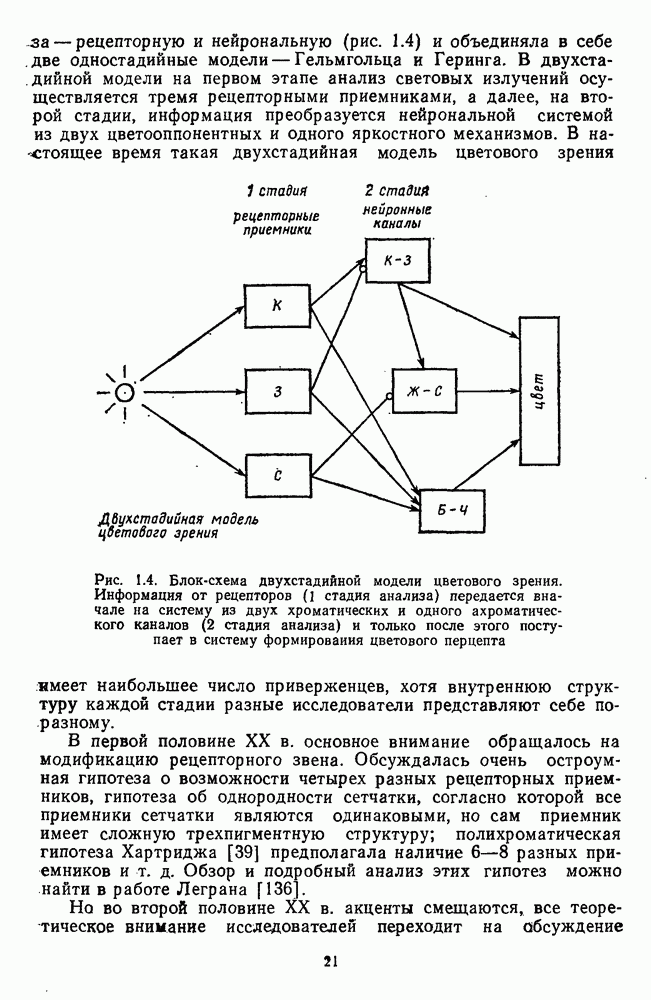
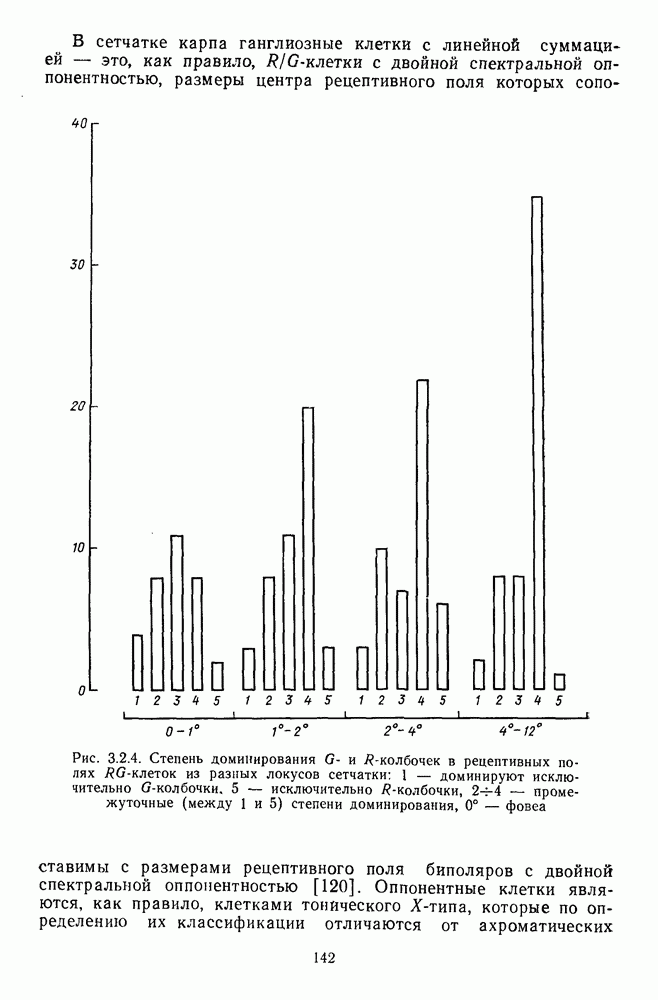
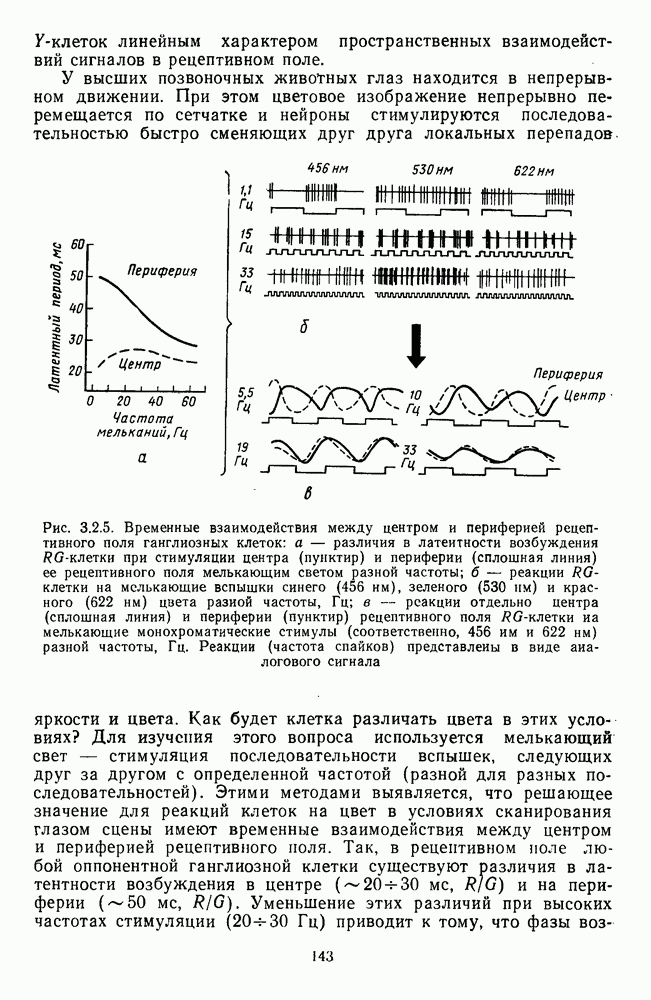
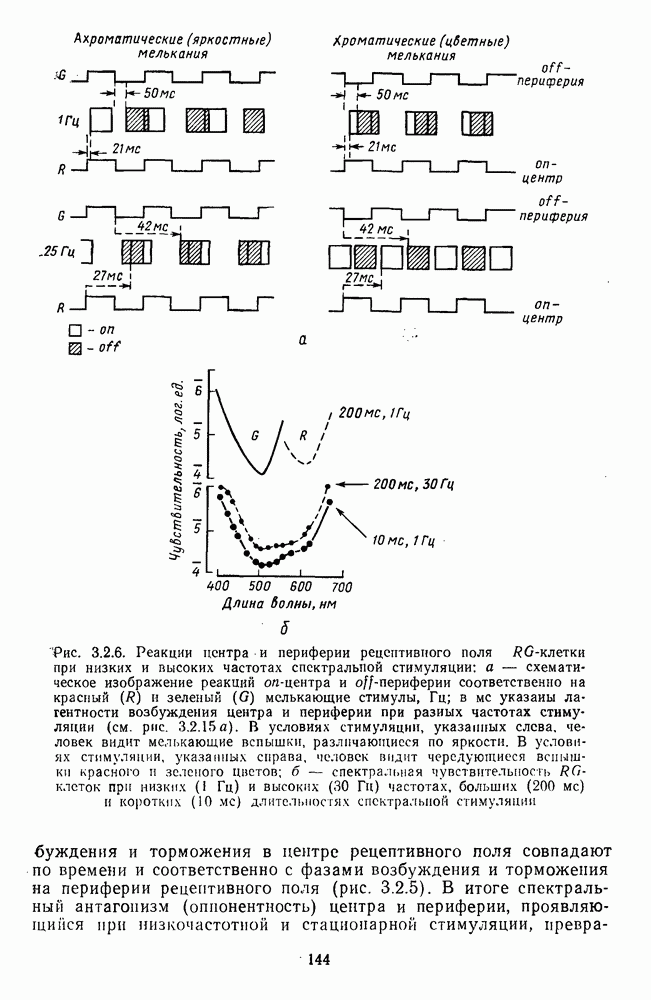
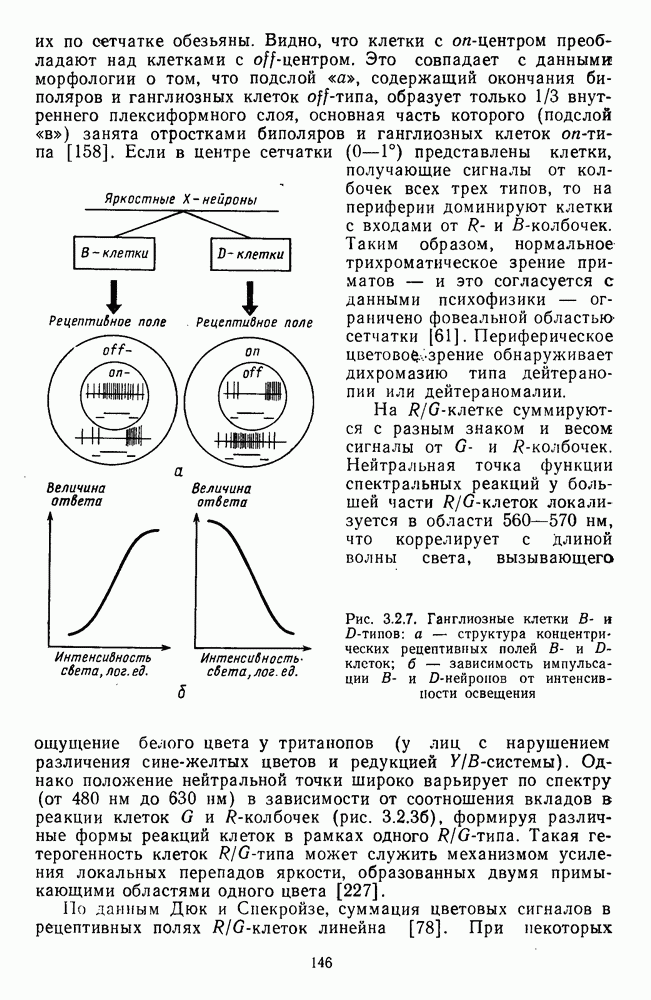
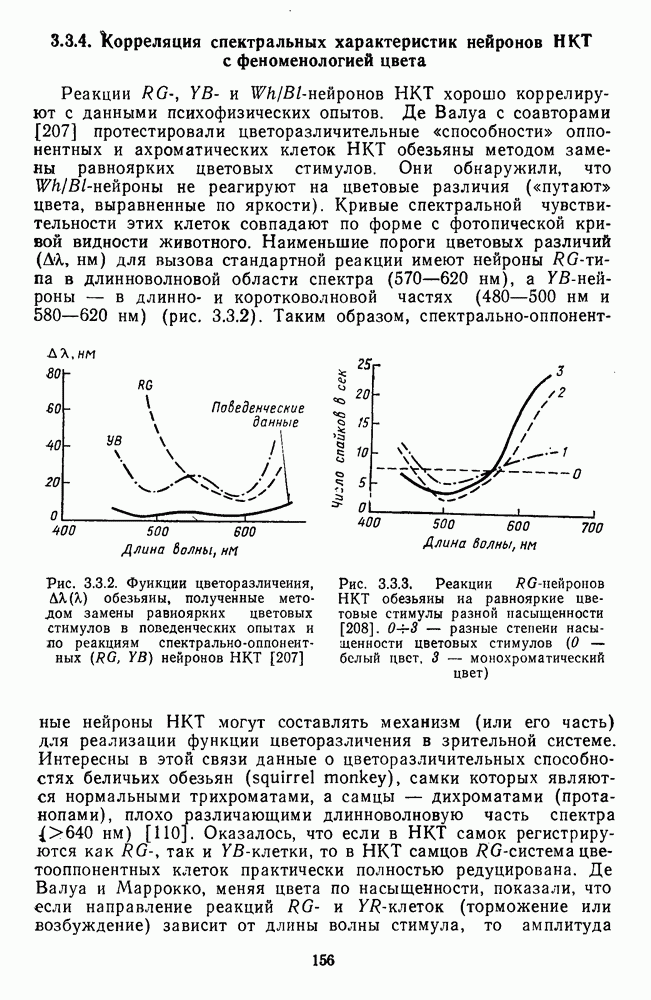
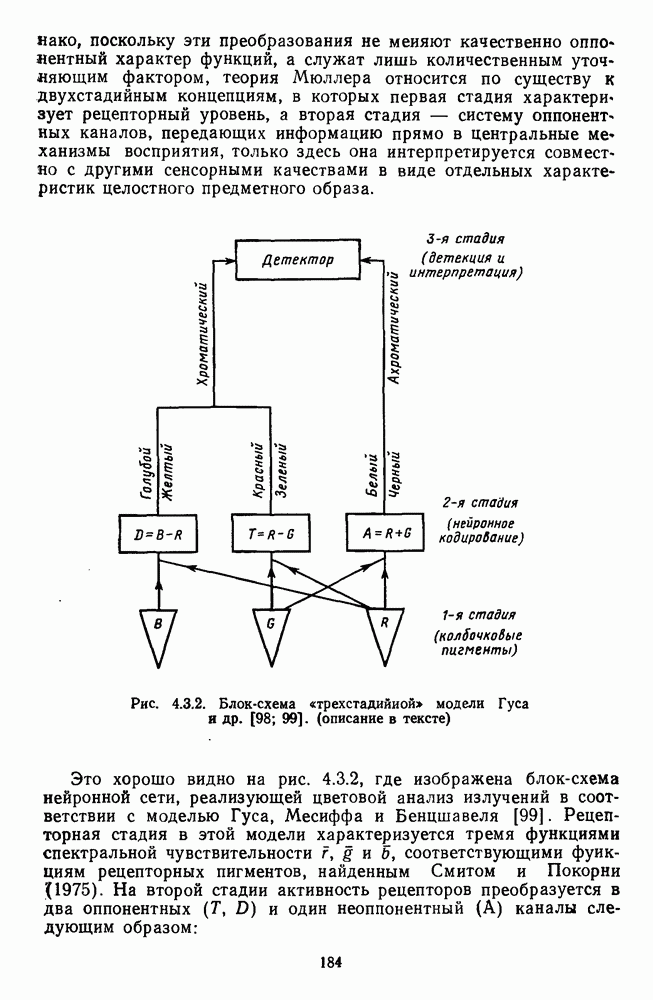
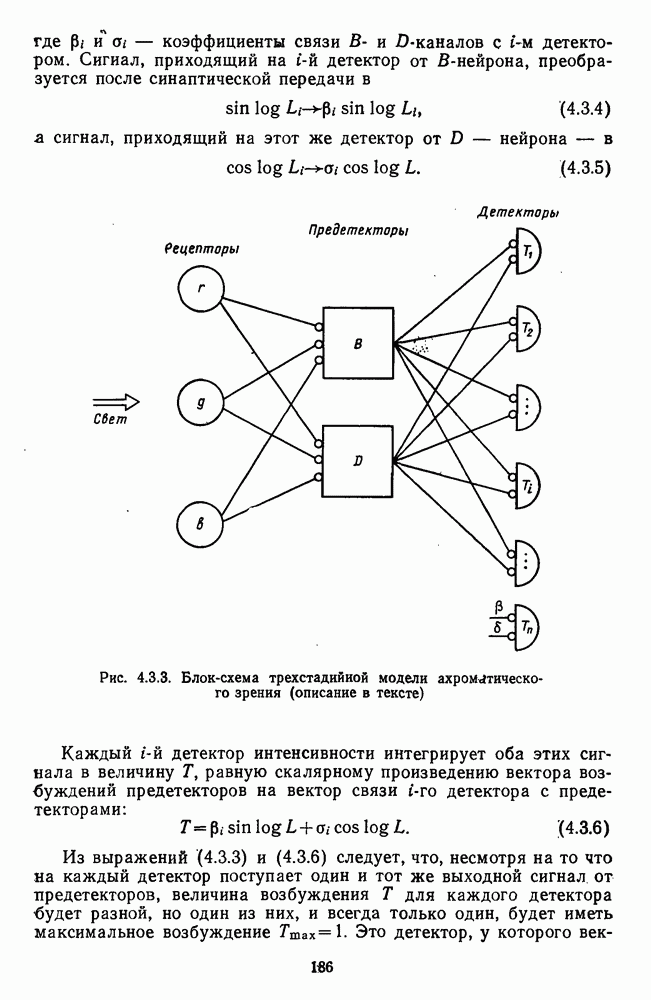
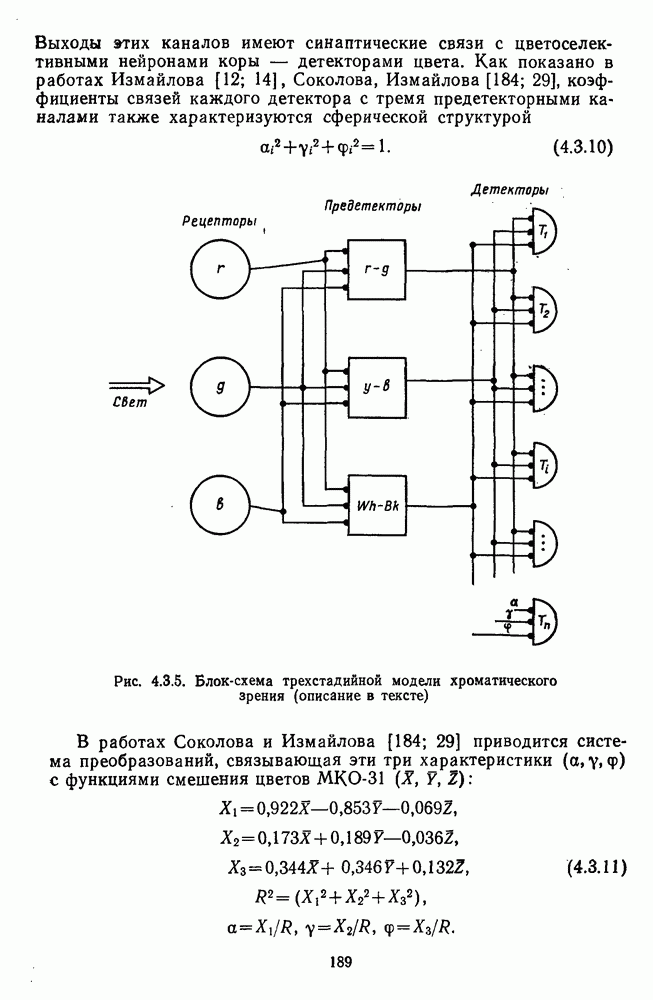
3.3.2. Спектрально-оппонентные и ахроматические нейроны
По реакциям на белый свет нейроны НКТ, как и ганглиозные клетки сетчатки, делятся на нейроны  на фазические и тонические. По реакциям на спектральные стимулы нейроны НКТ делят на спектрально-оппонентные и ахроматические. Наиболее подробно свойства этих клеток изучены в лабораториях известных американских ученых Хьюбела и Визеля и Де Валуа (1975) на НКТ обезьяны [40; 209]. Де Валуа с сотрудниками представили данные о наличии в НКТ разных типов оппонентных и ахроматических нейронов (рис. 3.3.1):
на фазические и тонические. По реакциям на спектральные стимулы нейроны НКТ делят на спектрально-оппонентные и ахроматические. Наиболее подробно свойства этих клеток изучены в лабораториях известных американских ученых Хьюбела и Визеля и Де Валуа (1975) на НКТ обезьяны [40; 209]. Де Валуа с сотрудниками представили данные о наличии в НКТ разных типов оппонентных и ахроматических нейронов (рис. 3.3.1):  увеличивающие частоту фоновых разрядов при стимуляции центра рецептивного поля длинноволновым излучением и уменьшающие ее при стимуляции периферии рецептивного поля средневолновым излучением, и
увеличивающие частоту фоновых разрядов при стимуляции центра рецептивного поля длинноволновым излучением и уменьшающие ее при стимуляции периферии рецептивного поля средневолновым излучением, и  обнаруживающие обратное направление реакций при той же стимуляции; клетки
обнаруживающие обратное направление реакций при той же стимуляции; клетки  -типа, тормозящиеся коротковолновыми и возбуждающиеся средне- и длинноволновыми излучениями, и
-типа, тормозящиеся коротковолновыми и возбуждающиеся средне- и длинноволновыми излучениями, и  реагирующие противоположным образом; неоппонентные
реагирующие противоположным образом; неоппонентные  возбуждающиеся стимулами любой длины волны с максимумом реакций при 560—580 нм, и
возбуждающиеся стимулами любой длины волны с максимумом реакций при 560—580 нм, и  тормозящиеся излучениями из любой области спектра с максимумом торможения при 560—580 нм. Де Валуа сгруппировал эти нейроны в три цветовых канала: красно-зеленый,
тормозящиеся излучениями из любой области спектра с максимумом торможения при 560—580 нм. Де Валуа сгруппировал эти нейроны в три цветовых канала: красно-зеленый,  желто-синий,
желто-синий,  и белочерный,
и белочерный,  Последний канал включает в себя
Последний канал включает в себя  -клетки, являющиеся аналогами ганглиозных клеток В- и
-клетки, являющиеся аналогами ганглиозных клеток В- и  -типов, кодирующих «свет» и «темноту». Типы функционально-пространственной организации рецептивных полей оппонентных нейронов НКТ приведены в табл. 3. Видно, что и по строению, и по частоте встречаемости разных типов нейроны НКТ мало отличаются от
-типов, кодирующих «свет» и «темноту». Типы функционально-пространственной организации рецептивных полей оппонентных нейронов НКТ приведены в табл. 3. Видно, что и по строению, и по частоте встречаемости разных типов нейроны НКТ мало отличаются от
ганглиозных клеток сетчатки. Нейроны с оппонентными рецептивными полями составляют 60—80% от общего числа клеток НКТ, где преобладают  -клетки (50-60% от общего числа оппонентных нейронов).
-клетки (50-60% от общего числа оппонентных нейронов).

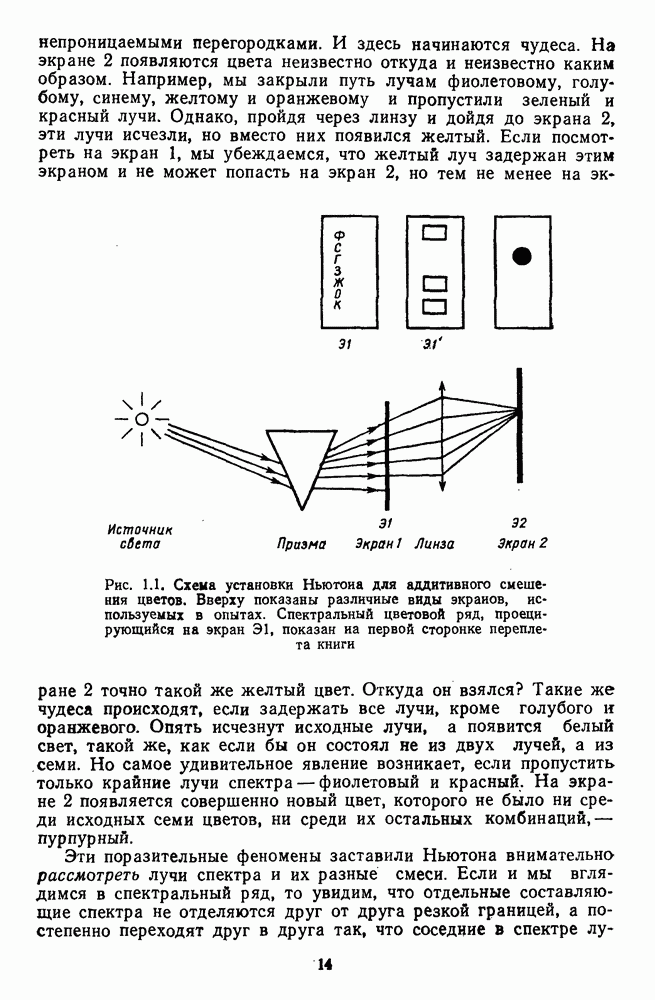
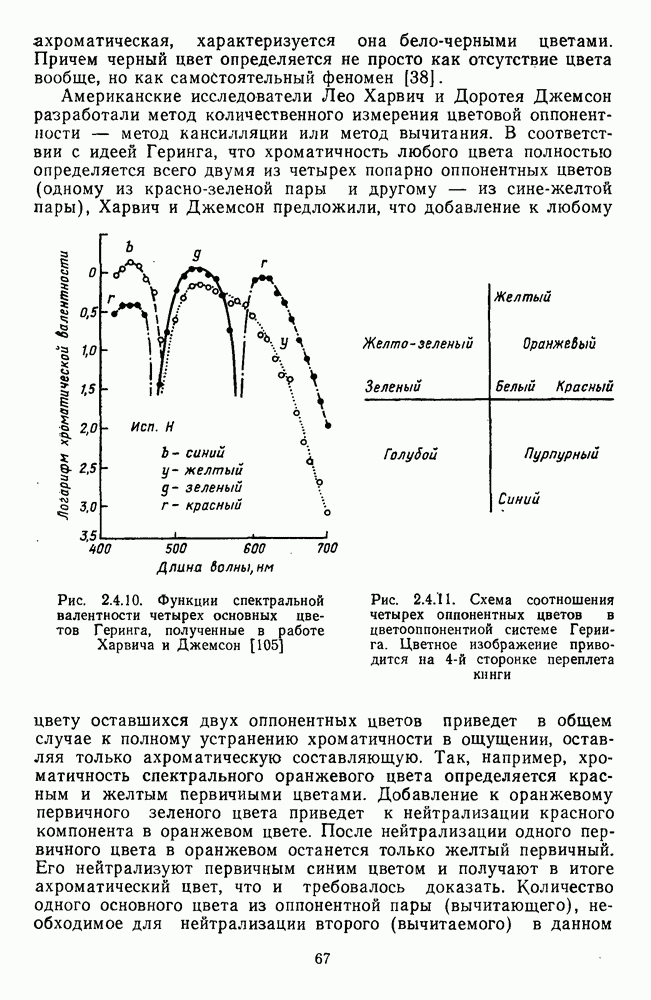
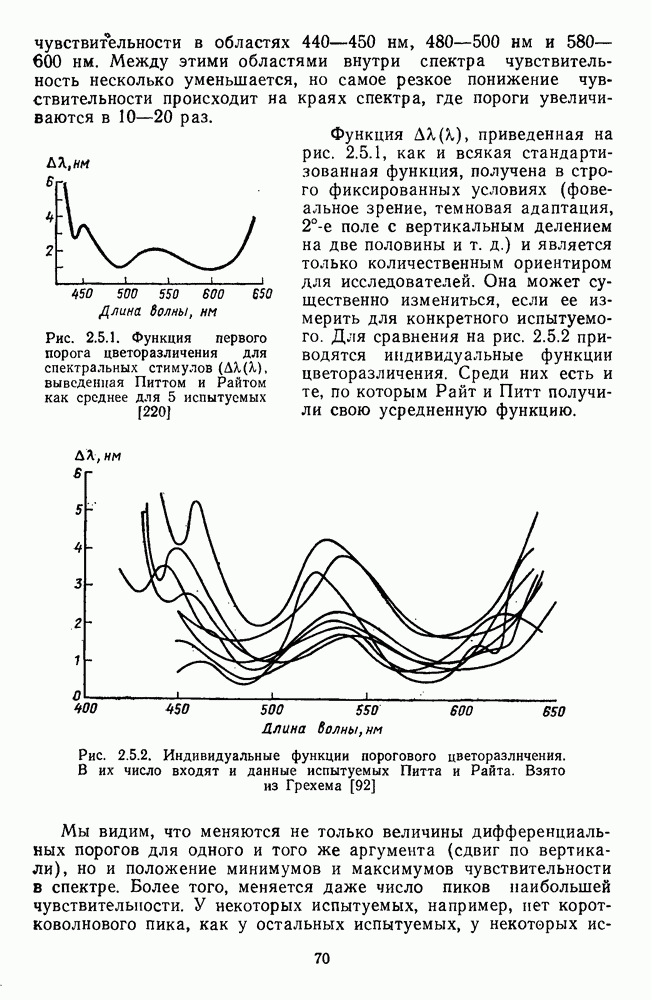
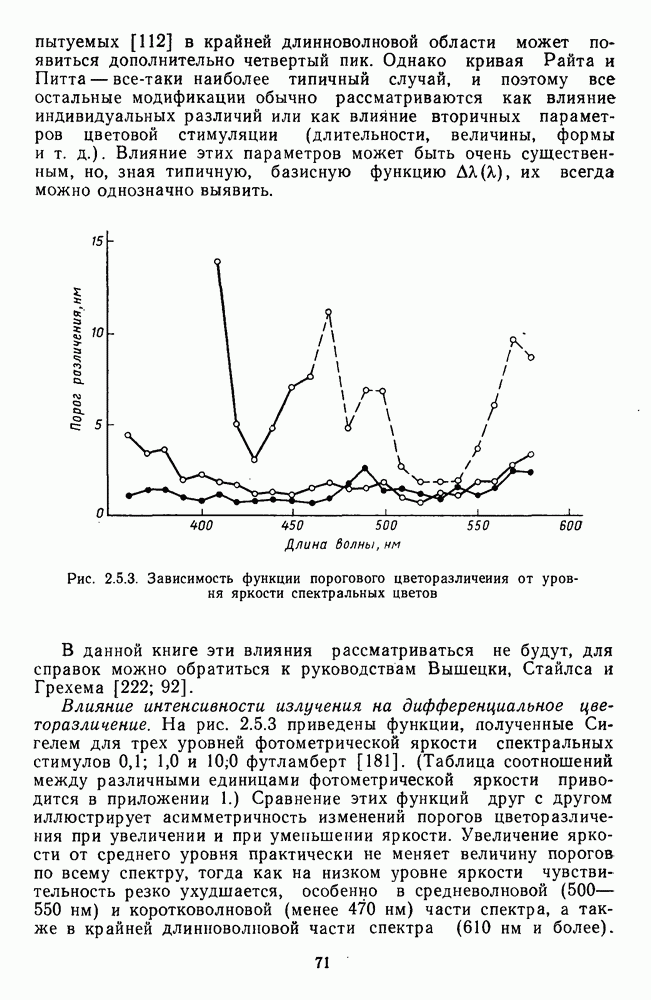
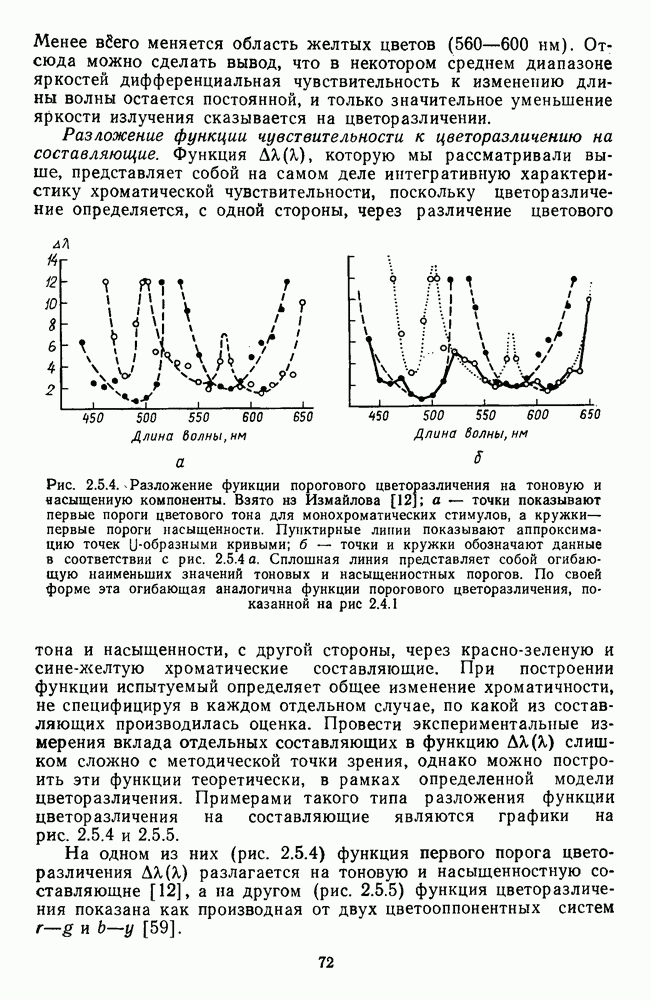
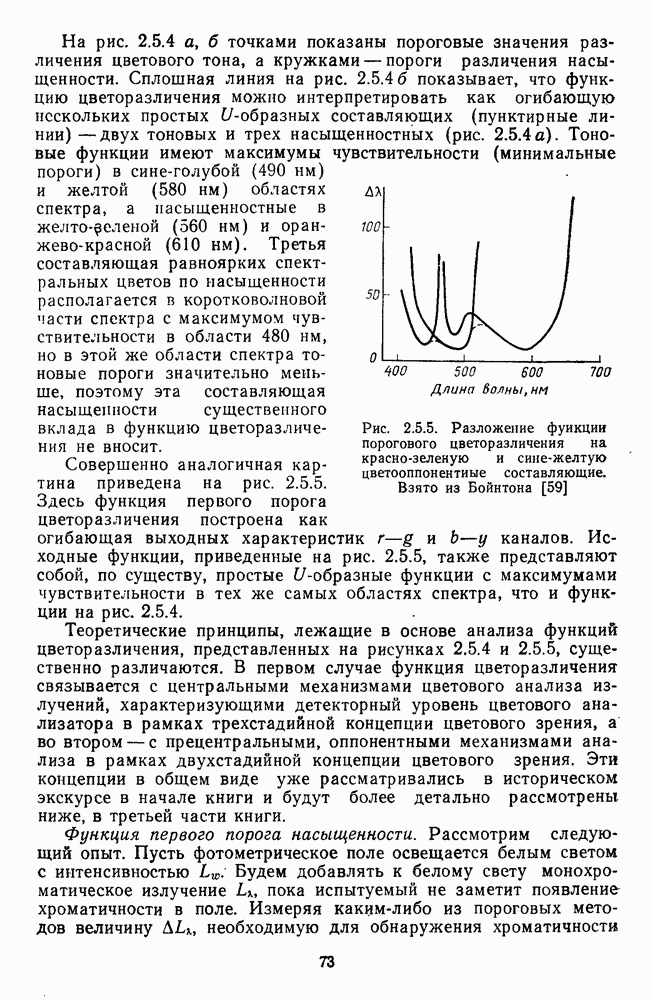
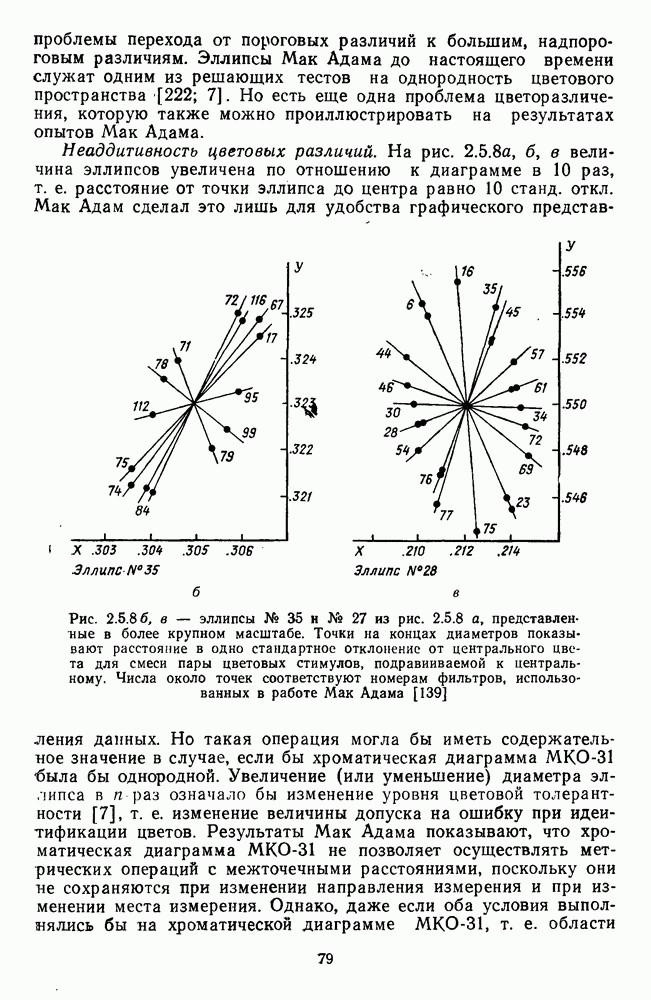
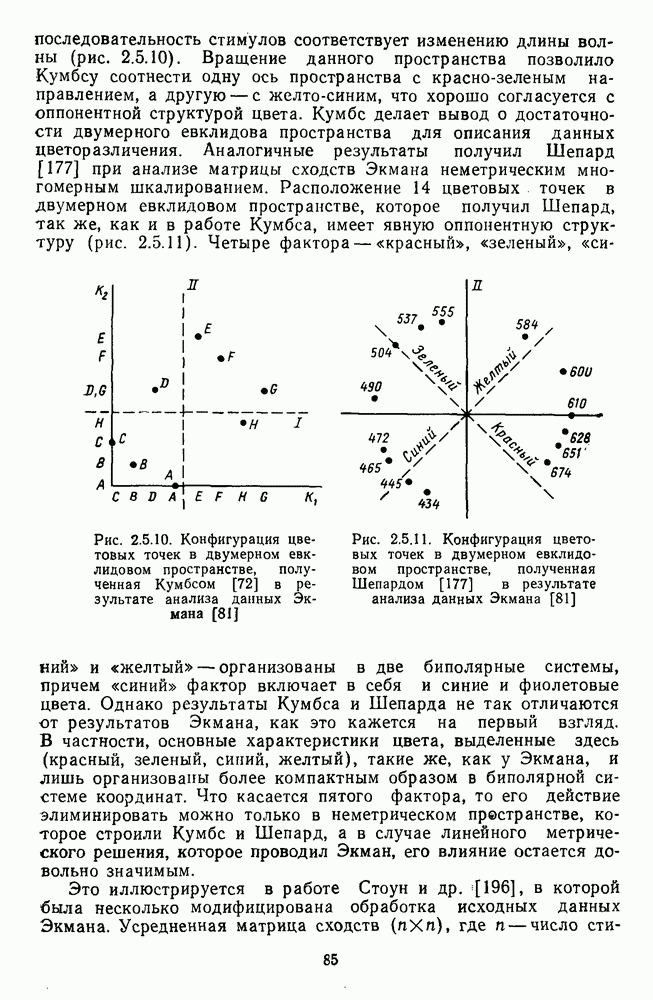
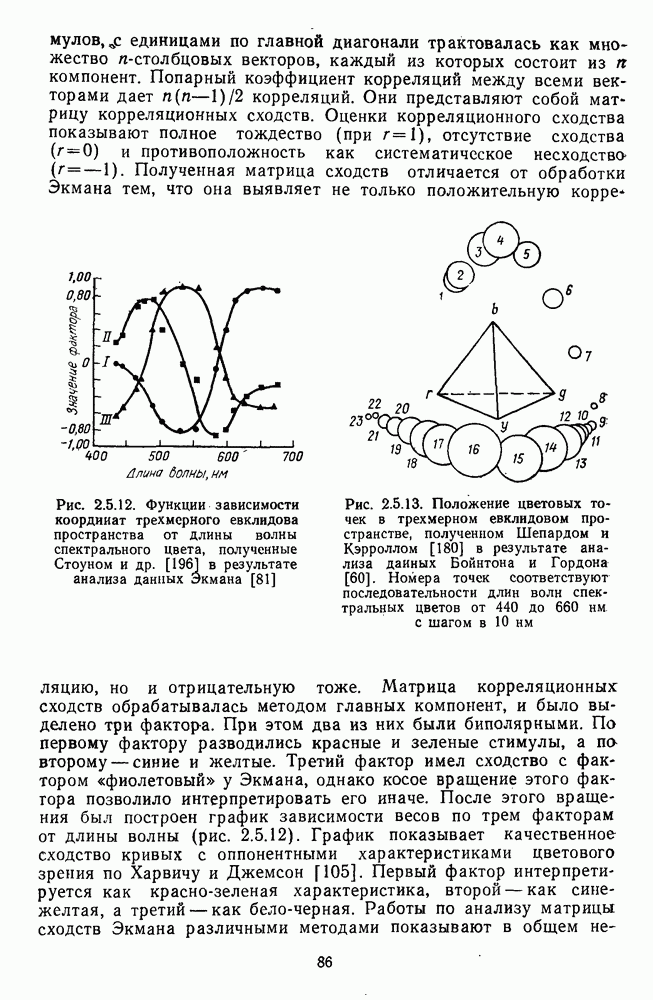
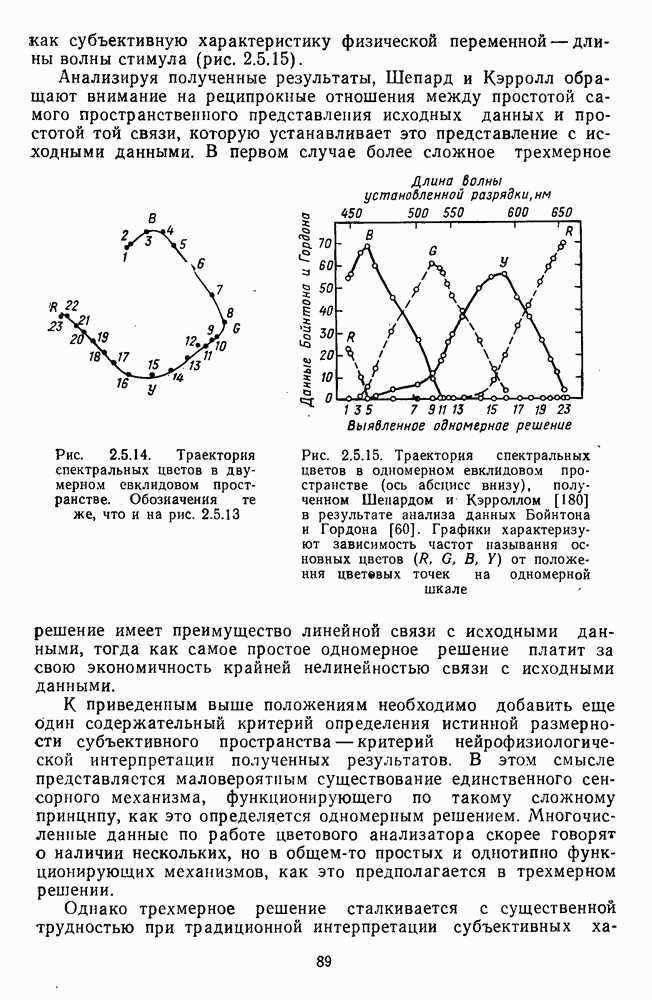
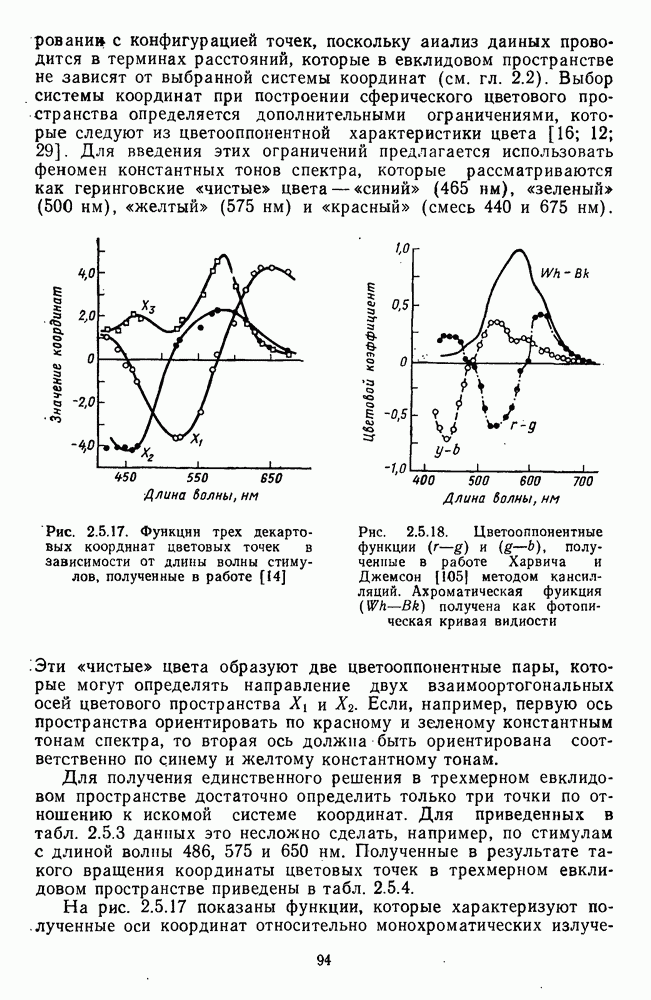
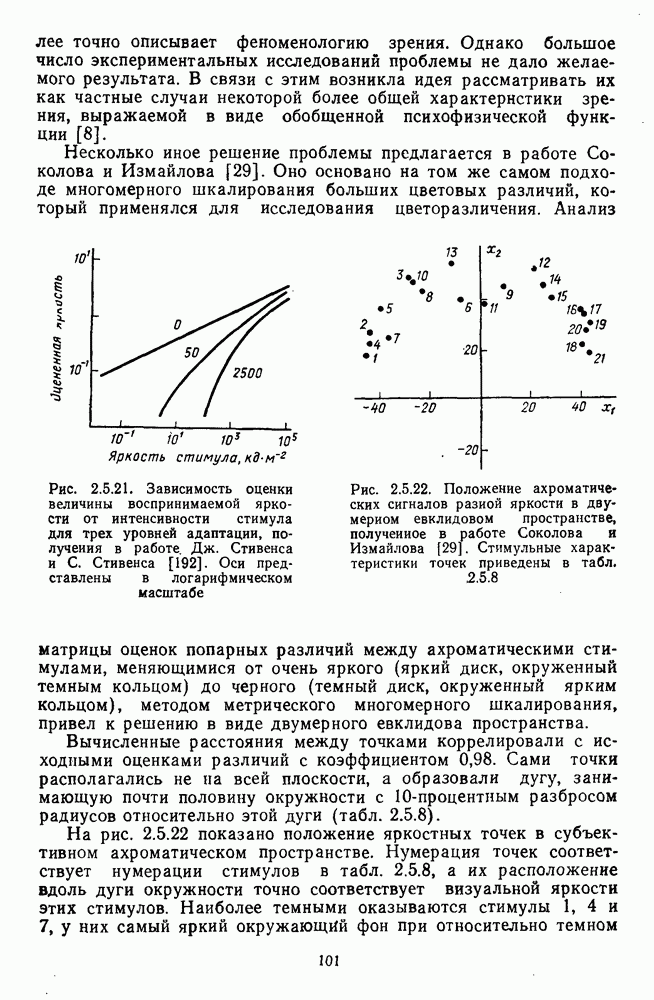
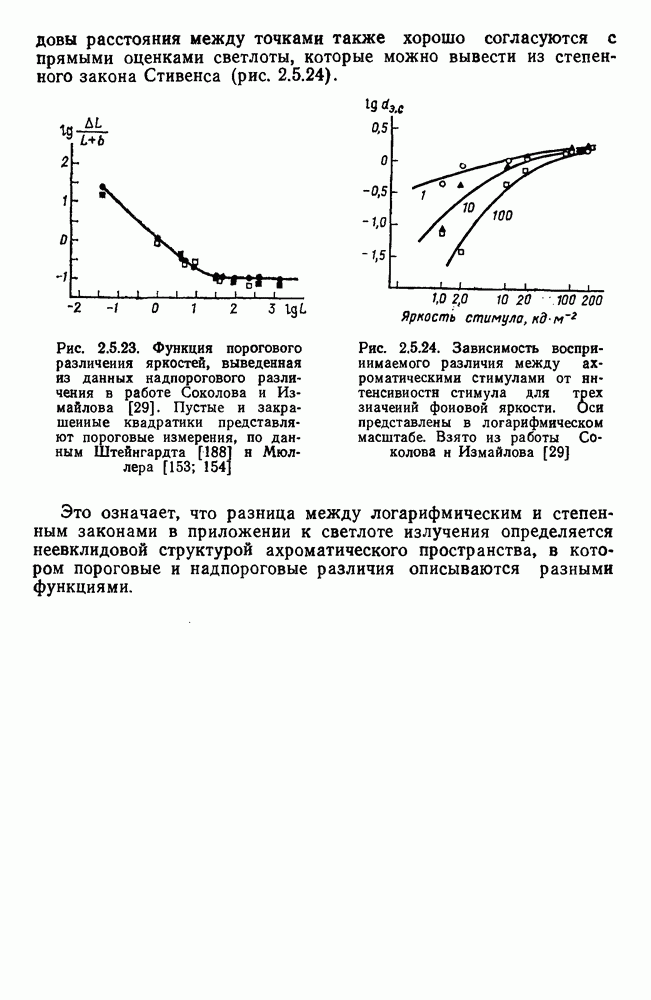
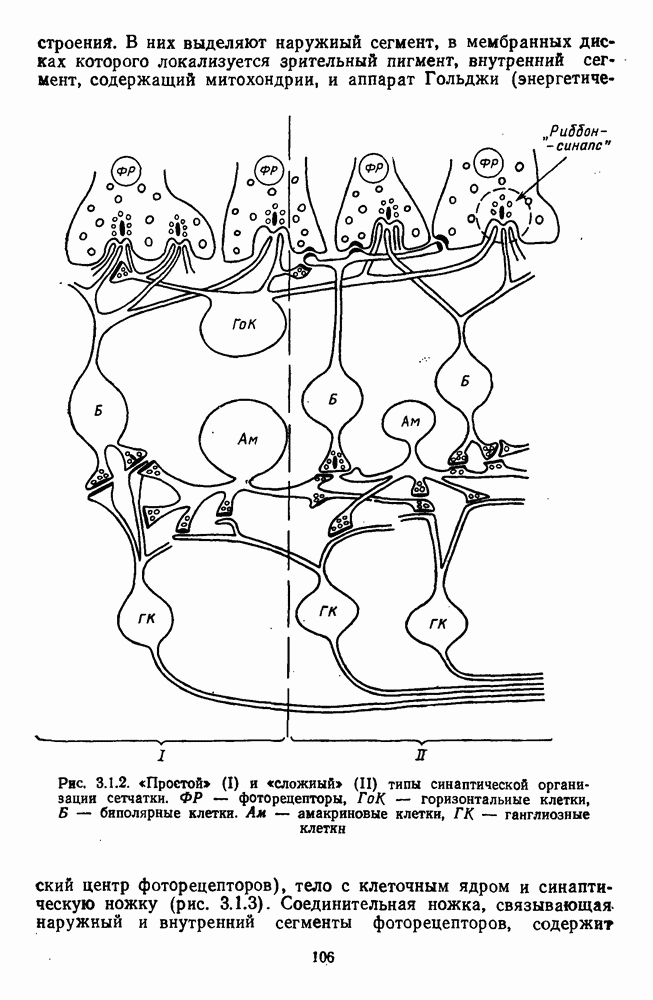
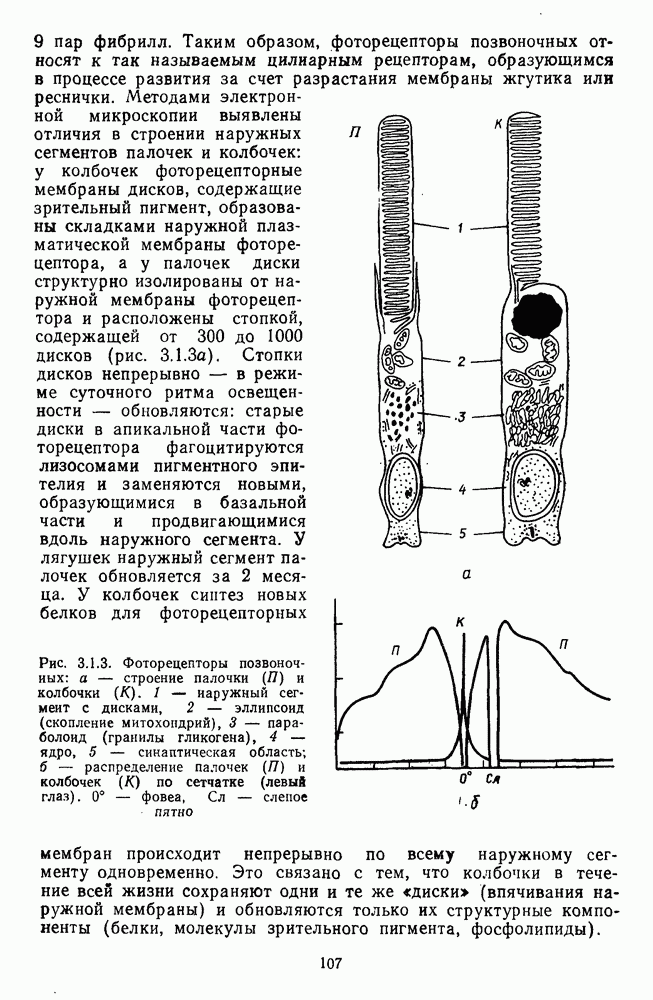
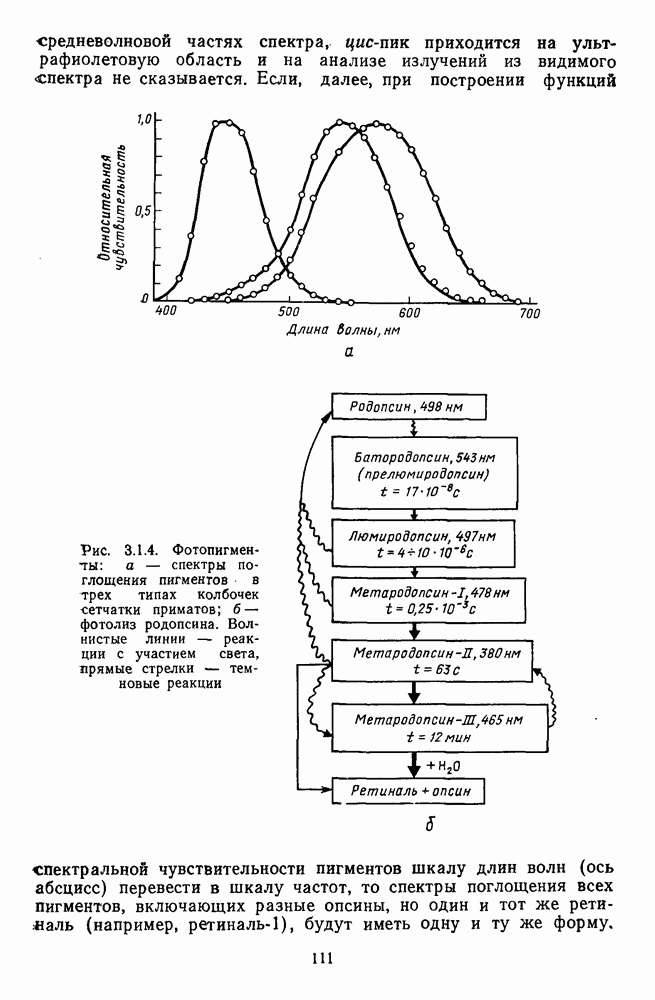
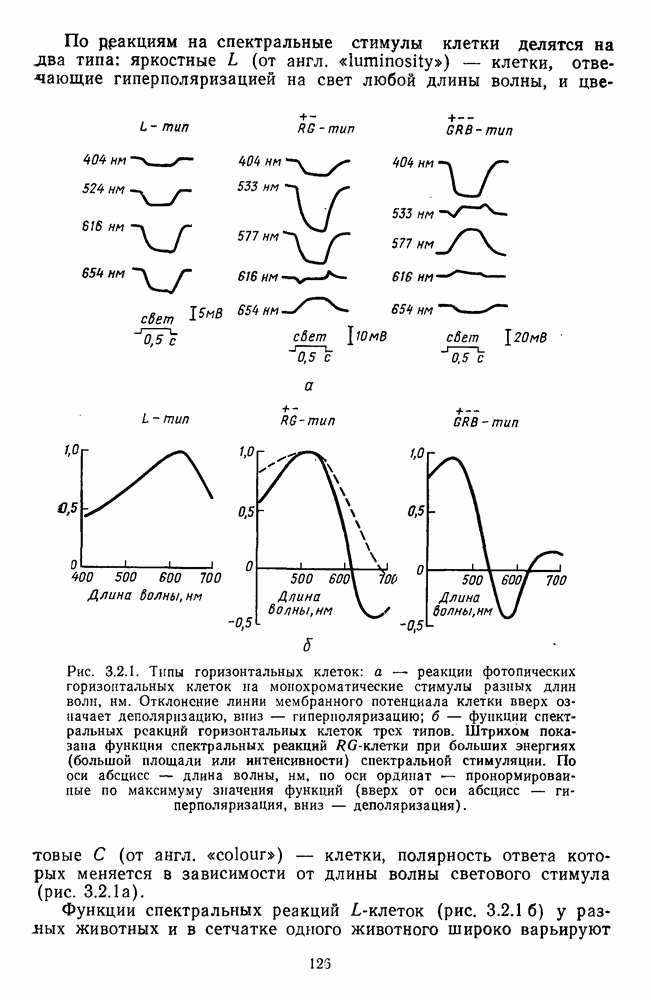
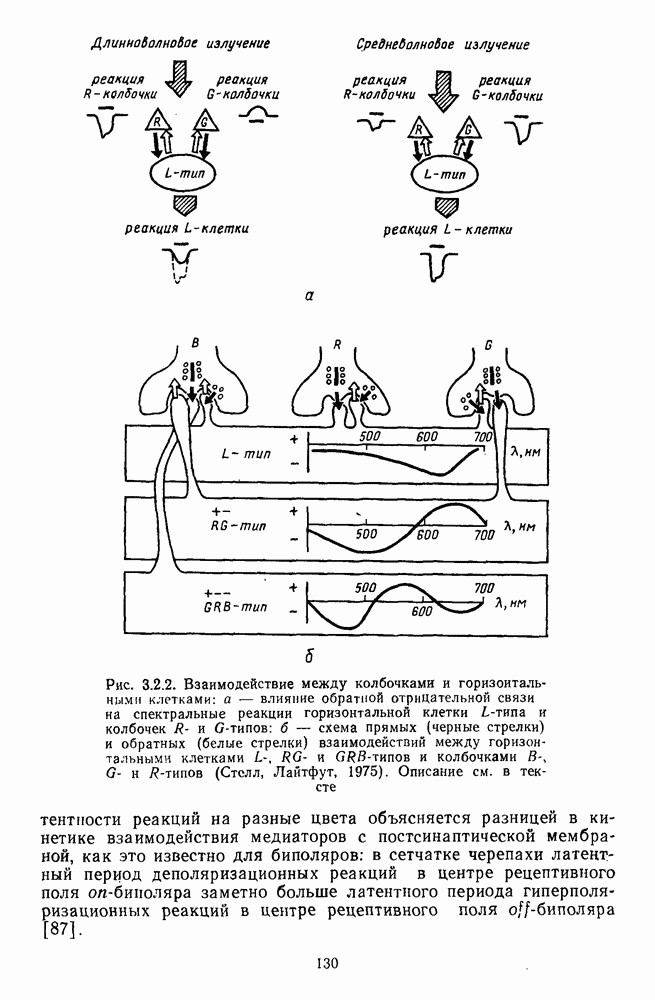
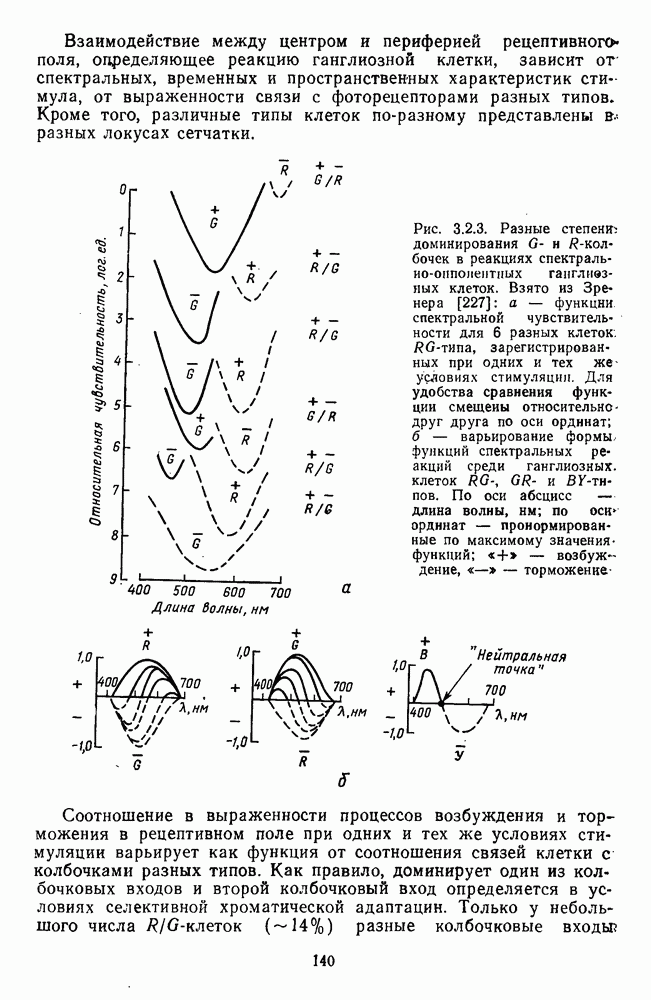
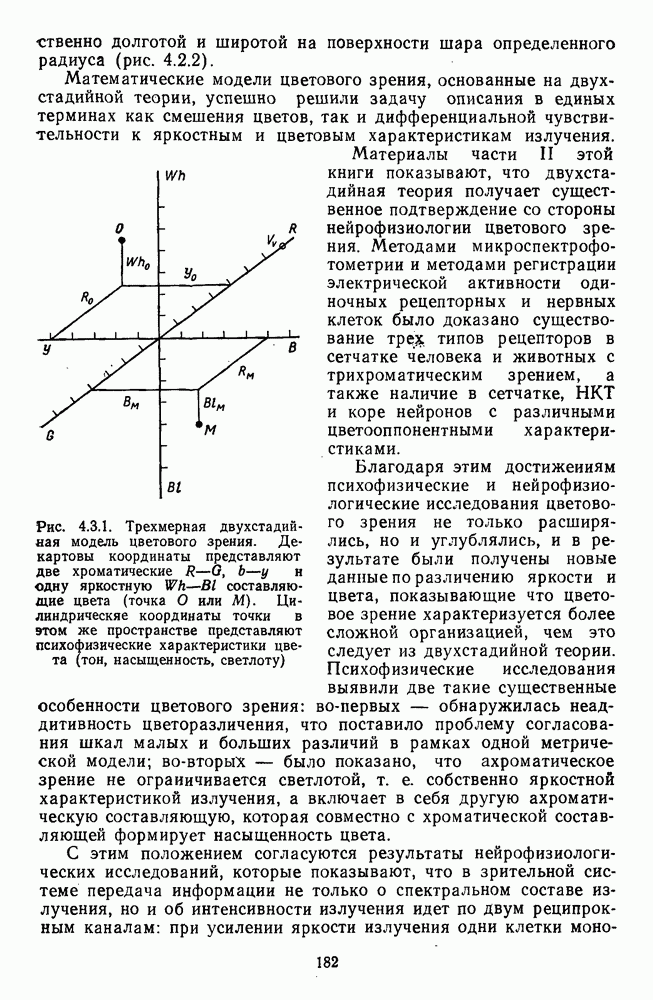
Рис. 3.3.1. Функции спектральных реакций оппонентных  и ахроматических
и ахроматических  нейронов НКТ обезьяны [209]. Три кривые на каждом графике представляют реакции клеток на спектральные стимулы разных интенсивностей (плотности нейтральных светофильтров указаны рядом, лог. ед.)
нейронов НКТ обезьяны [209]. Три кривые на каждом графике представляют реакции клеток на спектральные стимулы разных интенсивностей (плотности нейтральных светофильтров указаны рядом, лог. ед.)
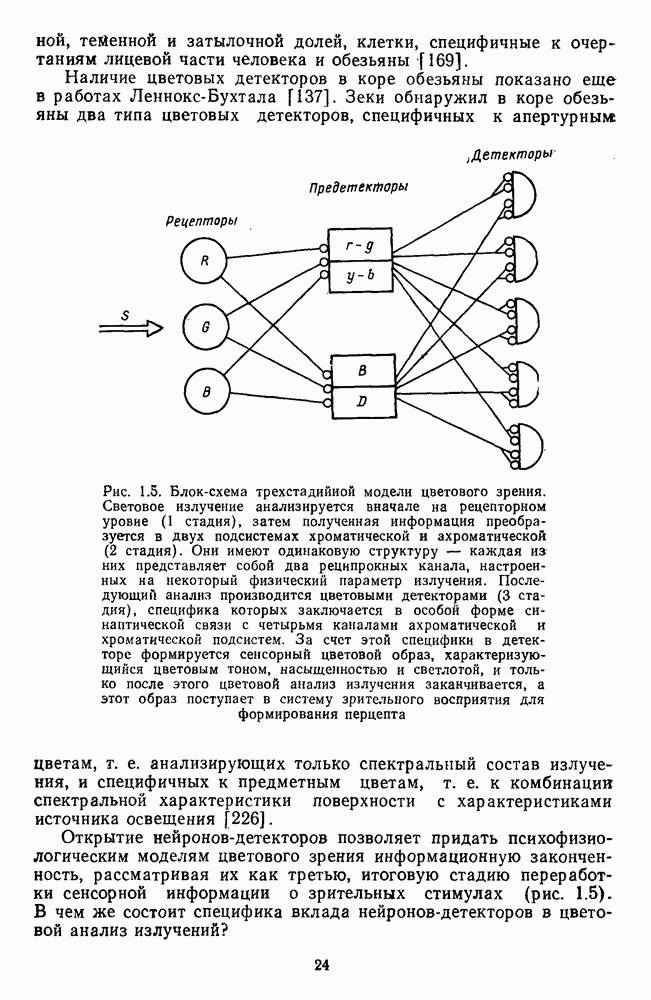
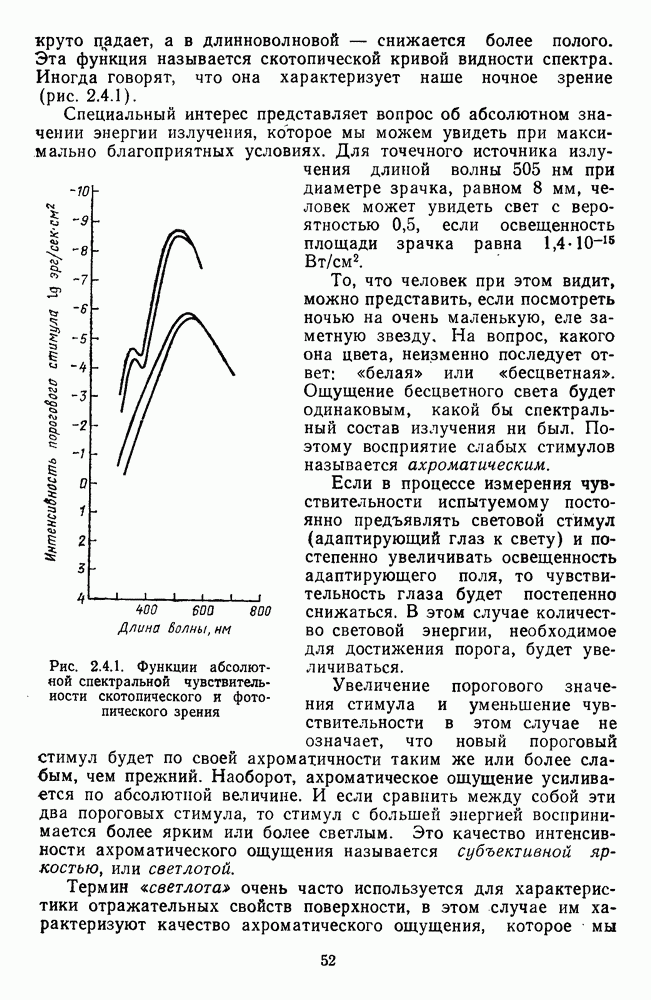
Рецептивные поля оппонентных и ахроматических нейронов НКТ формируются с участием тех же типов фоторецепторов, что и соответствующие типы ганглиозных
клеток. Одкако, в отличие от последних, многие и ахроматические и оппонентные клетки НКТ получают сигналы от палочек. У  -клеток это выражается в «разрушении» оппонентных свойств при темновой адаптации, что сопровождается увеличением абсолютной световой чувствительности в тысячи раз и сдвигом Пуркинье [40].
-клеток это выражается в «разрушении» оппонентных свойств при темновой адаптации, что сопровождается увеличением абсолютной световой чувствительности в тысячи раз и сдвигом Пуркинье [40].
В НКТ обезьяны имеются нейроны, у которых центр и периферия рецептивного поля связаны с одним и тем же рецептором: либо  либо
либо  -типа, Такое поле функционирует как обычное концентрическое: хорошо активируется локальным воздействием на центр и слабо — диффузным освещением или затемнением всей площади. Но это свойство сочетается с максимальной чувствительностью к свету с определенной длиной волны.
-типа, Такое поле функционирует как обычное концентрическое: хорошо активируется локальным воздействием на центр и слабо — диффузным освещением или затемнением всей площади. Но это свойство сочетается с максимальной чувствительностью к свету с определенной длиной волны.
Все нейроны с оппонентными рецептивными полями в НКТ принадлежат к нейронам тонического типа. Фазические клетки, как правило, не обнаруживают оппонентности.
RG-, YB- и  -типы нейронов описаны в НКТ и других животных — белки, суслика, кролика, кошки. В НКТ суслика обнаружены также клетки с двойной спектральной оппонентностью, которые у обезьяны имеются только в коре.
-типы нейронов описаны в НКТ и других животных — белки, суслика, кролика, кошки. В НКТ суслика обнаружены также клетки с двойной спектральной оппонентностью, которые у обезьяны имеются только в коре.