Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
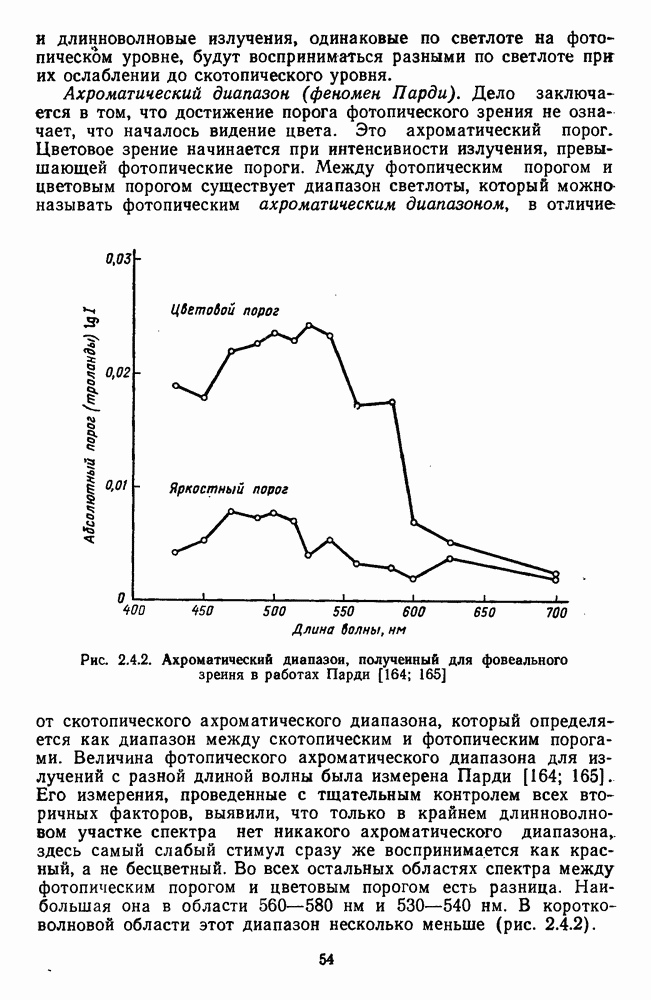
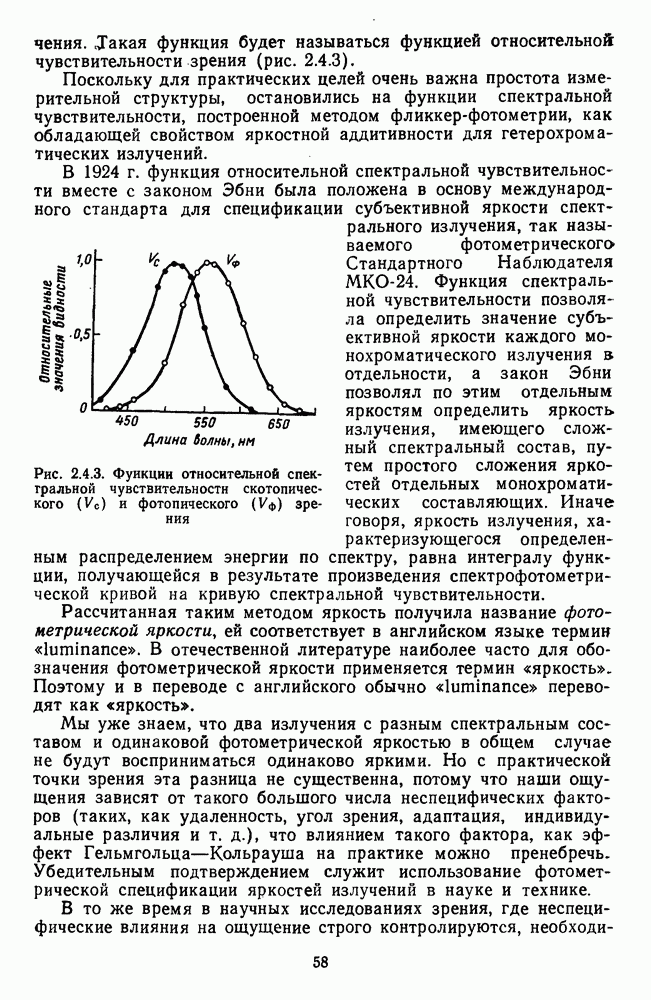
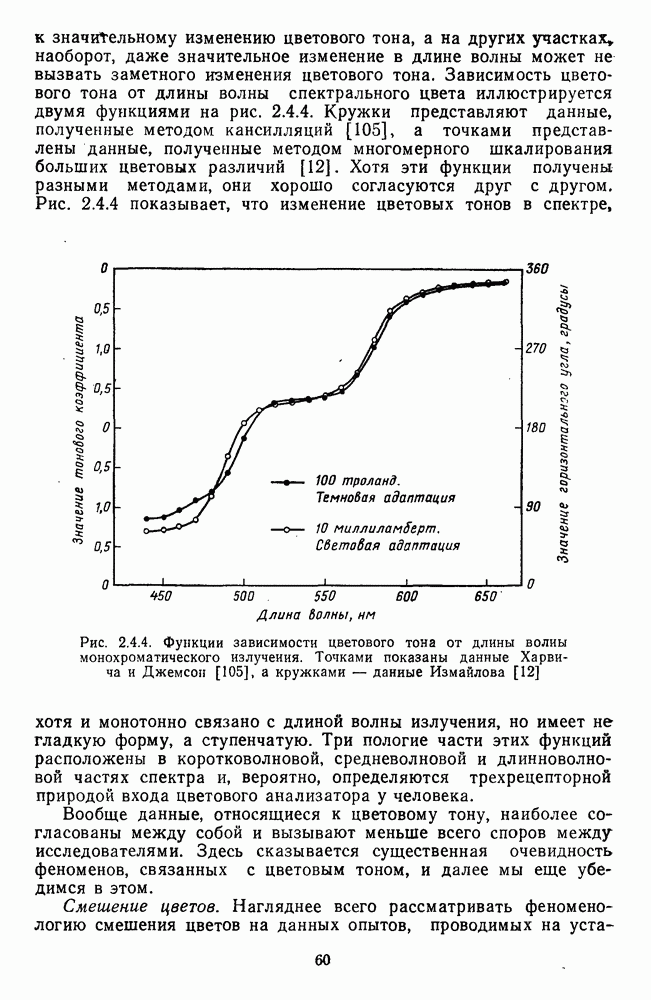
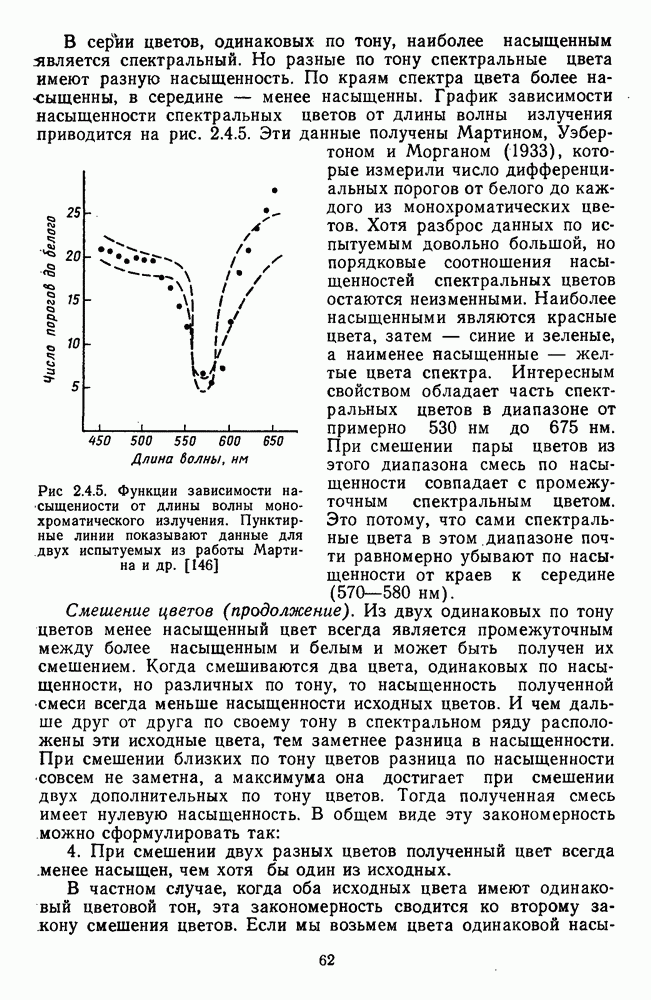
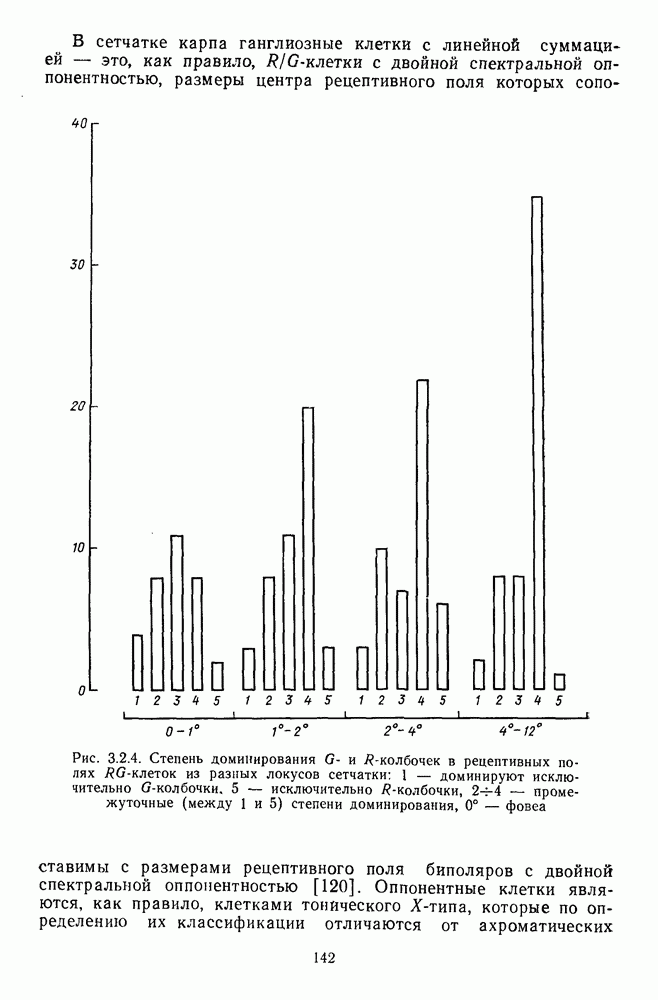
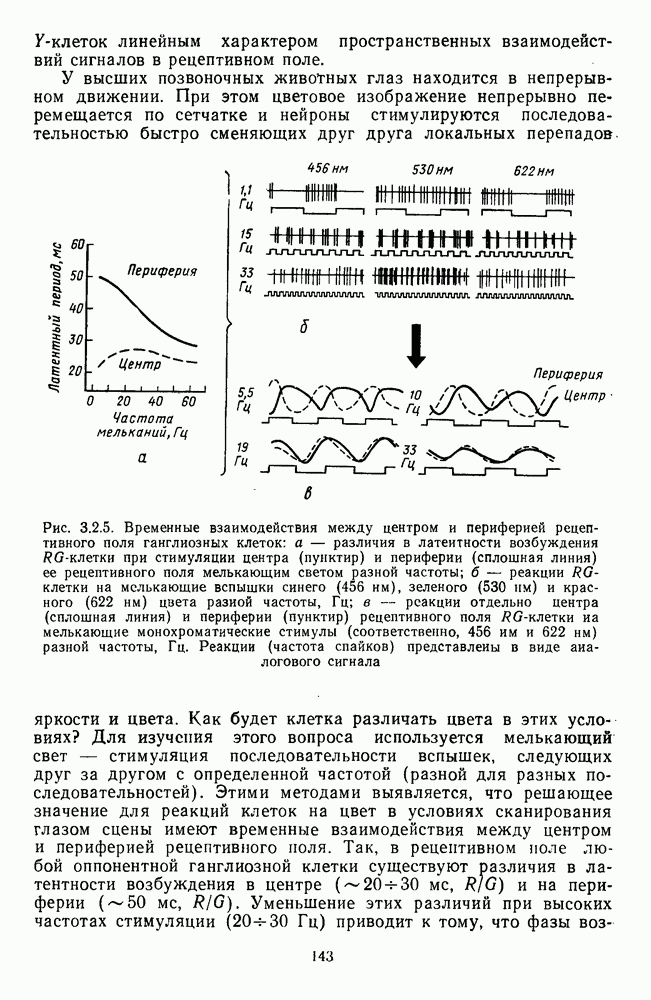
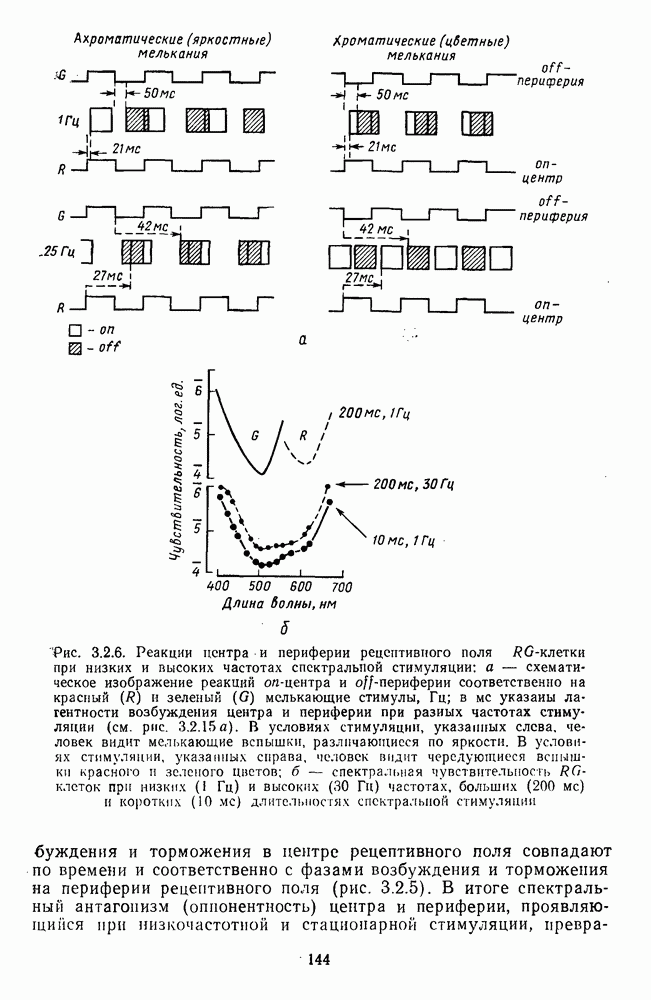
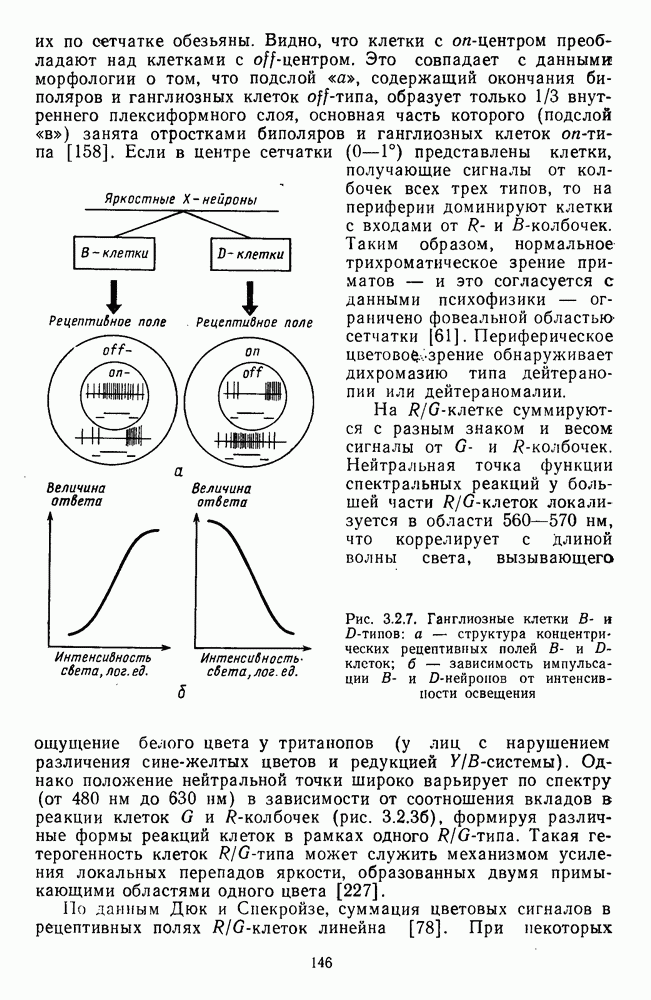
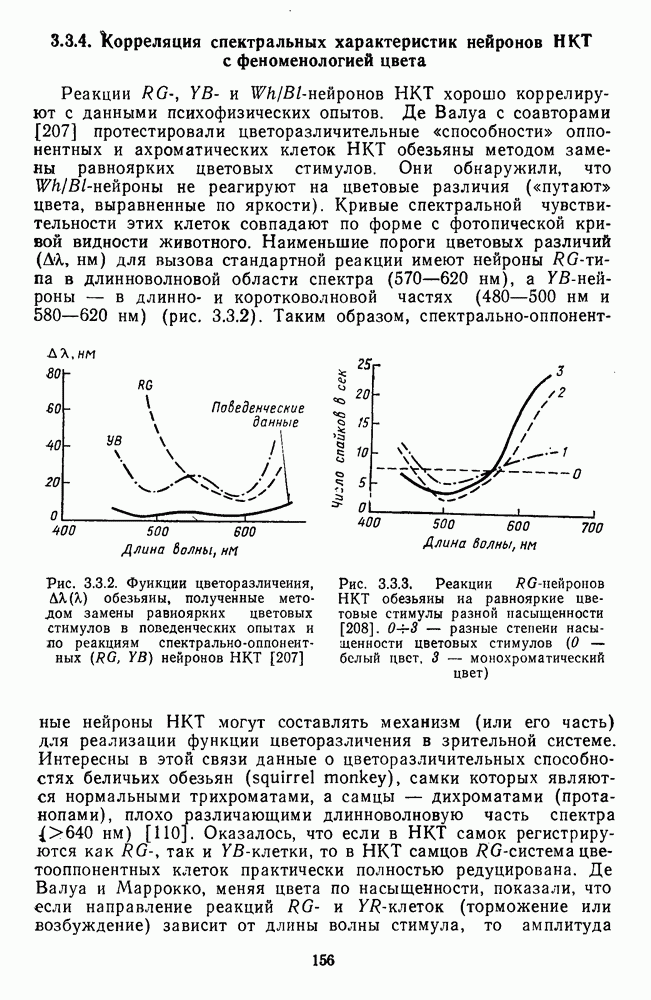
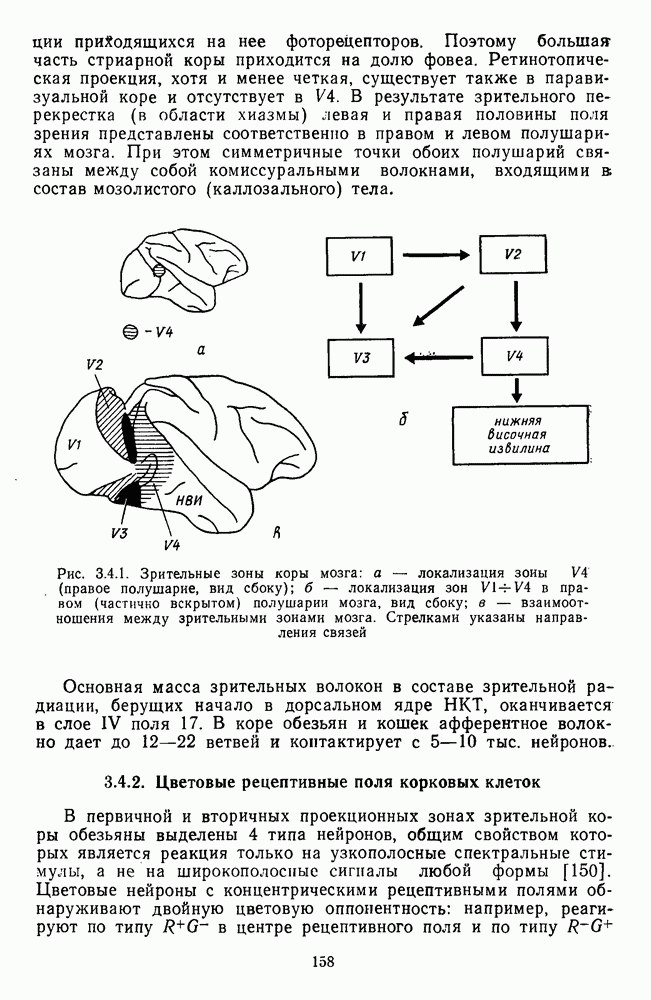
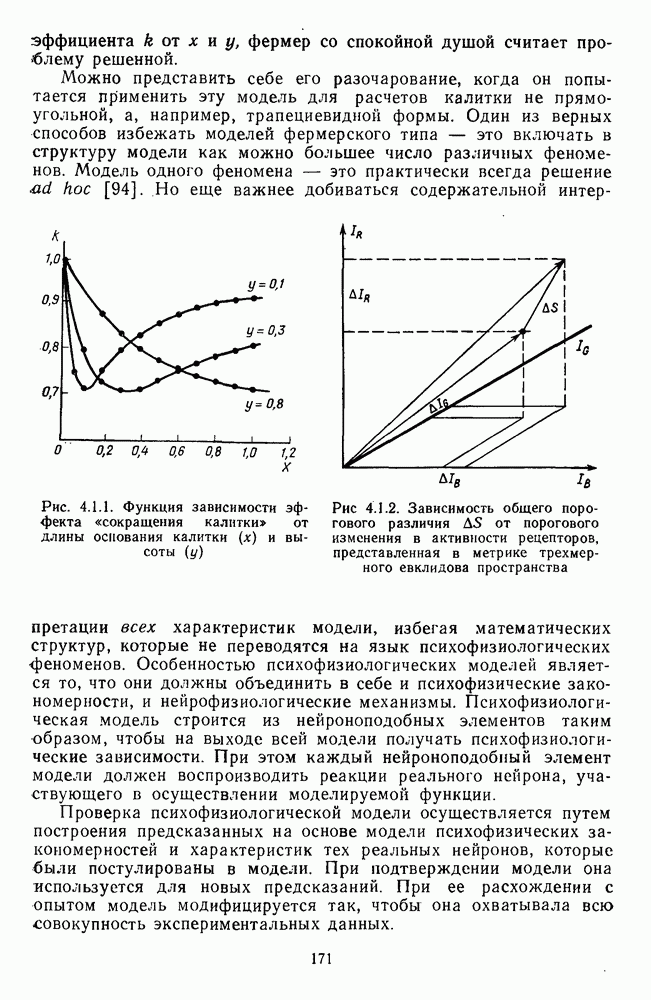
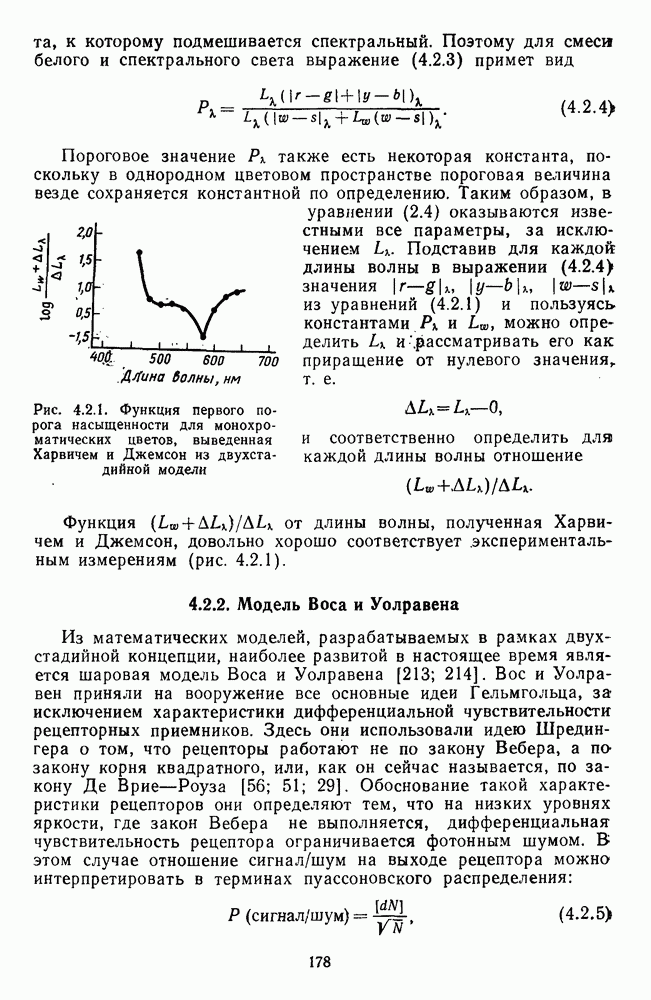
3.3.3. Особенности нейронов парво- и магноцеллюлярных слоевПо современным данным, нейроны НКТ, участвующие в кодировании цвета, локализуются в парвоцеллюлярных слоях [77; 74]. Здесь сконцентрированы тонические клетки спектральным составом излучения, но и яркостным контрастом стимула и фона. Интересно в этой связи, что максимально насыщенными цвета, кроме красных, воспринимаются не на темном фоне, а на светлом ахроматическом фоне [13]. Например, синий цвет наблюдается максимально насыщенным, тогда как его яркость составляет лишь Клетки Магноцеллюлярные слои также состоят из оппонентных нейронов с пространственно разнесенными в рецептивном поле входами от колбочек разного типа. Часть авторов связывают нейронные сети этих слоев с кодированием яркости [93], другая часть — с кодированием движущихся объектов [77]. В пользу последней точки зрения свидетельствуют такие свойства нейронов из крупноклеточных слоев, как выраженная чувствительность к высоким временным частотам, высокая контрастная чувствительность и большие рецептивные поля. Функциональные различия между нейронами из парво- и магноцеллюлярных слоев во многом определяются организацией их афферентных входов. В НКТ обезьяны ретинальные волокна от фазических и тонических ганглиозных клеток оканчиваются соответственно в крупно- и мелкоклеточных слоях. По свойствам реакций и строению РП фазические и тонические нейроны у обезьяны напоминают У- и Х-нейроны в зрительной системе кошки, у которой аксоны этих клеток также оканчиваются в разных слоях НКТ. Из различий в детекторных свойствах Анатомическая сегрегация нейронов с разными функциональными свойствами наблюдается уже в сетчатке, где отростки биполяров и ганглиозных клеток
|
 |
Оглавление
|

























































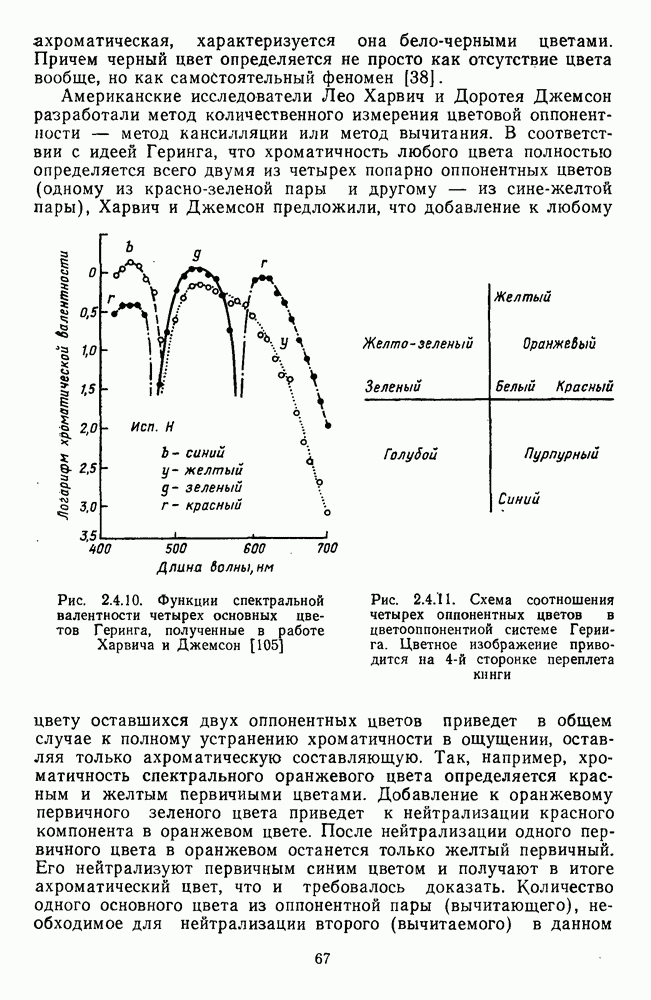


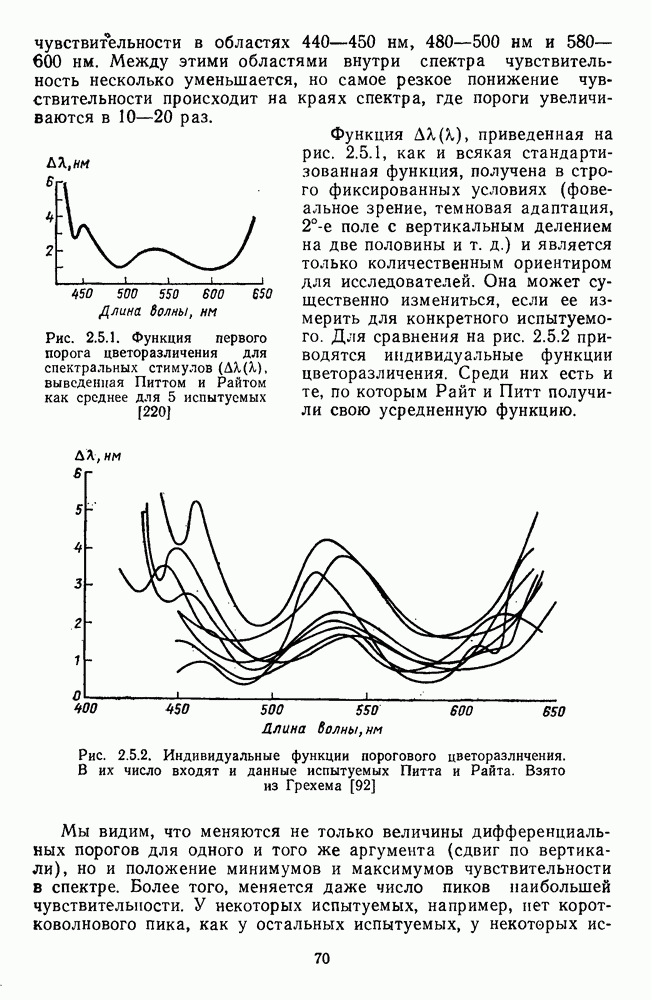
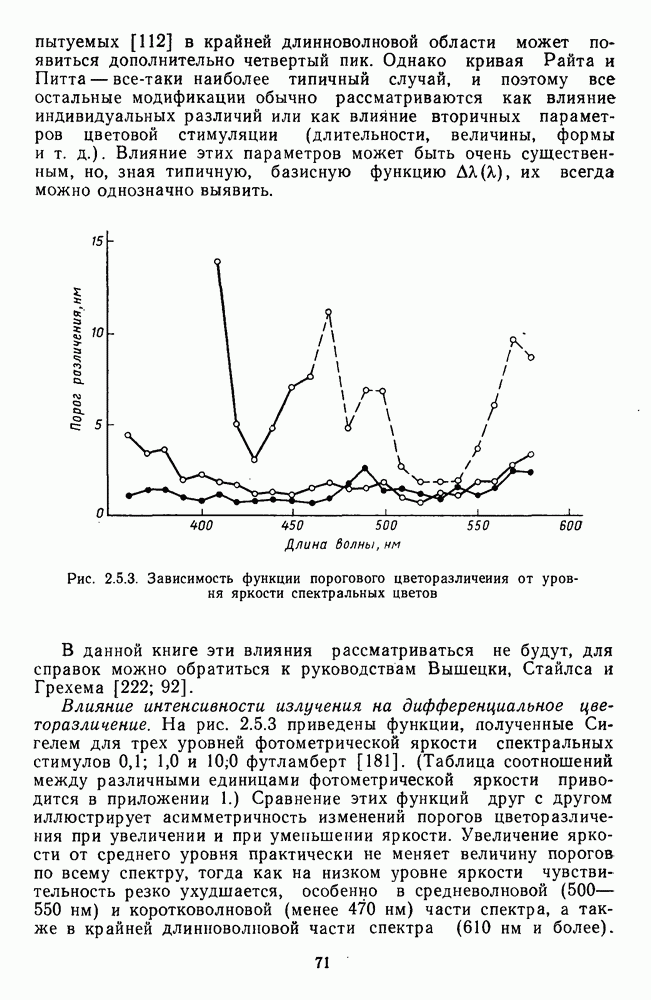
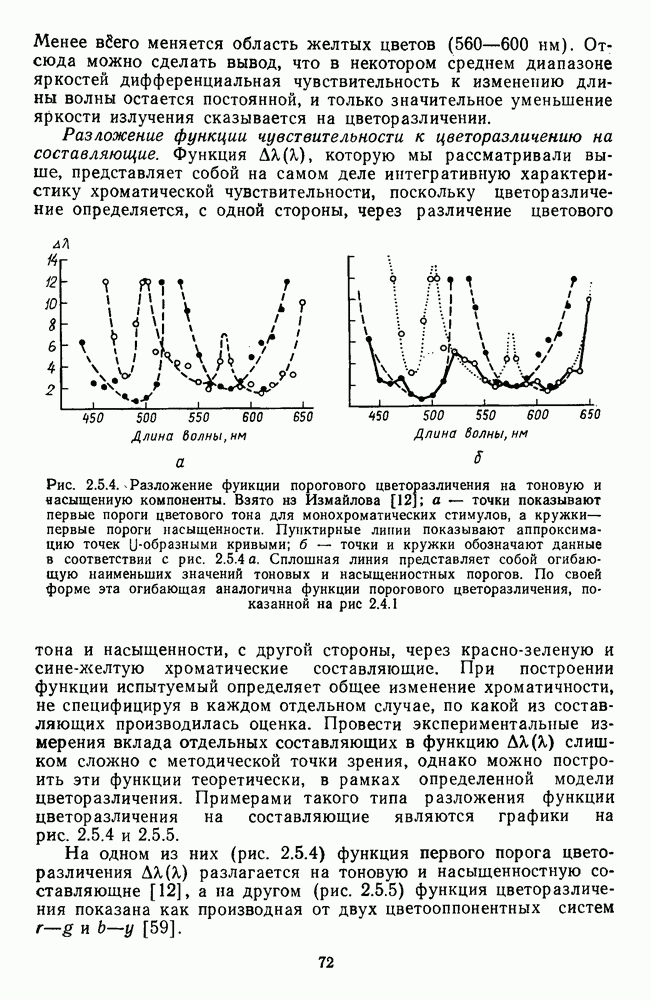







































































































 и
и  -типов, для которых характерна линейная суммация колбочковых сигналов по площади рецептивного поля. Нелинейность возникает, как и у оппонентных ганглиозных клеток сетчатки, лишь в специальных условиях адаптации. Среди нейронов этих слоев выделяют две большие группы: широкополосные
-типов, для которых характерна линейная суммация колбочковых сигналов по площади рецептивного поля. Нелинейность возникает, как и у оппонентных ганглиозных клеток сетчатки, лишь в специальных условиях адаптации. Среди нейронов этих слоев выделяют две большие группы: широкополосные  -клетки (от англ. «widp band») и узкополосные
-клетки (от англ. «widp band») и узкополосные  -клетки (от англ. «narrow band»).
-клетки (от англ. «narrow band»).  -клетки избирательно возбуждаются в узком диапазоне длин волн, как правило из коротко- или длинноволновой части спектра, и тормозятся на остальные.
-клетки избирательно возбуждаются в узком диапазоне длин волн, как правило из коротко- или длинноволновой части спектра, и тормозятся на остальные.  -клетки реагируют возбуждением в более широком диапазоне длин волн и торможением — в небольшой области спектра. При стимуляции белым светом
-клетки реагируют возбуждением в более широком диапазоне длин волн и торможением — в небольшой области спектра. При стимуляции белым светом  -нейроны ведут себя как ганглиозные клетки В-типа, а
-нейроны ведут себя как ганглиозные клетки В-типа, а  -нейропы — как клетки
-нейропы — как клетки  -типа. Оказалось, что реакции
-типа. Оказалось, что реакции  -клеток по-разному зависят от отношения яркости стимула, освещающего центр рецептивного поля, к яркости фона (ахроматического или хроматического): при яркостях центрального стимула, больших яркости фона, активируются
-клеток по-разному зависят от отношения яркости стимула, освещающего центр рецептивного поля, к яркости фона (ахроматического или хроматического): при яркостях центрального стимула, больших яркости фона, активируются  -клетки, при яркостях, меньших яркости фонового окружения, —
-клетки, при яркостях, меньших яркости фонового окружения, —  -клетки. Таким образом,
-клетки. Таким образом,  -нейроны реагируют на излучение, которое ярче фона, а
-нейроны реагируют на излучение, которое ярче фона, а  -нейроны — когда стимулы темнее фона. Эти нейрофизиологические данные хорошо коррелируют с феноменологией зрения, когда видение цвета определяется не только
-нейроны — когда стимулы темнее фона. Эти нейрофизиологические данные хорошо коррелируют с феноменологией зрения, когда видение цвета определяется не только
 от яркости фона, т. е. в условиях, оптимальных для активации
от яркости фона, т. е. в условиях, оптимальных для активации  -нейронов [227].
-нейронов [227].
 и Шьтипов не исчерпывают всей популяции нейронов в мелкоклеточных слоях, включающих также
и Шьтипов не исчерпывают всей популяции нейронов в мелкоклеточных слоях, включающих также  -ней-роны и нейроны, имеющие промежуточные между
-ней-роны и нейроны, имеющие промежуточные между  и
и  -типами свойства.
-типами свойства.
 -нейронов вытекают функциональные различия образуемых ими слоев НКТ: тонические нейроны мелкоклеточных слоев участвуют в анализе интенсивности и спектрального состава излучения в центре поля зрения, а фазические нейроны крупноклеточных слоев — резких перепадов в освещенности (например, при перемещении объекта) на периферии поля зрения. По-видимому, фазические ганглиозные клетки, нейроны верхнего двухолмия и крупноклеточных слоев НКТ включены в систему контроля быстрых саккадических движений глаз.
-нейронов вытекают функциональные различия образуемых ими слоев НКТ: тонические нейроны мелкоклеточных слоев участвуют в анализе интенсивности и спектрального состава излучения в центре поля зрения, а фазические нейроны крупноклеточных слоев — резких перепадов в освещенности (например, при перемещении объекта) на периферии поля зрения. По-видимому, фазические ганглиозные клетки, нейроны верхнего двухолмия и крупноклеточных слоев НКТ включены в систему контроля быстрых саккадических движений глаз.
 и
и  -типов локализуются в разных подслоях внутреннего синаптического слоя. Такое «анатомическое обособление» нейронных систем, образующих разные каналы передачи информации, является общим принципом в построении анализаторных структур и наиболее ярко выражено в колончатой организации коры.
-типов локализуются в разных подслоях внутреннего синаптического слоя. Такое «анатомическое обособление» нейронных систем, образующих разные каналы передачи информации, является общим принципом в построении анализаторных структур и наиболее ярко выражено в колончатой организации коры.