Пред.
След.
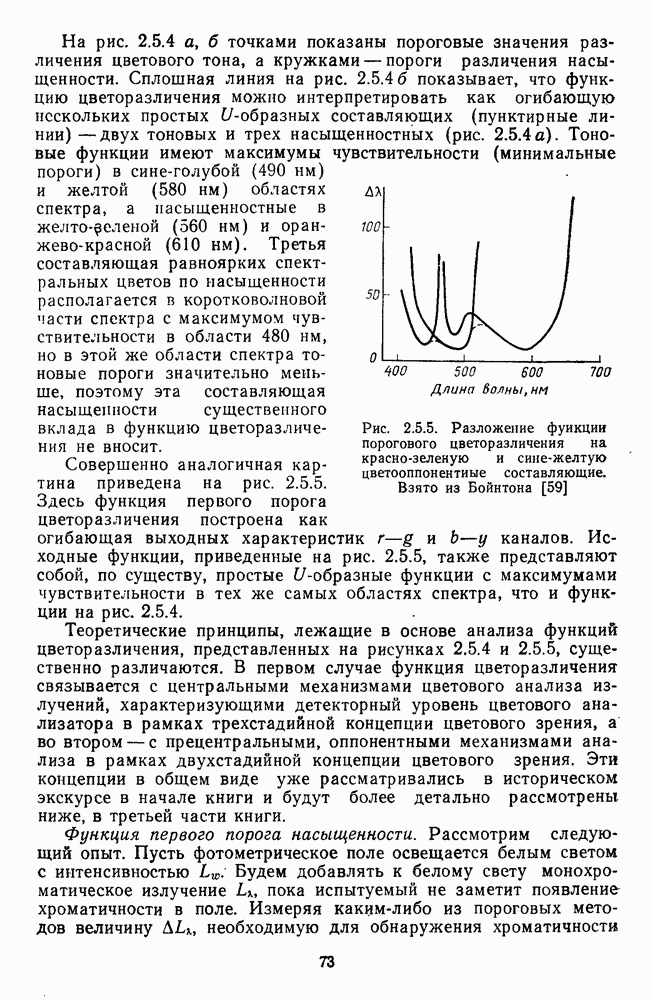
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
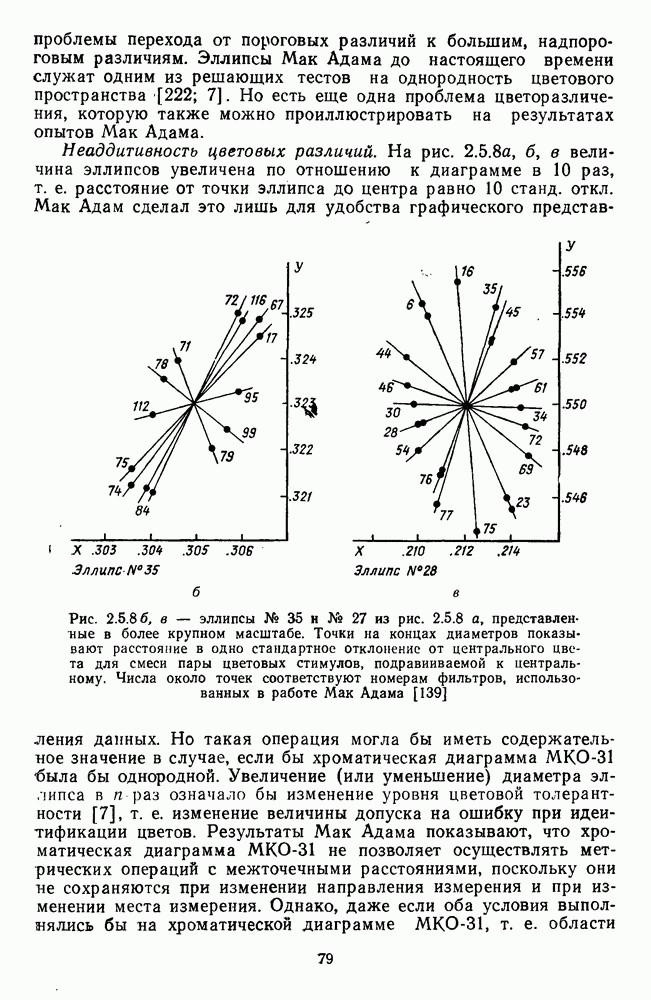
ZADANIA.TO
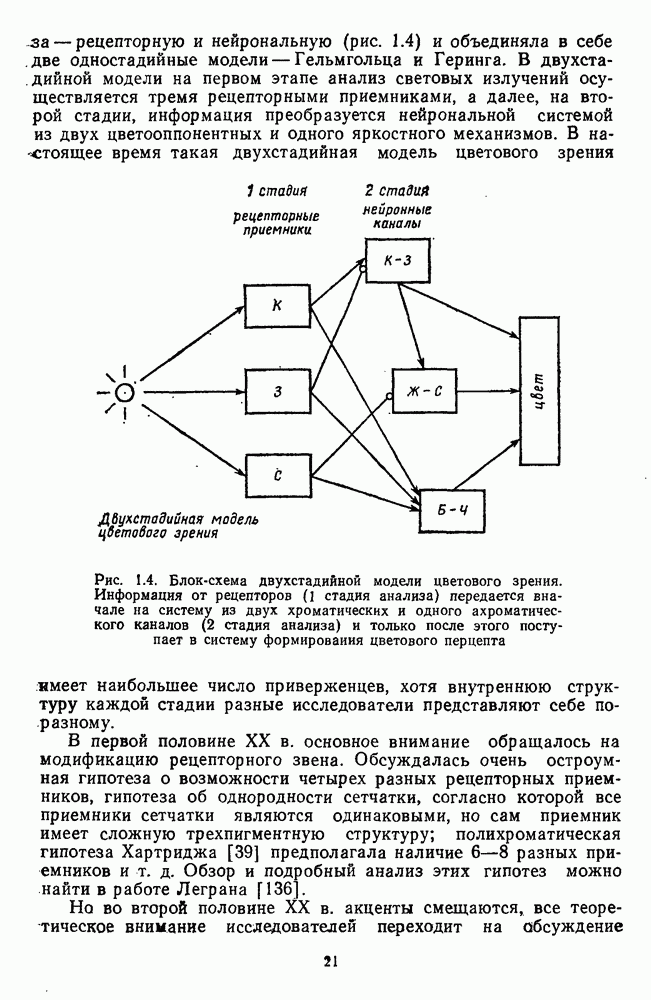
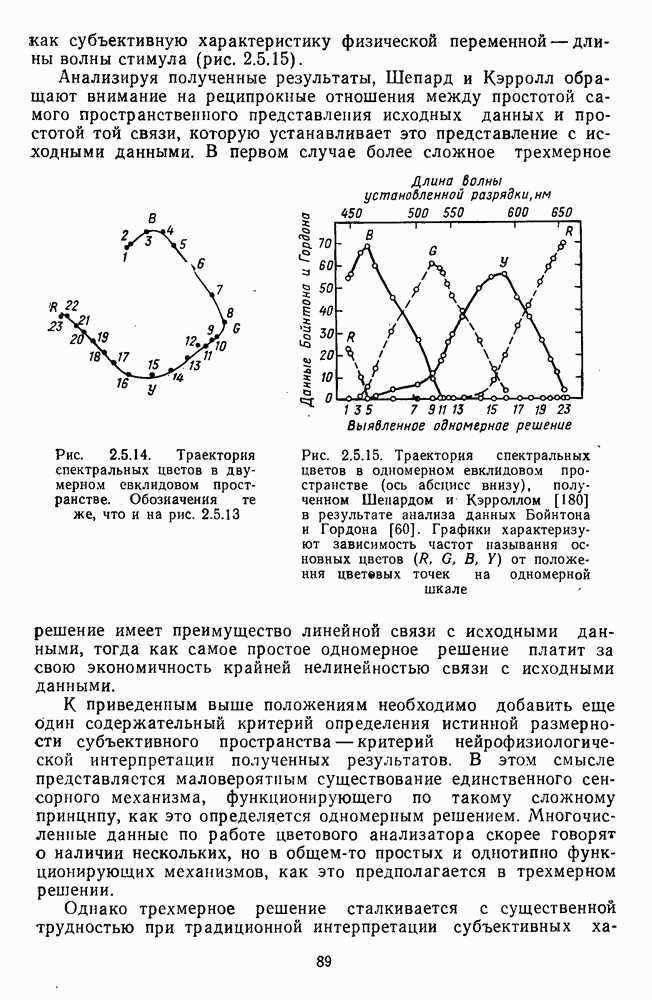
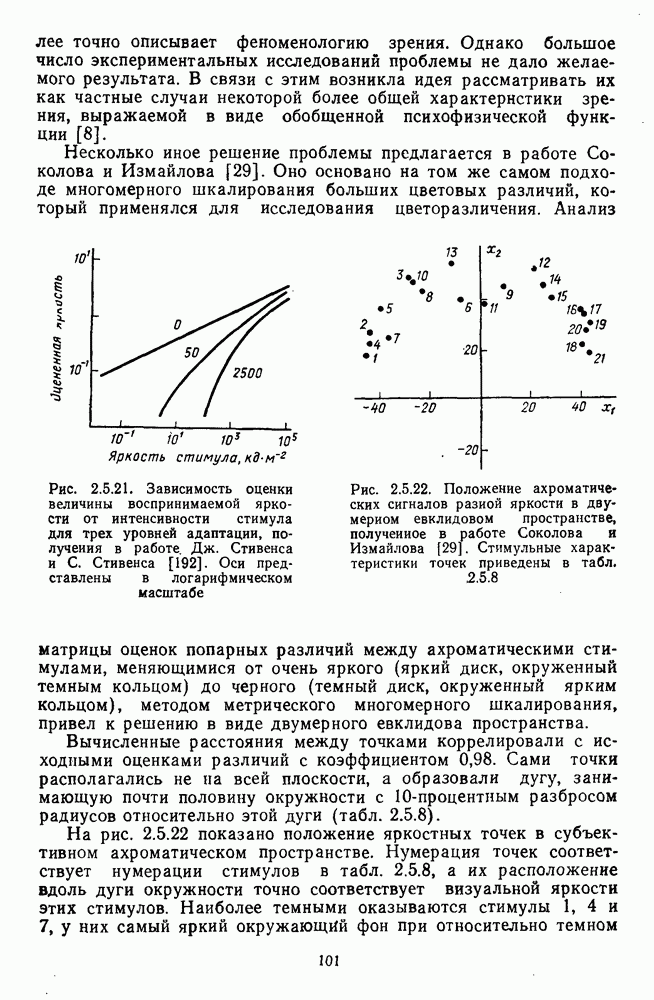
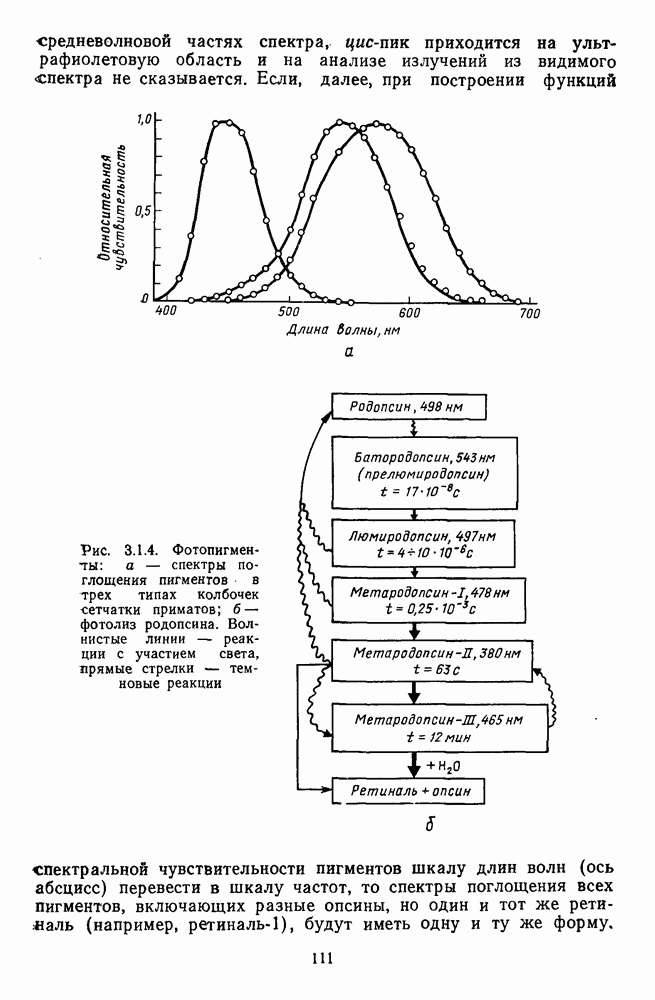
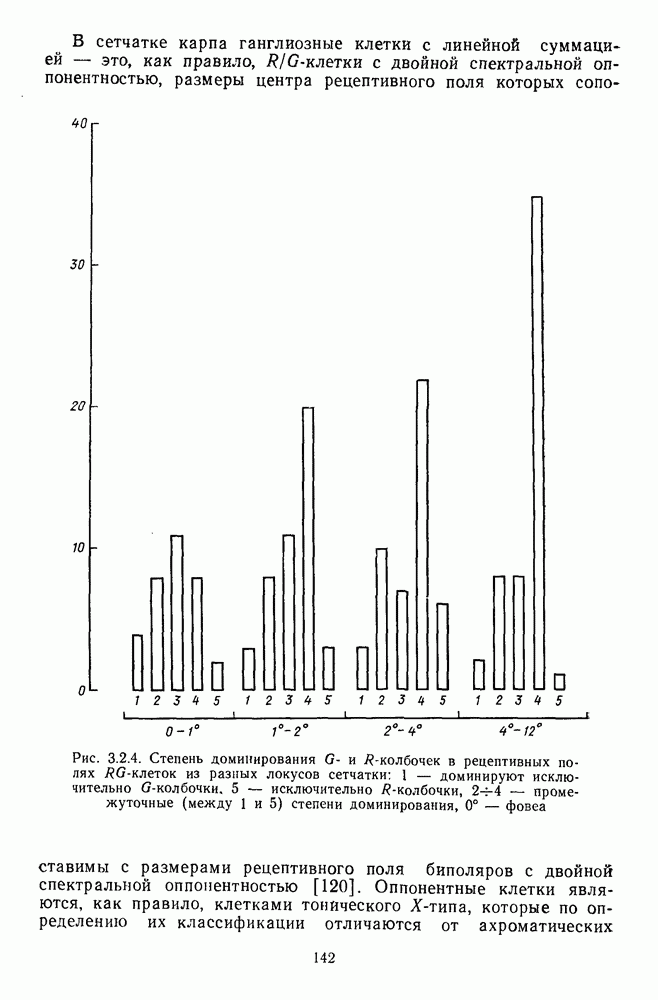
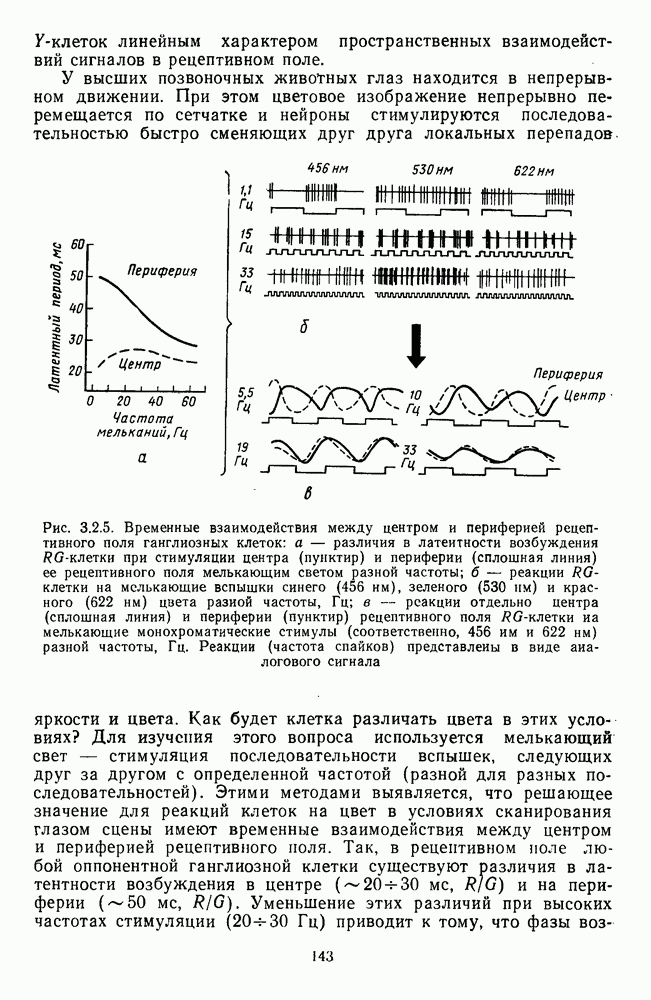
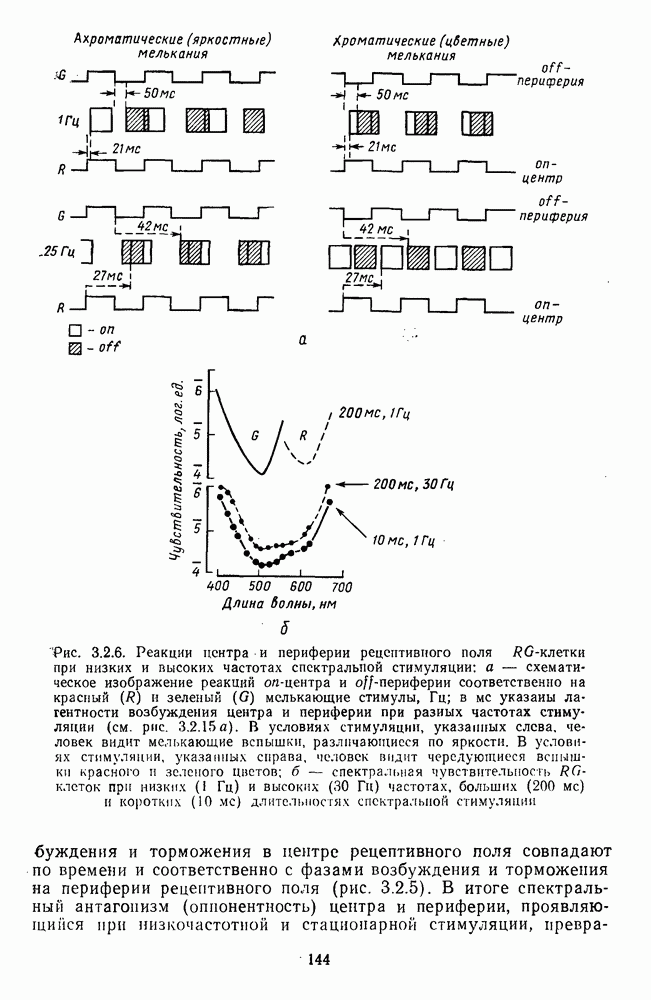
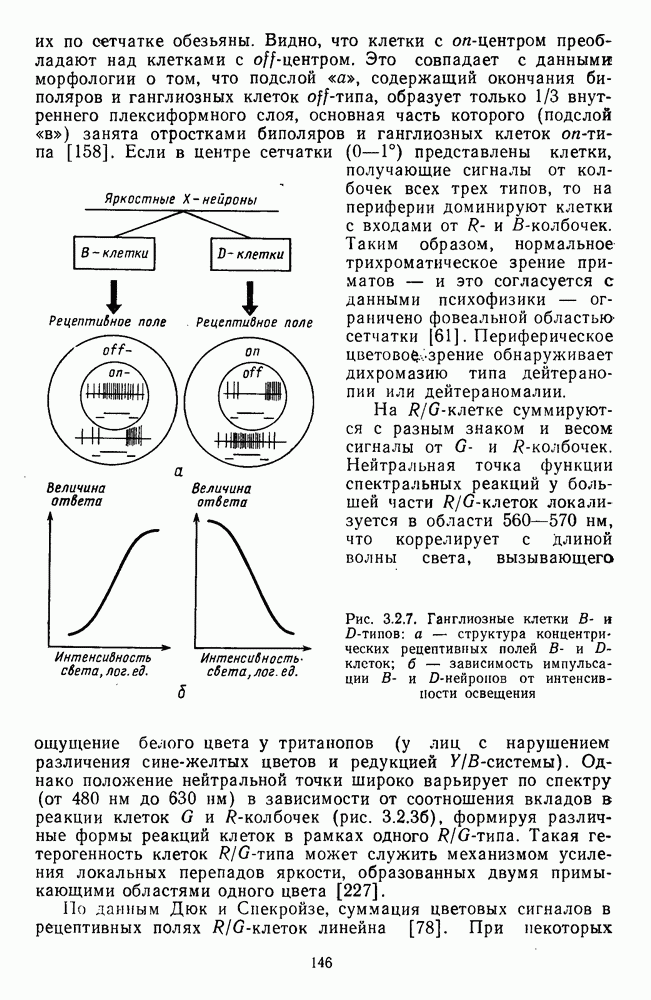
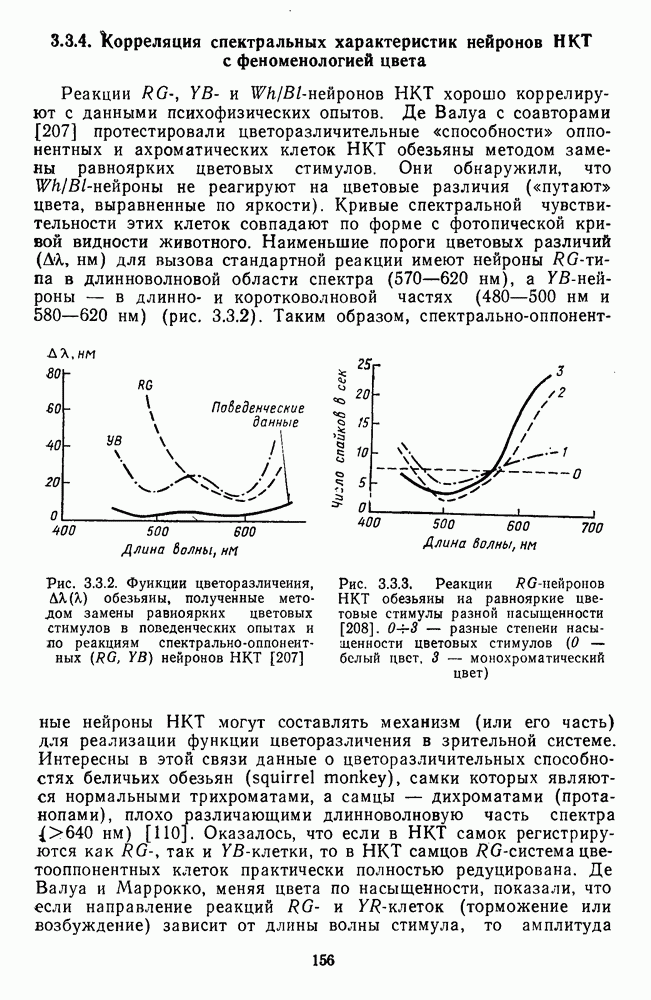
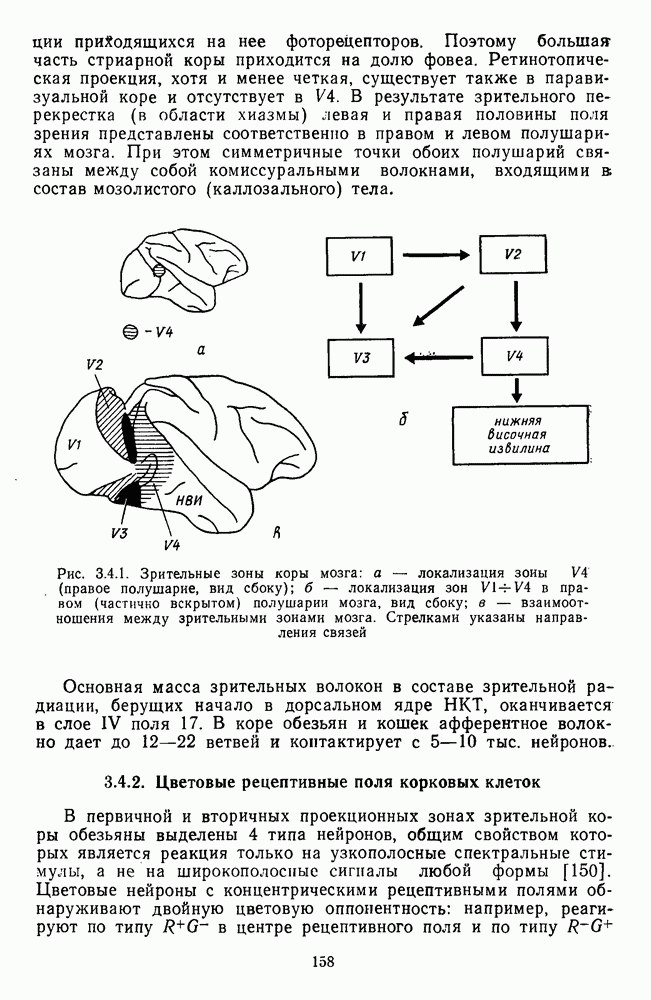
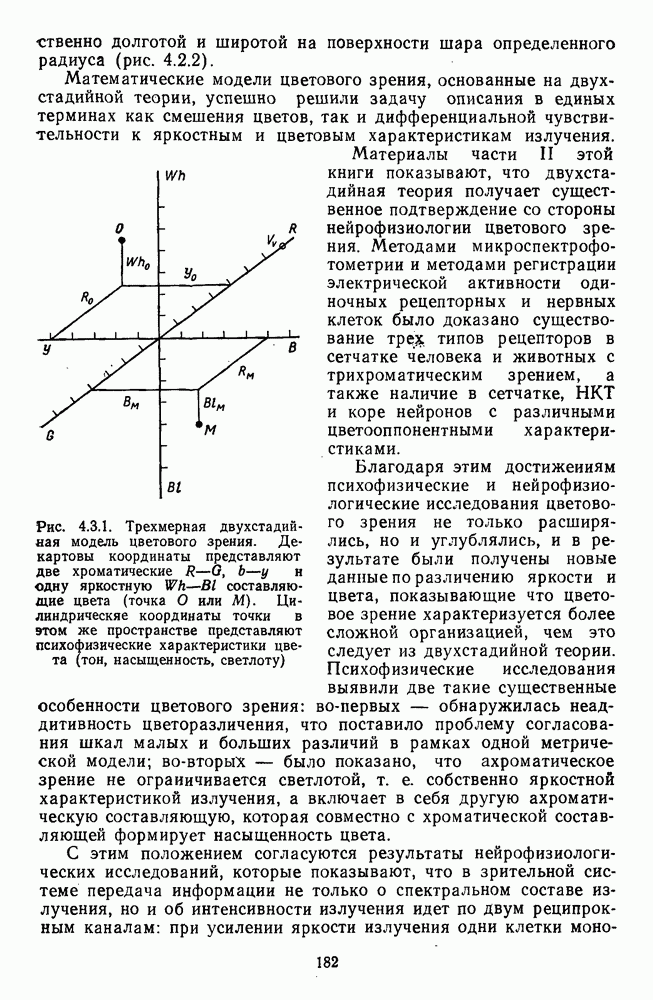
Часть 3. НЕЙРОФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ЦВЕТОВОГО ЗРЕНИЯГлава 3.1. ФОТОРЕЦЕПЦИЯВ настоящее время выделено два параллельных пути обработки зрительной информации: сетчатка — таламус — кора и сетчатка — переднее двухолмие — кора. Цветовой анализ излучения осуществляется последовательно нейронными структурами сетчатки, таламуса (наружное коленчатое тело) и коры больших полушарий головного мозга. Относительный вклад каждой из этих структур определяется уровнем развития животного. Наиболее близкими к человеку — анатомически и филогенетически — являются обезьяны, цветовое зрение которых, как показано в обширных поведенческих исследованиях Де Валуа [209], в основных своих проявлениях совпадает с цветовым зрением человека. Поэтому при рассмотрении нейронных механизмов цветового зрения акцент будет сделан — там, где это возможно, — на данных, полученных в опытах на обезьянах. 3.1.1. Морфология рецепторов сетчаткиОбщее строение сетчатки. Сетчатка глаза, выстилающая внутреннюю поверхность глазного яблока, в процессе онтогенеза развивается из передней стенки мозга. У позвоночных она инвертирована, и свет, прежде чем достичь слоя фоторецепторов, проходит через всю сетчатку. Принципиальная организация сетчатки одинакова у всех позвоночных (рис. 3.1.1). Однако структура синаптических связей между нейронами различна у животных разных видов. Наибольшее разнообразие в организации синаптических связей встречается во внутреннем плексиформном слое, образованном отростками биполярных, амакриновых и ганглиозных клеток. По характеру связей в этом слое выделяют «сложный» тип организации сетчатки, свойственный низшим позвоночным и птицам, и «простой» тип, свойственный млекопитающим (рис. 3.1.2). Так, у лягушек и птиц этот слой очень хорошо выражен. Он делится на ряд подслоев, в которых преобладают амакрино-амакриновые и амакрино-ганглиозные синапсы. У приматов этот слой меньше по размерам с нечетким делением на подслои и преобладанием прямых «биполярганглиозная клетка» синапсов. У низших позвоночных и птиц выделение простых признаков объекта происходит уже в сетчатке, что тесно связано с усложнением там синаптических взаимодействий нейронов и многочисленностью латеральных связей. Морфология сетчатки коррелирует с экологическими условиями обитания животного: варьируют типы фоторецепторов, выраженность слоев.
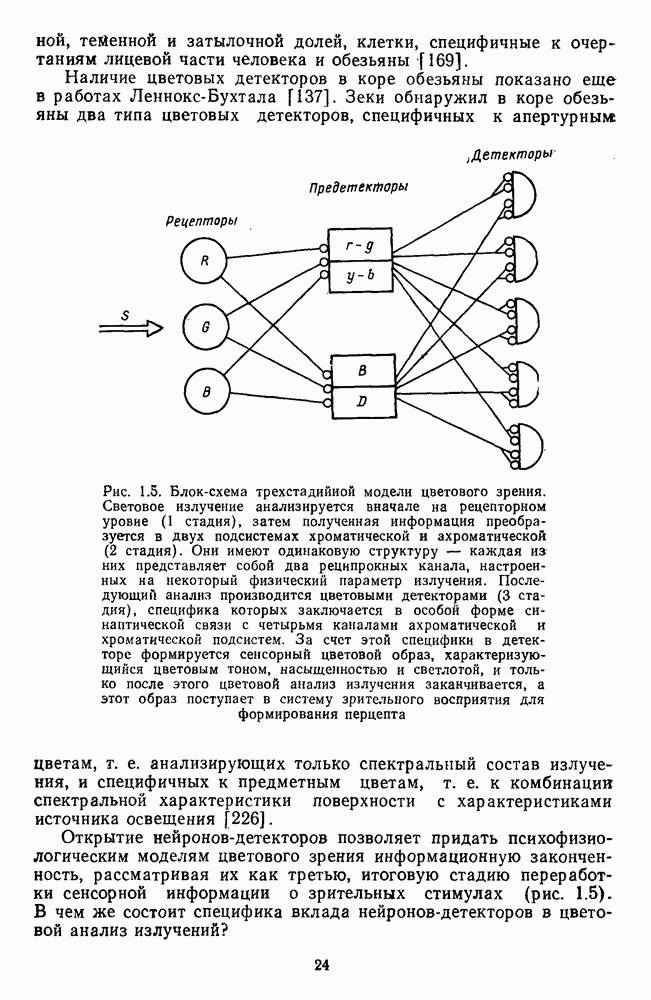
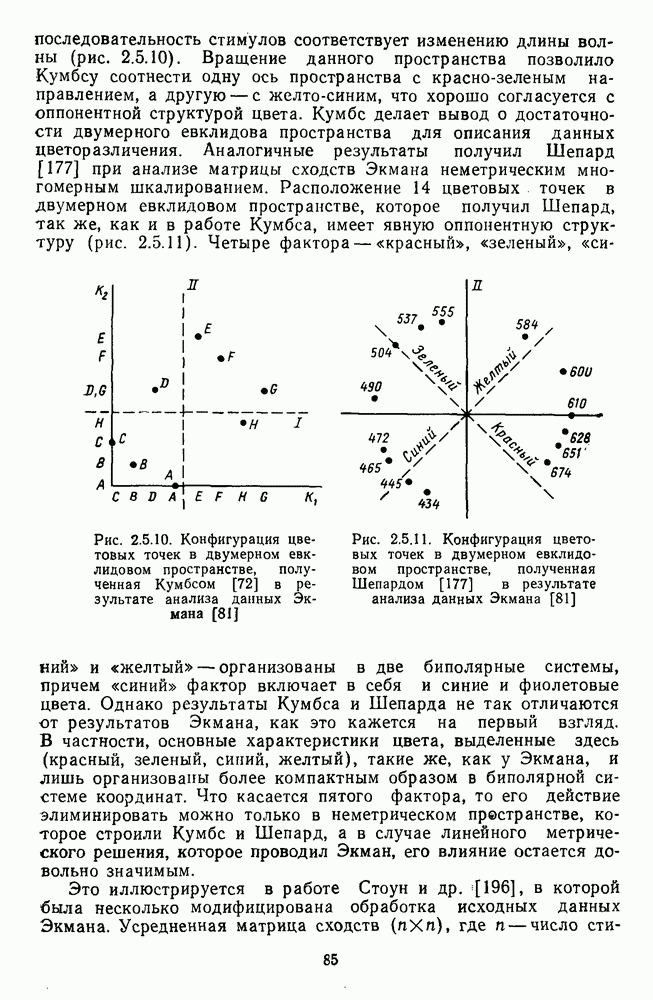
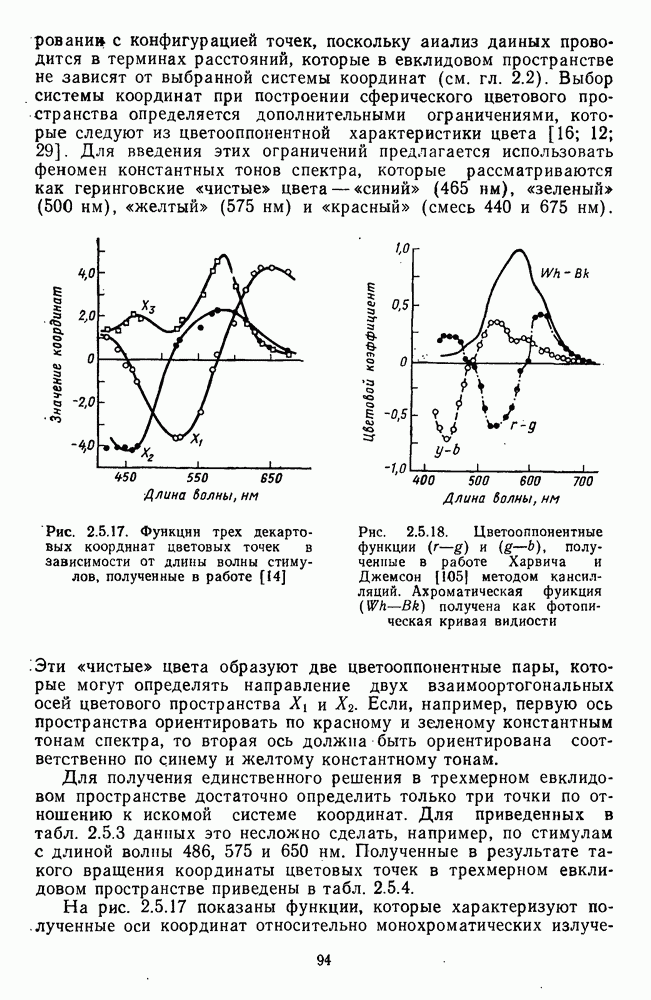
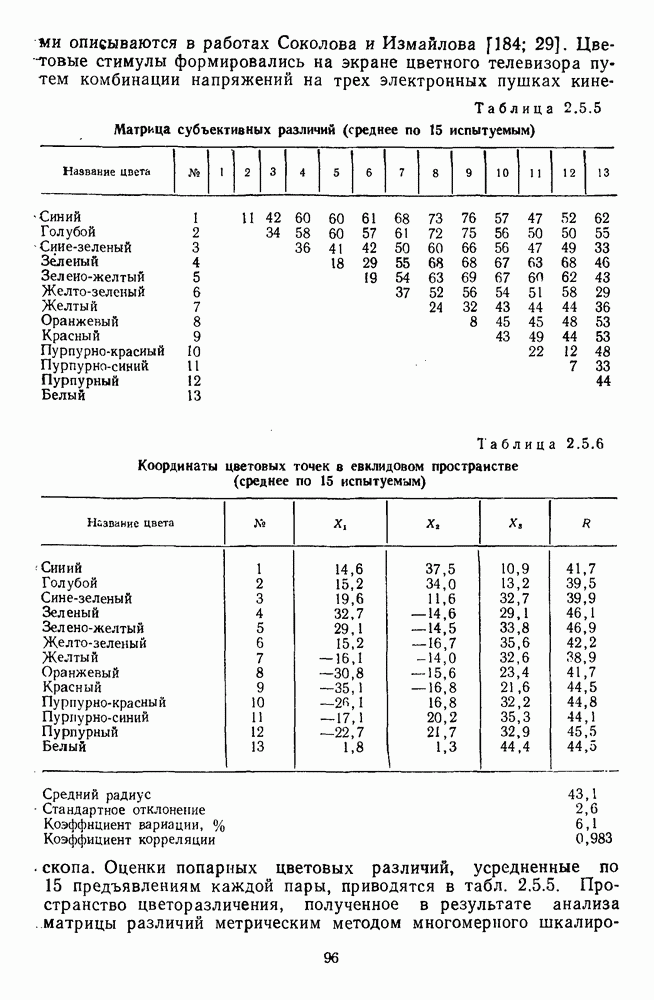
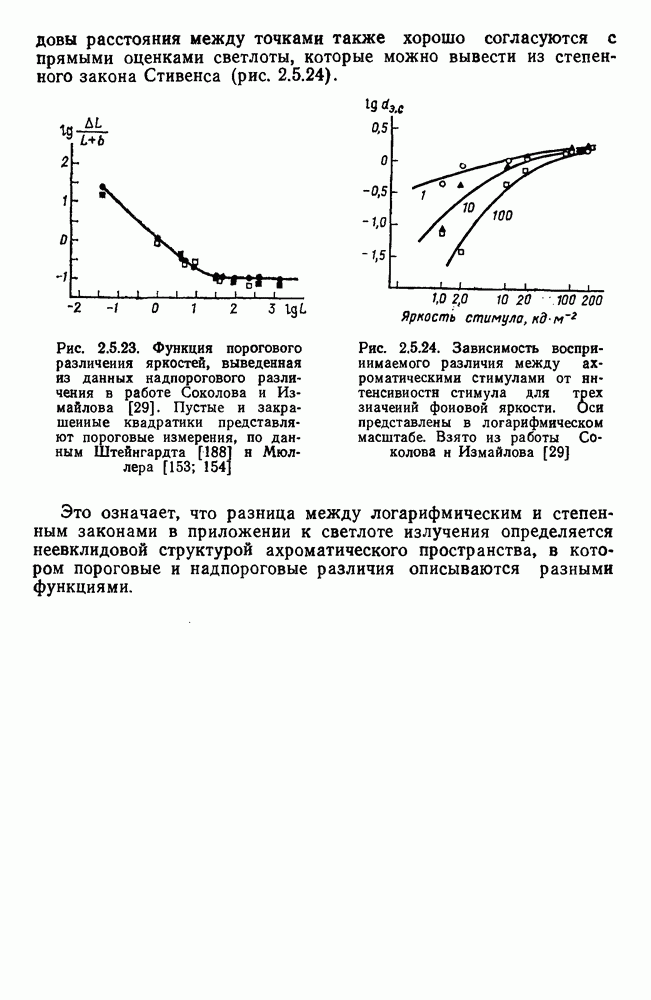
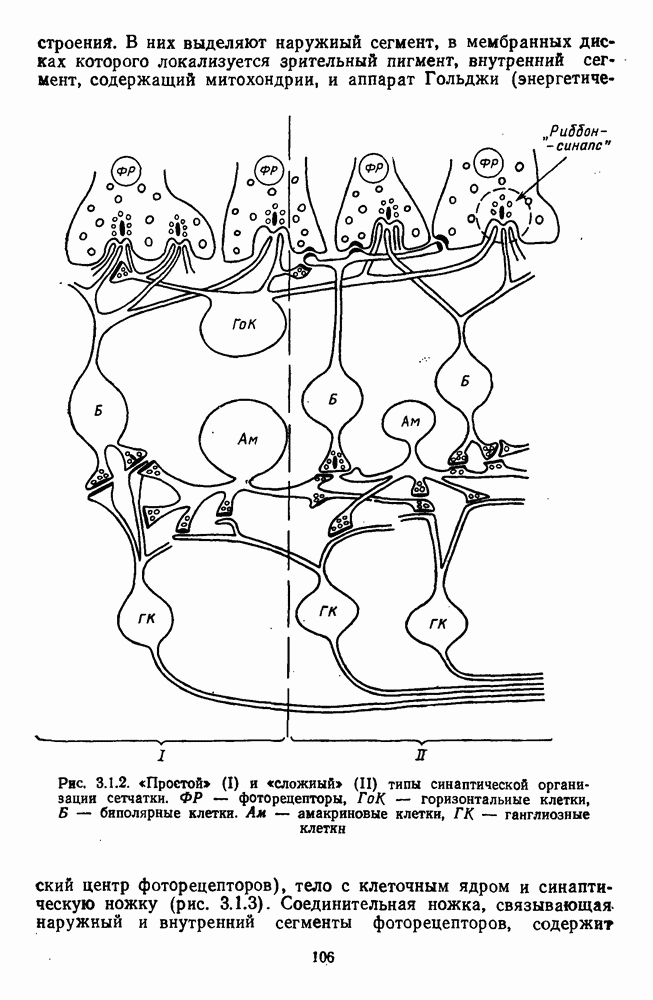
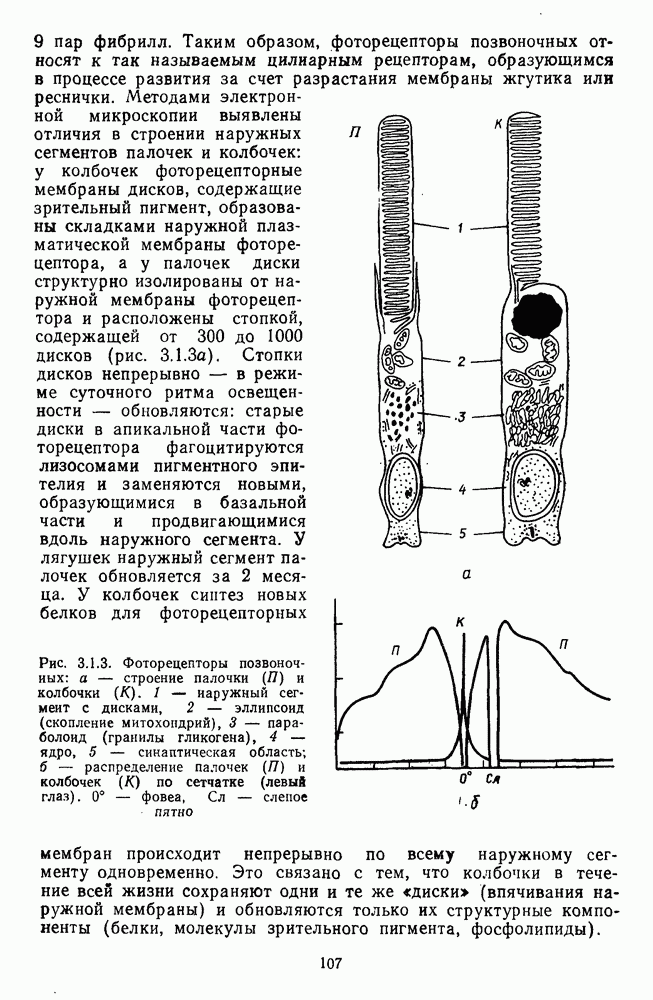
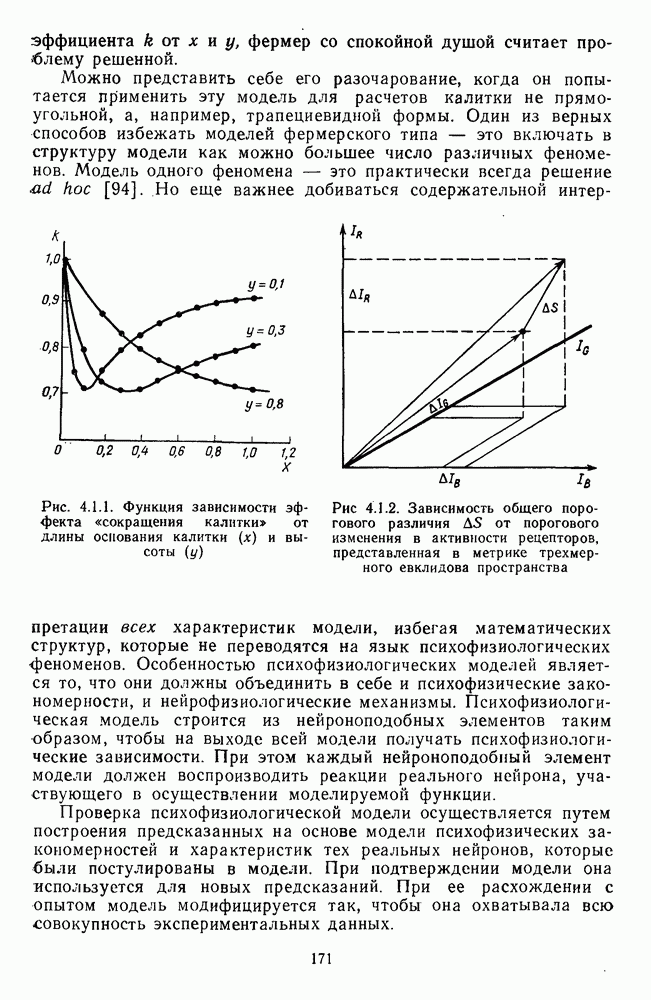
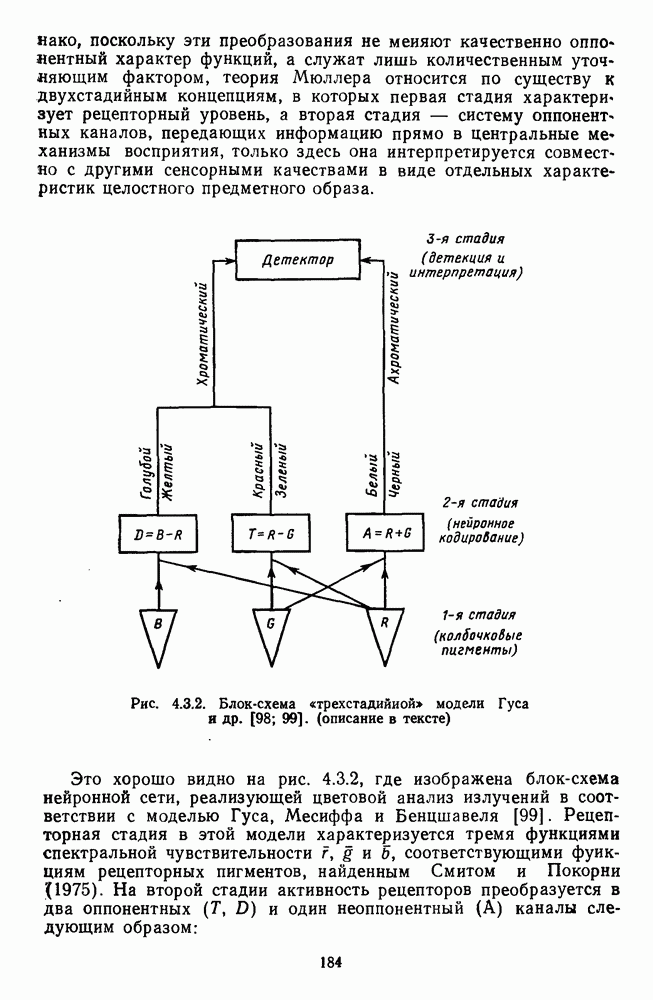
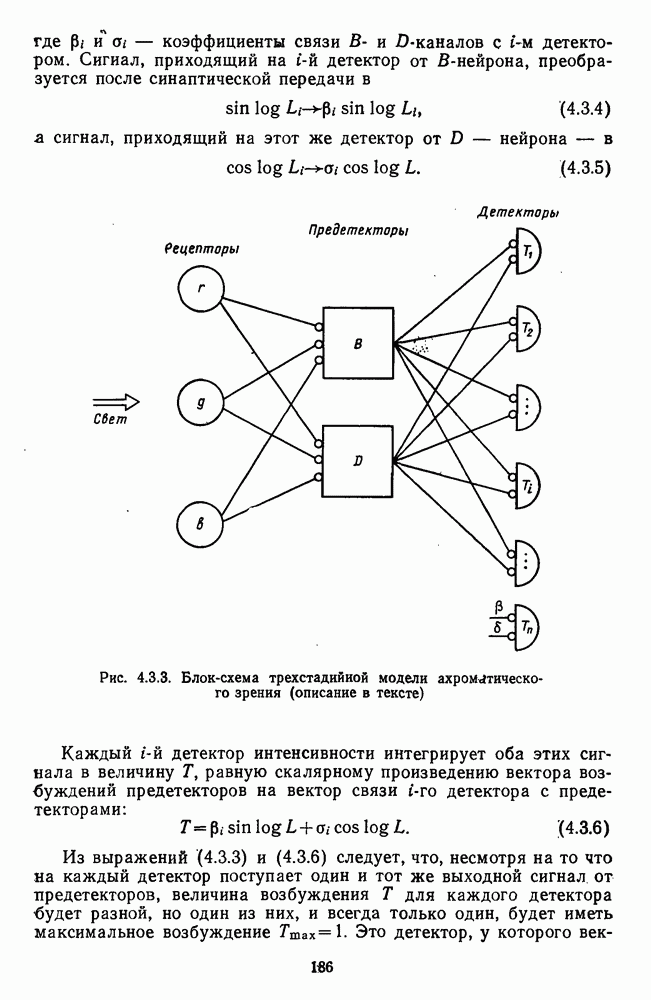
Рис. 3.1.1. Строение сетчатки позвоночных: I, II, III — первый, второй и третий нейроны зрительной системы; I — слой пигментного эпителия, 2 — слой палочек (П) и колбочек (К), 3 — наружная пограничная мембрана, 4 — наружный зернистый (ядериый) слой (клеточные ядра рецептора), 5 — наружный межзернистый (плексиформный) слой (синапсы рецепторов с нейронами второго порядка), 6 — внутренний зернистый (ядериый) слой (тела горизонтальных, биполярных и амакриновых клеток), 7 — внутренний межзернистый (плексиформный) слой (синапсы биполярных, амакриновых и ганглиозных клеток между собой), 8 — ганглиозные клетки, 9 — волокна зрительного нерва, 10 — внутренняя пограничная мембрана, М — волокно (клетка) Мюллера Строение, типы и распределение фоторецепторов по сетчатке. Сетчатки большинства позвоночных содержат рецепторы скотопической системы зрения — палочки и рецепторы фотопической системы — колбочки. Палочки и колбочки имеют общую схему строения. В них выделяют наружный сегмент, в мембранных дисках которого локализуется зрительный пигмент, внутренний сегмент, содержащий митохондрии, и аппарат Гольджи (энергетический центр фоторецепторов), тело с клеточным ядром и синаптическую ножку (рис. 3.1.3). Соединительная ножка, связывающая наружный и внутренний сегменты фоторецепторов, содержит Рис. 3.1.2. (см. скан) «Простой» (I) и «сложный» (II) типы синаптической организации сетчатки. 9 пар фибрилл. Таким образом, фоторецепторы позвоночных относят к так называемым цилиарным рецепторам, образующимся в процессе развития за счет разрастания мембраны жгутика или реснички. Методами электронной микроскопии выявлены отличия в строении наружных сегментов палочек и колбочек: у колбочек фоторецепторные мембраны дисков, содержащие зрительный пигмент, образованы складками наружной плазматической мембраны фоторецептора, а у палочек диски структурно изолированы от наружной мембраны фоторецептора и расположены стопкой, содержащей от 300 до 1000 дисков (рис. 3.1.3а). Стопки дисков непрерывно — в режиме суточного ритма освещенности — обновляются: старые диски в апикальной части фоторецептора фагоцитируются лизосомами пигментного эпителия и заменяются новыми, образующимися в базальной части и продвигающимися вдоль наружного сегмента. У лягушек наружный сегмент палочек обновляется за 2 месяца.
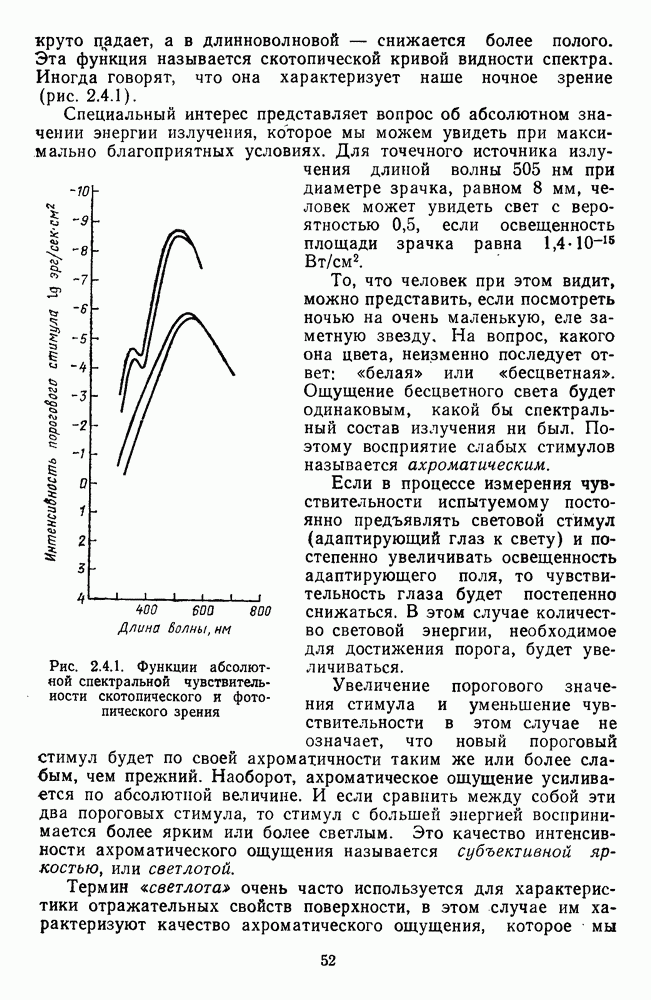
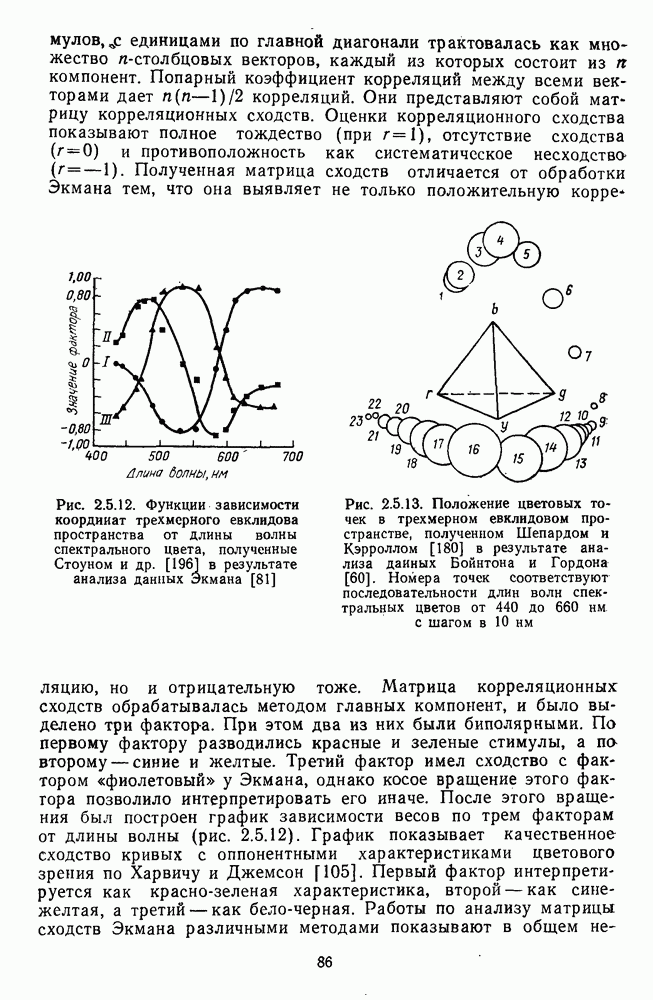
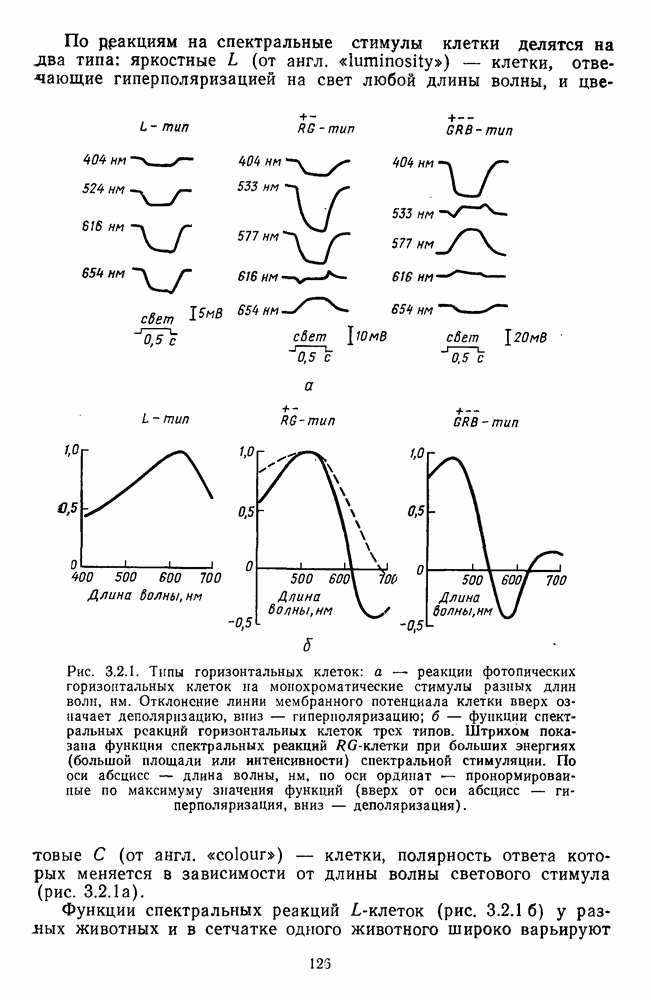
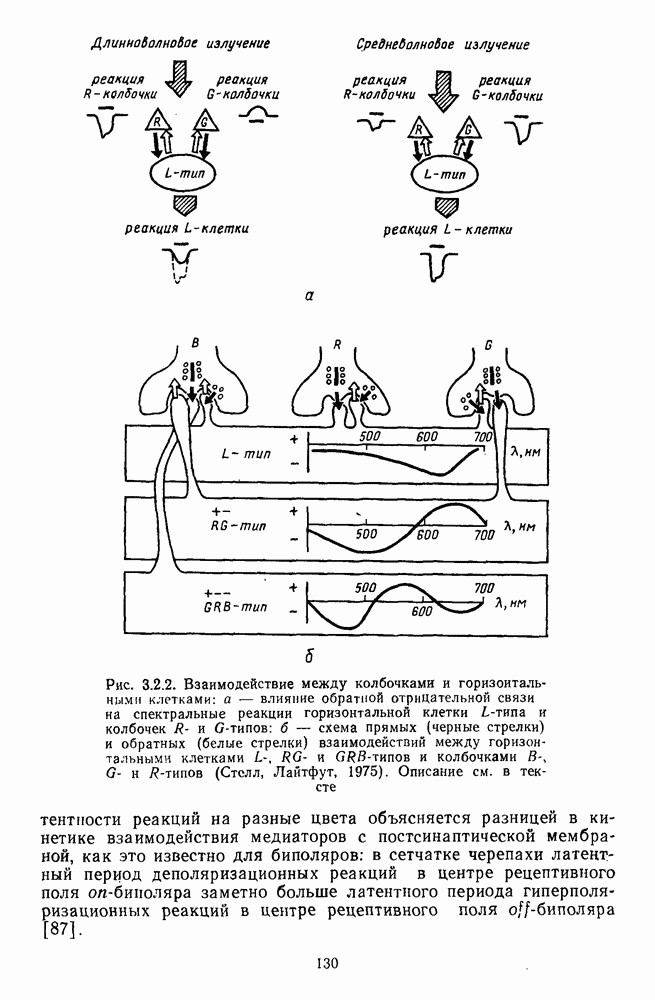
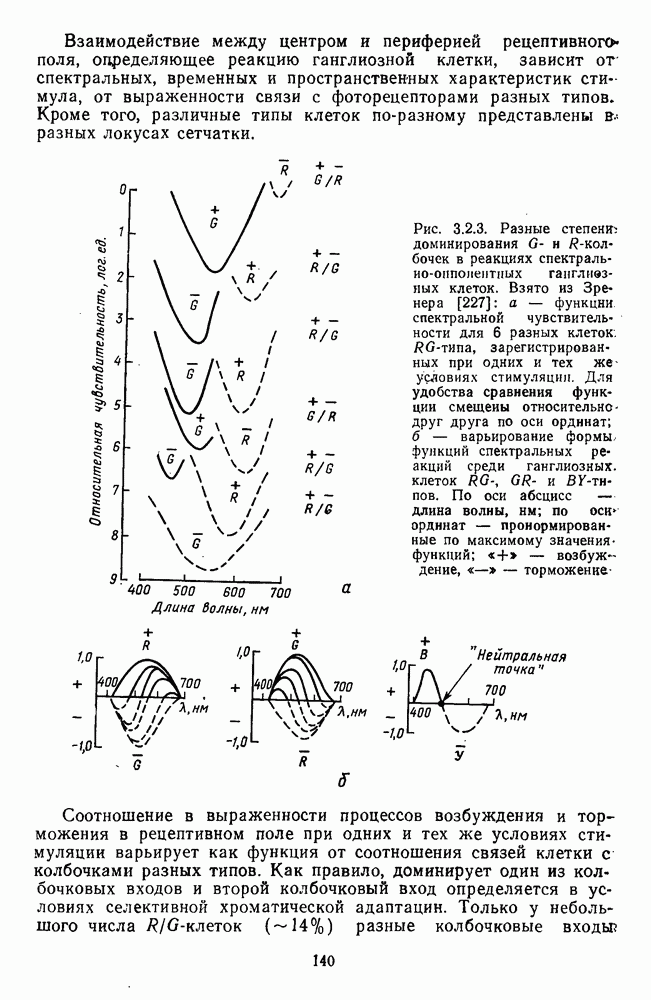
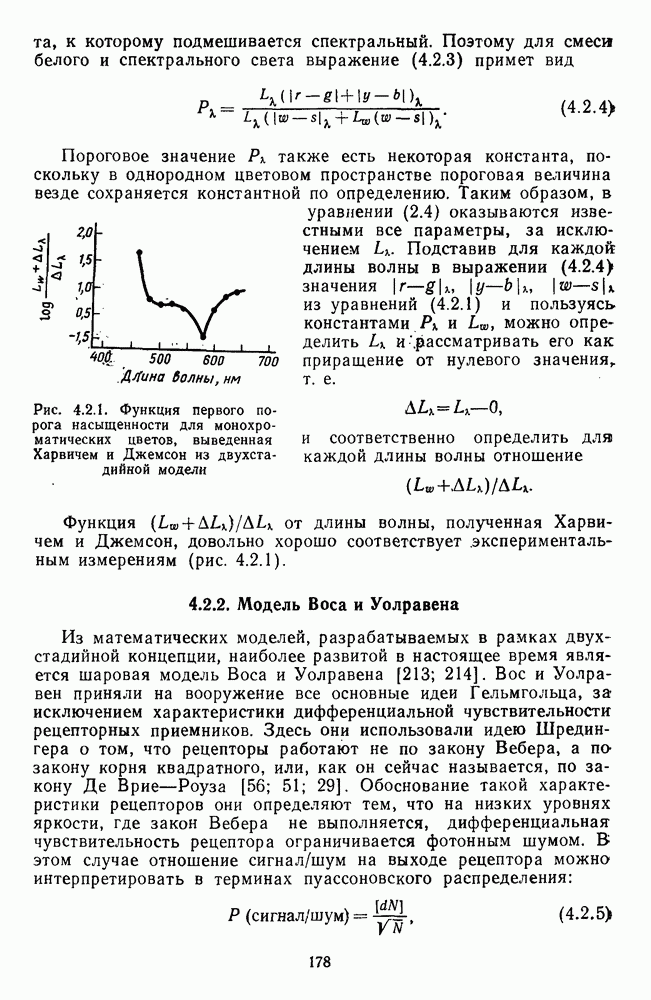
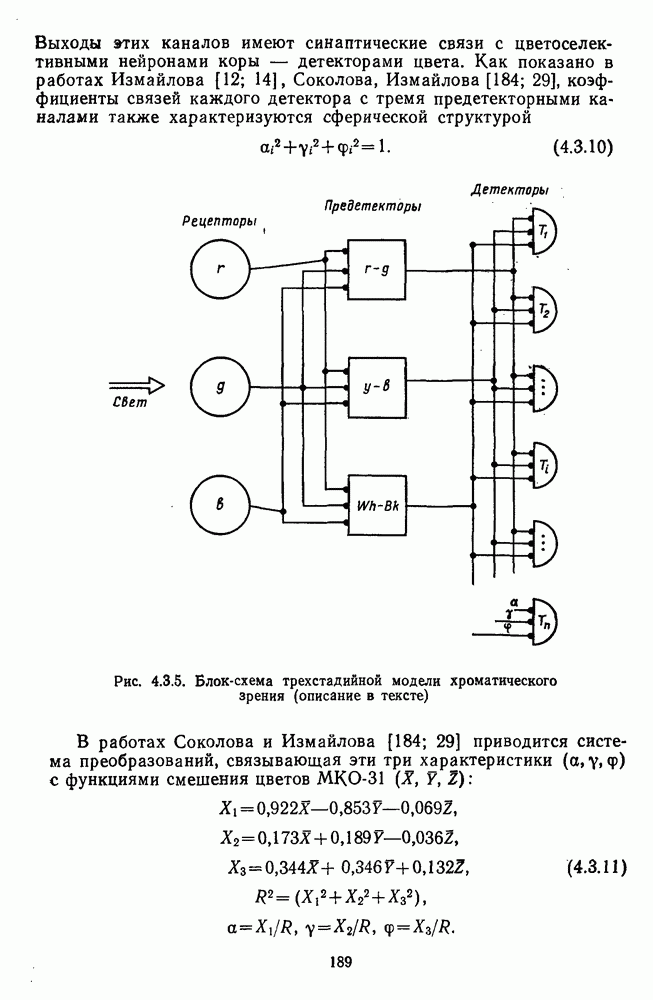
Рис. 3.1.3. Фоторецепторы позвоночных: а — строение палочки (П) и колбочки (К). 1 — наружный сегмент с дисками, 2 — эллипсоид (скопление митохондрий), 3 — параболоид (гранилы гликогена), 4 — ядро, 5 — синаптическая область; У колбочек синтез новых белков для фоторецепторных мембран происходит непрерывно по всему наружному сегменту одновременно. Это связано с тем, что колбочки в течение всей жизни сохраняют одни и те же «диски» (впячивания наружной мембраны) и обновляются только их структурные компоненты (белки, молекулы зрительного пигмента, фосфолипиды). У многих позвоночных (рептилии, птицы), кроме млекопитающих, во внутреннем сегменте колбочек содержатся масляные капли, могущие выполнять функции интраокулярных фильтров. У дневных животных эти капли окрашены каротиноидами в красный, желтый или желто-зеленый цвета, у ночных — они бесцветные, поглощающие в ультрафиолетовой области спектра. Цветные капли как светофильтры не пропускают синюю часть спектра, что может уменьшать хроматическую аберрацию и усиливать цветовой контраст. В «сложных» сетчатках низших позвоночных рецепторы характеризуются большим разнообразием, чем в «простых» сетчатках у млекопитающих. Так, в сетчатке рыб содержатся не только одиночные колбочки, но и отсутствующие у млекопитающих двойные, тройные и даже четверные колбочки, миниатюрные (карликовые) колбочки. Основной и дополнительный члены «составных» колбочек электрически связаны между собой и содержат разные фотопигменты. Не менее шести типов колбочек обнаружено в сетчатке птиц и черепах. Распределение фоторецепторов в «простых» и «сложных» сетчатках различно. Так, у рыб они распределены равномерно по сетчатке. У амфибий в центре сетчатки увеличена плотность всех клеток в 2—3 раза, и только у пресмыкающихся в центре сетчатки появляется область — «центральное пятно» или «центральная полоска» (у черепах) — с повышенным содержанием колбочек. У млекопитающих фоторецепторы распределены по сетчатке неравномерно (рис. 3.1.36). Плотность колбочек максимальна в области «желтого пятна» (макулы). Соотношение между колбочками и ганглиозными клетками достигает 1 : 1. У большинства птиц существует вторая фовеа в темпоральной (височной) области сетчатки на расстоянии около 6 мм от центральной. В сетчатке человека насчитывается 110 - 125 млн палочек и 4-7 колбочек. Связи фоторецепторов между собой и нейронами второго порядка (биполярными и горизонтальными клетками). В электронно-микроскопических исследованиях выявлены электрические синапсы щелевого типа (расстояние между примыкающими мембранами Синаптические окончания палочек и колбочек образуют специализированные соединения (т. е. «риббон—синапсы», или ленточные синапсы) с терминалями дендритов биполярных клеток, дендритов и аксонов горизонтальных клеток (рис. 3.1.2). Такие специализированные соединения представляют собой диады или триады, в которых центральный отросток — как правило, это дендрит биполяра — находится непосредственно под синаптической лентой рецептора, а по бокам расположены термннали дендритов горизонтальных клеток. Множество таких диад и триад группируется вокруг синаптических лент на синаптических ножках палочек и колбочек. Согласно одной из гипотез синаптического взаимодействия рецепторов с биполярами и горизонтальными клетками, основанной на теории локальных цепей, сигнал от рецепторов поступает на горизонтальную клетку и через локальный участок ее мембраны синаптически воздействует на биполяр. В результате биполяр имеет спектральную характеристику горизонтальной клетки. Кроме того, горизонтальная клетка выполняет функцию инвертора. Это приводит к удвоению типов биполярных клеток по отношению к числу типов горизонтальных клеток.
|
 |
Оглавление
|












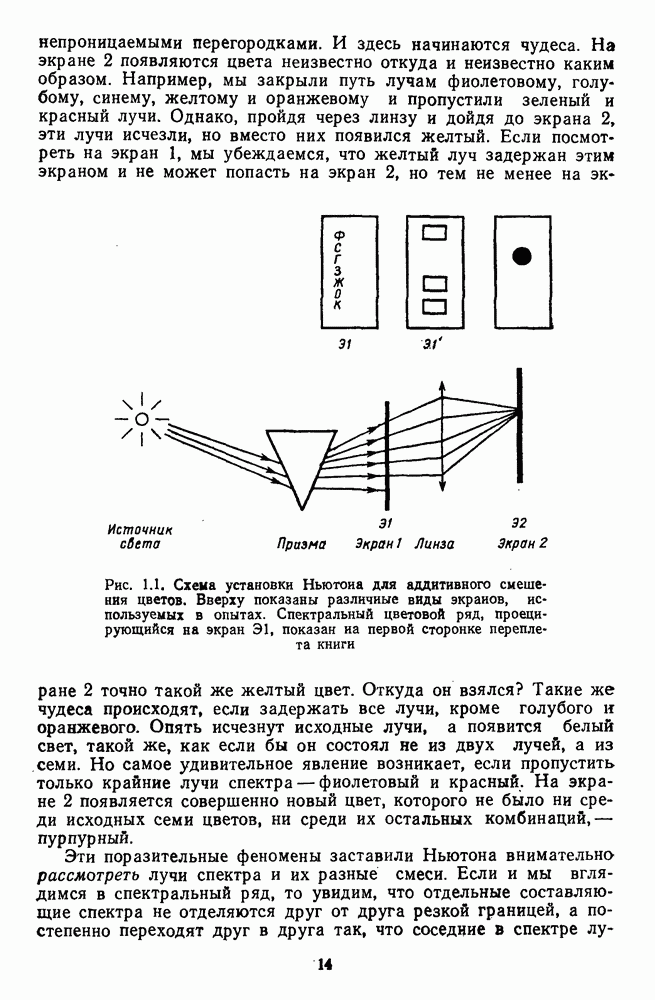























































































































































 — фоторецепторы,
— фоторецепторы,  — горизонтальные клетки, Б — биполярные клетки.
— горизонтальные клетки, Б — биполярные клетки.  — амакриновые клетки,
— амакриновые клетки,  — ганглиозные клеткн
— ганглиозные клеткн

 — распределение палочек
— распределение палочек  и колбочек (К) по сетчатке (левый глаз). 0° — фовеа,
и колбочек (К) по сетчатке (левый глаз). 0° — фовеа,  — слепое пятно
— слепое пятно
 нм) как между однотипными рецепторами (например, коротковолновыми колбочками), так и разных типов (например, между палочкой и колбочкой, между средне- и длинноволновой колбочками). Щелевые синапсы обнаружены и в сетчатке низших позвоночных и млекопитающих (у кошки). Электронно-микроскопическое изучение препаратов сетчатки черепахи, импрегнированных по методу Гольджи, выявило между рецепторами разного типа не только щелевые, но и электрические синапсы типа «плотный контакт (слияние мембраны), а также химические синапсы. Таким образом, уже на начальном этапе обработки сигнала может происходить взаимодействие между палочковой скотопической и колбочковой фотопической системами зрения, а также между колбочками с разной спектральной чувствительностью.
нм) как между однотипными рецепторами (например, коротковолновыми колбочками), так и разных типов (например, между палочкой и колбочкой, между средне- и длинноволновой колбочками). Щелевые синапсы обнаружены и в сетчатке низших позвоночных и млекопитающих (у кошки). Электронно-микроскопическое изучение препаратов сетчатки черепахи, импрегнированных по методу Гольджи, выявило между рецепторами разного типа не только щелевые, но и электрические синапсы типа «плотный контакт (слияние мембраны), а также химические синапсы. Таким образом, уже на начальном этапе обработки сигнала может происходить взаимодействие между палочковой скотопической и колбочковой фотопической системами зрения, а также между колбочками с разной спектральной чувствительностью.