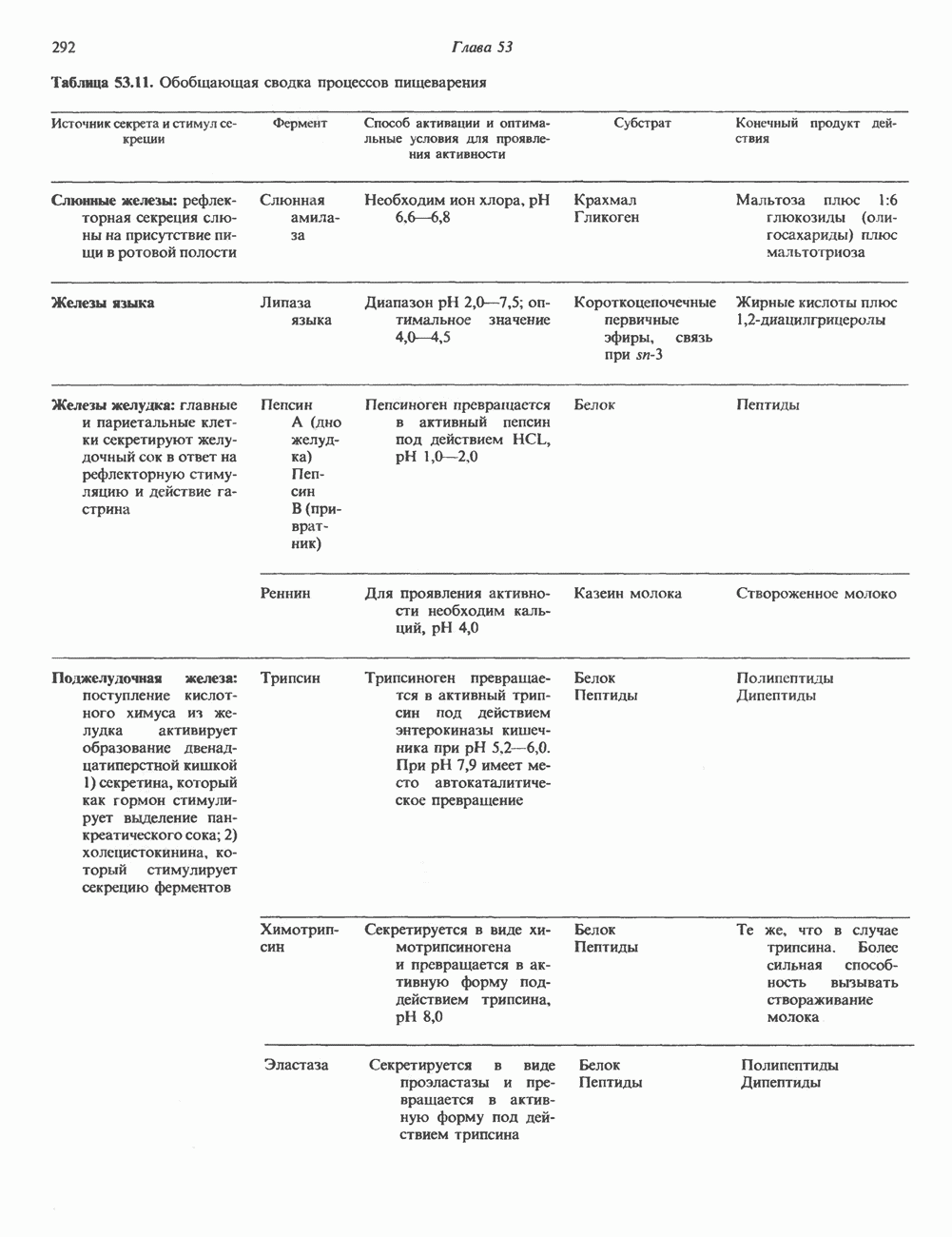
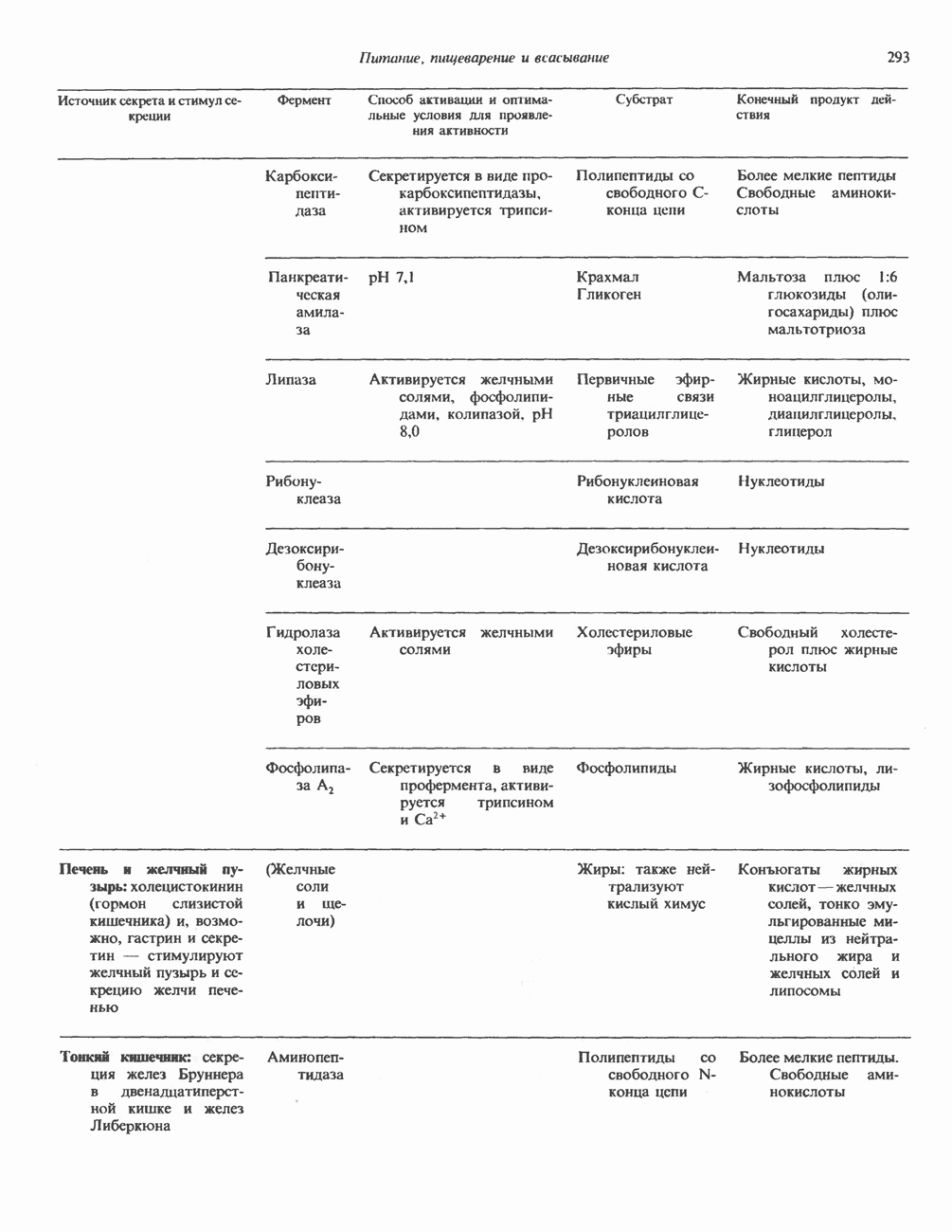
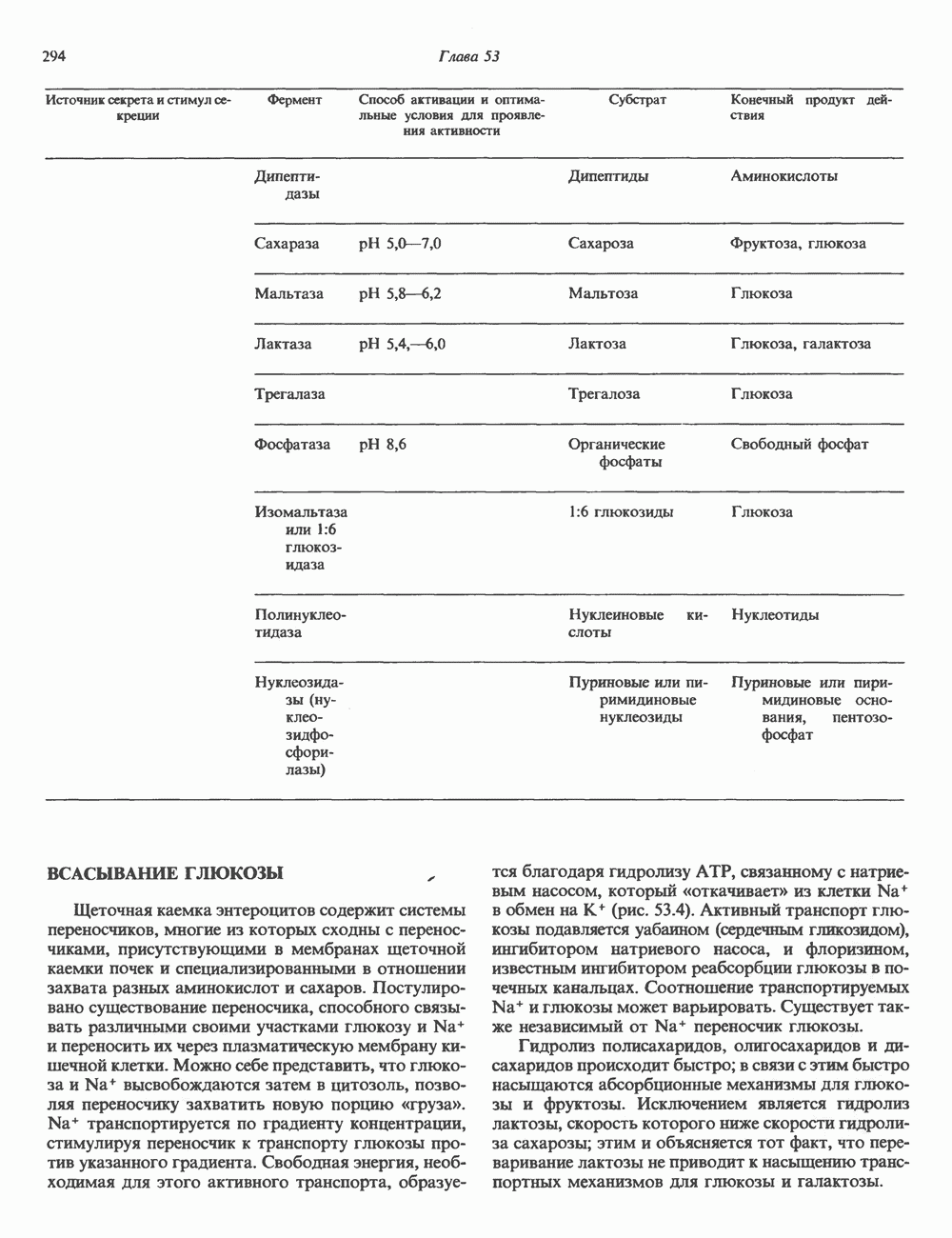
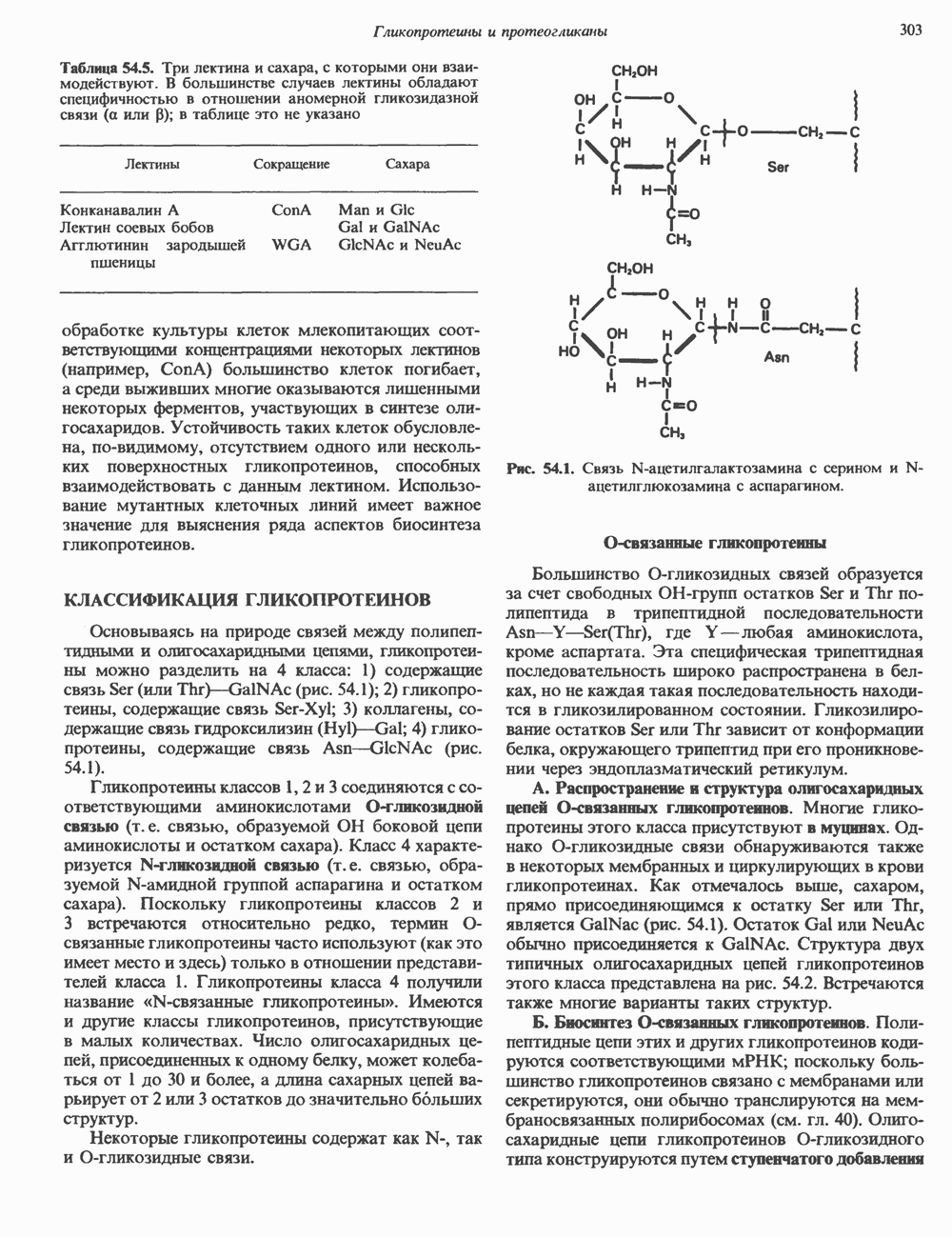
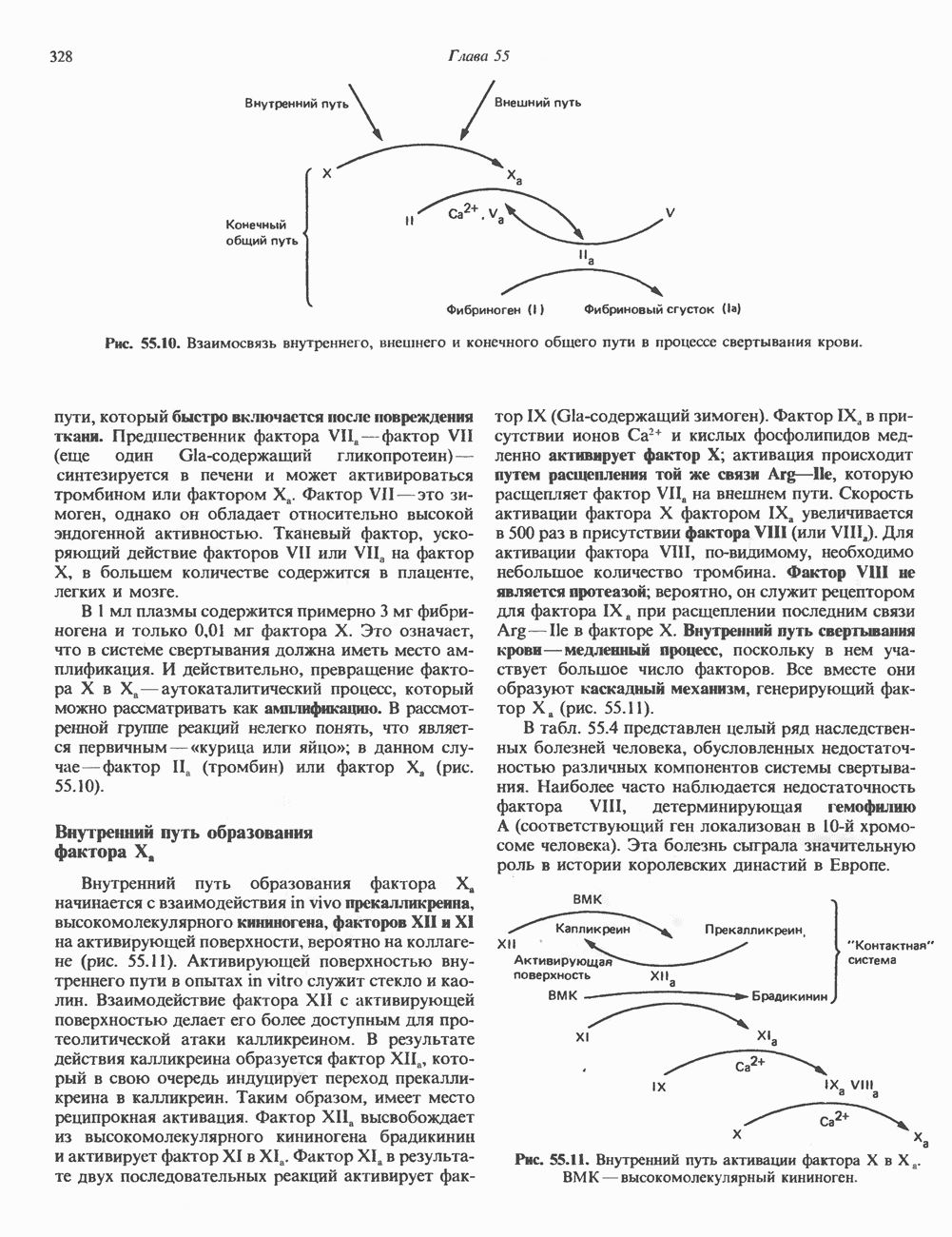
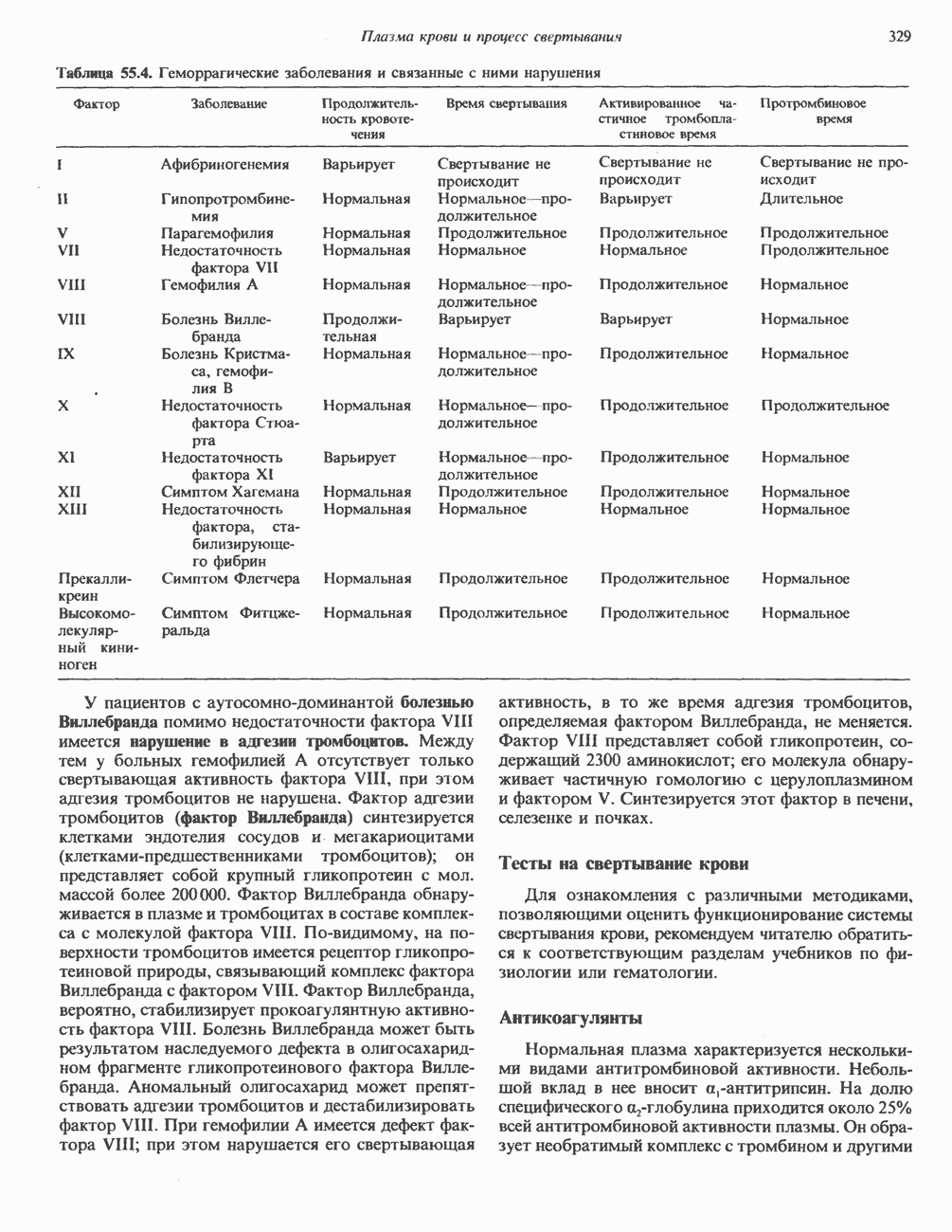
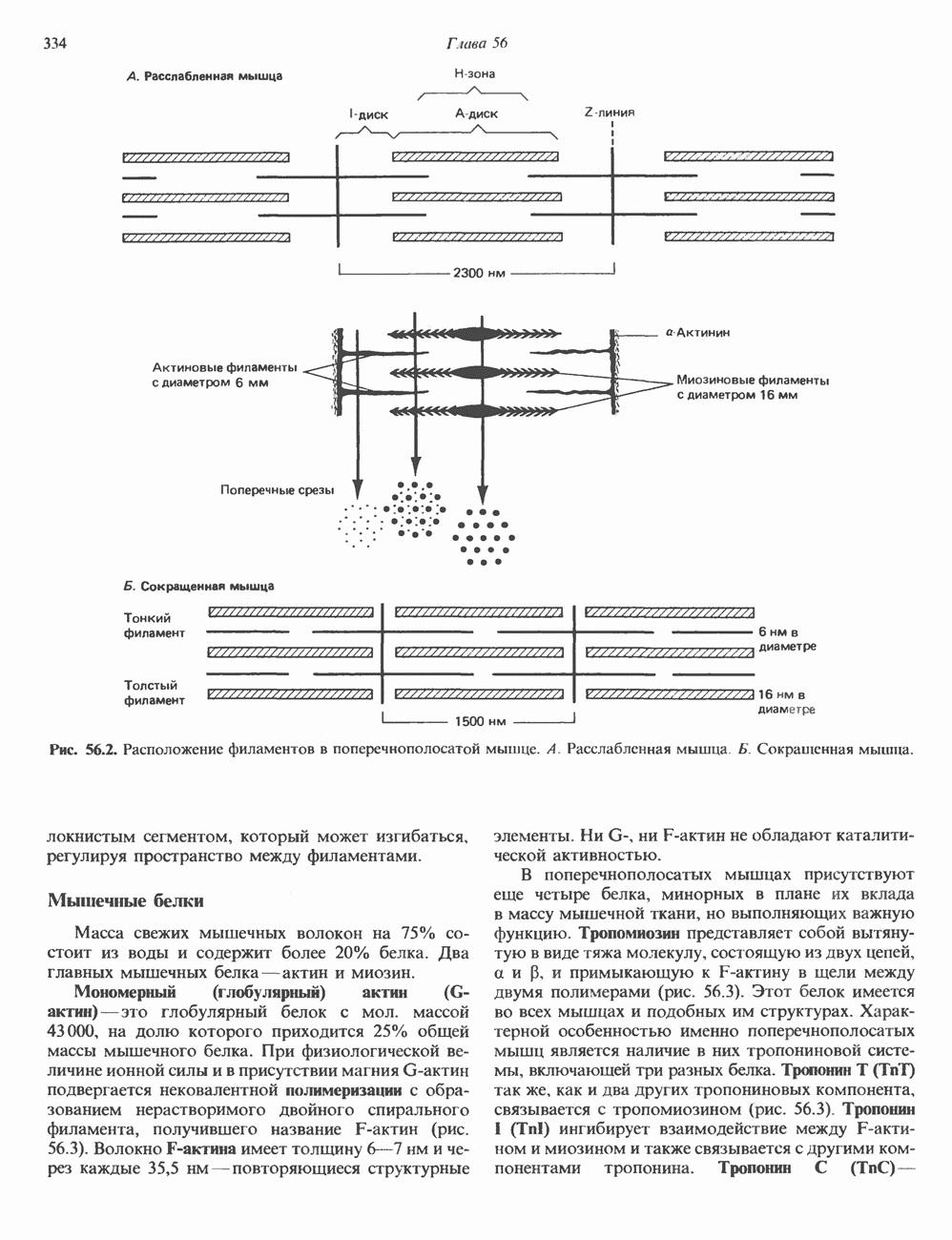
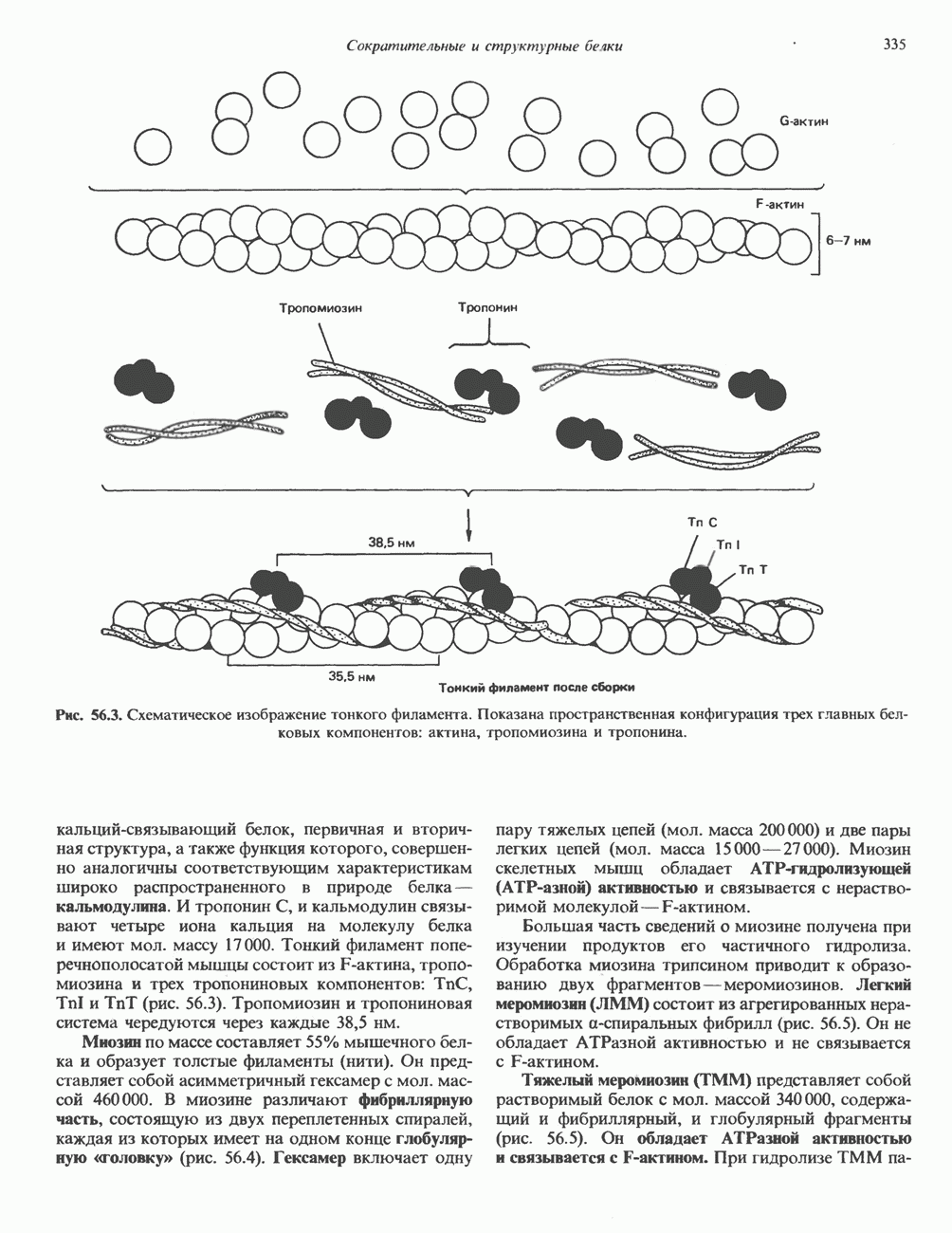
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
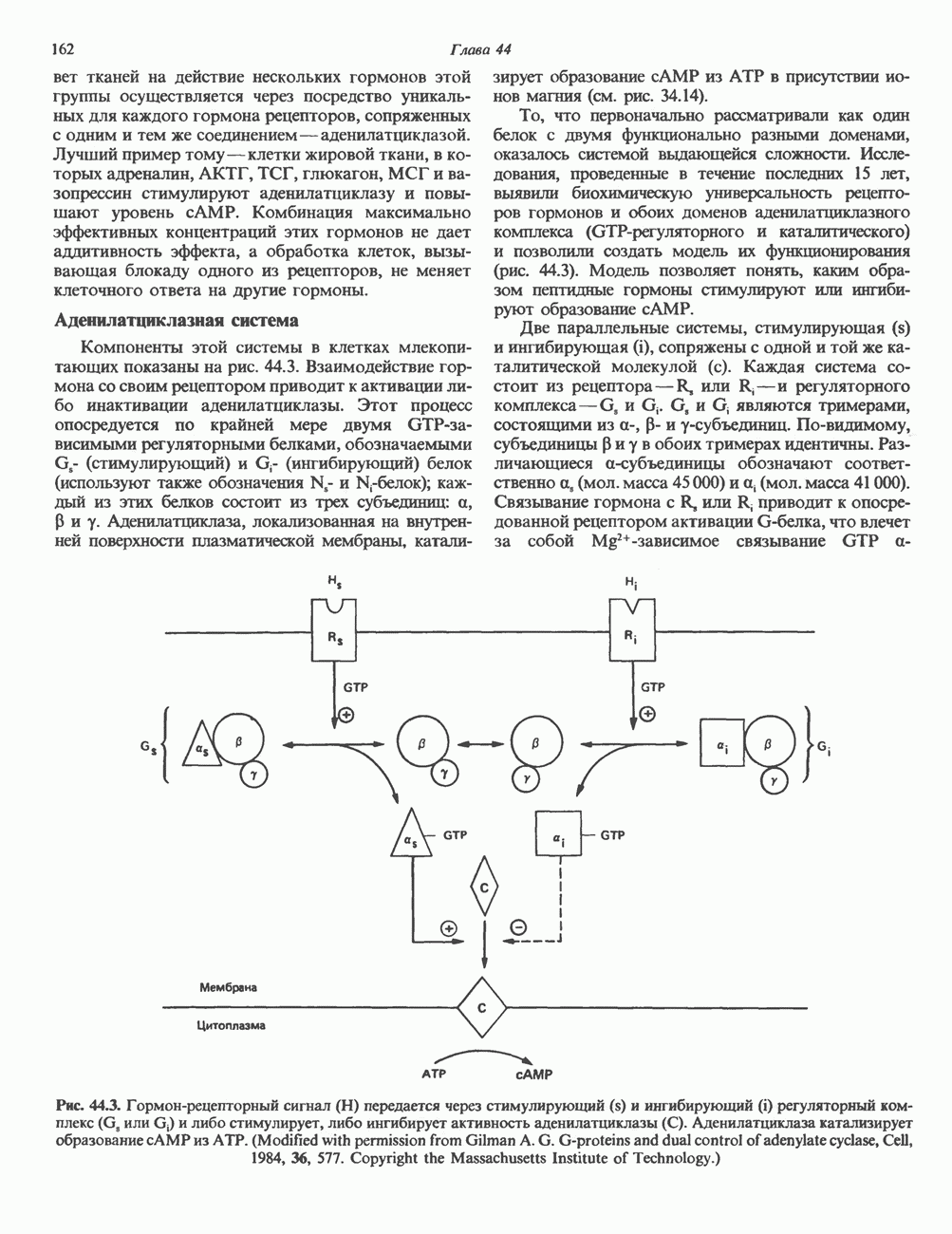
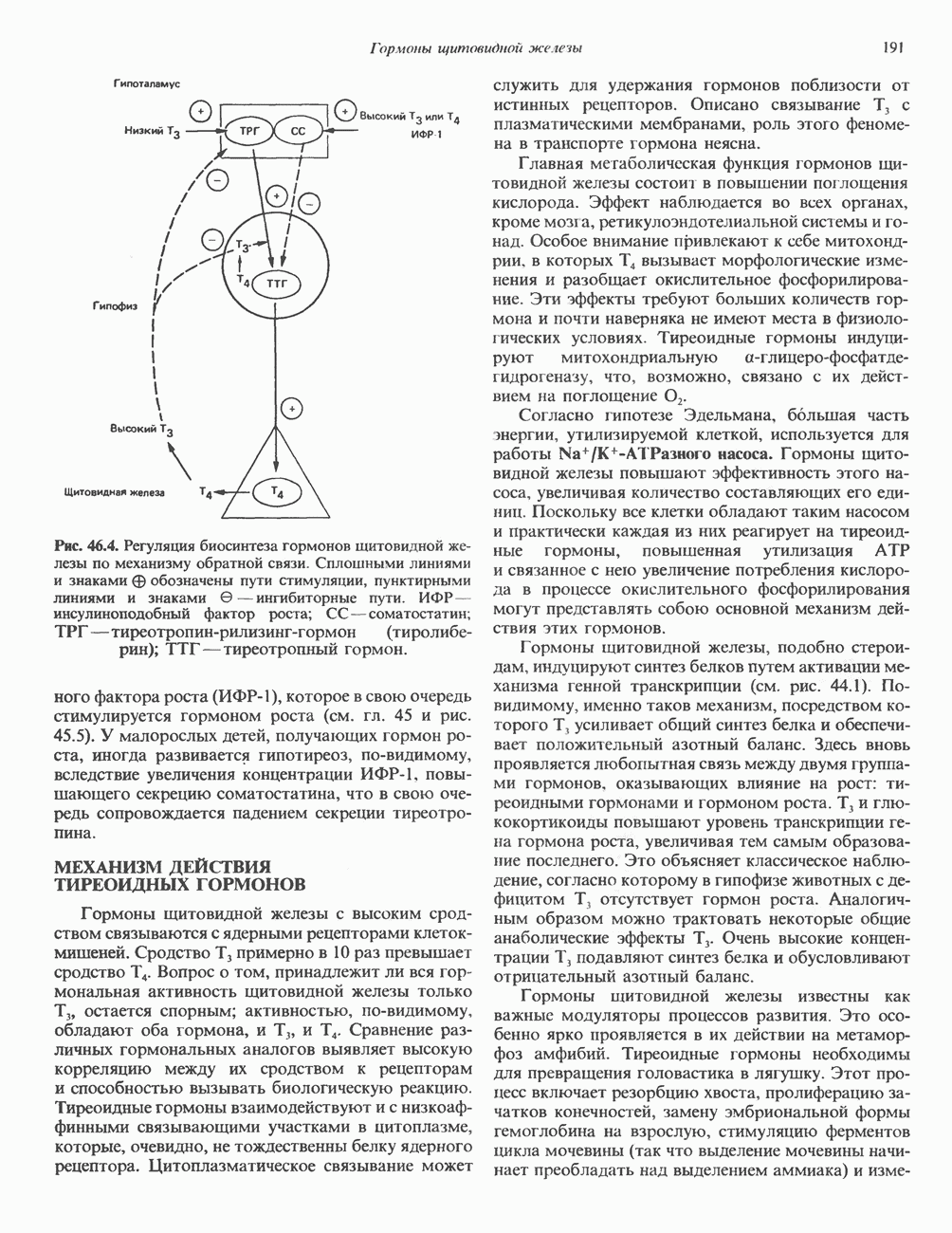
ИЗМЕНЕНИЯ И ПЕРЕСТРОЙКИ ГЕНЕТИЧЕСКОГО МАТЕРИАЛАИзменения последовательности пуриновых и пиримидиновых оснований, вызванные заменой, удалением или вставкой одного или более нуклеотидов, могут привести к изменению продукта данного гена в большинстве случаев белка. Последствия подобных изменений (мутаций) генетического материала описаны в гл. 40. Рекомбинация хромосомГомологичные хромосомы прокариот и эукариот могут обмениваться генетическим материалом. Обмен или рекомбинация происходит в клетках млекопитающих главным образом при мейозе. Этому событию предшествует попарное выстраивание гомологичных
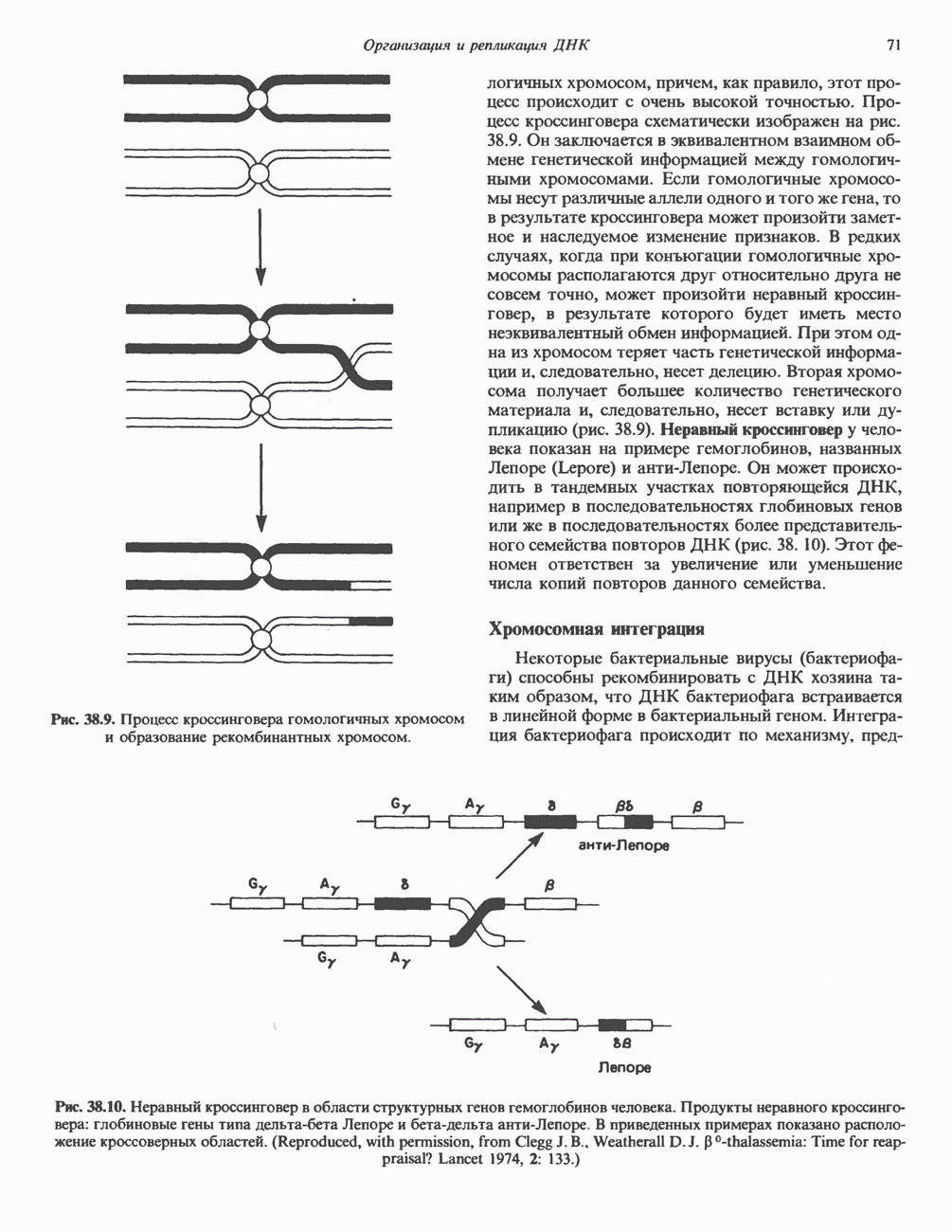
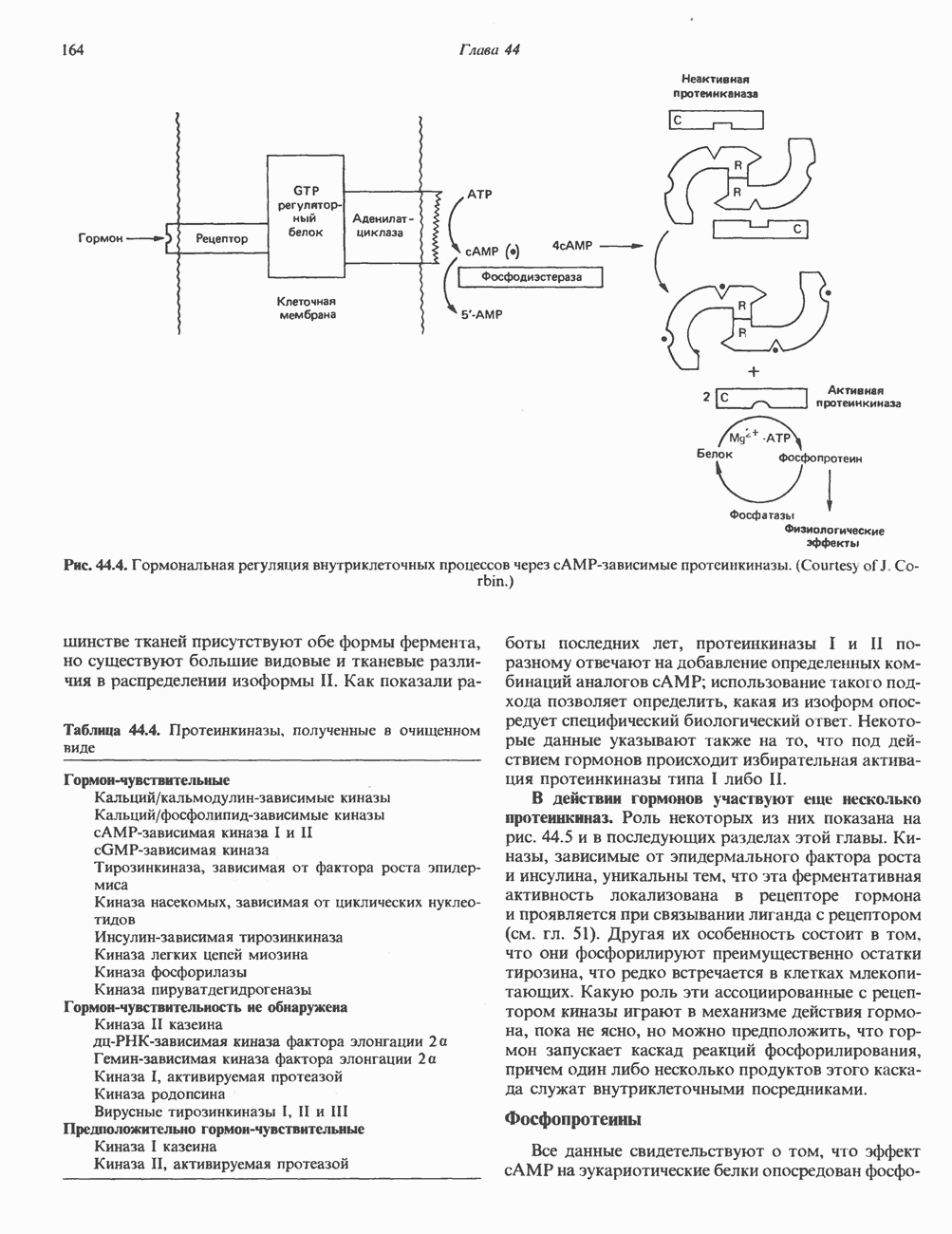
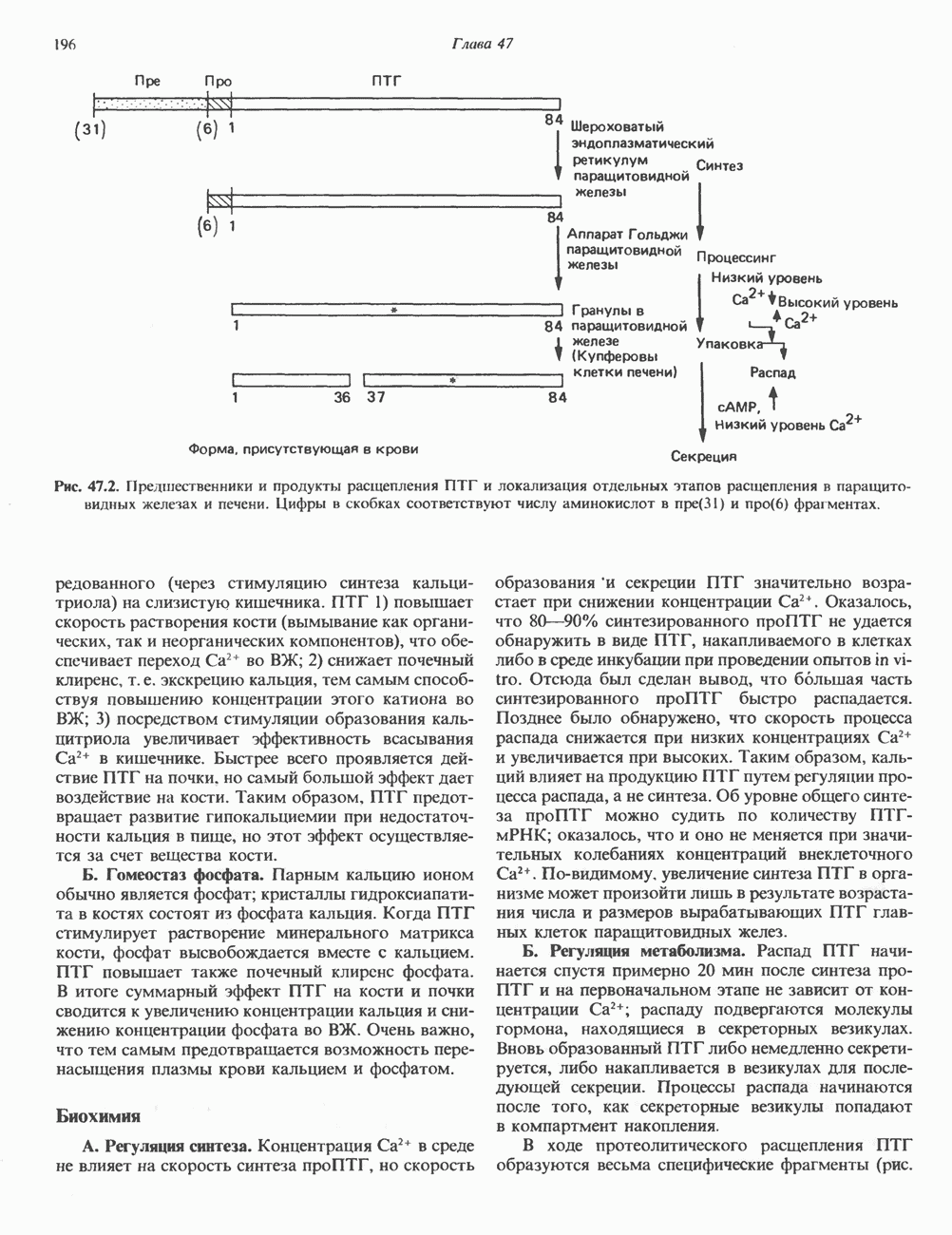
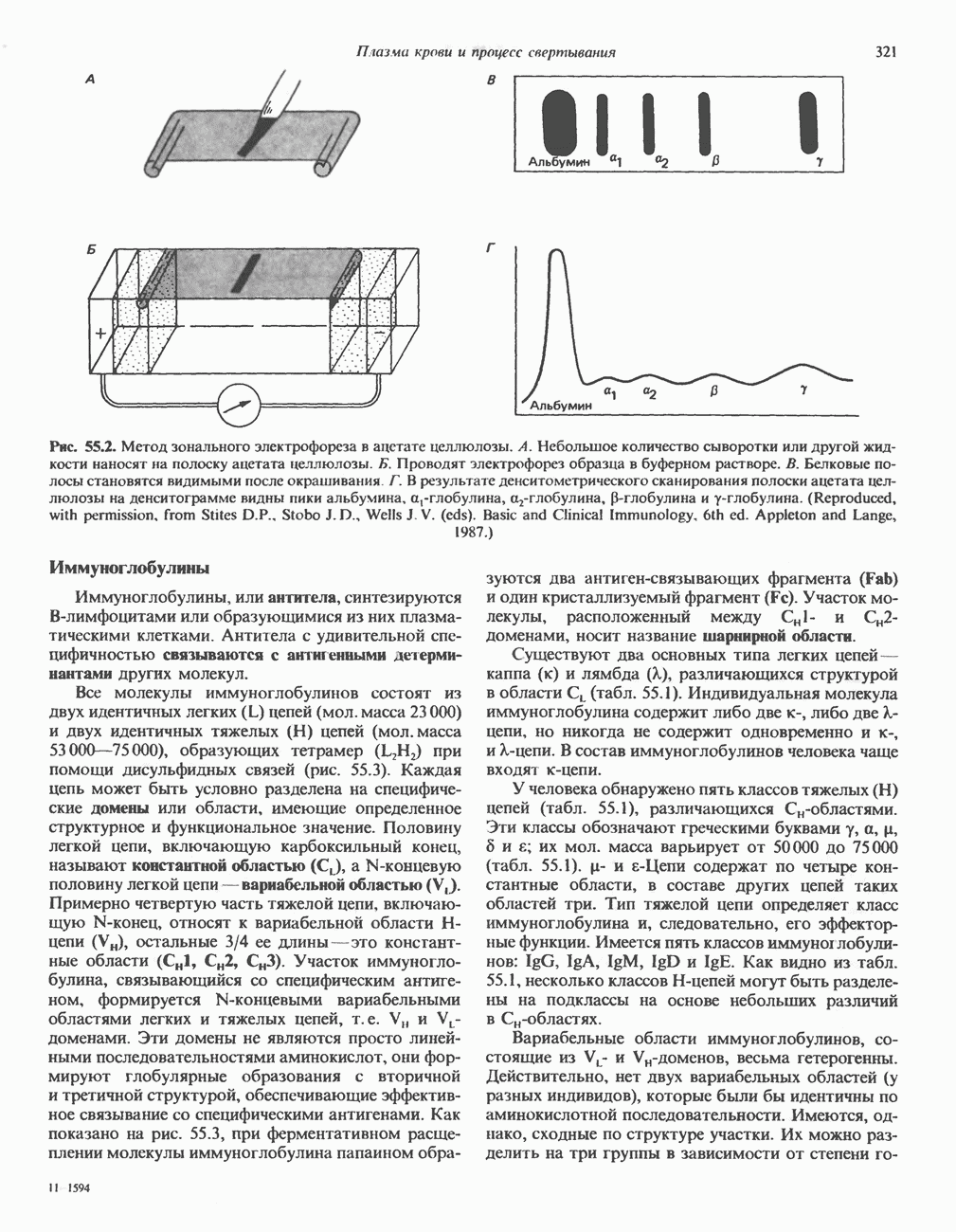
Рис. 38.9. Процесс кроссинговера гомологичных хромосом и образование рекомбинантных хромосом. хромосом, причем, как правило, этот процесс происходит с очень высокой точностью. Процесс кроссинговера схематически изображен на рис. 38.9. Он заключается в эквивалентном взаимном обмене генетической информацией между гомологичными хромосомами. Если гомологичные хромосомы несут различные аллели одного и того же гена, то в результате кроссинговера может произойти заметное и наследуемое изменение признаков. В редких случаях, когда при конъюгации гомологичные хромосомы располагаются друг относительно друга не совсем точно, может произойти неравный кроссинговер, в результате которого будет иметь место неэквивалентный обмен информацией. При этом одна из хромосом теряет часть генетической информации и, следовательно, несет делецию. Вторая хромосома получает большее количество генетического материала и, следовательно, несет вставку или дупликацию (рис. 38.9). Неравный кроссинговер у человека показан на примере гемоглобинов, названных Леноре (Lepore) и анти-Лепоре. Он может происходить в тандемных участках повторяющейся ДНК, например в последовательностях глобиновых генов или же в последовательностях более представительного семейства повторов ДНК (рис. 38. 10). Этот феномен ответствен за увеличение или уменьшение числа копий повторов данного семейства. Хромосомная интеграцияНекоторые бактериальные вирусы (бактериофаги) способны рекомбинировать с ДНК хозяина таким образом, что ДНК бактериофага встраивается в линейной форме в бактериальный геном. Интеграция бактериофага происходит по механизму, представленному
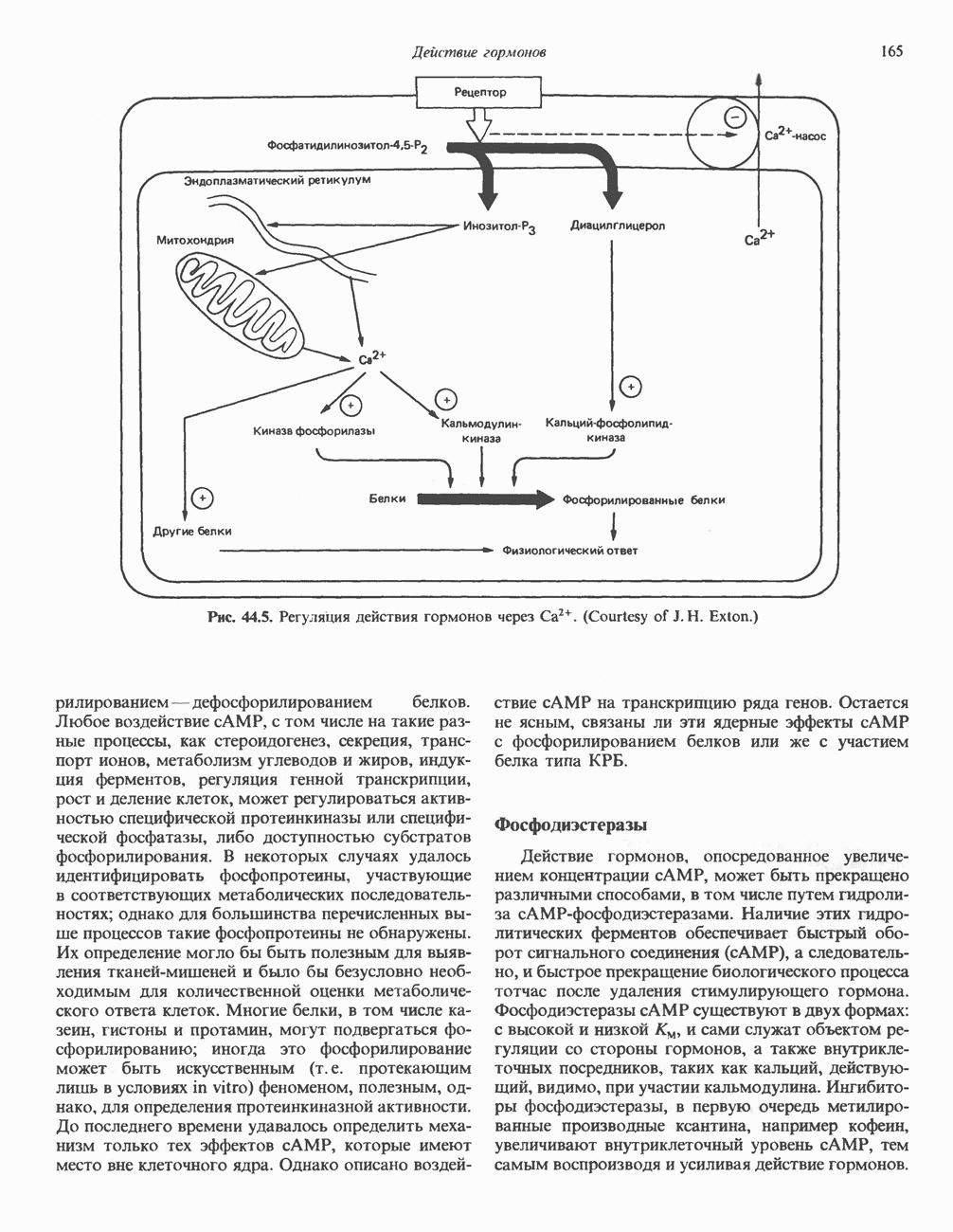
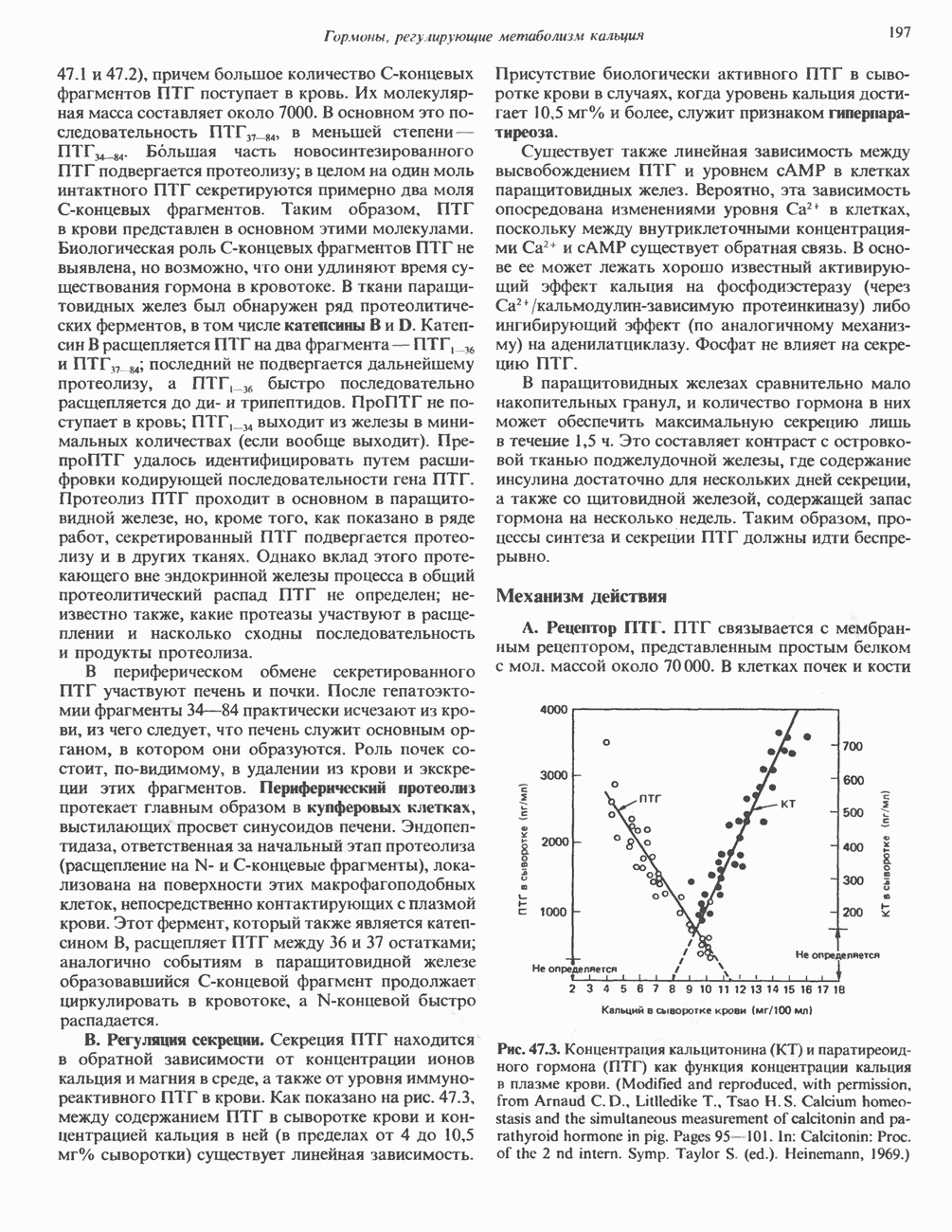
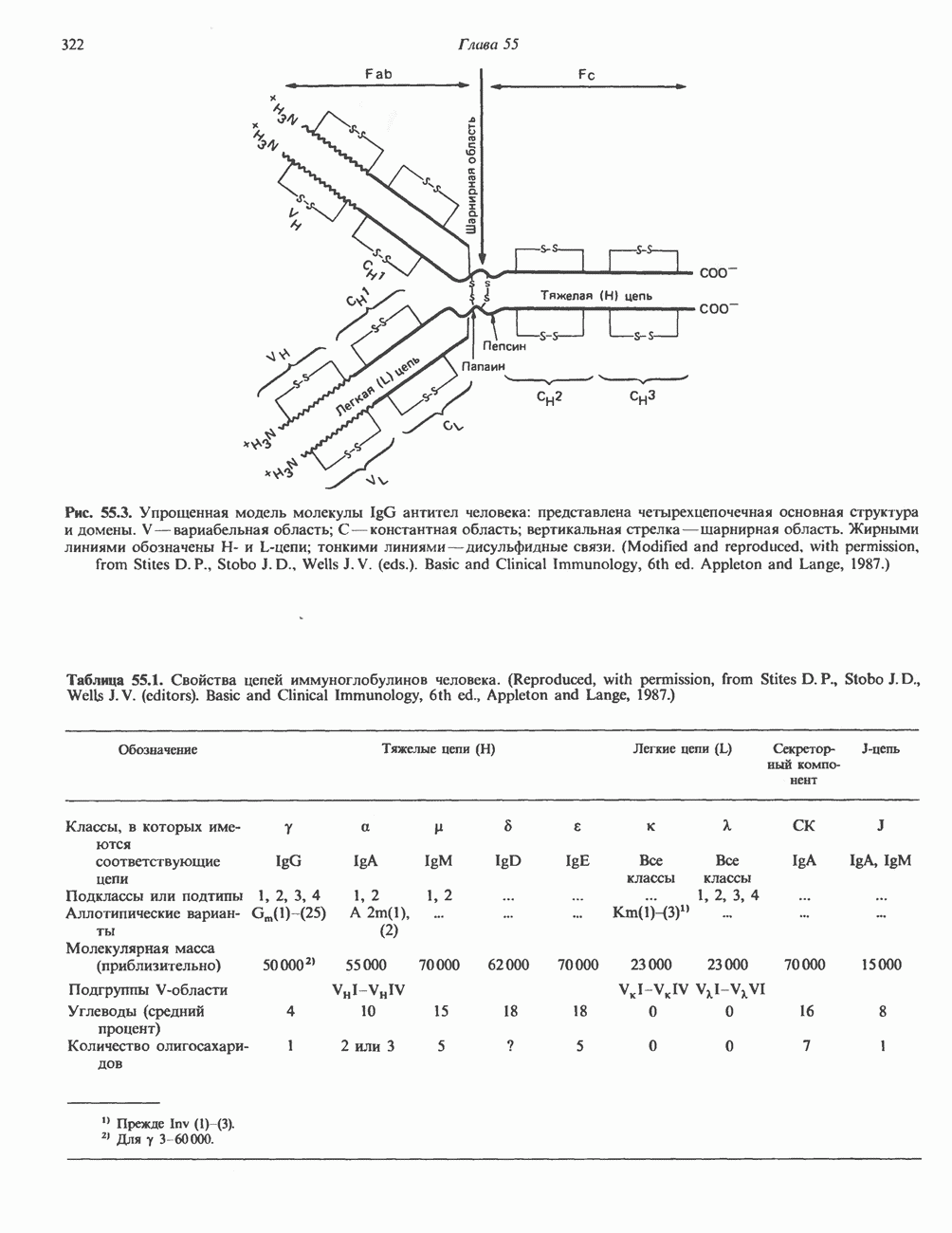
Рис. 38.10. Неравный кроссинговер в области структурных генов гемоглобинов человека. Продукты неравного кроссинговера: глобиновые гены типа дельта-бета Леноре и бета-дельта анти-Лепоре. В приведенных примерах показано расположение кроссоверных областей. (Reproduced, with permission, from Clegg J. B., Weatherall D. J. P - thalassemia: Time for reappraisal? Lancet 1974, 2: 133.) в упрощенном виде на рис. 38.11. При этом имеют место разрыв и соединение обеих молекул ДНК с соблюдением полярности. Следовательно, интеграция сопровождается линеаризацией — переходом кольцевой молекулы ДНК бактериофага в линейную форму. Известны два механизма интеграции генома бактериофага с бактериальным геномом. Если ДНК бактериофага содержит участки, гомологичные бактериальной ДНК, используется механизм, аналогичный рекомбинации гомологичных хромосом. Другой вариант интеграции осуществляется бактериофагами, которые синтезируют белки, направляющие процесс специфического связывания определенных участков последовательности (сайтов) бактериальной хромосомы с негомологичными сайтами в фаговой ДНК. Интеграция с помощью такого механизма носит название «сайт-специфической». Многие вирусы животных, особенно онкогенные вирусы, могут встраиваться в геном млекопитающих либо непосредственно, либо, в случае РНК-вирусов через ДНК-транскрипты. Интеграция вирусной ДНК в хромосомы животных, как правило, не является сайт-специфической.
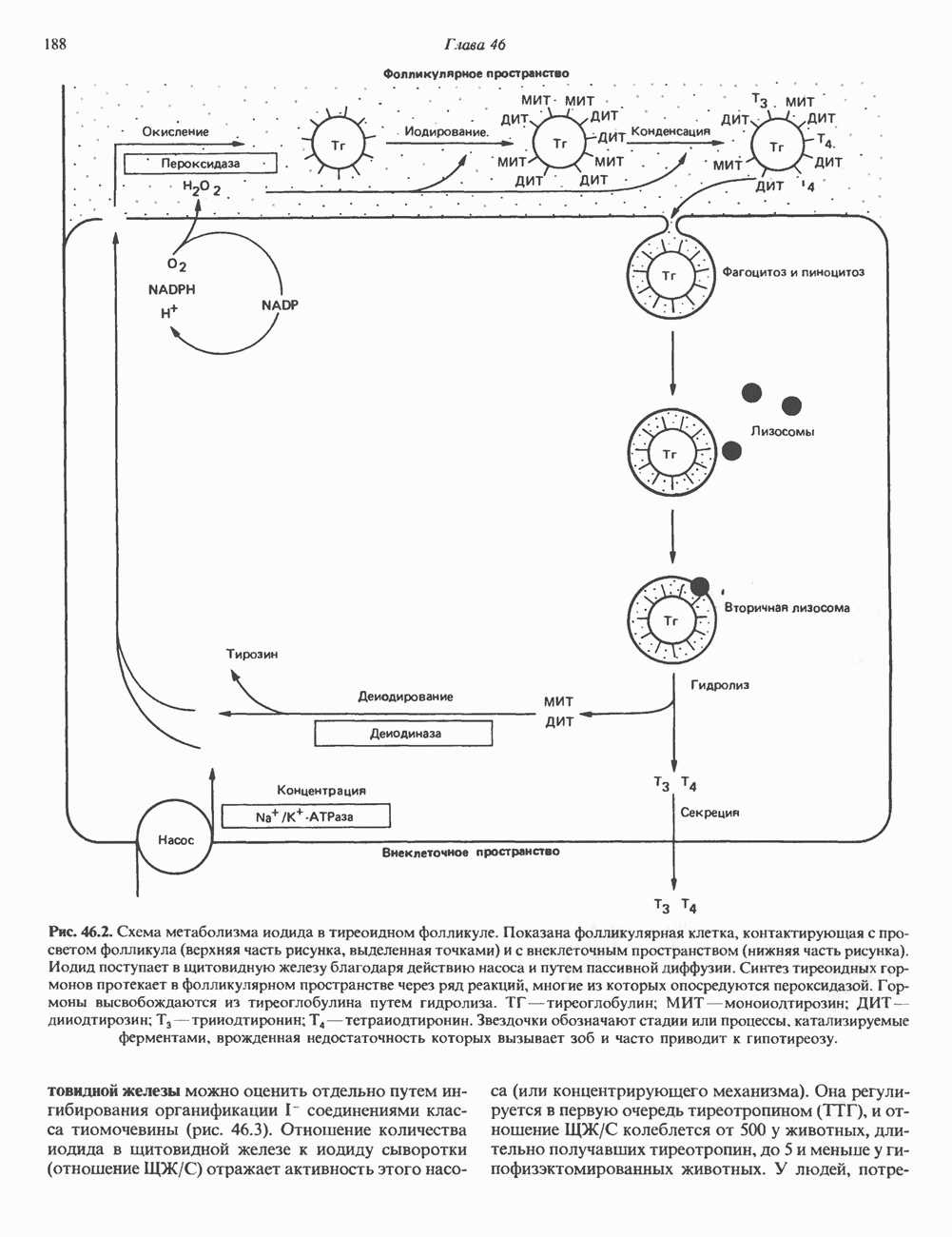
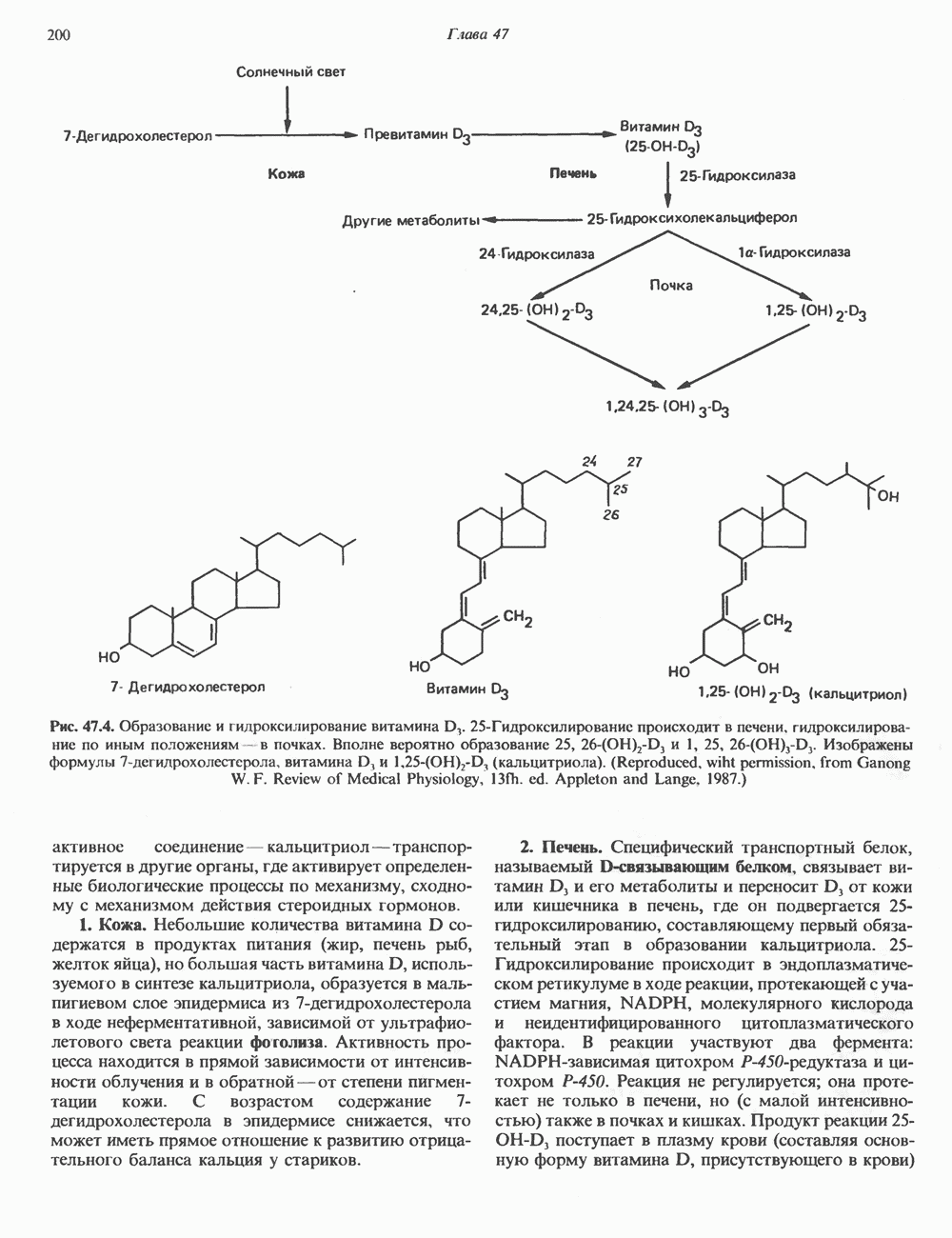
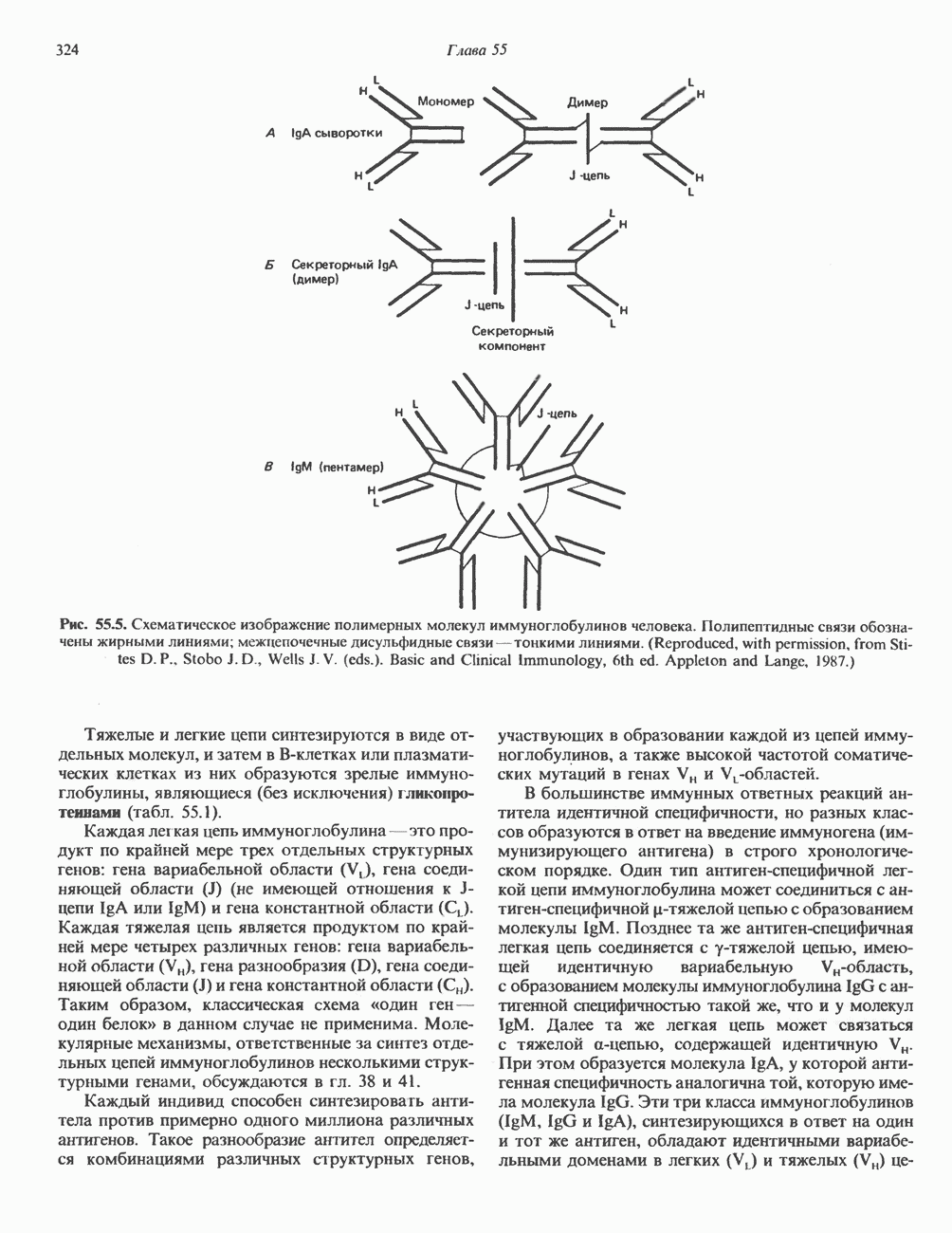
Рис. 38.11. Встраивание кольцевого генома (содержащего гены А, В, С) в хозяйскую молекулу ДНК (содержащую гены 1 и 2) и порядок чередования генов в рекомбинантной цепи ДНК. ТранспозицииВ эукариотическом геноме имеются небольшие элементы ДНК, не являющиеся провирусами, но способные самостоятельно вырезаться из хозяйского генома, а затем встраиваться в различные его участки, влияя при этом на функции прилегающих последовательностей ДНК. Эти подвижные (мобильные) элементы, которые иногда называют «прыгающая ДНК», могут перемещать фрагменты хромосомной ДНК и таким путем глубоко воздействовать на процессы эволюции генома. Как указывалось выше, семейство коротких Alu-повторов характеризуется наличием структурного сходства с концевыми последовательностями ретровирусов, благодаря которым последние могут встраиваться в геном млекопитающих и покидать его. Прямым доказательством транспозиций других небольших элементов ДНК в геноме человека явилось открытие так называемых «процесснрованных генов» иммуноглобулинов, а-глобинов и некоторых других. Процессированные гены идентичны или почти идентичны последовательностям зрелых мРНК данных генов. Они состоят из нетранскрибируемого 5-участка гена, кодирующей области без интронов и poly A-последовательности на З-конце. Появление процесснрованных генов можно объяснить только интеграцией обратных транскриптов соответствующих зрелых мРНК. Судя по всему, единственным возможным способом внедрения таких обратных транскриптов является транспозиция. Действительно, оба конца процесснрованных генов фланкируются короткими повторами, сходными с теми, которые имеют мобильные элементы низших организмов. Некоторые из процесснрованных генов содержат случайным образом распределенные изменения последовательности, накопившиеся в ходе эволюции. Подобные изменения часто приводят к образованию nonsense - (бессмысленных) - кодонов, препятствующих экспрессии (см. гл. 40). Такие процессированные гены называют псевдогенами. Генная конверсияКроме неравного кроссинговера и транспозиций существует и третий механизм быстрых изменений генетического материала. Одинаковые последовательности гомологичных или негомологичных хромосом могут формировать случайные пары, а несовпадающие участки — удаляться. В результате происходит закрепление определенного варианта повторов данного семейства. Этот процесс получил название генной конверсии. Диплоидные клетки эукариотических организмов (в том числе человека) после прохождения S-фазы клеточного цикла содержат тетраплоидный набор хромосом. Каждая из сестринских хроматид (хромосомных
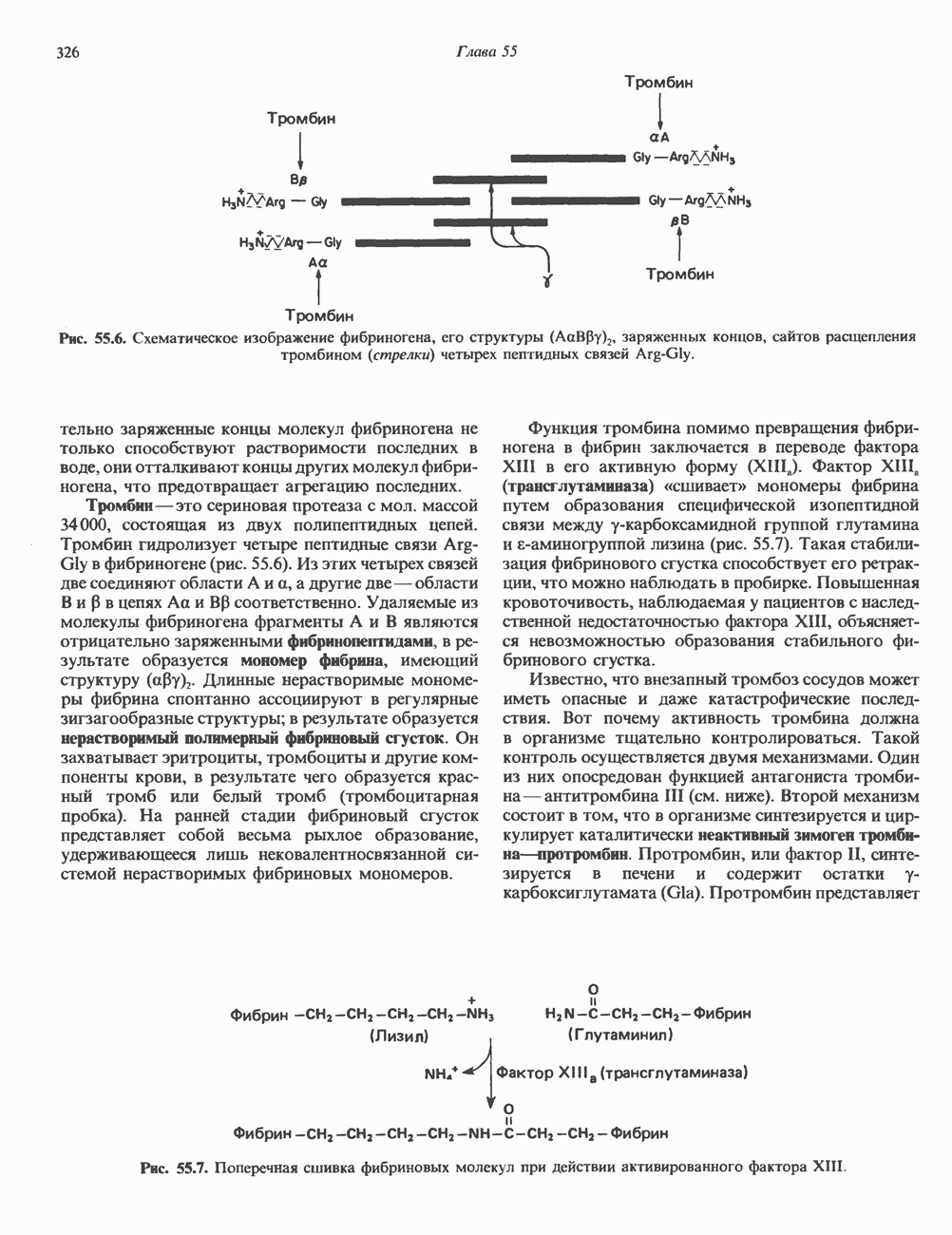
Рис. 38.12. Обмен между сестринскими хроматидами у человека. Окраска хромосом по Гимза после двух циклов репликации в присутствии бромдезоксиуридина. (Courtesy of S. Wolff and J. Bodycote.) пар) содержит одну и ту же генетическую информацию, поскольку обе они — результат полуконсервативной репликации родительских ДНК-молекул. Между этими генетически идентичными хроматидами может происходить кроссинговер. Обмен генетической информацией между сестринскими хроматидами (рис. 38.12) проявляется в форме равного кроссинговера и не имеет каких-либо генетических последствий. Некоторые интересные генетические перестройки происходят в клетках млекопитающих в ходе нормального развития и дифференцировки. Например, в клетках зародышевой линии мыши гены VL и CL, кодирующие единичную цепь молекулы иммуноглобулина (см. гл. 41), разнесены в геноме на значительное расстояние. В ДНК зрелых иммуноглобулин - продуцирующих (плазматических) клеток эти же гены оказываются на более близком расстоянии и транскрибируются в составе единого первичного транскрипта. Однако и после перестройки ДНК в ходе дифференцировки последовательности этих генов непосредственно не смыкаются. Между ними располагается промежуточная некодирующая последовательность (интрон) длиной около 1200 пар оснований, которая удаляется из первичного транскрипта при процессинге в ходе созревания мРНК (см. гл. 39 и 41).
|
 |
Оглавление
|