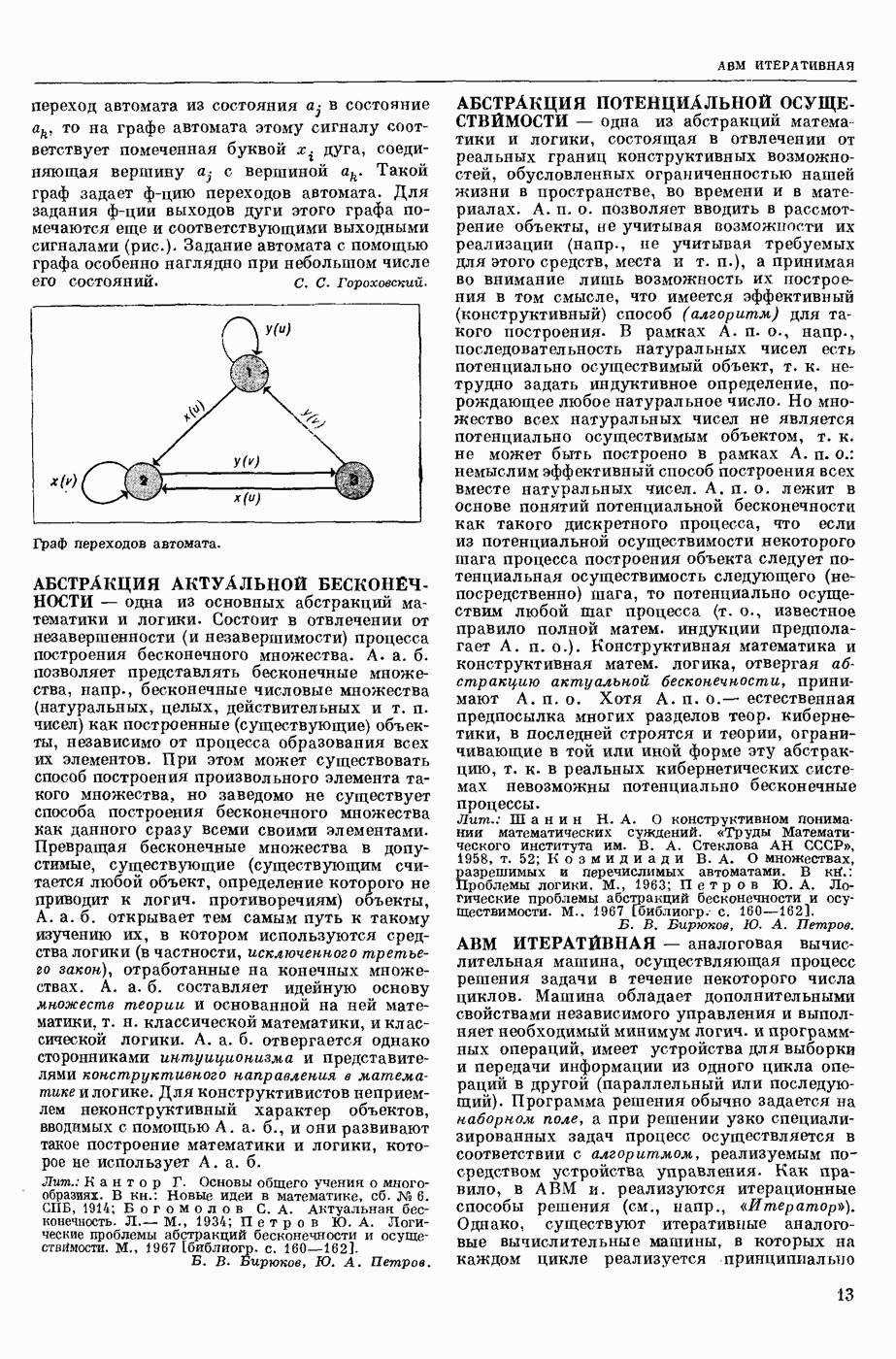
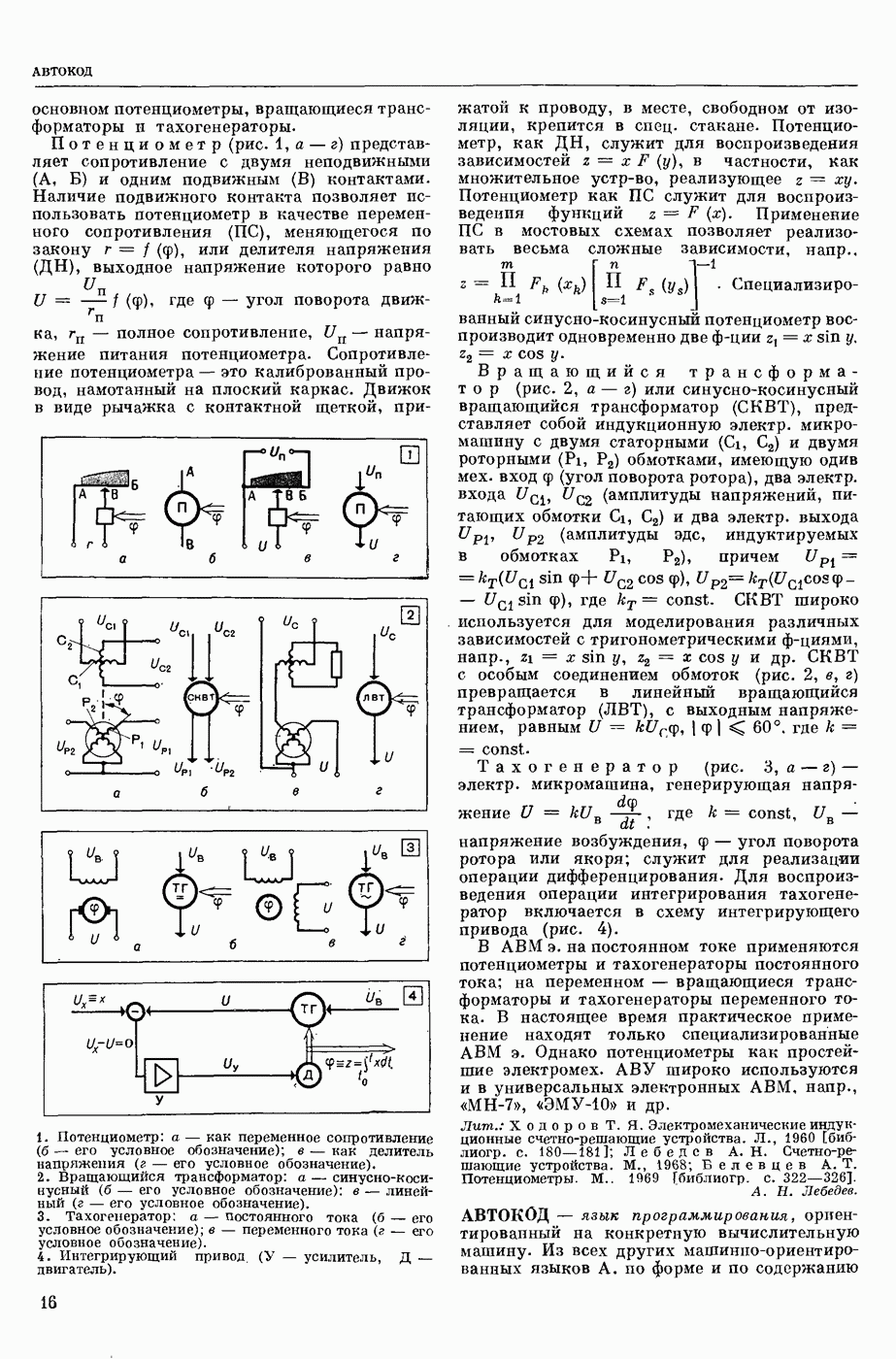
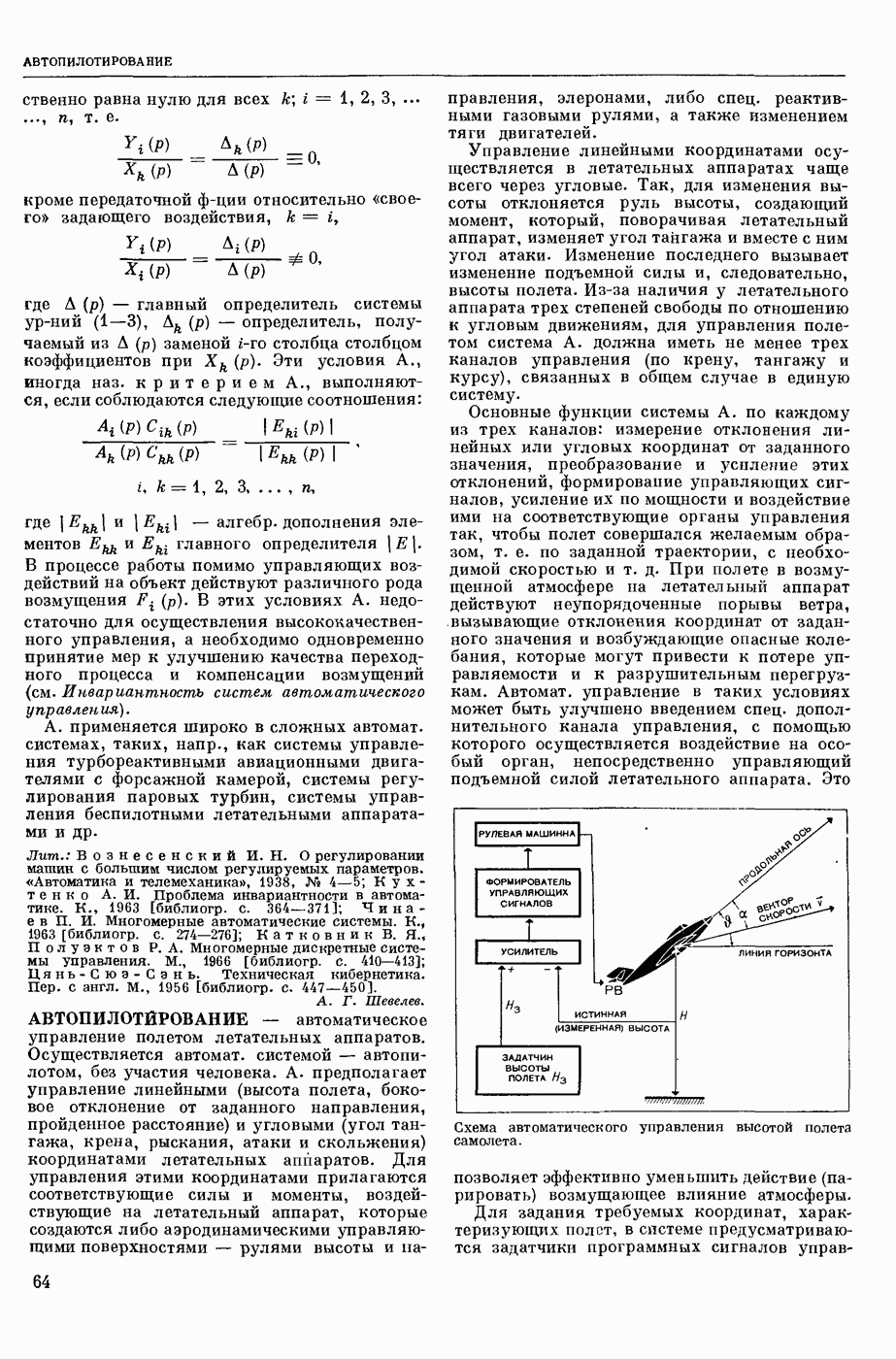
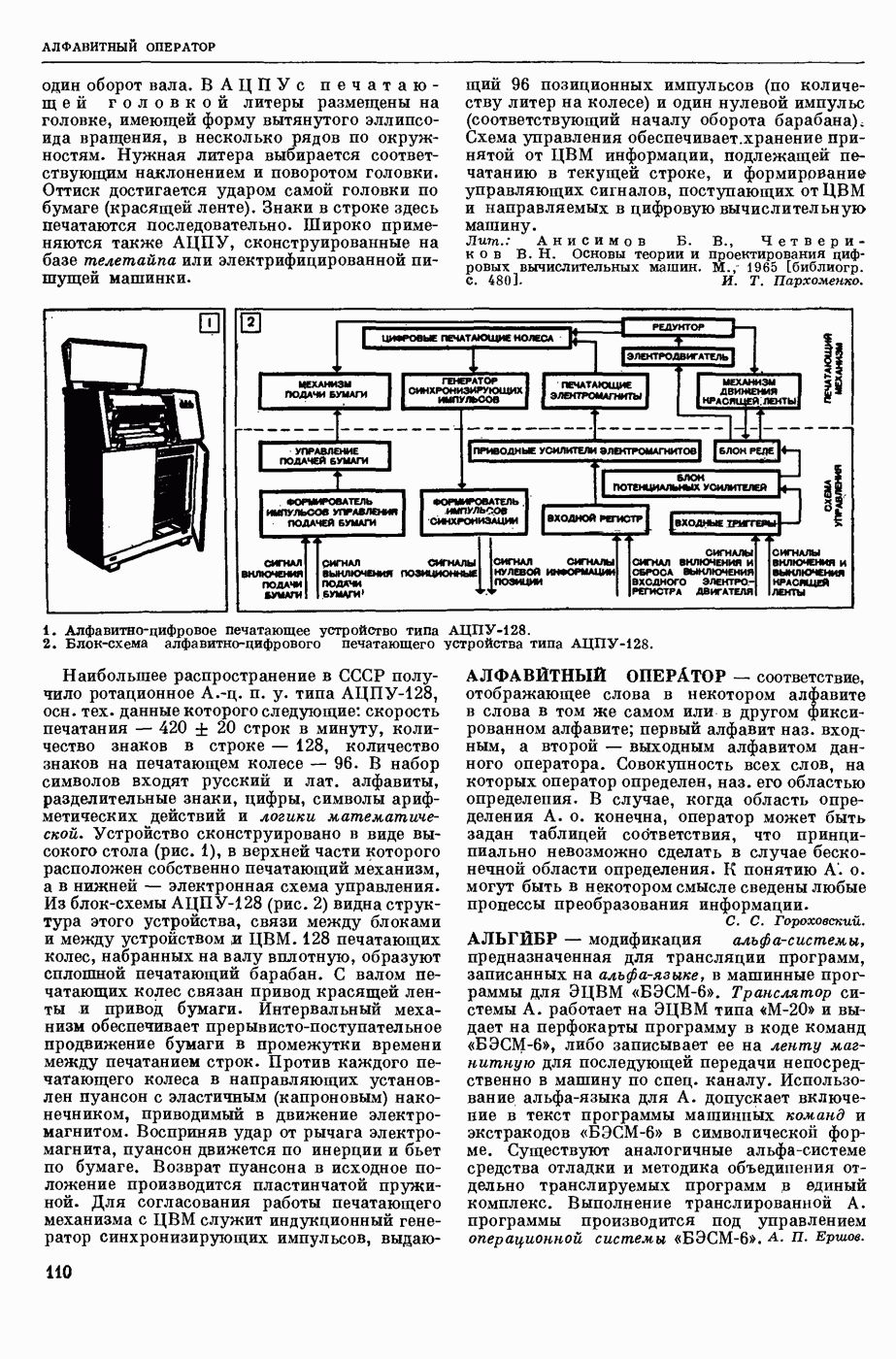
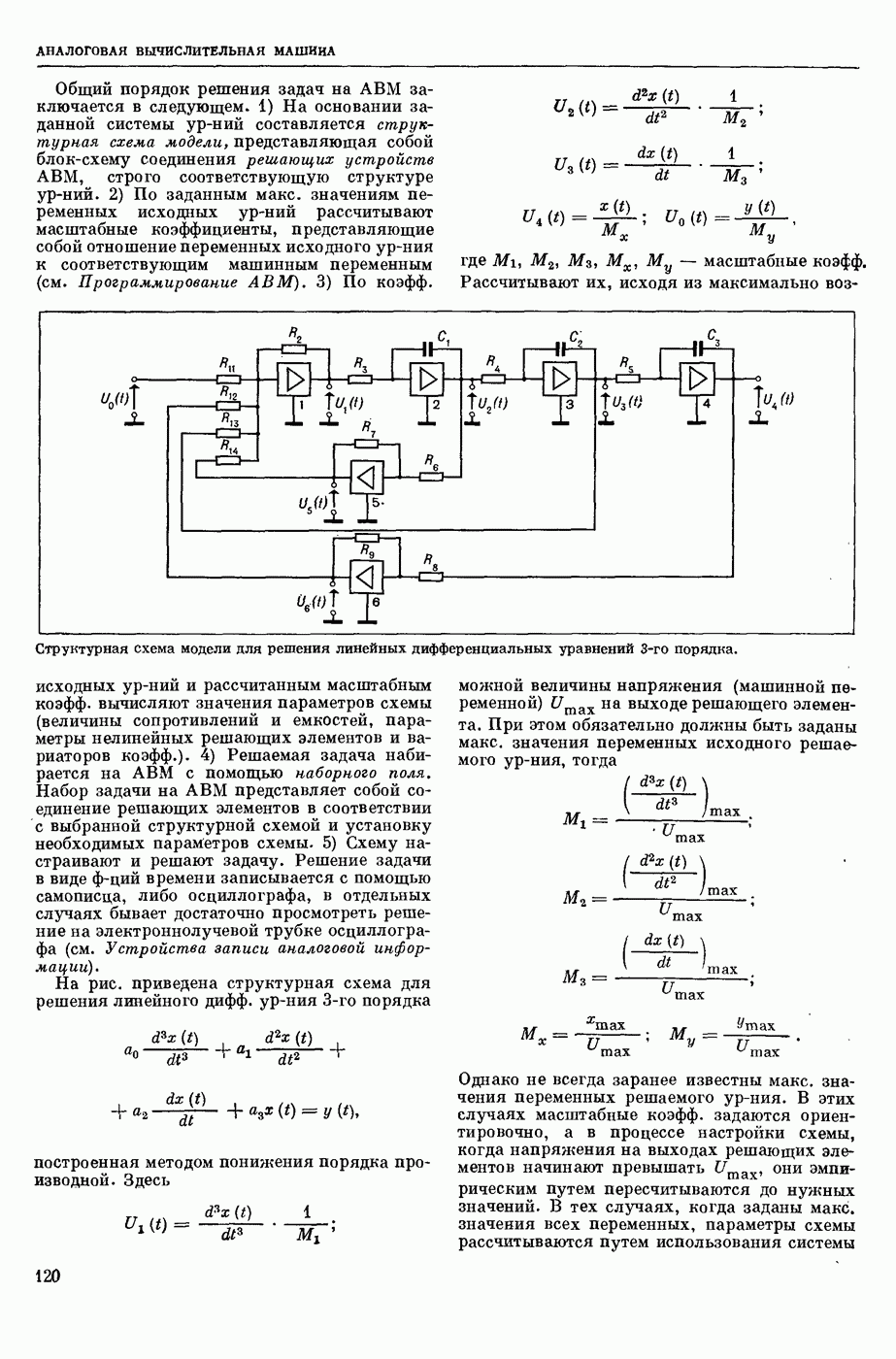
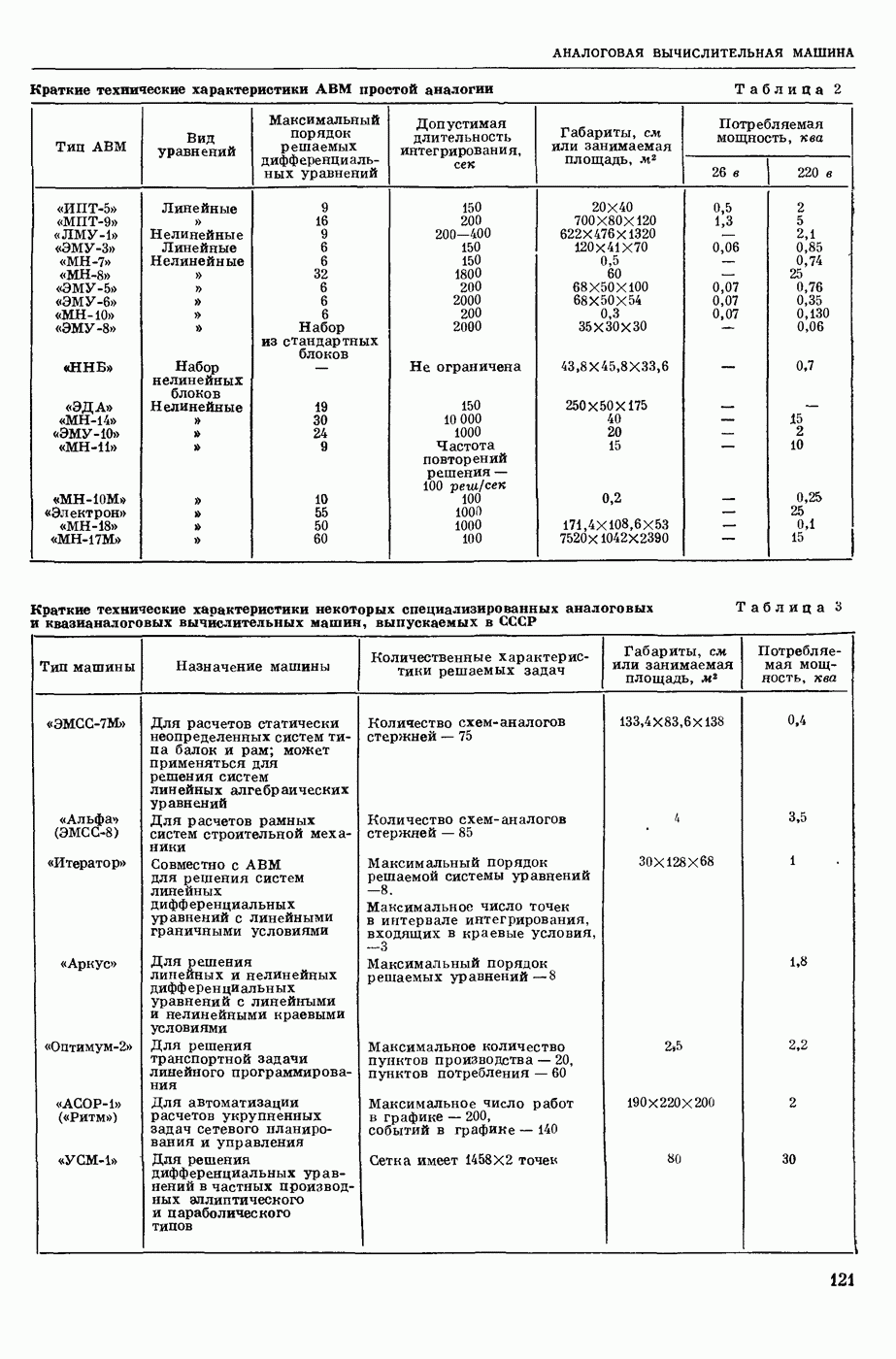
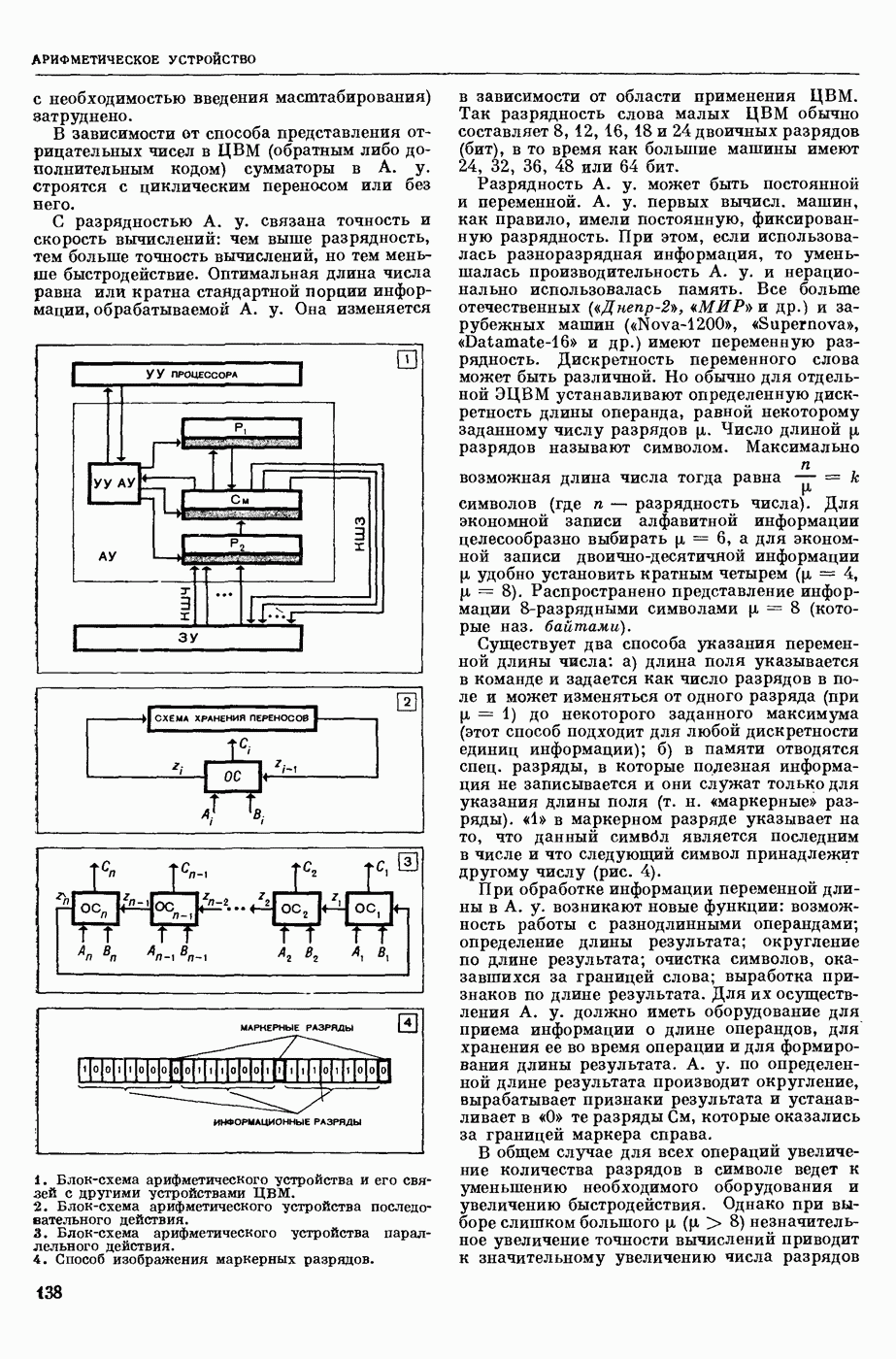
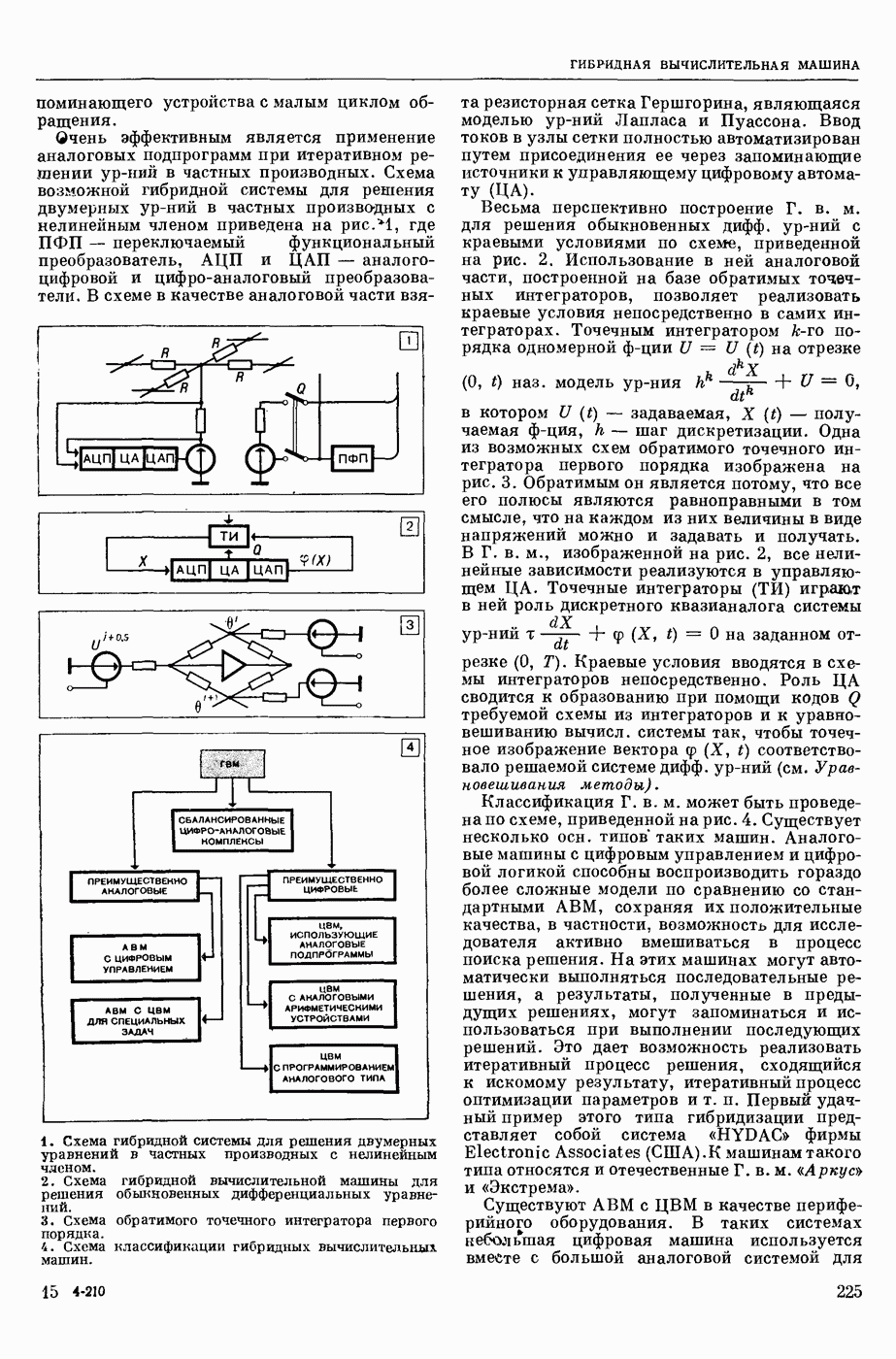
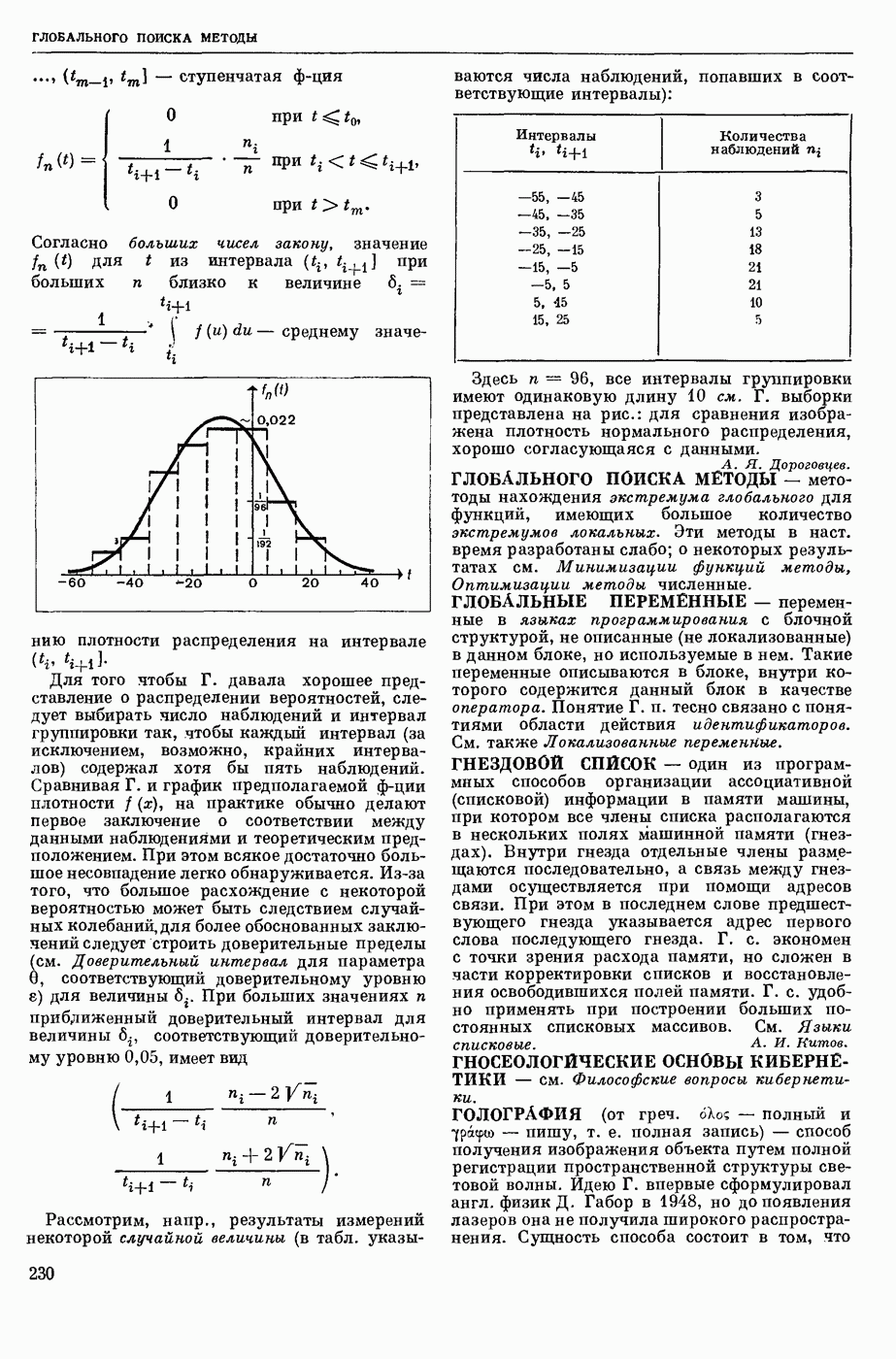
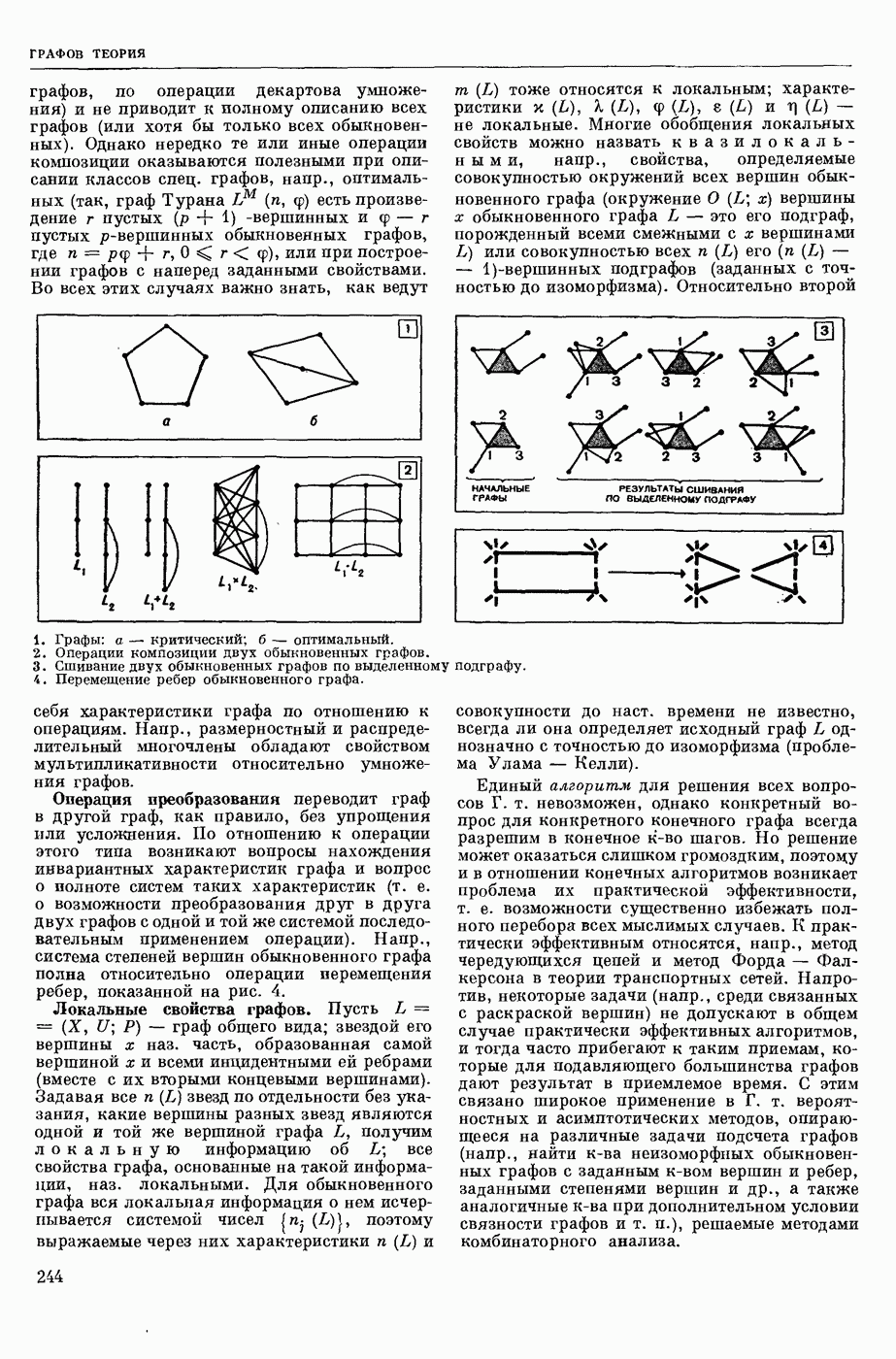
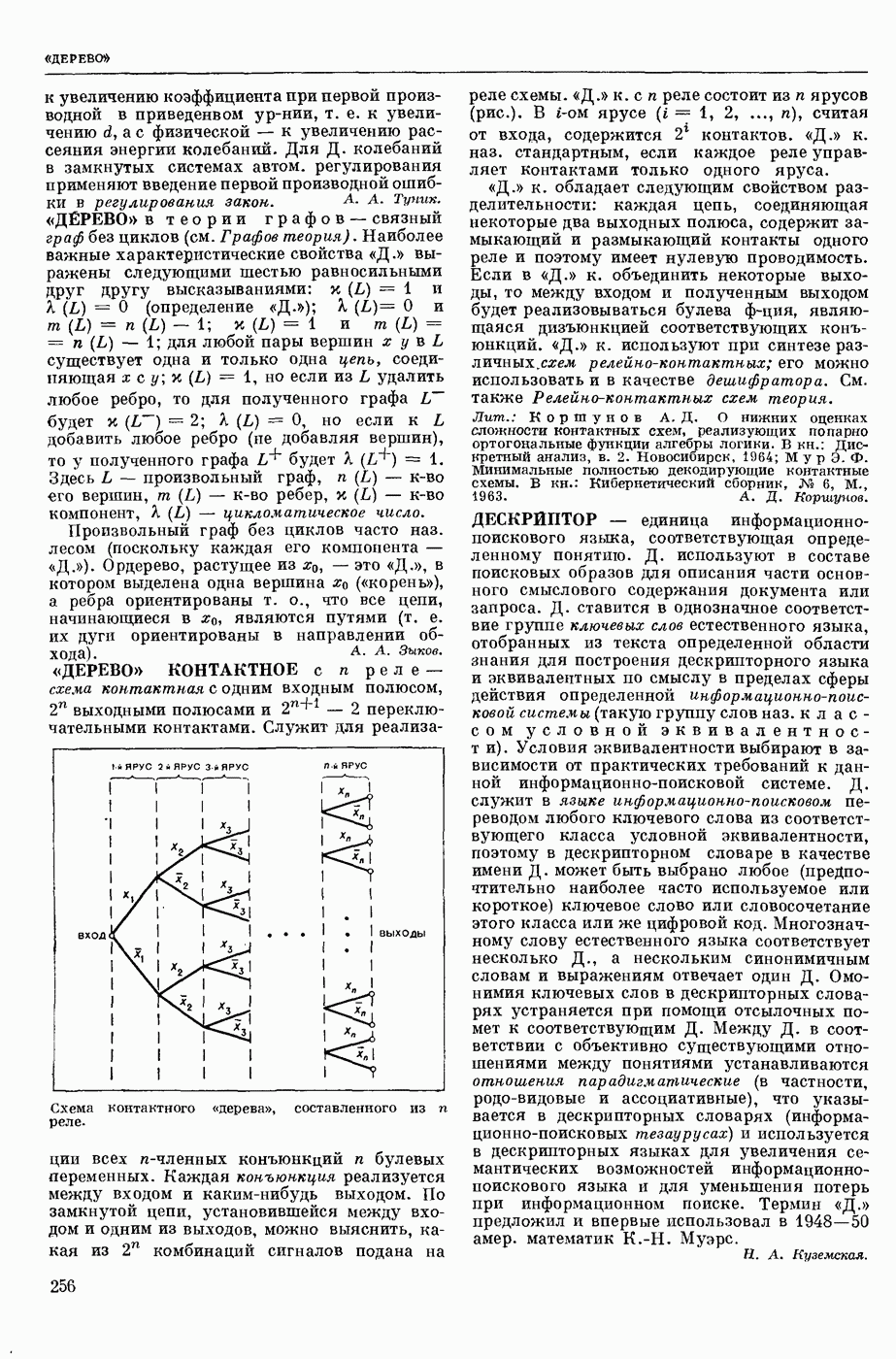
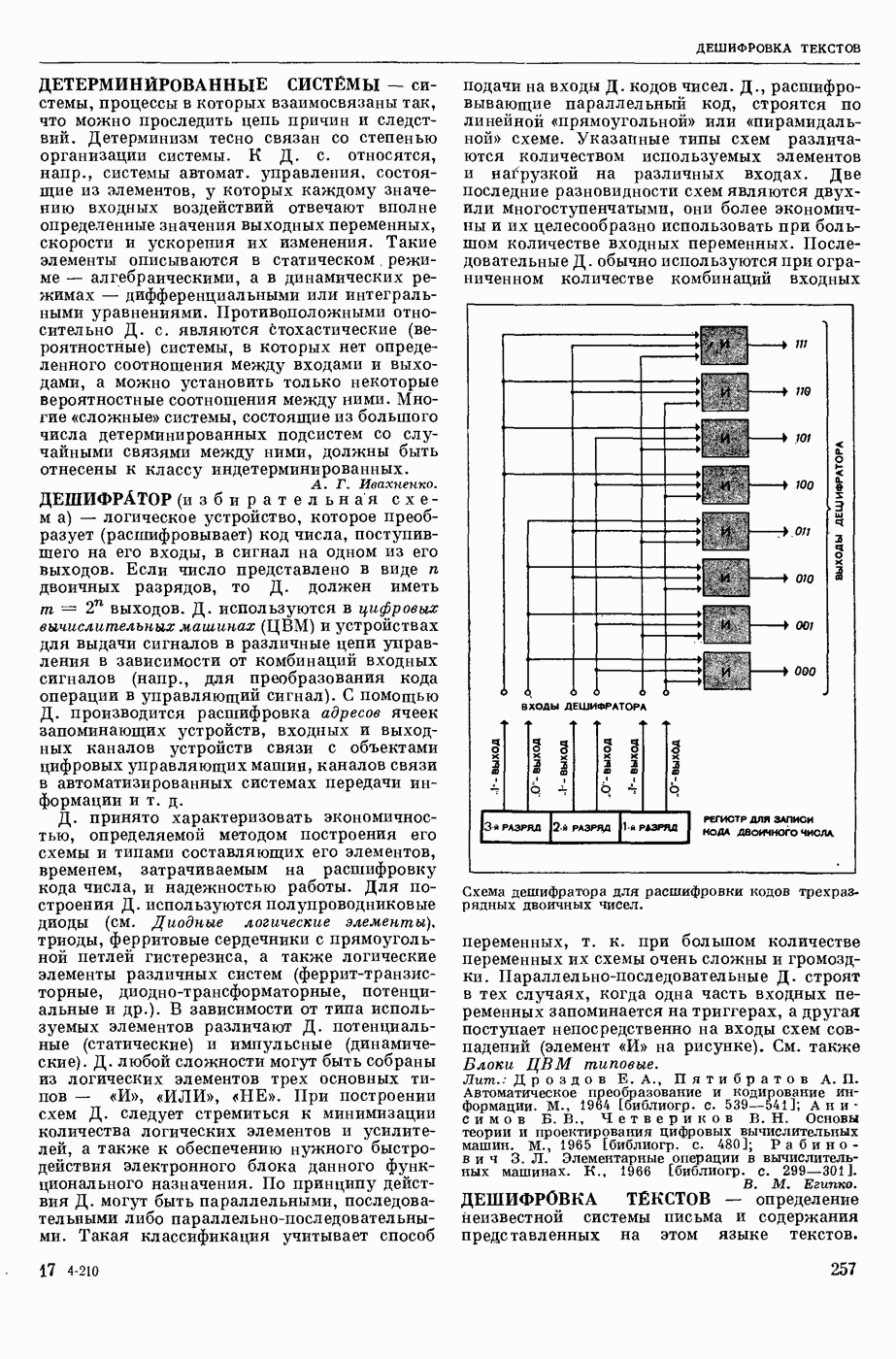
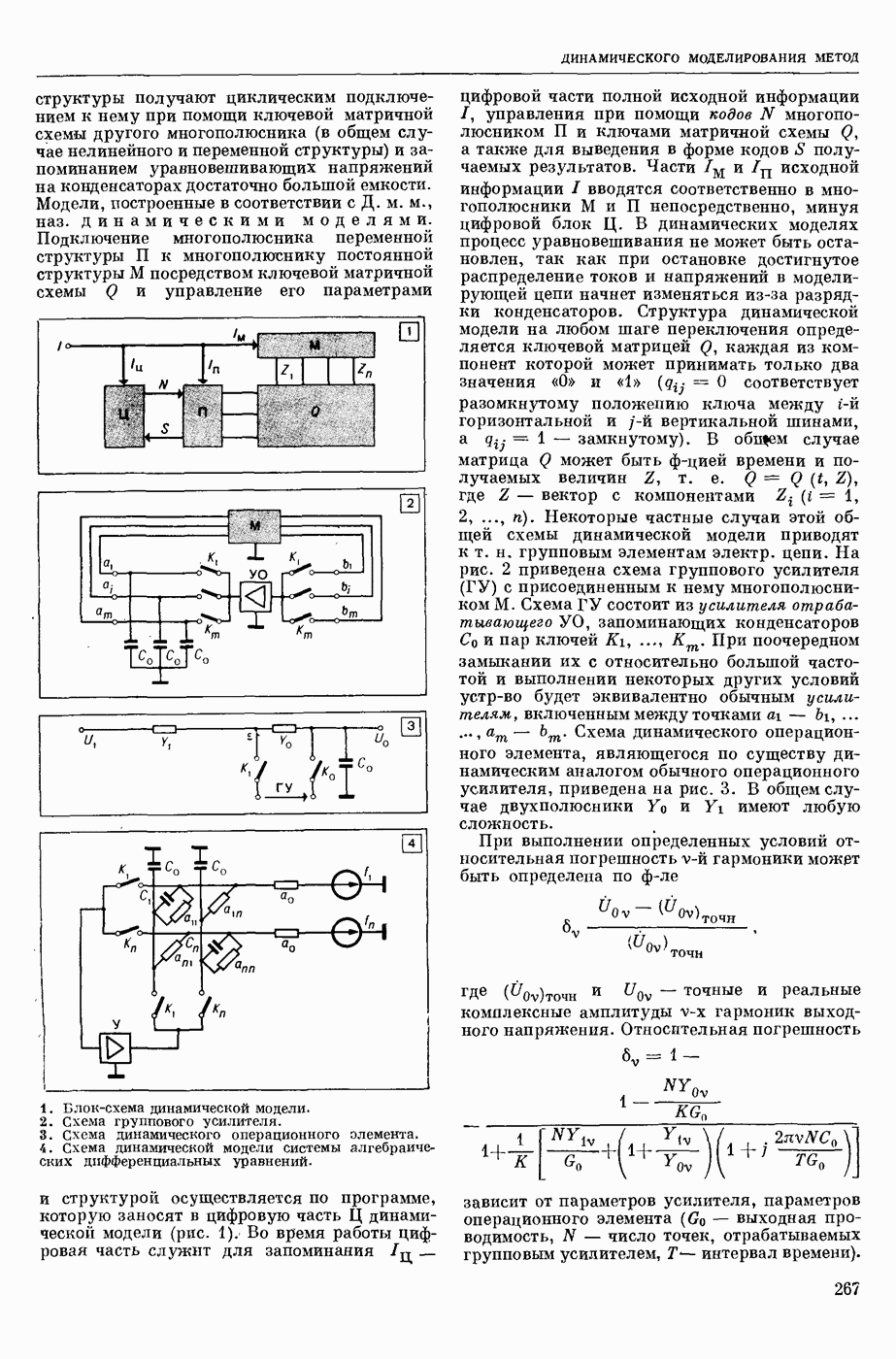
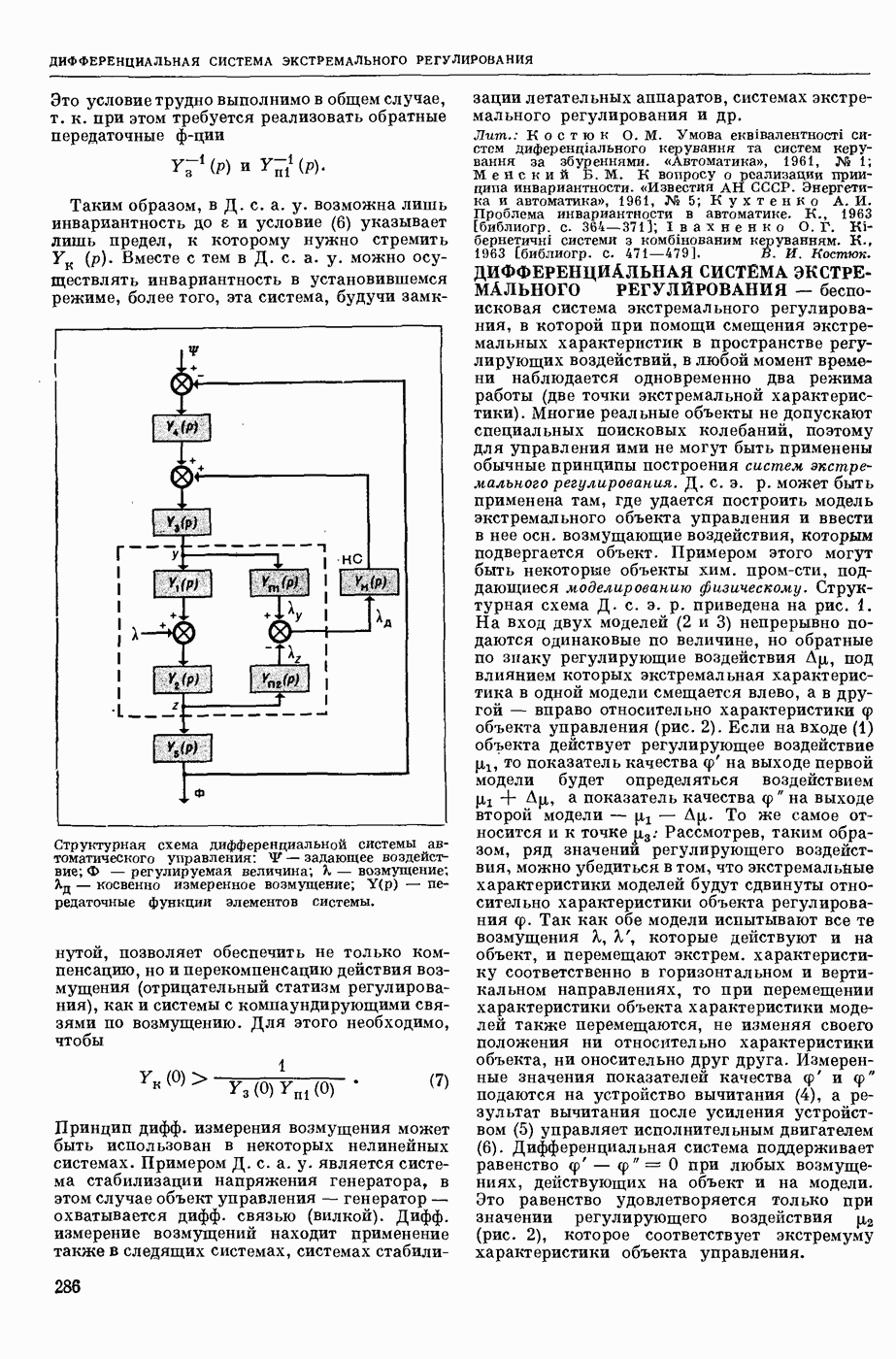
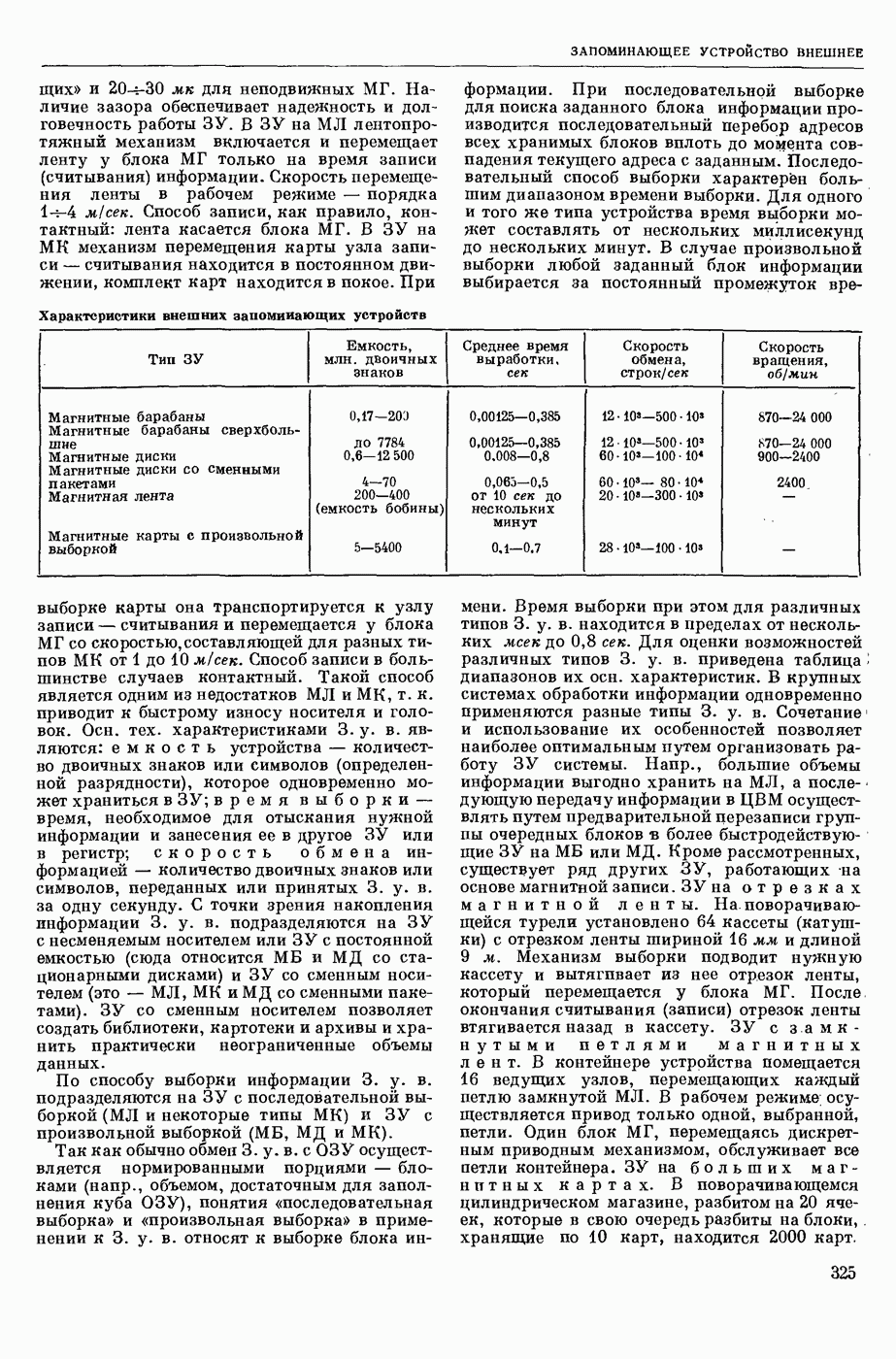
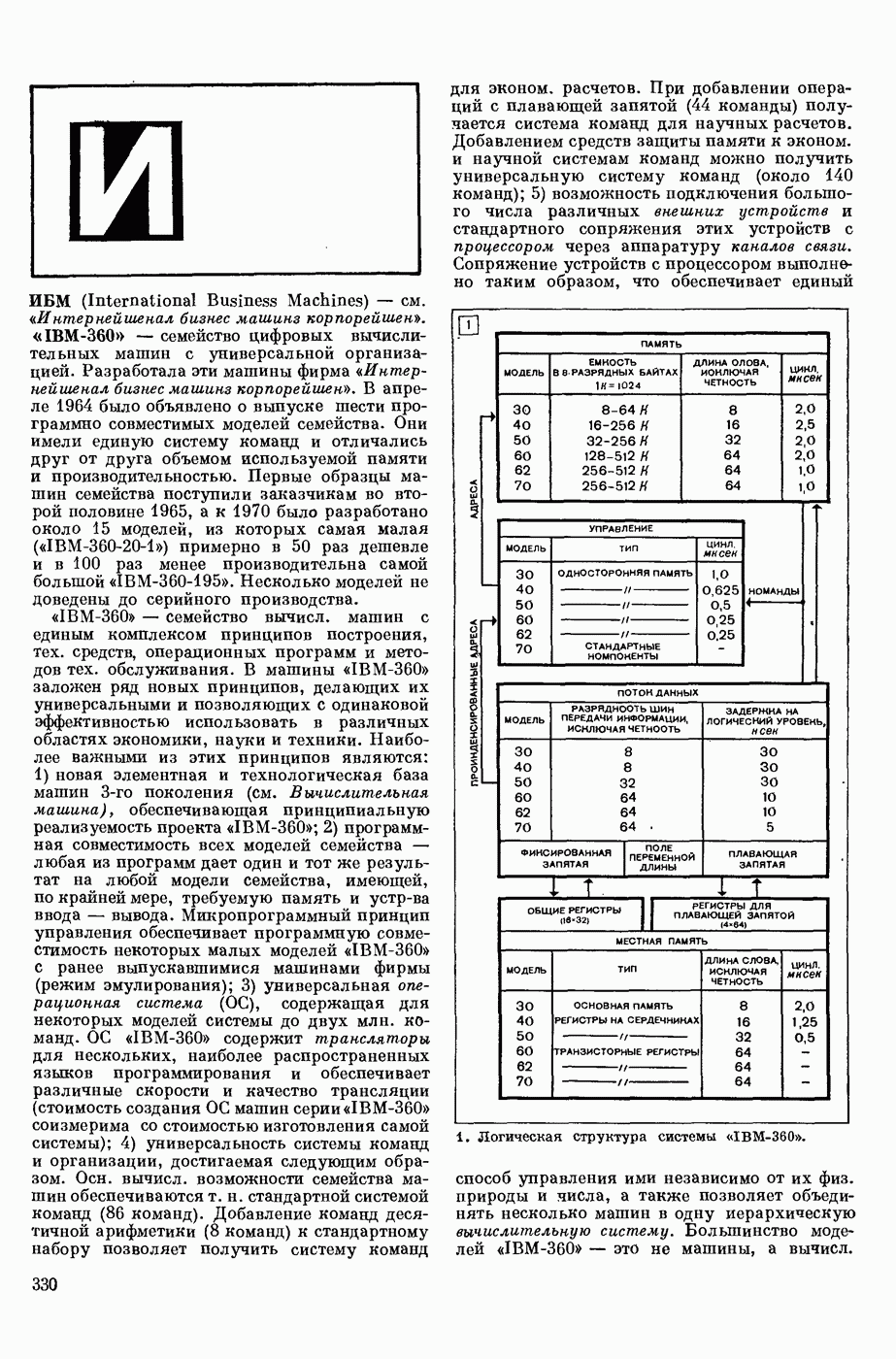
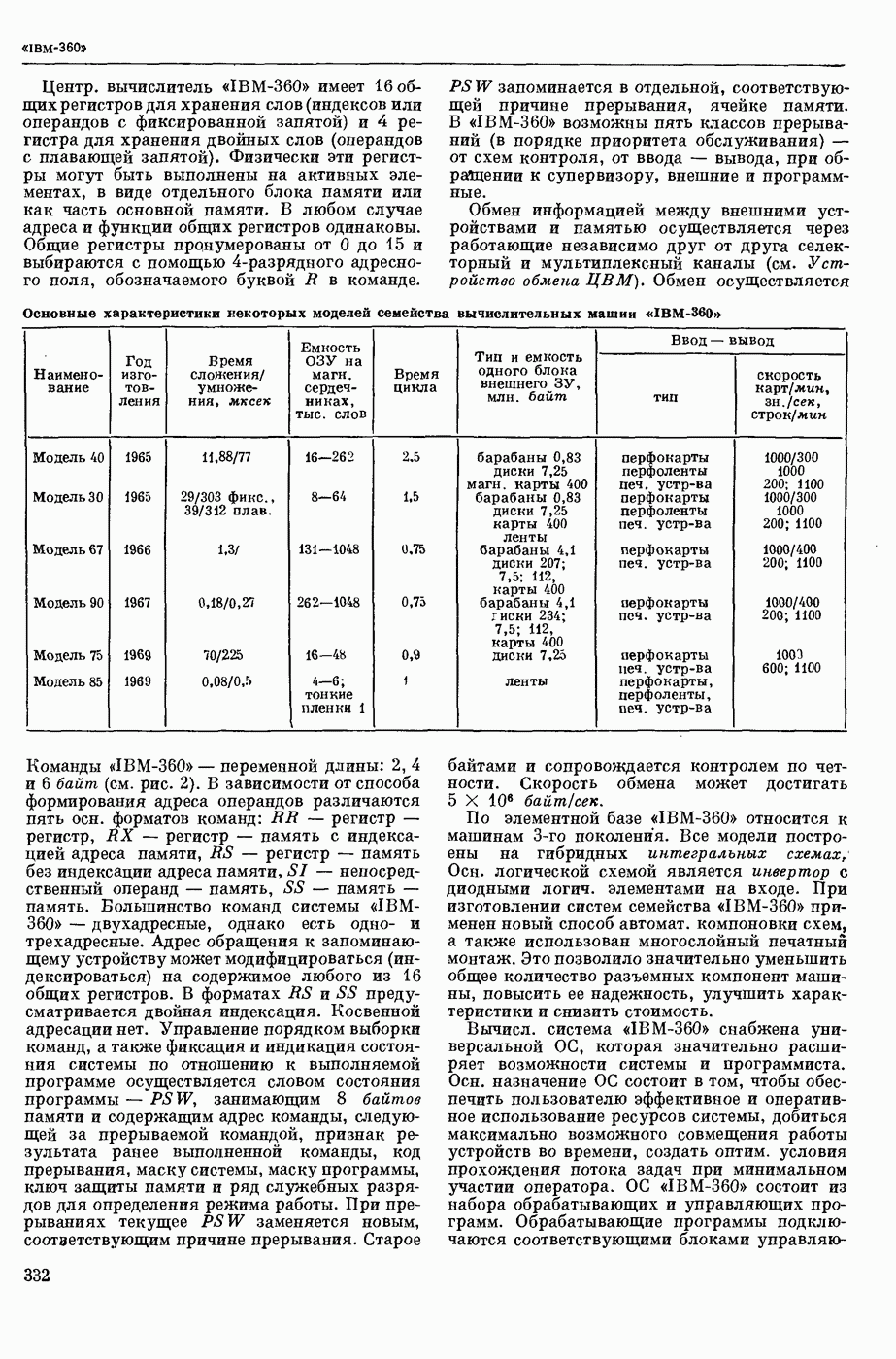
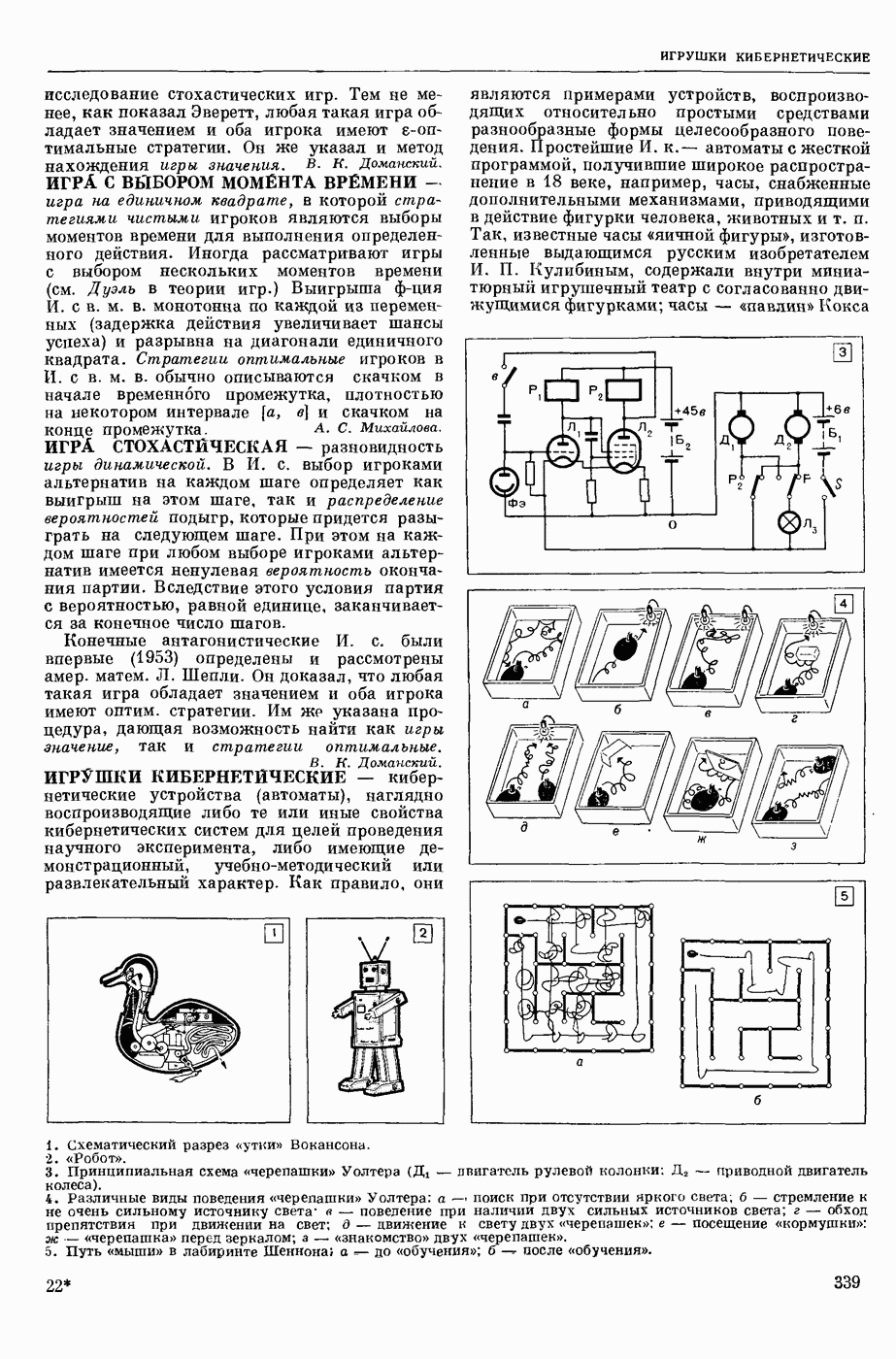
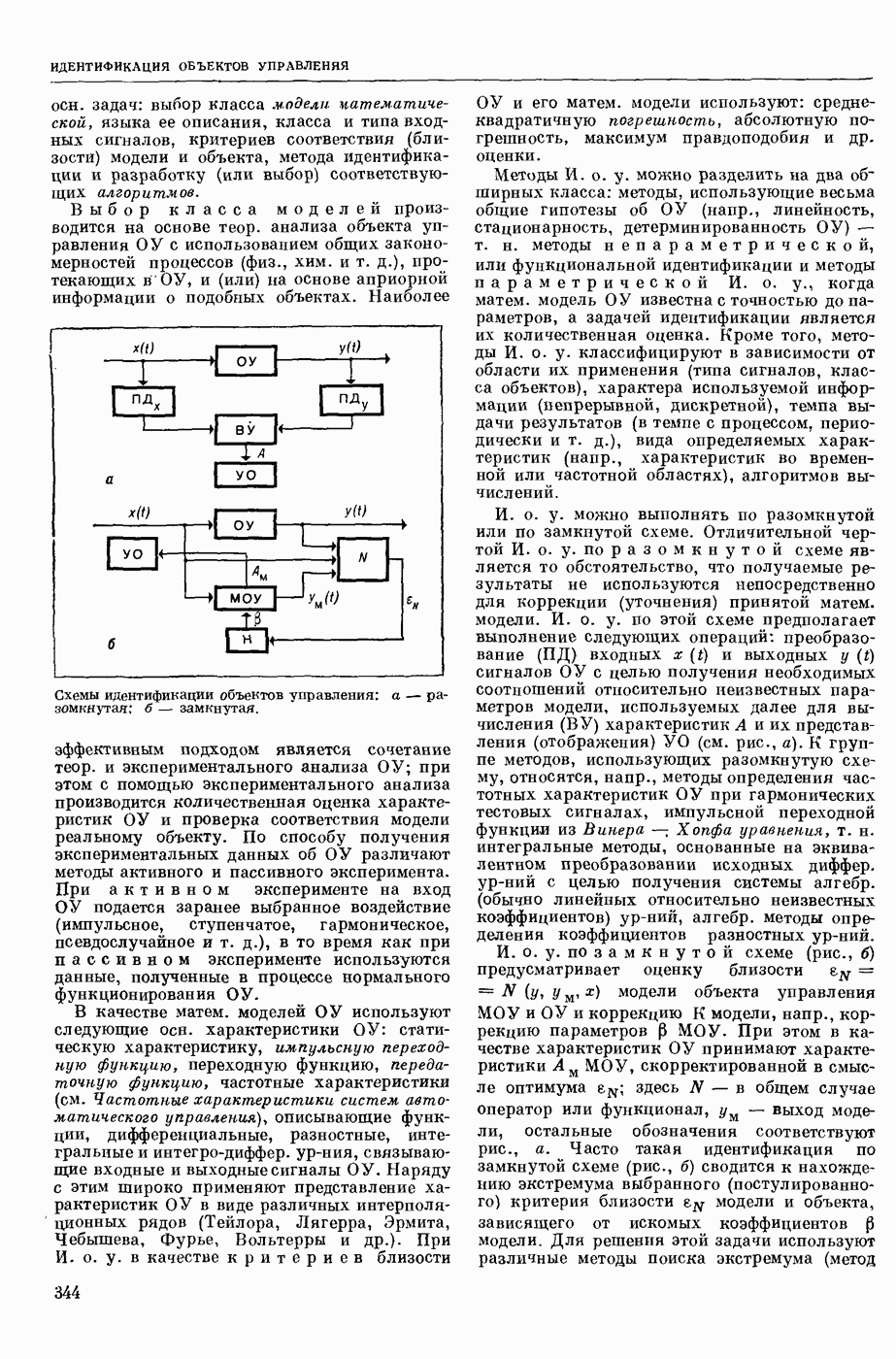
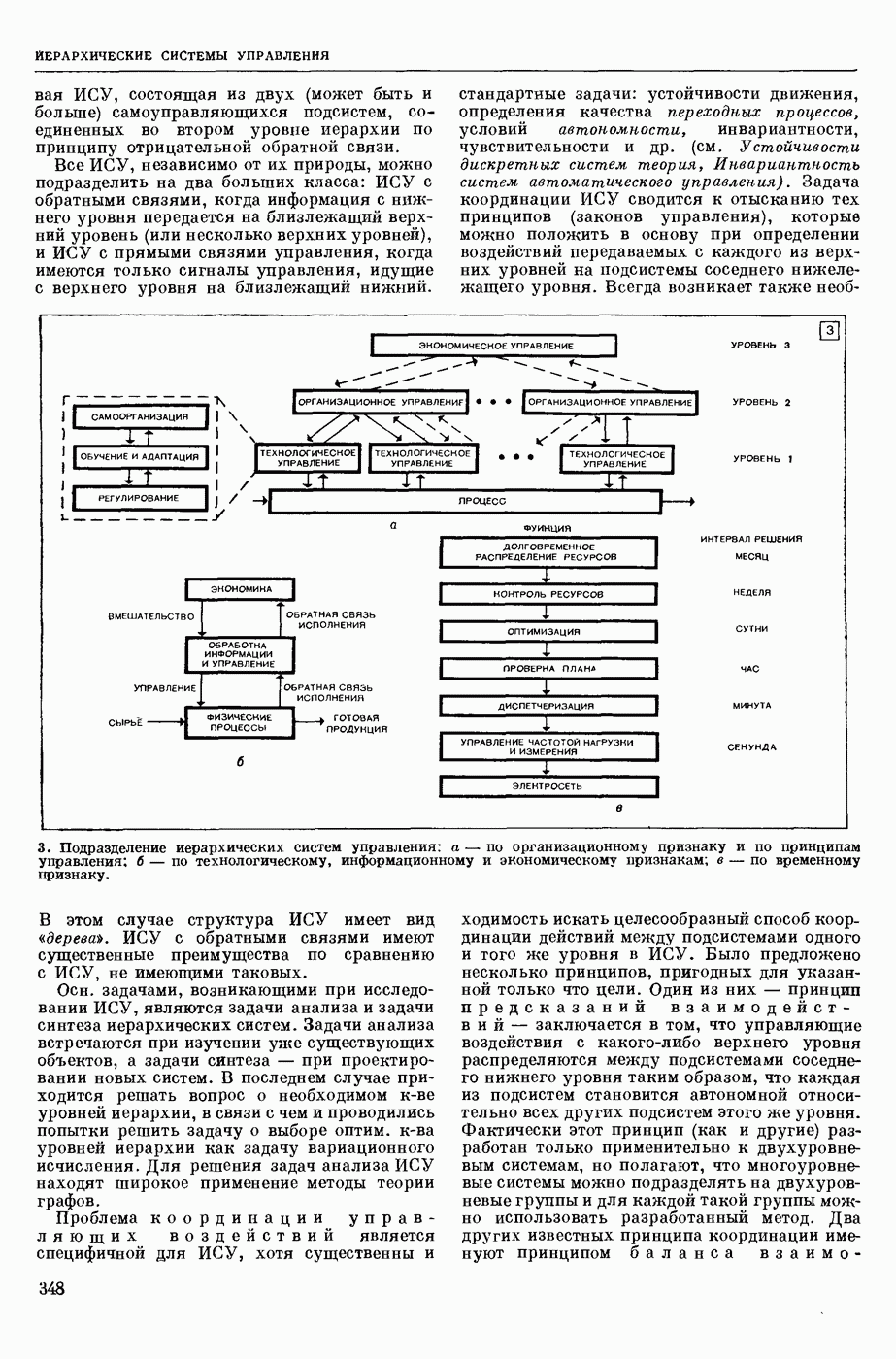
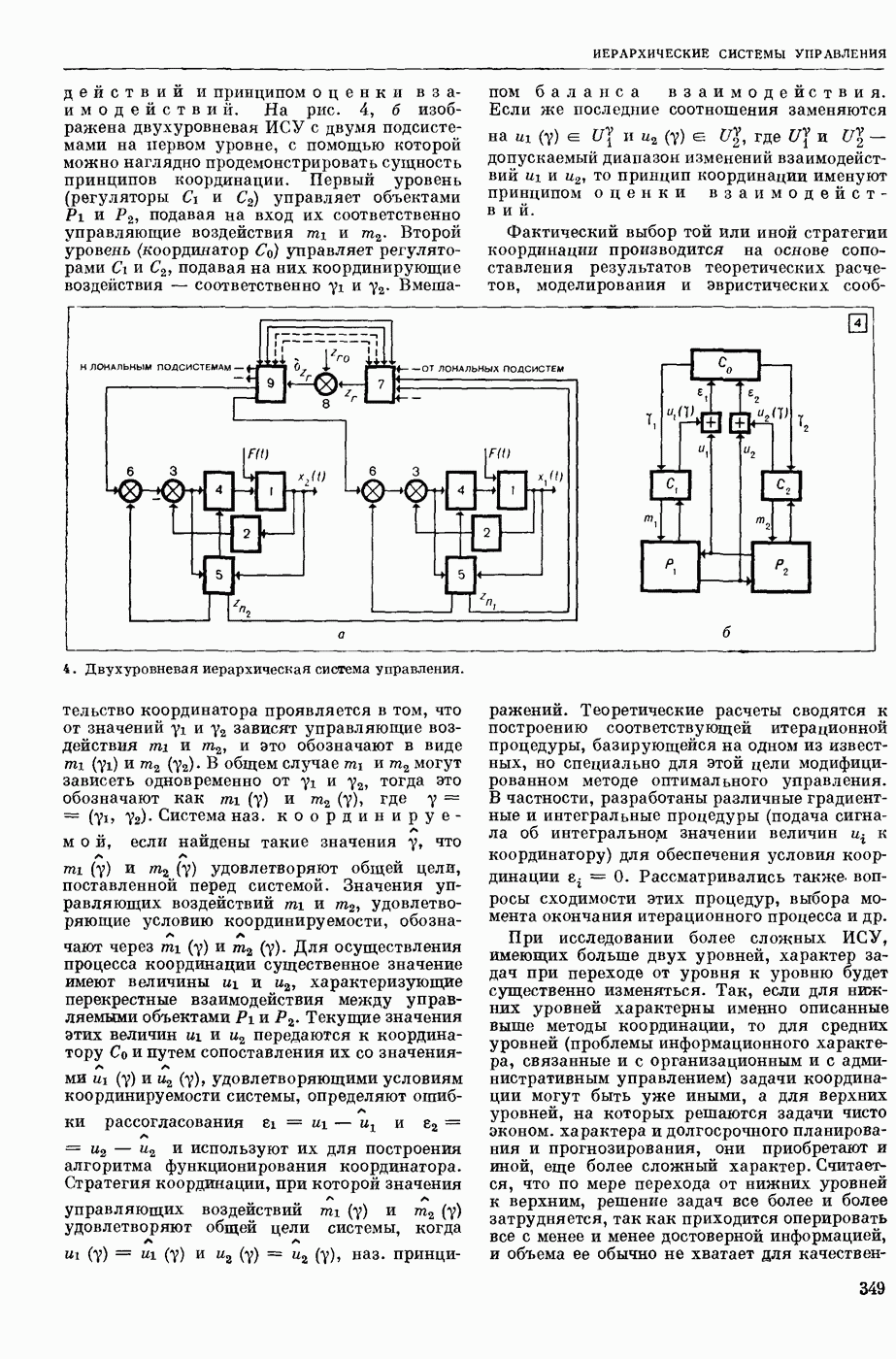
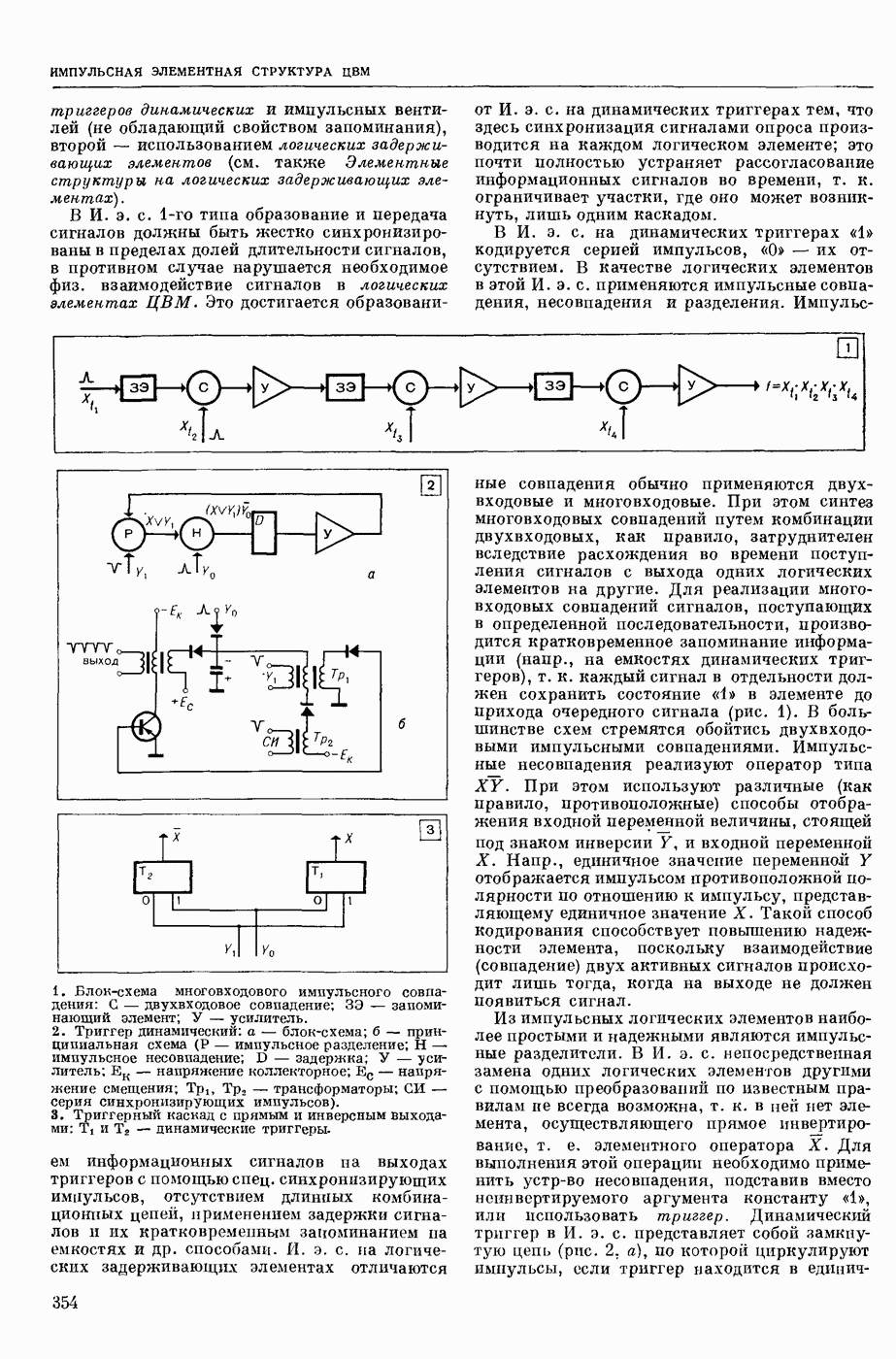
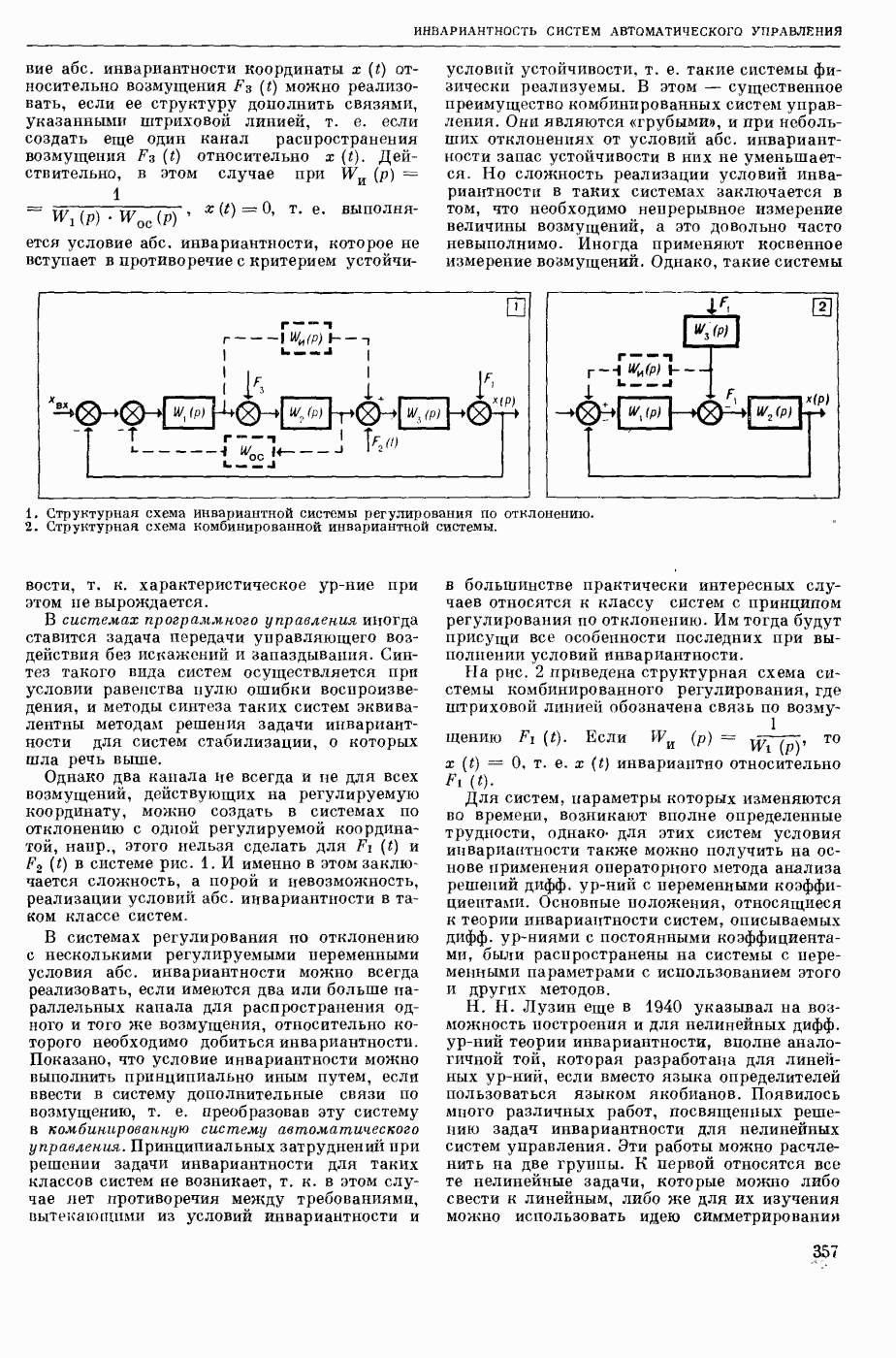
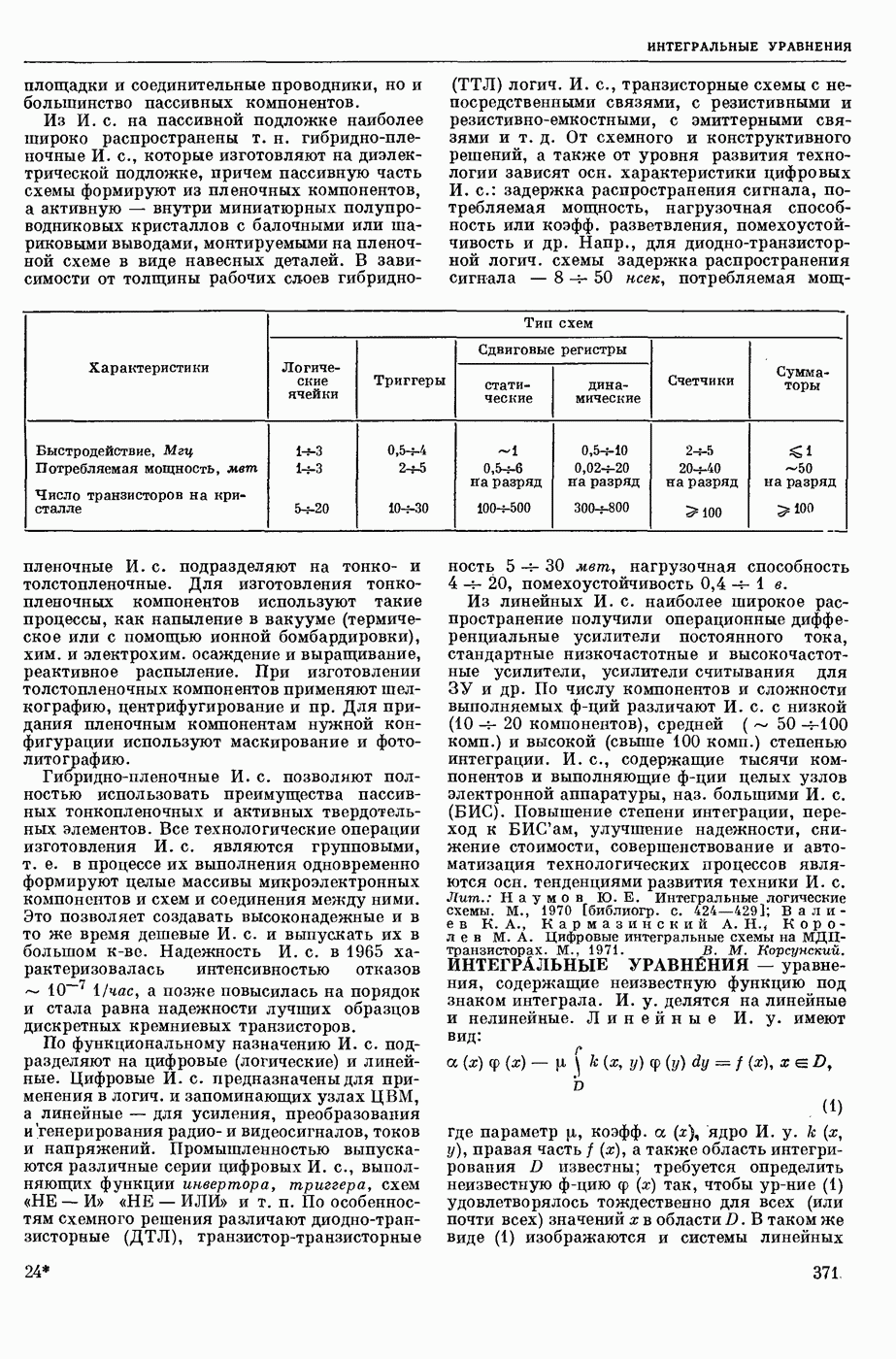
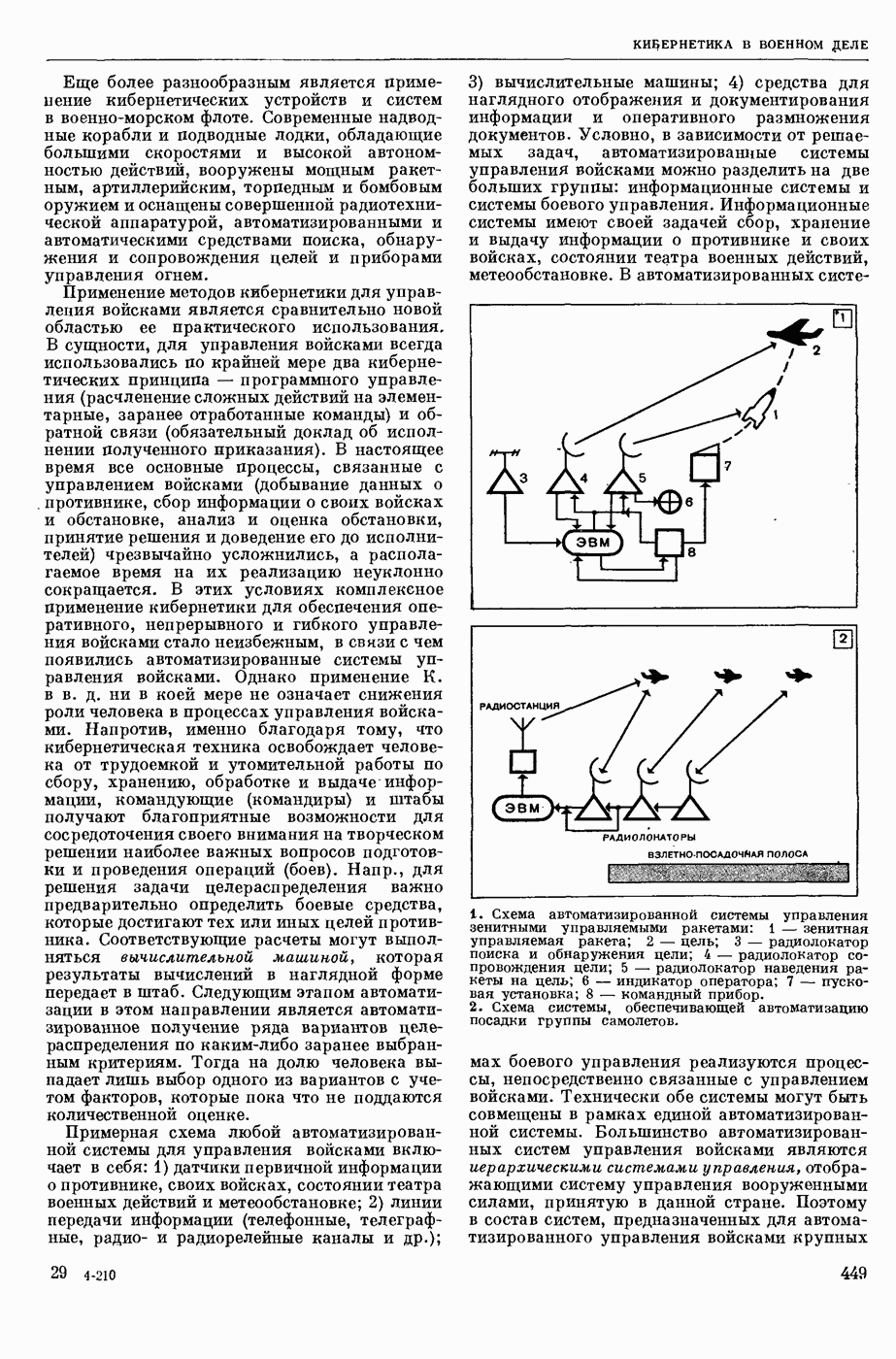
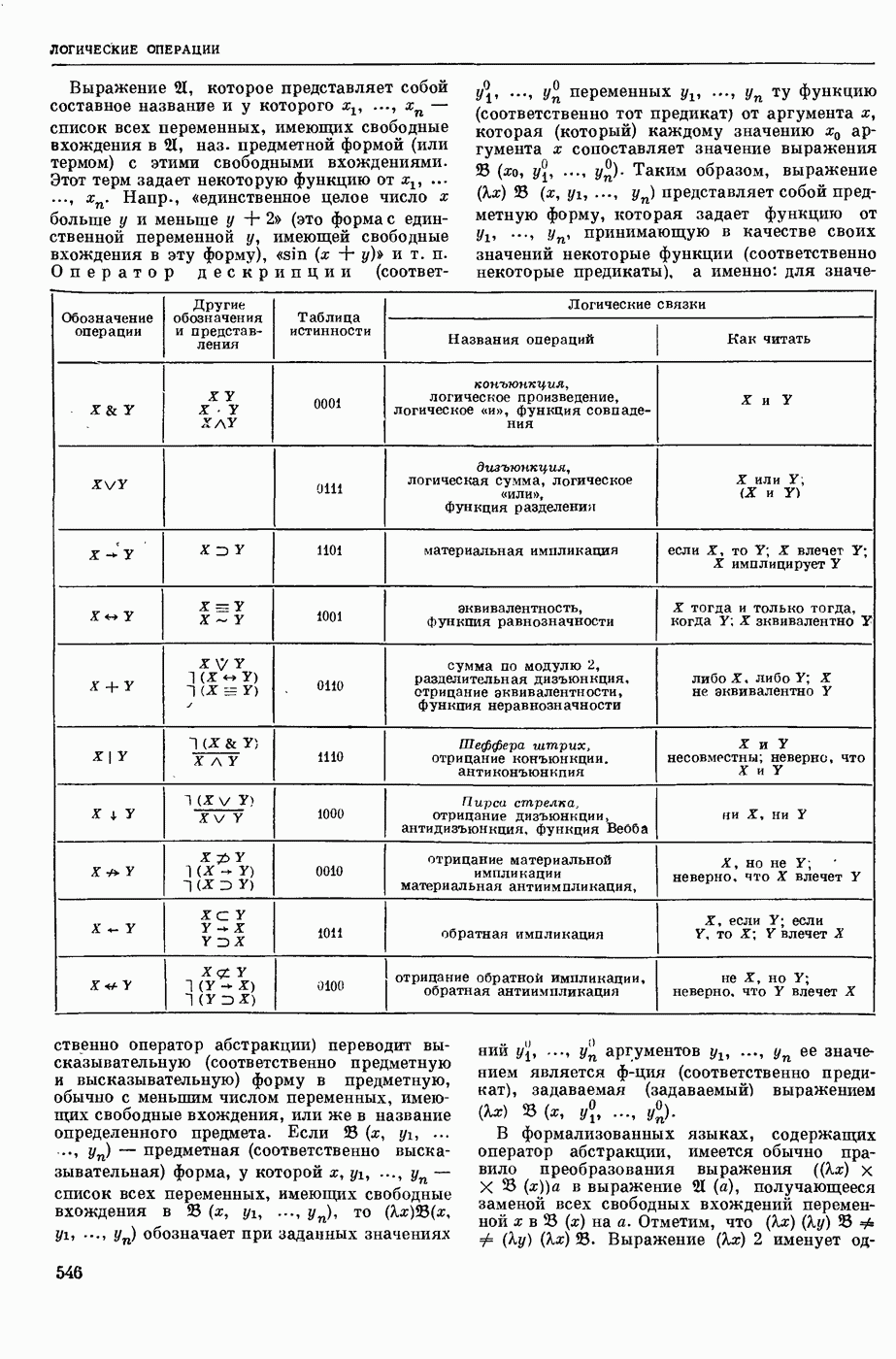
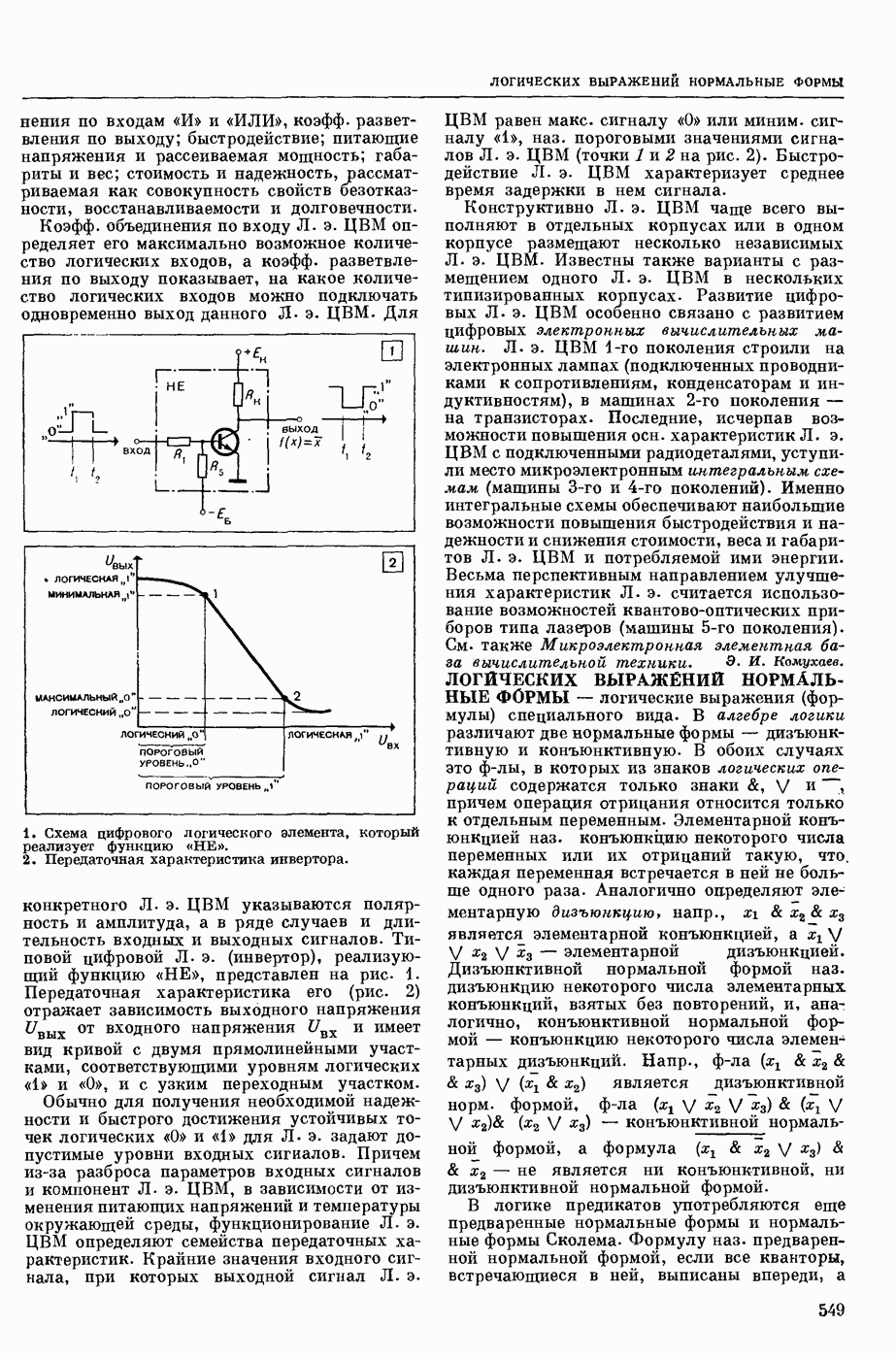
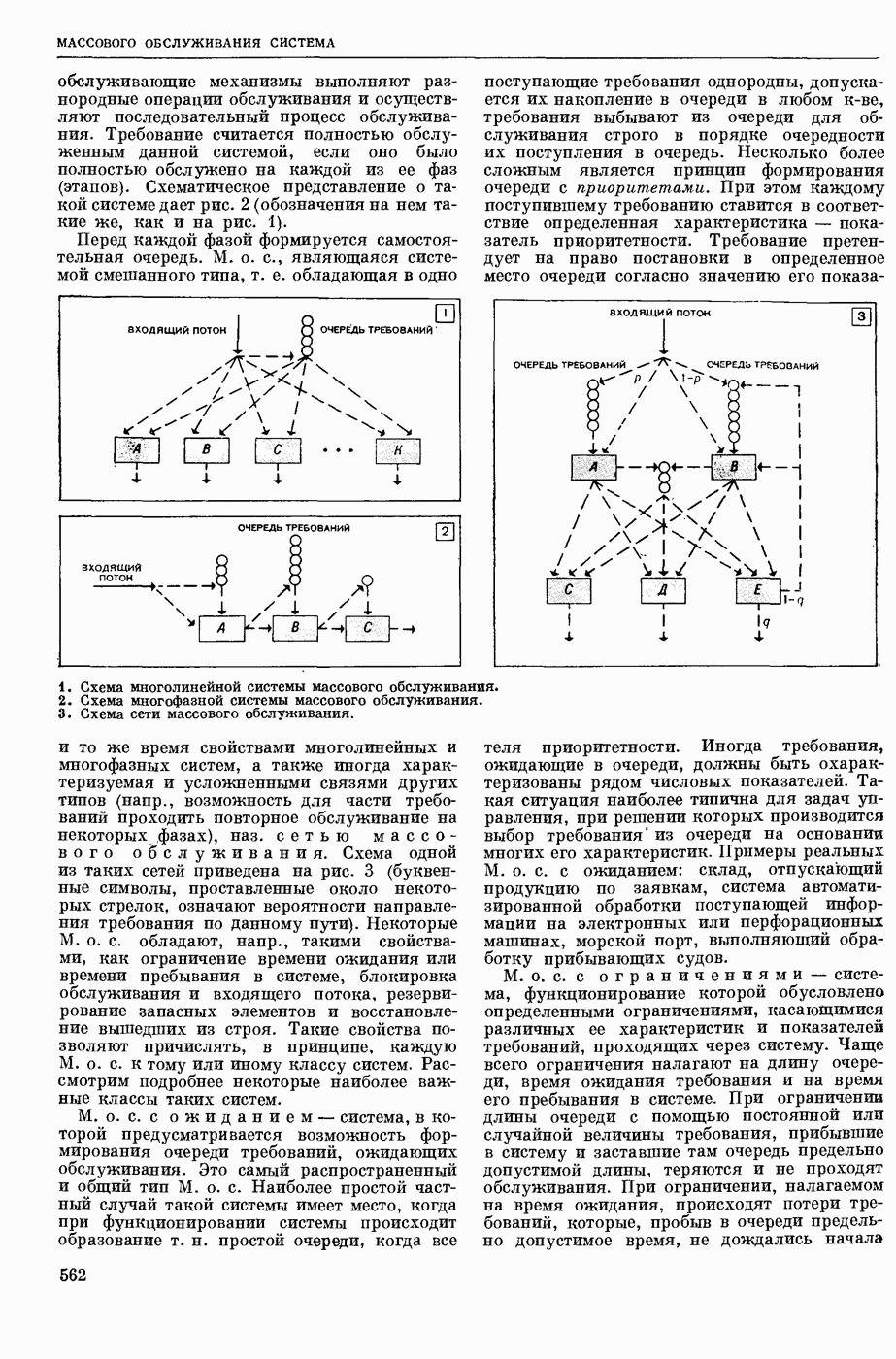
ВОЗБУЖДЕНИЯ КЛЕТКИ ТЕОРИЯ
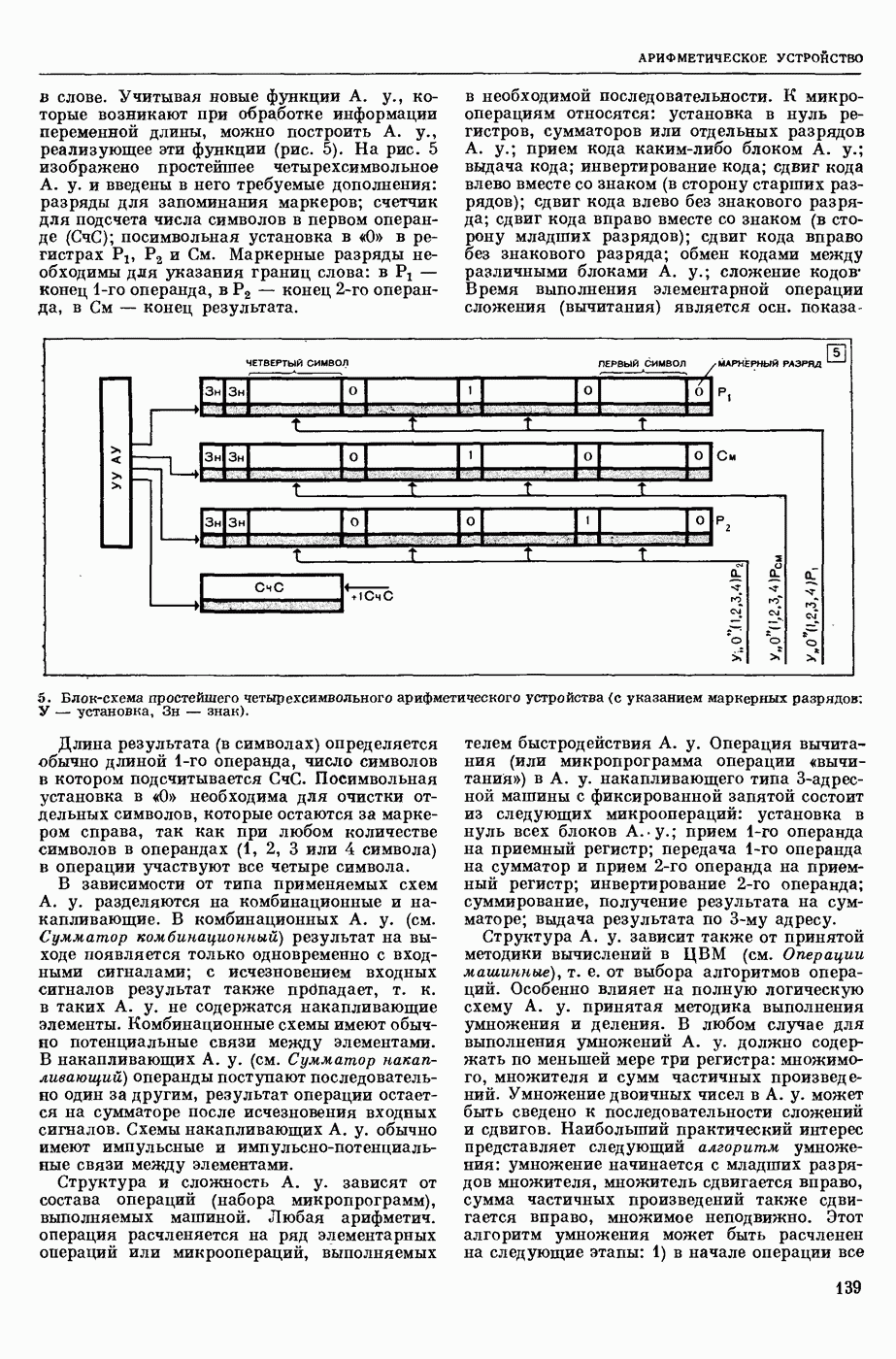
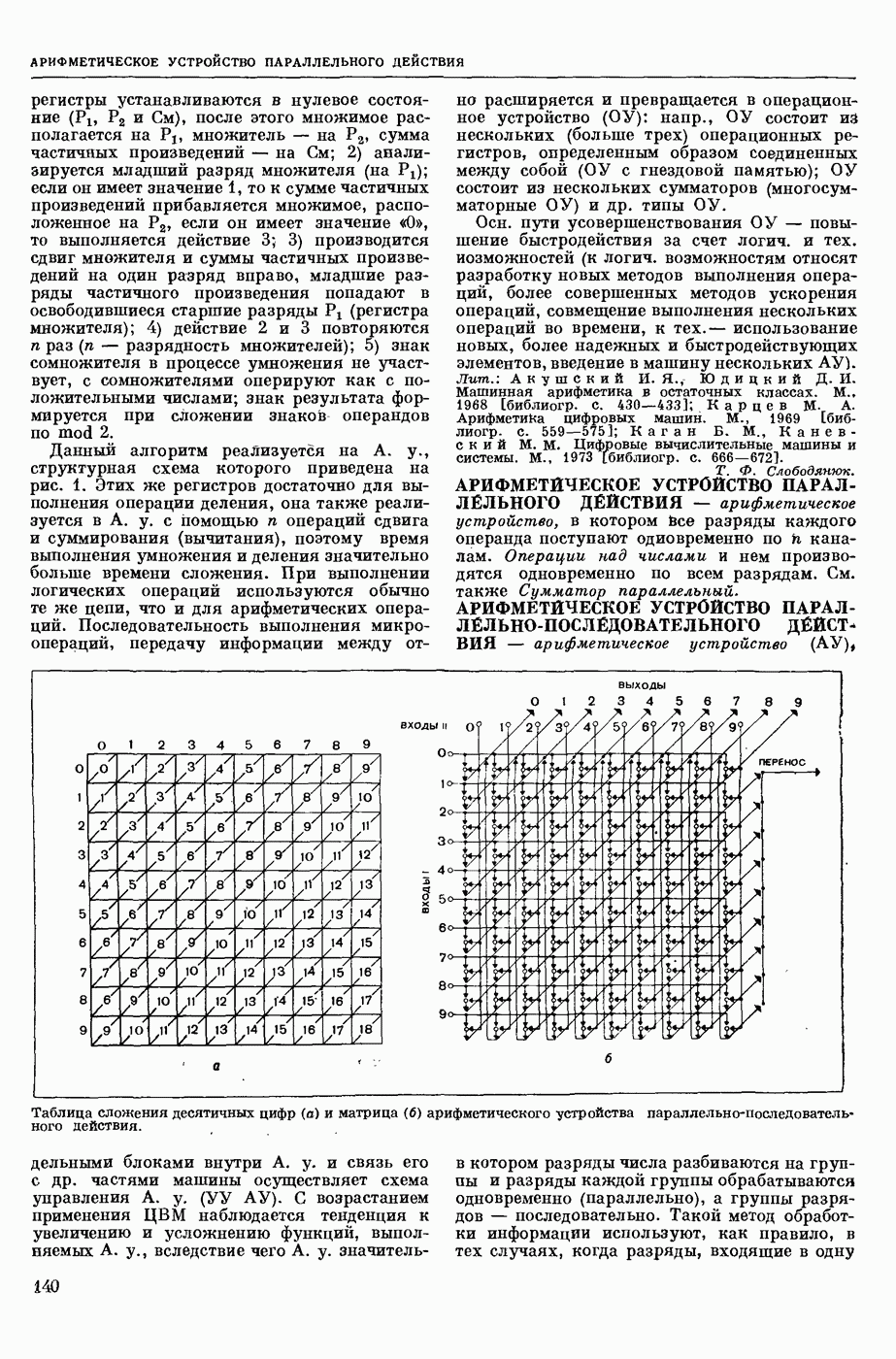
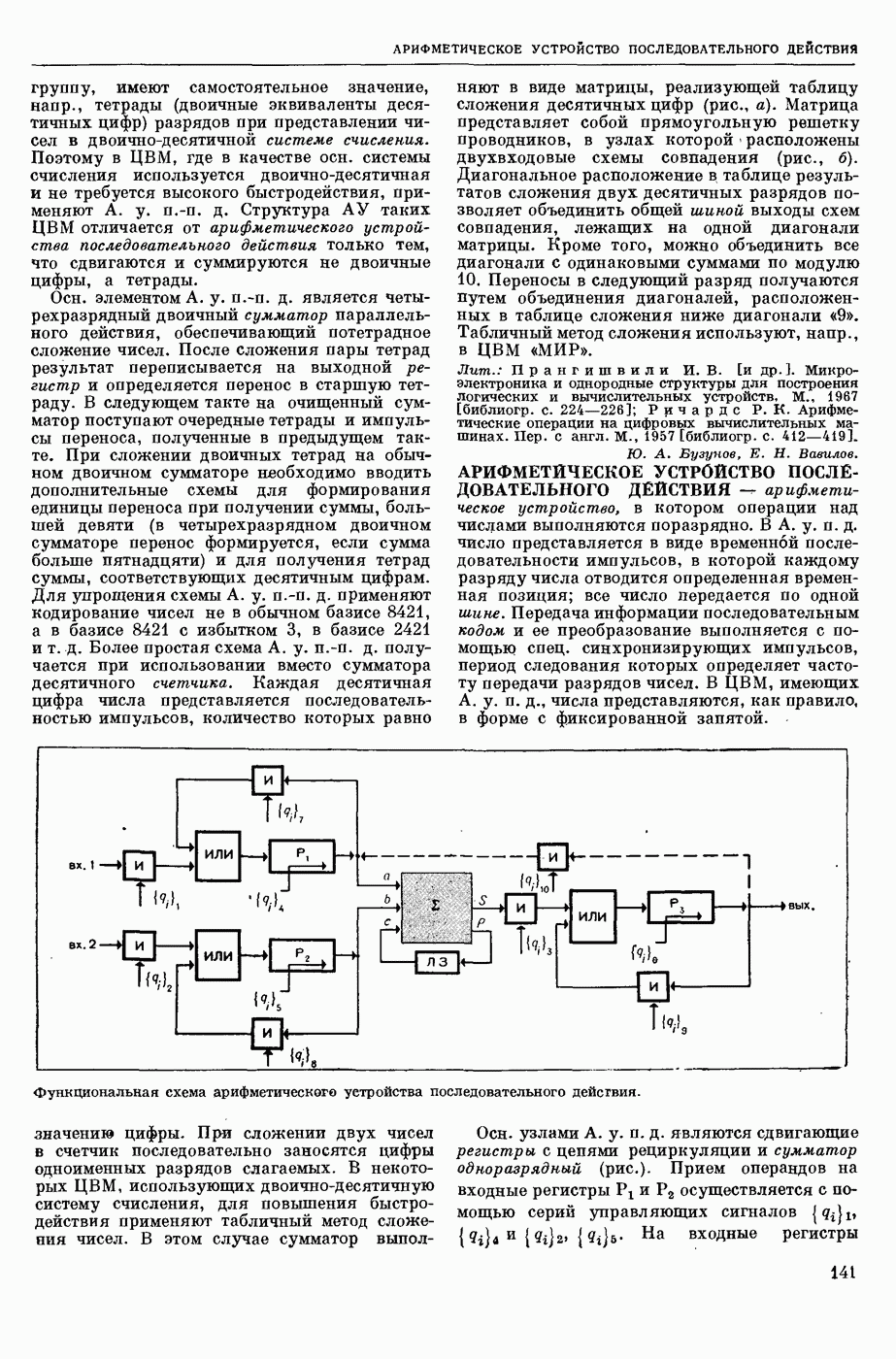
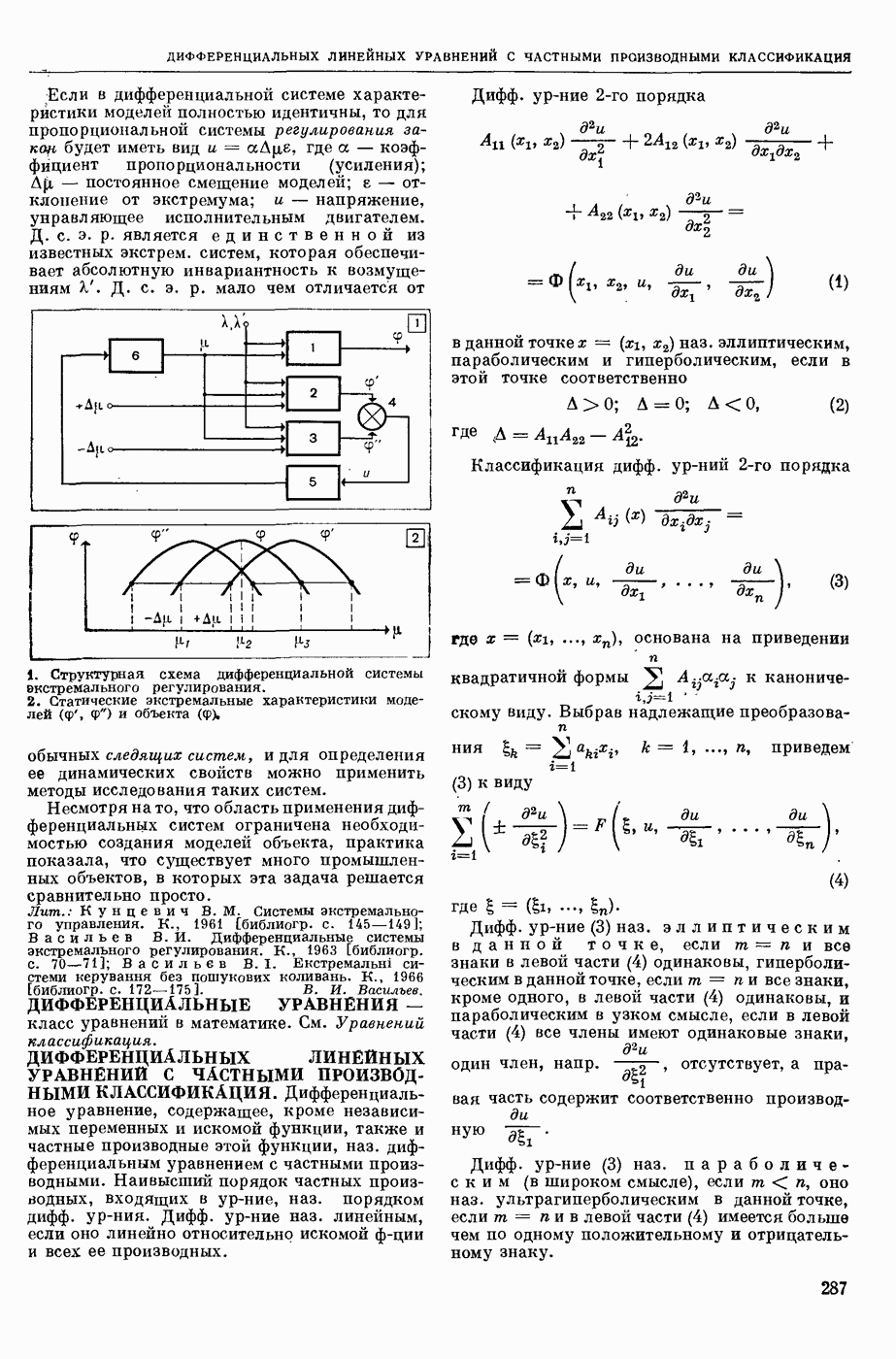
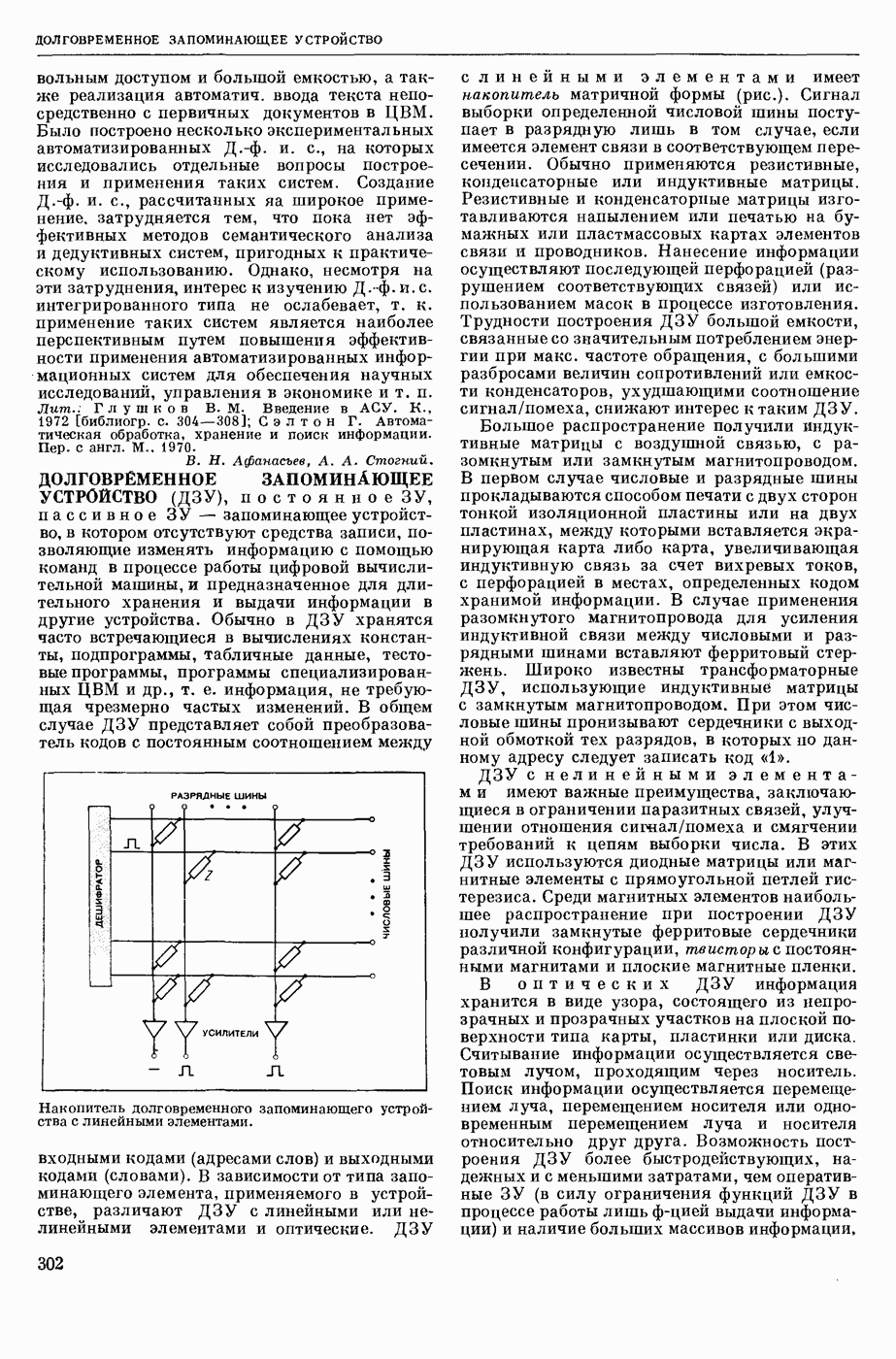
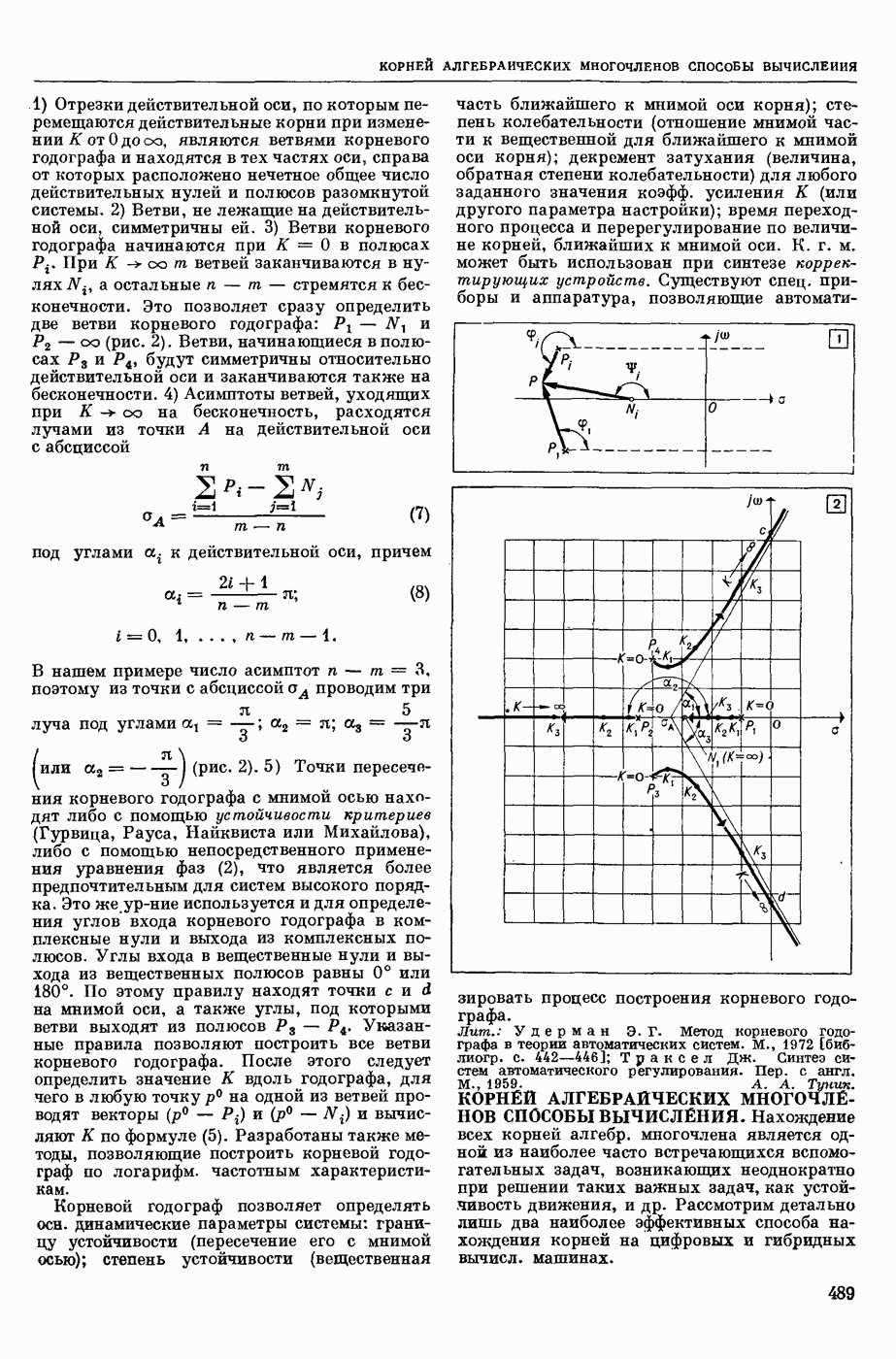
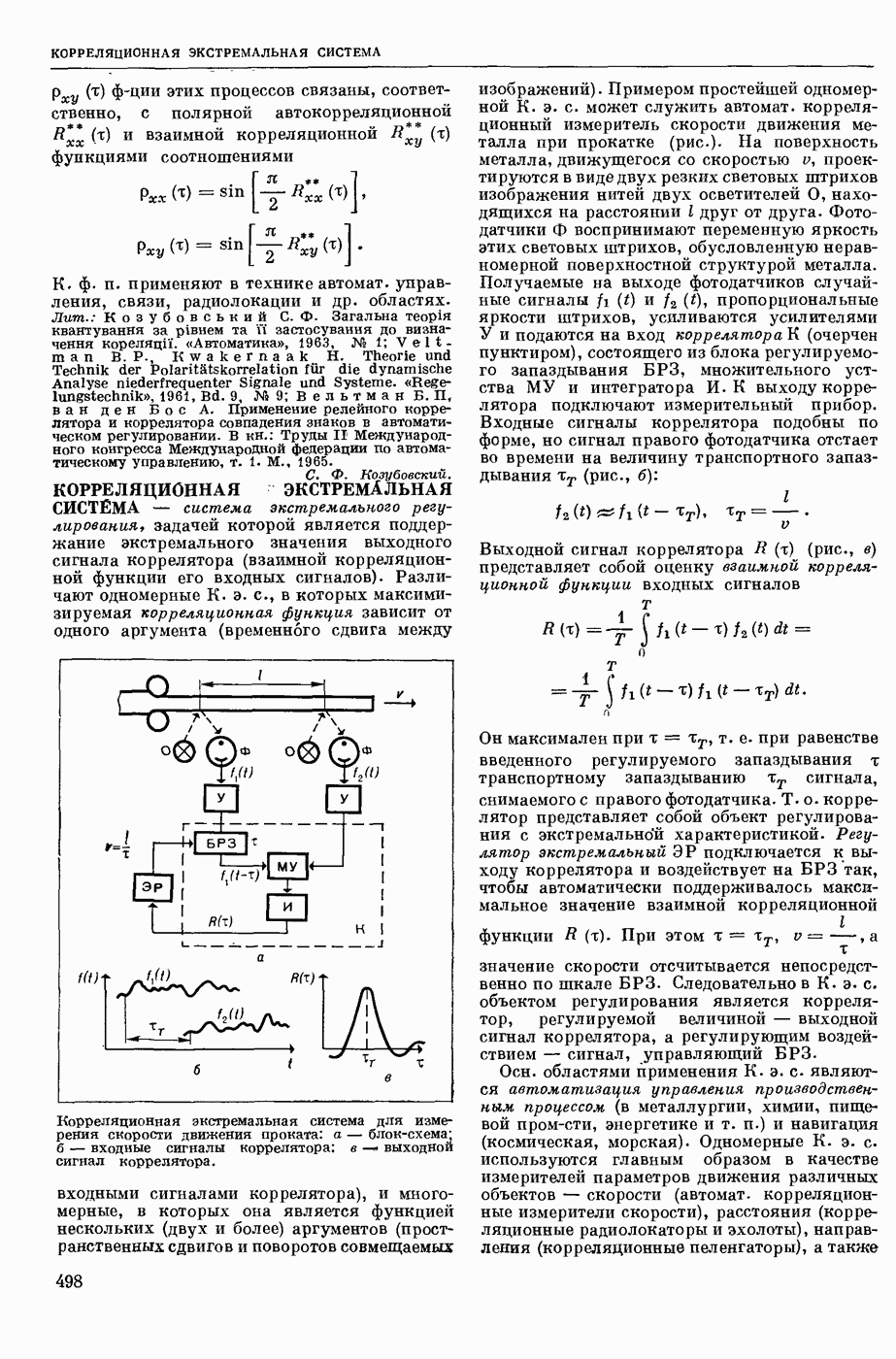
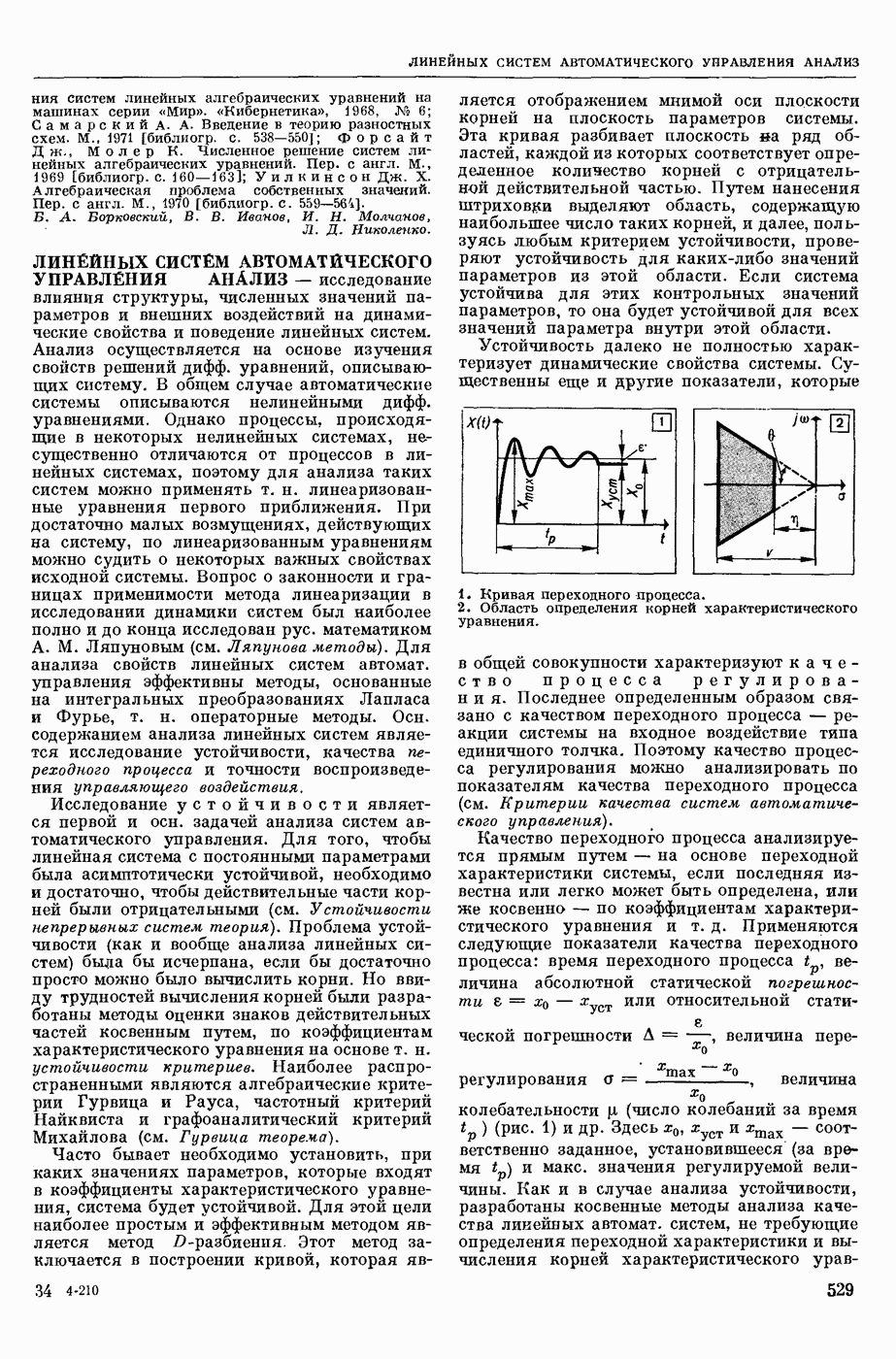
— теория процессов, возникающих в возбудимых клетках под действием раздражителей. Основой возбуждения являются определенные изменения физ.-хим. свойств поверхностных структур мышечных и нервных клеток, придающих возбуждению способность к самораспространению по клетке. Наиболее обоснованную В. к. т. разработали (1949) англ. физиологи А. Ходжкин и А. Хаксли. По этой теории, в поверхностной мембране клетки, находящейся в состоянии покоя, существует электрическая поляризация, обусловленная неравномерным распределением ионов натрия, калия и хлора между протоплазмой клетки и внеклеточной средой. Внеклеточная среда содержит в основном ионы натрия и хлора, а протоплазма — ионы калия. Электр, поля зарядов катионов внутри клетки компенсируются полями зарядов анионных групп аминокислот и белков. Т. к. в состоянии покоя поверхностная мембрана значительно лучше пропускает ионы калия, чем ионы натрия, то вследствие диффузии этих ионов изнутри наружу наружная поверхность мембраны заряжается положительно по отношению к ее внутр. поверхности. Величина возникающей разности потенциалов («мембранного потенциала») близка к потенциалу калиевого электрода


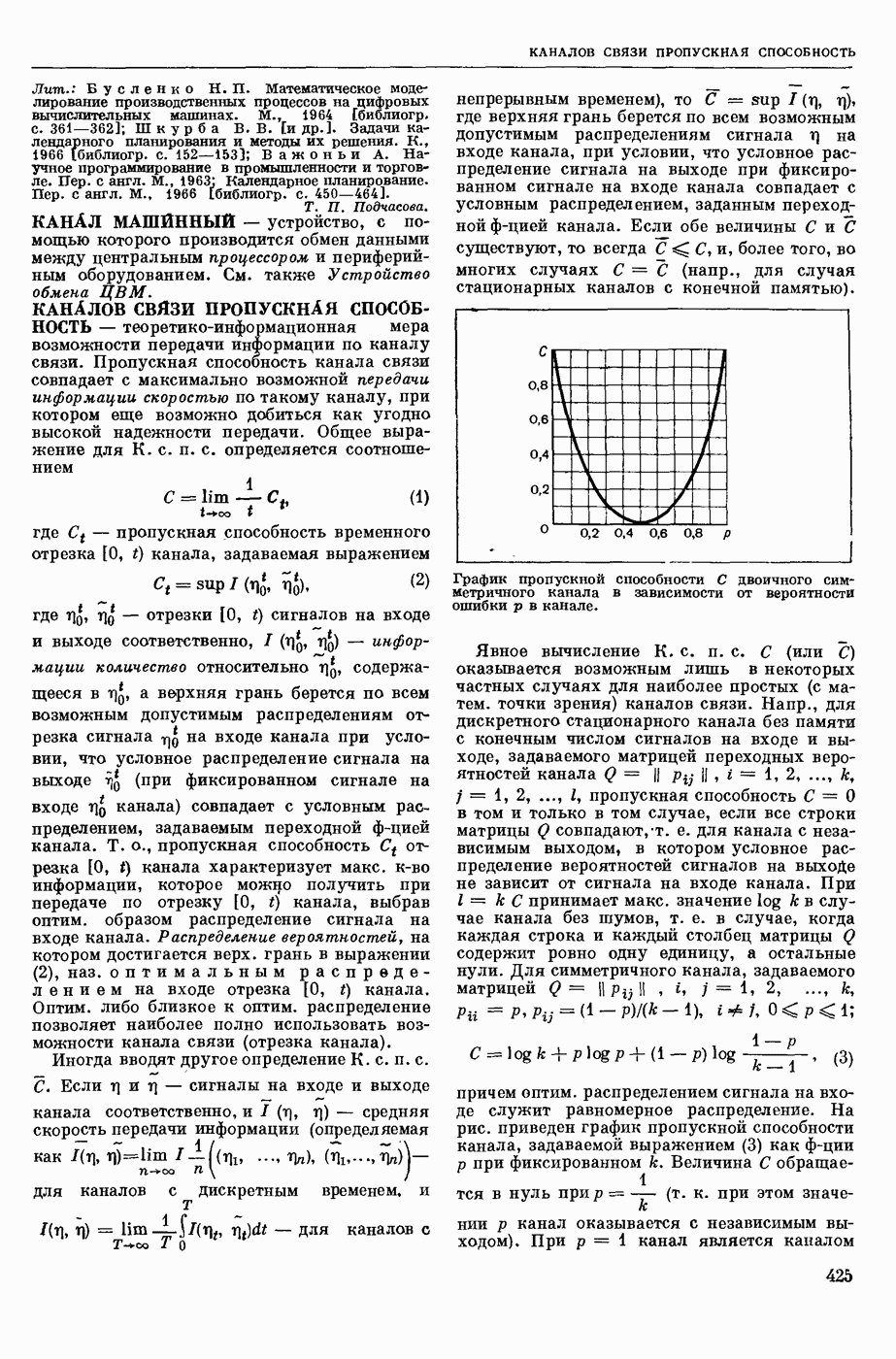
где R — универс. газовая постоянная,  абсол. т-ра,
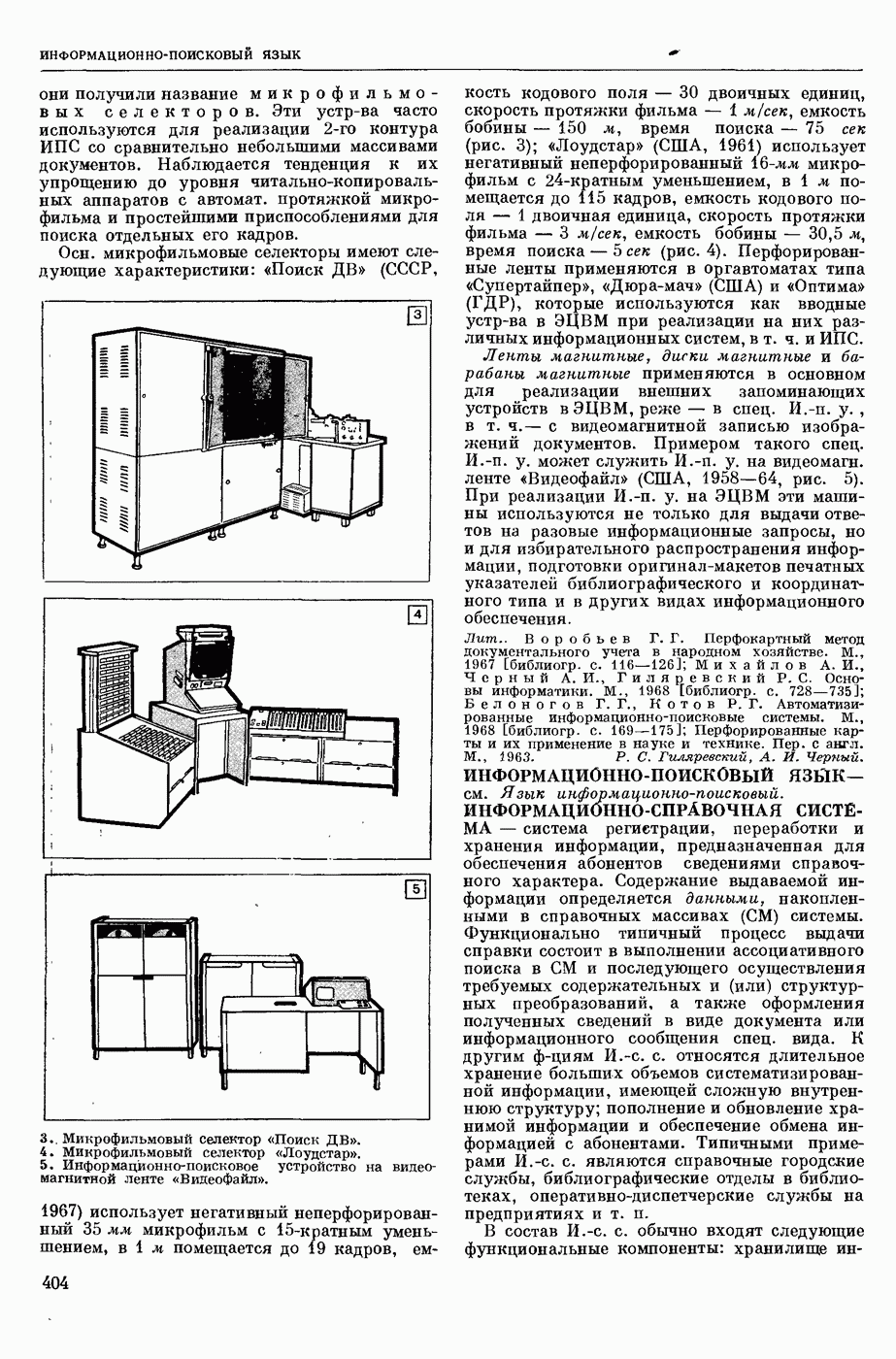
абсол. т-ра,  - число Фарадея,
- число Фарадея,  активность ионов калия внутри и снаружи клетки. Небольшая постоянная утечка ионов через мембрану компенсируется особыми «насосами», которые переносят ионы сквозь нее против градиента потенциала, используя энергию обмена веществ. Непременной причиной возникновения возбуждения является снижение под действием раздражителя мембранного потенциала до определенного критического уровня (порога). Это снижение потенциала (деполяризация) мембраны обусловлено действием на поверхностную мембрану клетки либо особых веществ, выделяемых окончаниями отростков других клеток, образующих с ней синаптические контакты, либо поступлением внеш. энергии от раздражителя и связано с изменениями ионной проницаемости мембраны. Когда деполяризация достигает порога, в мембране происходят структурные изменения (неясной пока природы), которые приводят к существенному повышению
активность ионов калия внутри и снаружи клетки. Небольшая постоянная утечка ионов через мембрану компенсируется особыми «насосами», которые переносят ионы сквозь нее против градиента потенциала, используя энергию обмена веществ. Непременной причиной возникновения возбуждения является снижение под действием раздражителя мембранного потенциала до определенного критического уровня (порога). Это снижение потенциала (деполяризация) мембраны обусловлено действием на поверхностную мембрану клетки либо особых веществ, выделяемых окончаниями отростков других клеток, образующих с ней синаптические контакты, либо поступлением внеш. энергии от раздражителя и связано с изменениями ионной проницаемости мембраны. Когда деполяризация достигает порога, в мембране происходят структурные изменения (неясной пока природы), которые приводят к существенному повышению
ее способности пропускать ионы натрия (в ней как бы открываются специфические «натриевые» каналы или поры). Вследствие этого возникает существенный ток их сквозь мембрану внутрь клетки, переносящий определенное к-во положительных зарядов и деполяризующий мембрану. Такой процесс является самоподдерживающимся, т. к. возникающая деполяризация еще больше увеличивает натриевую проницаемость и т. д. В итоге ток ионов натрия не только снижает существовавший на мембране мембранный потенциал, обусловленный ионами калия, но и создает разность потенциалов противоположного направления, близкую к потенциалу натриевого электрода:

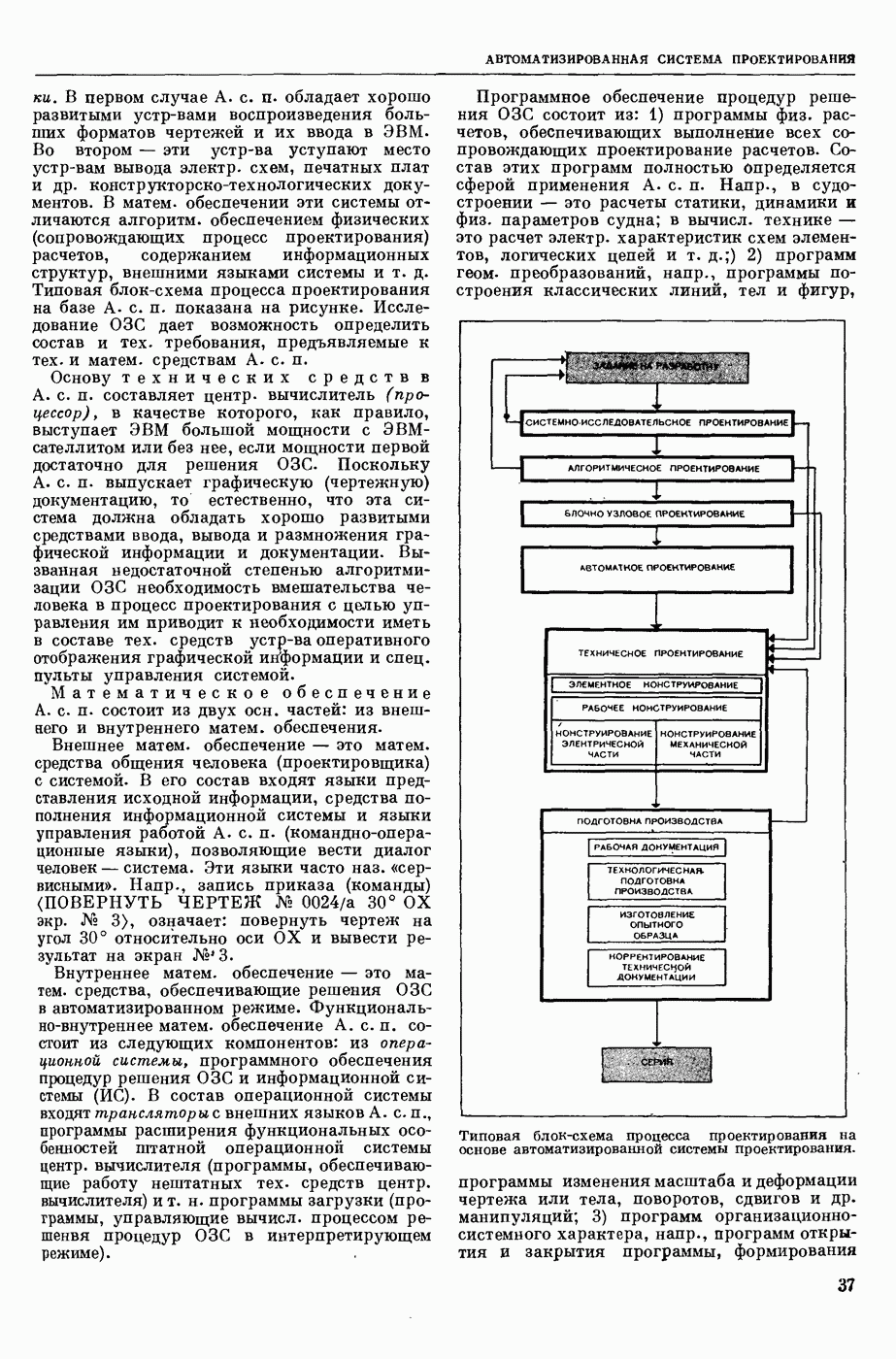
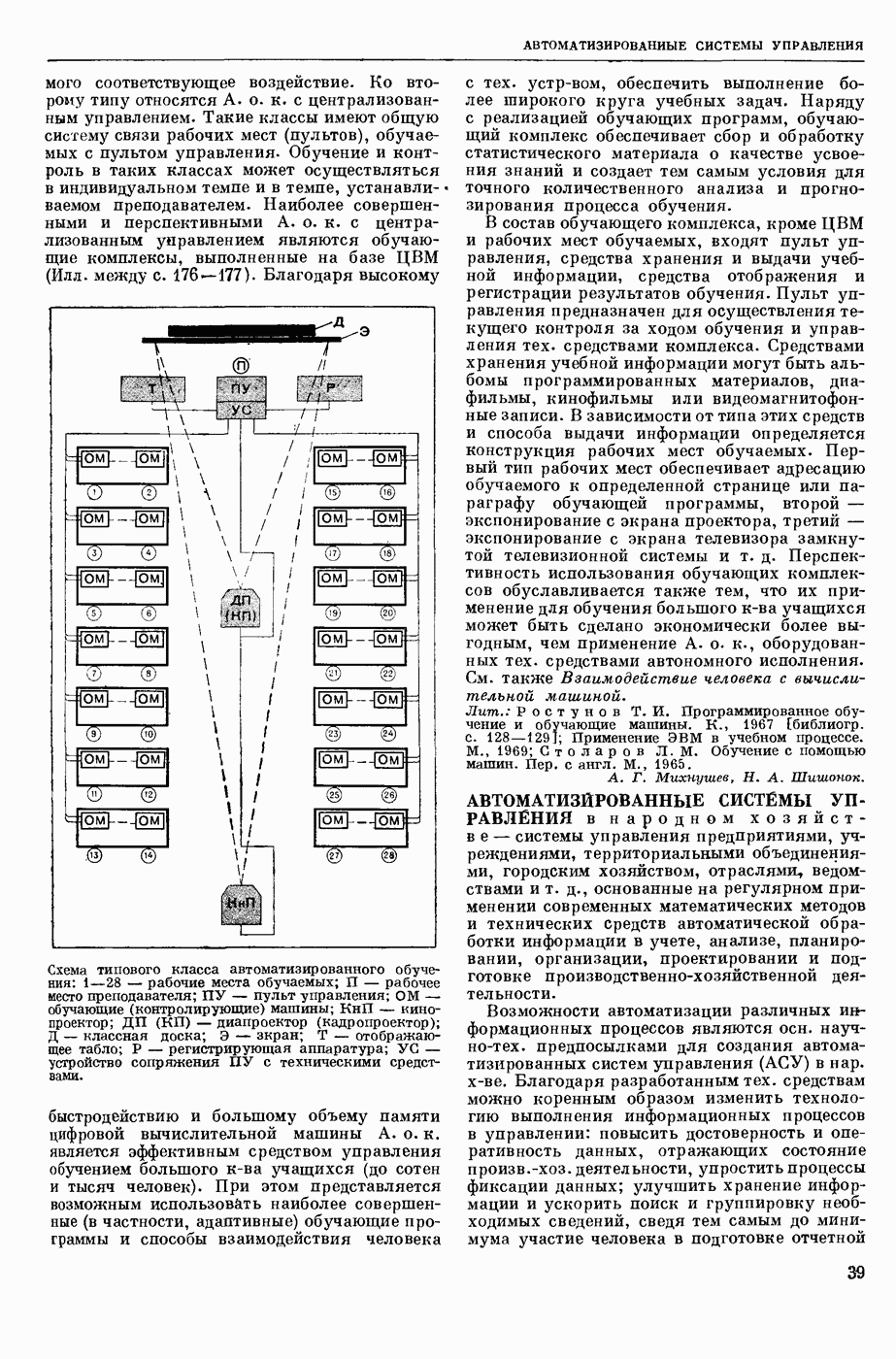
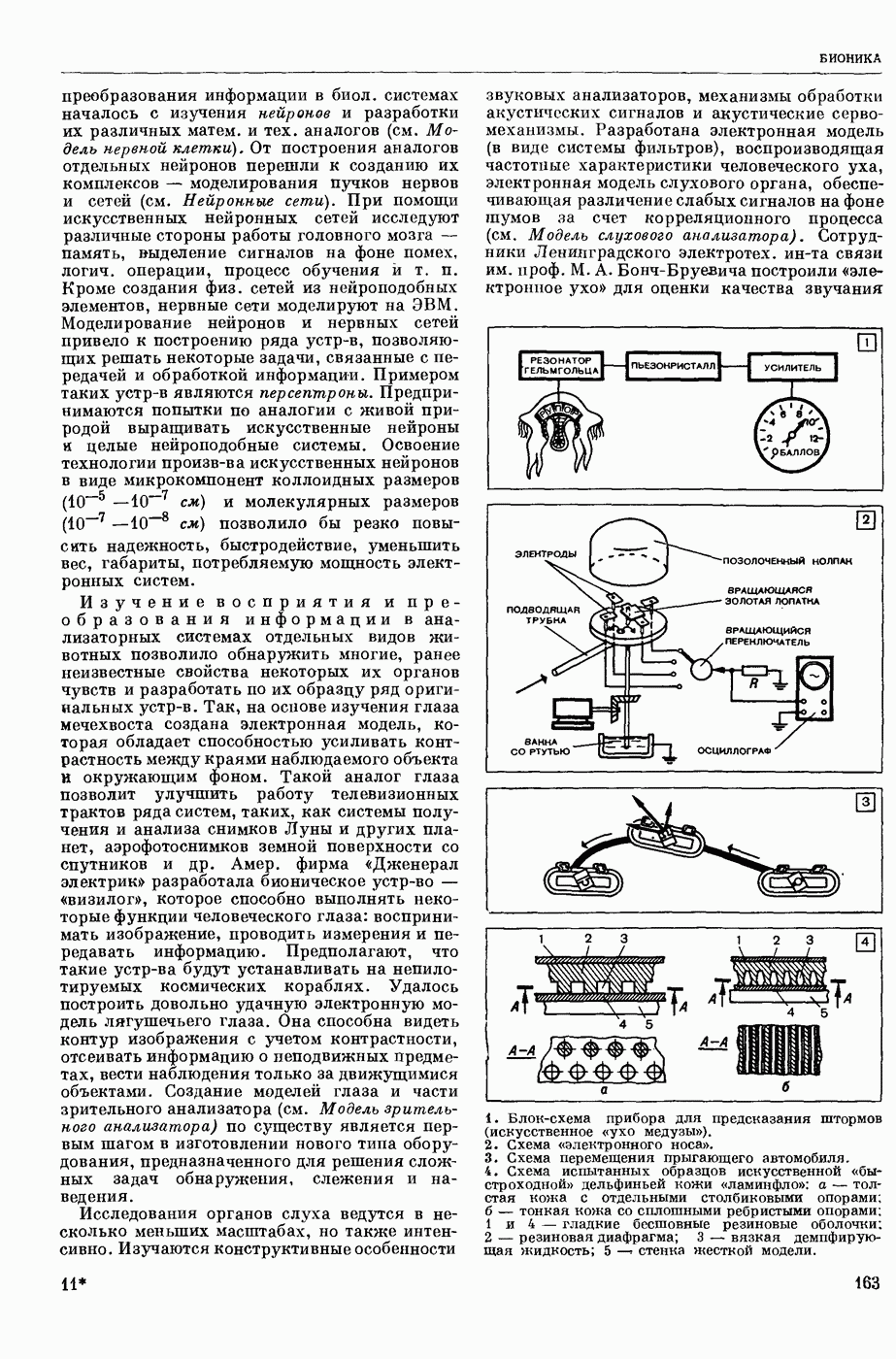
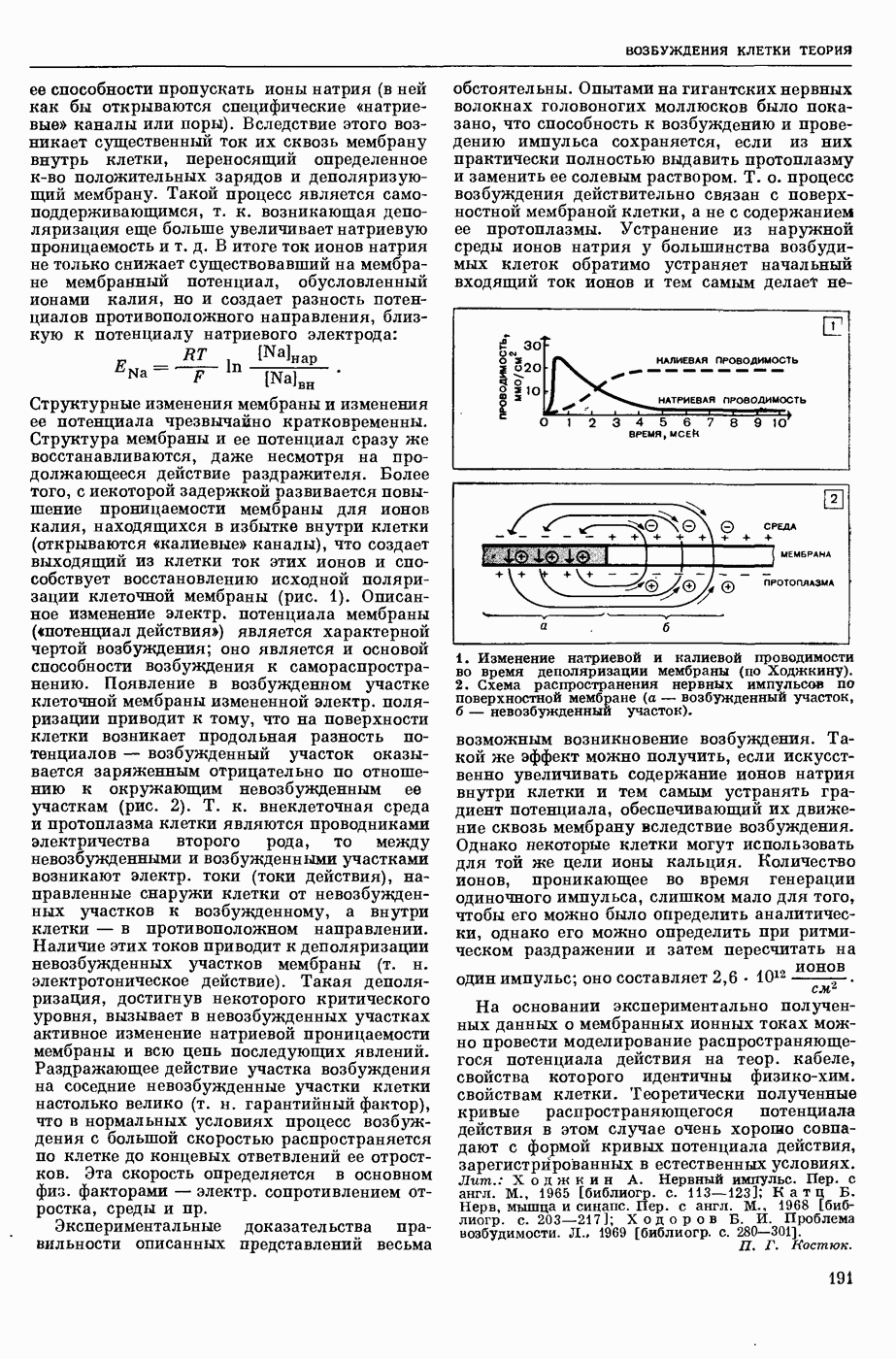
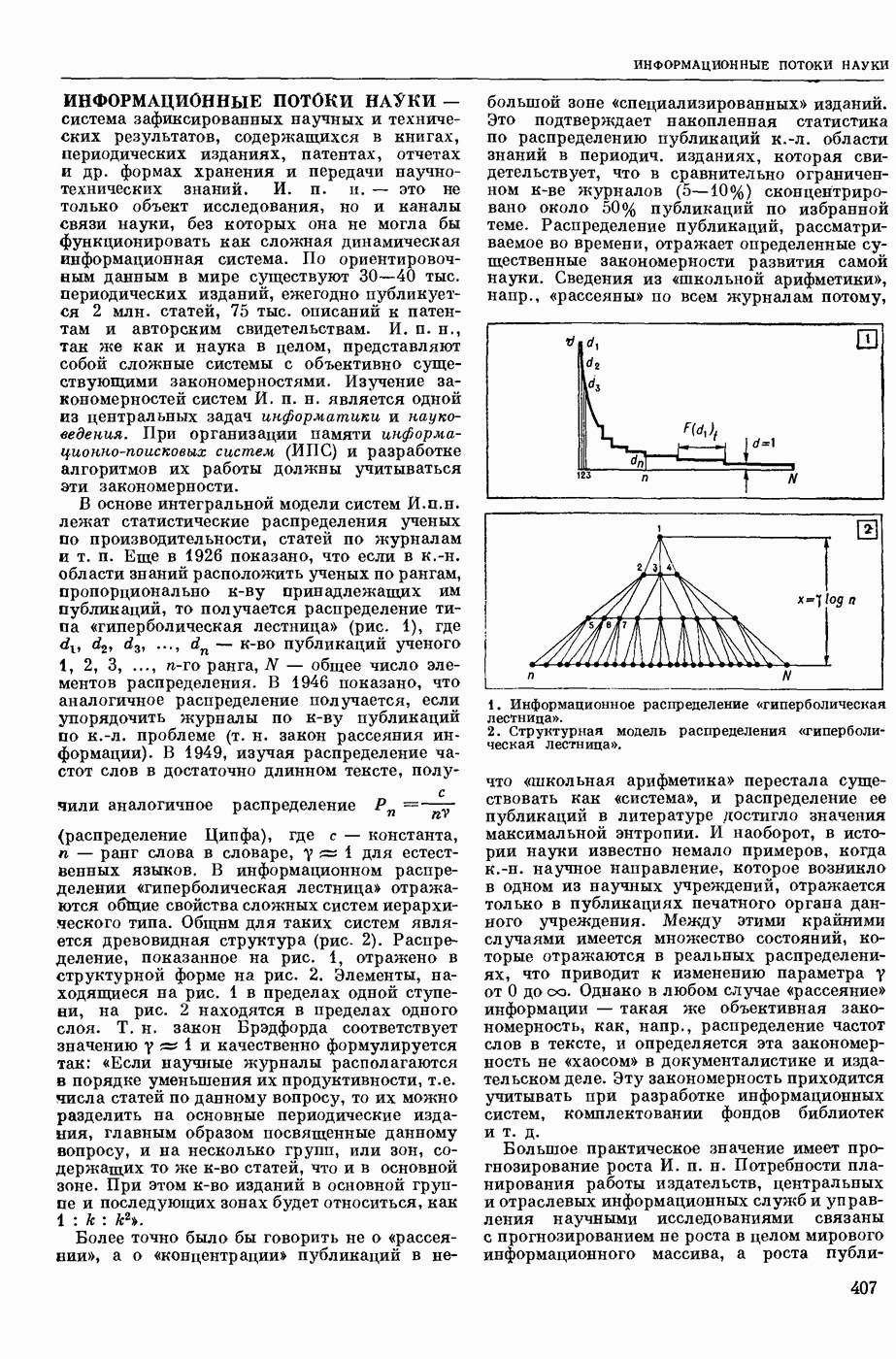
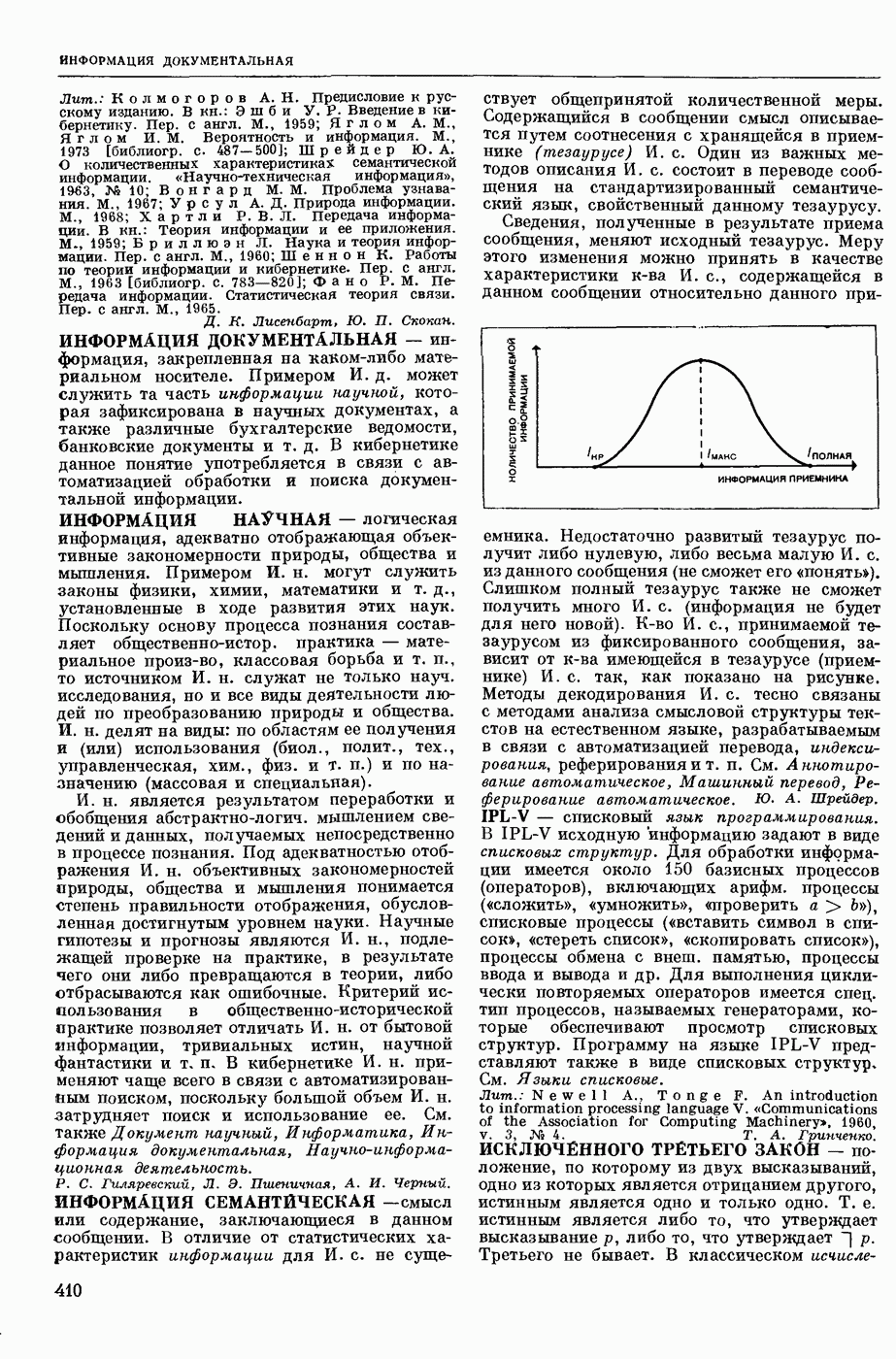
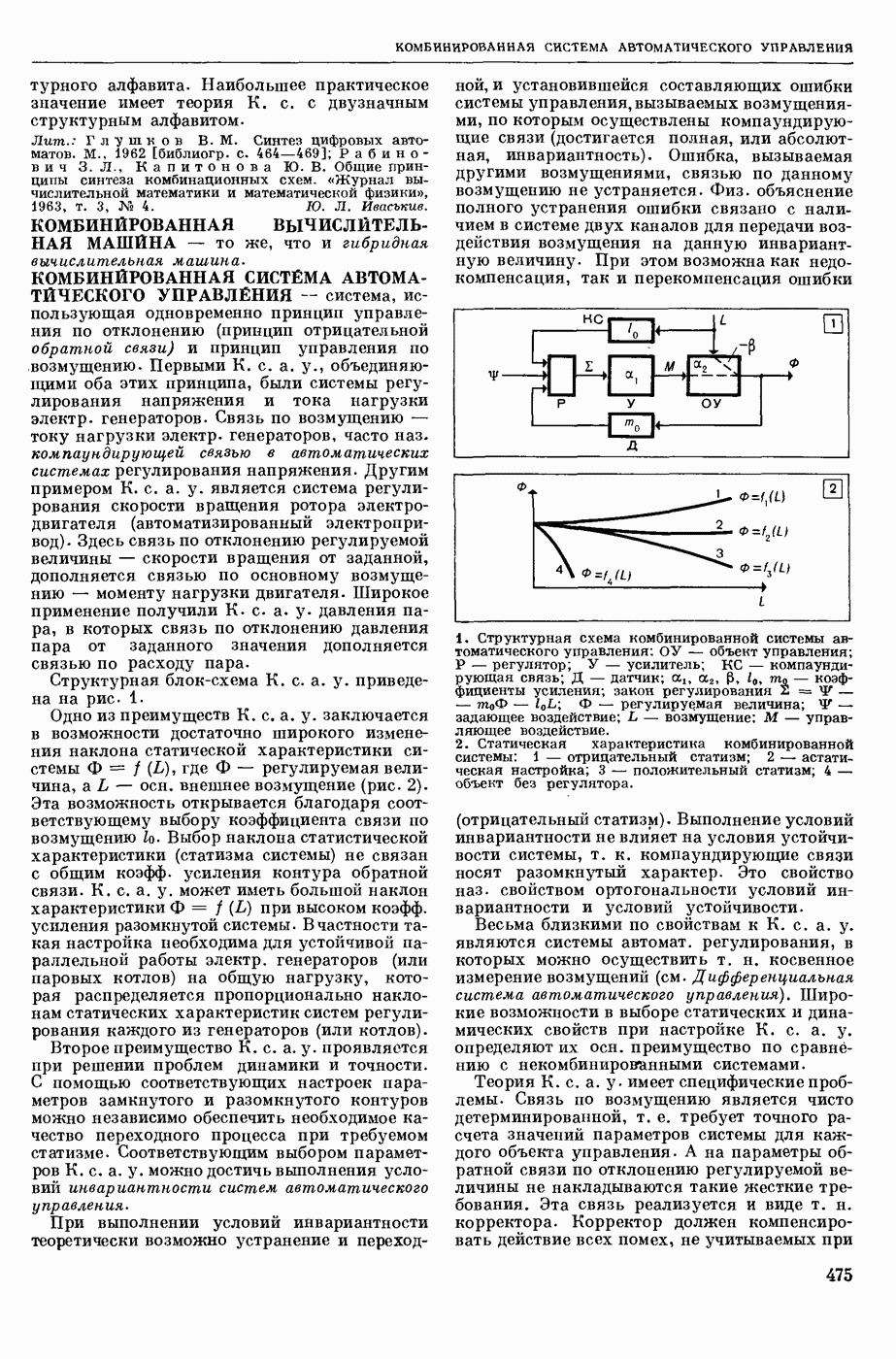
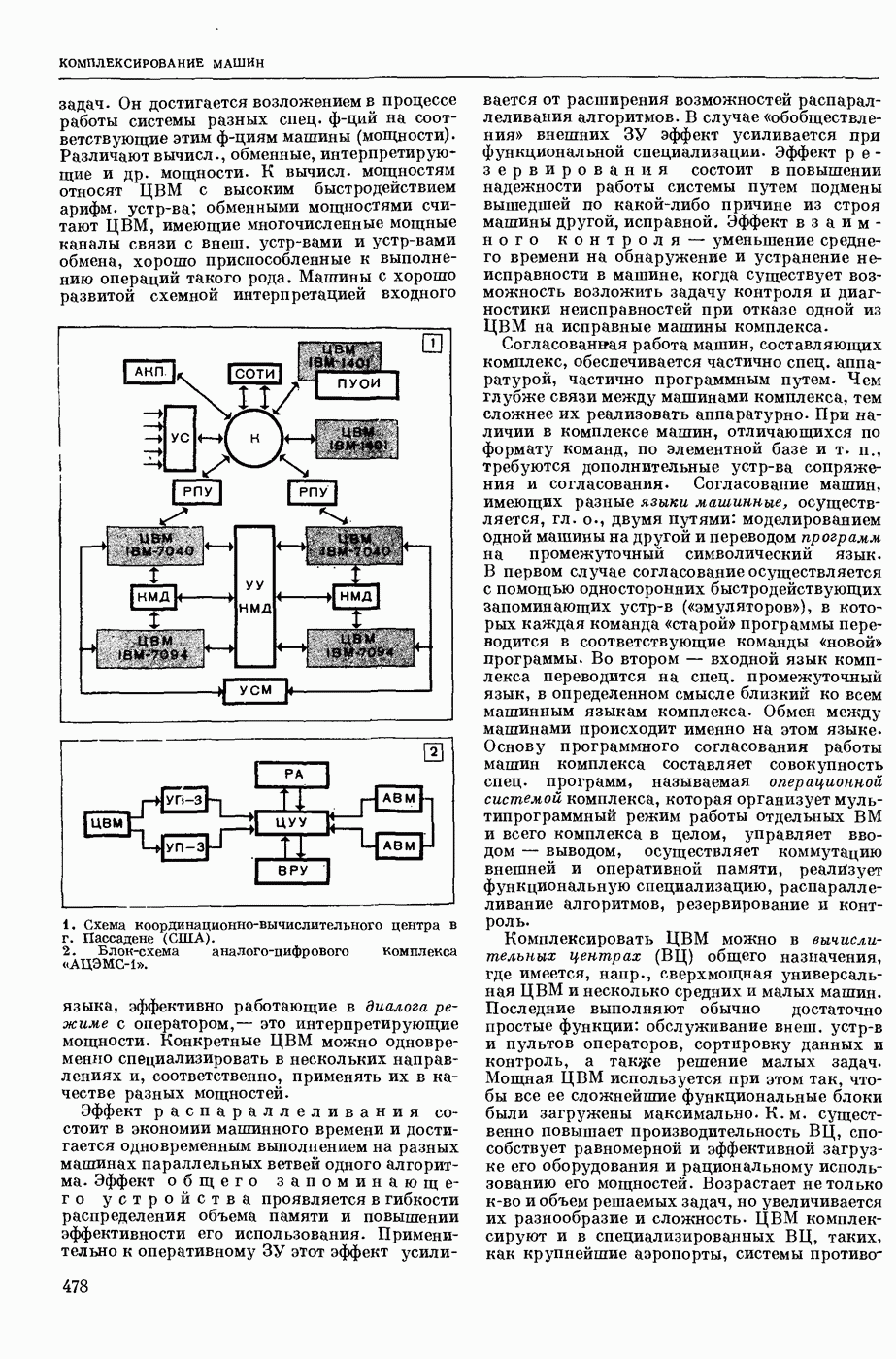
Структурные изменения мембраны и изменения ее потенциала чрезвычайно кратковременны. Структура мембраны и ее потенциал сразу же восстанавливаются, даже несмотря на продолжающееся действие раздражителя. Более того, с некоторой задержкой развивается повышение проницаемости мембраны для ионов калия, находящихся в избытке внутри клетки (открываются «калиевые» каналы), что создает выходящий из клетки ток этих ионов и способствует восстановлению исходной поляризации клеточной мембраны (рис. 1). Описанное изменение электр. потенциала мембраны («потенциал действия») является характерной чертой возбуждения; оно является и основой способности возбуждения к самораспространению. Появление в возбуяеденном участке клеточной мембраны измененной электр. поляризации приводит к тому, что на поверхности клетки возникает продольная разность потенциалов — возбужденный участок оказывается заряженным отрицательно по отношению к окружающим невозбужденным ее участкам (рис. 2). Т. к. внеклеточная среда и протоплазма клетки являются проводниками электричества второго рода, то между невозбужденными и возбужденными участками возникают электр. токи (токи действия), направленные снаружи клетки от невозбужденных участков к возбужденному, а внутри клетки — в противоположном направлении. Наличие этих токов приводит к деполяризации невозбужденных участков мембраны (т. н. электротоническое действие). Такая деполяризация, достигнув некоторого критического уровня, вызывает в невозбужденных участках активное изменение натриевой проницаемости мембраны и всю цепь последующих явлений. Раздражающее действие участка возбуждения на соседние невозбужденные участки клетки настолько велико (т. н. гарантийный фактор), что в нормальных условиях процесс возбуждения с большой скоростью распространяется по клетке до концевых ответвлений ее отростков. Эта скорость определяется в основном физ. факторами — электр. сопротивлением отростка, среды и пр.
Экспериментальные доказательства правильности описанных представлений весьма обстоятельны. Опытами на гигантских нервных волокнах головоногих моллюсков было показано, что способность к возбуждению и проведению импульса сохраняется, если из них практически полностью выдавить протоплазму и заменить ее солевым раствором. Т. о. процесс возбуждения действительно связан с поверхностной мембраной клетки, а не с содержанием ее протоплазмы. Устранение из наружной среды ионов натрия у большинства возбудимых клеток обратимо устраняет начальный входящий ток ионов и тем самым делает невозможным возникновение возбуяедения.

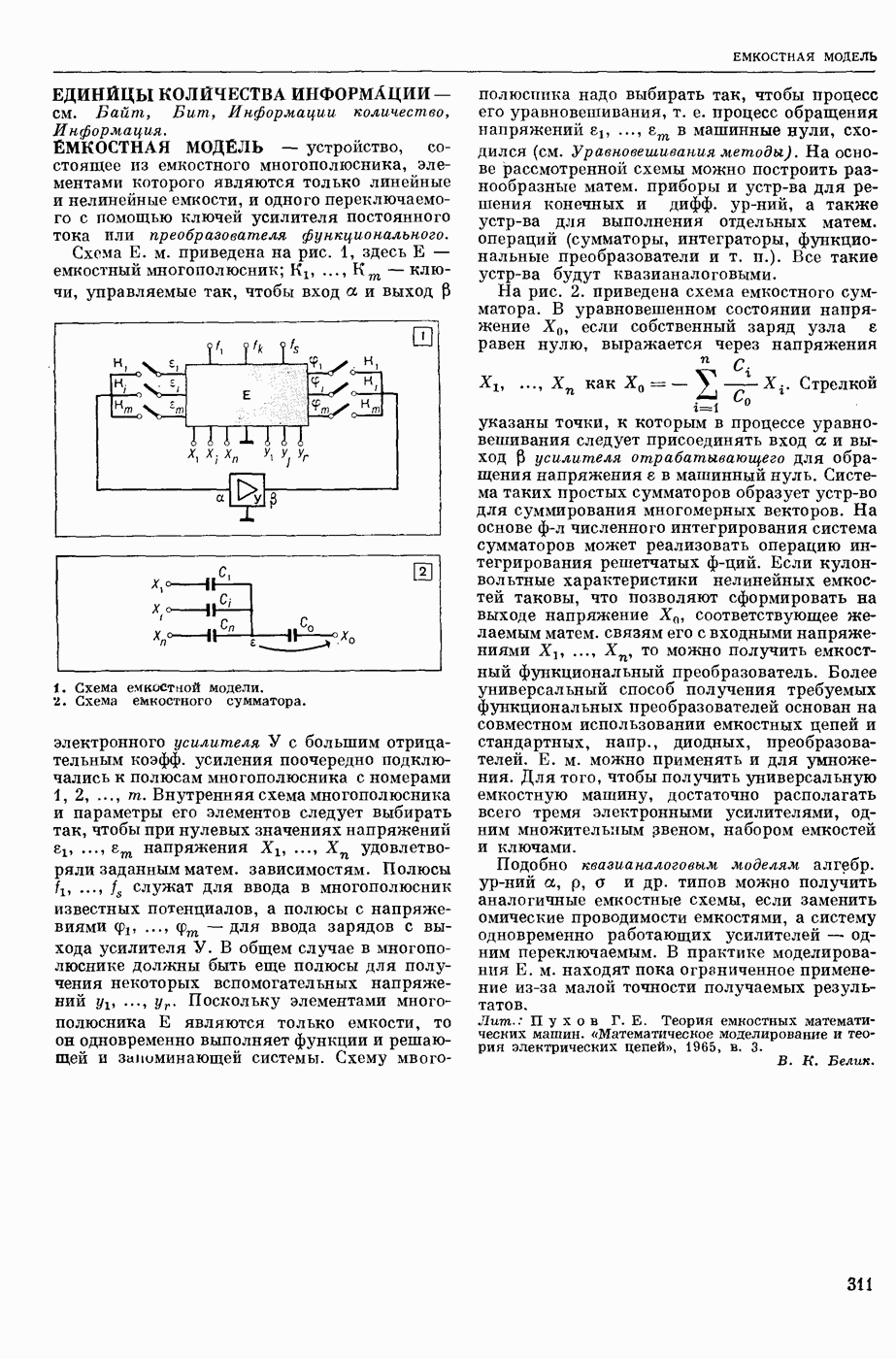
1. Изменение натриевой и калиевой проводимости во время деполяризации мембраны (по Ходжкину).
2. Схема распространения нервных импульсов по поверхностной мембране (а — возбужденный участок, б — невозбужденный участок).
Такой же эффект можно получить, если искусственно увеличивать содержание ионов натрия внутри клетки и тем самым устранять градиент потенциала, обеспечивающий их движение сквозь мембрану вследствие возбуждения. Однако некоторые клетки могут использовать для той же цели ионы кальция. Количество ионов, проникающее во время генерации одиночного импульса, слишком мало для того, чтобы его можно было определить аналитически, однако его можно определить при ритмическом раздражении и затем пересчитать на один импульс; оно составляет  . На основании экспериментально полученных данных о мембранных ионных токах можно провести моделирование распространяющегося потенциала действия на теор. кабеле, свойства которого идентичны физико-хим. свойствам клетки. Теоретически полученные кривые распространяющегося потенциала действия в этом случае очень хорошо совпадают с формой кривых потенциала действия, зарегистрированных в естественных условиях. Лит.: Ходжкин А. Нервный импульс. Пер. с англ. М., 1965 [библиогр. с. 113—123]; Катц Б. Нерв, мышца и синапс. Пер. с англ. М., 1968 [библиогр. с. 203—217]; Ходоров Б. И. Проблема возбудимости. Л., 1969 [библиогр. с. 280—301]. П. Г. Костюк.
. На основании экспериментально полученных данных о мембранных ионных токах можно провести моделирование распространяющегося потенциала действия на теор. кабеле, свойства которого идентичны физико-хим. свойствам клетки. Теоретически полученные кривые распространяющегося потенциала действия в этом случае очень хорошо совпадают с формой кривых потенциала действия, зарегистрированных в естественных условиях. Лит.: Ходжкин А. Нервный импульс. Пер. с англ. М., 1965 [библиогр. с. 113—123]; Катц Б. Нерв, мышца и синапс. Пер. с англ. М., 1968 [библиогр. с. 203—217]; Ходоров Б. И. Проблема возбудимости. Л., 1969 [библиогр. с. 280—301]. П. Г. Костюк.