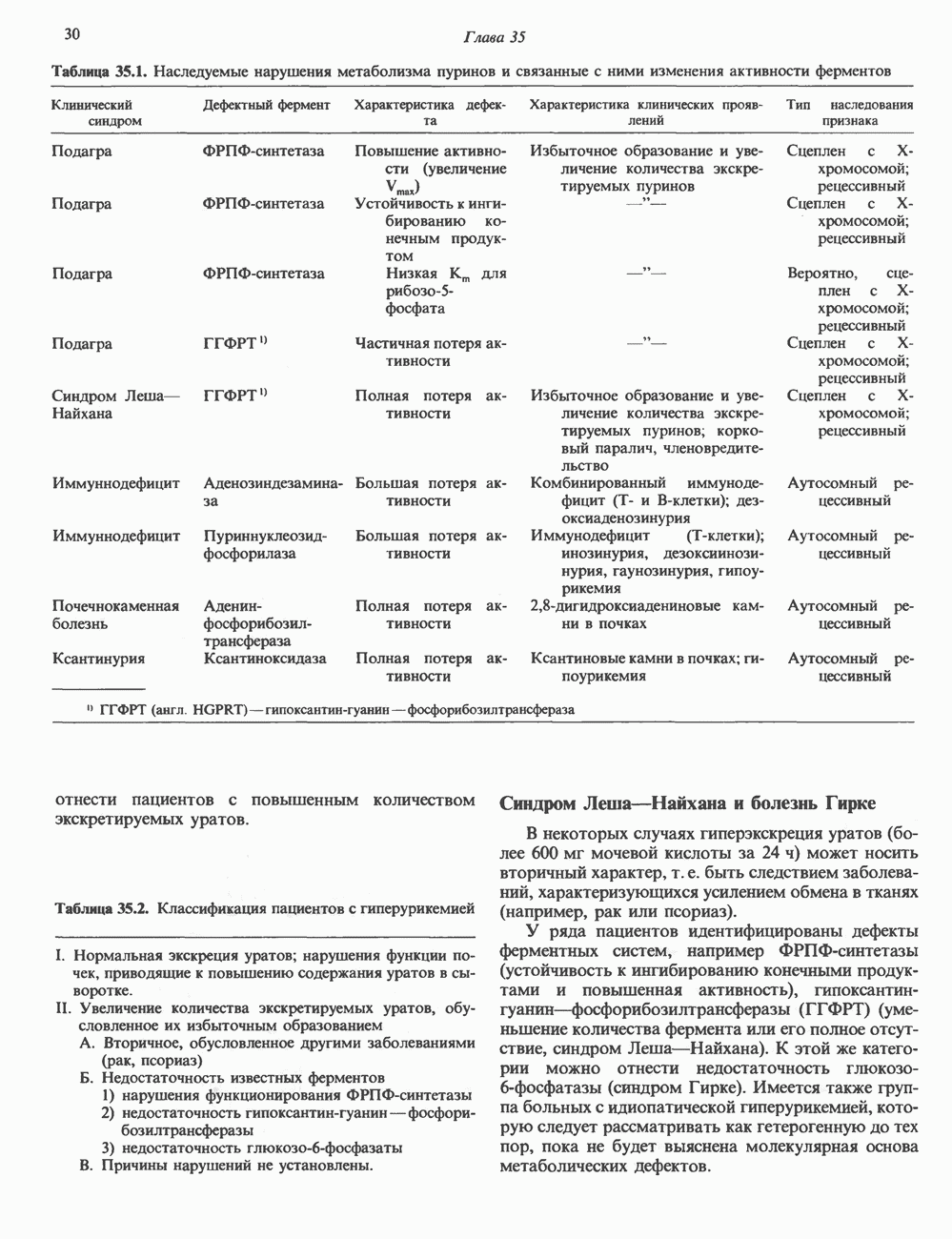
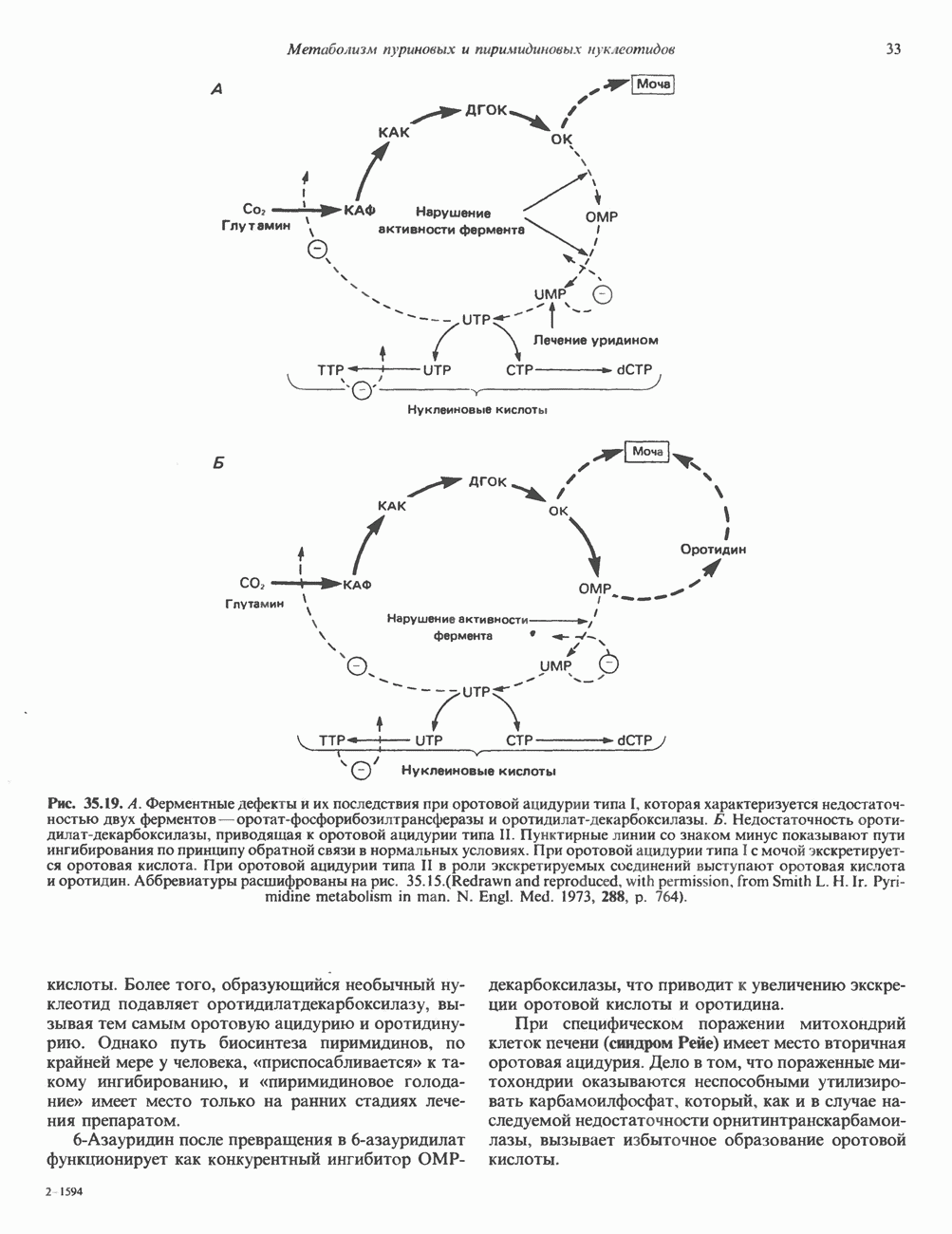
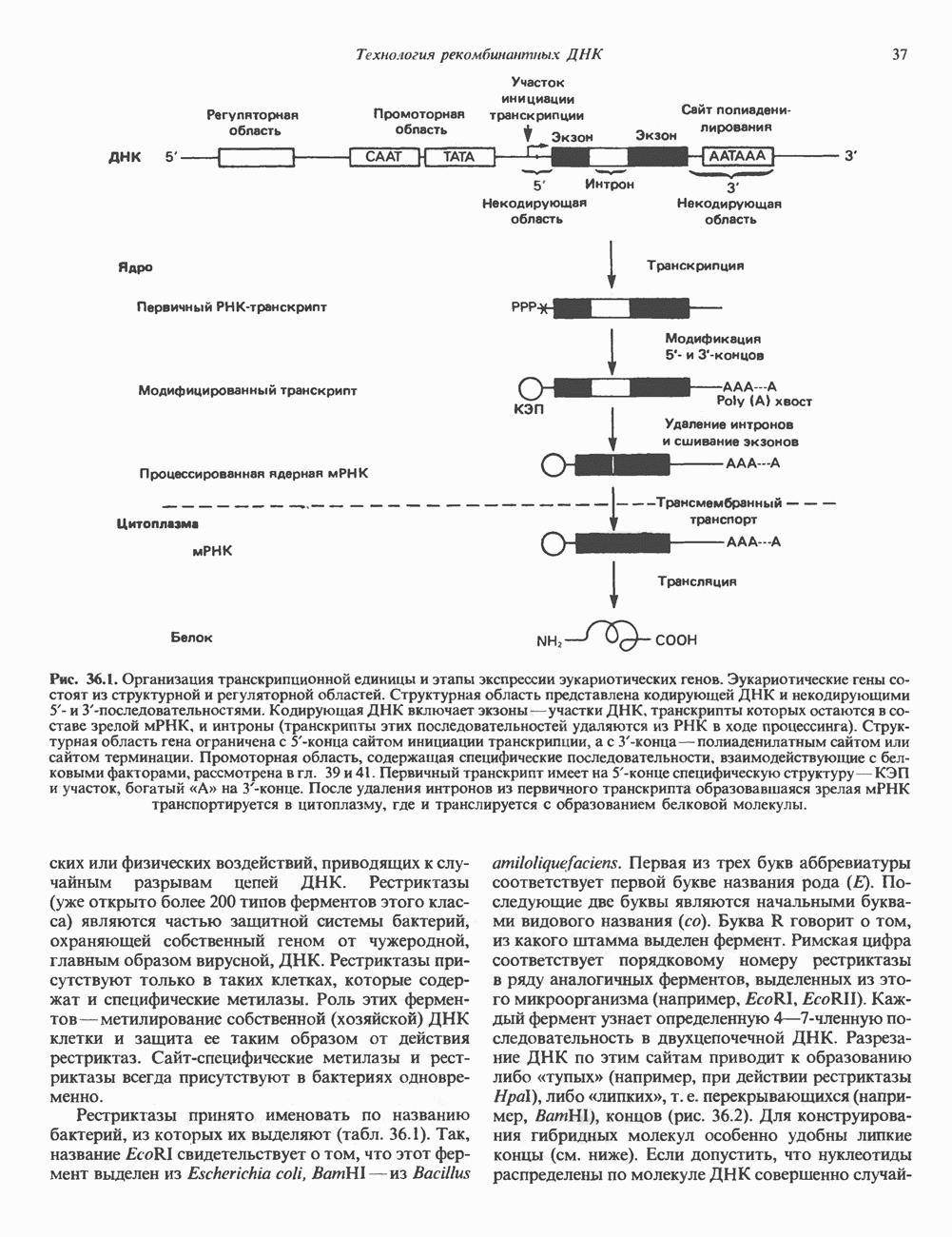
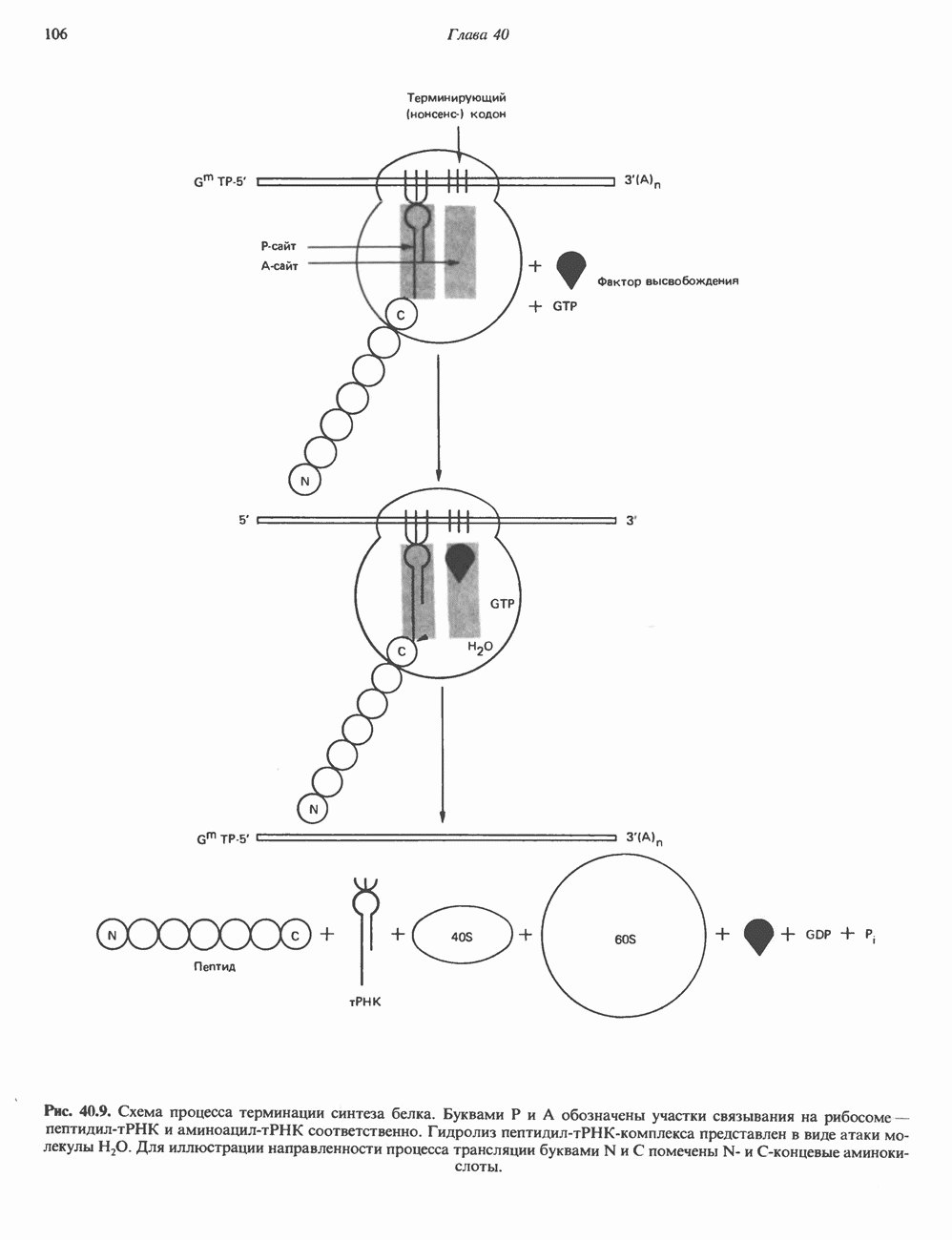
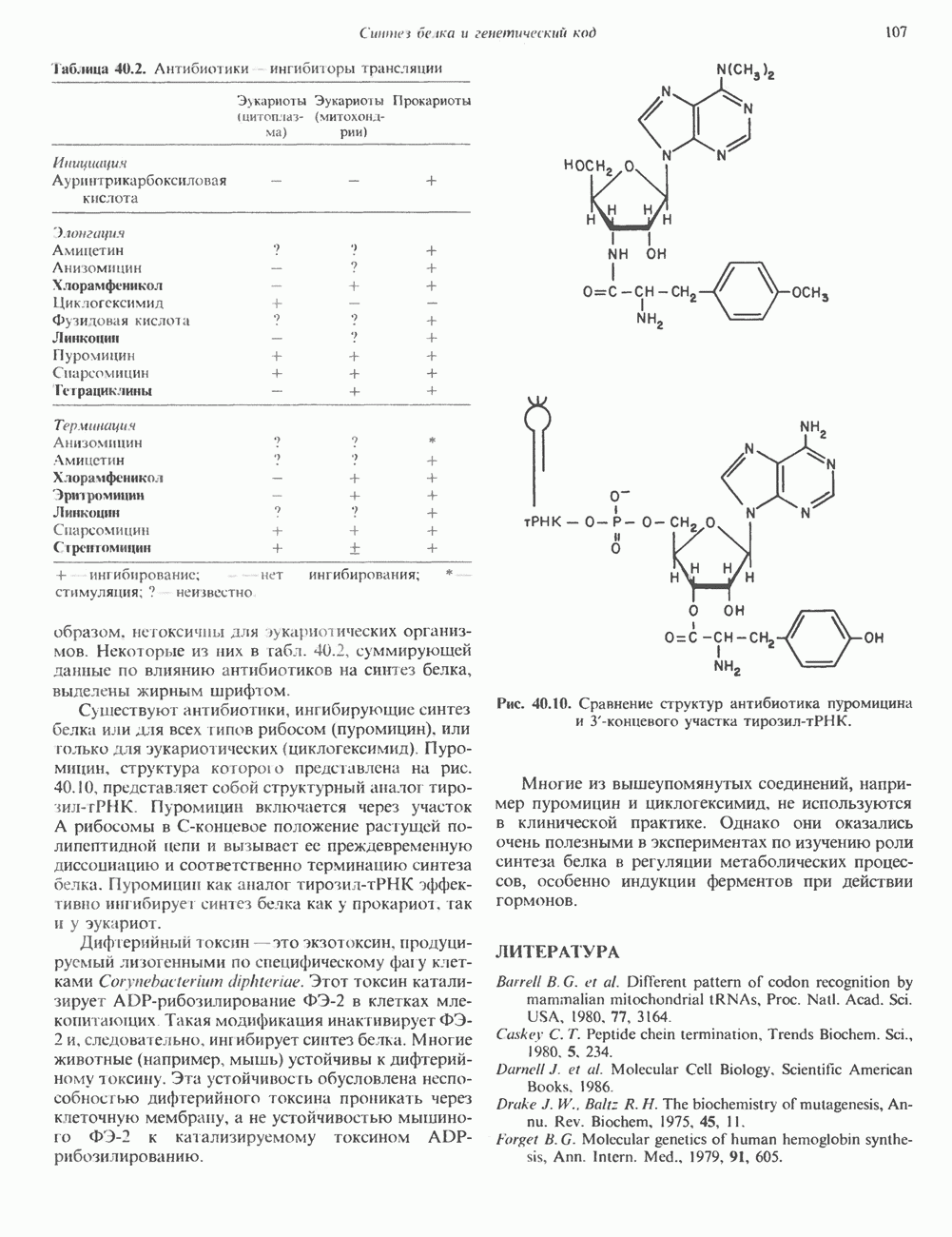
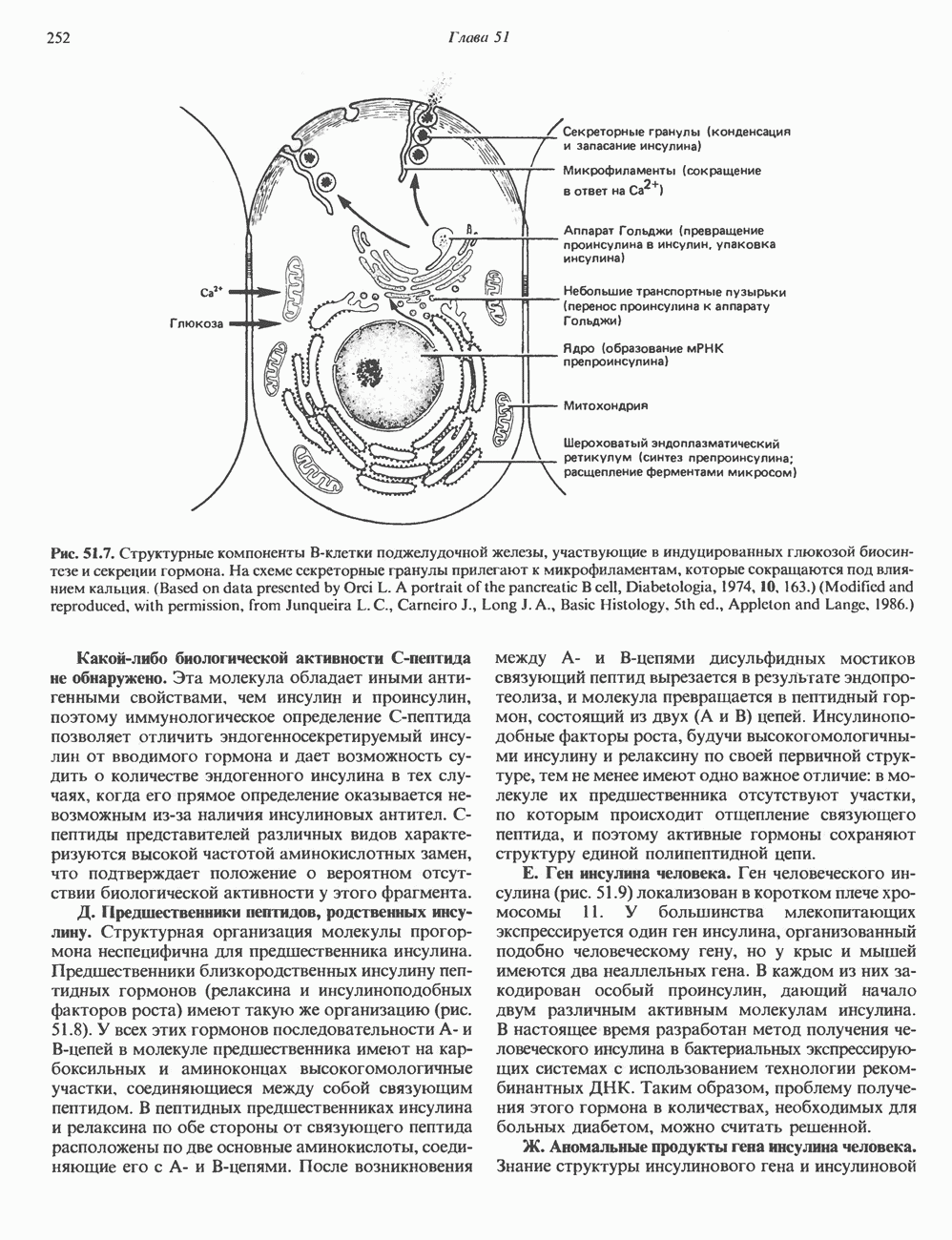
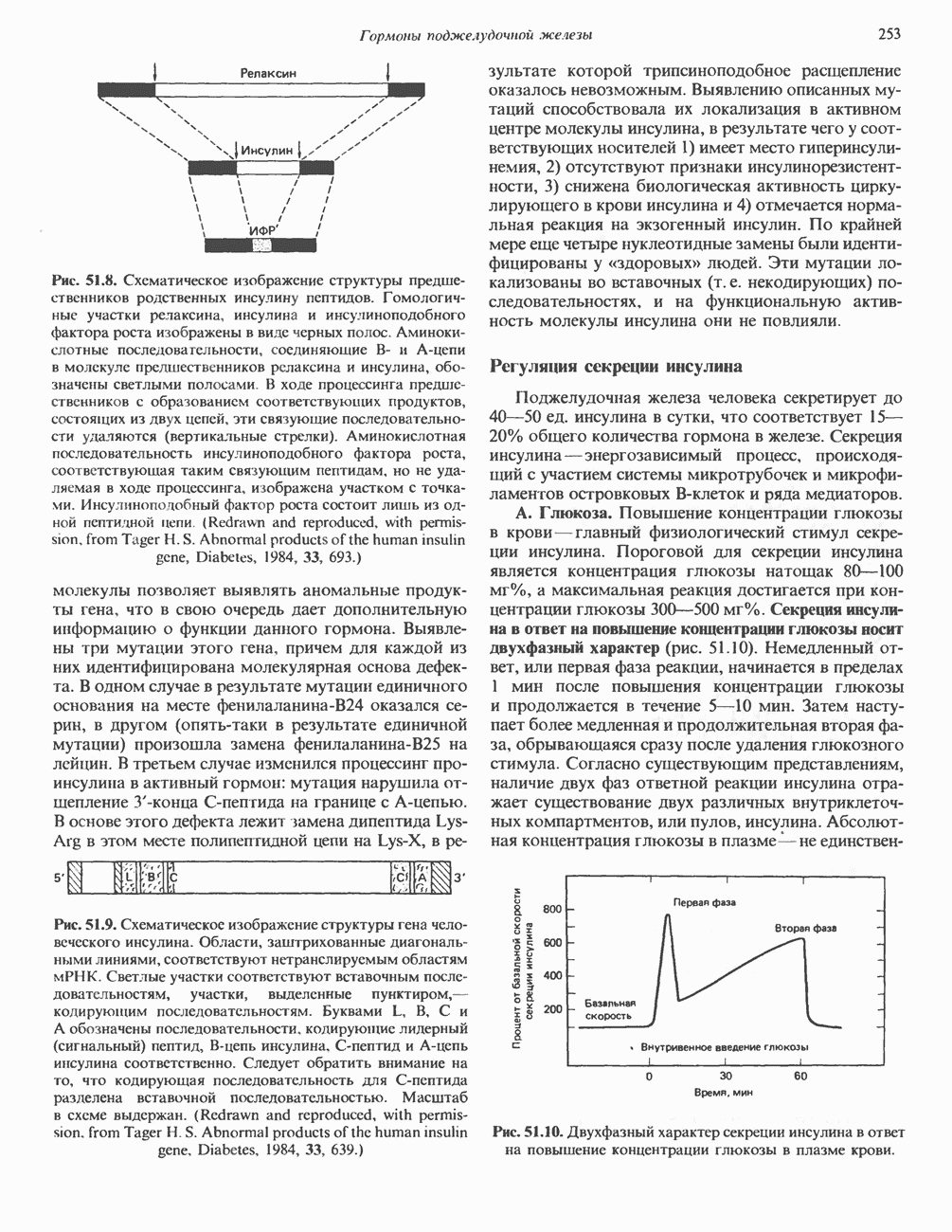
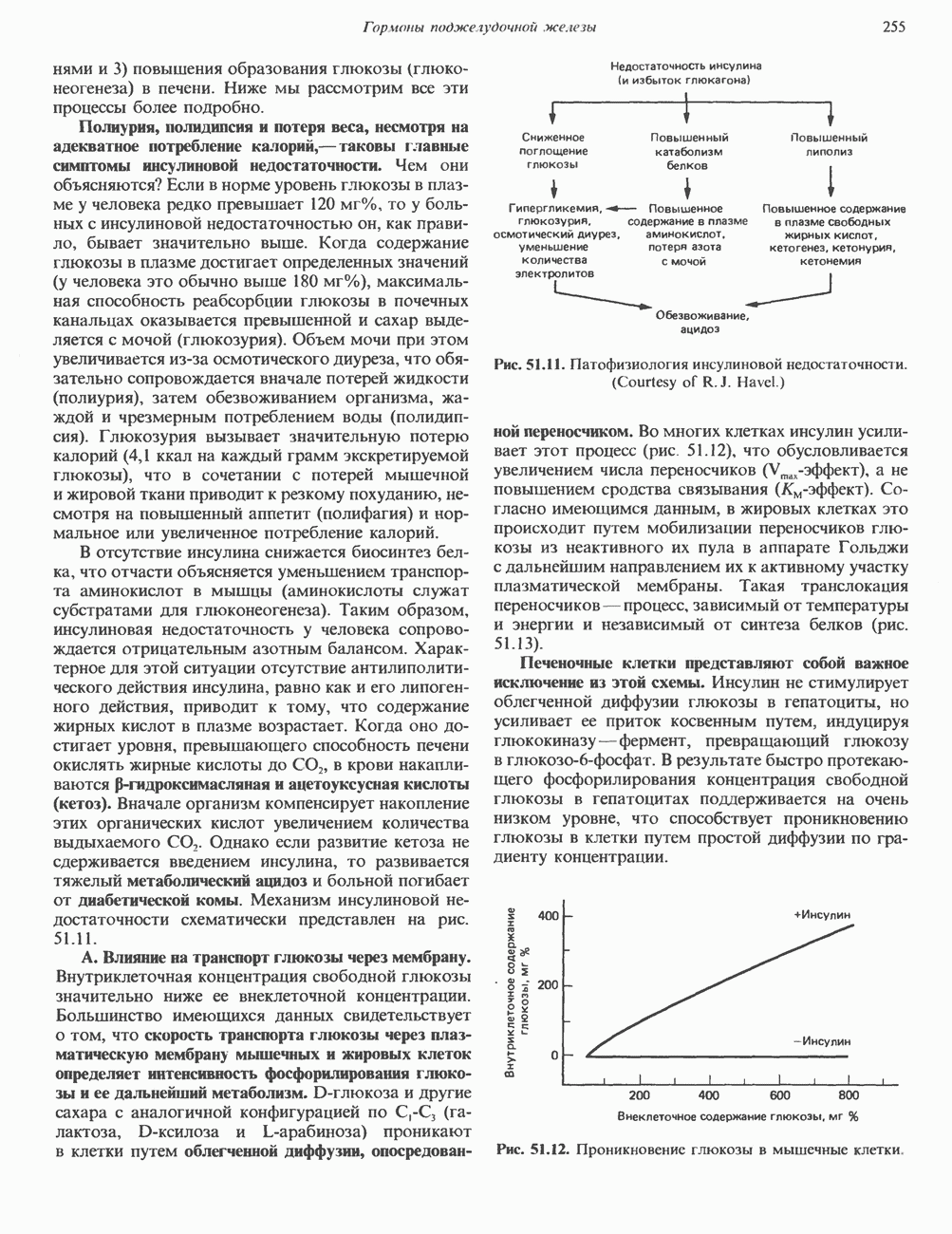
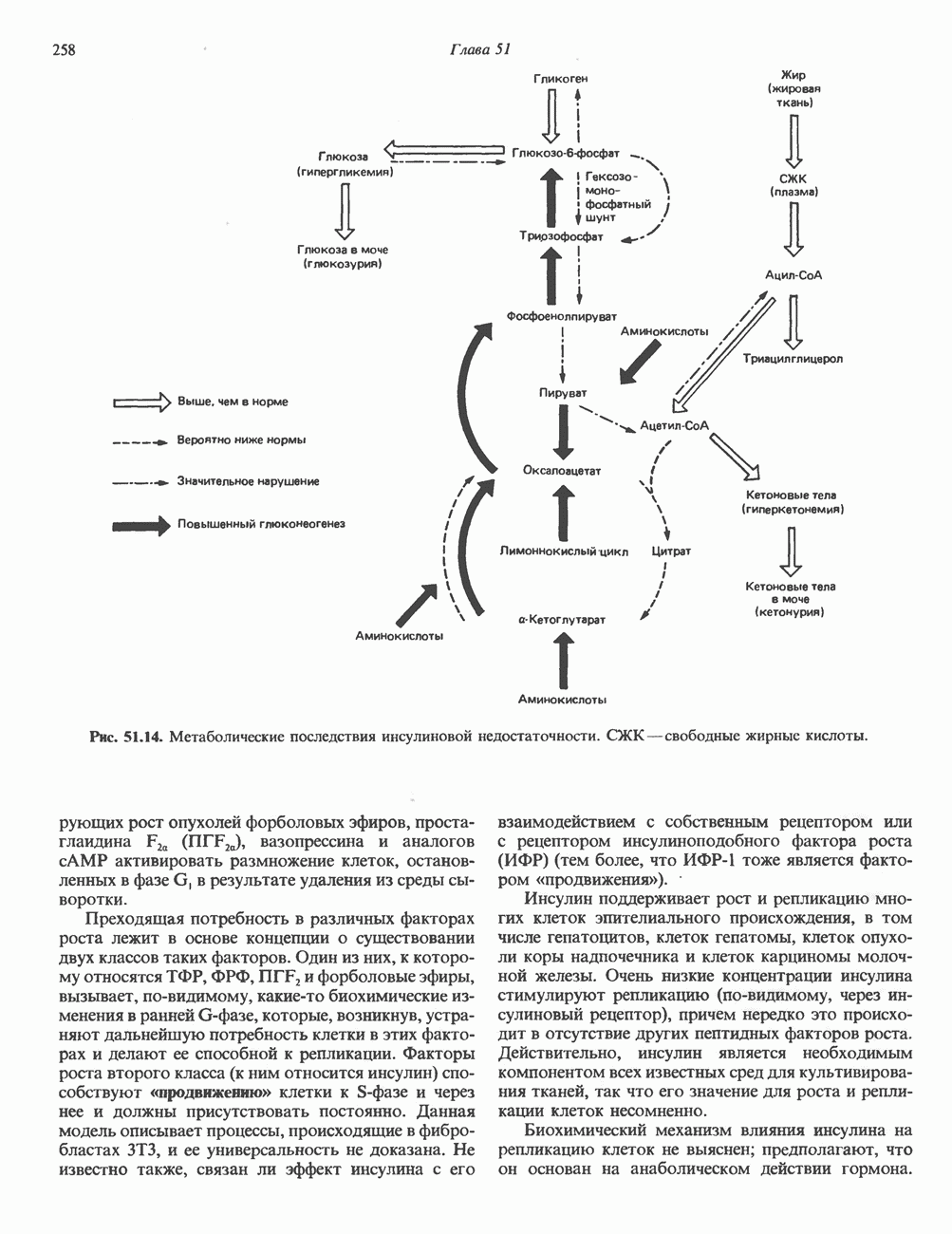
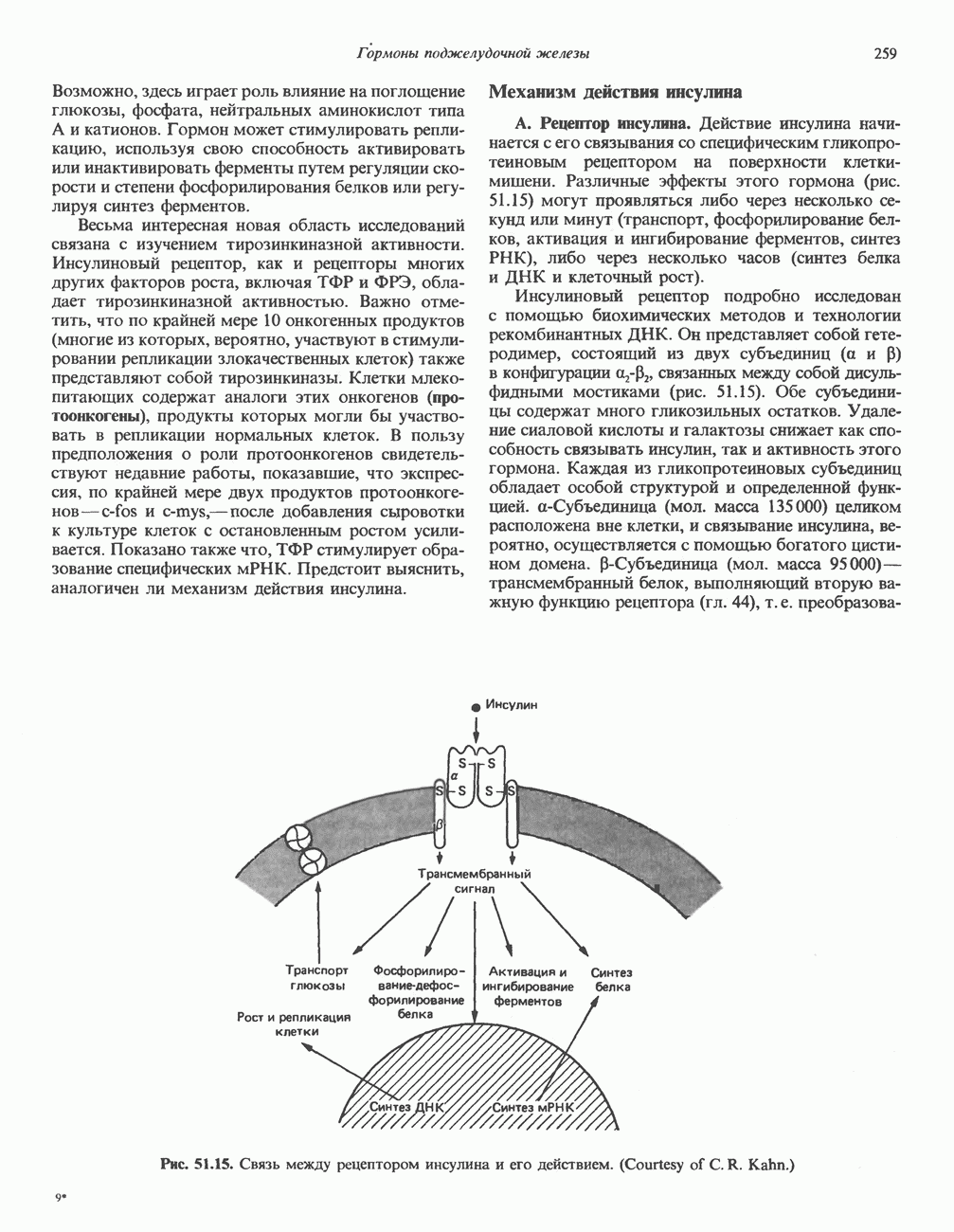
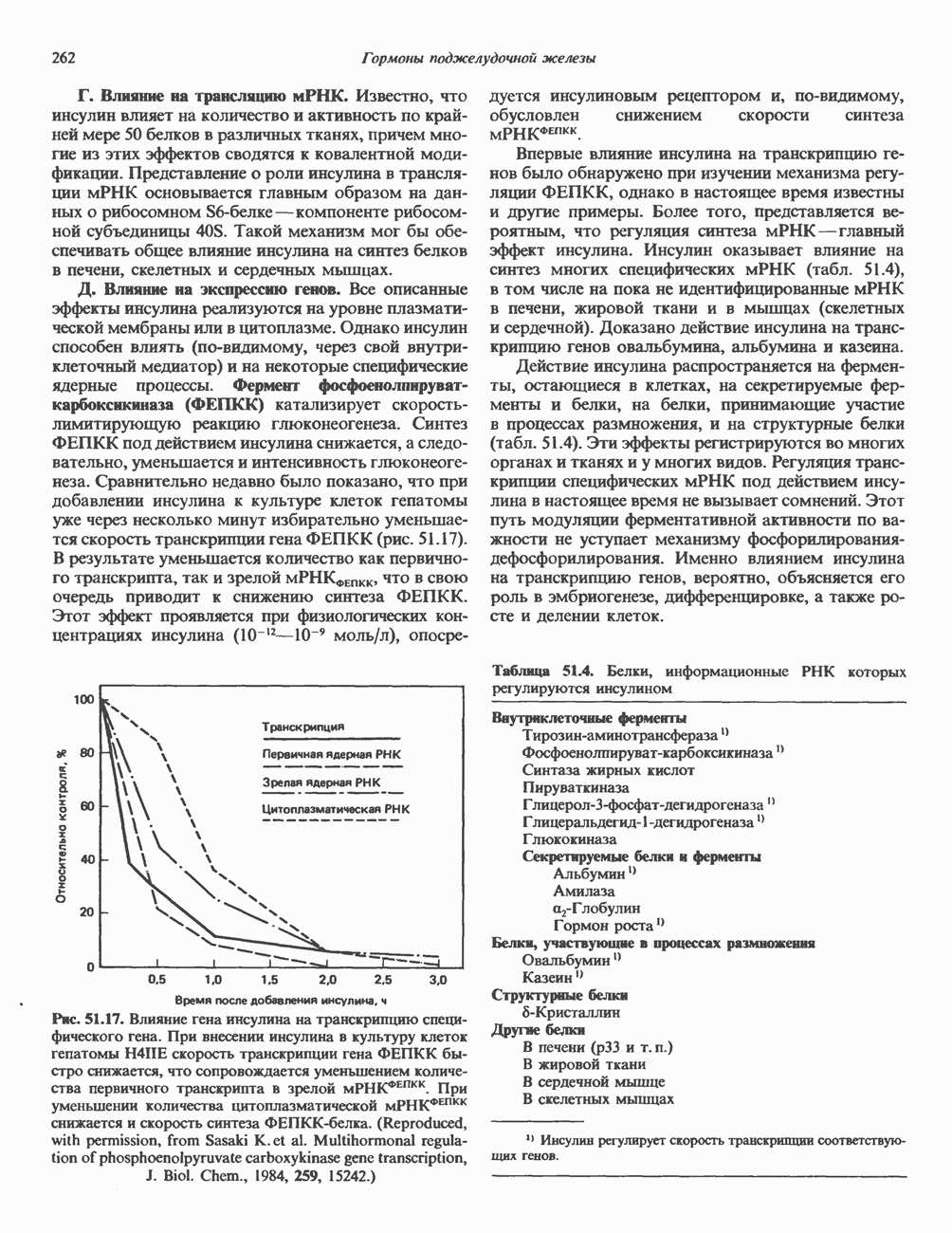
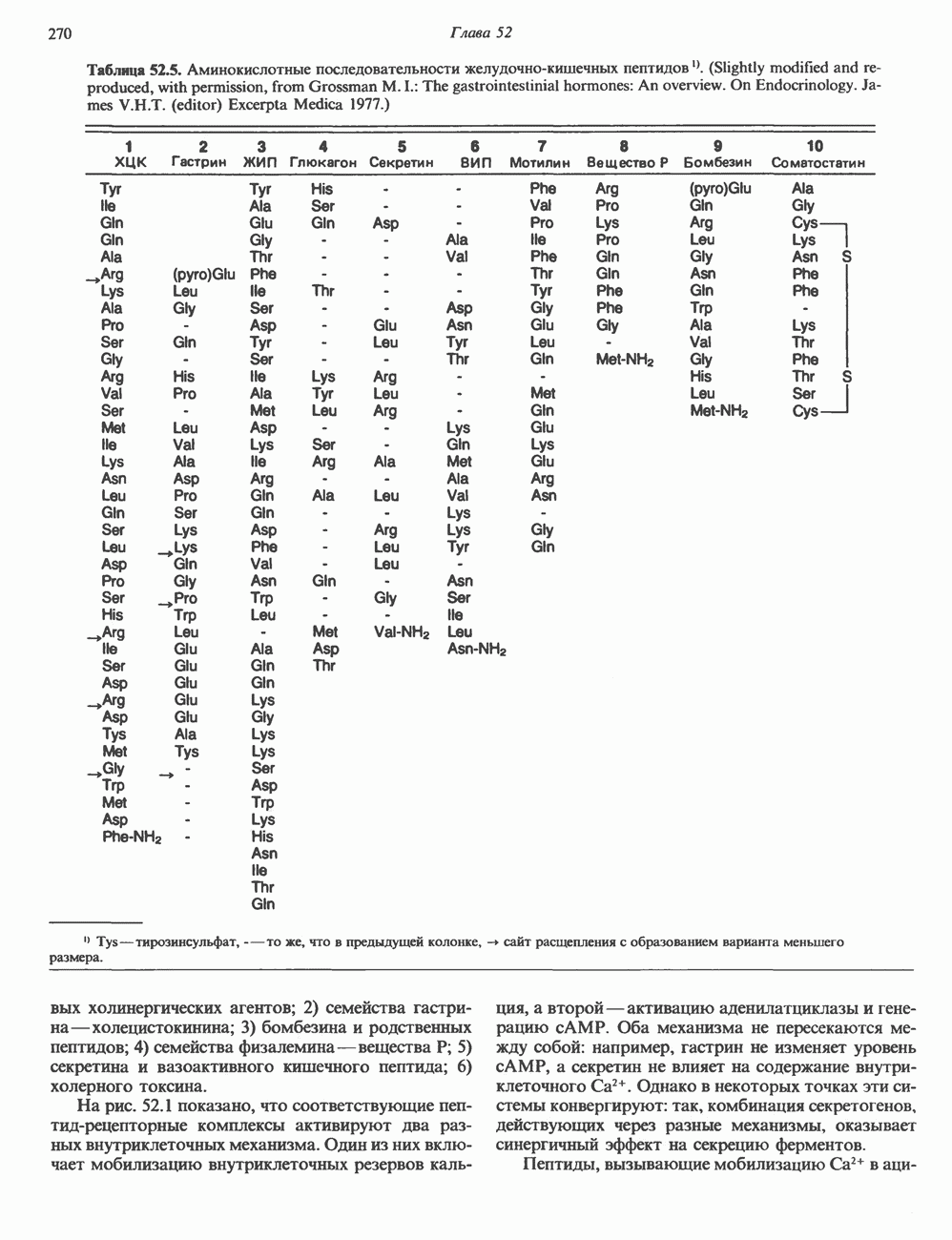
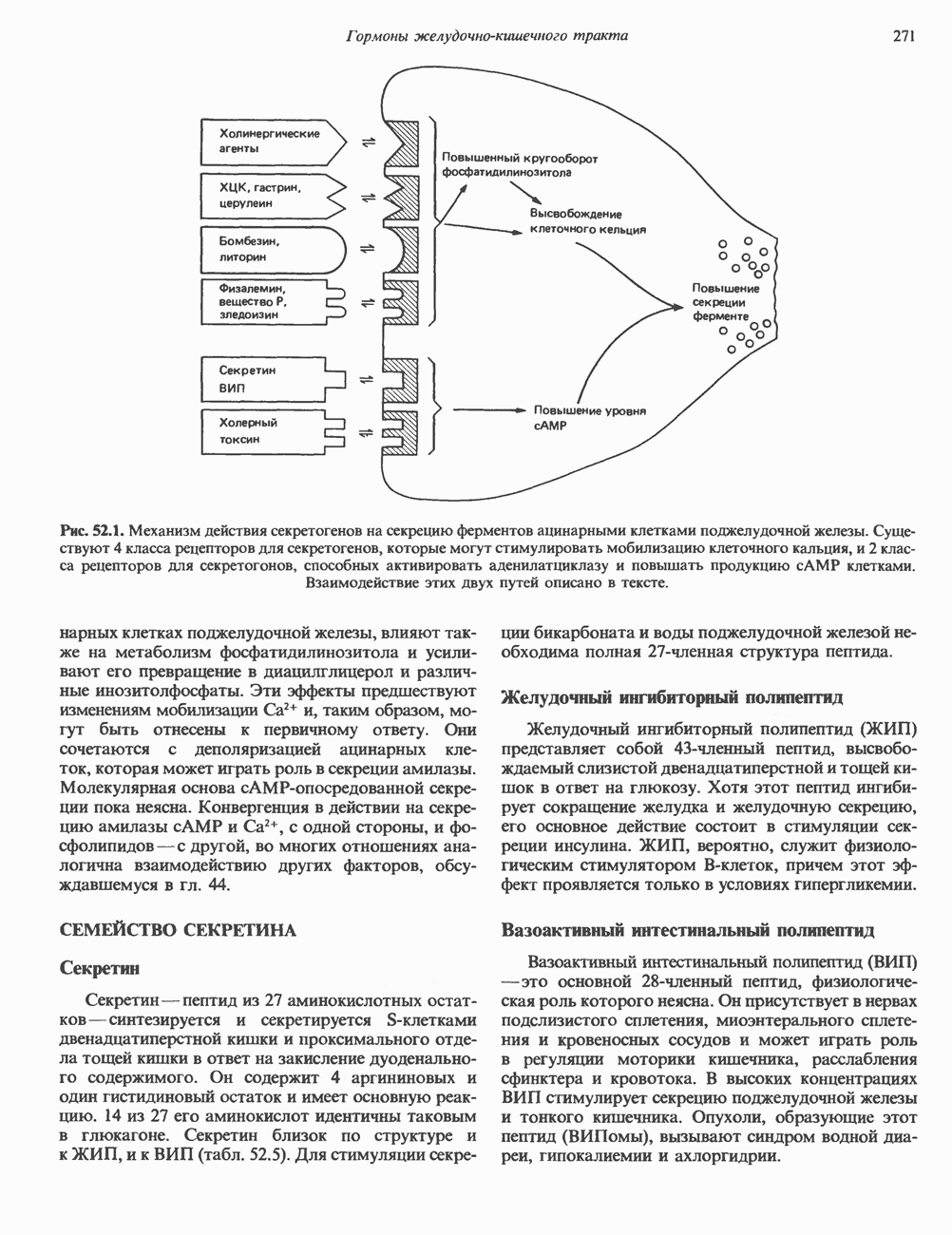
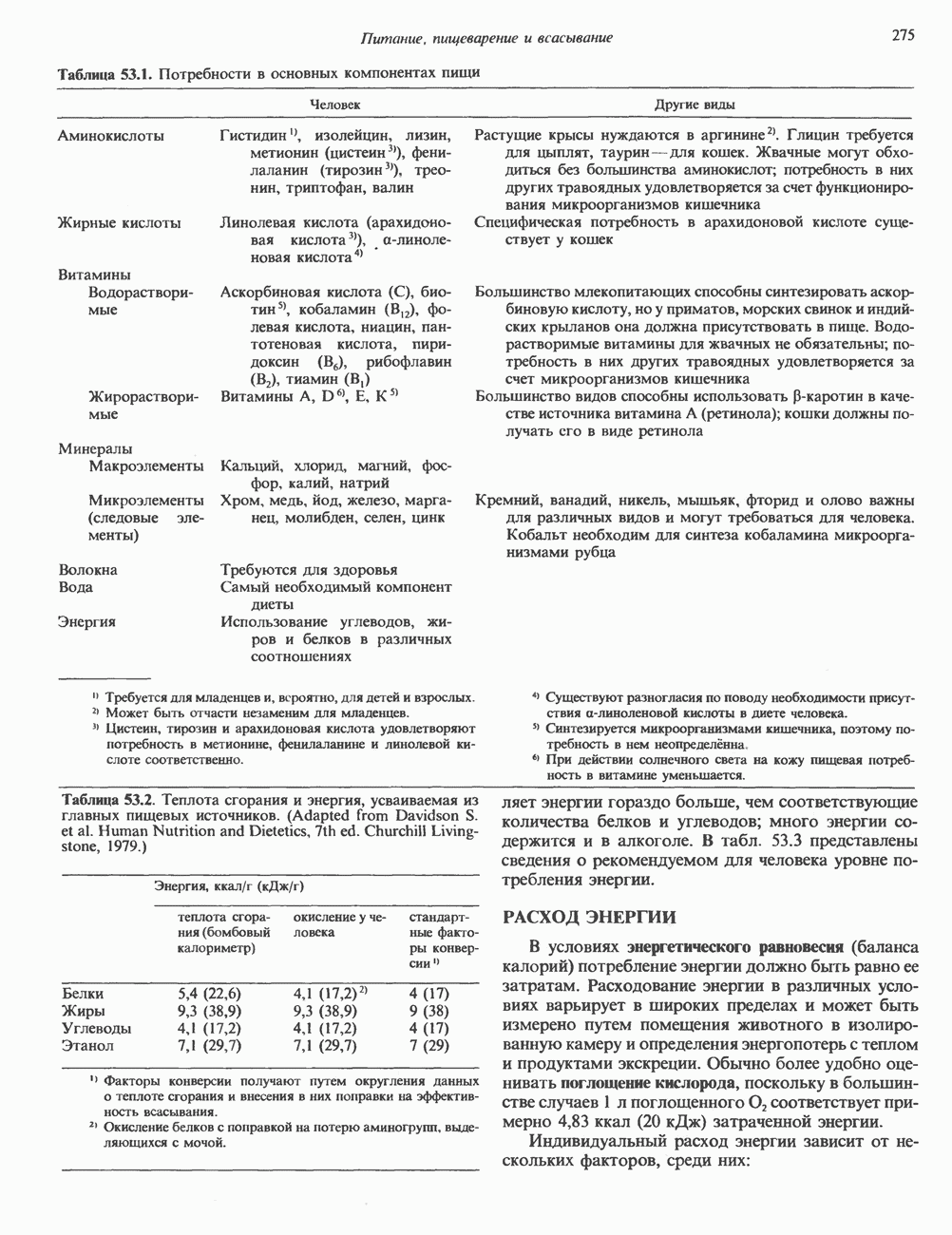
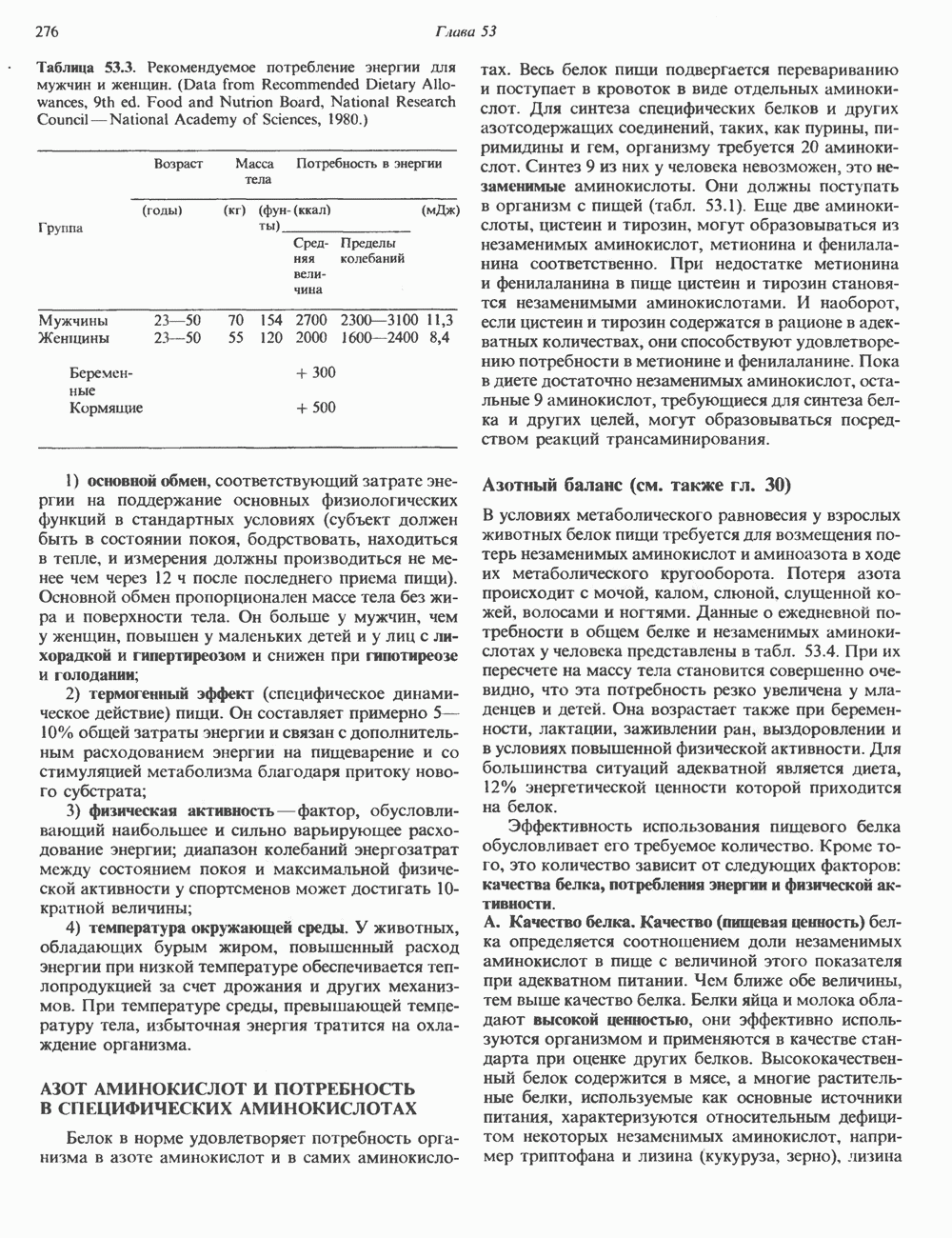
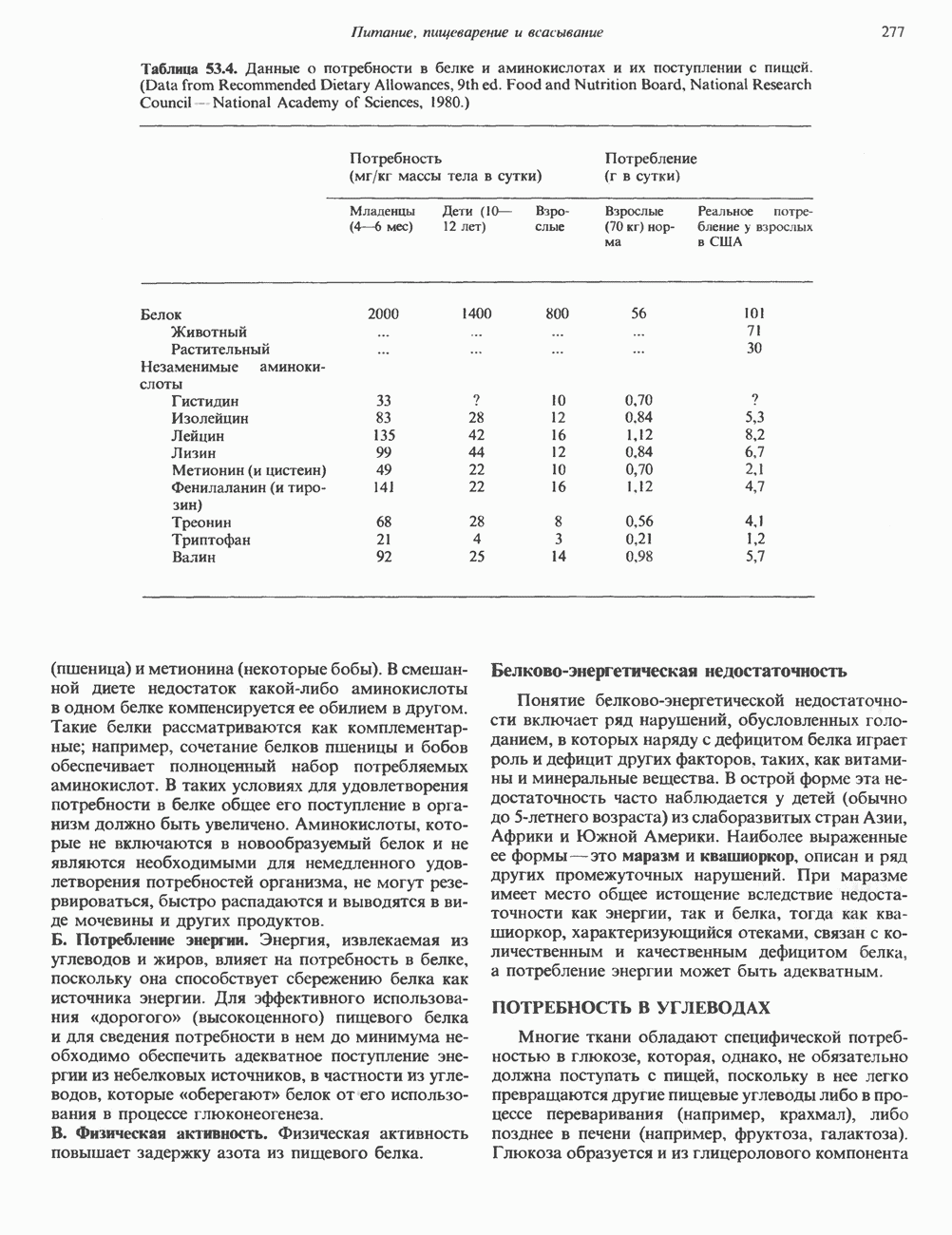
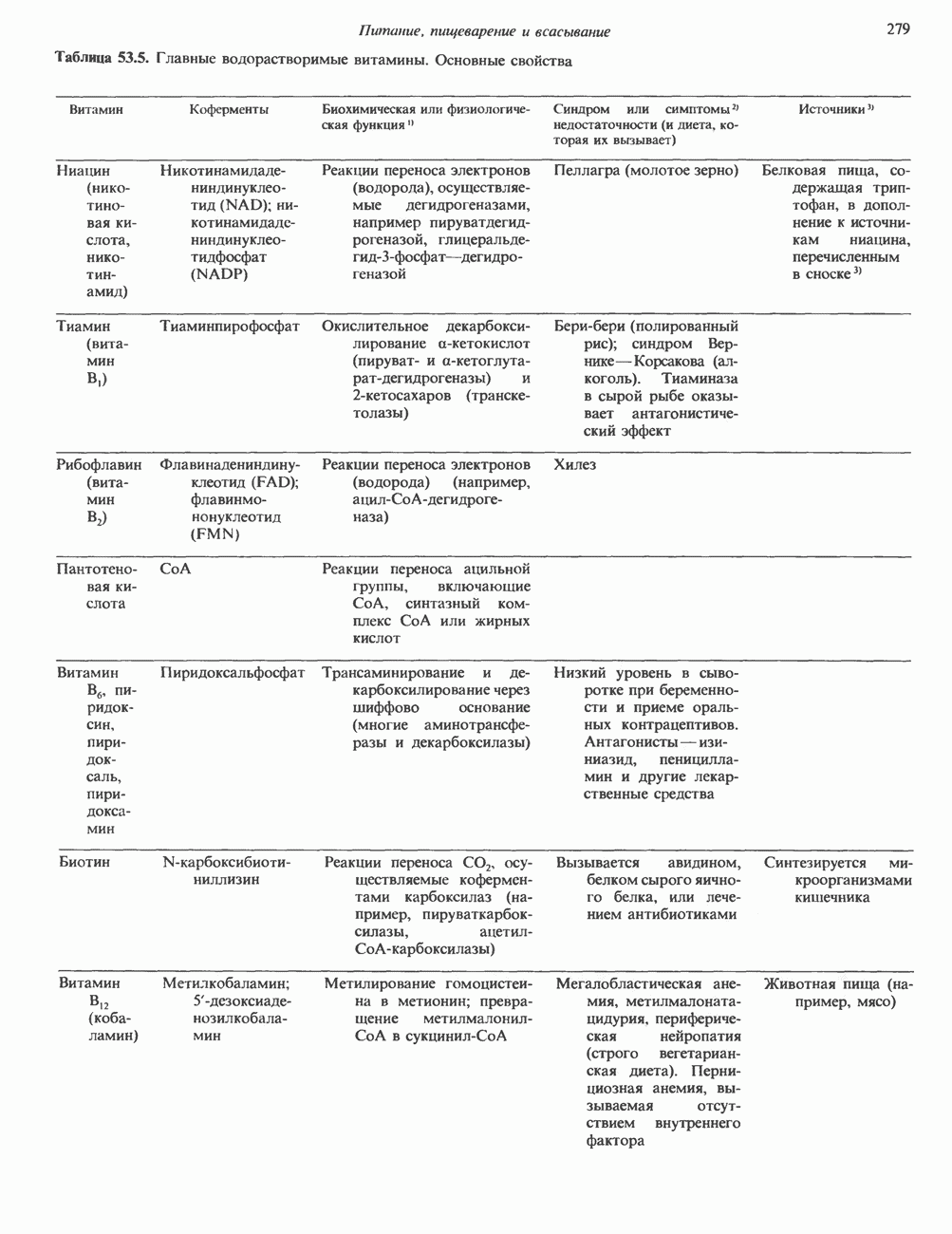
Пред.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 145 146 147 148 149 150 151 152 153 154 155 156 157 158 159 160 161 162 163 164 165 166 167 168 169 170 171 172 173 174 175 176 177 178 179 180 181 182 183 184 185 186 187 188 189 190 191 192 193 194 195 196 197 198 199 200 201 202 203 204 205 206 207 208 209 210 211 212 213 214 215 216 217 218 219 220 221 222 223 224 225 226 227 228 229 230 231 232 233 234 235 236 237 238 239 240 241 242 243 244 245 246 247 248 249 250 251 252 253 254 255 256 257 258 259 260 261 262 263 264 265 266 267 268 269 270 271 272 273 274 275 276 277 278 279 280 281 282 283 284 285 286 287 288 289 290 291 292 293 294 295 296 297 298 299 300 301 302 303 304 305 306 307 308 309 310 311 312 313 314 315 316 317 318 319 320 321 322 323 324 325 326 327 328 329 330 331 332 333 334 335 336 337 338 339 340 341 342 343 344 345 346 347 348 349 350 351 352 353 354 355 356 357 358 359 360 361 362 363 364 365 366 367 368 369 370 371 372 373 374 375 376 377 378 379 380 381 След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
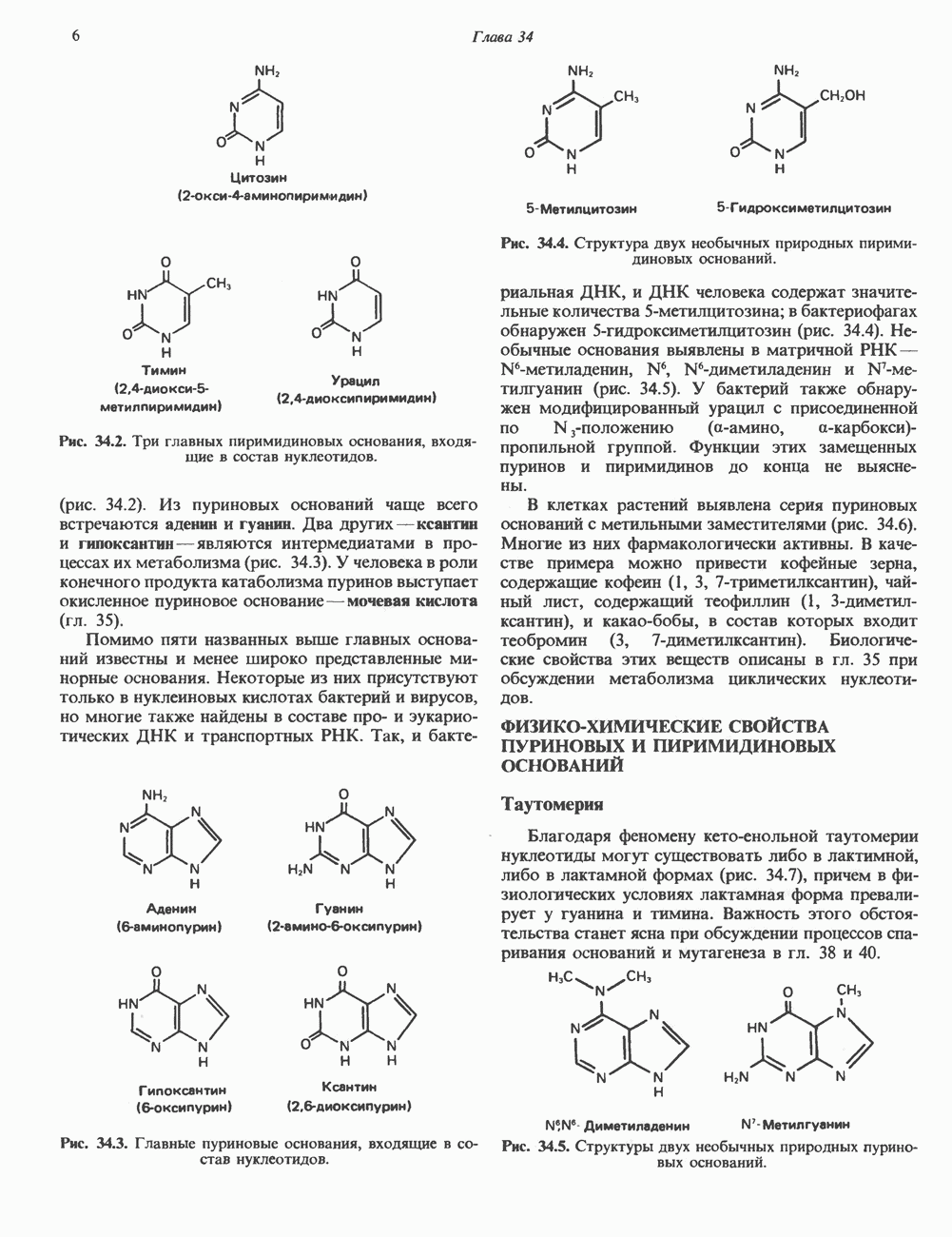
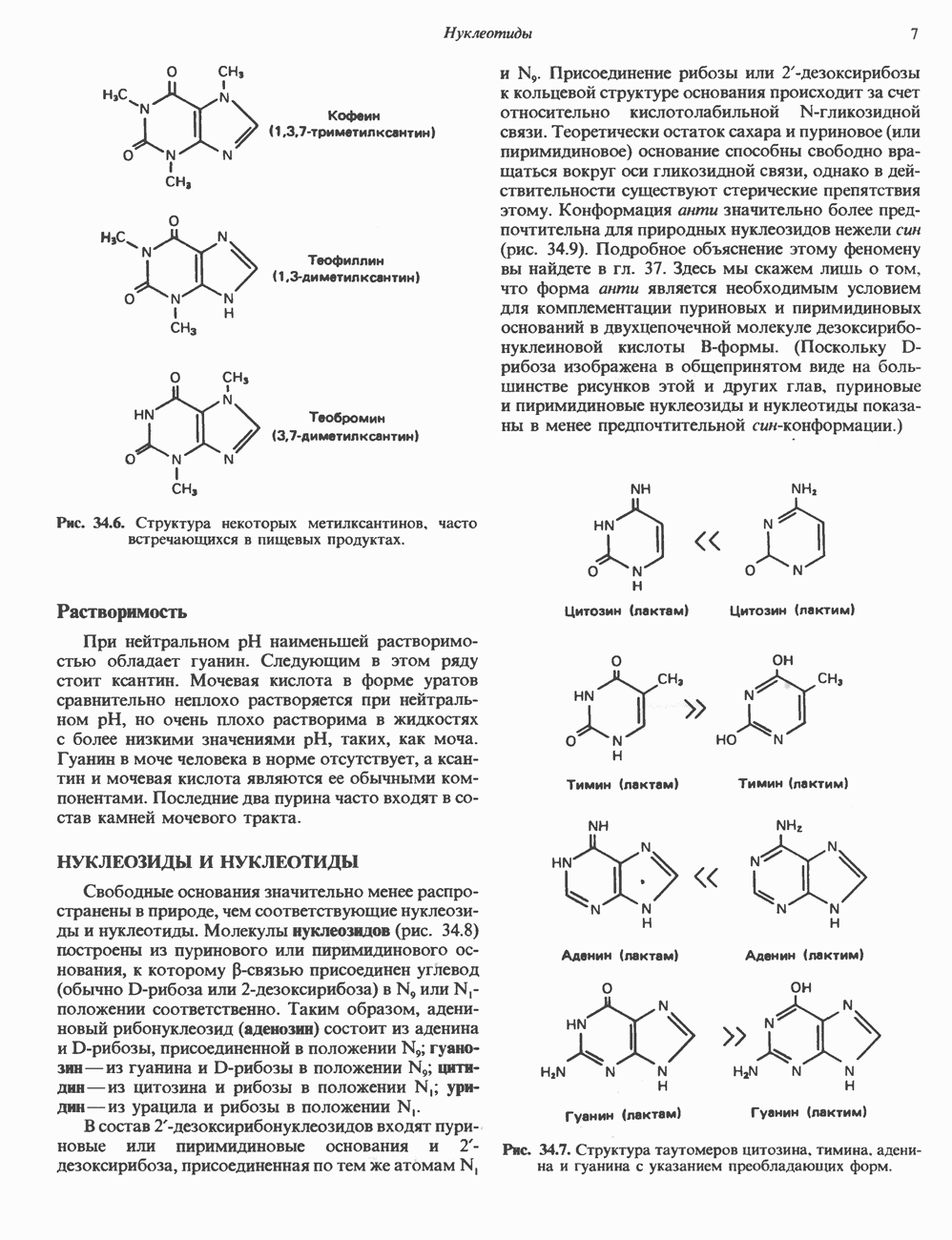
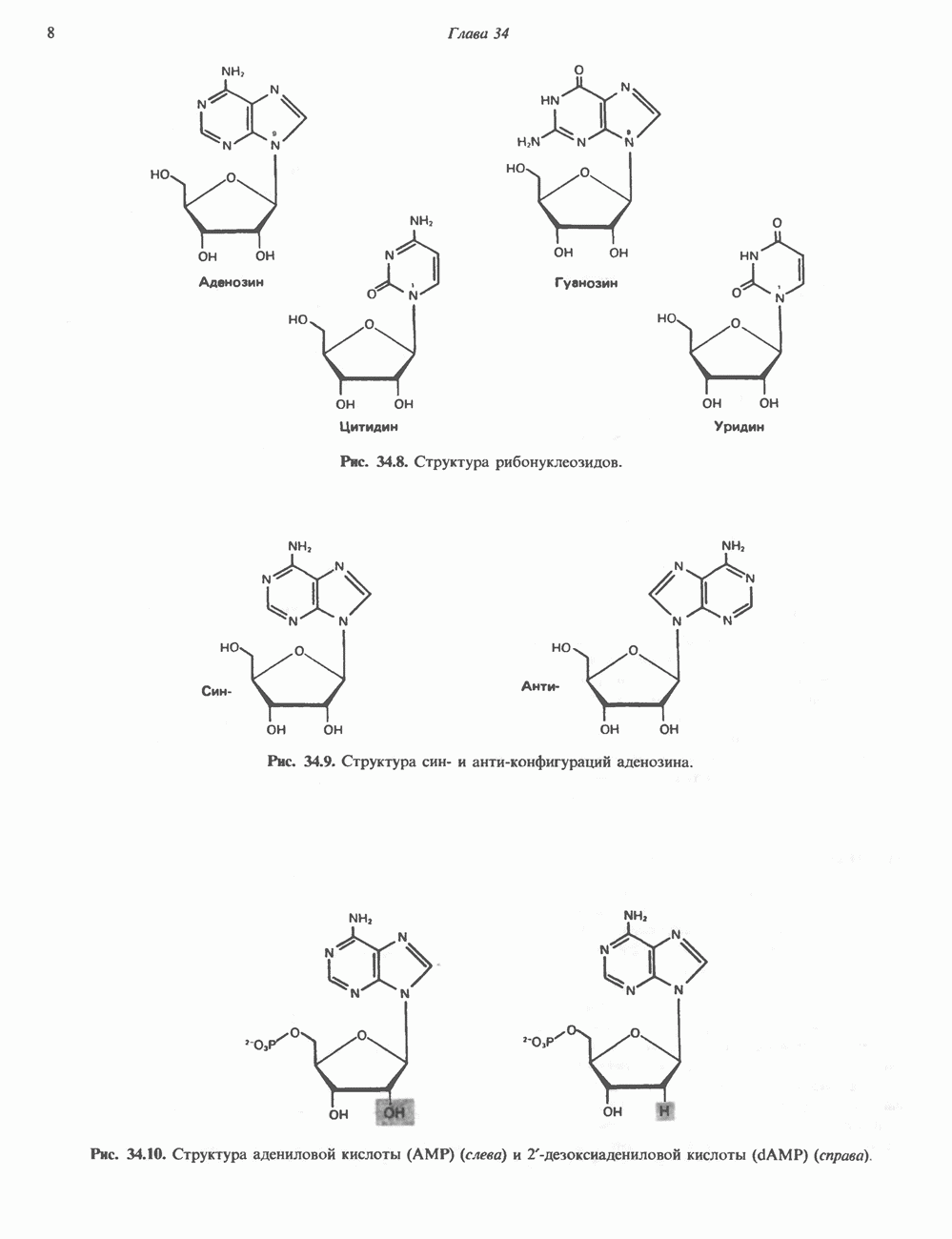
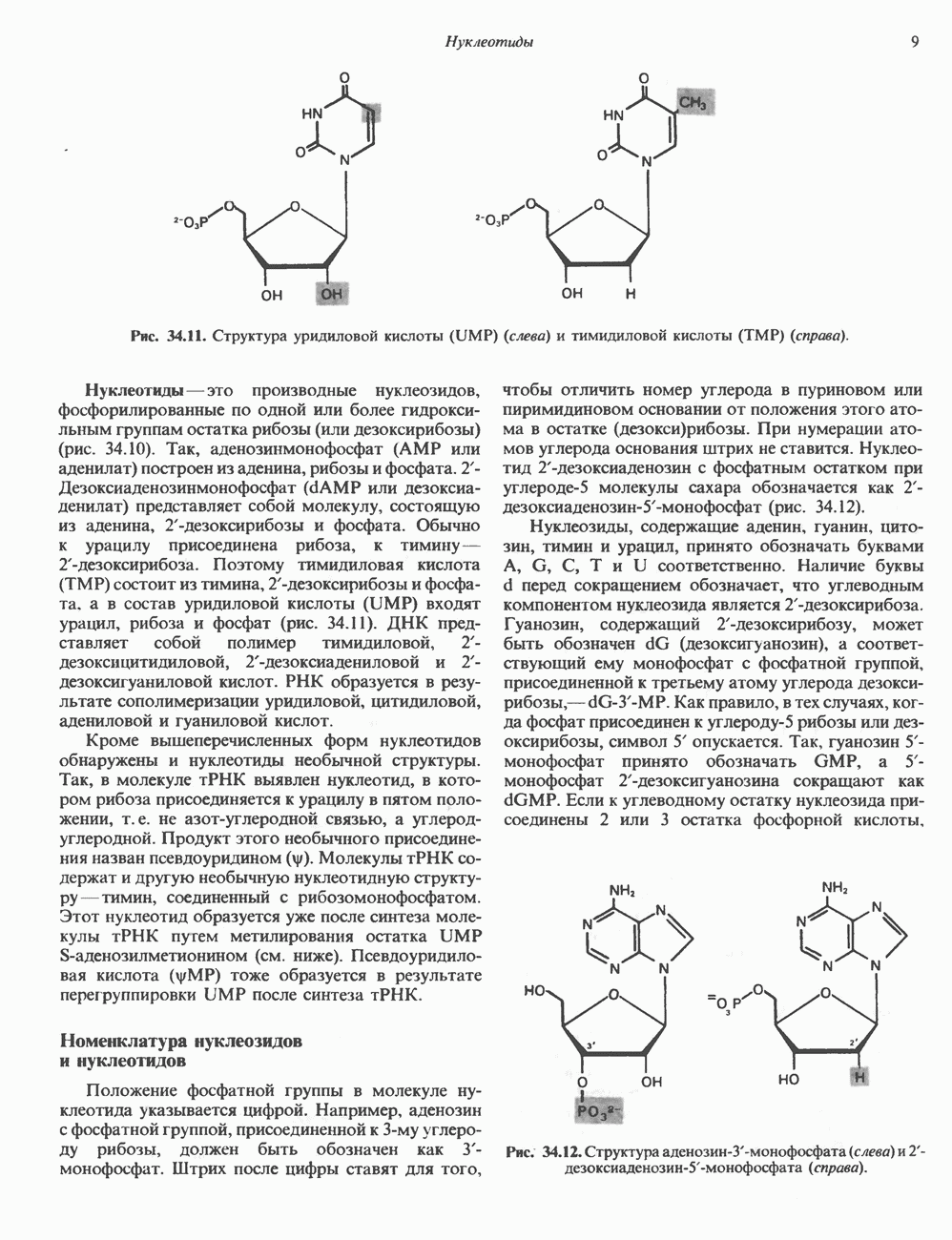
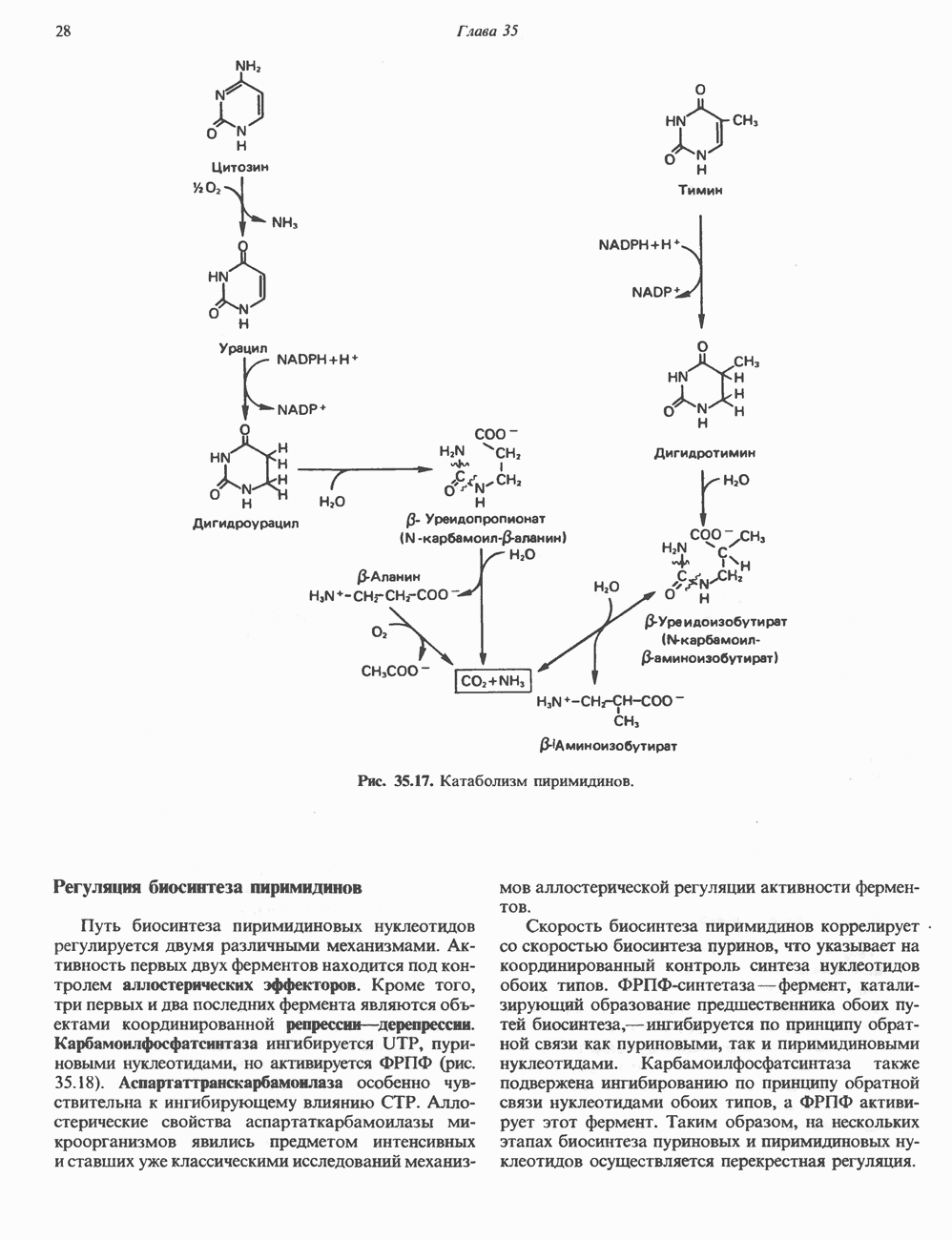
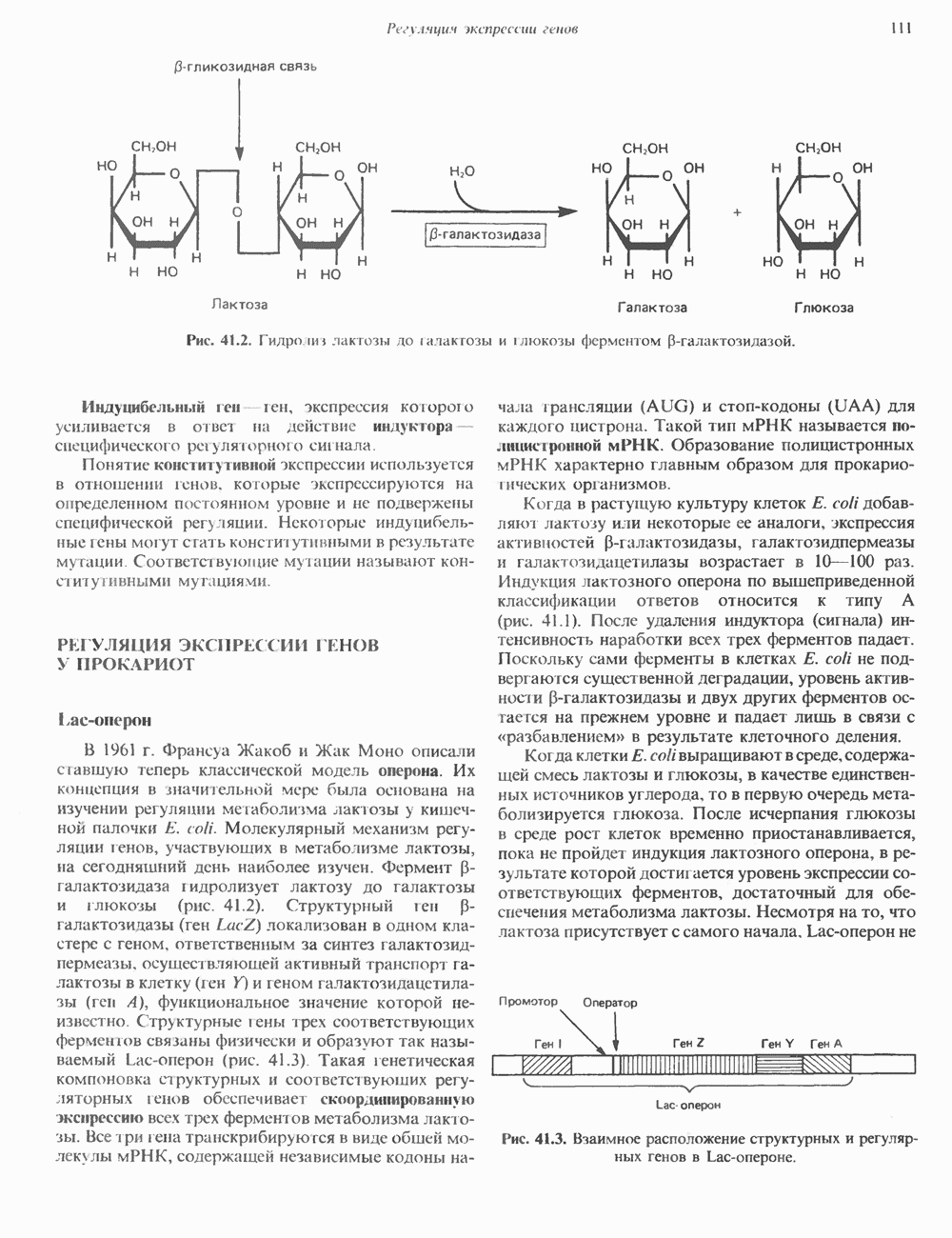
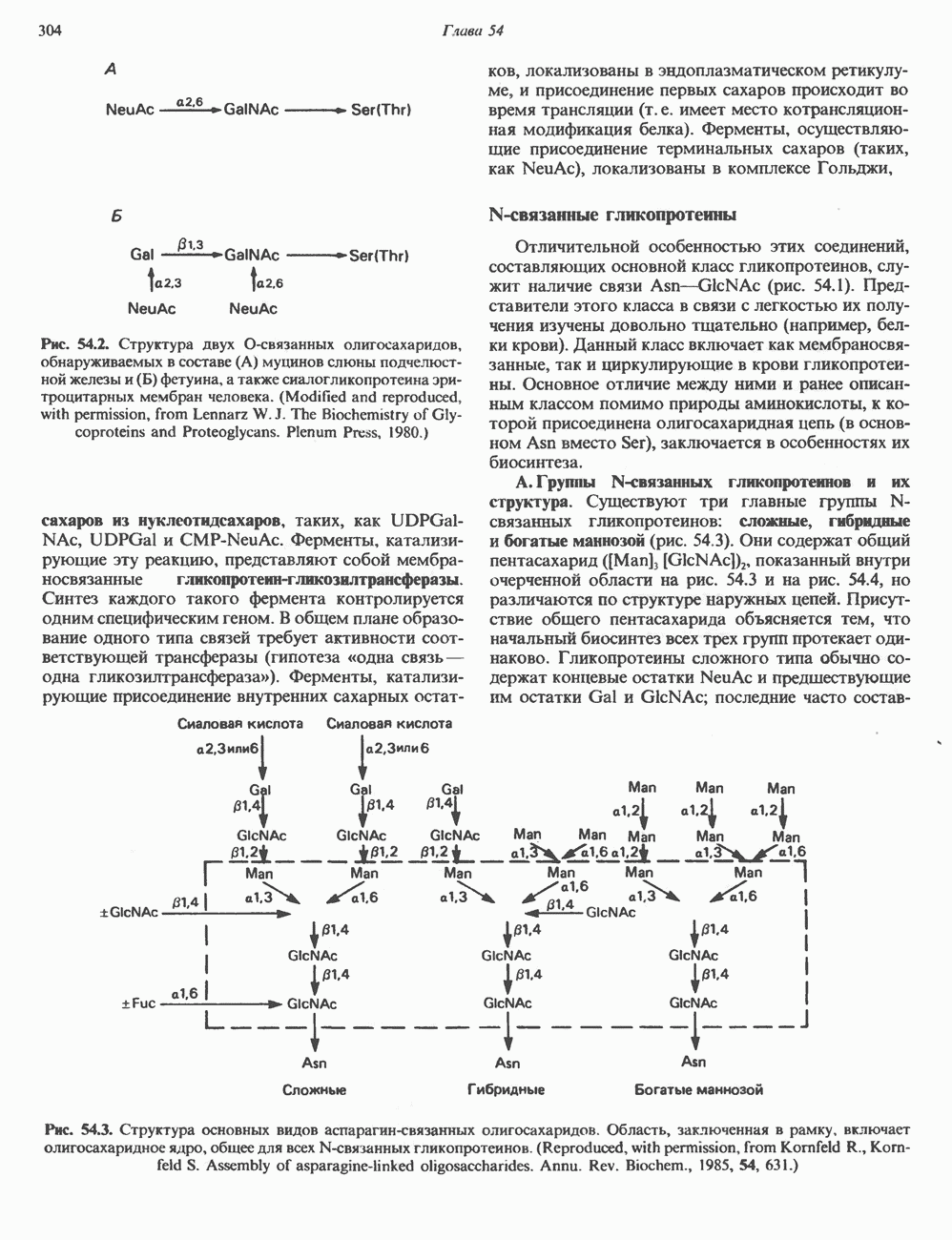
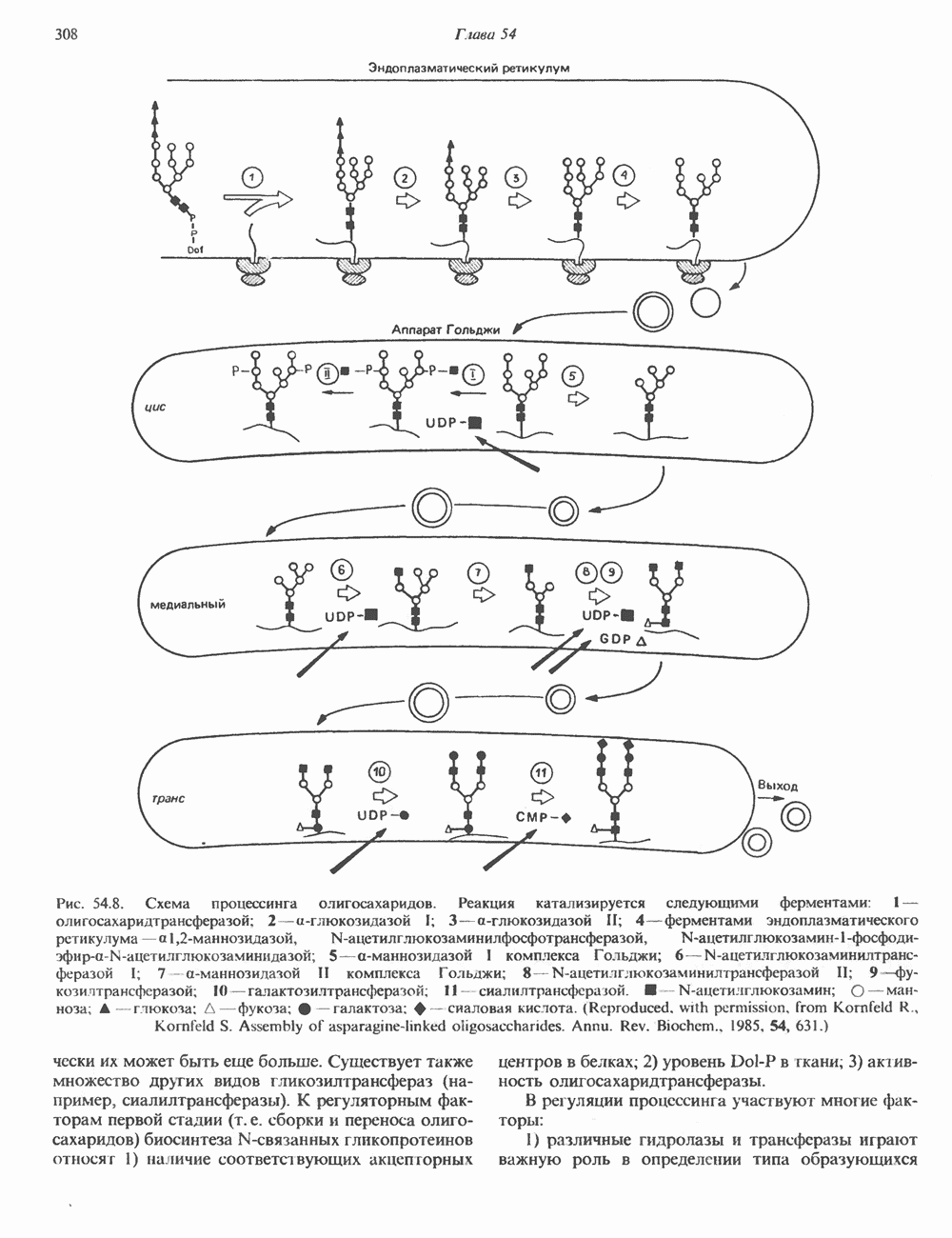
ПРОТЕОГЛИКАНЫ И ГЛИКОЗАМИНОГЛИКАНЫПротеогликаны и гликопротеины — это молекулы, состоящие из белков с ковалентно присоединенными к ним олигосахаридными или полисахаридными цепями. Различие между протеогликанами и гликопротеинами состоит в химической природе присоединенных к белку полисахаридов. В протеогликанах каждый полисахарид состоит из повторяющихся дисахаридных единиц, в которых всегда присутствуют Д-глюкозамин или Д-галактозамин. Каждый дисахаридный компонент протеогликановых полисахаридов (за исключением кератансульфата) содержит уроновую кислоту-L-глюкуроновую кислоту (GleUA) или ее 5-эпимер, Существуют 3 типа связей протеогликановых полисахаридов с их полипептидной цепью: 1) О-гликозидная связь между Пути образования полисахаридных цепей полностью идентичны путям, по которым происходит рост олигосахаридных цепей гликопротеинов. Процесс элонгации цепи протекает при участии нуклеотидсахаров, действующих в качестве доноров, Реакции регулируются в первую очередь субстратной специфичностью отдельных гликозилтрансфераз. Здесь вновь проявляется принцип «один фермент — одна связь». Специфичность реакций зависит от нуклеотидсахарного донора, акцепторного олигосахарида и от аномерной конфигурации и положения связи. Ферментативные системы, участвующие в элонгации цепи, обладают способностью очень точно воспроизводить сложные полисахариды. Завершение роста полисахаридной цепи является результатом следующих феноменов: 1) кэпирующего эффекта сиалилирования специфическими сиалил-трансферазами; 2) сульфирования, в особенности в После образования полисахаридной цепи происходят многочисленные химические модификации, такие, как включение сульфатных групп в Важным аспектом метаболизма протеогликанов является их деградация. Наследственные дефекты деградации полисахаридных цепей протеогликанов лежат в основе группы заболеваний, известных как му-копо лисахарцдозы и муколипидозы, которые обсуждаются ниже и в гл. 14. Эти нарушения катаболизма способствовали изучению специфических ферментов деградации и их субстратов. Существует ряд экзогликозидаз, осуществляющих ступенчатое отщепление сульфатных компонентов и гликозильных групп. Кроме того, имеются эндогликозидазы, которые присутствуют в нормальных условиях и обладают различной специфичностью. Например, широко распространенный фермент гиалуронидаза расщепляет Известно 7 типов полисахаридов (гликозамино-гликанов)
Рис. 54.9. Обобщенная структура протеогликанов и гликозаминогликанов соединяющихся с белками в составе протеогликанов. Шесть из них обладают родственной структурой и содержат остатки уроновой кислоты и гексозамина, которые перемежаются в повторяющихся дисахаридных компонентах; исключение составляет кератансульфат, лишенный уроновой кислоты. Все они, кроме гиалуроновой кислоты, содержат сульфированные сахара и ковалентно присоединены к белкам. Указанные 7 типов полисахаридов могут различаться по составу входящих в них мономеров, гликозидным связям, а также по количеству и локализации сульфатных заместителей. Все гликозаминогликаны являются полианионами благодаря присутствию в их структурах кислых сульфатных групп или карбоксильных групп уроновых кислот. С этой особенностью гликозаминогликанов связаны многие их функциональные свойства. Структуры 7 гликозаминогликанов, входящих в состав про еогликановых молекул, представлены на рис. 54.9. Гиалуроновая кислотаГиалуроновая кислота представляет собой неразветвленную цепь из повторяющихся дисахаридных компонентов, содержащих GlcUA и GlcNAc. Твердые доказательства связи гиалуроновой кислоты с молекулой белка (которые имеются для других полисахаридов соединительной ткани) отсутствуют, но, вероятно, эта кислота подобно другим гликозаминогликанам, синтезируется в составе протеогликанов. Гиалуроновая кислота присутствует у бактерий и широко распространена в различных тканях животных, включая синовиальную жидкость, стекловидное тело глаза и рыхлую соединительную ткань. ХондроитинсульфатыХондроитинсульфаты—это протеогликаны, являющиеся важнейшим компонентом хряща. Полисахарид связывается с белком О-гликозидной связью Xyl-Ser. Данные о структуре хондроитинсульфатов суммированы на рис. 54.9. Повторяющийся дисахаридный компонент очень сходен с таковым в гиалуроновой кислоте, за тем исключением, что гексозамин представлен GalNAc, а не GlcNAc. Однако и в хондроитинсульфатах, и в гиалуроновой кислоте уроновая кислота представлена GlcUA, а положения связей и аномерные конфигурации у них одни и те же. GalN Ас хондроитинсульфатов содержит сульфатный
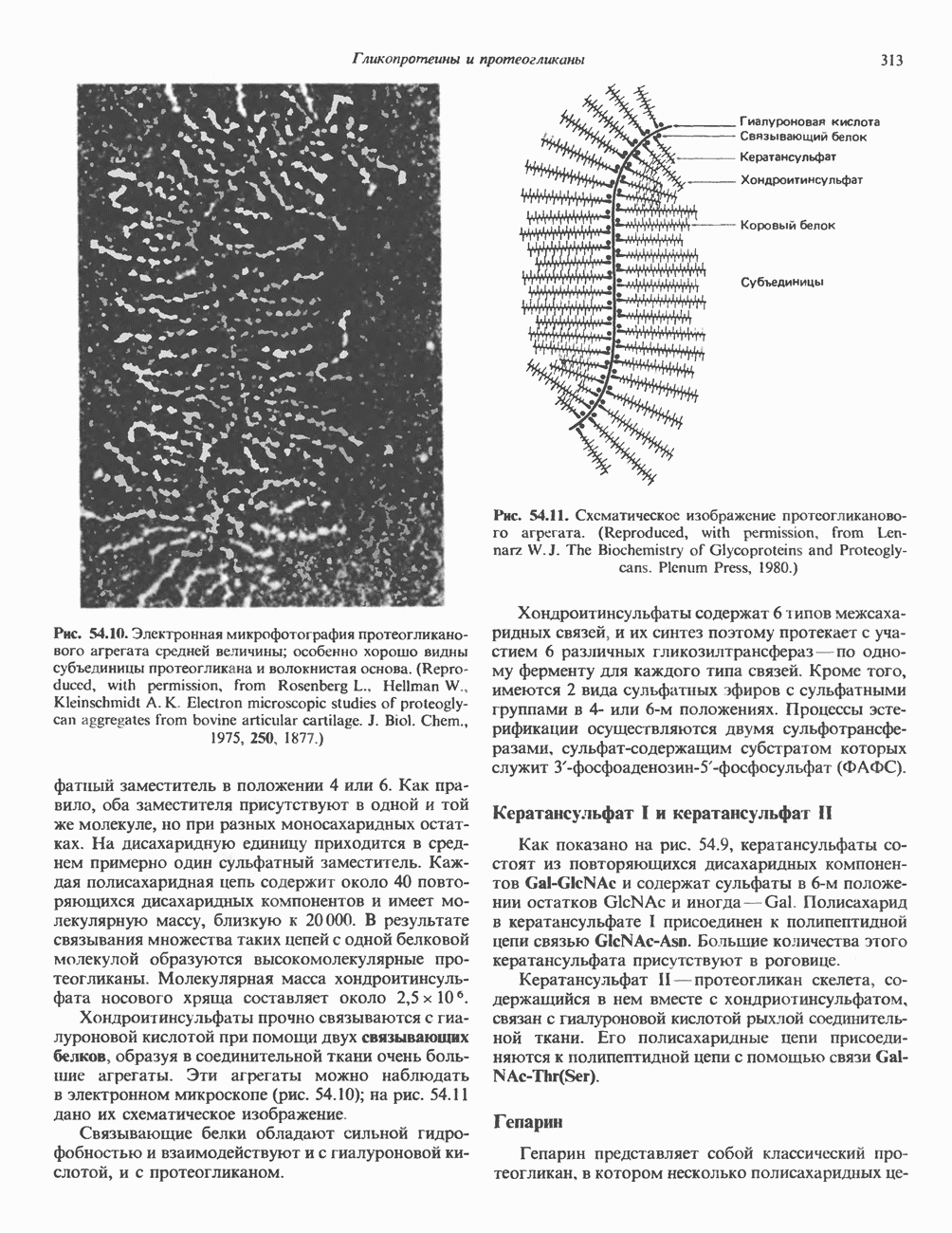
Рис. 54.10. Электронная микрофотография протеогликанового агрегата средней величины; особенно хорошо видны субъединицы протеогликана и волокнистая основа. (Reproduced, with permission, from Rosenberg L., Heilman W., Kleinschmidt A. K. Electron microscopic studies of proteoglycan aggregates from bovine articular cartilage. J. Biol. Chem., 1975, 250, 1877.) заместитель в положении 4 или 6. Как правило, оба заместителя присутствуют в одной и той же молекуле, но при разных моносахаридных остатках. На дисахаридную единицу приходится в среднем примерно один сульфатный заместитель. Каждая полисахаридная цепь содержит около 40 повторяющихся дисахаридных компонентов и имеет молекулярную массу, близкую к 20000. В результате связывания множества таких цепей с одной белковой молекулой образуются высокомолекулярные протеогликаны. Молекулярная масса хондроитинсульфата носового хряща составляет около Хондроитинсу льфаты прочно связываются с гиалуроновой кислотой при помощи двух связывающих белков, образуя в соединительной ткани очень большие агрегаты. Эти агрегаты можно наблюдать в электронном микроскопе (рис. 54.10); на рис. 54.11 дано их схематическое изображение. Связывающие белки обладают сильной гидрофобностью и взаимодействуют и с гиалуроновой кислотой, и с протеогликаном.
Рис. 54.11. Схематическое изображение протсогликаново-го агрегата. (Reproduced, with permission, from Len-narz W.J. The Biochemistry of Glycoproteins and Proteoglycans. Plenum Press, 1980.) Хондроитинсульфаты содержат 6 типов межсахаридных связей, и их синтез поэтому протекает с участием 6 различных гликозилтрансфераз — по одному ферменту для каждого типа связей. Кроме того, имеются 2 вида сульфатных эфиров с сульфатными группами в 4- или 6-м положениях. Процессы эстерификации осущес вляются двумя сульфотрансферазами, сульфатсодержащим субстратом которых служит 3'-фосфоаденозин-5'-фосфосульфат (ФАФС). Кератансульфат I и кератансульфат IIКак показано на рис. 54.9, кератансульфаты состоят из повторяющихся дисахаридных компонентов Gal-GlcNAc и содержат сульфаты в 6 м положении остатков GlcNAc и иногда - Gal. Полисахарид в кератансульфате I присоединен к полипептидной цепи связью GIcNAc-Asn. Большие количества этого кератансульфата присутствуют в роговице. Кератансульфат II — протеогликан скелета, содержащийся в нем вместе с хондриотинсульфатом, связан с гиалуроновои кислотой рыхлой соединительной ткани. Его полисахаридные цепи присоединяются к полипептидной цепи с помощью связи Gal-NAc-Thr(Ser). ГепаринГепарин представляет собой классический протеогликан в котором несколько полисахаридных
Рис. 54.12. Структура гепарина. Отрезок полимера иллюстрирует типичные структурные особенности гепарина, однако последовательность различным образом замещенных повторяющихся дисахаридных компонентов выбрана произвольно. Кроме того, могут присутствовать несульфированные или 3-О-сульфированные глюкозаминовые остатки. (Modified redrawn, and reproduced with permission, from Lindahl U. et al. Structure and biosynthesis of heparin-like polysaccharides. Fed. Proc., 1977, 36, 19.) цепей связаны с общим белковым ядром. Он обнаруживается в гранулах тучных клеток и, таким образом, локализован внутриклеточно. Гепарин обладает и другими структурными и функциональными особенностями, и некоторые из них имеют значение для медицины. Характерные особенности структуры гепарина представлены на рис. 54.12. Повторяющийся дисахаридный компонент содержит глюкозамин (GlcN) и уроновую кислоту. Большинство аминогрупп остатков GlcN присутствует в N-сульфированной форме, но имеется и небольшое количество ацетилированных аминогрупп. GlcN содержит также С6-сульфатный эфир. Около 90% уроновой кислоты — это IdUA и лишь 10% приходится на GlcUA. Первоначально остатки уроновой кислоты представлены в виде GlcUA, но в дальнейшем, после образования полисахарида 5-эпимераза превращает почти 90% остатков GlcUA в остатки IdUA, последние часто подвергаются сульфированию во 2-м положении. Белковая молекула протеогликана гепарина уникальна в том отношении, что она состоит только из сериновых и глициновых остатков. Приблизительно две трети сериновых остатков соединены с полисахаридными цепями; их молекулярная масса составляет обычно 5000—15000, но иногда достигает 100000. Полисахаридные цепи гепариновых протеогликанов после полимеризации претерпевают ряд модификаций: 1) первичный продукт не сульфируется, но полностью N-ацетилируется с образованием полимера (GlcUA-GlcNAc)n; 2) около 50% остатков GlcNAc подвергаются N-деацетилированию; 3) свободные аминогруппы GlcN сульфируются; затем происходит дальнейшее деацетилирование примерно половины остальных групп ГепарансульфатГепарансульфат присутствует на клеточных поверхностях, представляя собой внеклеточный протеогликан. Полипептидный остов гепарансульфатного протеогликана содержит дополнительный аминокислотный компонент, отличный от такового в гепарине. В процессе модификации его полисахаридных цепей деацетилирование остатков ДерматансульфатДерматансульфат представляет собой протеогликан, широко распространенный в тканях животных. По структуре он сходен и с хондроитинсульфатом, и с гепарансульфатом. Его отличие от хондроитинсульфата состоит в том, что вместо GlcUA, соединенной с
|
 |
Оглавление
|


























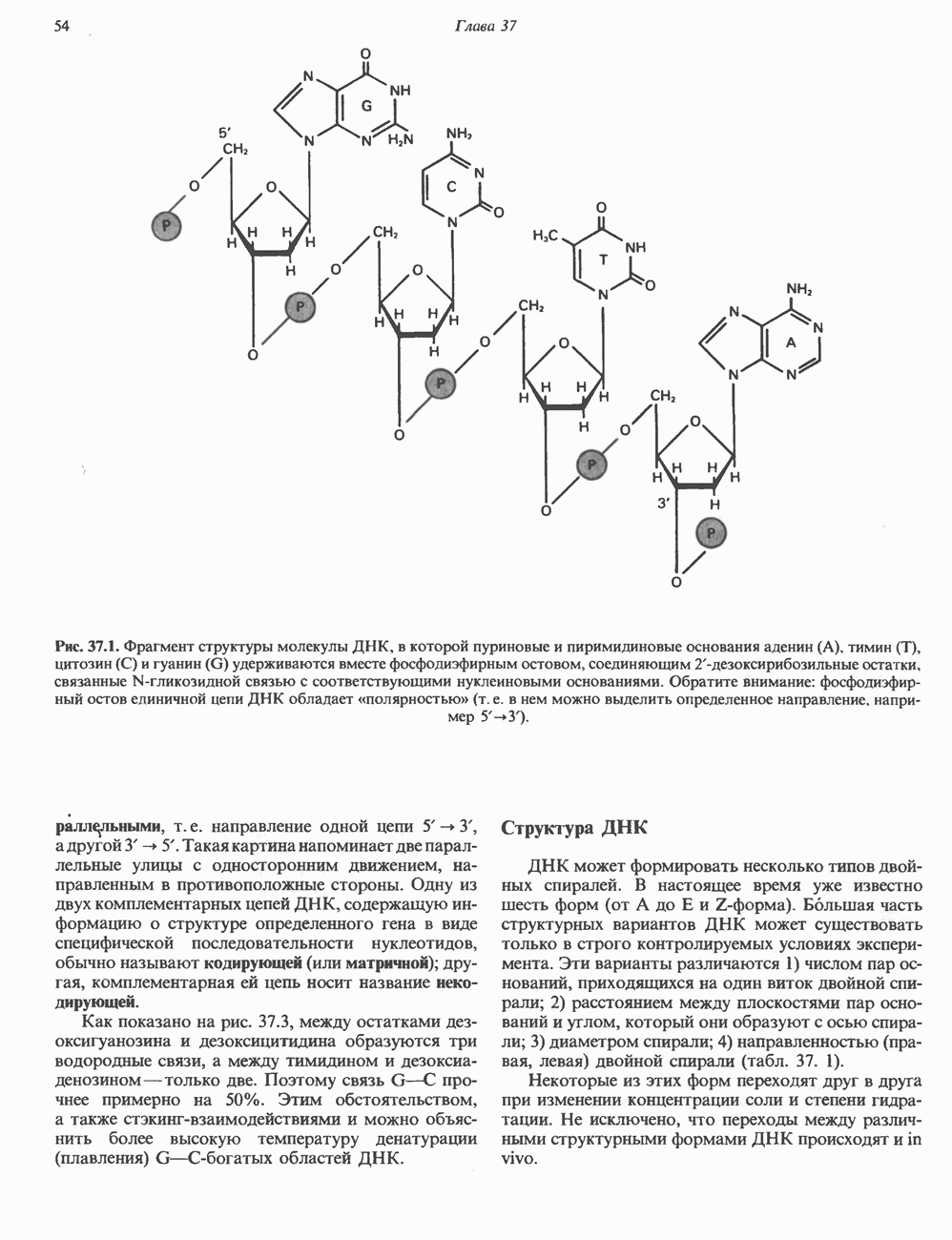
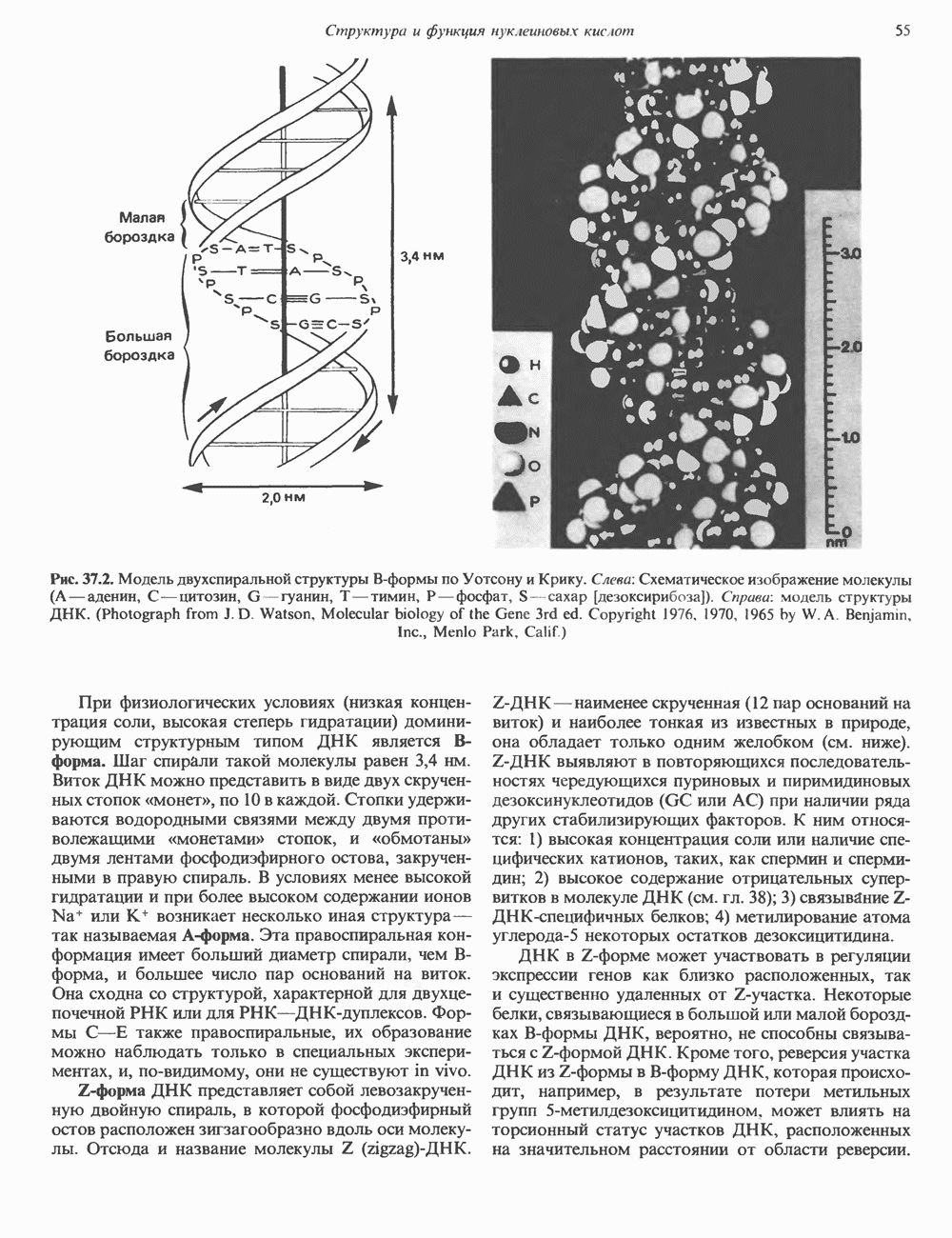
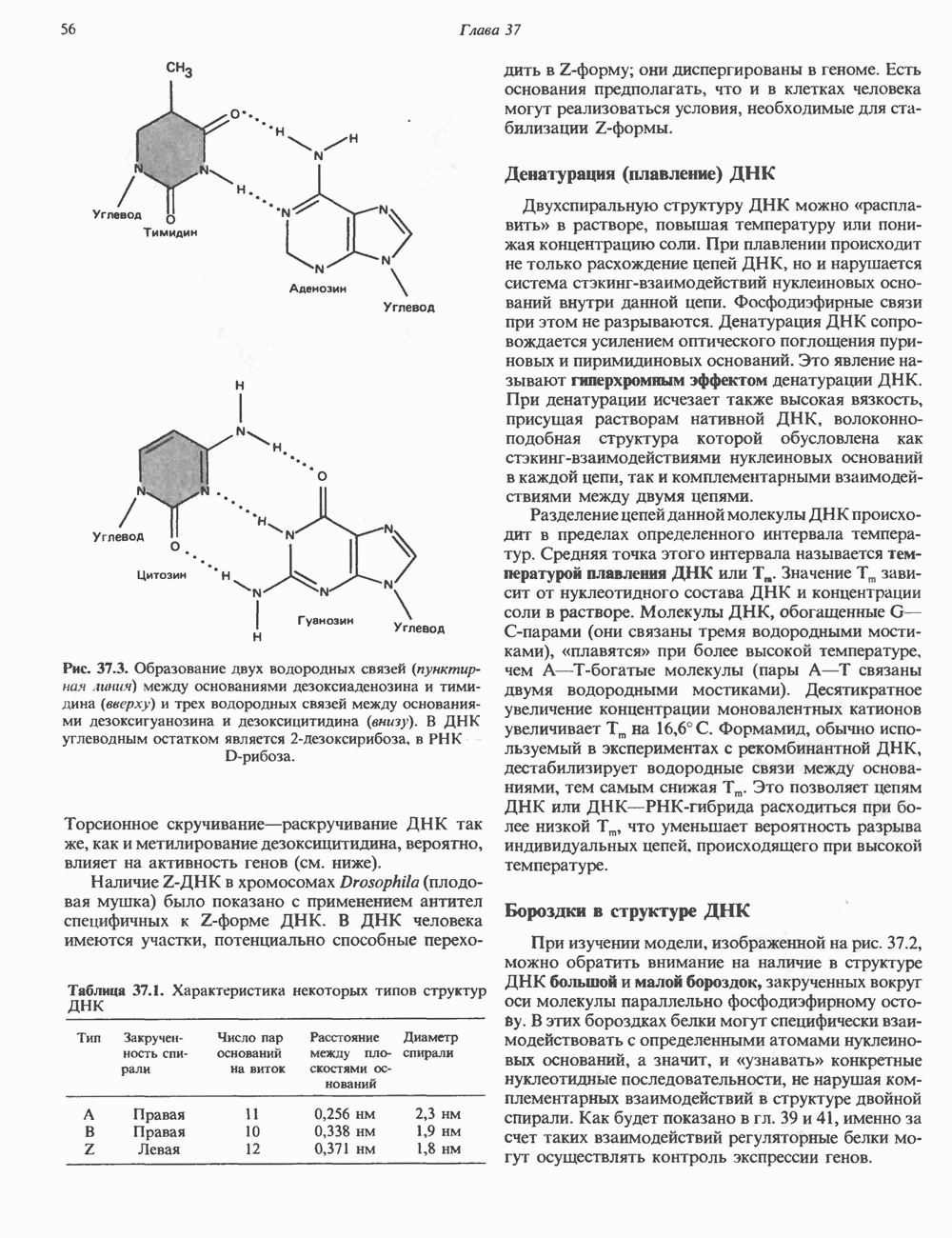

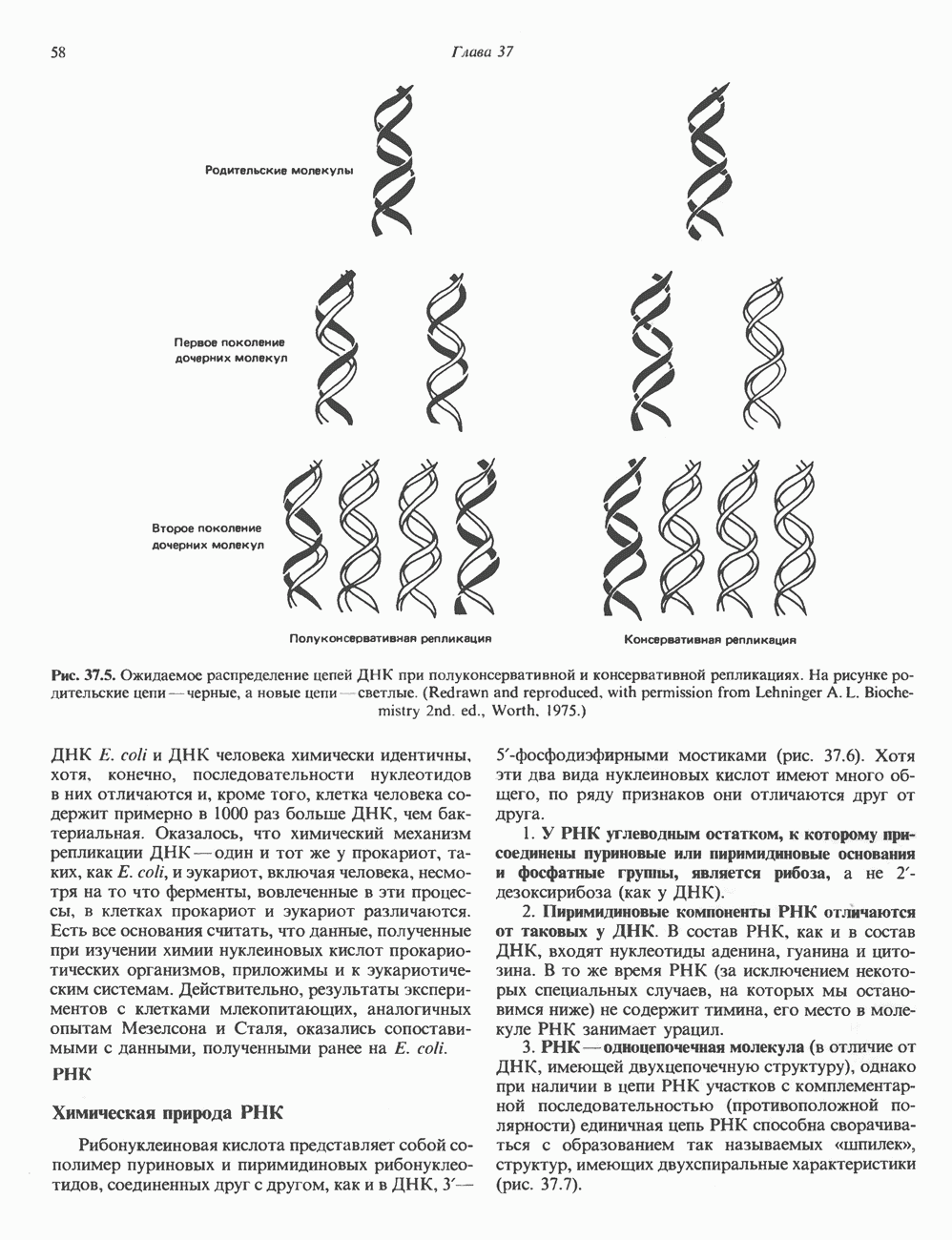








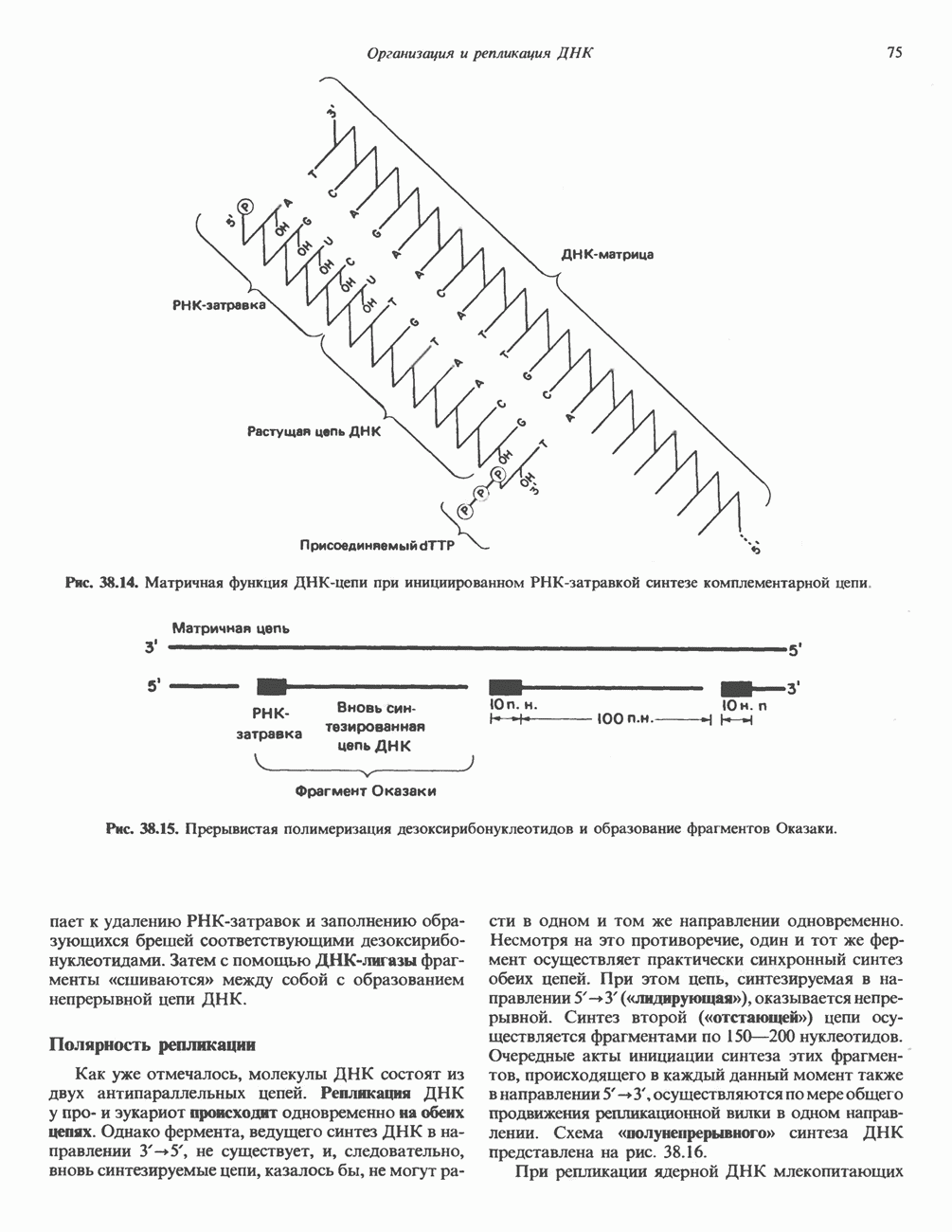
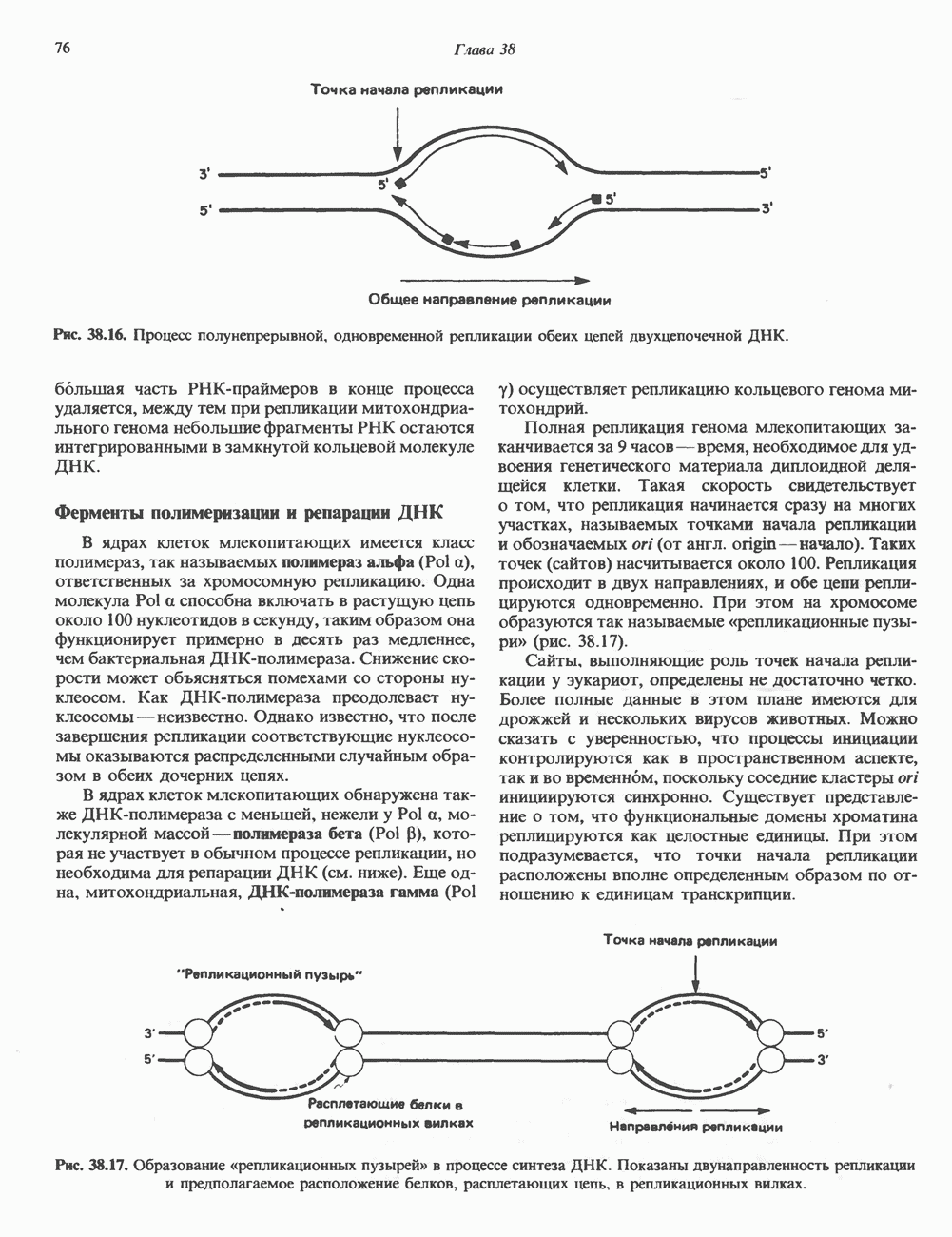
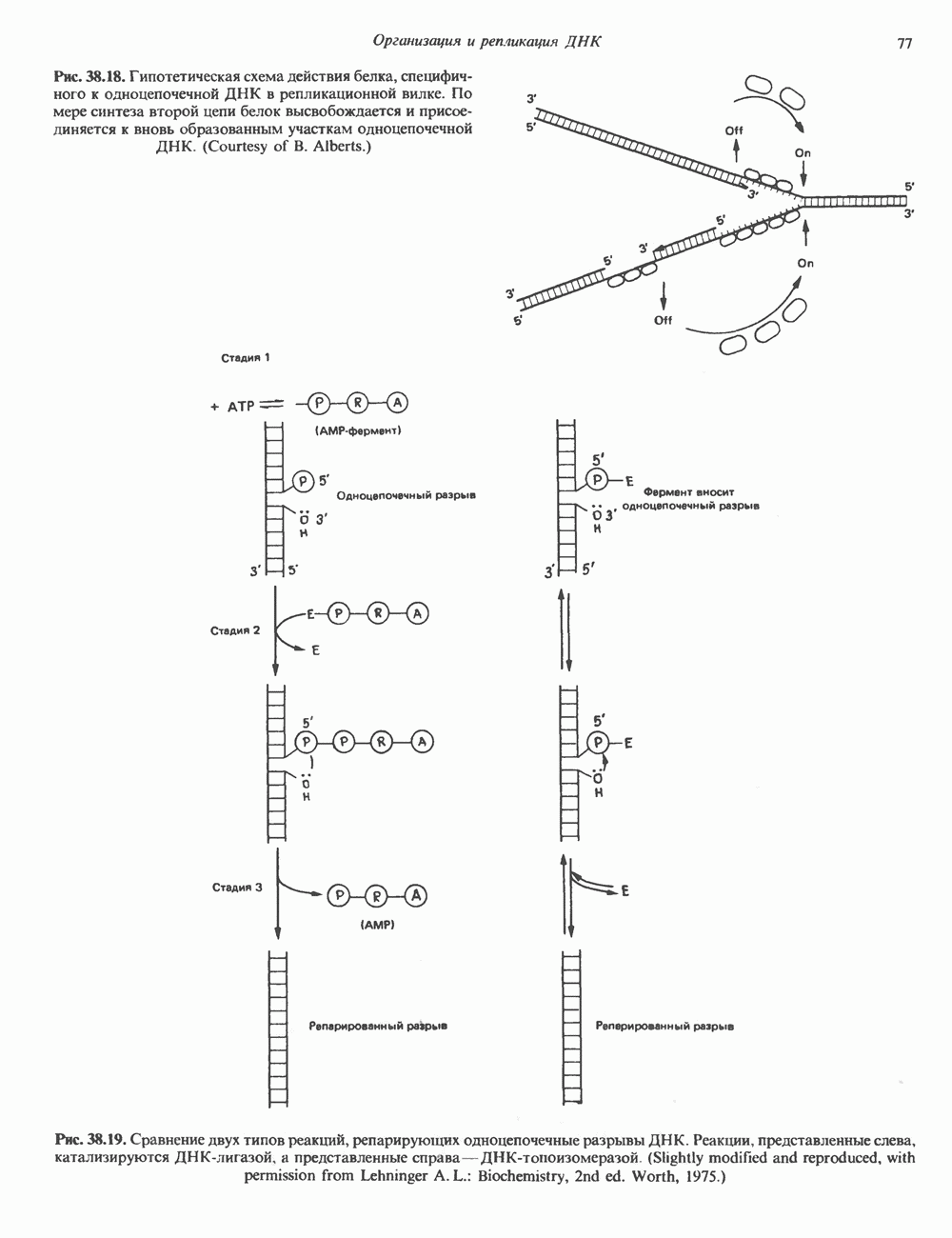






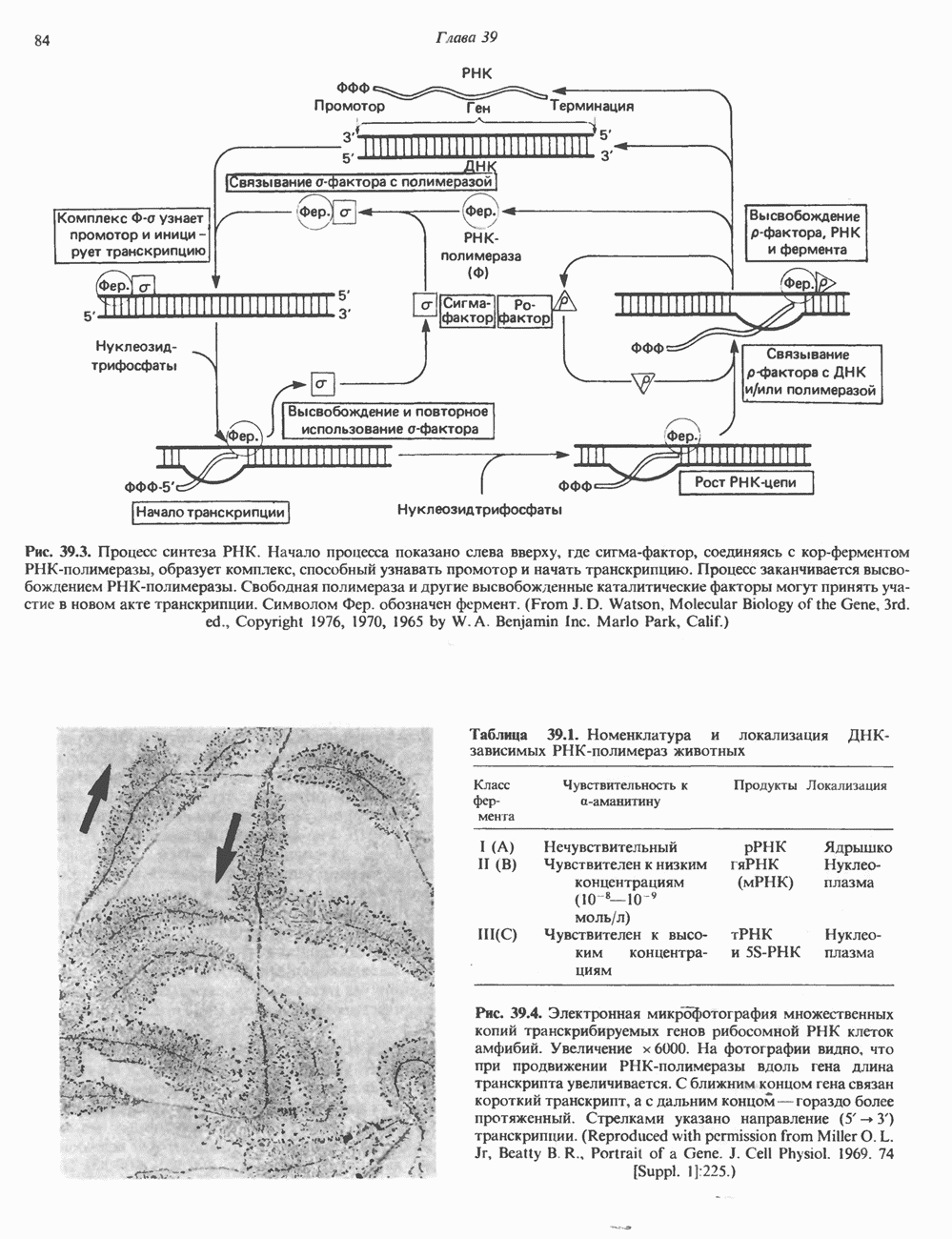


























































































































































































































 -идуроновую кислоту (IdUA). За исключением гиалуроновой
-идуроновую кислоту (IdUA). За исключением гиалуроновой  -сульфата (в гепарине или гепаринсульфате).
-сульфата (в гепарине или гепаринсульфате).
 характерная только для протеогликанов; 2) О-гликозидная связь между
характерная только для протеогликанов; 2) О-гликозидная связь между  присутствующая в кера-тансульфате II;
присутствующая в кера-тансульфате II;  связь между GlcNAc и амидным азотом
связь между GlcNAc и амидным азотом 
 присоединяет Ху1 нуклеотидного сахара к
присоединяет Ху1 нуклеотидного сахара к  с возникновением Ху1-Ser-O-гликозидной связи. Образование О-гликозидной связи между
с возникновением Ху1-Ser-O-гликозидной связи. Образование О-гликозидной связи между  вероятно, осуществляется аналогичной
вероятно, осуществляется аналогичной  -трансферазой.
-трансферазой.  -гликозидная связь между GlcNAc и амидным азотом
-гликозидная связь между GlcNAc и амидным азотом  почти наверняка образуется при участии липид-связанного полисахарида, долихол-Р— Р-полисахарида, который, как отмечалось выше, ответствен за транспорт предобразованного олиго- или полисахарида в процессе образования гликопротеинов. Однако детали этой реакции в ходе синтеза протеогликанов пока не установлены.
почти наверняка образуется при участии липид-связанного полисахарида, долихол-Р— Р-полисахарида, который, как отмечалось выше, ответствен за транспорт предобразованного олиго- или полисахарида в процессе образования гликопротеинов. Однако детали этой реакции в ходе синтеза протеогликанов пока не установлены.
 положении сахаров; 3) удаления данного полисахарида с того места на мембране, где протекает
положении сахаров; 3) удаления данного полисахарида с того места на мембране, где протекает  -компоненты хондроитинсульфата и дерматансульфата и эпимеризация остатка
-компоненты хондроитинсульфата и дерматансульфата и эпимеризация остатка  в остаток
в остаток  в гепарине и гепарансульфате.
в гепарине и гепарансульфате.
 -ацетилгексозаминовые связи в гиалуроновой кислоте и хондроитинсульфатах.
-ацетилгексозаминовые связи в гиалуроновой кислоте и хондроитинсульфатах.

 -гтюкуроновая
-гтюкуроновая 
 .
.



 вновь образованные остатки
вновь образованные остатки  далее подвергаются О-сульфированию в положениях
далее подвергаются О-сульфированию в положениях  модификация завершается О-сульфированием компонентов
модификация завершается О-сульфированием компонентов 
 происходит слабее, и поэтому он содержит меньше
происходит слабее, и поэтому он содержит меньше  -сульфатов. Поскольку
-сульфатов. Поскольку  -эпимераза (как отмечалось выше при описании модификаций гепарина) использует в качестве субстрата
-эпимераза (как отмечалось выше при описании модификаций гепарина) использует в качестве субстрата  -сульфатные заместители, гепарансульфат отличается от гепарина меньшим количеством остатков
-сульфатные заместители, гепарансульфат отличается от гепарина меньшим количеством остатков  и большим содержанием
и большим содержанием  . В соответствии с этим преобладающей уроновой кислотой в гепарансульфате является
. В соответствии с этим преобладающей уроновой кислотой в гепарансульфате является  а в гепарине
а в гепарине 
 он содержит IdUA, соединенную с GalN Ас а-1,3-связью. Образование IdUA, как и в случае гепарина и гепарансульфата, происходит путем 5-эпимеризации GlcUA. Как и при образовании гепарина, реакция эпимеризации тесно сопряжена с сульфированием гексозамина. Таким образом, дерматансульфат содержит 2 вида повторяющихся дисахаридных единиц:
он содержит IdUA, соединенную с GalN Ас а-1,3-связью. Образование IdUA, как и в случае гепарина и гепарансульфата, происходит путем 5-эпимеризации GlcUA. Как и при образовании гепарина, реакция эпимеризации тесно сопряжена с сульфированием гексозамина. Таким образом, дерматансульфат содержит 2 вида повторяющихся дисахаридных единиц: 