Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
3.4.3. Детекторы спектральных излученийПервые сообщения о цветовых нейронах детекторного типа сделаны Гранитом при исследовании аксонов зрительного нерва лягушки [6]. Гранит выявил, что аксоны ганглиозных клеток избирательно активировались на стимулы с узким диапазоном длин волн в разных участках спектра («модуляторы»). Вслед за Гранитом Де Валуа описал избирательные к цвету нейроны-модуляторы в НКТ макаки [209]. Однако в последующих работах Де Валуа показал, что эти нейроны являются на самом деле оппонентными и более широкополосными: низкий уровень спонтанной активности и высокий уровень шума в предыдущих опытах не позволили выделить тормозную периферию в РП «модуляторов». Сходные причины могли повлиять и на результаты Гранита. К тому же Гранит работал пороговыми методами, а в этих условиях форма функции спектральных реакций может сильно варьировать в зависимости от небольших колебаний в уровне адаптации. В то же время данные Гранита относятся к низшим позвоночным, у которых нейроны сетчатки обладают ярко выраженными детекторными свойствами [37]. Более определенные данные о детекторах цвета получены при изучении стриарной коры обезьяны, где цветоселективные нейроны не меняли своей спектральной избирательности с изменением условий световой адаптации и интенсивности стимуляции [48; 137]. В работе Ваутин и Доу обнаружены корковые нейроны поля 17, избирательно возбуждающиеся в узком диапазоне длин волн с максимумом реакций в области 450 нм, 506 нм, 577 и 656 нм (у каждой клетки свой максимум) [211]. Эти максимумы соответствуют положению в спектре «основных цветов» у человека и обезьяны — «синего» (450 нм), «зеленого» (506 нм),, «желтого» (577 нм) и «красного» (656 нм). Все эти узкоселективные нейроны имели рецептивное поле с оппонентной к центру периферией. По-разному они реагировали на белый свет: с пиками на 656 нм и 506 нм — слабо, на 450 нм и 577 нм — средне. В этой же зоне коры обнаружены клетки, реагирующие на широкий спектр монохроматических излучений с пиками возбуждения при 506 нм и 656 нм. Эти нейроны напоминают широкополосные Между цветочувствительными клетками поля 17 и полей 18—19 имеются определенные различия. Во-первых, в паравизуальной коре вообще больше нейронов со «сложными» и «сверхсложными» РП и соответственно больше и аналогичных цветовых нейронов. Во-вторых, клетки поля 18 с двойной цветовой оппонентностью и концентрическими РП реагируют на цветные пятна определенных размеров так же, как и «сложные» клетки на элементы контура, — независимо от положения стимула в РП. Часть этих клеток реагируют на световые пятна определенных размеров независимо от длины волны излучения и в пределах участка поля зрения, превышающего оптимальные размеры светового стимула. В соответствии с такими свойствами данные нейроны были отнесены к клеткам со «сложными РП без ориентационной избирательности» («complex-unoriented») [107]. Они отличаются от обычных концентрических с центр-периферической организацией РП так же, как «простые» клетки отличаются от «сложных». Усложнение цветовых свойств нейронов в полях 18—19 может быть результатом того, что информация, выделенная нейронами поля; 17, проходит в них дальнейшую обработку.
|
 |
Оглавление
|












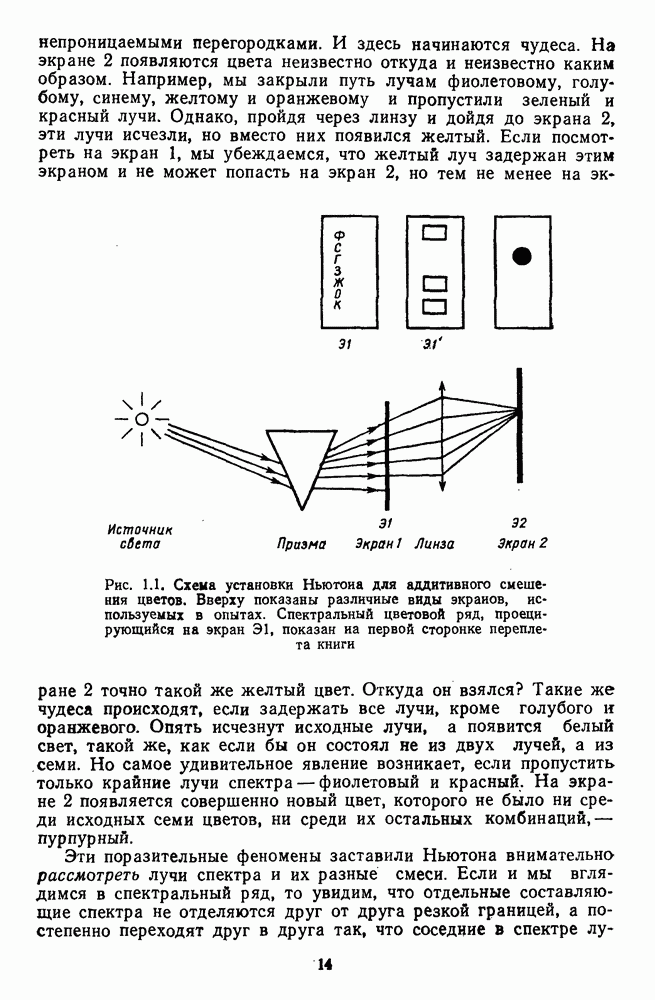






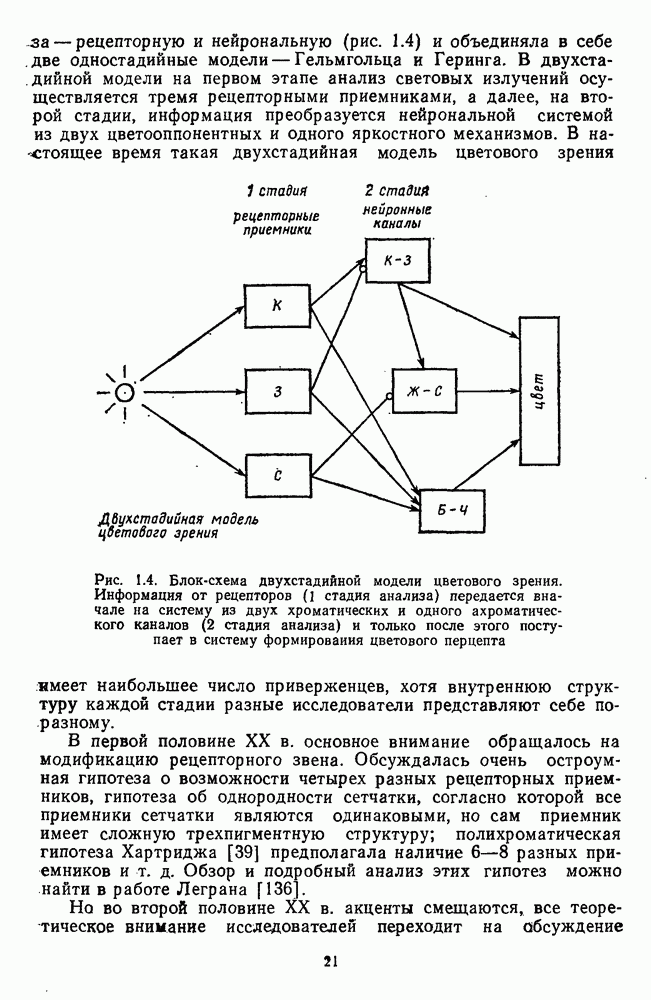


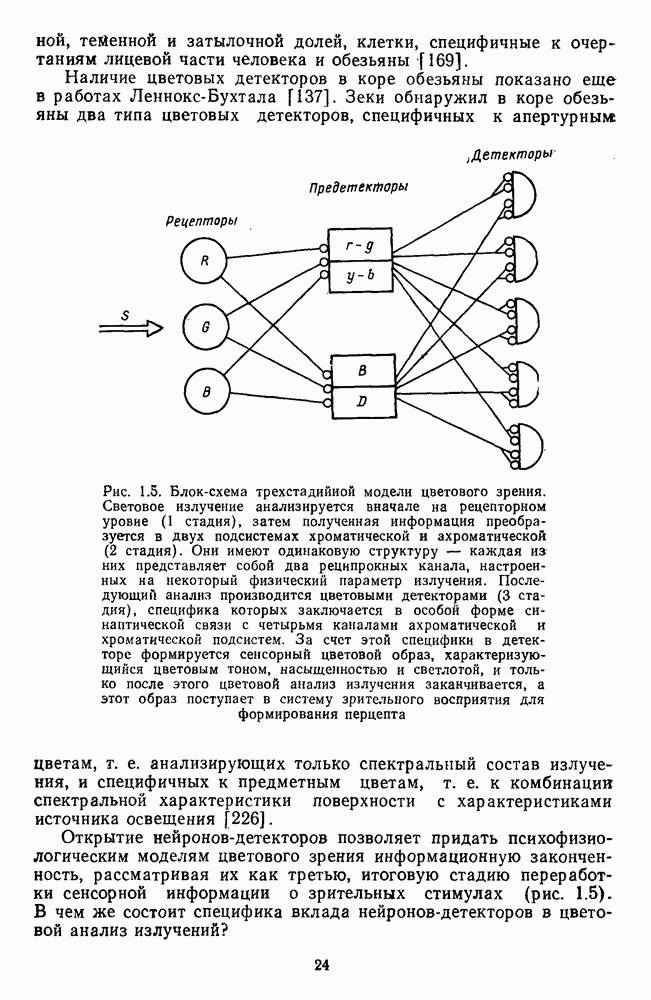



























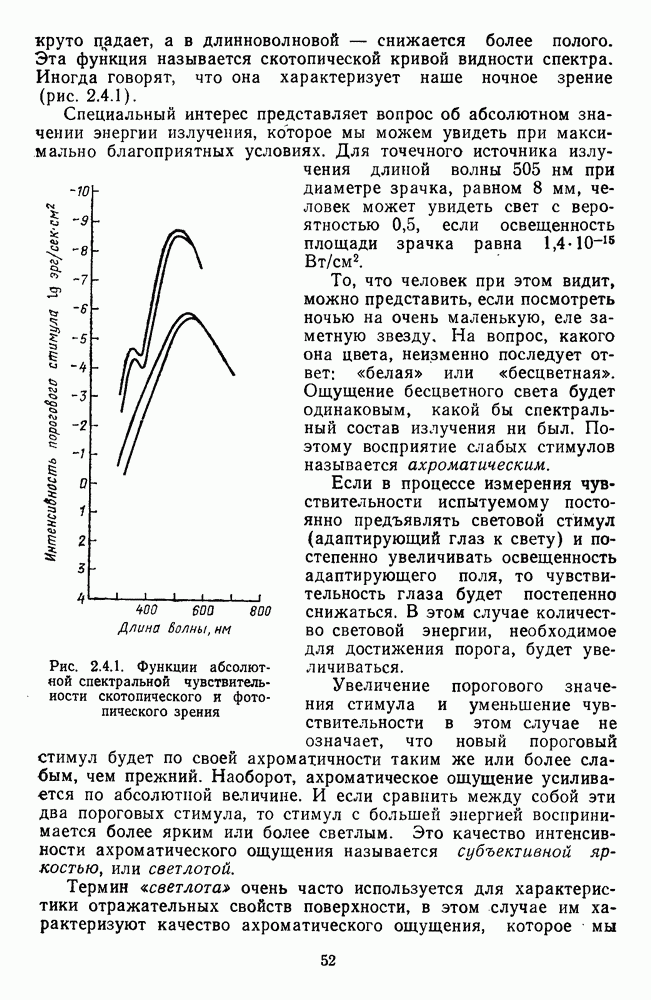










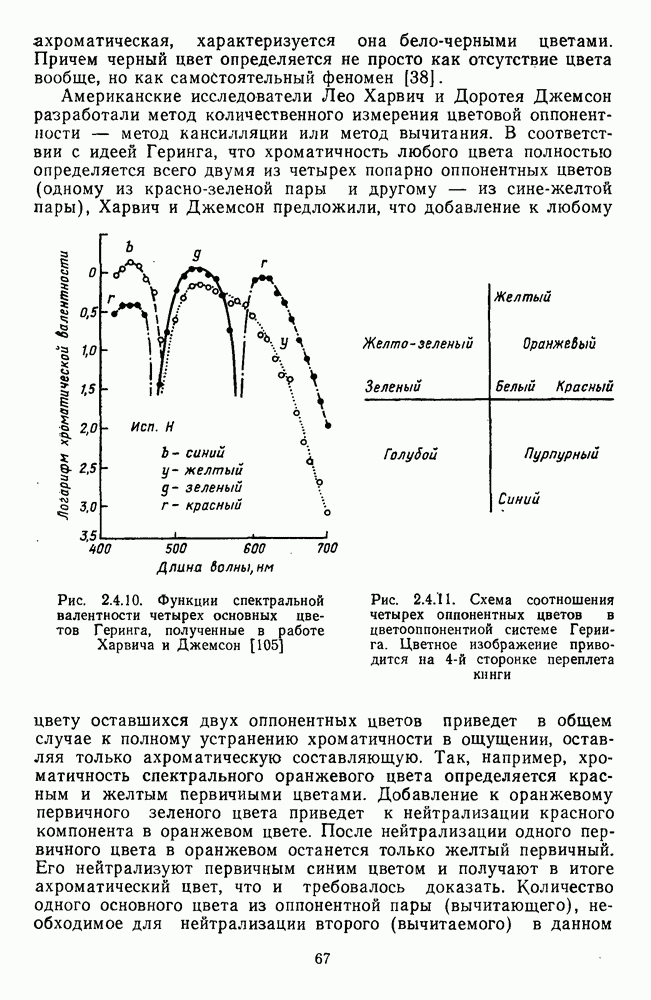


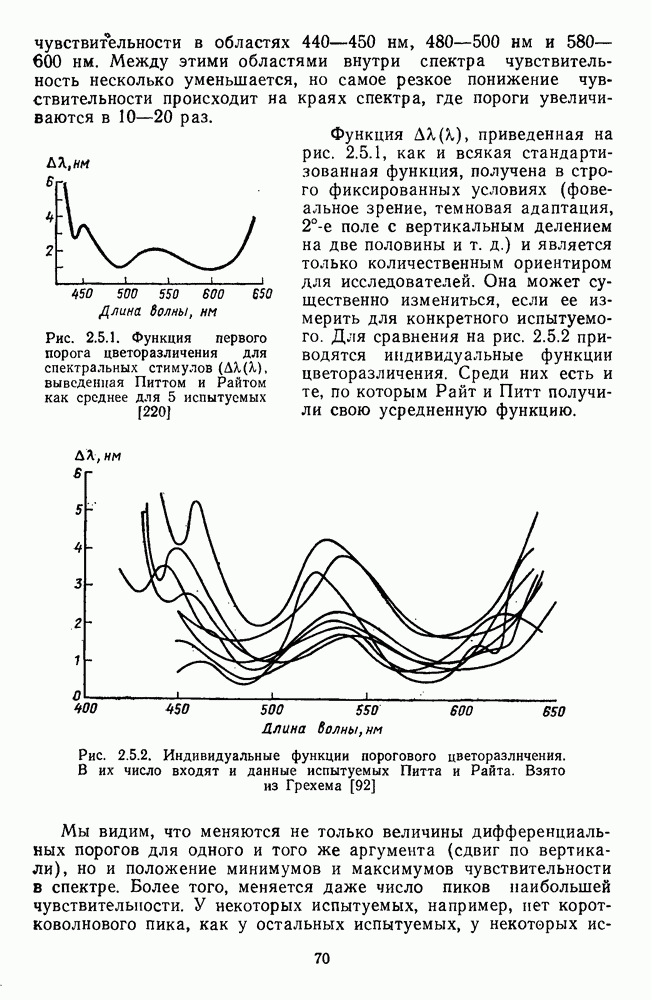
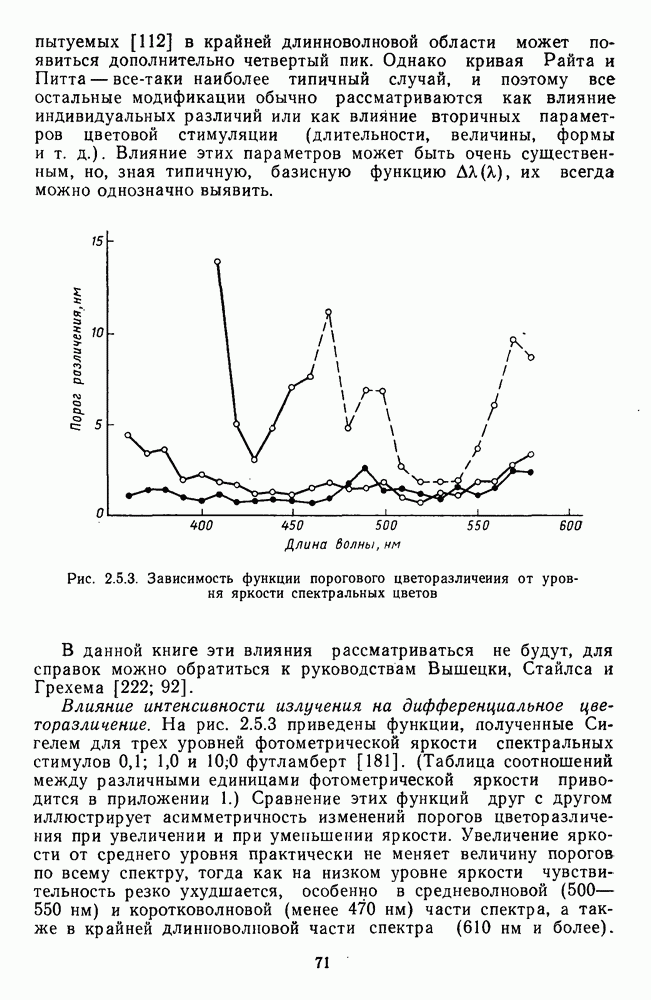
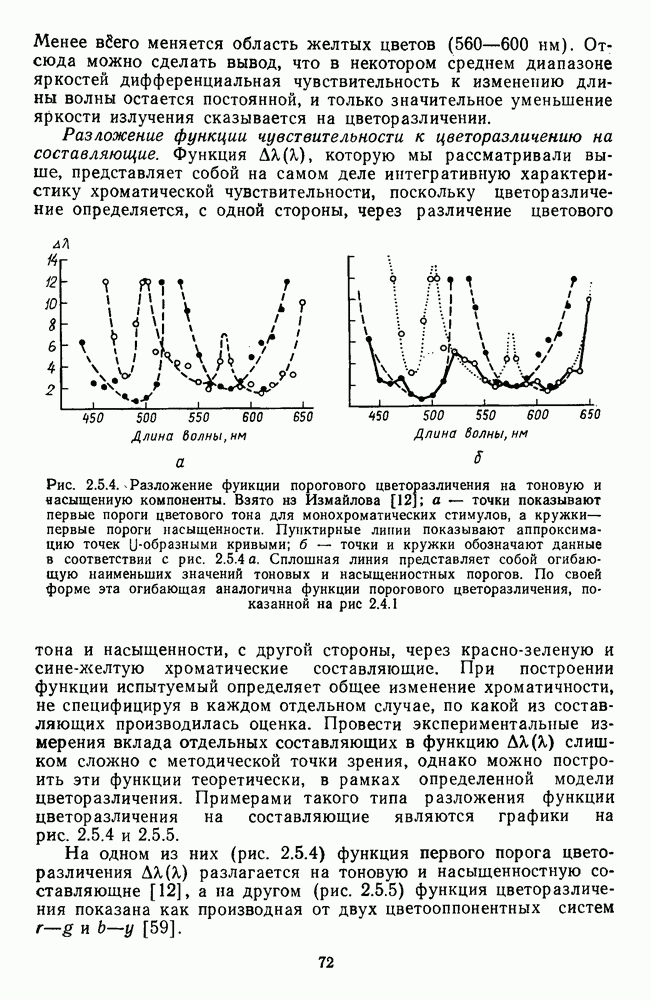
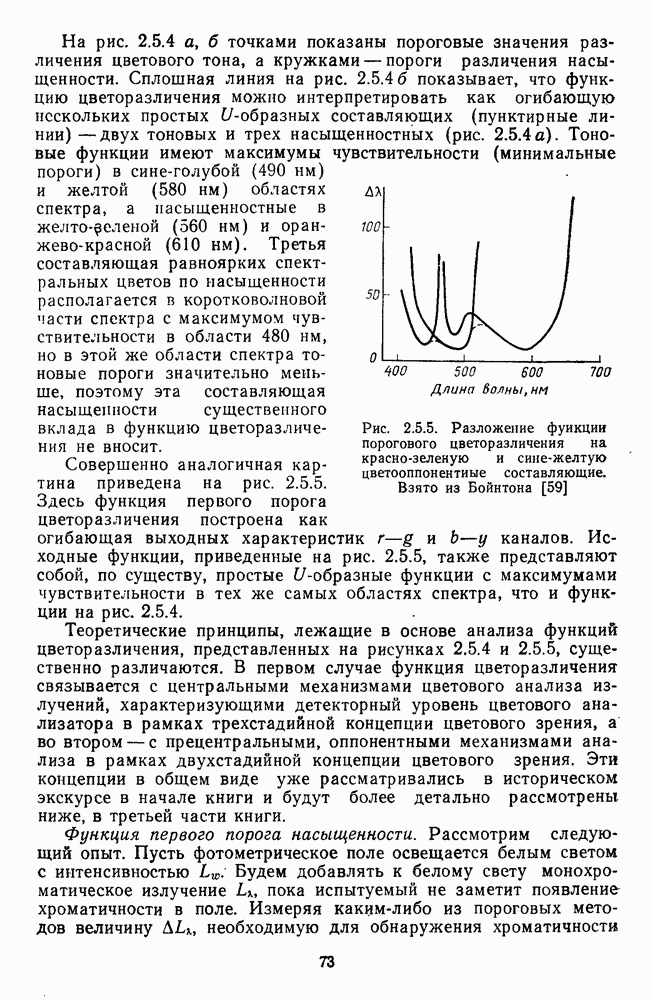





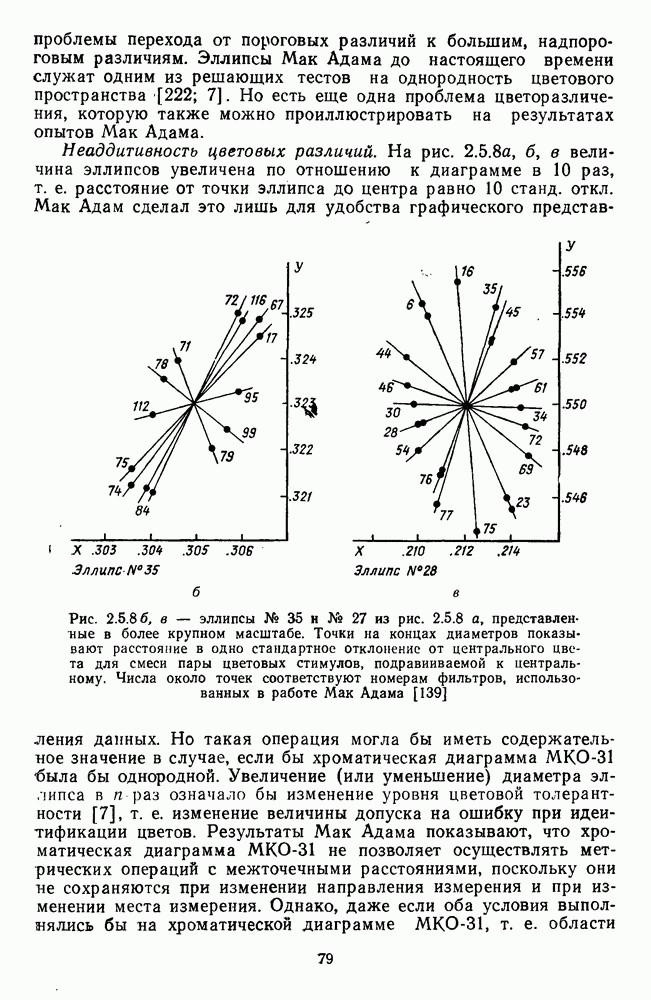





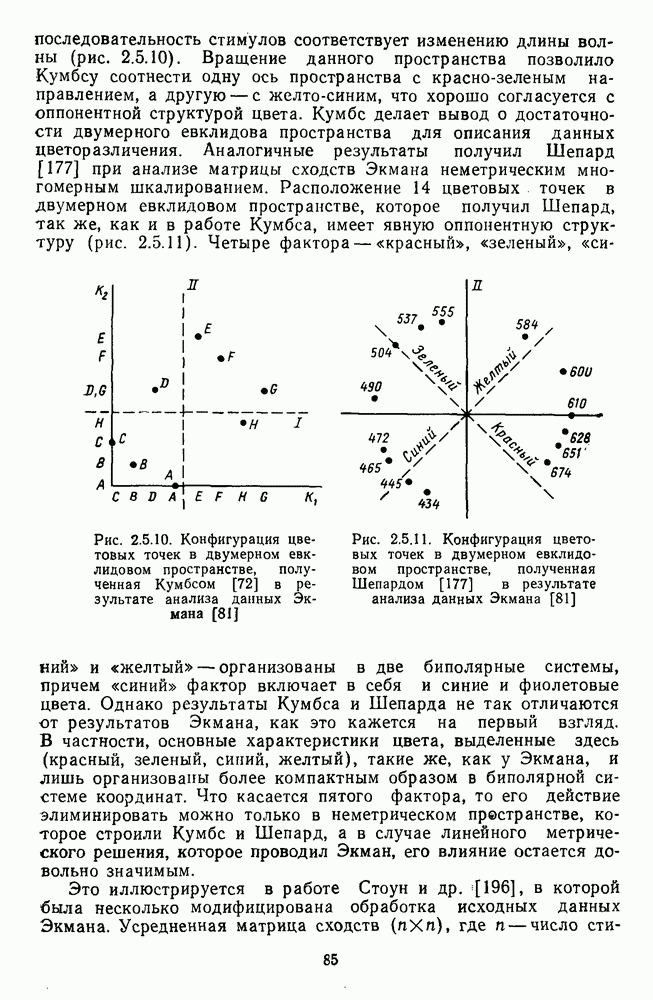
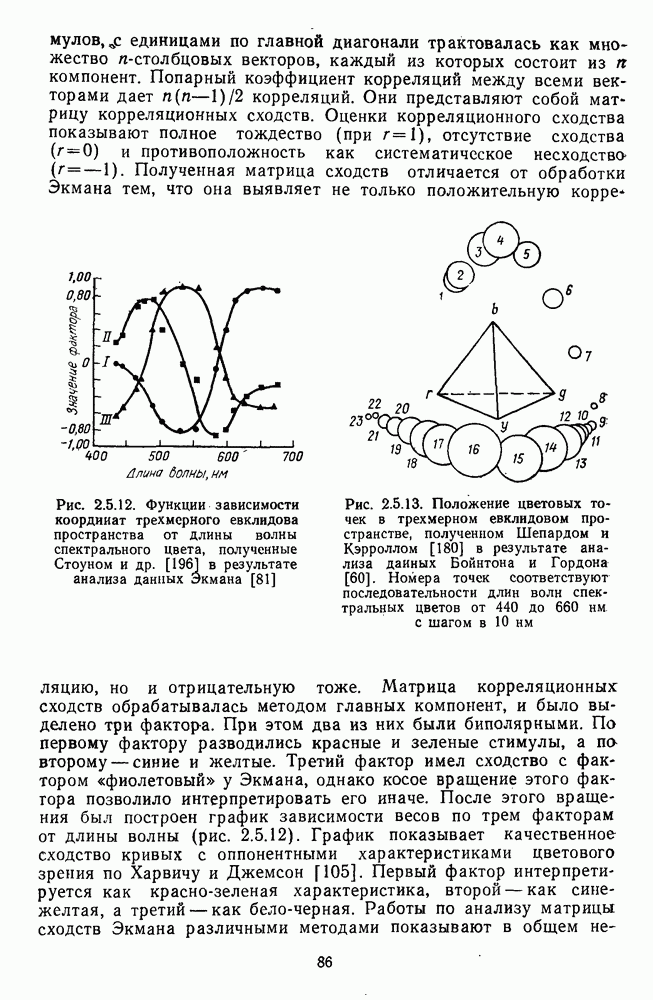


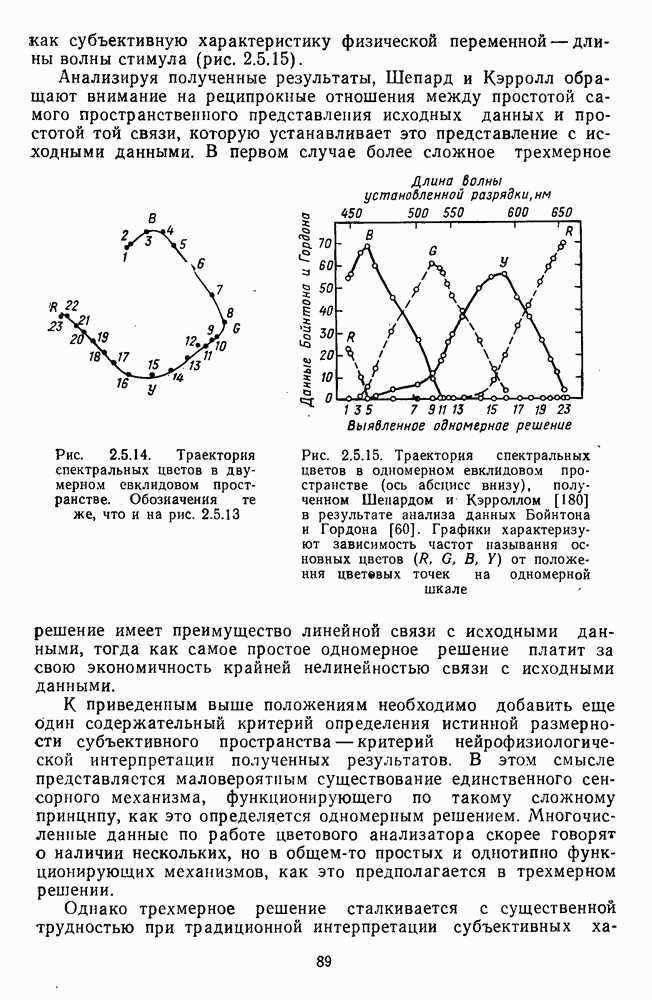




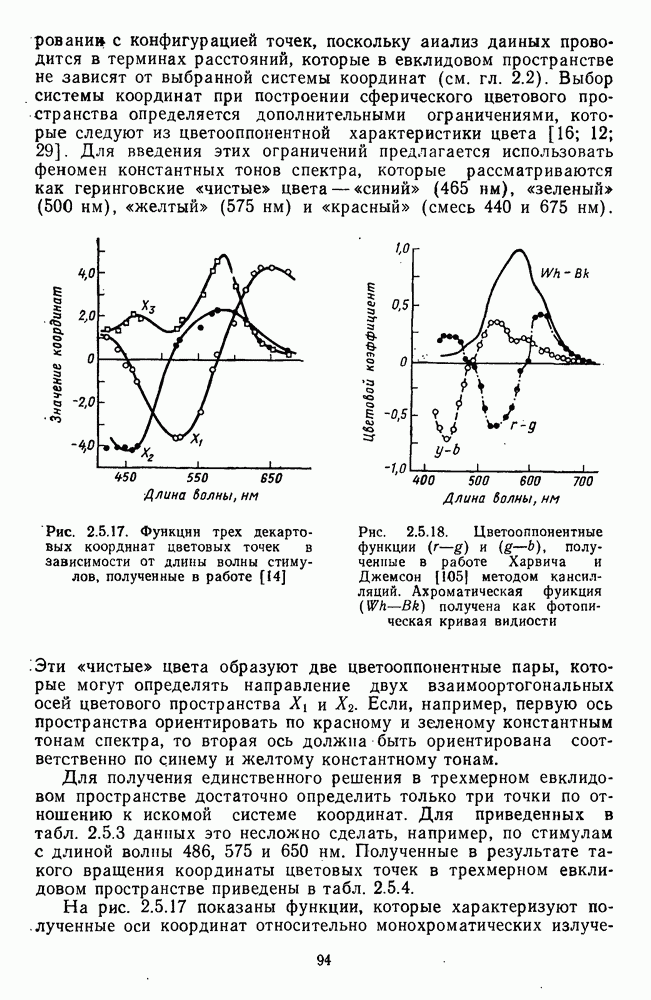

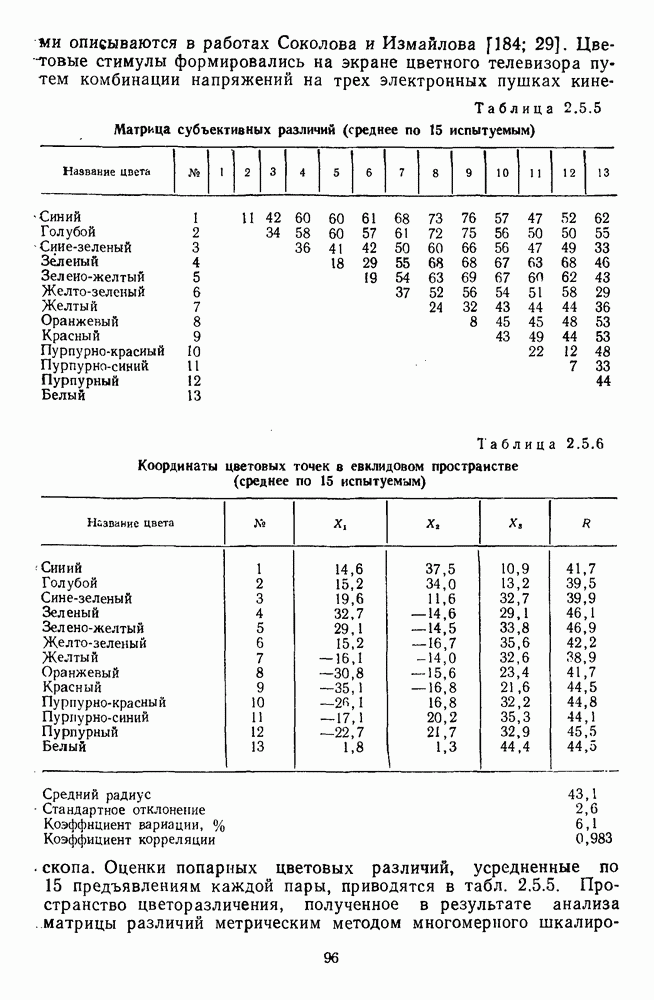


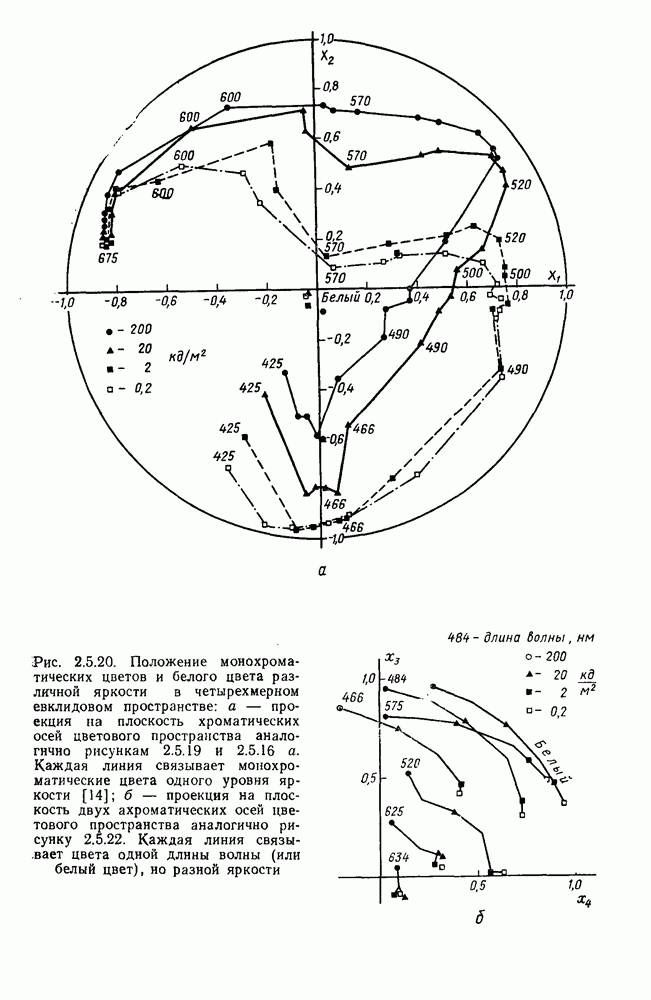







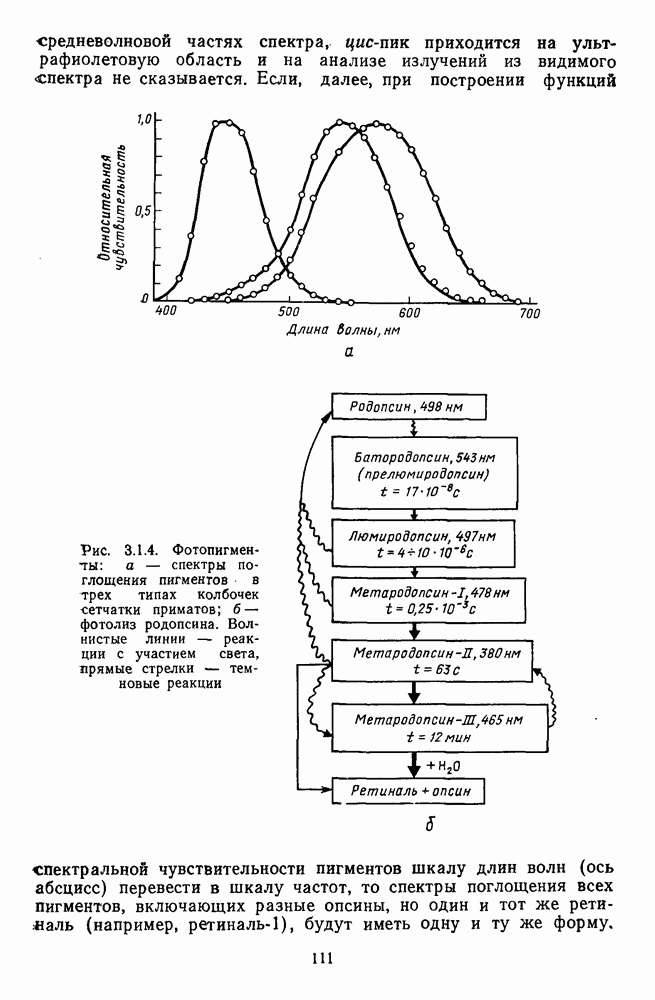














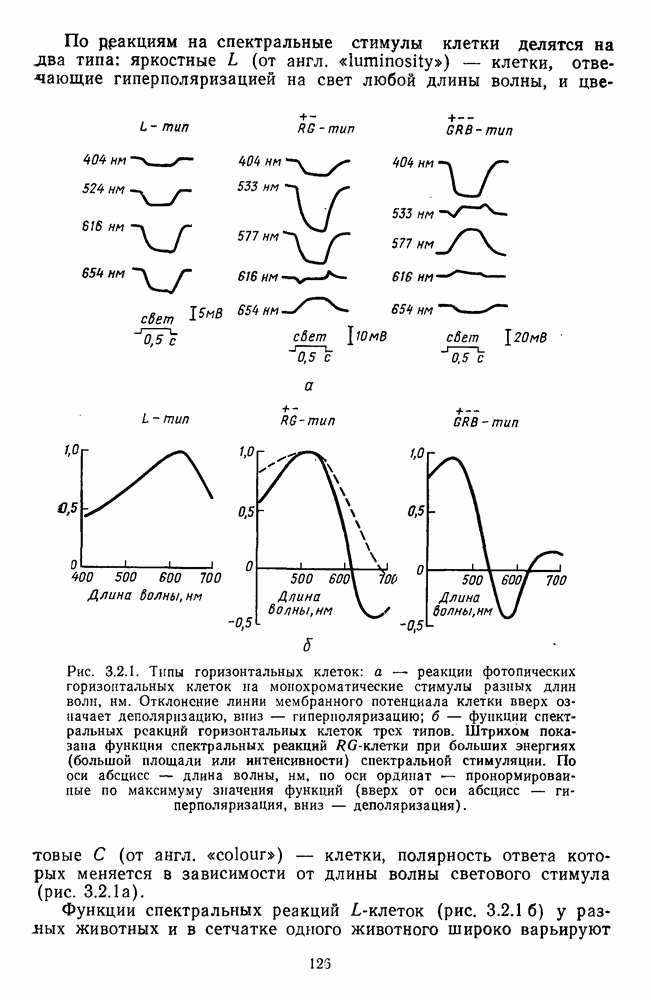



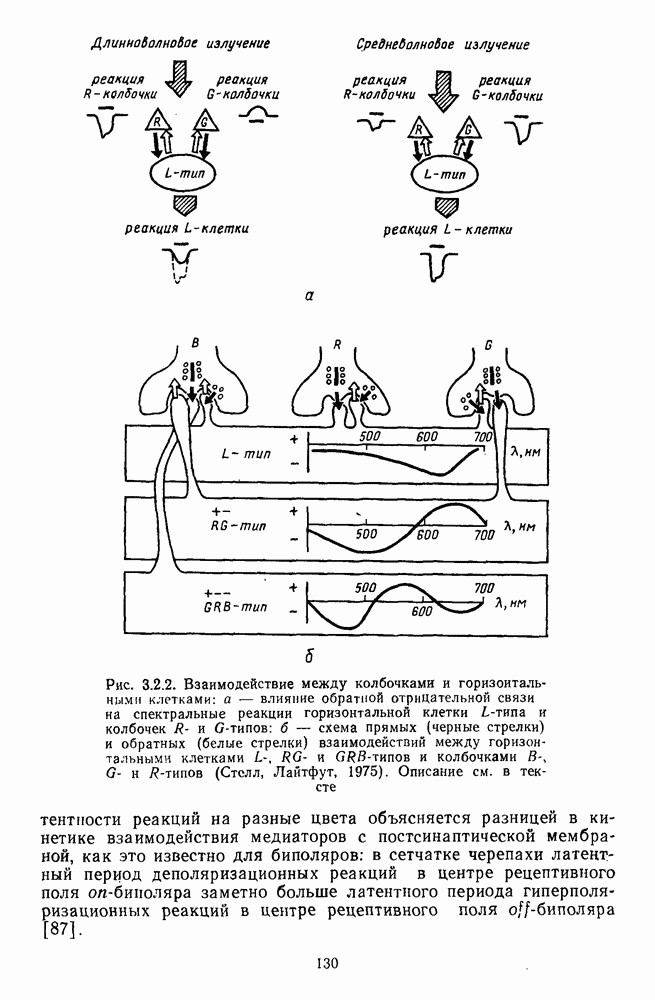









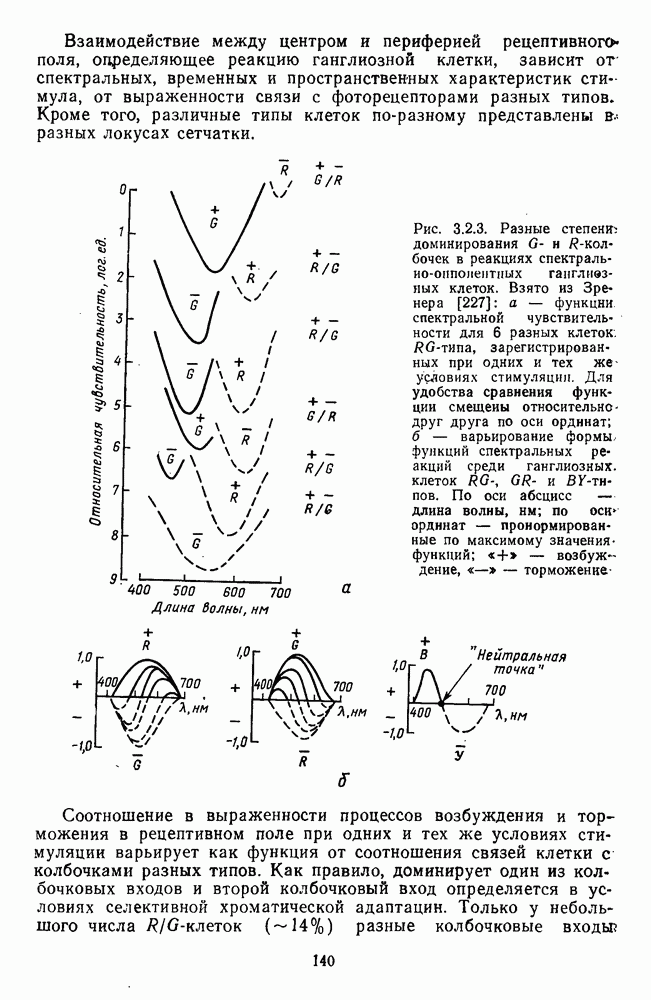

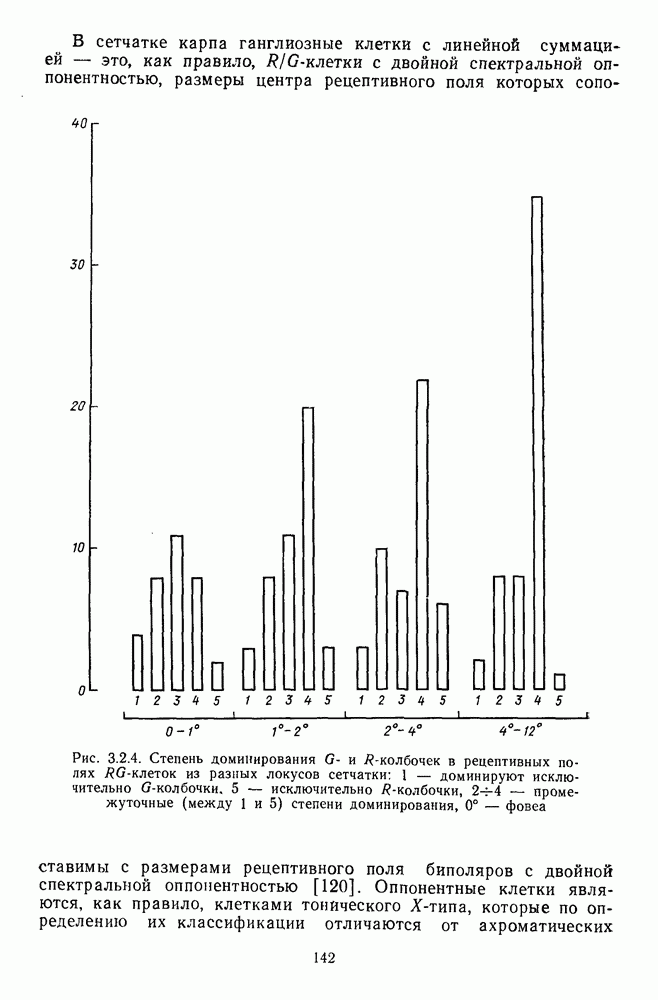
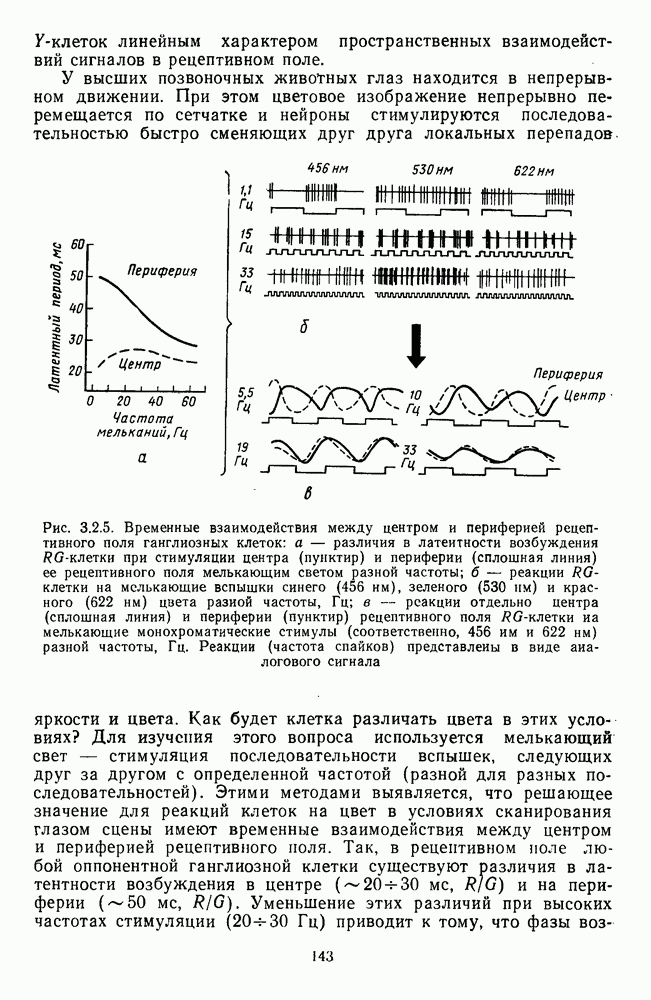
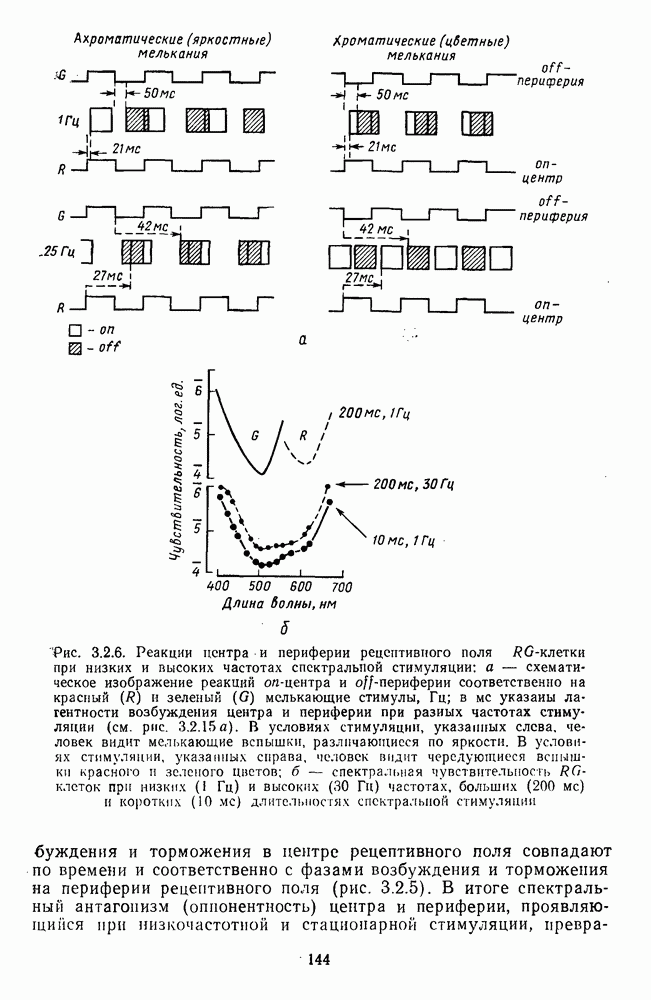

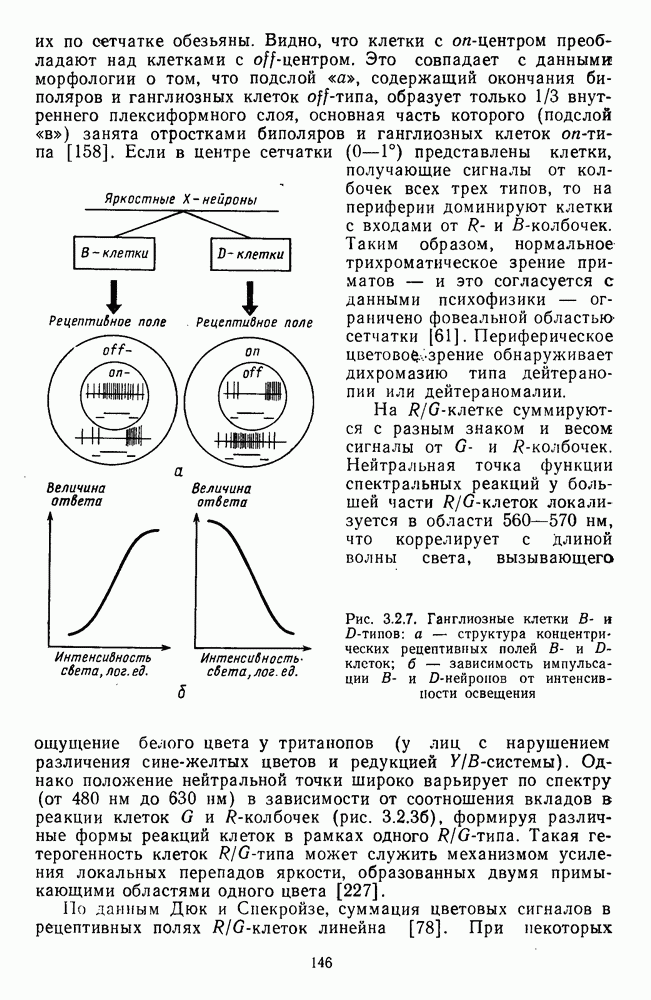









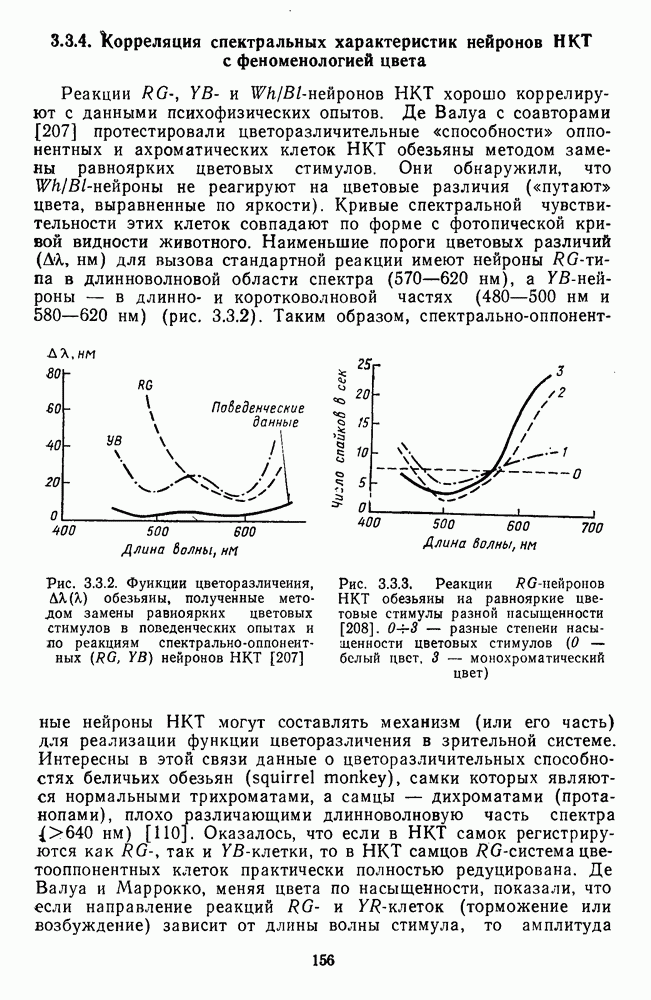

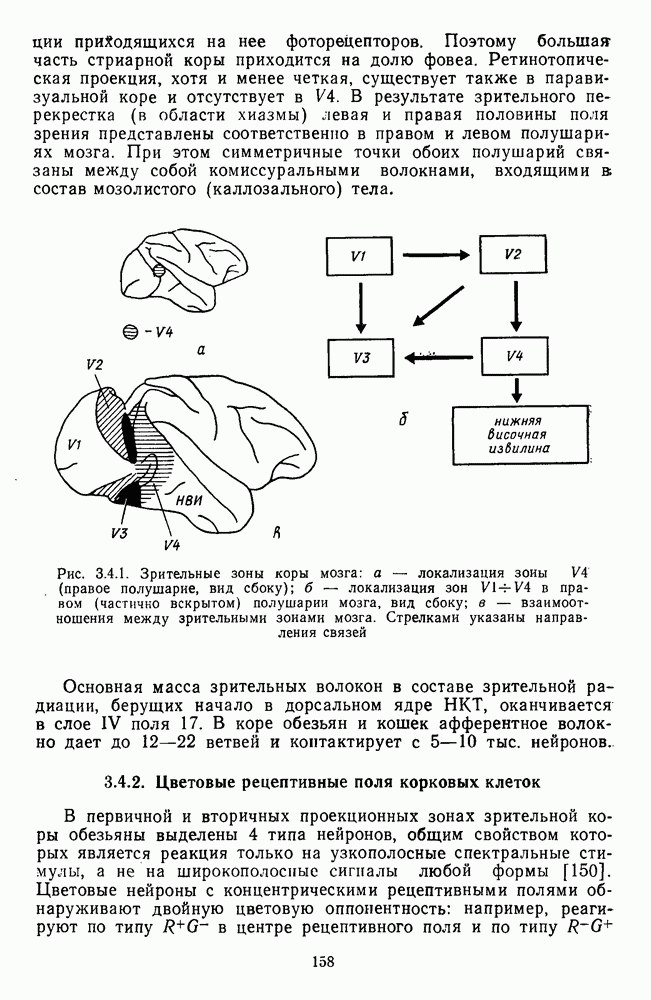












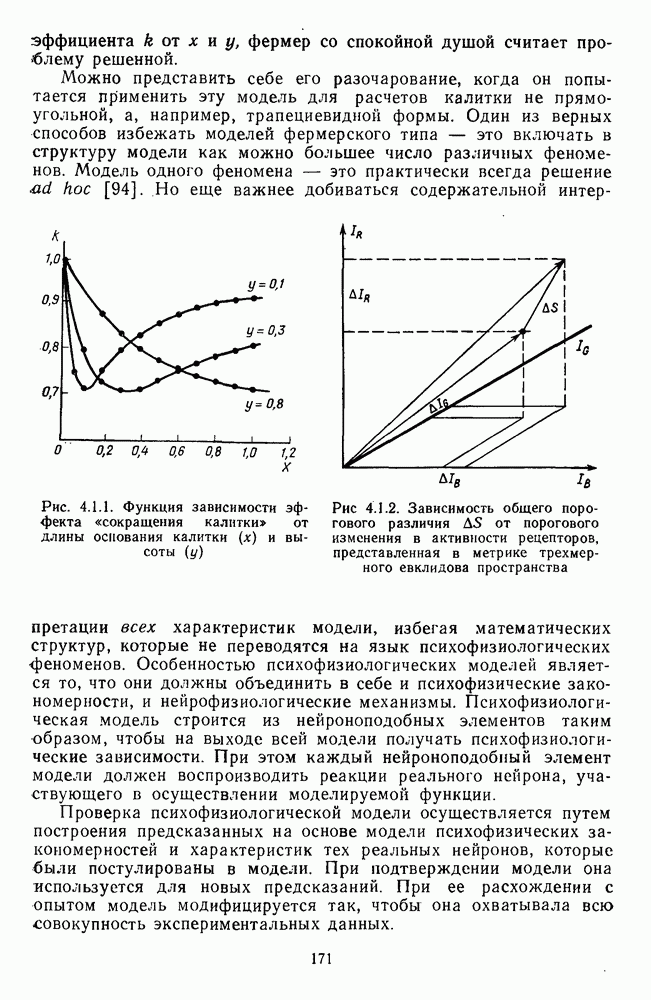






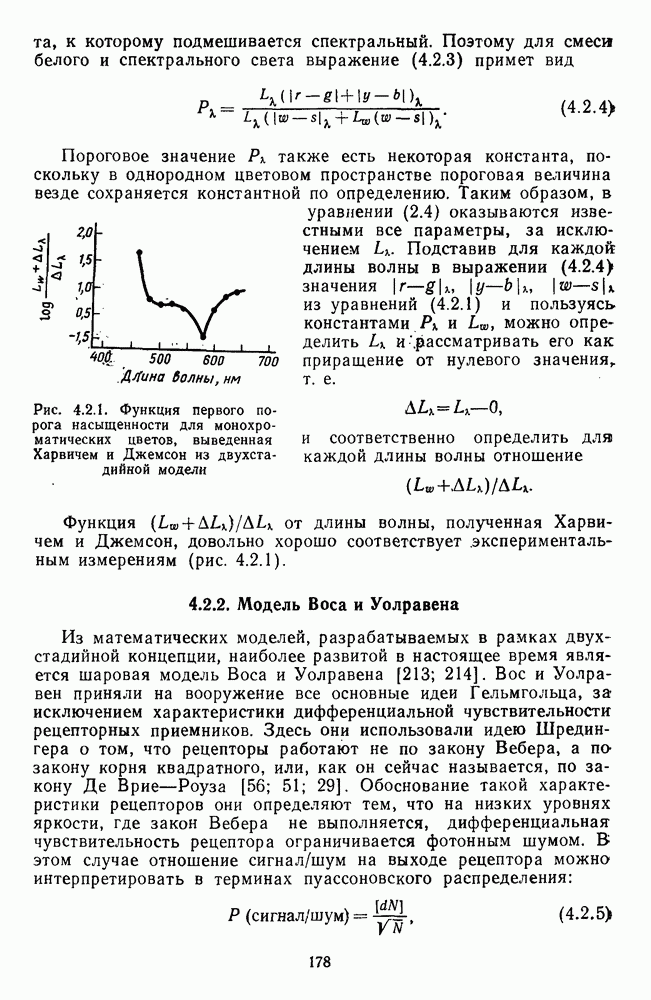





















 -клетки в НКТ. Широкополосные нейроны максимально возбуждались при предъявлении белого света. Таким образом, в стриарной коре имеется два типа спектрально-селективных детекторов: 1) с концентрическими рецептивными полями и 2) с «простыми», «сложными» или «сверхсложными» полями.
-клетки в НКТ. Широкополосные нейроны максимально возбуждались при предъявлении белого света. Таким образом, в стриарной коре имеется два типа спектрально-селективных детекторов: 1) с концентрическими рецептивными полями и 2) с «простыми», «сложными» или «сверхсложными» полями.