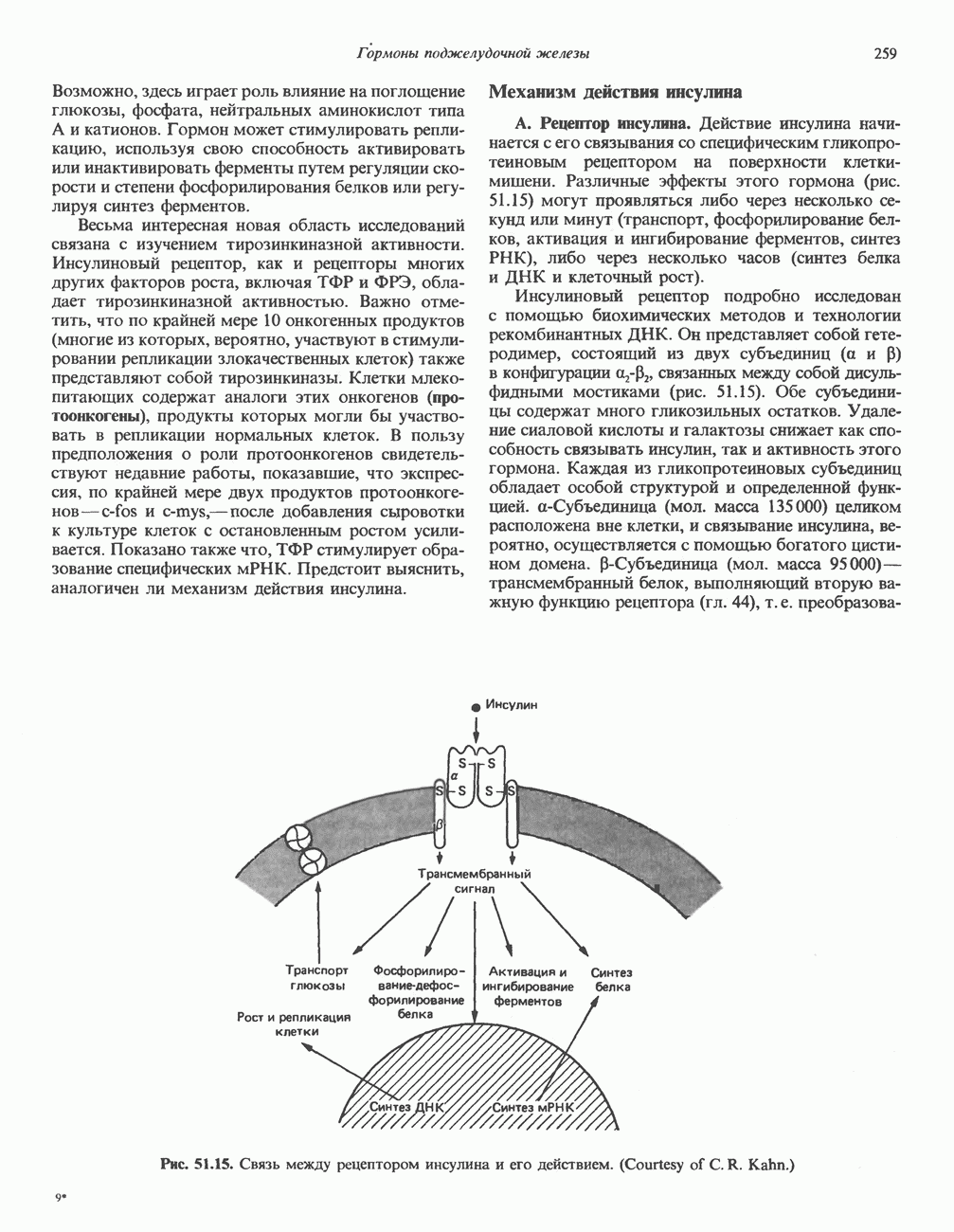
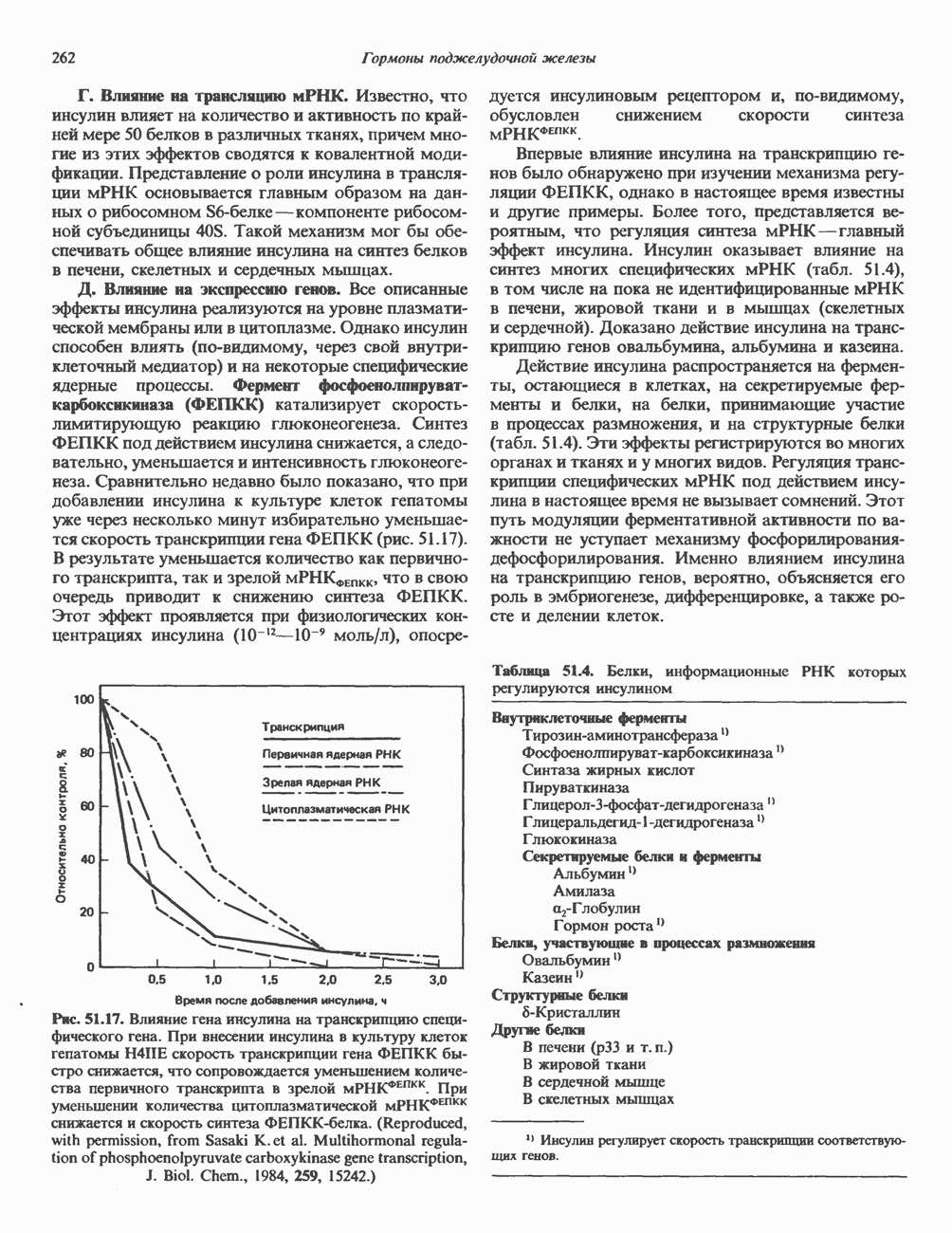
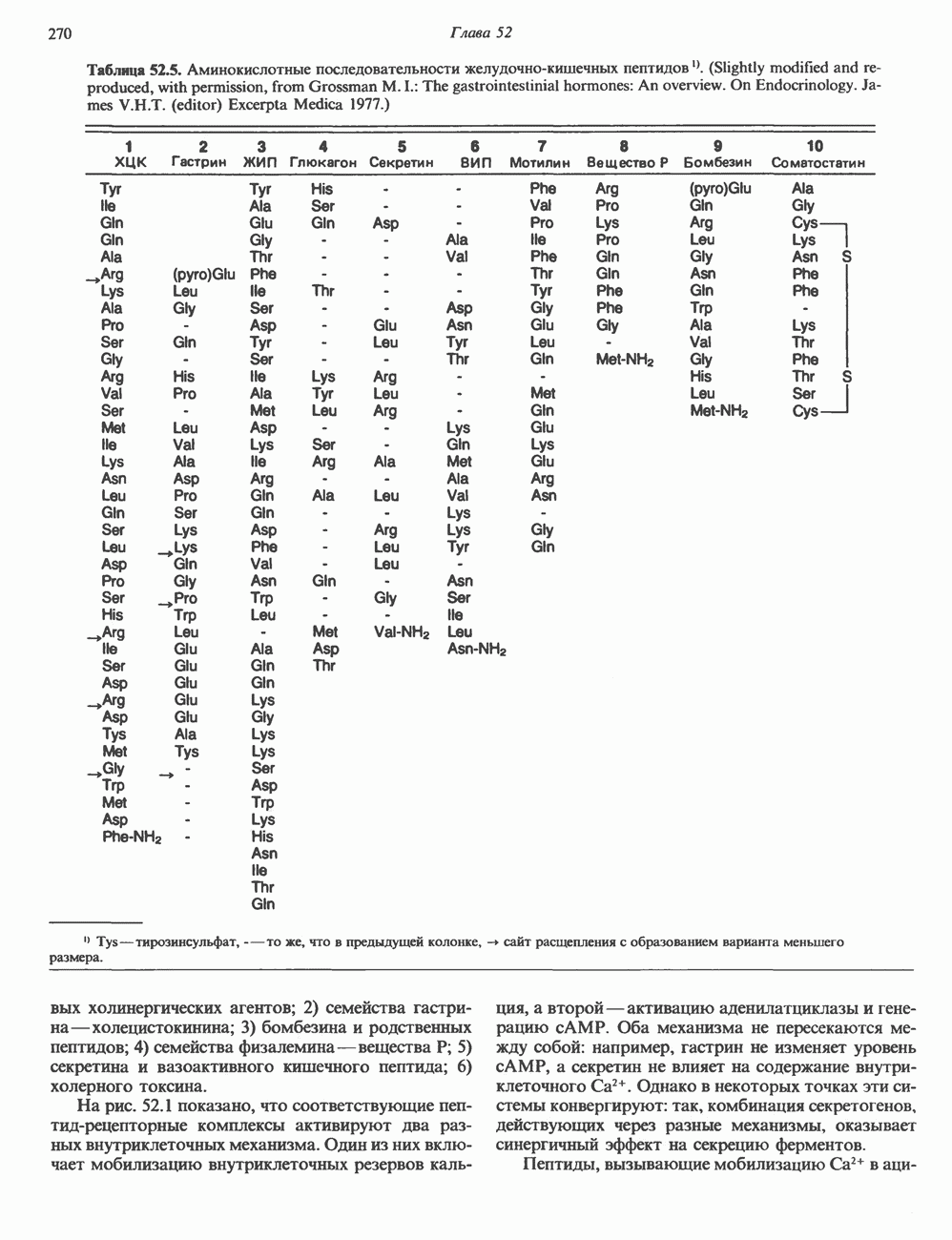
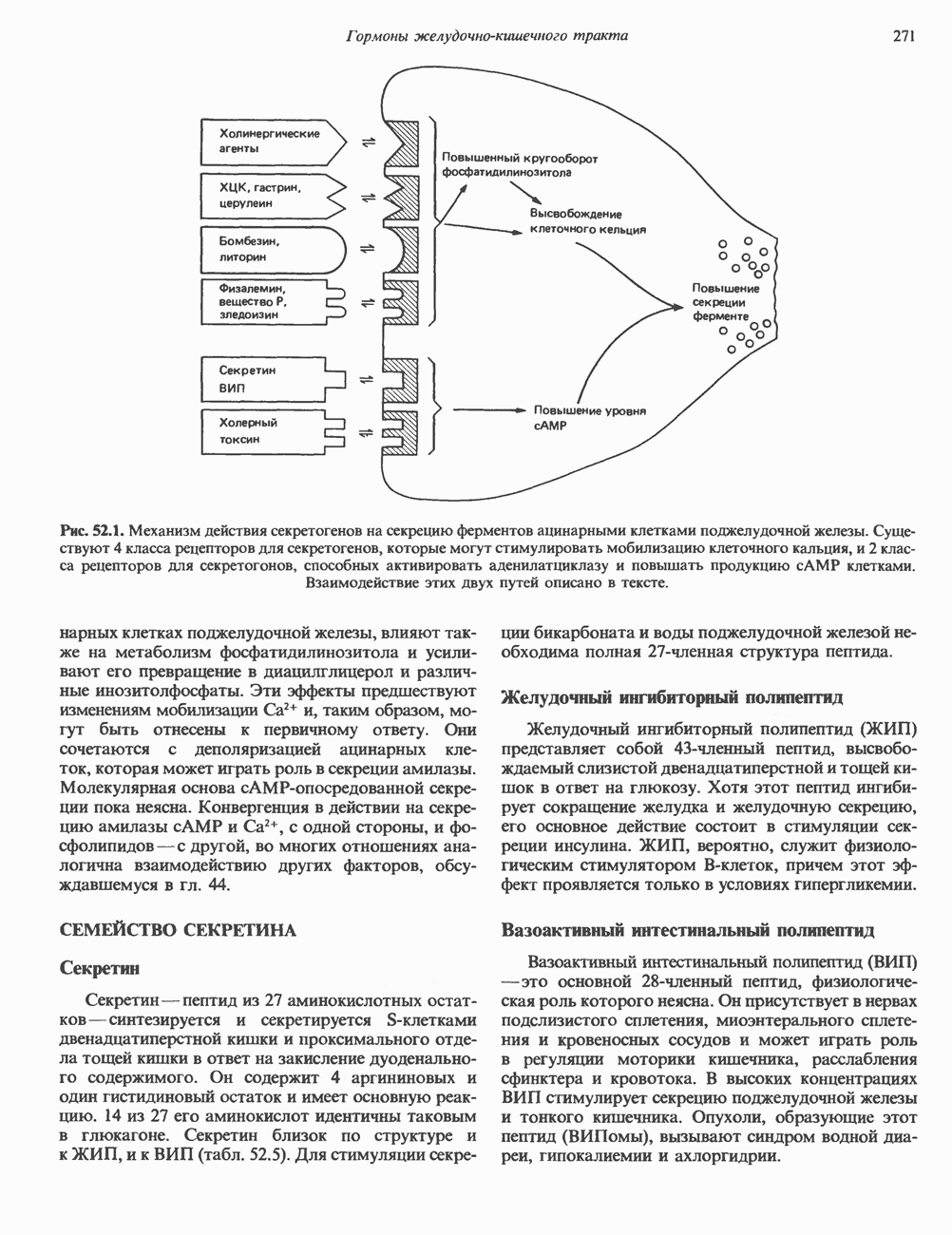
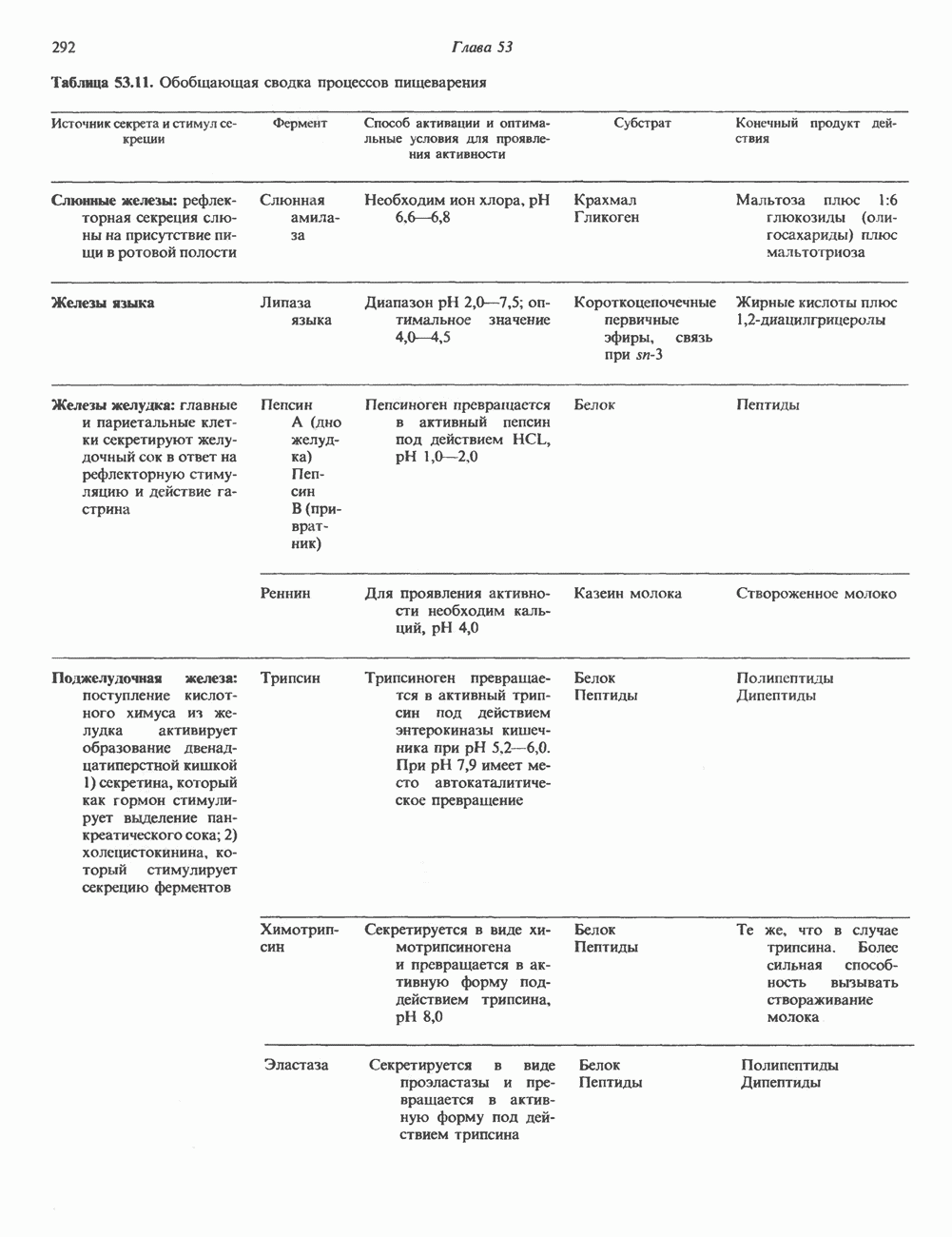
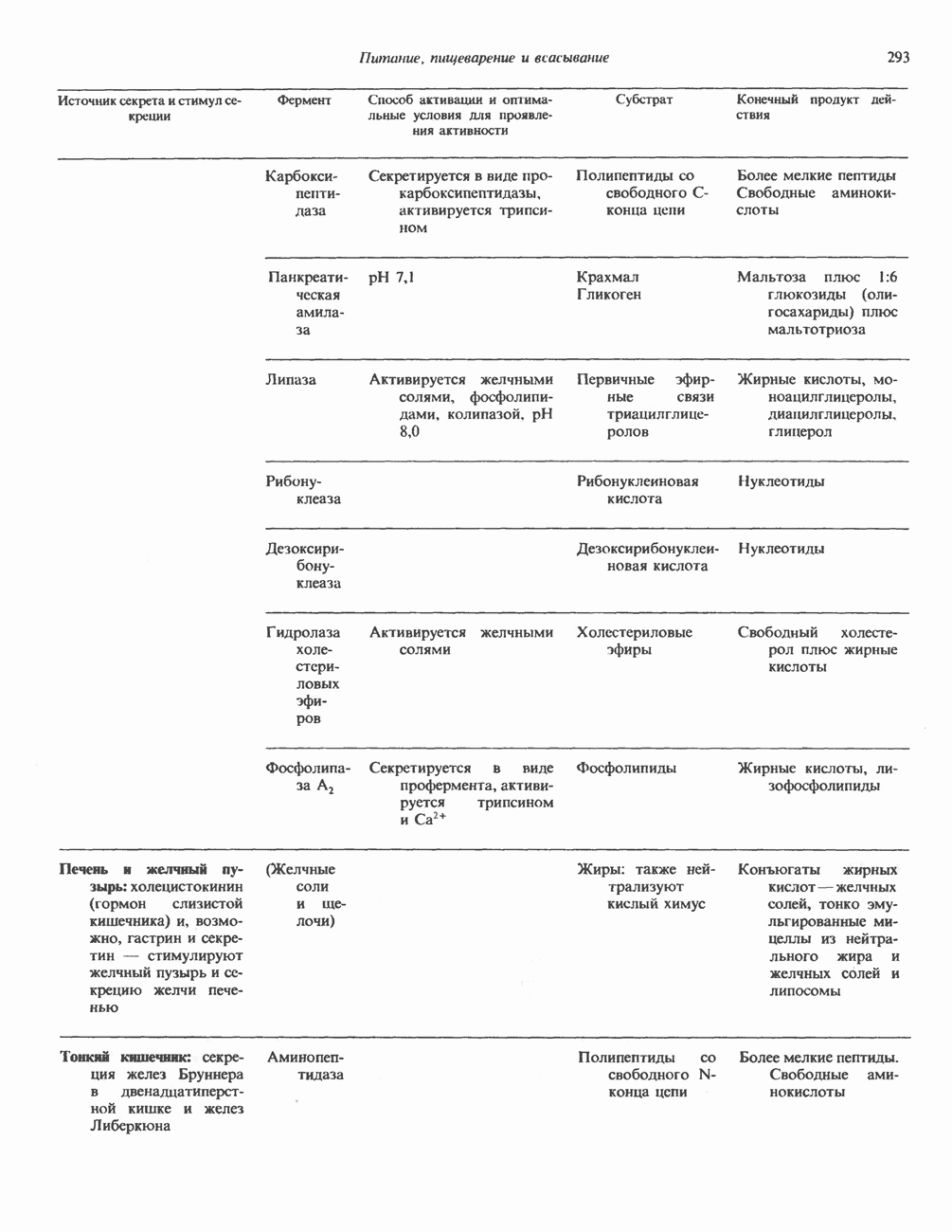
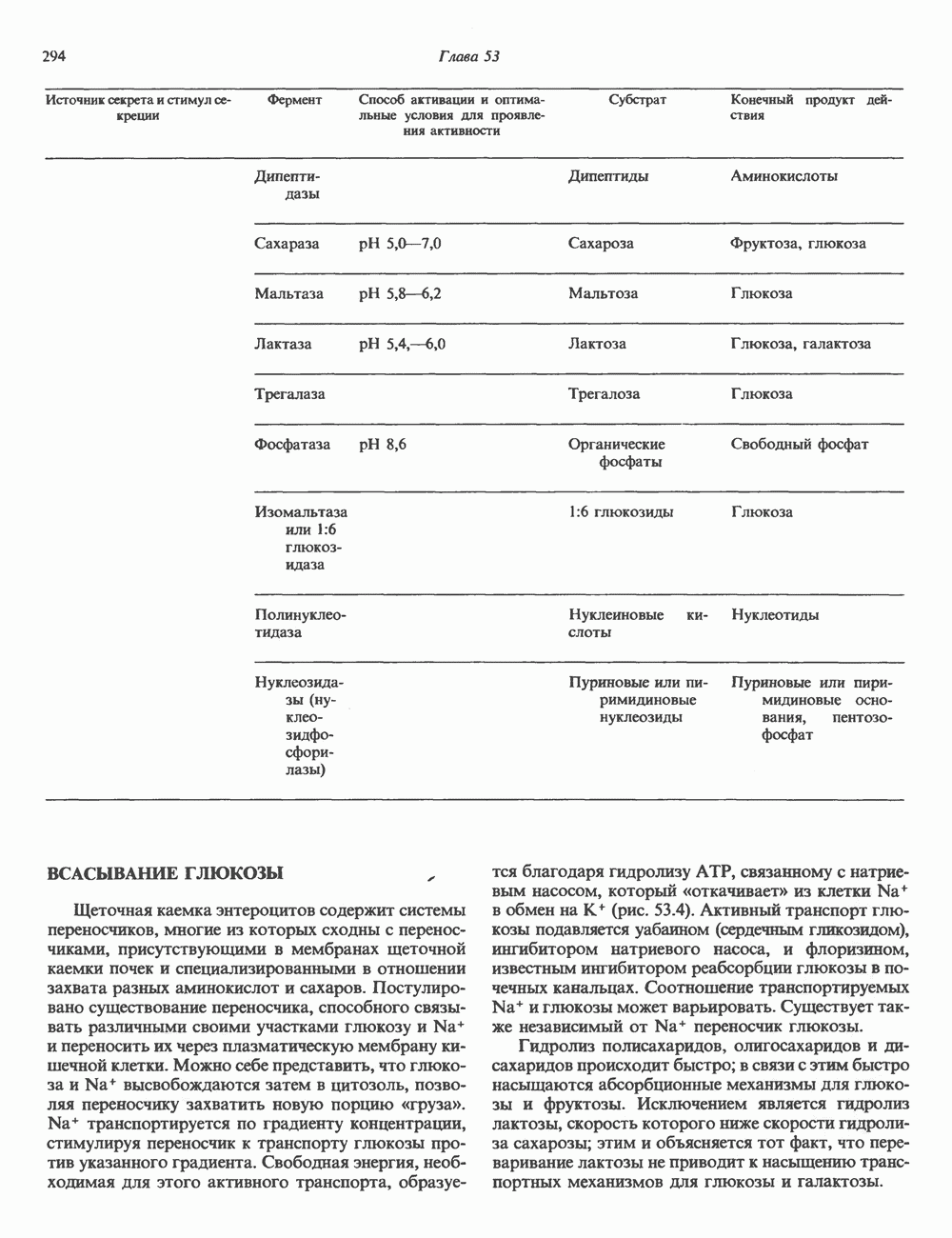
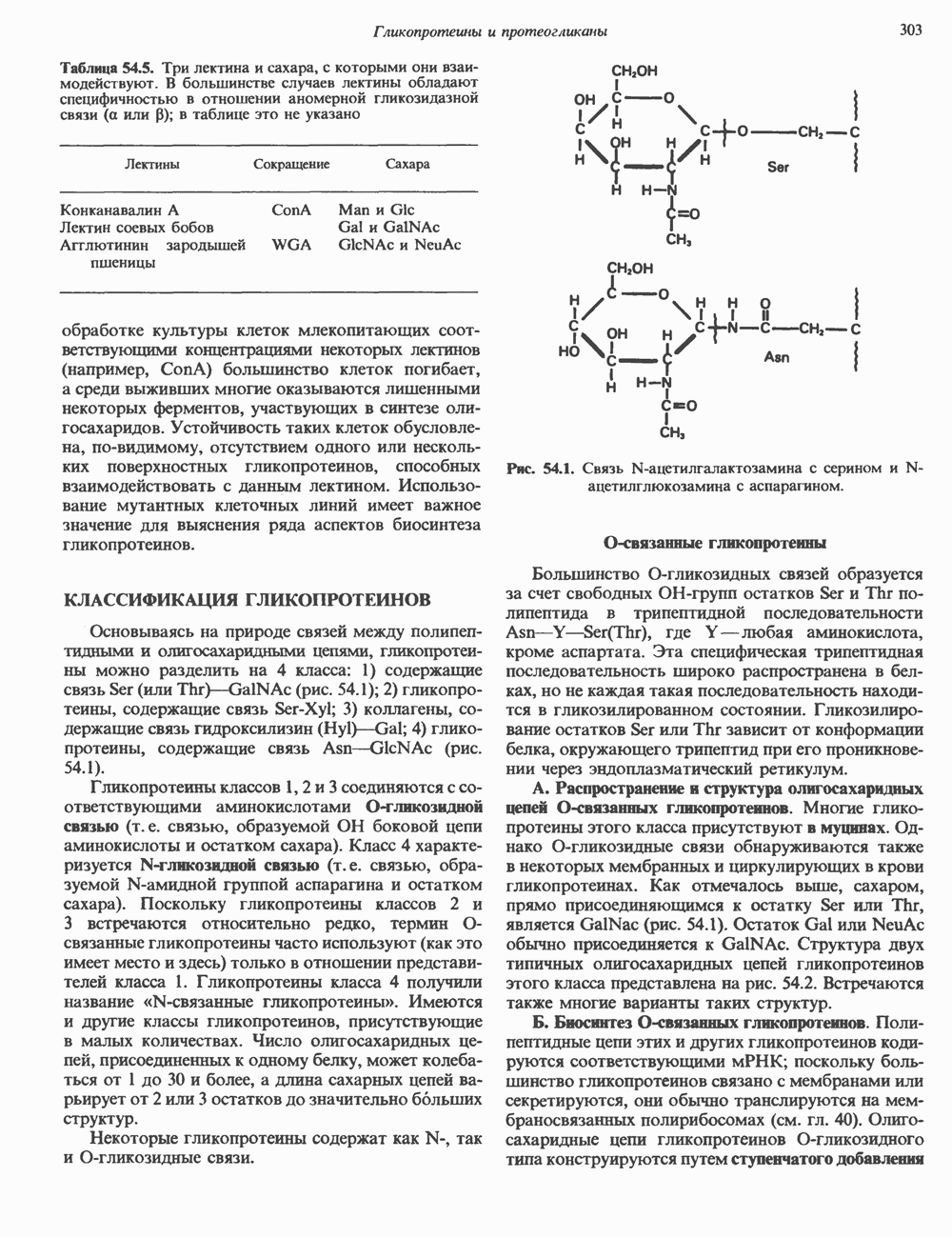
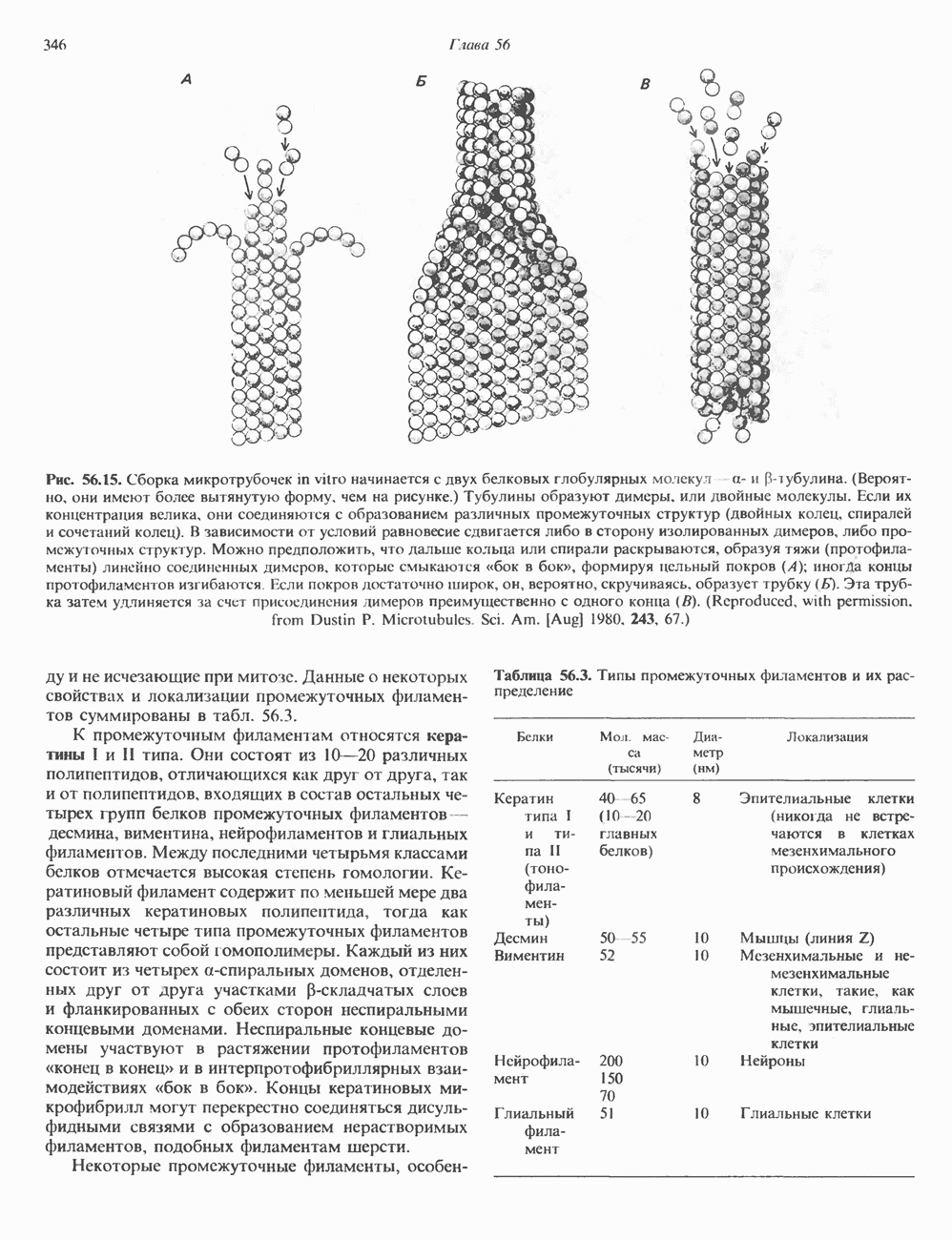
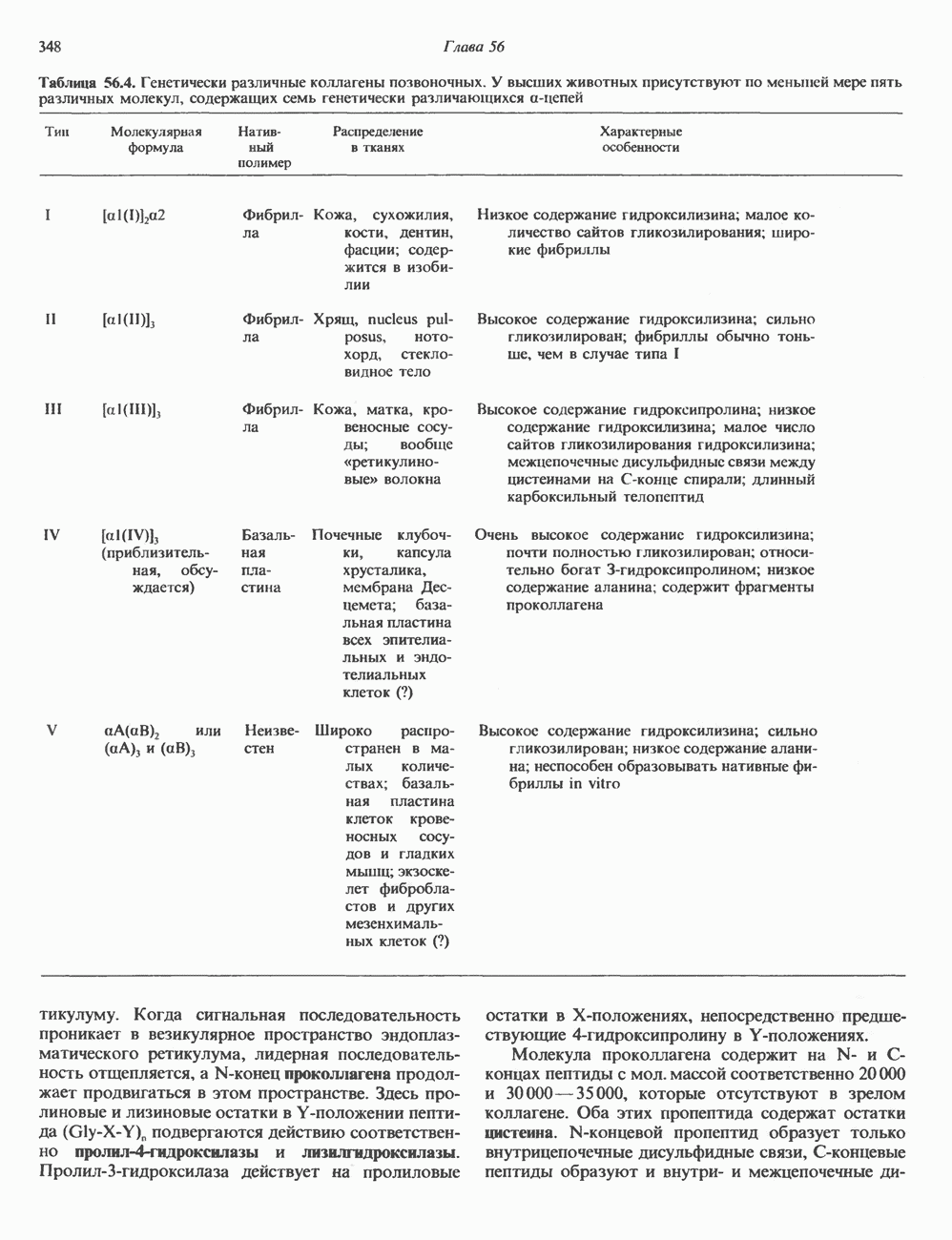
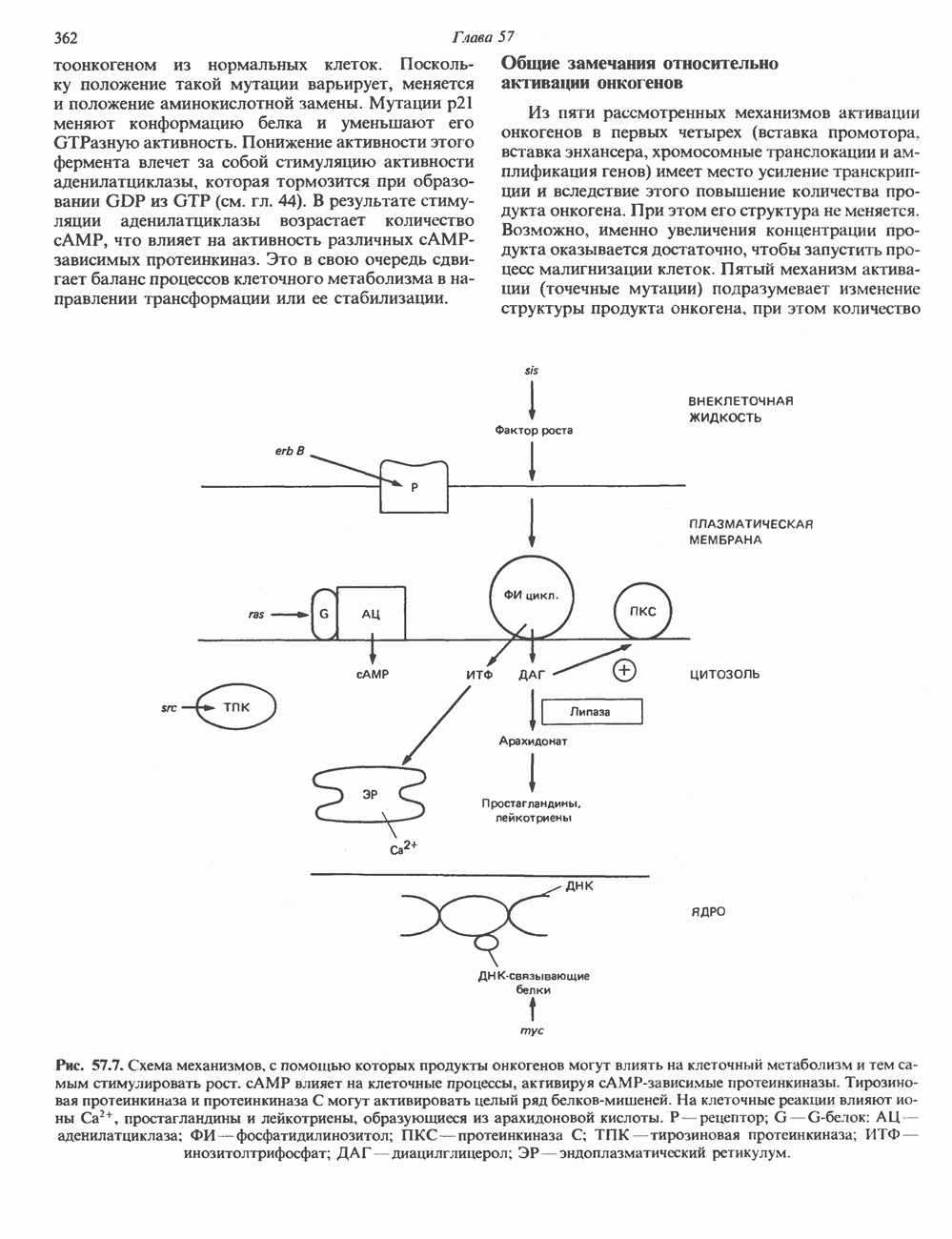
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
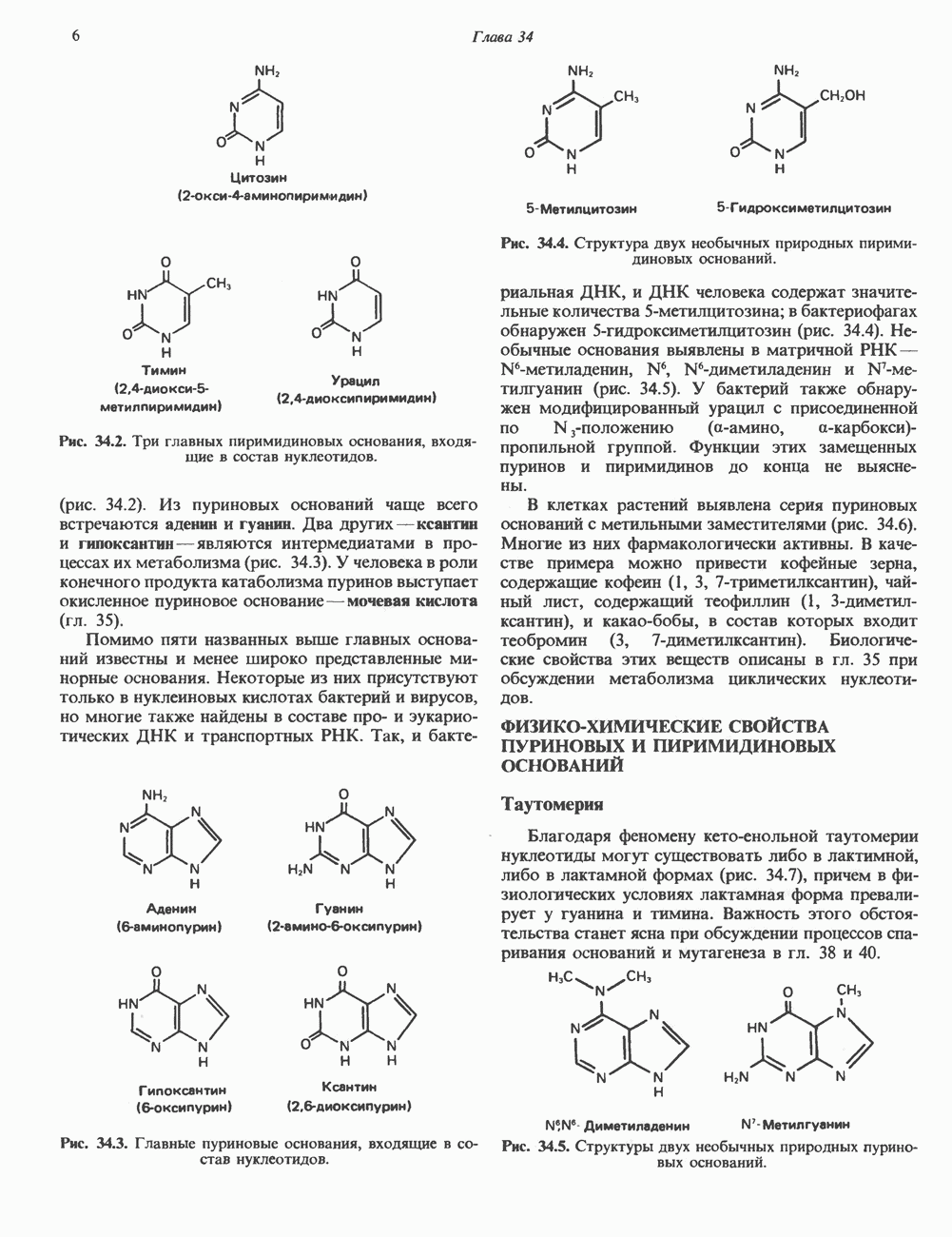
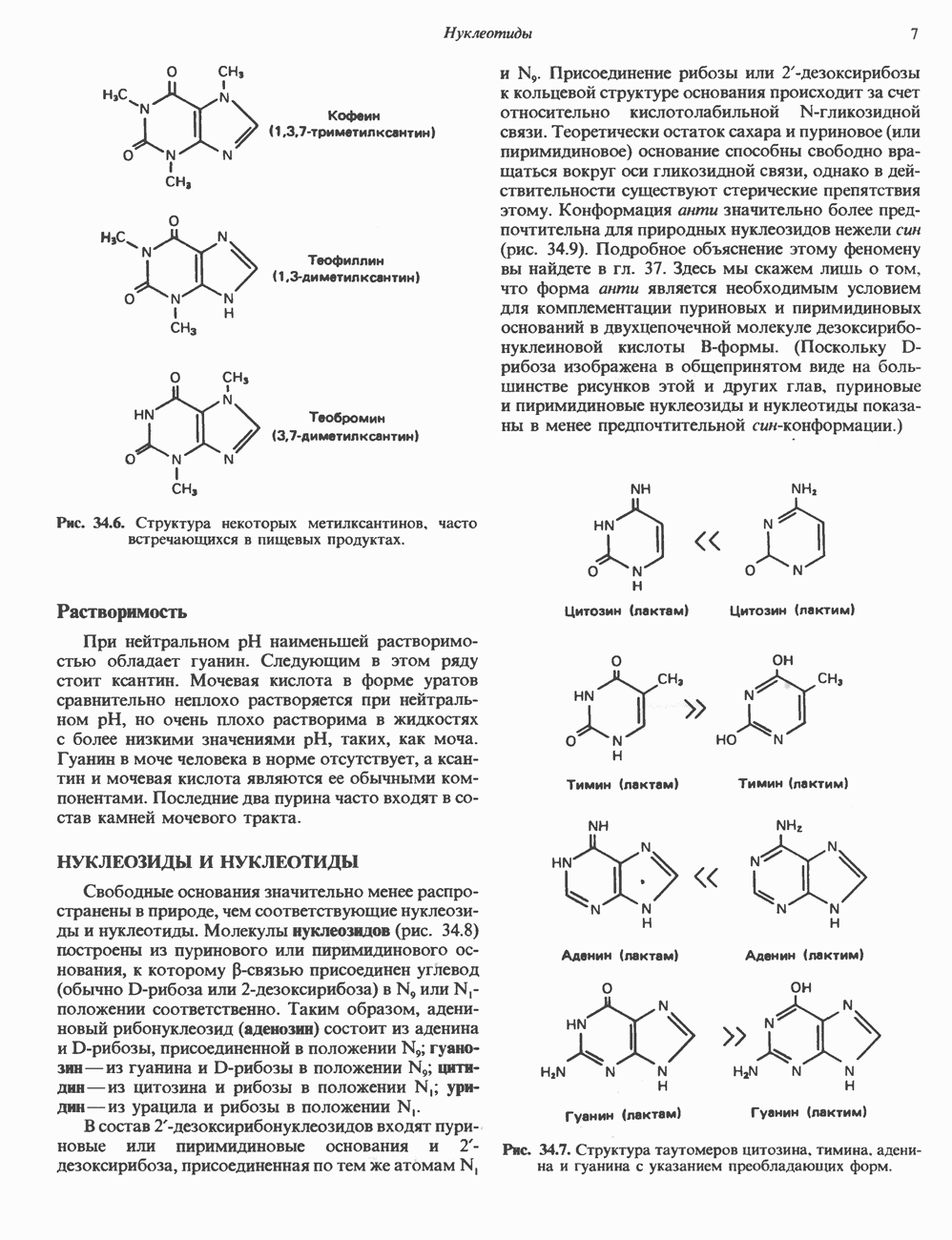
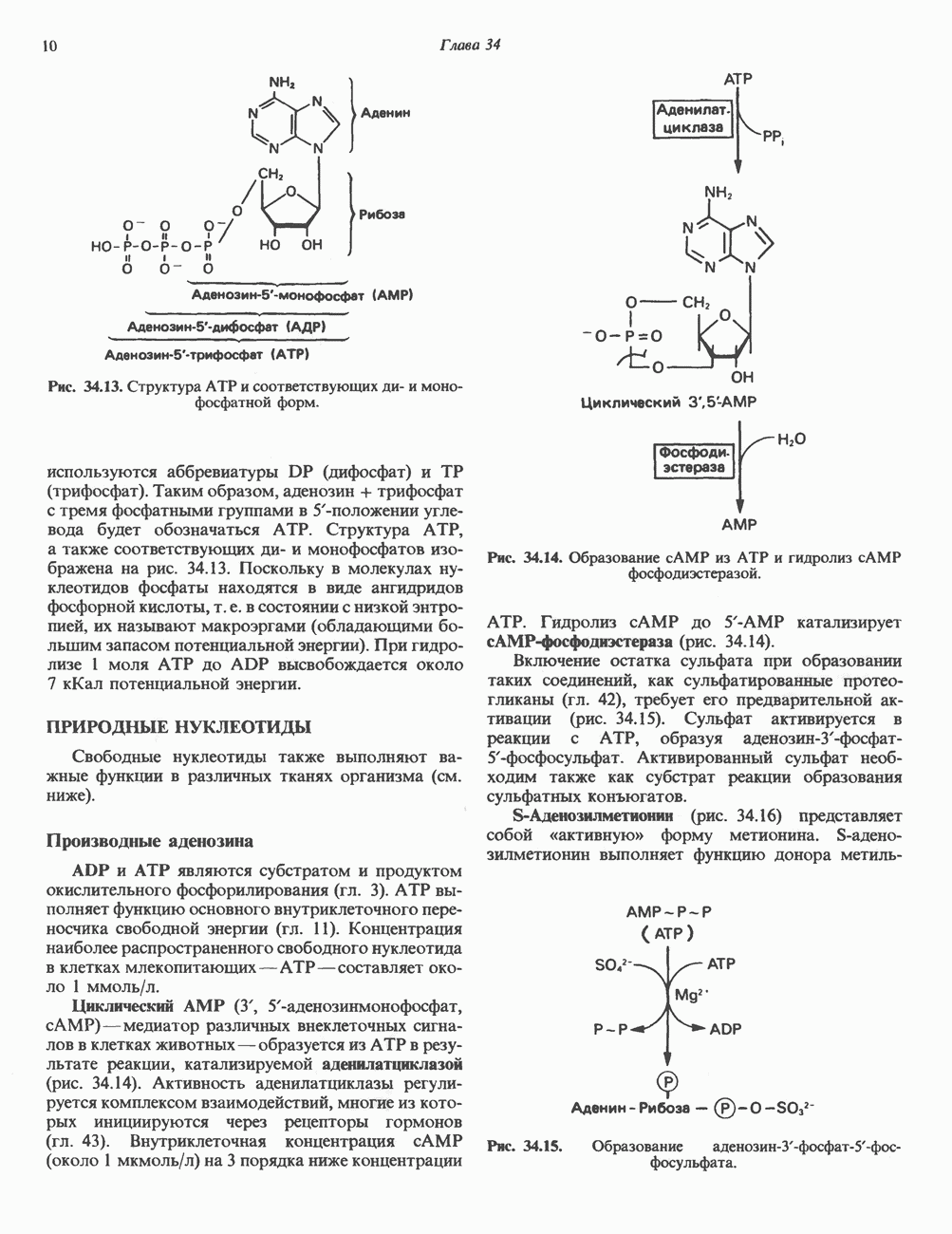
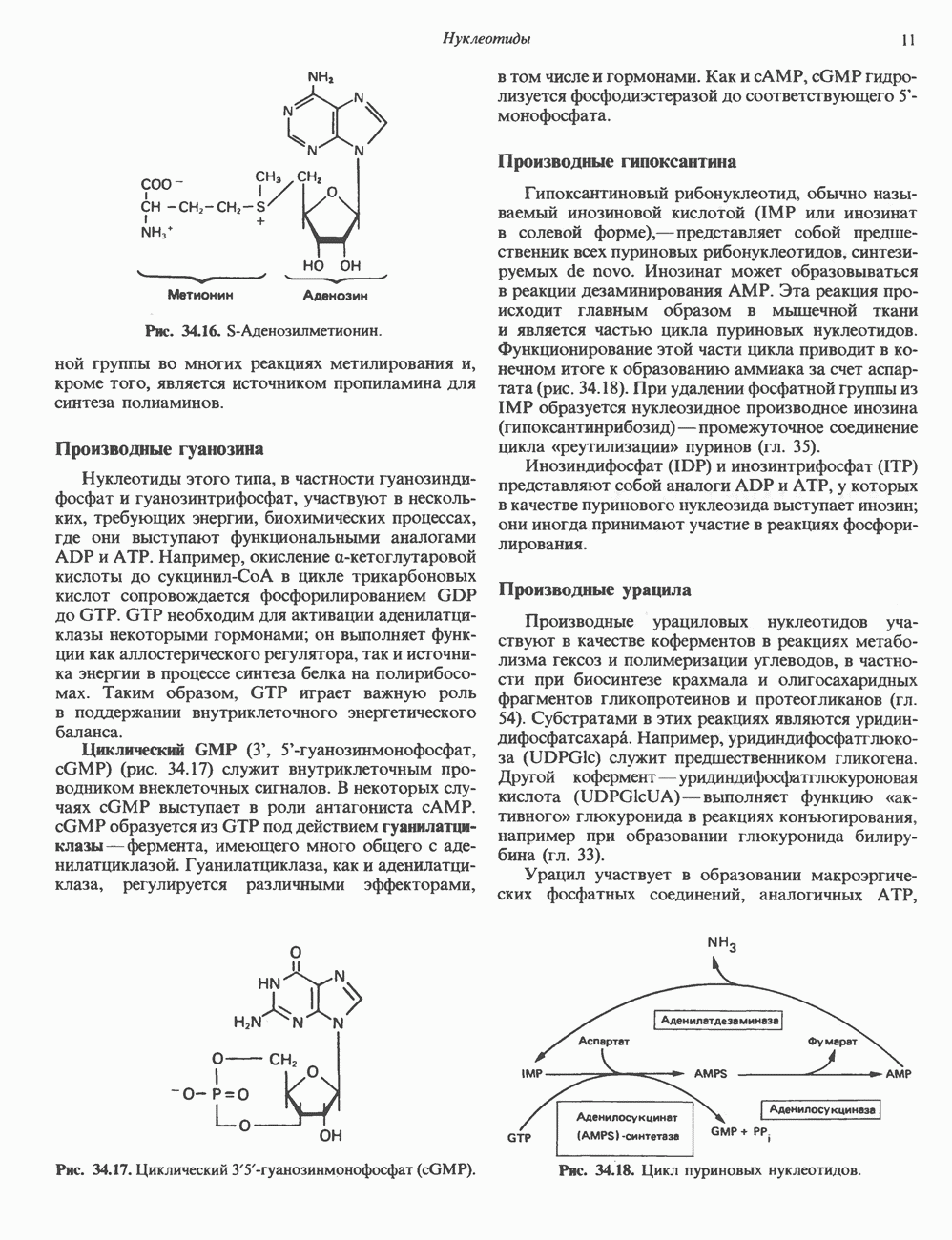
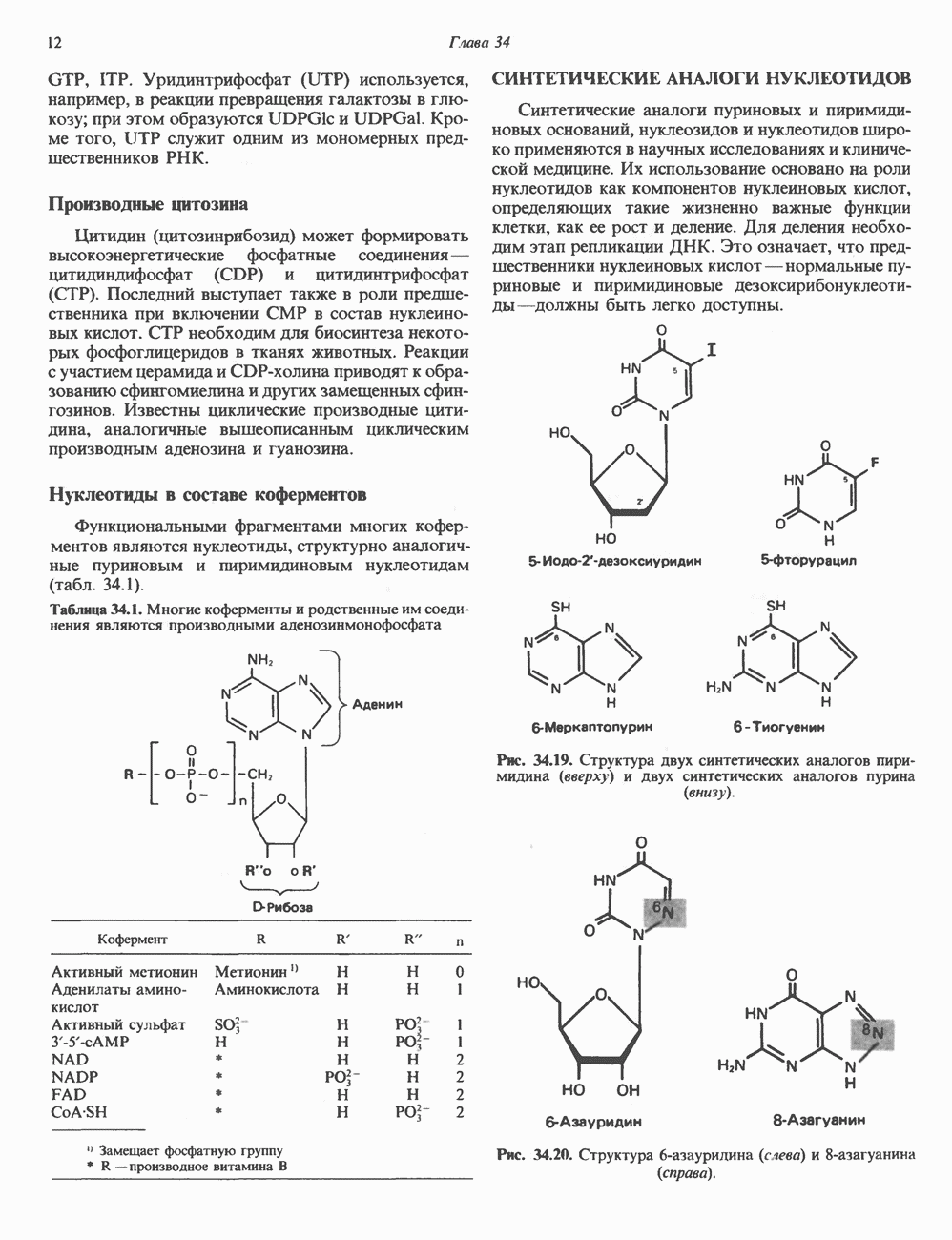
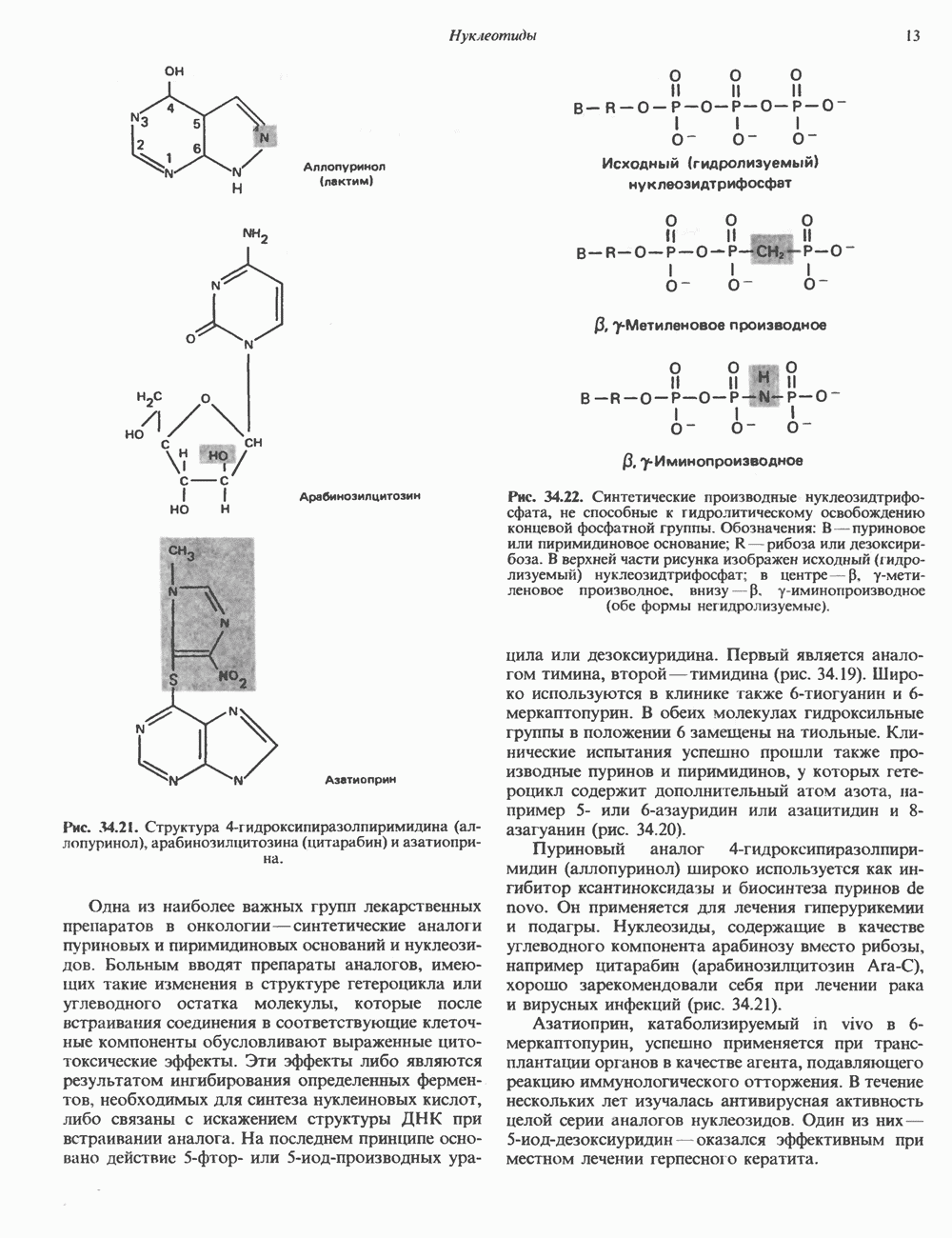
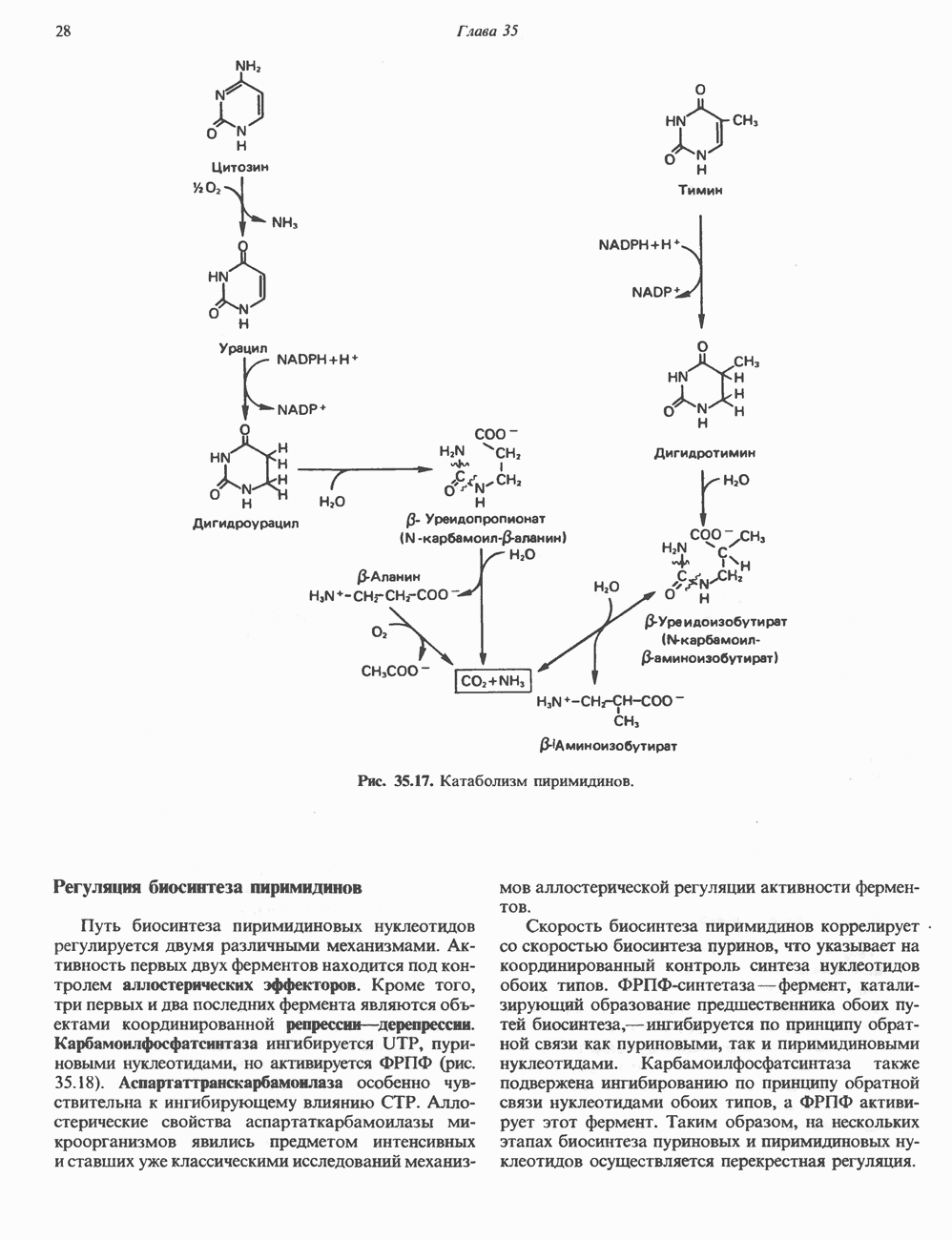
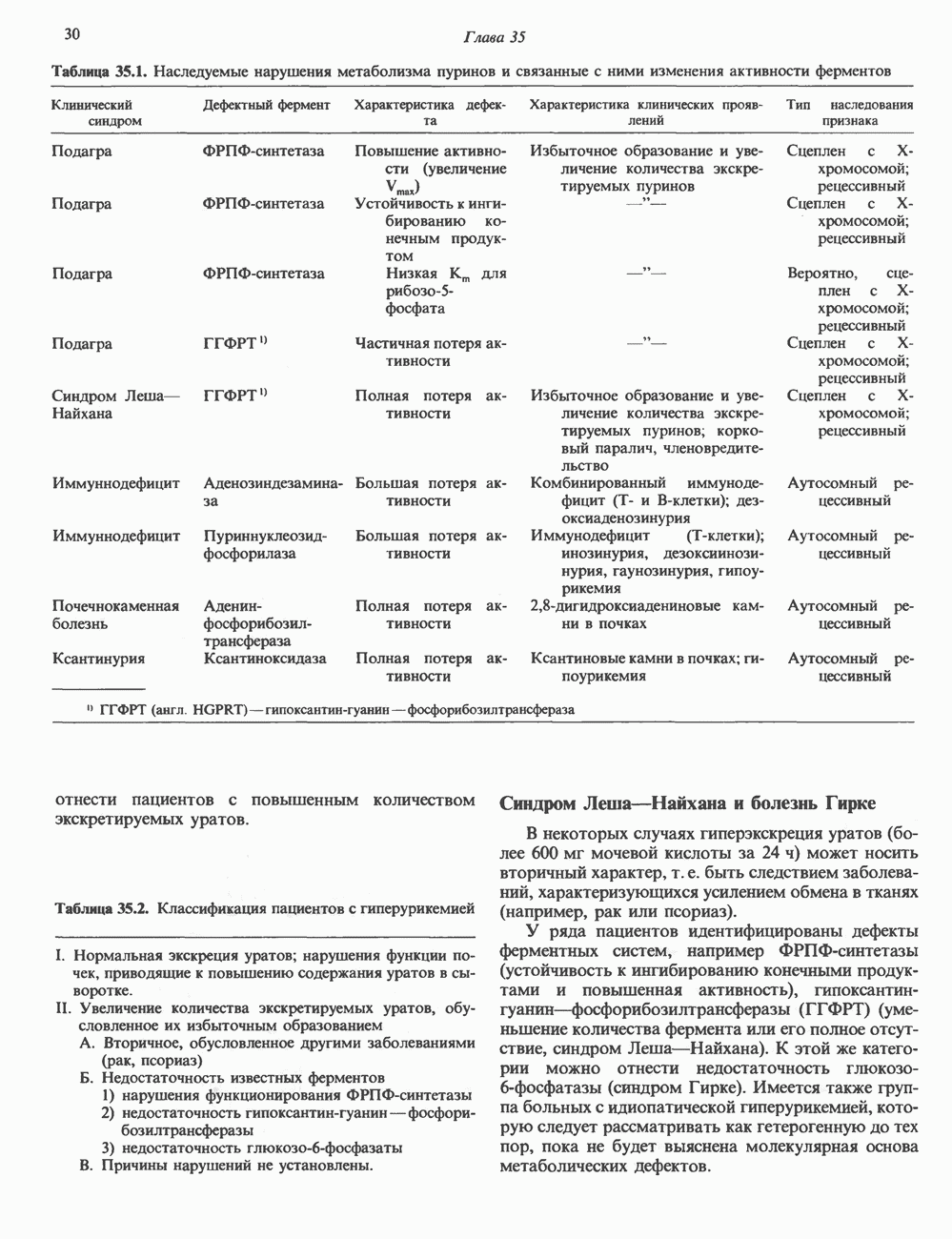
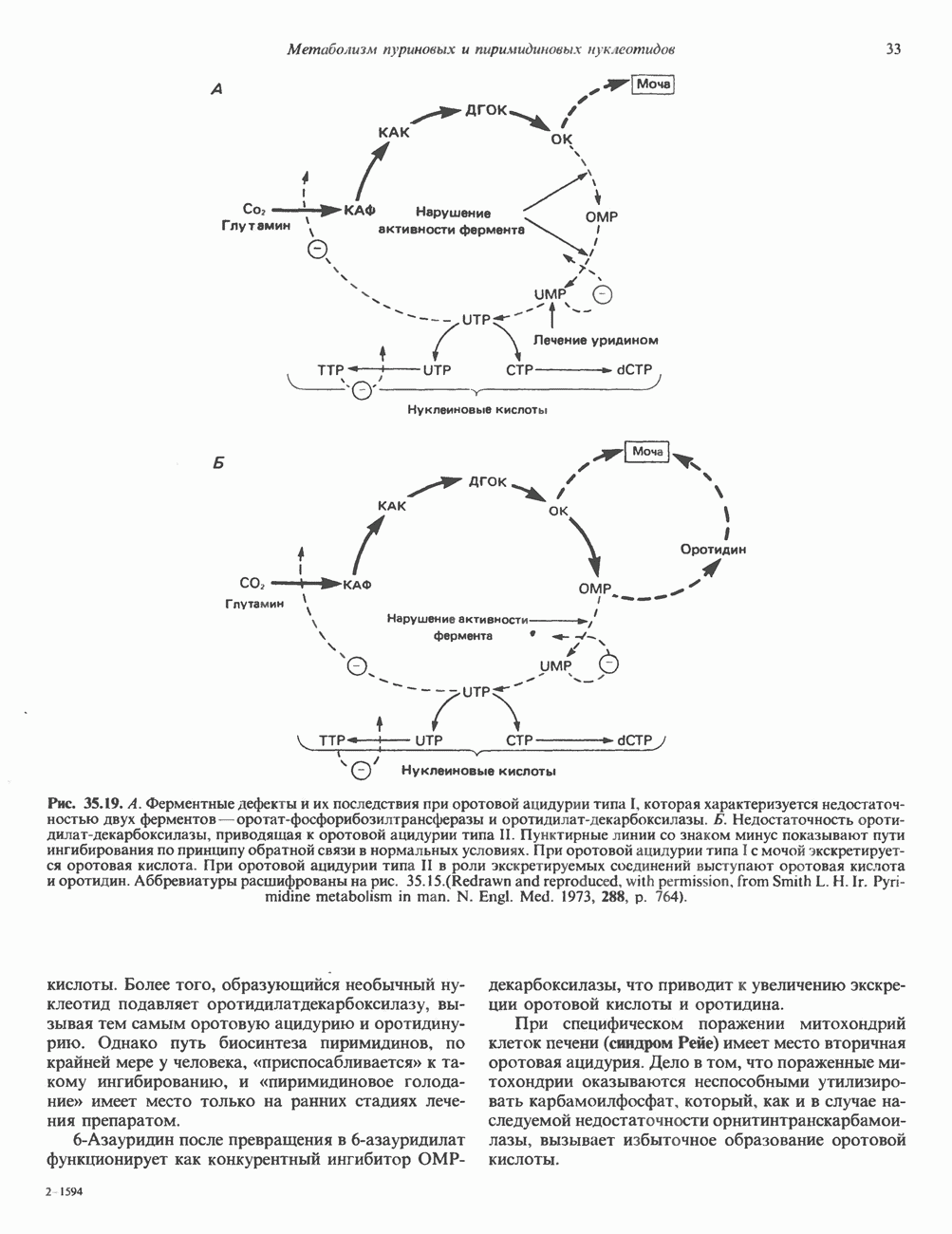
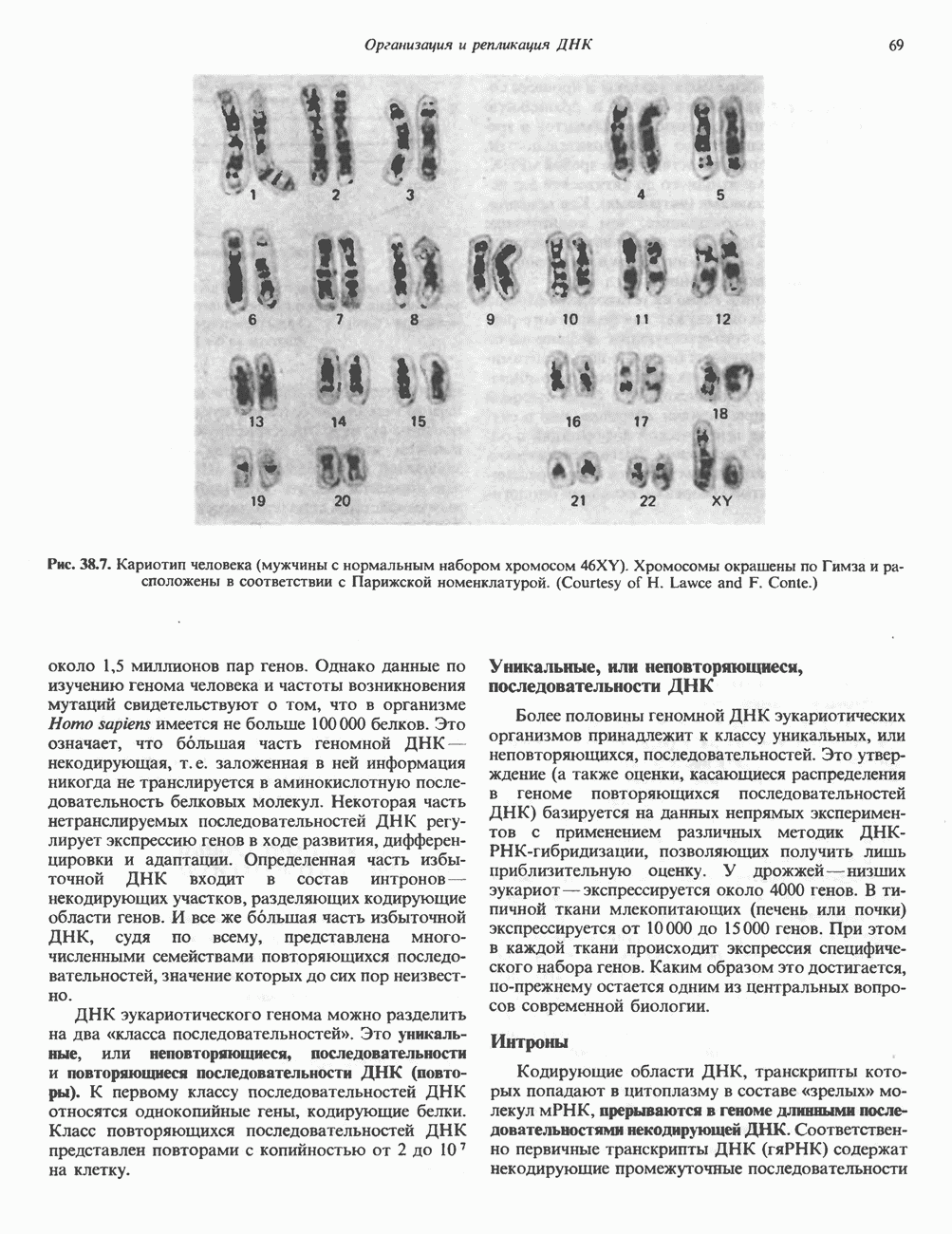
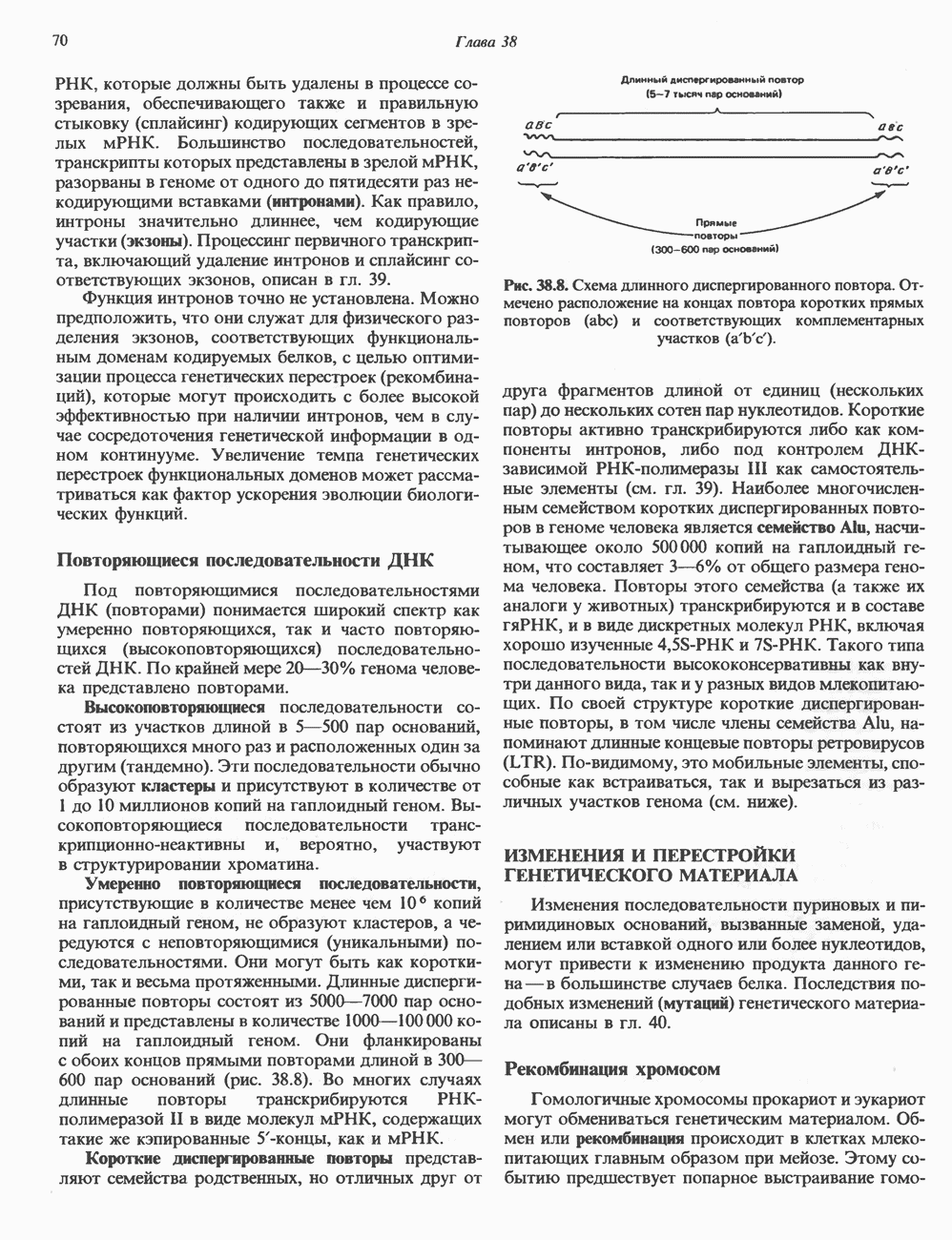
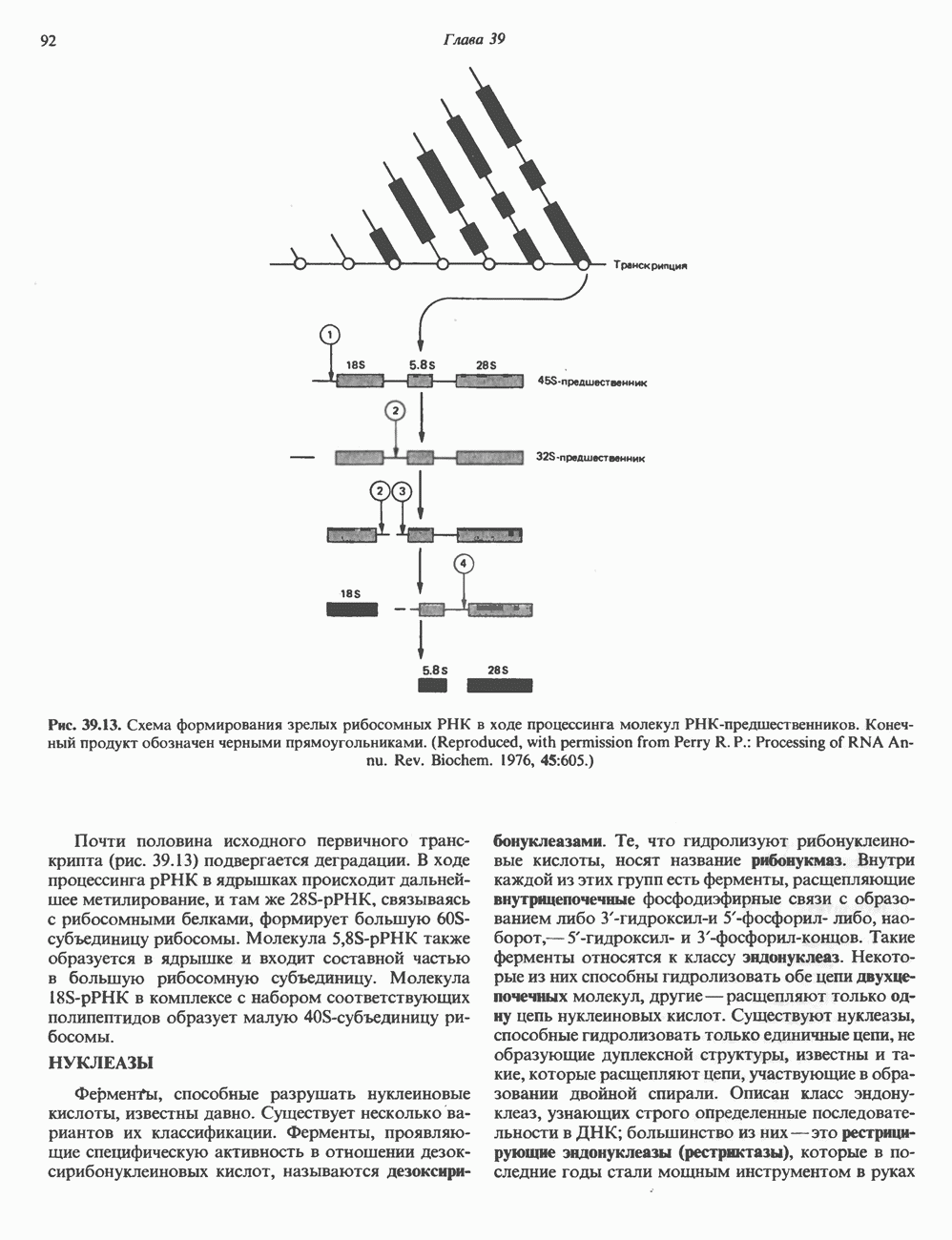
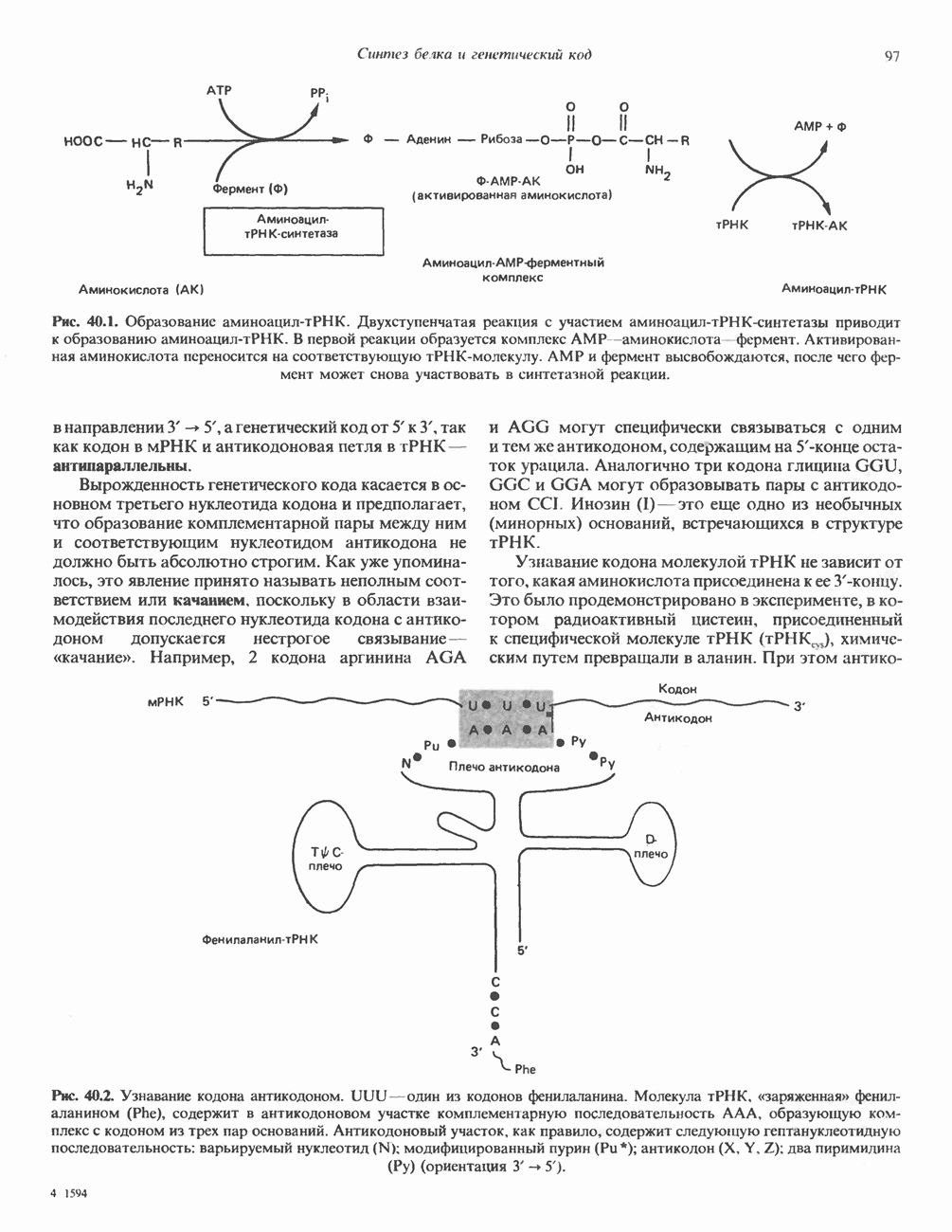
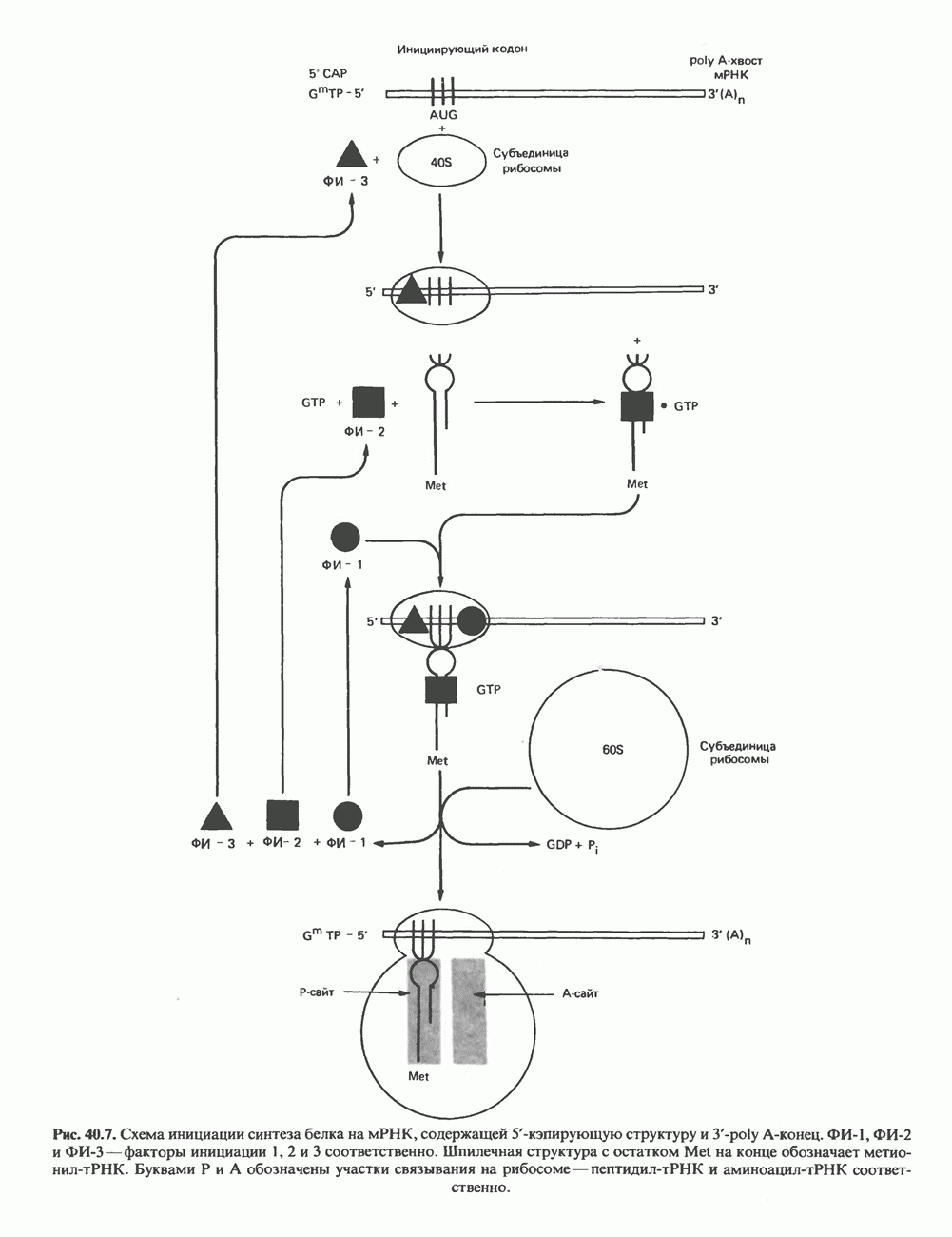
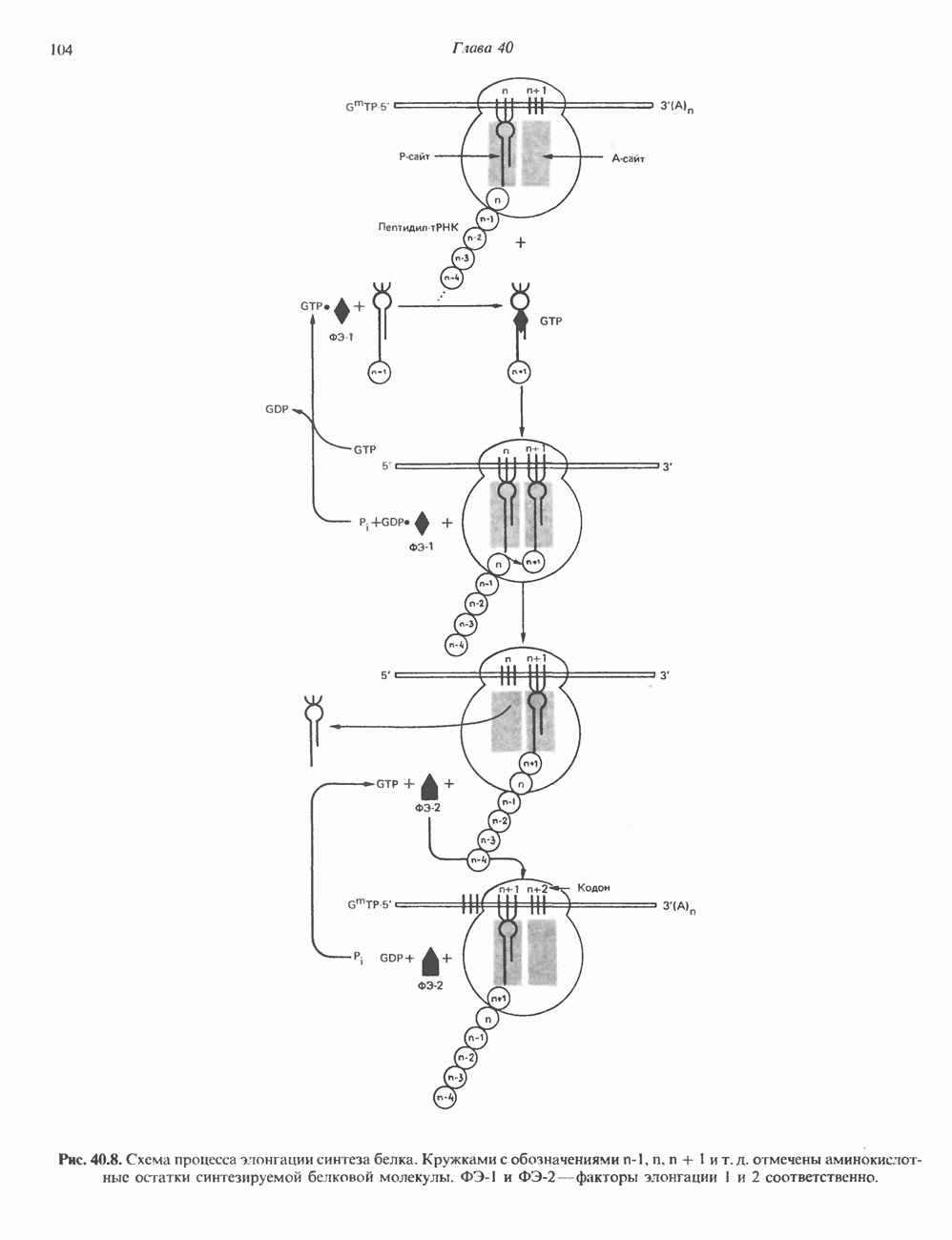
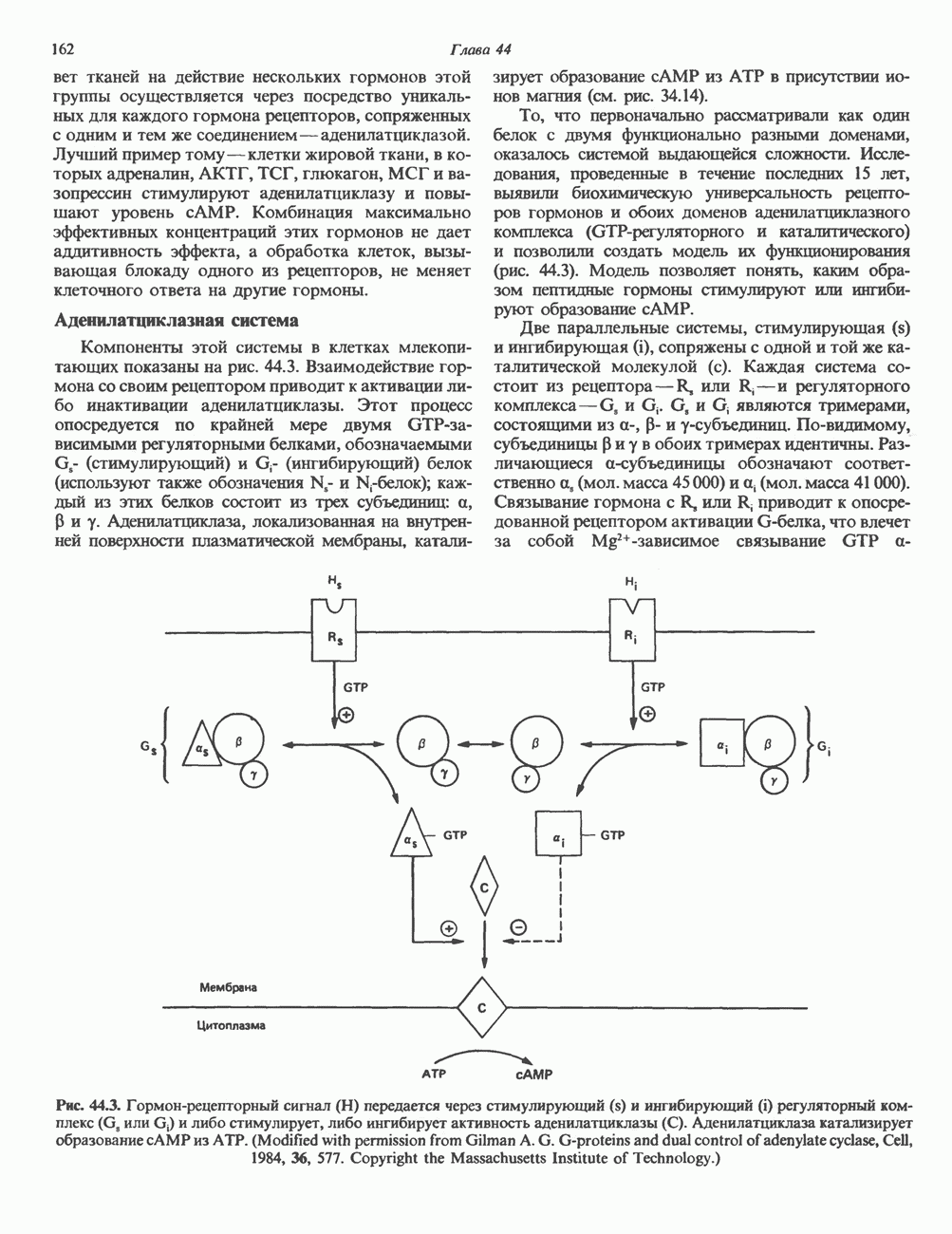
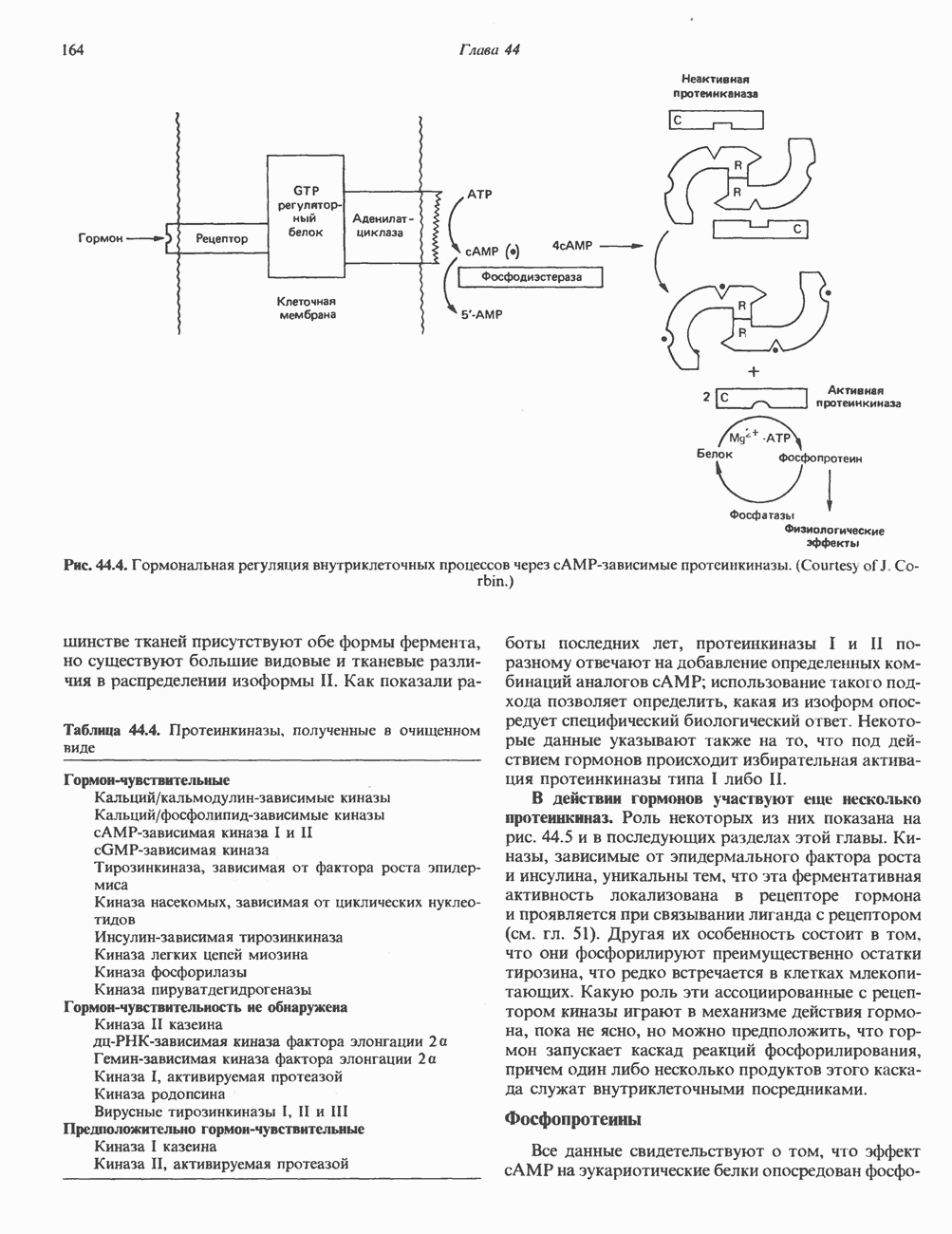
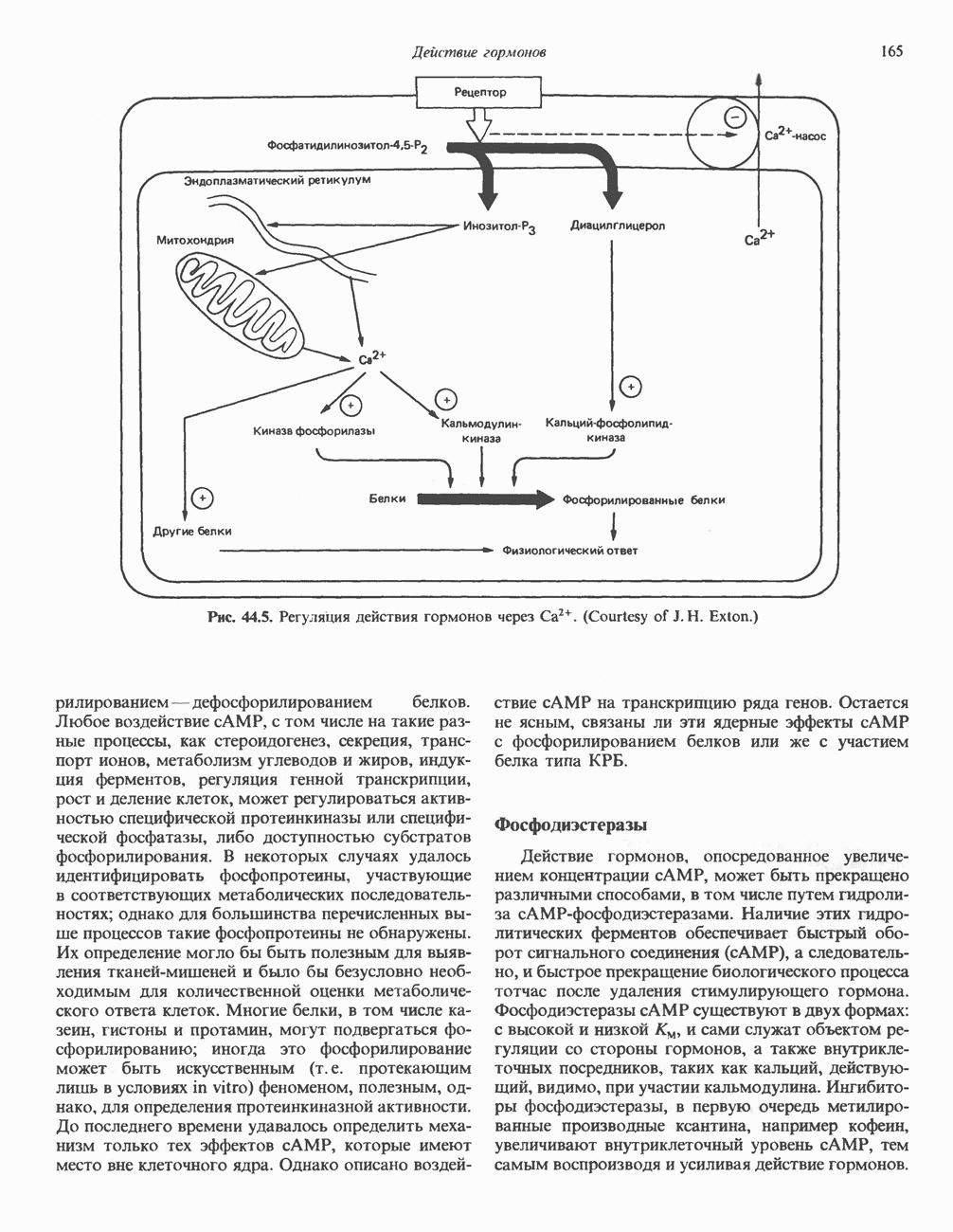
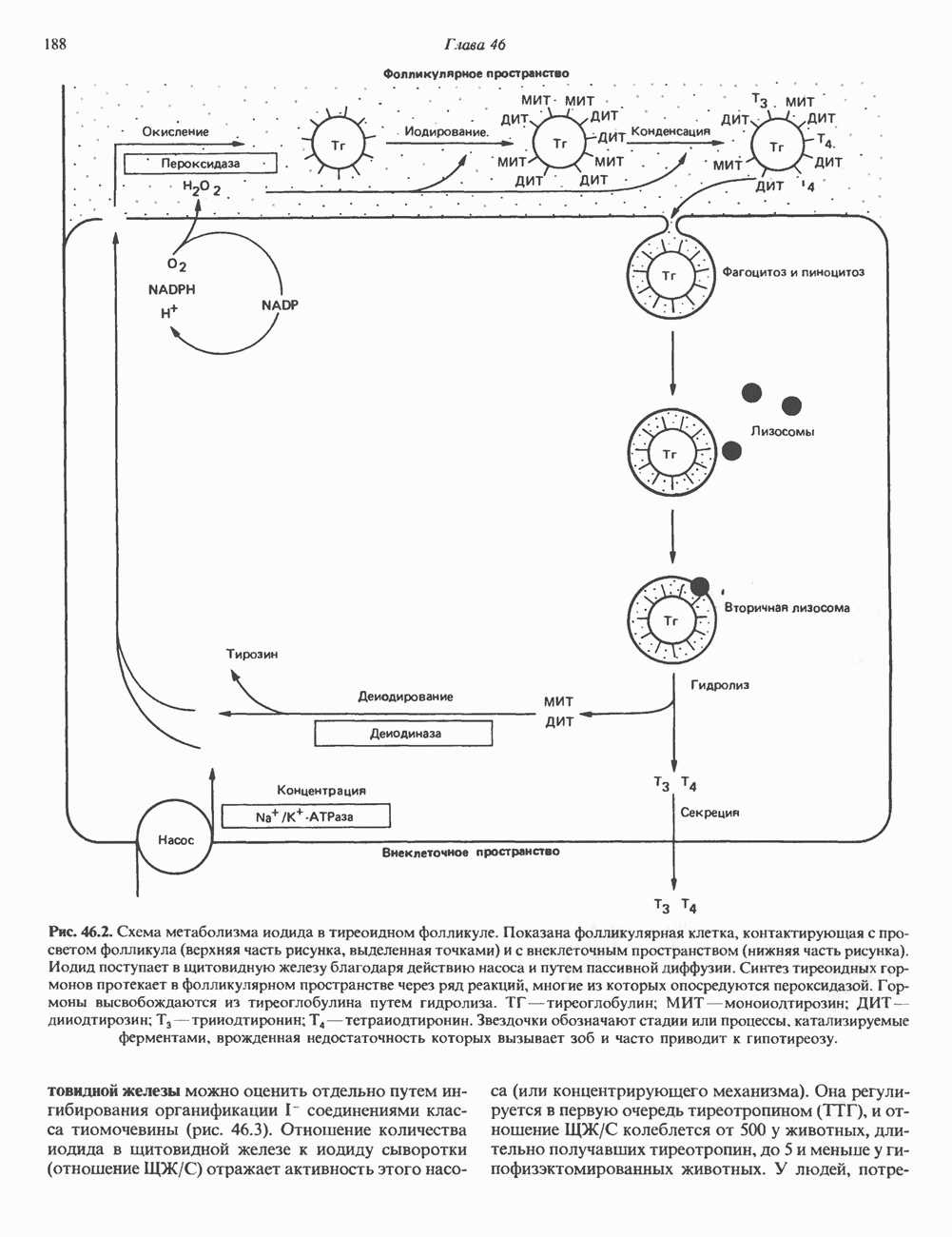
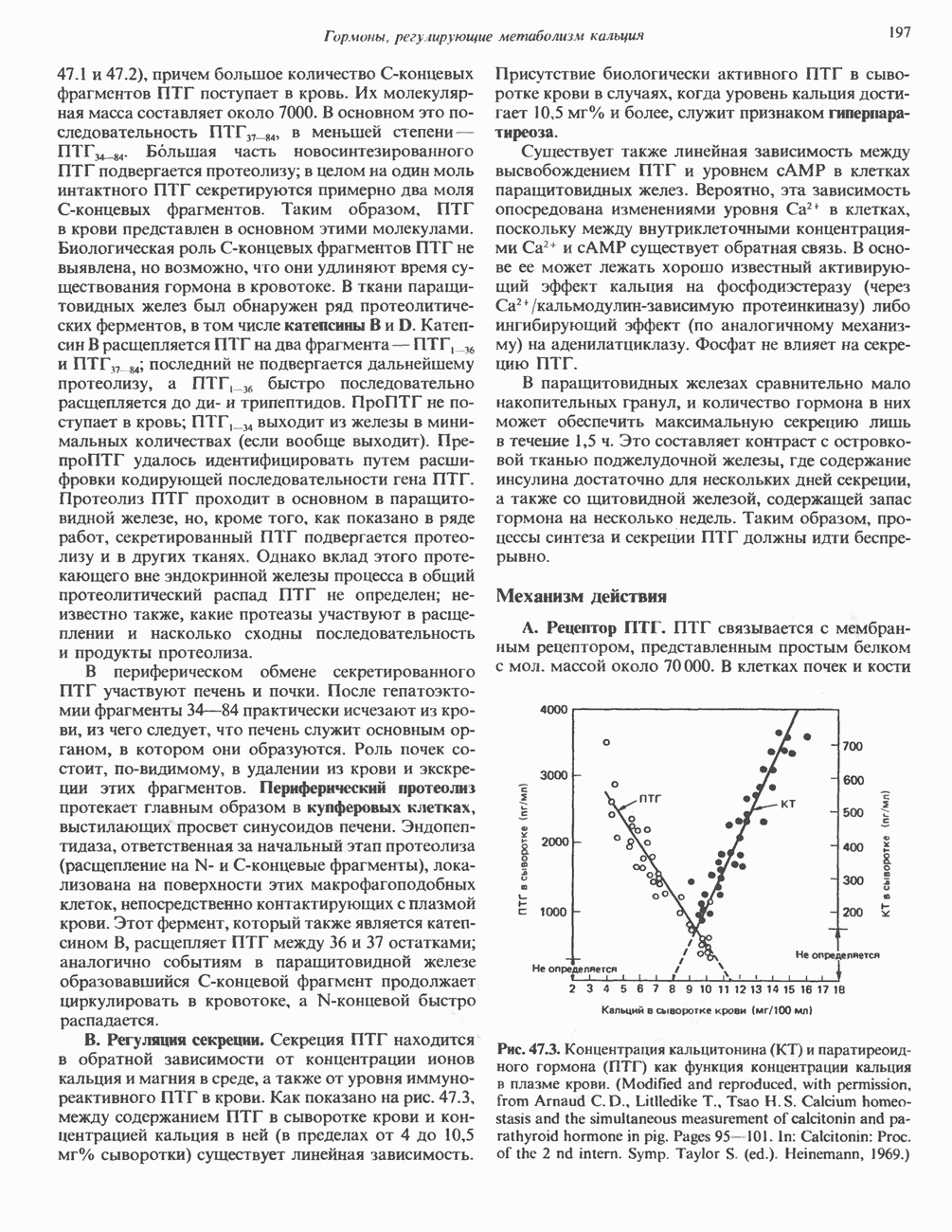
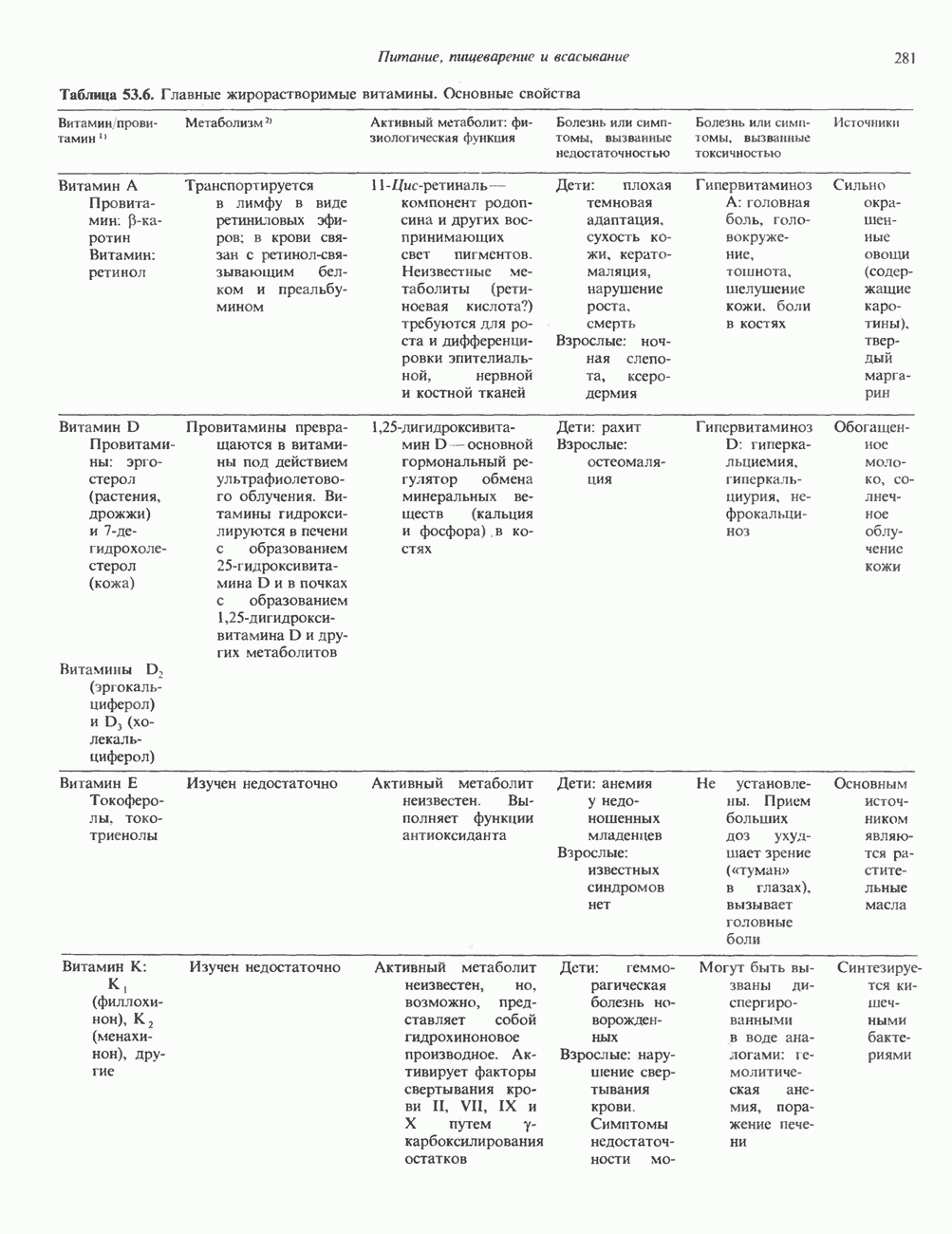
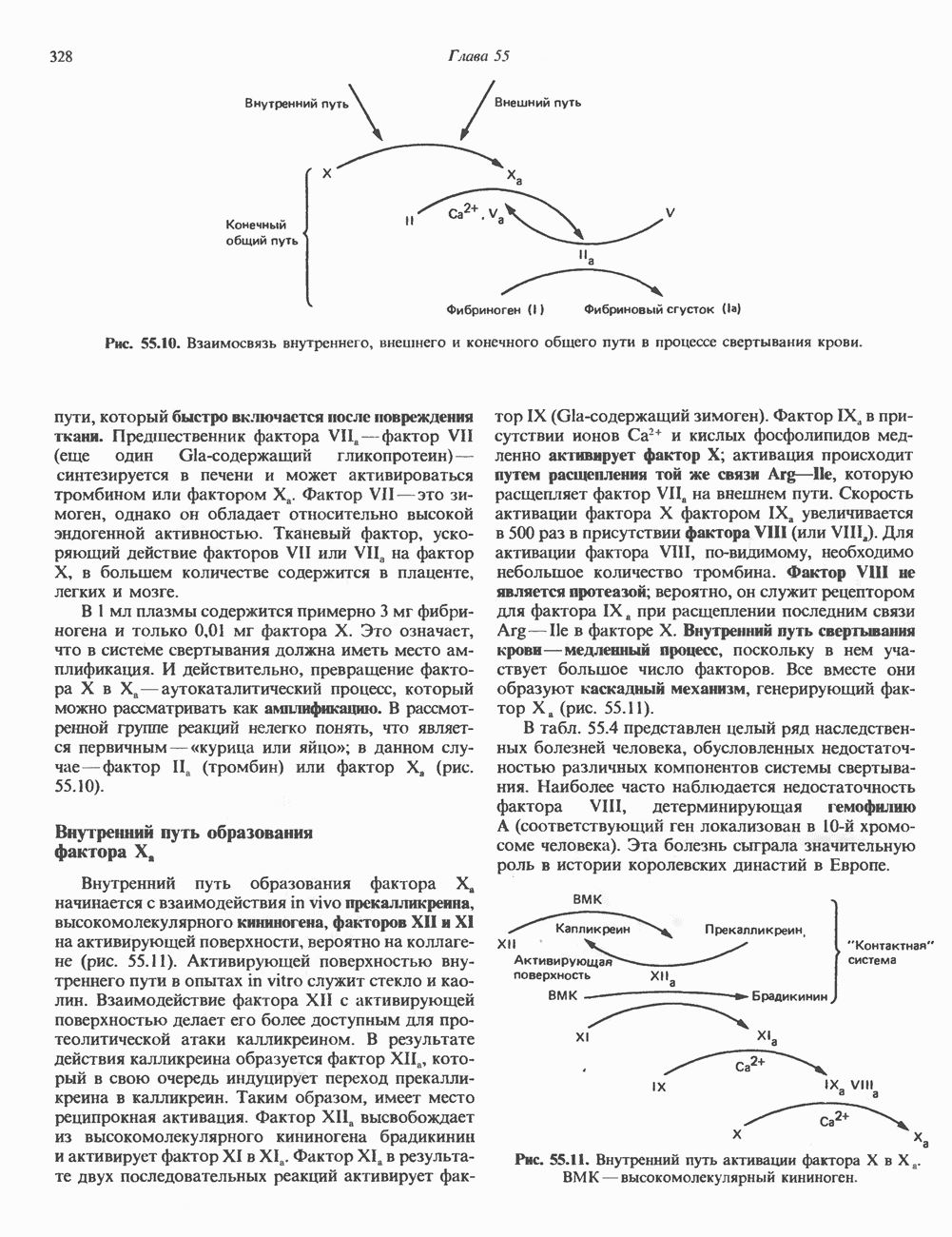
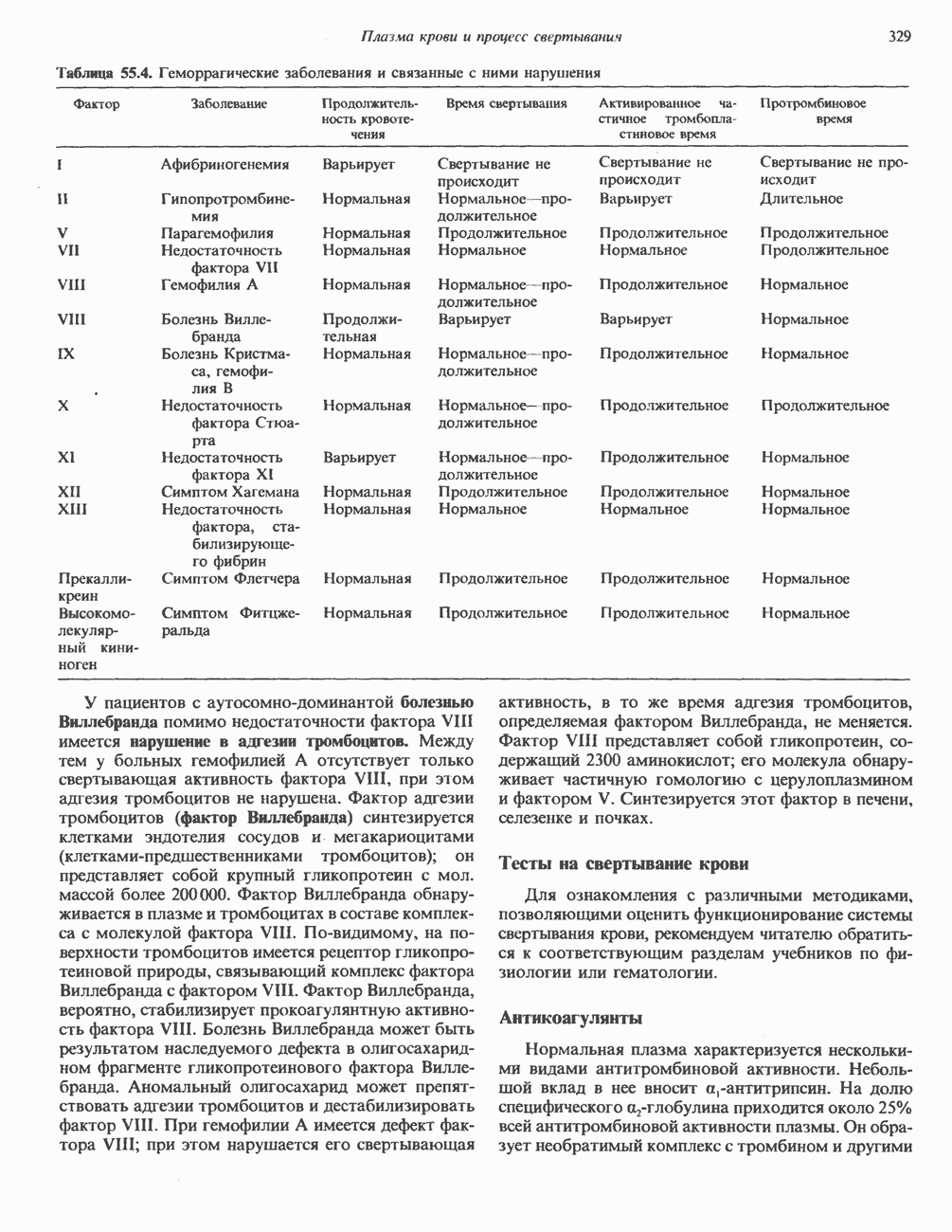
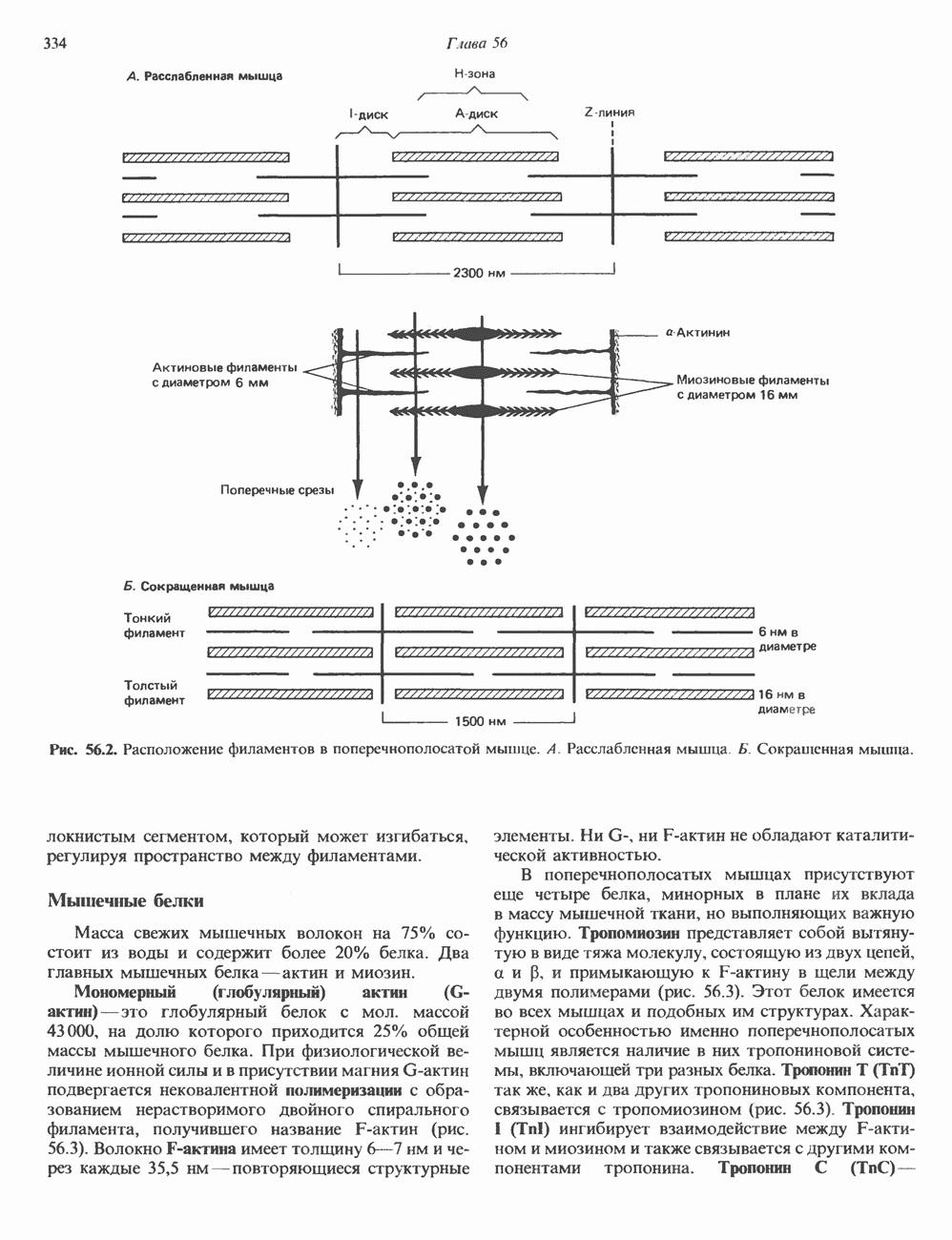
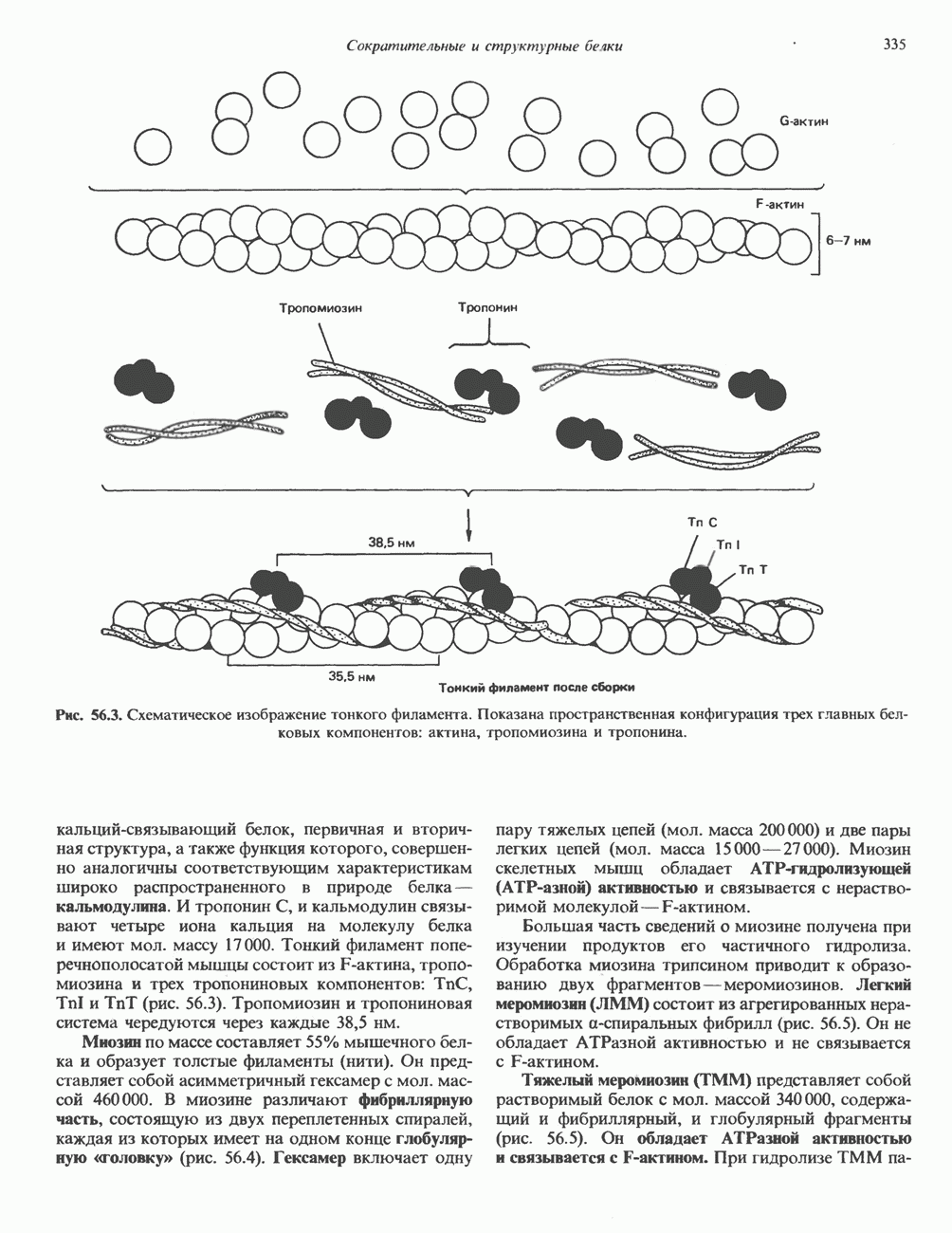
ПИРИМИДИНЫБиосинтез пиримидиновСтруктура ядра пиримидинов проще и путь их биосинтеза короче, чем у пуринов. В то же время оба пути имеют ряд общих предшественников. ФРПФ, глутамин, Синтез пиримидинового кольца (рис. 35.15) начинается с образования карбамонлфосфата из глутамина, АТР и Первый уникальный для биосинтеза пиримидинов этап — образование карбамоиласпартата в реакции конденсации карбамонлфосфата и аспартата катализируется аепартаттранскарбамоилазой (реакция 2). Затем в реакции, катализируемой дигидрооротазой, выщепляется На следующем этапе происходит дегидрогенирование под действием дигидрооротатдегидрогеназы с использованием NAD в качестве кофактора, при этом образуется оротовая кислота (реакция 4). В реакции 5 к оротовой кислоте присоединяется остаток рибозофосфата с образованием оротидилата (оротидинмонофосфат, ОМР). Этот процесс осуществляется оро гат-фосфорибозилтрансферазой — ферментом, аналогичным гипоксантин-гуанин— фосфорибозилтрансферазе и аденин-фосфори-бозилтрансферазе, которые участвуют в фосфорибозилировании пуриновых колец. Первый истинный пиримидиновый рибонуклеотид—уридилат (уридинмонофосфат, UMP) образуется при декарбоксилировании оротидилата (реакция 6). Таким образом, только на предпоследней стадии образования UMP происходит фосфорибозилирование гетероцикла. Дигидрооротатдегидрогеназа — митохондриальный фермент. Все остальные ферменты, участвующие в синтезе пиримидинов de novo, локализуются в цитозоле. Фосфорилирование пиримидиновых нуклеозид-монофосфатов до соответствующих ди- и трифосфатов происходит аналогично тому, как это описано для пуриновых нуклеозидмонофосфатов (реакции 7-12). UTP аминируется до СТР; в реакции участвуют глутамин и АТР (реакция 9). Механизм восстановления пиримидиннуклеозидцифосфатов до соответствующих Образование тимидилата (тимндинмонофосфат; ТМР) (реакция 12) - единственная реакция на пути биосинтеза пиримидиновых нуклеотидов, требующая участия производного тетрагидрофолата в качестве донора одноуглеродного фрагмента. 2-Дезокси-UMP метилируется тимидилатсинтазой, использующей (см. скан) Рис. 35.15. Путь биосинтеза пиримидиновых нуклеотидов. метильной с одновременным окислением тетрагидрофолата до дигидрофолата. Для того чтобы фолатный переносчик и далее мог функционировать, необходимо восстановить дигидрофолат до тетрагидрофолата. Эту реакцию катализирует дигидрофолатредуктаза. Именно поэтому делящиеся клетки, вынужденные синтезировать ТМР с образованием дигидрофолата, оказываются особенно чувствительны к ингибиторам дигидрофолатредуктазы. Один из таких ингибиторов — метотрексат (аметоптерин) широко используется как противоопухолевый препарат. Пути регенерации пиримидиновых нуклеотидовКлетки млекопитающих не располагают эффективными средствами использования свободных пиримидиновых оснований для синтеза нуклеотидов. В то же время они обладают способностью утилизировать пиримидиновые рубонуклеозиды уридин и цитидин, а также Фермент оротат-фосфорибозилтрансфераза, необходимый для синтеза пиримидинов de novo, отвечает за фосфорибозилирование оротовой кислоты с образованием ОМР, хотя оротовая кислота, строго говоря, и не является истинным пиримидиновым основанием. Оротатфосфорибозилтрансфераза не может использовать в качестве субстратов нормальные пиримидиновые основания, но способна фосфорибозилировать аллопуринол ( Катаболизм пиримидиновВ результате катаболизма пиримидинов, протекающего в основном в печени, образуются хорошо растворимые конечные продукты (рис. 35.17). Именно этим они отличаются от конечных продуктов катаболизма пуринов (мочевая кислота и ее натриевая соль обладают слабой растворимостью). Выделение Тимин выступает в роли предшественника р-аминоизобутирата у человека и обычных лабораторных животных.
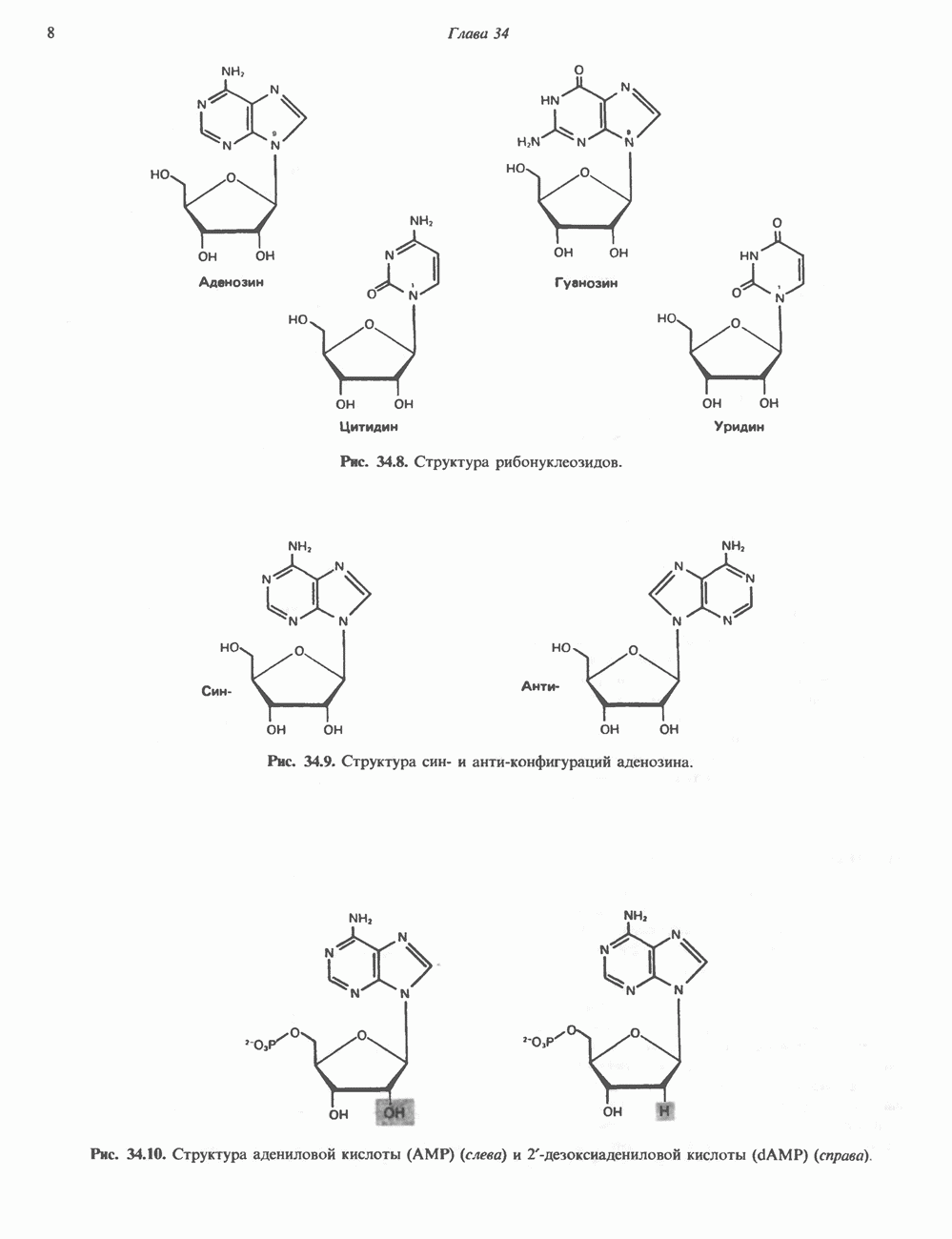
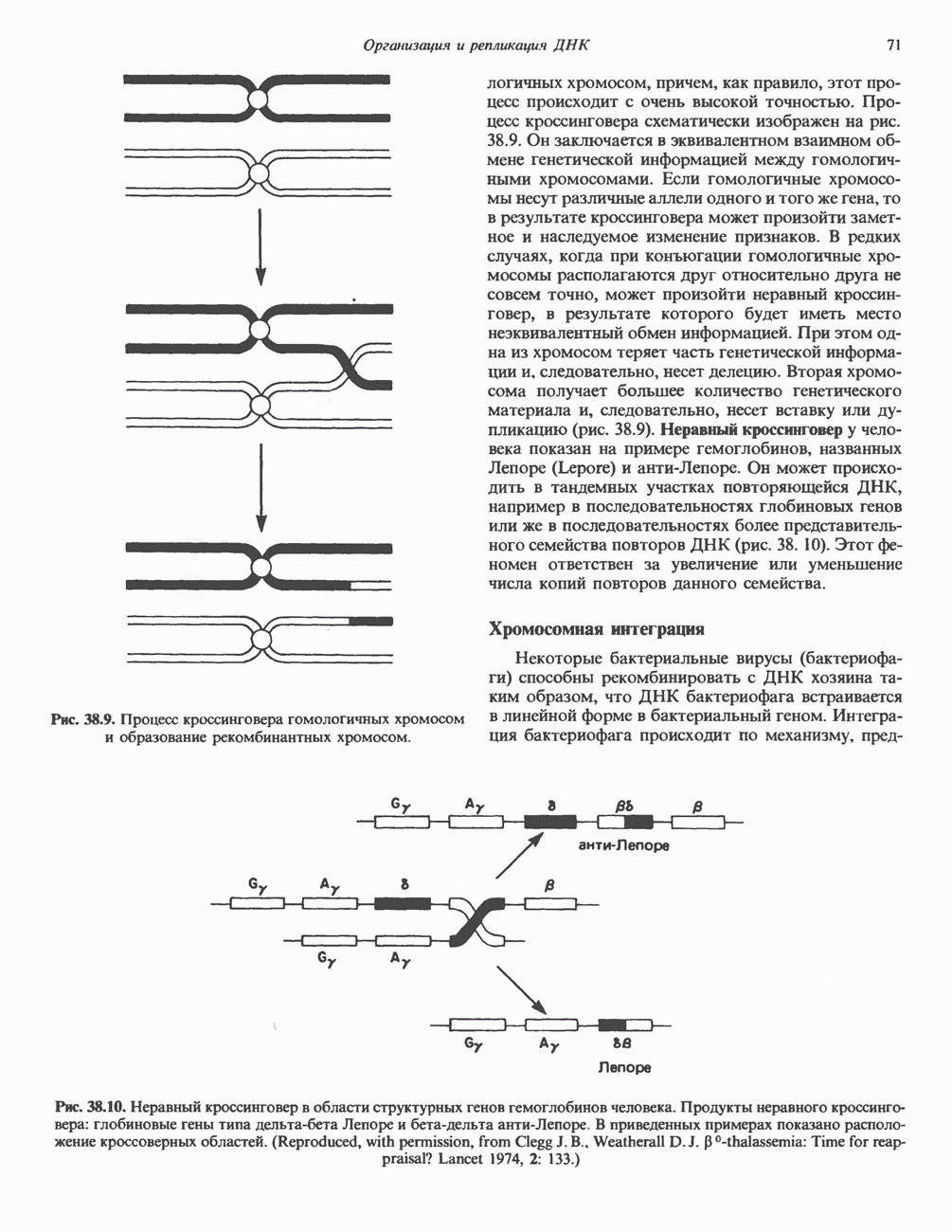
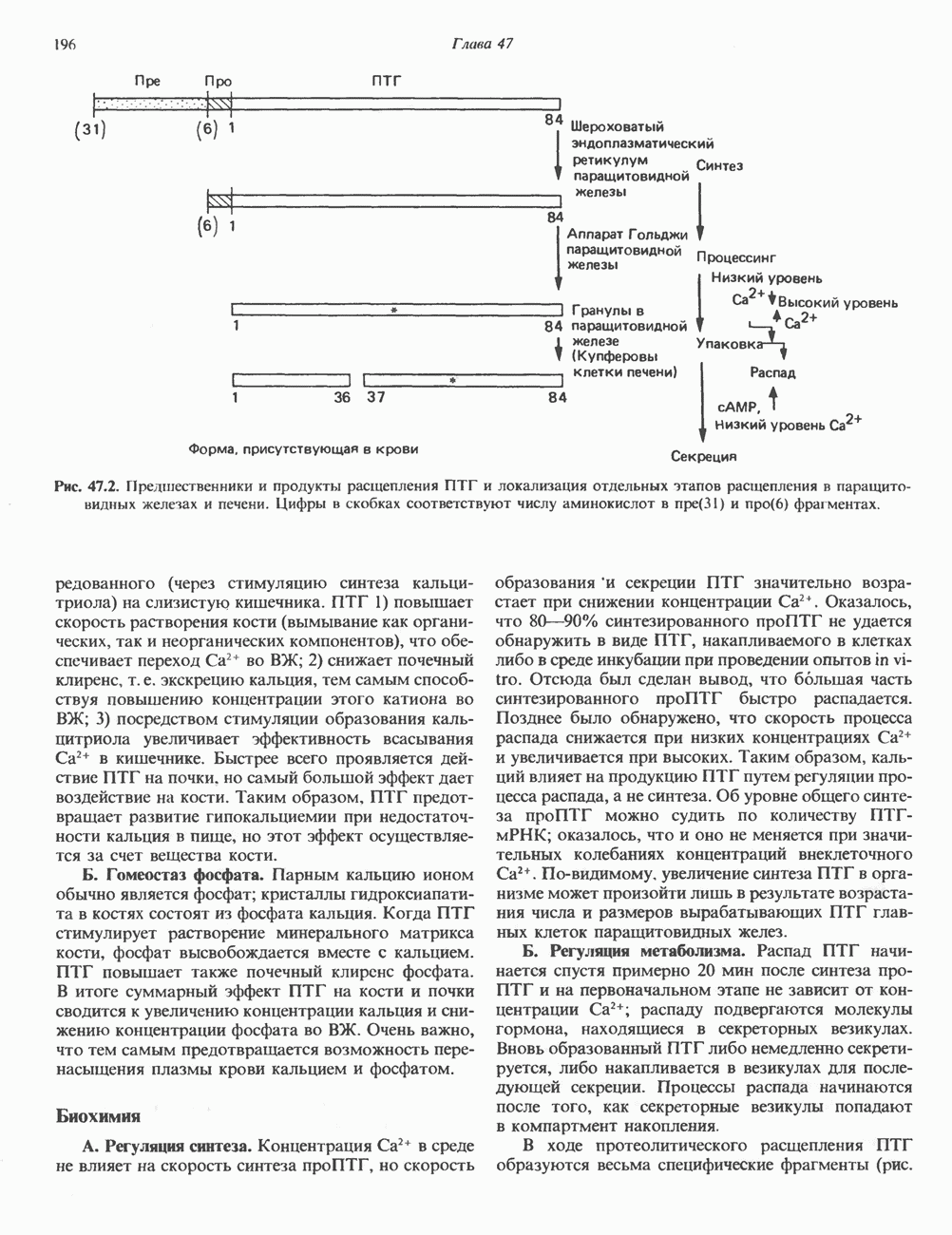
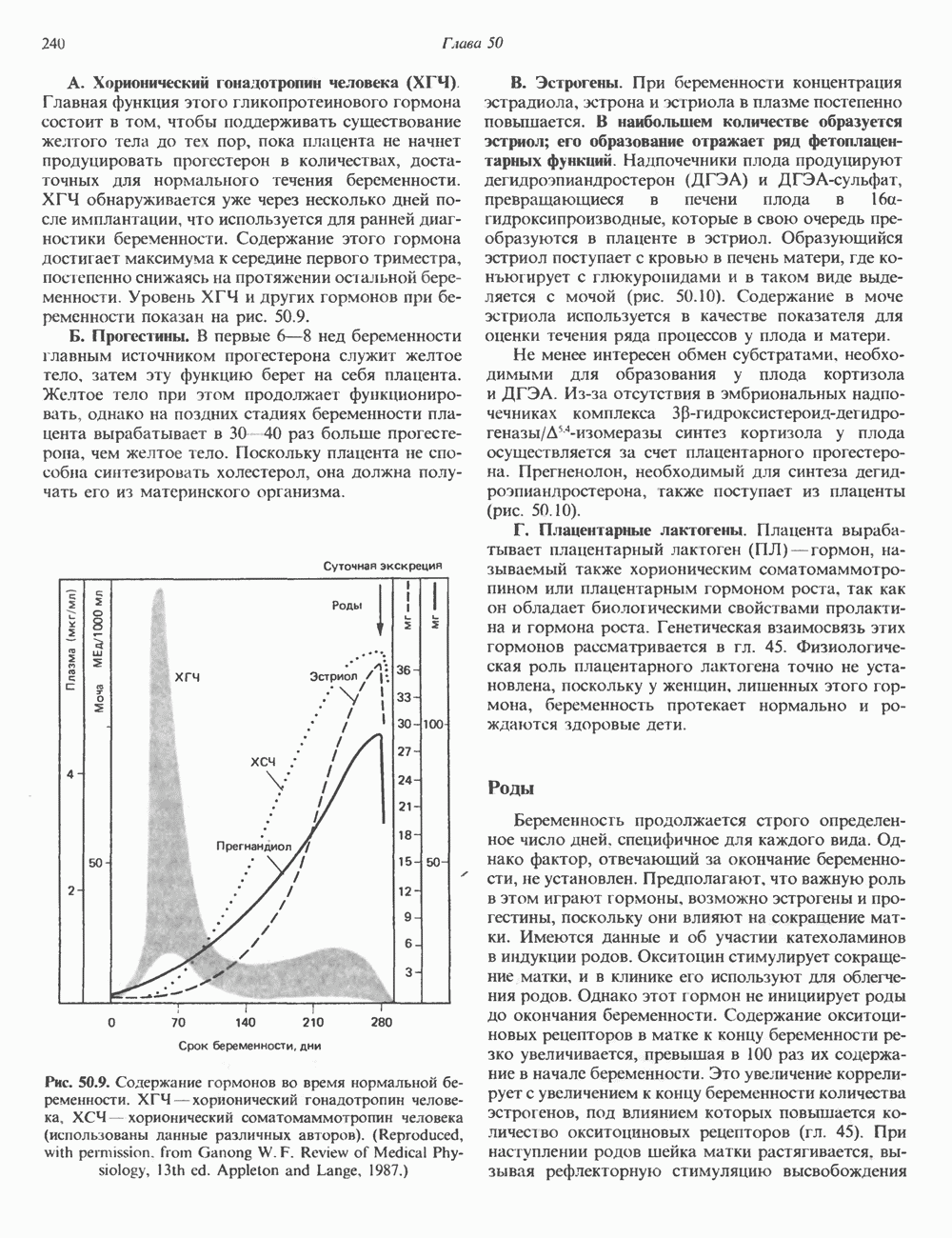
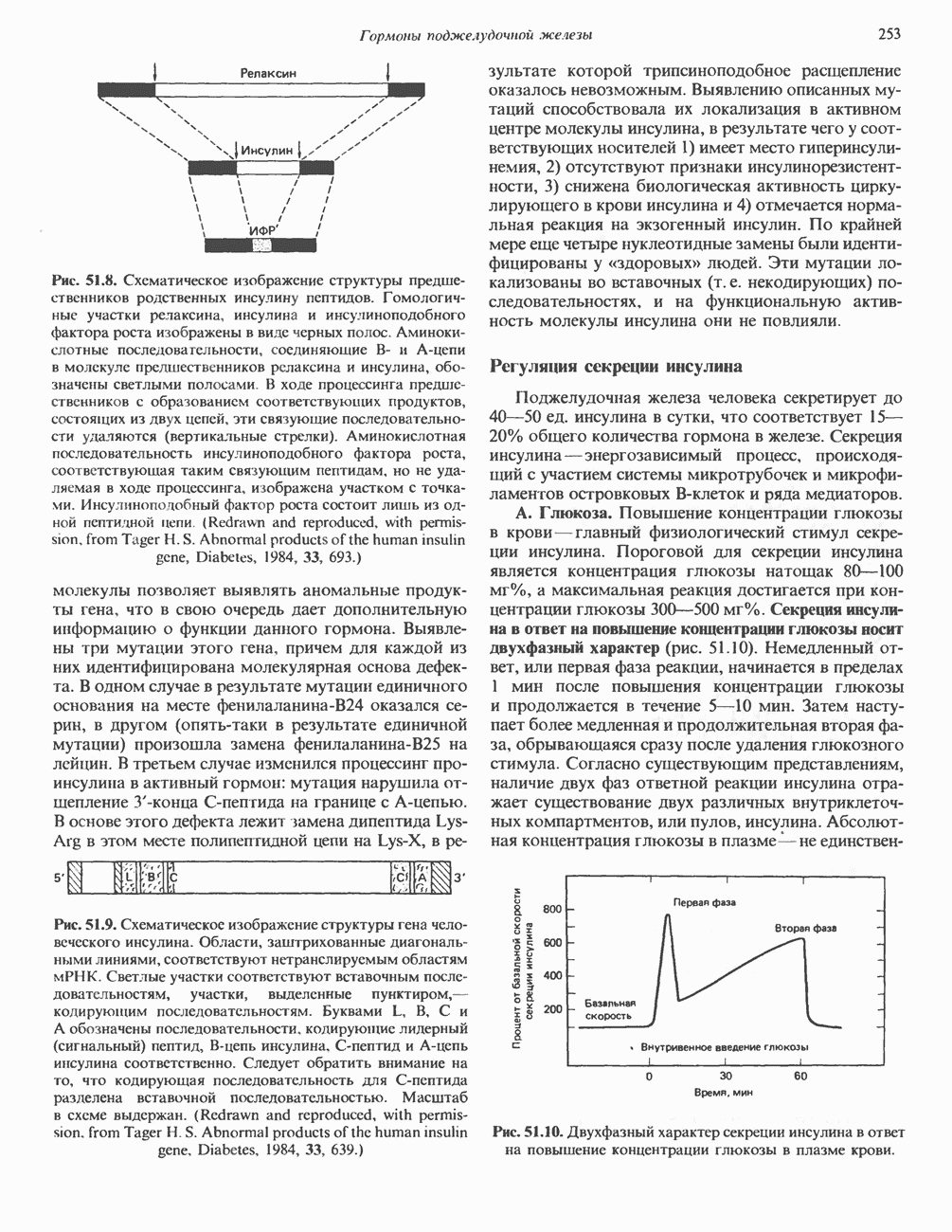
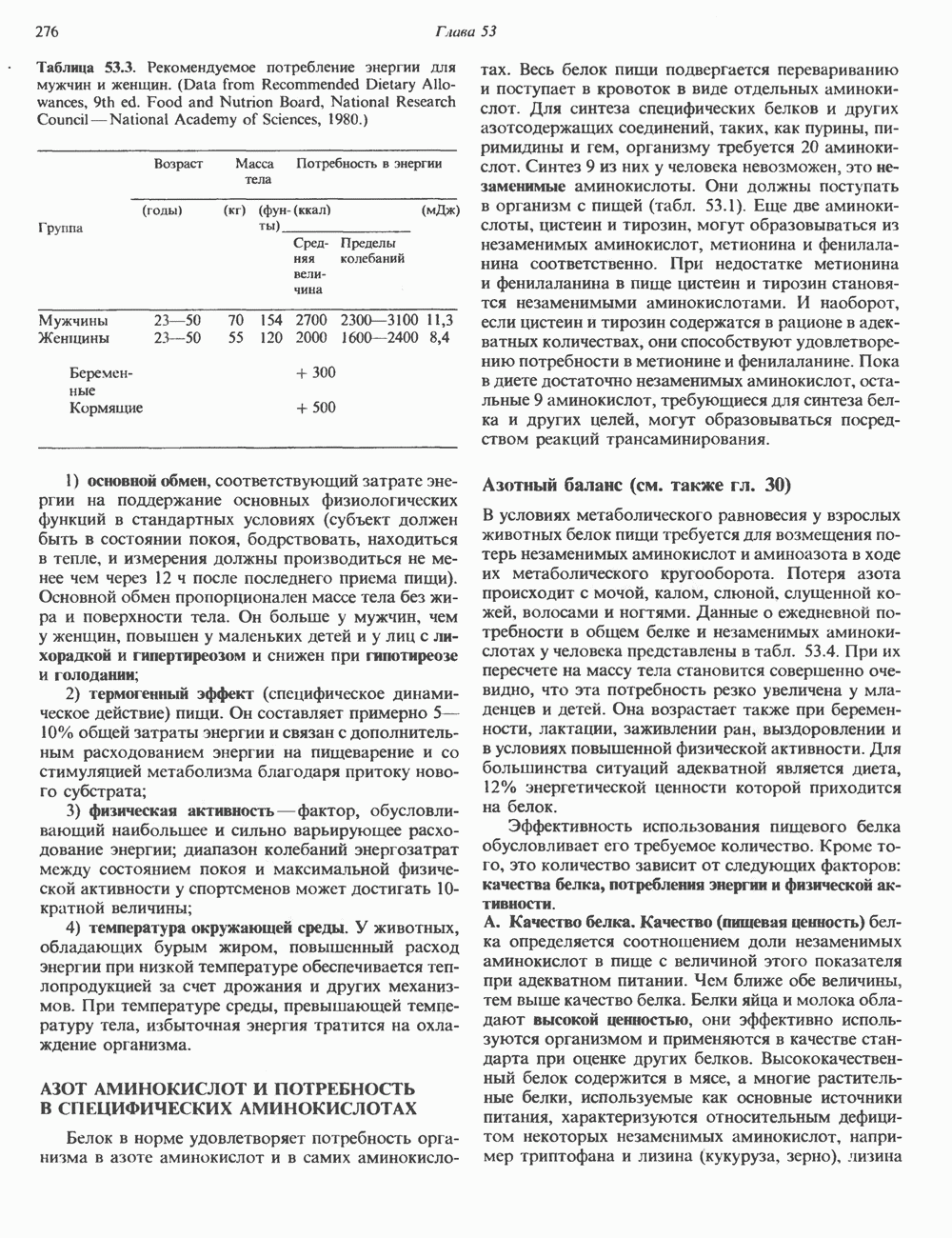
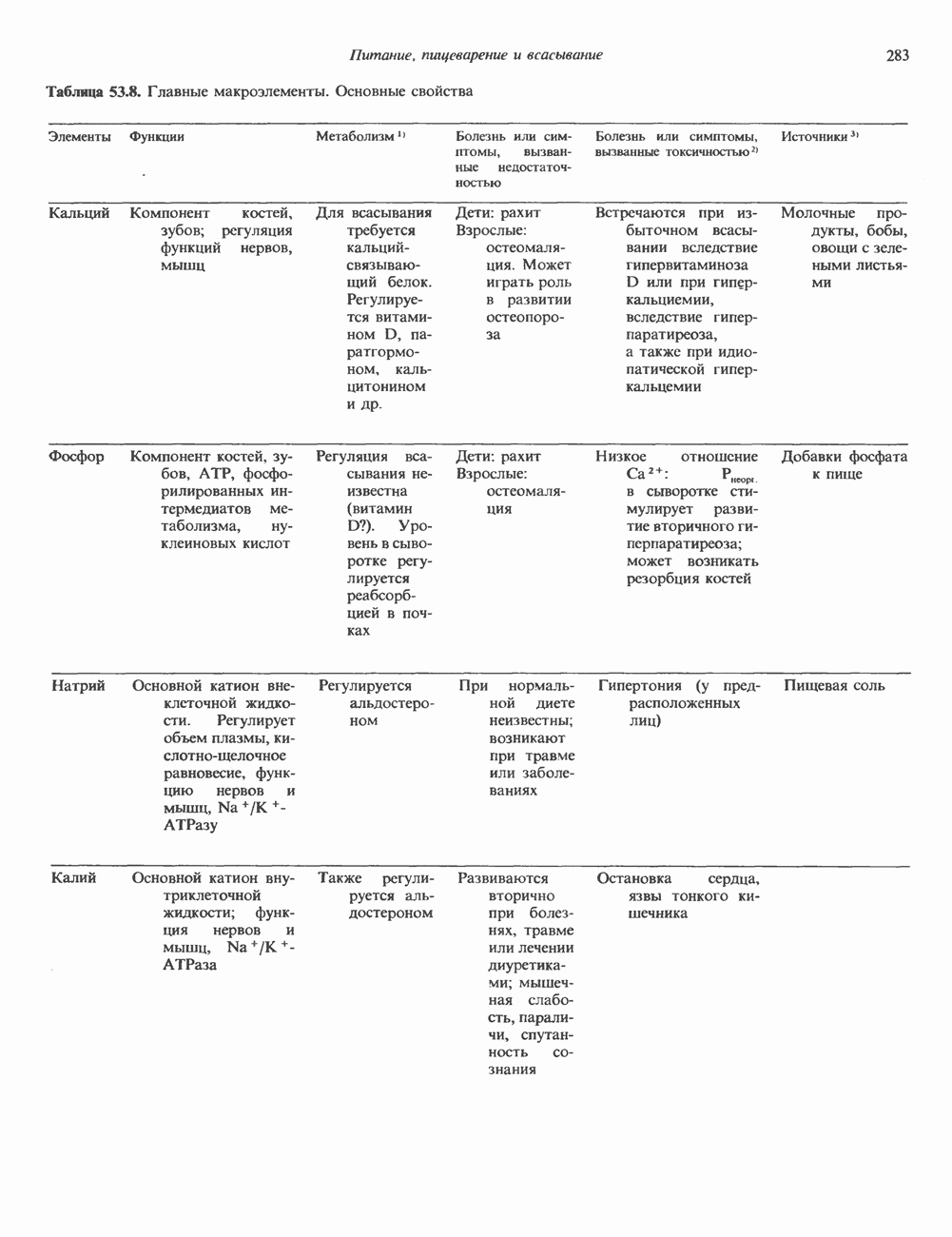
Рис. 35.16. Реакции образования пиримидиновых нуклео-зидмонофосфатов из соответствующих пиримидиновых нуклеозидов, катализируемые нуклеозидкиназой. Экскреция Р-аминоизобутирата увеличивается при лейкемии и после рентгеновского облучения, что, без сомнения, отражает ускорение гибели клеток и деструкцию их ДНК. Выделение аномально больших количеств Р-аминоизобутирата может наблюдаться и у здоровых во всех остальных отношениях людей. Этот признак наследуется как рецессивный и, следовательно, проявляется только у гомозигот по соответствующему аллелю. Примерно у 25% обследованных индивидов (японцев и китайцев по происхождению) обнаружено повышение уровня экскреции Р-аминоизобутирата. Сравнительно немного известно о механизме деградации Р-аминоизобутирата в организме человека. Фермент, катализирующий обратимое трансаминирование этого соединения, обнаружен в почках свиньи. Р-Аминоизобутират превращается в метилмалоновый полуальдегид, затем в пропионат, который в свою очередь преобразуется в сукцинат. Начальные стадии деградации пиримидиновых нуклеотидов, включающие этап отщепления углеводфосфатного фрагмента по N-гликозидной связи, весьма напоминают обращенные последние стадии пути биосинтеза. Для псевдоуридина, образующегося in situ в результате внутренней перестройки, не существует механизма гидролиза или фосфоролиза до урацила. Соответственно этот необычный нуклеотид у здоровых индивидов экскретируется с мочой неизмененным.
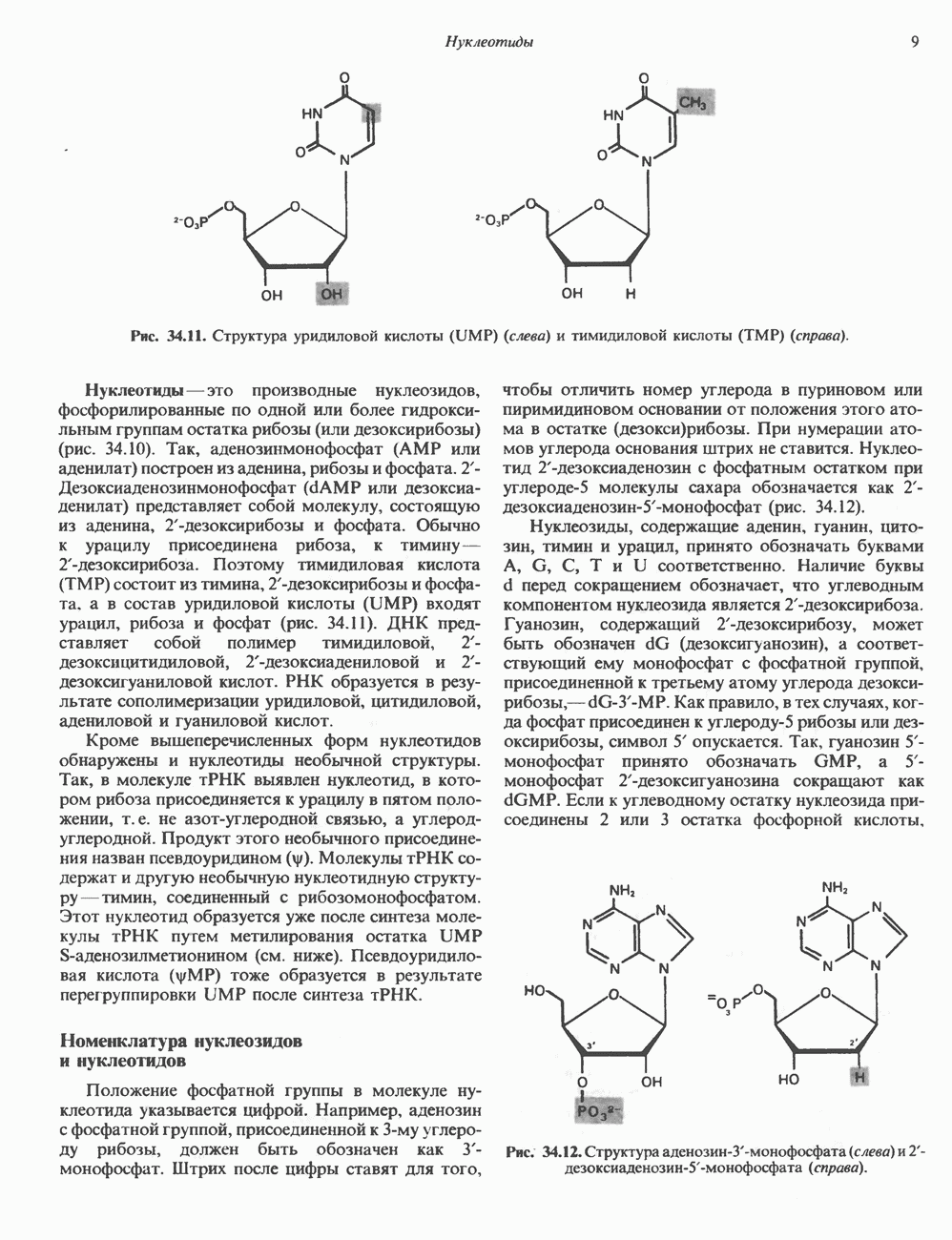
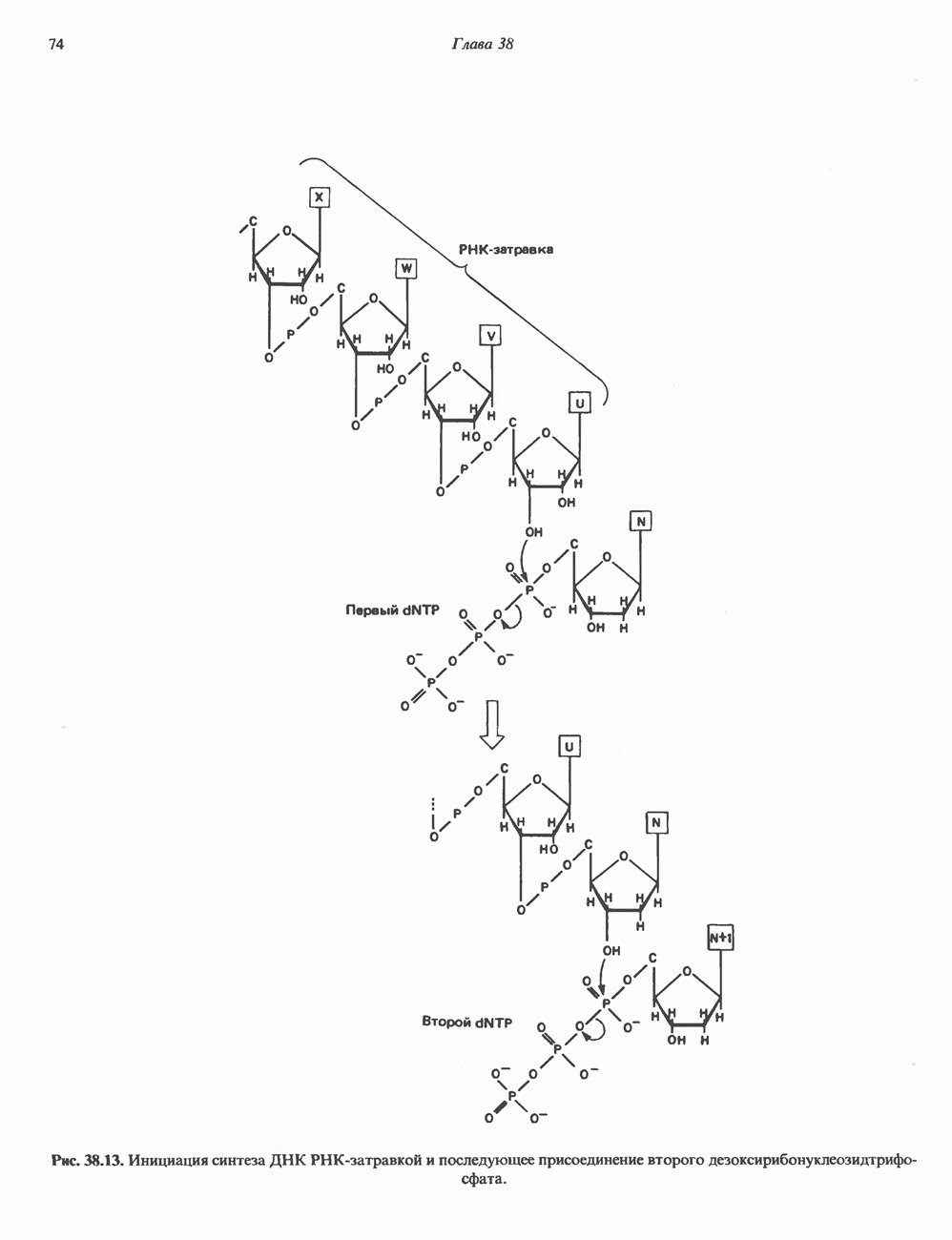
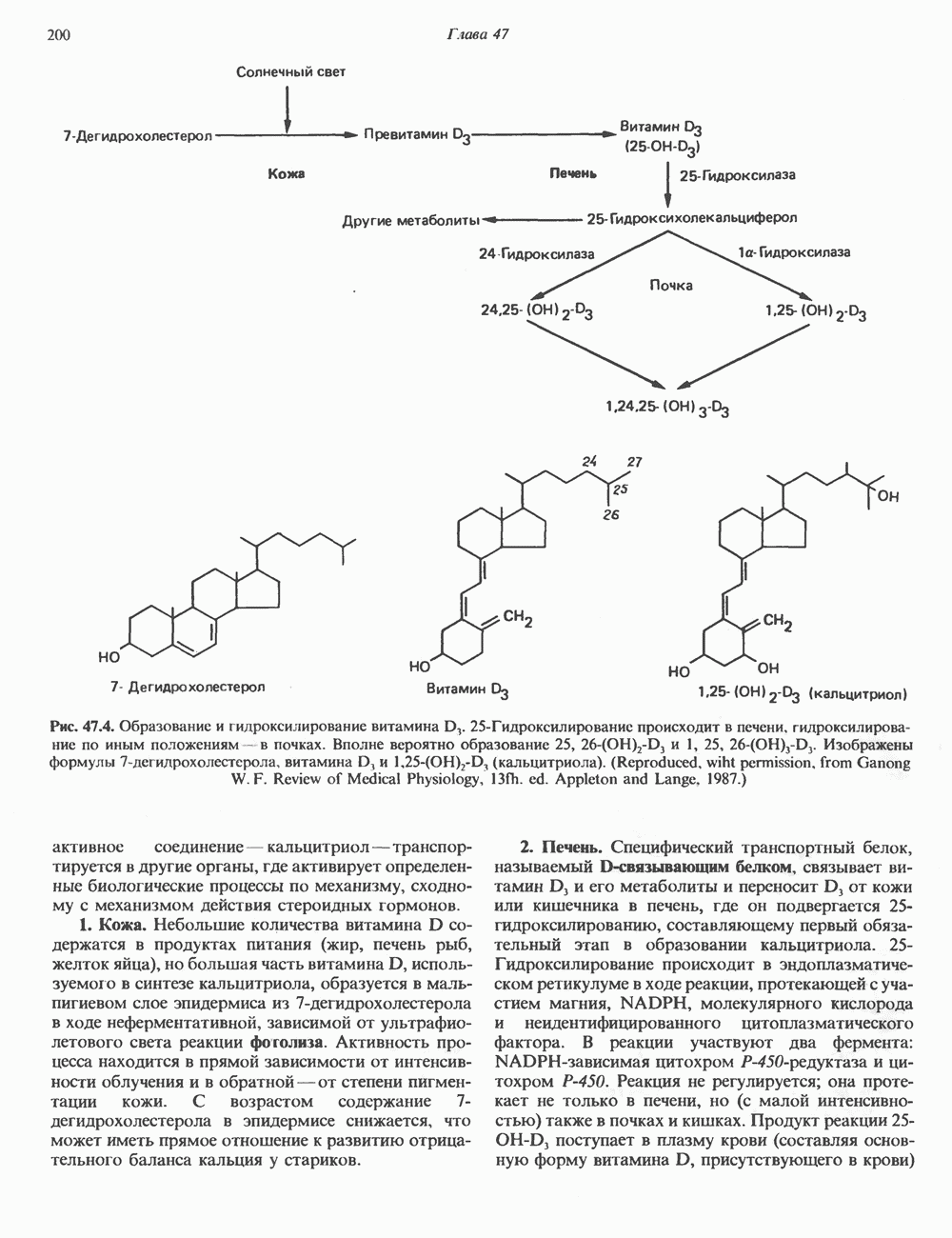
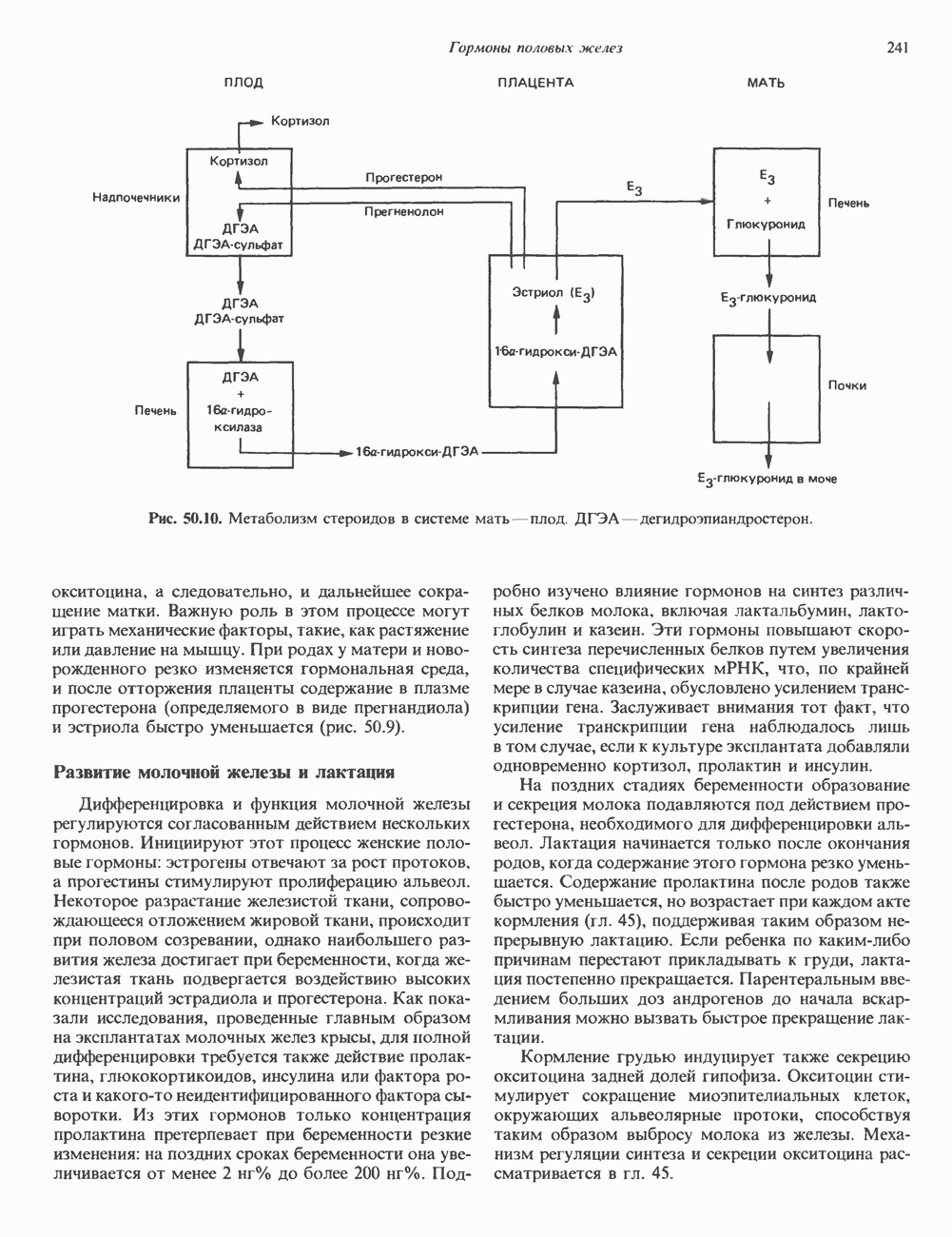
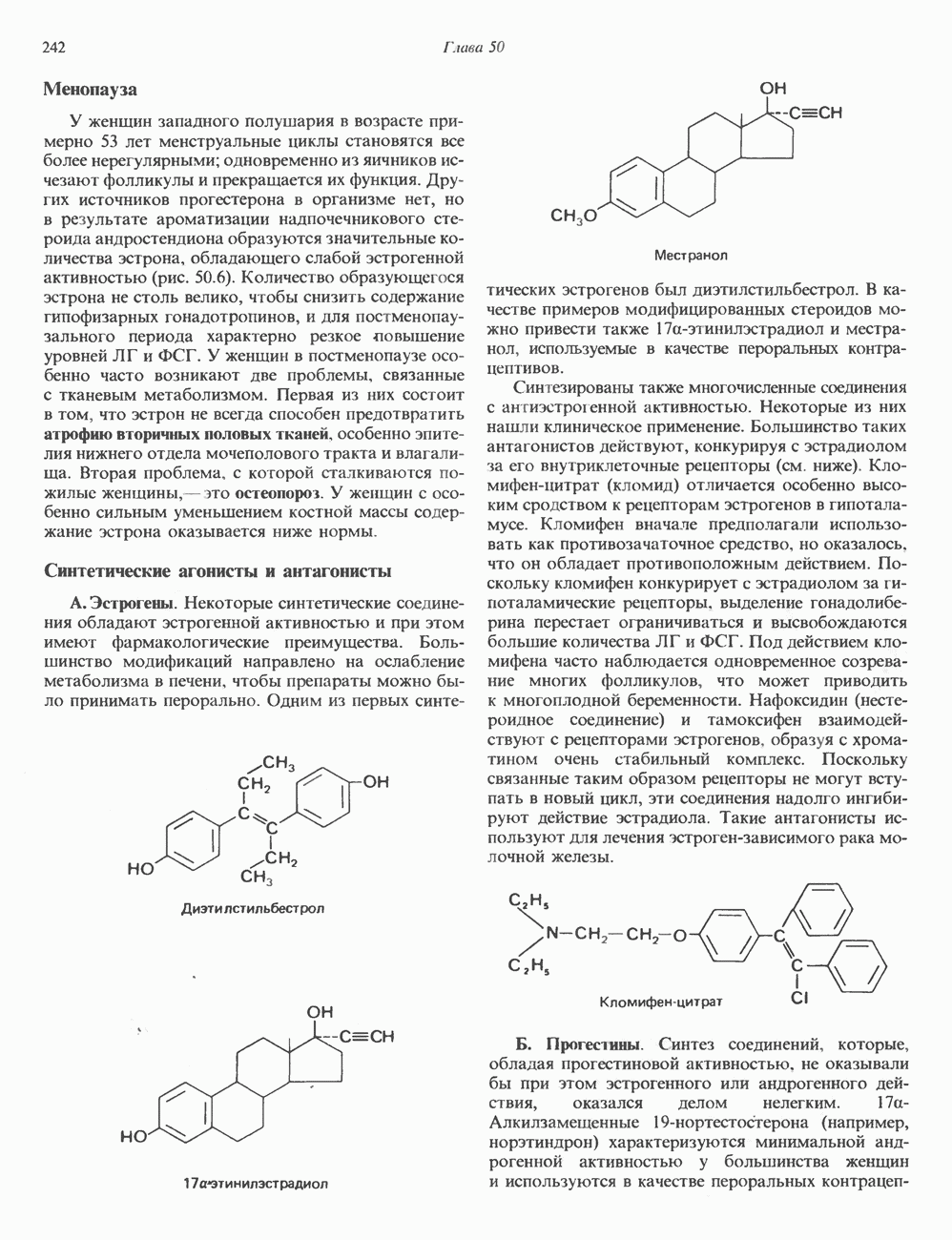
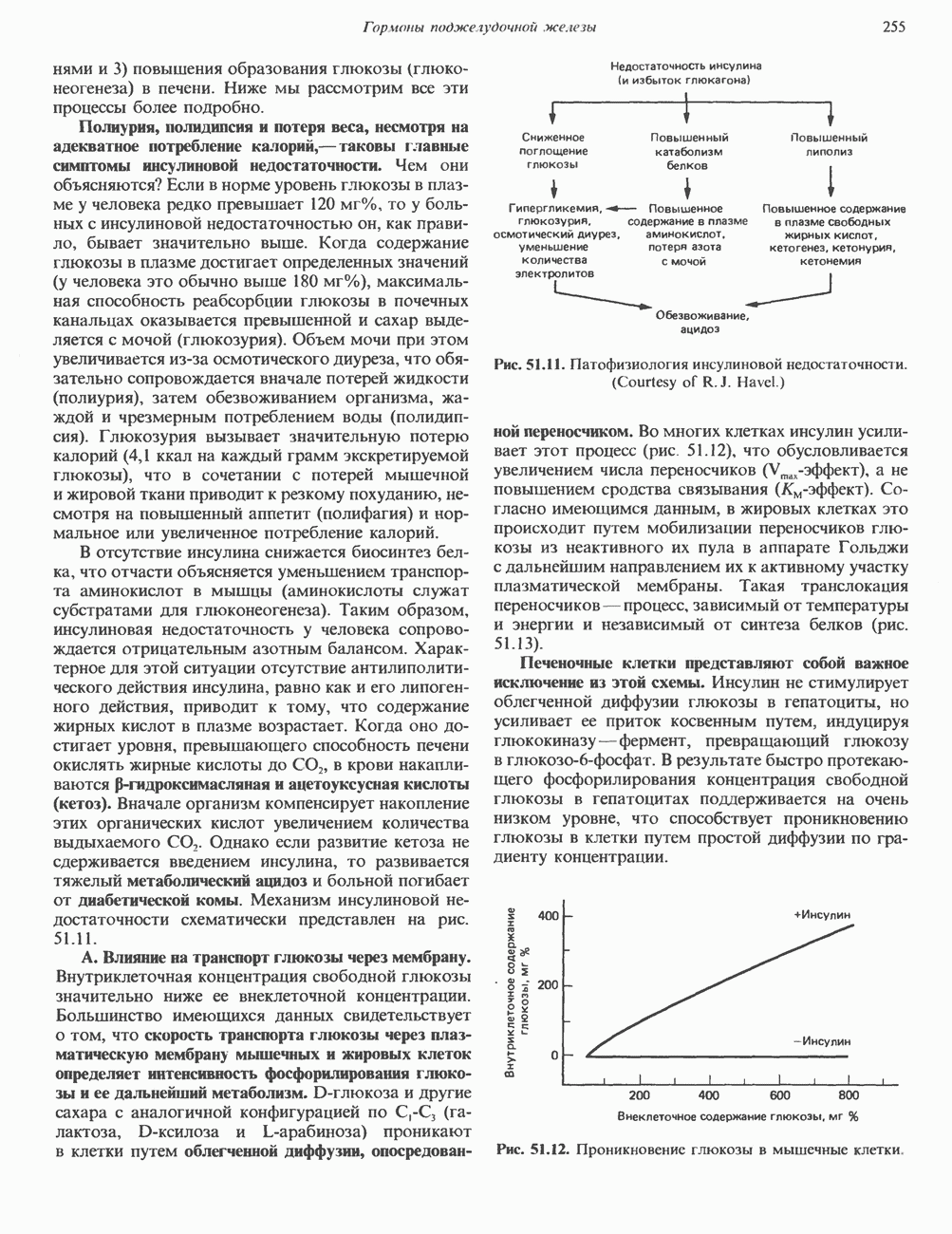
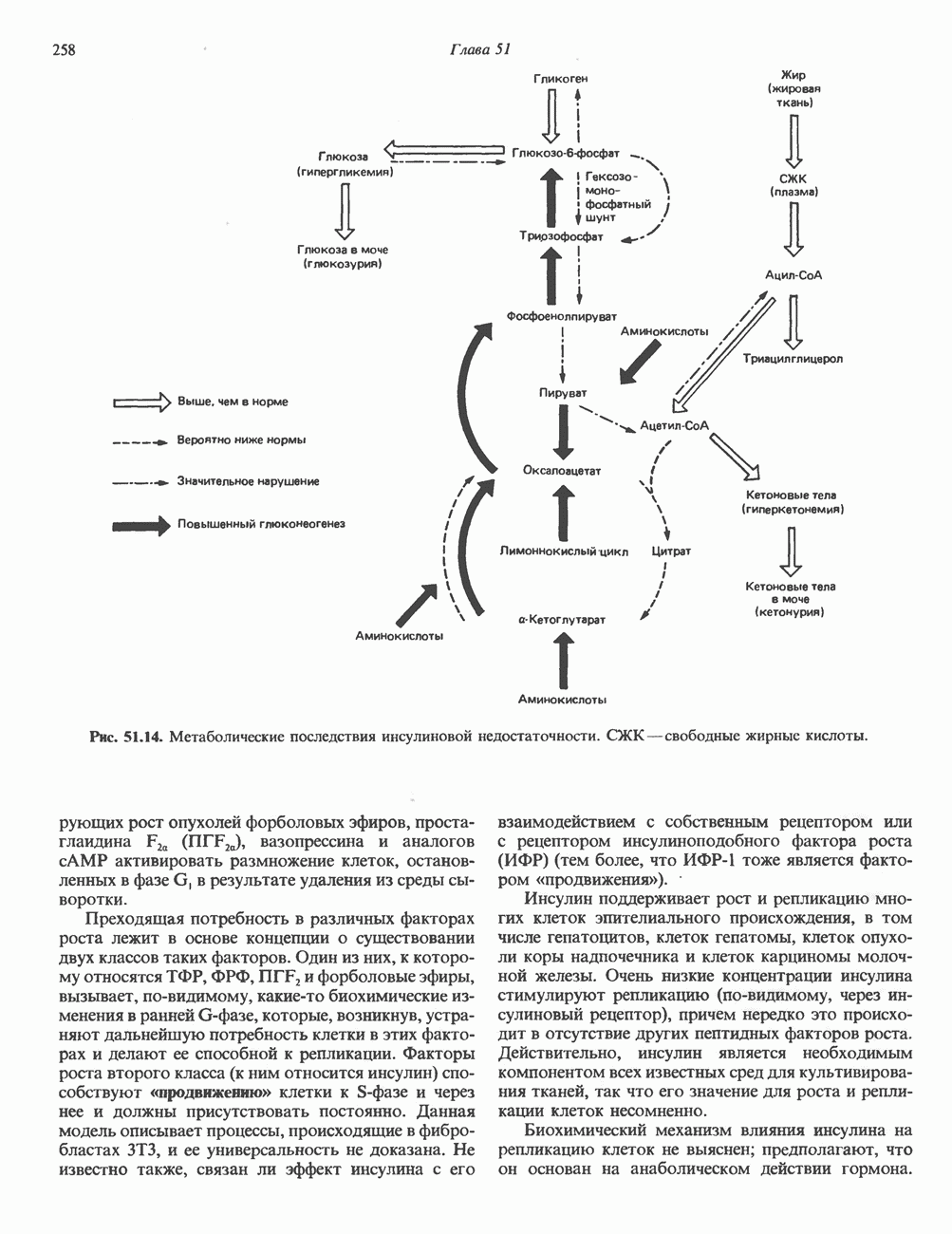
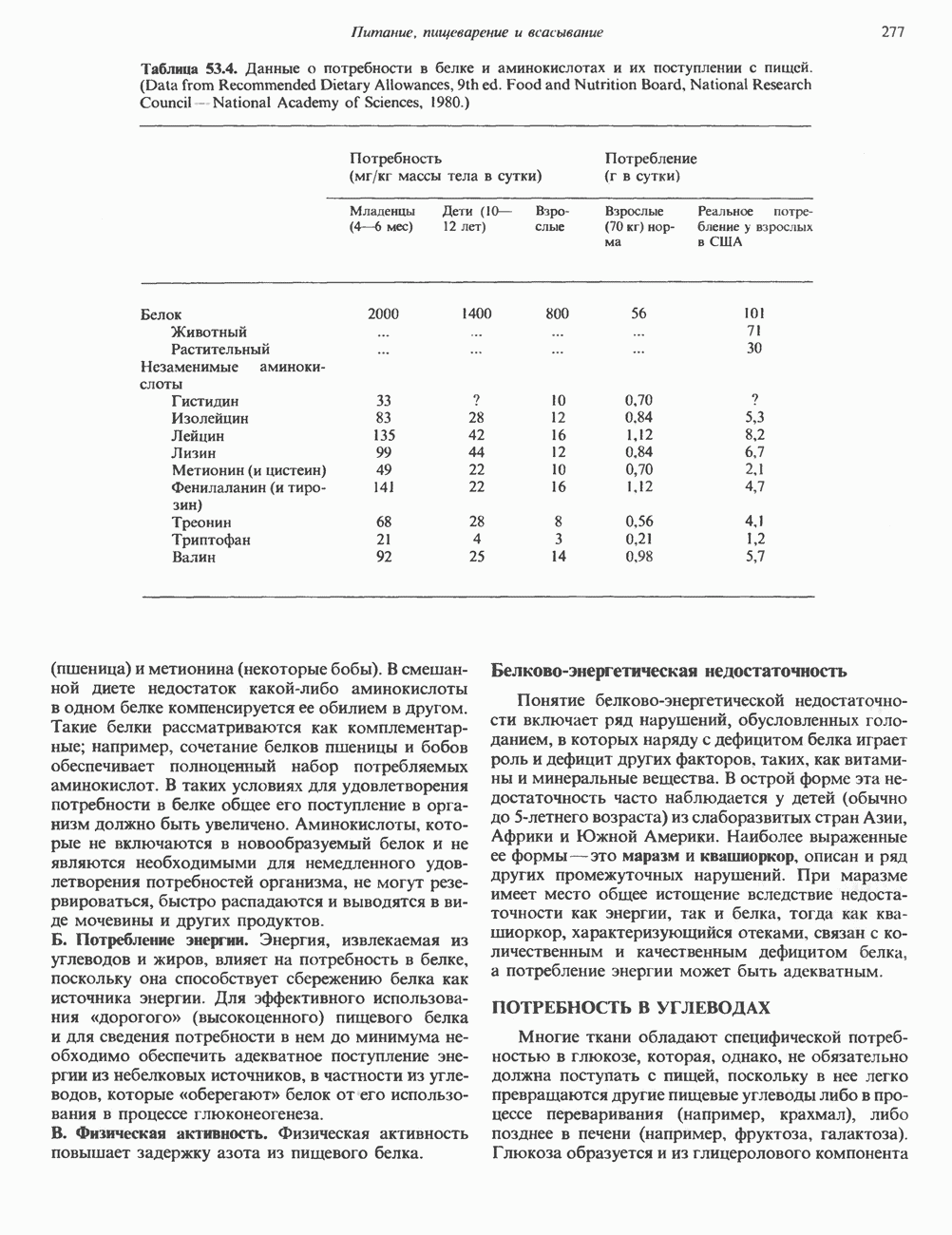
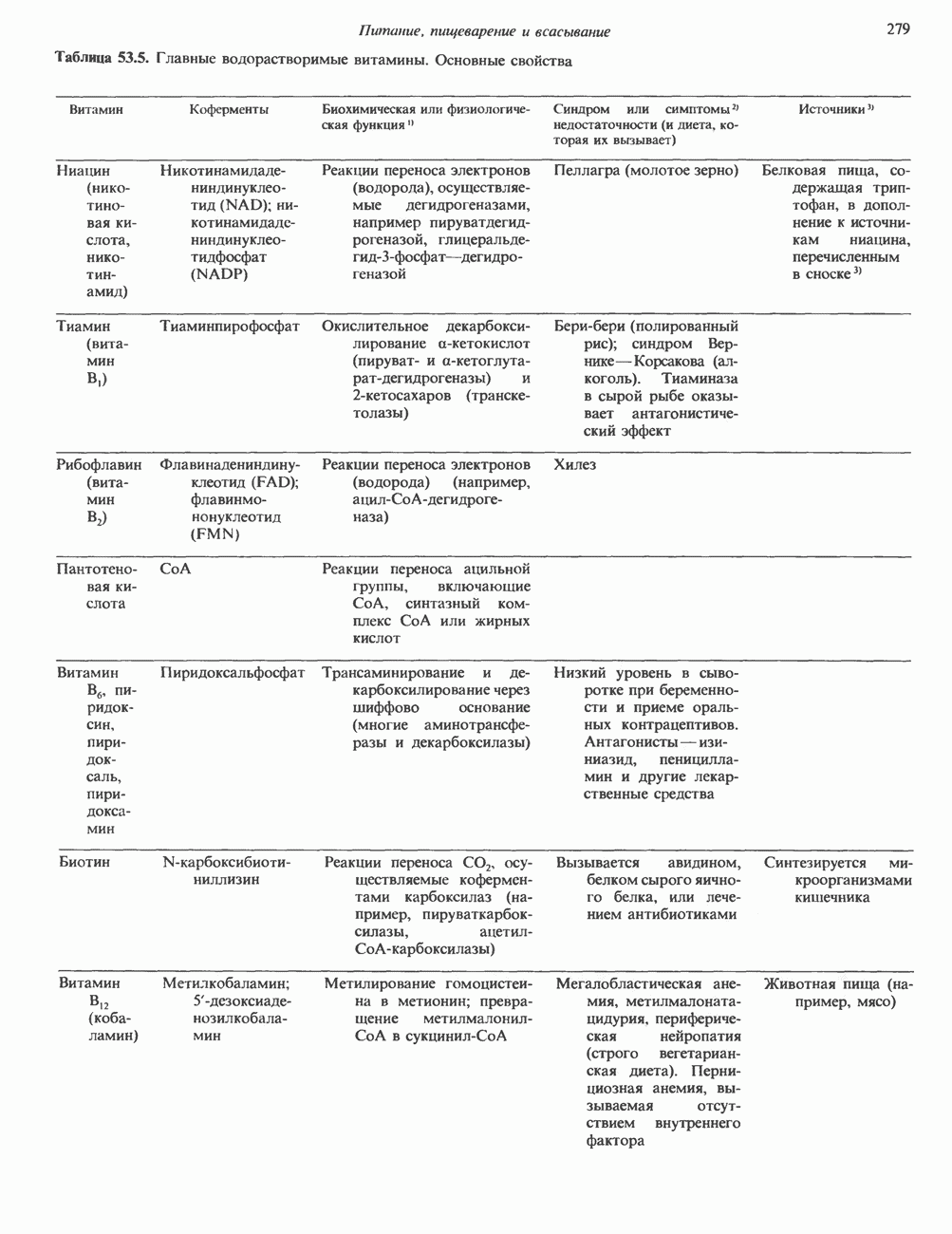
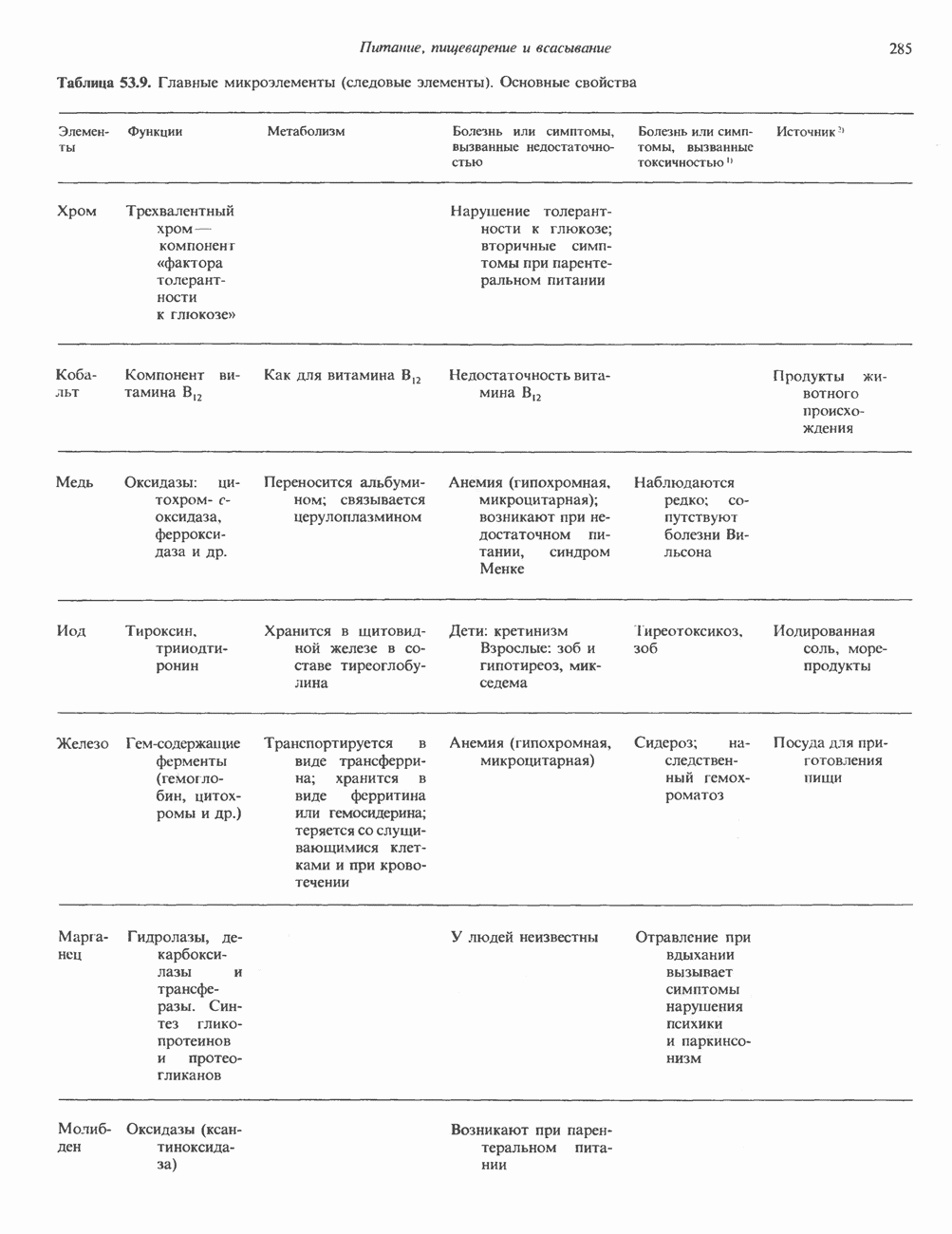
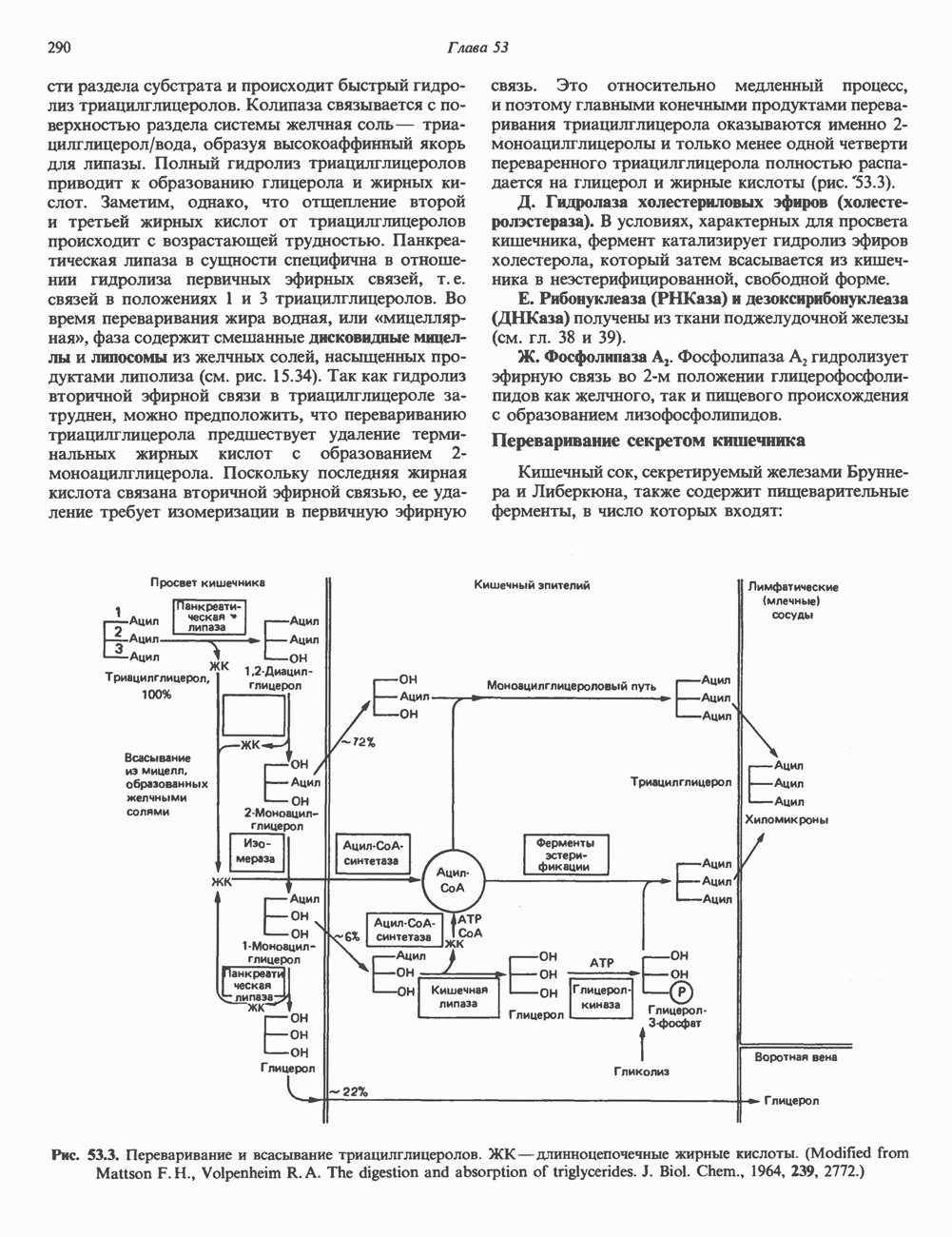
Рис. 35.17. Катаболизм пиримидинов. Регуляция биосинтеза пиримидиновПуть биосинтеза пиримидиновых нуклеотидов регулируется двумя различными механизмами. Активность первых двух ферментов находится под контролем аллостерических эффекторов. Кроме того, три первых и два последних фермента являются объектами координированной репрессии—дерепрессин. Карбамоилфосфатсинтаза ингибируется UTP, пуриновыми нуклеотидами, но активируется ФРПФ (рис. 35.18). Аспартаттранскарбамонлаза особенно чувствительна к ингибирующему влиянию СТР. Аллостерические свойства аспартаткарбамоилазы микроорганизмов явились предметом интенсивных и ставших уже классическими исследований механизмов аллостерической регуляции активности ферментов. Скорость биосинтеза пиримидинов коррелирует со скоростью биосинтеза пуринов, что указывает на координированный контроль синтеза нуклеотидов обоих типов. ФРПФ-синтетаза—фермент, катализирующий образование предшественника обоих путей биосинтеза, - ингибируется по принципу обратной связи как пуриновыми, так и пиримидиновыми нуклеотидами. Карбамоилфосфатсинтаза также подвержена ингибированию по принципу обратной связи нуклеотидами обоих типов, а ФРПФ активирует этот фермент. Таким образом, на нескольких этапах биосинтеза пуриновых и пиримидиновых нуклеотидов осуществляется перекрестная регуляция.
Рис. 35.18. Регуляция пути биосинтеза пиримидиновых нуклеотидов. Сплошные линии указывают путь химических превращений. Пунктирные линии обозначают положи гельную
|
 |
Оглавление
|




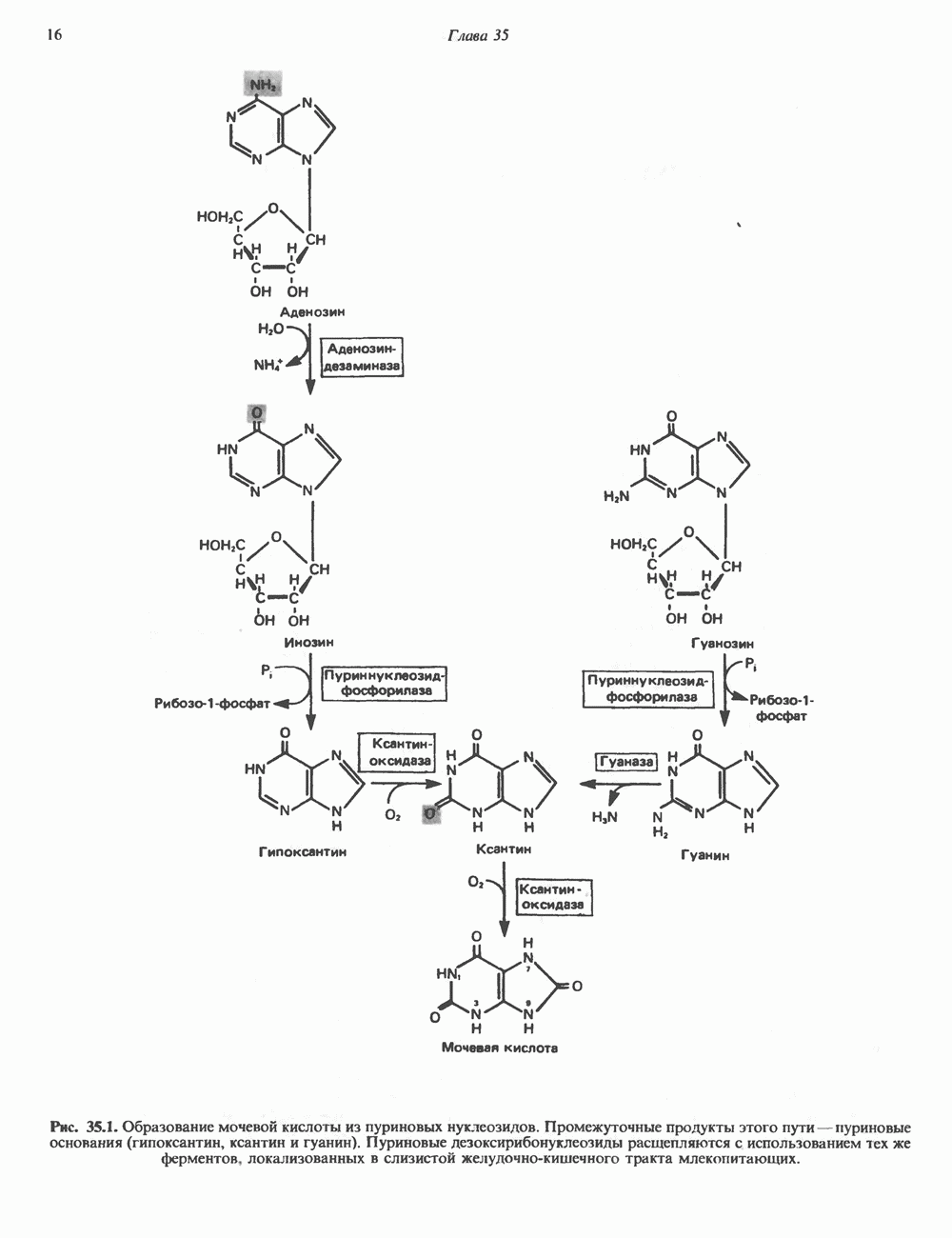

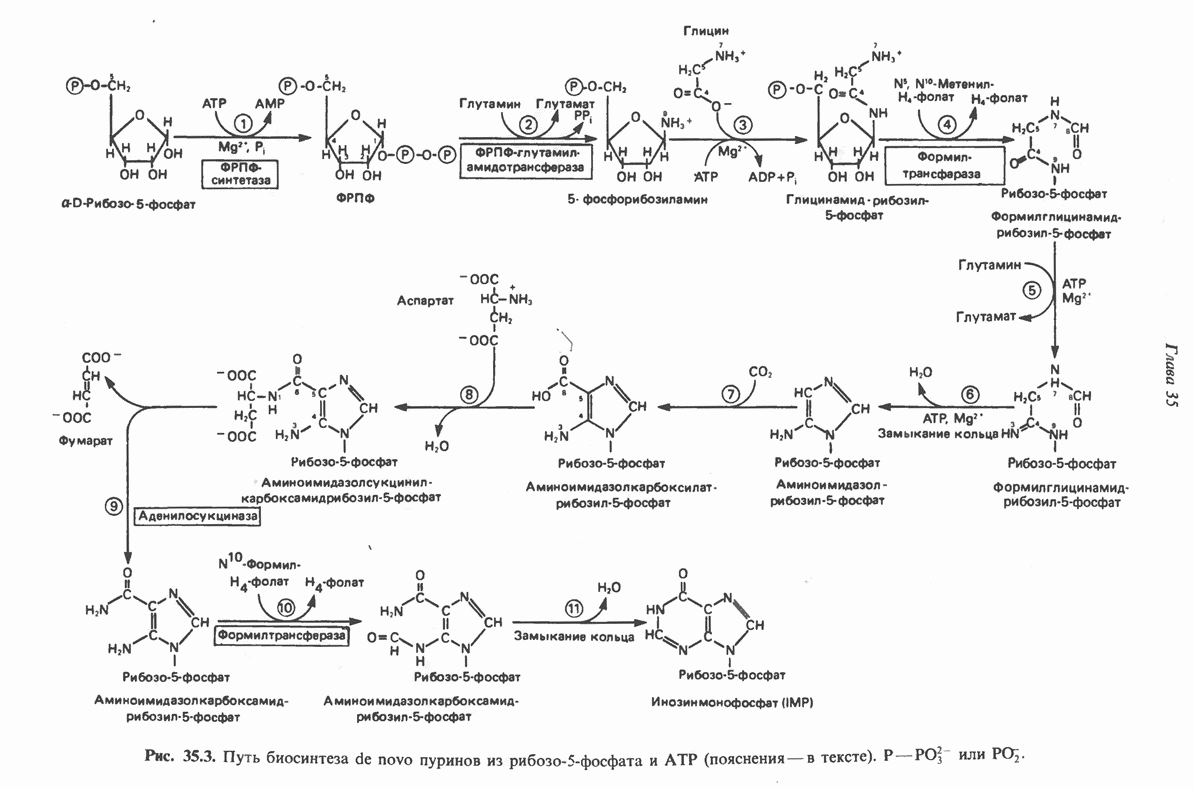
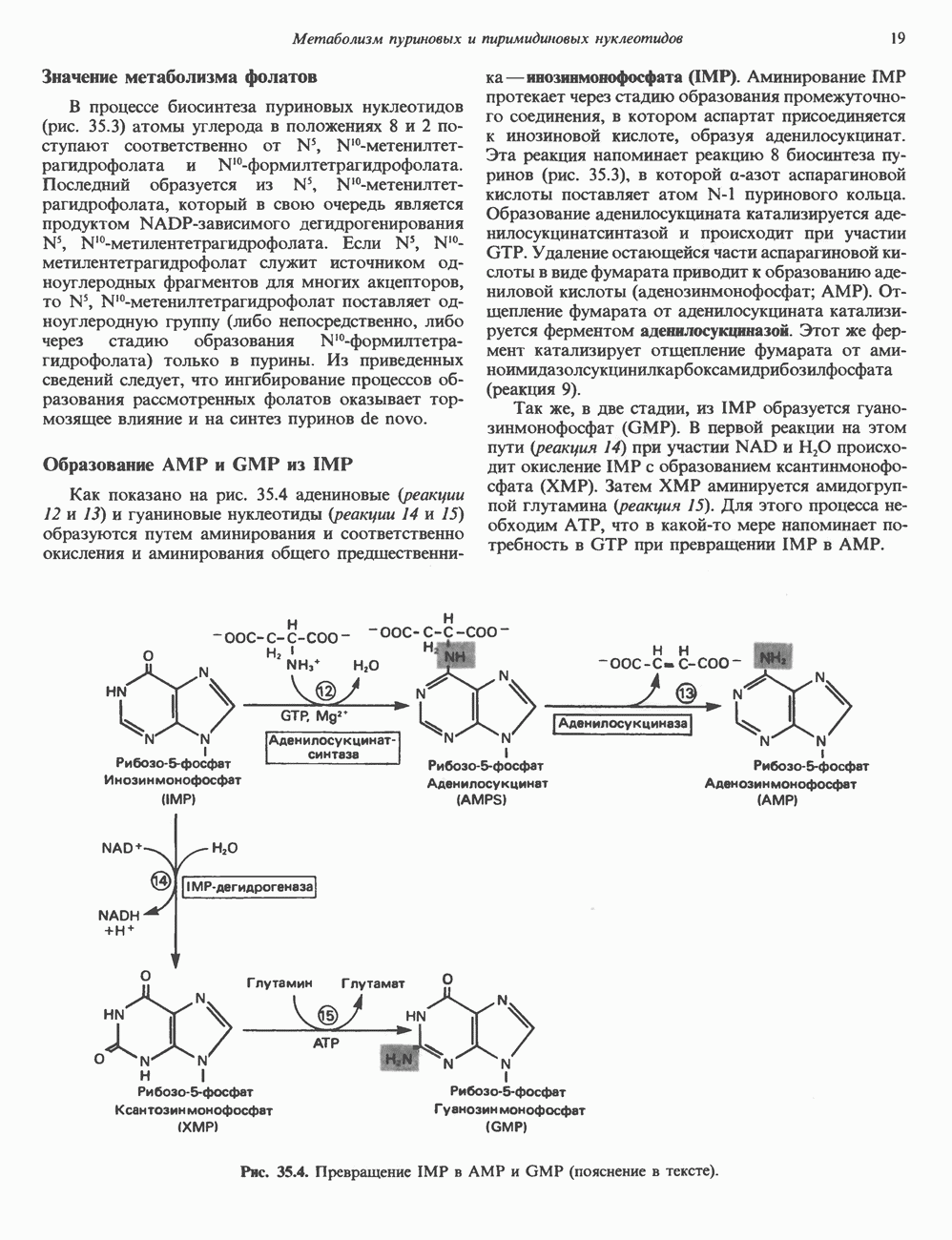

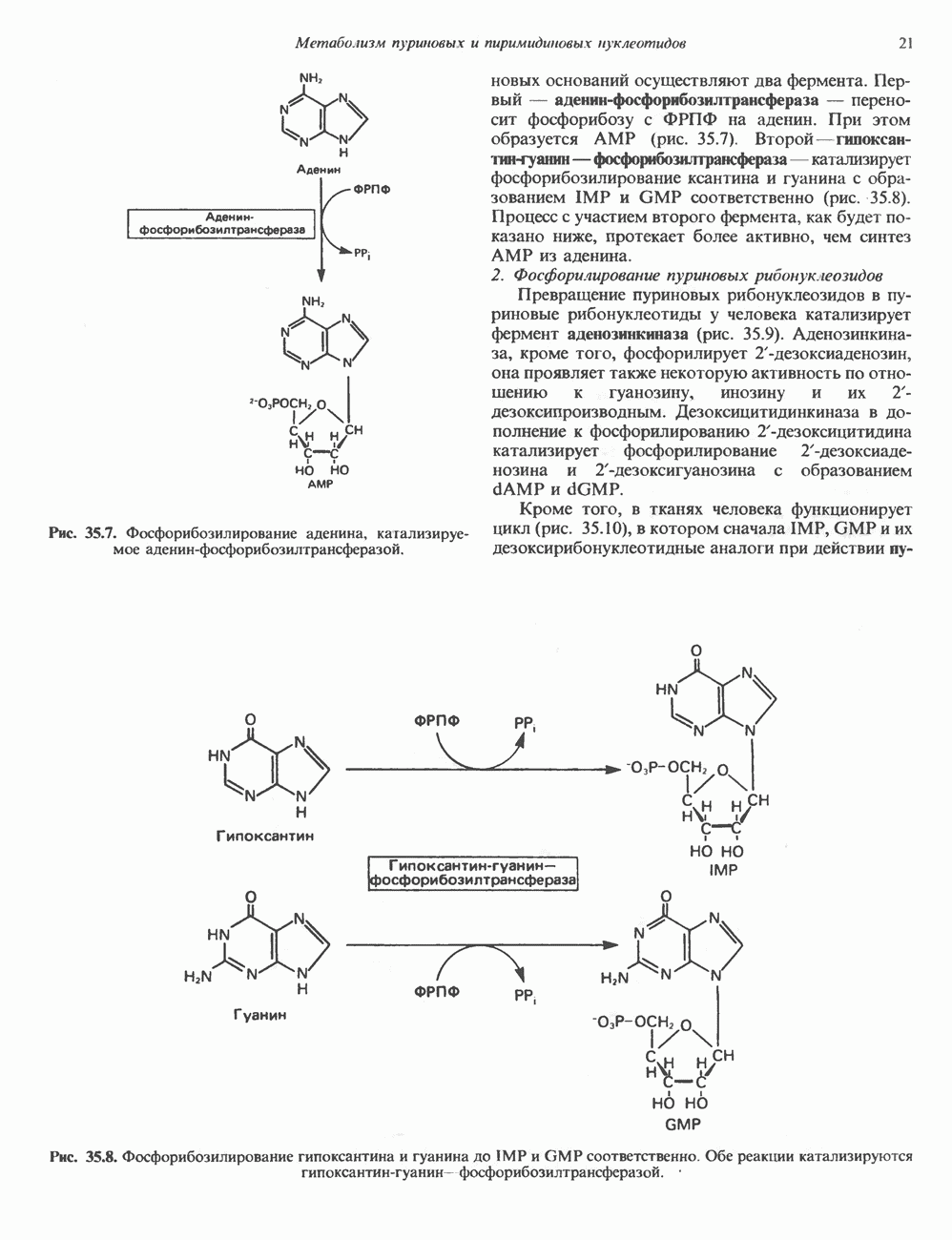

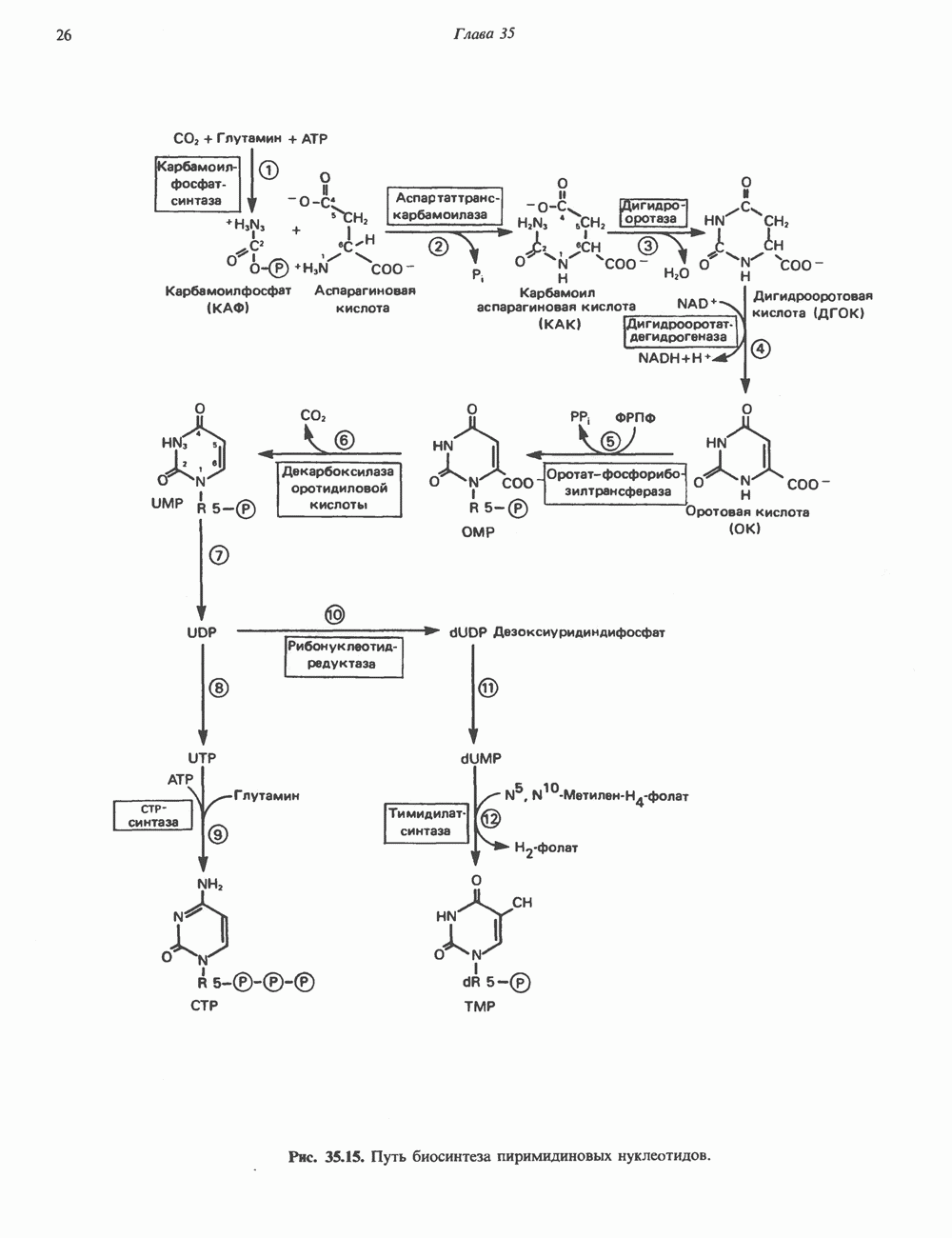







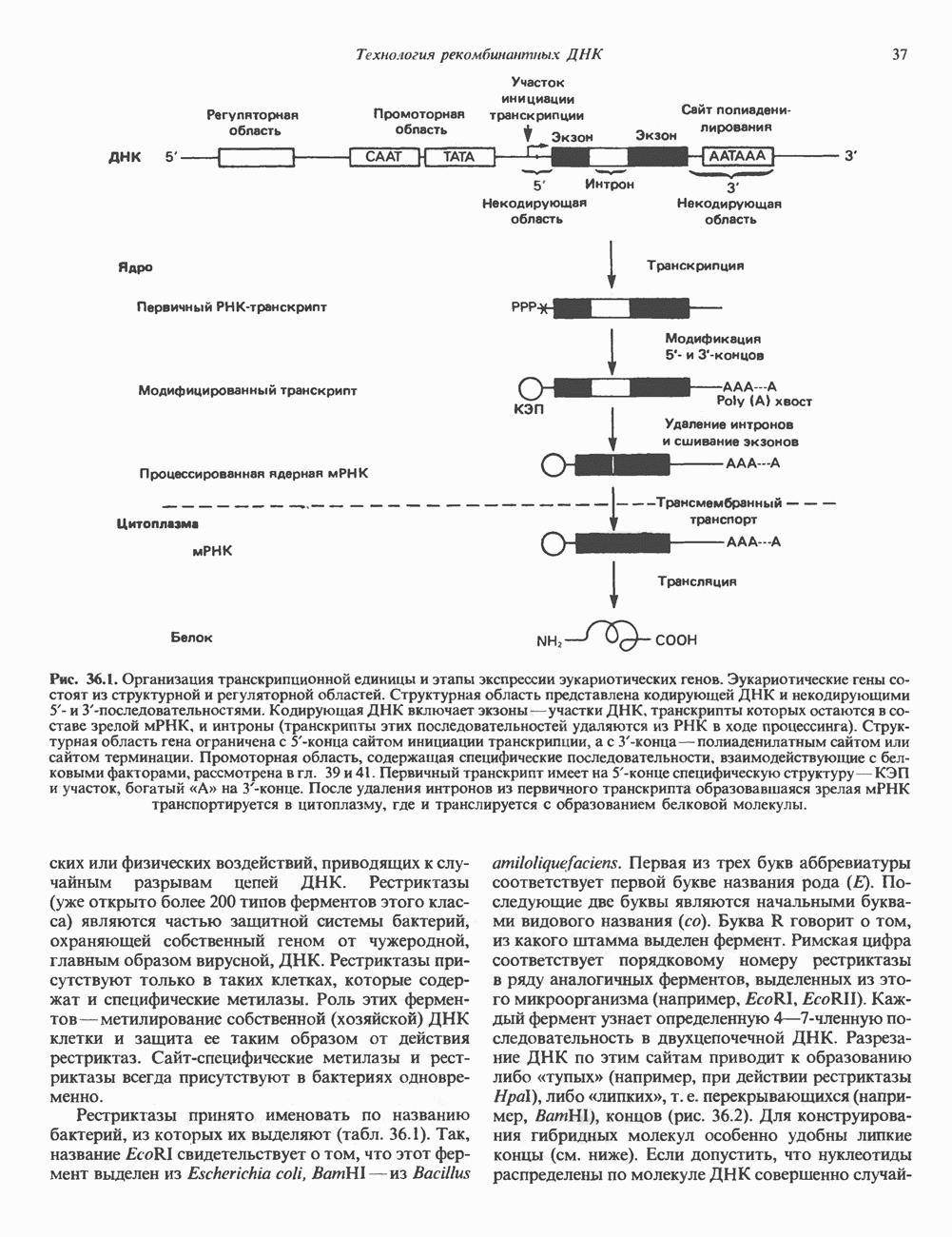












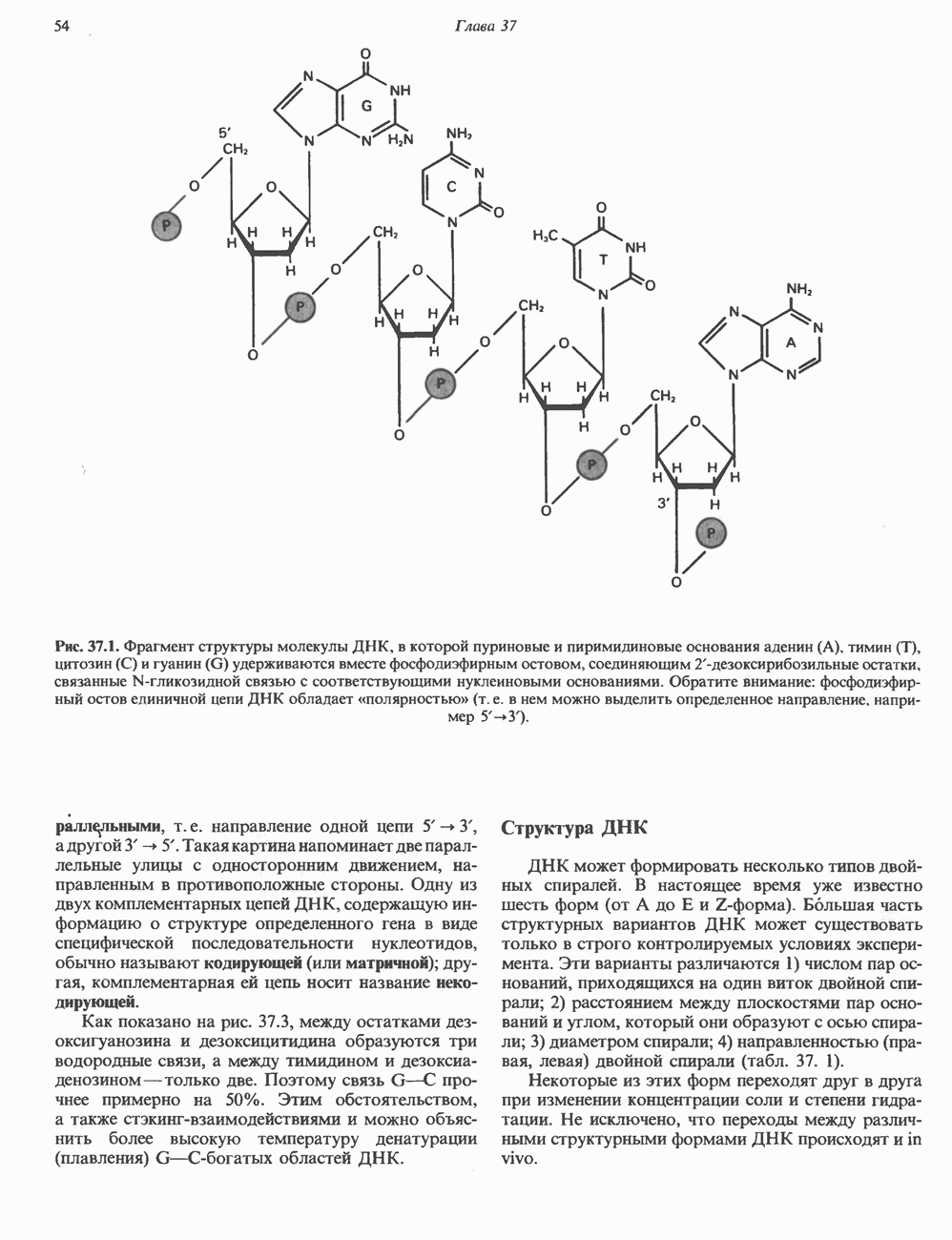
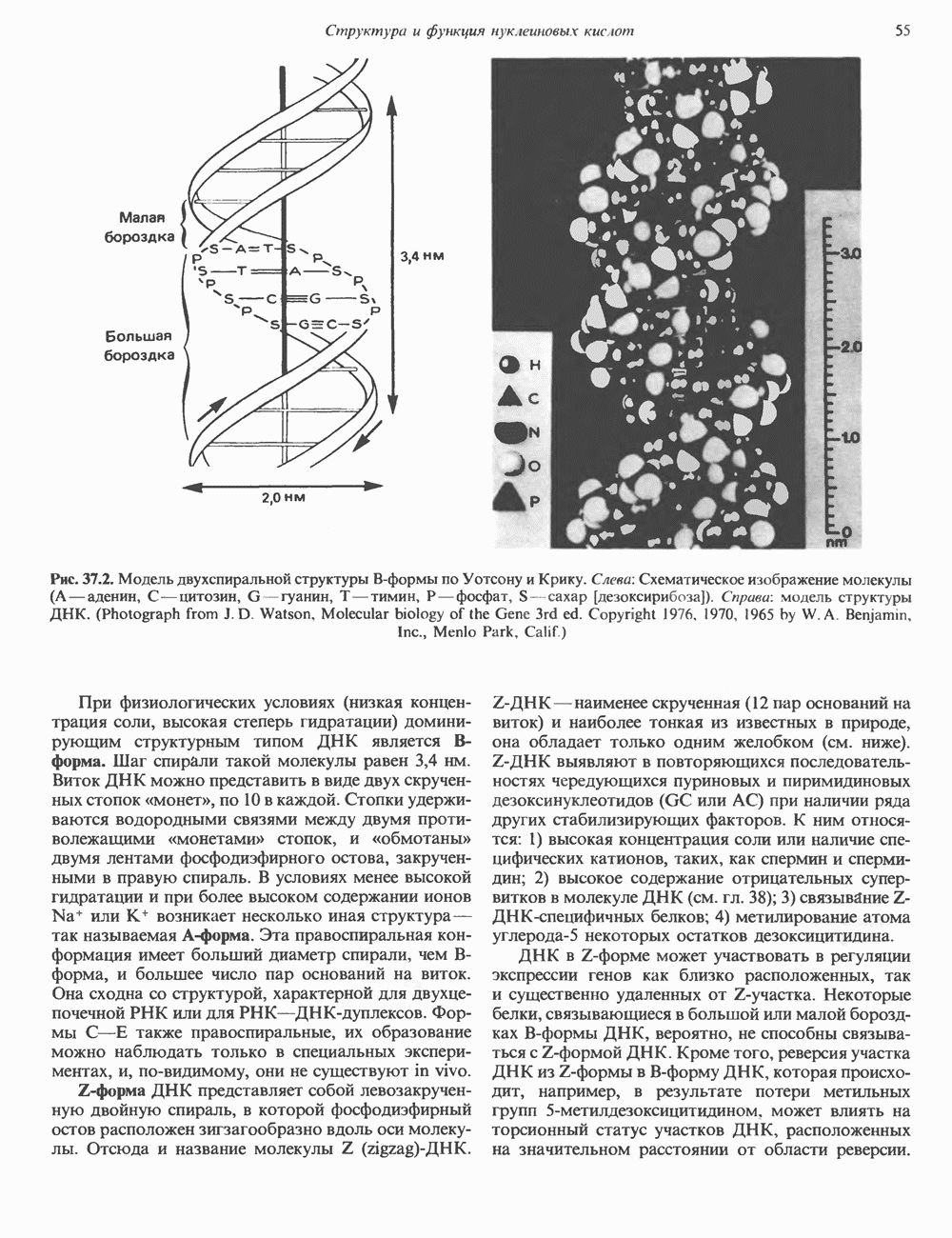
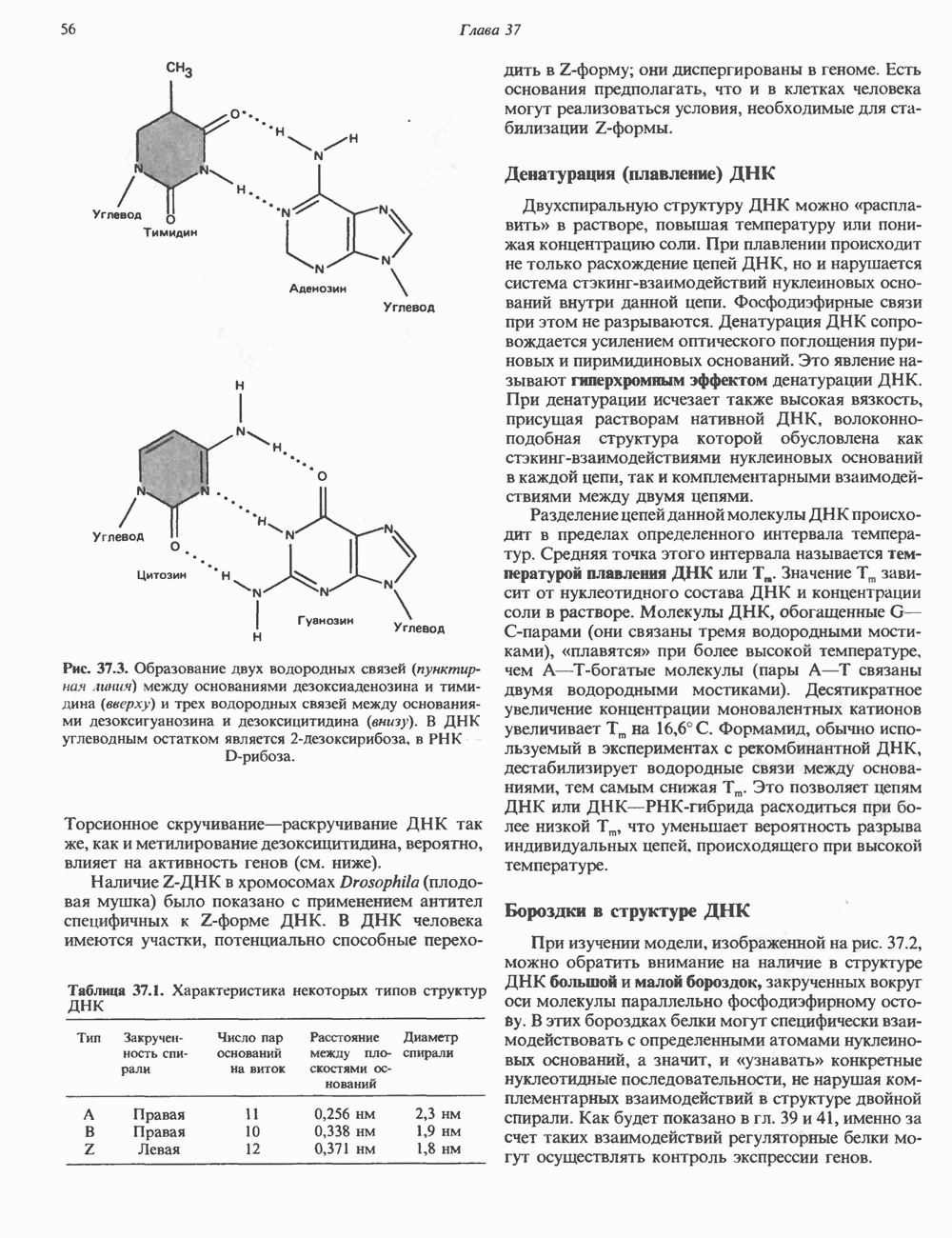

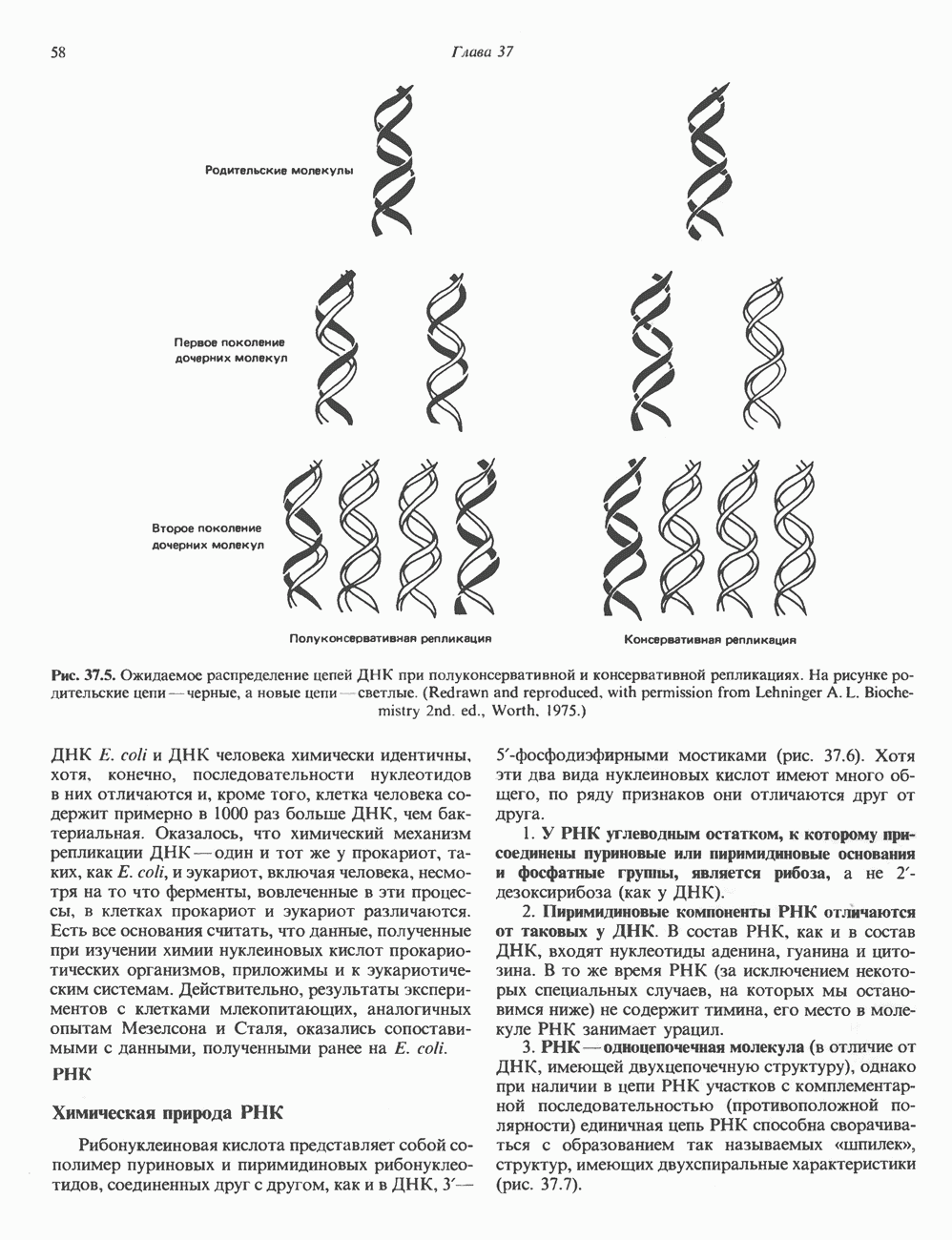
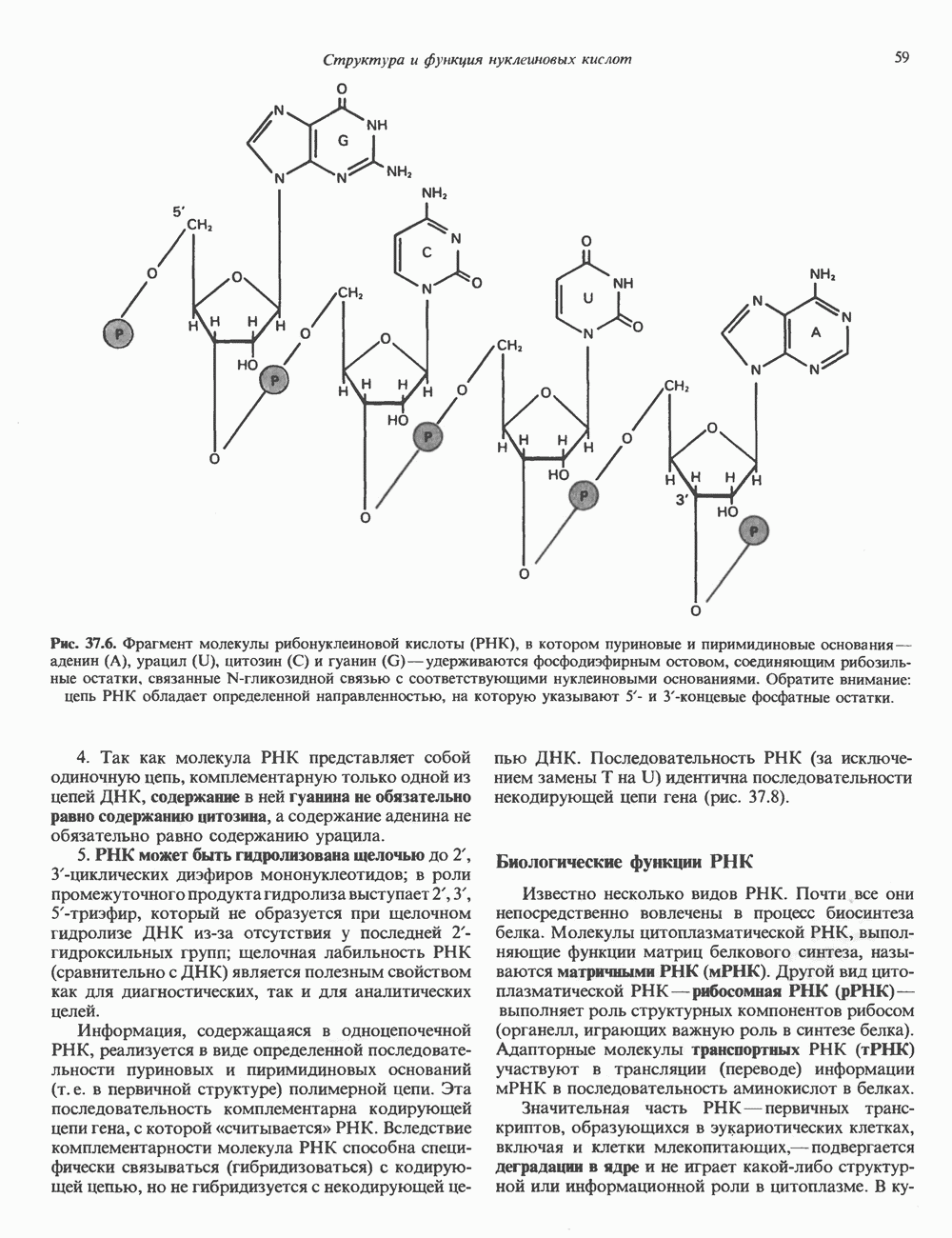

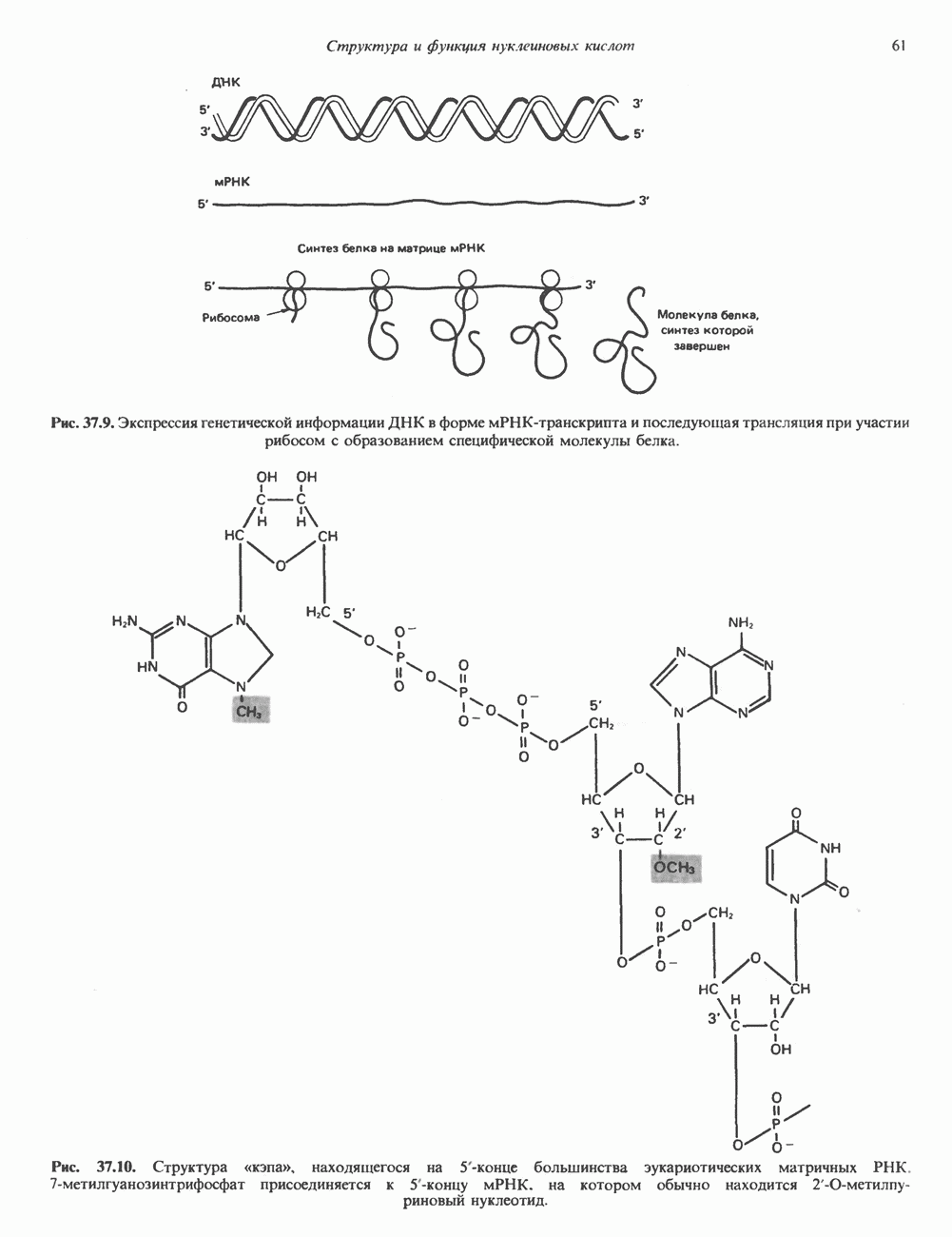

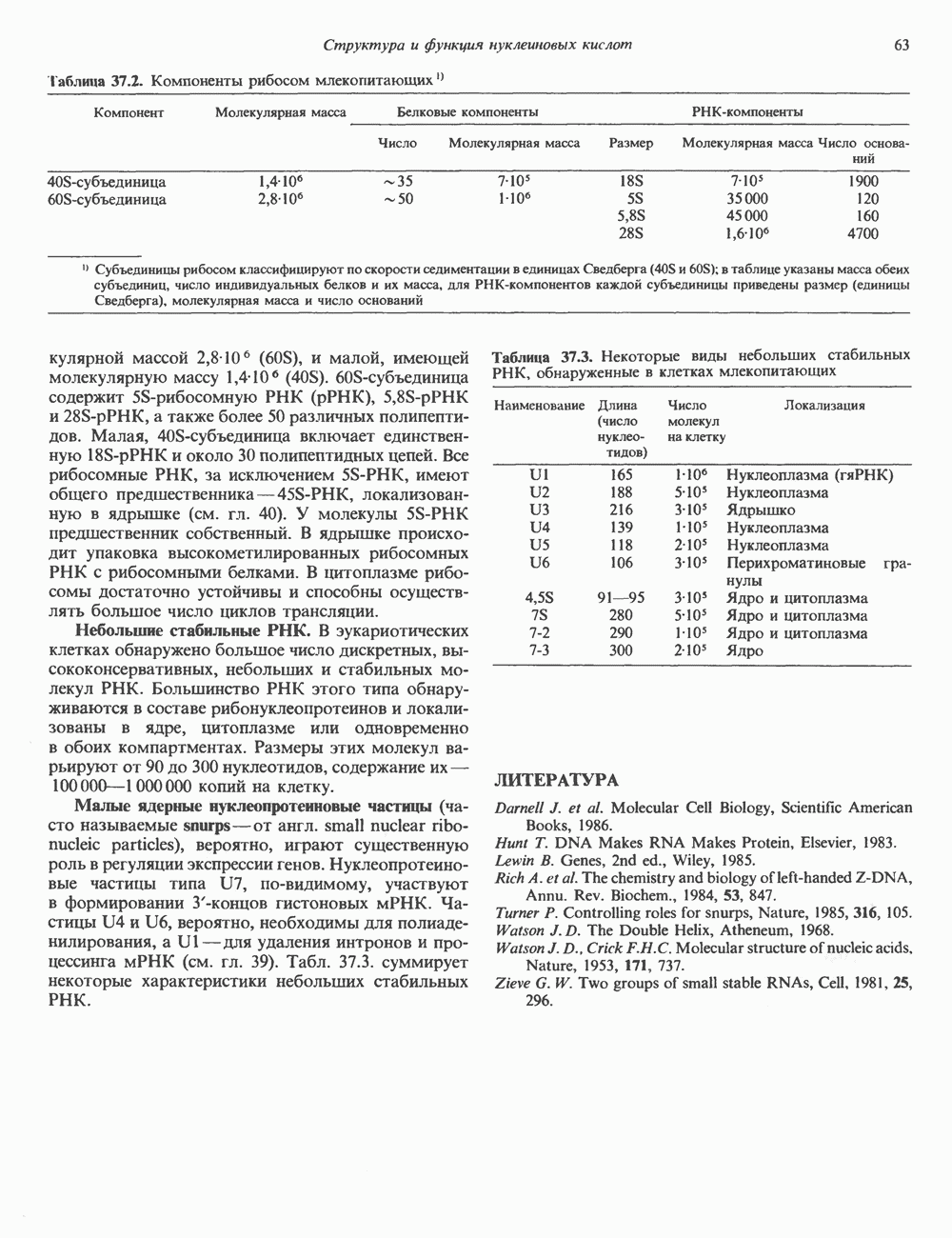


























































































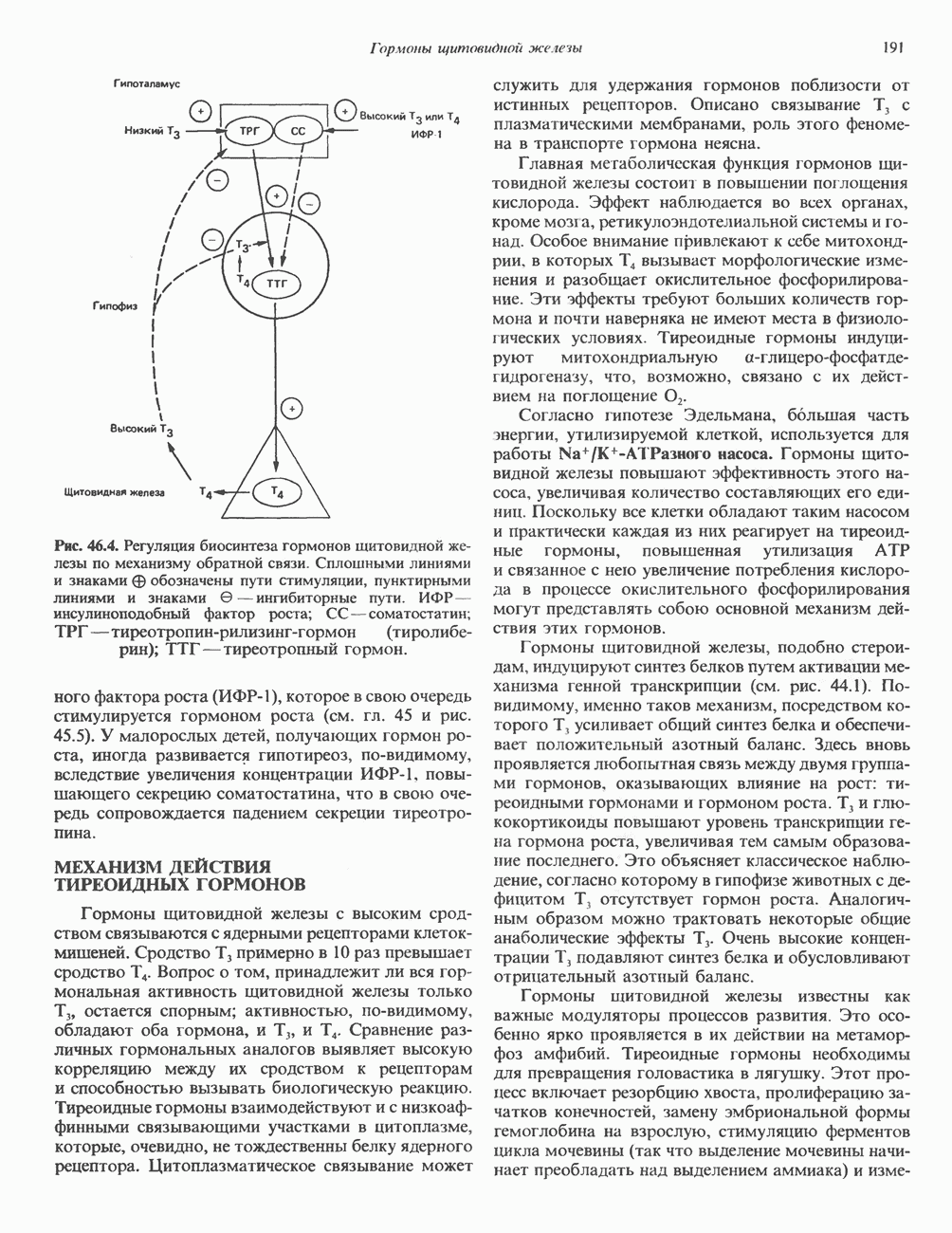




































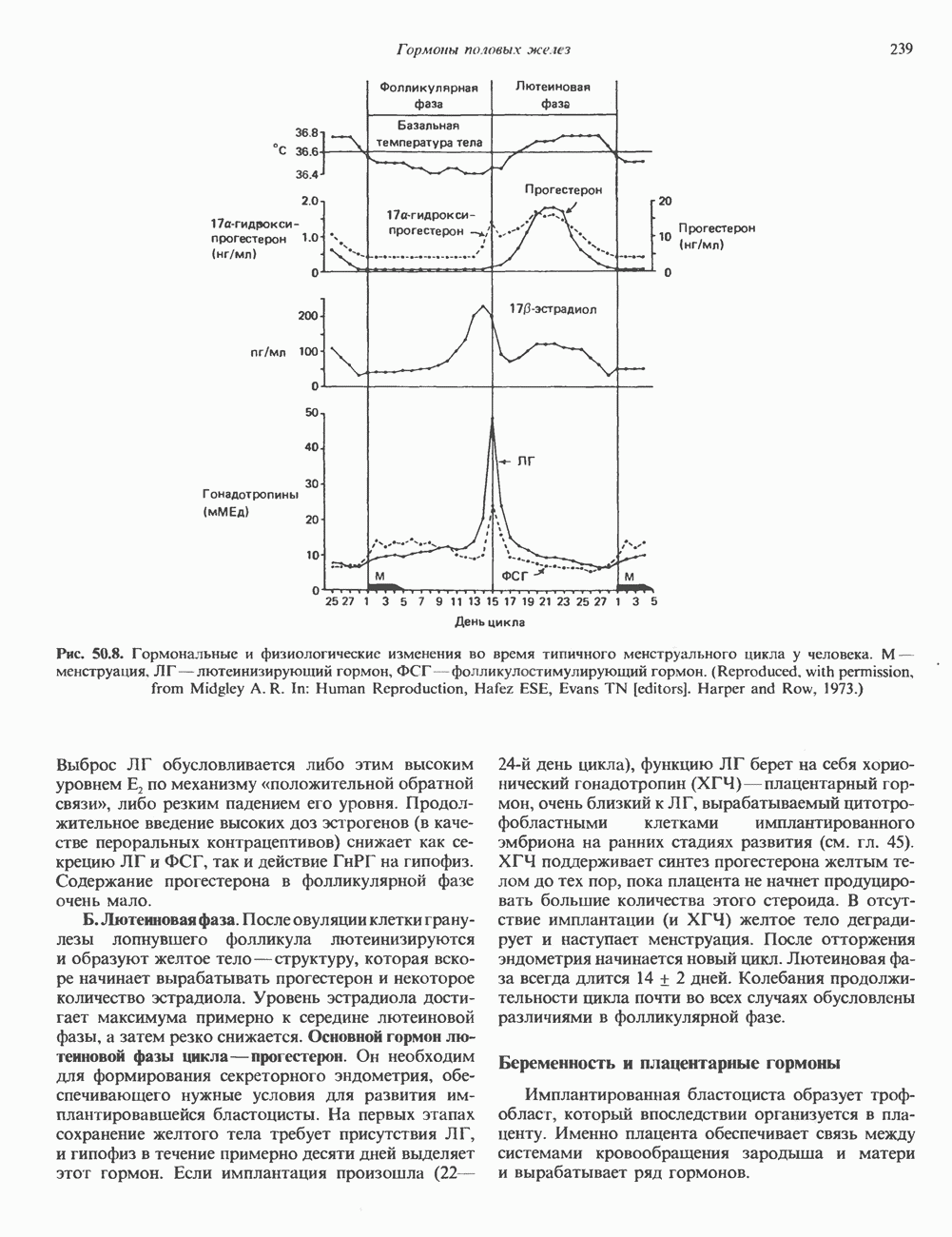









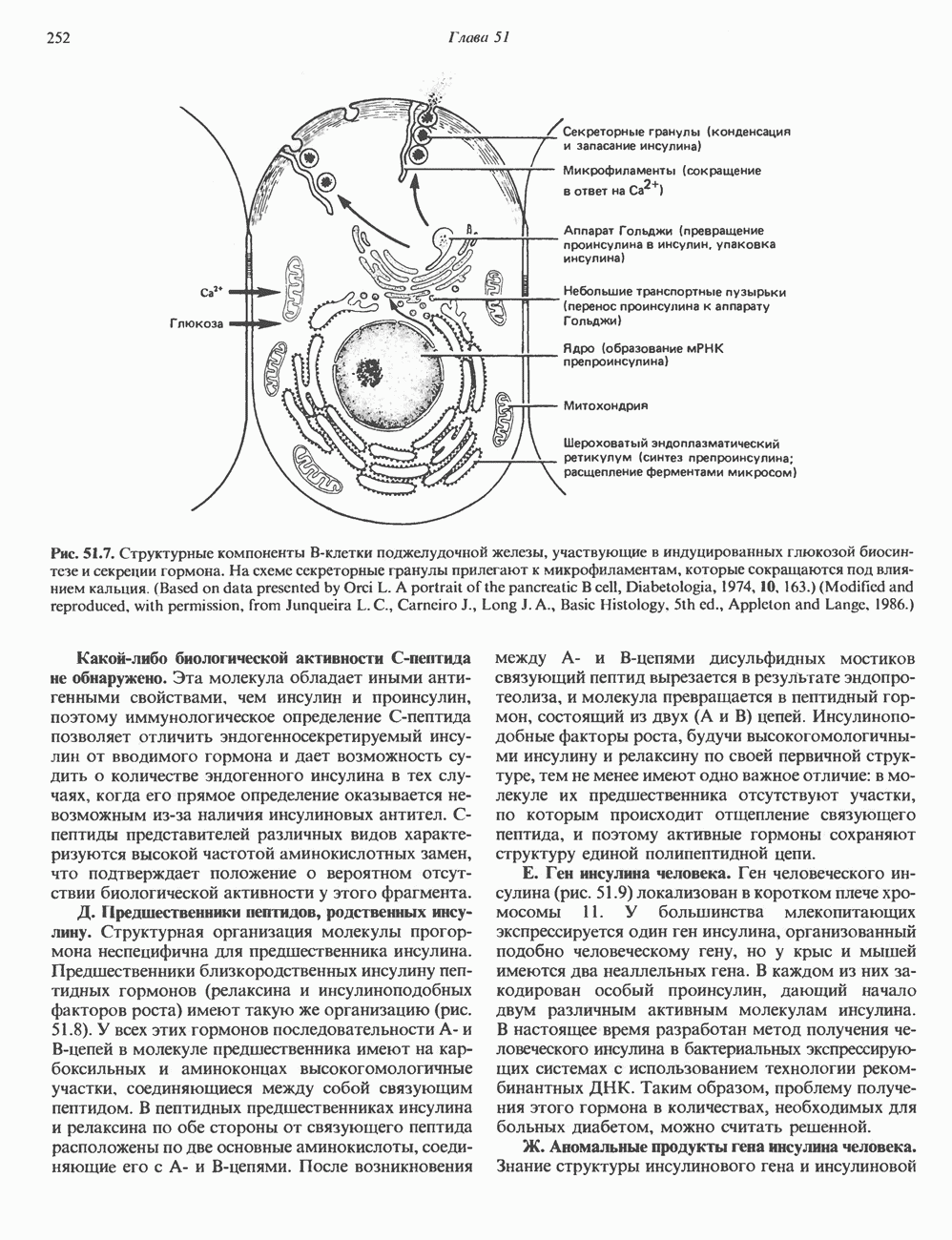















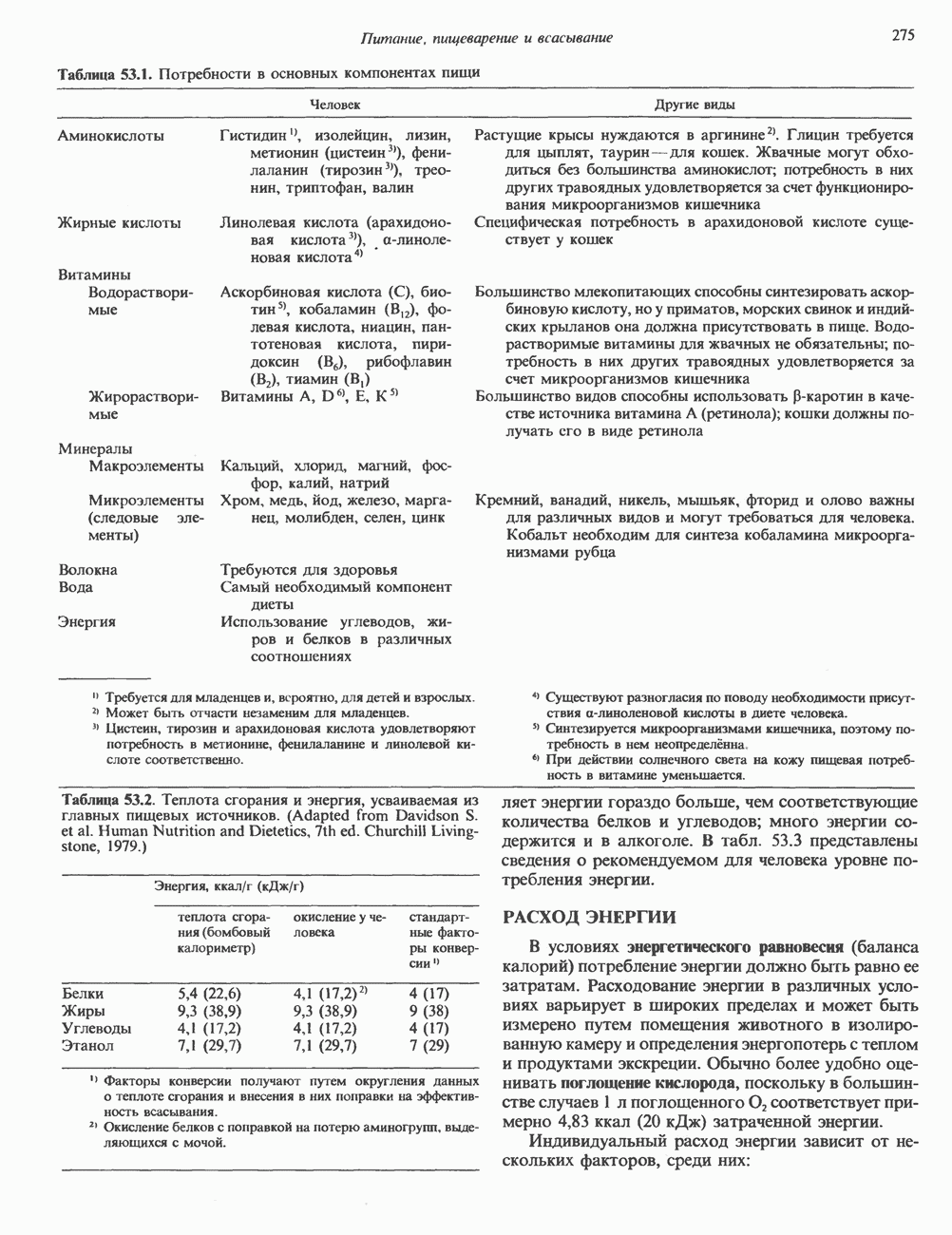





































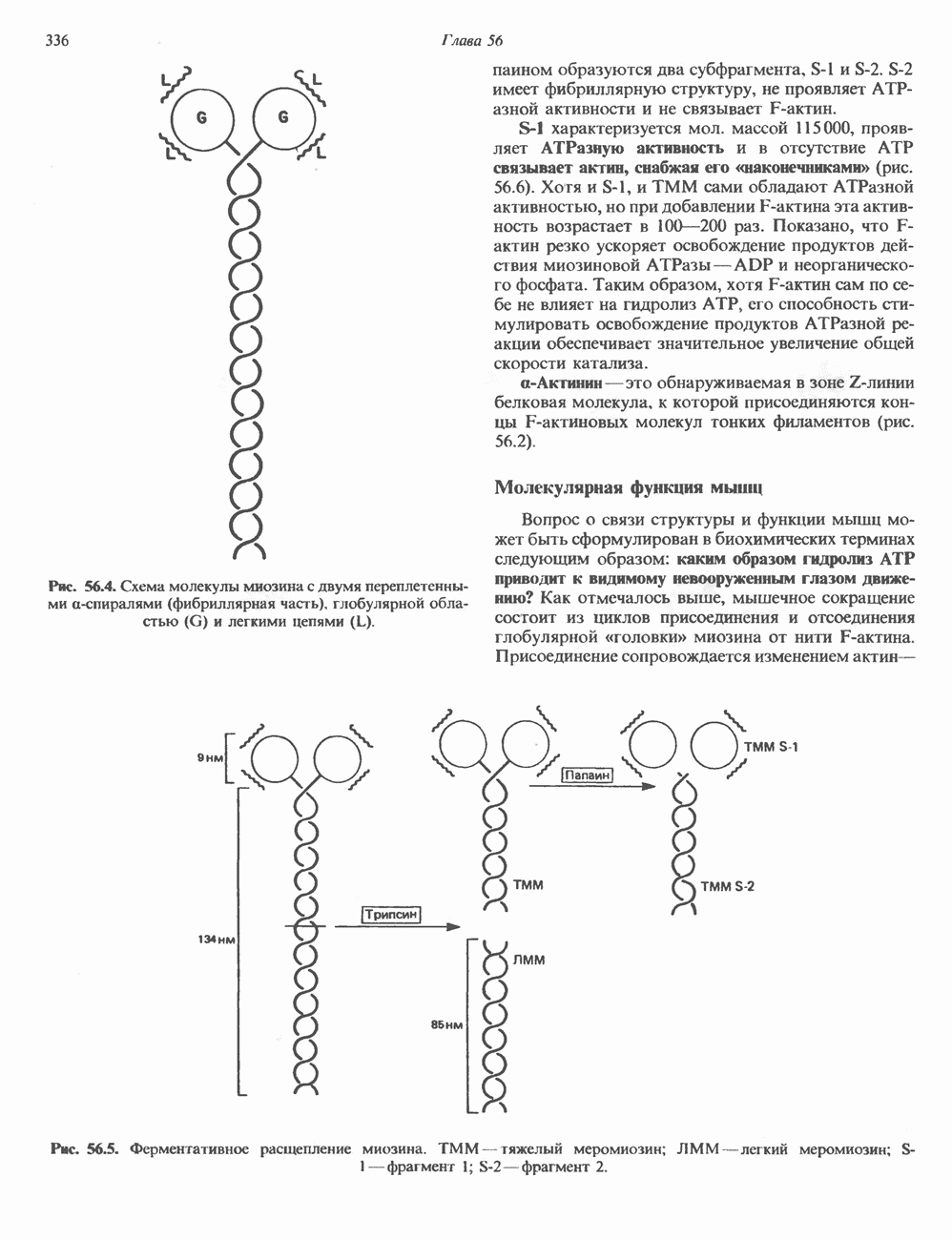


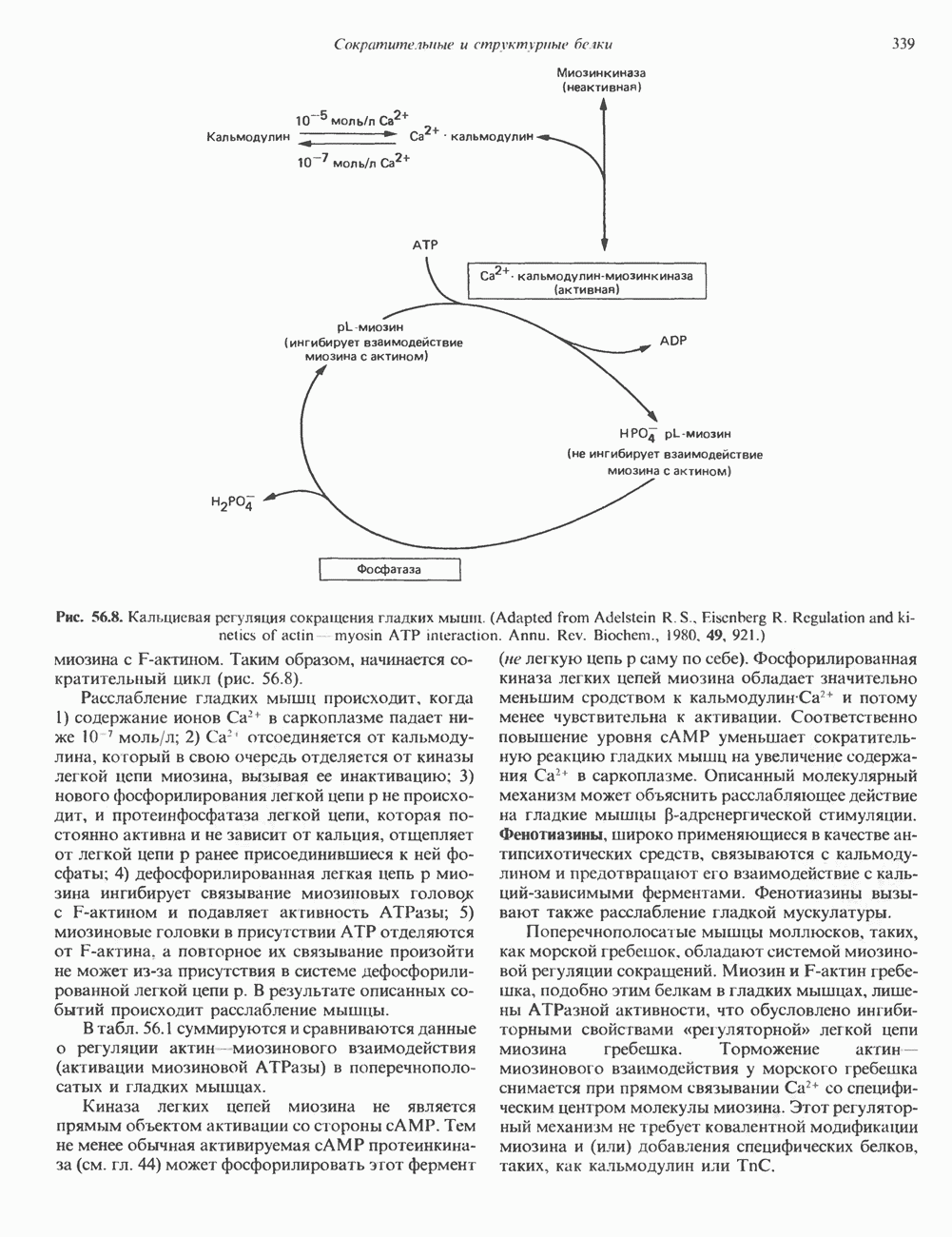



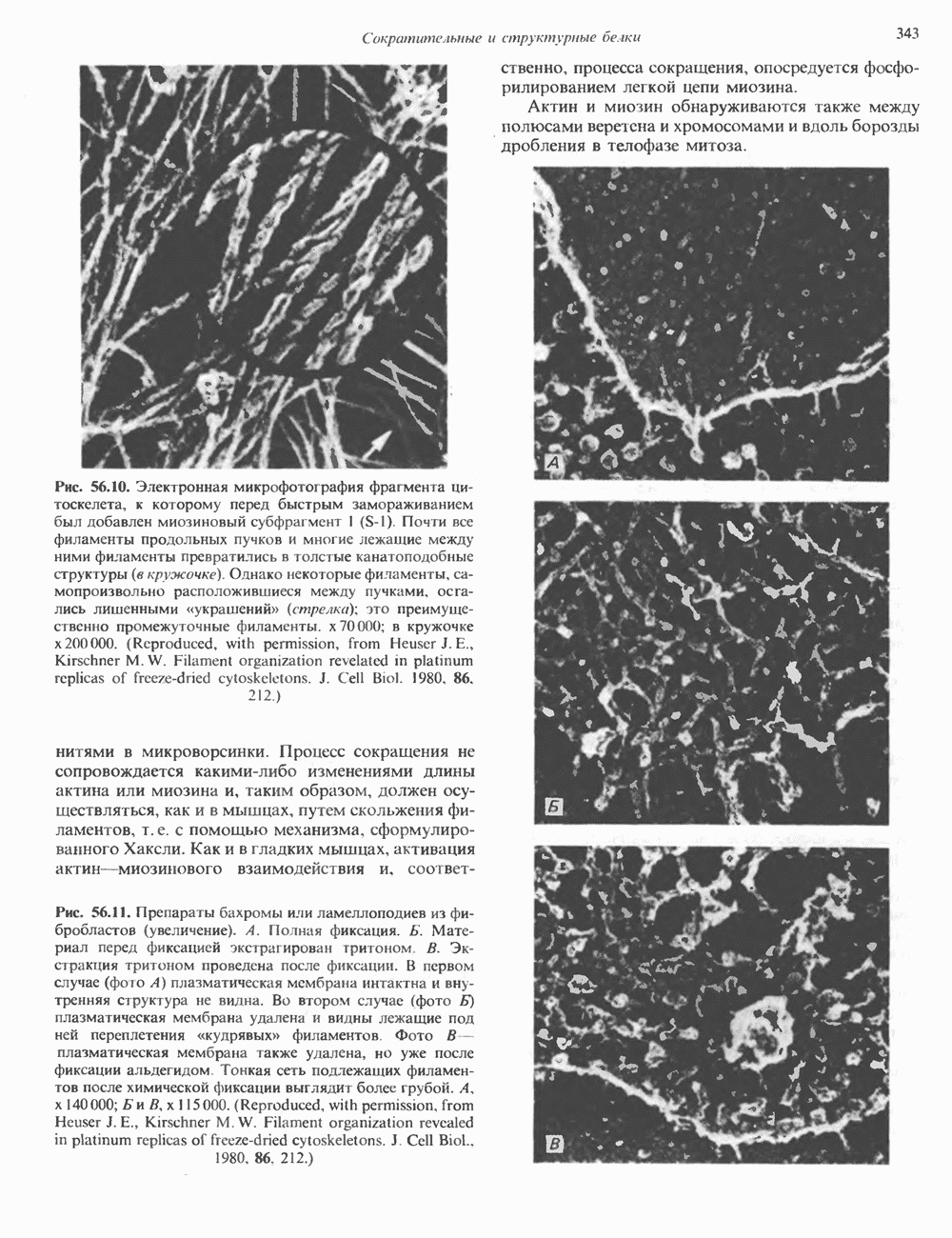
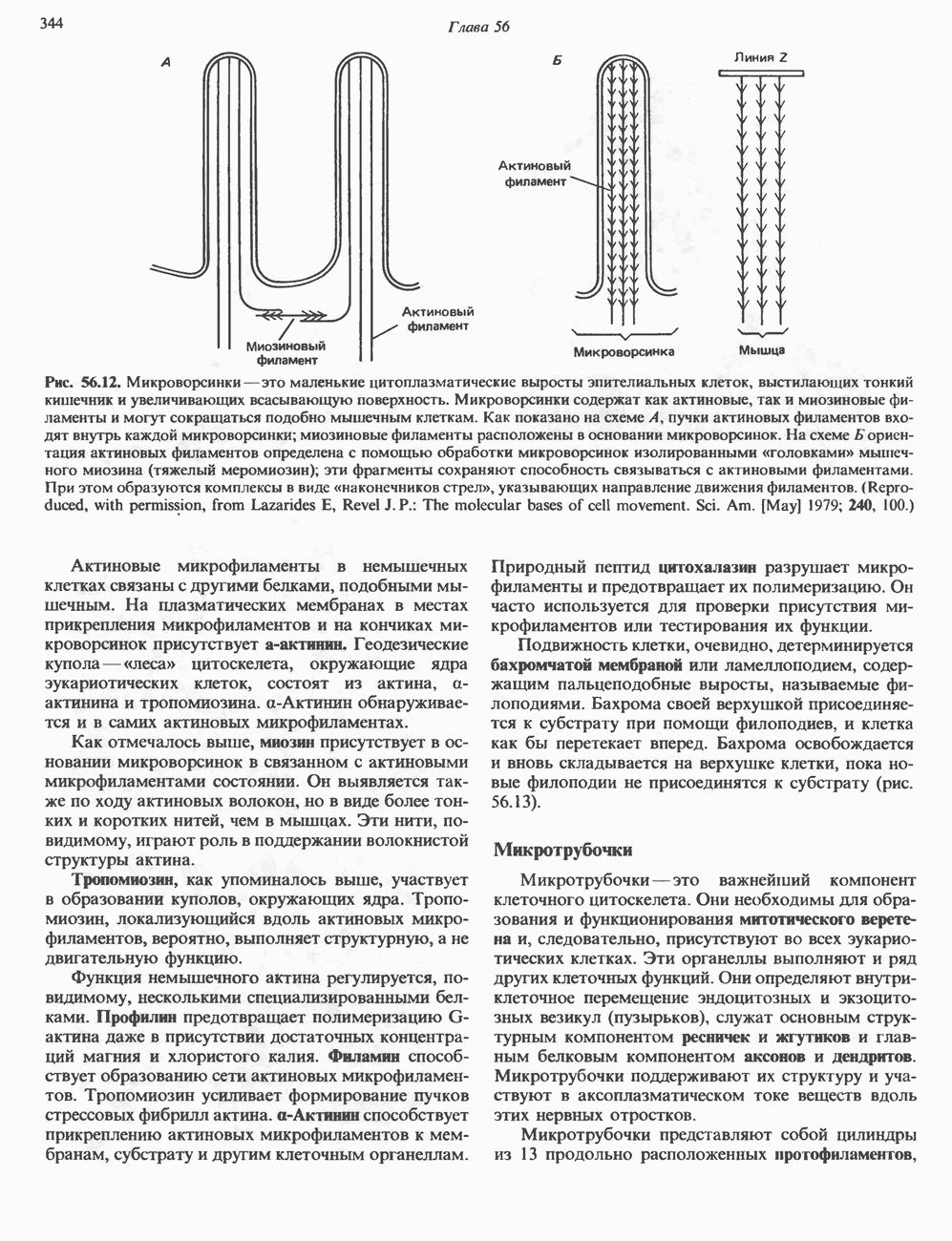






































 и аспартат необходимы для синтеза всех пиримидиновых и пуриновых нуклеотидов. Синтез тимидиновых нуклеотидов, а также всех пуриновых нуждается в присутствии производных тетрагидрофолата. Можно отметить одно существенное различие в путях биосинтеза пуриновых и пиримидиновых нуклеотидов. В первом случае синтез начинается с молекулы рибозофосфата как интегральной части будущей молекулы предшественника нуклеотида, во втором случае сначала синтезируется пиримидиновое основание и только на последних стадиях присоединяется остаток рибозофосфата.
и аспартат необходимы для синтеза всех пиримидиновых и пуриновых нуклеотидов. Синтез тимидиновых нуклеотидов, а также всех пуриновых нуждается в присутствии производных тетрагидрофолата. Можно отметить одно существенное различие в путях биосинтеза пуриновых и пиримидиновых нуклеотидов. В первом случае синтез начинается с молекулы рибозофосфата как интегральной части будущей молекулы предшественника нуклеотида, во втором случае сначала синтезируется пиримидиновое основание и только на последних стадиях присоединяется остаток рибозофосфата.
 в реакции, катализируемой в цитозоле карбамоилфосфатсинтазой (реакция 1). Отметим, что карбамоилфосфатсинтаза, ответственная за ранние стадии синтеза мочевины, локализована в митохондриях.
в реакции, катализируемой в цитозоле карбамоилфосфатсинтазой (реакция 1). Отметим, что карбамоилфосфатсинтаза, ответственная за ранние стадии синтеза мочевины, локализована в митохондриях.
 и образуется кольцевая структура (реакция 3).
и образуется кольцевая структура (реакция 3).
 -дезоксинуклеозиддифосфатов (реакция 10) также аналогичен тому, который описан для пуриновых нуклеозидцифосфатов (рис. 35.6 и 35.13).
-дезоксинуклеозиддифосфатов (реакция 10) также аналогичен тому, который описан для пуриновых нуклеозидцифосфатов (рис. 35.6 и 35.13).
 как донор метильной группы. Метиленовая группа
как донор метильной группы. Метиленовая группа  в ходе реакции восстанавливается до метильной и присоединяется к атому
в ходе реакции восстанавливается до метильной и присоединяется к атому  Процесс сопровождается окислением тетрагидрофолатного переносчика до дигидрофолата. Можно считать, что в результа
Процесс сопровождается окислением тетрагидрофолатного переносчика до дигидрофолата. Можно считать, что в результа  метилирования dUMP с образованием ТМР происходит полное восстановление гидроксиметильной группы серина (переносимой на тетрагидрофолат при образовании
метилирования dUMP с образованием ТМР происходит полное восстановление гидроксиметильной группы серина (переносимой на тетрагидрофолат при образовании  до
до
 -дезоксирибонуклеозиды тимидин и дезоксицитидин путем превращения их в соответствующие нуклеотиды (рис. 35.16). 2-Дезоксицитидин фосфорилируется дезоксицитидин-киназой—ферментом, способным также фосфорилировать дезоксигуанозин и дезоксиаденозин.
-дезоксирибонуклеозиды тимидин и дезоксицитидин путем превращения их в соответствующие нуклеотиды (рис. 35.16). 2-Дезоксицитидин фосфорилируется дезоксицитидин-киназой—ферментом, способным также фосфорилировать дезоксигуанозин и дезоксиаденозин.
 -гидроксипиразо-лопиримидин) до нуклеотидного производного, в котором рибозилфосфат присоединен к атому
-гидроксипиразо-лопиримидин) до нуклеотидного производного, в котором рибозилфосфат присоединен к атому  пиримидинового кольца этого лекарственного соединения. Противоопухолевый препарат
пиримидинового кольца этого лекарственного соединения. Противоопухолевый препарат  -фторурацил также фосфорибозилируется этим ферментом.
-фторурацил также фосфорибозилируется этим ферментом.
 происходящего из уреидного углерода
происходящего из уреидного углерода  пиримидинового кольца, представляет собой главный путь катаболизма урацила, цитозина и тимина. Основные конечные продукты катаболизма этих оснований — (З-аланин и (
пиримидинового кольца, представляет собой главный путь катаболизма урацила, цитозина и тимина. Основные конечные продукты катаболизма этих оснований — (З-аланин и ( -аминоизобутират.
-аминоизобутират.



 и отрицательную
и отрицательную  регуляцию по принципу обратной связи. Сокращения расшифрованы на рис. 35.15.
регуляцию по принципу обратной связи. Сокращения расшифрованы на рис. 35.15.