Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
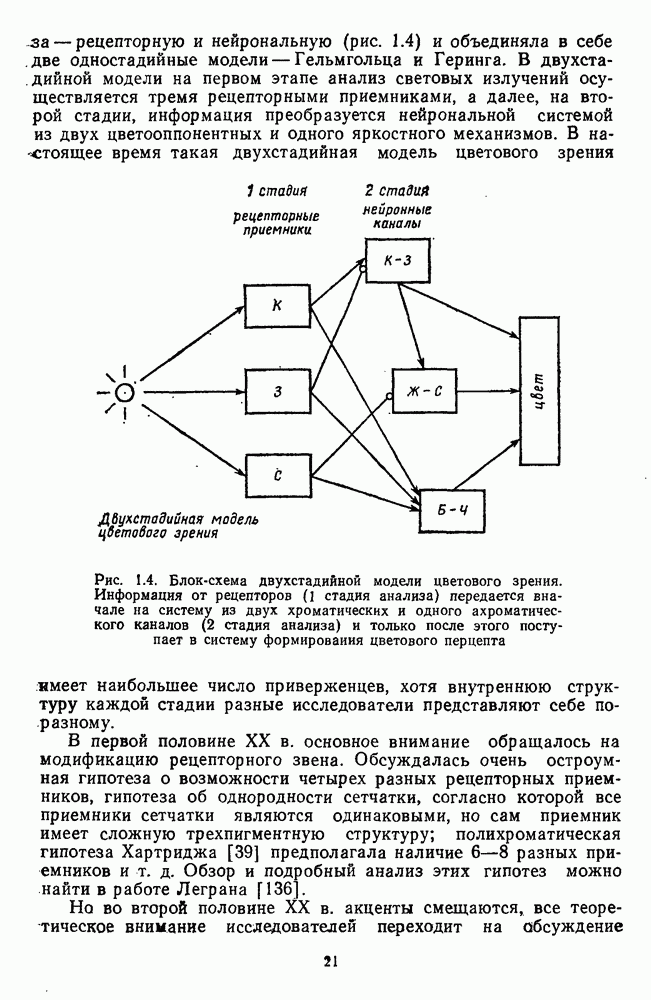
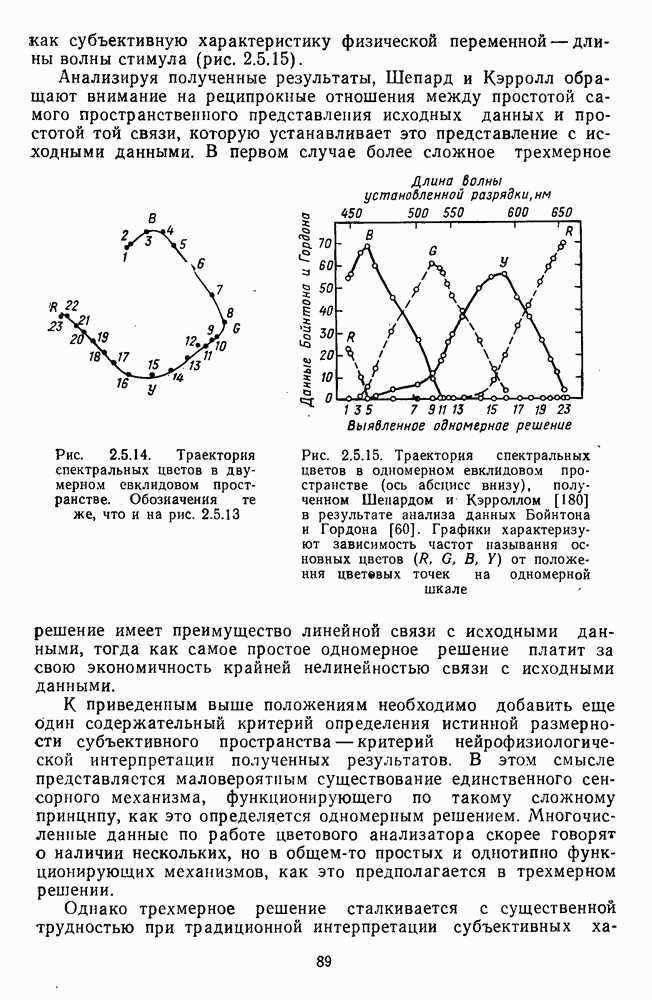
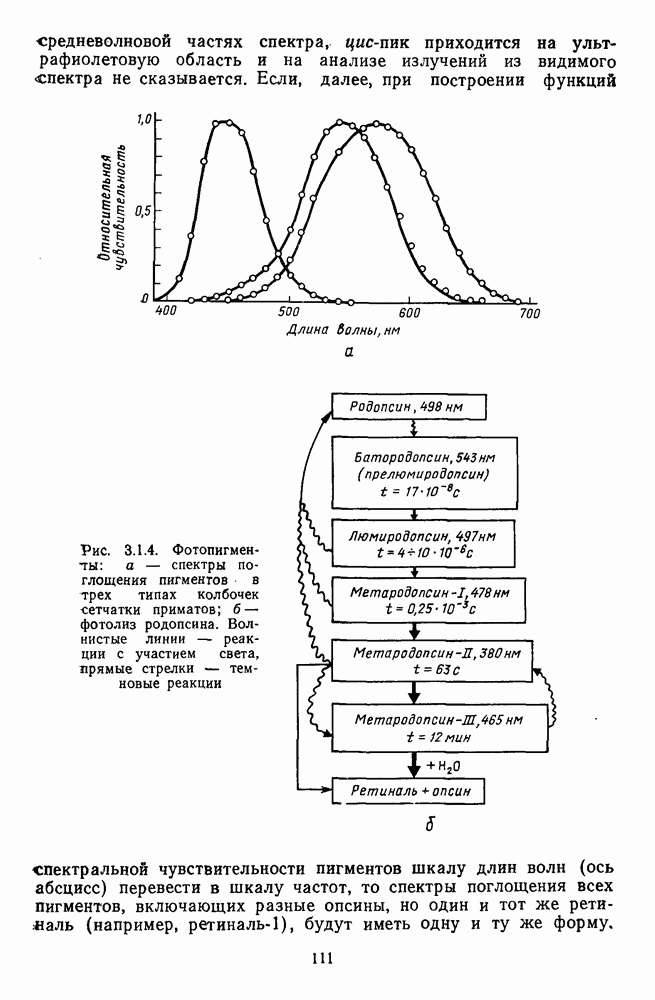
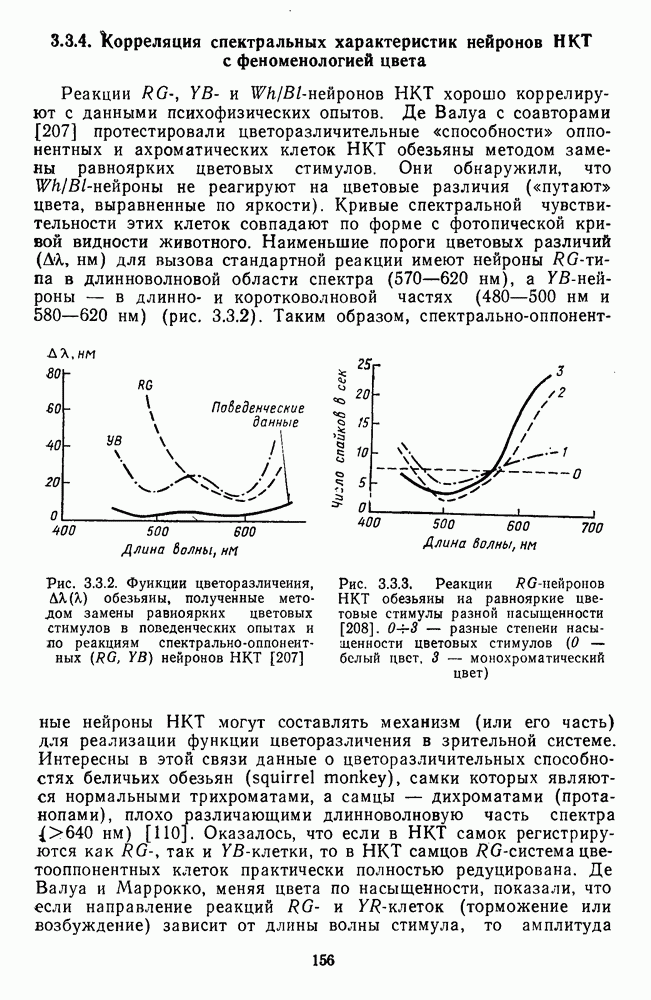
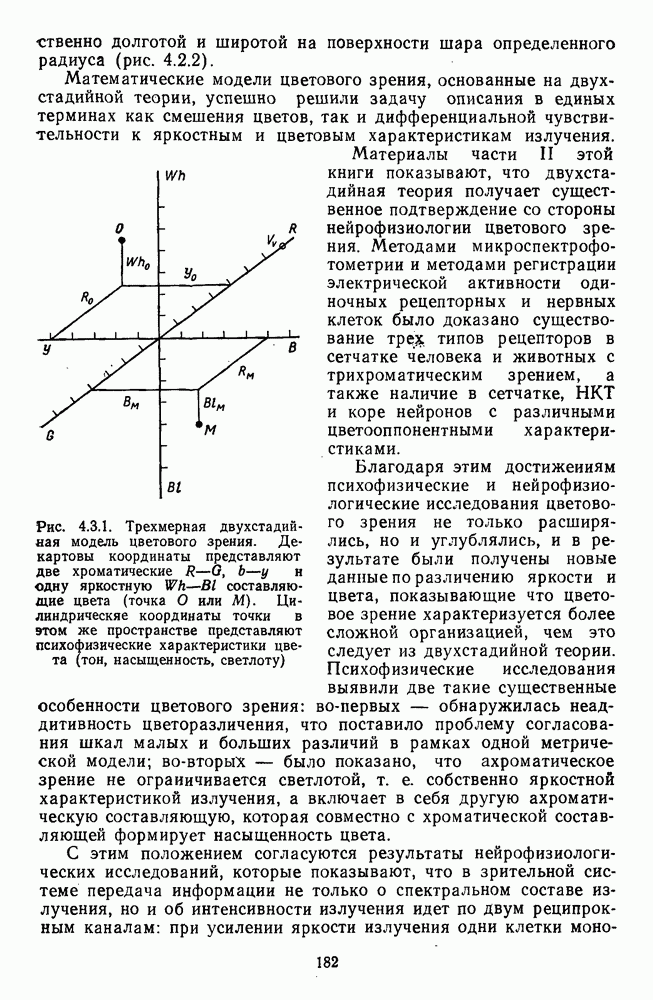
3.3.4. Корреляция спектральных характеристик нейронов НКТ с феноменологией цветаРеакции
Рис. 3.3.2. Функции цветоразличения,
Рис. 3.3.3. Реакции Таким образом, спектрально-оппонентные нейроны НКТ могут составлять механизм (или его часть) для реализации функции цветоразличения в зрительной системе. Интересны в этой связи данные о цветоразличительных способностях беличьих обезьян (squirrel monkey), самки которых являются нормальными трихроматами, а самцы — дихроматами (прота-нопами), плохо различающими длинноволновую часть спектра ответа зависит от насыщенности при условии, что интенсивность по-разному насыщенных цветов при этом не меняется (рис. 3.3.3) [208]. Имеется положительная корреляция между степенью развитости цветового зрения животного и выраженностью системы оппонентных нейронов в НКТ. Так, в НКТ макаки, цветовое зрение которой близко к цветовому зрению человека, оппонентные клетки составляют 60—70% от общего числа регистрируемых нейронов. А в НКТ кошки, цветоразличительные способности которой крайне ограничены, спектрально-оппонентные клетки составляют всего несколько процентов [161]. Итак, система спектрально-оппонентных нейронов в НКТ мало отличается от таковой в слое ганглиозных клеток. Сходная организация рецептивных полей нейронов сетчатки и нейронов НКТ является основанием для широко распространенного мнения о том, что НКТ — это релейная (передающая) «станция». Однако с учетом усиления латерального торможения в НКТ можно думать, что смысл такого дублирования состоит в необходимости чисто количественного усиления тех эффектов, которые обеспечиваются концентрической организацией рецептивного поля: выделение и обострение контрастов [32; 33]. Возможна также «коррекция» на уровне НКТ афферентного потока со стороны других отделов мозга [32].
|
 |
Оглавление
|












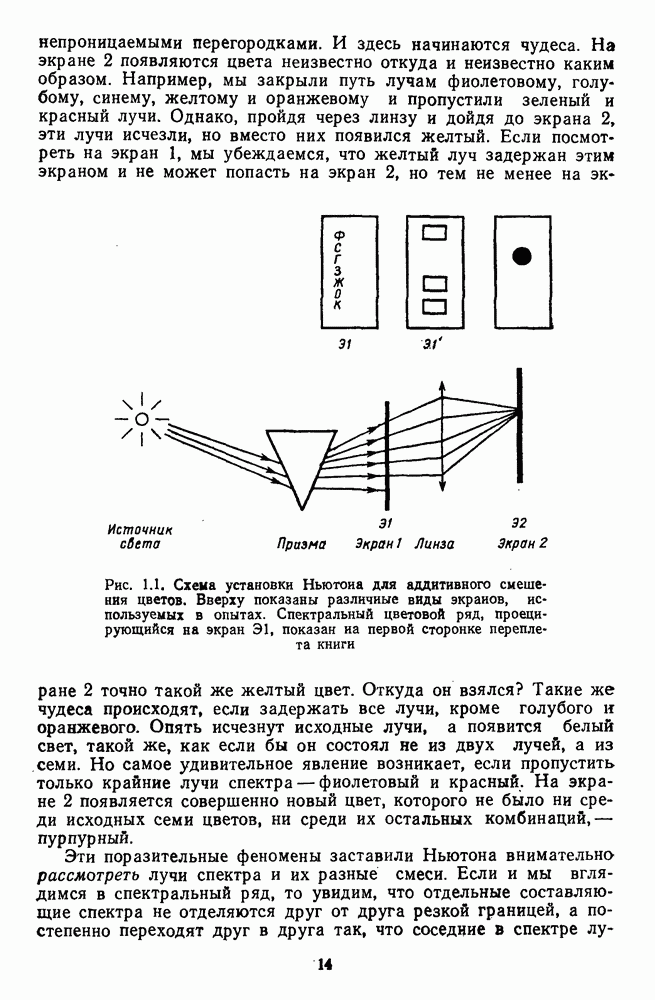




































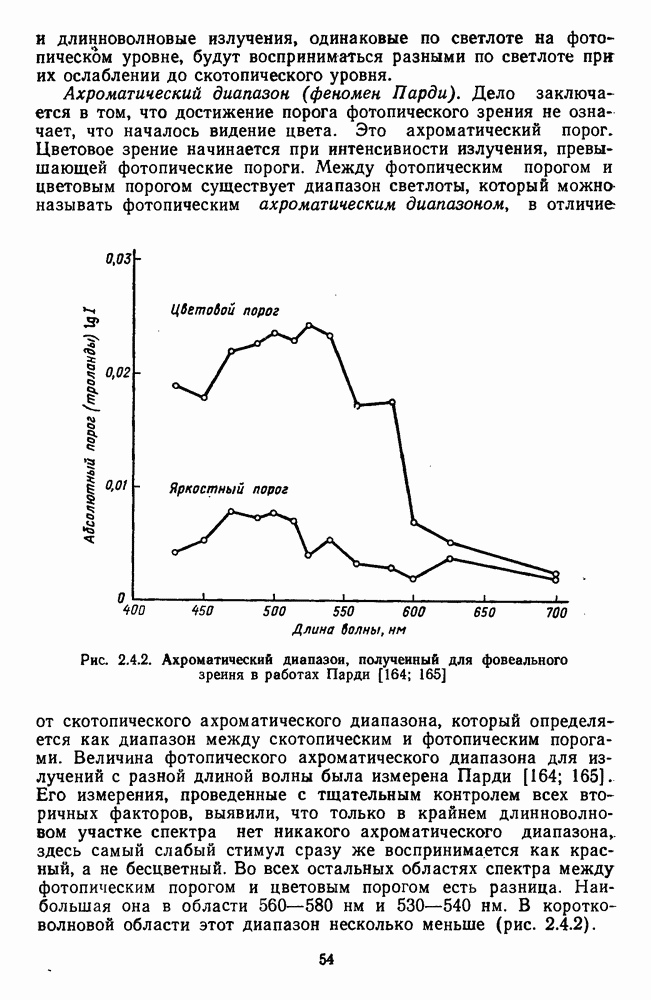



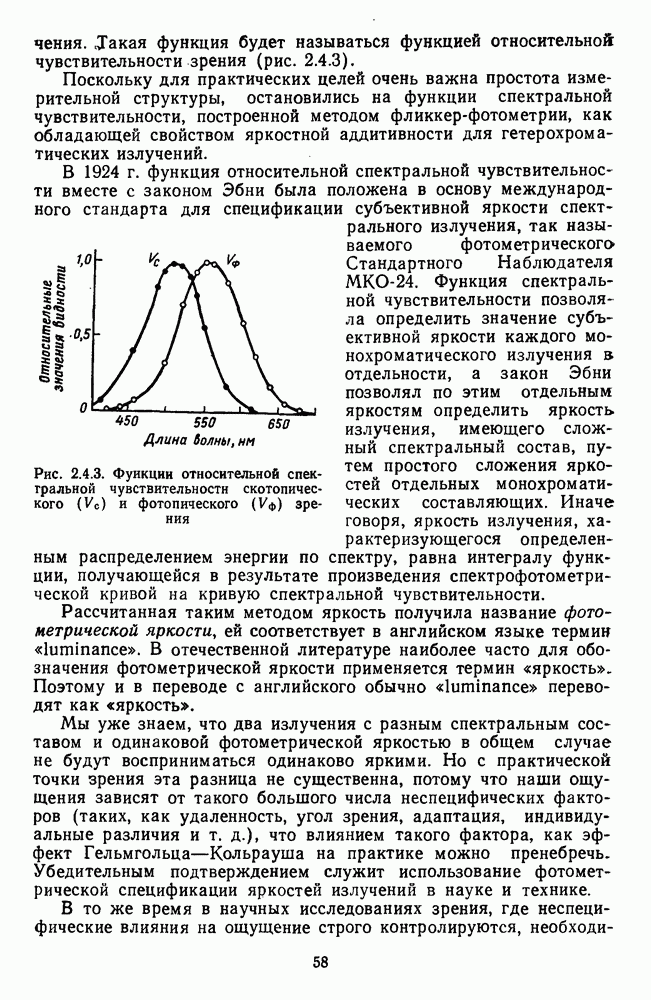

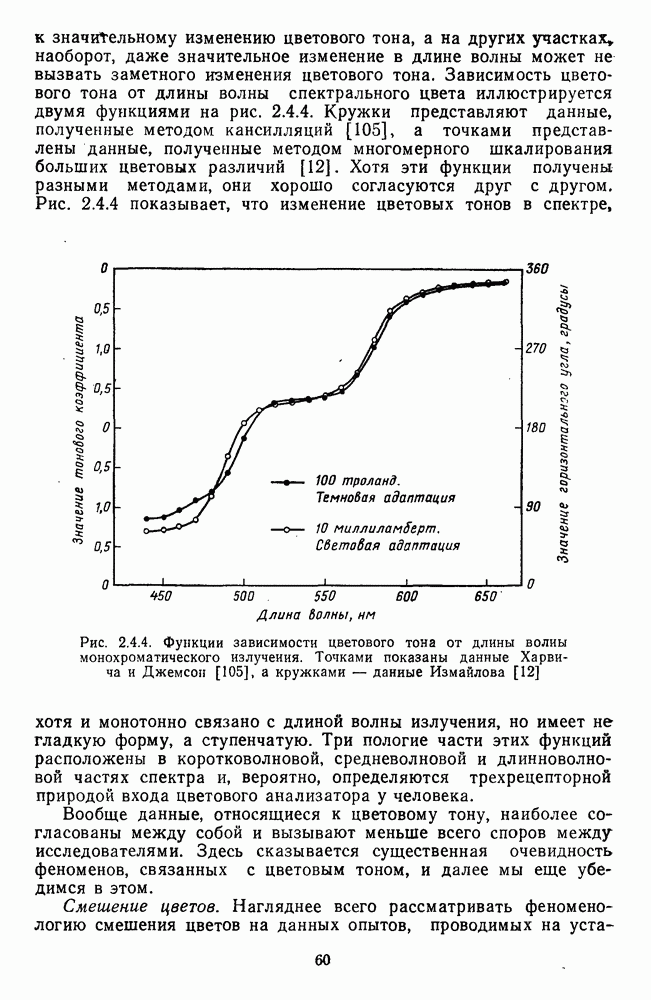







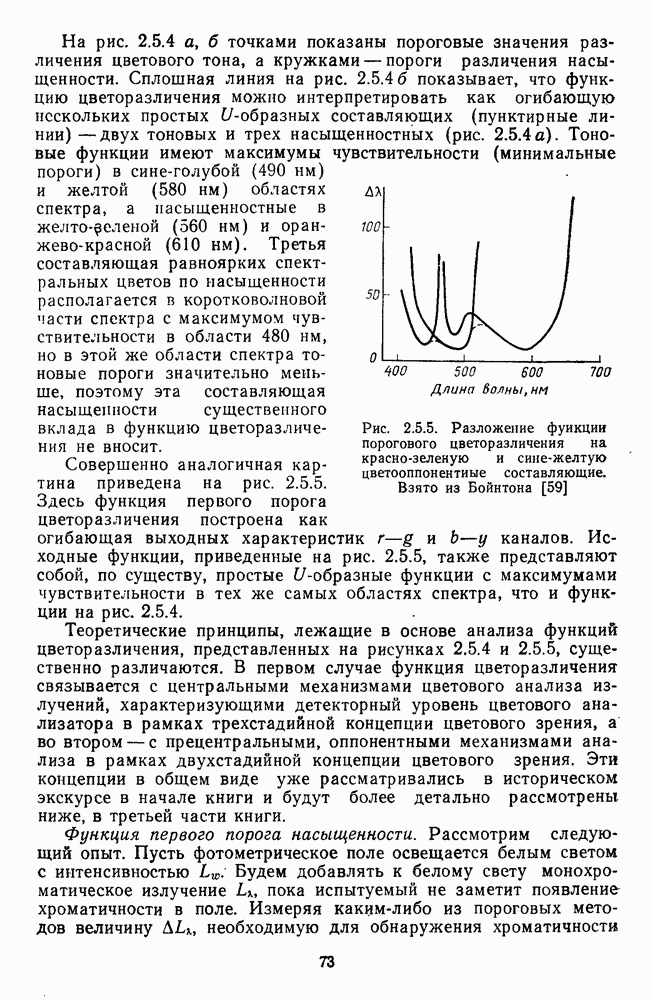





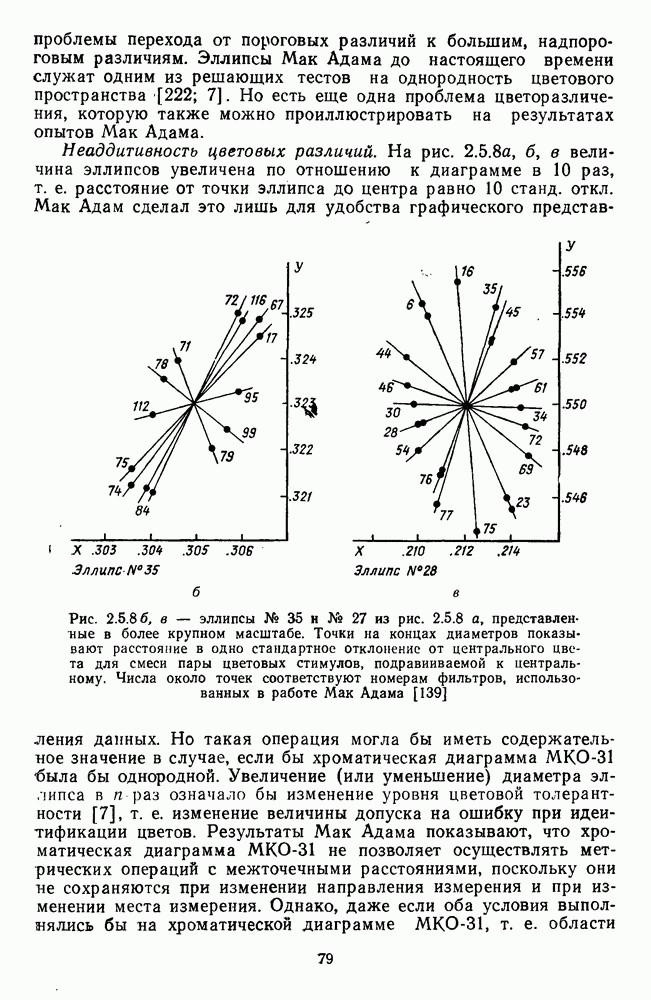





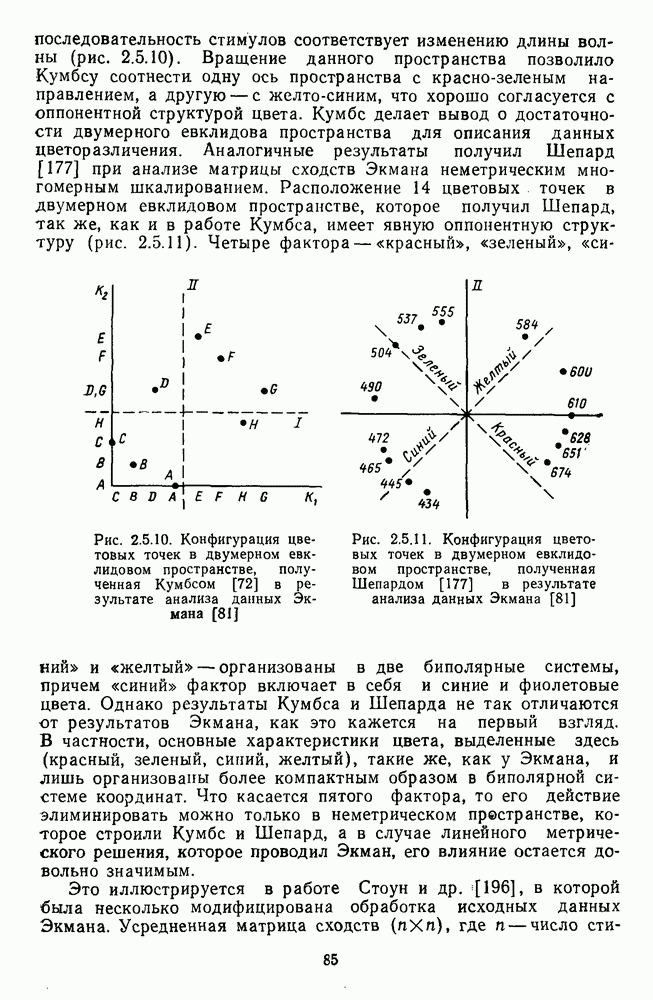
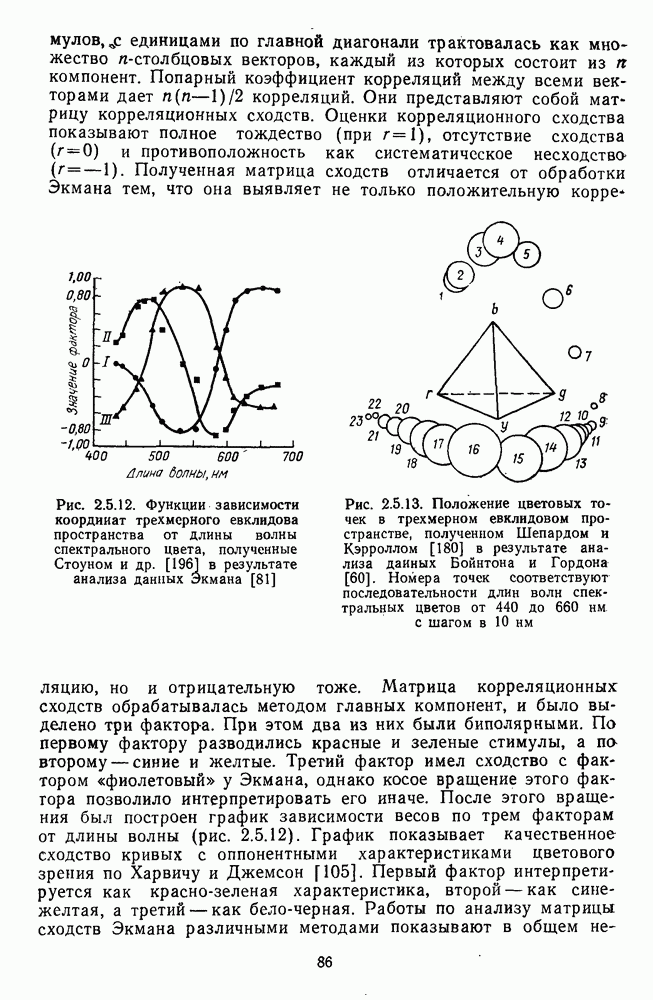










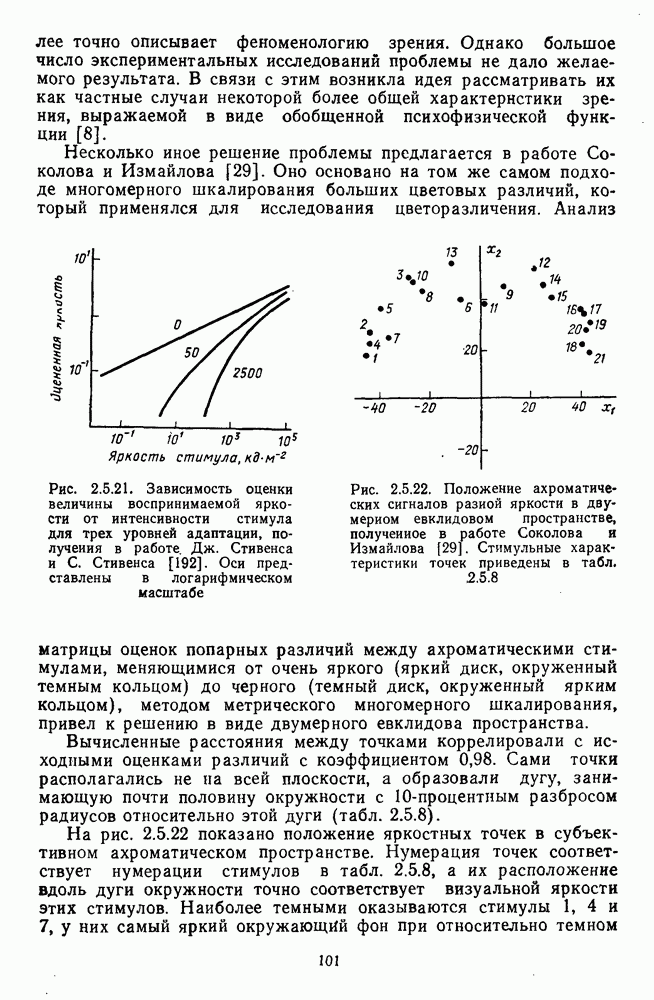

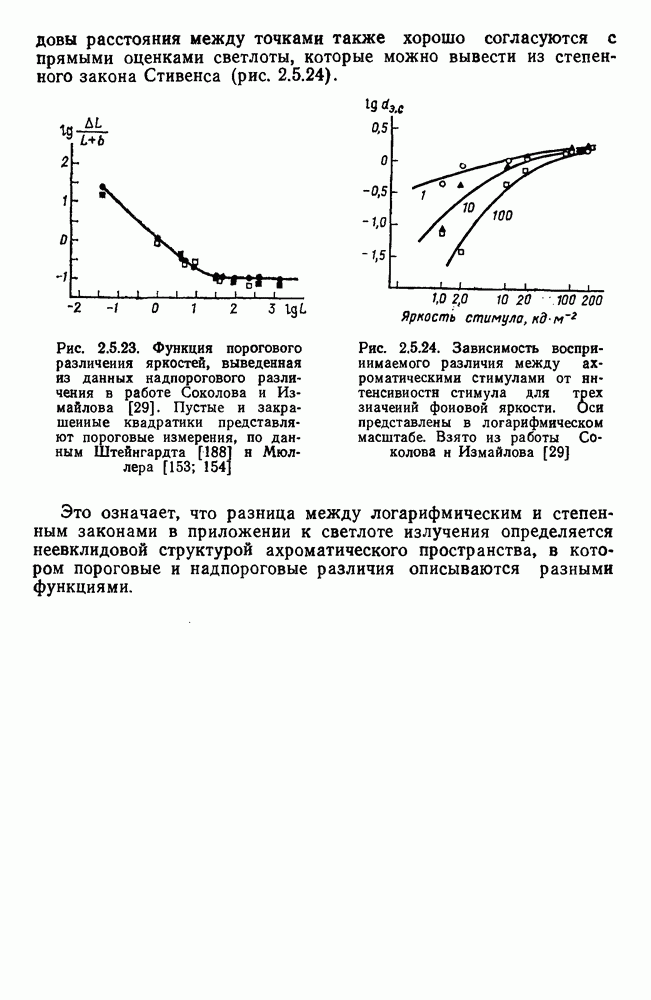


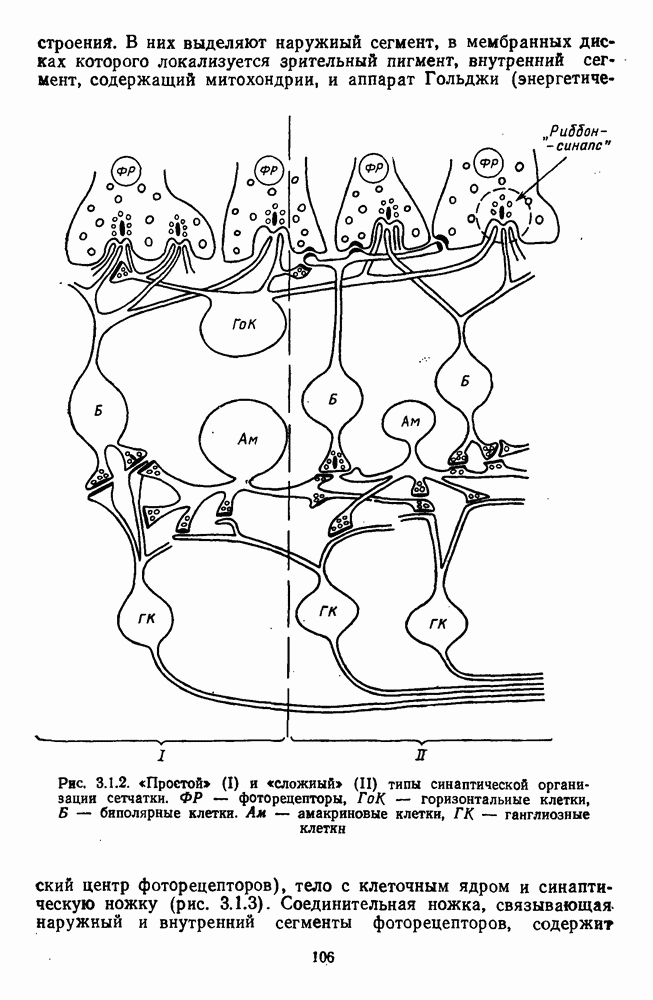
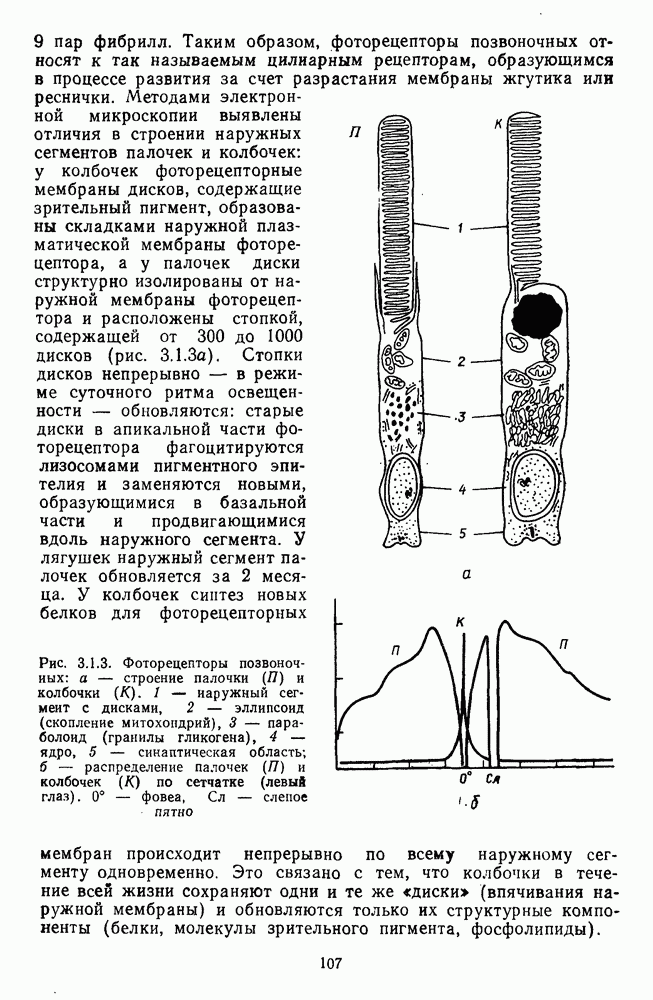
















































































 и
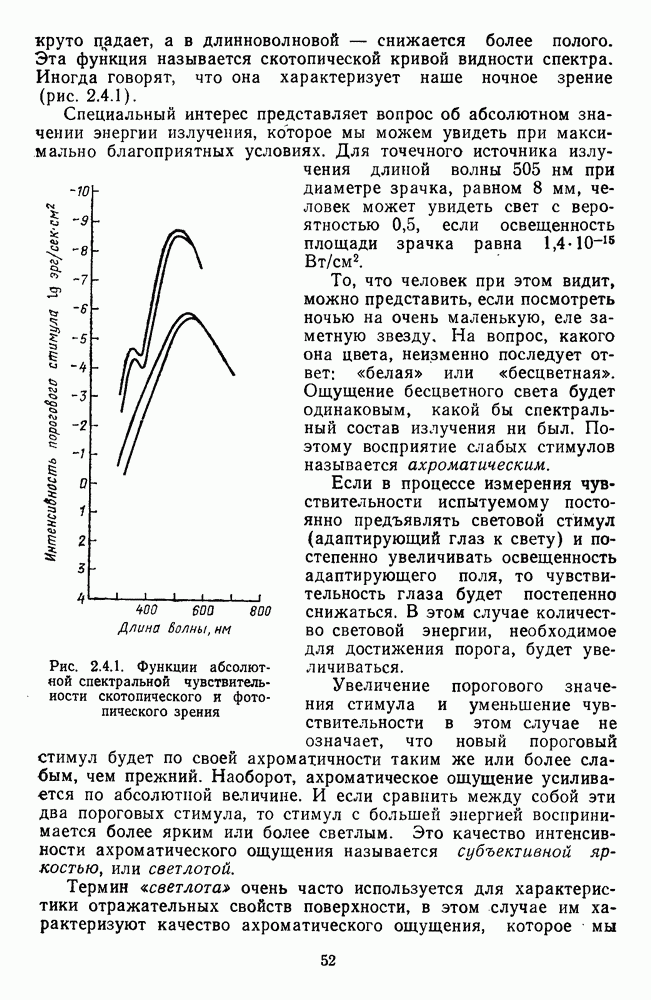
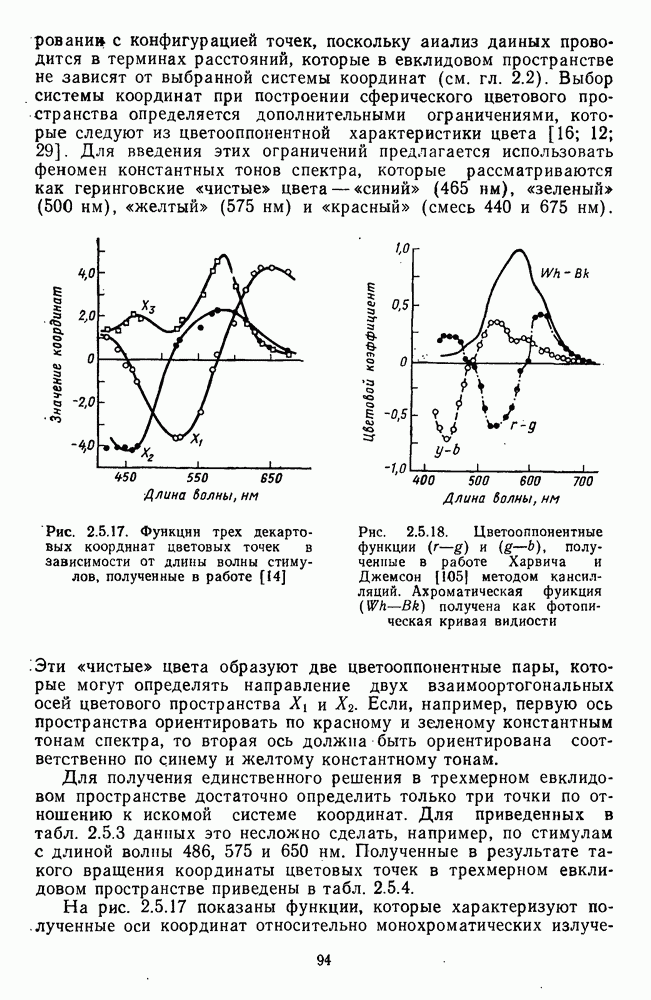
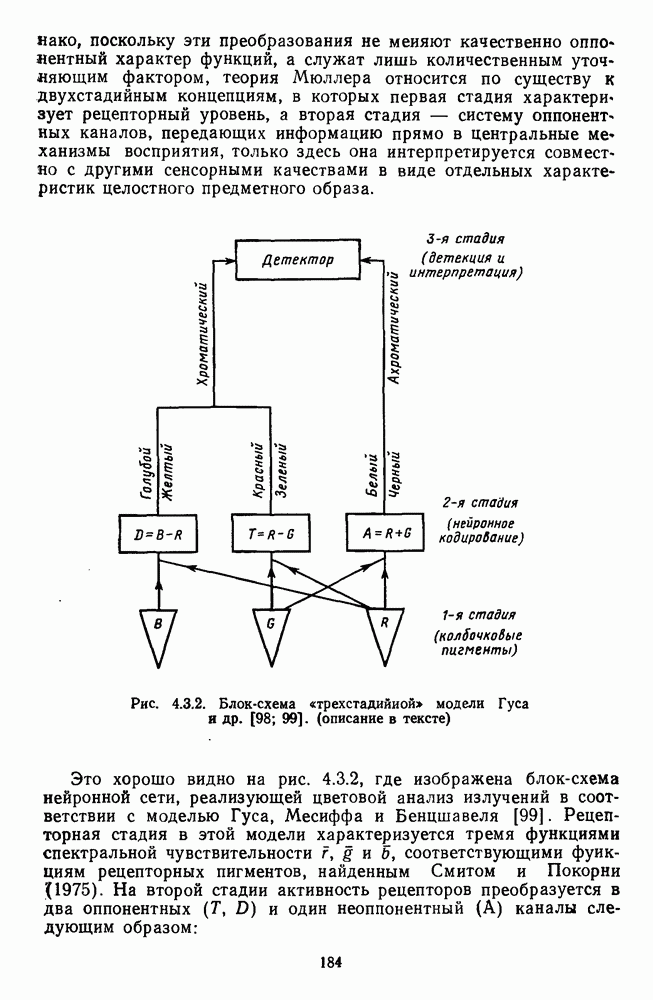
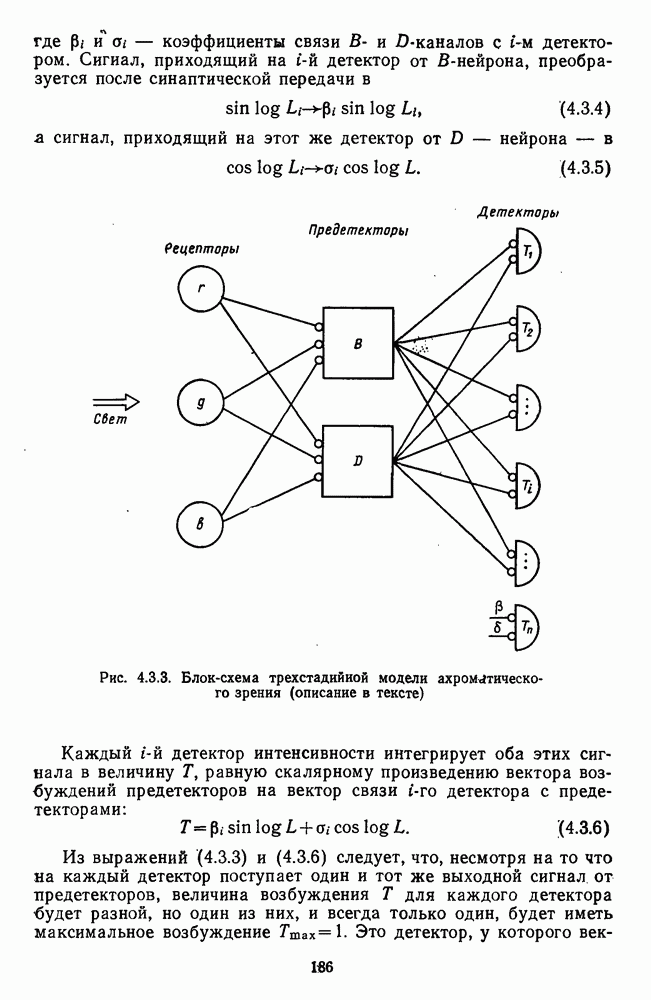
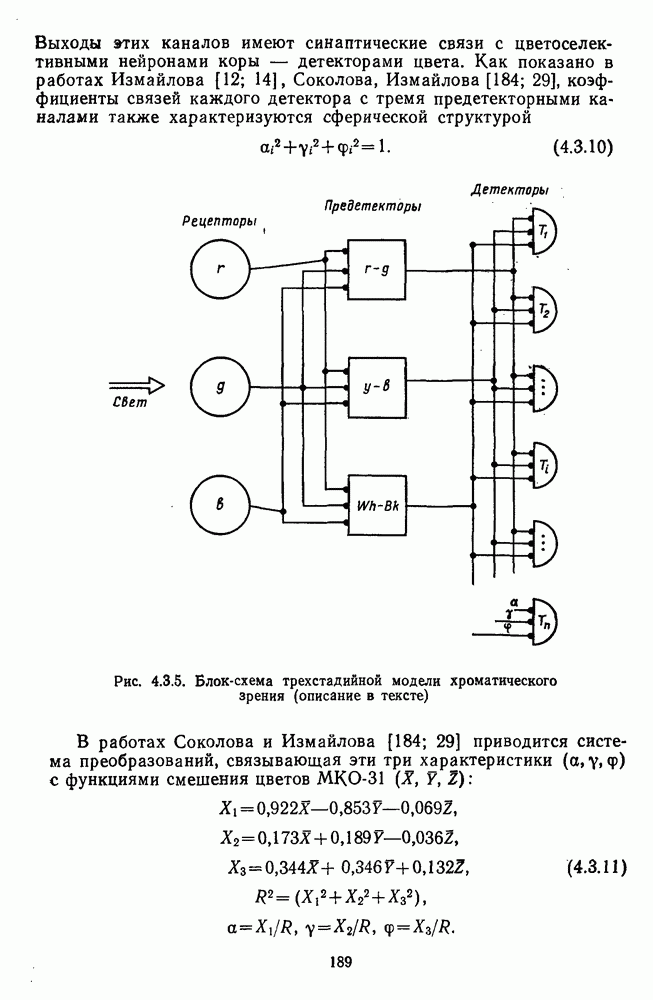
и  -нейронов НКТ хорошо коррелируют с данными психофизических опытов. Де Валуа с соавторами (207] протестировали цветоразличительные «способности» оппонентных и ахроматических клеток НКТ обезьяны методом замены равноярких цветовых стимулов. Они обнаружили, что
-нейронов НКТ хорошо коррелируют с данными психофизических опытов. Де Валуа с соавторами (207] протестировали цветоразличительные «способности» оппонентных и ахроматических клеток НКТ обезьяны методом замены равноярких цветовых стимулов. Они обнаружили, что  -нейроны не реагируют на цветовые различия («путают» цвета, выравненные по яркости). Кривые спектральной чувствительности этих клеток совпадают по форме с фотопической кривой видности животного. Наименьшие пороги цветовых различий
-нейроны не реагируют на цветовые различия («путают» цвета, выравненные по яркости). Кривые спектральной чувствительности этих клеток совпадают по форме с фотопической кривой видности животного. Наименьшие пороги цветовых различий  для вызова стандартной реакции имеют нейроны
для вызова стандартной реакции имеют нейроны  -типа в длинноволновой области спектра (570—620 нм), а
-типа в длинноволновой области спектра (570—620 нм), а  -ней-роны — в длинно- и коротковолновой частях (480—500 нм и 580—620 нм) (рис. 3.3.2).
-ней-роны — в длинно- и коротковолновой частях (480—500 нм и 580—620 нм) (рис. 3.3.2).

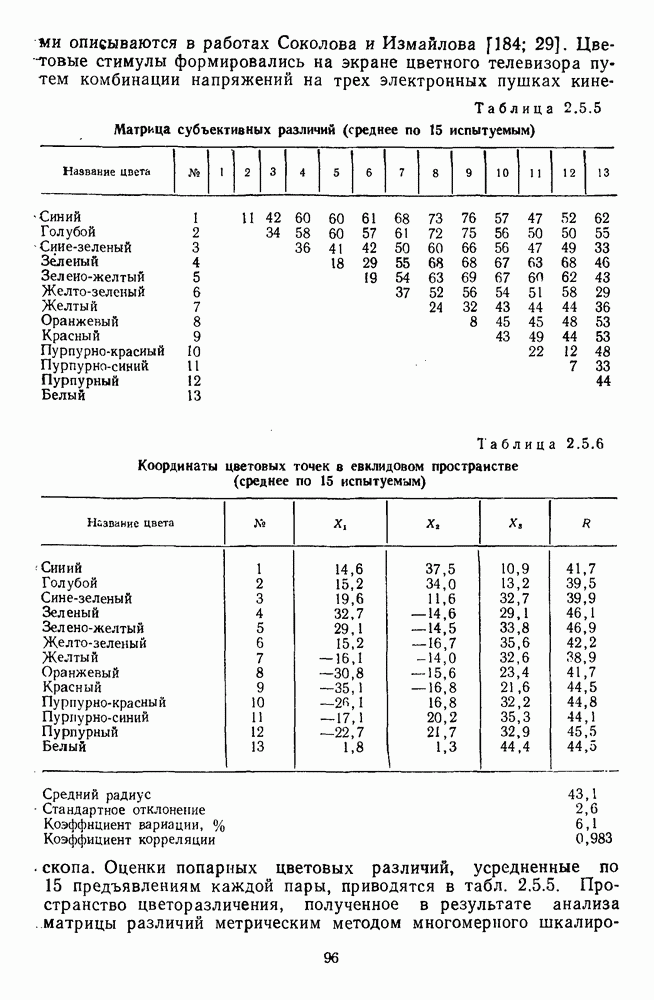
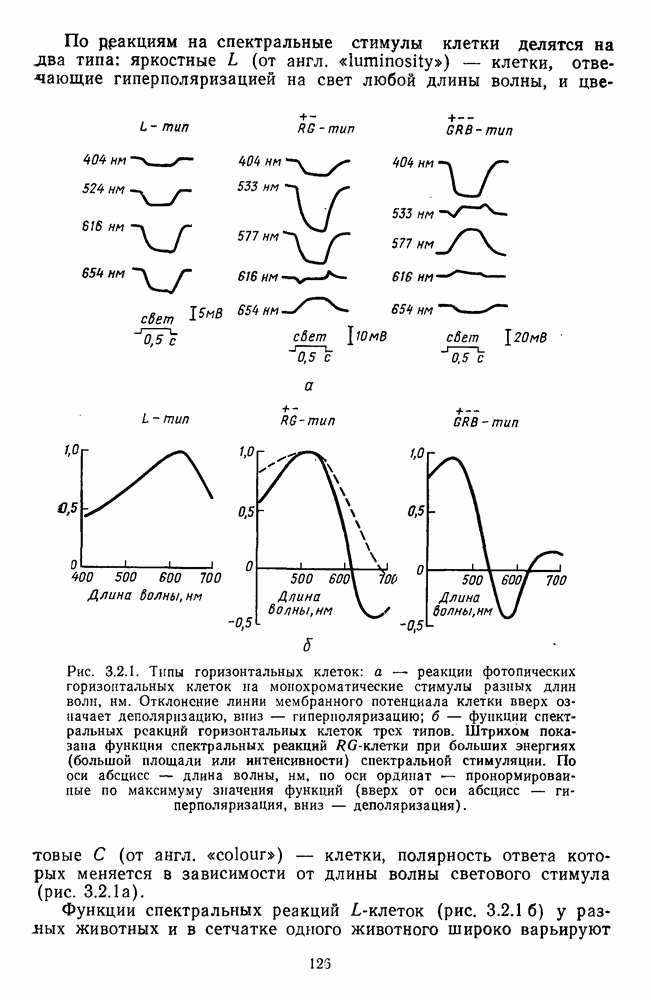
 обезьяны, полученные методом замены равиоярких цветовых стимулов в поведенческих опытах и по реакциям спектрально-оппонентных
обезьяны, полученные методом замены равиоярких цветовых стимулов в поведенческих опытах и по реакциям спектрально-оппонентных  нейронов
нейронов 

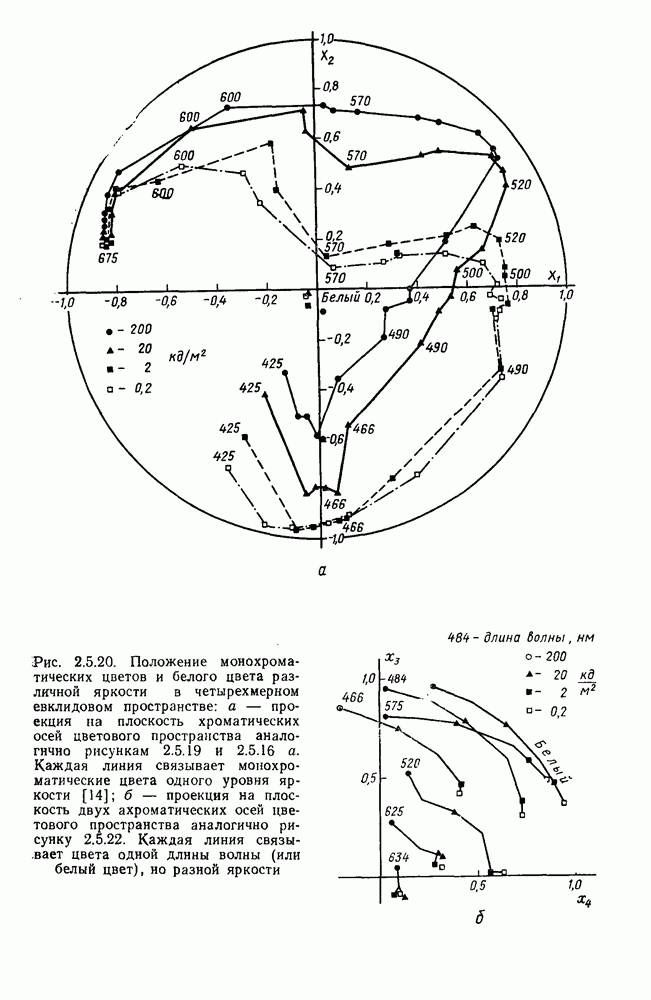
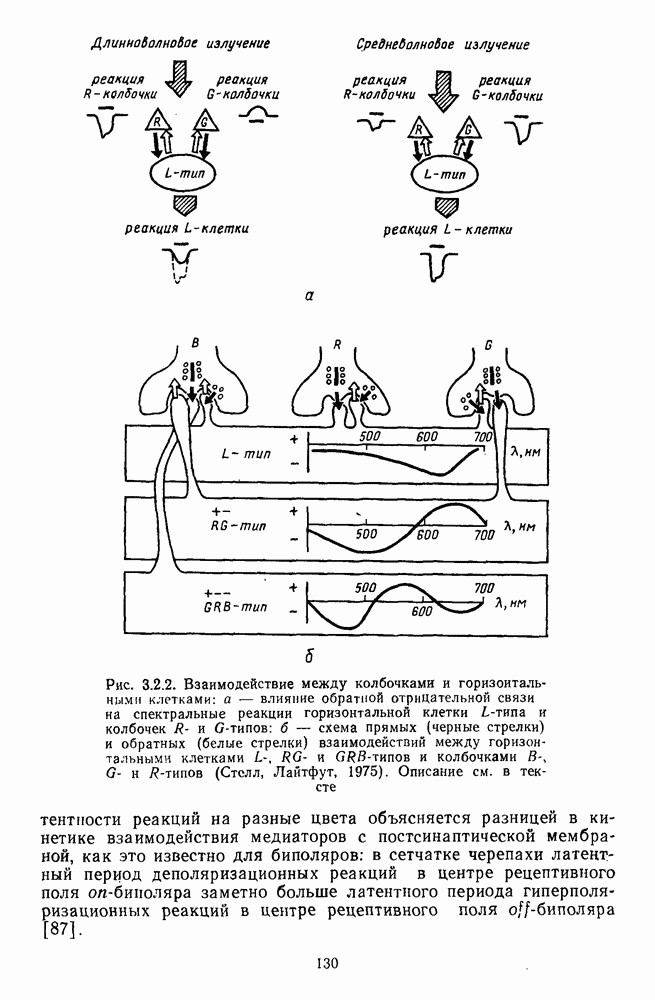
 -нейронов НКТ обезьяны на равнояркие цветовые стимулы разной насыщенности [208].
-нейронов НКТ обезьяны на равнояркие цветовые стимулы разной насыщенности [208].  — разные степени насыщенности цветовых стимулов (0 — белый цвет, 3 — монохроматический цвет)
— разные степени насыщенности цветовых стимулов (0 — белый цвет, 3 — монохроматический цвет)
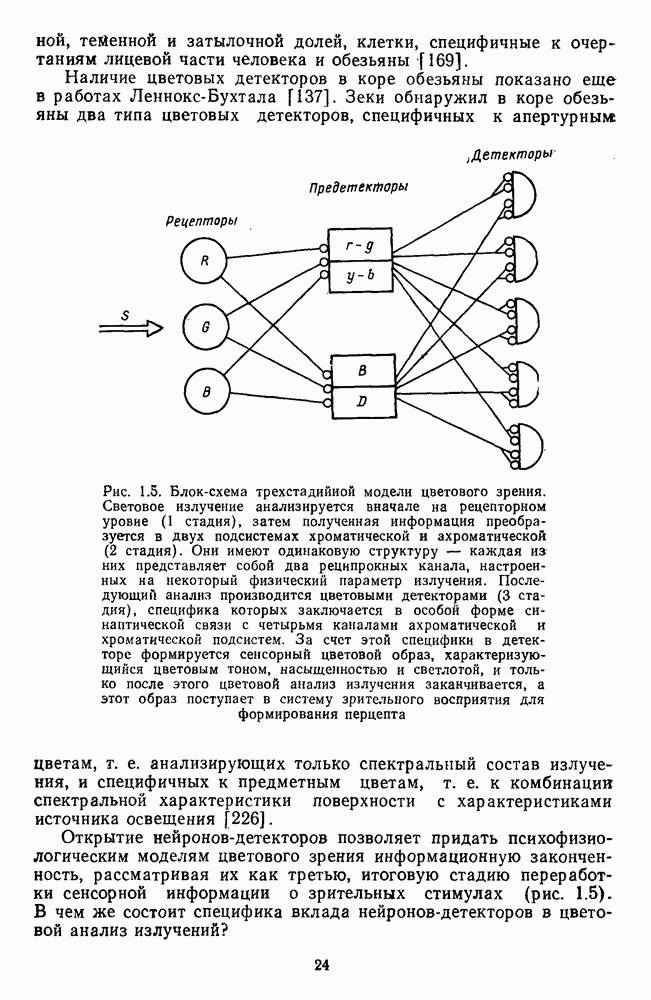
 [110]. Оказалось, что если в НКТ самок регистрируются как
[110]. Оказалось, что если в НКТ самок регистрируются как  так и
так и  -клетки, то в НКТ самцов
-клетки, то в НКТ самцов  -система цветооппонентных клеток практически полностью редуцирована. Де Валуа и Маррокко, меняя цвета по насыщенности, показали, что если направление реакций
-система цветооппонентных клеток практически полностью редуцирована. Де Валуа и Маррокко, меняя цвета по насыщенности, показали, что если направление реакций  и
и  -клеток (торможение или возбуждение) зависит от длины волны стимула, то амплитуда
-клеток (торможение или возбуждение) зависит от длины волны стимула, то амплитуда