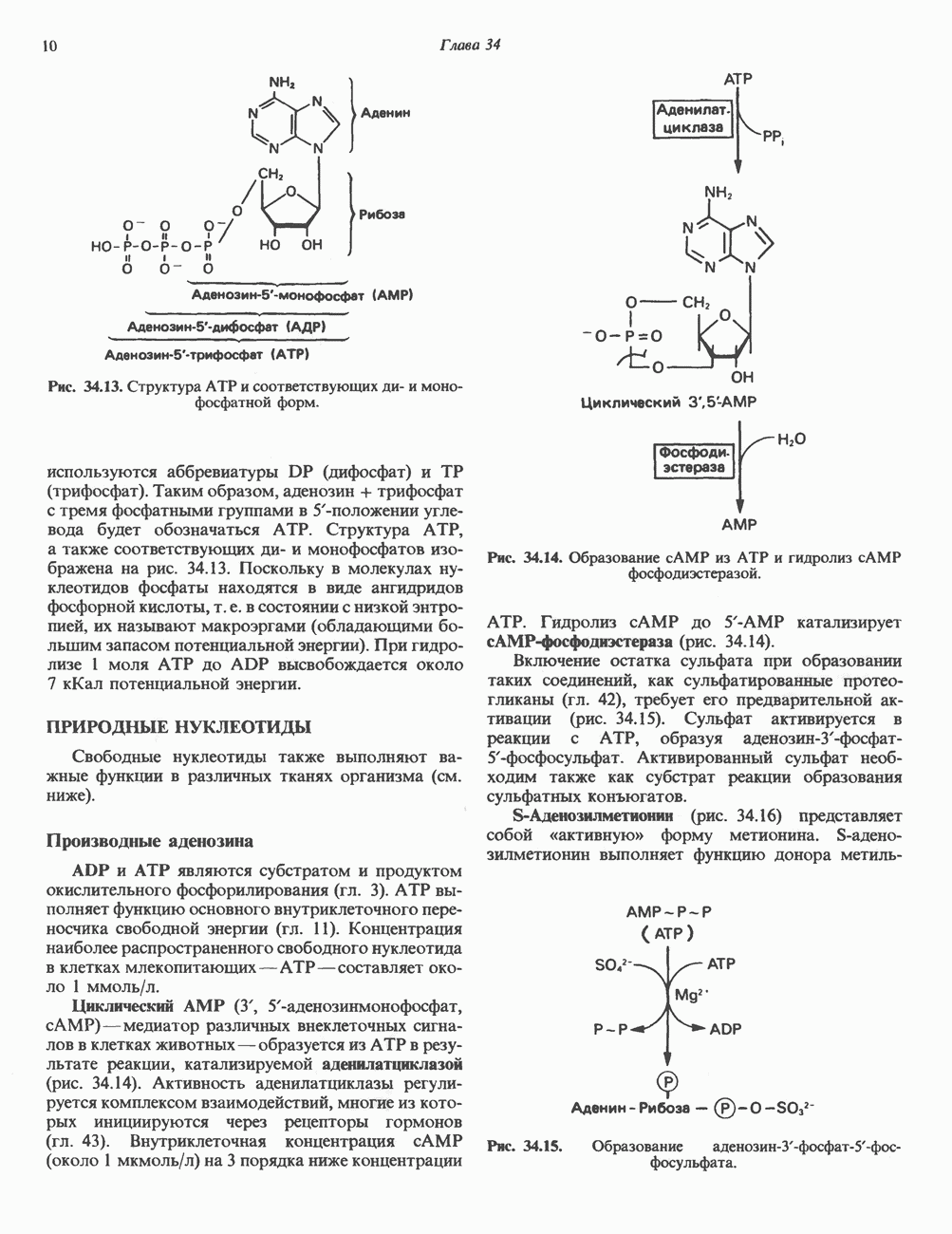
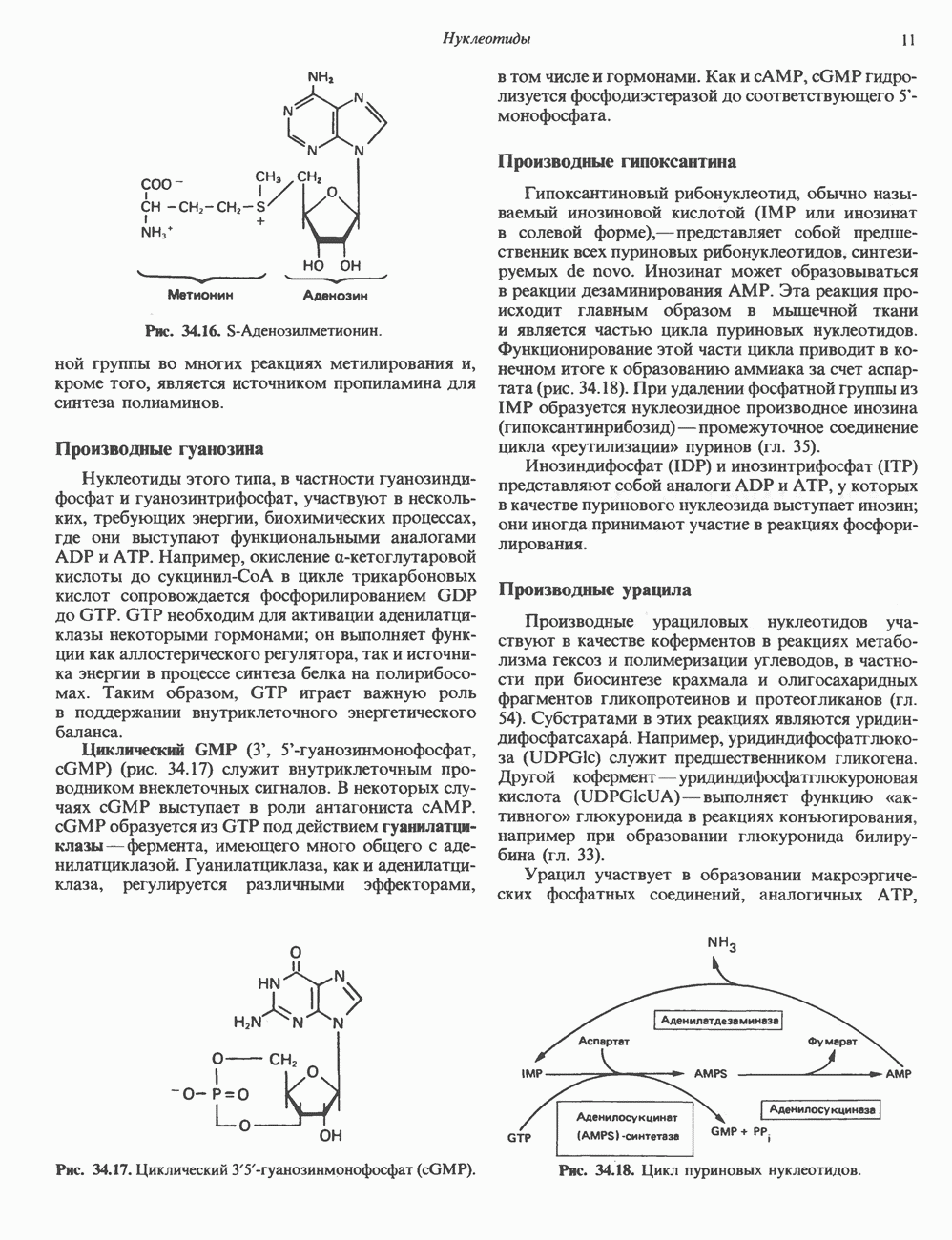
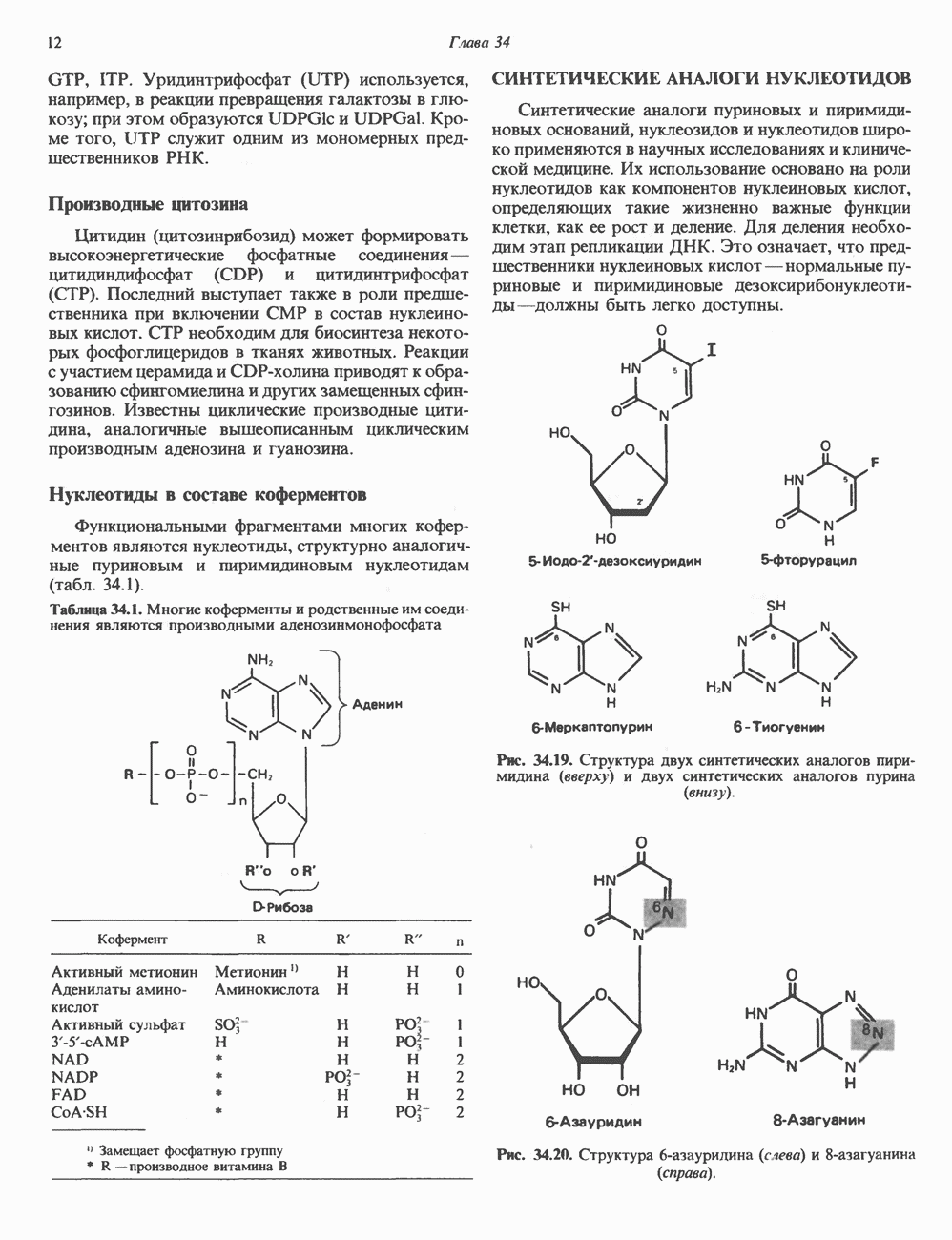
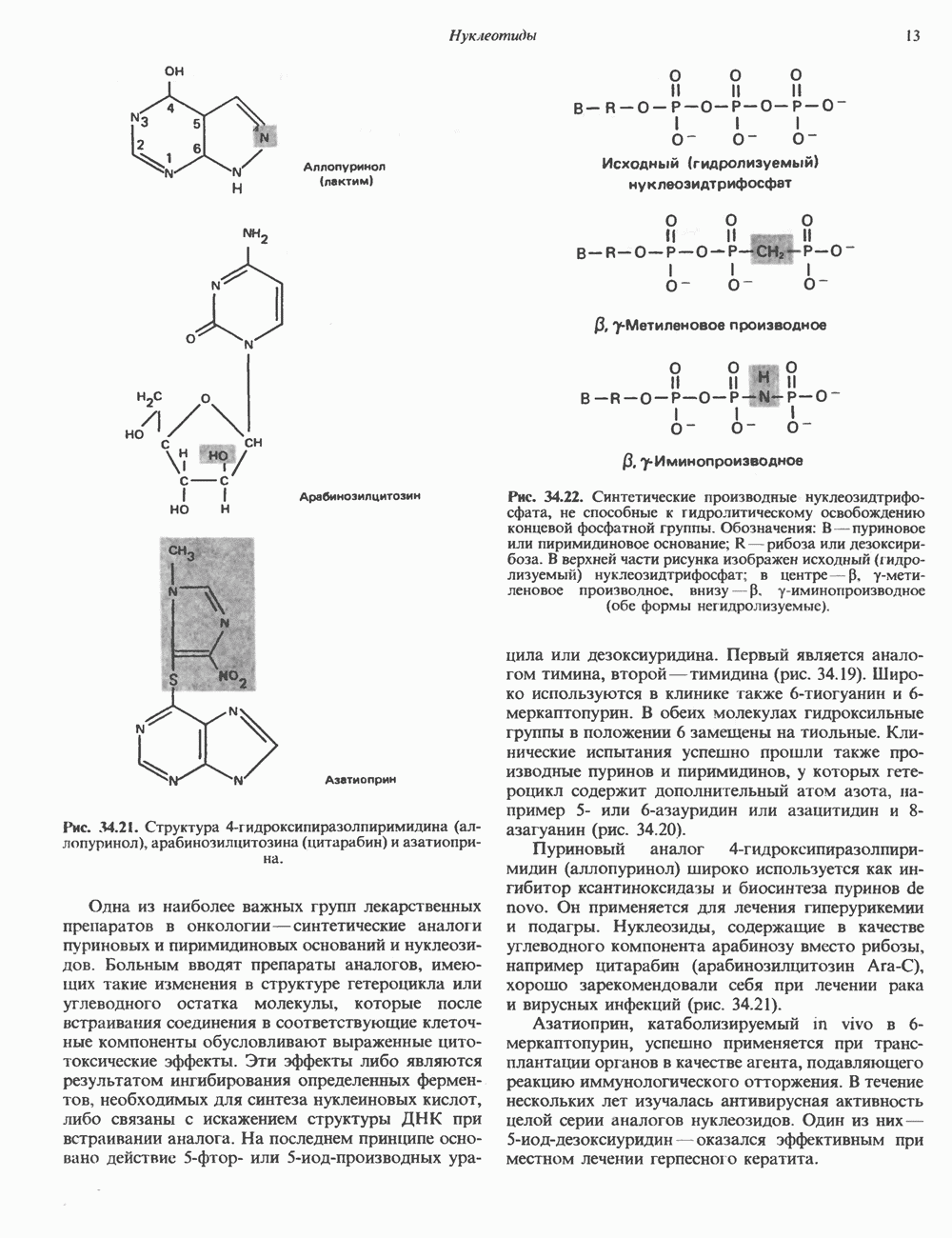
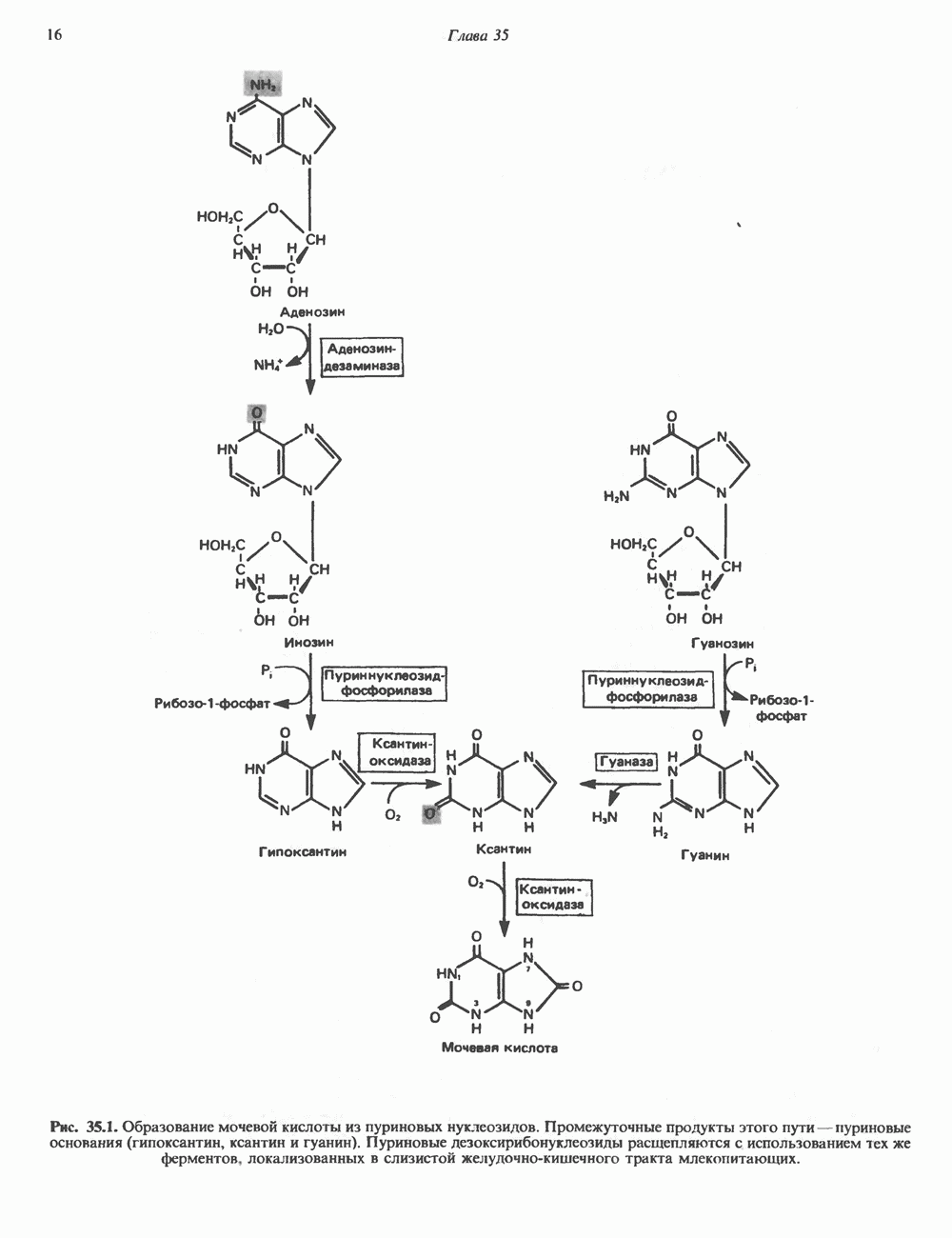
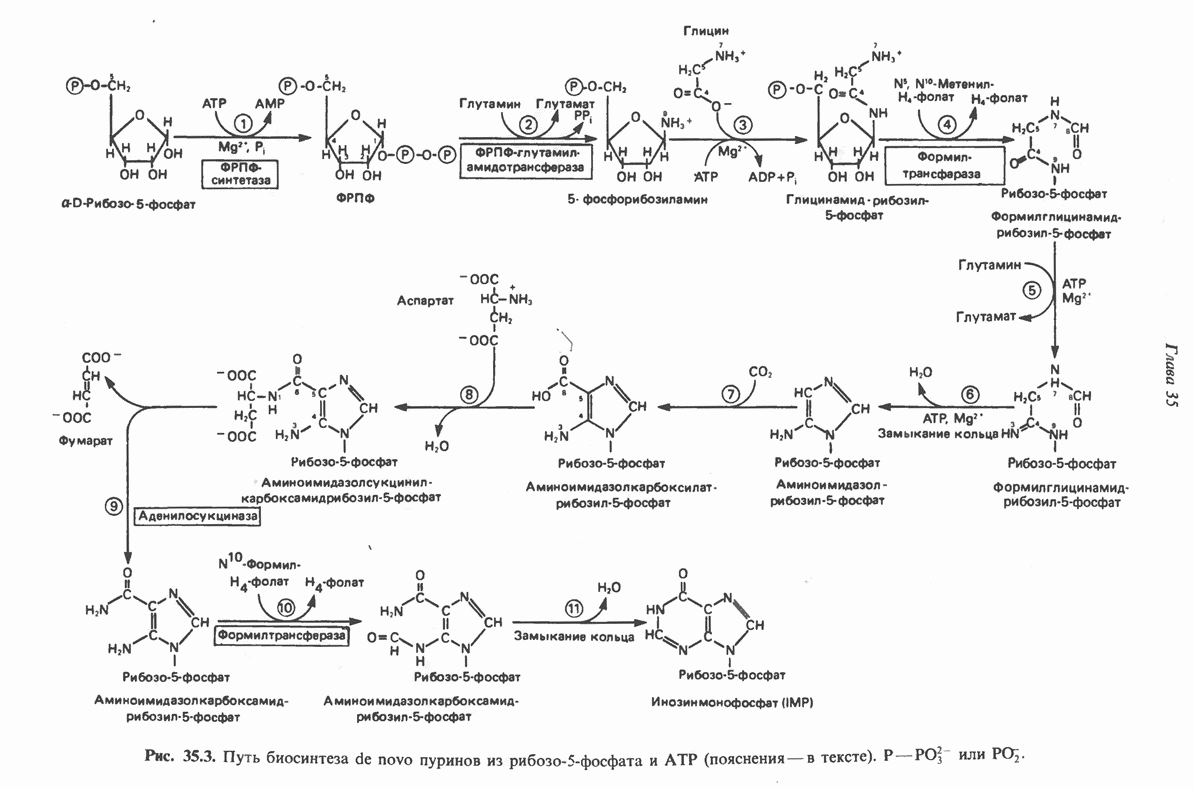
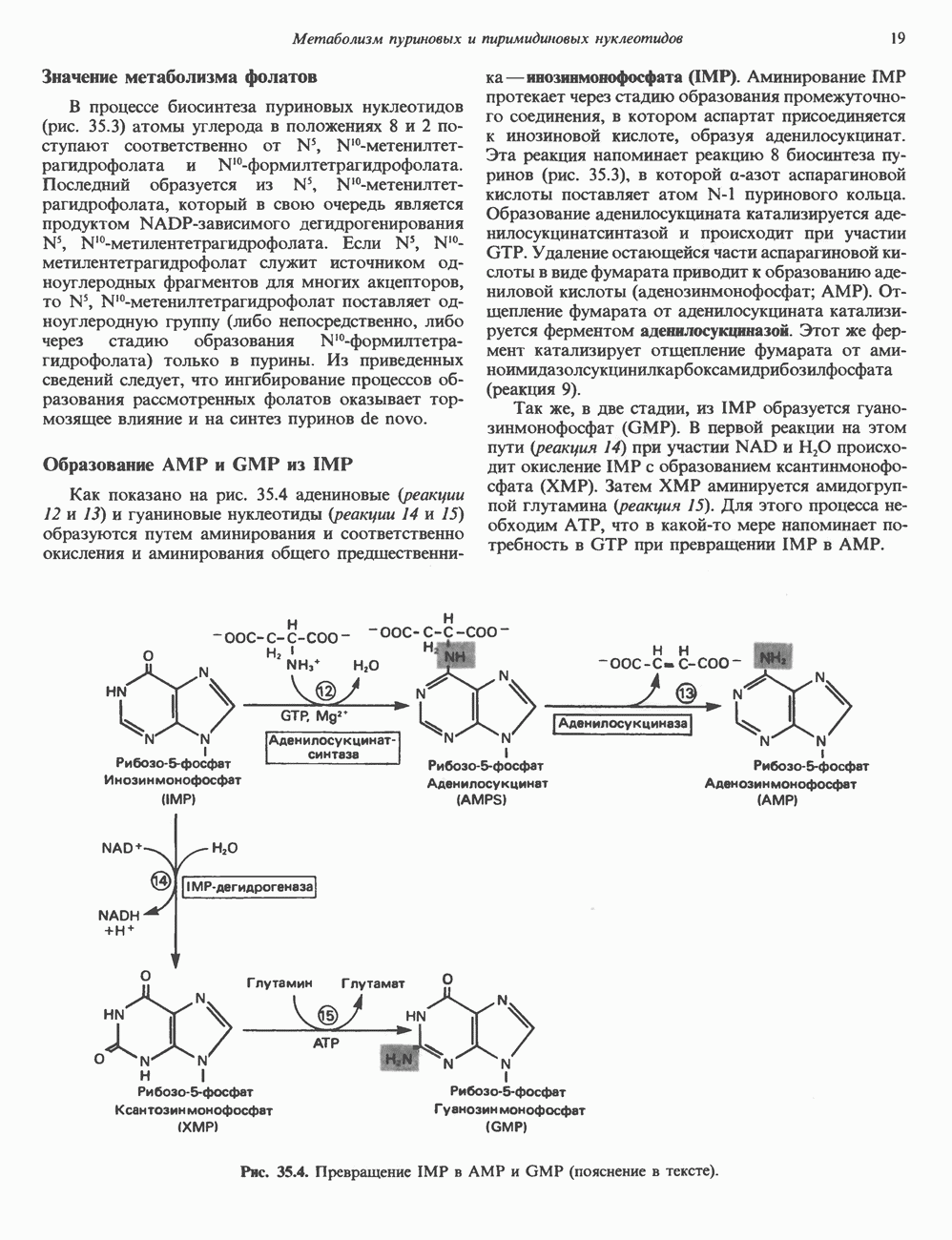
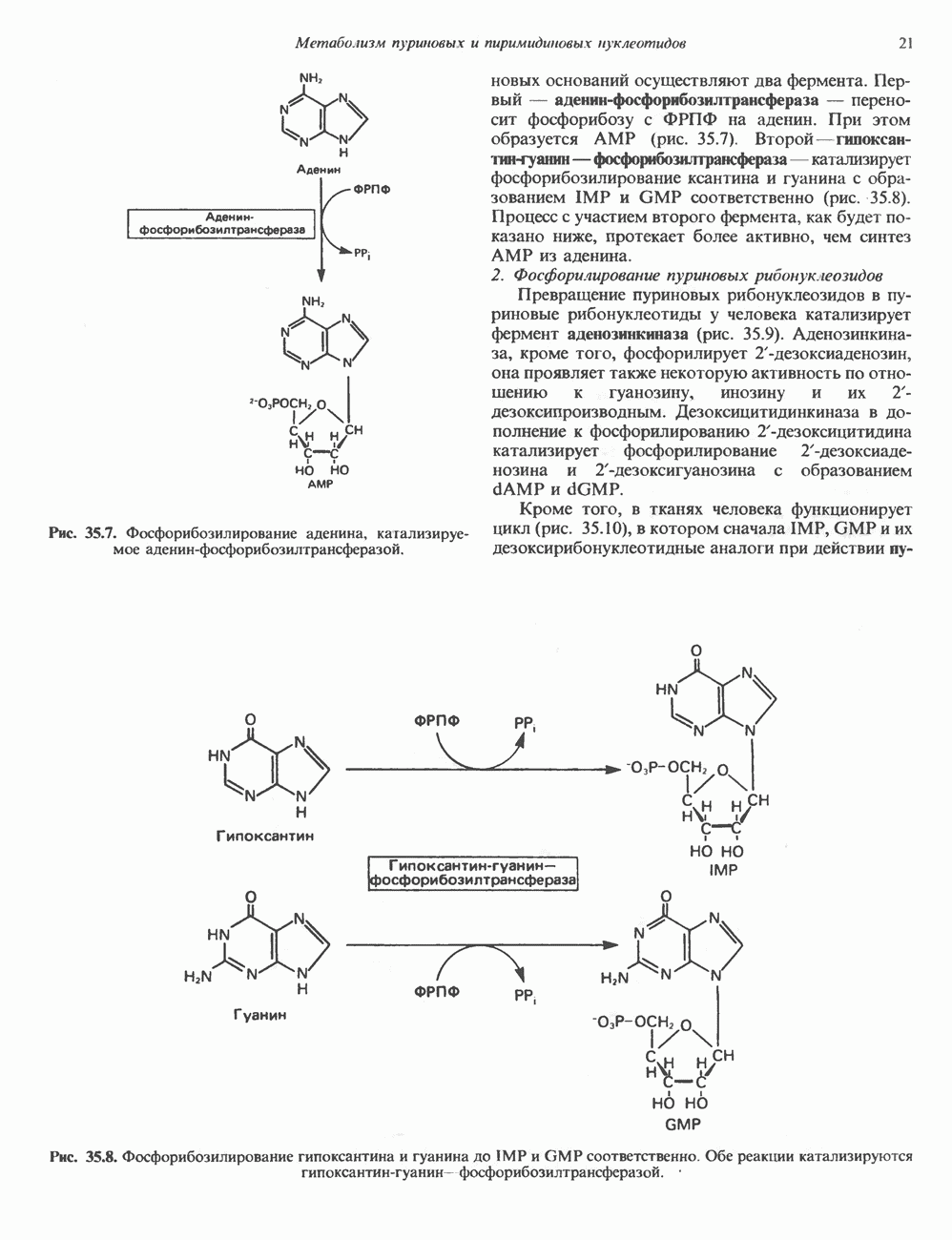
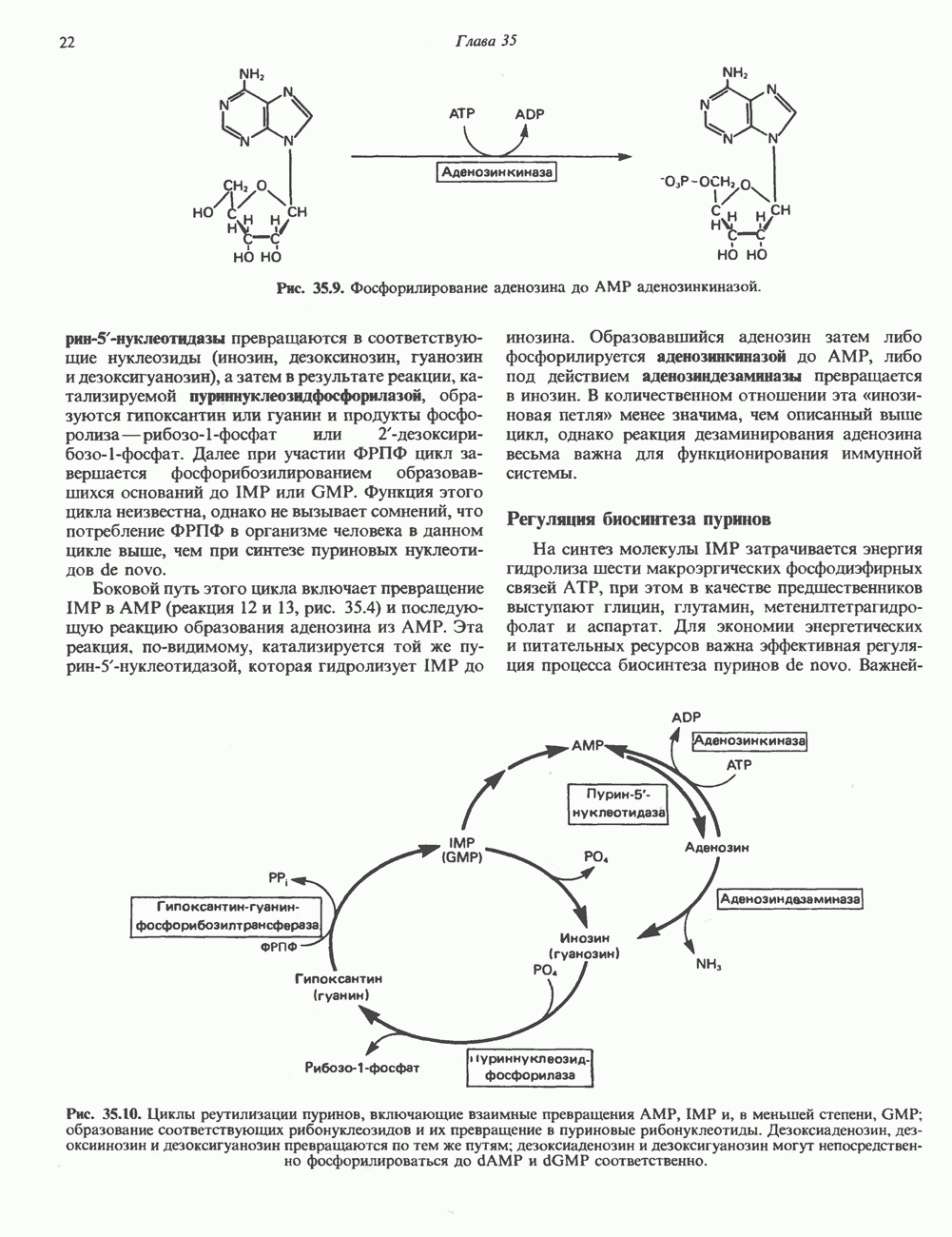
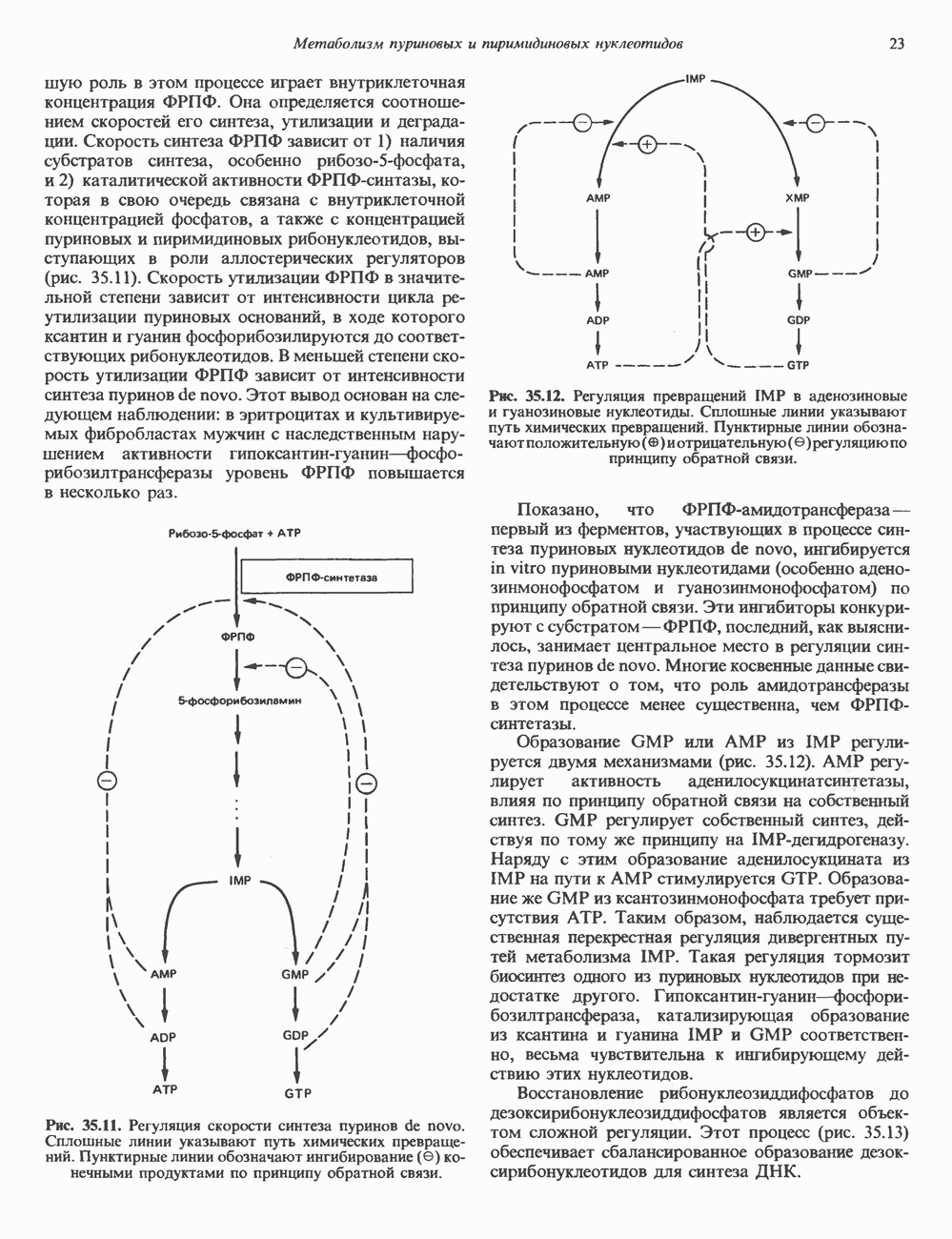
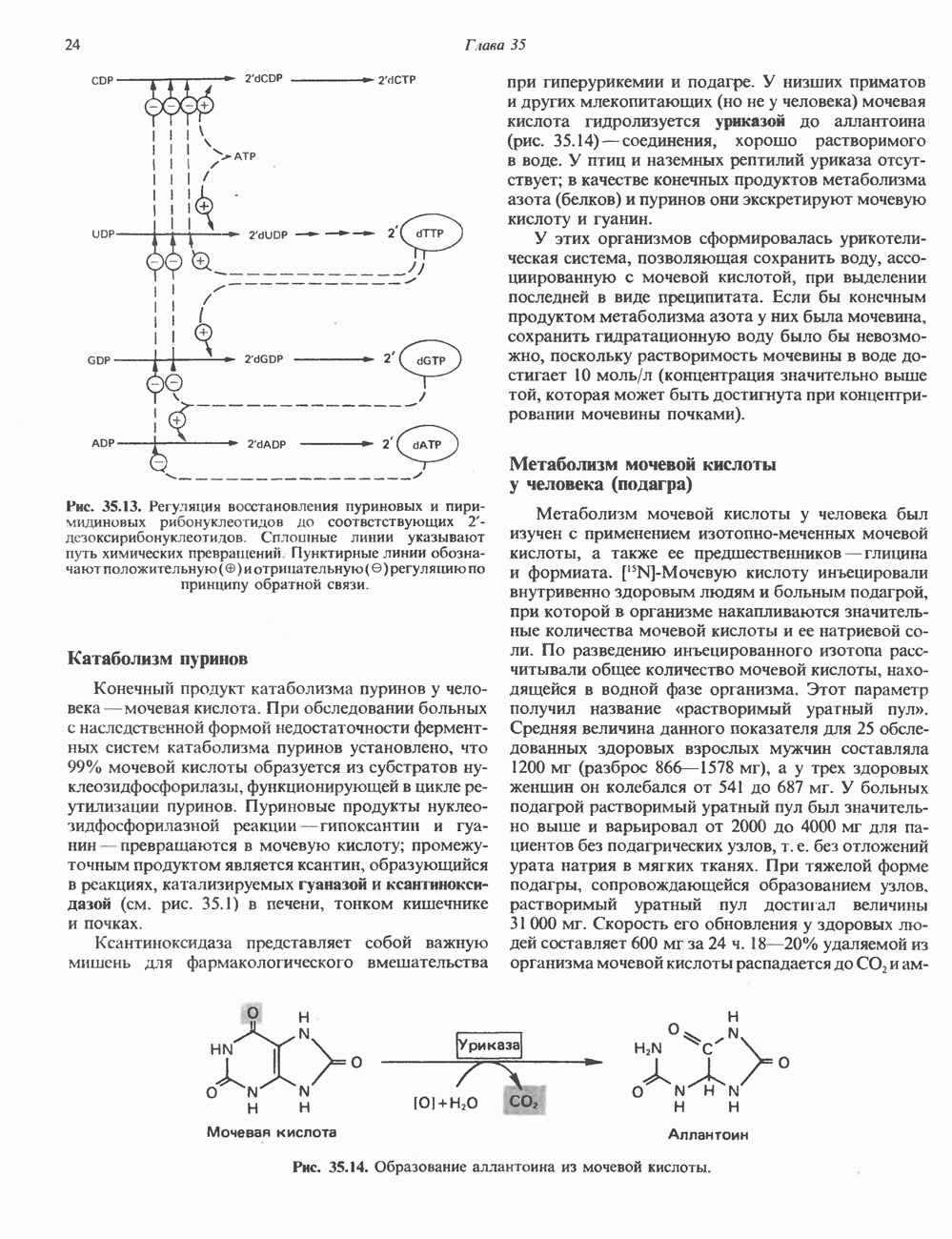
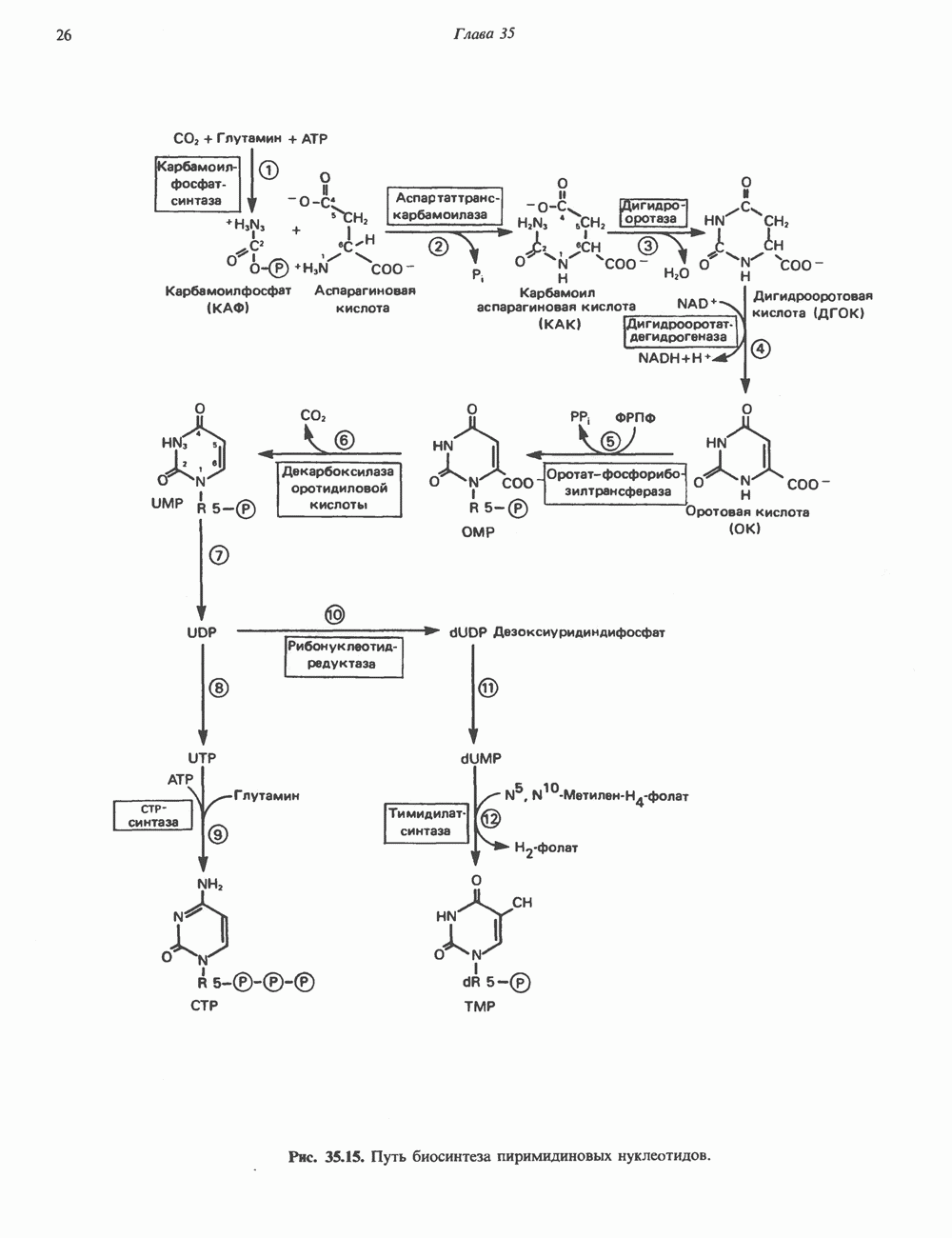
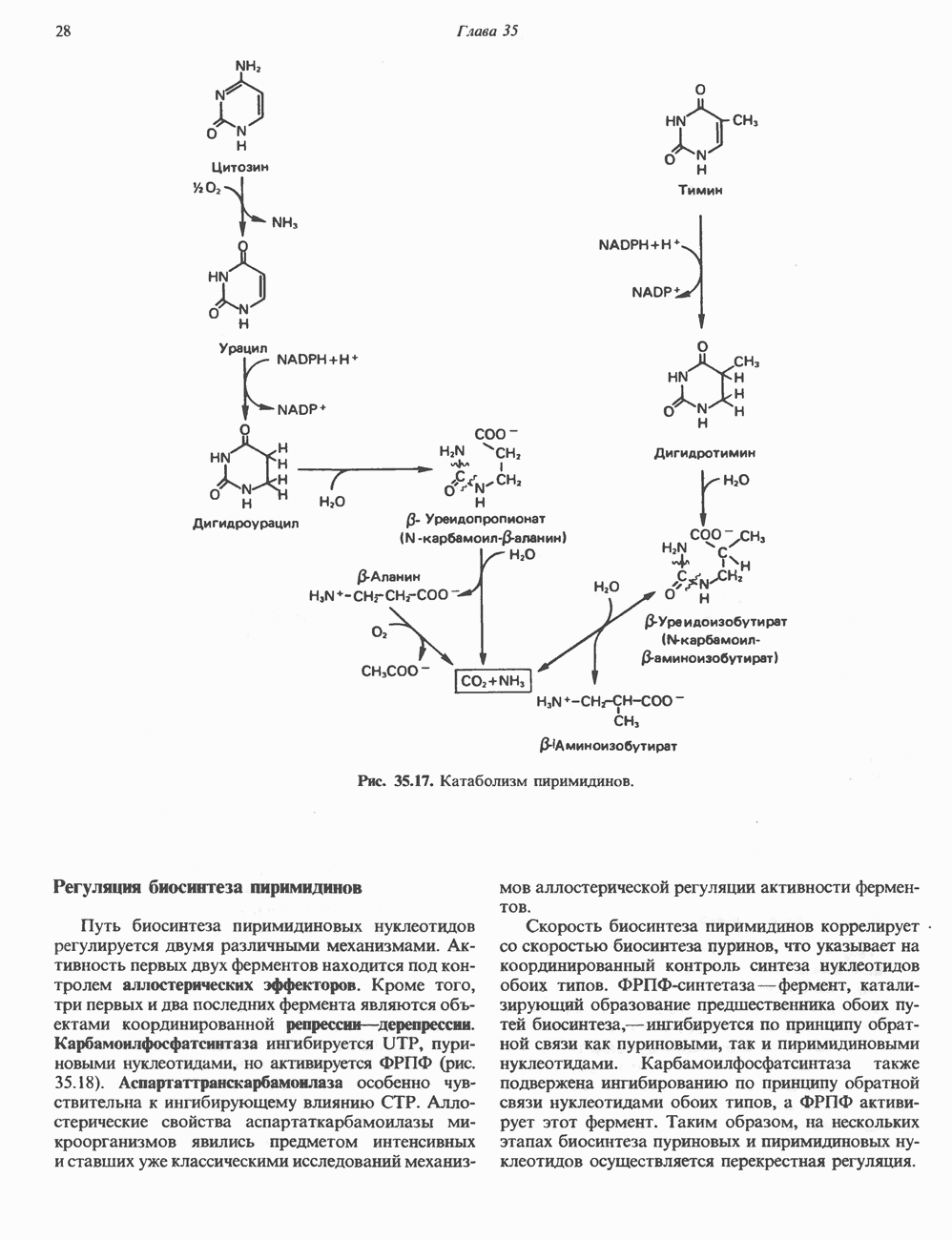
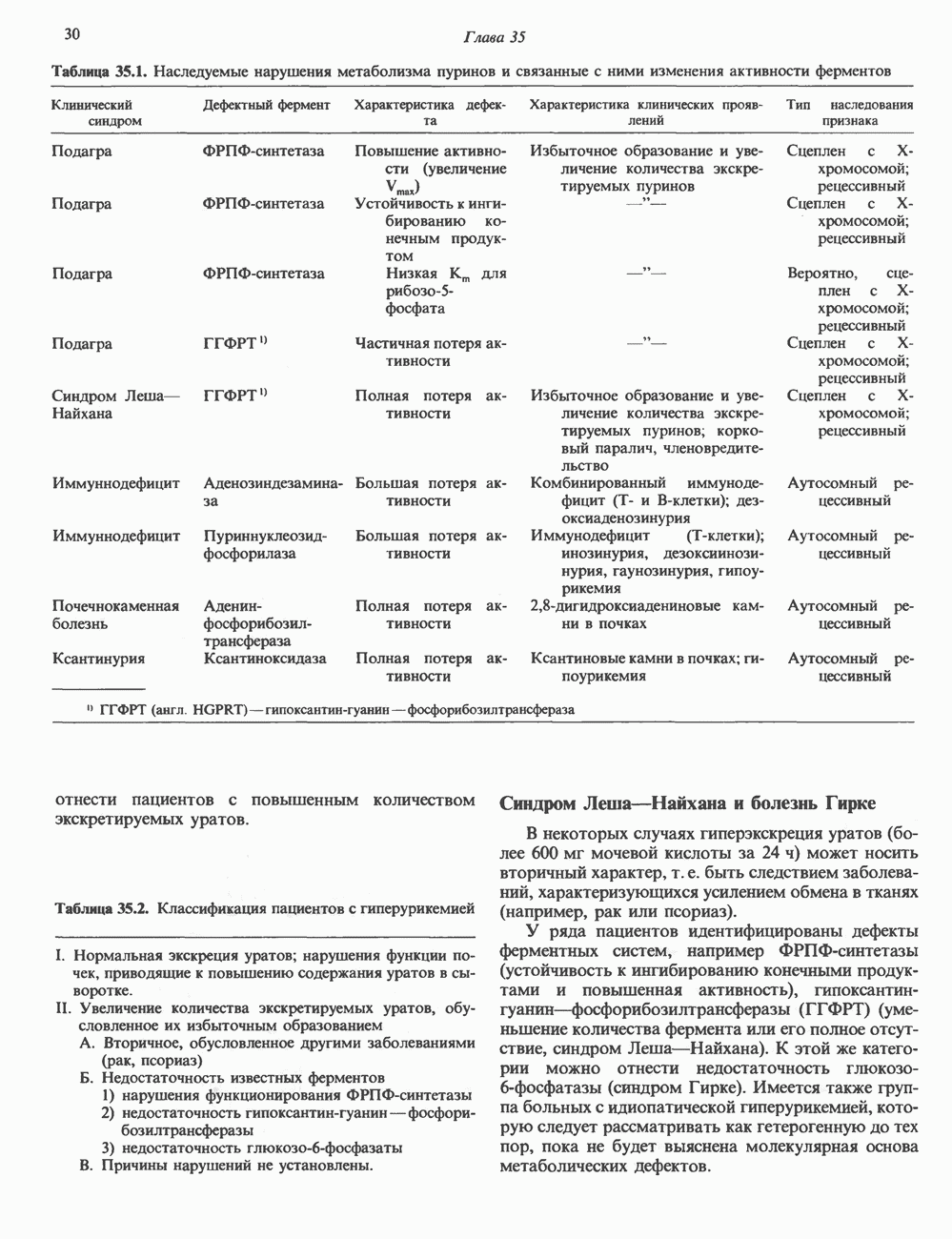
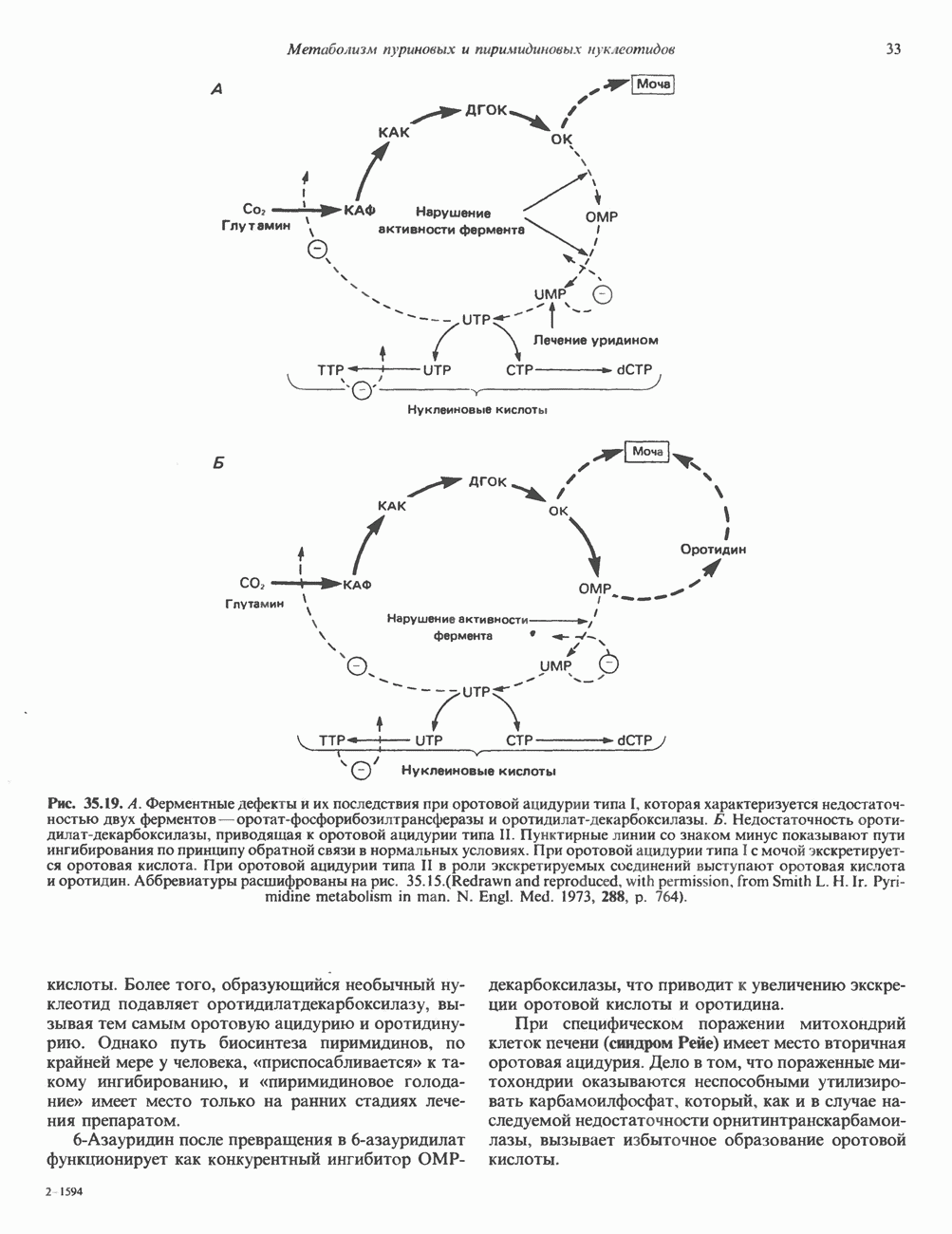
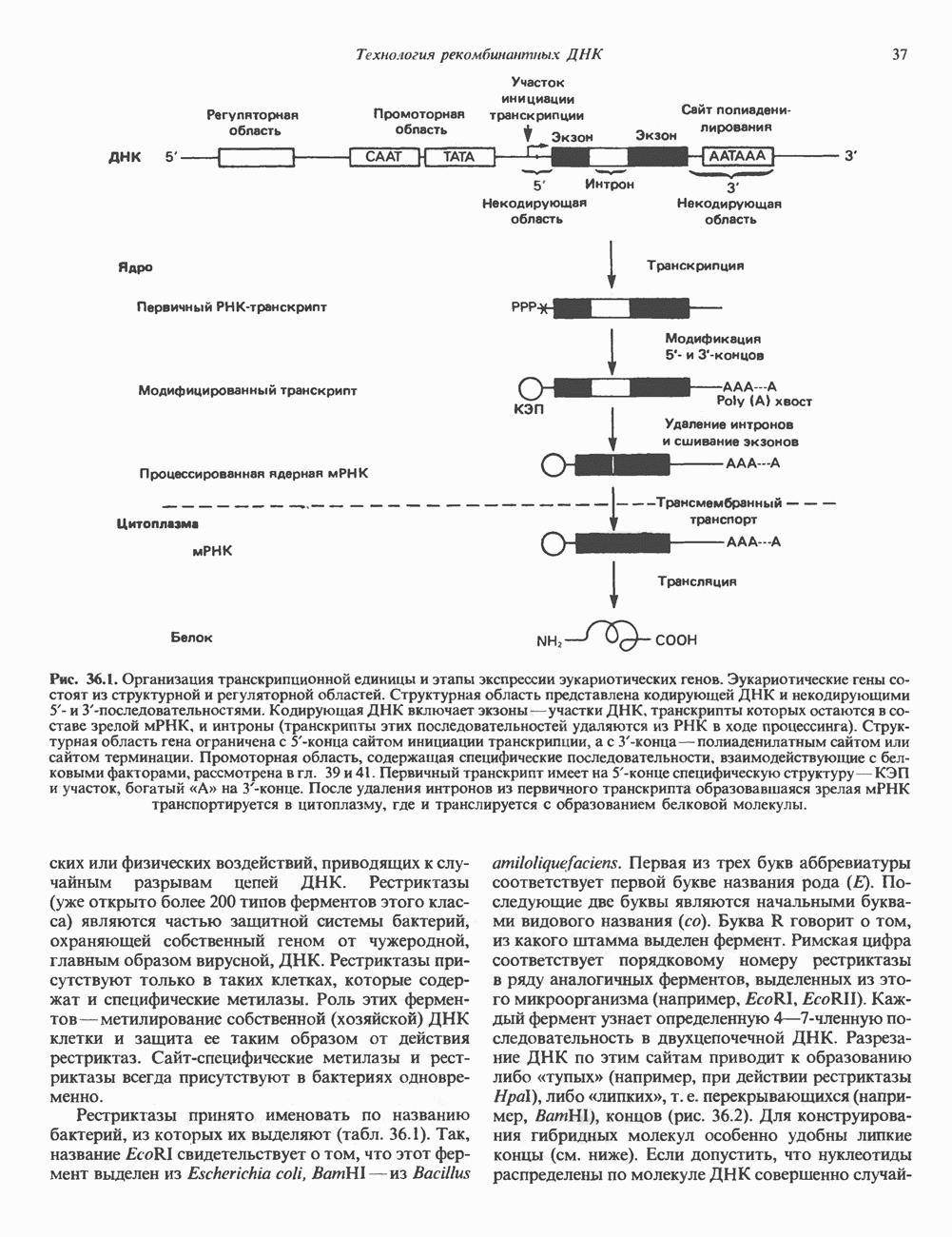
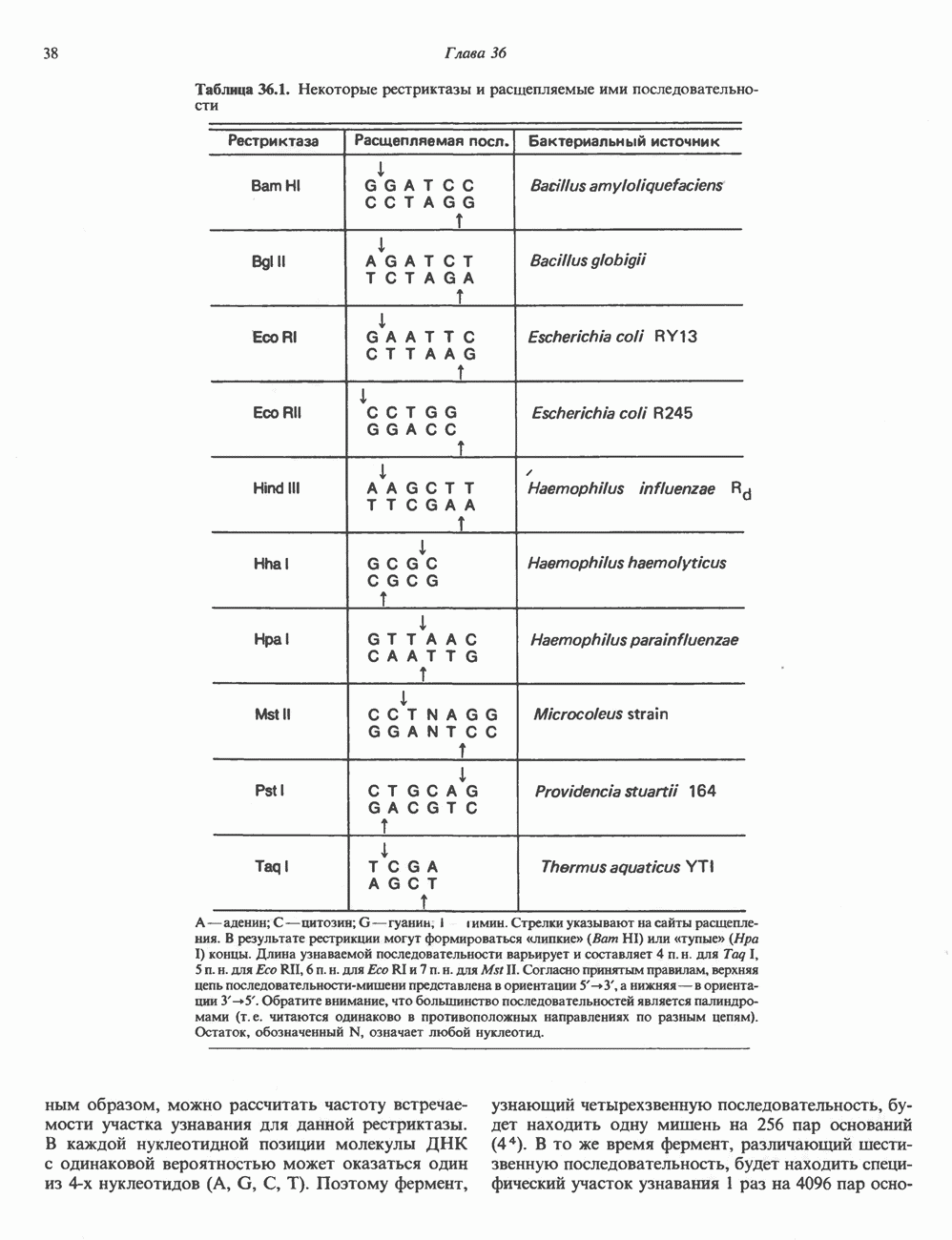
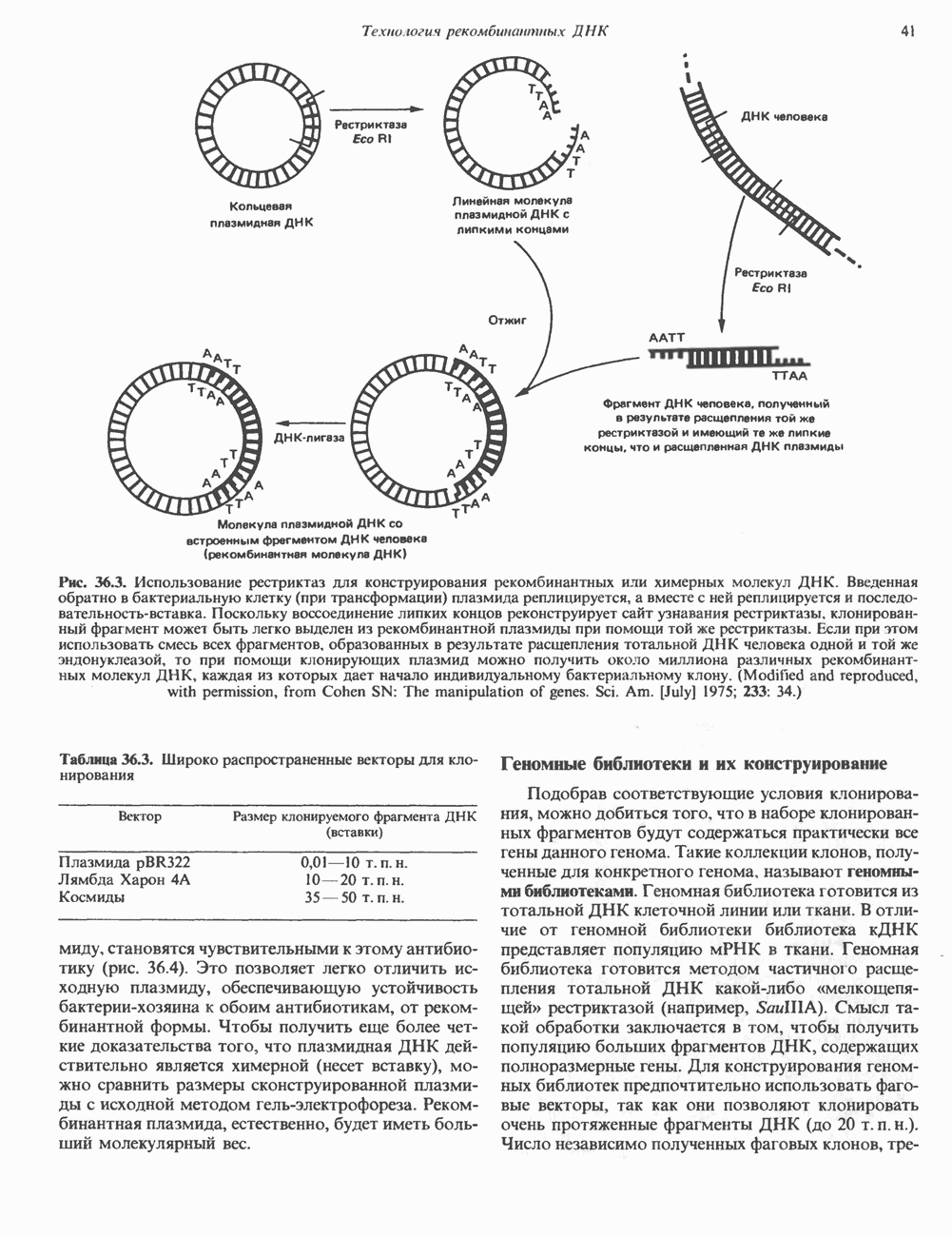
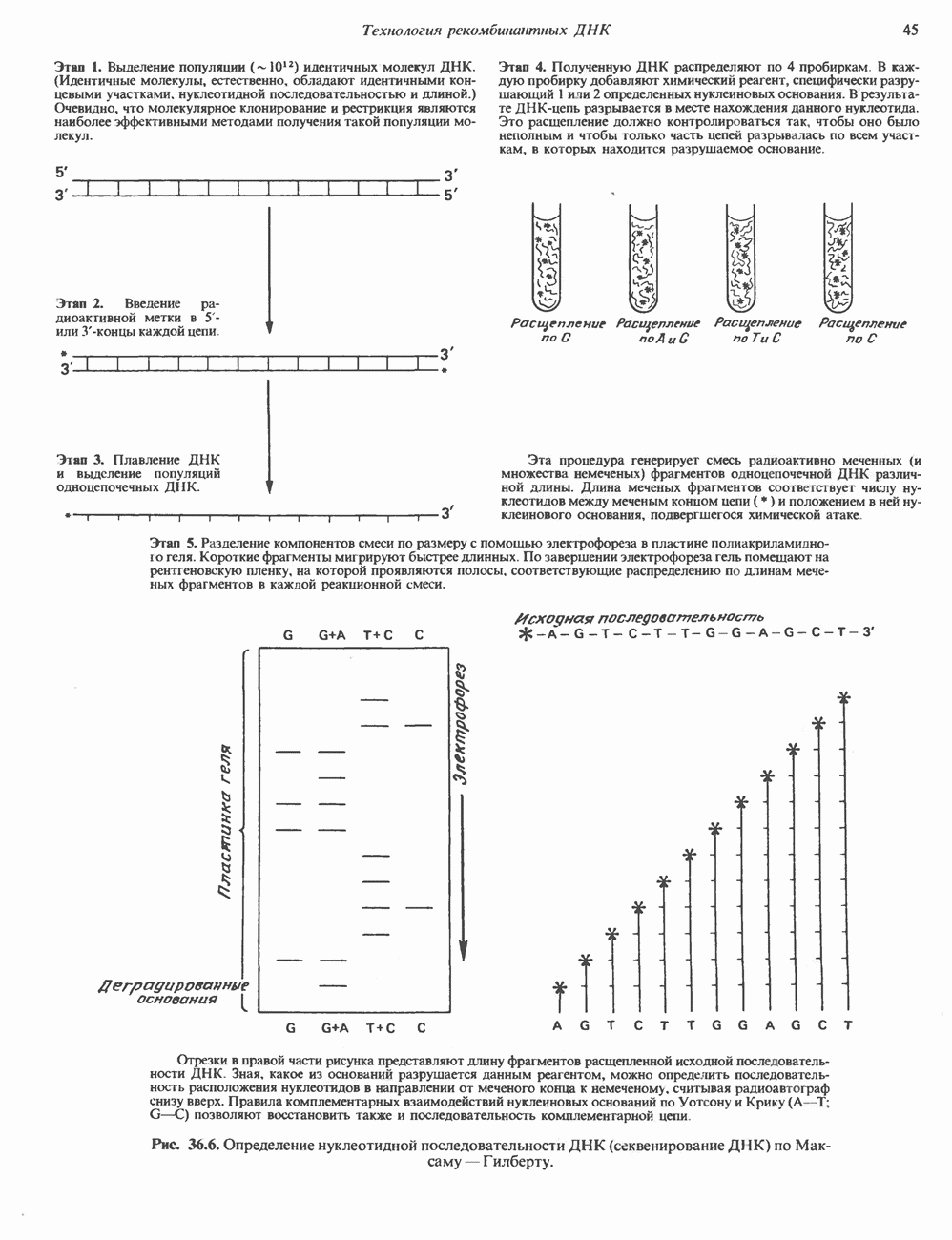
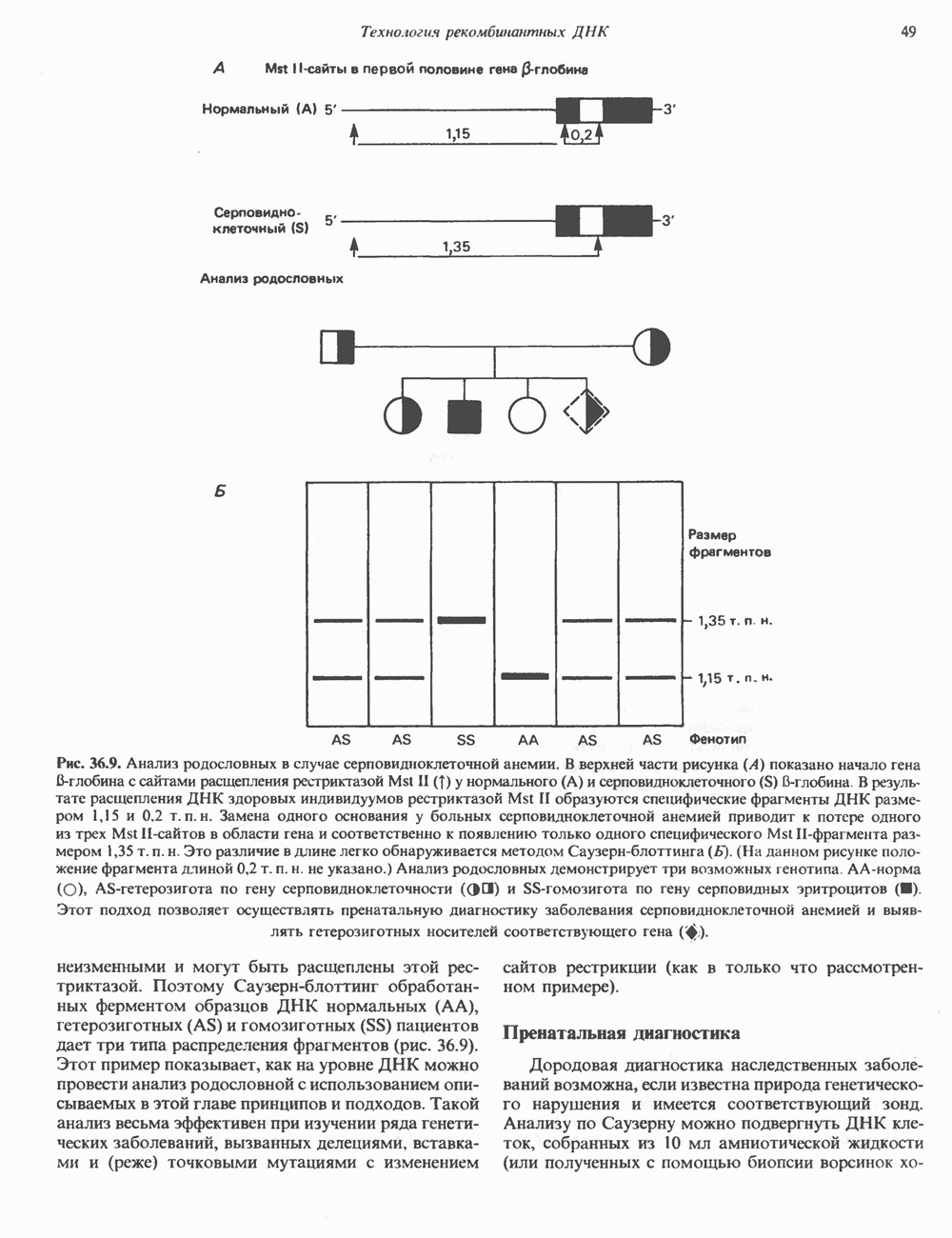
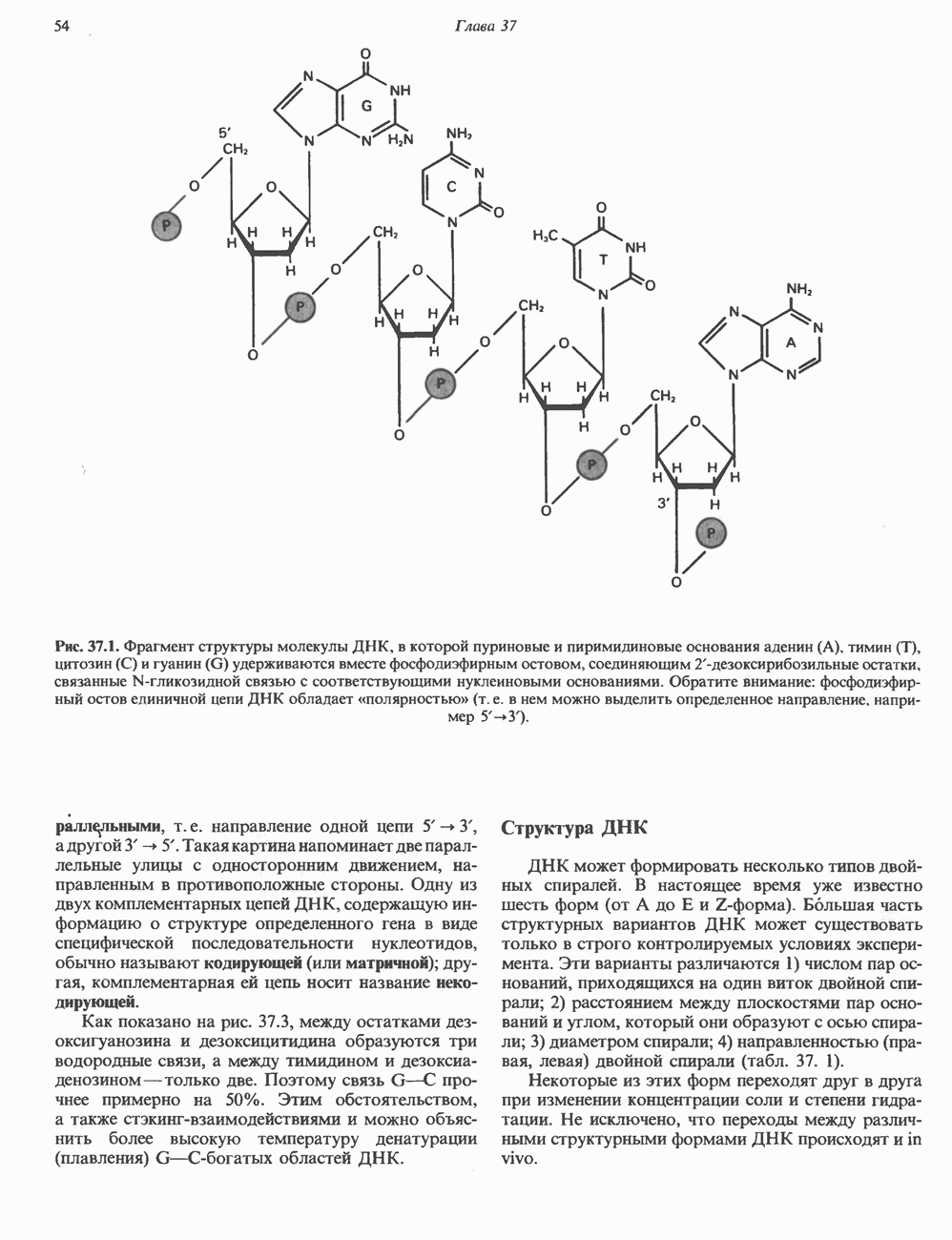
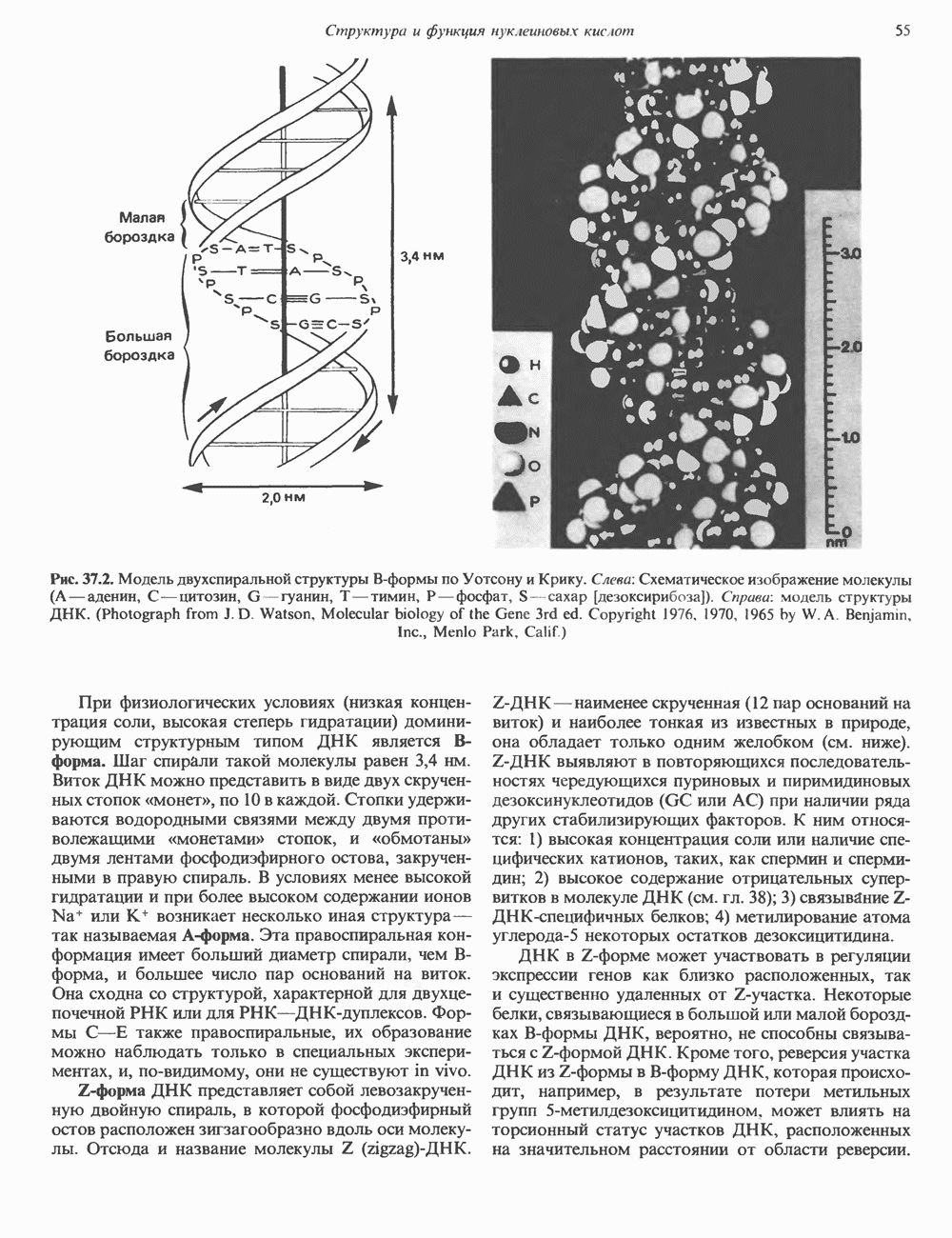
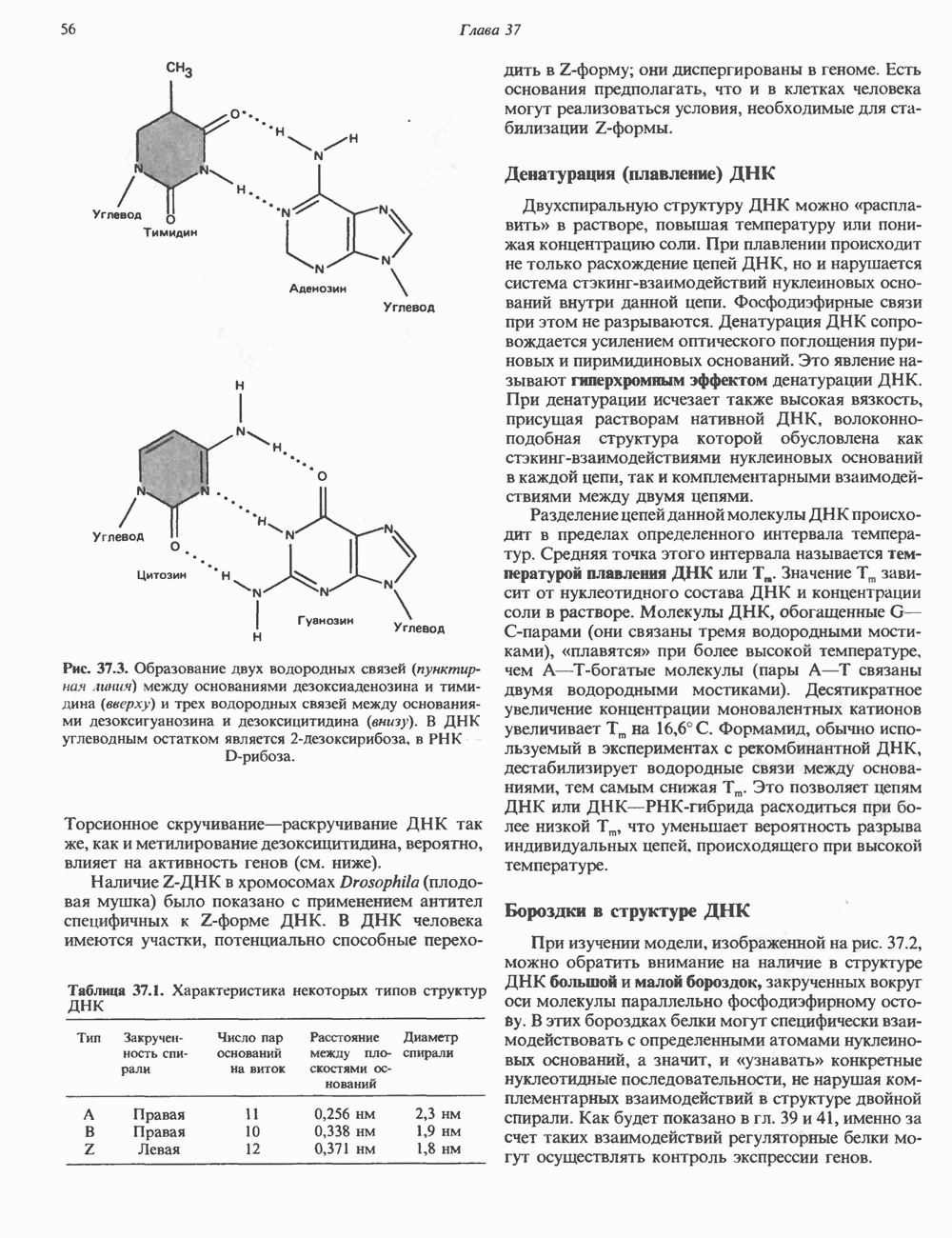
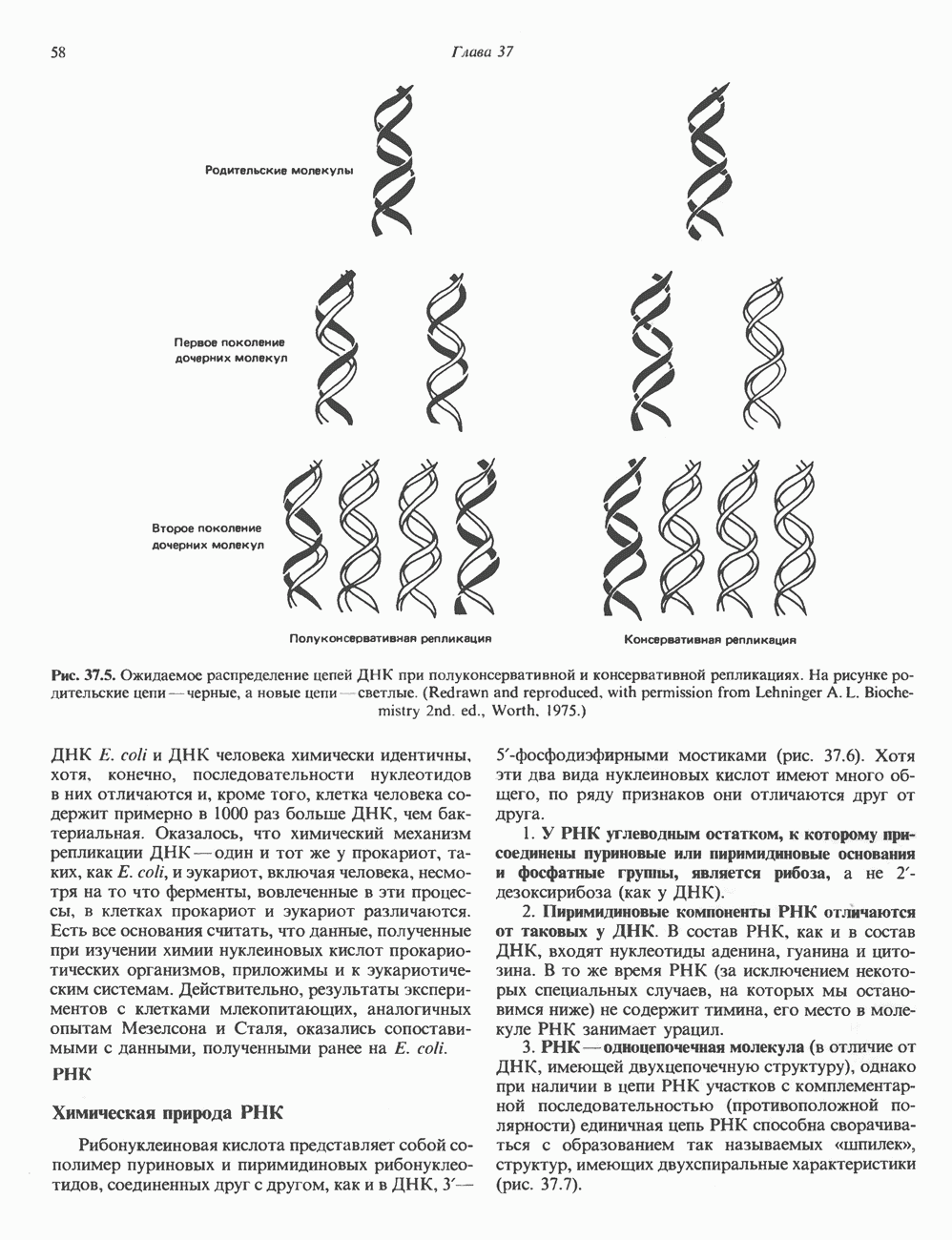
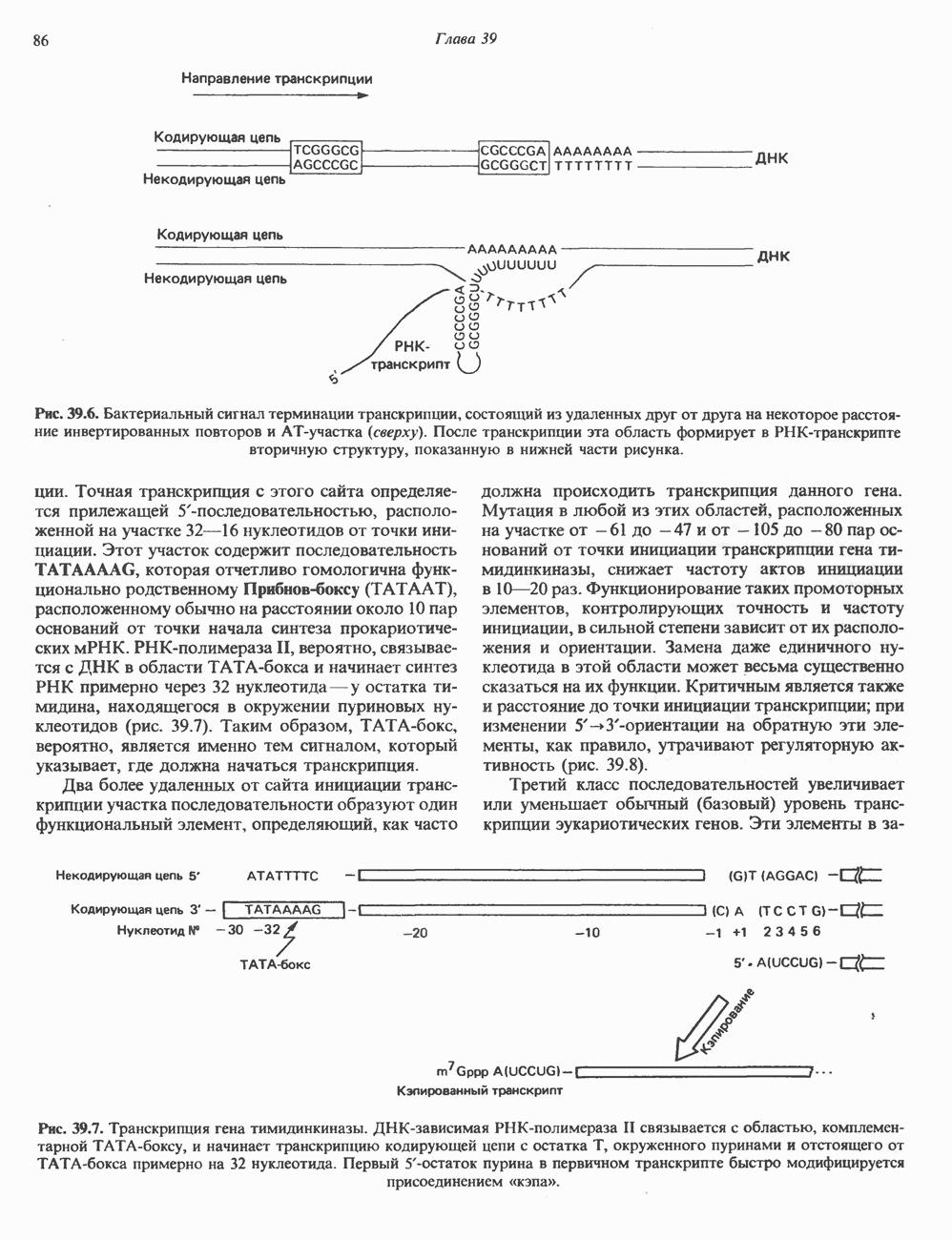
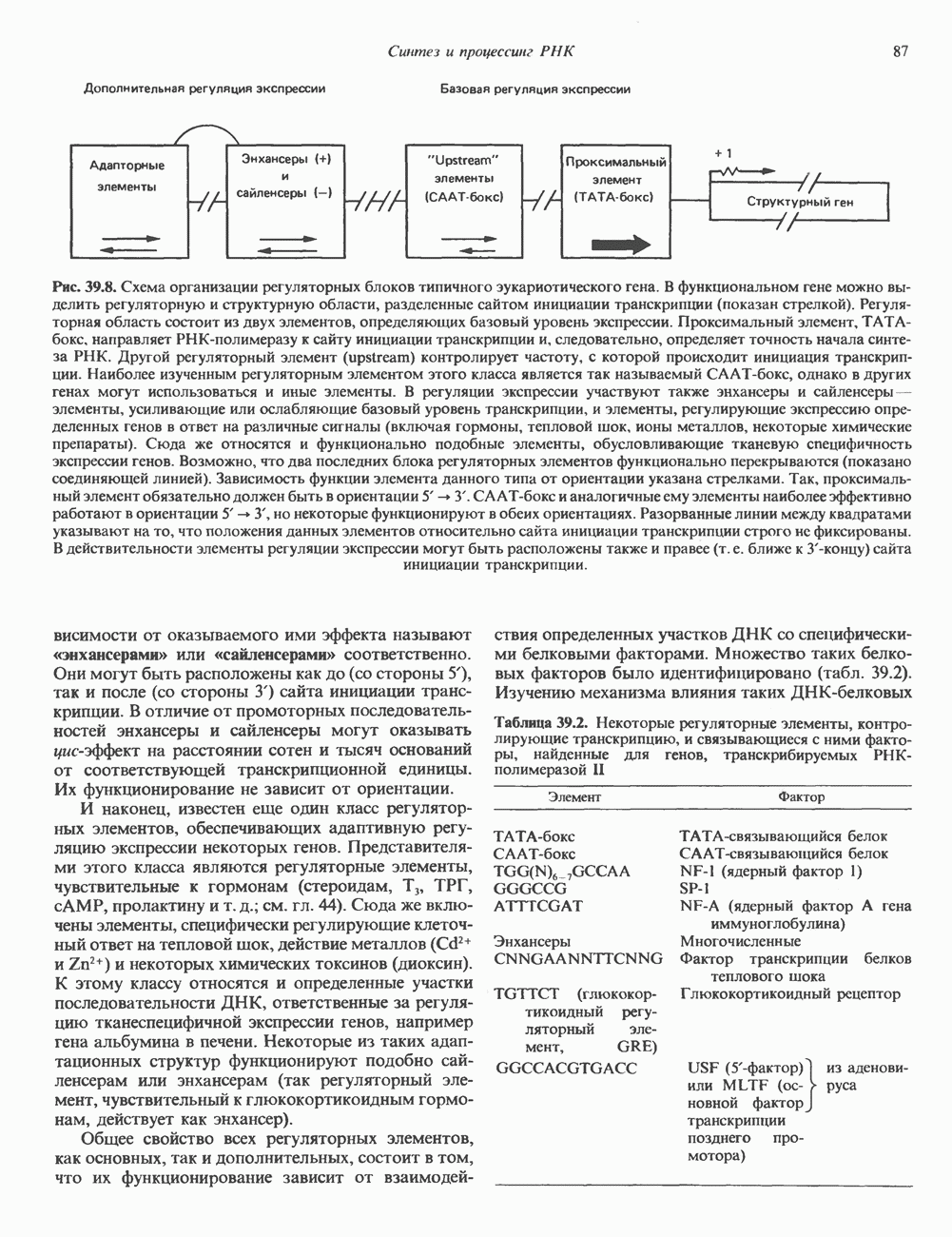
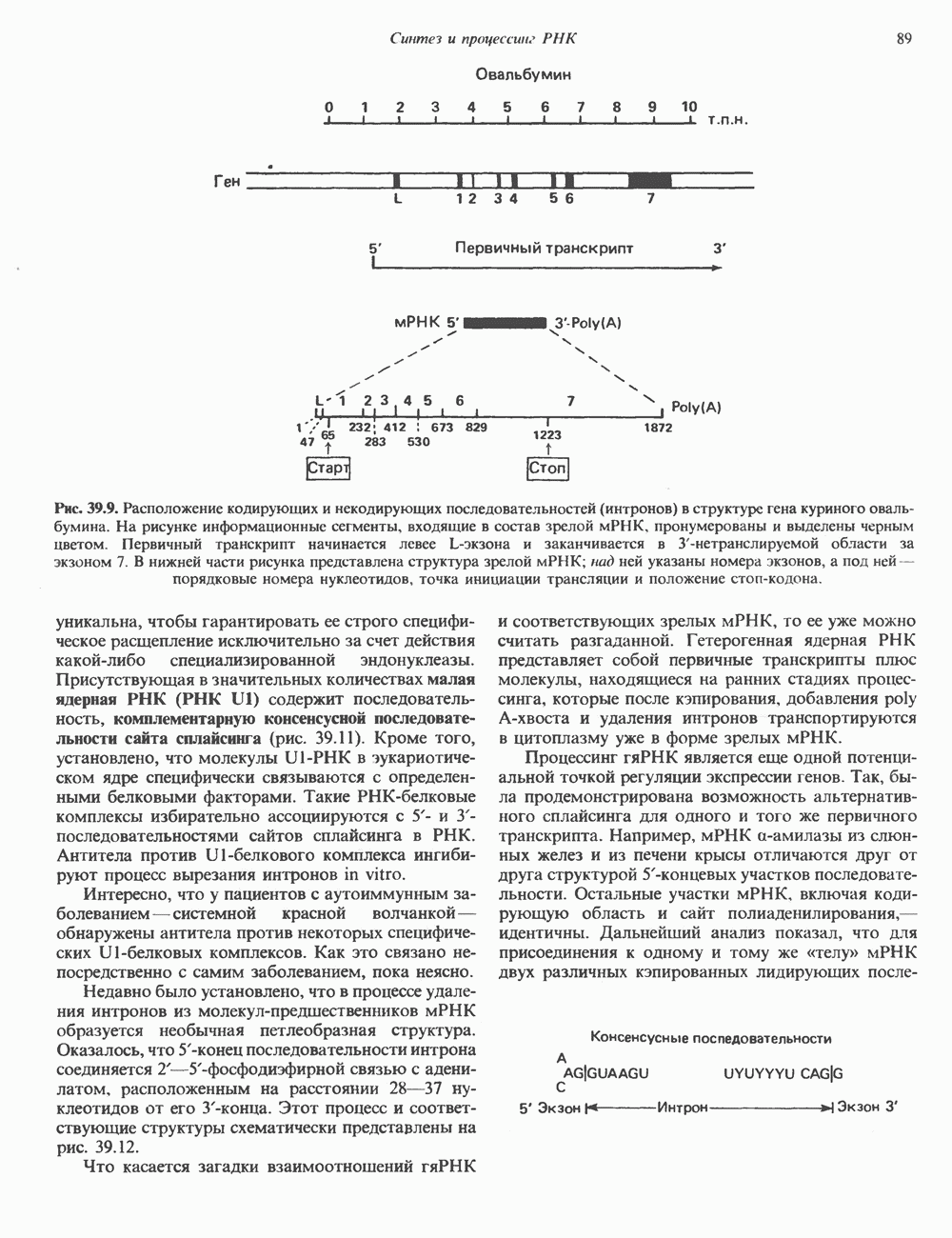
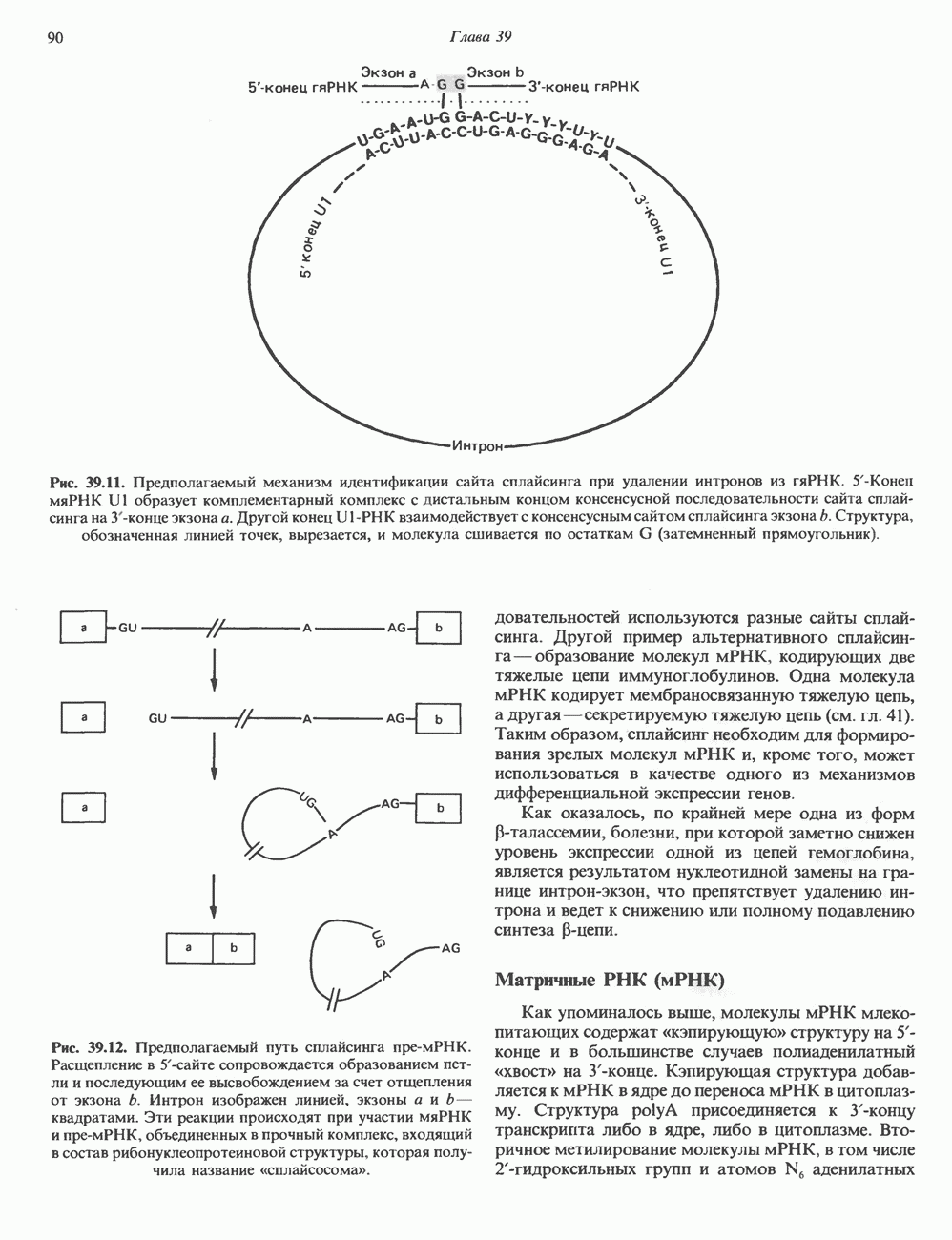
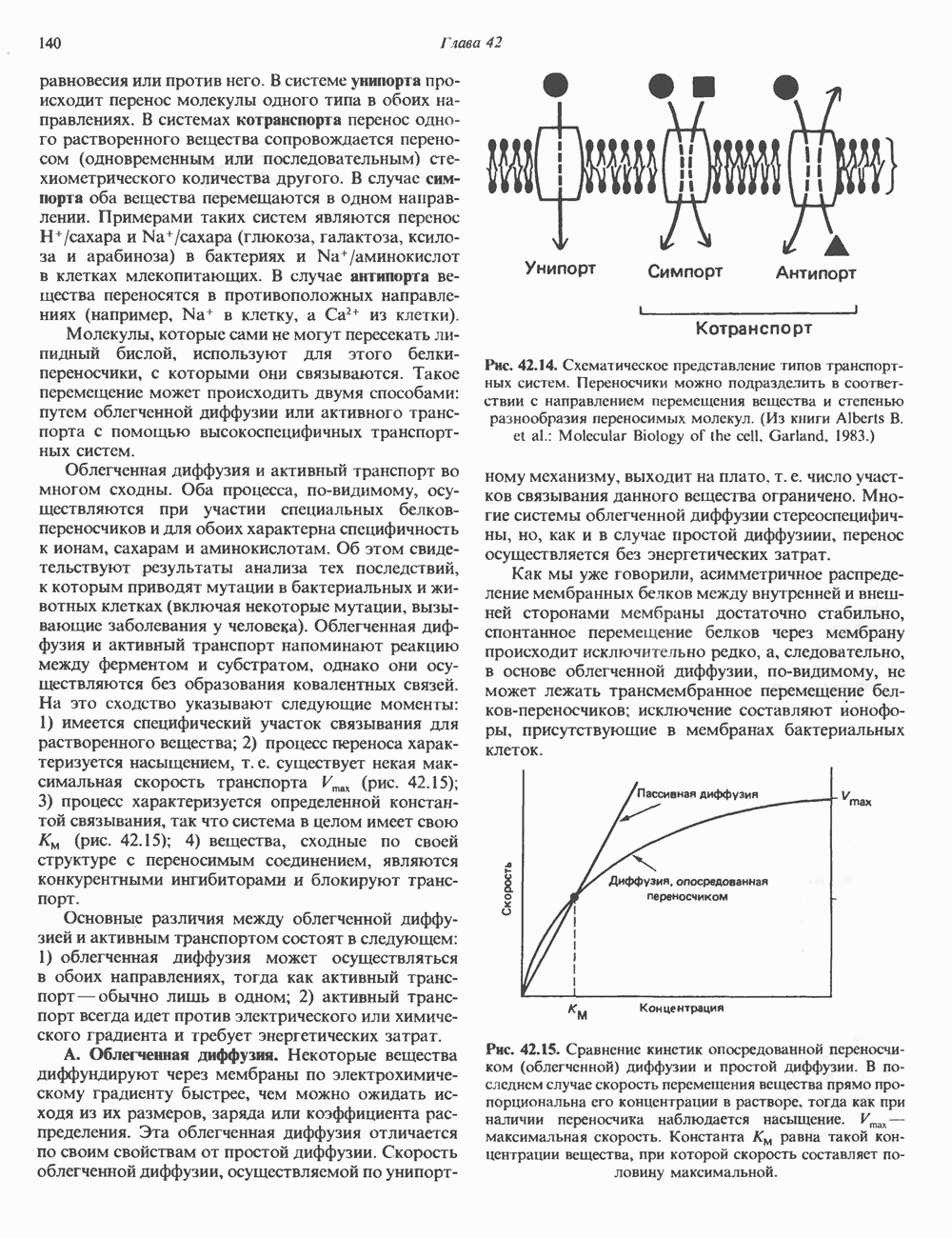
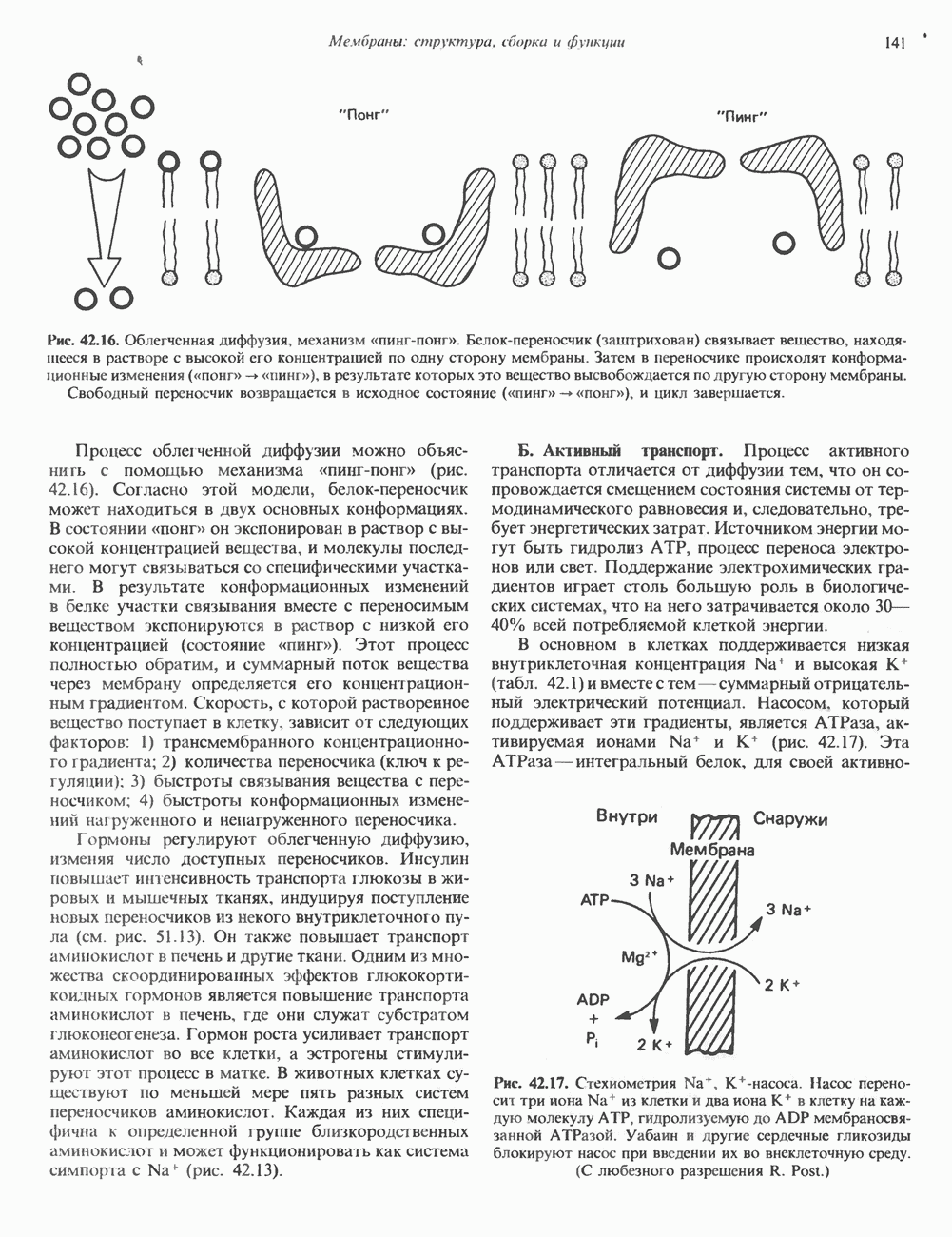
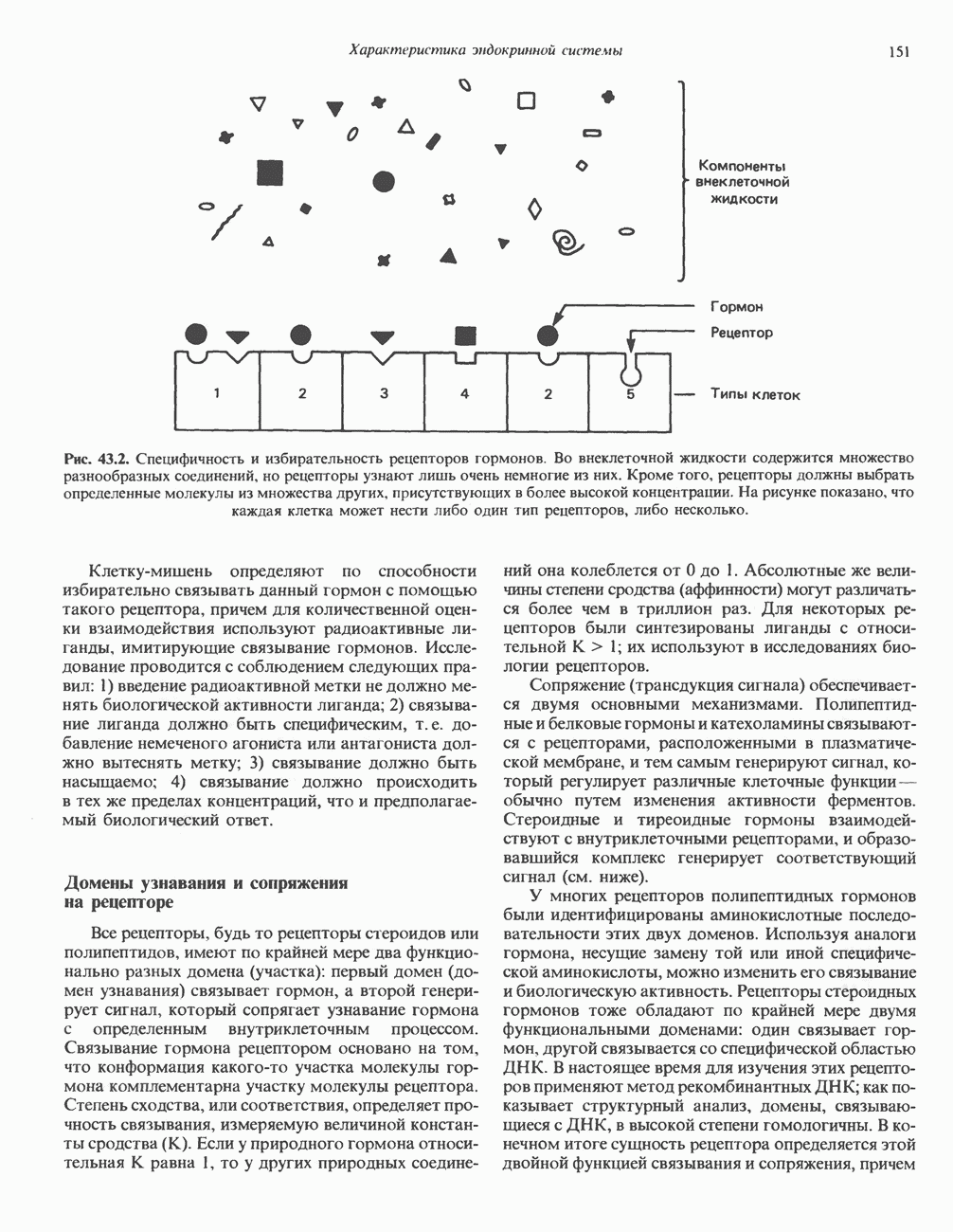
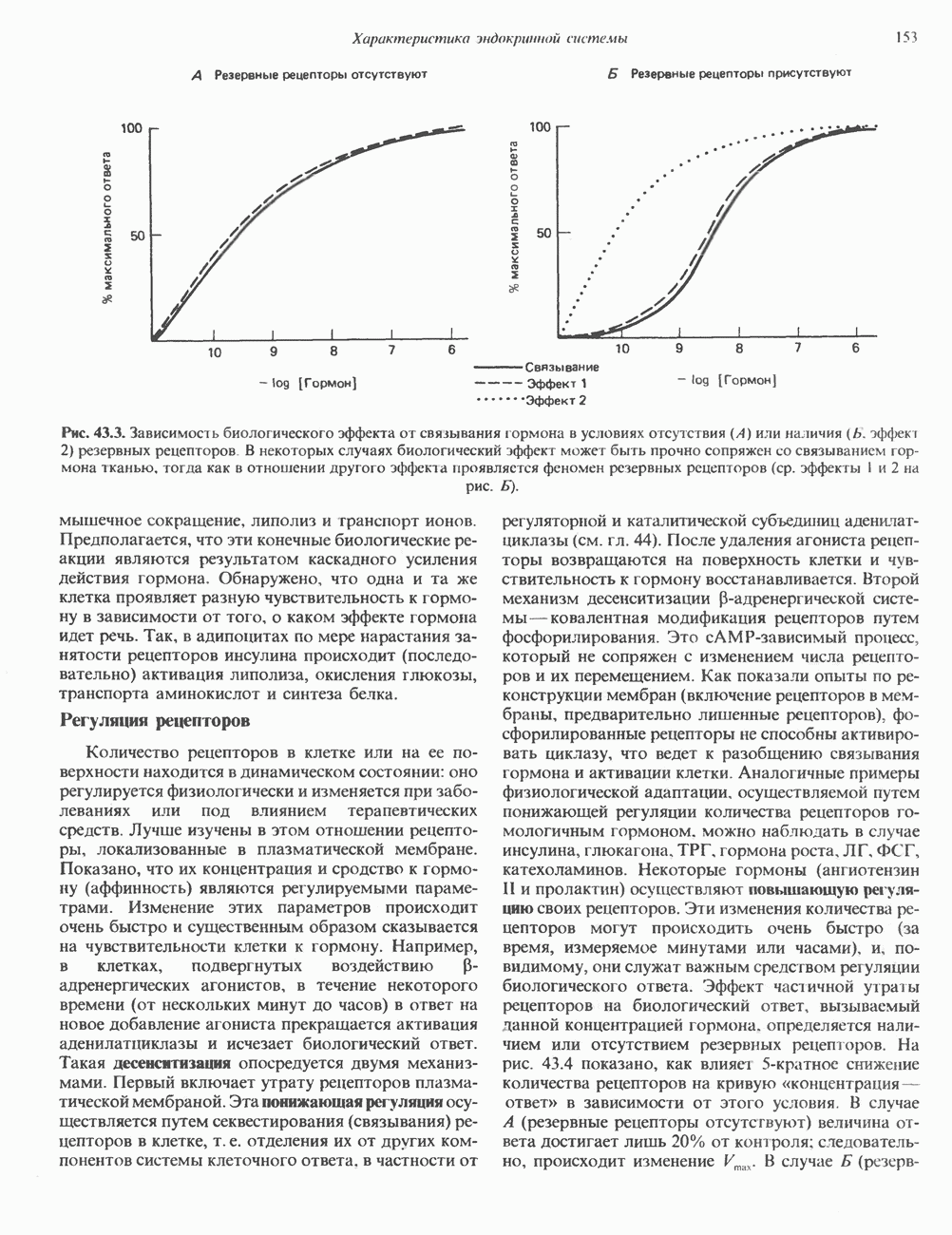
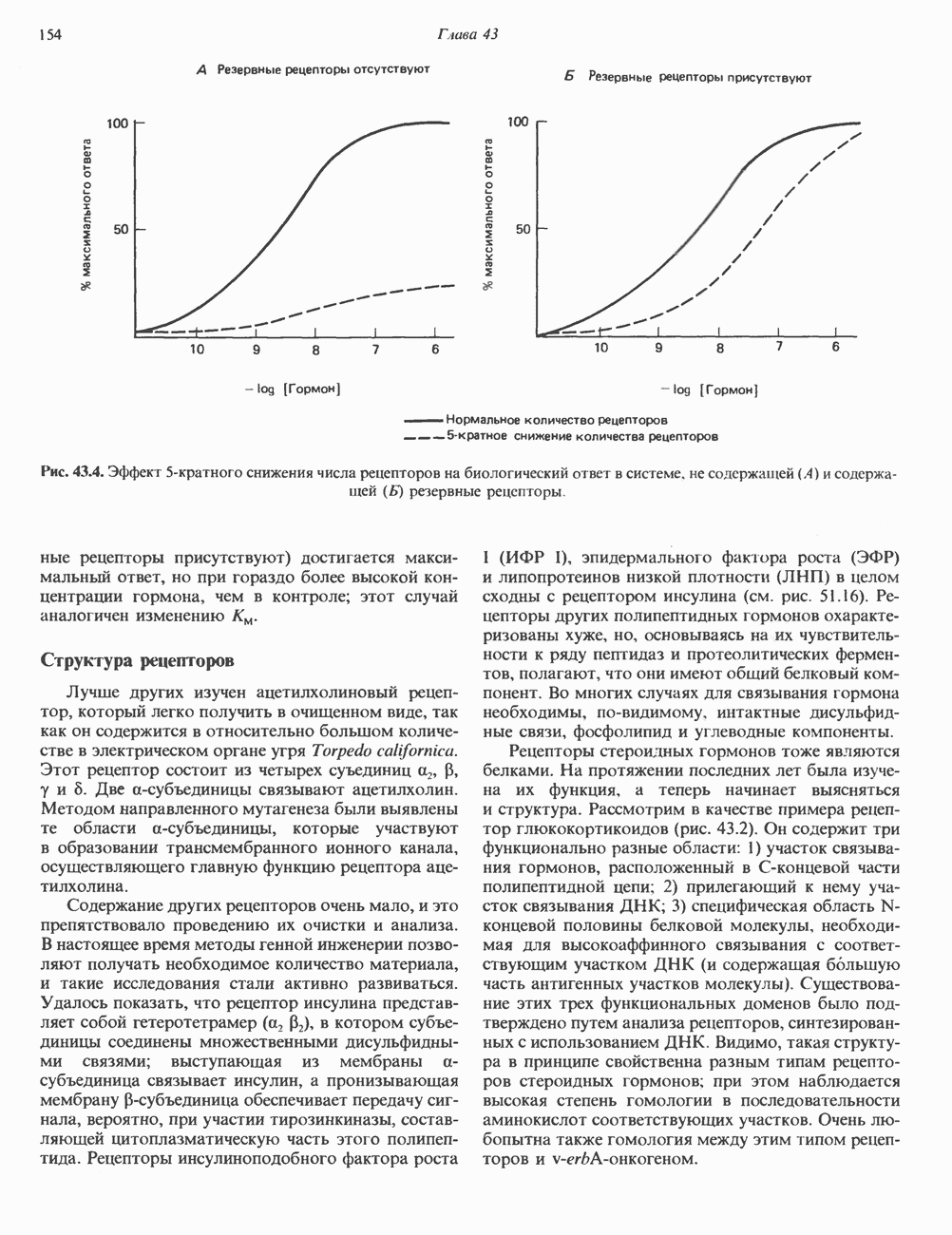
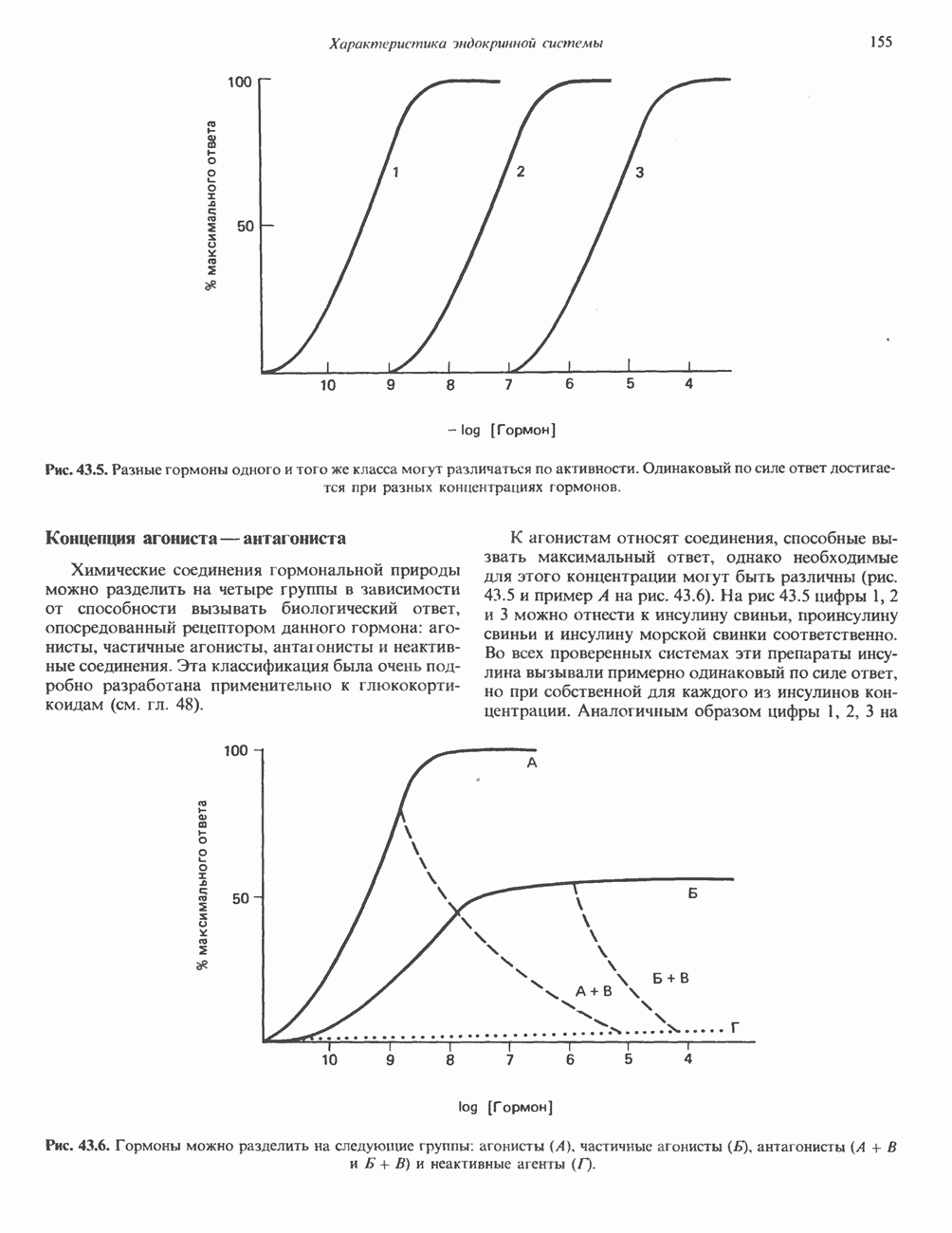
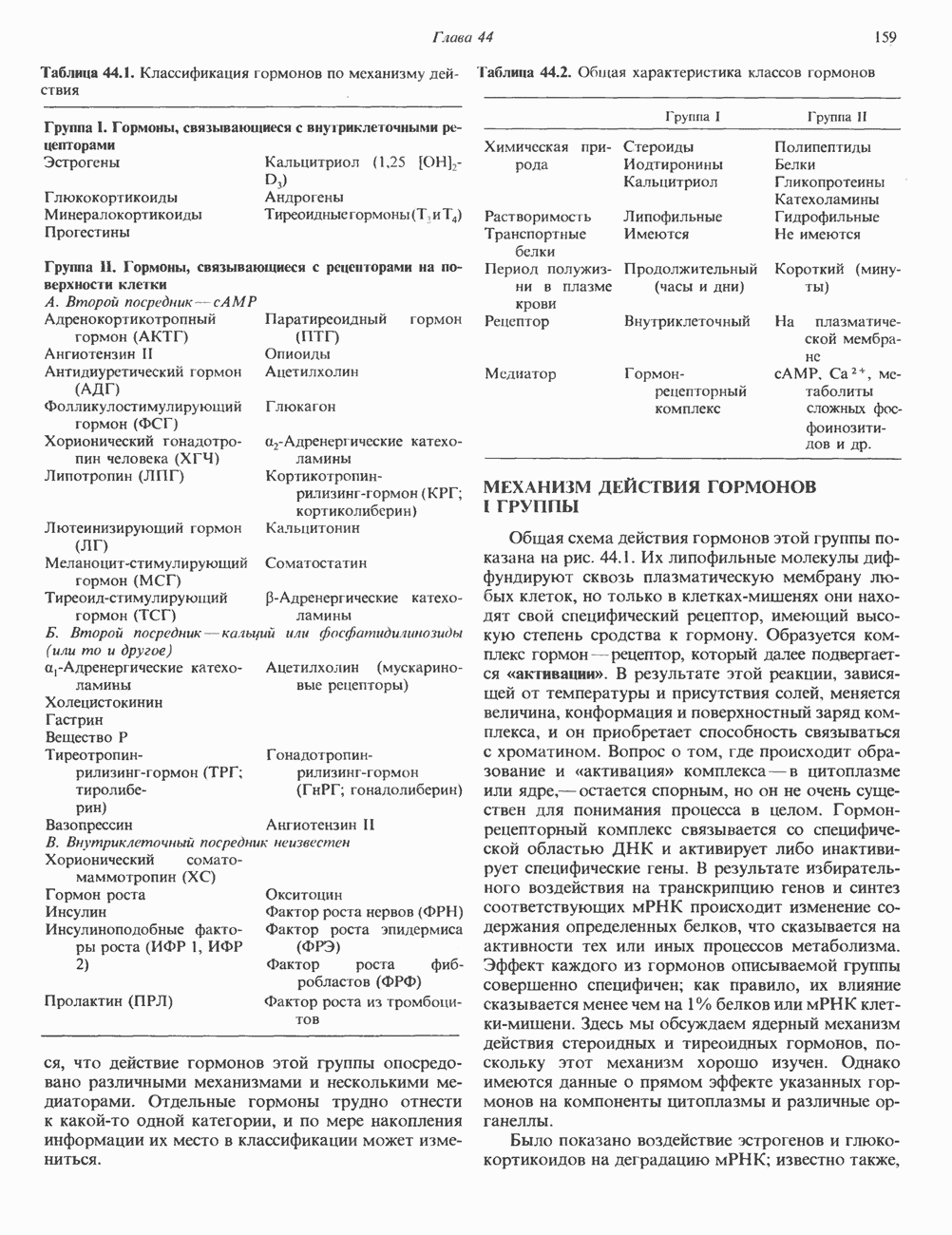
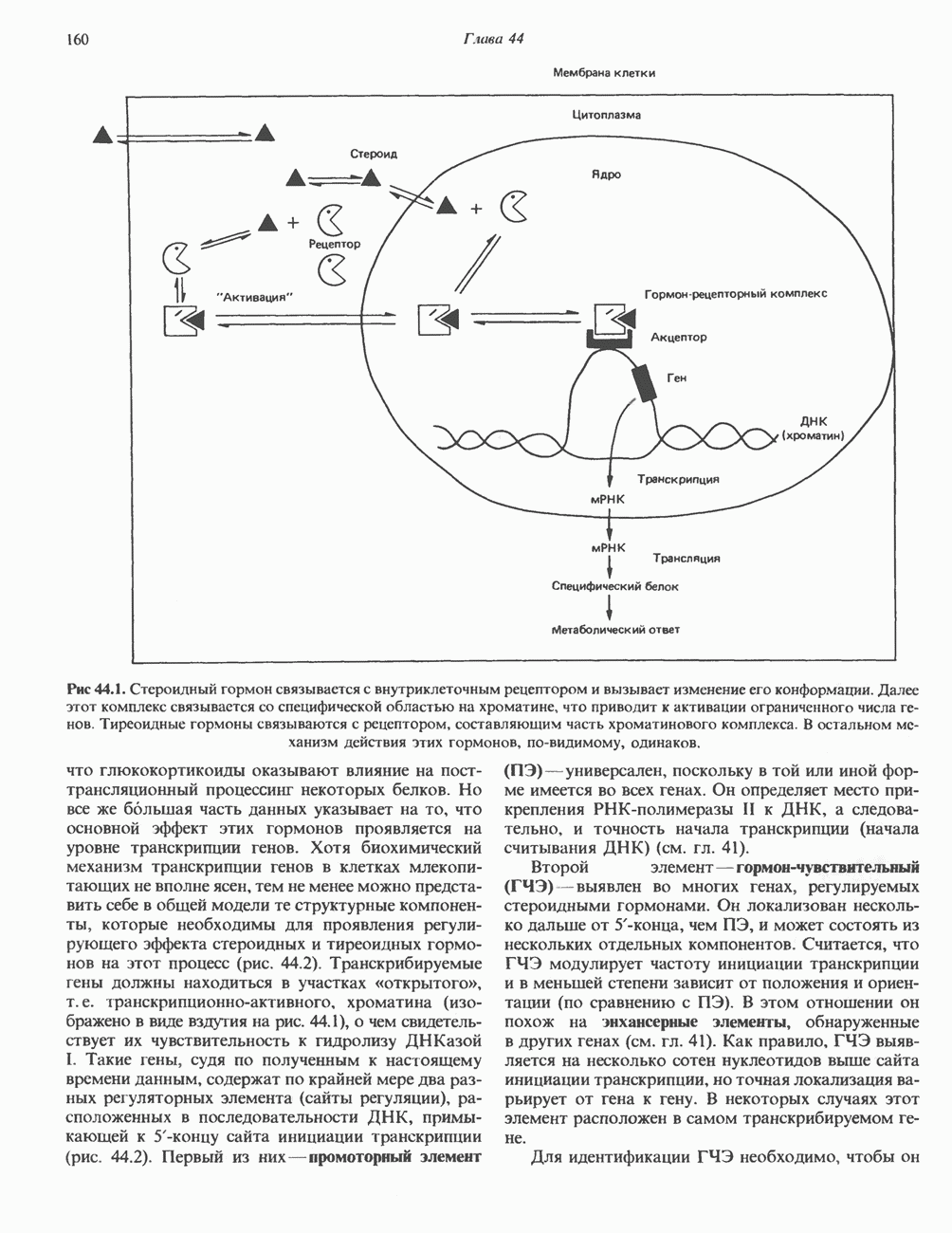
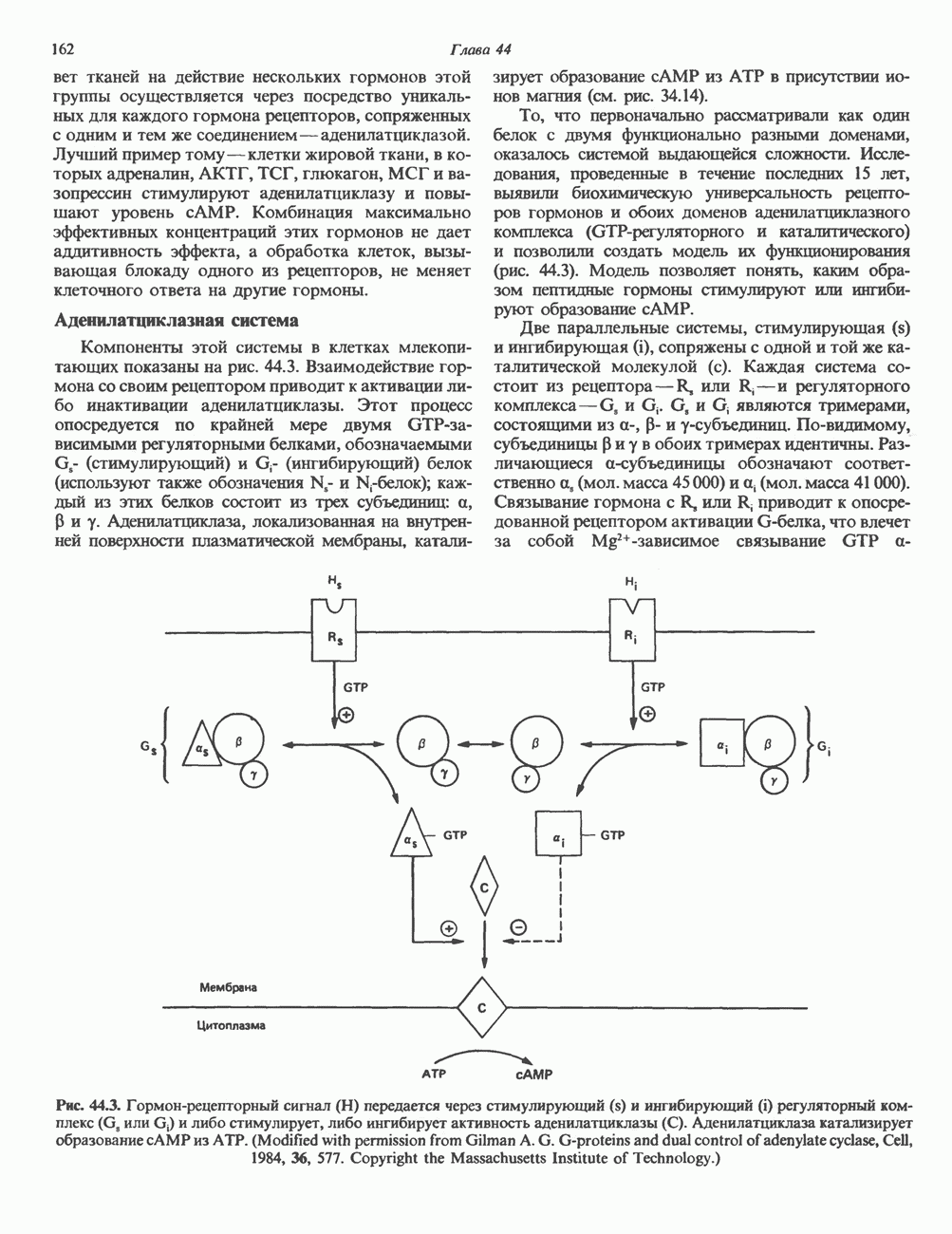
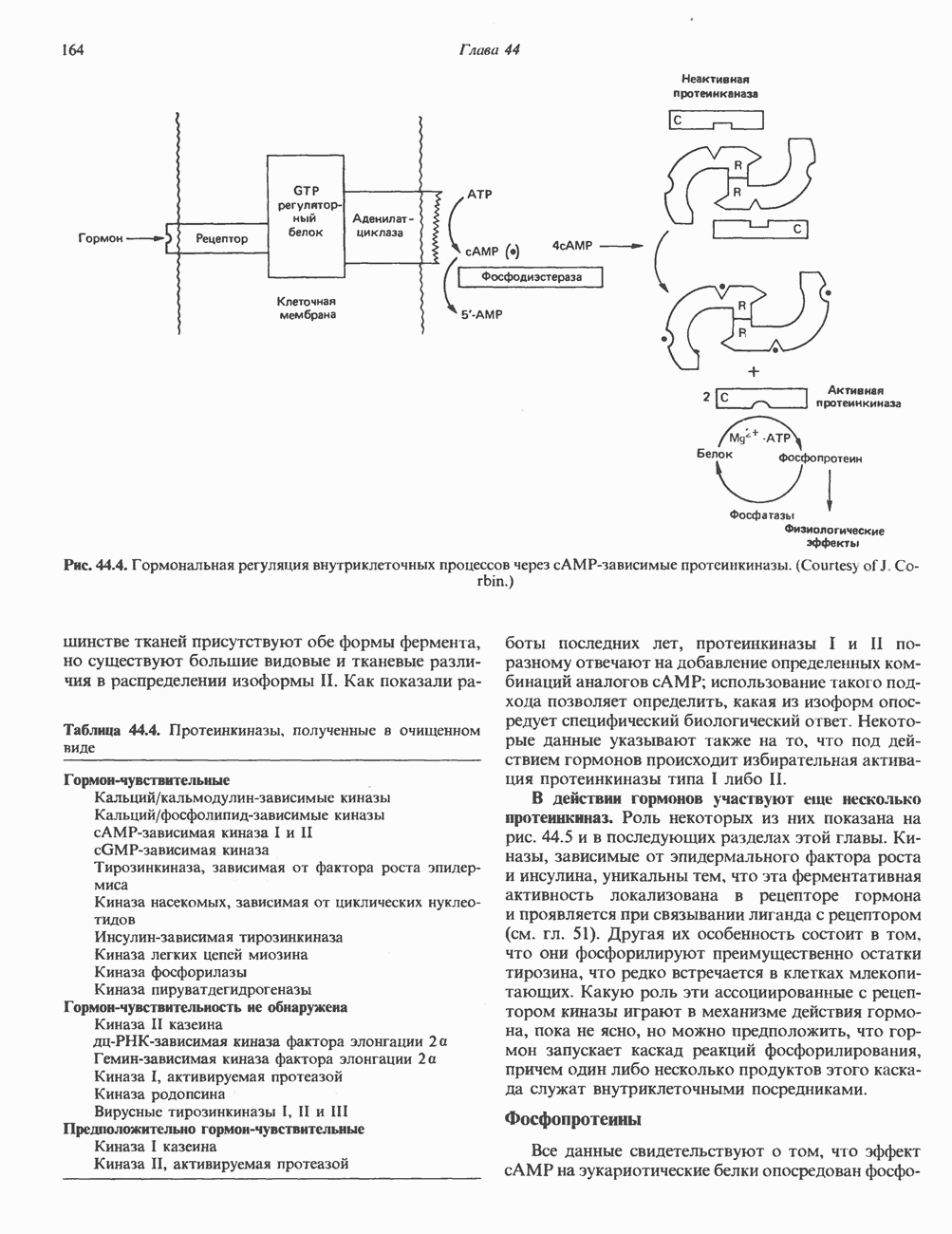
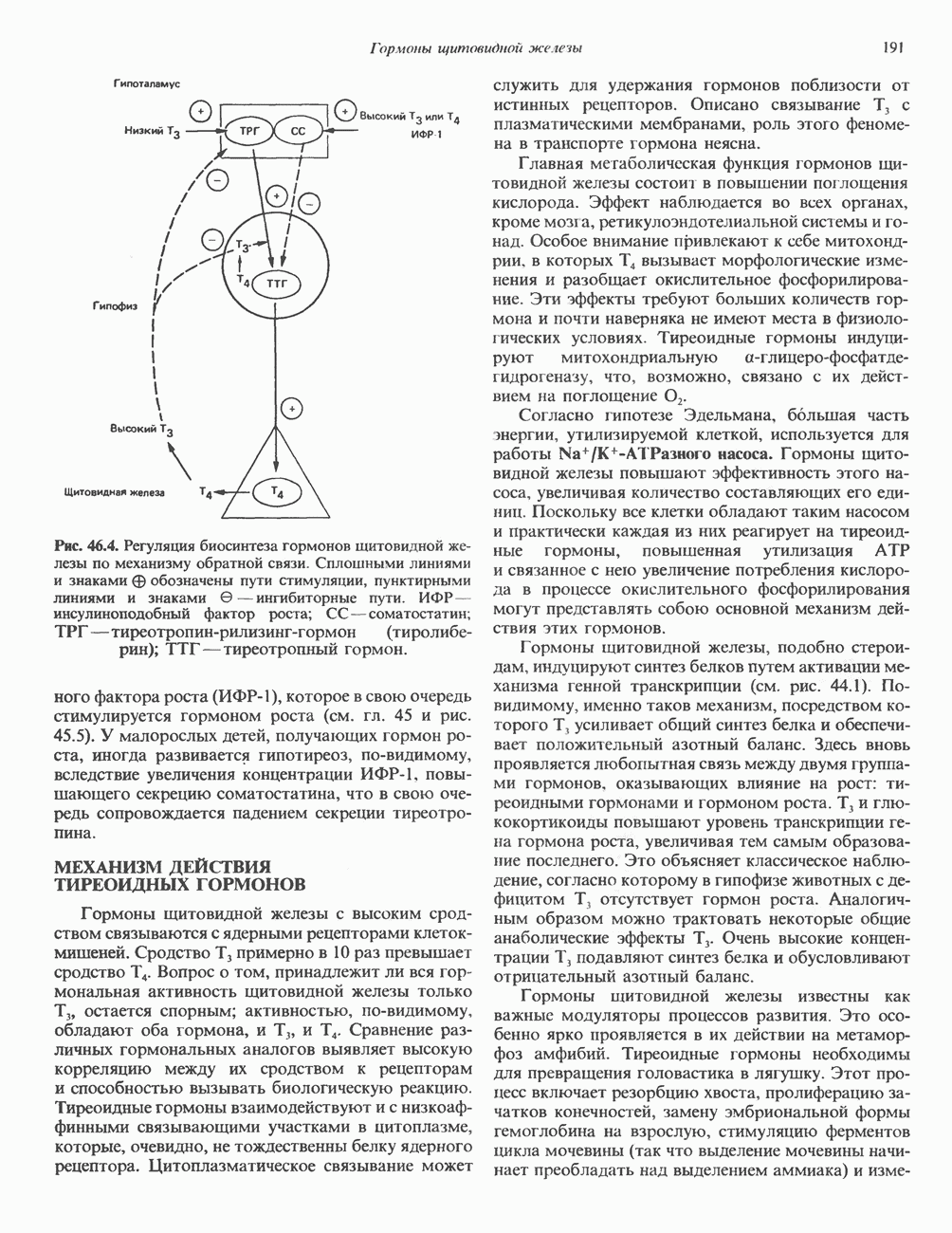
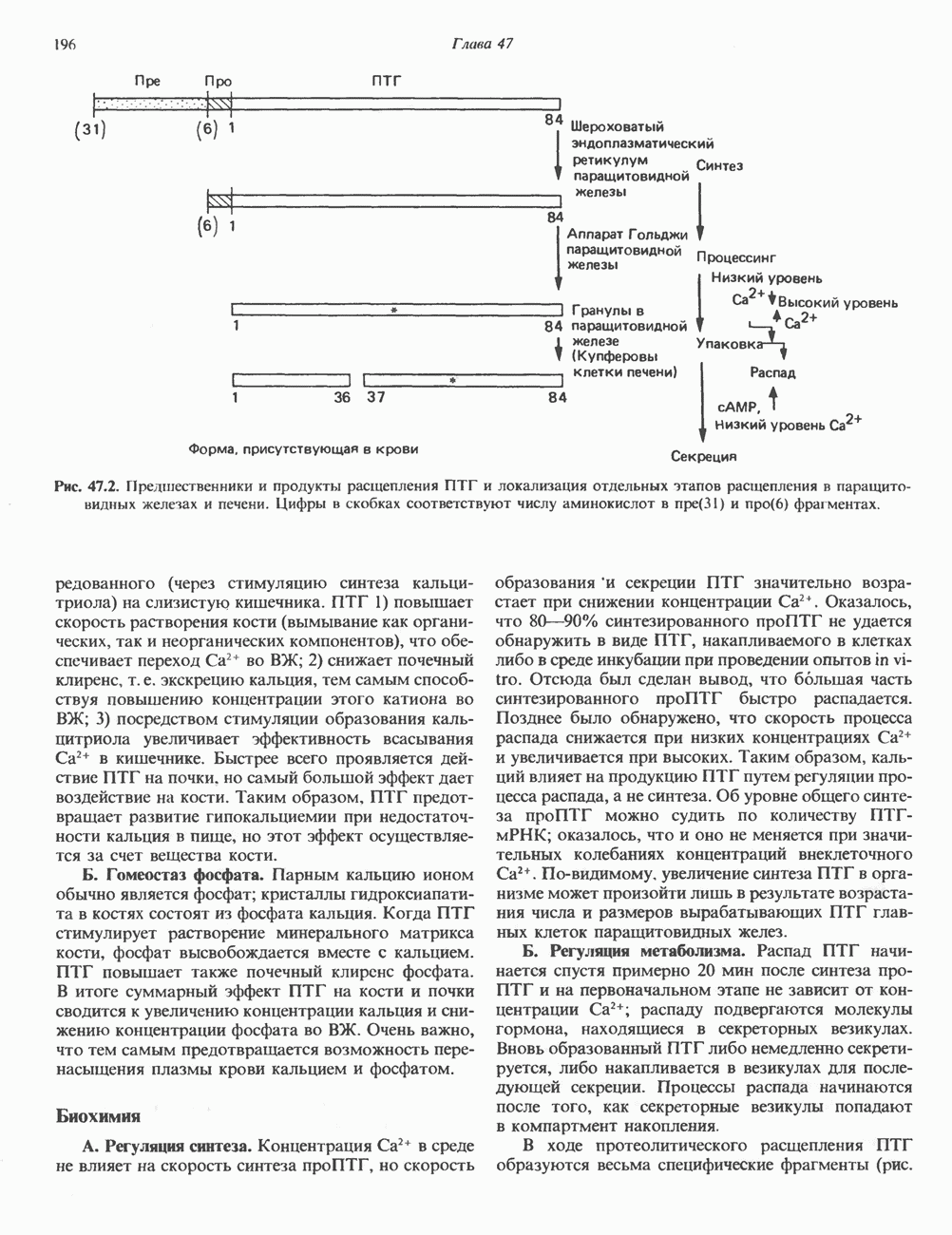
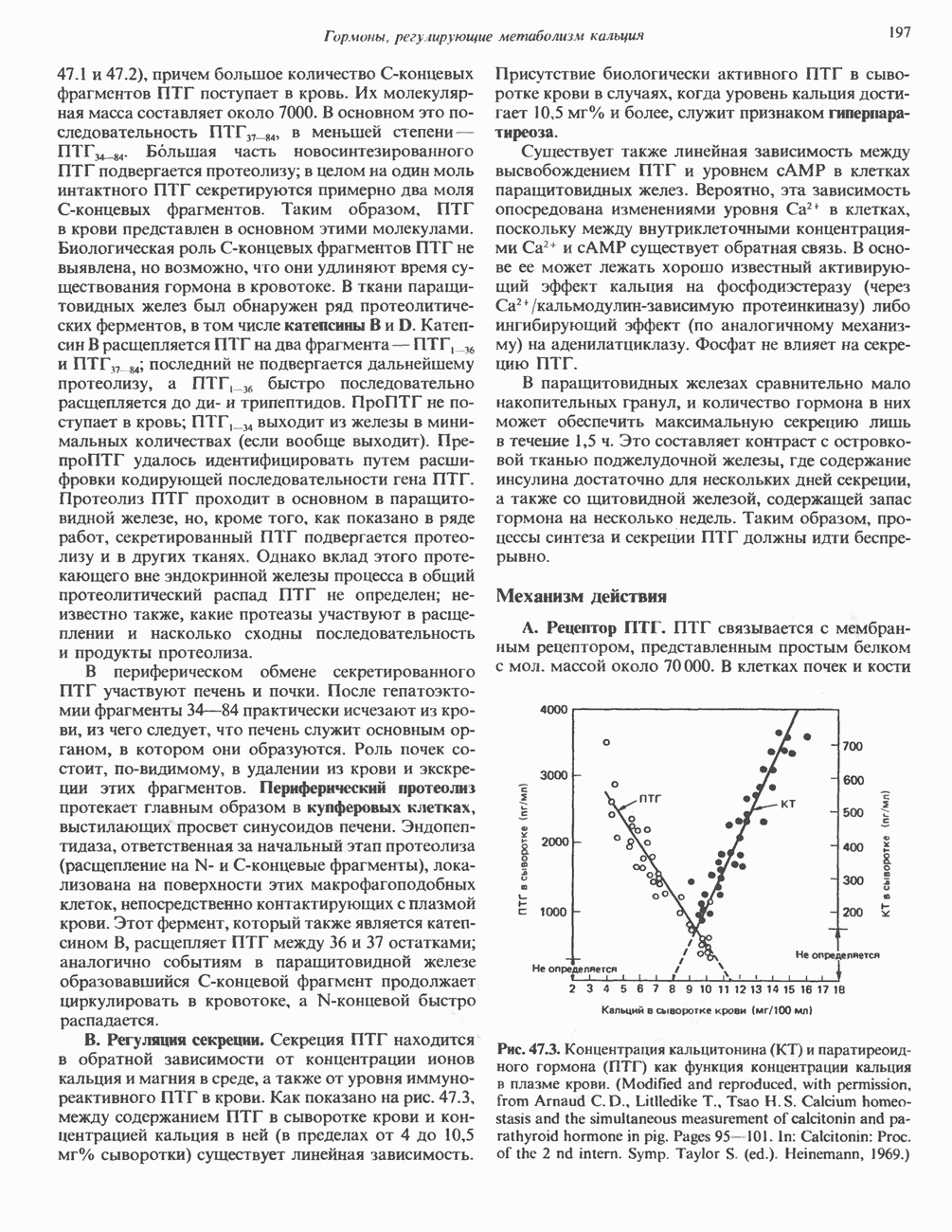
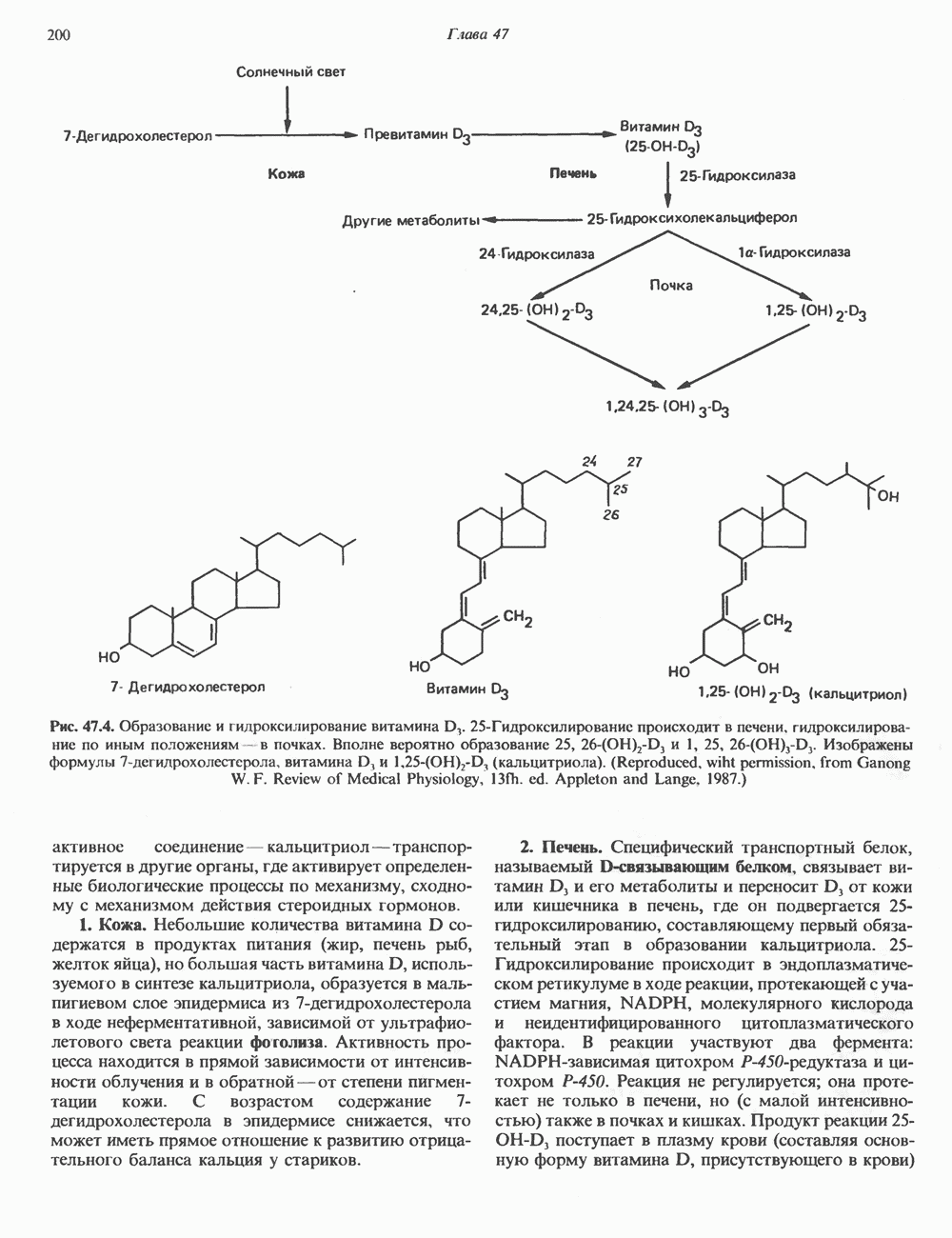
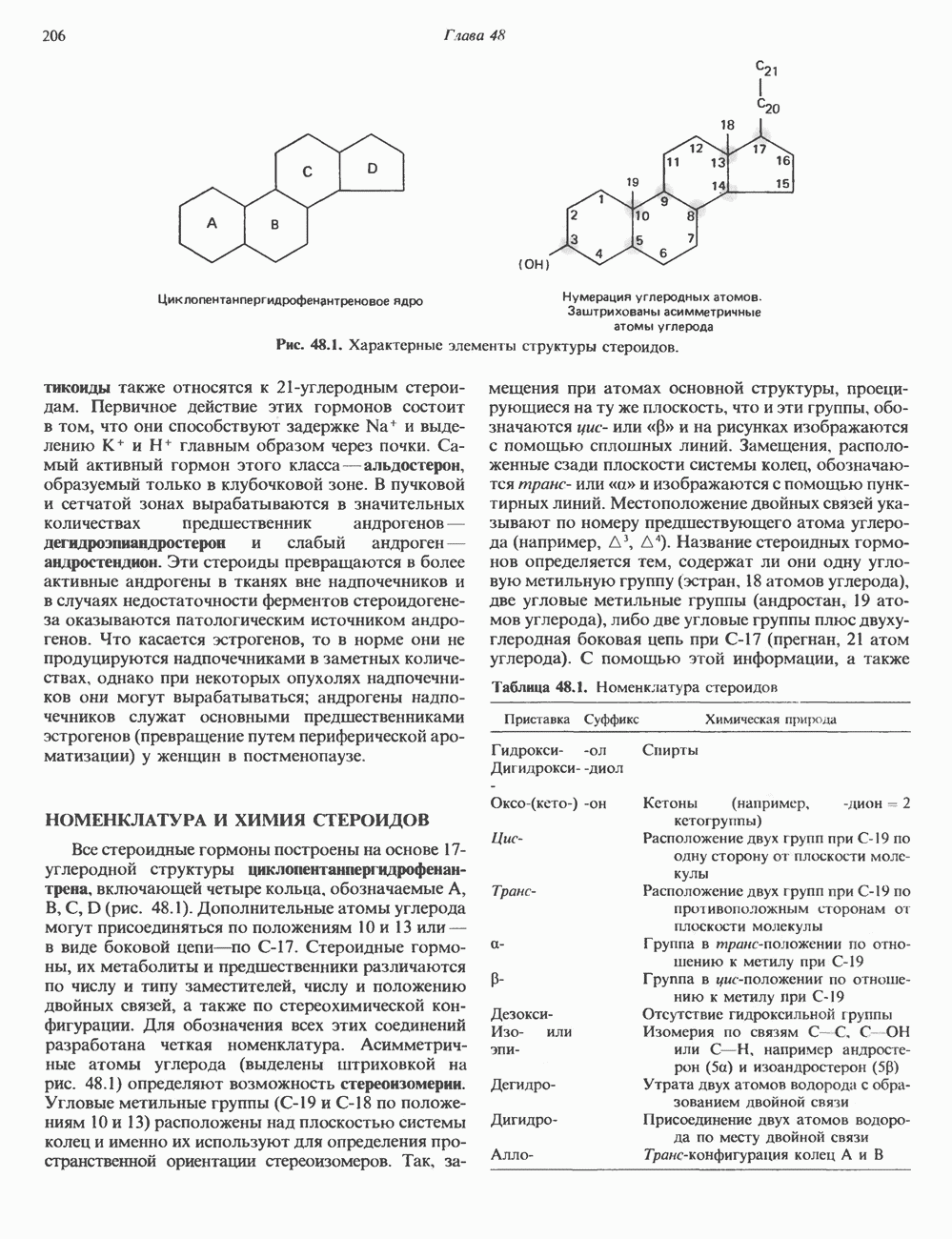
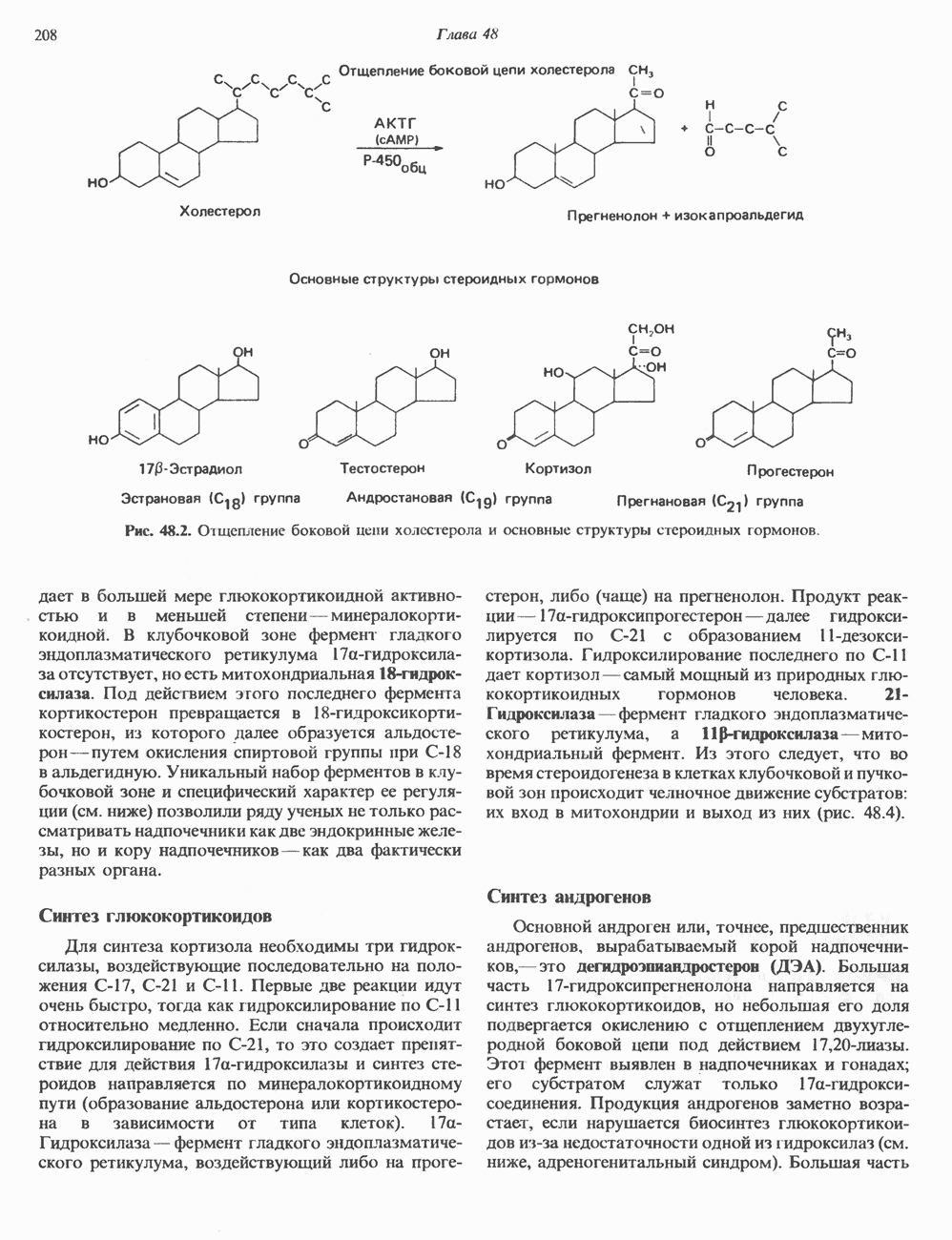
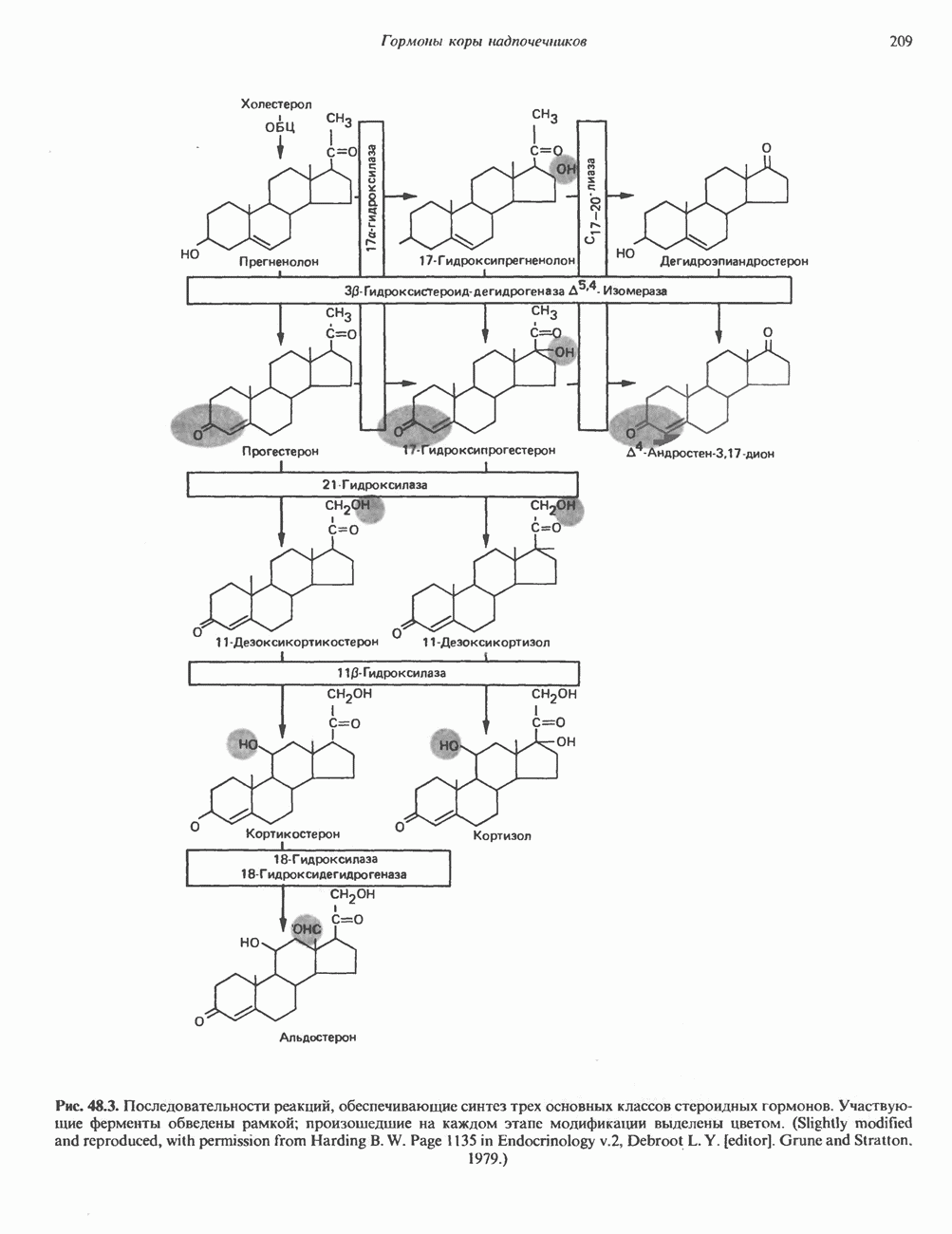
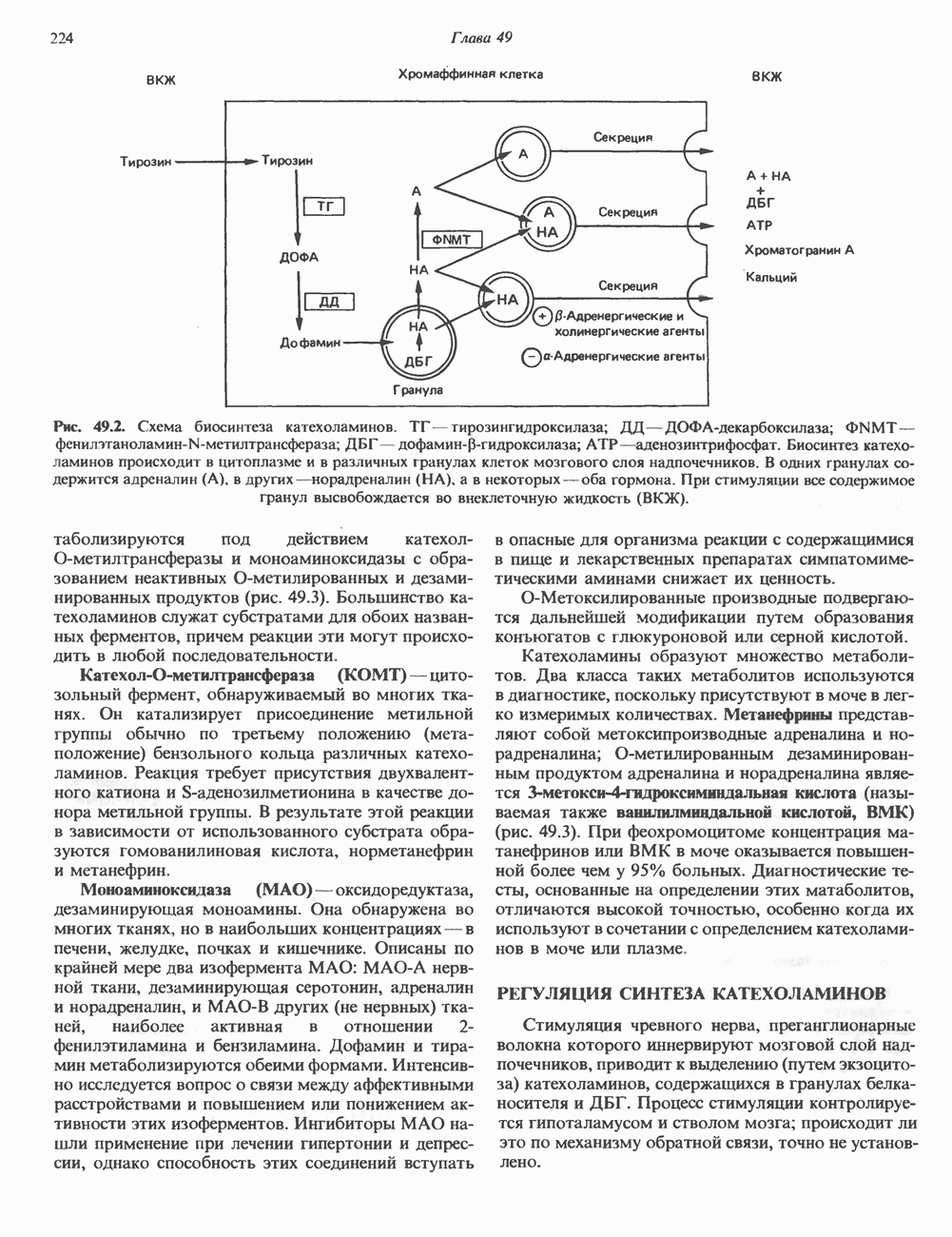
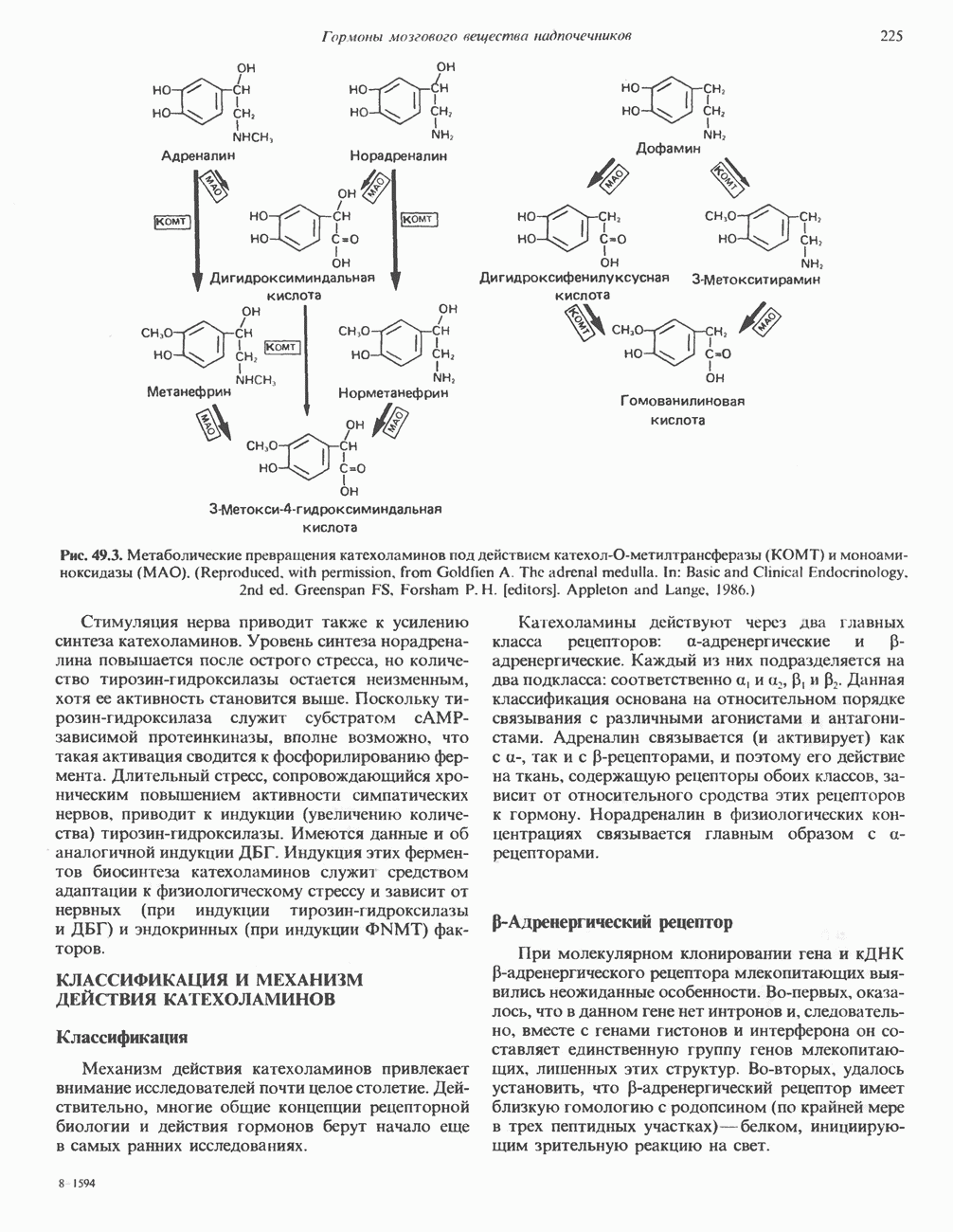
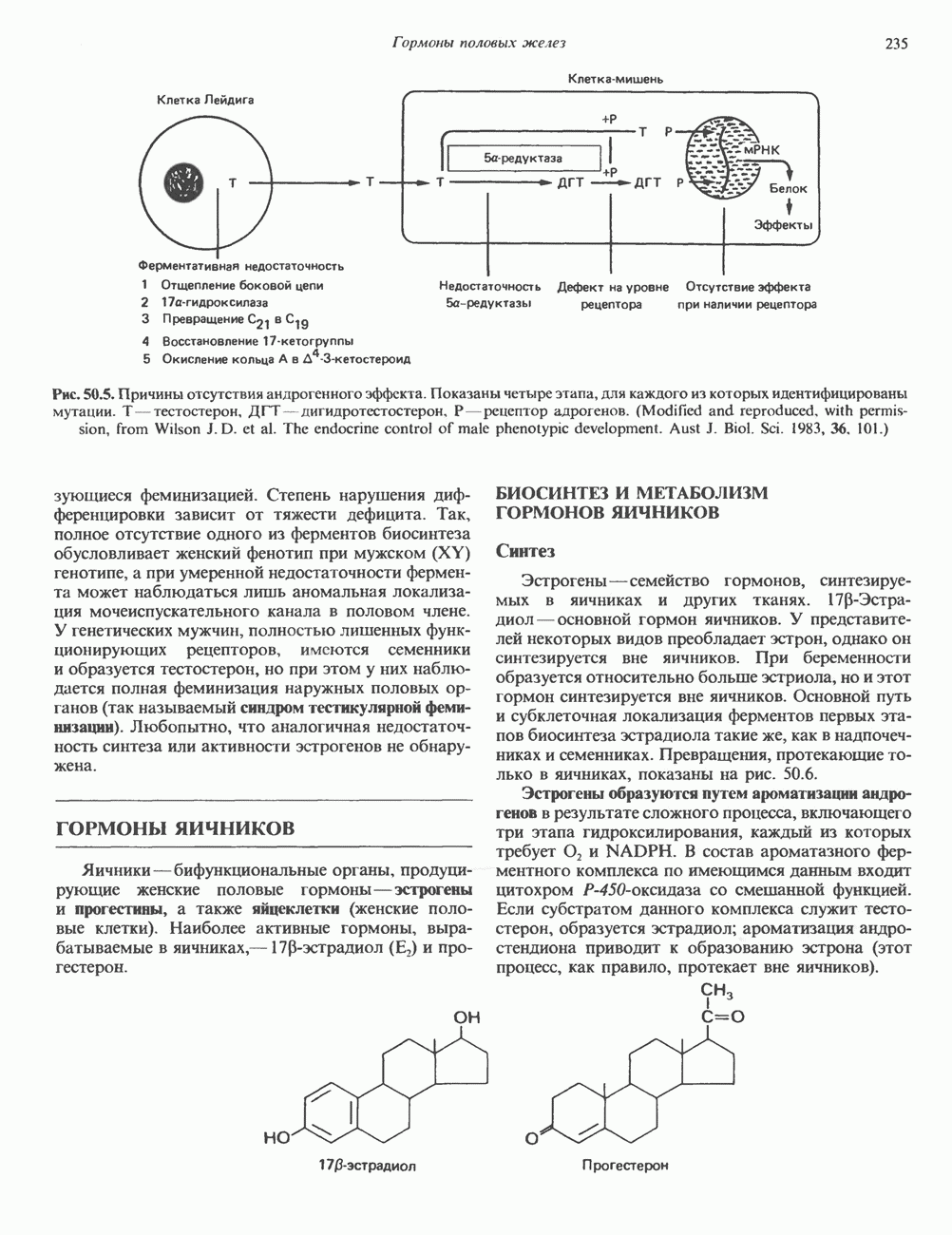
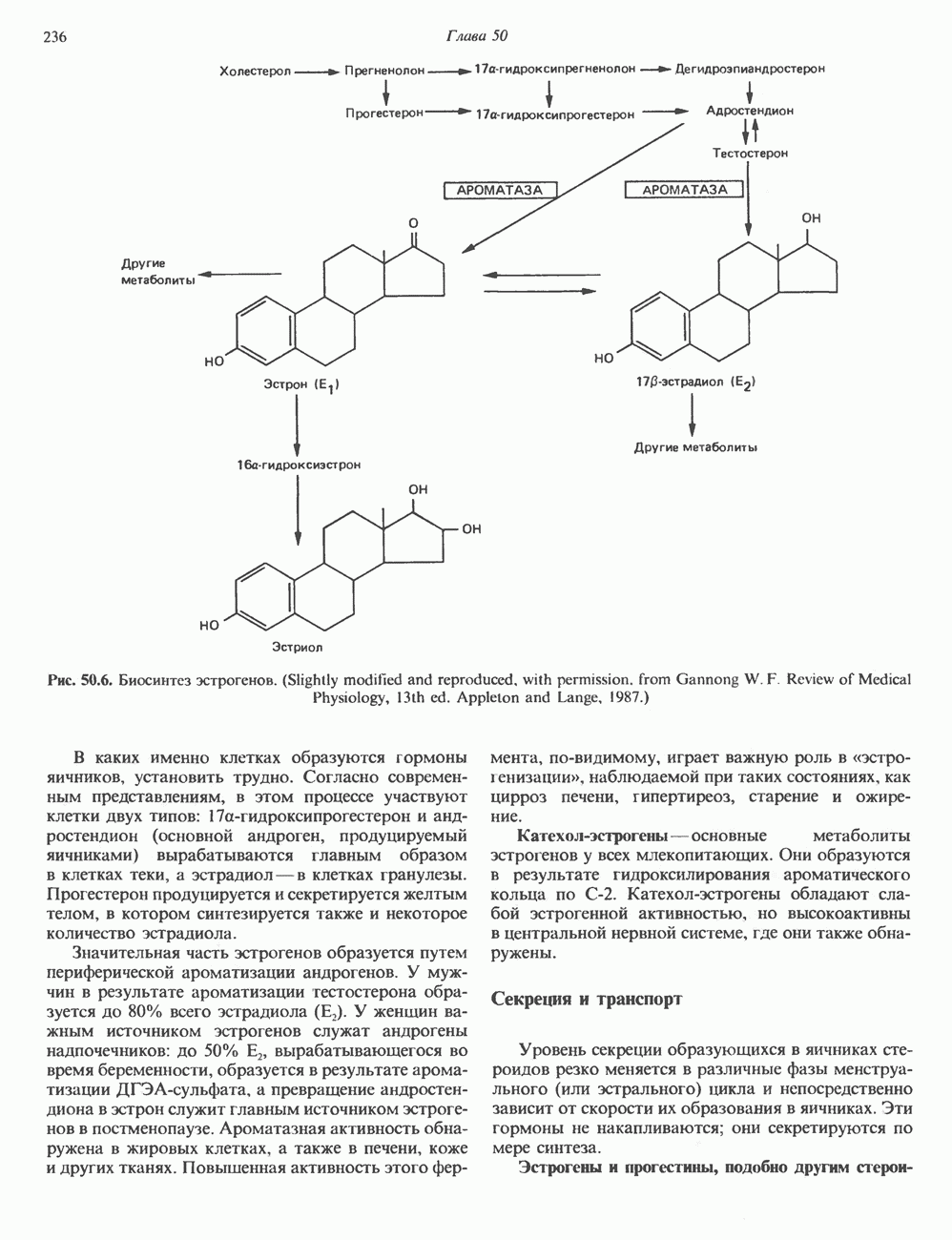
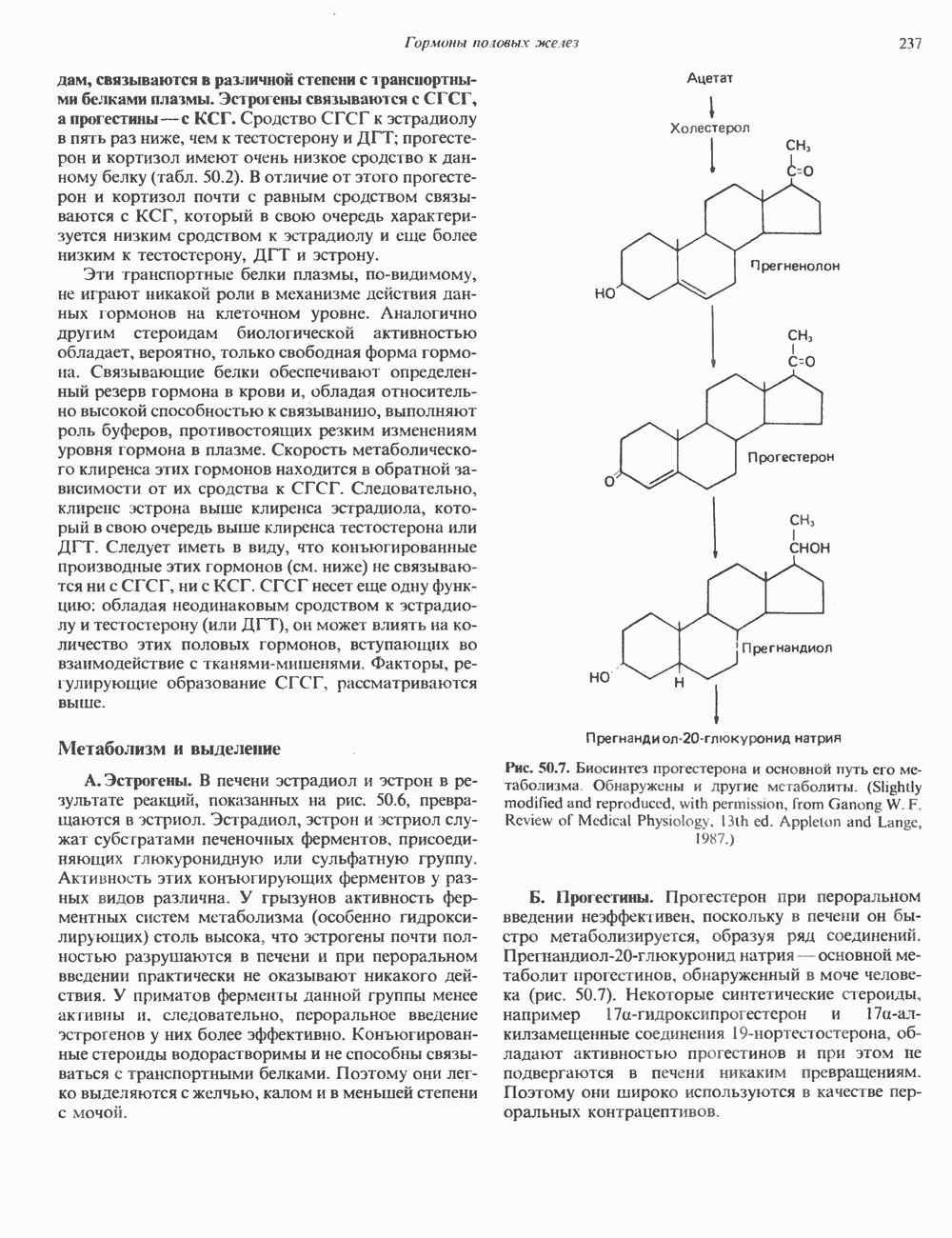
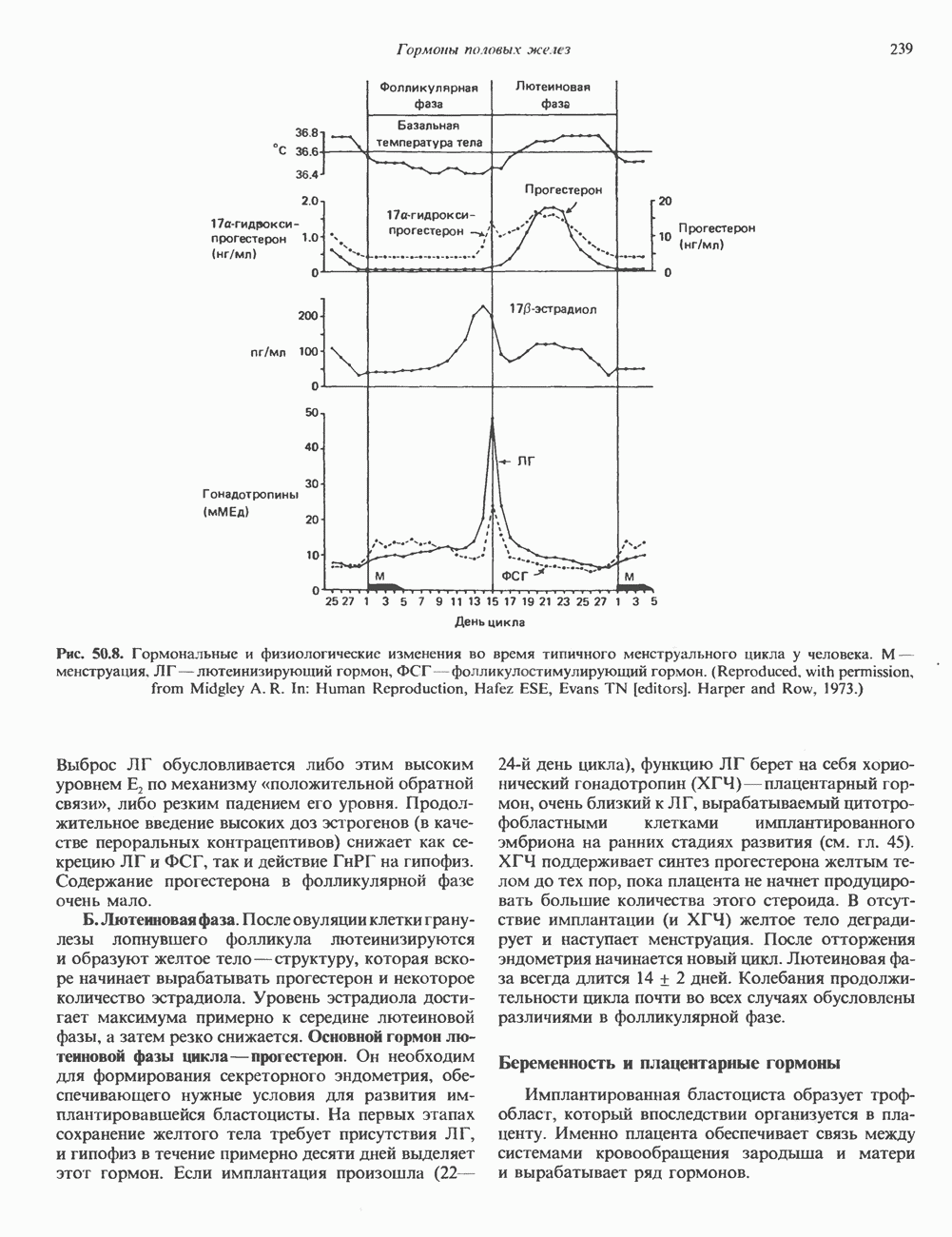
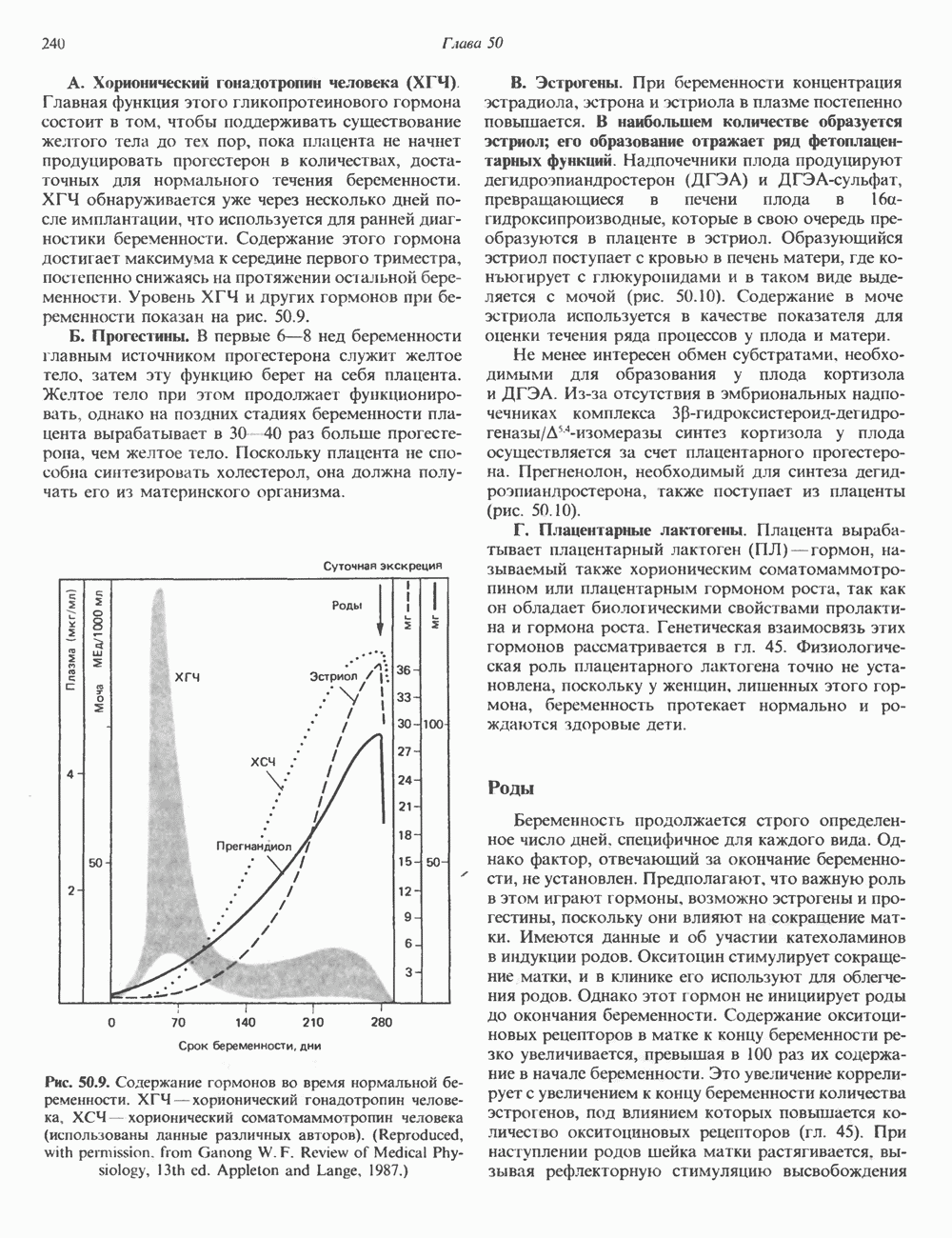
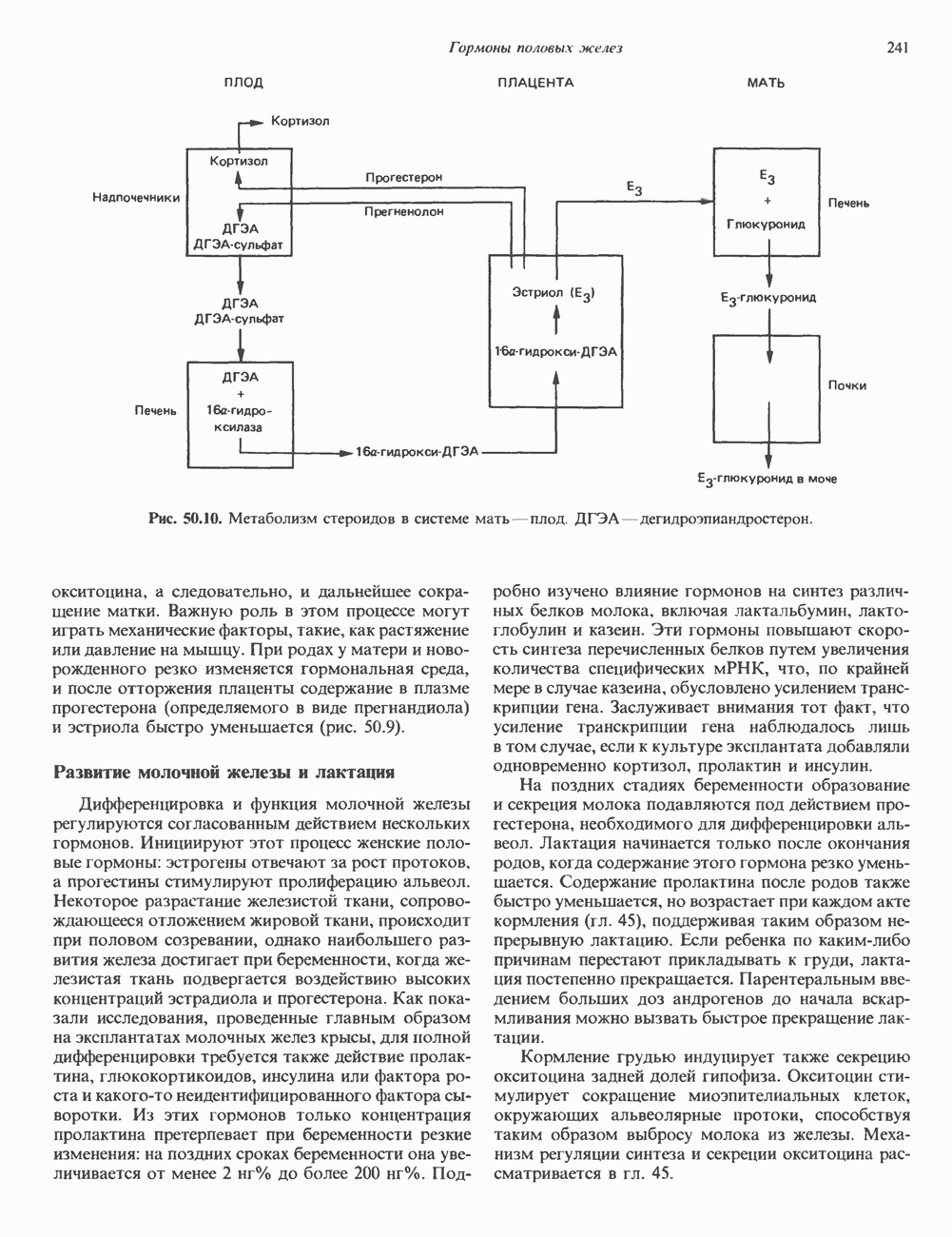
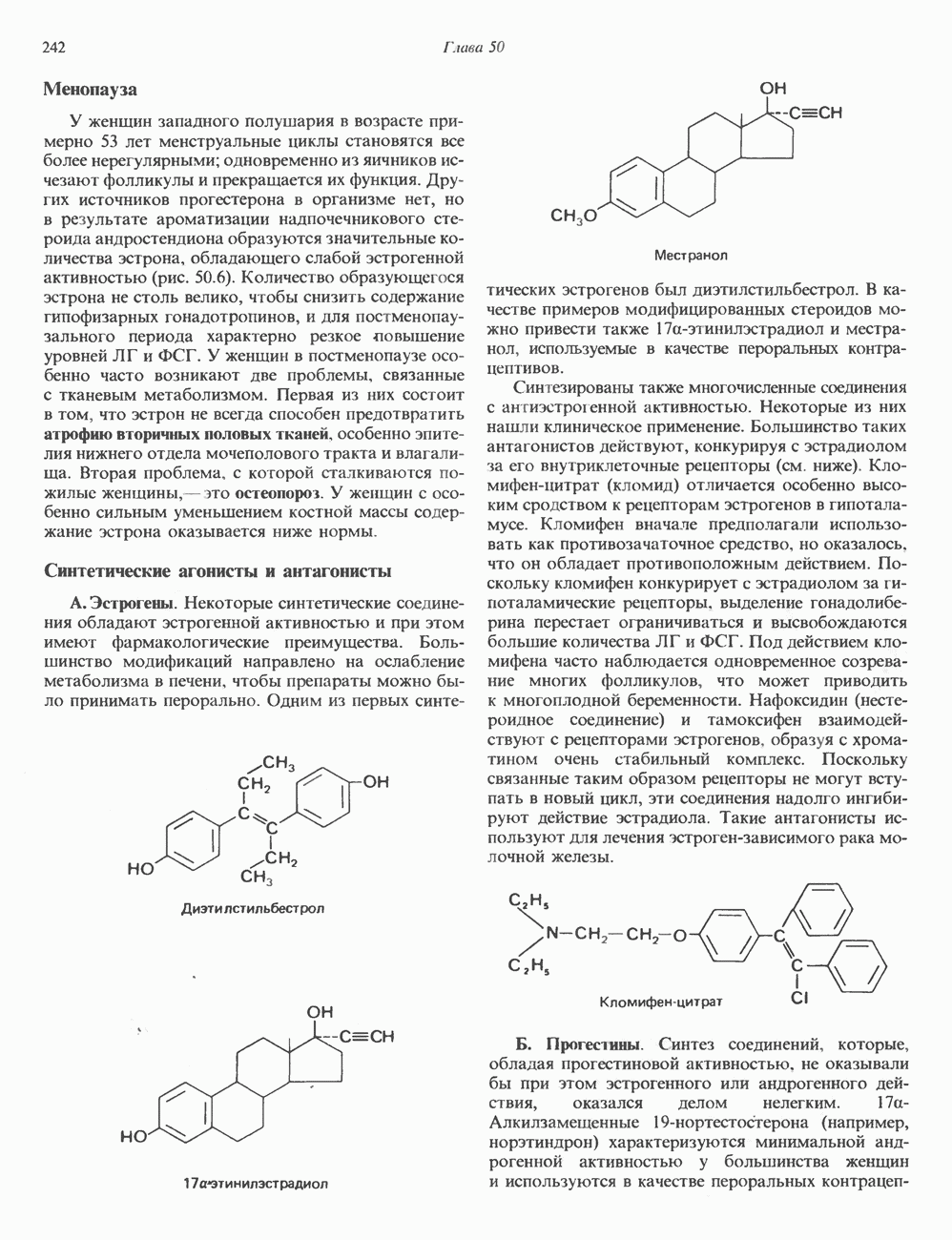
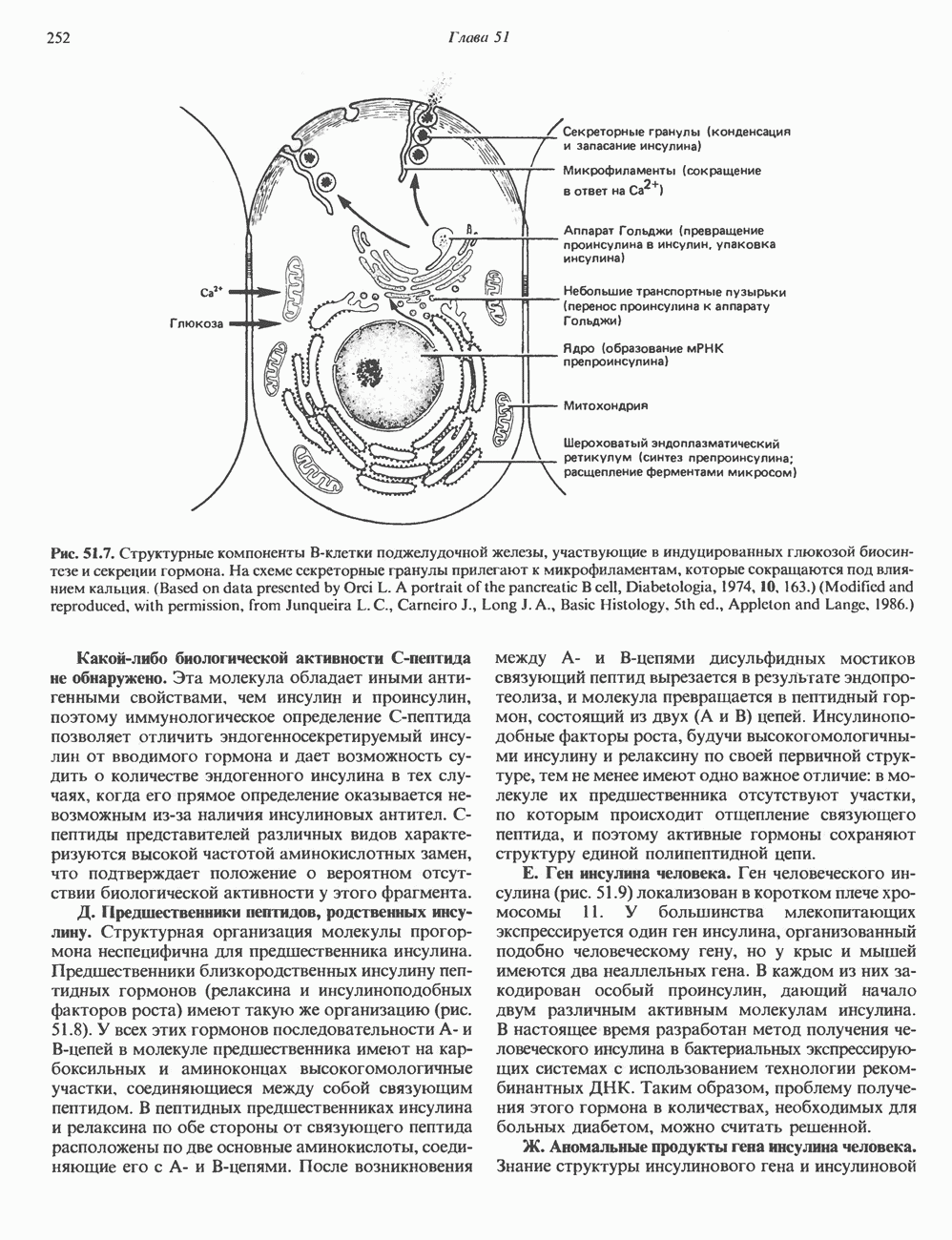
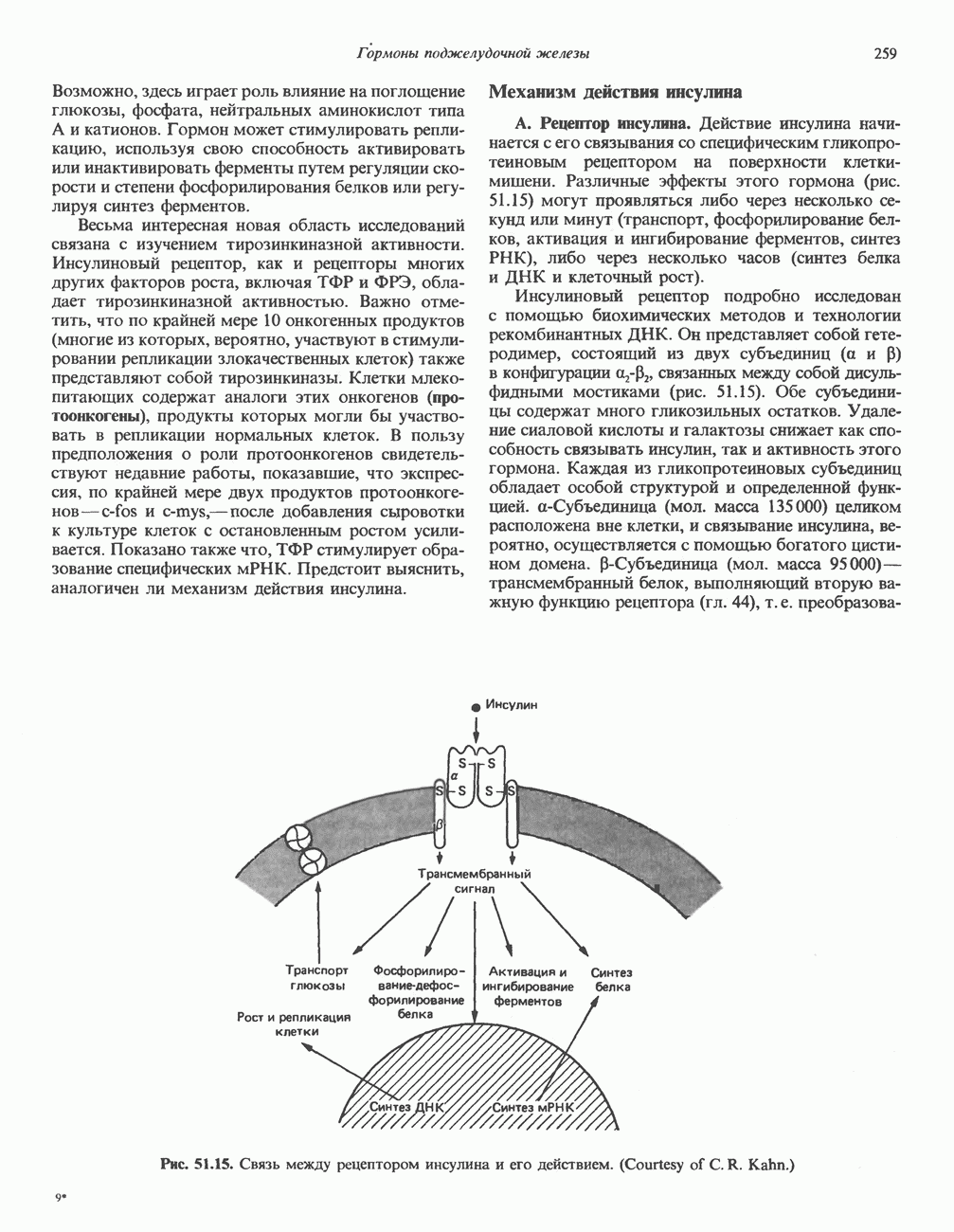
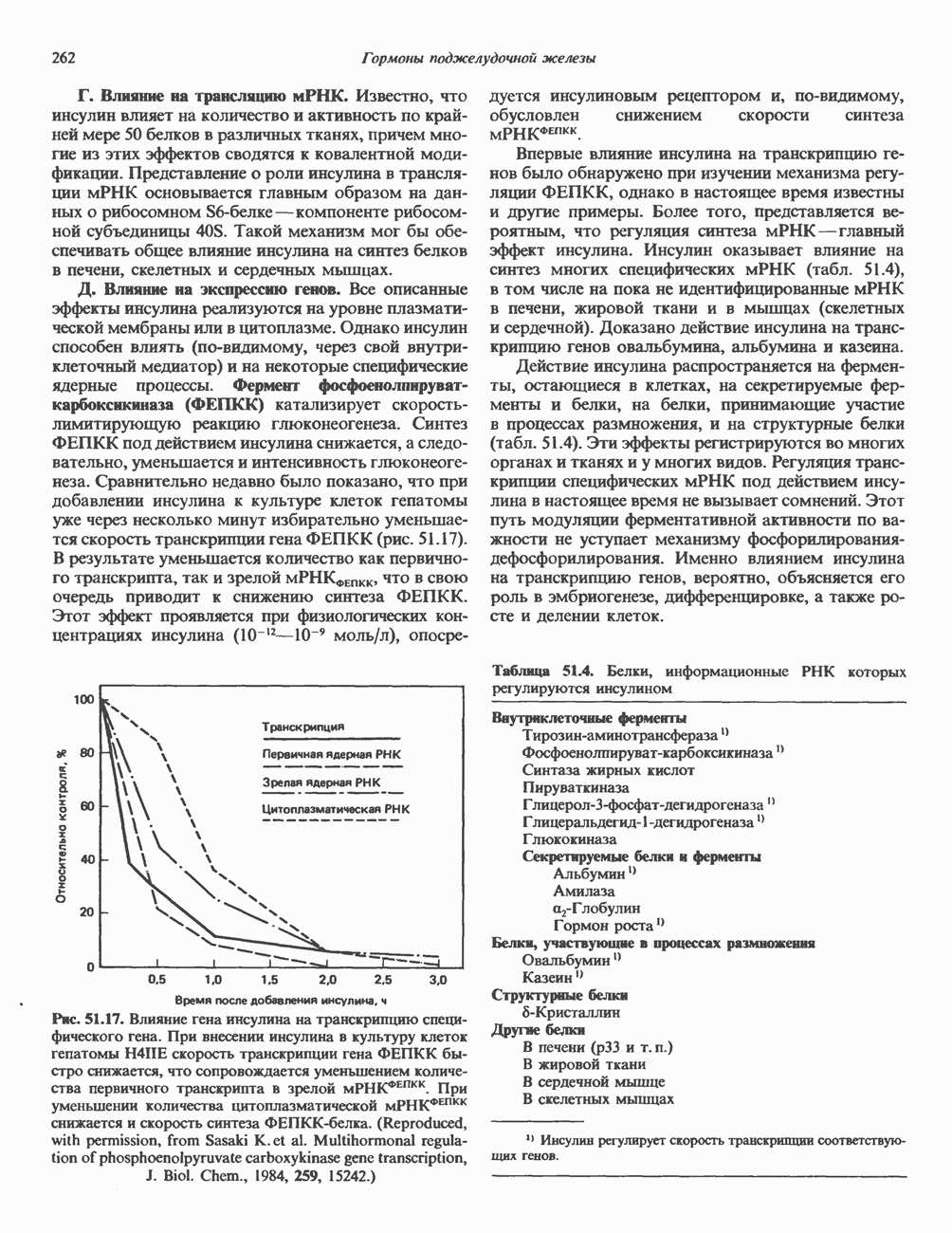
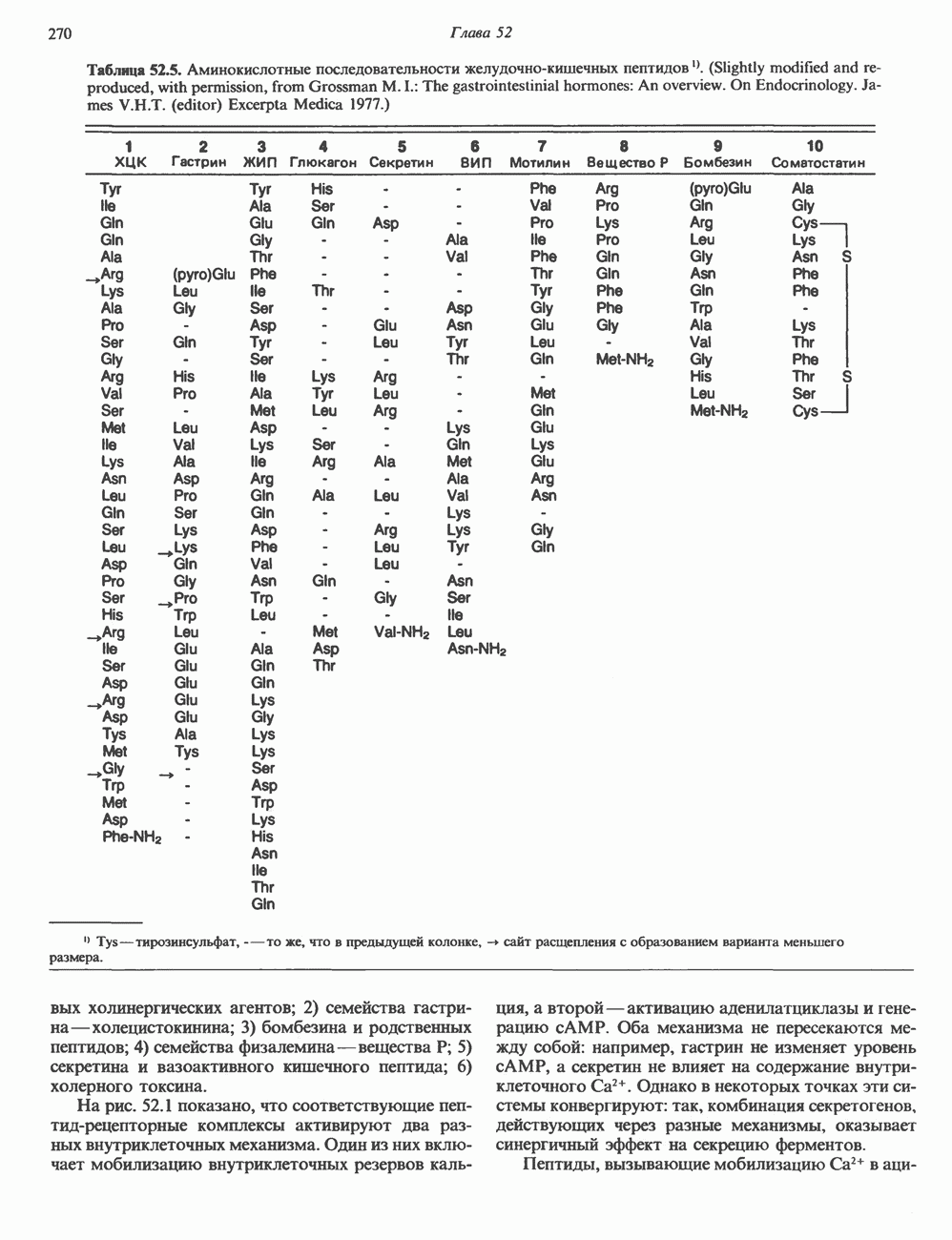
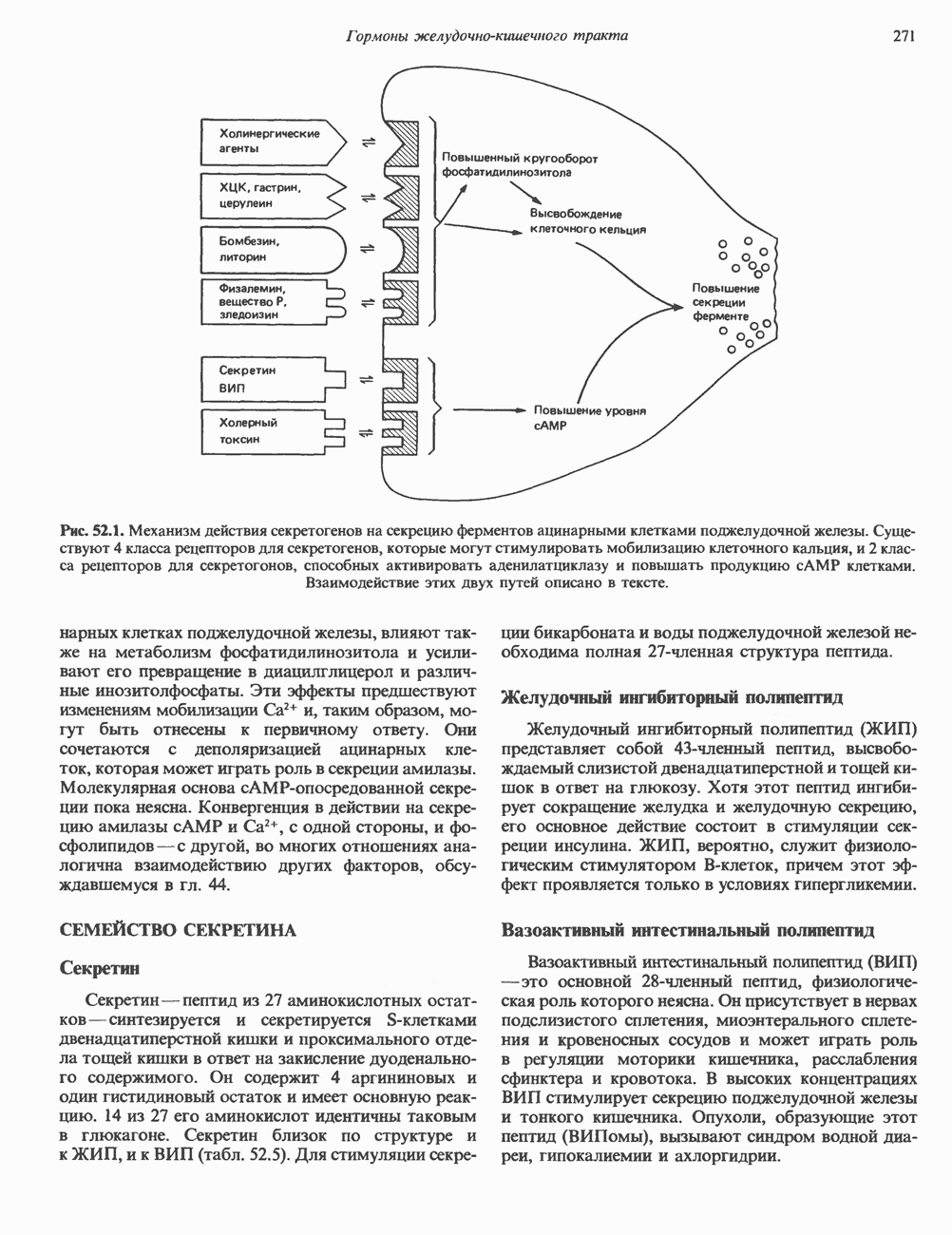
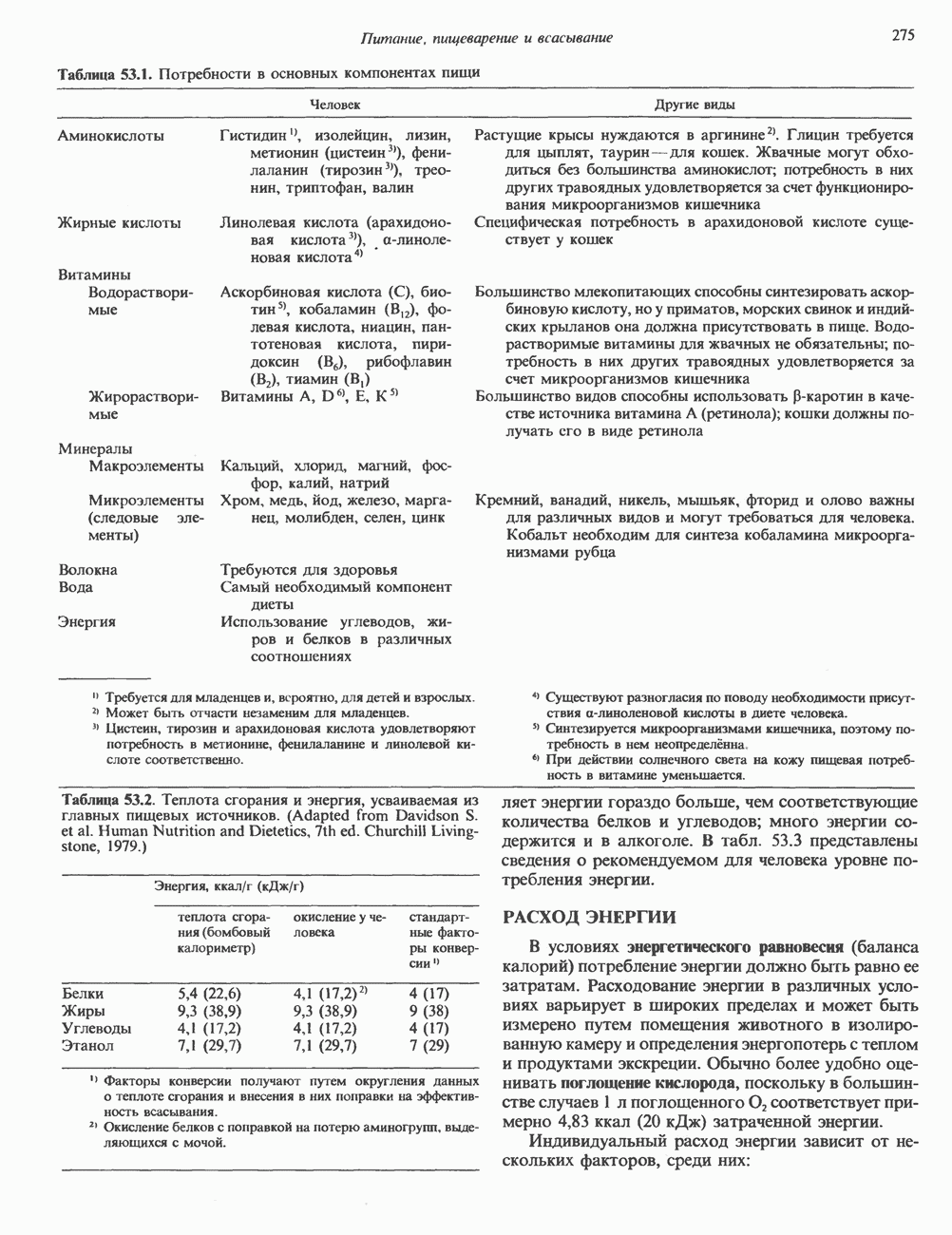
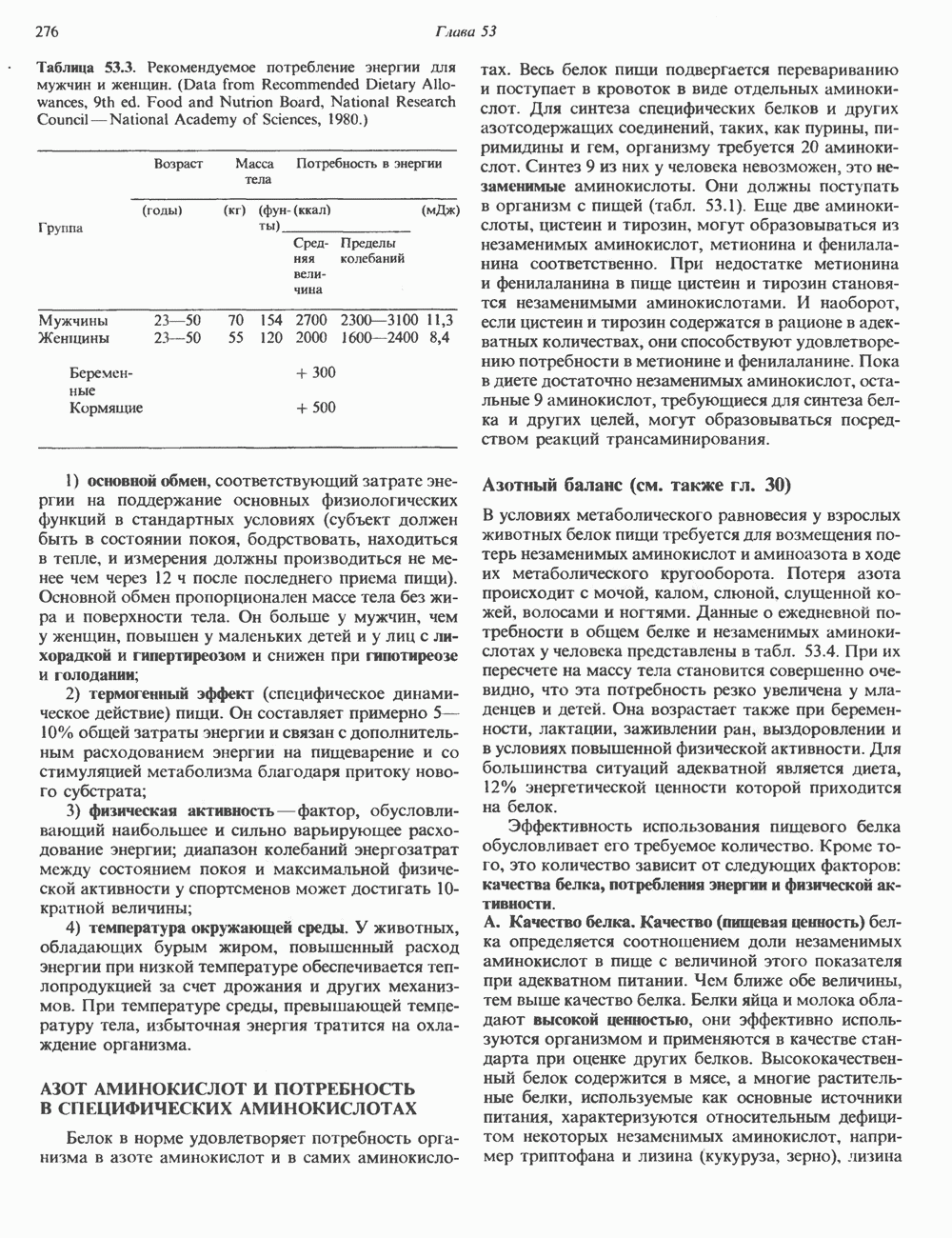
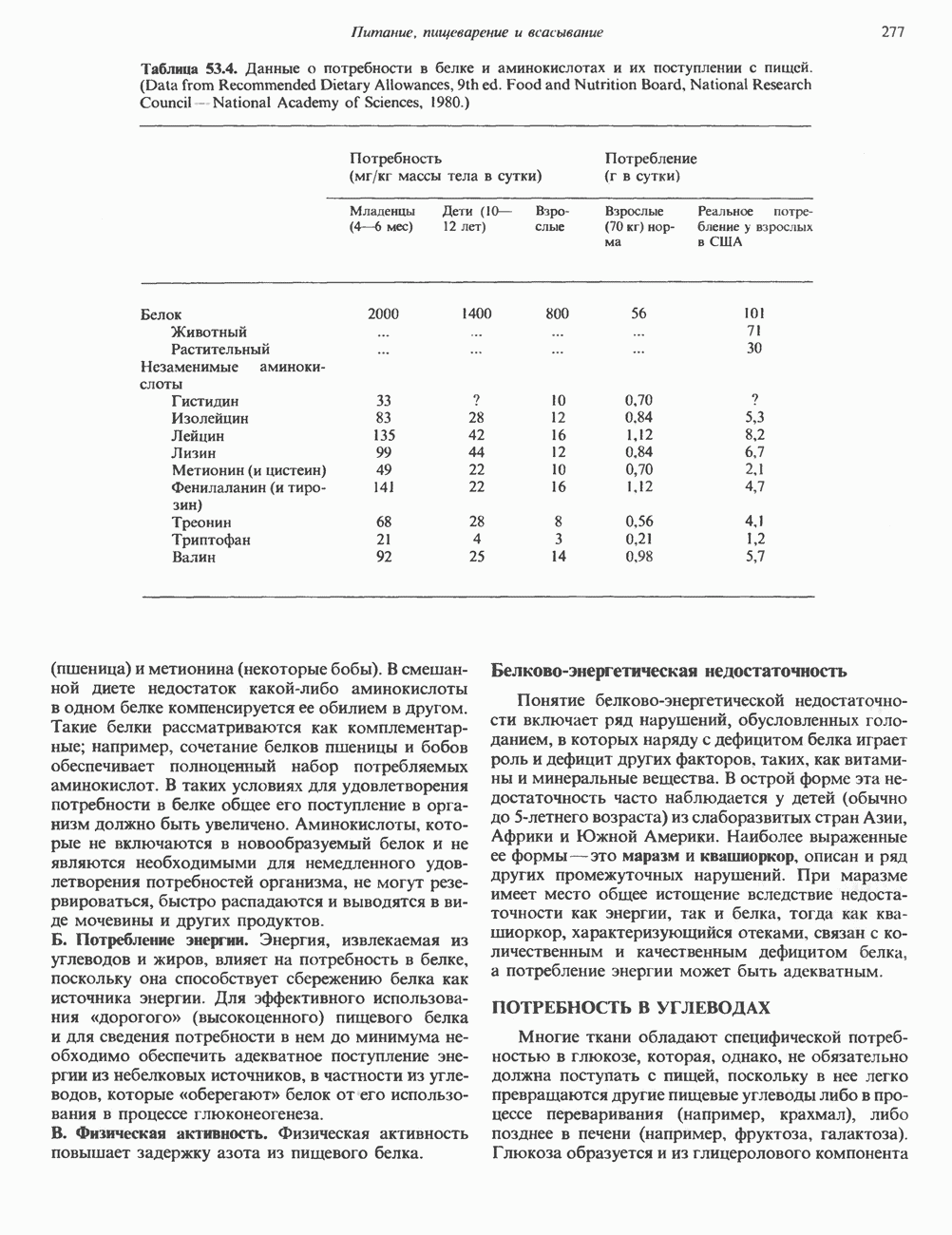
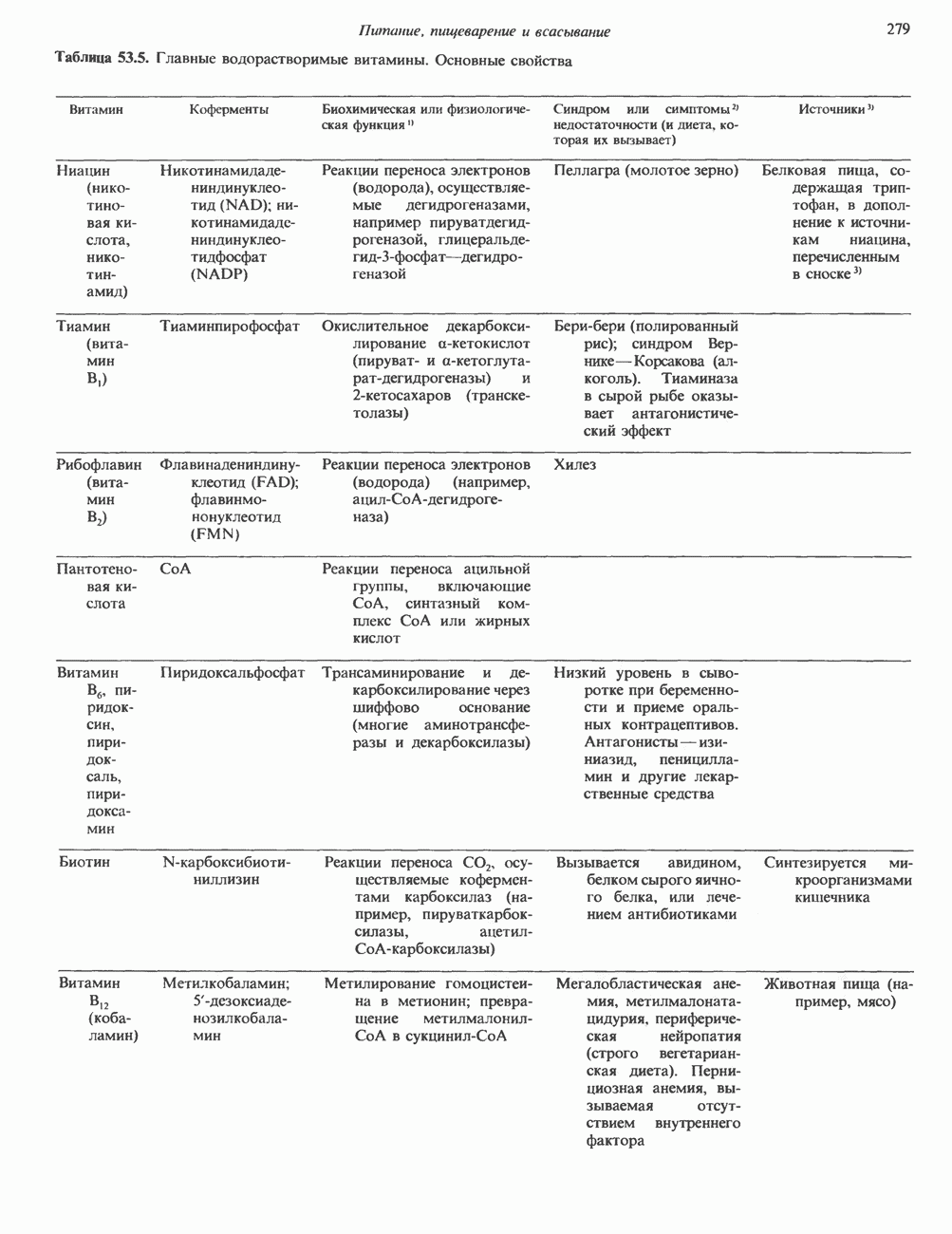
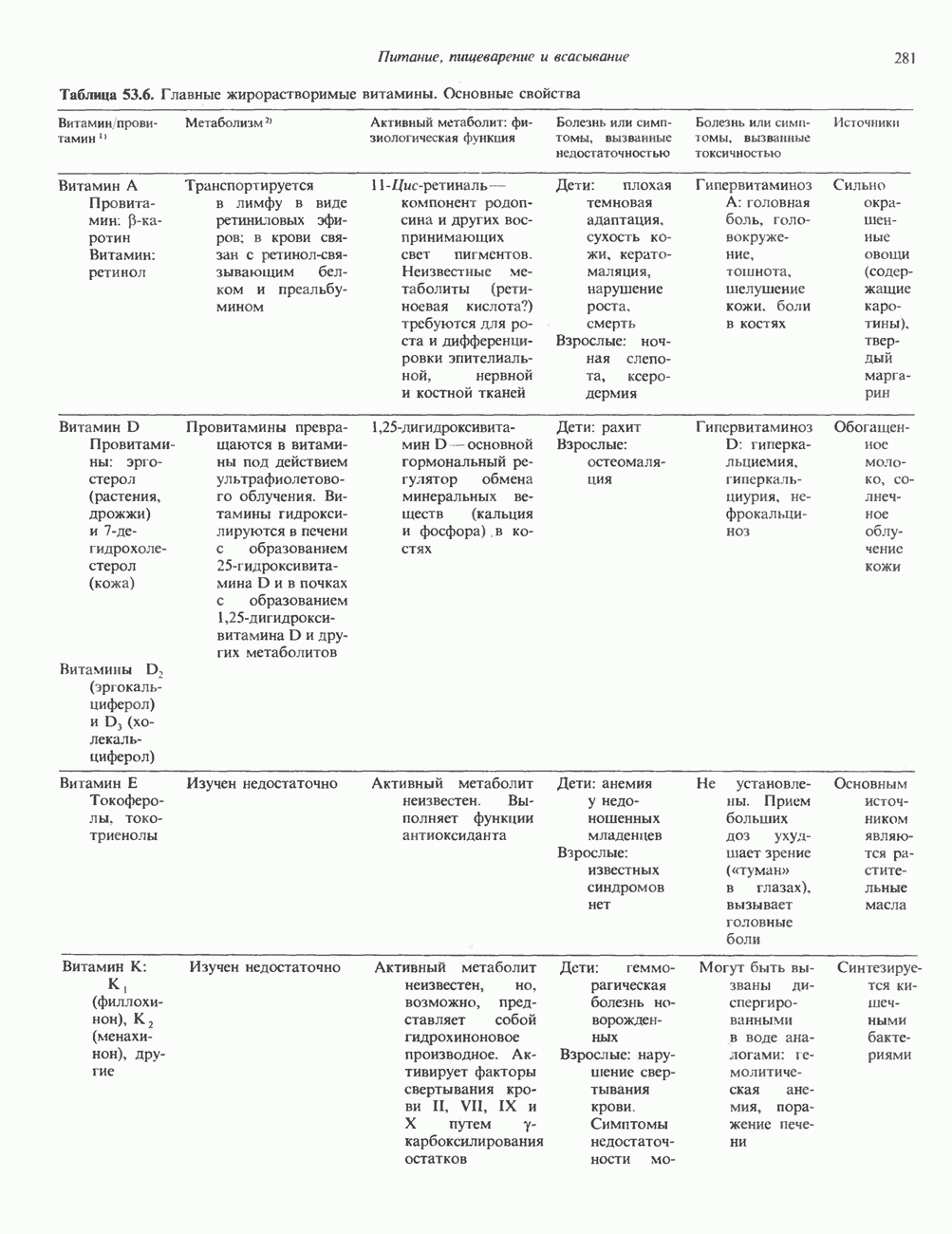
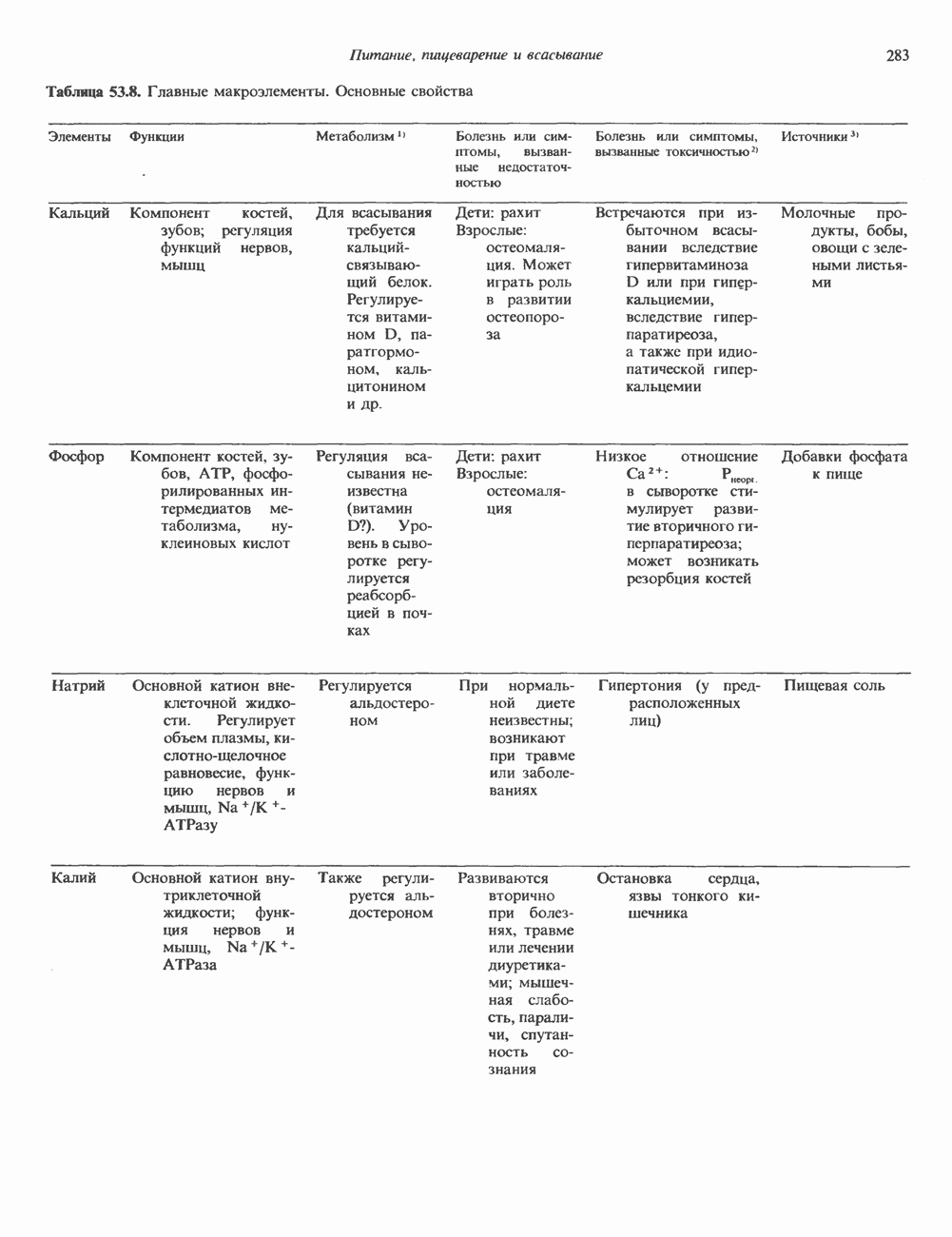
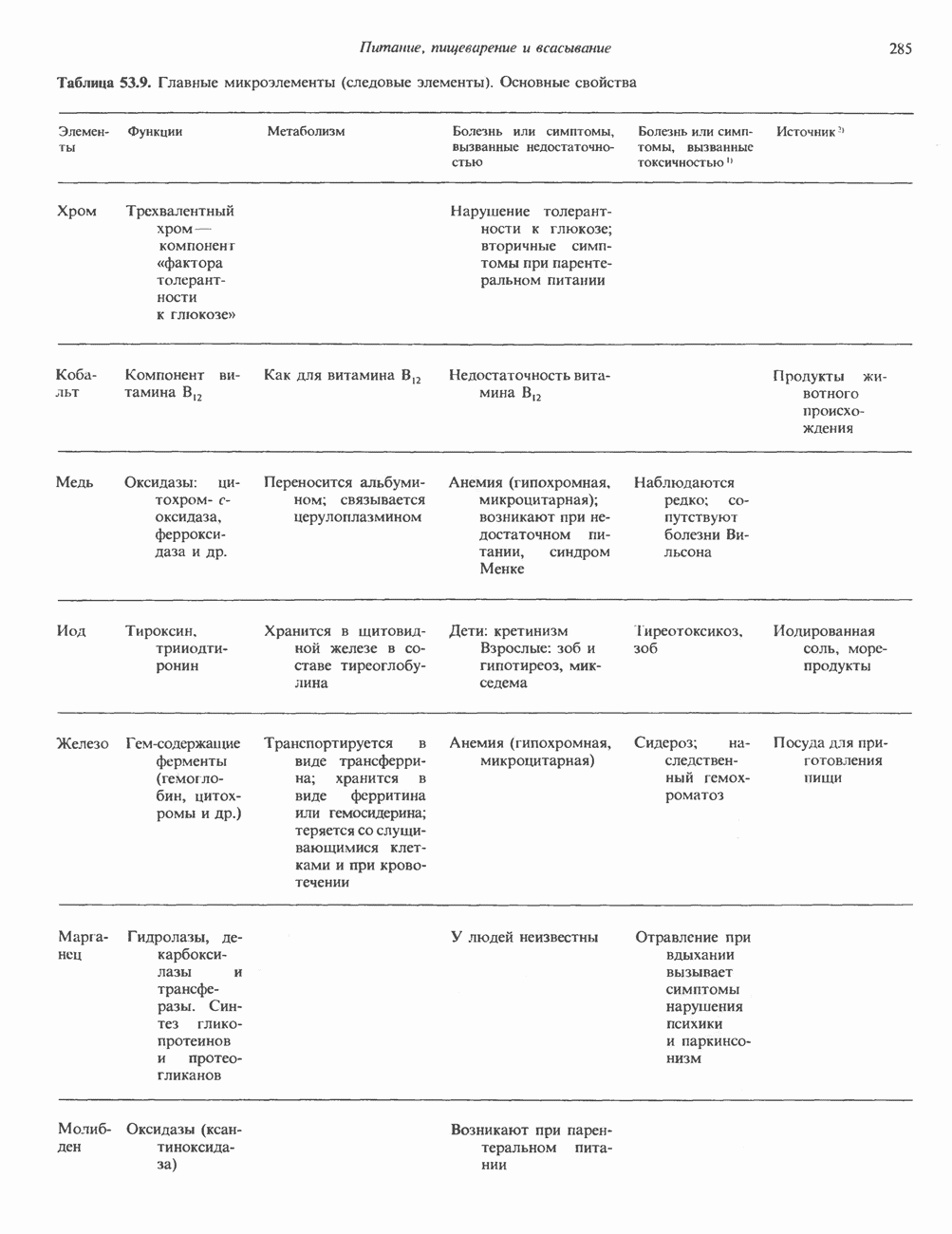
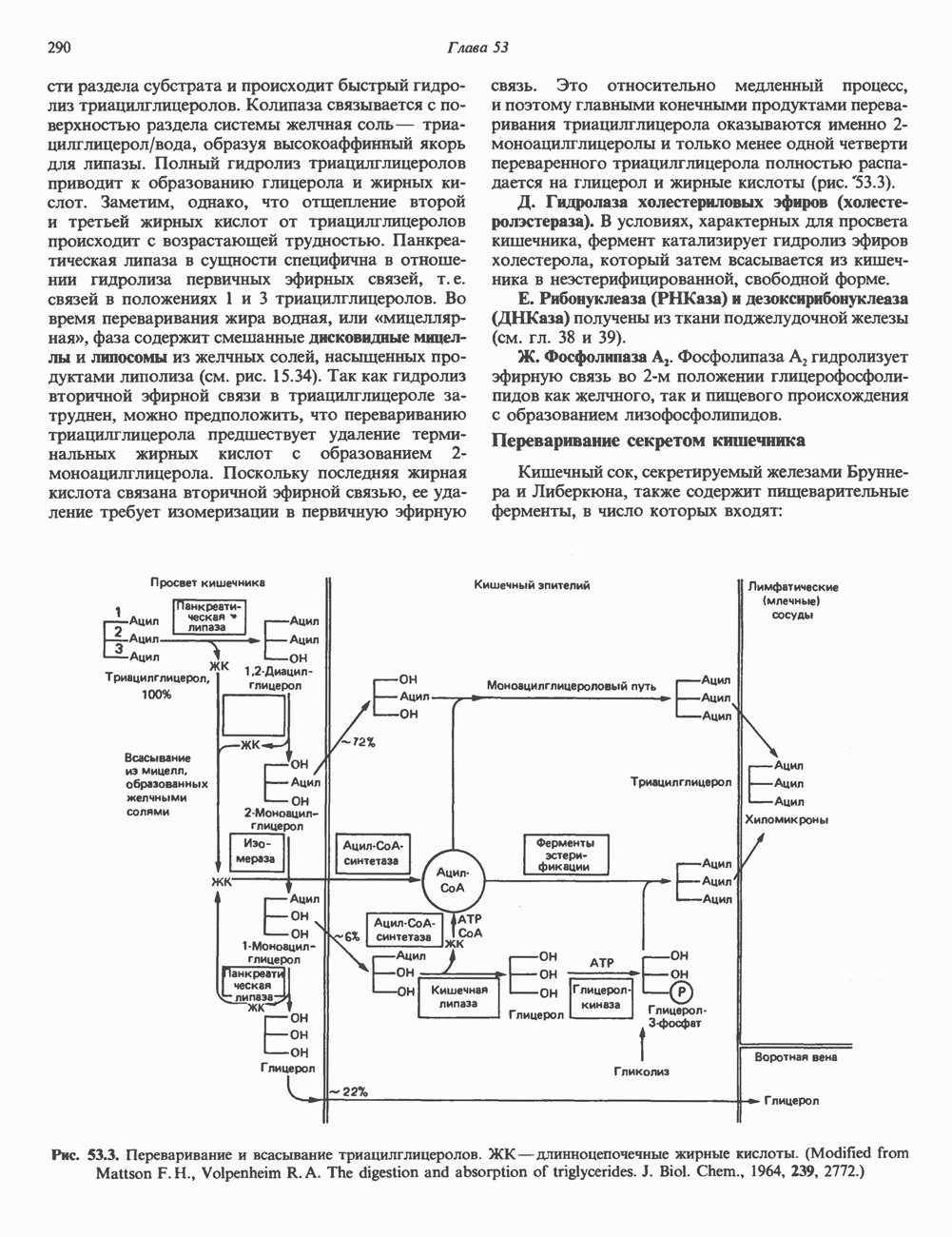
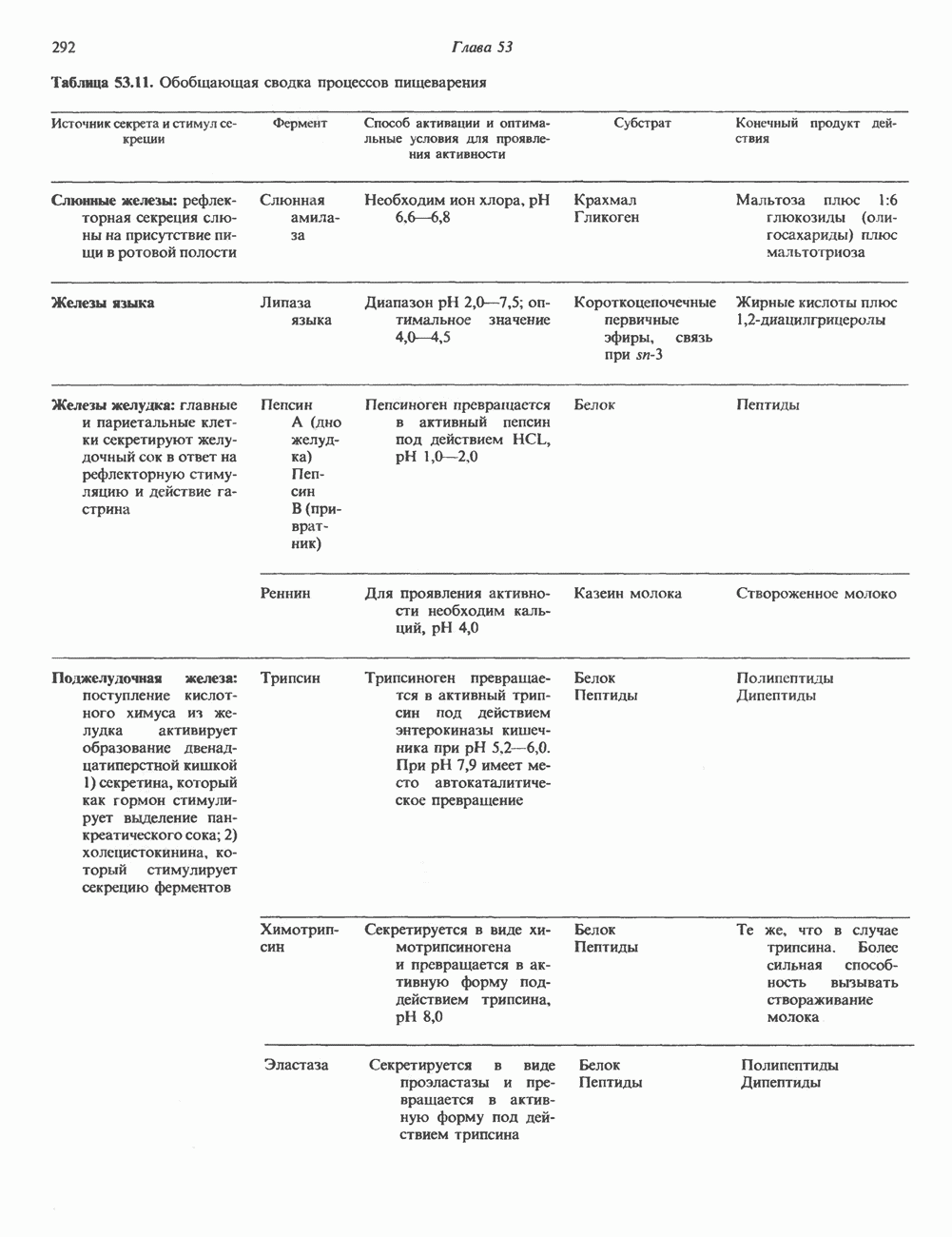
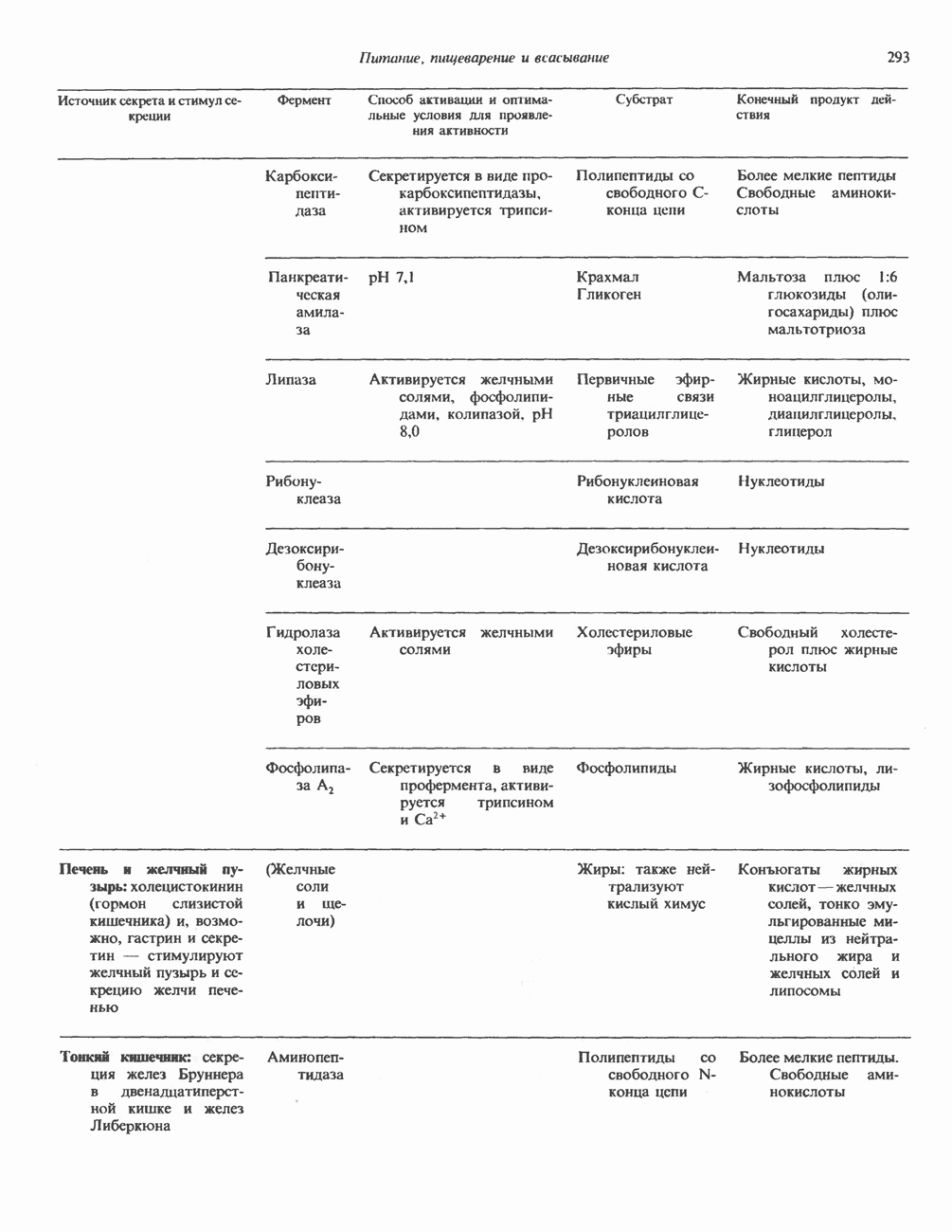
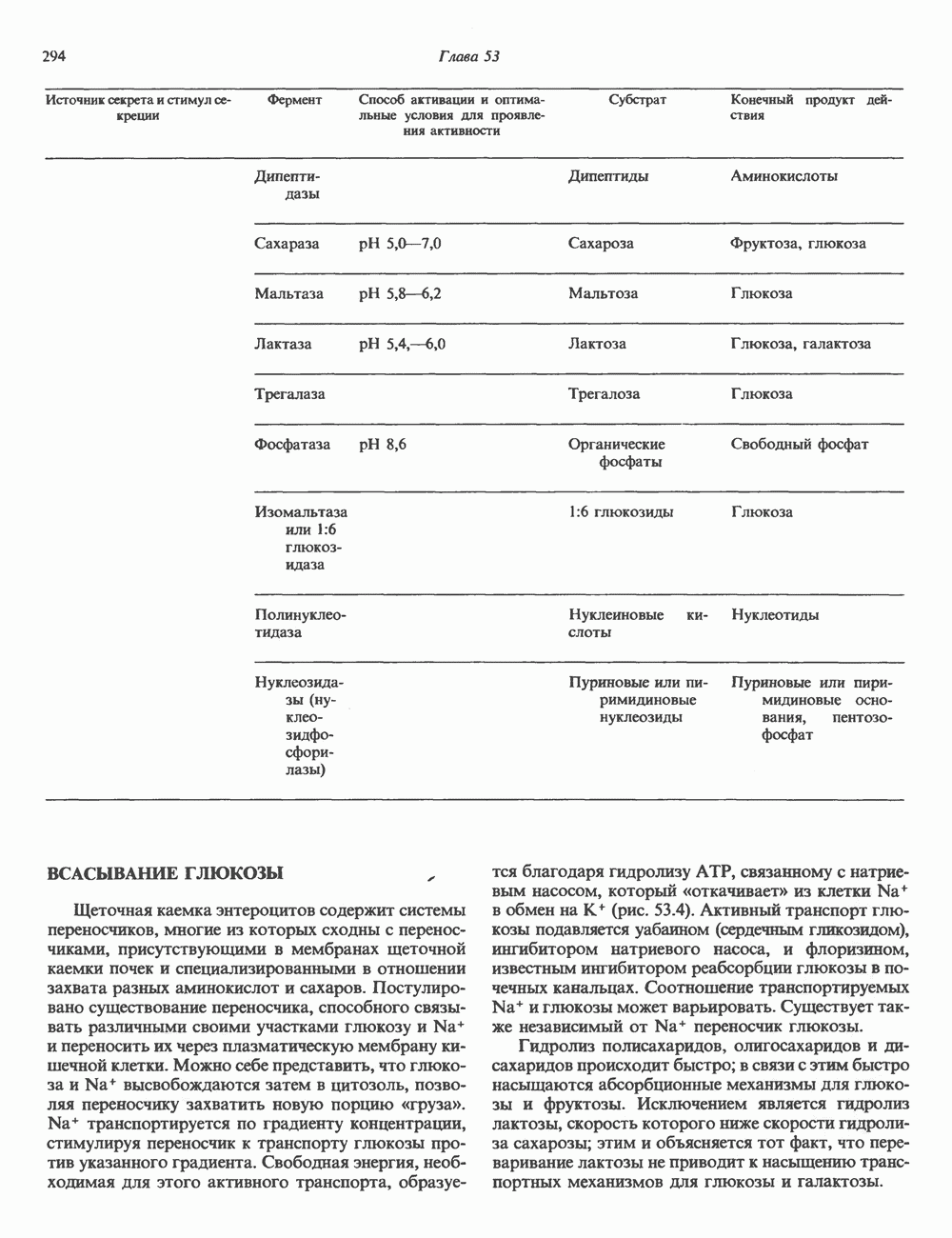
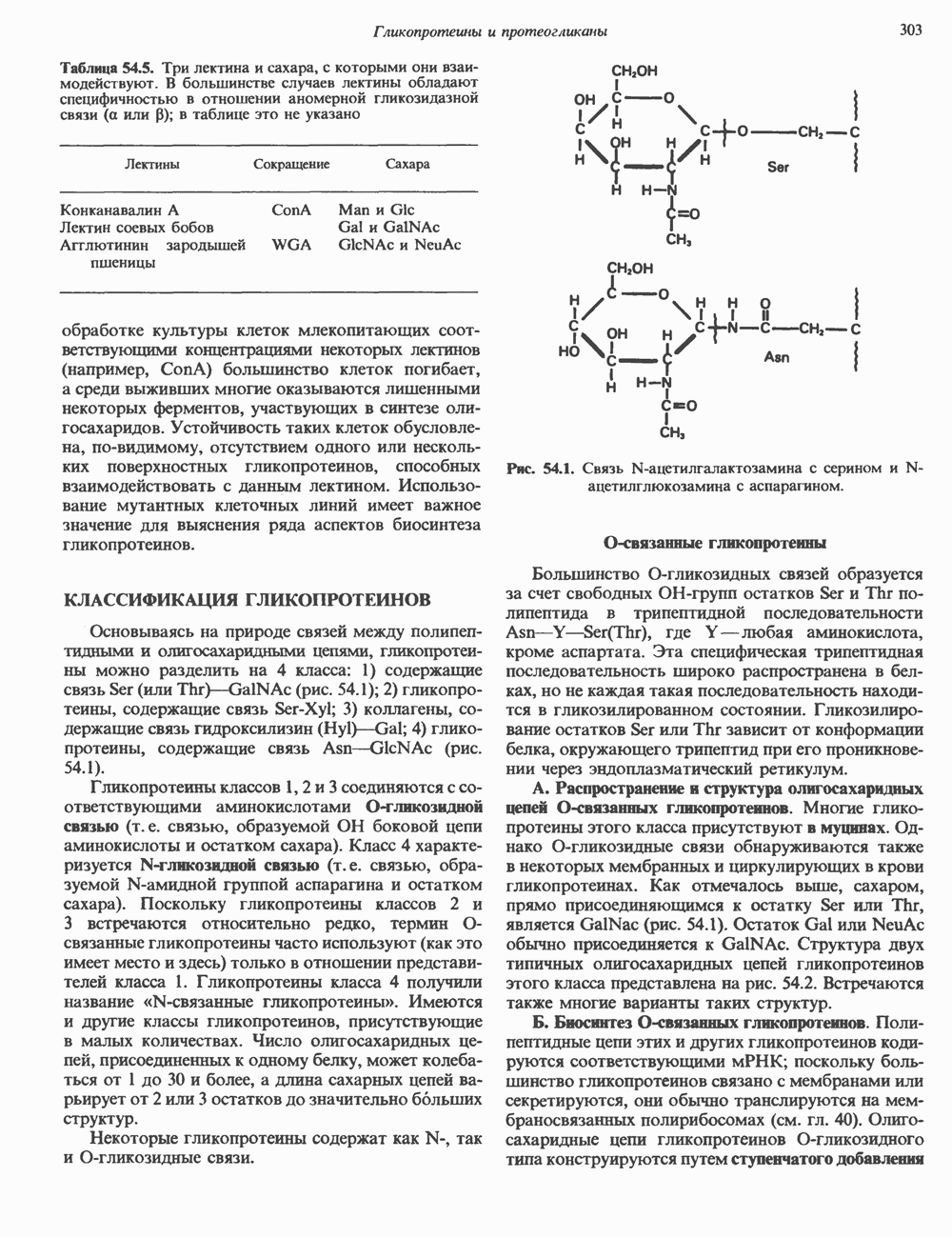
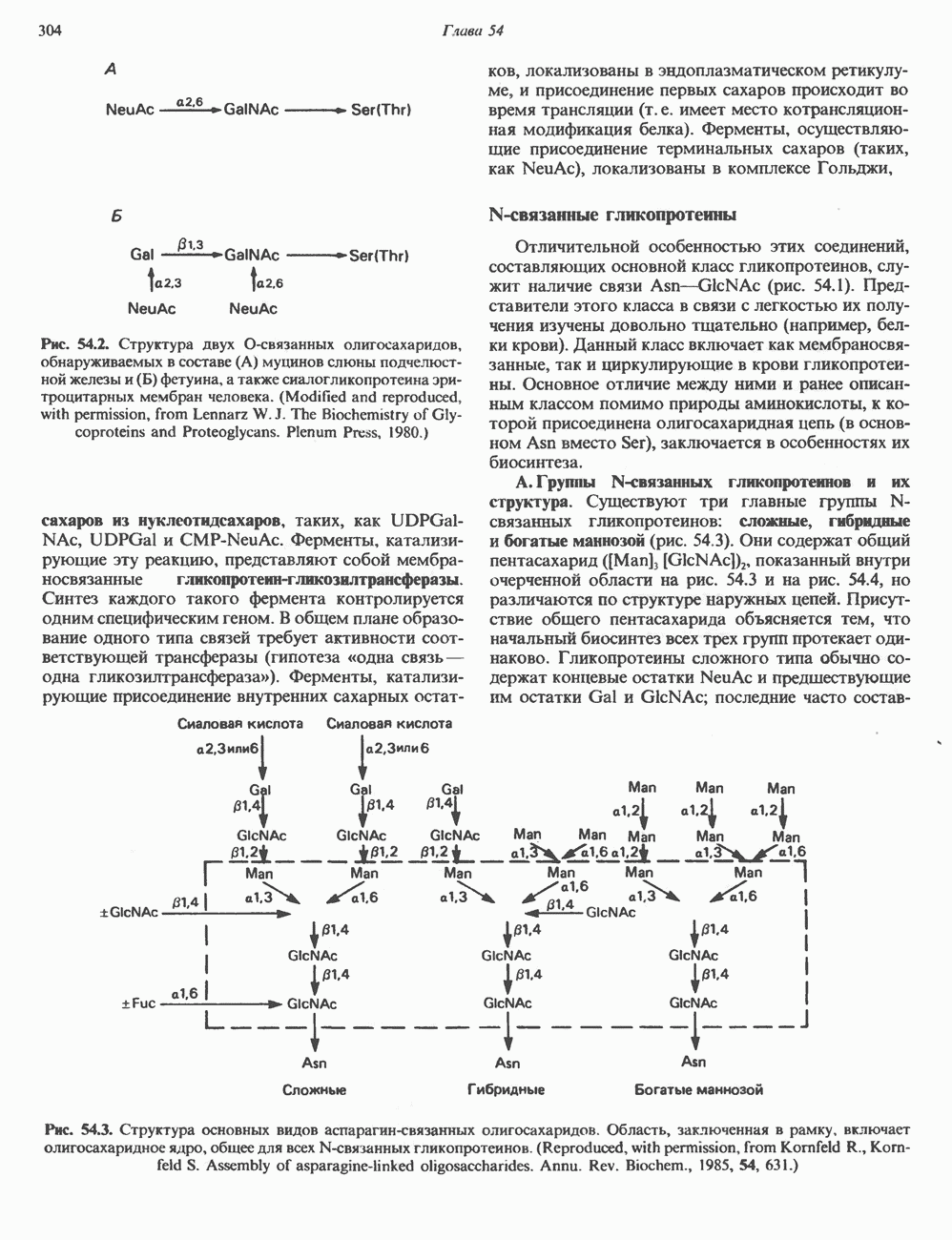
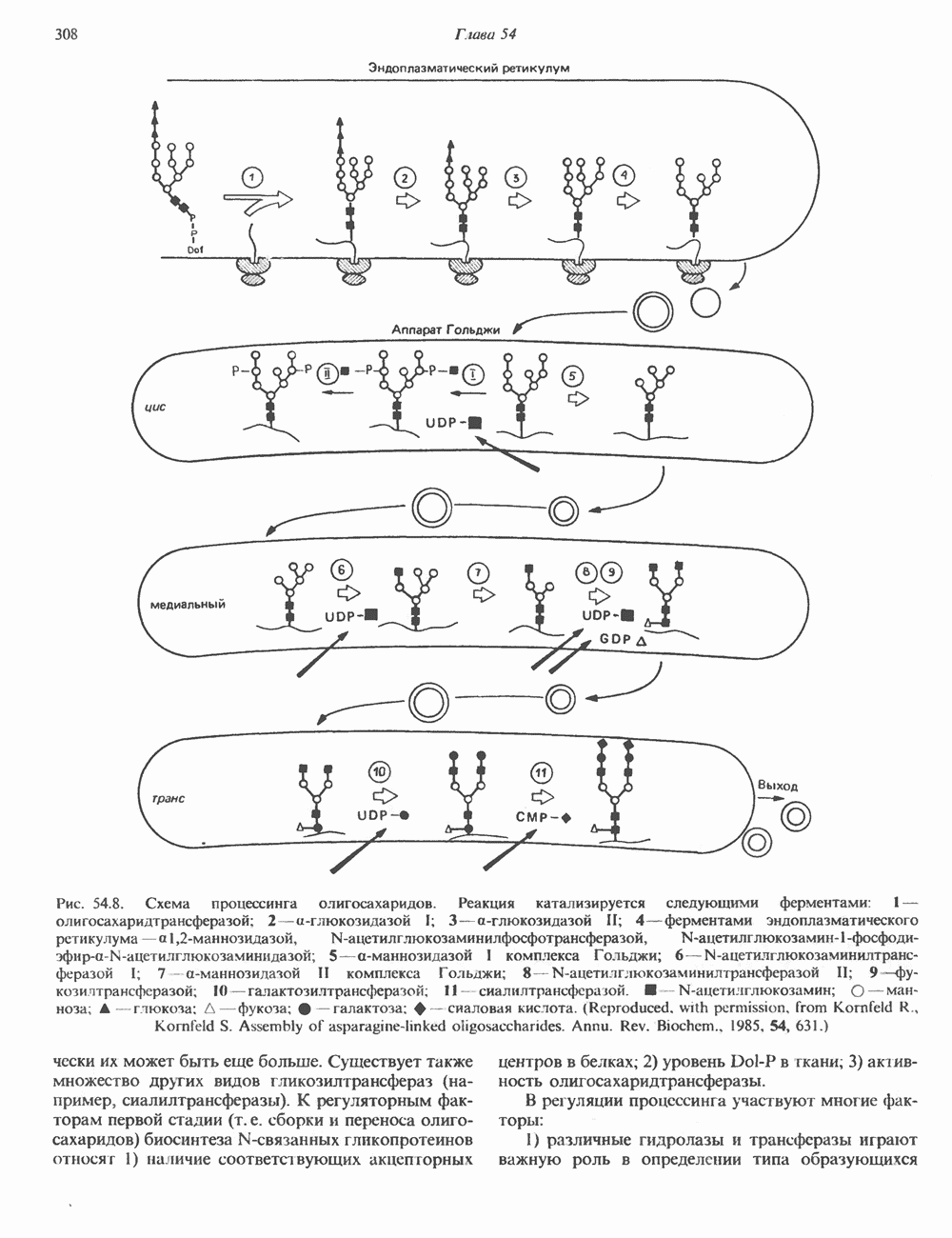
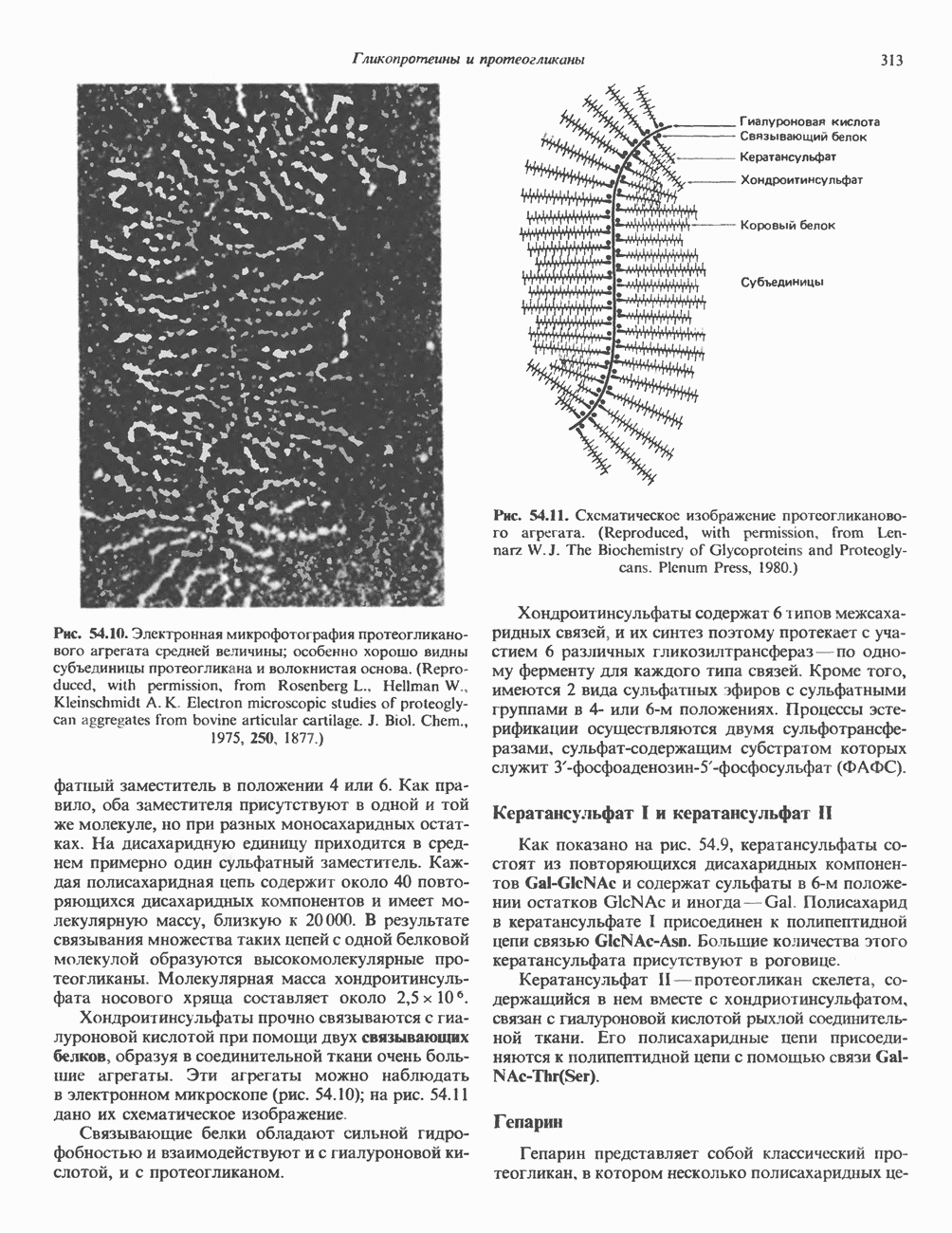
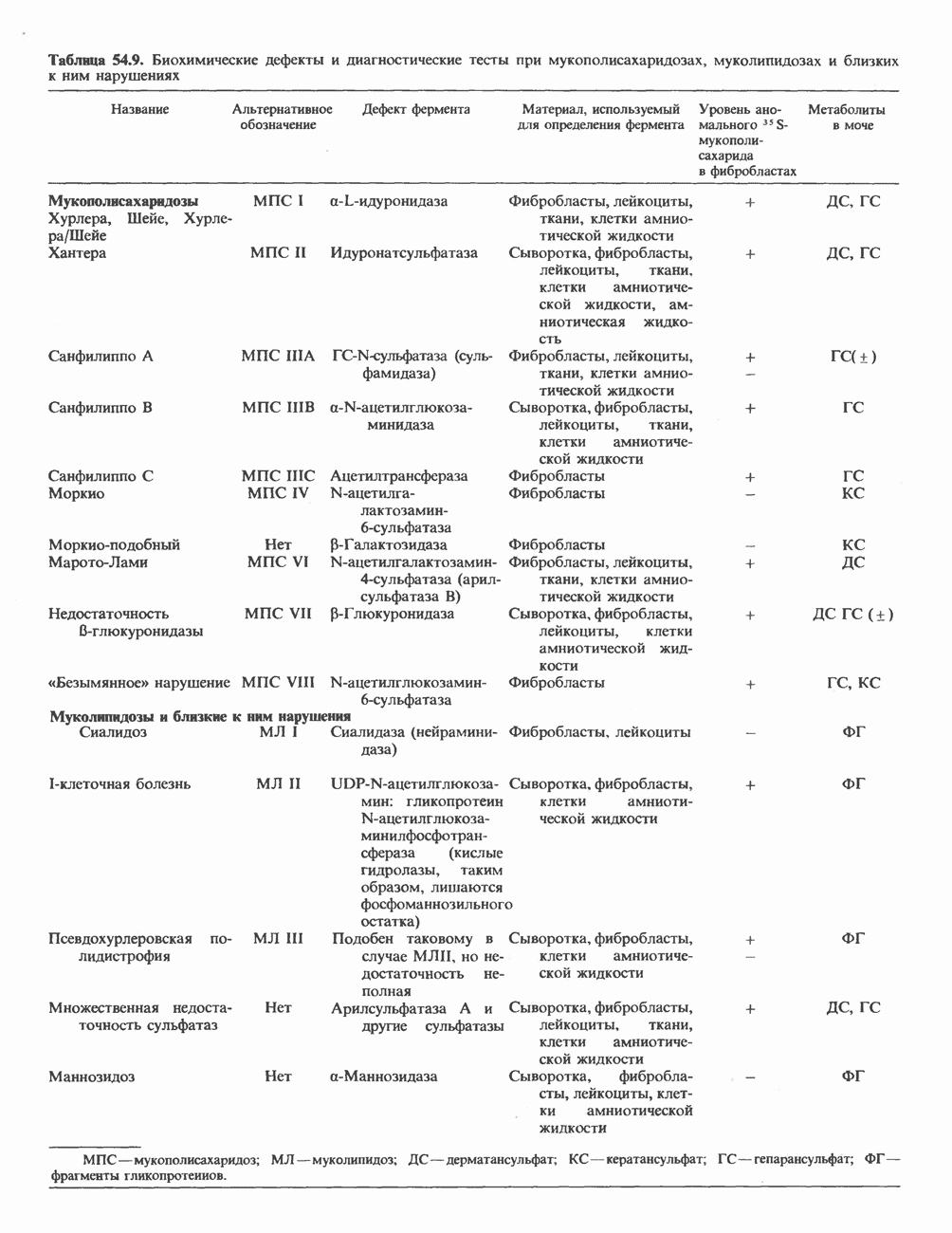
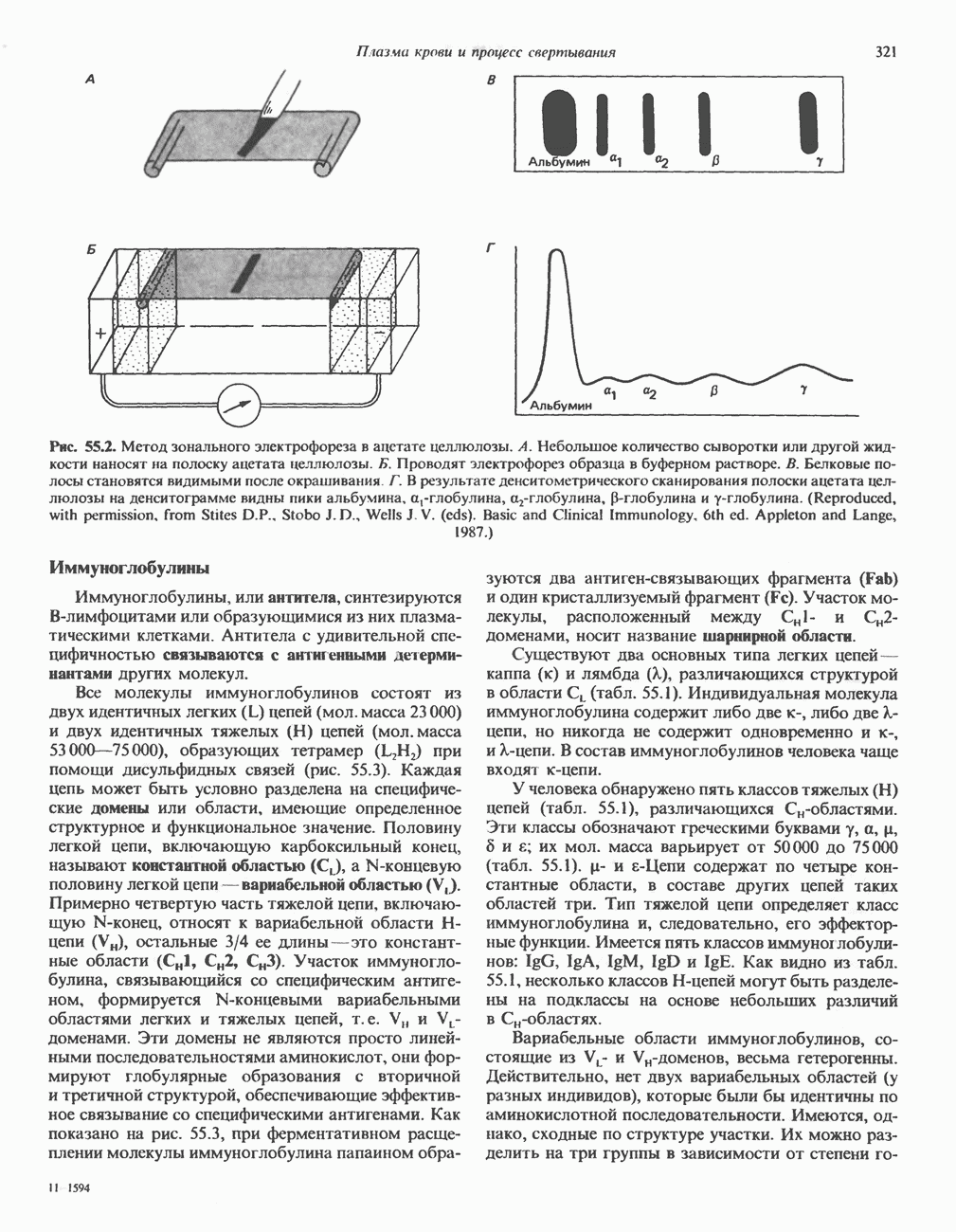
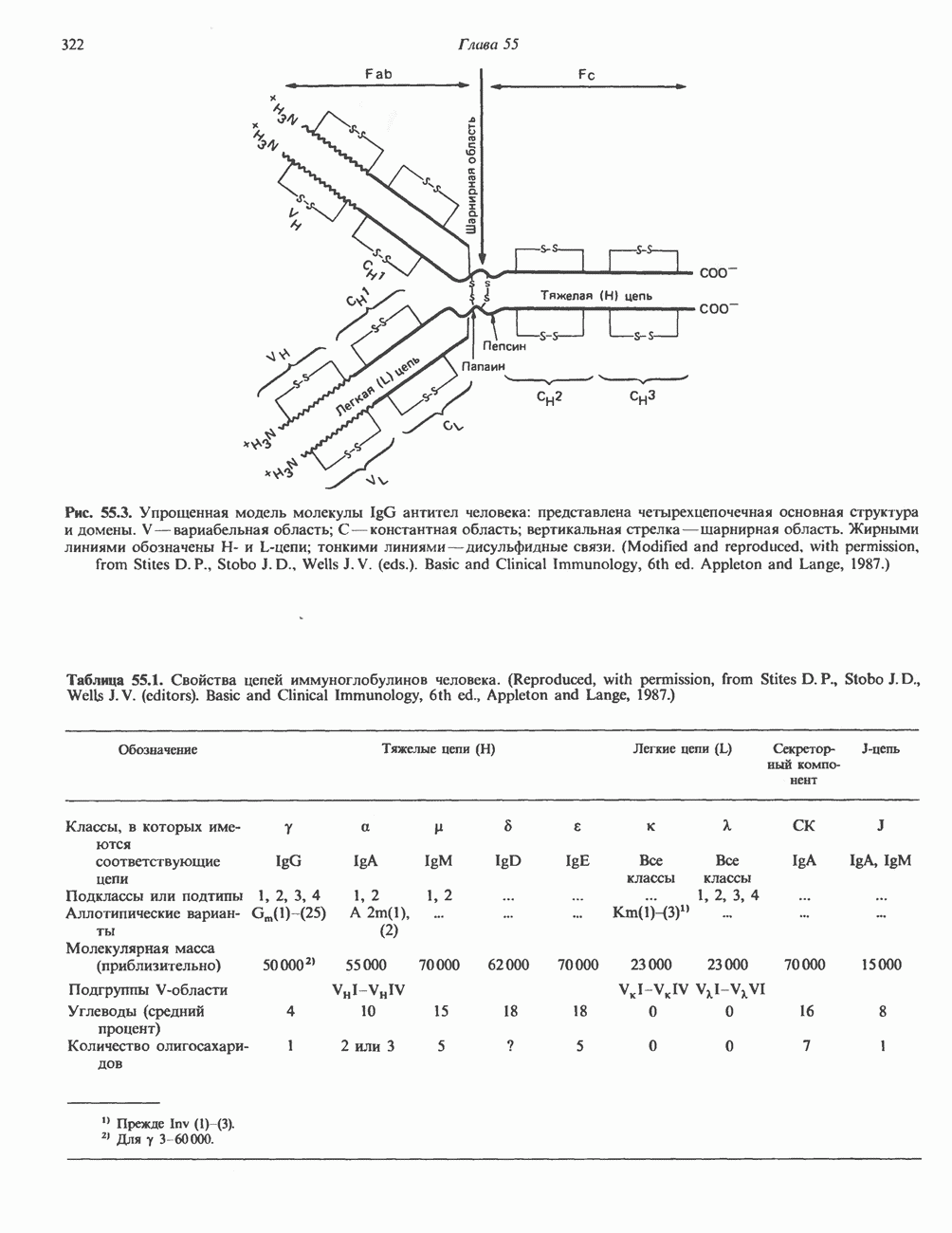
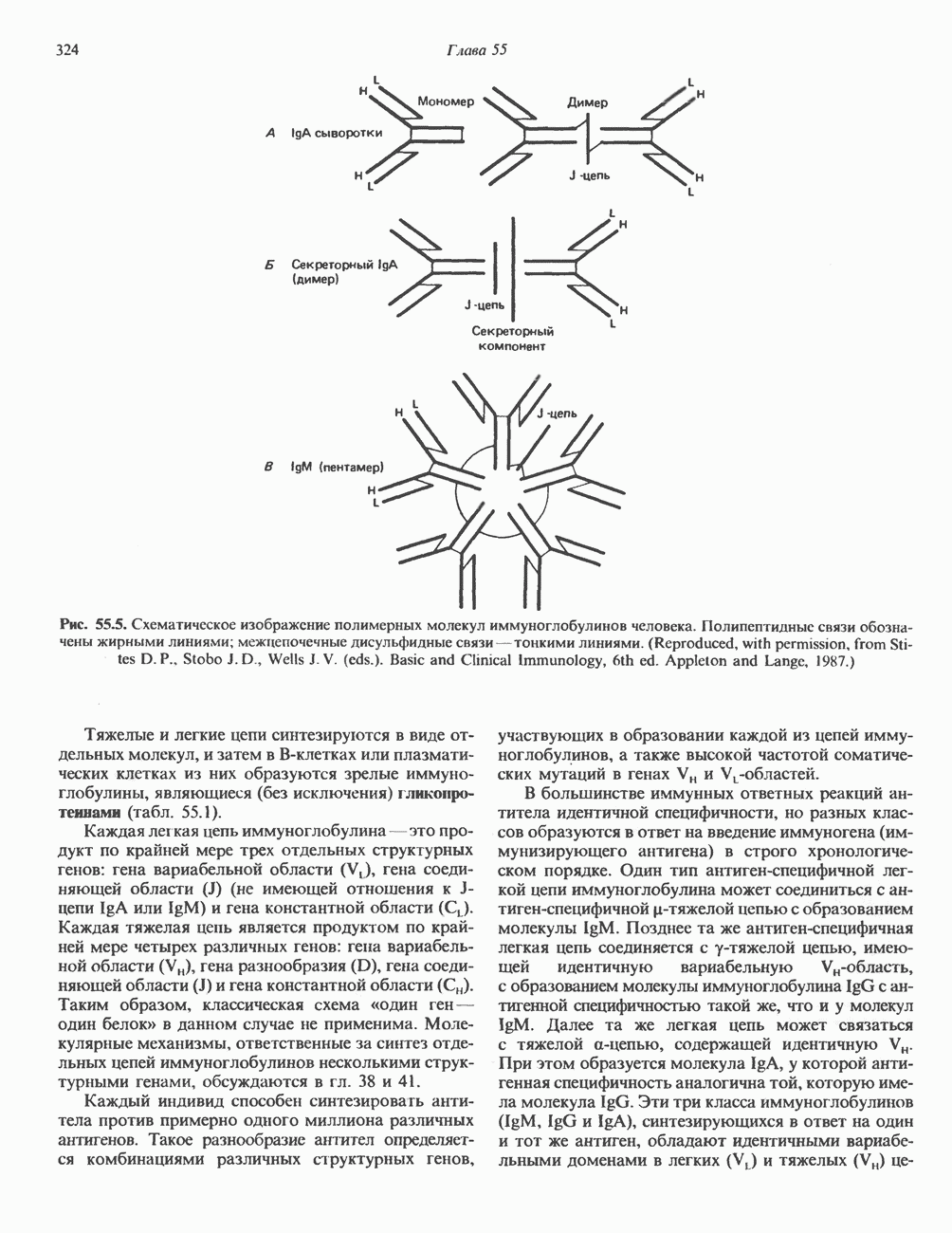
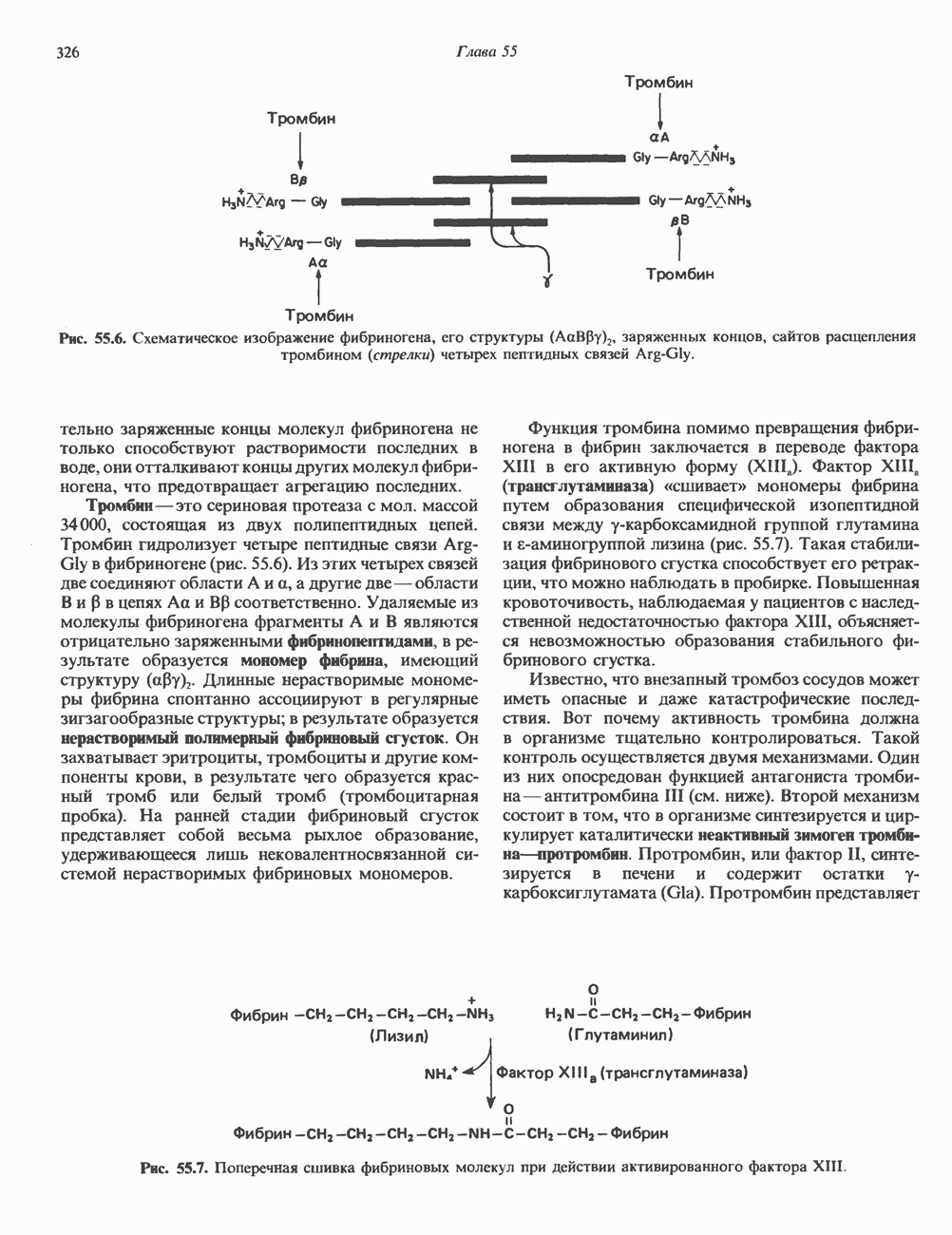
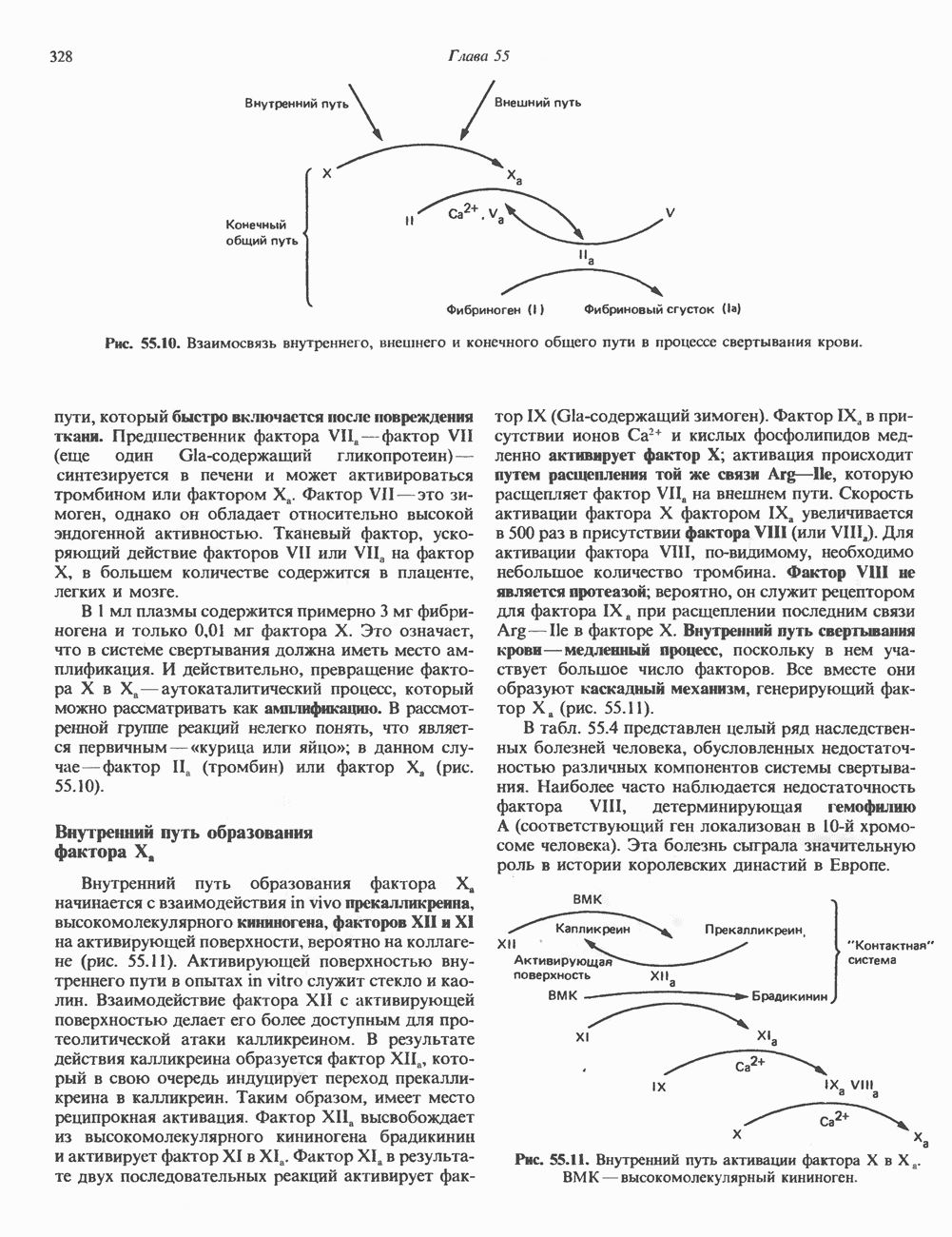
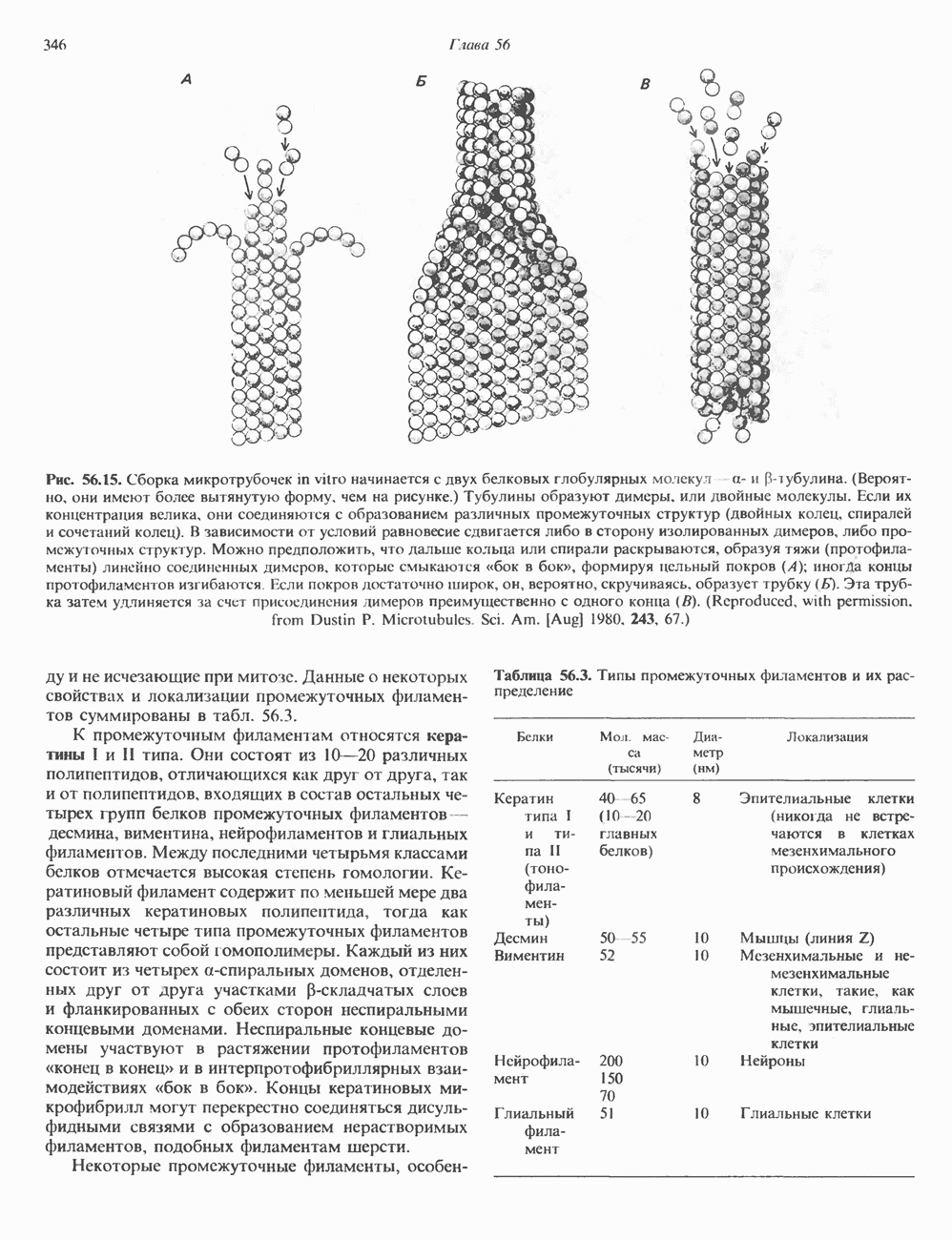
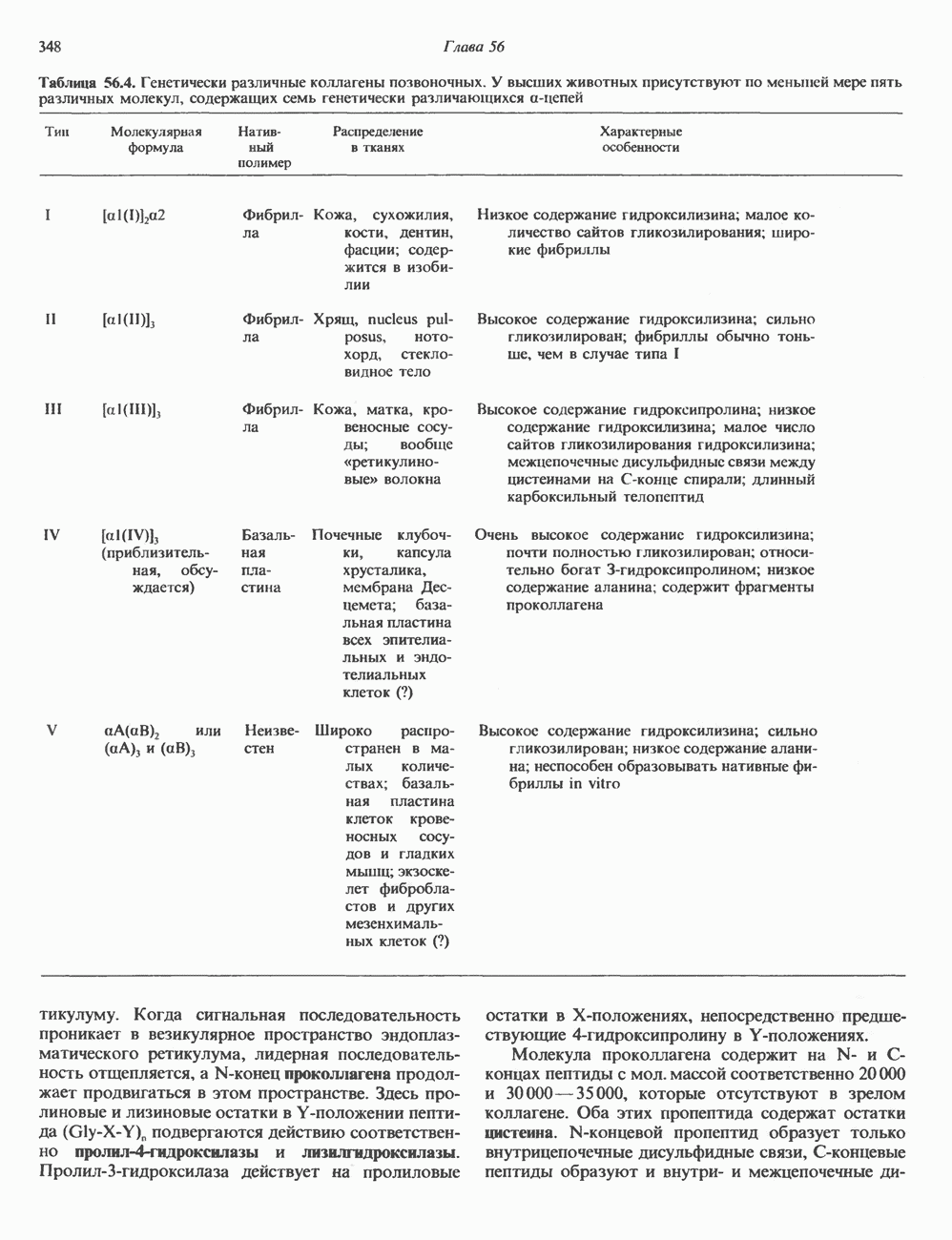
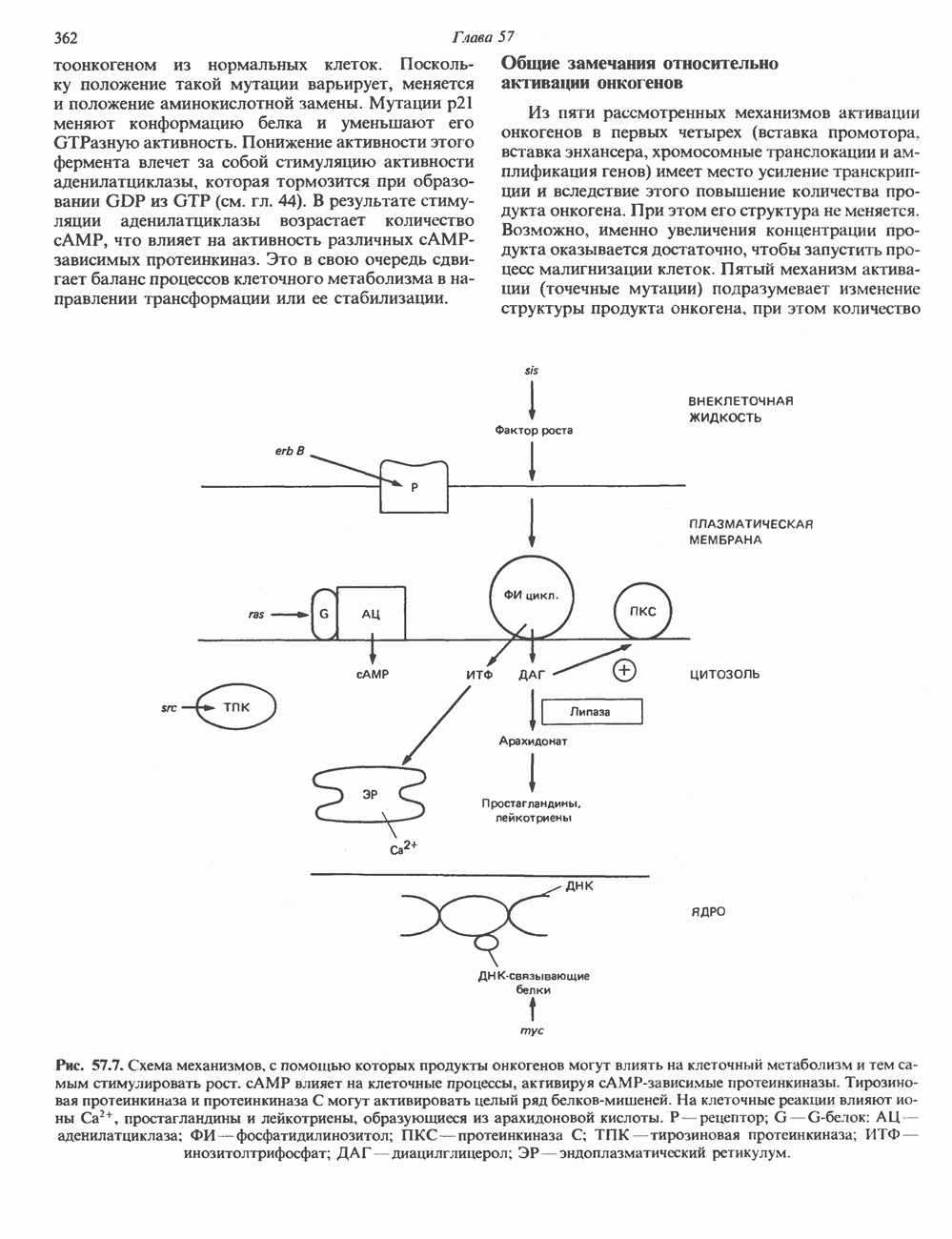
2. ДЕЙСТВИЕ ГОРМОНОВ, ОПОСРЕДОВАННОЕ КАЛЬЦИЕМ И ФОСФОИНОЗИТИДАМИ
Ионизированный кальций служит важнейшим регулятором разнообразных процессов, таких, как мышечное сокращение, сопряжение стимул-секреция, последовательность реакций свертывания крови, активность многих ферментов, возбудимость клеточных мембран. Он является также внутриклеточным посредником действия ряда гормонов.
Метаболизм кальция
Концентрация внеклеточного кальция (Са2+) составляет 5 ммоль/л и регулируется очень строго (см. гл. 47). Внутриклеточная концентрация свободных ионов кальция гораздо ниже - 0,1-10 мкмоль/л, а количество  связанного с внутриклеточными органеллами (митохондриями и эндоплазматическим ретикулумом), колеблется в пределах 1-20 мкмоль/л. Несмотря на этот 5000-10000-кратный концентрационный градиент и благоприятствующий проникновению Са2+ трансмембранный электрический градиент, вход Са2+ в клетки резко ограничен. Изменение концентрации Са2+ в цитозоле происходит по трем механизмам. Ряд гормонов (класса II. Б) повышает проницаемость мембраны для Са2+ и тем самым увеличивает вход в клетку Са2+. Это может осуществляться по механизму
связанного с внутриклеточными органеллами (митохондриями и эндоплазматическим ретикулумом), колеблется в пределах 1-20 мкмоль/л. Несмотря на этот 5000-10000-кратный концентрационный градиент и благоприятствующий проникновению Са2+ трансмембранный электрический градиент, вход Са2+ в клетки резко ограничен. Изменение концентрации Са2+ в цитозоле происходит по трем механизмам. Ряд гормонов (класса II. Б) повышает проницаемость мембраны для Са2+ и тем самым увеличивает вход в клетку Са2+. Это может осуществляться по механизму  -обмена, обладающему большой емкостью, но низким сродством к Са2+. Существует также зависимый от АТРазы
-обмена, обладающему большой емкостью, но низким сродством к Са2+. Существует также зависимый от АТРазы  -насос, обеспечивающий выведение Са2+ из клетки в обмен на Н+. Этот механизм характеризуется высоким сродством к Са2+, но малой емкостью и, по-видимому, ответствен за тонкую настройку уровня Са2+ в цитозоле. Наконец, возможна
-насос, обеспечивающий выведение Са2+ из клетки в обмен на Н+. Этот механизм характеризуется высоким сродством к Са2+, но малой емкостью и, по-видимому, ответствен за тонкую настройку уровня Са2+ в цитозоле. Наконец, возможна
как мобилизация  митохондрий и эндоплазматического ретикулума, так и накопление
митохондрий и эндоплазматического ретикулума, так и накопление  в этих органеллах.
в этих органеллах.
Современные представления о роли  как внутриклеточного посредника в действии гормонов основаны на двух наблюдениях. Во-первых, удалось количественно определить быстрые изменения внутриклеточной концентрации
как внутриклеточного посредника в действии гормонов основаны на двух наблюдениях. Во-первых, удалось количественно определить быстрые изменения внутриклеточной концентрации  такие изменения соответствуют роли внутриклеточного посредника. Эти данные были получены разными методами, в том числе путем использования флуоресцирующих хелаторов
такие изменения соответствуют роли внутриклеточного посредника. Эти данные были получены разными методами, в том числе путем использования флуоресцирующих хелаторов  и фура-2 (Fura 2). С помощью этих соединений можно количественно оценить быстрые изменения концентрации
и фура-2 (Fura 2). С помощью этих соединений можно количественно оценить быстрые изменения концентрации  на субмикромолярном уровне. Второе важное наблюдение, указывающее на связь
на субмикромолярном уровне. Второе важное наблюдение, указывающее на связь  с эффектом гормонов, состояло в определении внутриклеточных мишеней действия этого иона: был обнаружен
с эффектом гормонов, состояло в определении внутриклеточных мишеней действия этого иона: был обнаружен  -зависимый регулятор фосфодиэстеразной активности, и это послужило основой для понимания того, каким образом
-зависимый регулятор фосфодиэстеразной активности, и это послужило основой для понимания того, каким образом  взаимодействуют внутри клетки.
взаимодействуют внутри клетки.
Кальмодулин
Кальций-зависимый регуляторный белок назван кальмодулином; его мол. масса 17000, по структуре и функции он гомологичен мышечному белку тропонину С. Кальмодулин содержит четыре участка связывания  Связывание
Связывание  по всем четырем участкам ведет к заметному изменению конформации белка: большая часть молекулы приобретает структуру а-спирали. Эти конформационные переходы определяют, видимо, способность кальмодулина активировать или инактивировать определенные ферменты. Взаимодействие ионов кальция с кальмодулином (и соответствующее изменение активности последнего) в принципе сходно с процессом связывания
по всем четырем участкам ведет к заметному изменению конформации белка: большая часть молекулы приобретает структуру а-спирали. Эти конформационные переходы определяют, видимо, способность кальмодулина активировать или инактивировать определенные ферменты. Взаимодействие ионов кальция с кальмодулином (и соответствующее изменение активности последнего) в принципе сходно с процессом связывания  с протеинкиназой, обеспечивающим активацию этого фермента. Кальмодулин часто оказывается одной из многочисленных субъединиц сложных белков и, как правило, участвует в регуляции активности различных киназ, а также ферментов синтеза и распада циклических нуклеотидов. Список некоторых ферментов, прямо или косвенно (по-видимому, через кальмодулин) регулируемых
с протеинкиназой, обеспечивающим активацию этого фермента. Кальмодулин часто оказывается одной из многочисленных субъединиц сложных белков и, как правило, участвует в регуляции активности различных киназ, а также ферментов синтеза и распада циклических нуклеотидов. Список некоторых ферментов, прямо или косвенно (по-видимому, через кальмодулин) регулируемых  приведен в табл. 44.5.
приведен в табл. 44.5.
Са2+-кальмодулин оказывает регуляторное влияние не только на активность ферментов и транспорт ионов, но и на функционирование многих структурных элементов в клетке. К числу последних относится актинмиозиновый комплекс гладких мыщц, находящийся под Р-адренергическим контролем, а в неконтрактильных клетках — микрофиламенты, опосредующие такие процессы, как клеточная подвижность, изменение формы клеток, митоз, высвобождение гранул, эндоцитоз.
Таблица 44.5. Ферменты, регулируемые комплексом кальций-кальмодулин

Кальций как медиатор действия гормонов
Роль ионизированного кальция в действии гормонов доказывается следующими наблюдениями: эффект многих гормонов 1) исчезает в бескальциевой среде или при истощении внутриклеточных запасов  может быть имитирован с помощью агентов, увеличивающих концентрацию
может быть имитирован с помощью агентов, увеличивающих концентрацию  в цитозоле, например
в цитозоле, например  сопряжен с транспортом
сопряжен с транспортом  в клетку. Все эти явления были довольно подробно изучены на клетках гипофиза, гладких мышц, слюнных желез и на тромбоцитах; наиболее полно исследован механизм регуляции метаболизма гликогена в печени вазопрессином и
в клетку. Все эти явления были довольно подробно изучены на клетках гипофиза, гладких мышц, слюнных желез и на тромбоцитах; наиболее полно исследован механизм регуляции метаболизма гликогена в печени вазопрессином и  -адренергическими катехоламинами. Указанный механизм схематически представлен на рис. 19.5 и 19.7.
-адренергическими катехоламинами. Указанный механизм схематически представлен на рис. 19.5 и 19.7.
Добавление  -агонистов или вазопрессина к изолированным гепатоцитам уже через несколько секунд вызывает
-агонистов или вазопрессина к изолированным гепатоцитам уже через несколько секунд вызывает  -кратное увеличение содержания
-кратное увеличение содержания  в цитозоле (с 0,2 до 0,6 мкмоль/л). Это увеличение предшествует такому же возрастанию активности фосфорилазы; изучение каждого из указанных эффектов показало, что они наблюдаются при сопоставимых концентрациях гормона,
в цитозоле (с 0,2 до 0,6 мкмоль/л). Это увеличение предшествует такому же возрастанию активности фосфорилазы; изучение каждого из указанных эффектов показало, что они наблюдаются при сопоставимых концентрациях гормона,  -Антагонисты ингибируют повышение
-Антагонисты ингибируют повышение  в цитозоле; удаление гормона приводит к быстрому снижению как концентрации
в цитозоле; удаление гормона приводит к быстрому снижению как концентрации  в цитозоле, так и количества фосфорилазы а. Первоначально
в цитозоле, так и количества фосфорилазы а. Первоначально  поступает, очевидно, из клеточных органелл, причем запасенного в них
поступает, очевидно, из клеточных органелл, причем запасенного в них  видимо, достаточно для того, чтобы мог проявиться немедленный эффект гормона. Для более продолжительного действия необходим либо вход
видимо, достаточно для того, чтобы мог проявиться немедленный эффект гормона. Для более продолжительного действия необходим либо вход  в клетку, либо торможение его выхода, осуществляемого
в клетку, либо торможение его выхода, осуществляемого  -насосом. Последний процесс зависит от происходящего одновременно возрастания концентрации
-насосом. Последний процесс зависит от происходящего одновременно возрастания концентрации 
Активация фосфорилазы происходит путем превращения фосфорилазы b в фосфорилазу а под
действием фермента киназы фосфорилазы b. В состав этого фермента (в качестве его 5-субъединицы) входит кальмодулин, и активность фермента возрастает при увеличении концентрации Са2+ в пределах 0,1-1 мкмоль/л, т. е. в тех же пределах, в каких содержание ионов кальция в цитозоле печени повышается в присутствии гормона. Связь между Са2+ и активацией фосфорилазы совершенно определенна.
С помощью Са2+ или путем фосфорилирования или обоими способами одновременно осуществляется регуляция целого ряда ключевых ферментов метаболизма; к их числу относятся гликогенсинтаза, глицерол-3-фосфат-дегидрогеназа, пируватдегидрогеназа, пируваткиназа и пируваткарбоксилаза. Остается не ясным, прямо ли участвует в этой регуляции кальмодулин или же основная роль принадлежит недавно открытым протеинкиназам ( -модулин-зависимой либо
-модулин-зависимой либо  -зависимой).
-зависимой).
Роль продуктов превращения фосфоинозитидов в зависимом действии гормонов
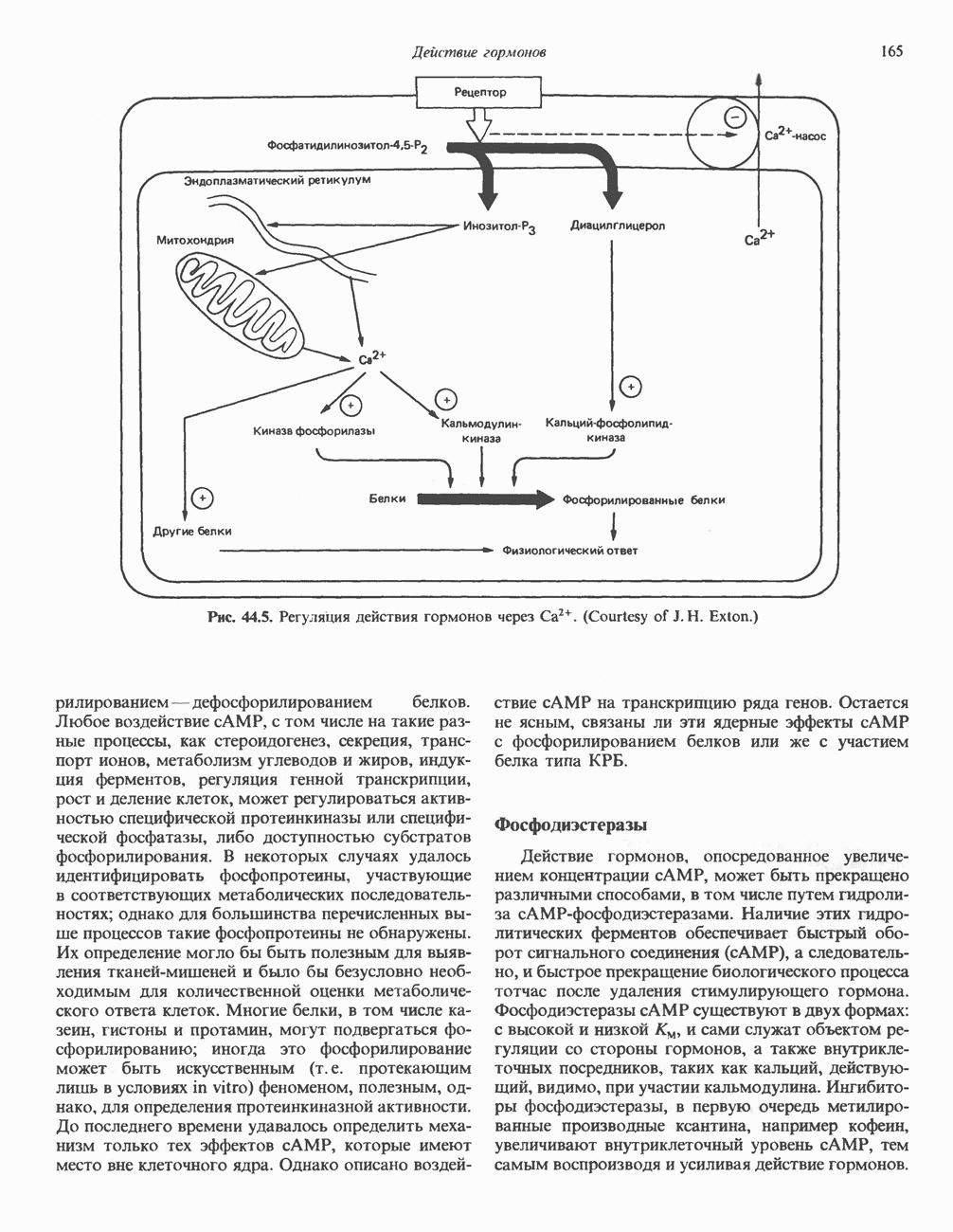
Очевидно, что коммуникация между рецептором гормона на плазматической мембране и внутриклеточными резервуарами  должна осуществляться с помощью какого-то сигнала. Наиболее вероятные кандидаты на роль такого сигнала — продукты превращения фосфоинозитидов. Фосфатидилинозитол-4,5-бисфосфат под действием фосфолипазы С гидролизуется до мионнозигол-1,4,5-трифосфата и диацилглицерола (рис. 44.5). В гепатоцитах эта реакция наблюдается через несколько секунд после добавления вазопрессина или адреналина. Как показано на различных препаратах мембран и целых органелл,
должна осуществляться с помощью какого-то сигнала. Наиболее вероятные кандидаты на роль такого сигнала — продукты превращения фосфоинозитидов. Фосфатидилинозитол-4,5-бисфосфат под действием фосфолипазы С гидролизуется до мионнозигол-1,4,5-трифосфата и диацилглицерола (рис. 44.5). В гепатоцитах эта реакция наблюдается через несколько секунд после добавления вазопрессина или адреналина. Как показано на различных препаратах мембран и целых органелл,  в концентрациях
в концентрациях  вызывает очень быстрое высвобождение
вызывает очень быстрое высвобождение  Попытки воспроизвести действие гормона посредством этого соединения (а это важный этап установления взаимосвязи между ними) были не совсем успешными, вероятно потому, что трудно добиться проникновения миоинозитола-Р3 в клетку, а внутри клетки он очень быстро гидролизуется. Другой продукт гидролиза фосфоинозитида-1,2-диацилгли-церол—активирует
Попытки воспроизвести действие гормона посредством этого соединения (а это важный этап установления взаимосвязи между ними) были не совсем успешными, вероятно потому, что трудно добиться проникновения миоинозитола-Р3 в клетку, а внутри клетки он очень быстро гидролизуется. Другой продукт гидролиза фосфоинозитида-1,2-диацилгли-церол—активирует  -фосфолипид-зависимую протеинкиназу за счет увеличения АГМ-фермента по отношению к
-фосфолипид-зависимую протеинкиназу за счет увеличения АГМ-фермента по отношению к  Вопрос о том, какую роль играет этот процесс в действии
Вопрос о том, какую роль играет этот процесс в действии  -зависимых гормонов, в настоящее время исследуется.
-зависимых гормонов, в настоящее время исследуется.
Действие стероидогенных агентов, в том числе АКТГ и  в коре надпочечников; ангиотензина
в коре надпочечников; ангиотензина  серотонина, АКТГ и дибутирил-сАМР в клубочковой зоне надпочечников; ЛГ в яичниках; ЛГ и
серотонина, АКТГ и дибутирил-сАМР в клубочковой зоне надпочечников; ЛГ в яичниках; ЛГ и  в клетках Лейдига (в семенниках), сопряжено с возрастанием концентраций фосфатидной кислоты, фосфоинозитола и полифосфоинозитидов в соответствующих тканях-мишенях.
в клетках Лейдига (в семенниках), сопряжено с возрастанием концентраций фосфатидной кислоты, фосфоинозитола и полифосфоинозитидов в соответствующих тканях-мишенях.
Можно привести еще несколько примеров. Так, через 5—10 с после добавления ТРГ к клетках гипофиза в них заметно возрастает расщепление фосфоинозитидов фосфолипазой С; при этом повышается уровень инозитолди- и трифосфатов в клетках и в результате происходит мобилизация внутриклеточного кальция. Это ведет к активации  -зависимой протеинкиназы, которая в свою очередь фосфорилирует ряд белков (один из них, вероятно, участвует в высвобождении ТСГ). Кальций, по-видимому, служит также внутриклеточным медиатором действия
-зависимой протеинкиназы, которая в свою очередь фосфорилирует ряд белков (один из них, вероятно, участвует в высвобождении ТСГ). Кальций, по-видимому, служит также внутриклеточным медиатором действия  на высвобождение Л. Г. Полагают, что в этом процессе участвует также кальмодулин.
на высвобождение Л. Г. Полагают, что в этом процессе участвует также кальмодулин.
Роль кальция и продуктов расщепления полифосфоинозитидов в действии гормонов представлена на рис. 44.5. Как видно из схемы, продукты гидролиза фосфоинозитидов служат вторыми посредниками, а  фактически третьим посредником. Схему можно было бы дополнить тем, что в сопряжении событий, происходящих на мембране, с высвобождением
фактически третьим посредником. Схему можно было бы дополнить тем, что в сопряжении событий, происходящих на мембране, с высвобождением  участвует, возможно,
участвует, возможно,  -белок. Такая сложнейшая сеть внутриклеточных посредников, по-видимому, не уникальное явление.
-белок. Такая сложнейшая сеть внутриклеточных посредников, по-видимому, не уникальное явление.