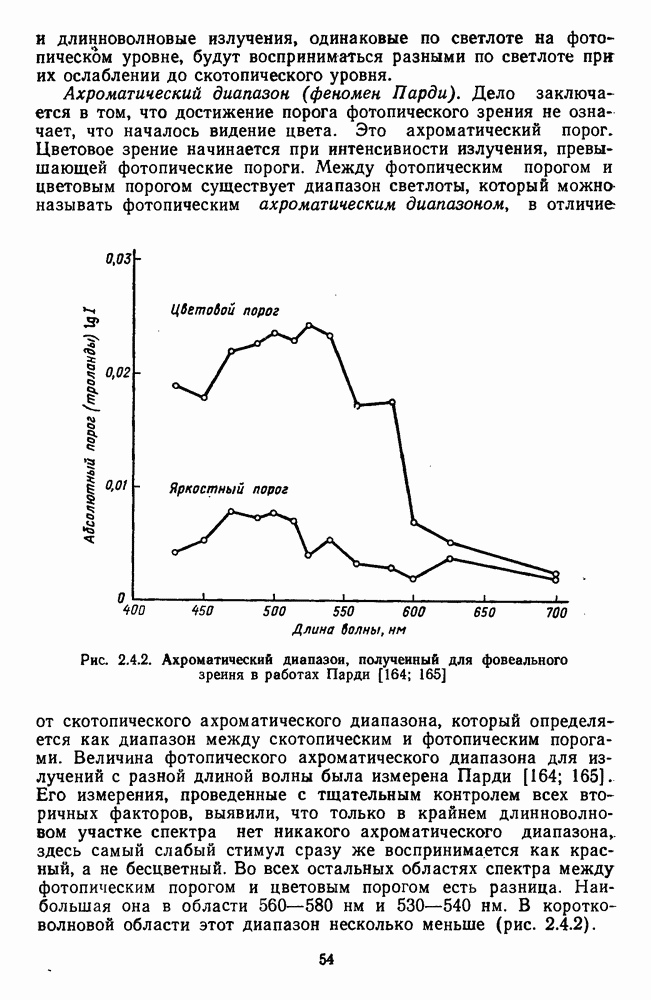
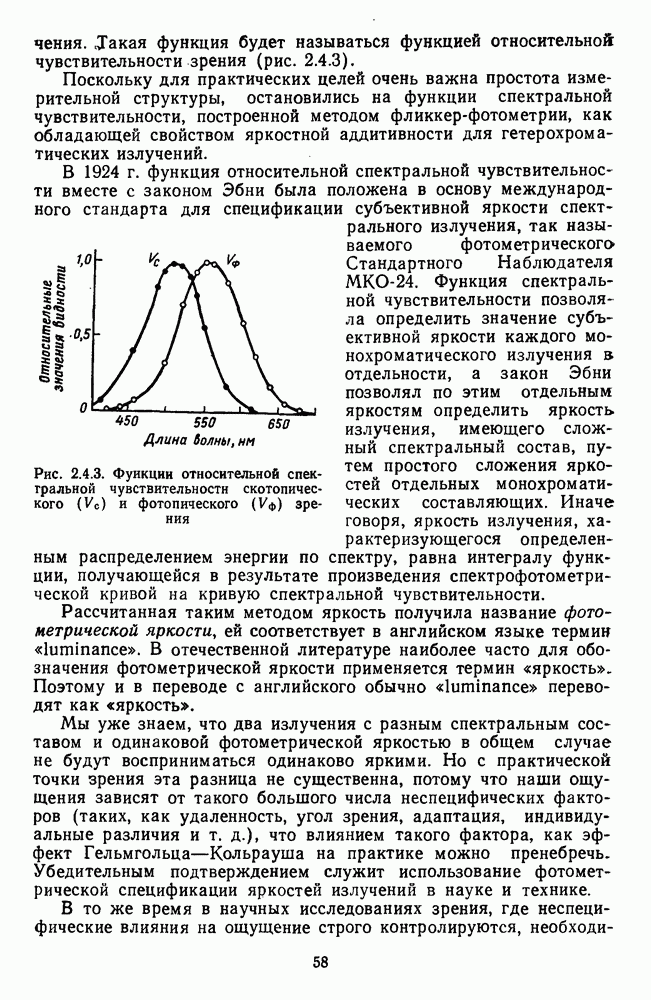
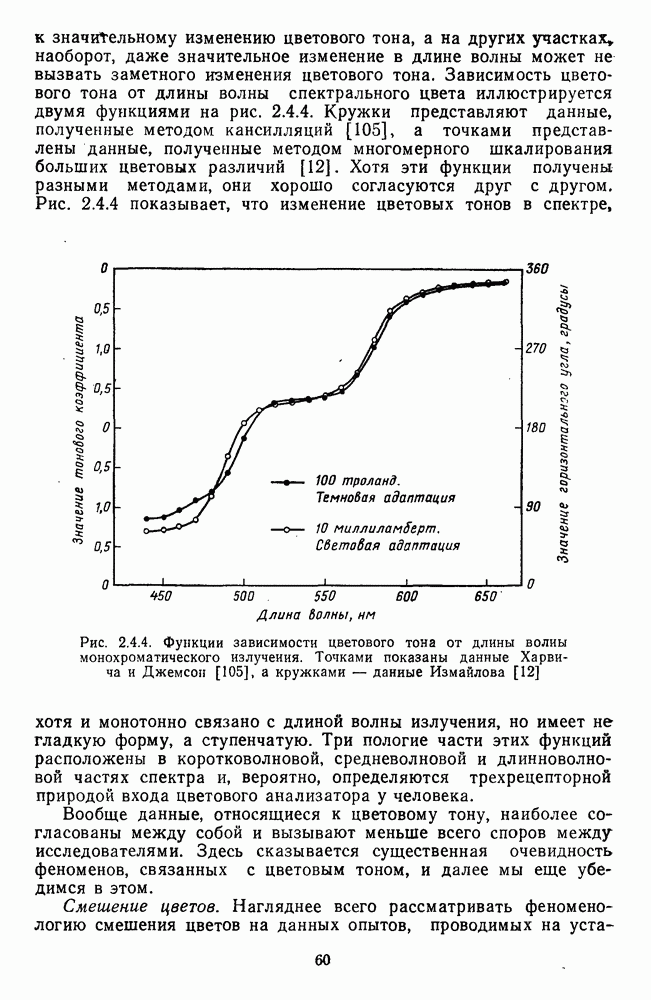
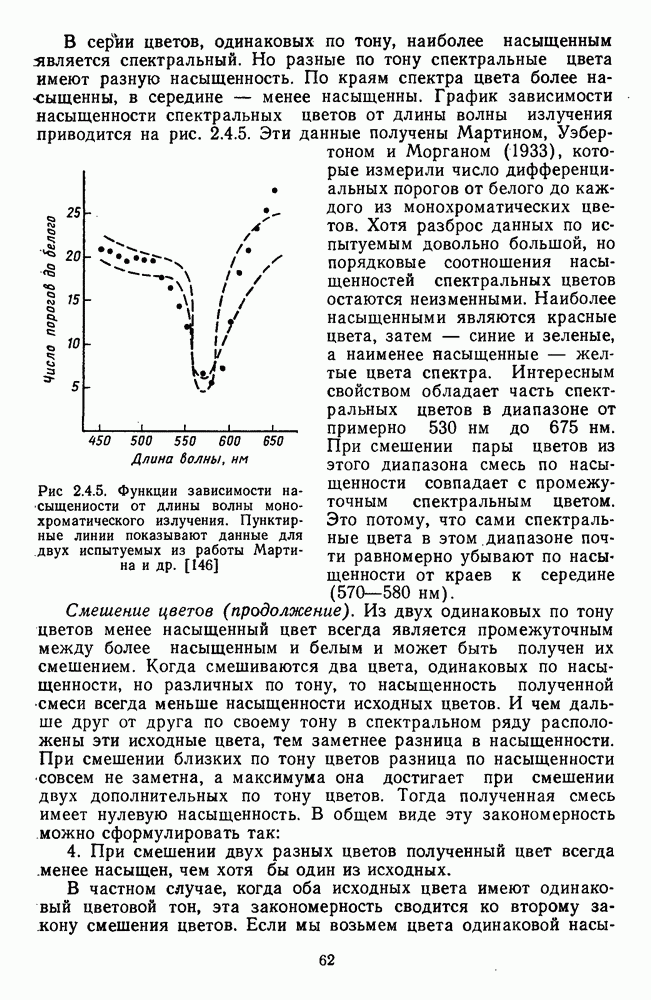
Пред.
След.
Макеты страниц
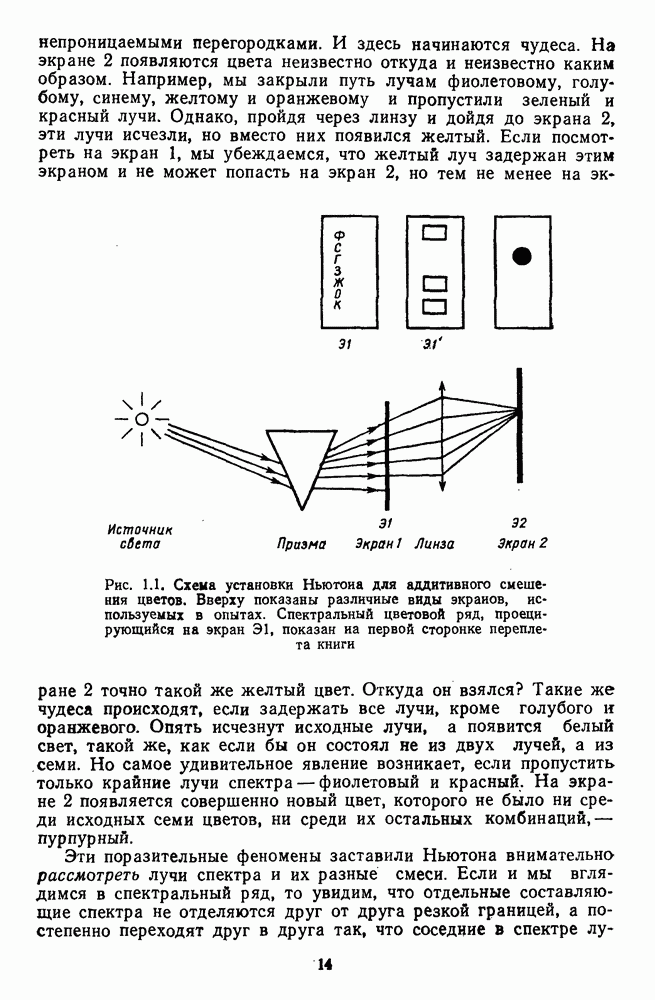
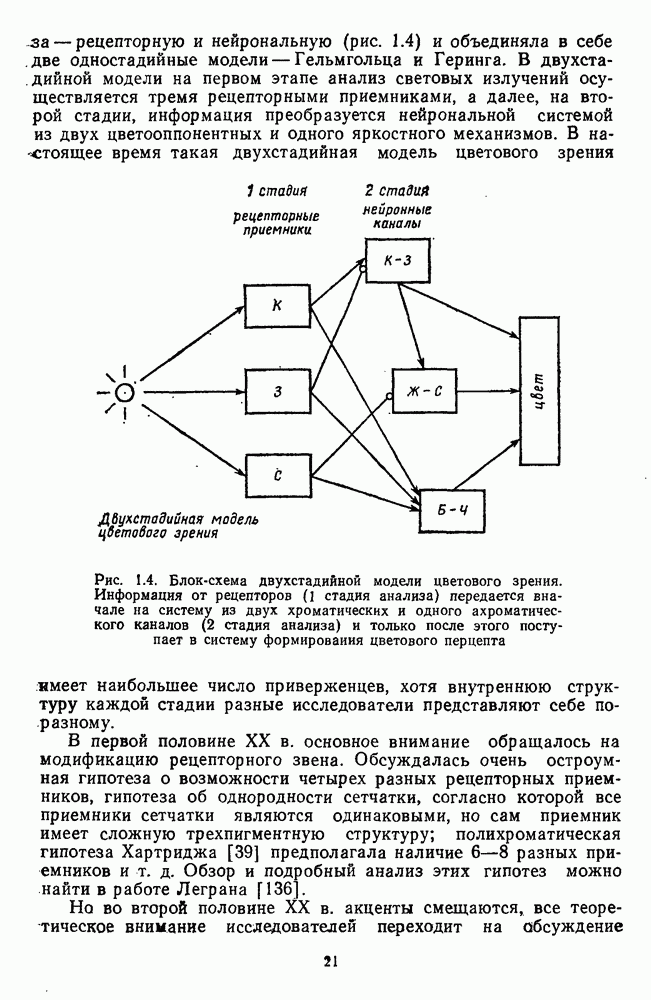
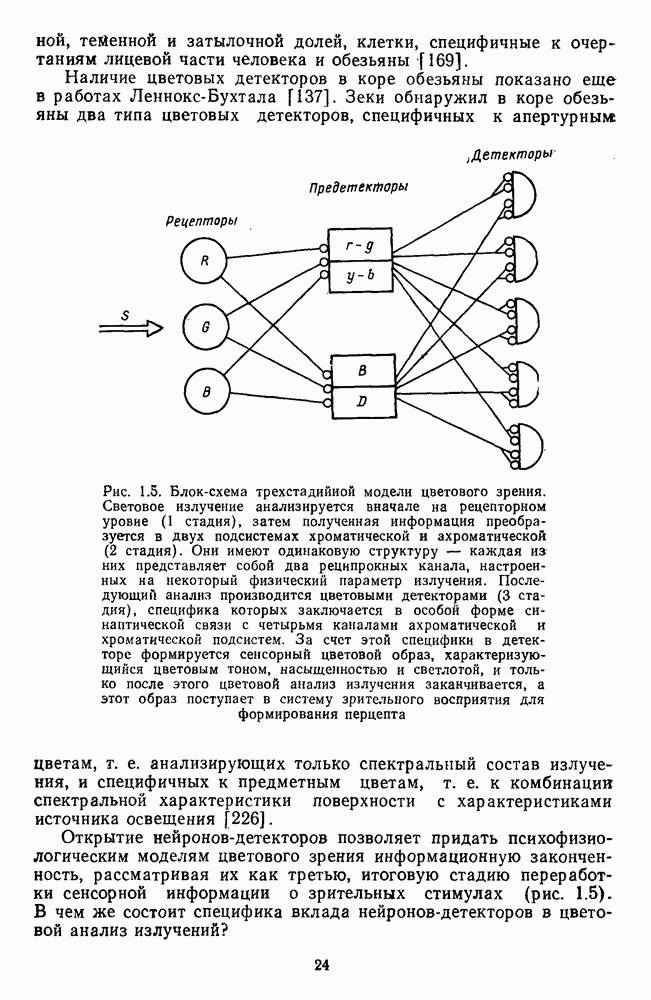
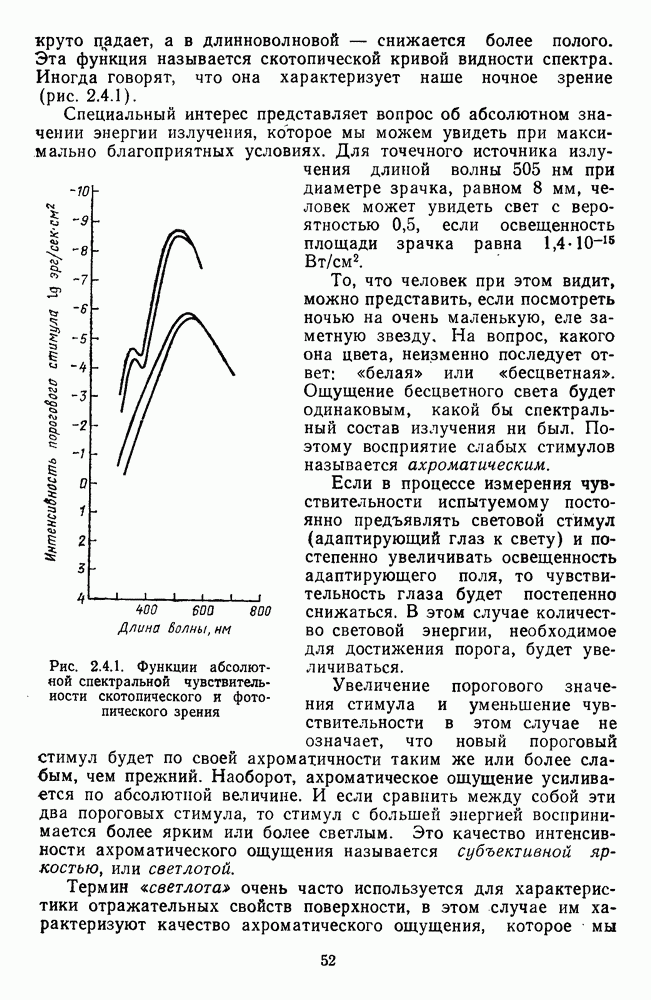
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
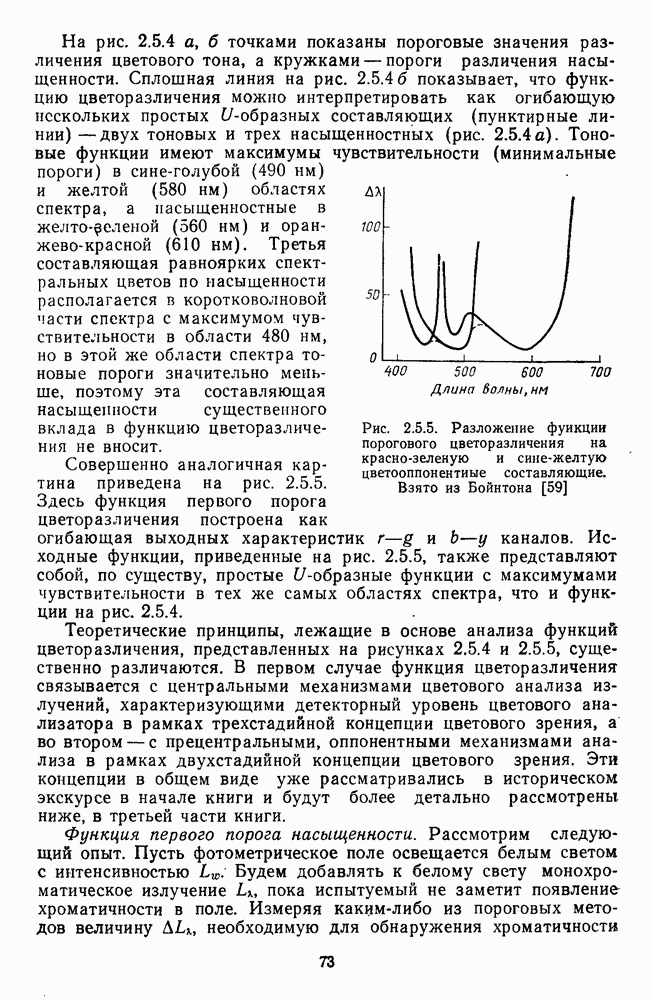
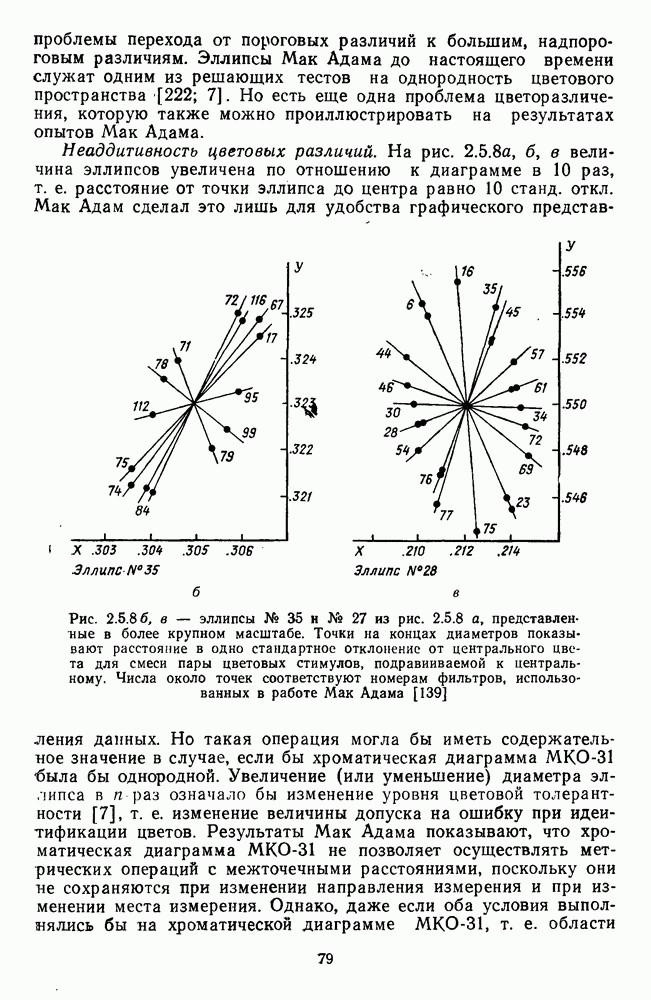
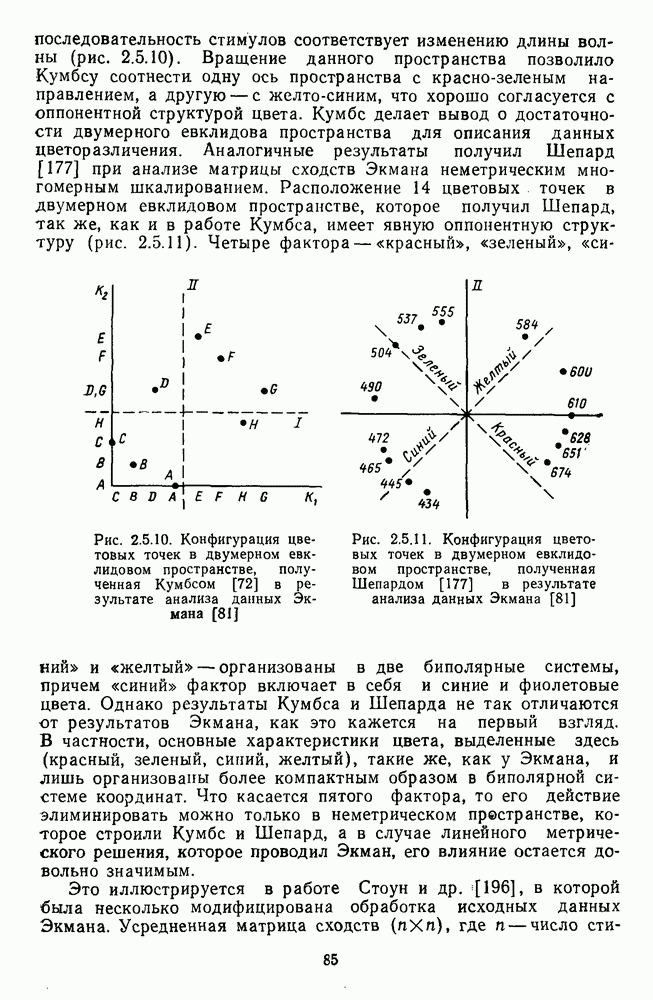
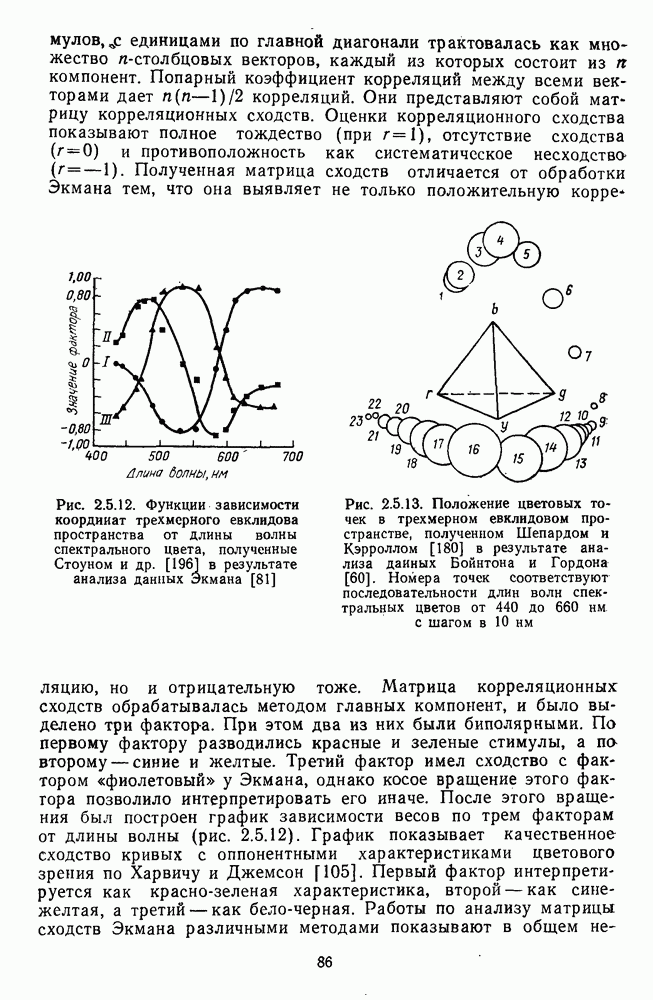
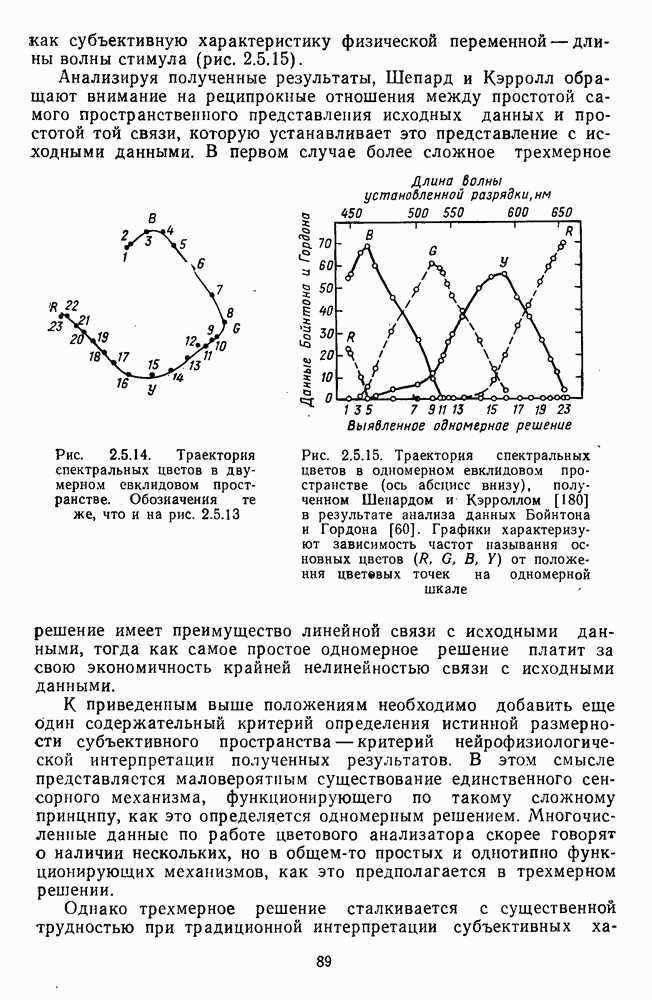
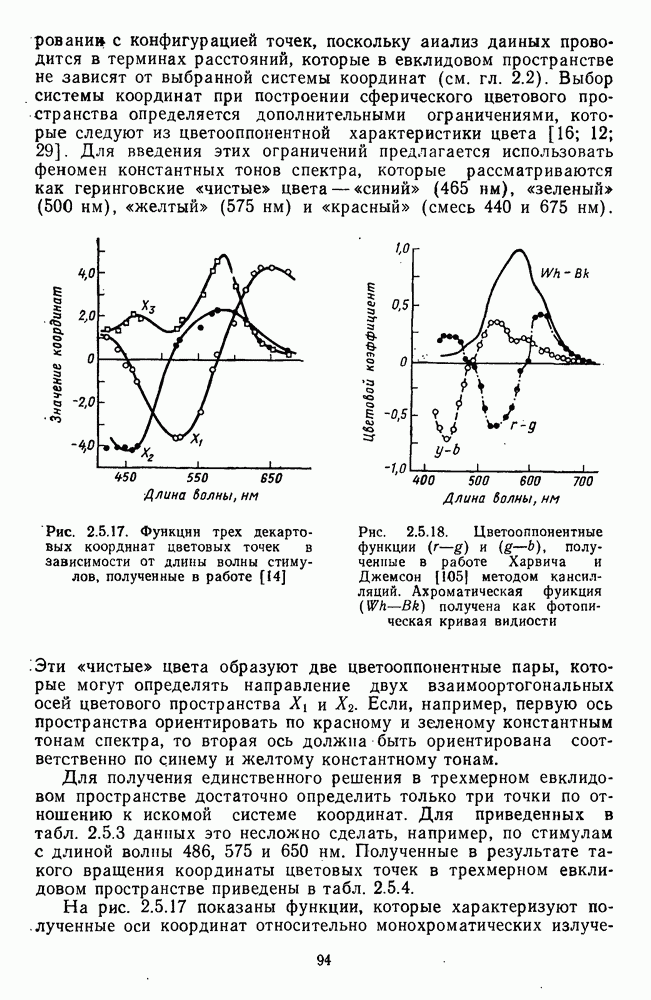
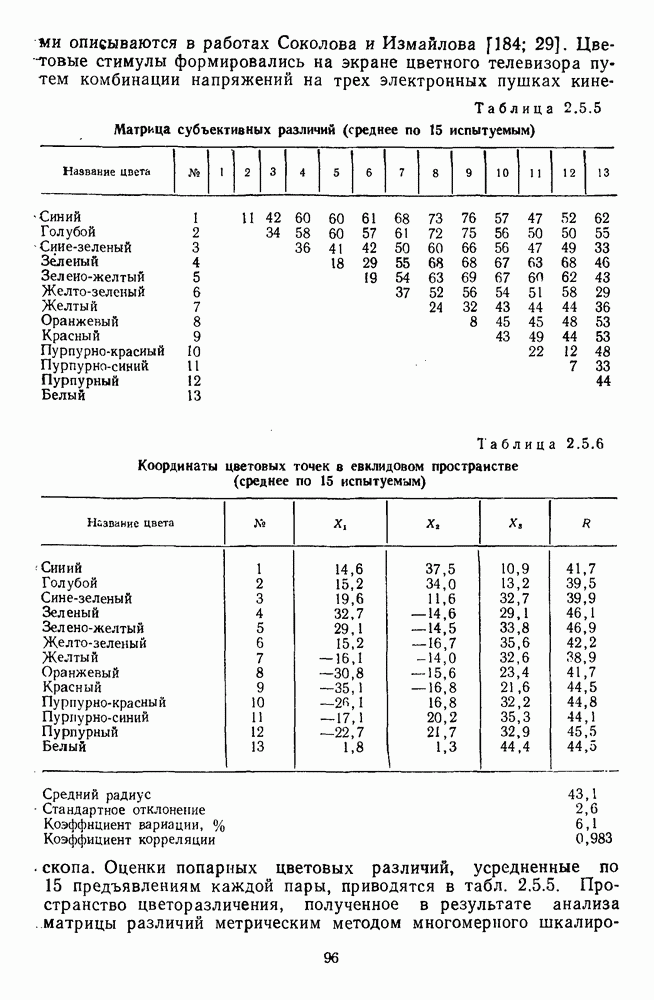
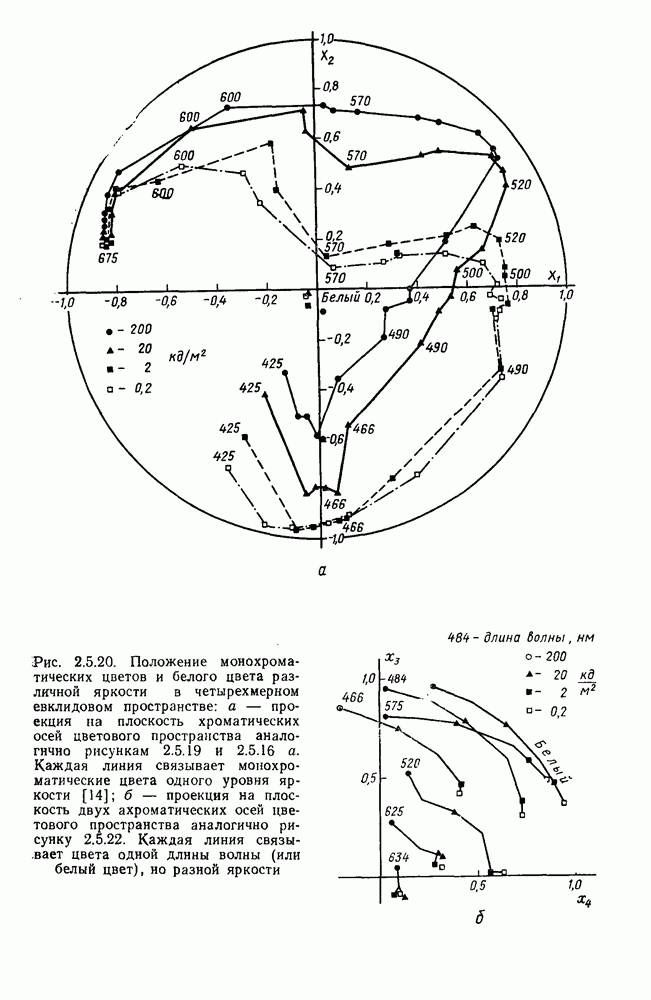
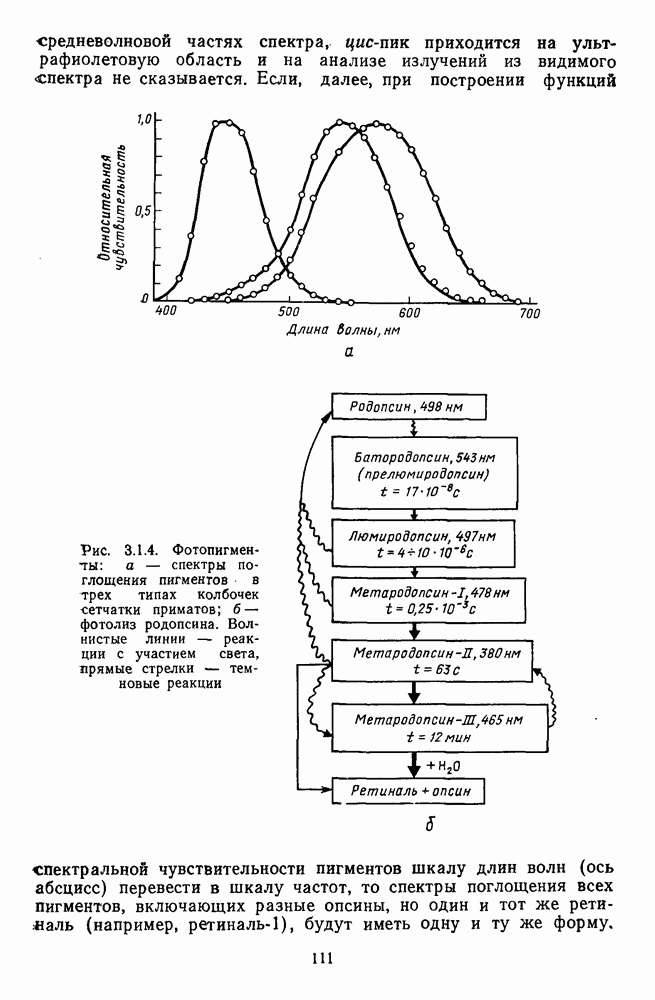
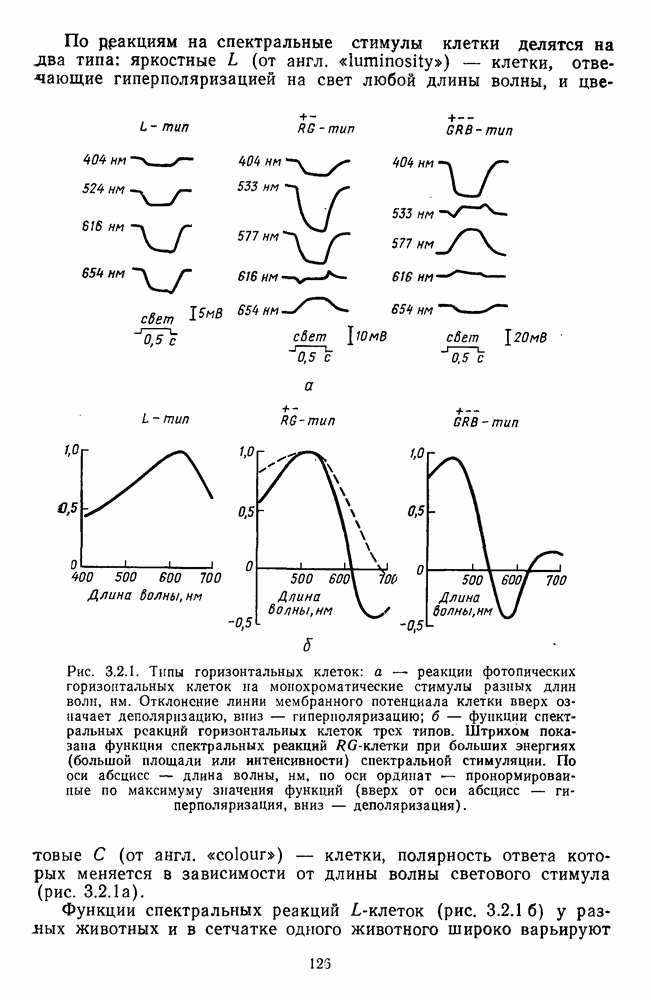
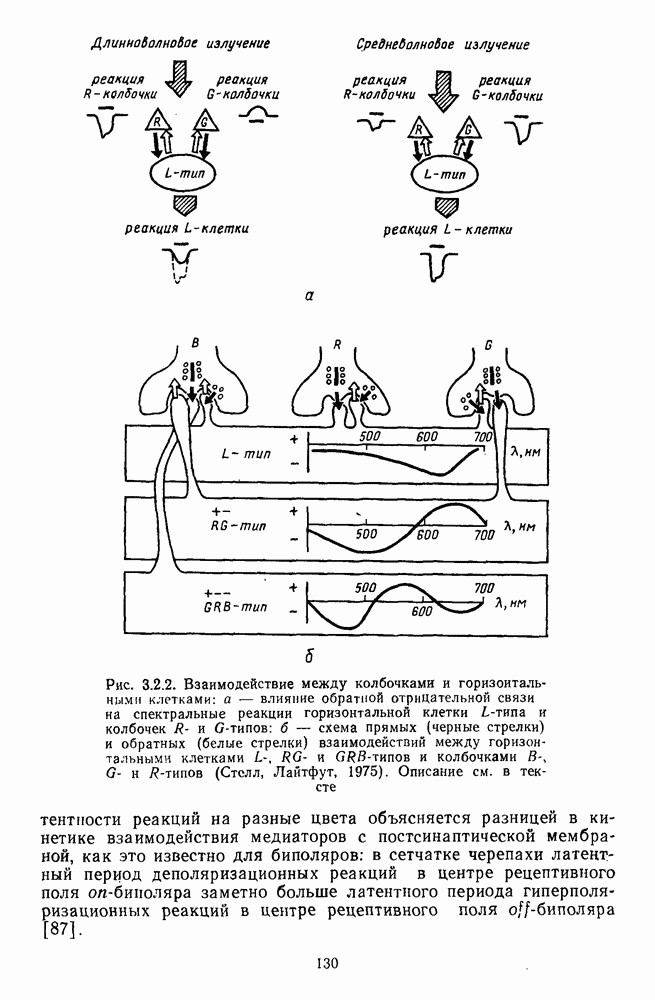
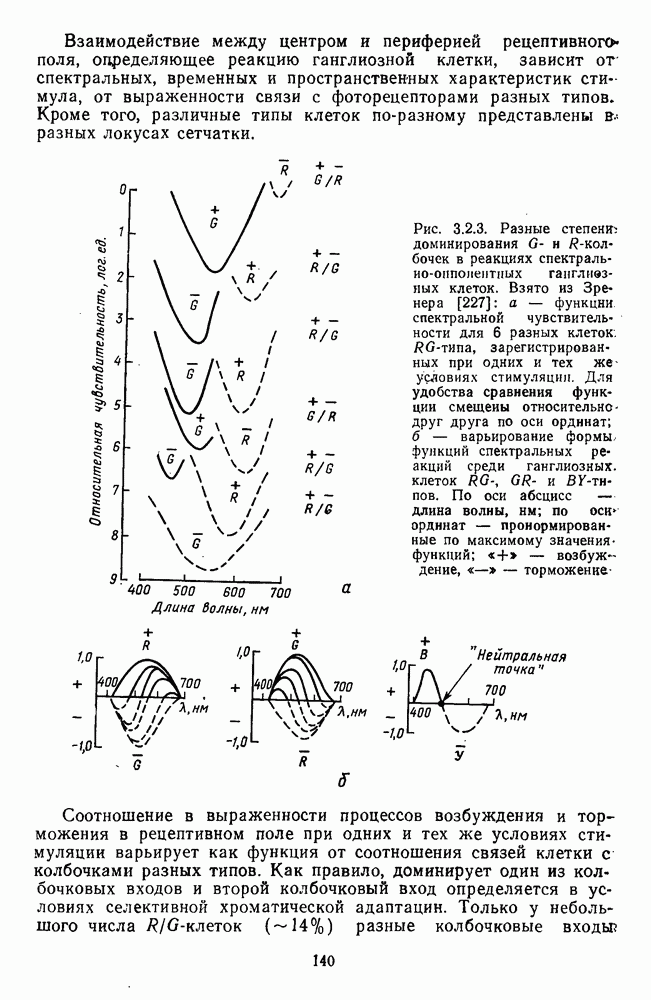
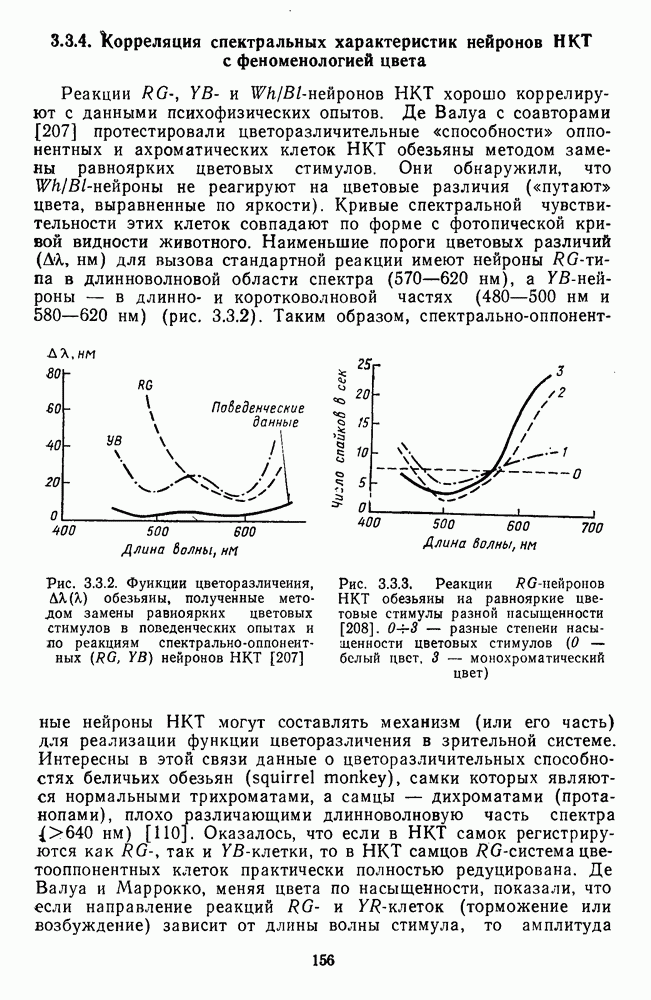
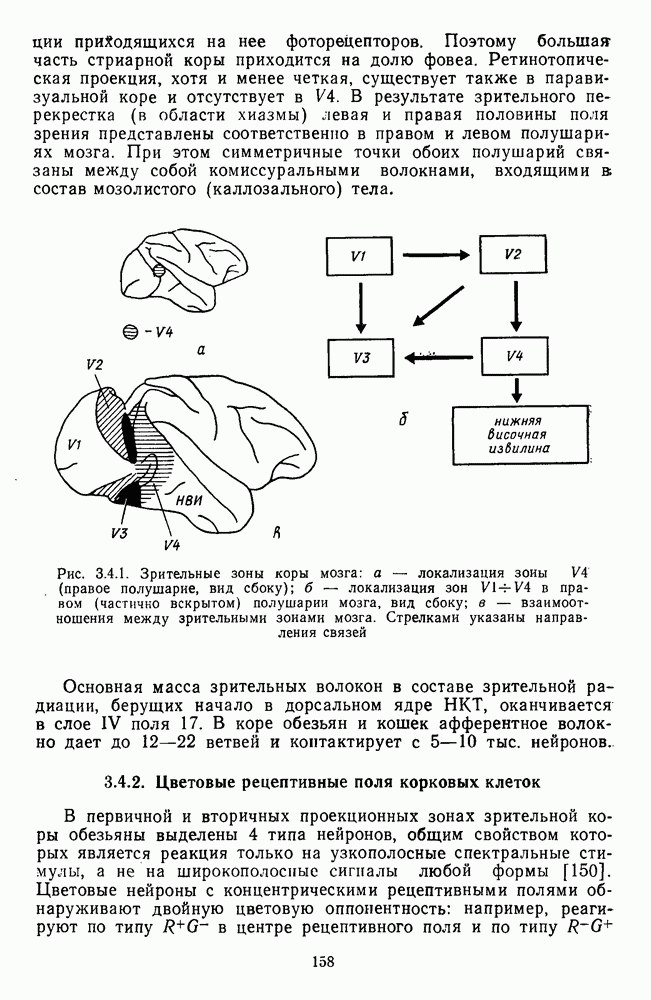
3.4.6. Особенности процессов кодирования цвета в престриарной зоне V4В престриарной коре обезьяны выделяют зону V4, специфическим образом связанную с кодированием цвета (рис. 3.4.1). V4 получает основные афферентные входы от полей 18 и 19 и частично от поля 17. Из V4 сигналы поступают в нижневисочную кору. В V4 представлена только центральная часть поля зрения (20-30°). РП нейронов этой зоны имеют небольшой по размерам Свойства цветовых клеток различных зон коры, включая зону V4, подробно изучены английским исследователем С. Зеки [226]. Им описаны три типа корковых нейронов: цветооппонентные нейроны и нейроны, избирательные к определенному узкому диапазону длин волн в зоне VI, и цветокодирующие — в зоне V4. Цветоппонентные клетки генерируют оя-ответы на один диапазон длин волн и освощалйсь излучениями с разными длинами волн, реакции цветооппонентных и спектрально-избирательных нейронов при этом могли резко меняться. По данным Зеки, клетки первых двух групп составляли большинство среди цветочувствительных клеток стриарной коры и меньшинство — среди нейронов V4. Основную часть цветовых нейронов V4 образовывали клетки третьей группы — цветокодирующие клетки. Эти нейроны реагировали избирательно на тот или иной цвет поверхности независимо от спектрального состава освещения и независимо от последовательности, в которой внешние источники света освещают изображение стимула. Однако, важным условием их максимальной активации является освещение объекта излучениями из всех трех областей спектра — коротко-, средне- и длинноволновой. При монохроматическом освещении эти клетки не реагируют на оптимальный для них стимул. Ответы этих нейронов не зависели от величины и формы цветового сигнала, от скорости и направления его движения. Ширина полосы спектра, в которой активируются нейроны поля V4, составляла в среднем 10—20 нм. Чаще всего встречались клетки с чувствительностью в полосе 480, 500 и 620 нм, реже — в полосе 550—570 нм. Отмечены нейроны, реагировавшие только на пурпурный и белый цвета. В совокупности спектральная чувствительность всей популяции нейронов V4 перекрывает диапазон види мого спектра. Клетки V4 организованы в вертикальные колонки, каждая из которых сигнализирует свой цвет: имеются «красные», «синие», «зеленые», «белые» и другие колонки. Таким образом, реакции цветокодирующих нейронов коррелируют с субъективно воспринимаемым человеком цветом, т. е. с цветом как с перцептом. Связь нейронов поля V4 с цветокодированием является в настоящее время предметом интенсивной научной дискуссии. По данным Шейн и др., цветоспецифичные нейроны в V4 составляют всего обездвижено, вследствие чего экстраретинальные входы коры были заторможены и функционировали только сетчаточные, чисто афферентные входы. Не исключены и другие, не менее важные причины, объясняющие расхождения в данных, — трудности анатомической идентификации области V4, уровень анестезин, влияние светового или хроматического фона, способ стимуляции и выбор стимулов. Например, Зеки в своих работах использовал «мон-дриан» — цветную матрицу на экране дисплея, освещаемую тремя источниками света из длинно-, средне- и коротковолновых областей спектра [135]. Далее, авторы не указывают полушарие мозга, в котором осуществлялась регистрация, хотя из литературы известны различия между правым и левым полушариями в способах обработки сенсорной информации. В правом полушарии локализуют механизмы детального описания образа, а в левом — ?механизмы схематического (обобщенного) описания, связанного с выделением ограниченного числа главных информативных признаков [22]. Не исключено, что такая специализация полушарий распространяется и на цветовое зрение. В пользу этого могут свидетельствовать данные о различиях в восприятии яркости и насыщенности цветов людьми с доминированием правого или левого глаза [170]. Все вышеперечисленное затрудняет сравнение данных разных авторов и не позволяет категорично отрицать роль V4 в цветовом зрении. Обобщая, можно выделить в ретино-таламо-кортикальном пути преобразования зрительной информации две подсистемы Х-ней-ронов, ответственных, по крайней мере, за восприятие цвета и формы, и У-нейронов, связанных с восприятием движения. Цветовое кодирование в X- и У-нейронах выполняет разные функции. Х-нейроны выделяют цветовые свойства как таковые и, кроме то-то, участвуют в выделении цветовых контуров. В У-нейронах цветовое кодирование используется для выделения движущихся объектов на цветовом фоне. Х-нейроны образуют основу для трех каналов параллельной обработки информации: для выделения элементов контура (в том числе с использованием цветового кодирования) с последующим опознанием изображения в клетках инферотемпоральной коры; для цветокодирующих нейронов константного типа в V4; и для пространственной константности в нейронах париетальной коры. Собственно цветовое кодирование образует отдельный канал, который начинается на уровне колбочек и включает, далее, фотопические горизонтальные клетки, биполяры и ганглиозные клетки Х-типа и Х-нейроны с двойной оппонентностью наружного коленчатого тела. Завершается система кодирования цвета спектрально-селективными нейронами зрительной коры аконстантного типа. Информация, выделенная из светового излучения этой системой цветового анализатора, распределяется далее в различные зоны коры, где она используется для построения более сложных психических феноменов. Например, можно предположить, что включение цвета в образ восприятия — перцепт — осуществляется через цветоконстантные нейроны поля V4, а включение цвета в мнемические феномены (например, образы представления цвета) осуществляется с помощью клеток лобной коры, которая обеспечивает сохранение «контекста, временной организации событий» [23]. Такая же информация может поступать в зоны формирования речи, где она используется для генерации цветовых названий. Современная психофизиология еще только подходит к установлению связей между нейронными механизмами различных психических функций, но можно с большой долей уверенности полагать, что именно в этом направлении будут сосредоточены усилия психофизики, нейрофизиологии и психофизиологии в ближайшие десятилетия.
|
 |
Оглавление
|















































































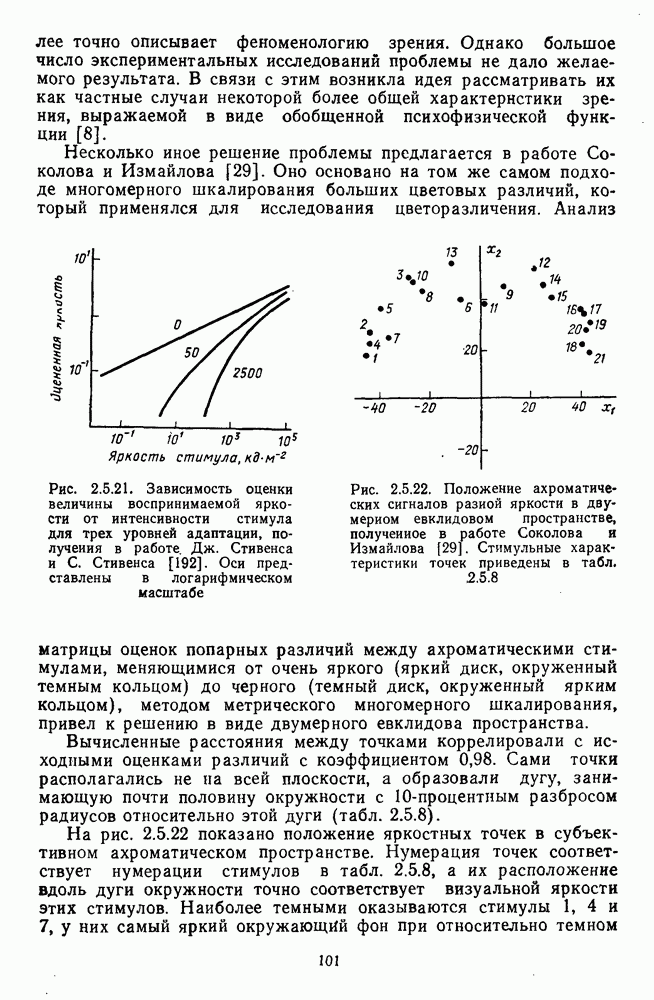

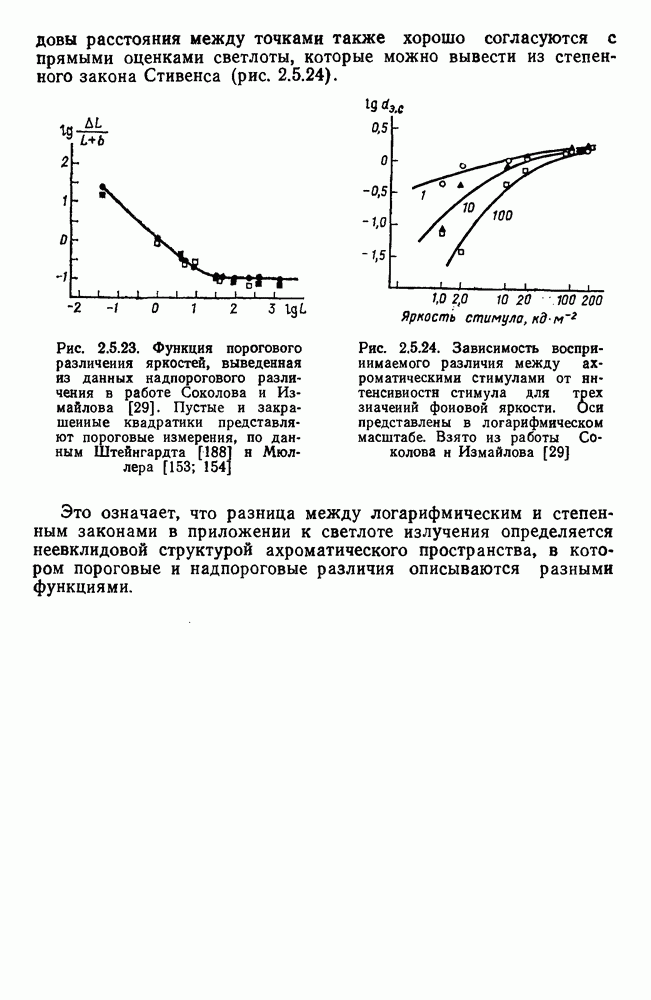


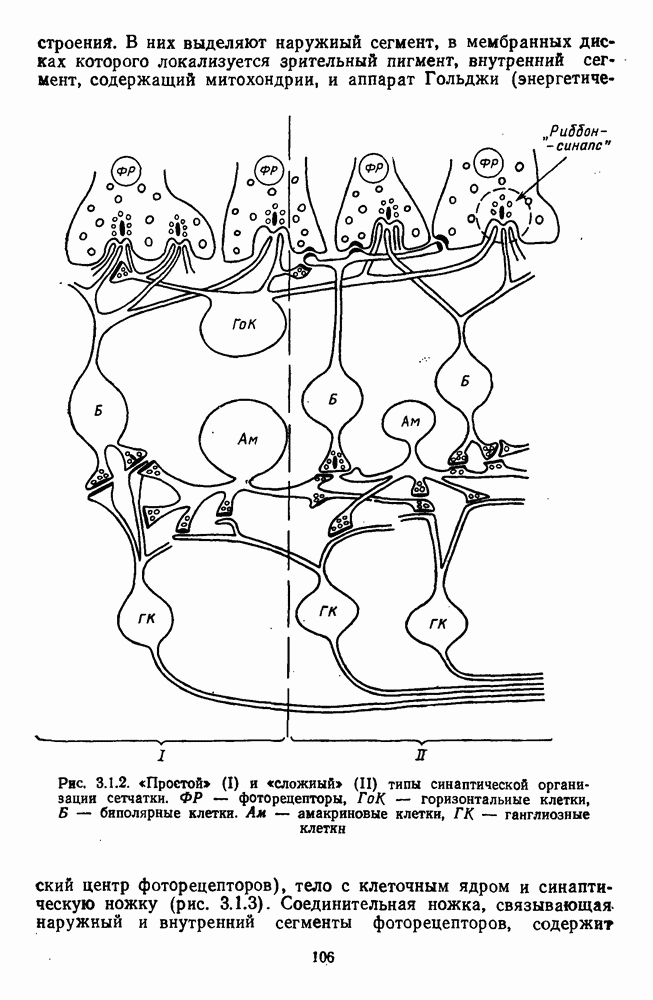
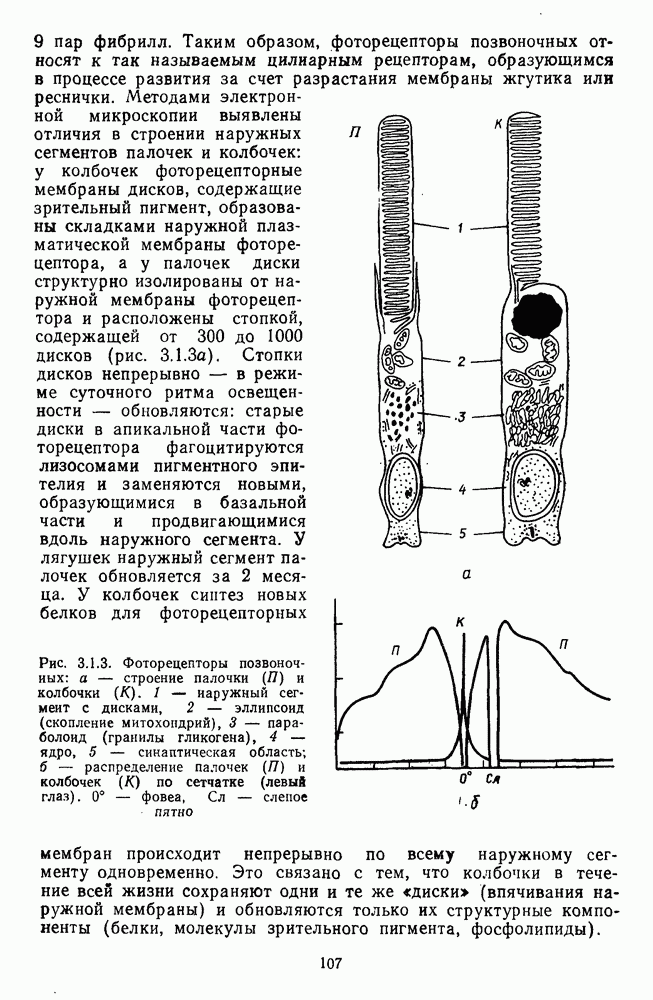






























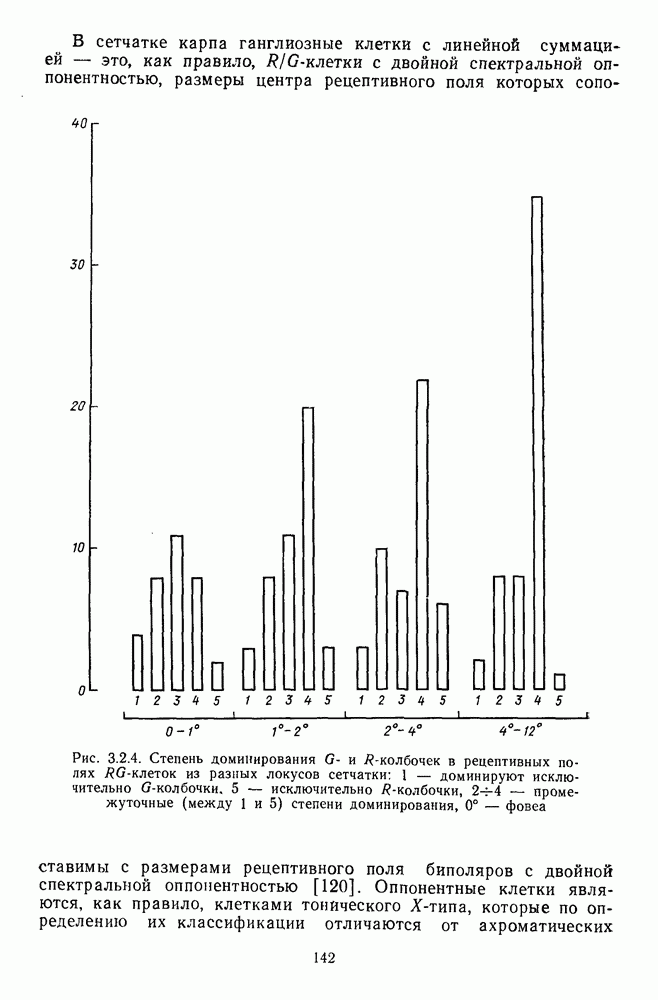
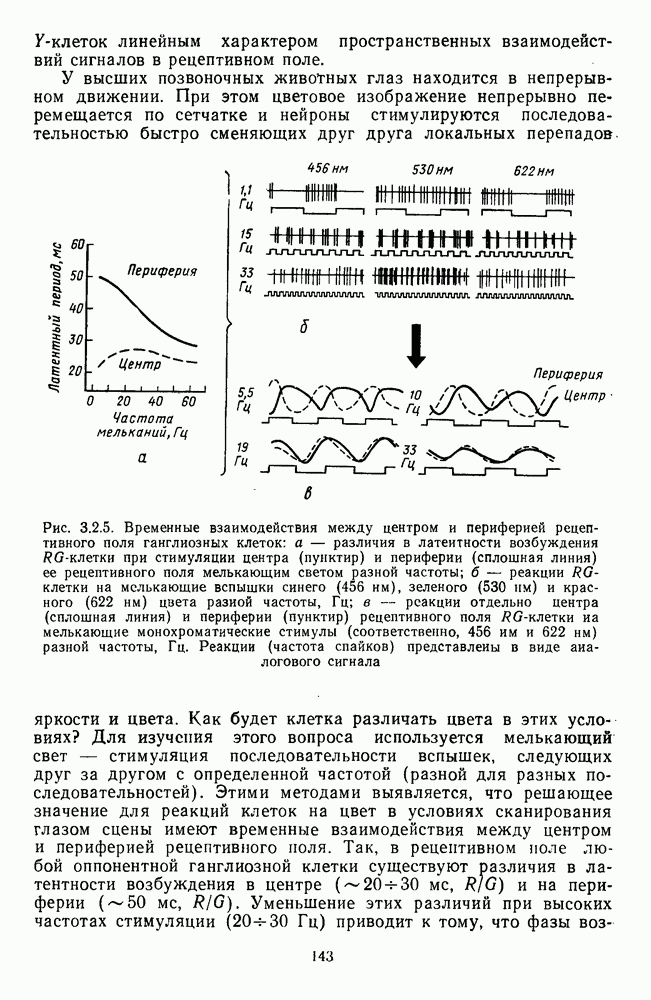
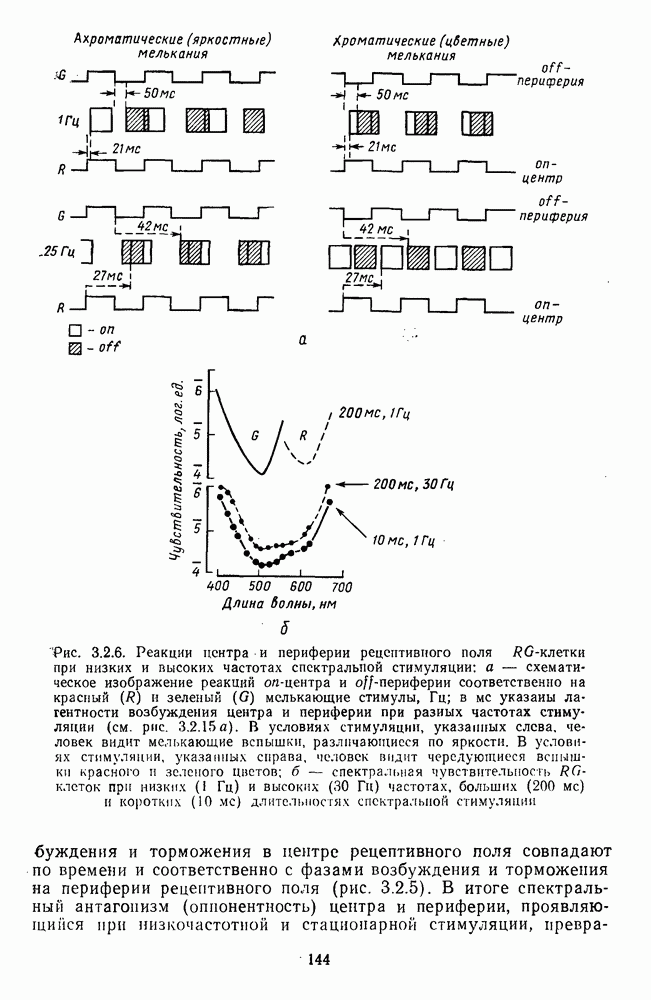

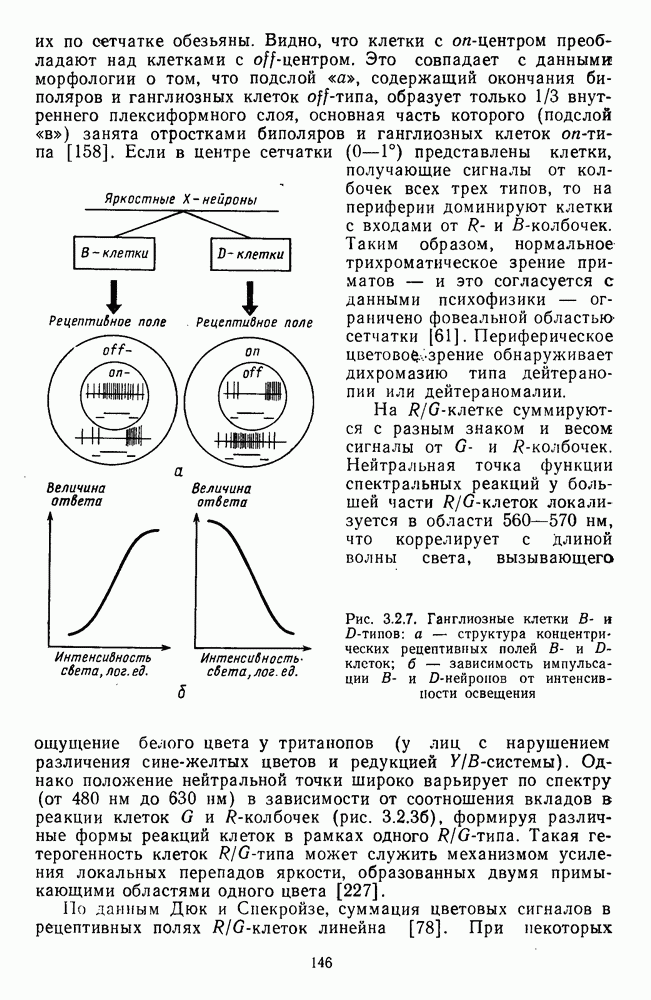

















































 возбуждающий центр и обширную «молчащую» периферию
возбуждающий центр и обширную «молчащую» периферию  наличие которой можно выявить только по ее тормозному влиянию на реакцию центра. За счет больших размеров периферии рецептивные поля нейронов V4 в среднем в 30 раз превышают по площади РП нейронов стриарной коры. Тормозное воздействие периферии РП максимально, если свойства проецируемого на нее стимула совпадают (или близки) со свойствами стимула, возбуждающего центр (размеры, спектральный состав и др.). В зоне V4 отсутствует ретинотопическая проекция.
наличие которой можно выявить только по ее тормозному влиянию на реакцию центра. За счет больших размеров периферии рецептивные поля нейронов V4 в среднем в 30 раз превышают по площади РП нейронов стриарной коры. Тормозное воздействие периферии РП максимально, если свойства проецируемого на нее стимула совпадают (или близки) со свойствами стимула, возбуждающего центр (размеры, спектральный состав и др.). В зоне V4 отсутствует ретинотопическая проекция.
 -ответы — на другой. Свойства большинства этих клеток совпадают со свойствами аналогичных нейронов НКТ и сетчатки (за исключением клеток с двойной цветовой оппонентностью). Нейроны из второй группы избирательно реагировали на узкий диапазон длин волн в спектре. Реакции этих клеток на цветные поверхности, освещаемые различным светом, зависели от общего спектрального состава отраженного от поверхности света: чем больше в отраженном от объекта свете было длин волн, к которым был чувствителен данный нейрон, тем больше была его реакция. Активация нейронов этого типа, как и цветоппонентных клеток, зависела от порядка, в котором стимулы разных длин волн вводились в РП. Например, если клетке, чувствительной к длинноволновому излучению, предъявляли в ее рецептивное поле квадратик красного цвета, освещенный длинноволновым излучением, то клетка активировалась, а затем, когда к освещению красного квадратика добавляли другие длины волн, то эта клетка тормозилась. При таком добавлении средне- и коротковолнового освещения количество красного цвета в отраженном от экрана дисплея свете не изменялось и стимул по-прежнему был виден как красный. Однако если освещали красный квадратик в обратном порядке — сначала синим и зеленым, а потом добавляли красный цвет, то вначале клетка не отвечала, а возбуждалась только при добавке красного освещения. Таким образом, несмотря на то что окончательный цвет поверхности экрана оставался неизменным, а менялась очередность, с которой его поверхность
-ответы — на другой. Свойства большинства этих клеток совпадают со свойствами аналогичных нейронов НКТ и сетчатки (за исключением клеток с двойной цветовой оппонентностью). Нейроны из второй группы избирательно реагировали на узкий диапазон длин волн в спектре. Реакции этих клеток на цветные поверхности, освещаемые различным светом, зависели от общего спектрального состава отраженного от поверхности света: чем больше в отраженном от объекта свете было длин волн, к которым был чувствителен данный нейрон, тем больше была его реакция. Активация нейронов этого типа, как и цветоппонентных клеток, зависела от порядка, в котором стимулы разных длин волн вводились в РП. Например, если клетке, чувствительной к длинноволновому излучению, предъявляли в ее рецептивное поле квадратик красного цвета, освещенный длинноволновым излучением, то клетка активировалась, а затем, когда к освещению красного квадратика добавляли другие длины волн, то эта клетка тормозилась. При таком добавлении средне- и коротковолнового освещения количество красного цвета в отраженном от экрана дисплея свете не изменялось и стимул по-прежнему был виден как красный. Однако если освещали красный квадратик в обратном порядке — сначала синим и зеленым, а потом добавляли красный цвет, то вначале клетка не отвечала, а возбуждалась только при добавке красного освещения. Таким образом, несмотря на то что окончательный цвет поверхности экрана оставался неизменным, а менялась очередность, с которой его поверхность
 от общего числа регистрируемых клеток, что почти не отличается от содержания таких нейронов в других зрительных областях [172]. В опытах Крюгер и Фишера [132] 90% клеток из 300, зарегистрированных в зоне V4 макаки-резуса, хорошо реагировали на нейтральный свет. Остальные 10% не реагировали ни на белый, ни на спектральный цвета. Из 90% клеток, реагировавших на нейтральный свет,
от общего числа регистрируемых клеток, что почти не отличается от содержания таких нейронов в других зрительных областях [172]. В опытах Крюгер и Фишера [132] 90% клеток из 300, зарегистрированных в зоне V4 макаки-резуса, хорошо реагировали на нейтральный свет. Остальные 10% не реагировали ни на белый, ни на спектральный цвета. Из 90% клеток, реагировавших на нейтральный свет,  селективно выделяли свою специфическую область спектра (красную, зеленую, синюю и желтую) и слабо возбуждались белым и другими цветами. Из этих 7% большая часть нейронов реагировала на красный цвет — цвет точки фиксации глаз обезьяны в опытах. Таким образом, Крюгер и Фишер не обнаружили в этой области нейронов с такой цветовой избирательностью, как это описано у Зеки. Причиной расхождения последних работ с работой Зеки, регистрировавшего в V4 до 90% цветоселективных клеток, могут быть различия в условиях содержания животного во время эксперимента. В опытах Зеки животное было анестезировано и
селективно выделяли свою специфическую область спектра (красную, зеленую, синюю и желтую) и слабо возбуждались белым и другими цветами. Из этих 7% большая часть нейронов реагировала на красный цвет — цвет точки фиксации глаз обезьяны в опытах. Таким образом, Крюгер и Фишер не обнаружили в этой области нейронов с такой цветовой избирательностью, как это описано у Зеки. Причиной расхождения последних работ с работой Зеки, регистрировавшего в V4 до 90% цветоселективных клеток, могут быть различия в условиях содержания животного во время эксперимента. В опытах Зеки животное было анестезировано и