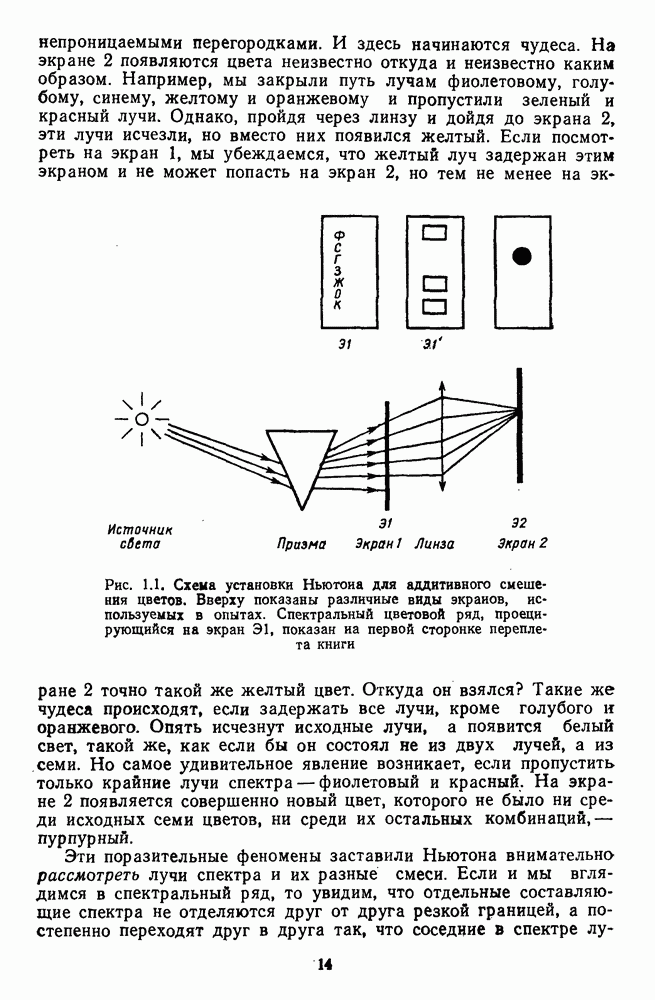
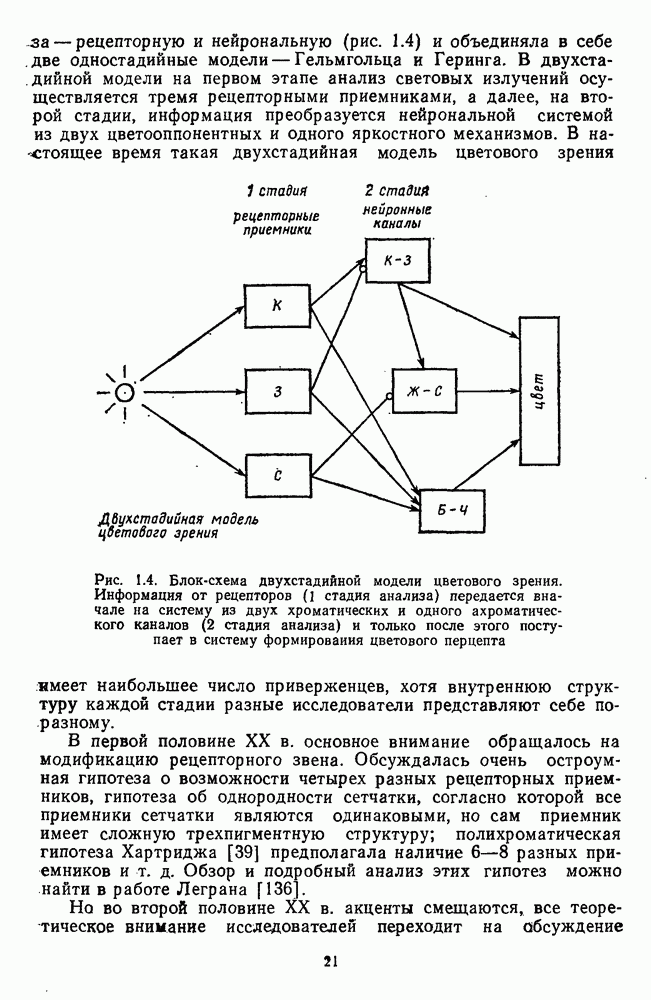
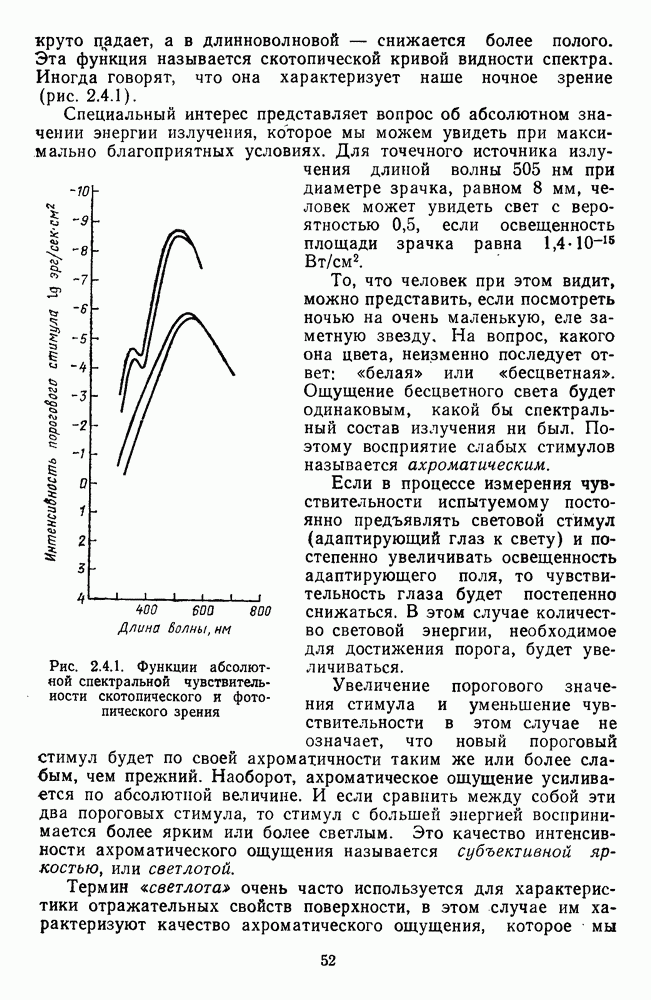
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
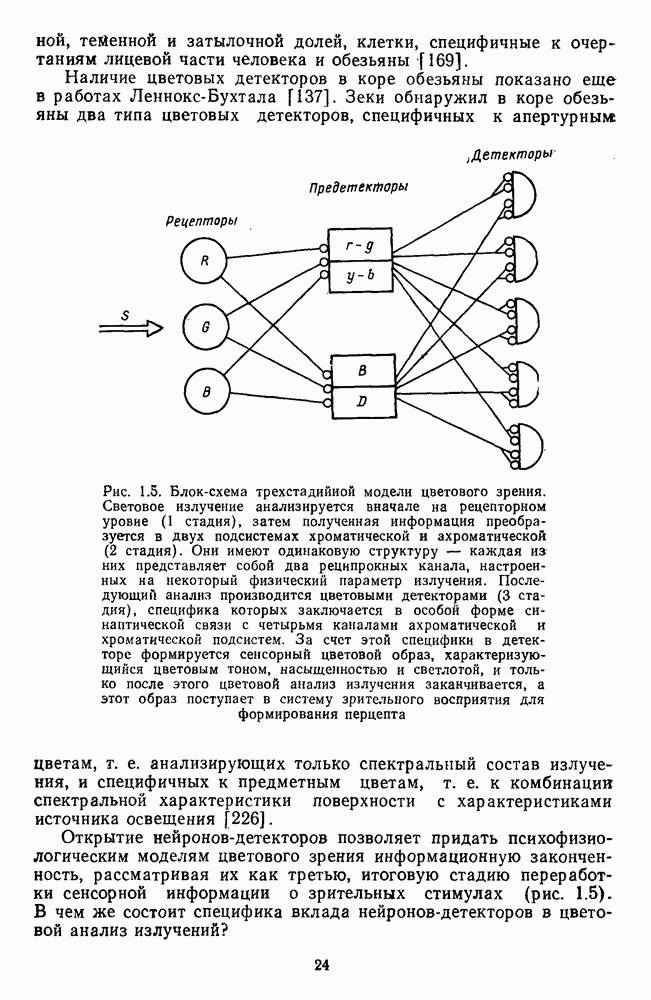
1.9. Трехстадийная концепция цветового зренияПринципиальное значение для ответа на этот вопрос имеет одно из важнейших открытий нейрофизиологии XX в. — обнаружение в сенсорных системах у животных нейронов—детекторов. Первые исследования в этом направлении, проведенные на низших животных (лягушках) [17] и высших животных (кошках) [116; 117], выявили, что в сетчатке у лягушки и в коре у кошки существуют клетки, селективно реагирующие либо на освещение (on - клетки по Кафлеру или В — клетки по Юнгу), либо на затемнение (off- клетки по Кафлеру или D — клетки по Юнгу) локальных участков рецепторной поверхности сетчатки. Их назвали соответственно детекторами света и детекторами темноты. Последовавшие затем серии электрофизиологических исследований Хьюбела и Визеля на мозге кошки и обезьяны взбудоражили научный мир открытием целого ряда детекторов, настроенных на самые различные характеристики сигналов. Наиболее эффектными были открытия детекторов ориентации линий, которые являются теперь классическим примером для иллюстрации принципов переработки сенсорной информации в коре больших полушарий. Хьюбел и Визел показали, что детектирование ориентации линии происходит со значительной селективностью: диапазон углов оптимальной для данной клетки ориентации не превышает 10—15°. Клетки, детектирующие одну и ту же ориентацию, сгруппированы вертикально по слоям в виде «колонки» нейронов, причем соседние колонки детектируют «соседние» ориентации. Последовательное смещение микроэлектрода параллельно поверхности коры обнаруживает серию колонок, последовательно настроенных на все больший и больший угол поворота линии в поле зрения. Набор колонок, необходимых для детектирования всех ориентаций от 0 до 180° (макроколонка), занимает на поверхности коры участок длиной 800 микрон. Эти исследования показали, что набор отдельных детекторов, каждый из которых настроен «а свой угол ориентации, объединяется в нейронную систему, детектирующую всевозможные ориентации (макроколонку). Такая «ейронная система оказывается характеризуется не только функциональной, но и морфологической структурой. Ориентация линий не является привилегированным признаком детектирования. Выявлены детекторы направления движения, доминантности левого или правого глаза, детекторы удаленности объекта (глубины), детекторы углов и др. Показательно, что чем более удалена от сетчатки область мозга, в которой обнаружены те или иные детекторы, тем более избирательны, тем более специфичны они по детектируемым характеристикам стимула. Например, клетки II и IV слоев 17 поля коры по Бродману (которое называется стриарной корой или первичной проекционной зоной), на которых непосредственно оканчиваются волокна, идущие из подкорковых центров, детектируют такие простые признаки, как граница света и тени на данном участке сетчатки, а клетки, следующие за ними в 17 и 18 полях, выделяют уже ориентацию линии независимо от местоположения на сетчатке и углы, следующие за ними клетки 18 и 19 полей уже специфичны не только к ориентации линии, но и ее ширине и длине и т. д. В самые последние годы в Англии, ФРГ, Шотландии исследователи обнаружили у обезьян в задней височной извилине, лежащей на границе височной, теменной и затылочной долей, клетки, специфичные к очертаниям лицевой части человека и обезьяны [169].
Рис. 1.5. Блок-схема трехстадийиой модели цветового зрения. Световое излучение анализируется вначале на рецепторном уровне (1 стадия), затем полученная информация преобразуется в двух подсистемах хроматической и ахроматической (2 стадия). Они имеют одинаковую структуру — каждая из них представляет собой два реципрокных канала, настроенных на некоторый физический параметр излучения. Последующий анализ производится цветовыми детекторами (3 стадия), специфика которых заключается в особой форме синаптической связи с четырьмя каналами ахроматической и хроматической подсистем. За счет этой специфики в детекторе формируется сенсорный цветовой образ, характеризующийся цветовым тоном, насыщенностью и светлотой, и только после этого цветовой анализ излучения заканчивается, а этот образ поступает в систему зрительного восприятия для формирования перцепта Наличие цветовых детекторов в коре обезьяны показано еще в работах Леннокс-Бухтала [137]. Зеки обнаружил в коре обезьяны два типа цветовых детекторов, специфичных к апертурным цветам, т. е. анализирующих только спектральный состав излучения, и специфичных к предметным цветам, т. е. к комбинации спектральной характеристики поверхности с характеристиками источника освещения [226]. Открытие нейронов-детекторов позволяет придать психофизиологическим моделям цветового зрения информационную законченность, рассматривая их как третью, итоговую стадию переработки сенсорной информации о зрительных стимулах (рис. 1.5). В чем же состоит специфика вклада нейронов-детекторов в цветовой анализ излучений? Этот вопрос на самом деле может быть задан и в более общей форме: в чем состоит специфика вклада нейронов-детекторов вообще в анализ зрительных стимулов? Исследованием этой общей проблемы в настоящее время занято значительное число исследователей [40; 43; 33; 117; 23; 45]. Но мы рассмотрим здесь только одну из гипотез, касающуюся цветового зрения. Эта гипотеза разрабатывается авторами данной книги в течение последних десяти лет на базе экспериментальных исследований цветового зрения человека и животных [30; 12; 31; 29; 14]. В основании гипотезы лежат две новые идеи о функциональной организации цветового анализатора излучений. Для иллюстрации этих идей можно обратиться к схеме трехстадийной модели цветового зрения, приведенной на рис. 1.5. Первая идея заключается в том, что пострецепторная нейронная сеть представляет собой набор попарно реципрокных каналов передачи информации. Цветовой анализатор содержит в своем составе только две пары таких каналов — хроматическую и ахроматическую. Каждая пара каналов специализируется на свой физический параметр излучения. Хроматические (оппонентные) каналы анализируют частотный состав излучения, который затем в детекторе преобразуется в такую сенсорную характеристику, как цветовой тон. Ахроматические (неоппонентные каналы) анализируют интенсивность излучения, которая в детекторе цвета преобразуется в яркость или светлоту. Если бы зрительная система анализировала еще какие-либо физические параметры излучения (например, фазу электромагнитной волны), то в составе нейронной сети анализатора появилась бы еще одна пара таких же реципрокных каналов для анализа фазовых характеристик излучения. Две нейрональные подсистемы — хроматическая и ахроматическая — получают одну и ту же информацию от рецепторов, но перерабатывают ее независимо друг от друга и каждая по своему. Переработанная в них информация поступает на множество цветовых детекторов, каждый из которых, возбуждаясь, вызывает в коре сенсорный образ отдельного апертурного цвета. Вторая идея заключается в том, что нейроны-детекторы имеют специфическую организацию синаптических входов, через которые поступают возбуждения от хроматических и ахроматических каналов. Эта специфика во взаимной зависимости коэффициентов синаптической передачи на входе нейрона-детектора, которые подобраны так, что сумма квадратов этих коэффициентов всегда постоянна. В математическом смысле значения коэффициентов передачи информации от цветовых каналов к нейронам-детекторам удовлетворяют условию расположения точек на сфере. Отсюда и яазвание этой трехстадийной модели — сферическая модель цветового зрения. Преобразование возбуждений четырех цветовых каналов системой четырех нормированных по своим весам синапсов каждого цветового детектора приводит к тому, что возбуждение детектора цвета на поверхности сферы определяется окончательно только через три независимые угловые характеристики, представляющие тон, светлоту и насыщенность. Причем если цветовой тон формируется в подсистеме хроматических каналов, светлота — в подсистеме ахроматических каналов, то насыщенность формируется целиком на третьей стадии анализа, на уровне цветовых детекторов как следствие нормировки, обеспечивающей сферическую организацию цветов. Этим же объясняется отсутствие однозначной связи между насыщенностью и каким-либо физическим параметром излучения. Целью этого раздела было формирование у читателя исторического контекста, без которого трудно установить масштабные соотношения между различными теоретическими и экспериментальными работами в области цветового зрения. На обложках книги приводятся цветные иллюстрации, которые дают некоторое представление о моделях цветового зрения, характерных для разных исторических периодов. На лицевой стороне обложки внизу показан линейный цветовой спектр, в котором предполагается прямая связь между длиной волны излучения и цветом, а наверху — цветовой круг Ньютона, который объясняет, почему нельзя однозначно соотносить цвета непосредственно с отдельными излучениями. На четвертой странице обложки приводятся цветные иллюстрации геометрических моделей цветового зрения, основанных на различных концепциях строения цветового анализатора (сверху вииз): цветовой треугольник, евклидова плоскость оппонентных цветов и сферическая модель цвета.
|
 |
Оглавление
|