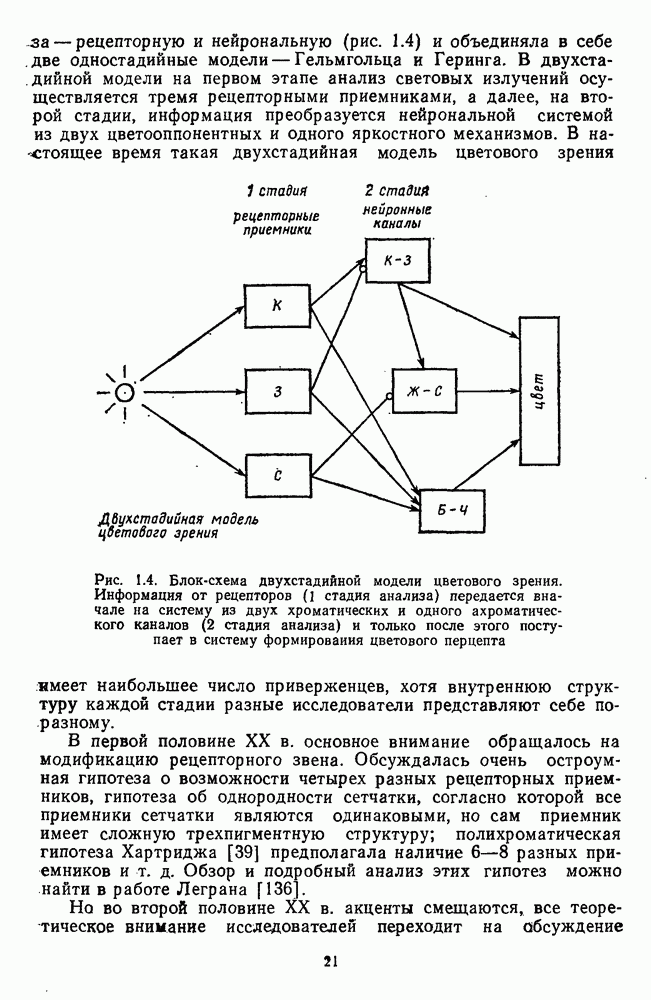
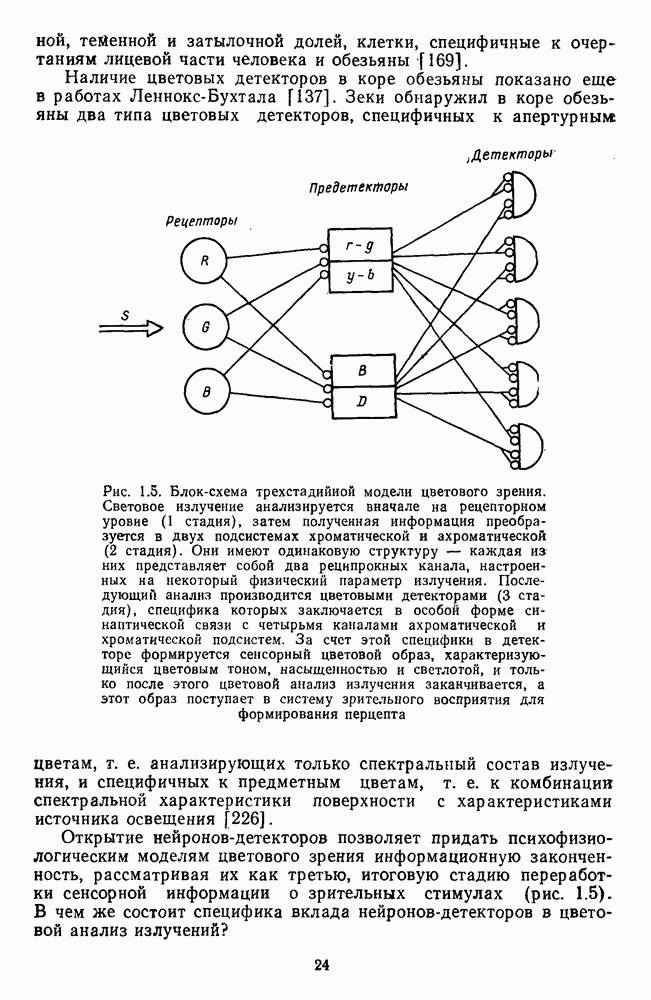
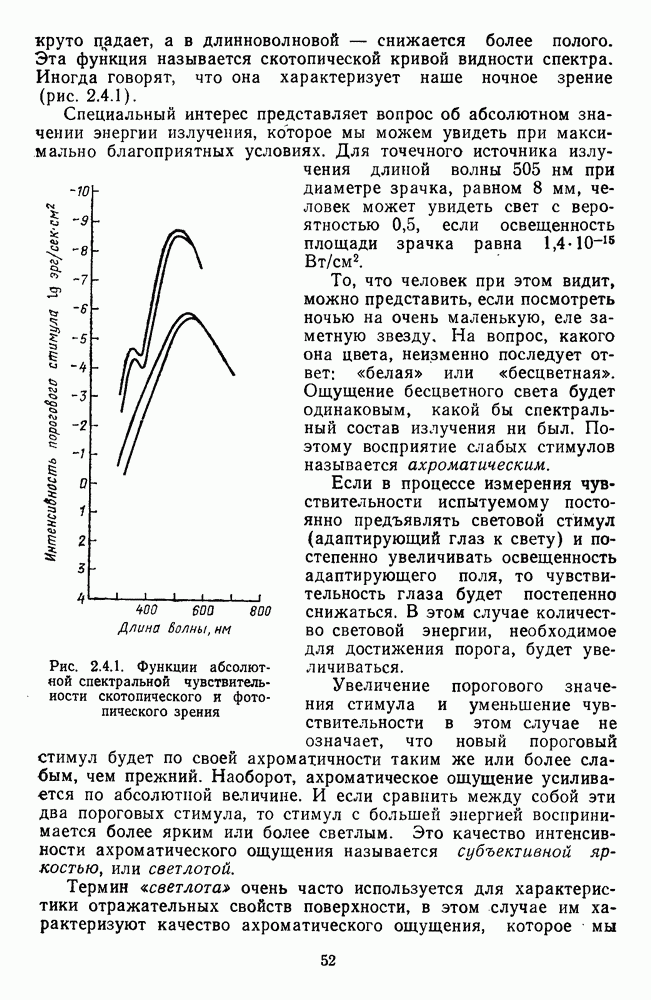
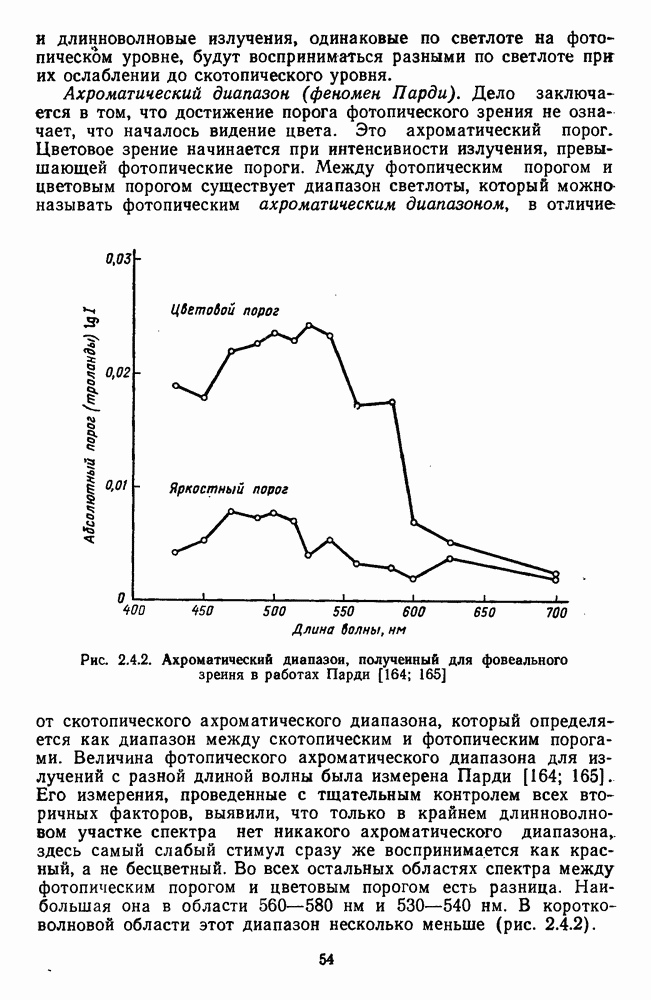
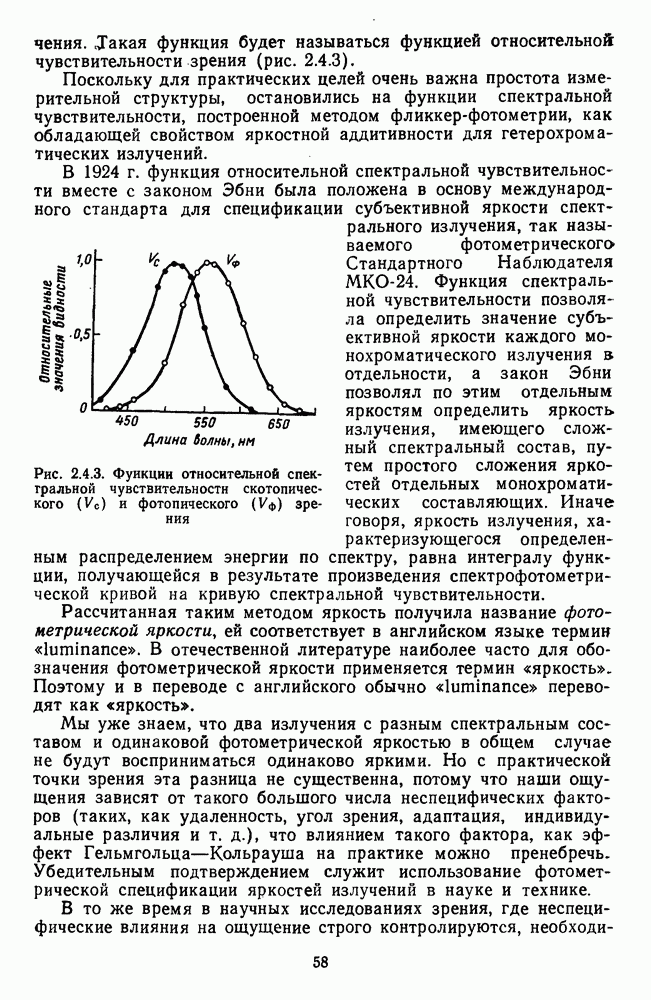
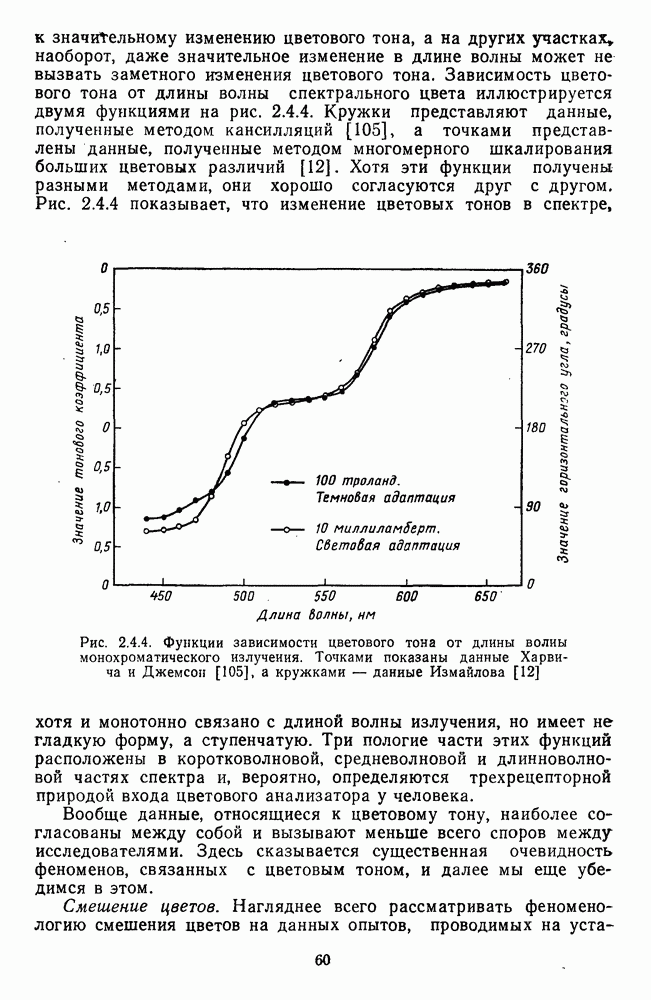
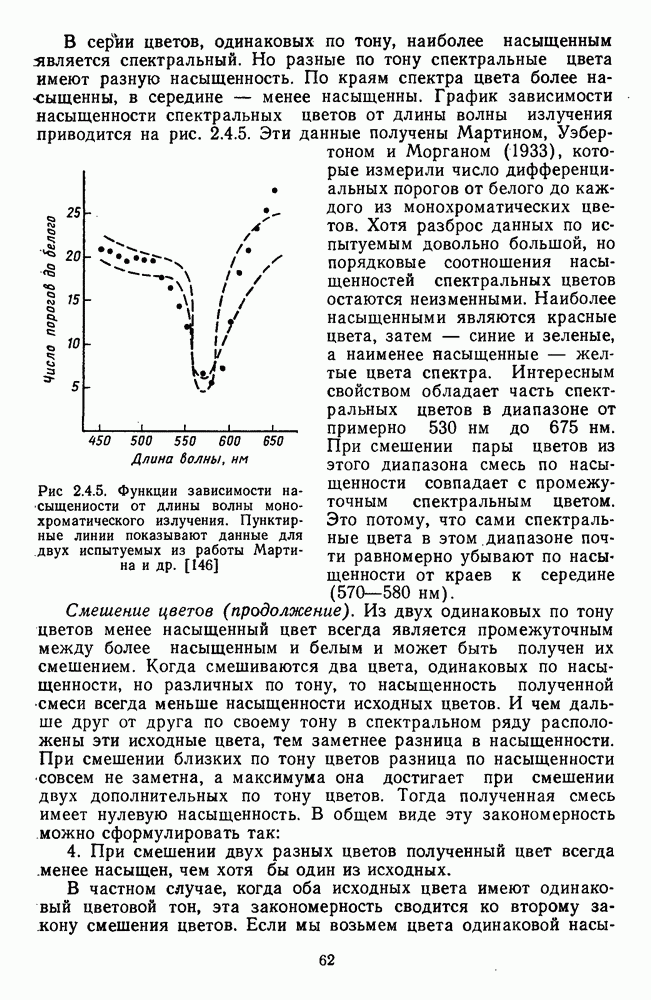
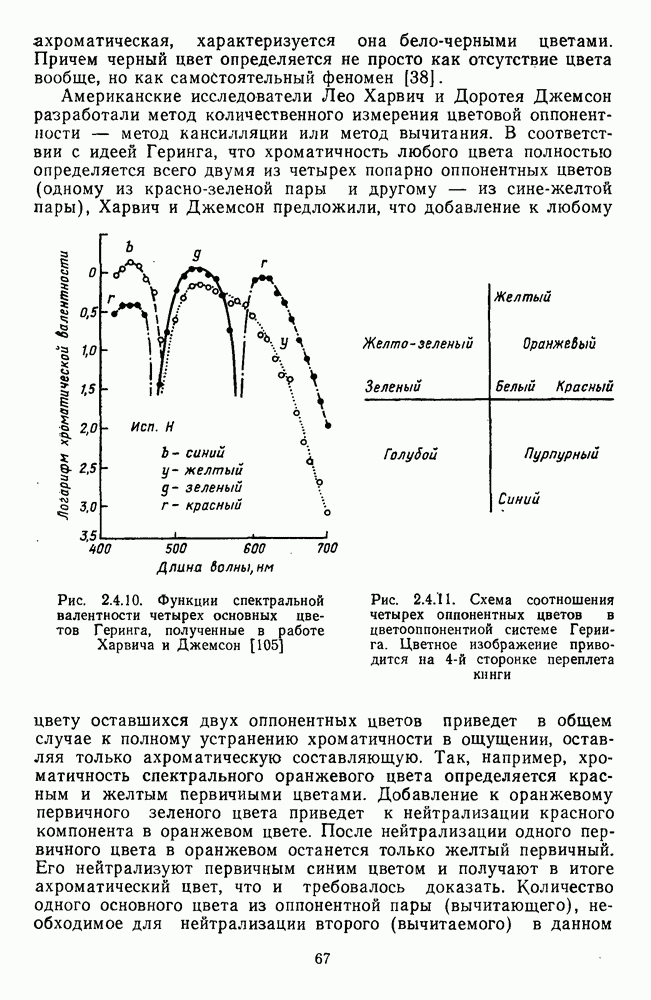
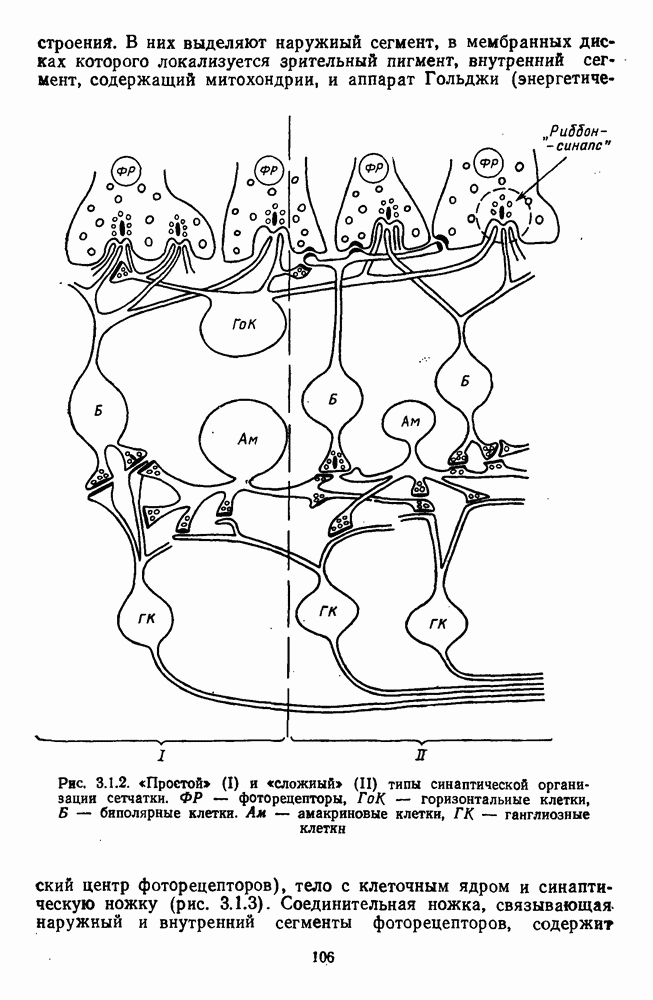
3.2.4. Ганглиозные клетки
Анализ излучения в сетчатке завершается в слое ганглиозных клеток, реакции которых, состоящие из последовательностей импульсов, передаются в подкорковые зрительные центры — верхнее двухолмие и наружное коленчатое тело.
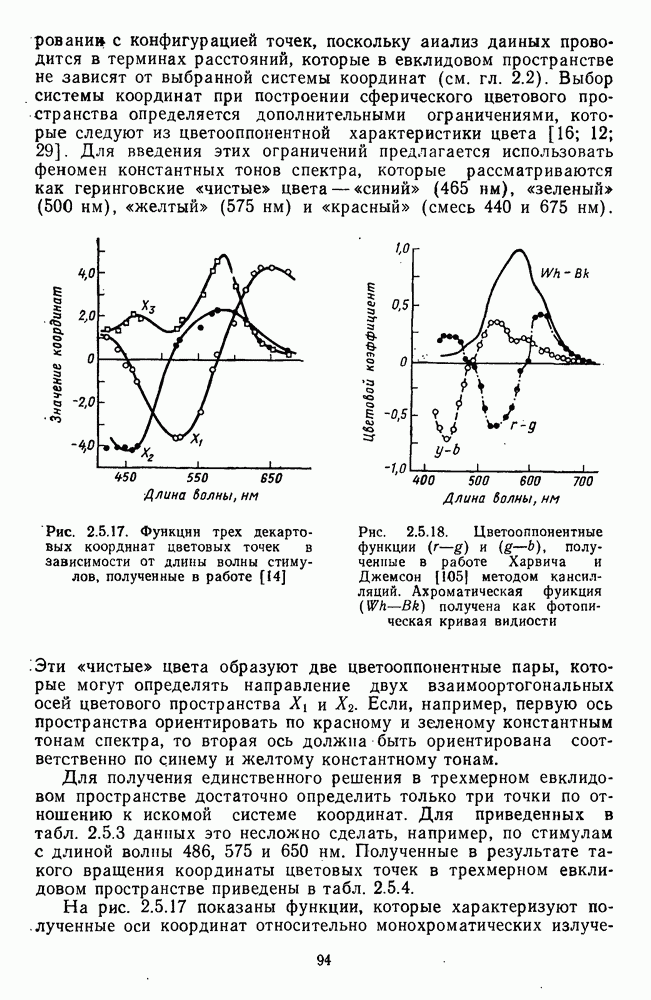
Морфологические типы ганглиозных клеток. В течение последних десятилетий проведено большое число исследований морфологии ганглиозных клеток сетчатки у животных разного уровня эволюционного развития [42]. Данные этих исследований предоставляют интерес в связи с интенсивным развитием в современных нейронауках структурно-функционального подхода,
основанного на выявлении закономерных связей между морфологией и функцией нейронных образований мозга.
Морфологические классификации клеток делаются на основе признаков, которые наиболее информативны с точки зрения описания возможных функций. Это: 1) геометрия дендритного дерева (форма, размах); 2) уровень ветвления дендритов во внутреннем плексиформном слое, от которого зависят их контакты с определенными типами нижележащих нейронов и соответственно-свойства рецептивных полей; 3) проекция аксонов в вышележащие структуры мозга.
Наиболее распространена морфологическая классификация ганглиозных клеток на  и
и  соответствующая физиологической классификации на
соответствующая физиологической классификации на  - и
- и  -тнпы. Существует, однако, ряд переходных форм, что говорит об определенной условности этой классификации ганглиозных клеток.
-тнпы. Существует, однако, ряд переходных форм, что говорит об определенной условности этой классификации ганглиозных клеток.
Электрофизиологическая классификация ганглиозных клеток. Существует три основные системы классификации ганглиозных: клеток:  -клетки;
-клетки;
2) фазические и тонические клетки;

Фазические клетки генерируют короткие по времени (фазные)  или
или  -разряды импульсов, а тонические клетки отвечают длительным изменением своей импульсной активности (возбуждением или торможением) в ответ на изменение интенсивности или спектрального состава излучения. В сетчатке обезьяны фазические клетки имеют концентрические рецептивные поля, центр и периферия которых имеют один и тот же колбочко-вый вход от
-разряды импульсов, а тонические клетки отвечают длительным изменением своей импульсной активности (возбуждением или торможением) в ответ на изменение интенсивности или спектрального состава излучения. В сетчатке обезьяны фазические клетки имеют концентрические рецептивные поля, центр и периферия которых имеют один и тот же колбочко-вый вход от  или
или  но не
но не  -колбочек [89]. Локализуются фазические клетки, как правило, на периферии сетчатки. Их аксоньр проецируются в крупноклеточные (магноцеллюлярные) слои НКТ, а аксоны тонических клеток — в мелкоклеточные (парвоцеллю-лярные) слои НКТ. Афферентация верхних бугров четверохолмия, где локализованы нейронные механизмы движений глаз, осуществляется исключительно фазическими клетками, которые, таким образом, выполняют специальные функции в окуломоторной: системе. Включенностью фазических клеток в систему детекции движения и управления движениями глаз объясняется, по-видимому, и тот факт, что скорость проведения возбуждения по аксону фазической клетки выше скорости проведения в аксоне спектрально-оппонентной тонической клетки [173].
-колбочек [89]. Локализуются фазические клетки, как правило, на периферии сетчатки. Их аксоньр проецируются в крупноклеточные (магноцеллюлярные) слои НКТ, а аксоны тонических клеток — в мелкоклеточные (парвоцеллю-лярные) слои НКТ. Афферентация верхних бугров четверохолмия, где локализованы нейронные механизмы движений глаз, осуществляется исключительно фазическими клетками, которые, таким образом, выполняют специальные функции в окуломоторной: системе. Включенностью фазических клеток в систему детекции движения и управления движениями глаз объясняется, по-видимому, и тот факт, что скорость проведения возбуждения по аксону фазической клетки выше скорости проведения в аксоне спектрально-оппонентной тонической клетки [173].
Классификация ганглиозных клеток на  и
и  -типы предложена Энрот-Кугелем и Робсоном по результатам работ на кошке [84]. Их отличителыше свойства представлены в табл. 3.2.2: Х-клетки во многом сходны с тоническими клетками, а У-клет-ки — с фазическими клетками сетчатки приматов. Предполагается соответствие между морфологическими классами
-типы предложена Энрот-Кугелем и Робсоном по результатам работ на кошке [84]. Их отличителыше свойства представлены в табл. 3.2.2: Х-клетки во многом сходны с тоническими клетками, а У-клет-ки — с фазическими клетками сетчатки приматов. Предполагается соответствие между морфологическими классами  а и у и. физиологическими типами
а и у и. физиологическими типами  и
и 
Существует еще одна классификация ганглиозных клеток — на А- и В-типы, основанная на характеристике входов: клетки;
Таблица 3.2.2. (см. скан) Ганглиозные клетки  и
и  -типов
-типов
А-типа получают сигналы преимущественно от биполяров, а В-ти-па — от биполяров и амакринов. Клетки А- и В-типов образуют два различных функциональных класса. А-клетки имеют небольшие по размеру рецептивные поля и обладают высокой абсолютной чувствительностью к свету, а В-клетки имеют сложные рецептивные поля с хорошо выраженной избирательностью к направлению движения стимула [144].
В терминологии перечисленных выше классификаций к спектральному анализу излучения имеют отношение следующие типы ганглиозных клеток: тонические on-, off- или  -клетки; клетки Х-типа и клетки А-типа.
-клетки; клетки Х-типа и клетки А-типа.
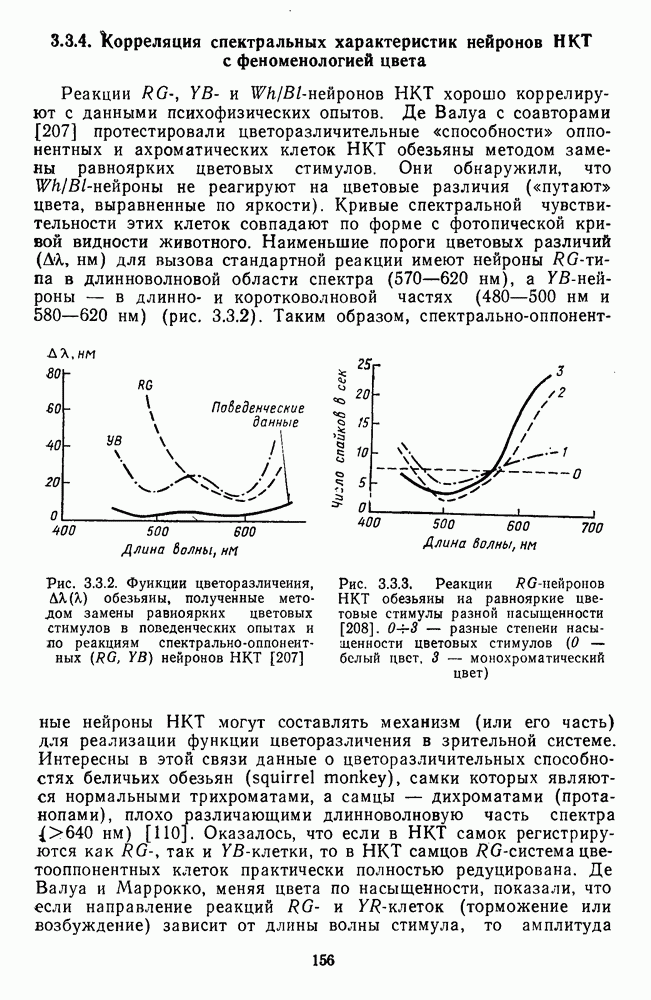
Кодирование цвета ганглиозными клетками сетчатки. Гранит (1955) первым использовал микроэлектроды для анализа механизмов кодирования цвета в слое ганглиозных клеток. Он выделил клетки, реагирующие на широкий спектр излучений (домина-торы), и клетки, селективно настроенные на узкий диапазон длин волн (модуляторы). Доминаторам приписывалась функция
поля которых устроены аналогично рецептивным полям биполяров с двойной спектральной оппонентностью. Клетки этого типа дают  -ответы на длинноволновый стимул и
-ответы на длинноволновый стимул и  -ответы на средневолновое освещение центра рецептивного поля и обратные реакции на аналогичную стимуляцию периферии [76; 148]. В сетчатке обезьяны описан небольшой процент клеток
-ответы на средневолновое освещение центра рецептивного поля и обратные реакции на аналогичную стимуляцию периферии [76; 148]. В сетчатке обезьяны описан небольшой процент клеток  селективно настроенных на выделение длинноволновых излучений [227]. Их спектральная чувствительность к длинноволновому свету в
селективно настроенных на выделение длинноволновых излучений [227]. Их спектральная чувствительность к длинноволновому свету в  раз превышает чувствительность к свету в средне- и коротковолновой частях спектра.
раз превышает чувствительность к свету в средне- и коротковолновой частях спектра.
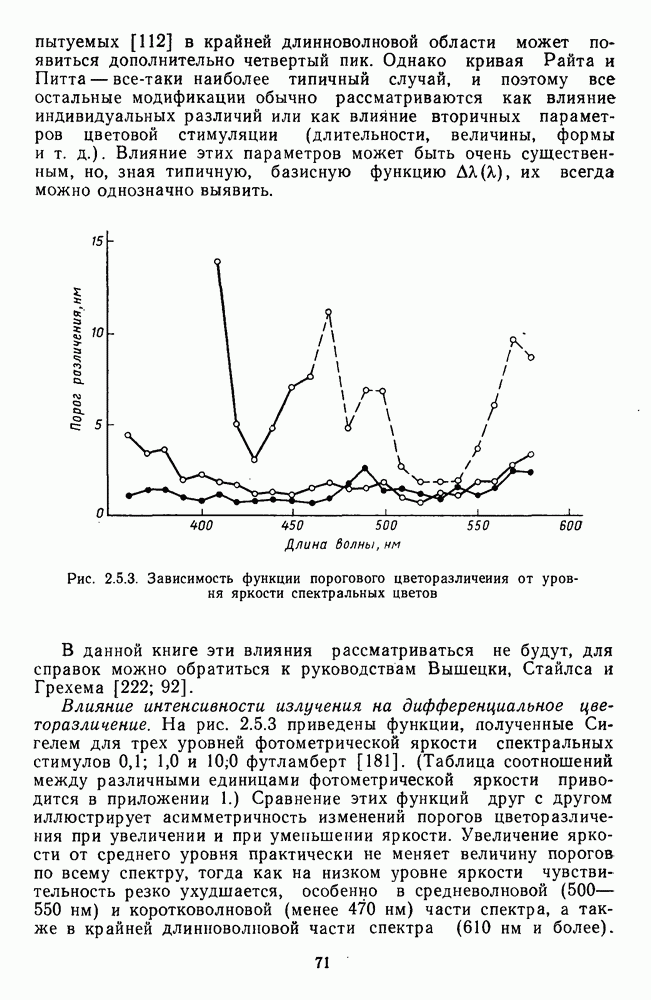
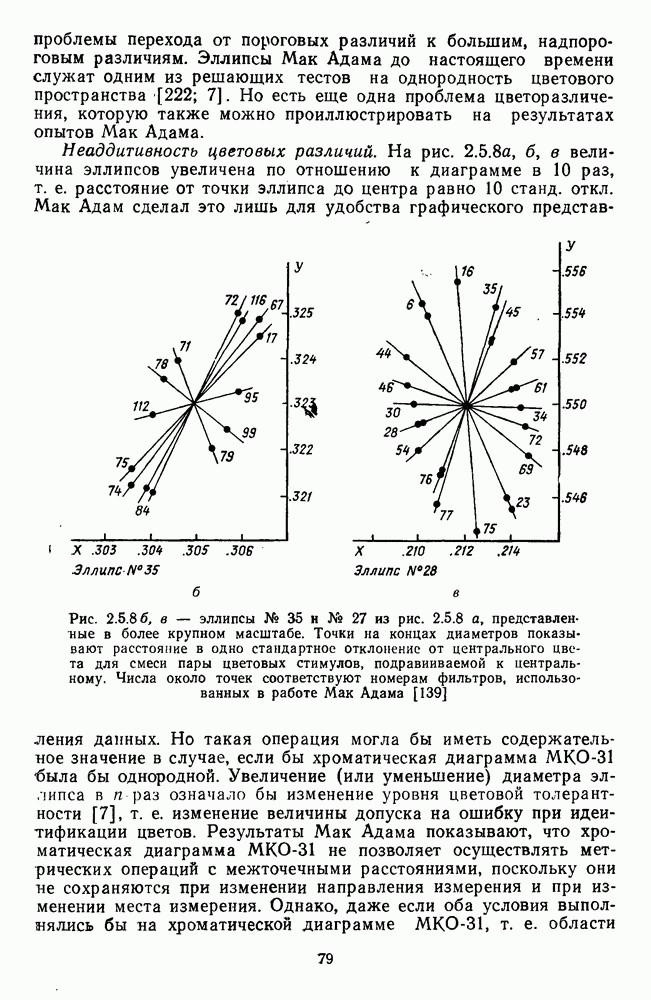
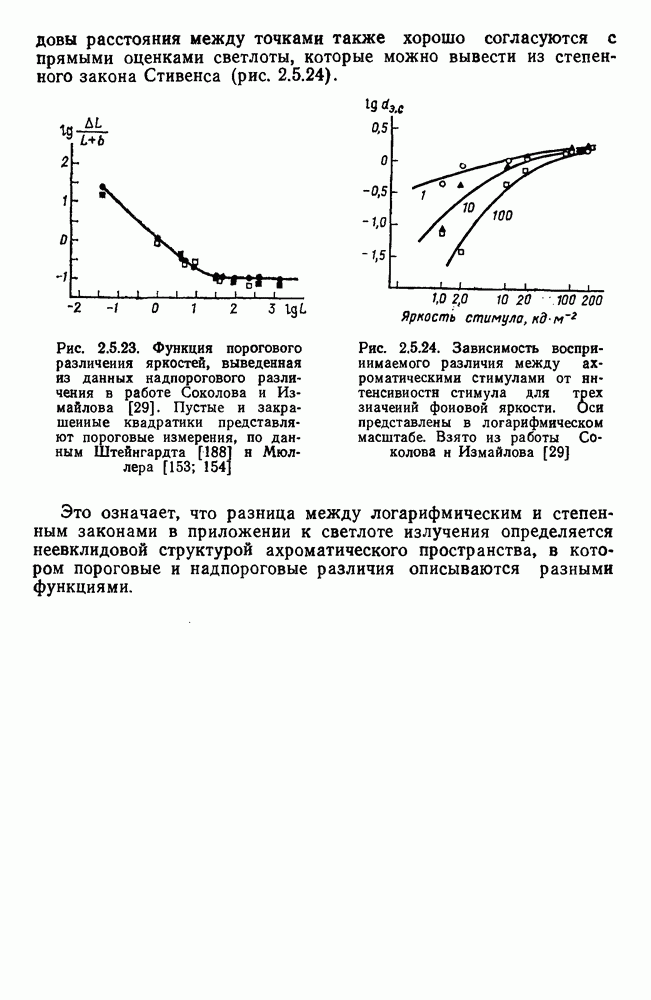
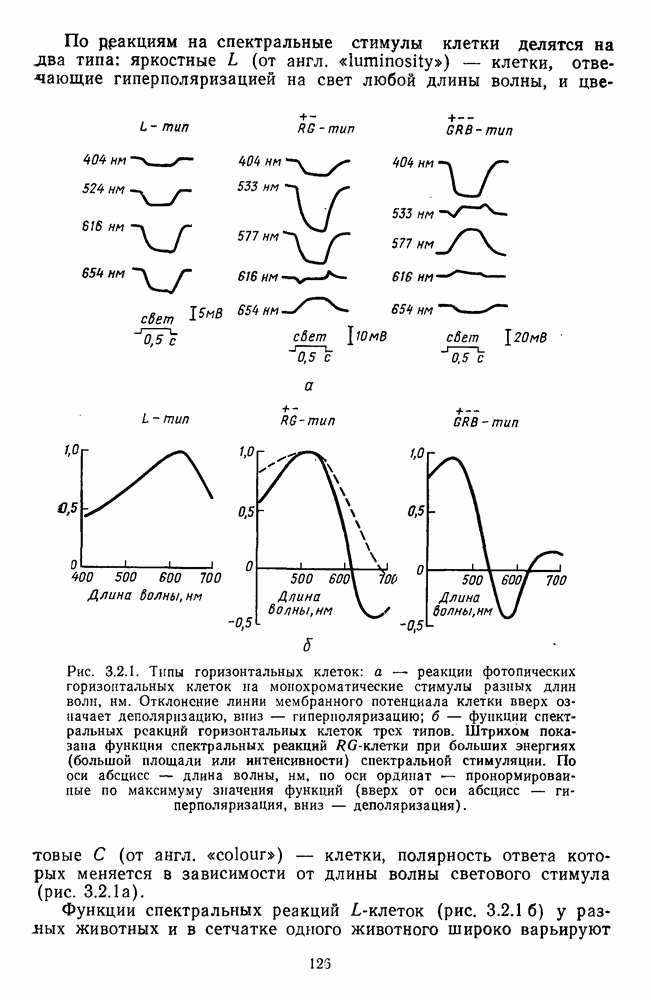
Аналогично цветовым горизонтальным и биполярным клеткам каждый тип оппонентных ганглиозных клеток характеризуется своей формой функций спектральных реакций, где информативным является положение максимумов и «нейтральной точки» (рис. 3.2.3а, б). В области нейтральной точки клетка меняет характер своих реакций с возбуждения на торможение или наоборот. В этой точке спектра реакции клеток практически не отличимы от фона, что свидетельствует об одинаковой выраженности здесь процессов возбуждения и торможения, запускаемых фоторецепторами разного типа.
Фоторецепторные входы ганглиозных клеток. Большая часть клеток  и
и  -типов имеют концентрически организованные рецептивные поля с делением на центр и периферию, которые получают сигналы от колбочек разного типа. В рецептивных полях
-типов имеют концентрически организованные рецептивные поля с делением на центр и периферию, которые получают сигналы от колбочек разного типа. В рецептивных полях  -клеток входами служат
-клеток входами служат  и
и  -колбочки, а в рецептивных полях
-колбочки, а в рецептивных полях  -клеток — с одной стороны, В-колбочки, а с другой —
-клеток — с одной стороны, В-колбочки, а с другой —  или
или  (или
(или  и
и  одновременно). У небольшой части клеток, роль которых в цветовом зрении неясна, центр рецептивного поля представлен
одновременно). У небольшой части клеток, роль которых в цветовом зрении неясна, центр рецептивного поля представлен  -колбочками, а периферия —
-колбочками, а периферия —  -колбочками [152].
-колбочками [152].
В сетчатке обезьяны оппонентные клетки  -типов составляют
-типов составляют  от общего числа ганглиозных клеток (остальные
от общего числа ганглиозных клеток (остальные  являются ахроматическими). В сетчатке карповых рыб доминируют
являются ахроматическими). В сетчатке карповых рыб доминируют  -клетки с двойной спектральной оппонентностью, которые у приматов регистрируются только на уровне коры [90].
-клетки с двойной спектральной оппонентностью, которые у приматов регистрируются только на уровне коры [90].
Среди оппонентных клеток с сооотношением  преобладают клетки с
преобладают клетки с  -центром по сравнению с
-центром по сравнению с  -центром. Однако в отдельном локусе сетчатки регистрируются, как правило, оппонентные клетки всех типов [227]. Размеры центра рецептивного поля
-центром. Однако в отдельном локусе сетчатки регистрируются, как правило, оппонентные клетки всех типов [227]. Размеры центра рецептивного поля  -клеток варьируют в пределах
-клеток варьируют в пределах  -клеток — в пределах
-клеток — в пределах  -клеток латентный период реакций в центре рецептивного поля составляет
-клеток латентный период реакций в центре рецептивного поля составляет  а на периферии —
а на периферии —  Латентность реакций
Латентность реакций  -клеток соответственно на
-клеток соответственно на  больше. Кроме того, у
больше. Кроме того, у  -клеток наблюдаются длительные разряды последствия (200-400 мс). Все это ограничивает временные разрешающие способности
-клеток наблюдаются длительные разряды последствия (200-400 мс). Все это ограничивает временные разрешающие способности  -канала: так, ганглиозные клетки
-канала: так, ганглиозные клетки  -типа «отслеживают» мелькания света с частотой 60-70 Гц, а
-типа «отслеживают» мелькания света с частотой 60-70 Гц, а  -типа — с частотой 35-40 Гц [227].
-типа — с частотой 35-40 Гц [227].
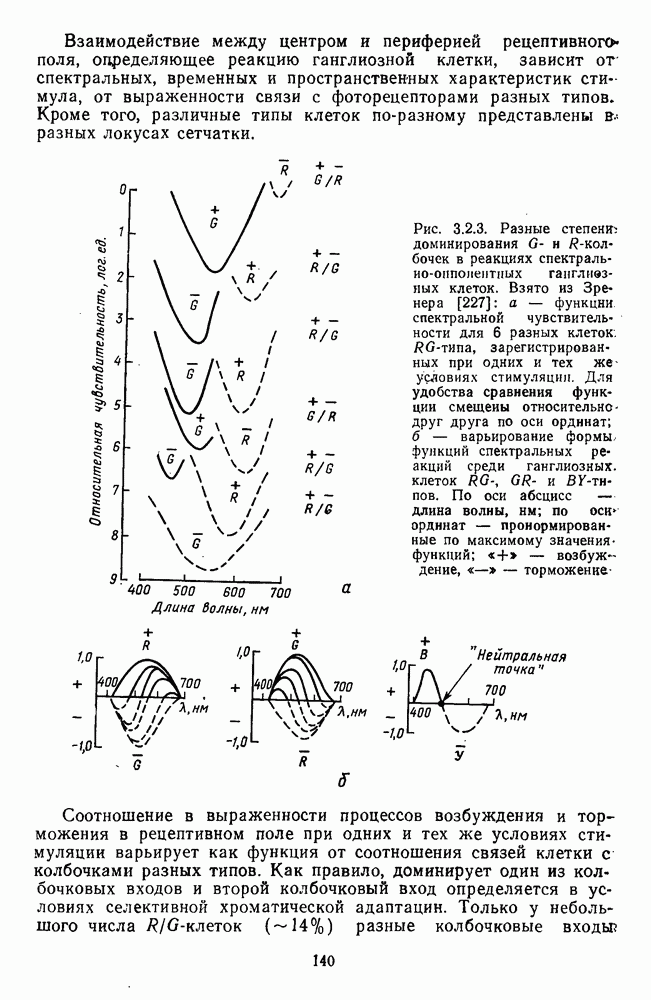
Взаимодействие между центром и периферией рецептивного поля, определяющее реакцию ганглиозной клетки, зависит  спектральных, временных и пространственных характеристик стимула, от выраженности связи с фоторецепторами разных типов. Кроме того, различные типы клеток по-разному представлены
спектральных, временных и пространственных характеристик стимула, от выраженности связи с фоторецепторами разных типов. Кроме того, различные типы клеток по-разному представлены  разных локусах сетчатки.
разных локусах сетчатки.
Рис. 3.2.3. (см. скан) Разные степени: доминирования  и
и  -колбочек в реакциях спектрально-оппонентных ганглиозных клеток. Взято из Зренера [227]: а — функции спектральной чувствительности для 6 разных клеток,
-колбочек в реакциях спектрально-оппонентных ганглиозных клеток. Взято из Зренера [227]: а — функции спектральной чувствительности для 6 разных клеток,  -типа, зарегистрированных при одних и тех же условиях стимуляции. Для удобства сравнения функции смещены относительно друг друга по оси ординат;
-типа, зарегистрированных при одних и тех же условиях стимуляции. Для удобства сравнения функции смещены относительно друг друга по оси ординат;  — варьирование формы функций спектральных реакций среди ганглиозных, клеток
— варьирование формы функций спектральных реакций среди ганглиозных, клеток  -типов. По оси абсцисс — длина волны, нм; по
-типов. По оси абсцисс — длина волны, нм; по  ординат — пронормированные по максимому значения функций; «+» — возбуждение, «-» — торможение.
ординат — пронормированные по максимому значения функций; «+» — возбуждение, «-» — торможение.
Соотношение в выраженности процессов возбуждения и торможения в рецептивном поле при одних и тех же условиях стимуляции варьирует как функция от соотношения связей клетки с колбочками разных типов. Как правило, доминирует один из колбочковых входов и второй колбочковый вход определяется в условиях селективной хроматической адаптации. Только у небольшого числа  -клеток
-клеток  разные колбочковые входы?
разные колбочковые входы?
представлены в РП одинаково эффективно [227]. Степень доминирования того или иного входа определяет форму функций спектральных реакций клетки. Так, при доминировании  -колбочек нейтральная точка функции клетки
-колбочек нейтральная точка функции клетки  -типа локализуется в области 480 нм, а при доминировании
-типа локализуется в области 480 нм, а при доминировании  -колбочек — в области 630 нм. Наличие разных степеней доминирования между колбочками разных типов объясняет практически непрерывную градацию цветооппонентности среди клеток
-колбочек — в области 630 нм. Наличие разных степеней доминирования между колбочками разных типов объясняет практически непрерывную градацию цветооппонентности среди клеток  -типа (рис. 3.2.3 6). Одновременно варьируют максимумы функции спектральных реакций: при доминировании
-типа (рис. 3.2.3 6). Одновременно варьируют максимумы функции спектральных реакций: при доминировании  -колбочек
-колбочек  -клеток может формироваться
-клеток может формироваться  -оппонентность, и только хроматическая адаптация позволяет выяснить, образуется ли эта оппонентность с участием
-оппонентность, и только хроматическая адаптация позволяет выяснить, образуется ли эта оппонентность с участием  -колбочек. Такая гетерогенность клеток
-колбочек. Такая гетерогенность клеток  -тцпа может служить механизмом усиления локальных перепадов слабой яркости, образованных двумя соседними областями разного цвета. В отличие от клеток
-тцпа может служить механизмом усиления локальных перепадов слабой яркости, образованных двумя соседними областями разного цвета. В отличие от клеток  -типа
-типа  -клетки не обнаруживают заметной вариабельности в коэффициентах связи с колбочками и их функции спектральных реакций стабильны.
-клетки не обнаруживают заметной вариабельности в коэффициентах связи с колбочками и их функции спектральных реакций стабильны.
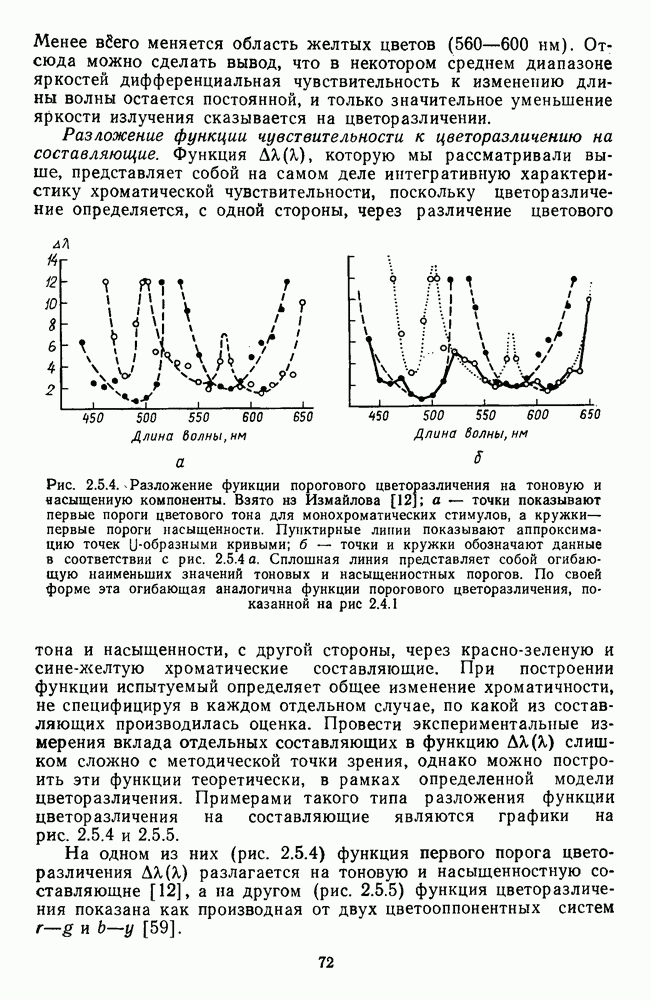
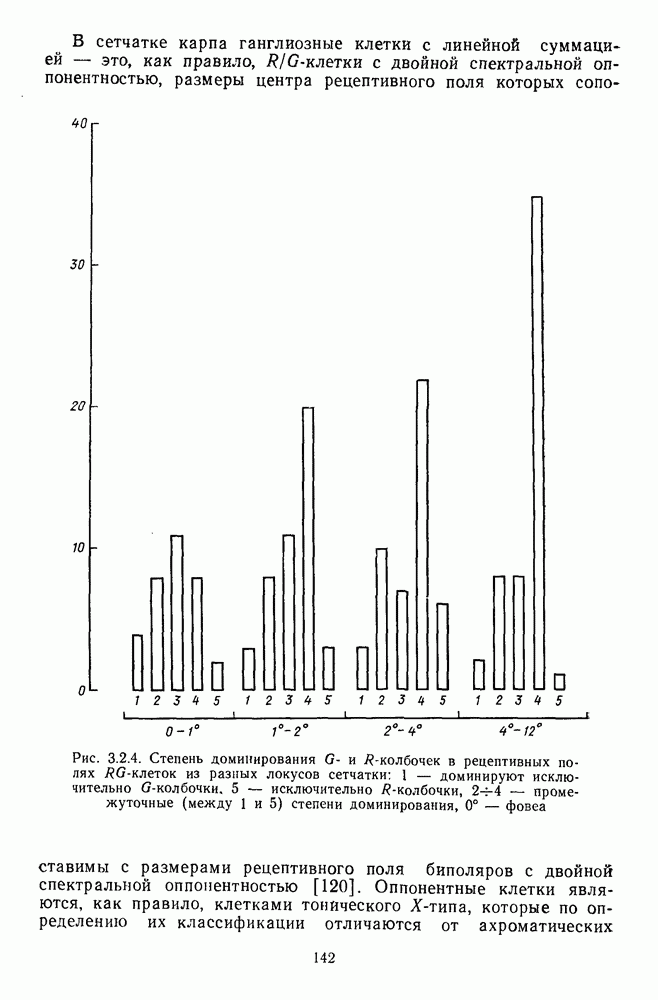
В каждом локусе сетчатки представлены  -клетки с разным соотношением вкладов
-клетки с разным соотношением вкладов  и
и  -колбочек. Однако степень доминирования тех или иных колбочек в рецептивных полях клеток меняется систематическим образом от фовеа к периферии: в центре преобладают
-колбочек. Однако степень доминирования тех или иных колбочек в рецептивных полях клеток меняется систематическим образом от фовеа к периферии: в центре преобладают  -клетки, реакции которых определяются в основном
-клетки, реакции которых определяются в основном  -колбочками, а на периферии —
-колбочками, а на периферии —  -клетки, в реакциях которых доминируют вклады от
-клетки, в реакциях которых доминируют вклады от  -колбочек (рис. 3.2.4). И хотя
-колбочек (рис. 3.2.4). И хотя  -колбочки доминируют по всей сетчатке в количественном отношении, это не означает их доминирование в функциональном смысле [142].
-колбочки доминируют по всей сетчатке в количественном отношении, это не означает их доминирование в функциональном смысле [142].
По небольшому числу данных, форма функции спектральных реакций и абсолютная чувствительность  -клеток не меняются с изменением локализации в сетчатке [227].
-клеток не меняются с изменением локализации в сетчатке [227].
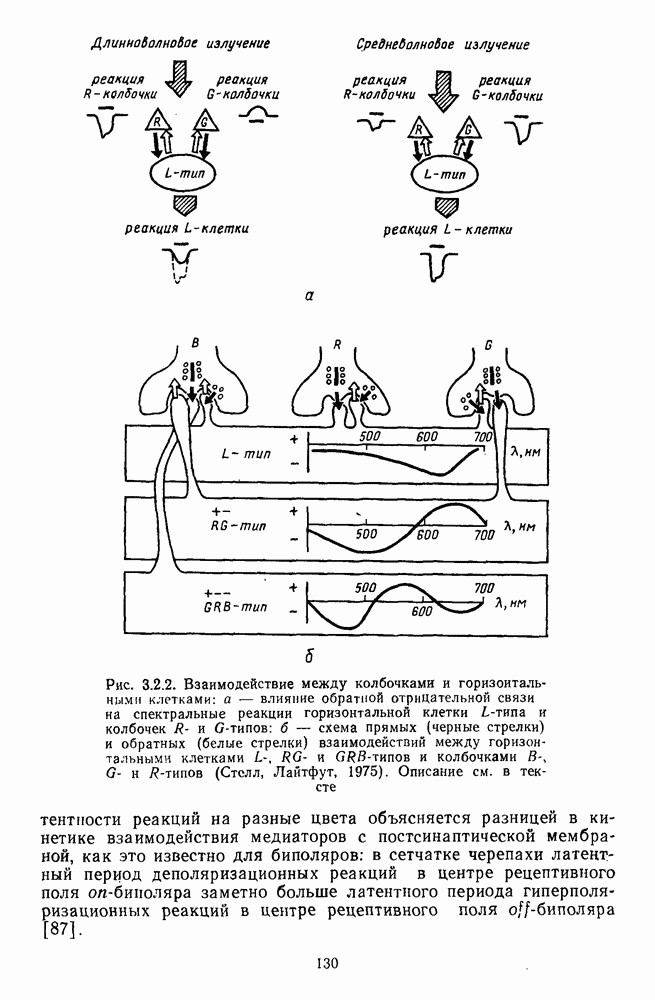
Динамические характеристики рецептивных полей ганглиозных клеток. Спектральная оппонентность ганглиозных клеток зависит от размеров цветового сигнала. При небольших размерах стимула, сравнимых с центром рецептивного поля клетки, оппонентность нарушается и клетка становится ахроматической (не-оппонентной) с доминированием в реакциях вклада колбочек одного типа в центре рецептивного поля. Ослабление онпонентности происходит и при стимуляции цветами небольших интенсивностей, вызывающих реакции колбочек одного типа преимущественно в центре рецептивного поля, где клетка имеет более высокую абсолютную чувствительность по сравнению с периферией.
Как суммируются сигналы от колбочек разного типа в площади рецептивного поля ганглиозной клетки? По данным Дюк и Спекройзе (1984), такая суммация у большей части  -клеток посит линейный характер. Это означает, что реакцию клетки
-клеток посит линейный характер. Это означает, что реакцию клетки  на комбинацию цветовых стимулов
на комбинацию цветовых стимулов  и можно воспроизвести простой суперпозицией реакций отдельно на
и можно воспроизвести простой суперпозицией реакций отдельно на 

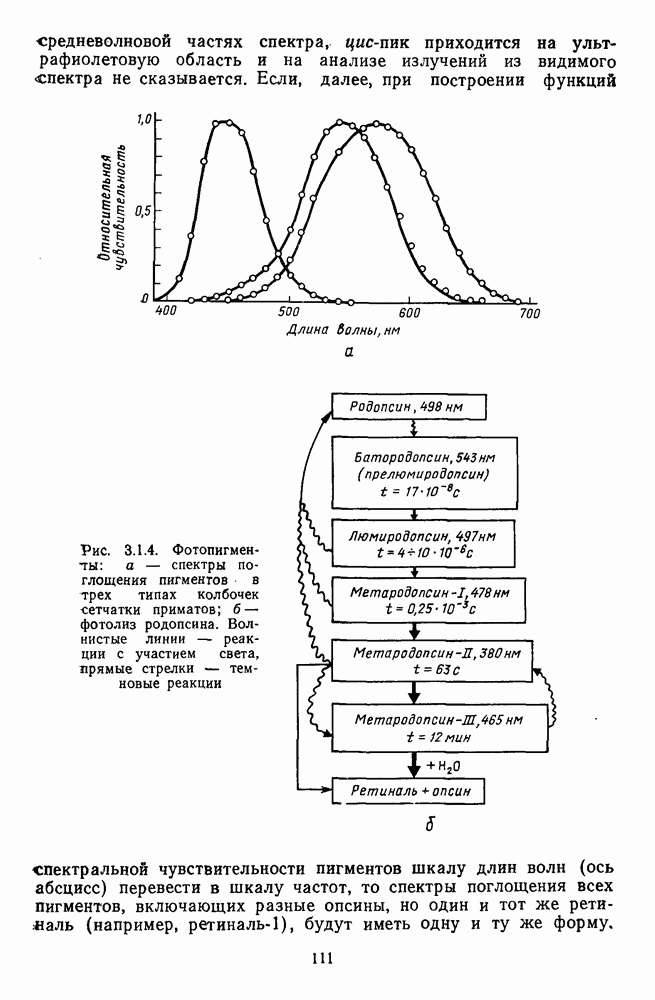
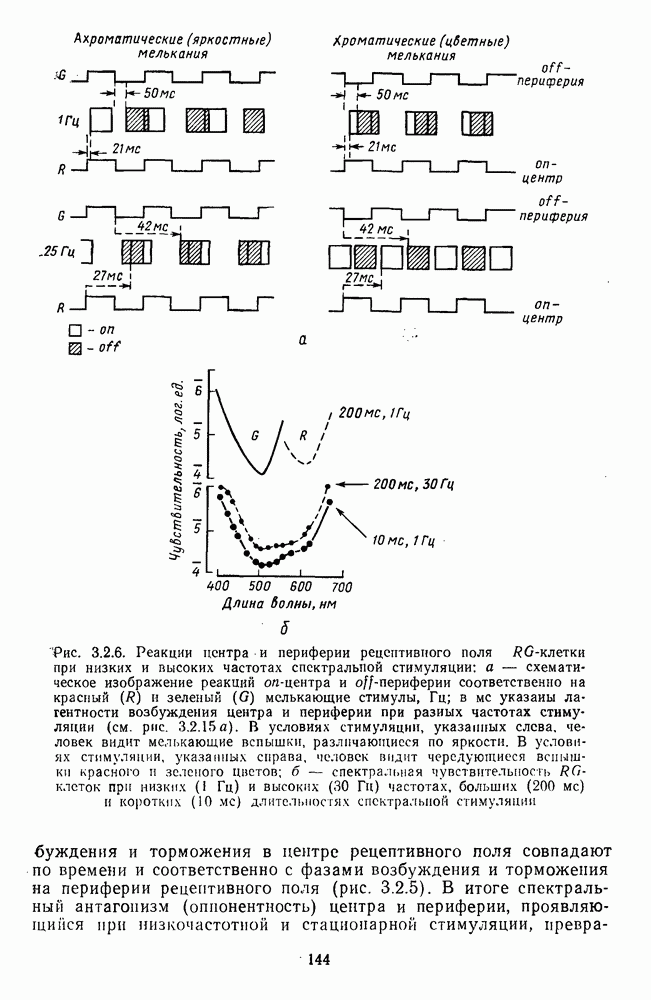
Рис. 3.2.6. (см. скан) Реакции центра и периферии рецеитивиого поля 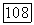 -клетки при низких и высоких частотах спектральной стимуляции: а — схематическое изображение реакций
-клетки при низких и высоких частотах спектральной стимуляции: а — схематическое изображение реакций 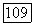 -центра и
-центра и 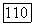 -периферии соответственно на красный (R) и зеленый (G) мелькающие стимулы, Гц; в мс указаны латентности возбуждения центра и периферии при разных частотах стимуляции (см. рис. 3.2.15а). В условиях стимуляции, указанных слева, человек видит мелькающие вспышки, различающиеся по яркости. В условиях стимуляции, указанных справа, человек видит чередующиеся вспышки красного и зеленого цветов; б - спектральная чувствительность RG-клеток при низких (I Гц) и высоких (30 Гц) частотах, больших (200 мс) и коротких (10 мс) длительностях спектральной стимуляции
-периферии соответственно на красный (R) и зеленый (G) мелькающие стимулы, Гц; в мс указаны латентности возбуждения центра и периферии при разных частотах стимуляции (см. рис. 3.2.15а). В условиях стимуляции, указанных слева, человек видит мелькающие вспышки, различающиеся по яркости. В условиях стимуляции, указанных справа, человек видит чередующиеся вспышки красного и зеленого цветов; б - спектральная чувствительность RG-клеток при низких (I Гц) и высоких (30 Гц) частотах, больших (200 мс) и коротких (10 мс) длительностях спектральной стимуляции
возбуждения и торможения в центре рецептивного поля совпадают по времени и соответственно с фазами возбуждения и торможения на периферии рецептивного поля (рис. 3.2.5). В итоге спектральный антагонизм (опнонентность) центра и периферии, проявляющийся при низкочастотной и стационарной стимуляции,
превращается в синергизм при высоких частотах стимуляции. Оппонентная клетка теряет спектральную избирательность и становится ахроматической (рис. 3.2.6). Оптимальные условия синхронизации в работе центра и периферии рецептивного поля индивидуальны для каждой оппонентной клетки и зависят от интенсивности и спектрального состава фонового освещения, от цвета тестовых стимулов. Перестройка временных взаимодействий в рецептивных полях оппонентных клеток при высоких частотах стимуляции может иметь адаптивное значение [227]. Стационарный объект (вариант низкочастотной стимуляции) выделяется из равного по яркости фонового окружения путем анализа цветовых различий с участием оппонентных нейронов. При наблюдении быстродвижущегося объекта первостепенное значение приобретает анализ яркостных различий, к выделению которых в этой ситуации и подключается наряду с системой К-нейронов система оппонентных ганглиозных клеток. Таким образом, спектрально-оппонентные ганглиозные клетки обнаруживают функциональную пластичность: в зависимости от условий восприятия они могут участвовать в выделении как цветовых (стационарный объект), так и яркостных (движущийся объект) контрастов. Это объясняет, почему движущийся объект видится нами бесцветным.
Ахроматические спектрально-неоппонентные ганглиозные клетки. В сетчатке обезьяны около 40% ганглиозных клеток не обнаруживают спектральной оппонентности [227]. Часть из них являются фазическими (25%), часть — тоническими 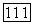 Функции спектральной чувствительности и тонических и фазических ахроматических клеток широкоплосны с пиком на длине волны 550 нм. До настоящего времени неясно, с колбочками каких типов связаны эти клетки — только с
Функции спектральной чувствительности и тонических и фазических ахроматических клеток широкоплосны с пиком на длине волны 550 нм. До настоящего времени неясно, с колбочками каких типов связаны эти клетки — только с 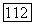 только с
только с 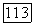 или с
или с 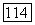 и
и 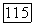 одновременно. При отсутствии спектральной оппонентности ахроматические клетки обнаруживают пространственную оппонентность в рецептивном поле, где центр и периферия запускаются колбочками одного типа (например,
одновременно. При отсутствии спектральной оппонентности ахроматические клетки обнаруживают пространственную оппонентность в рецептивном поле, где центр и периферия запускаются колбочками одного типа (например, 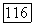 или
или 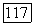
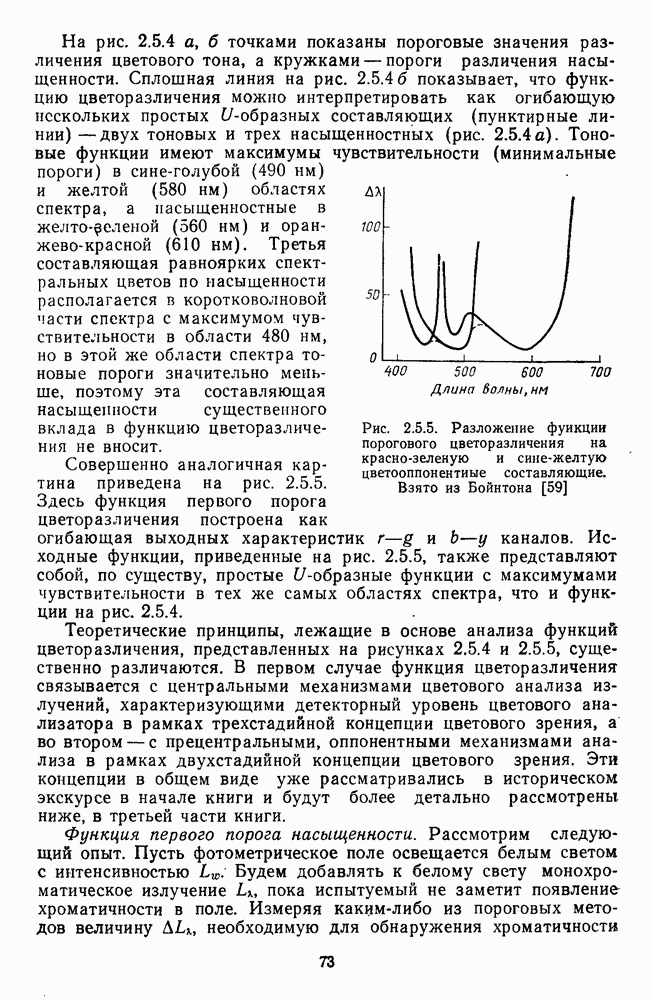
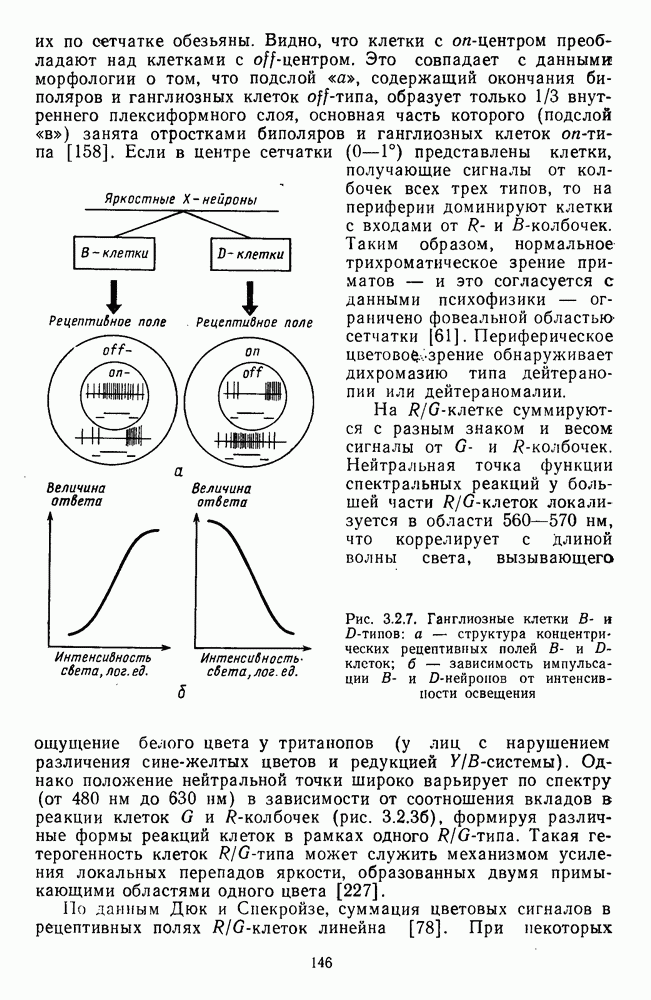
Среди ахроматических клеток тонического типа выделяют 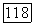 -нейроны, возбуждающиеся при увеличении интенсивности света, и
-нейроны, возбуждающиеся при увеличении интенсивности света, и 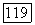 -нейроны, возбуждающиеся при затемнении (от англ. «bright» — световой и «dark» — темновой) [116; 117]. При постепенном увеличении интенсивности освещения частота импульсации В-нейронов монотонно увеличивается, а
-нейроны, возбуждающиеся при затемнении (от англ. «bright» — световой и «dark» — темновой) [116; 117]. При постепенном увеличении интенсивности освещения частота импульсации В-нейронов монотонно увеличивается, а 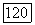 -нейронов монотонно уменьшается (рис. 3.2.7). Каждому участку сетчатки соответствует свой набор В- и
-нейронов монотонно уменьшается (рис. 3.2.7). Каждому участку сетчатки соответствует свой набор В- и 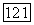 -нейронов. В итоге в любом локусе сетчатки действие света вызывает одновременное возбуждение .В-нейронов и торможение
-нейронов. В итоге в любом локусе сетчатки действие света вызывает одновременное возбуждение .В-нейронов и торможение 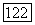 -нейронов, а темнота — возбуждение
-нейронов, а темнота — возбуждение 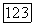 -нейронов и торможение В-нейронов. Ответы В- и
-нейронов и торможение В-нейронов. Ответы В- и 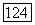 -нейронов устойчивы и сохраняются в течение всего времени действия стимула.
-нейронов устойчивы и сохраняются в течение всего времени действия стимула.
На рис. 3.28 представлены все типы описанных выше хроматических и ахроматических клеток в соответствии с распределением
их по сетчатке обезьяны. Видно, что клетки с 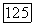 -центром преобладают над клетками с
-центром преобладают над клетками с 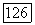 -центром. Это совпадает с данными морфологии о том, что подслой
-центром. Это совпадает с данными морфологии о том, что подслой 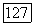 содержащий окончания биполяров и ганглиозных клеток
содержащий окончания биполяров и ганглиозных клеток 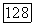 -типа, образует только 1/3 внутреннего плексиформного слоя, основная часть которого (подслой «в») занята отростками биполяров и ганглиозных клеток
-типа, образует только 1/3 внутреннего плексиформного слоя, основная часть которого (подслой «в») занята отростками биполяров и ганглиозных клеток 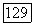 Если в центре сетчатки
Если в центре сетчатки 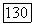 представлены клетки, получающие сигналы от колбочек всех трех типов, то на периферии доминируют клетки с входами от
представлены клетки, получающие сигналы от колбочек всех трех типов, то на периферии доминируют клетки с входами от 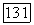 и В-колбочек. Таким образом, нормальное трихроматическое зрение приматов — и это согласуется с данными психофизики — ограничено фовеальной областью сетчатки [61]. Периферическое цветовое зрение обнаруживает дихромазию типа дейтеранопии или дейтераномалии.
и В-колбочек. Таким образом, нормальное трихроматическое зрение приматов — и это согласуется с данными психофизики — ограничено фовеальной областью сетчатки [61]. Периферическое цветовое зрение обнаруживает дихромазию типа дейтеранопии или дейтераномалии.

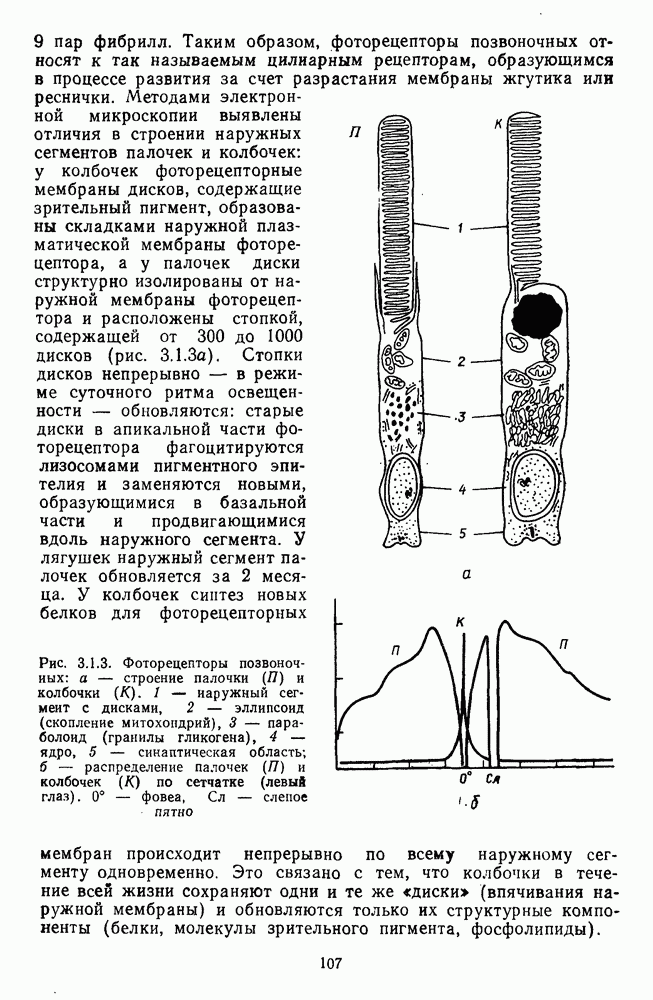
Рис. 3.2.7. Ганглиозные клетки В- и  -типов: а — структура концентрических рецептивных полей В- и
-типов: а — структура концентрических рецептивных полей В- и  -клеток;
-клеток;  — зависимость импульсации В- и
— зависимость импульсации В- и  -нейронов от интенсивности освещения
-нейронов от интенсивности освещения
На  -клетке суммируются с разным знаком и весом сигналы от
-клетке суммируются с разным знаком и весом сигналы от 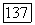 и
и 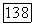 -колбочек. Нейтральная точка функции спектральных реакций у большей части
-колбочек. Нейтральная точка функции спектральных реакций у большей части 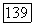 -клеток локализуется в области 560—570 нм, что коррелирует с длиной волны света, вызывающего ощущение белого цвета у тританопов (у лиц с нарушением различения сине-желтых цветов и редукцией
-клеток локализуется в области 560—570 нм, что коррелирует с длиной волны света, вызывающего ощущение белого цвета у тританопов (у лиц с нарушением различения сине-желтых цветов и редукцией 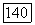 -системы). Однако положение нейтральной точки широко варьирует по спектру (от 480 нм до 630 нм) в зависимости от соотношения вкладов в реакции клеток
-системы). Однако положение нейтральной точки широко варьирует по спектру (от 480 нм до 630 нм) в зависимости от соотношения вкладов в реакции клеток 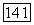 и
и 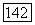 -колбочек (рис. 3.2.36), формируя различные формы реакций клеток в рамках одного
-колбочек (рис. 3.2.36), формируя различные формы реакций клеток в рамках одного 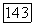 -типа. Такая гетерогенность клеток
-типа. Такая гетерогенность клеток 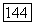 -типа может служить механизмом усиления локальных перепадов яркости, образованных двумя примыкающими областями одного цвета [227].
-типа может служить механизмом усиления локальных перепадов яркости, образованных двумя примыкающими областями одного цвета [227].
По данным Дюк и Снекройзе, суммация цветовых сигналов в рецептивных полях 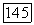 -клеток линейна [78]. При некоторых
-клеток линейна [78]. При некоторых
уровнях фонового освещения у части 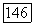 -клеток обнаруживаются отклонения от линейности, проявляющиеся в эффектах «адаптационного торможения» и «адаптационного растормаживания» [32; 144]. Освещение периферии ганглиозной клетки цветом, который тормозит ее, растормаживает (облегчает) ответ центральной зоны, если он был заторможен до этого фоновым цветом в центре поля («адаптационное растормаживание»). Если один участок центра освещен ранее, а второй участок освещается позднее, то ответ на засвет второго локуса уменьшается («адаптационное торможение»). Процессы адаптации усложняют антагонистические отношения между центром и периферией рецептивного поля, вызывая отклонения от линейности в суммации цветовых сигналов.
-клеток обнаруживаются отклонения от линейности, проявляющиеся в эффектах «адаптационного торможения» и «адаптационного растормаживания» [32; 144]. Освещение периферии ганглиозной клетки цветом, который тормозит ее, растормаживает (облегчает) ответ центральной зоны, если он был заторможен до этого фоновым цветом в центре поля («адаптационное растормаживание»). Если один участок центра освещен ранее, а второй участок освещается позднее, то ответ на засвет второго локуса уменьшается («адаптационное торможение»). Процессы адаптации усложняют антагонистические отношения между центром и периферией рецептивного поля, вызывая отклонения от линейности в суммации цветовых сигналов.

Рис. 3.2.8. Распределение спектрально-оппонентных и ахроматических ганглиозных клеток по сетчатке обезьяны (Зреннер, 1983). Заштрихованные блоки — клетки с 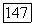 -центром рецептивного поля
-центром рецептивного поля 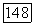 белые блоки — клетки с
белые блоки — клетки с 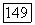 -центром (—), блоки с точками — клетки, у которых не удалось определить центр и которые реагировали возбуждением в любой точке своего
-центром (—), блоки с точками — клетки, у которых не удалось определить центр и которые реагировали возбуждением в любой точке своего 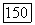 и Я-типы колбочек, доминирующих в центре РП. Общее число клеток, по которому составлена диаграмма, — 385. Эксц. — эксцентриситет на сетчатке, в градусах (0° — фовеа)
и Я-типы колбочек, доминирующих в центре РП. Общее число клеток, по которому составлена диаграмма, — 385. Эксц. — эксцентриситет на сетчатке, в градусах (0° — фовеа)
Временные взаимодействия между центром и периферией рецептивного поля играют существенную роль в формировании спектральных реакций 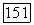 -клеток на мелькающие цветовые стимулы, имитирующие в эксперименте смещения цветного изображения при движении глаз. Они объясняют возникновение у человека ощущений цвета в условиях гетерохромной фликерметрии (рис. 3.2.6). Так как нарушение спектральной оппонентности ганглиозных клеток при высоких частотах стимуляции связано с синергизмом в работе центра и периферии рецептивного поля, то логично думать, что стимульные условия, десинхронизирующие их взаимодействие, будут приводить к возникновению ощущения
-клеток на мелькающие цветовые стимулы, имитирующие в эксперименте смещения цветного изображения при движении глаз. Они объясняют возникновение у человека ощущений цвета в условиях гетерохромной фликерметрии (рис. 3.2.6). Так как нарушение спектральной оппонентности ганглиозных клеток при высоких частотах стимуляции связано с синергизмом в работе центра и периферии рецептивного поля, то логично думать, что стимульные условия, десинхронизирующие их взаимодействие, будут приводить к возникновению ощущения
цвета. Действительно, такое наблюдается при вращении диска «Фехнера — Бенхэма». Черно-белые полосы диска при вращении с частотой 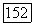 Гц вызывают ощущения разных цветов. Механизмом этой иллюзии может быть десинхронизация в реакциях центра и периферии рецептивного поля (фазовый сдвиг) оппонентной ганглиозной клетки, вызываемая полосами разной длины и ширины и аналогичная той, что наблюдается при низкочастотной гетерохромной стимуляции (рис. 3.2.6) [227].
Гц вызывают ощущения разных цветов. Механизмом этой иллюзии может быть десинхронизация в реакциях центра и периферии рецептивного поля (фазовый сдвиг) оппонентной ганглиозной клетки, вызываемая полосами разной длины и ширины и аналогичная той, что наблюдается при низкочастотной гетерохромной стимуляции (рис. 3.2.6) [227].
На 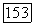 -клетках антагонистически взаимодействуют сигналы от колбочек
-клетках антагонистически взаимодействуют сигналы от колбочек 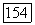 -типа, с одной стороны, и
-типа, с одной стороны, и 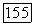 или
или 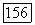 -типов (или
-типов (или 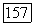 -типа одновременно) — с другой. В сетчатке приматов клетки этого типа редки
-типа одновременно) — с другой. В сетчатке приматов клетки этого типа редки 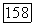 Их рецептивные поля, как правило, не имеют деления на центр и периферию. У большинства
Их рецептивные поля, как правило, не имеют деления на центр и периферию. У большинства 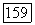 -клеток
-клеток 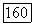 вход от рецепторов В-типа — возбуждающий [227]. Нейтральная точка функций спектральных реакций
вход от рецепторов В-типа — возбуждающий [227]. Нейтральная точка функций спектральных реакций 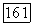 -клеток локализуется в области 490—500 нм, соответствующей области белого цвета для протанопов и дейтеранопов (редукция
-клеток локализуется в области 490—500 нм, соответствующей области белого цвета для протанопов и дейтеранопов (редукция 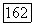 -системы). По сравнению с .
-системы). По сравнению с .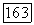 -клетками клетки
-клетками клетки 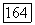 -типа генерируют длиннолатентные реакции на включение света и длительные разряды последействия на выключение света. В отличие от
-типа генерируют длиннолатентные реакции на включение света и длительные разряды последействия на выключение света. В отличие от 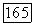 -клеток их функции спектральных реакций стабильны и не меняются с эксцентриситетом на сетчатке (рис. 3.2.3). Большая латентность реакций
-клеток их функции спектральных реакций стабильны и не меняются с эксцентриситетом на сетчатке (рис. 3.2.3). Большая латентность реакций 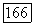 -клеток проявляется в психофизических опытах по наблюдению движущейся полоски, одна половина которой голубого цвета, а другая — красного: возникает ощущение, что синяя часть отстает от красной [151]. Временными параметрами реакций
-клеток проявляется в психофизических опытах по наблюдению движущейся полоски, одна половина которой голубого цвета, а другая — красного: возникает ощущение, что синяя часть отстает от красной [151]. Временными параметрами реакций 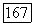 -клеток определяются особенности временной суммации сигналов в их рецептивных полях: если
-клеток определяются особенности временной суммации сигналов в их рецептивных полях: если 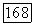 -клетки «отлеживают» вспышки света с частотой до 60—70 Гц, то
-клетки «отлеживают» вспышки света с частотой до 60—70 Гц, то 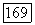 -клетки — с частотой до 35 Гц [91]. Особенности
-клетки — с частотой до 35 Гц [91]. Особенности 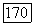 -системы ганглиозных клеток связывают с особенностями В-колбочек, отличающими их от колбочек
-системы ганглиозных клеток связывают с особенностями В-колбочек, отличающими их от колбочек 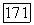 и
и 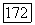 -типов и сближающими с палочками [227].
-типов и сближающими с палочками [227].
Таким образом, 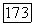 -система ганглиозных клеток обладает рядом специфических черт по сравнению с
-система ганглиозных клеток обладает рядом специфических черт по сравнению с 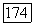 -системой. Если считать, что передача информации связана с фазой возбуждения нейрона, то
-системой. Если считать, что передача информации связана с фазой возбуждения нейрона, то 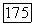 -клетки передают информацию только о синем цвете, так как являются в своем большинстве клетками
-клетки передают информацию только о синем цвете, так как являются в своем большинстве клетками 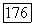 -типа. И тогда сигналы о желтом цвете формируются или в НКТ, или в системе ганглиозных клеток
-типа. И тогда сигналы о желтом цвете формируются или в НКТ, или в системе ганглиозных клеток  -типа (рис. 3.2.3). Это справедливо по крайней мере для сетчатки приматов [227],
-типа (рис. 3.2.3). Это справедливо по крайней мере для сетчатки приматов [227],  -клетки плохо возбуждаются излучениями с длиной волны больше 500 нм и хорошо — белым светом.
-клетки плохо возбуждаются излучениями с длиной волны больше 500 нм и хорошо — белым светом.
Рассмотренные выше типы рецептивных полей ганглиозных клеток обнаруживают двойственную типологию. В сетчатке низших позвоночных (рыб, лягушек) наблюдаются два типа спектрально-оппонентных клеток. Одни характеризуются пространственной спектральной оппонентностью рецептивного поля, когда пространственно разные части рецептивного поля (центр и
периферия) представляют разные типы рецепторов, а другие — функциональной спектральной оппонентностью, когда одна и та же часть рецептивного поля представлена разными рецепторами, — это клетки с двойной спектральной оппонентностью.
Показательно, что в сетчатке высших позвоночных (например, обезьян) клеток с двойной спектральной оппонентностью практически нет, зато, как мы увидим далее, они появляются на более высших (корковых) уровнях цветового анализатора излучений.
Все это свидетельствует о том, что только клетки второго типа могут участвовать в цветовом анализе излучений, тогда как клетки первого типа настроены скорее всего на выделение пространственных характеристик стимуляции (края, контрасты  и т. д.).
и т. д.).



































































































































































 клетки
клетки

 и
и  -типы (табл. 3.2.3).
-типы (табл. 3.2.3).  -клетка возбуждается в ответ на длинноволновое излучение и тормозится в ответ на средневолновое излучение.
-клетка возбуждается в ответ на длинноволновое излучение и тормозится в ответ на средневолновое излучение.  -клетки возбуждаются средне- и длинноволновым излучением и тормозятся коротковолновым. Обратный тип реакций наблюдается
-клетки возбуждаются средне- и длинноволновым излучением и тормозятся коротковолновым. Обратный тип реакций наблюдается  и
и  -клеток. Сигналы от фоторецепторов разного типа представлены в рецептивных полях оппонентных ганглиозных клеток упорядоченно и, как правило, пространственно разнесены. В сетчатке карповых рыб и суслика обнаружены ганглиозные «летки с двойной спектральной оппонентностью, рецептивные
-клеток. Сигналы от фоторецепторов разного типа представлены в рецептивных полях оппонентных ганглиозных клеток упорядоченно и, как правило, пространственно разнесены. В сетчатке карповых рыб и суслика обнаружены ганглиозные «летки с двойной спектральной оппонентностью, рецептивные
 -клетки с двойной спектральной оппонентностью, размеры центра рецептивного поля которых сопоставимы с размерами рецептивного поля биполяров с двойной спектральной оппоментностью [120].
-клетки с двойной спектральной оппонентностью, размеры центра рецептивного поля которых сопоставимы с размерами рецептивного поля биполяров с двойной спектральной оппоментностью [120].

 -колбочек в рецептивных полях
-колбочек в рецептивных полях  -клеток из разных локусов сетчатки: 1 — доминируют исключительно
-клеток из разных локусов сетчатки: 1 — доминируют исключительно  -колбочки. 5 — исключительно
-колбочки. 5 — исключительно  -колбочки,
-колбочки,  — промежуточные (между 1 и 5) степени доминирования, 0° — фовеа
— промежуточные (между 1 и 5) степени доминирования, 0° — фовеа
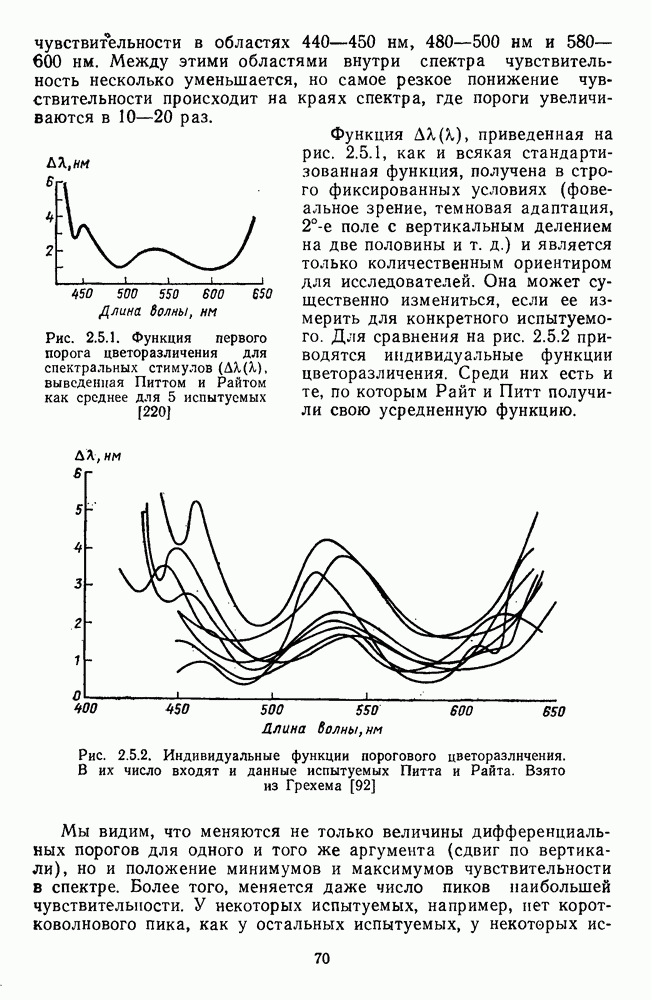
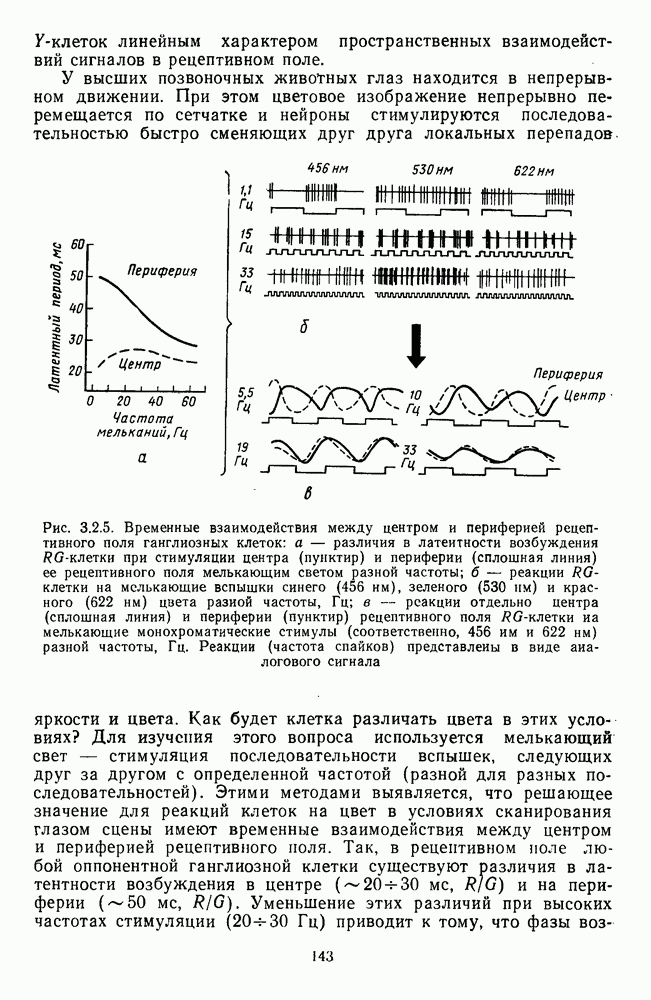
 -клеток линейным характером пространственных взаимодействий сигналов в рецептивном поле.
-клеток линейным характером пространственных взаимодействий сигналов в рецептивном поле.

 -клетки при стимуляции центра (пунктир) и периферии (сплошная линия) ее рецептивного поля мелькающим светом разной частоты;
-клетки при стимуляции центра (пунктир) и периферии (сплошная линия) ее рецептивного поля мелькающим светом разной частоты;  — реакции
— реакции  -клетки на мелькающие вспышки синего (456 нм), зеленого (530 нм) и красного (622 нм) цвета разной частоты, Гц; в — реакции отдельно центра (сплошная линия) и периферии (пунктир) рецептивного поля
-клетки на мелькающие вспышки синего (456 нм), зеленого (530 нм) и красного (622 нм) цвета разной частоты, Гц; в — реакции отдельно центра (сплошная линия) и периферии (пунктир) рецептивного поля  -клетки на мелькающие монохроматические стимулы (соответственно, 456 им и 622 нм) разной частоты, Гц. Реакции (частота спайков) представлены в виде аналогового сигнала
-клетки на мелькающие монохроматические стимулы (соответственно, 456 им и 622 нм) разной частоты, Гц. Реакции (частота спайков) представлены в виде аналогового сигнала
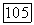 и на периферии
и на периферии 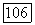 Уменьшение этих различий при высоких частотах стимуляции
Уменьшение этих различий при высоких частотах стимуляции 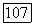 приводит к тому, что фазы
приводит к тому, что фазы