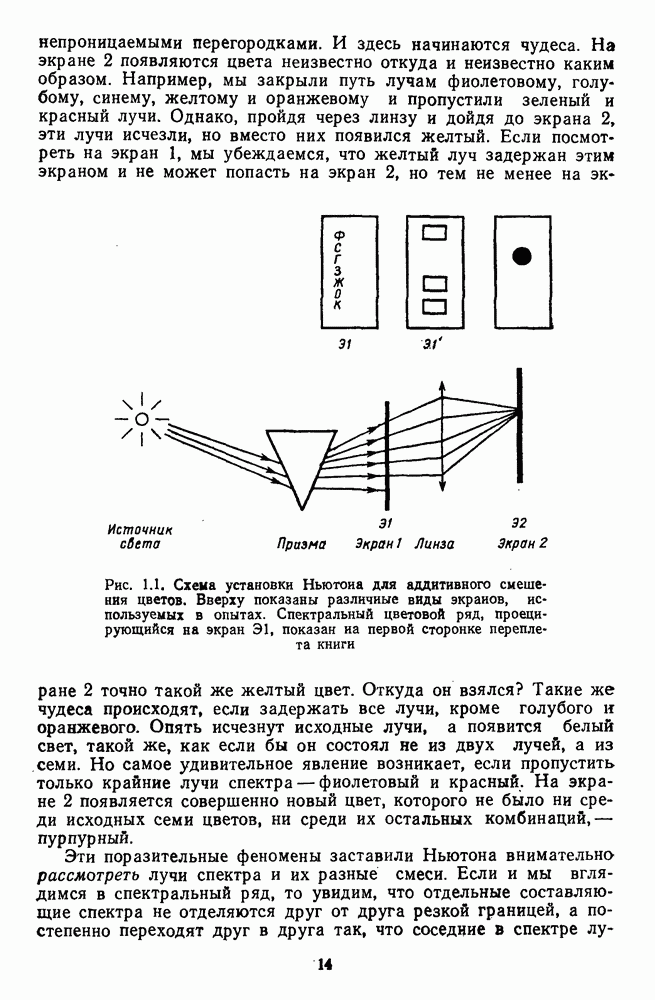
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
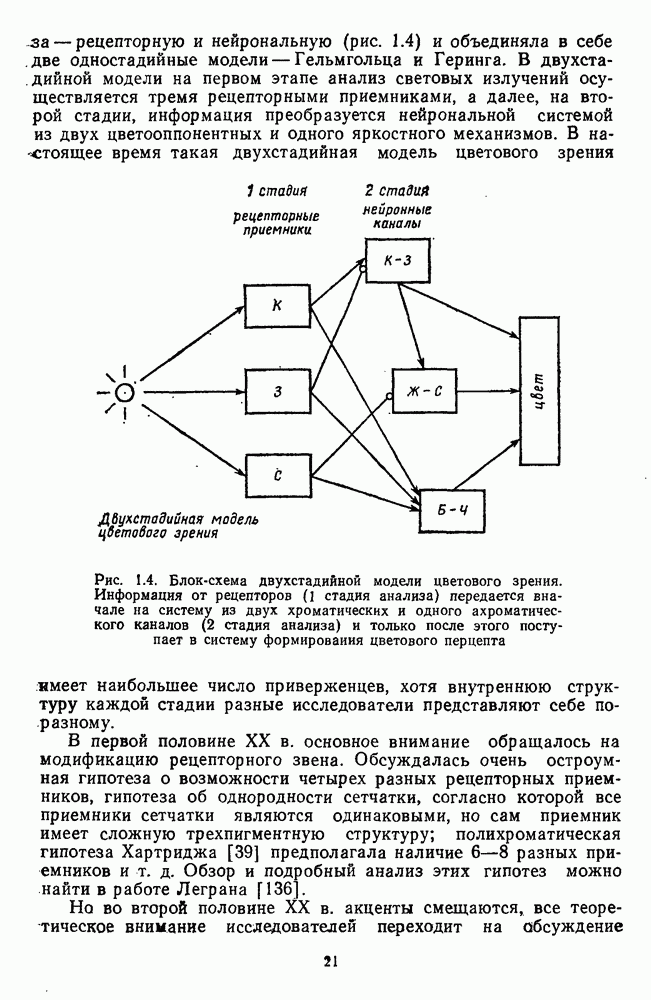
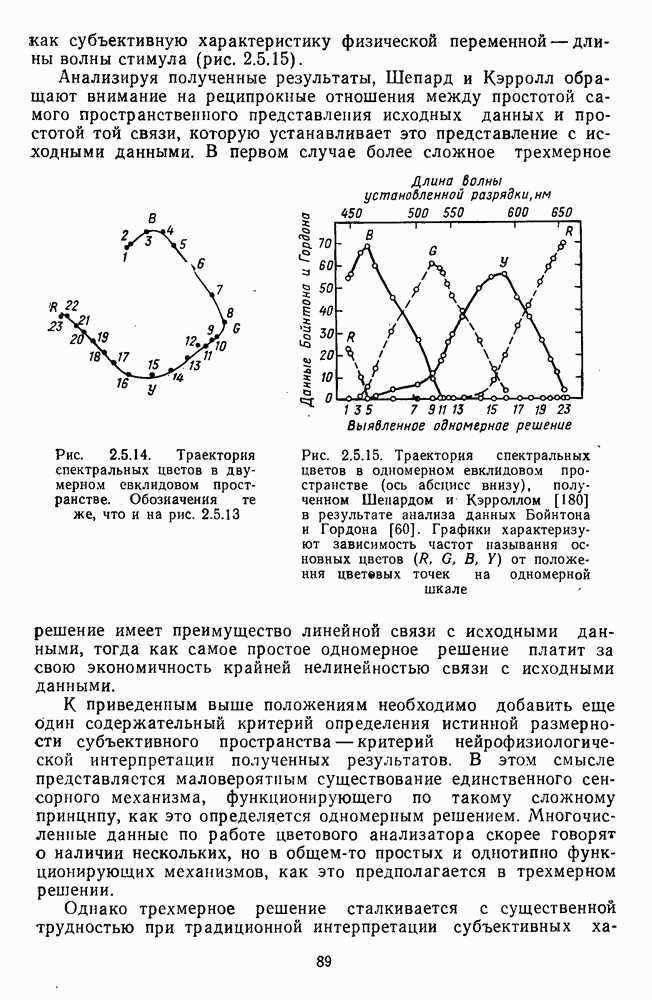
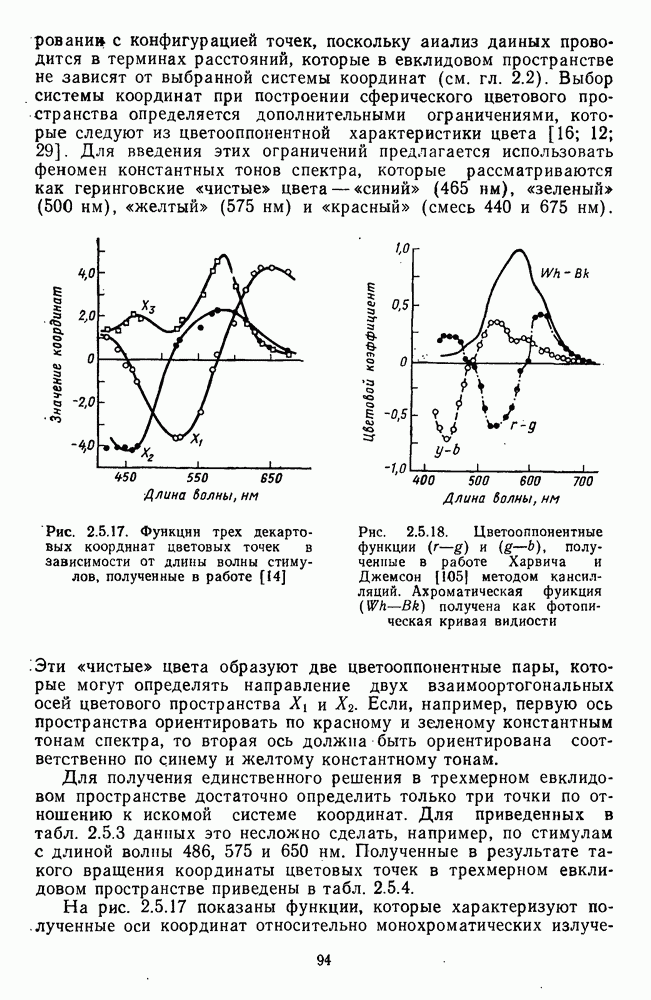
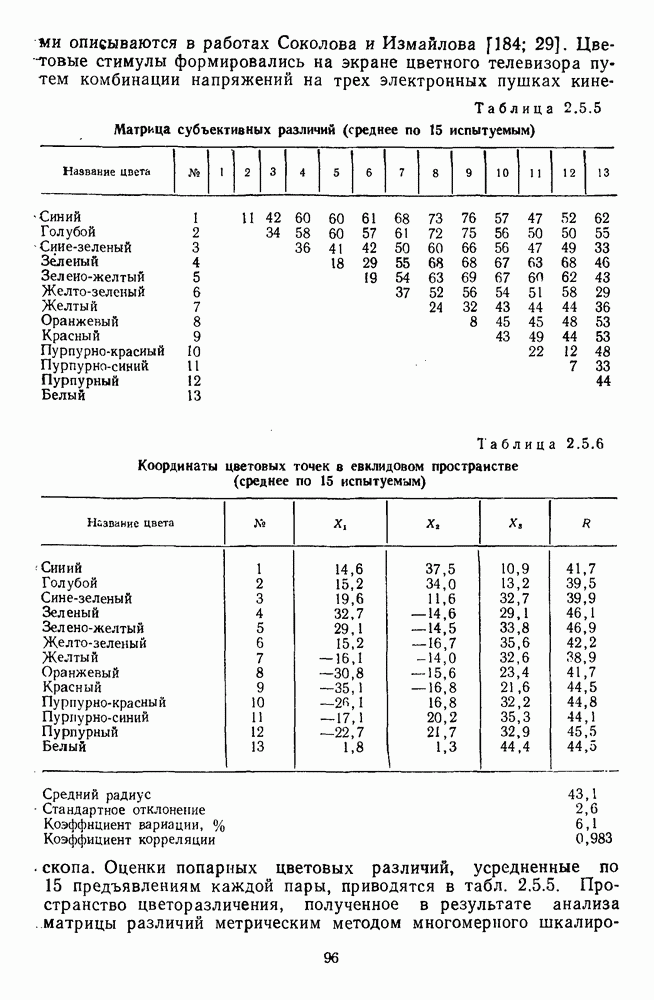
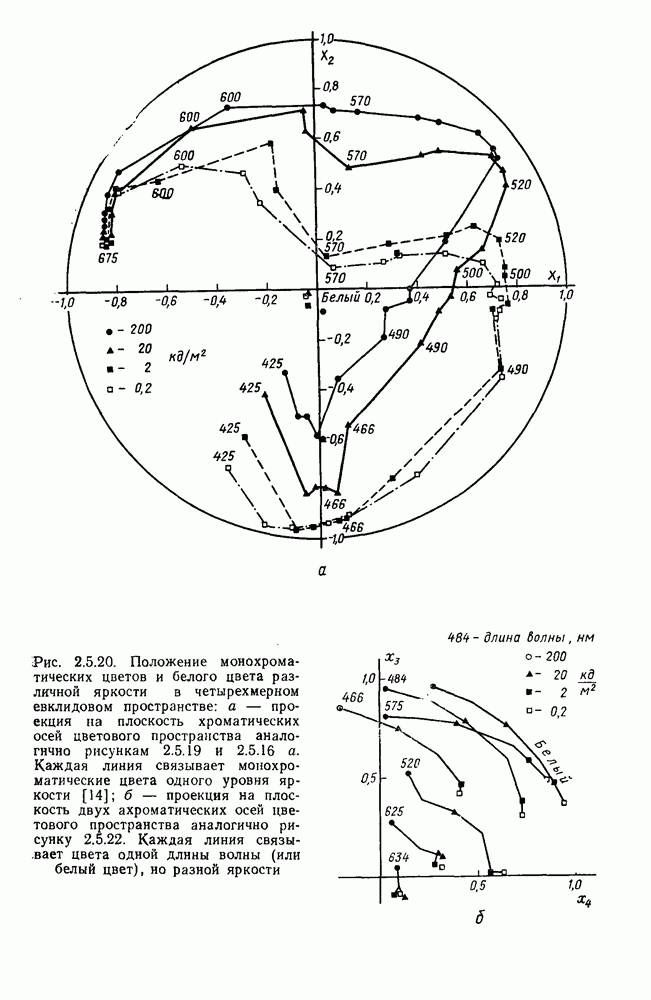
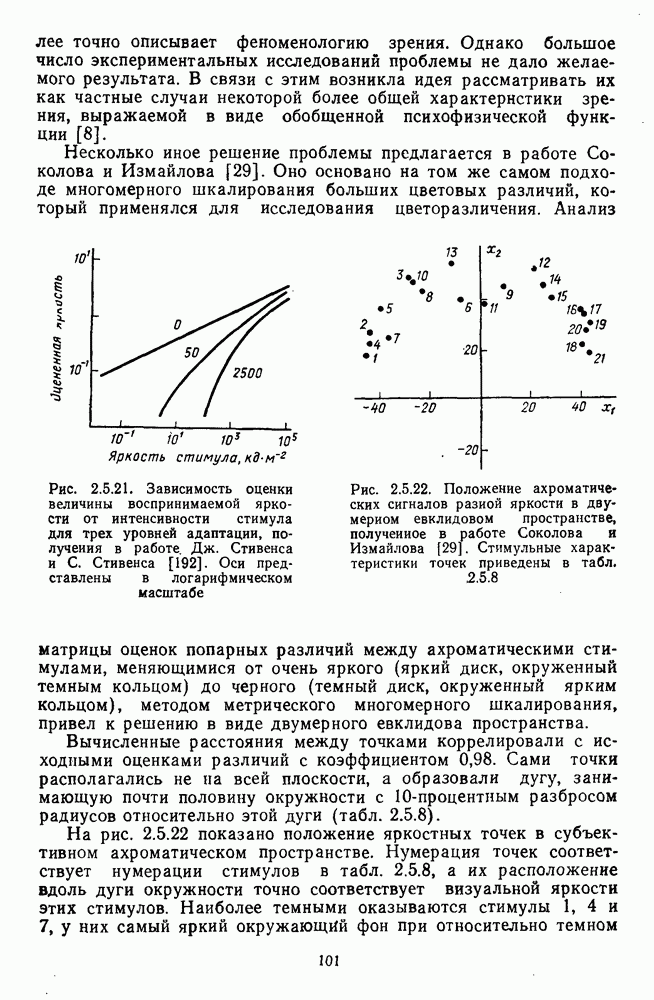
4.3.5. Общая трехстадийная модель цветового зренияРезультаты экспериментального исследования взаимосвязи между нейрональной сетью ахроматического зрения, представленной на рис. 4.3.3, и нейрональной сетью хроматического зрения, представленной на рис. 4.3.5, было проведено в работе Измайлова [14]. С этой целью методом многомерного шкалирования была построена пространственная модель различения монохроматических и ахроматических цветов разной яркости. В результате анализа оказалось, что множество цветов разной яркости можно расположить только в четырехмерном евклидовом пространстве, если исходить из условия линейных соотношений между цветовыми различиями и межточечными расстояниями. Как и в случае только ахроматических или только хроматических стимулов, цвета разной яркости заполняют не все четырехмерное пространство, а образуют в нем гиперсферу. Случайные отклонения цветовых точек от гиперсферы, измеряемые как вариативность радиуса гиперсферы, не превышают 14%. Сферическая модель цветового зрения, основанная на этих данных, позволила установить, что каждый цвет может быть представлен как в системе четырех нейрофизиологических характеристик (красно-зеленый, сине-желтый, световой и темновой каналы зрительной системы), определяемых декартовыми координатами цветовой точки, так и в системе психофизических характеристик (цветовой тон, насыщенность, светлота), определяемых сферическими координатами этой же цветовой точки. При этом две хроматические характеристики цвета (красно-зеленая и сине-желтая) будучи представлены как функции двух первых координат спектральных цветов от длины волны, полностью совпадают с аналогичными характеристиками, полученными для равноярких цветов, (рис. 2.5.17), а две другие, ахроматические характеристики (световая и темновая), будучи представлены как функции третьей и, четвертой координат ахроматических цветов от яркости, полностью совпадают с аналогичными характеристиками, полученными ранее только для ахроматических стимулов [29; 14]. Кроме того, оказалось, что ахроматическая координата хроматического пространства, представленная в уравнении 4.3.10 символом
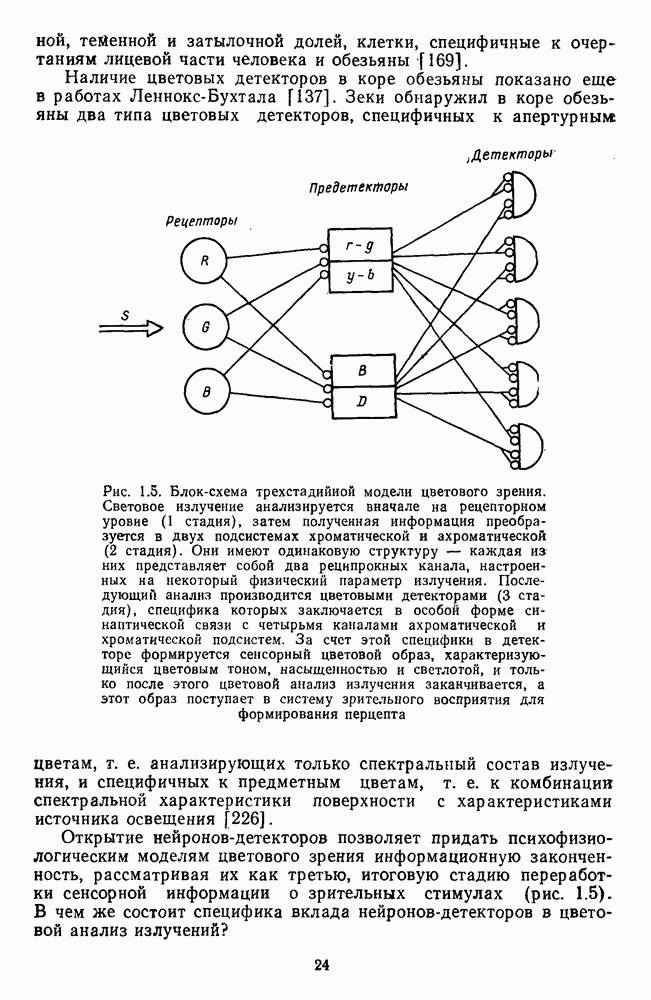
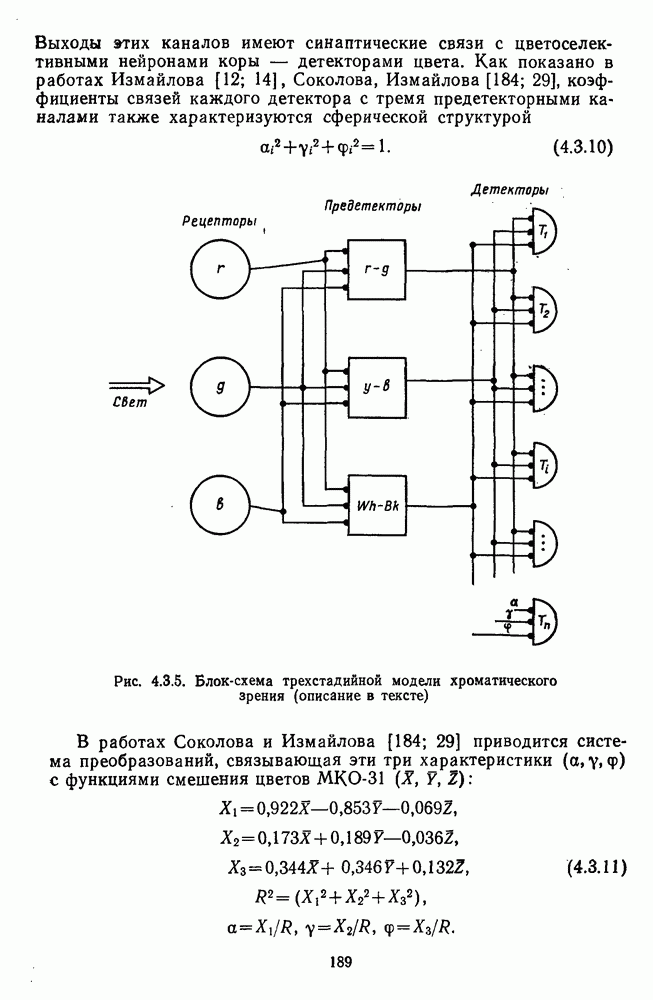
Таким образом, удалось выявить, что общая нейрональная сеть, осуществляющая в зрительной системе цветовой анализ спектрального состава и интенсивности излучения, представляет собой трехзвенную структуру, блок-схема которой уже была приведена на рис. 1.5 в историческом обзоре. Входное рецепторное звено этой сети остается неизменным при анализе как спектрального состава, так и интенсивности излучения. Второе, пострецепторное (или предетекторное) звено представляет собой две однотипные подсистемы (хроматическую и ахроматическую), каждая из которых состоит из двух реципрокно активирующихся каналов Информация из этих четырех каналов передается на множество цветовых детекторов, представляющих третье звено нейрональной сети. Синаптические входы каждого детектора характеризуются сферической структурой так, что
где При анализе ахроматических сигналов активируются только световой и темновой каналы, которые передают возбуждения на входы При анализе равноярких цветов активируются все четыре канала пострецепторной сети, но, хотя абсолютные значения возбуждений в каналах В и И только когда излучение меняется и по спектральному составу, и по интенсивности, можно наблюдать общую работу всех элементов цветового анализатора. Представленная на рис. 1.5 модель цветового анализатора предусматривает три стадии анализа излучения. На первой, рецепторной стадии излучения классифицируются только по качественному признаку: одинаковые — разные. На второй, предетекторной стадии осуществляется дальнейшая дифференциация излучений на хроматические и ахроматические за счет функционирования двух независимых механизмов для кодирования сенсорного образа. Один из них служит нейрофизиологической основой такой сенсорной характеристики цвета, как цветовой тон, а другой — основой светлоты цвета. На третьей стадии — детекторной — осуществляется интеграция этих характеристик — хроматической и ахроматической — в целостный сенсорный образ. В процессе этой специфически нейрональной интеграции формируются не только две основные сенсорные характеристики стимула — тон и светлота но дополнительно, за счет сферической структуры синапсов детектора, образуется еще одна цветовая характеристика — насыщенность. Это означает, что насыщенность, в отличие от цветового тона и светлоты, является исключительно психофизиологической характеристикой и объясняет, почему она не имеет своего физического коррелята. В заключение необходимо отметить, что специфика трехстадийной концепции цветового зрения, развиваемой авторами этой книги, заключается в том, что обоснование необходимости третьей стадии анализа светового излучения прямо следует из структуры сферической модели различения цветовых стимулов, в которой набор оппонентных (хроматических) и неоппонентных (ахроматических) характеристик излучения преобразуется в систему тон — насыщенность — светлота за счет специальной сферической структуры взаимоотношения между этими характеристиками. А структура сферической модели в свою очередь следует непосредственно из экспериментальных измерений больших" надпороговых различий. Таким образом, если одностадийная концепция базировалась на опытах по смешению цветов, двухстадийная концепция — на исследованиях цветовой оппонентности и порогового цветоразличения, то трехстадийная концепция базируется на исследованиях надпорогового цветоразличения.
|
 |
Оглавление
|


































































































































































 является производной от двух ахроматических координат четырехмерного цветояркостного пространства так, что
является производной от двух ахроматических координат четырехмерного цветояркостного пространства так, что

 в хроматической подсистеме и В и
в хроматической подсистеме и В и  в ахроматической).
в ахроматической).

 — индекс детектора.
— индекс детектора.
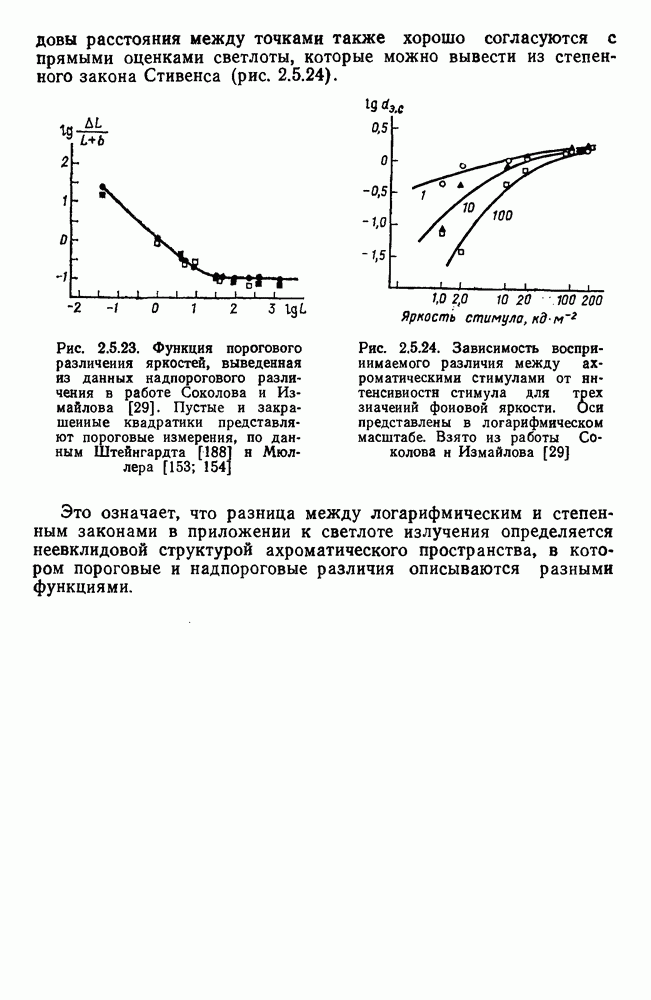
 детекторов. Хроматические планы не активируются, через входы а и у идет нулевая информация, и поэтому общая нейрональная сеть функционирует в редуцированном виде, как это и наблюдается в опытах с ахроматическими сигналами (рис. 2.5.22 и уравнение 4.3.3).
детекторов. Хроматические планы не активируются, через входы а и у идет нулевая информация, и поэтому общая нейрональная сеть функционирует в редуцированном виде, как это и наблюдается в опытах с ахроматическими сигналами (рис. 2.5.22 и уравнение 4.3.3).
 меняются при изменении стимулов, отношение возбуждений остается константным, поэтому общая ней рональная сеть также функционирует в редуцированном виде, хотя и иначе, чем в предыдущем случае (уравнение 4.3.10 и 4.3.12).
меняются при изменении стимулов, отношение возбуждений остается константным, поэтому общая ней рональная сеть также функционирует в редуцированном виде, хотя и иначе, чем в предыдущем случае (уравнение 4.3.10 и 4.3.12).