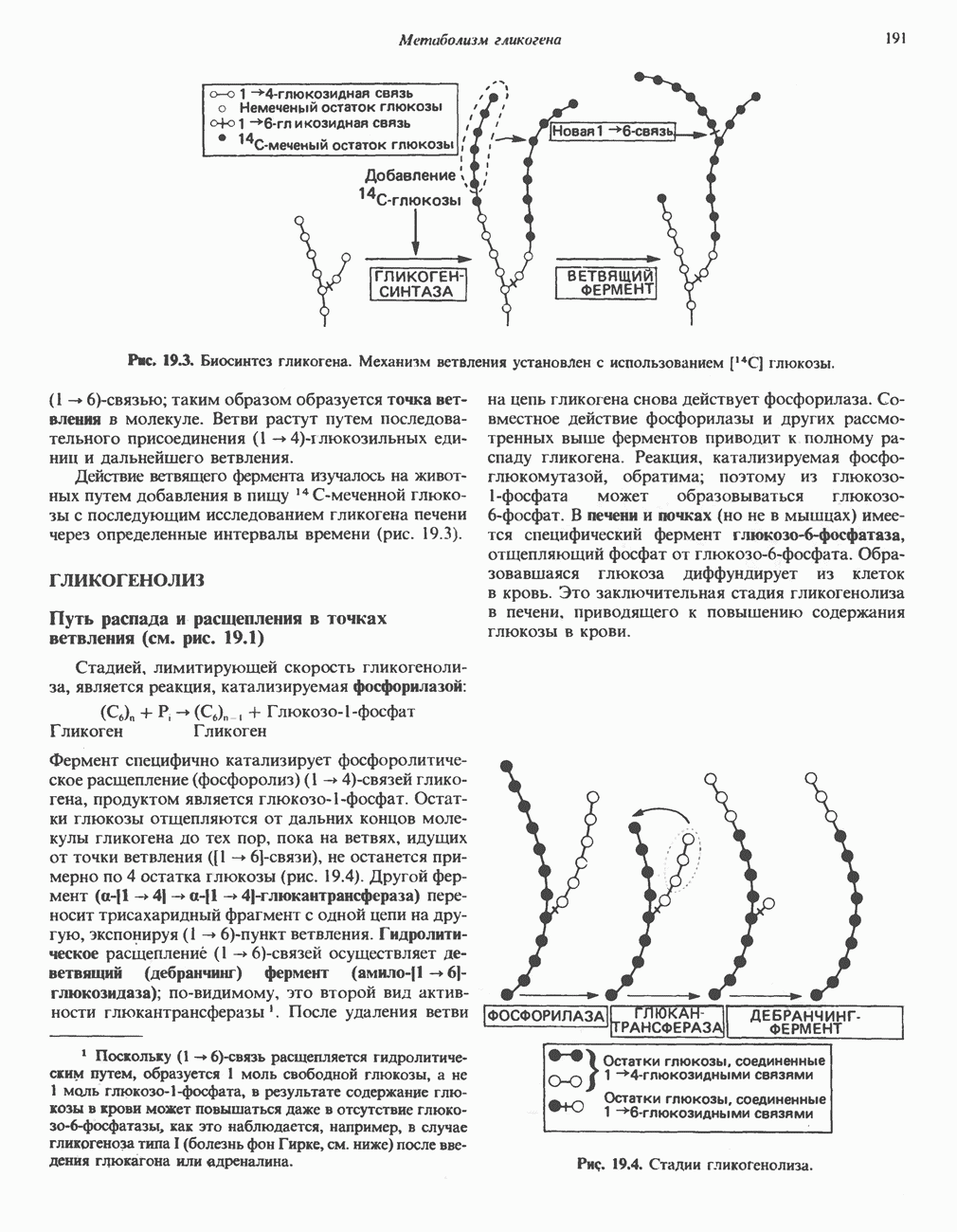
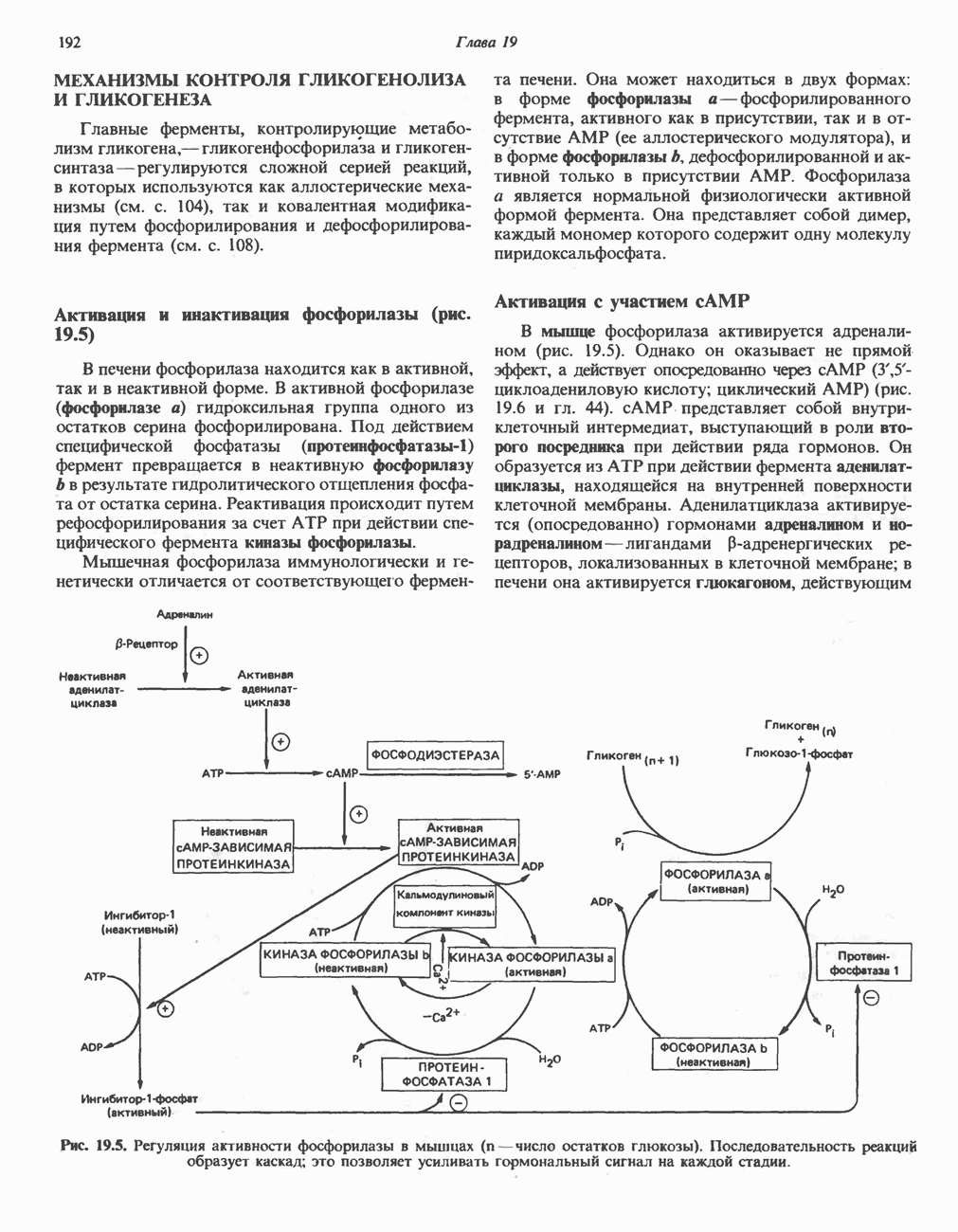
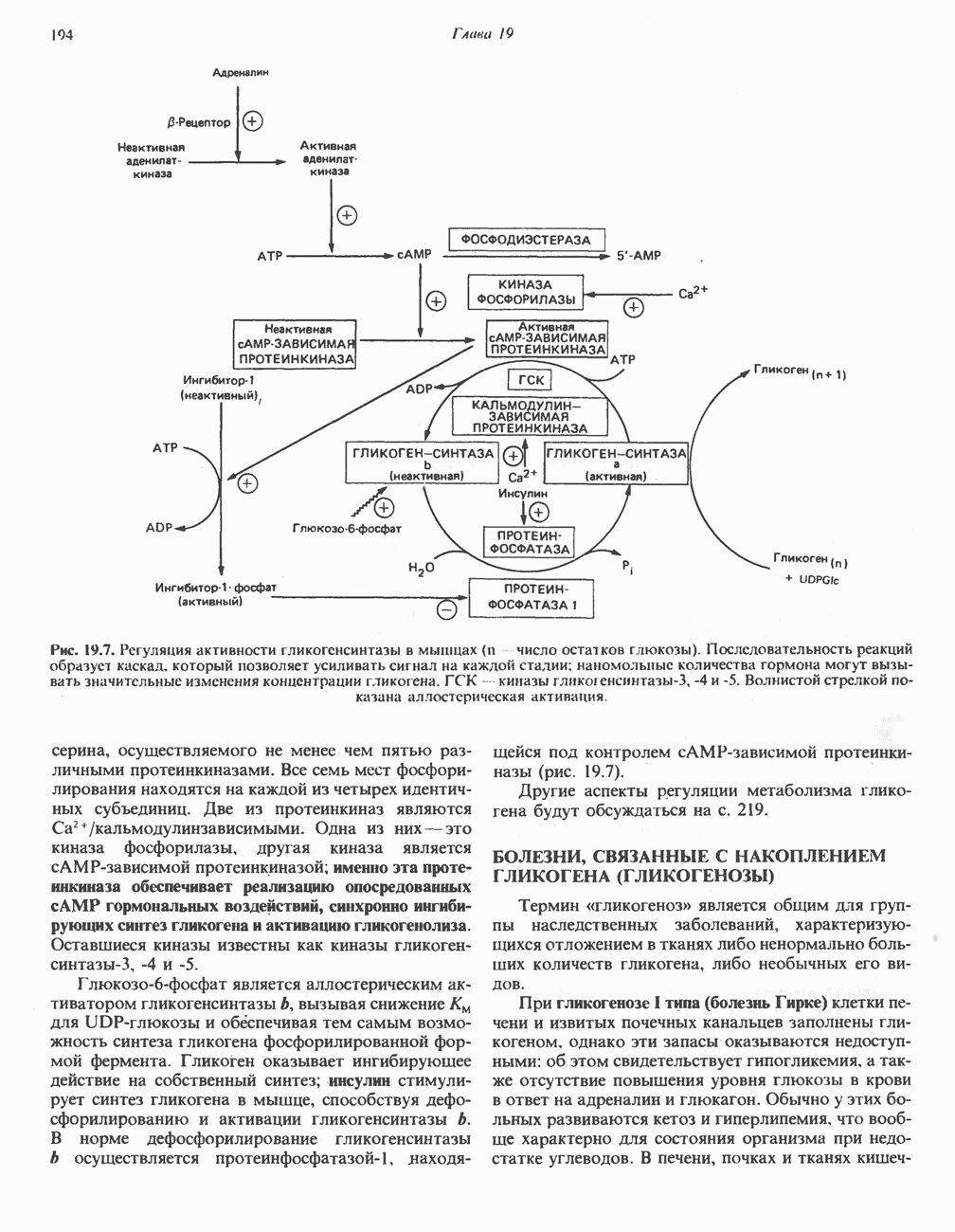
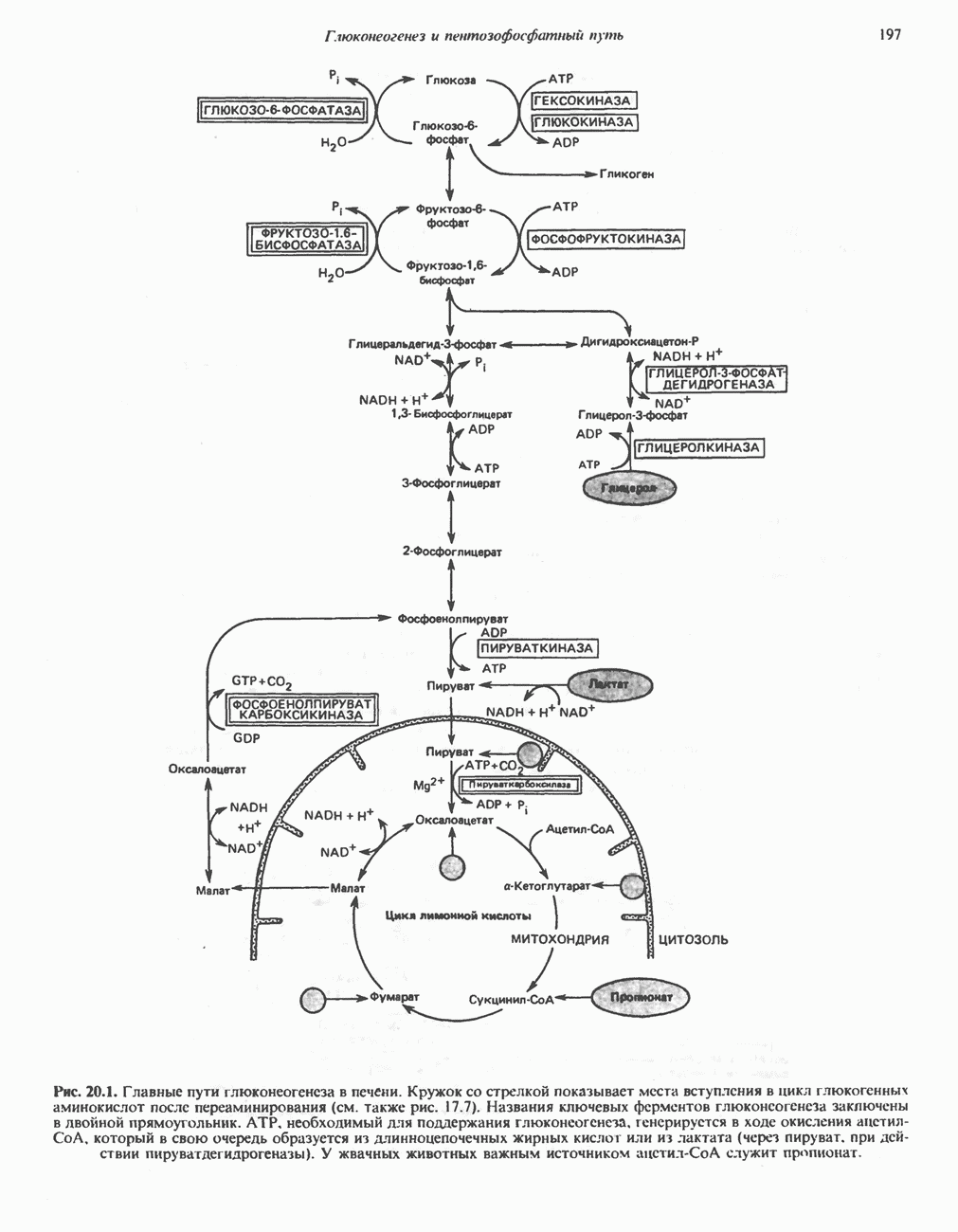
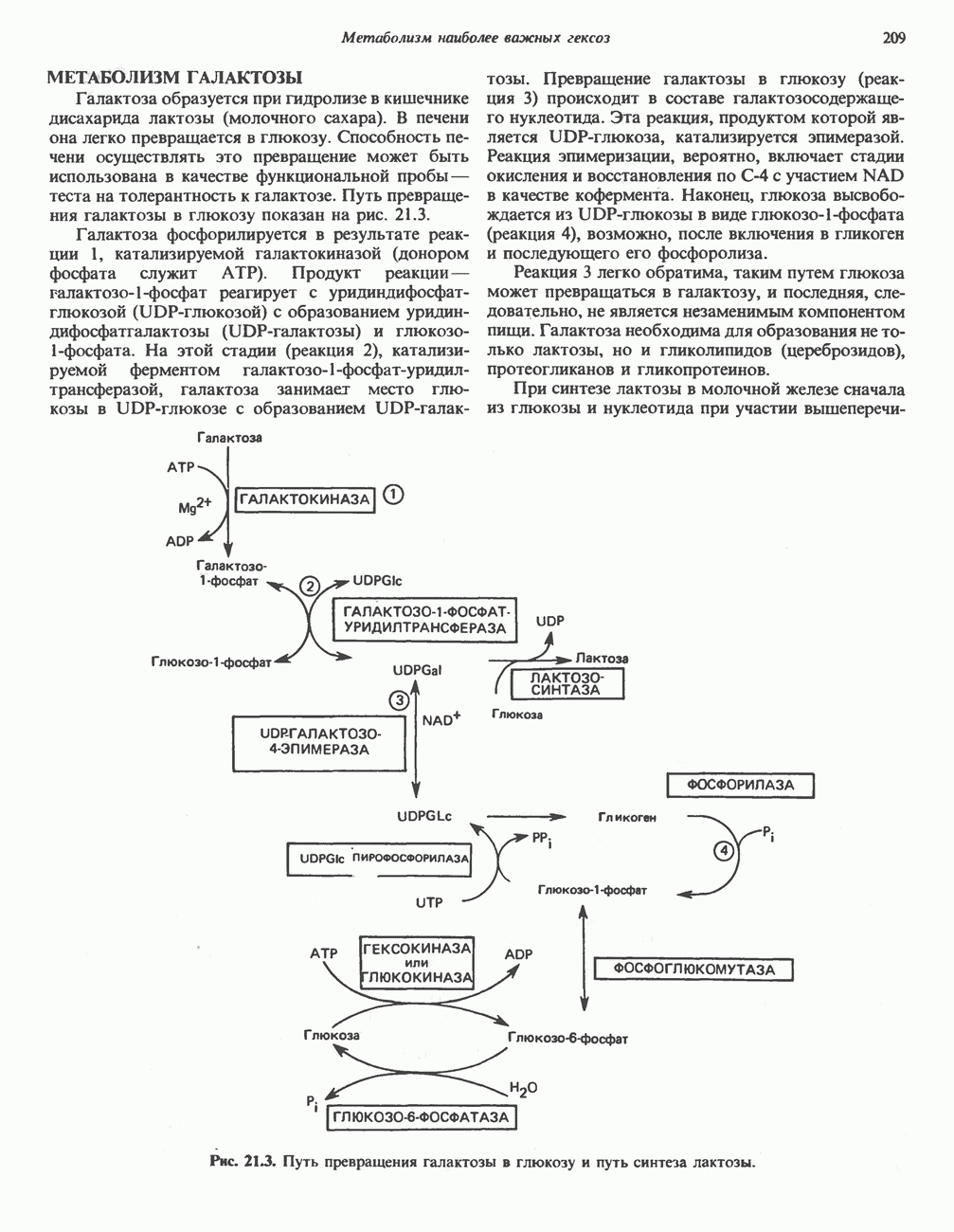
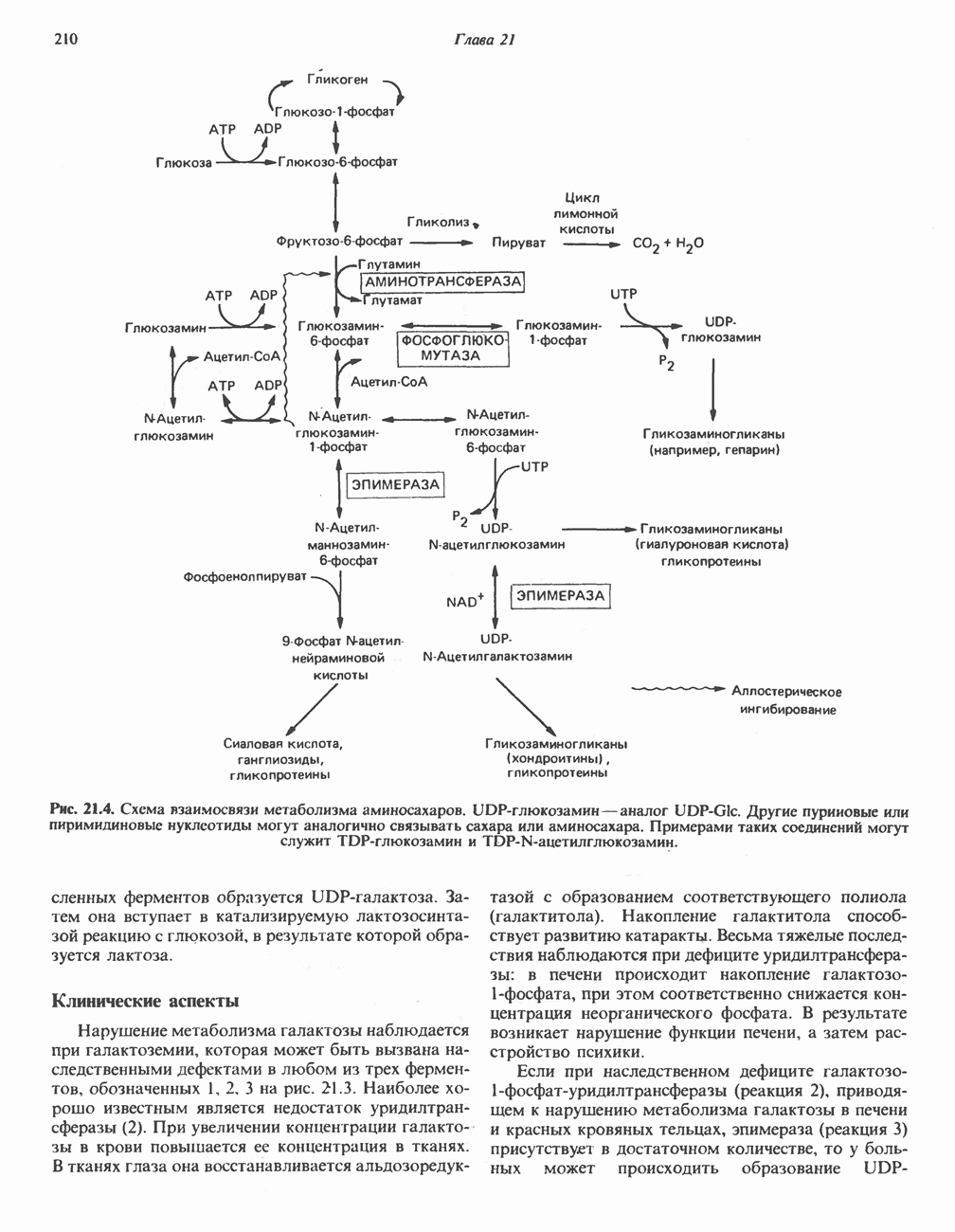
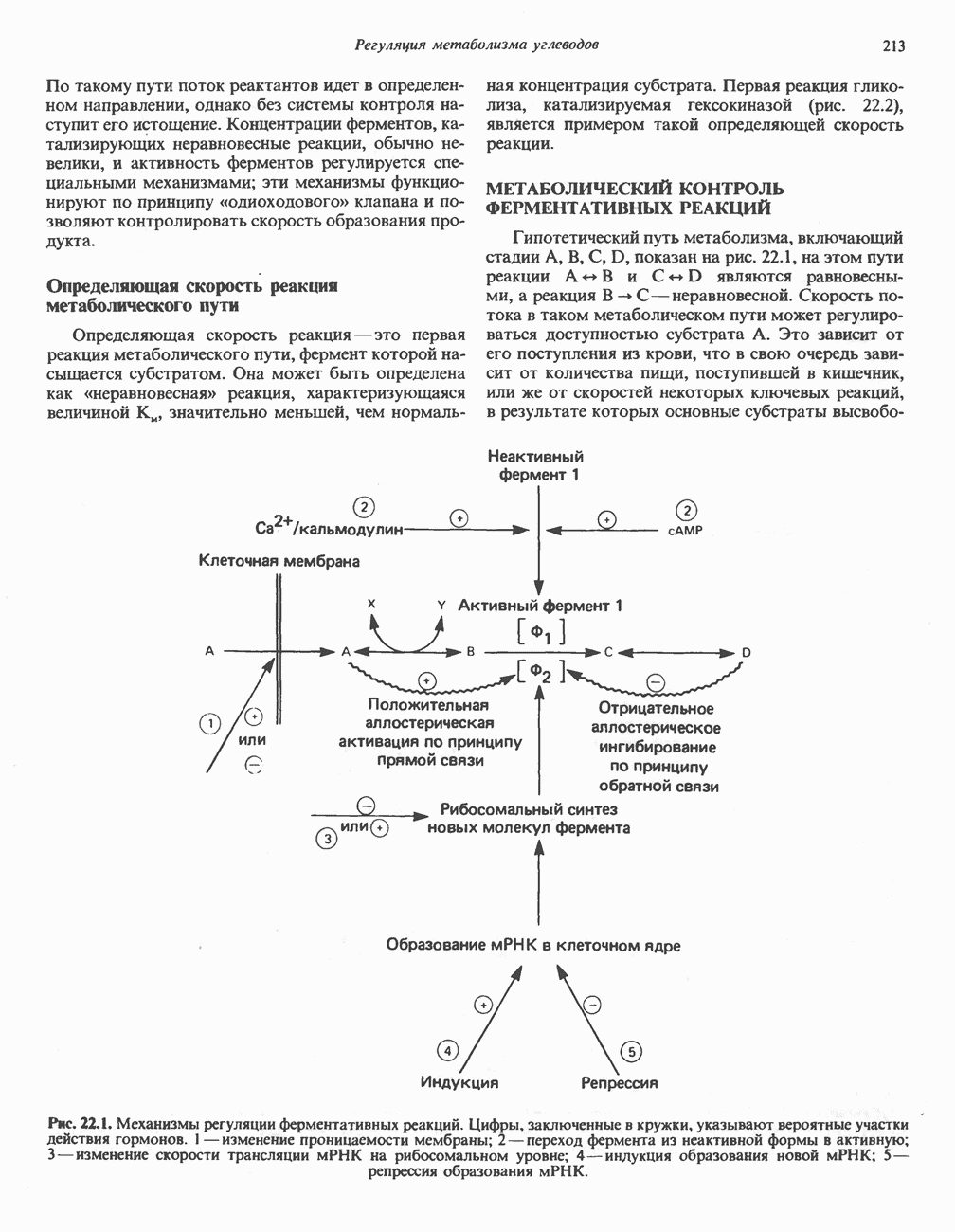
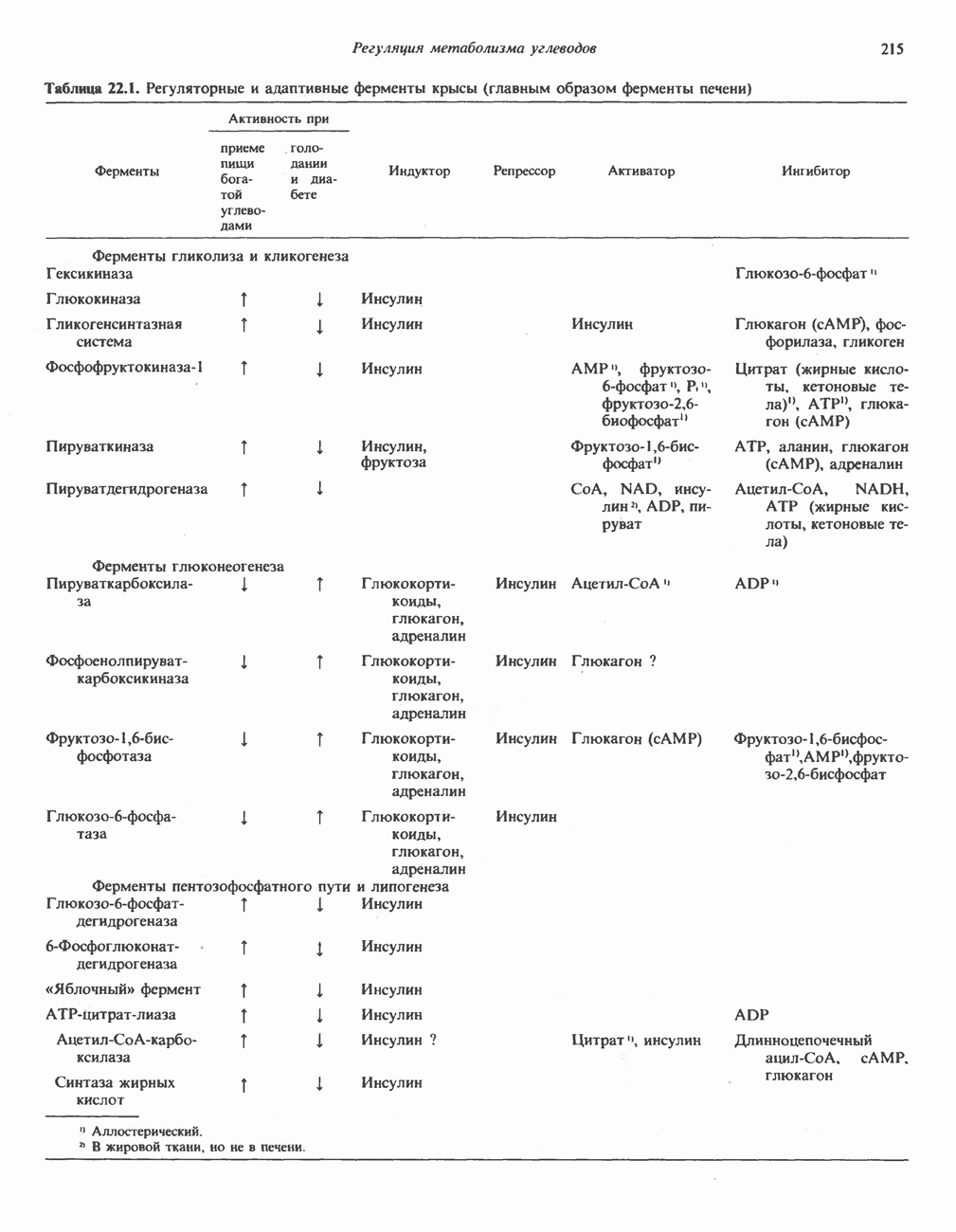
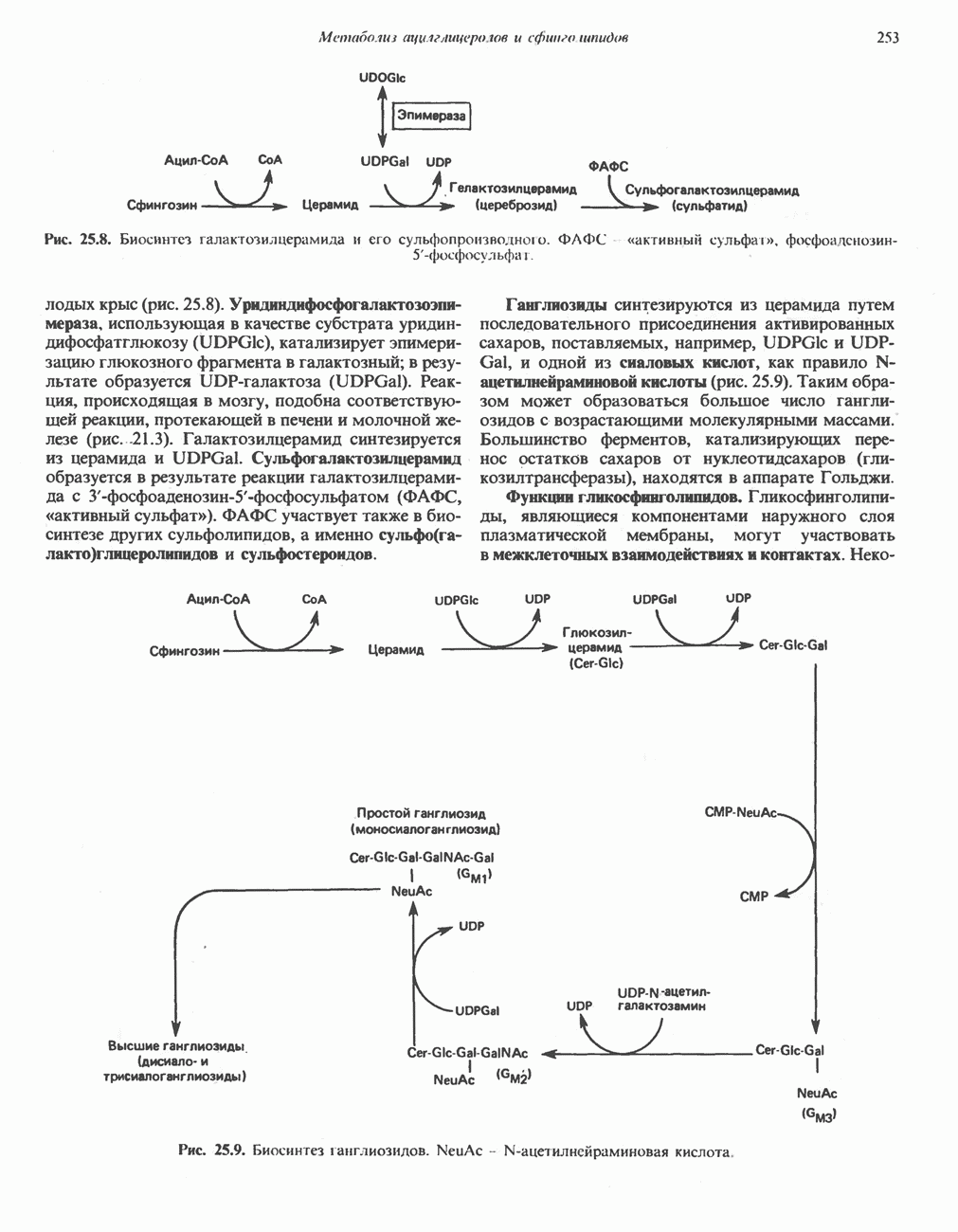
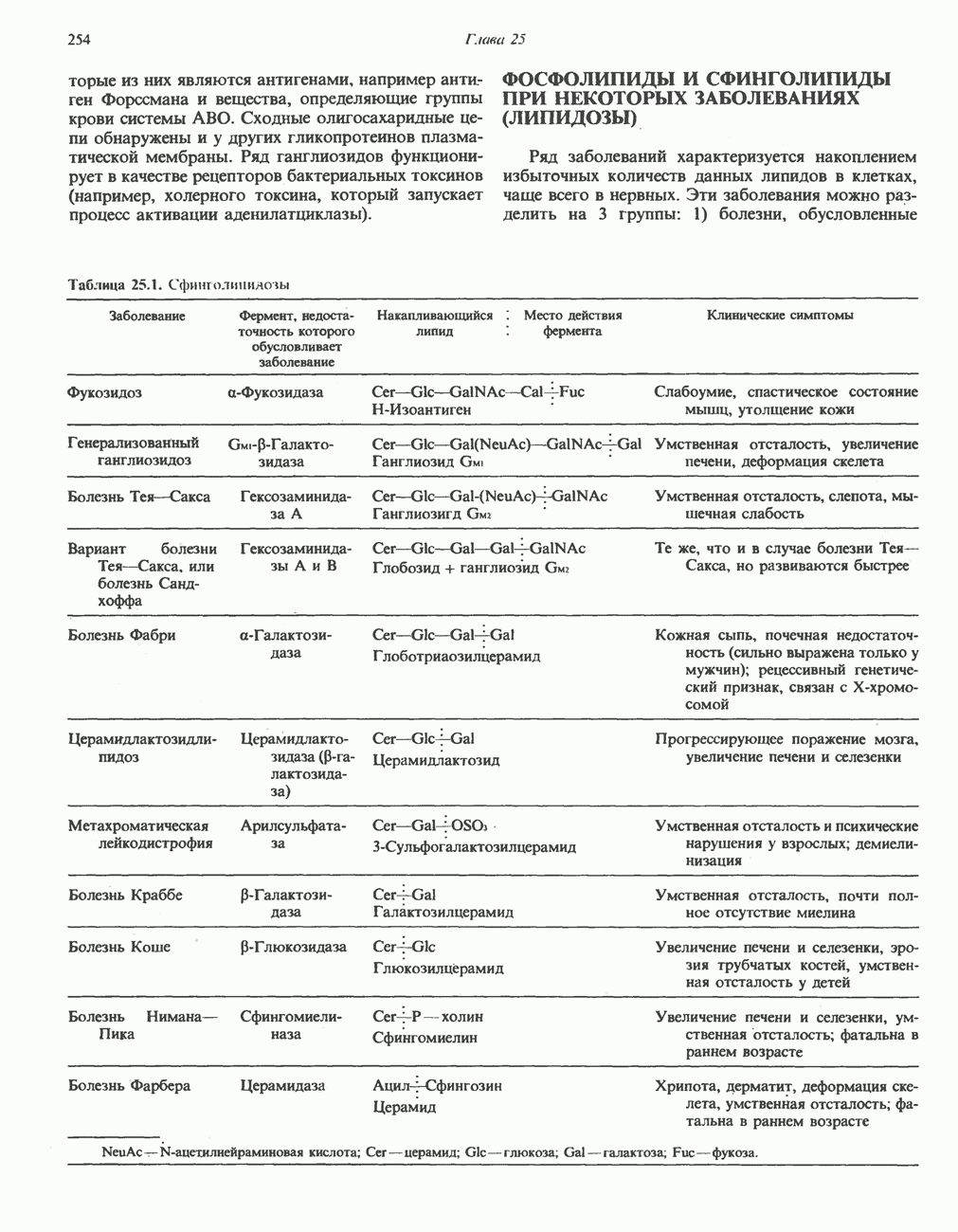
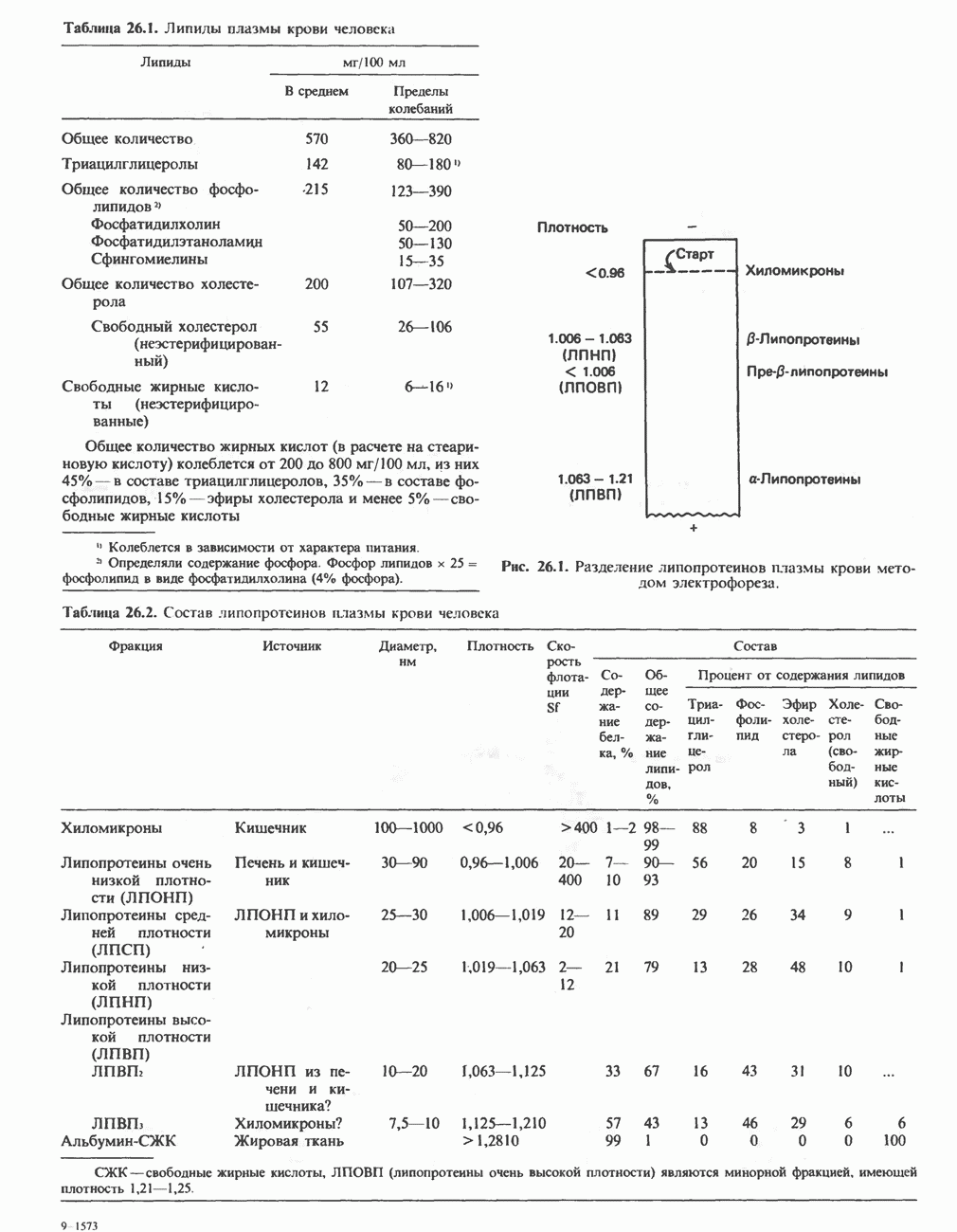
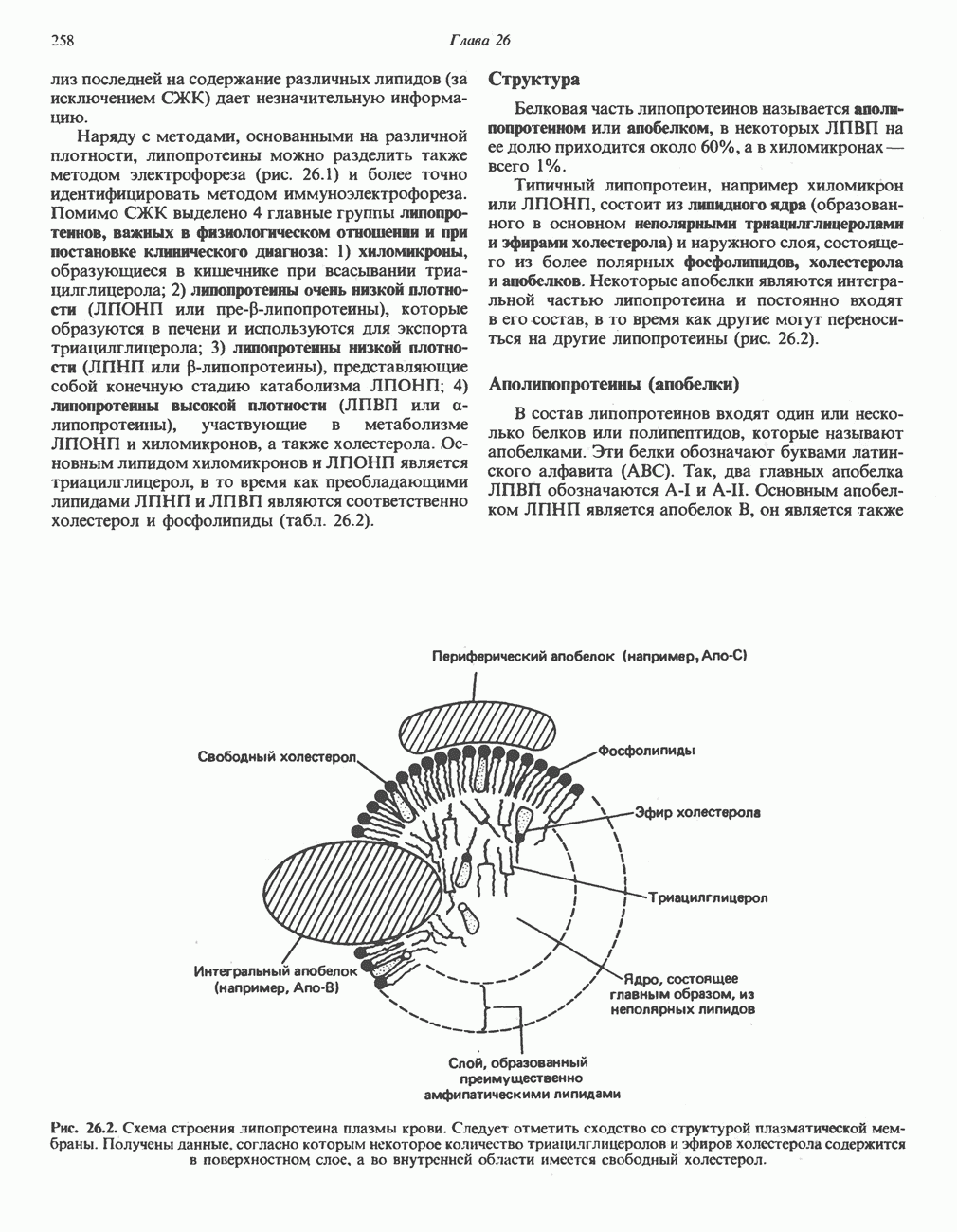
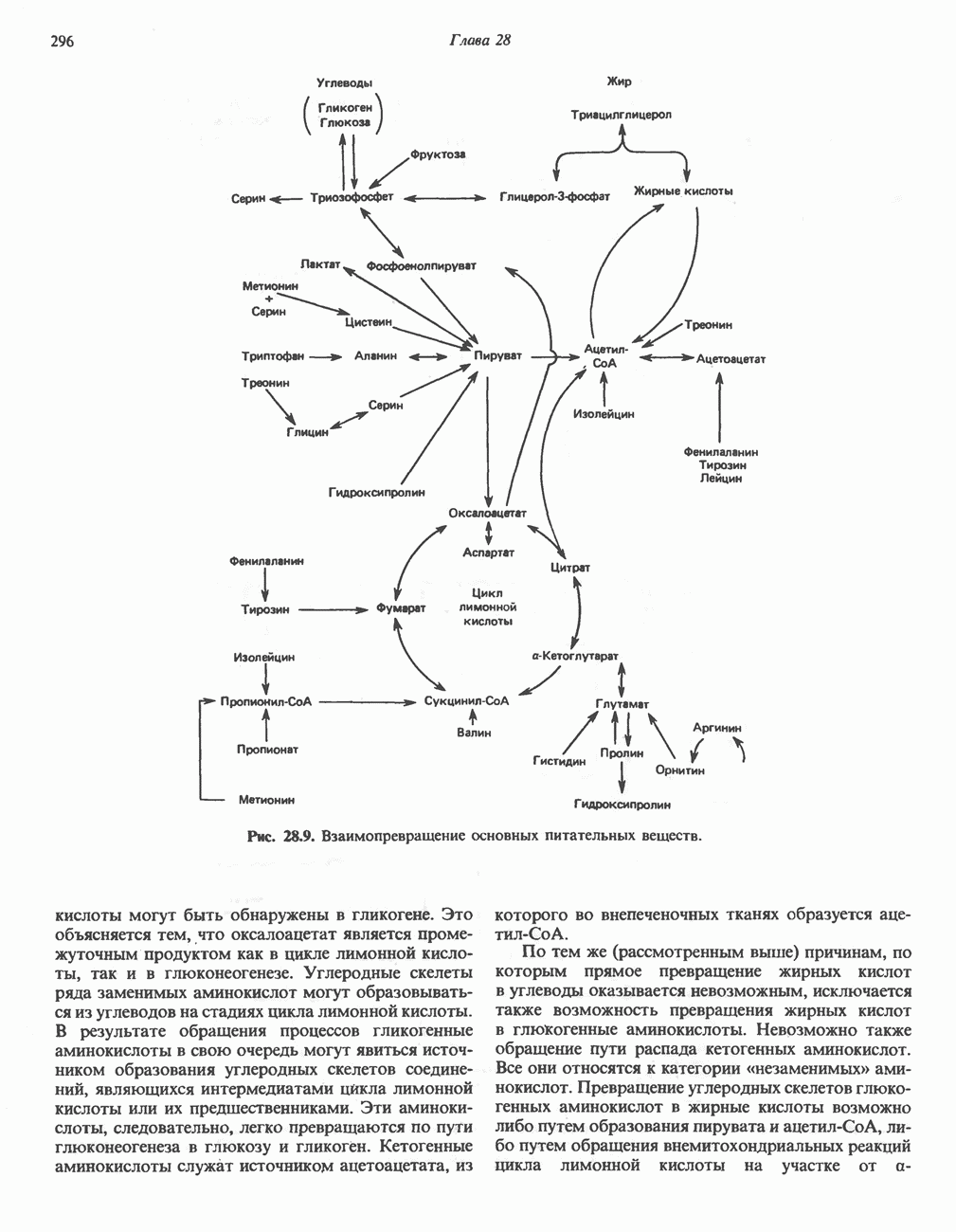
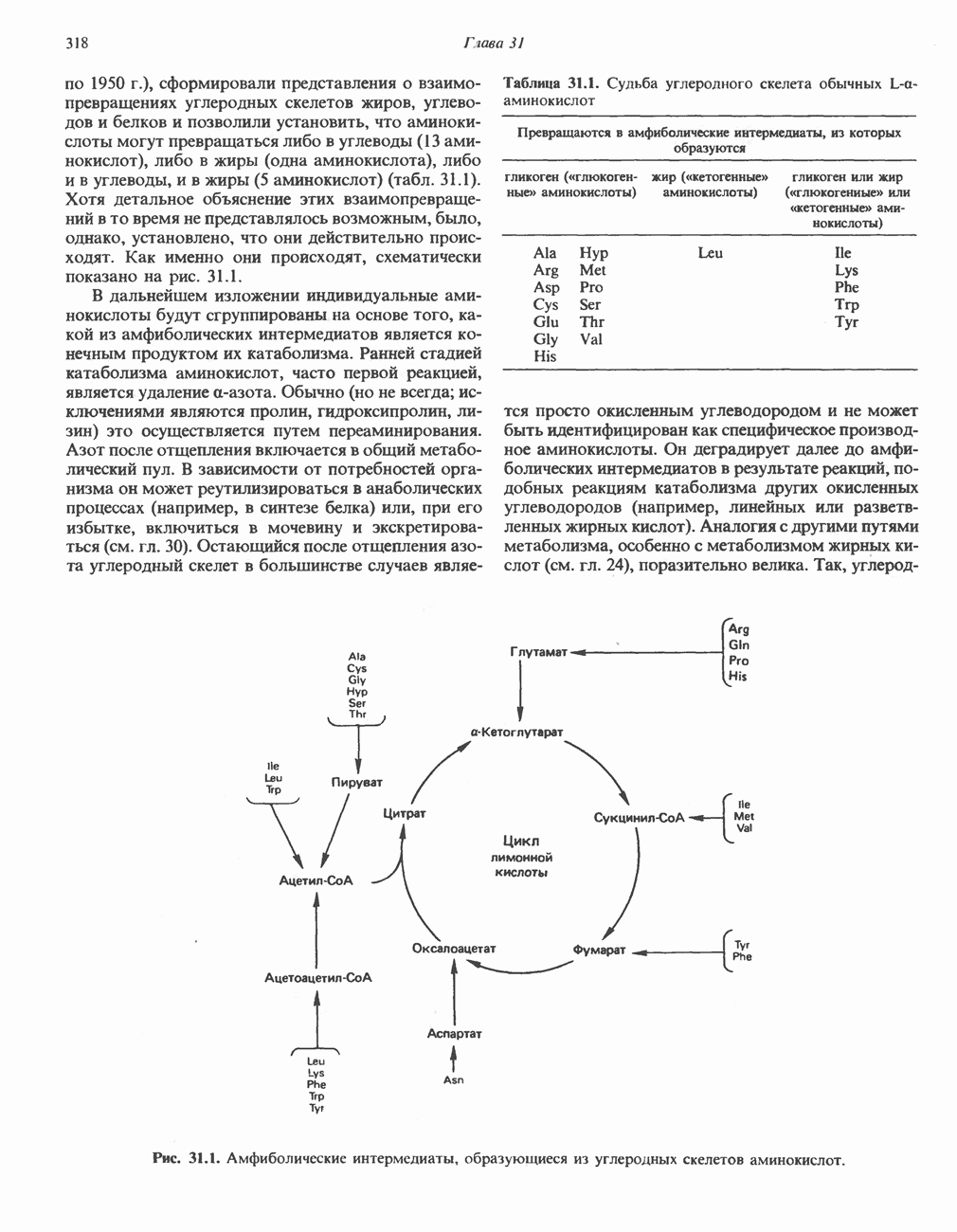
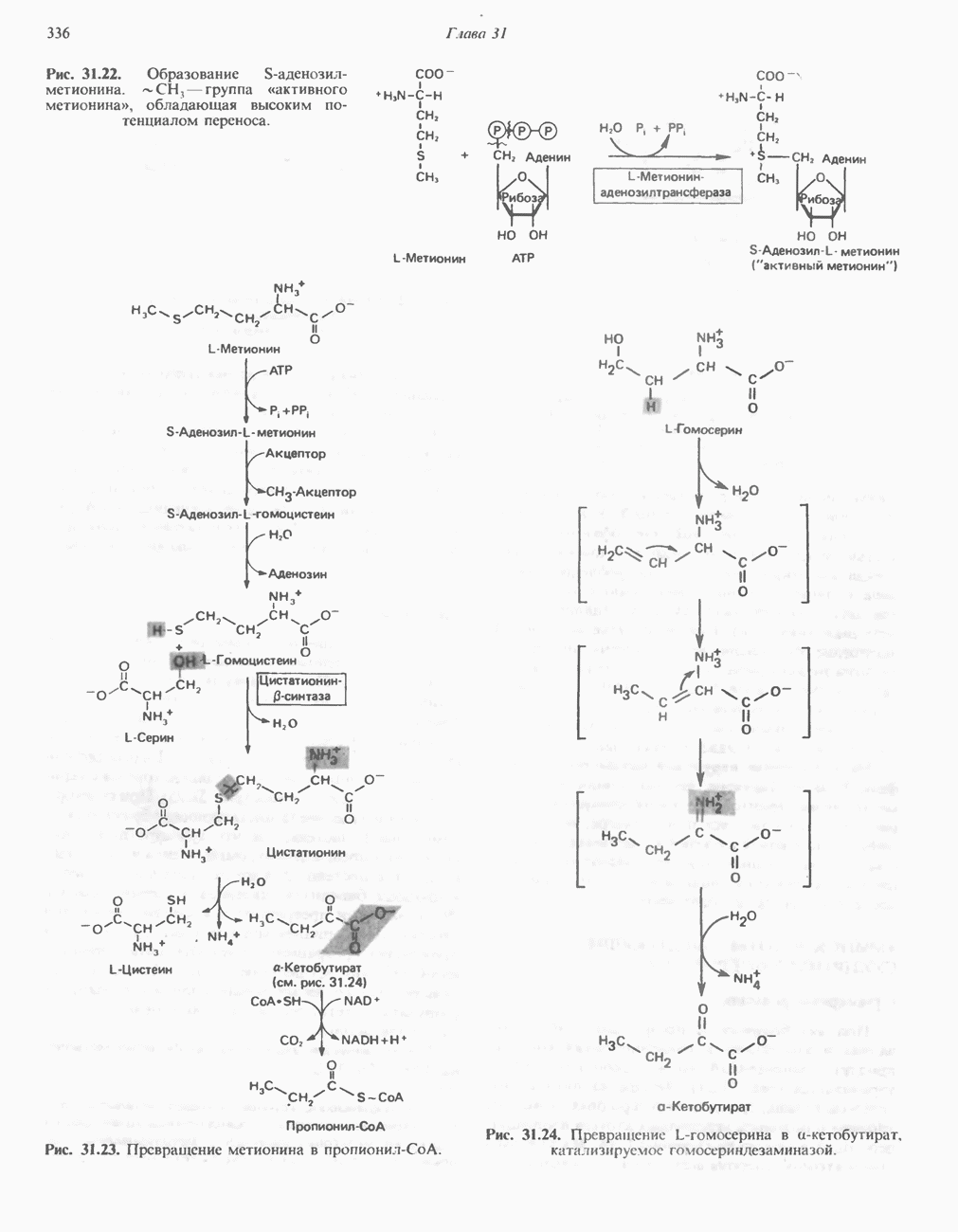
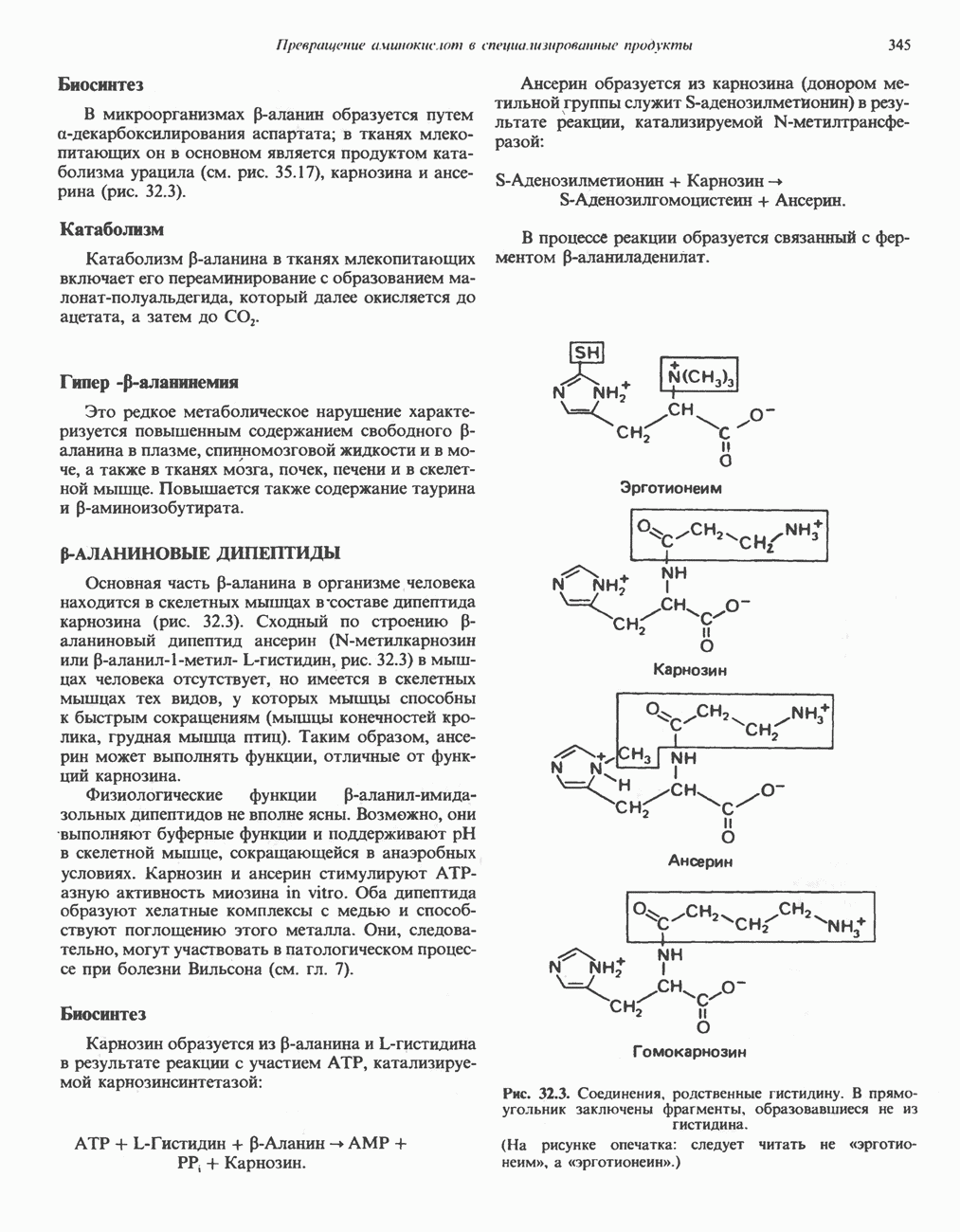
АМИНОКИСЛОТЫ, ОБРАЗУЮЩИЕ АЦЕТИЛ-КОФЕРМЕНТ АВсе аминокислоты, образующие пируват (аланин, цистеин, цистин, глицин, гидроксипролин, серин и треонин), могут превращаться в ацетил-СоА. Кроме того, 5 аминокислот образуют ацетил-СоА без промежуточного образования пирувата. К числу этих аминокислот относятся ароматические аминокислоты фенилаланин, тирозин и триптофан, основная аминокислота лизин и нейтральная аминокислота с разветвленной цепью лейцин. ТирозинА. Общая последовательность реакций. Пять последовательных ферментативных реакций превращают тирозин в фумарат и ацетоацетат (рис. 31.13): 1) реакция переаминирования с образованием п-гидроксифенилпирувата; 2) окисление с миграцией трехуглеродной боковой цепи и декарбоксилирова ние, приводящее к образованию гомогентизата; 3) окисление гомогентизата до малеилацетоацетата; 4) изомеризация малеилацетоацетата в фумарилацетоацетат и 5) гидролиз фумарилацетоацетата с образованием фумарата и ацетоацетата. Ацетоацетат может далее подвергнуться тиолитическому расщеплению на ацетат и ацетил-СоА. Несколько интермедиатов метаболизма тирозина
Рис. 31.12. Интермедиаты катаболизма а-гидроксипролина в тканях млекопитающих.
Рис. 31.13. Интермедиаты катаболизма тирозина. За исключением реакции, кат ализируемой р-кетотиолазой, все остальные реакции обсуждаются в тексте. Чтобы читателю было легче проследить за судьбой каждого атома углерода, они пронумерованы (см. также рис. было идентифицировано при изучении генетического заболевания человека алкаптонурии. Больные алкаптонурией экскретируют с мочой гомогентизат и немало полезной информации было получено при добавлении в пищу пациентов предполагаемых предшественников гомогентизата. Б. Переаминирование тирозина. Переаминирование тирозина с образованием « В. Окисление л-гидрокснфенилпирувата в гомогентизат. Хотя реакция (рис. 31.13) выглядит как гид-роксилирование Г. Превращение гомогентизата в фумарат и ацетоацетат. Окислительная реакция, приводящая к разрыву бензольного кольца гомогентизата и образованию малоилацетоацетата, катализируется гомогентизат-оксидазой—железосодержащим металлопротеином печени млекопитающих. Превращение малеилацетоацетата в фумарила-цетоацетат является цис-транс-изомеризацией относительно двойной связи и катализируется маленлацетоацетат- Нарушения катаболизма тирозина. Тяжелые нарушения катаболизма приводят к тирозинемии, тирозинурии и фенолацидурии. А. Тирозинемии типа I (тирозиноз). Тирозиноз характеризуется накоплением метаболитов, снижающих активность ряда ферментов и транспортных систем. Патофизиология этого нарушения является, следовательно, весьма сложной. Дефектными ферментами являются, вероятно, фумарилацетоацетат-гидролаза (рис. 31.13) и малеилацетоацетат-гидролаза. Известны острая и хроническая формы тирозино-за. Острая форма характерна для младенческого возраста, ее признаками являются понос, рвота, «капустный» запах, задержка в развитии. Если не проводится лечение, летальный исход наступает в возрасте 6—8 месяцев из-за недостаточности печени. Хроническая тирозинемия характеризуется сходными, но более умеренно выраженными симптомами; летальный исход наступает в возрасте примерно 10 лет. Содержание тирозина в плазме повышается до 6-12 мг/100 мл, повышено содержание и некоторых других аминокислот, особенно метионина. Лечение Б. Тирозинемия типа II (синдром Рихнера—Ханхарта). Предполагаемой причиной метаболических нарушений при тирозинемии типа II является недостаточность тирозин-трансаминазы печени (рис. 31.13). Клинические проявления включают повышение содержания тирозина в плазме (4-5 мг/100 мл), характерные поражения глаз и кожи и умеренную умственную отсталость. Отмечались также случаи членовредительства, нарушения тонкой координации движений. Тирозин является единственной аминокислотой, концентрация которой в моче повышена. Тем не менее клубочковая фильтрация и обратное всасывание тирозина остаются в пределах нормы. В число метаболитов, экскретируемых с мочой, входят и-гидроксифенилпируват, и-гидроксифениллактат, В. Тирозинемия новорожденных. Причиной болезни считается недостаточность « Г. Алкаптонурия. Это наследственное метаболическое (см. скан) Рис. 31.14. Альтернативные катаболиты тирозина, и-Гидроксифенилацетальдегид является интермедиатом стадии окисления тирамина в и-гидроксифенилацетат. нарушение описано в медицинской литературе еще в XVI в., оно было охарактеризовано в 1859 г. Болезнь представляет значительный исторический интерес, поскольку именно на основе ее изучения Гаррод выдвинул идею о наследственных метаболических нарушениях. Наиболее ярким клиническим проявлением этой болезни является потемнение мочи при условиях доступа воздуха. На более поздних стадиях заболевания наблюдается общая пигментация соединительной ткани (охроноз) и развивается артрит. Причиной болезни является недостаточность гомогеитизат-оксидазы (рис. 31.13). Субстрат фермента, гомогентизат, экскретнруется с мочой; при окислении на воздухе он образует темно-коричневый пигмент. Зарегистрировано свыше 600 случаев заболевания, частота заболевания оценивается в 2—5 случаев на миллион новорожденных. Алкаптонурия наследуется по аутосомно-рецессивному типу. В настоящее время не найдено диагностических методов выявления гетерозигот. Хотя механизм охроноза не установлен, полагают, что он обусловлен окислением гомогентизата полифенолоксидазой, приводящим к образованию бензохиноацетата, который далее полимеризуется и связывается с макромолекулами соединительной ткани.
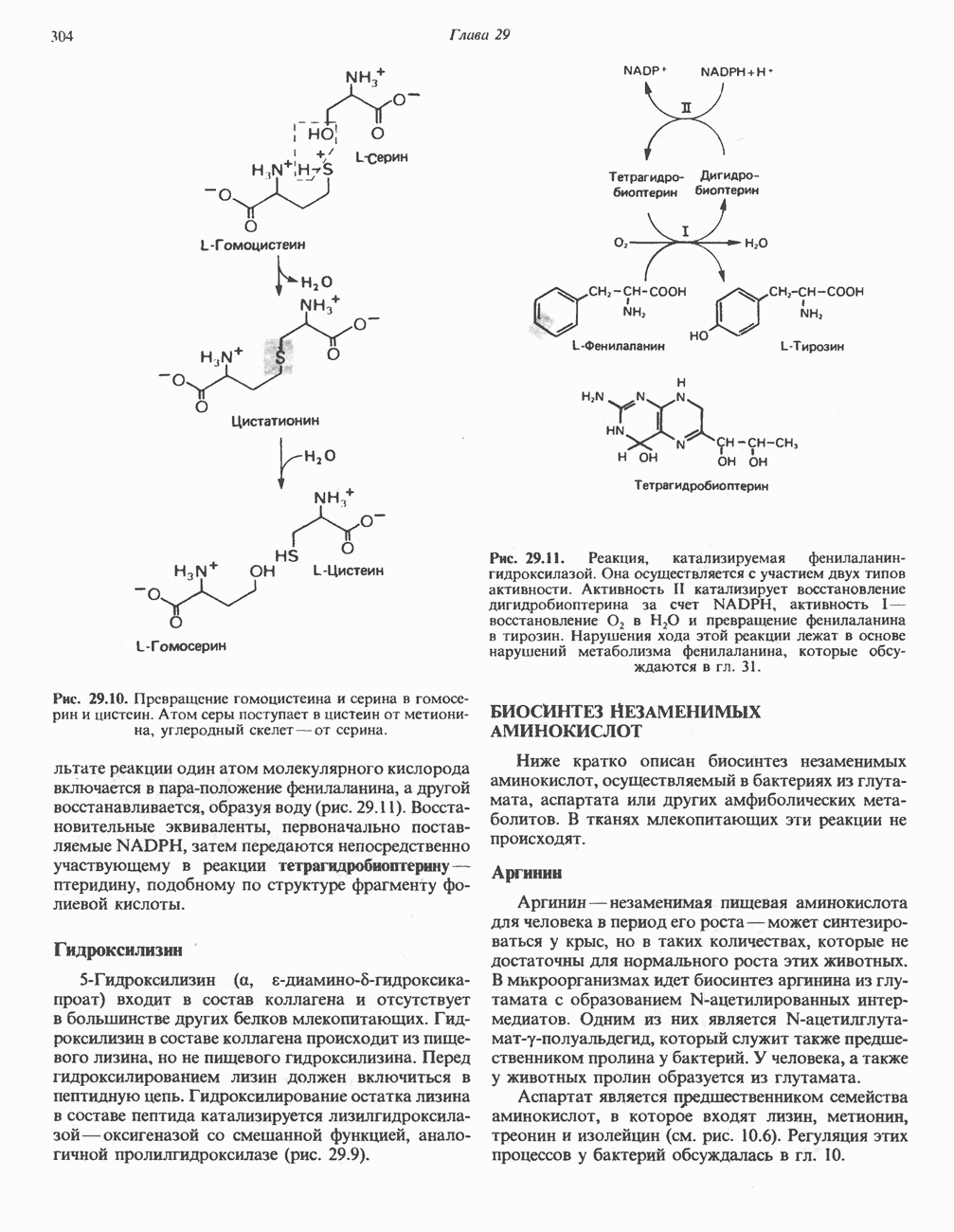
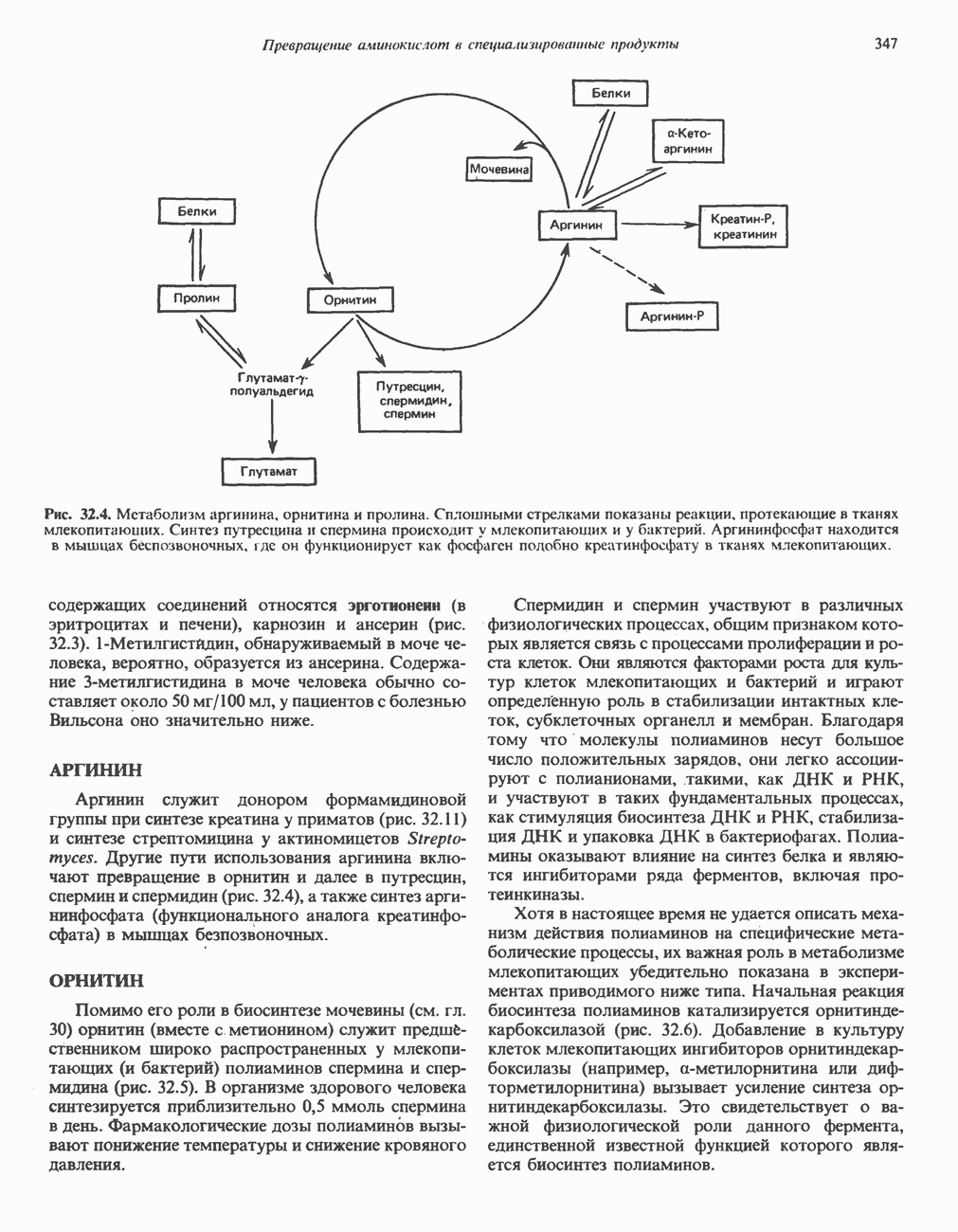
ФенилаланинФенилаланин сначала превращается в тирозин при участии фенилаланингидроксилазы (см. рис. 29.11). Распределение изотопной метки в амфиболических продуктах фумарате и ацетоацетате (рис. 31.15) такое же, как и в соответствующих продуктах катаболизма тирозина (рис. 31.13). Метаболические нарушения катаболизма фенилаланина. Главные Метаболические нарушения состоят в блокировании превращения фенилаланина в тирозин (см. рис. 29.12). Можно выделить три причины нарушений: недостаточность фенилаланингидроксилазы (гиперфенилаланинемия типа I, или классическая фенилкетонурия), недостаточность дигидробиоптеринредуктазы (гиперфенилаланинемия типов II и III) и нарушения биосинтеза дигидробиоптерина (гиперфенилаланинемия типов IV и V). Были также зарегистрированы нарушения других типов (табл. 31.3).
Рис. 31.15. Катаболическая судьба атомов углерода фенилаланина. Распределение изотопной метки в конечных катаболитах фенилаланина (и тирозина). Главным последствием гиперфенилаланинемии типа 1 (классическая фенилкетонурия, ФКУ) при отсутствии лечения является умственная отсталость (ниже 70 баллов по стандартному тесту для детей старшего возраста). Дополнительными клиническими симптомами являются припадки, психозы, экзема и «мышиный» запах. При раннем диагнозе и своевременном лечении эти симптомы могут не развиться. В связи с наличием экспериментальных моделей заболевания, а также благодаря возможности путем регулирования диеты предотвращать неизбежное в другом случае состояние, характеризующееся умственной отсталостью, ФКУ служит моделью для изучения этого состояния при других метаболических нарушениях. При классической ФКУ, которая является наследственным нарушением, встречающимся с частотой 1 на 10000 новорожденных, содержание в печени компонента 1 фенилаланингидроксилазы (см. рис. 29.12) составляет около 25% от нормы; при этом фермент оказывается нечувствительным к регуляторному действию фенилаланина. Организм пациента оказывается не способным превращать фенилаланин в тирозин, в результате образуются альтернативные катаболиты фенилаланина (рис. 31.16). В их число входят фенилпировиноградная кислота (продукт дезаминирования фенилаланина), фенилмолочная кислота (продукт восстановления фенилпировиноградной кислоты) и фенилуксусная кислота, образующаяся путем декарбоксилирования и окисления фенилпировиноградной кислоты. Большая часть фенилацетата в печени конъюгирует с глутамином и экскретнруется с мочой в виде конъюгата фенилацетилглутамина. В табл. 31.4 приведены концентрации метаболитов фенилаланина в крови и моче у пациентов с фенилкетонурией. Присутствие в моче кетокислоты фенилпирувата определило и само название болезни — фенилкетонурия. Таблица 31.3. Типы гиперфенилаланинемии (см. скан) При блокировании нормальных путей катаболизма фенилаланина на первый план выходят несколько катаболических реакций, которые протекают в нормальной печени, но обычно имеют второстепенное значение. У больных фенилкетонурией в крови и моче появляются фенилпируват, фениллактат, фенилацетат и фенилацетилглутамин (рис. 31.16). Хотя присутствие фенилпирувата в моче больных фенилкетонурией может быть установлено путем простого биохимического анализа, для убедительного диагноза необходимо зарегистрировать повышение содержания фенилаланина в плазме крови. Прогрессирующее развитие нарушений умственного развития у детей, больных фенилкетонурией, можно предотвратить путем назначения диеты с очень низким содержанием фенилаланина. По достижении шестилетнего возраста эту диету можно отменить, поскольку к этому времени повышенные концентрации фенилаланина и его производных перестают оказывать неблагоприятное действие на мозг. Содержание фенилаланина в плазме можно измерить с помощью автоматизированного микрометода, для которого требуются пробы крови не более 20 мкл. Следует отметить, что ненормально высокое содержание фенилаланина у детей, больных фенилкетонурией, обнаруживается лишь на третий или четвертый день жизни, поскольку они потребляют незначительное количество пищевого белка. Далее, у недоношенных детей можно получить ложную положительную реакцию в связи с замедленным образованием ряда ферментов метаболизма фенилаланина. Полезным, но не вполне надежным массовым тестом может служить обнаружение повышенного Таблица 31.4. Метаболиты фенилаланина, накапливающиеся в плазме и моче больных фенилкетонурией
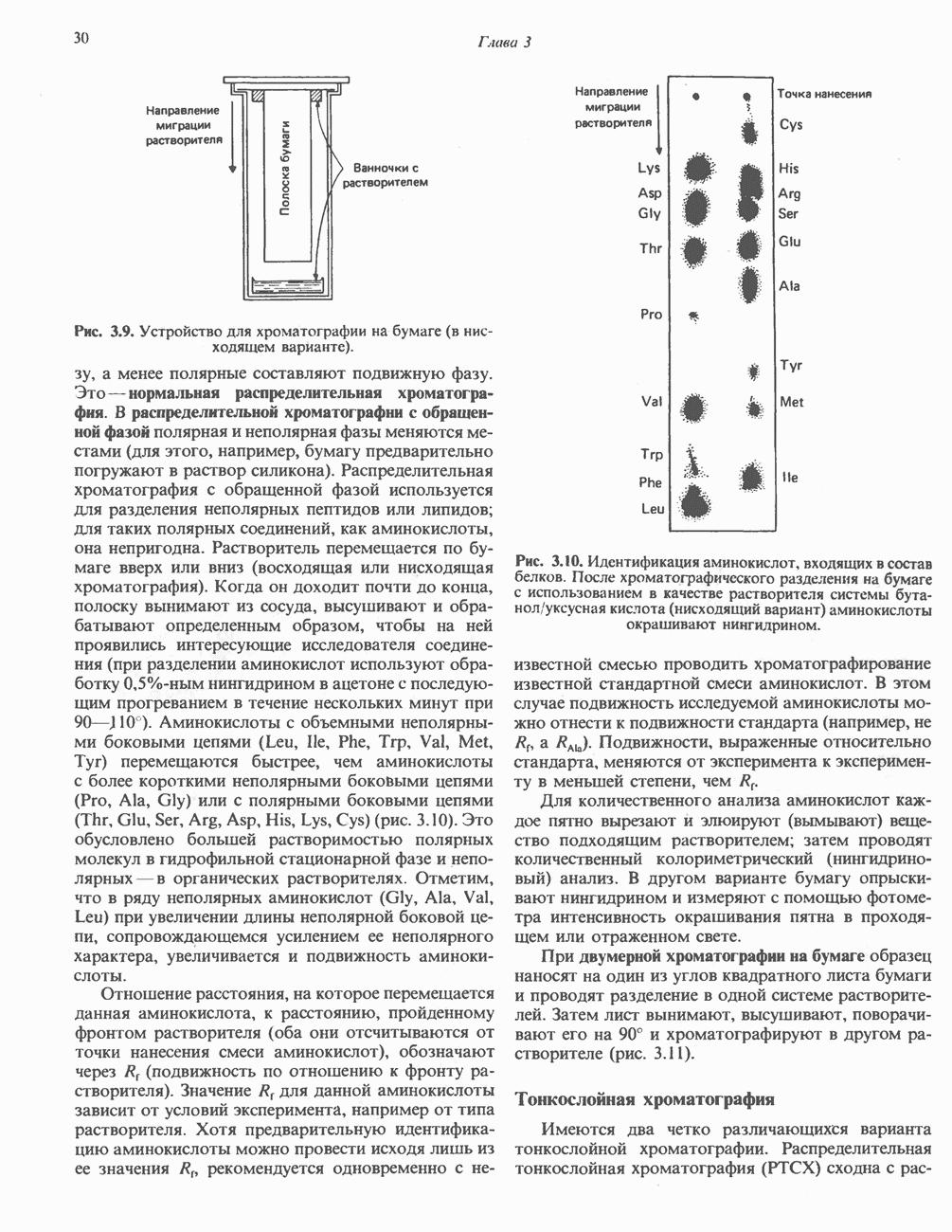
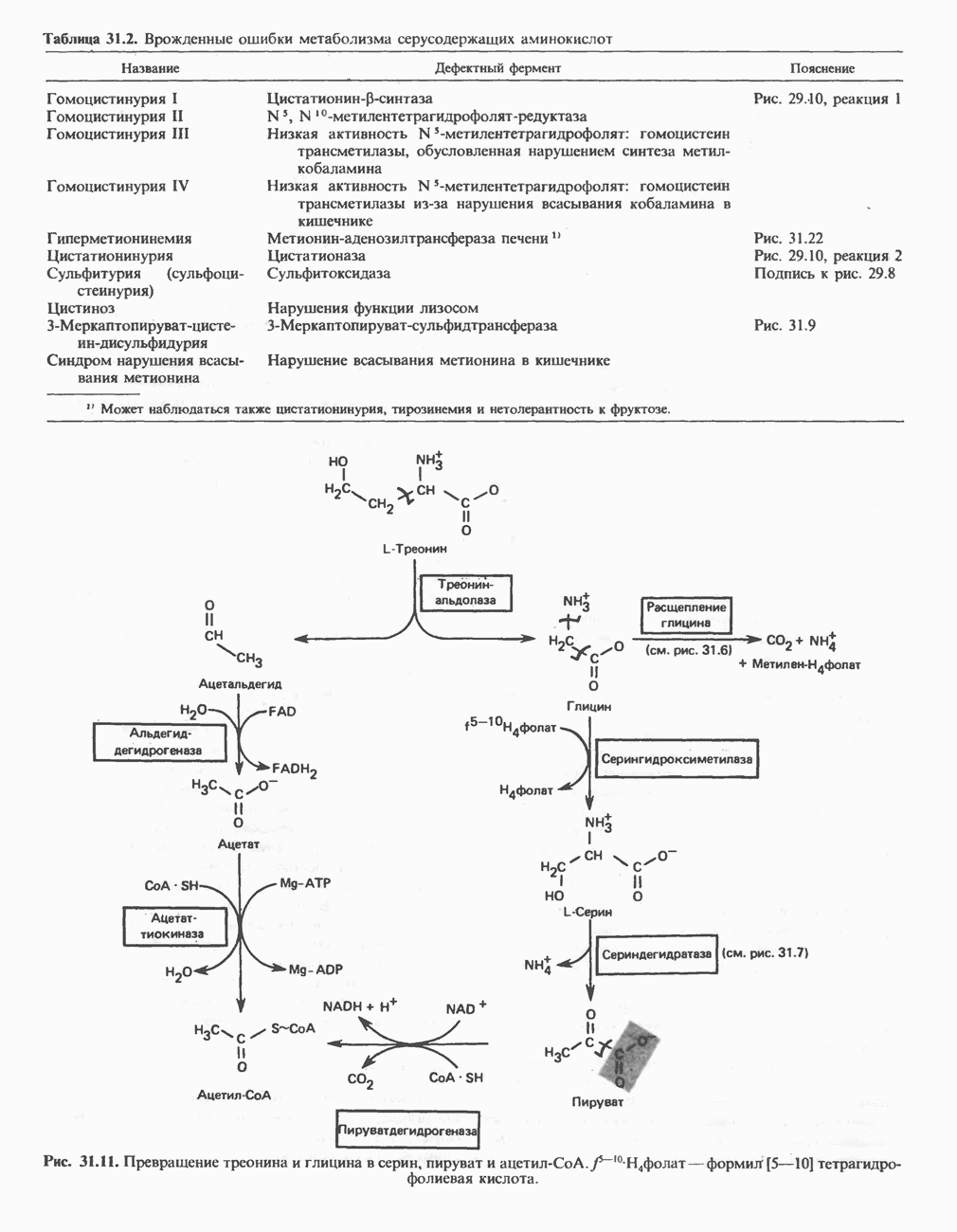
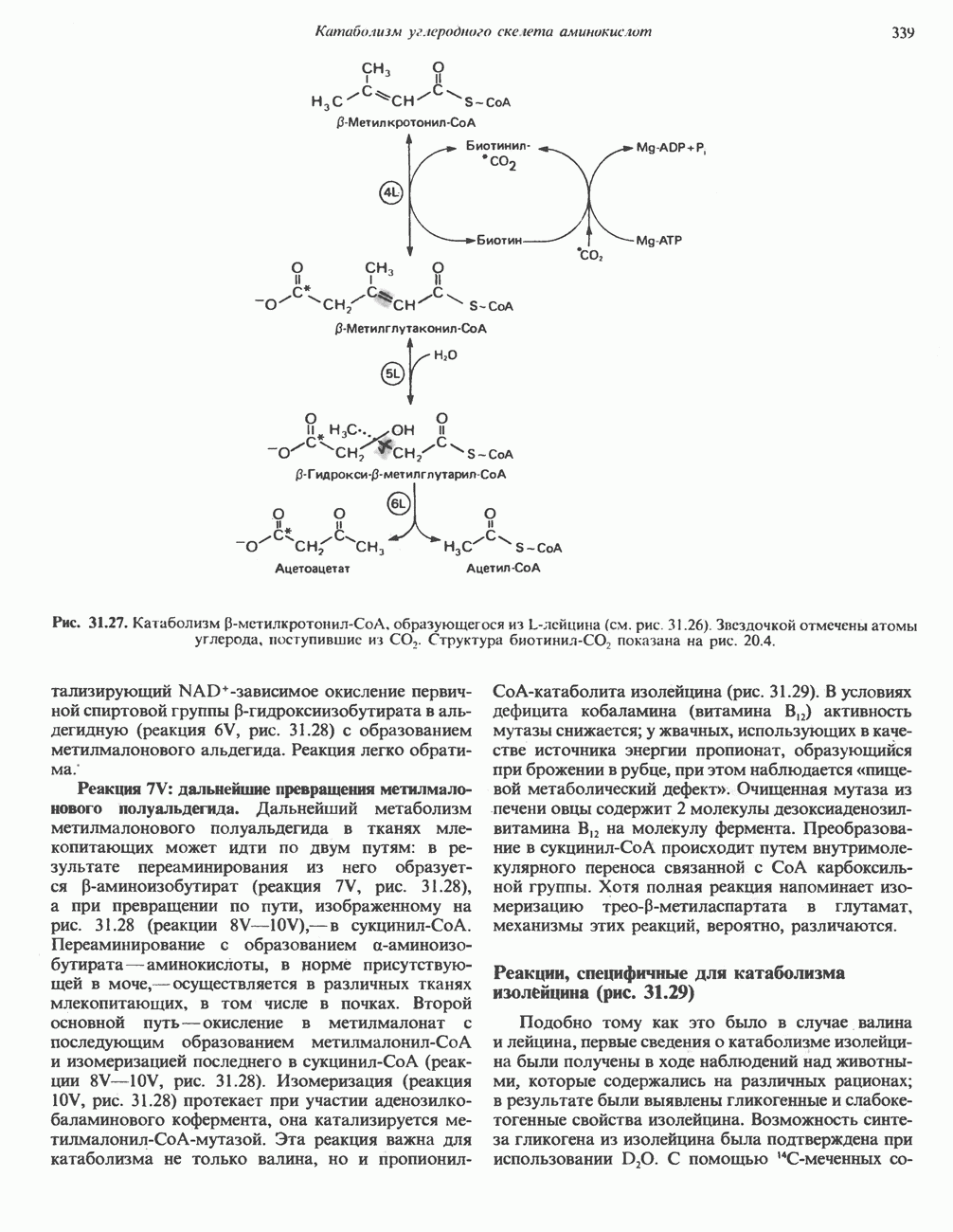
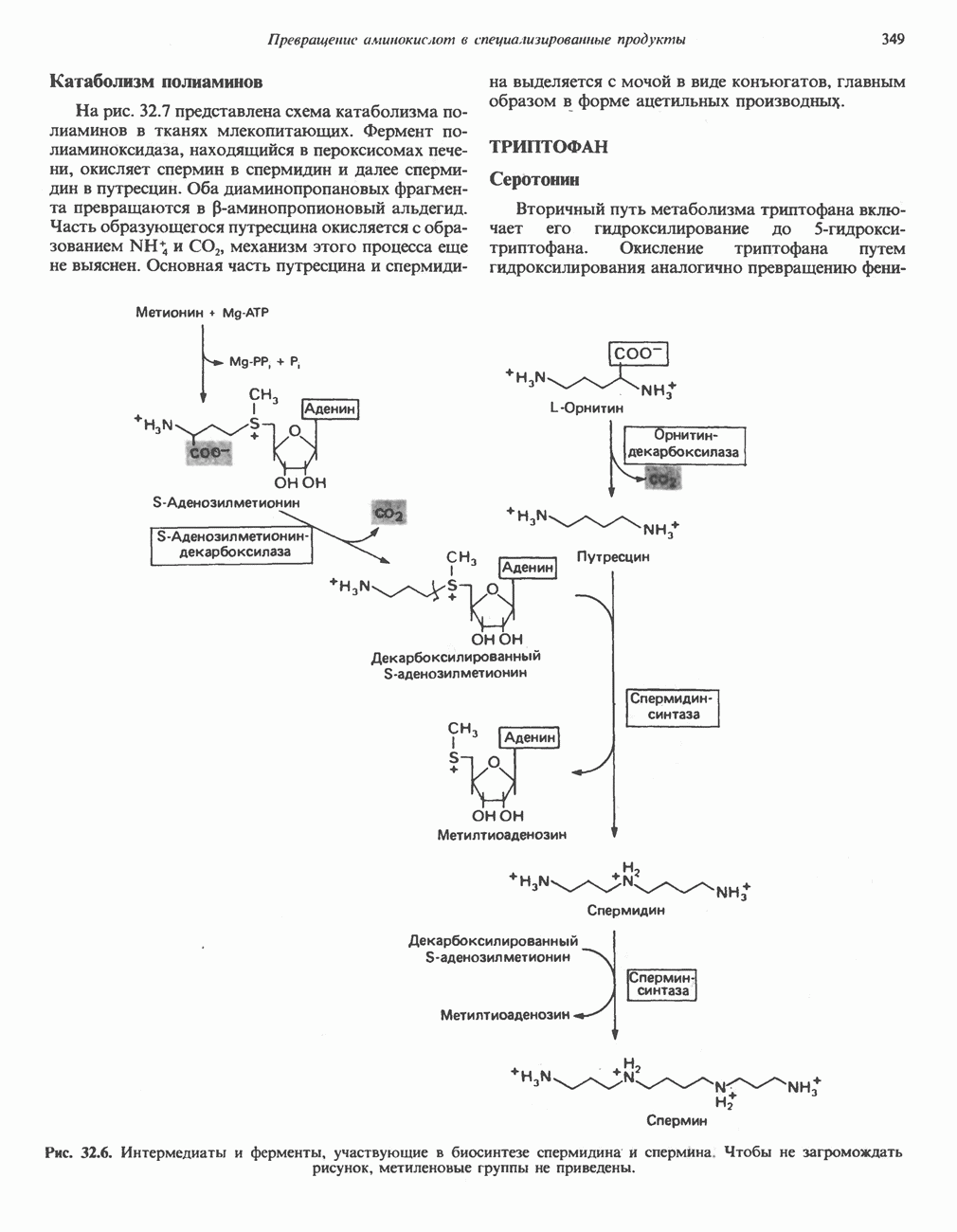
Рис. 31.16. Альтернативные пути катаболизма фенилаланина, имеющие особое значение при фенилкетонурии. Указанные реакции протекают также и в печени здоровых людей, но при нормальном функционировании фенилаланин-гидроксилазы они не имеют существенного значения. Glu — глутамат. Gin - глутамин. содержания фенилпирувата с моче с использованием в качестве реагента хлорида железа. После введения фенилаланина пациентам с фенилкетонурией содержание этой аминокислоты в крови долгое время остается высоким, что свидетельствует о снижении толерантности к фенилаланину. Вместе с тем ненормально низкая толерантность к введенному фенилаланину и повышенное содержание последнего в период между приемами пищи характерно также и для родителей, больных фенилкетонурией. Таким образом, дефектный ген, ответственный за фенилкетонурию, может быть выявлен и у фенотипически нормальных гетерозиготных родителей. ЛизинЛизин составляет исключение из общего правила, согласно которому первой стадией катаболизма аминокислоты является удаление а-аминогруппы путем переаминировання. В тканях млекопитающих ни а-, ни е-аминогруппы L-лизина не участвуют в переаминировании. В организме млекопитающих углеродный скелет L-лизина переходит в состав а-аминоадипата и а-кетоадипата (рис. 31.17). Первоначально полагали, что деградация L-лизина идет через стадию образования пипеколиновой кислоты — циклической иминокислоты. В печени D-лизин действительно превращается в пипеколат, однако распад L-лизина происходит через стадию образования сахаропина (рис. 31.18) - интермедиата биосинтеза лизина у грибов. L-лизин сначала конденсируется с а-кетоглутаратом, при этом отщепляется молекула воды и образуется шиффово основание. Далее происходит восстановление этого соединения в сахаропин при участии соответствующей дегидрогеназы (рис. 31.18), а затем окисление сахаропина другой дегидрогеназой. Расщепление продукта водой приводит к образованию В ходе дальнейшего катаболизма а-аминоадипат путем переаминировання превращается в а-кетоадипат, после чего, вероятно, происходит окислительное декарбоксилирование а-кетоадипата с образованием Метаболические нарушения катаболизма лизина. Описаны два редких метаболических нарушения катаболизма лизина. Оба они являются следствием дефектности ферментов, осуществляющих катаболизм
Рис. 31.17. Превращение
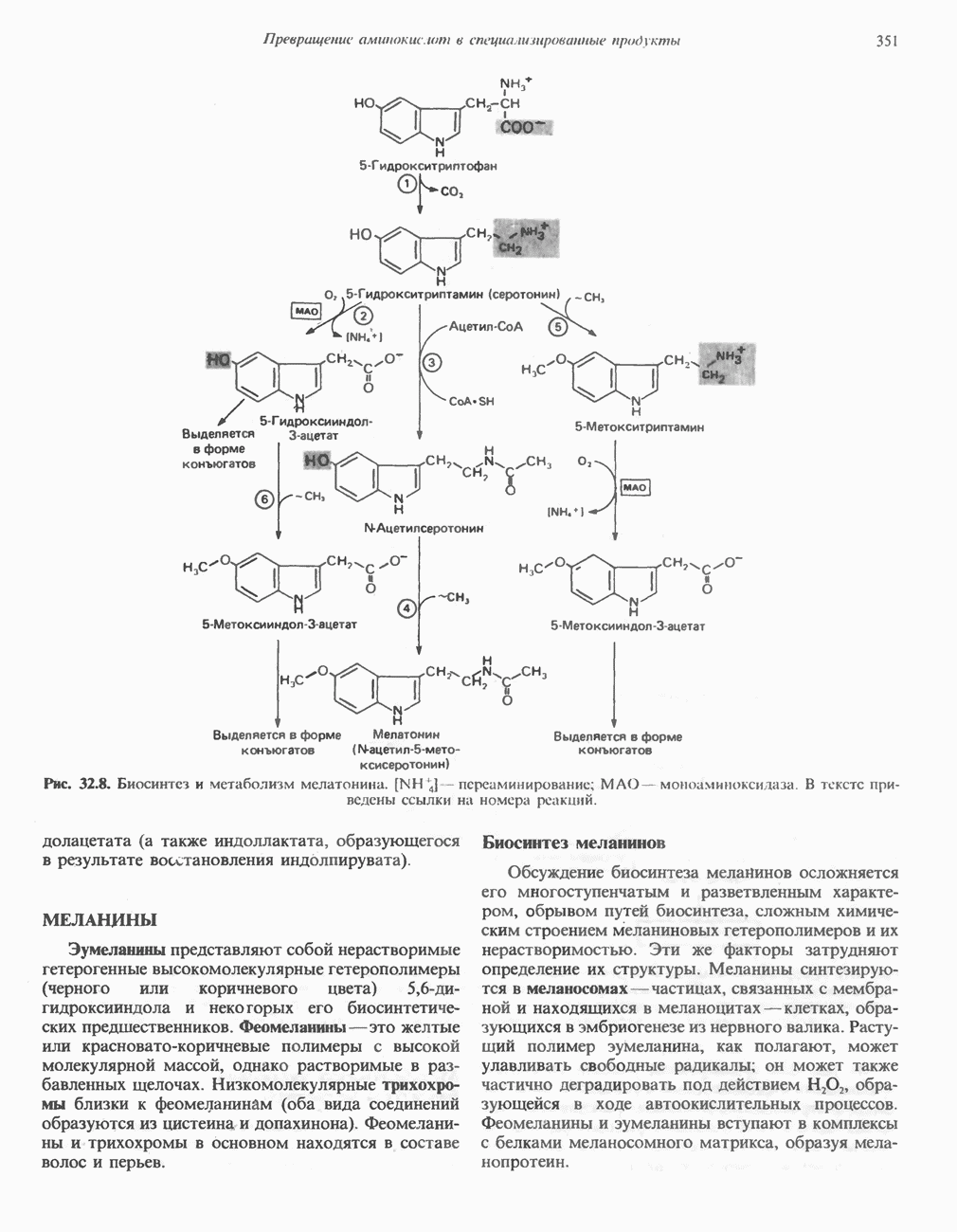
Рис. 31.18. Катаболизм лизина до А. Периодическая гиперлизинемия и сопутствующая гипераммониемия. При периодической гиперлизинемии даже потребление белка в нормальных количествах вызывает повышение концентрации лизина в тканях белка. Вторичной по отношению к гиперлизинемии является гипераммониемия, развивающаяся вследствие конкурентного ингибирования лизином активности печеночной аргиназы (см. рис. 30.13). Увеличение потребления жидкости и снижение содержания лизина в диете понижают уровень гипераммониемии и ослабляют ее клинические проявления. И наоборот, нагрузка лизином вызывает тяжелый криз и коматозное состояние. Пока не имеется данных о генетической природе болезни. Б. Стойкая гиперлизинемия без сопутствующей гипераммониемии. Клинические и биохимические данные оказались весьма варьирующими у 12 пациентов со стойкой гиперлизинемией. У некоторых больных, но не у всех, наблюдалась умственная отсталость. Сопутствующей гипераммониемии не отмечалось даже после лизиновой нагрузки. В ряде случаев наблюдалось накопление катаболитов лизина в биологических жидкостях. Устойчивую гипер-лизинемию рассматривают как аутосомно-рецессивный признак. Наряду с блокированием процесса превращения лизина и а-кетоглутарата в сахаропин у некоторых пациентов, по-видимому, имелось нарушение превращения сахаропина в ТриптофанТриптофан вследствие многообразия связанных с ним метаболических реакции и продуктов был одной из первых аминокислот, которые были отнесены к незаменимым. Исследования, проведенные на мутантах Neurospora и на бактериях Pseudomonas, а также выделение метаболитов триптофана из мочи, оказали неоценимую помощь в выяснении деталей метаболизма триптофана. При введении с пищей [ Триптофаноксигеназа (триптофанпирролаза) катализирует раскрытие индольного кольца с включением двух атомов молекулярного кислорода в образующийся (см. скан) Рис. 31.19. Катаболизм L-триптофана PLP - пиридоксальфосфат. является металлопротеином, содержащим железо-порфирин; синтез его в печени индуцируется адрено-кортикостероидами и триптофаном. Значительная часть синтезированного фермента находится в латентной форме и требует активации. Триптофан стабилизирует оксигеназу по отношению к протеолити ческим ферментам. Она ингибируется по принципу обратной связи производными никотиновой кислоты, в том числе NADPH. Гидролитическое удаление формильной группы N-формилкинуренина катализируется в печени млекопитающих кинуренин-формилазой. Гидролиз в Продуктом реакции, катализируемой кинуренин-формилазой, является кинуренин (рис. 31.19). Он может быть дезаминирован в результате реакции переаминирования с переносом аминогруппы боковой цепи на а-кетоглутарат. Образующееся при этом кетопроизводное претерпевает спонтанную циклизацию превращаясь в кинуреновую кислоту. Это соединение является побочным продуктом катаболизма кинуренин и не относится к катаболитам, образующимся на главном пути (рис. 31.19). Дальнейший ход метаболизма кинуренина вклю чает его превращение в З-гидроксикинуренин и далее в 3-гидроксиантранилат. Гидроксилированйе происходит при участии молекулярного кислорода и осуществляется в Кинуренин и гидрок икинуренин превращаются в гидроксиантранилат при участии пиридоксальфосфат-содержащего фермента кинурениназы. Недостаток витамина В6 приводит к частичной утрате способности к катаболизму этих кинурениновых производных; во внепеченочных тканях они превращаются в ксантуренат (рис. 31.20). Этот в норме отсутствующий катаболит появляется в моче человека обезьян и крыс при недостаточном содержании в пище витамина В6. В этих условиях введение избыточных количеств триптофана приводит к экскреции ксантурената с мочой. У многих животных превращение триптофана
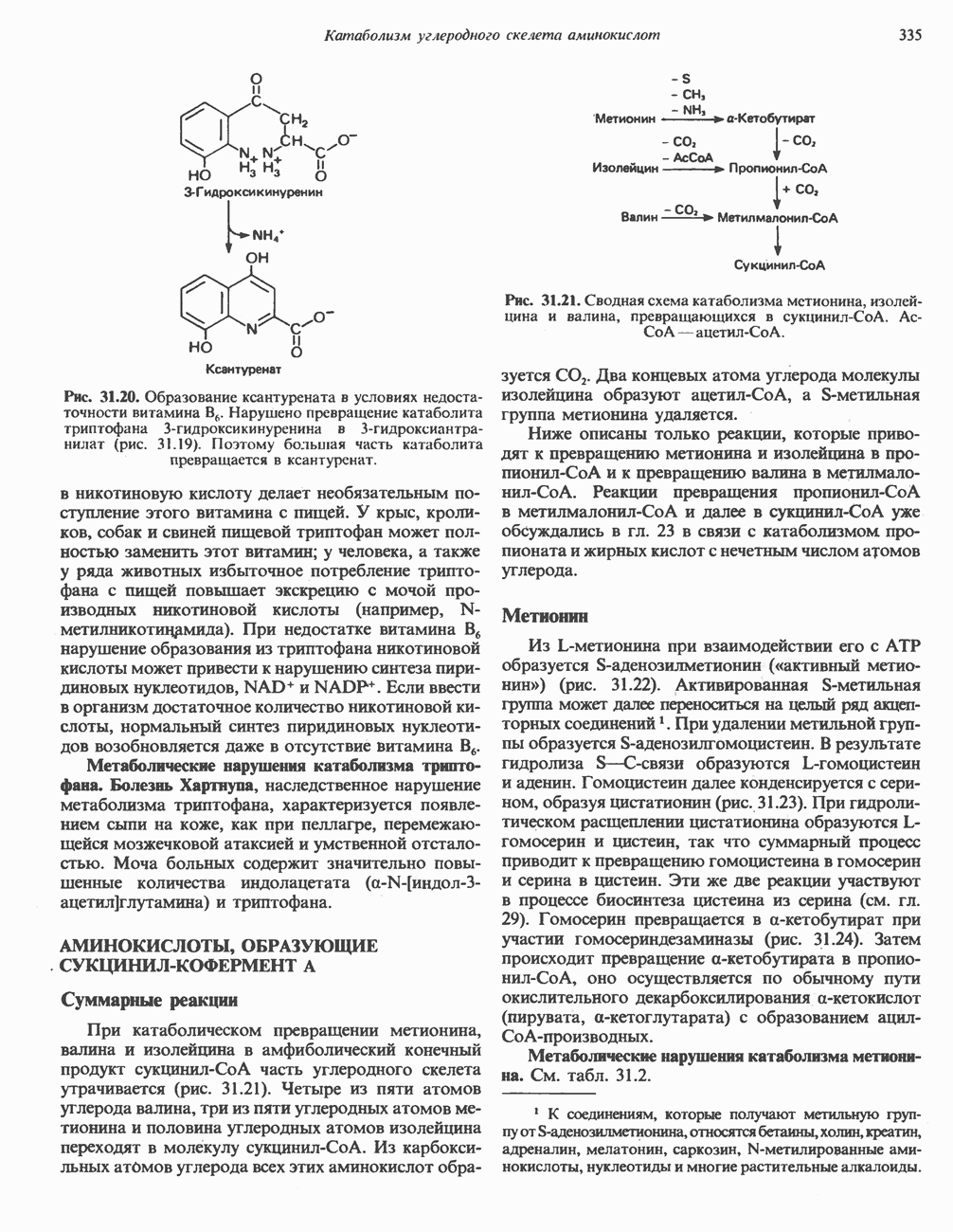
Рис. 31.20. Образование ксантурената в условиях недостаточности витамина В6. Нарушено превращение катаболита триптофана 3-гидроксикинуренина в 3-гидроксиантра-нилат (рис. 31.19). Поэтому большая часть катаболита превращается в ксантуренат. в никотиновую кислоту делает необязательным поступление этого витамина с пищей. У крыс, кроликов, собак и свиней пищевой триптофан может полностью заменить этот витамин; у человека, а также у ряда животных избыточное потребление триптофана с пищей повышает экскрецию с мочой производных никотиновой кислоты (например, N-метилникотивдмида). При недостатке витамина В6 нарушение образования из триптофана никотиновой кислоты может привести к нарушению синтеза пиридиновых нуклеотидов, NAD+ и NADP+. Если ввести в организм достаточное количество никотиновой кислоты, нормальный синтез пиридиновых нуклеотидов возобновляется даже в отсутствие витамина В6. Метаболические нарушения катаболизма триптофана. Болезнь Хартнупа, наследственное нарушение метаболизма триптофана, характеризуется появлением сыпи на коже, как при пеллагре, перемежающейся мозжечковой атаксией и умственной отсталостью. Моча больных содержит значительно повышенные количества индолацетата
|
 |
Оглавление
|







































































































































































































































 а-кетокислота,
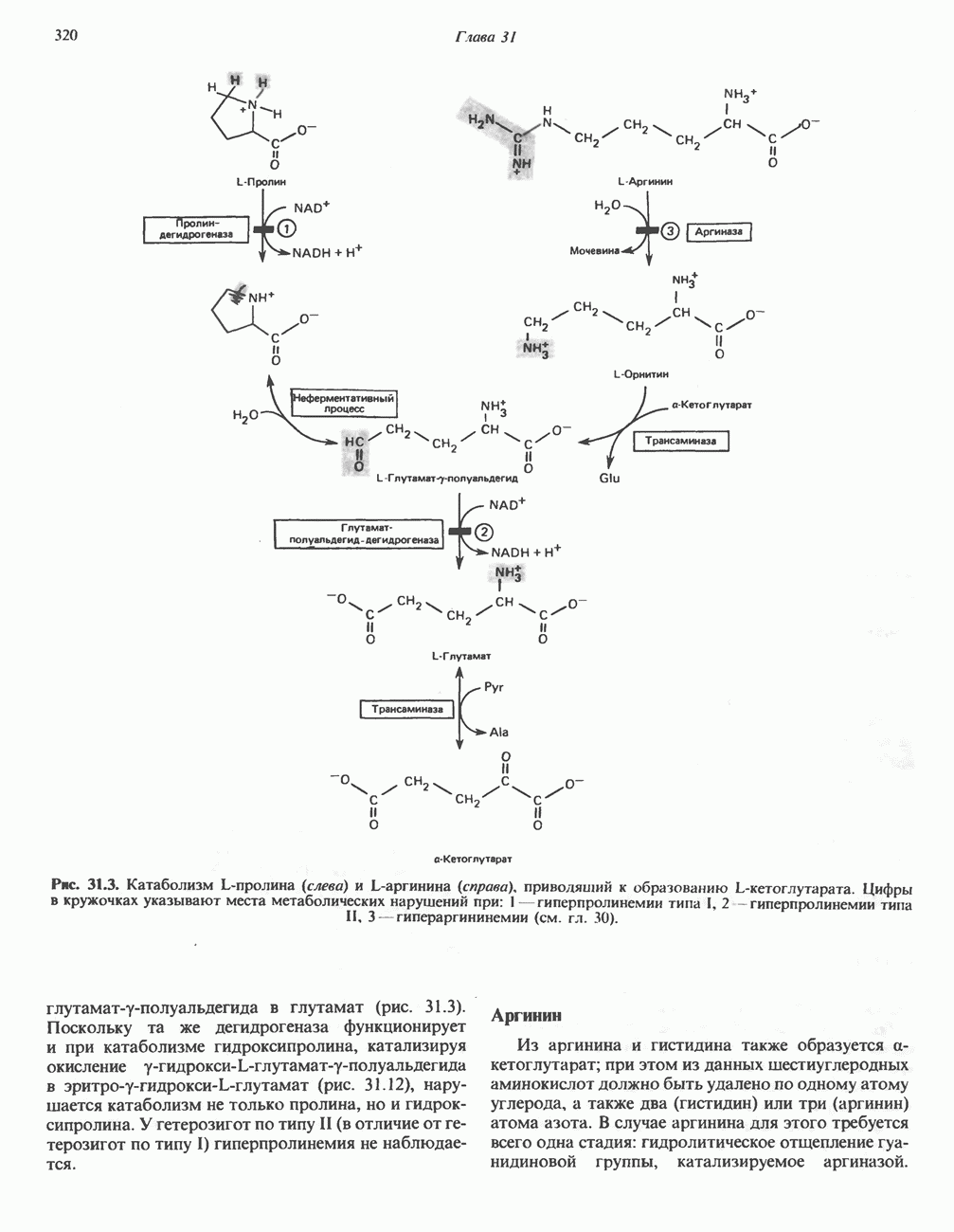
а-кетокислота,  а-аминокислота. Цифры в кружочках указывают вероятные места метаболических нарушений при:
а-аминокислота. Цифры в кружочках указывают вероятные места метаболических нарушений при:  — гиперпролинемии типа II.
— гиперпролинемии типа II.

 а-кетоглутарат, Glu - глутамат,
а-кетоглутарат, Glu - глутамат,  -пиридоксальфосфат. Цифры в кружочках указывают вероятные места метаболических нарушений при 1 — тирозинемии типа II, 2-тирозинемии новорожденных, 3-алкаптонурии и 4-тирозинемии типа I, или тирозинозе.
-пиридоксальфосфат. Цифры в кружочках указывают вероятные места метаболических нарушений при 1 — тирозинемии типа II, 2-тирозинемии новорожденных, 3-алкаптонурии и 4-тирозинемии типа I, или тирозинозе.
 -гидроксифенил-пирувата катализируется тирозин-а-кетоглутарат-трансаминазой—индуцируемым ферментом печени млекопитающих.
-гидроксифенил-пирувата катализируется тирозин-а-кетоглутарат-трансаминазой—индуцируемым ферментом печени млекопитающих.
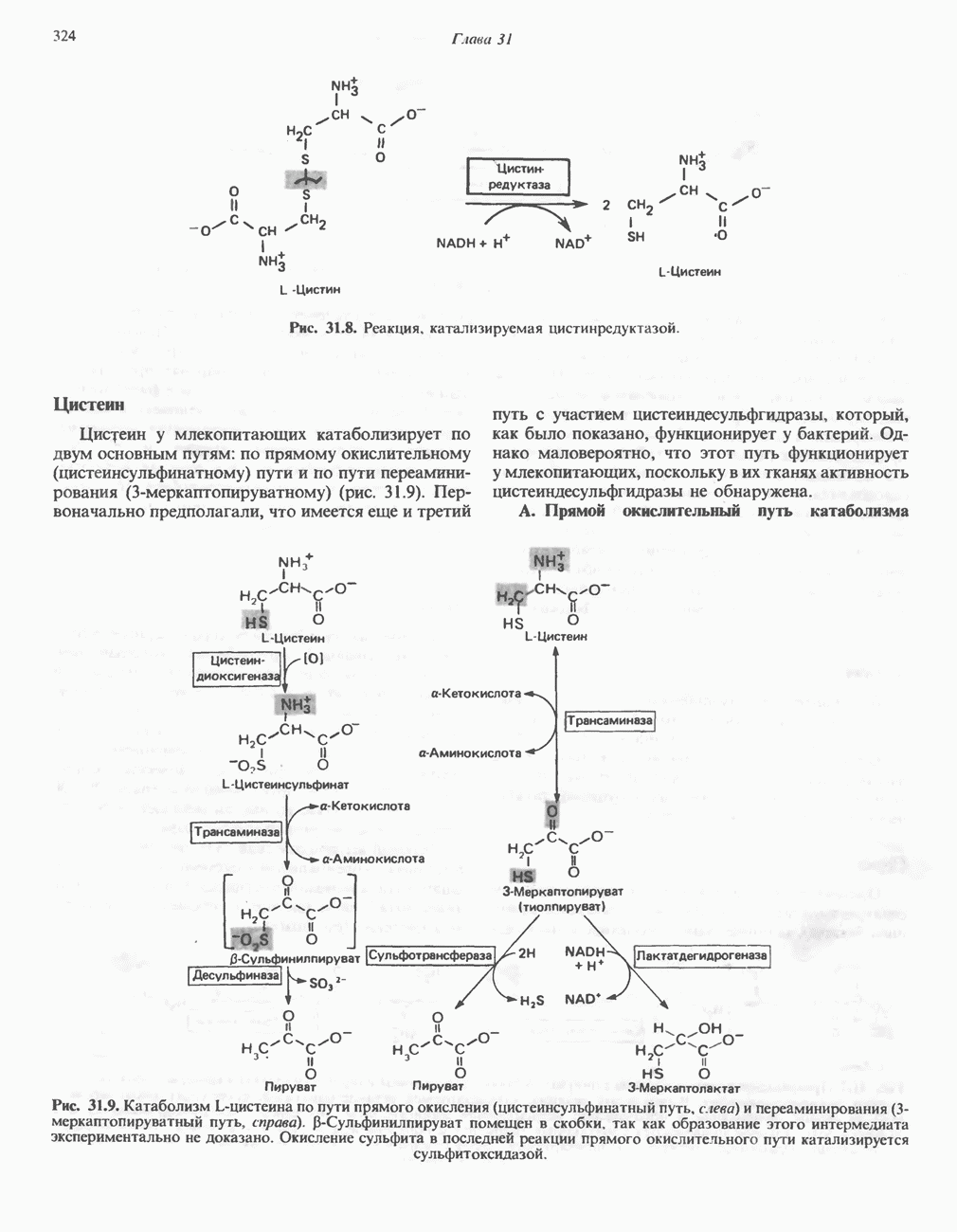
 -гидроксифенилпирувата, сопровождающееся окислительным удалением карбоксильного углерода, в действительности сочетанно происходит миграция боковой цепи и гидроксилирование кольца. л-Гидроксифенилпируват-гидролаза является медьсодержащим металлопротеином, сходным с тирозиназой. Хотя in vitro кофактором реакции могут служить не только аскорбат, но и другие восстанавливающие агенты, больные цингой экскретируют с мочой неполностью окисленные продукты метаболизма тирозина.
-гидроксифенилпирувата, сопровождающееся окислительным удалением карбоксильного углерода, в действительности сочетанно происходит миграция боковой цепи и гидроксилирование кольца. л-Гидроксифенилпируват-гидролаза является медьсодержащим металлопротеином, сходным с тирозиназой. Хотя in vitro кофактором реакции могут служить не только аскорбат, но и другие восстанавливающие агенты, больные цингой экскретируют с мочой неполностью окисленные продукты метаболизма тирозина.
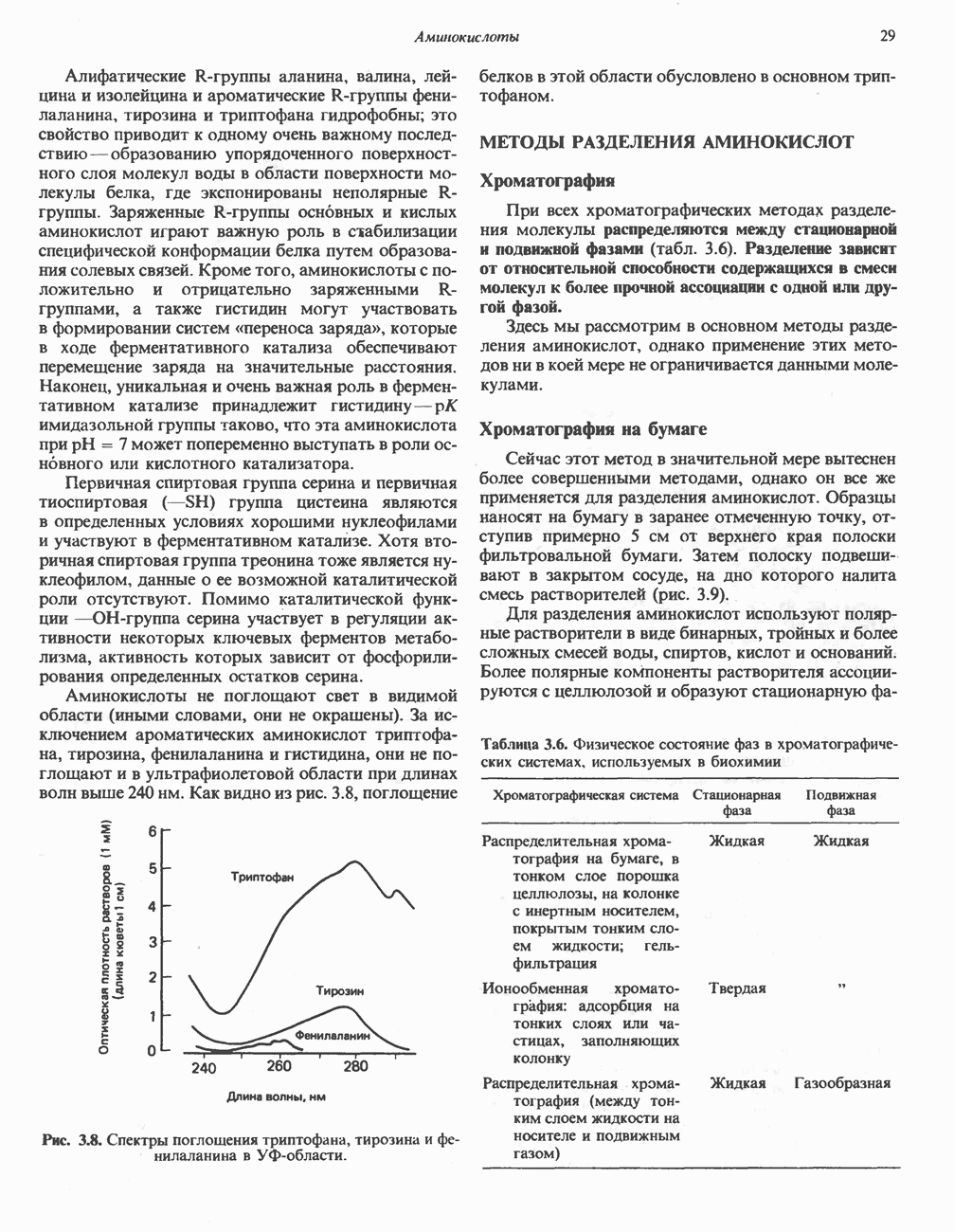
 -изомеразой -
-изомеразой -  зависимым ферментом печени млекопитающих. Гидролиз фумарилацетоацетата, катализируемый фумарилацетоацетат-гидролазой, приводит к образованию фумарата и ацетоацетата. Последний затем может быть превращен в ацетил-СоА и ацетат в реакции, катализируемой р-кетотиолазой (см. гл. 23).
зависимым ферментом печени млекопитающих. Гидролиз фумарилацетоацетата, катализируемый фумарилацетоацетат-гидролазой, приводит к образованию фумарата и ацетоацетата. Последний затем может быть превращен в ацетил-СоА и ацетат в реакции, катализируемой р-кетотиолазой (см. гл. 23).
 включает диету с пониженным содержанием тирозина и фенилаланина, а в ряде случаев также и метионина.
включает диету с пониженным содержанием тирозина и фенилаланина, а в ряде случаев также и метионина.
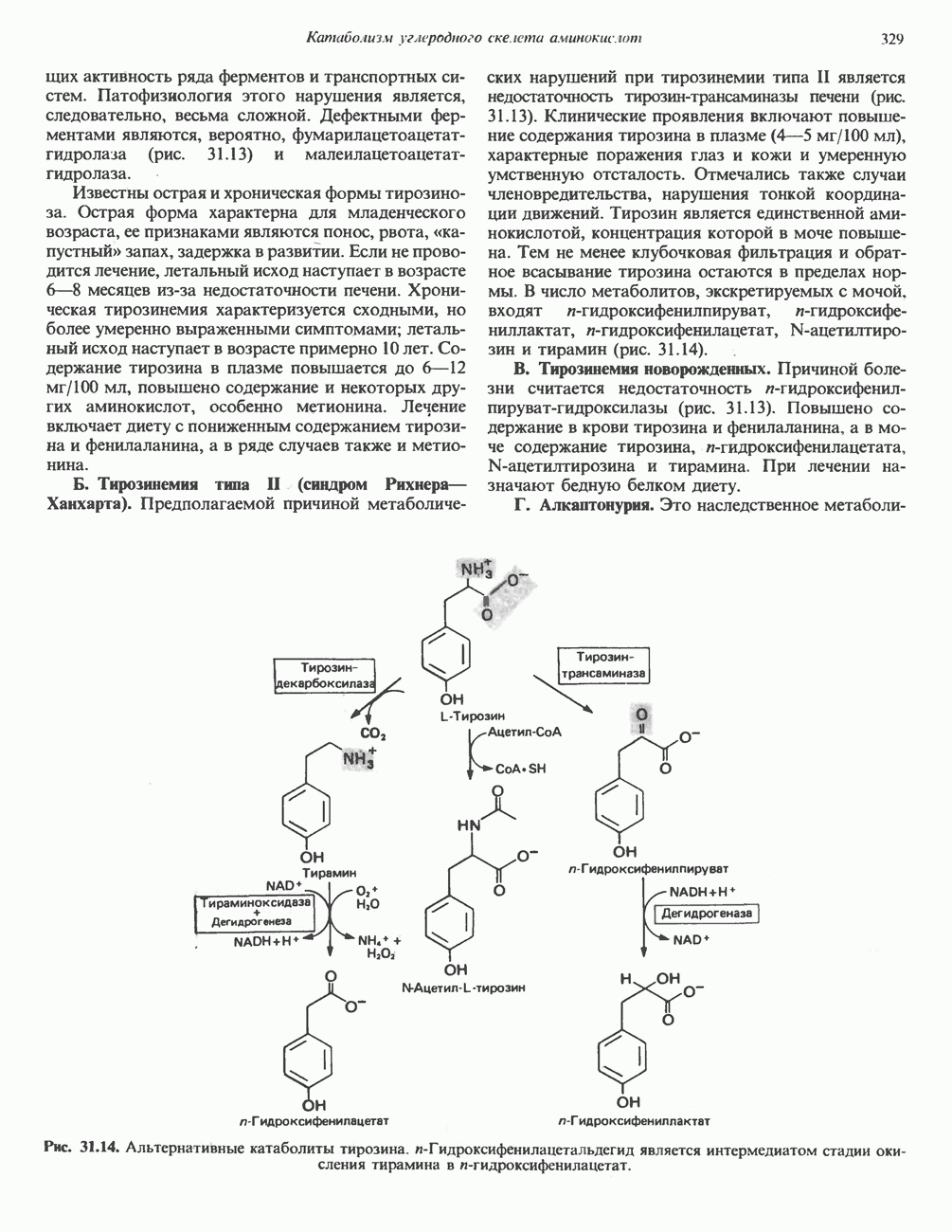
 -гидроксифенилацетат,
-гидроксифенилацетат,  -ацетилтирозин и тирамин (рис. 31.14).
-ацетилтирозин и тирамин (рис. 31.14).
 -гидроксифенил-пируват-гидроксилазы (рис. 31.13). Повышено содержание в крови тирозина и фенилаланина, а в моче содержание тирозина, и-гидроксифенилацетата, N-ацетилтирозина и тирамина. При лечении назначают бедную белком диету.
-гидроксифенил-пируват-гидроксилазы (рис. 31.13). Повышено содержание в крови тирозина и фенилаланина, а в моче содержание тирозина, и-гидроксифенилацетата, N-ацетилтирозина и тирамина. При лечении назначают бедную белком диету.




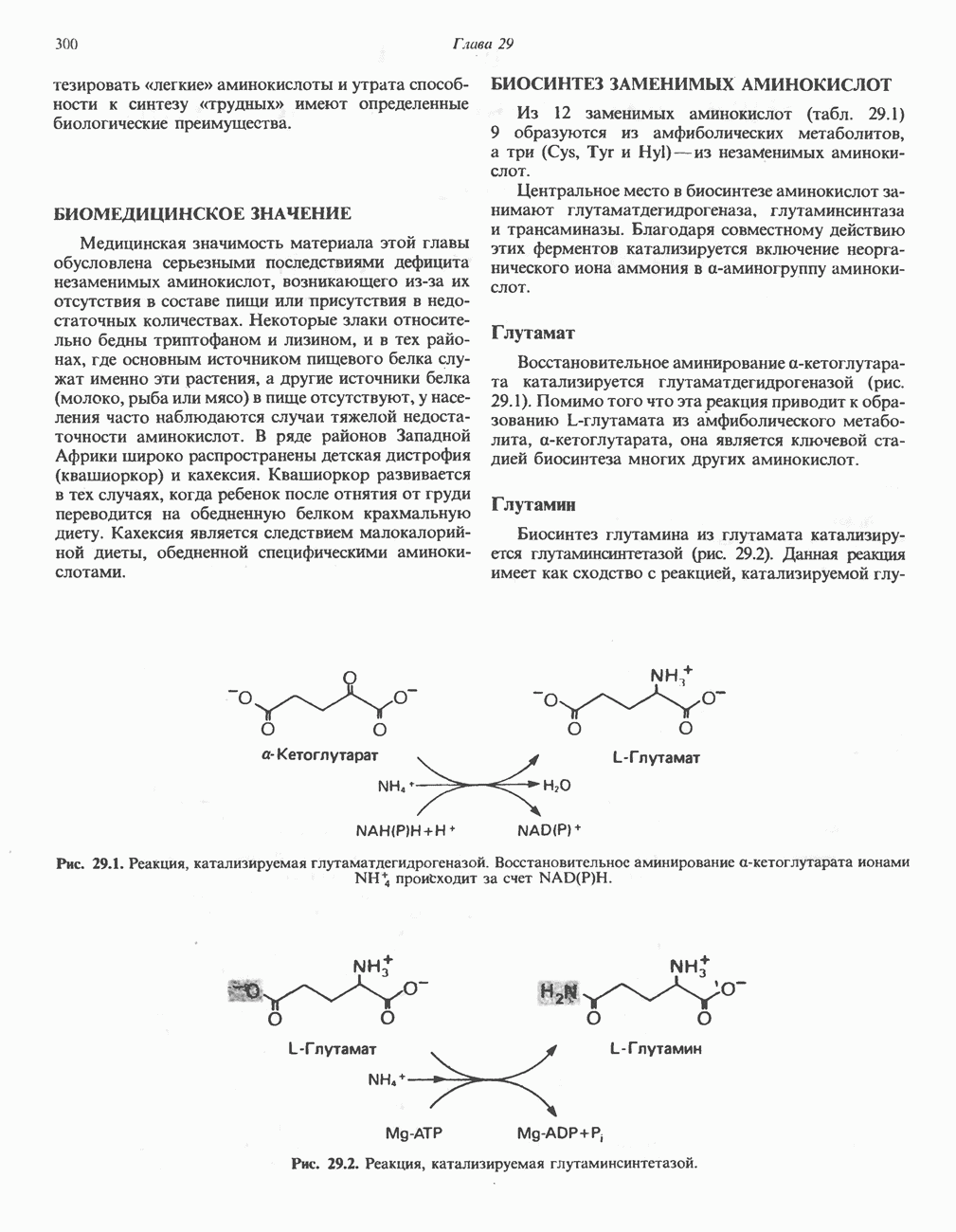
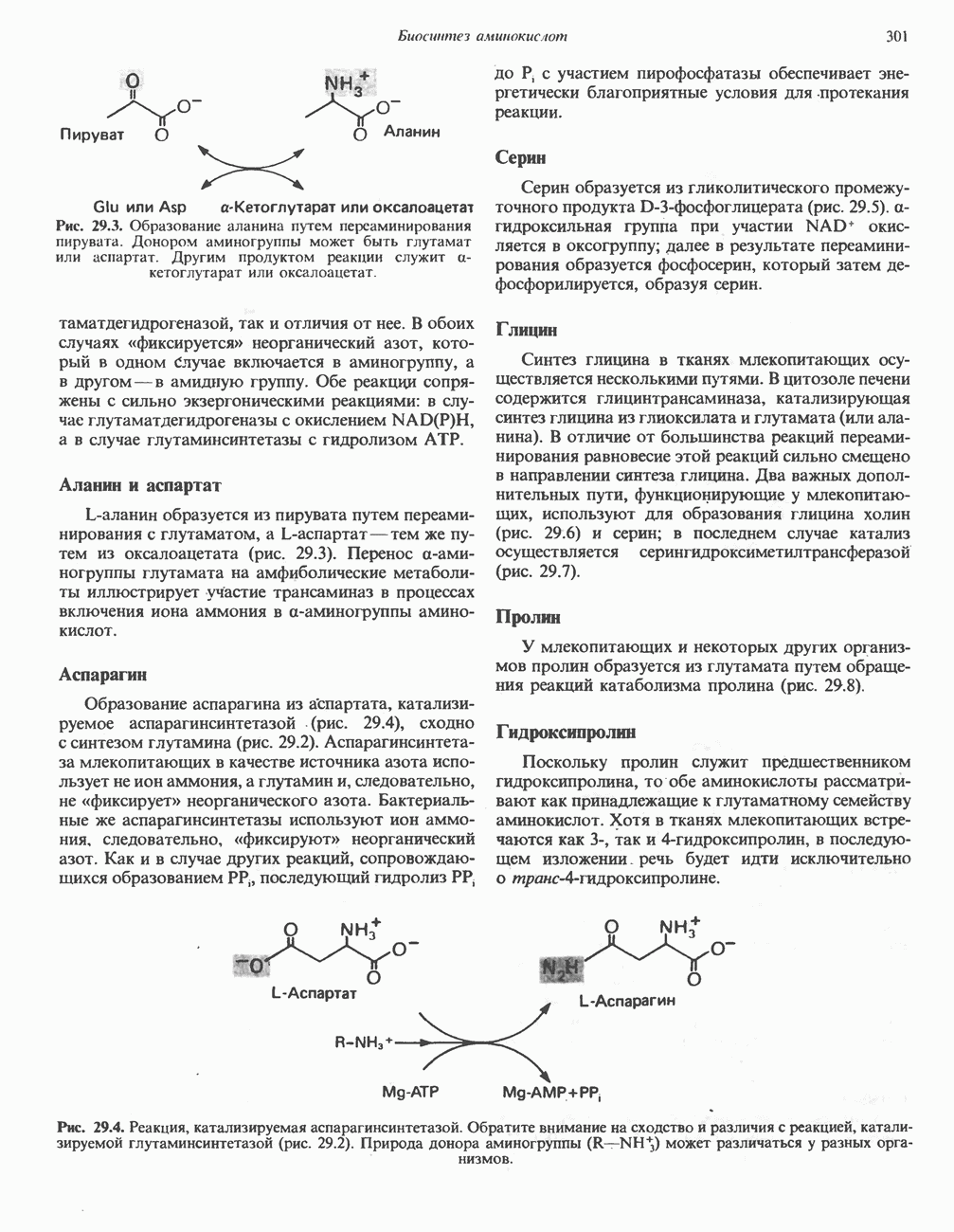
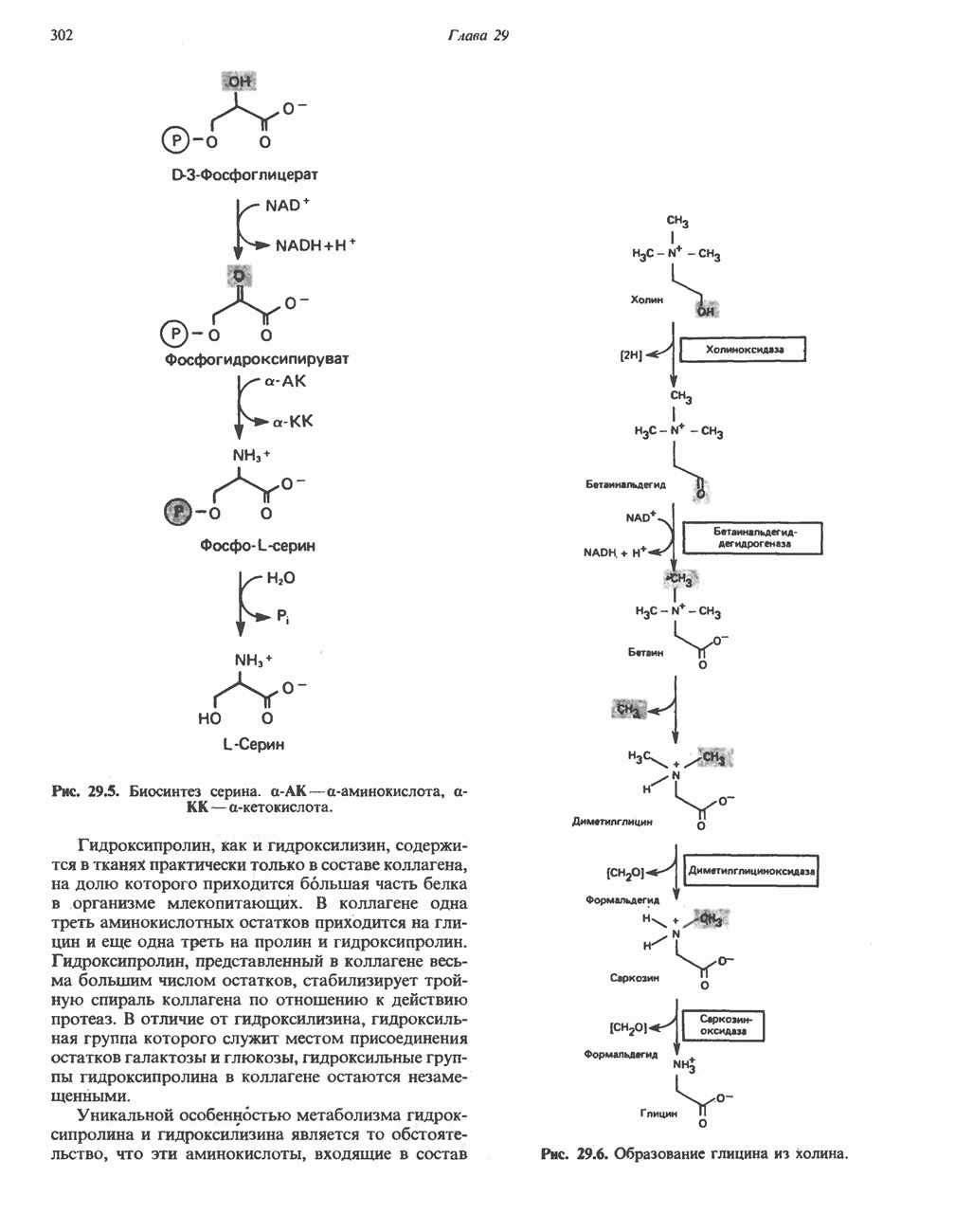
 -глутамата и
-глутамата и  Суммарный эффект этой серии реакций эквивалентен удалению е-аминогруппы лизина путем переаминировання;
Суммарный эффект этой серии реакций эквивалентен удалению е-аминогруппы лизина путем переаминировання;  -лизин и а-кетоглутарат превращаются в
-лизин и а-кетоглутарат превращаются в  и глутамат. В качестве кофакторов в процессе участвуют NAD и NADH, при этом в итоге не происходит ни окисления, ни восстановления.
и глутамат. В качестве кофакторов в процессе участвуют NAD и NADH, при этом в итоге не происходит ни окисления, ни восстановления.
 Лизин является одновременно и гликогенной, и кетогенной аминокислотой; природа же катаболитов
Лизин является одновременно и гликогенной, и кетогенной аминокислотой; природа же катаболитов  образующихся в организме млекопитающих, не установлена.
образующихся в организме млекопитающих, не установлена.

 -лизина в
-лизина в  -аминоадипат и
-аминоадипат и  -кстоадипат. Несколько стрелок подряд означают промежуточные реакции.
-кстоадипат. Несколько стрелок подряд означают промежуточные реакции.

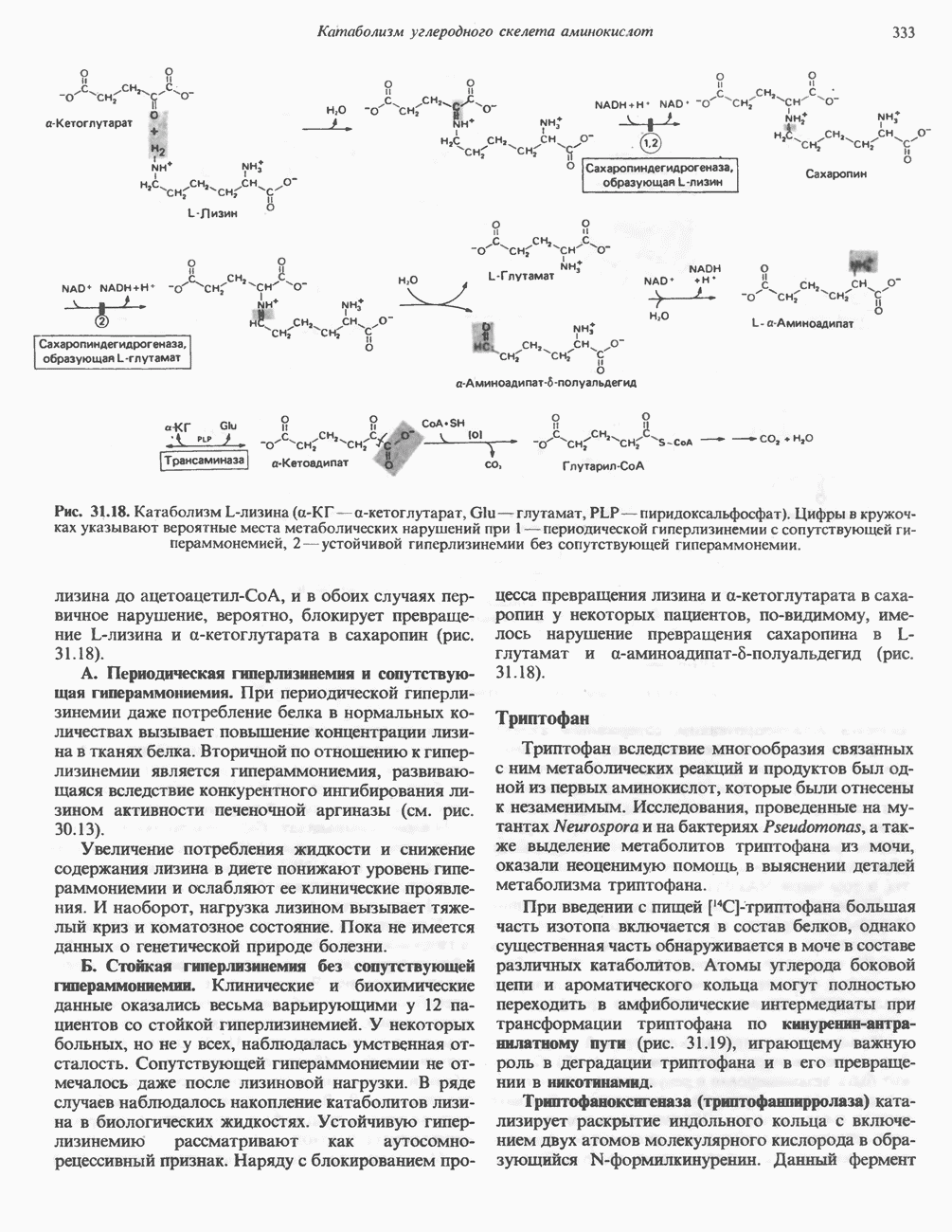
 -лизина (а-КГ—а-кетоглутарат,
-лизина (а-КГ—а-кетоглутарат,  -глутамат,
-глутамат,  -пиридоксальфосфат). Цифры в кружочках указывают вероятные места метаболических нарушений при 1 — периодической гиперлизинемии с сопутствующей гипераммонемией,
-пиридоксальфосфат). Цифры в кружочках указывают вероятные места метаболических нарушений при 1 — периодической гиперлизинемии с сопутствующей гипераммонемией,  -устойчивой гиперлизинемии без сопутствующей гипераммонемии.
-устойчивой гиперлизинемии без сопутствующей гипераммонемии.
 и в обоих случаях первичное нарушение, вероятно, блокирует превращение
и в обоих случаях первичное нарушение, вероятно, блокирует превращение  -лизина и а-кетоглутарата в сахаропин (рис. 31.18)
-лизина и а-кетоглутарата в сахаропин (рис. 31.18)
 -глутамат и
-глутамат и 
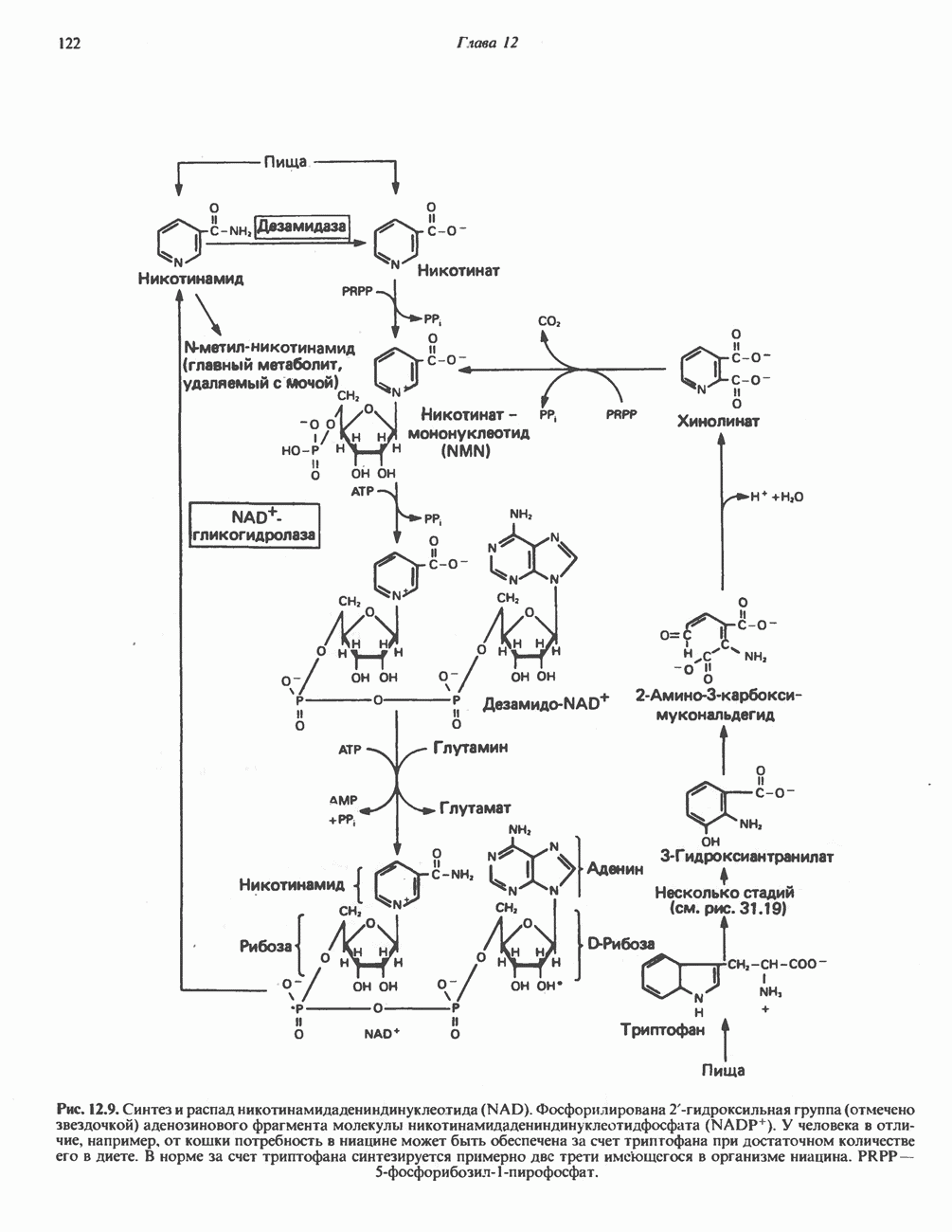
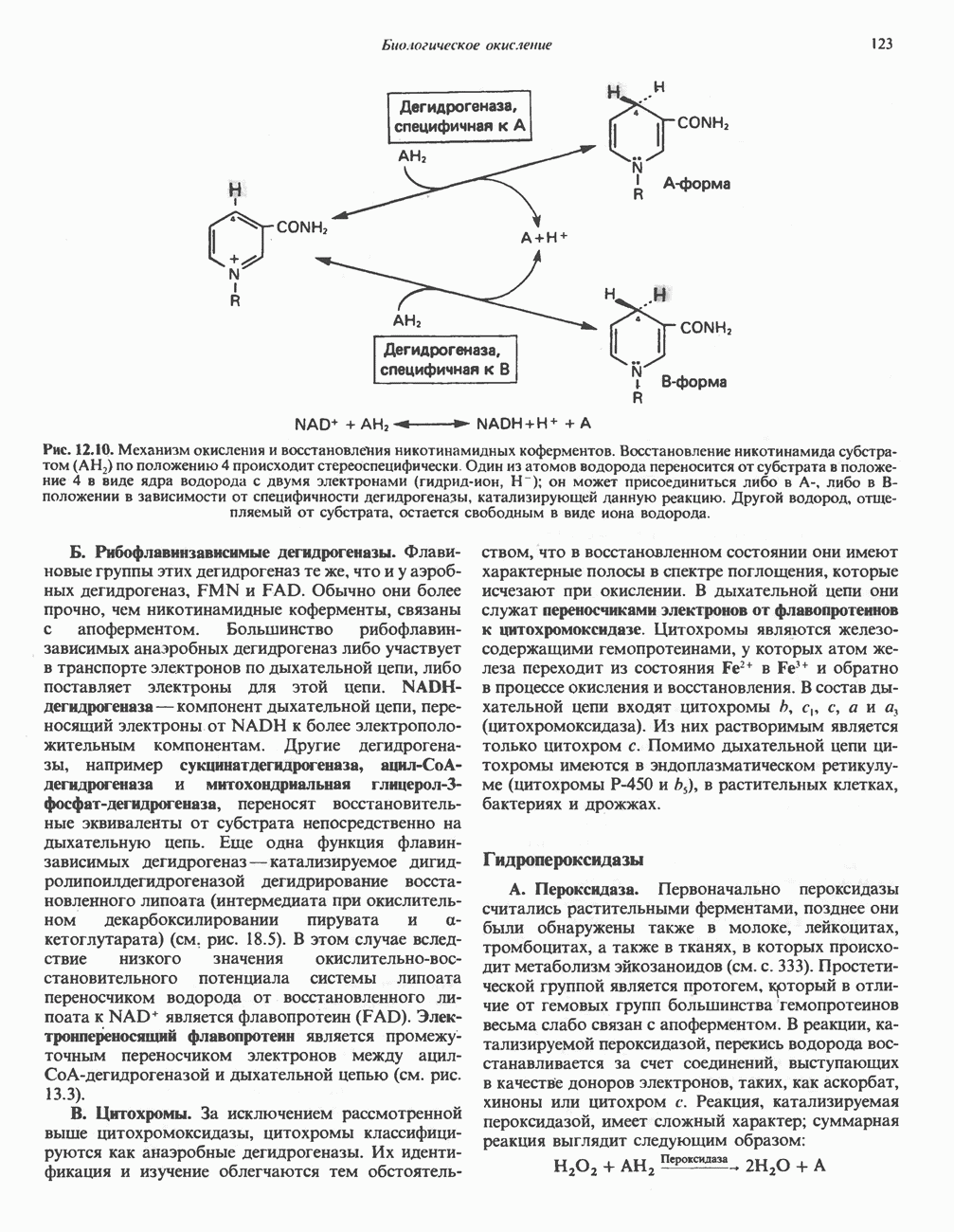
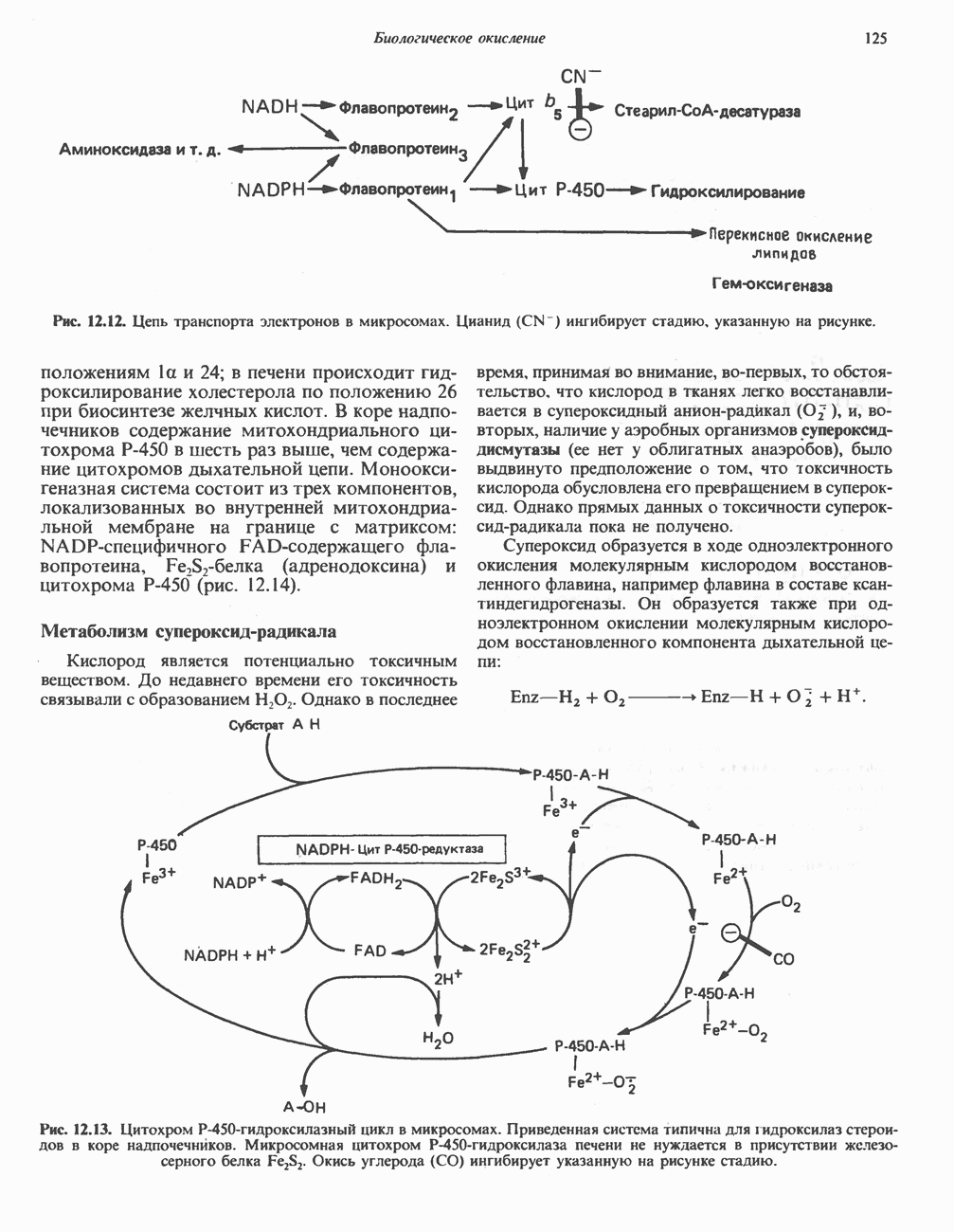
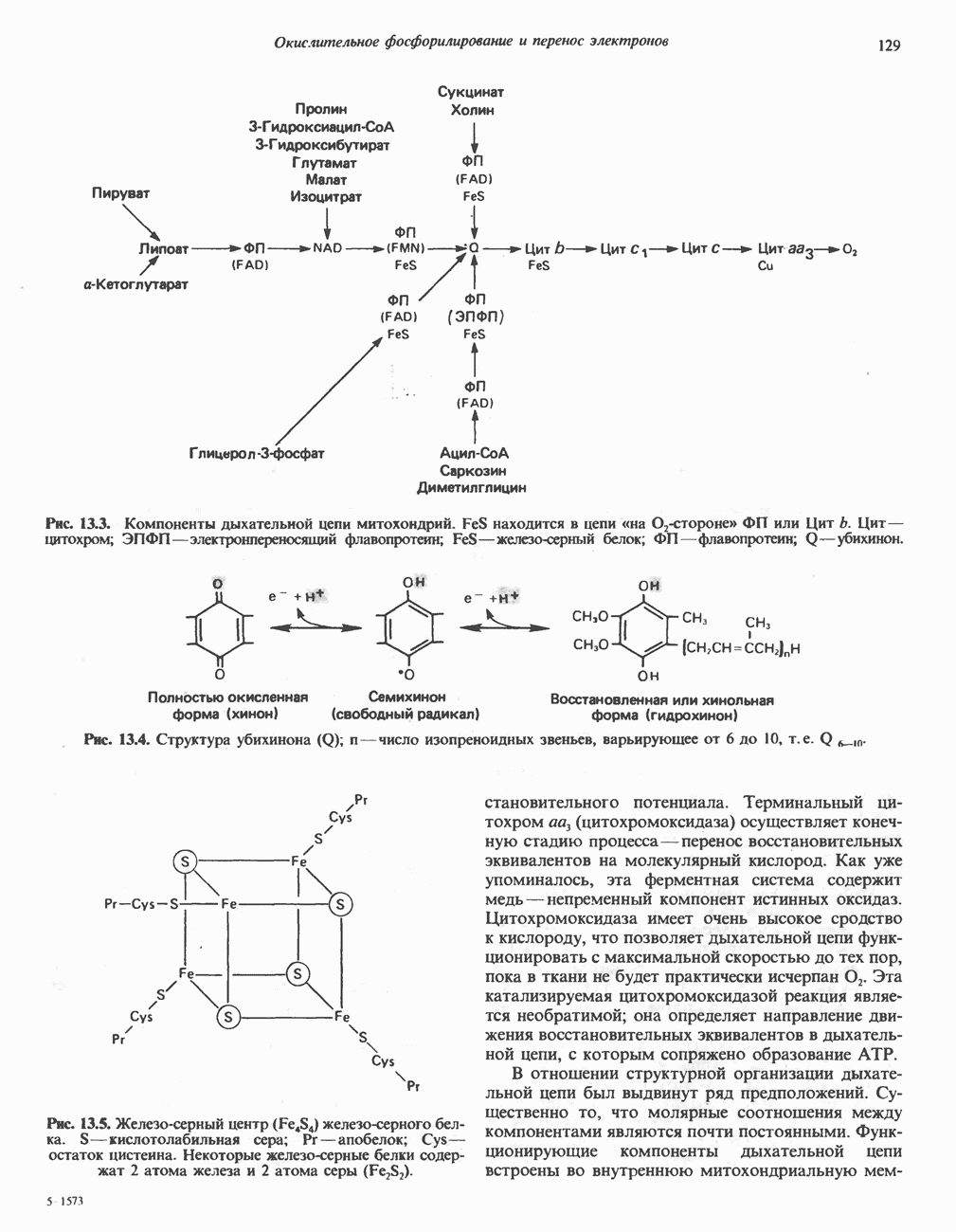
 -триптофана большая часть изотопа включается в состав белков, однако существенная часть обнаруживается в моче в составе различных катаболитов. Атомы углерода боковой цепи и ароматического кольца могут полностью переходить в амфиболические интермедиаты при трансформации триптофана по кинуренин-антранилатному пути (рис. 31.19), играющему важную роль в деградации триптофана и в его превращении в никотинамид.
-триптофана большая часть изотопа включается в состав белков, однако существенная часть обнаруживается в моче в составе различных катаболитов. Атомы углерода боковой цепи и ароматического кольца могут полностью переходить в амфиболические интермедиаты при трансформации триптофана по кинуренин-антранилатному пути (рис. 31.19), играющему важную роль в деградации триптофана и в его превращении в никотинамид.
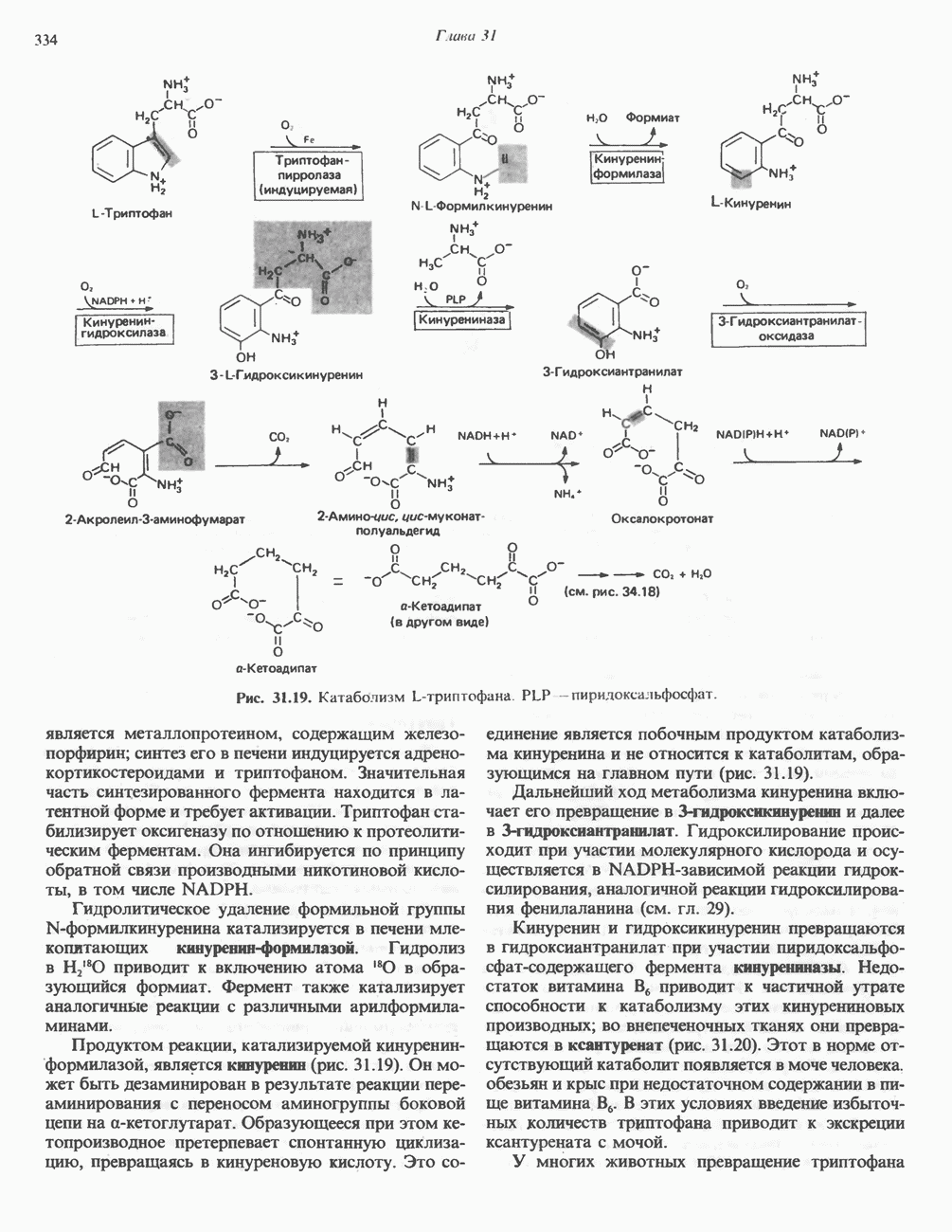
 -формилкинуренин. Данный фермент
-формилкинуренин. Данный фермент
 приводит к включению атома
приводит к включению атома  в образующийся формиат. Фермент также катализирует аналогичные реакции с различными арилформиламинами.
в образующийся формиат. Фермент также катализирует аналогичные реакции с различными арилформиламинами.
 -зависимой реакции гидроксилирования, аналогичной реакции гидроксилирования фенилаланина (см. гл. 29).
-зависимой реакции гидроксилирования, аналогичной реакции гидроксилирования фенилаланина (см. гл. 29).

 и триптофана.
и триптофана.