Пред.
След.
Макеты страниц
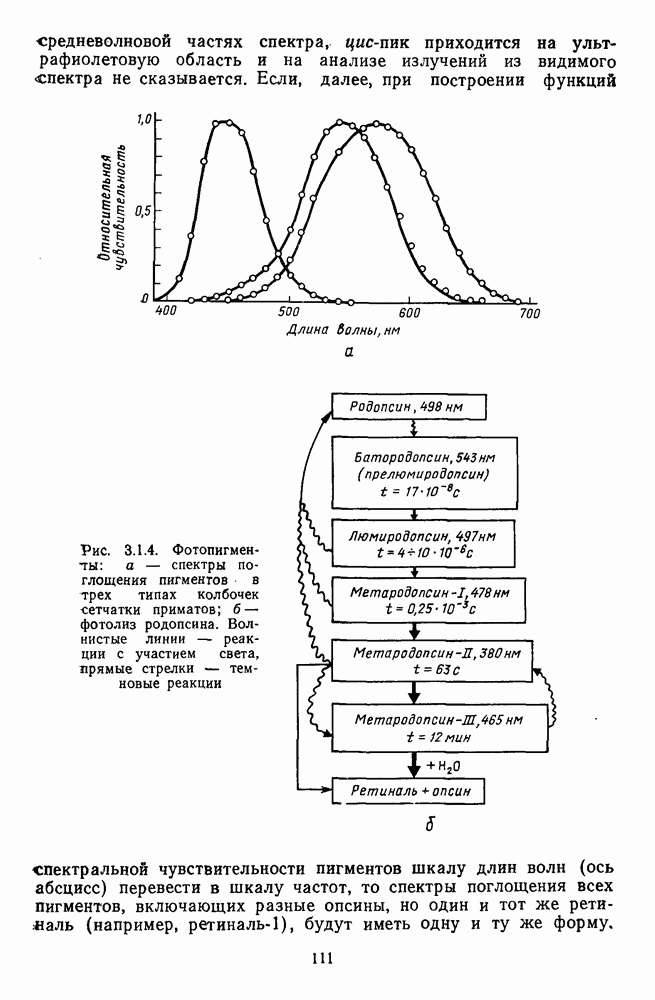
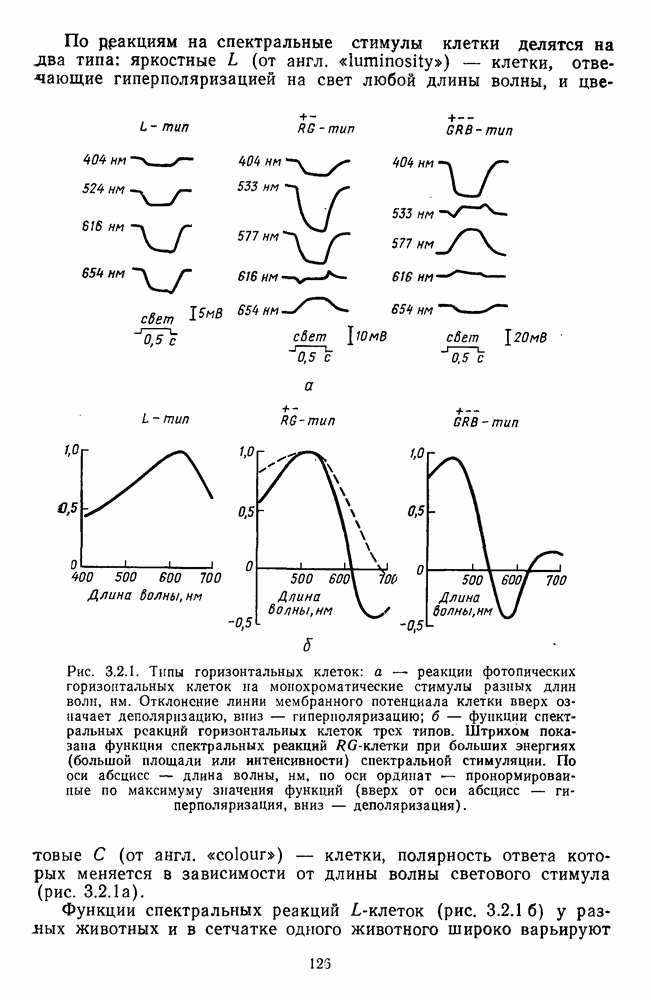
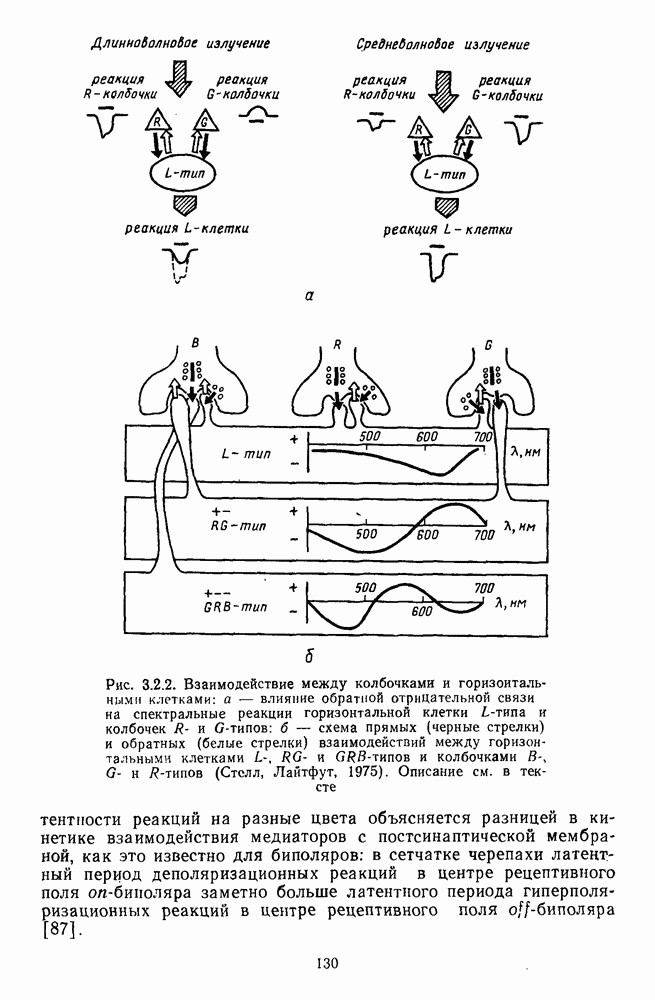
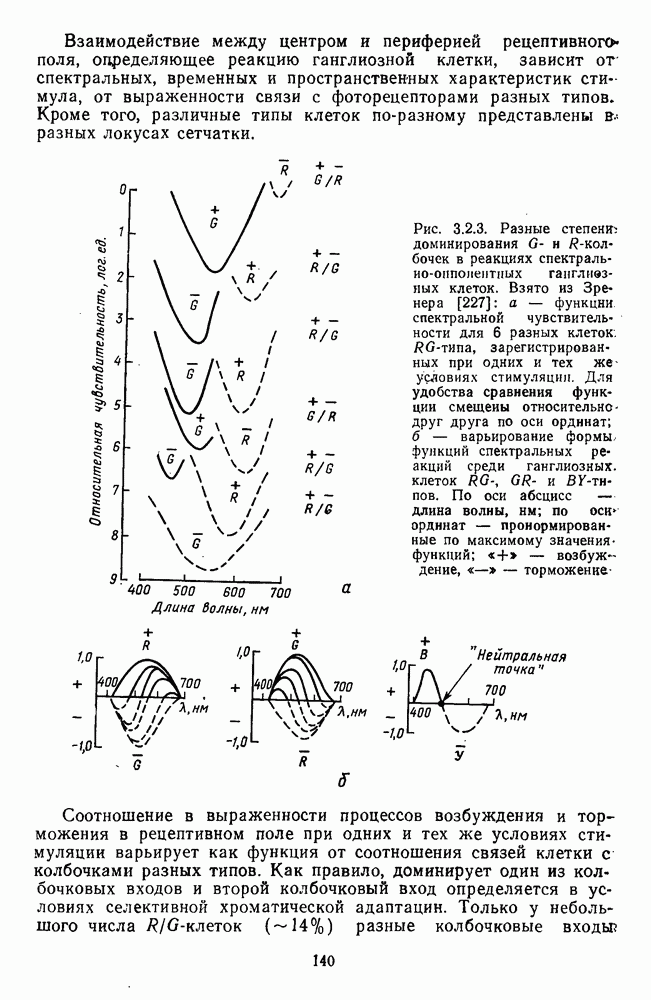
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
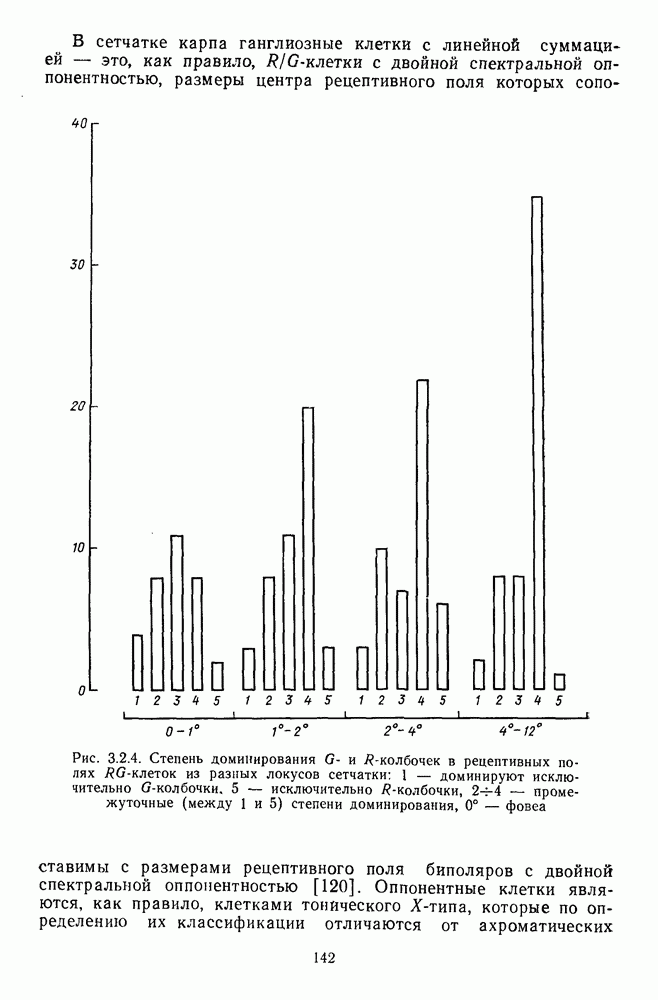
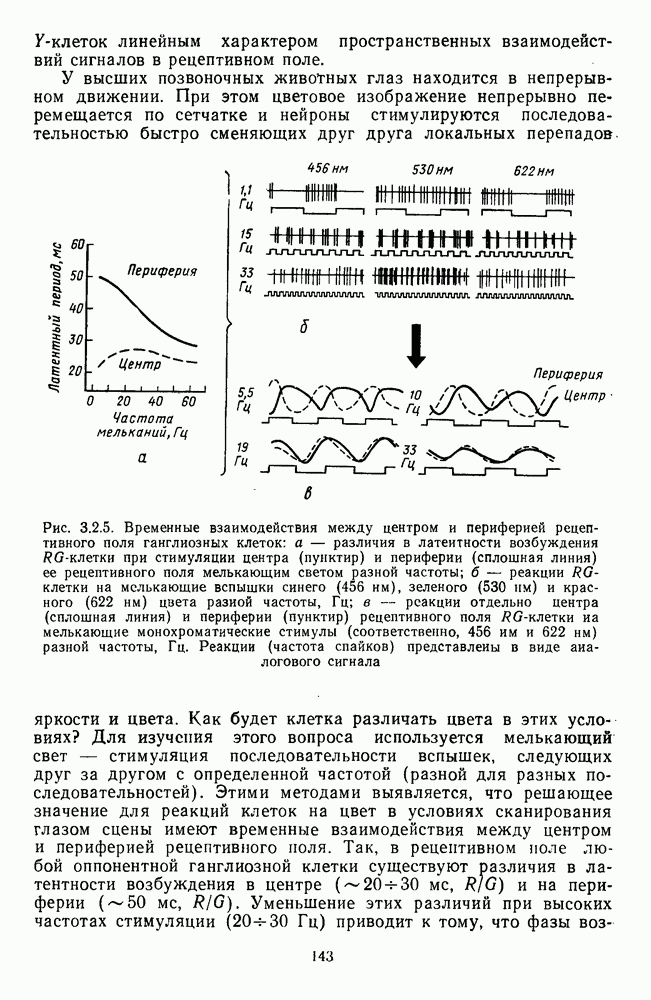
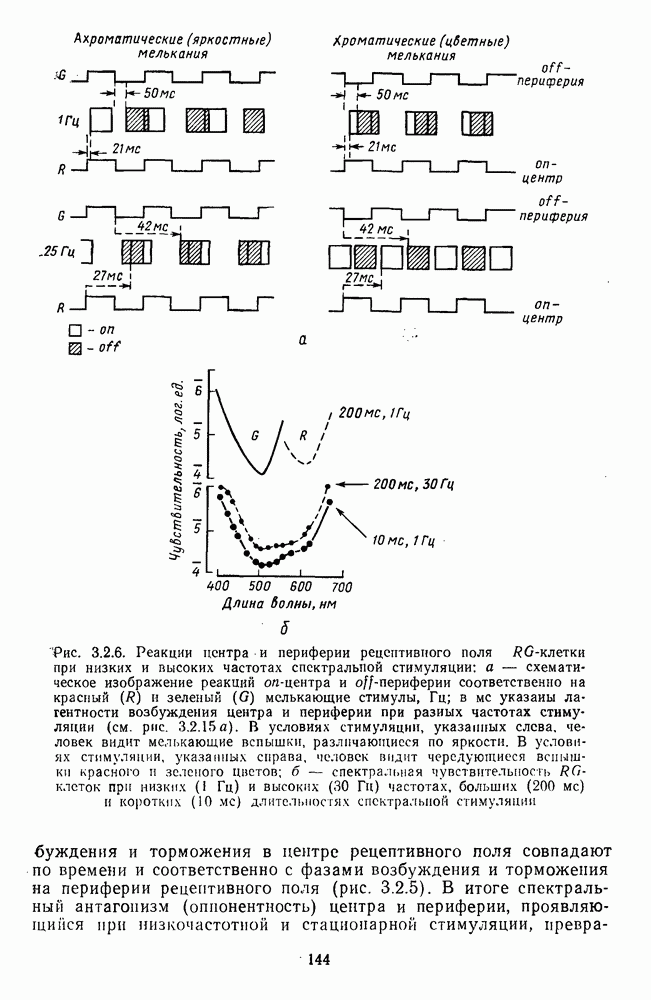
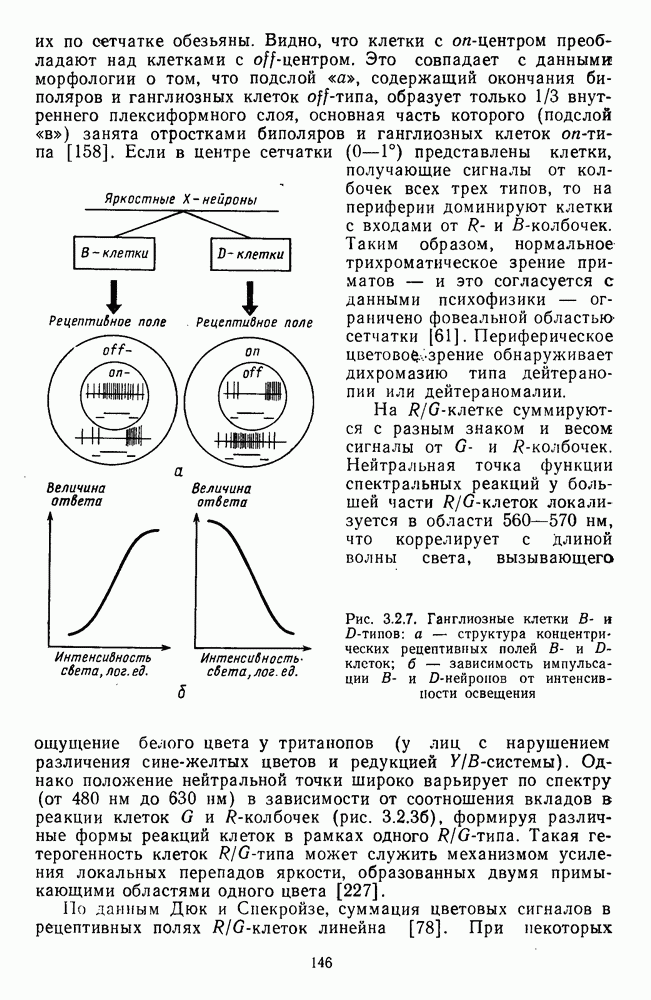
3.2.2. Биполярные клеткиБиполяры При всем многообразии морфологических типов физиологически выделяют два основных типа этих клеток: деполяризационные (off-клетки) и гиперполяризационные ( В наружном плексиформном слое биполяры связаны с фоторецептором синаптическими контактами двух типов: базальными (или поверхностными) и ленточными (или инвагинирующими) (рис. 3.1.2). Возможно, что базальные контакты принадлежат Биполярные клетки представляют в сетчатке прямой, или вертикальный путь прохождения сигнала от рецепторов к ганглиозным клеткам. Спектральные характеристики биполярных клеток. Характеристики рецептивных полей биполяров по отношению к стимулам разного спектрального состава приведены в табл. 3.2.1. Здесь показаны ахроматические клетки, которые получают сигналы разного знака от колбочек одного и того же типа, затем цветовые клетки, у которых пространственная организация рецептивного поля связана с разными рецепторами, и, наконец, клетки, рецептивные поля которых характеризуются двойной спектральной оппонентностью: они реагируют деполяризацией на стимуляцию Таблица 3.2.1 (см. скан) Тилы колбочковых биполяров центра РП длинноволновым излучением и гиперполяризацией на стимуляцию центра РП средневолновым излучением. Такая же стимуляция периферии дает противоположные по знаку реакции. В сетчатке карпа число таких клеток составляет ~25% от общего числа зарегистрированных биполяров [120]. Необычно организованы РП цветовых биполяров в сетчатке шпорцевой лягушки [223]. Во-первых, они однородны, без деления на центр и периферию. Во-вторых, они обнаруживают необычный тип оппонентности — между длинноволновыми и коротковолновым излучениями У черепахи цветовые биполяры отвечают деполяризацией на красное пятно и гиперполяризацией на зеленое, а при освещении кольцом гиперполяризуются (или деполяризуются) как на красный, так и на зеленый свет [224]. Таким образом, организация рецепторных входов биполярных клеток обеспечивает одним клеткам синергизм реакций на разные излучения, а другим — антагонизм. У низших позвоночных биполяры непосредственно участвуют в формировании цветоселективных свойств ганглиозных клеток. Так, у шпорцевой лягушки гиперполяризация биполяров на коротковолновое излучение сопровождается Механизмы генерации реакций биполяров. Реакция биполяра на свет в центре рецептивного поля является результатом прямой передачи сигнала от рецепторов, синаптически связанных с его дендритными отростками. При этом биполяры ведут себя сходно с горизонтальными клетками, т. е. в темноте медиатор фоторецептора деполяризует их, увеличивая проводимость по натрию, а гиперполяризация на свету возникает в результате уменьшения поступления этого медиатора. Не до конца ясны ионные механизмы генерации реакций Реакция биполяра на освещение периферии рецептивного поля определяется горизонтальными клетками. Так, искусственная гиперполяризация горизонтальной клетки вызывает в биполярах тот же ответ, что и освещение периферии, а деполяризация — реакцию обратного знака [37]. Функция спектральной чувствительности, измеренная при стимуляции периферии рецептивного поля, аппроксимируется линейной комбинацией функций спектральной чувствительности горизонтальных клеток Биполяры
|
 |
Оглавление
|












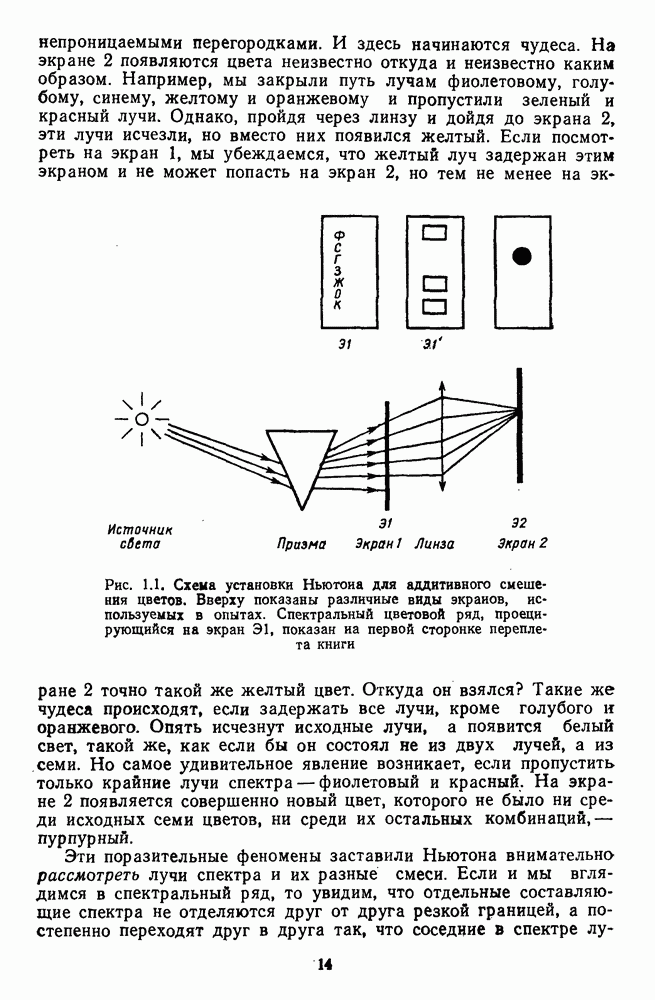






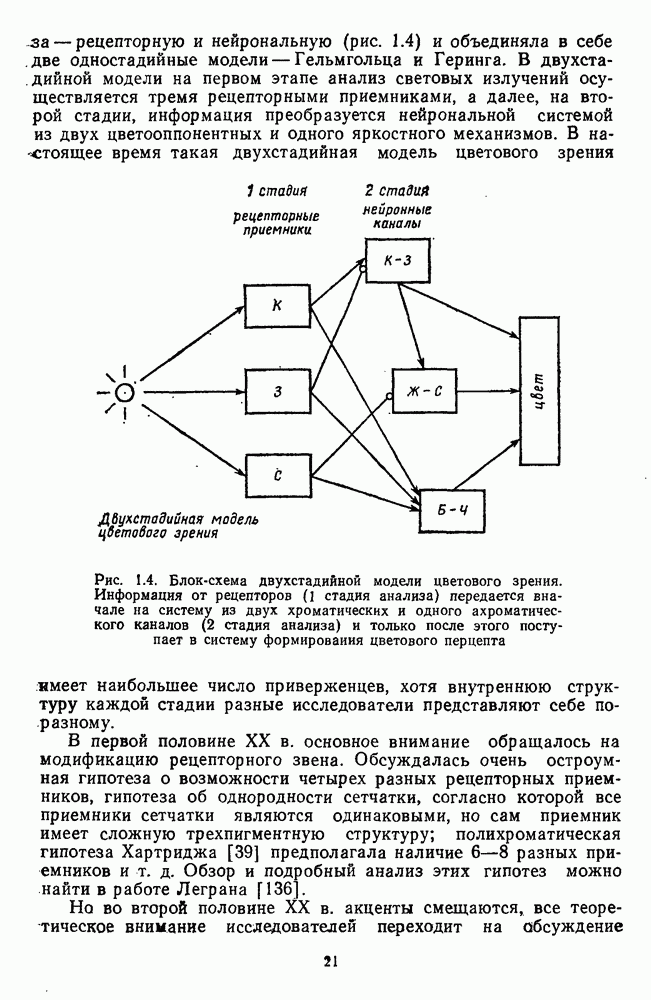


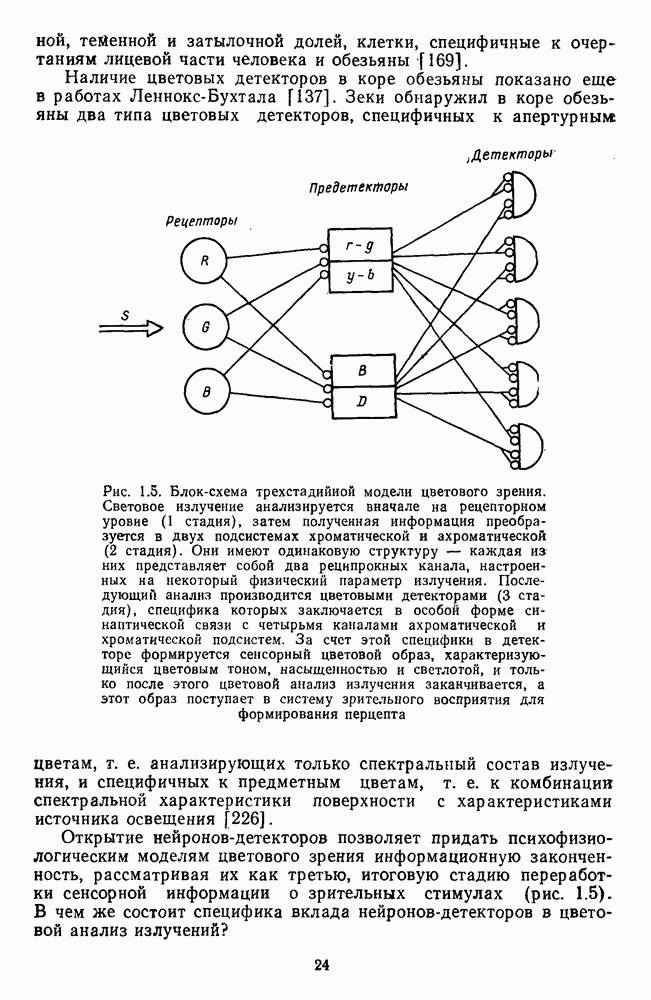



























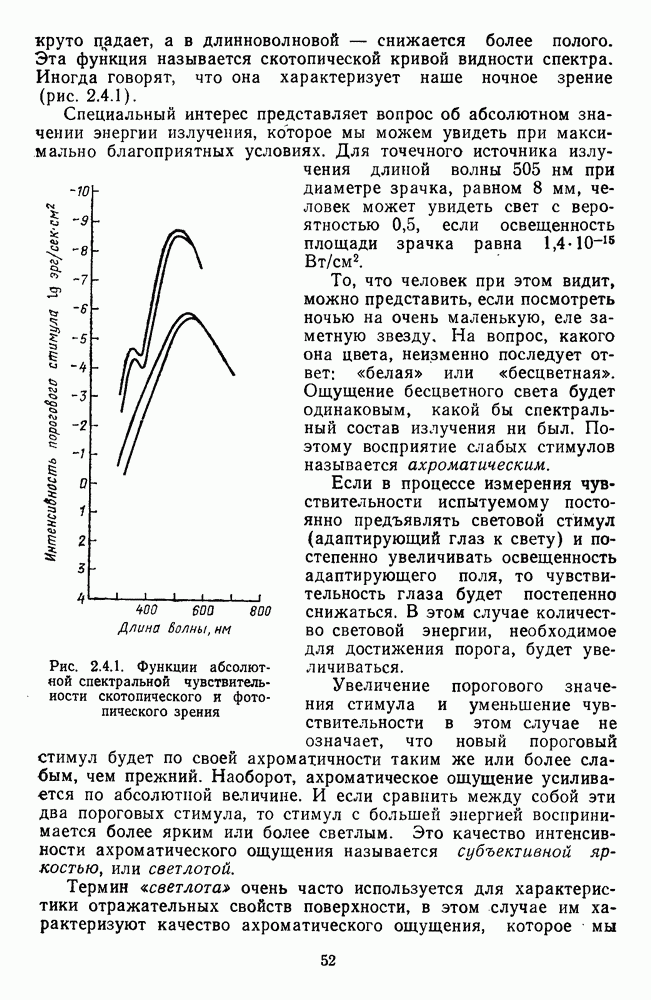










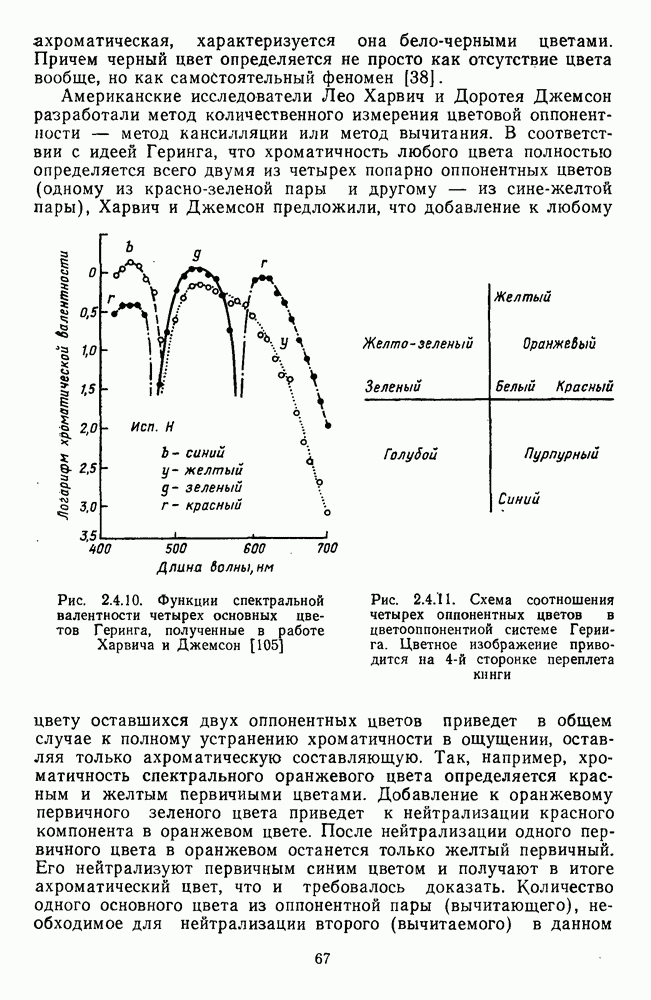


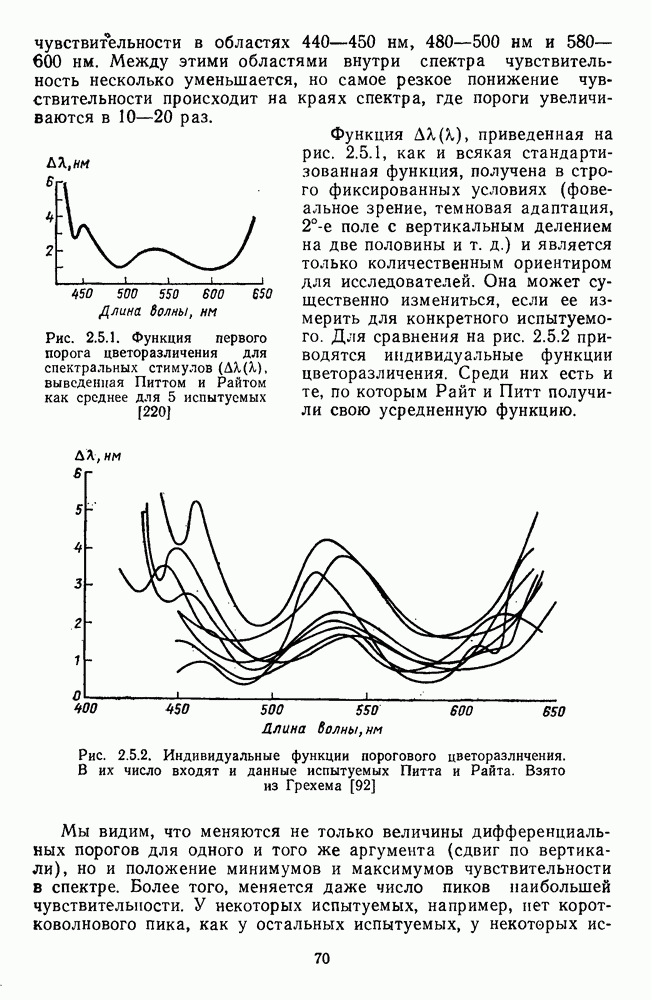
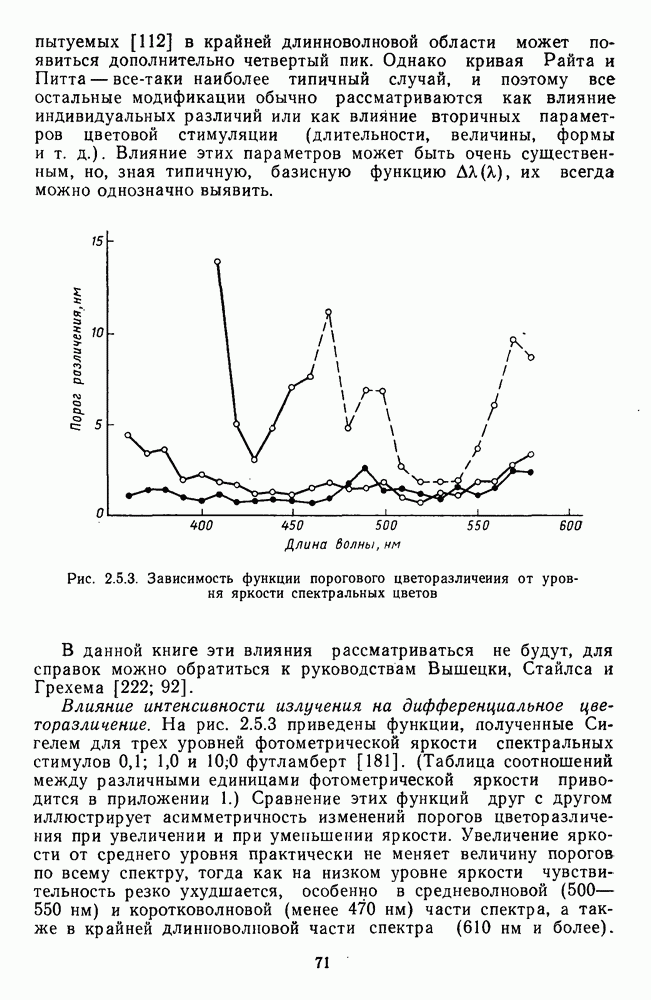
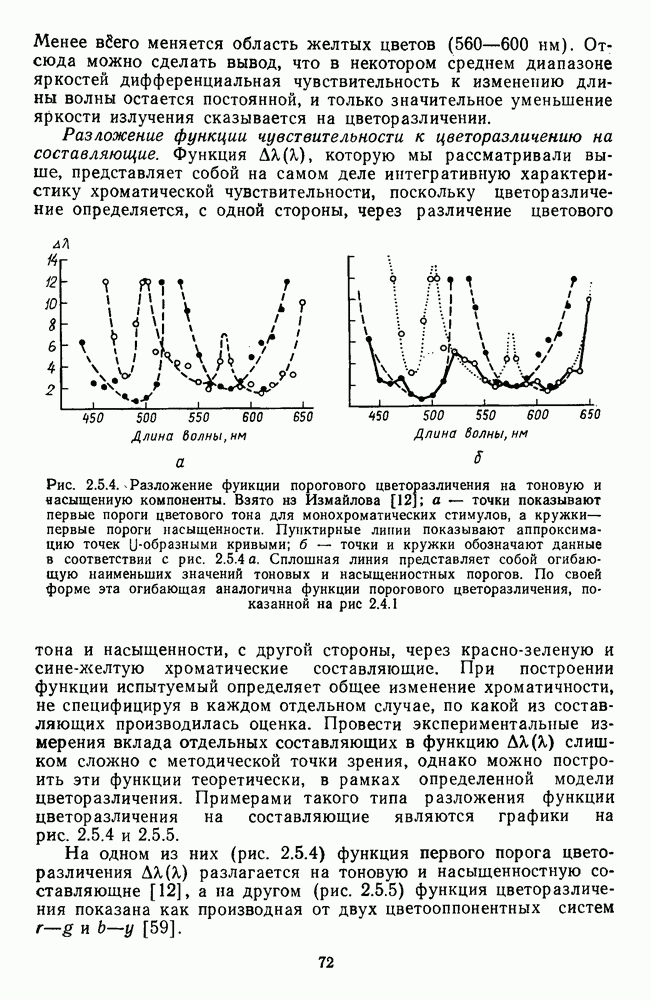







































































































 и
и  Биполярные клетки связывают рецепторные входы сетчатки с ее выходными элементами — ганглиозными клетками. Через биполяры каждая ганглиозная клетка соединена со многими тысячами фоторецепторов и каждый рецептор — с несколькими ганглиозными клетками.
Биполярные клетки связывают рецепторные входы сетчатки с ее выходными элементами — ганглиозными клетками. Через биполяры каждая ганглиозная клетка соединена со многими тысячами фоторецепторов и каждый рецептор — с несколькими ганглиозными клетками.
 -клетки)
-клетки)  биполяры реагируют деполяризацией на освещение центра рецептивного поля пятном света с диаметром 100-5-200 мкм (это примерно соответствует области ветвления дендритов биполярной клетки), а
биполяры реагируют деполяризацией на освещение центра рецептивного поля пятном света с диаметром 100-5-200 мкм (это примерно соответствует области ветвления дендритов биполярной клетки), а  -биполяры на тот же стимул реагируют гиперполяризацией [119]. Освещение периферии рецептивного поля вызывает в клетках обоих типов обратные реакции. По характеру связей с фоторецепторами биполяры делятся на палочковые и колбочковые. Одна часть колбочковых биполяров избирательно связана с
-биполяры на тот же стимул реагируют гиперполяризацией [119]. Освещение периферии рецептивного поля вызывает в клетках обоих типов обратные реакции. По характеру связей с фоторецепторами биполяры делятся на палочковые и колбочковые. Одна часть колбочковых биполяров избирательно связана с  -колбочками, другая часть — с В-колбочками. «Смешанные» биполяры контактируют с двумя: либо с
-колбочками, другая часть — с В-колбочками. «Смешанные» биполяры контактируют с двумя: либо с  и
и  -колбочками, либо с
-колбочками, либо с  и В-колбочками.
и В-колбочками.
 -биполярам, а инвагинирующие — on-биполярам [166]. Во внутреннем плексиформном слое биполяры контактируют с отростками амакриновых и ганглиозных клеток через ленточные синапсы с двумя постсинаптическими элементами (диады). В большинстве диад у лягушек оба постсинаптических нейрона — отростки амакринов. У приматов доминируют диады, в которых один постсинаптический элемент — отросток амакриновой, а другой — отросток ганглиозной клетки. Аксоны
-биполярам, а инвагинирующие — on-биполярам [166]. Во внутреннем плексиформном слое биполяры контактируют с отростками амакриновых и ганглиозных клеток через ленточные синапсы с двумя постсинаптическими элементами (диады). В большинстве диад у лягушек оба постсинаптических нейрона — отростки амакринов. У приматов доминируют диады, в которых один постсинаптический элемент — отросток амакриновой, а другой — отросток ганглиозной клетки. Аксоны  и
и  -биполяров оканчиваются в разных подслоях внутреннего синаптического слоя — проксимальной и дистальной частях соответственно. В разных отделах внутреннего нлексиформного слоя оканчиваются и отростки палочковых и колбочковых биполяров.
-биполяров оканчиваются в разных подслоях внутреннего синаптического слоя — проксимальной и дистальной частях соответственно. В разных отделах внутреннего нлексиформного слоя оканчиваются и отростки палочковых и колбочковых биполяров.
 гиперполяризуются на коротковолновый свет, возбуждающий «зеленые» палочки
гиперполяризуются на коротковолновый свет, возбуждающий «зеленые» палочки  , и деполяризуются на длинноволновый свет, возбуждающий колбочки
, и деполяризуются на длинноволновый свет, возбуждающий колбочки  . При мезопическом уровне освещения они начинают деполяризоваться в ответ на средневолновое излучение, возбуждающее «розовые» палочки
. При мезопическом уровне освещения они начинают деполяризоваться в ответ на средневолновое излучение, возбуждающее «розовые» палочки  .
.
 -реакцией спектрально-оппонентных ганглиозных кйеток, а большое число
-реакцией спектрально-оппонентных ганглиозных кйеток, а большое число  биполяров с двойной спектральной оппонентностью в сетчатке карпа хорошо коррелирует с большим числом ганглиозных клеток с аналогичными свойствами
биполяров с двойной спектральной оппонентностью в сетчатке карпа хорошо коррелирует с большим числом ганглиозных клеток с аналогичными свойствами 
 -биполяров. По данным Тойода и др., деполяризация
-биполяров. По данным Тойода и др., деполяризация  -биполяра на освещение центра его рецептивного поля является результатом действия двух различных механизмов: увеличения проницаемости для ионов натрия (в синапсах с палочками) и уменьшения проницаемости для ионов калия (в синапсах с колбочками) [204]. Об отличии синаптических входов
-биполяра на освещение центра его рецептивного поля является результатом действия двух различных механизмов: увеличения проницаемости для ионов натрия (в синапсах с палочками) и уменьшения проницаемости для ионов калия (в синапсах с колбочками) [204]. Об отличии синаптических входов  -биполяров от входов
-биполяров от входов  -биполяров свидетельствует и более медленная кинетика взаимодействия медиатора с постсинаптическими каналами в
-биполяров свидетельствует и более медленная кинетика взаимодействия медиатора с постсинаптическими каналами в  -биполярах [73].
-биполярах [73].
 и С-типов [224].
и С-типов [224].
 и
и  -типов образуют множество параллельных каналов прямой передачи информации от фоторецепторов на ганглиозные клетки. Сложная организация рецептивных полей — сочетание пространственной оппонентности с цветовой — делает их реакции на спектральный состав излучения качественно отличными от реакций фоторецепторов и горизонтальных клеток.
-типов образуют множество параллельных каналов прямой передачи информации от фоторецепторов на ганглиозные клетки. Сложная организация рецептивных полей — сочетание пространственной оппонентности с цветовой — делает их реакции на спектральный состав излучения качественно отличными от реакций фоторецепторов и горизонтальных клеток.