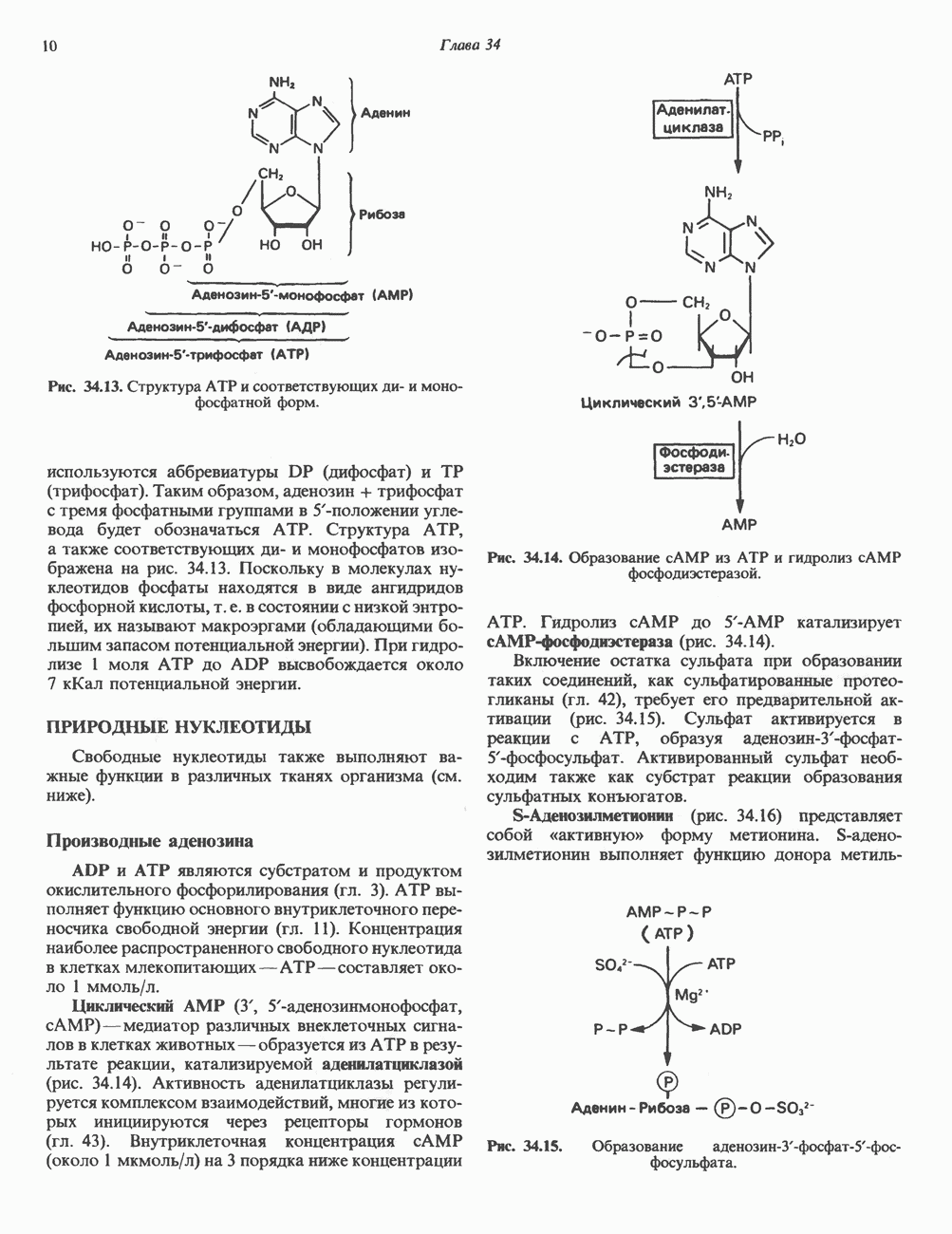
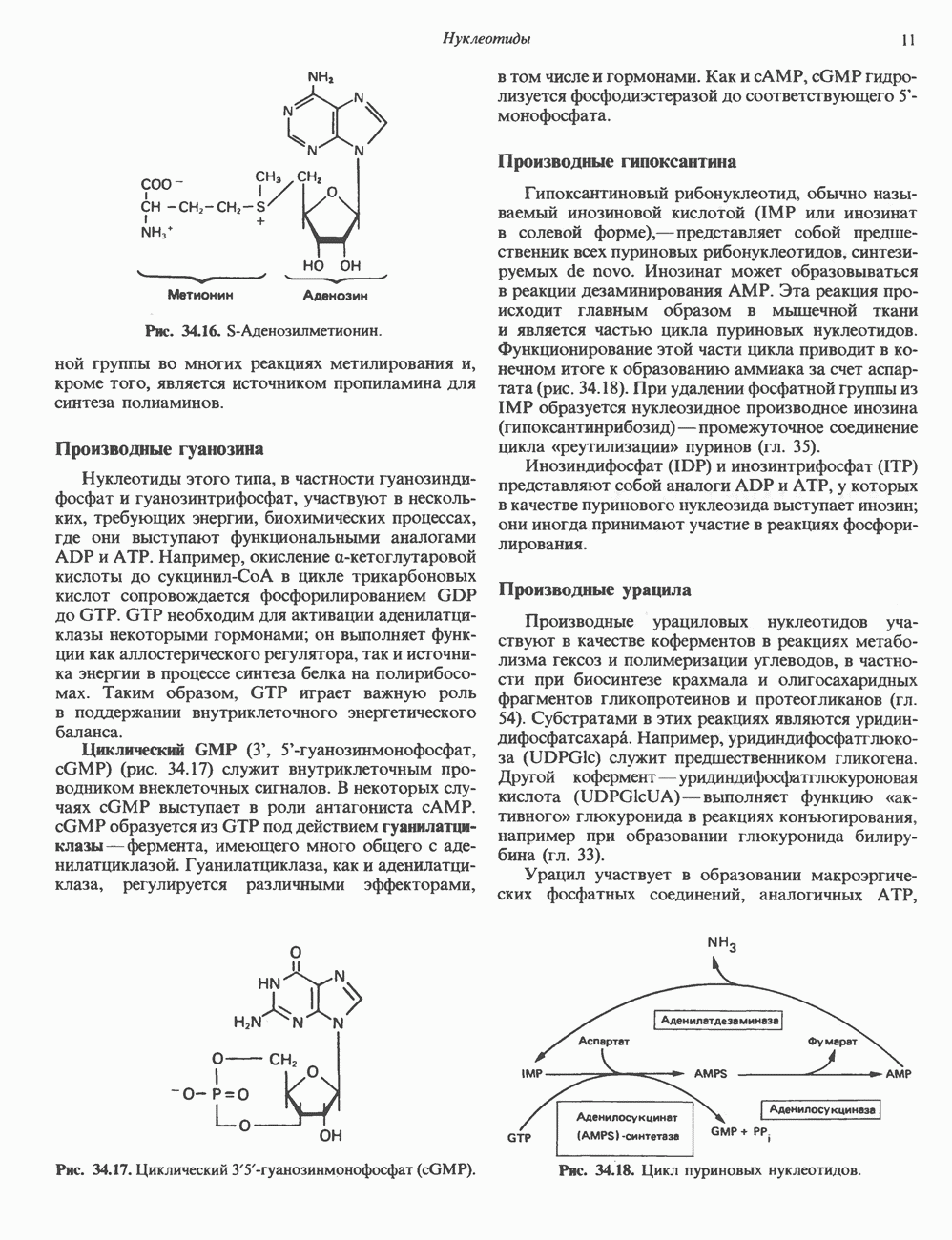
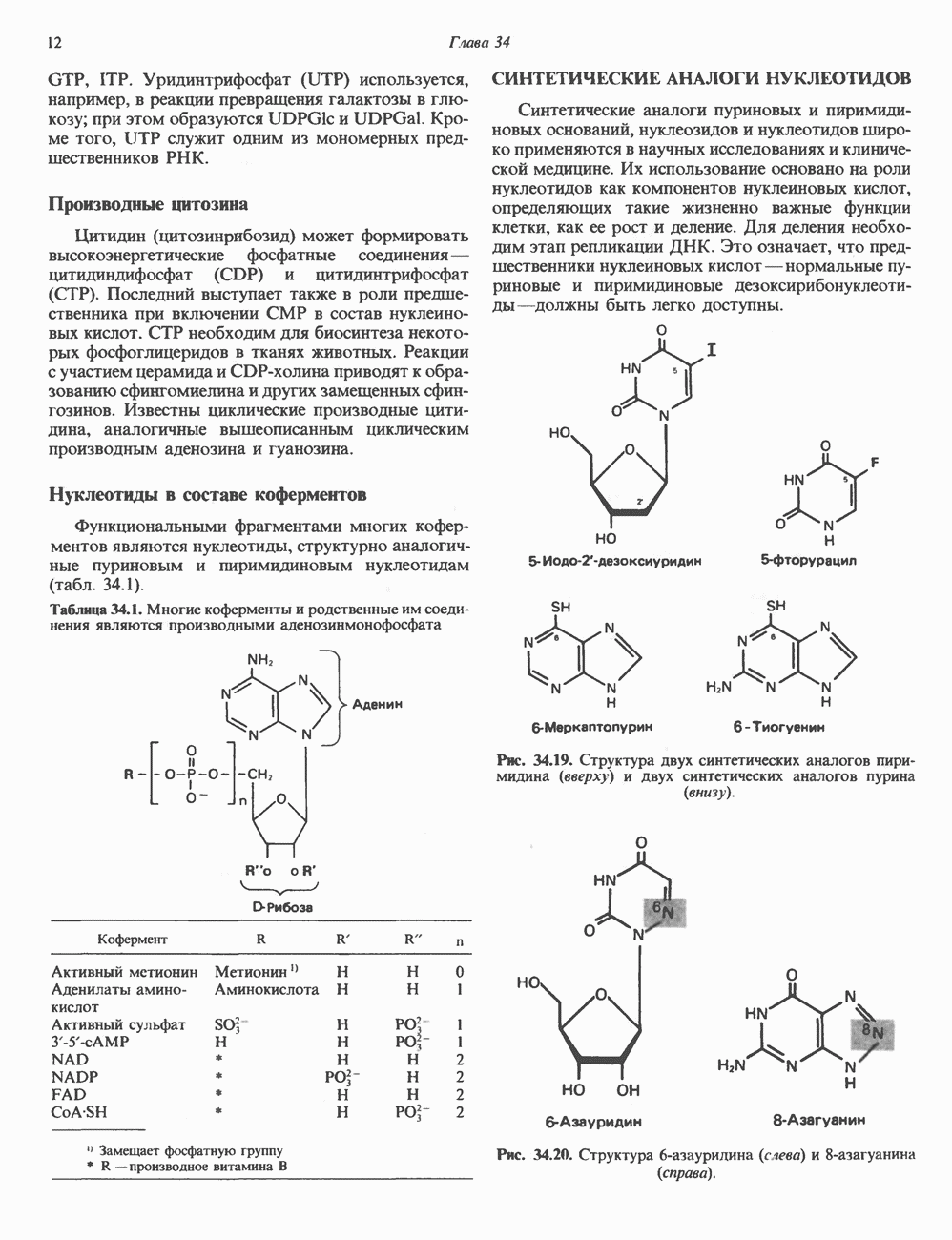
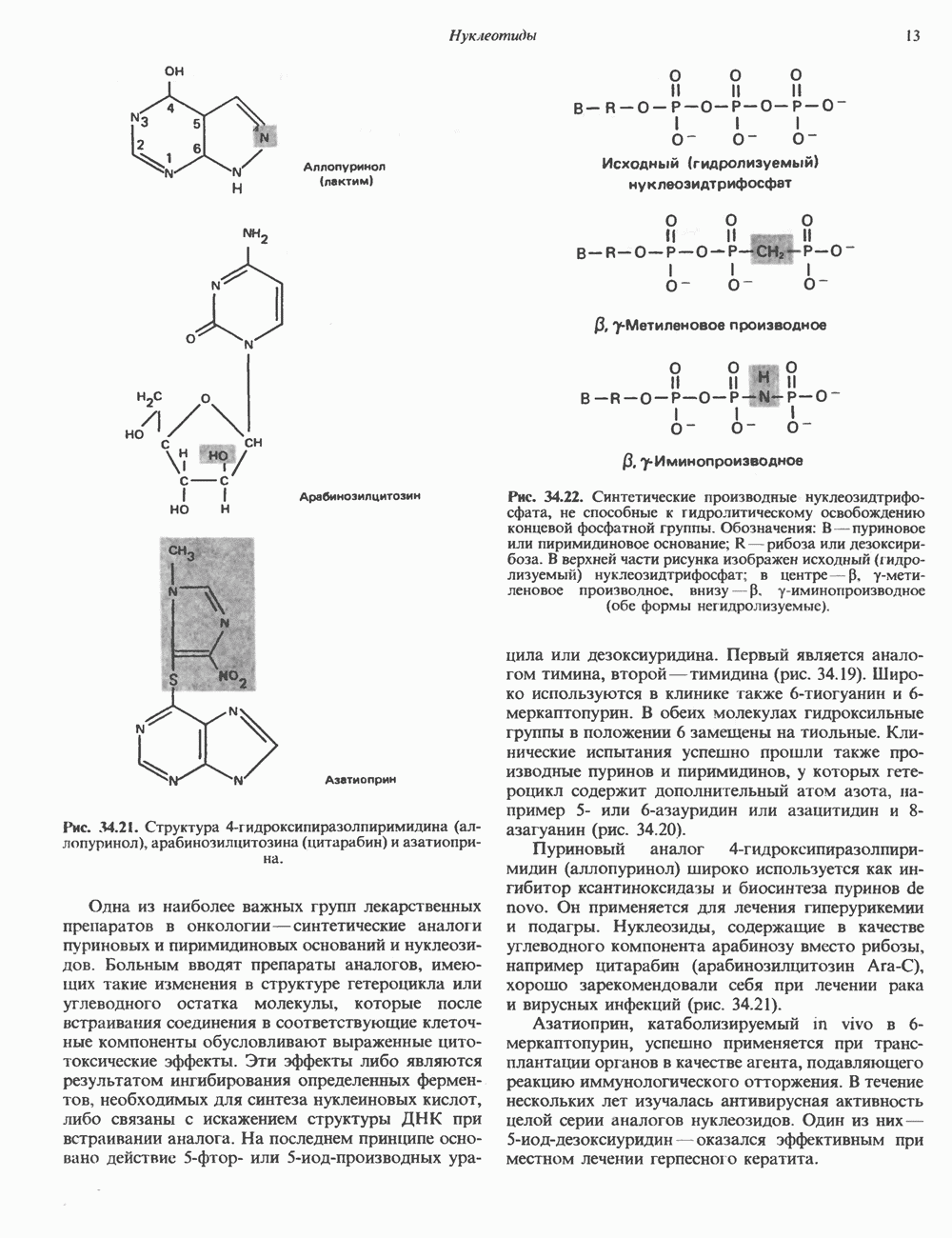
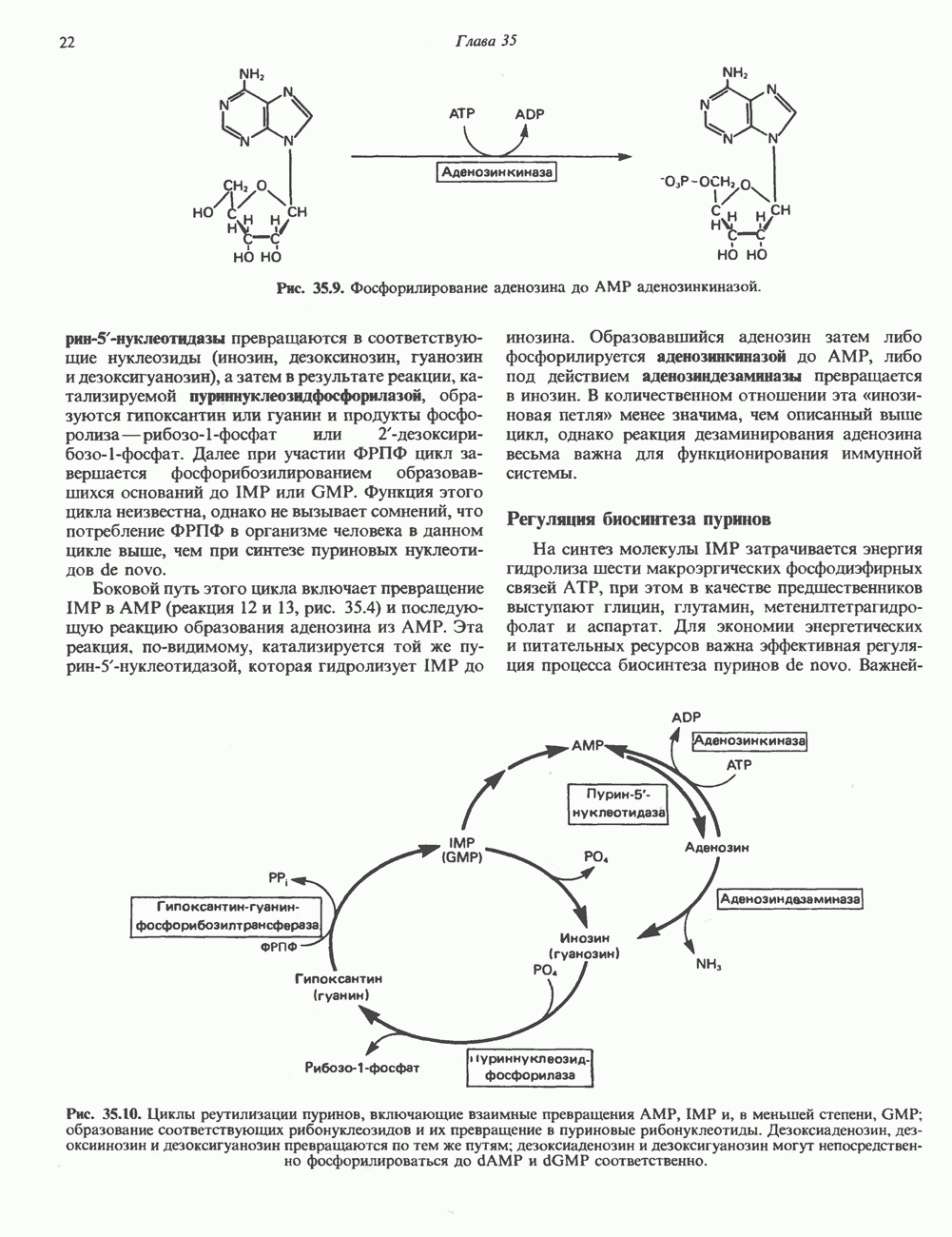
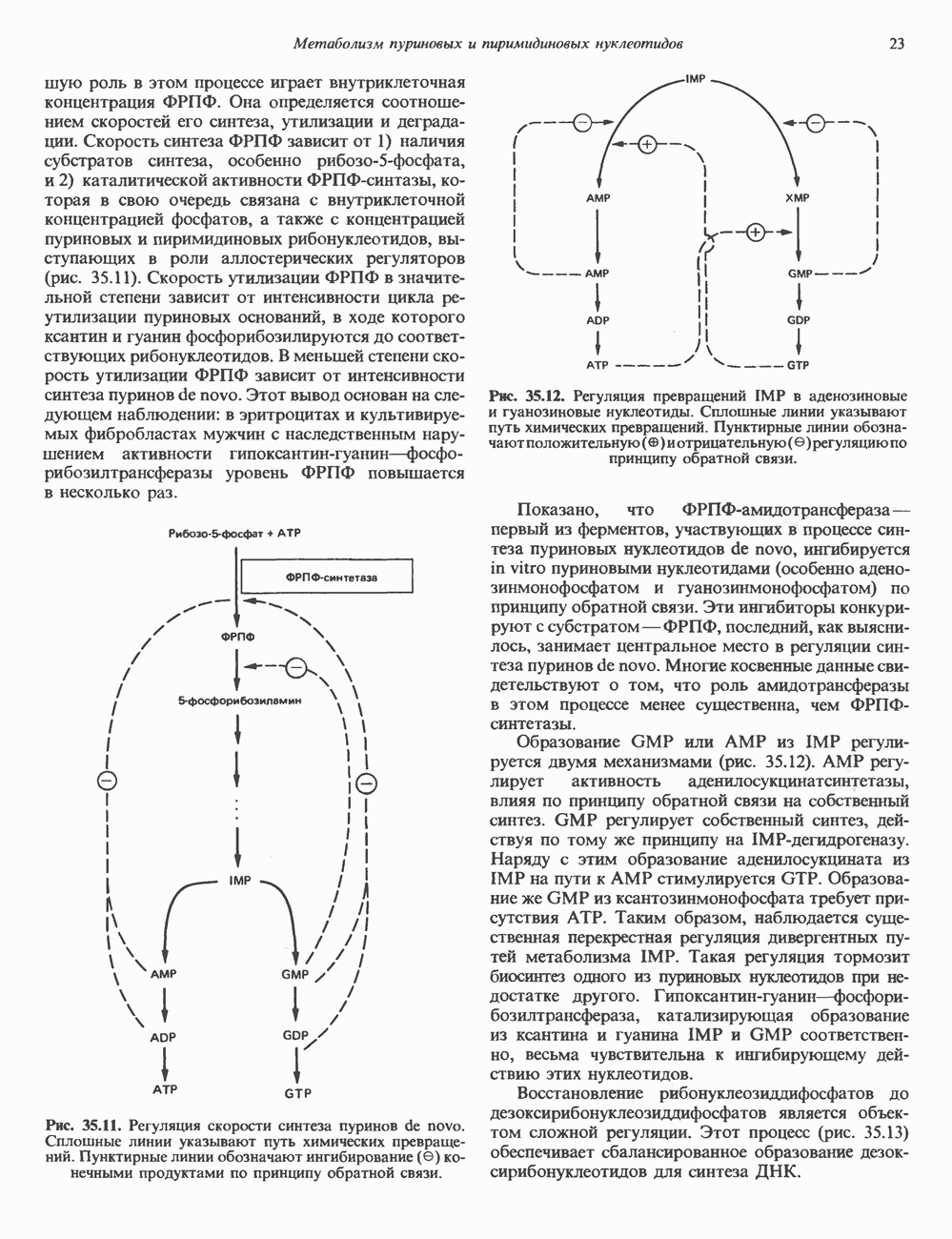
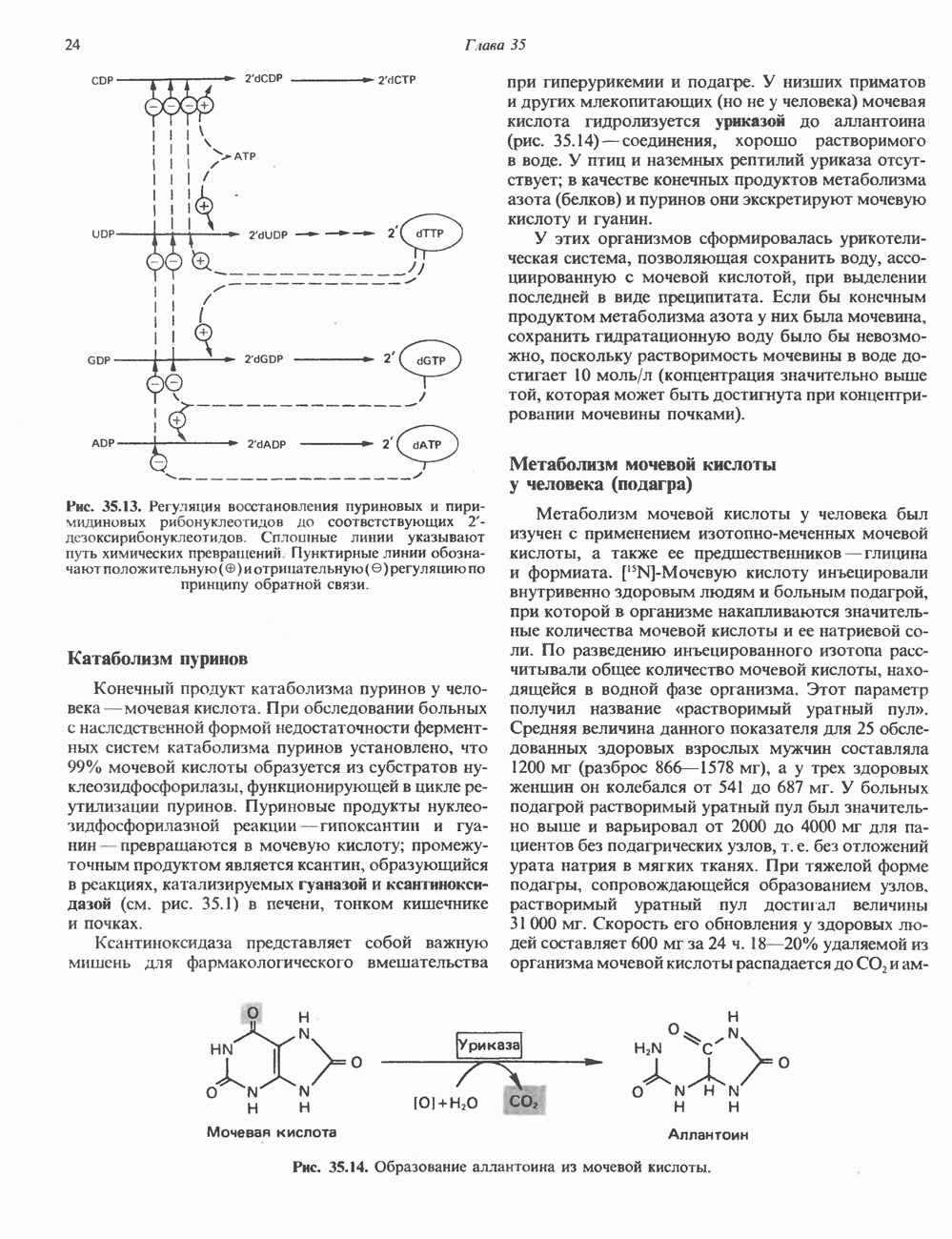
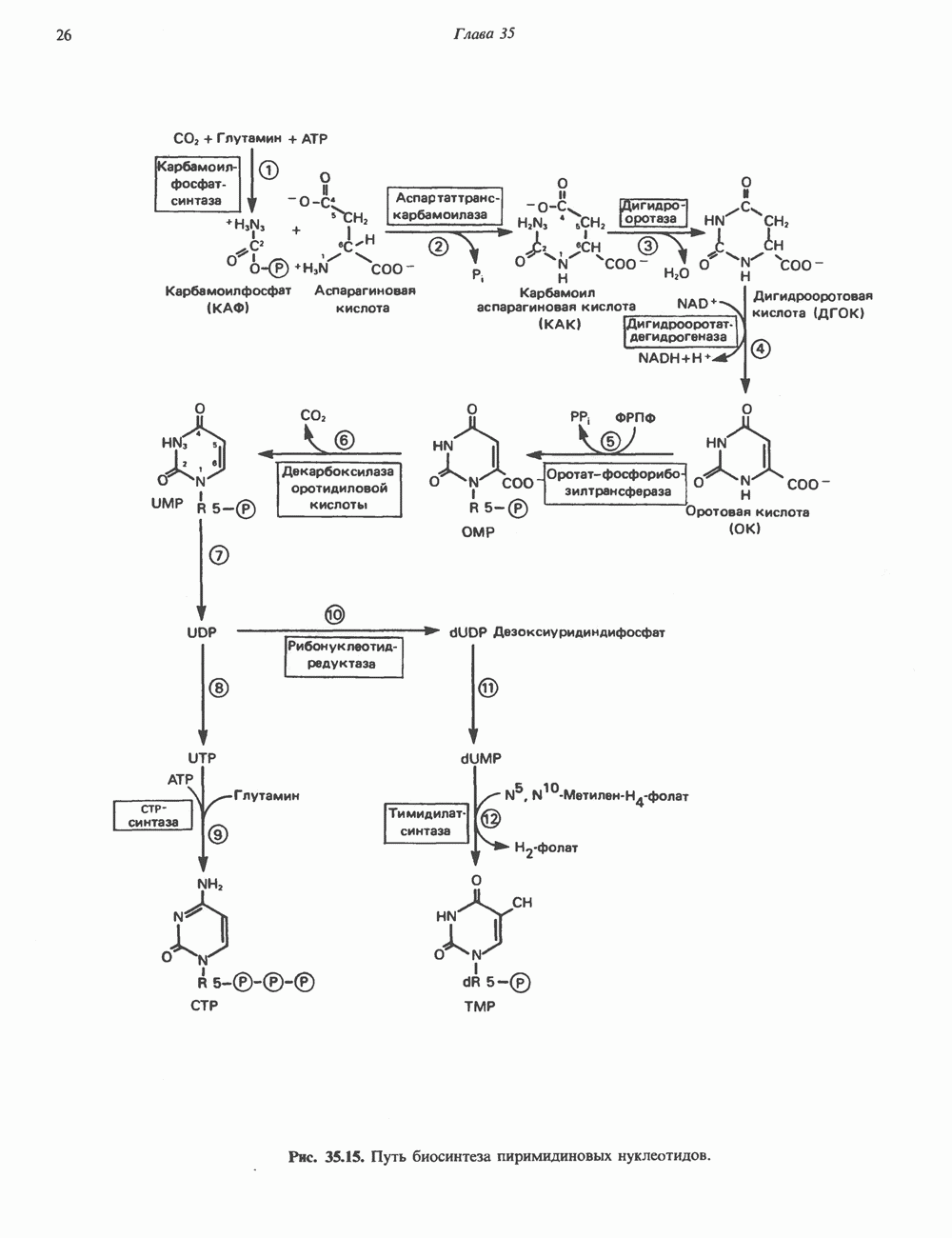
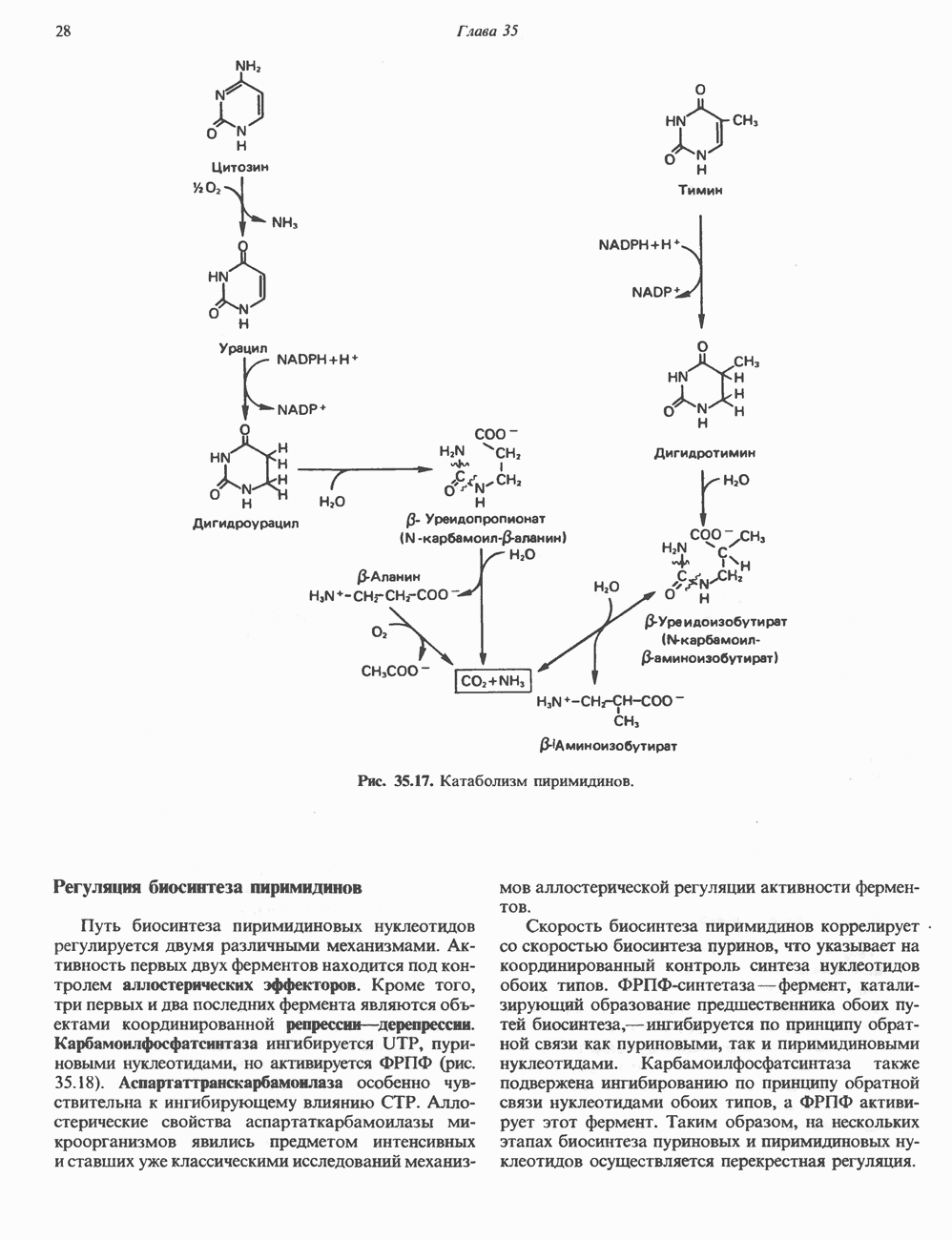
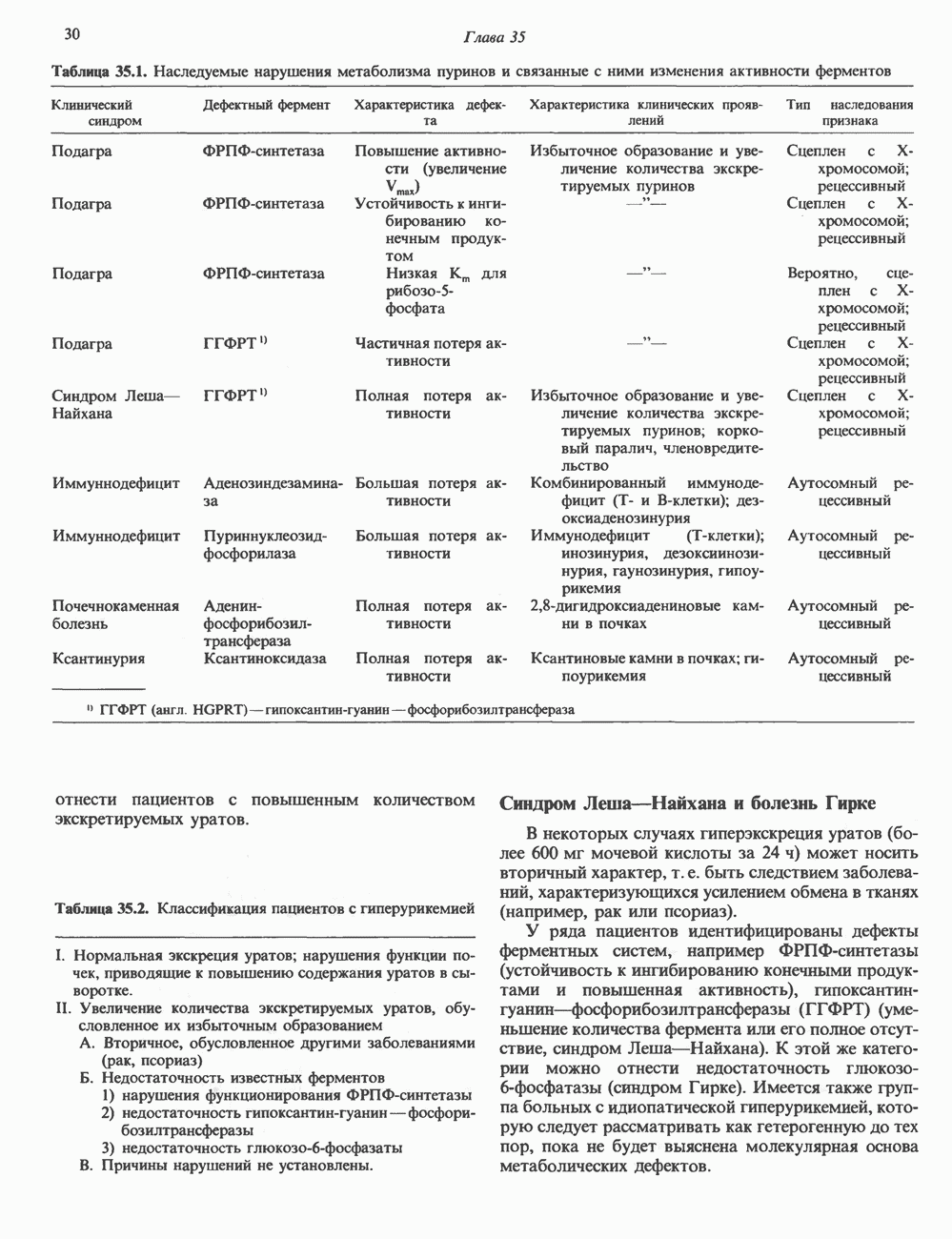
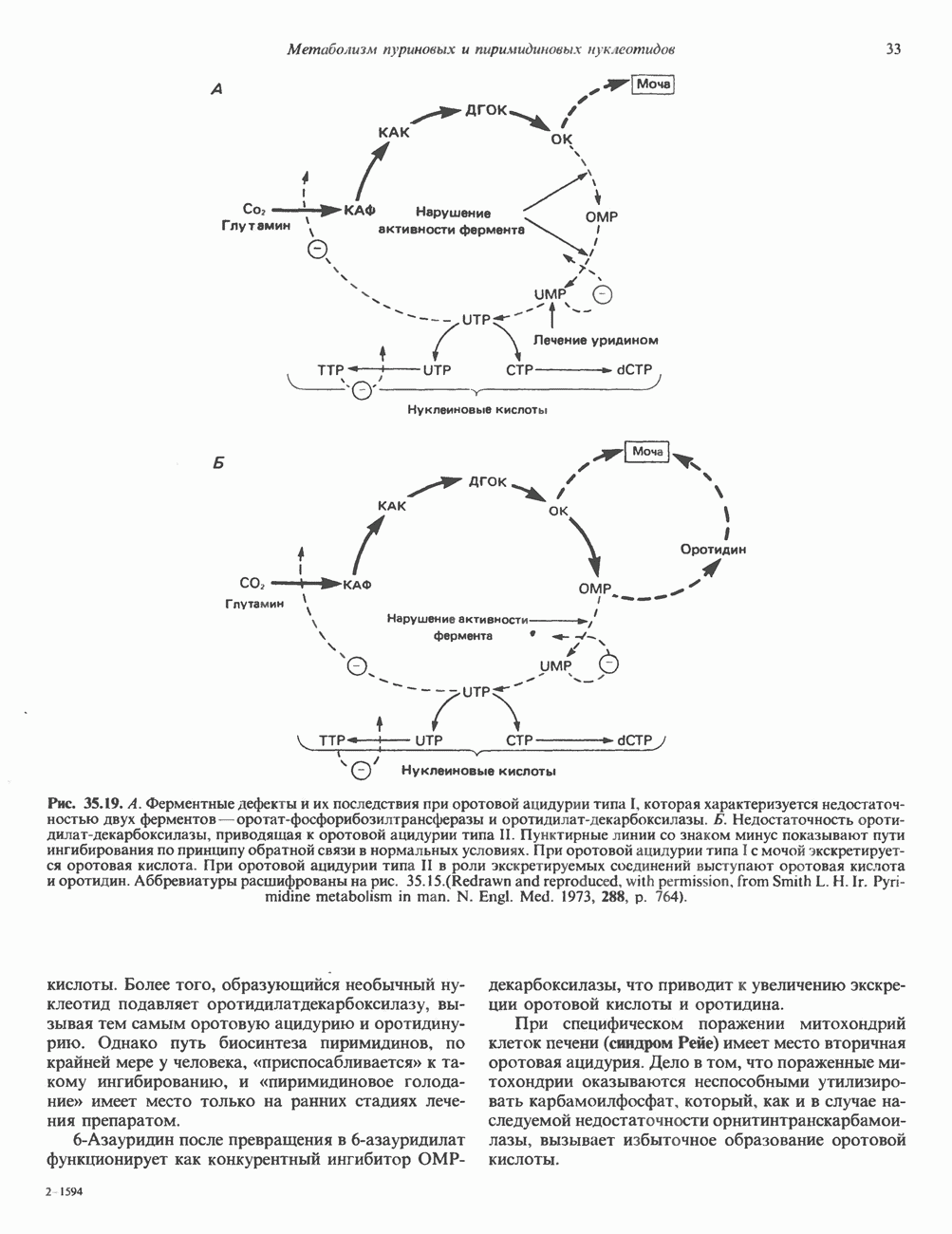
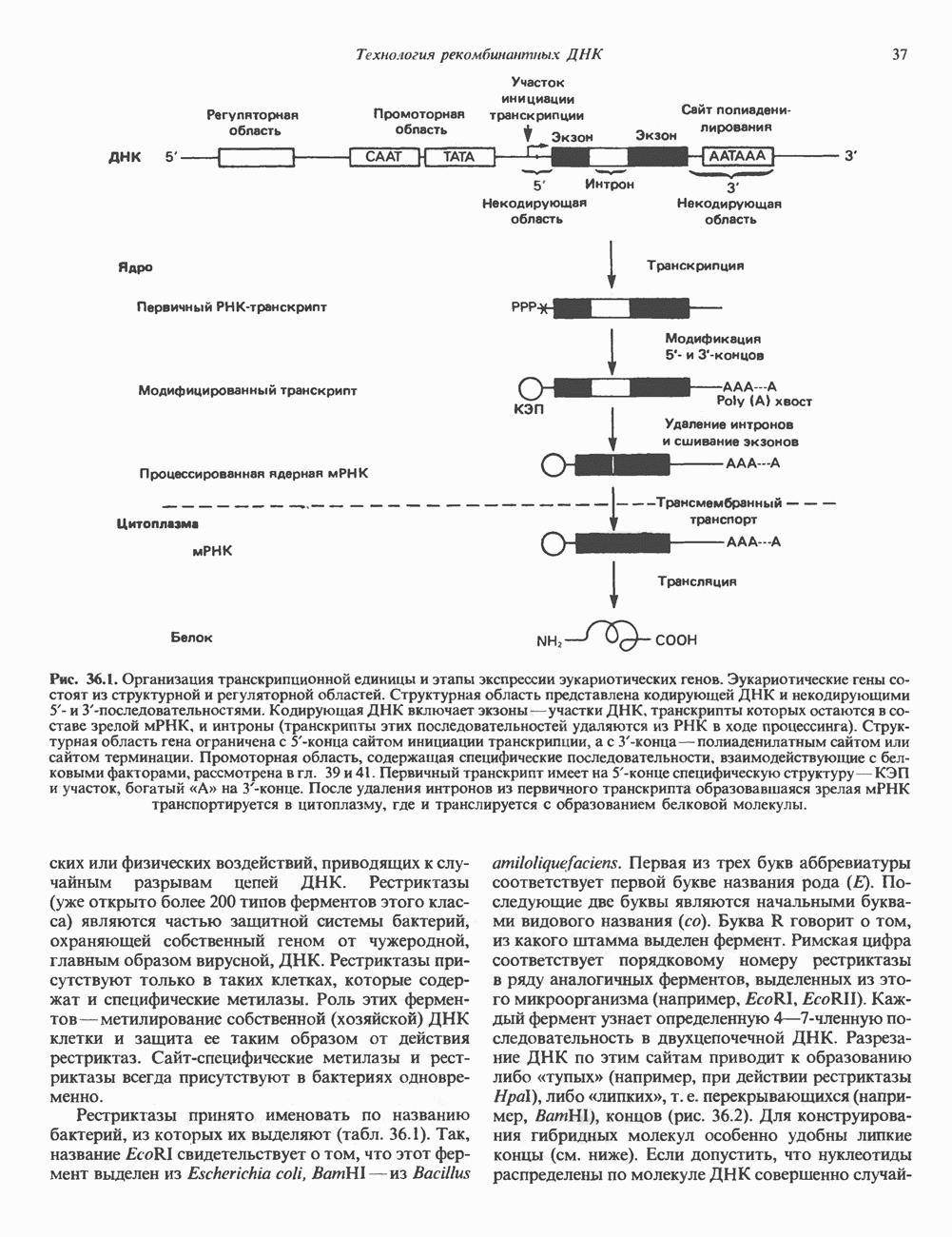
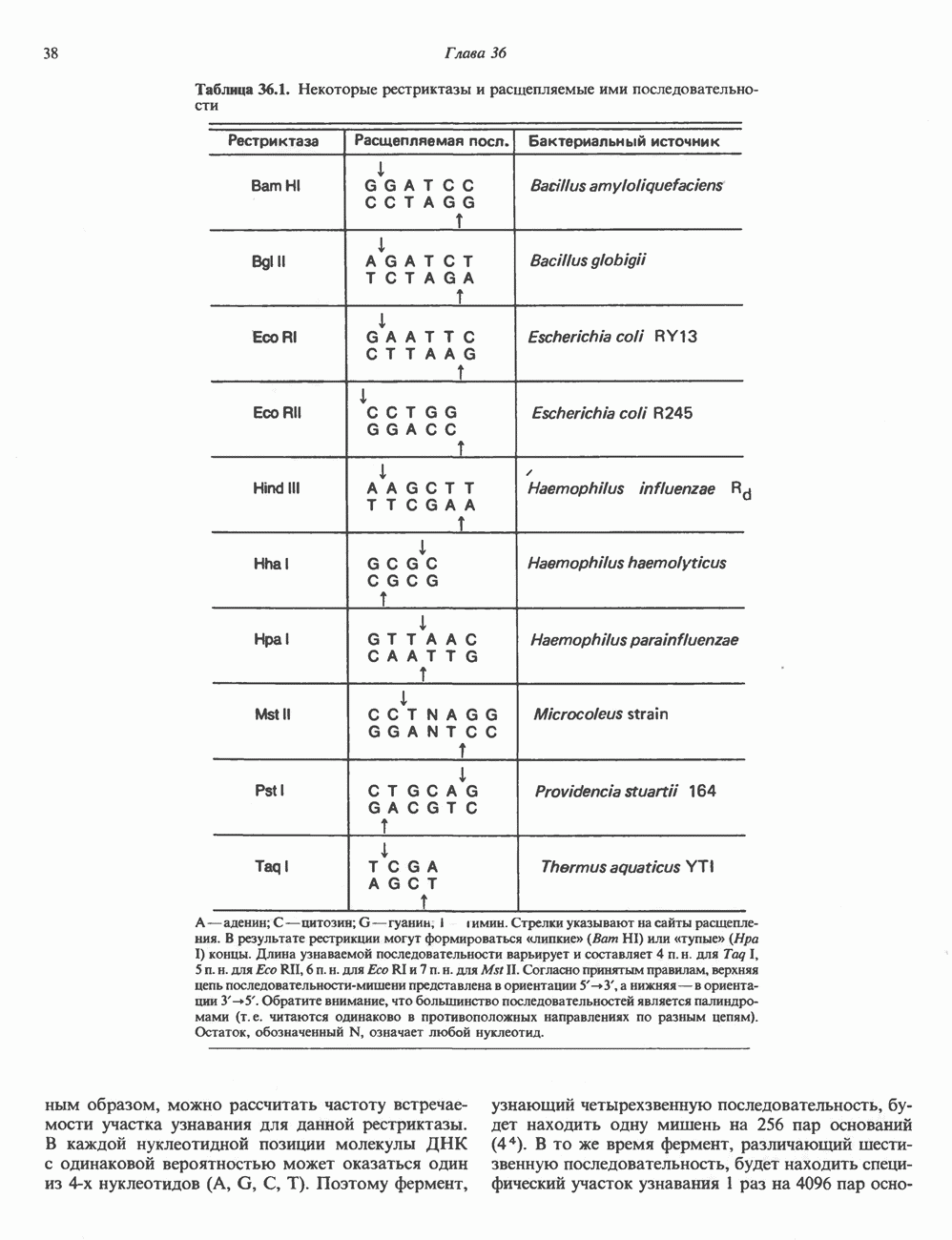
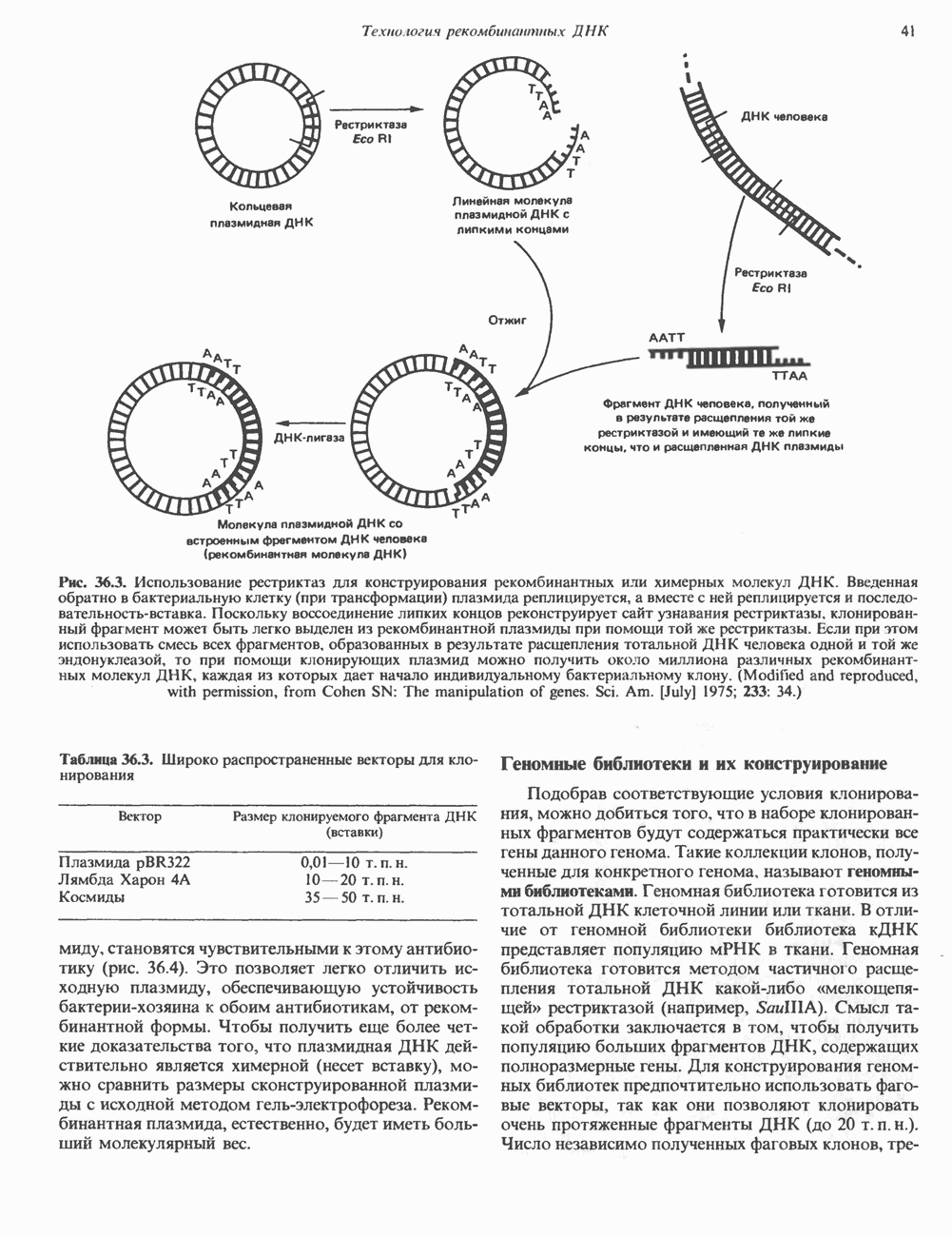
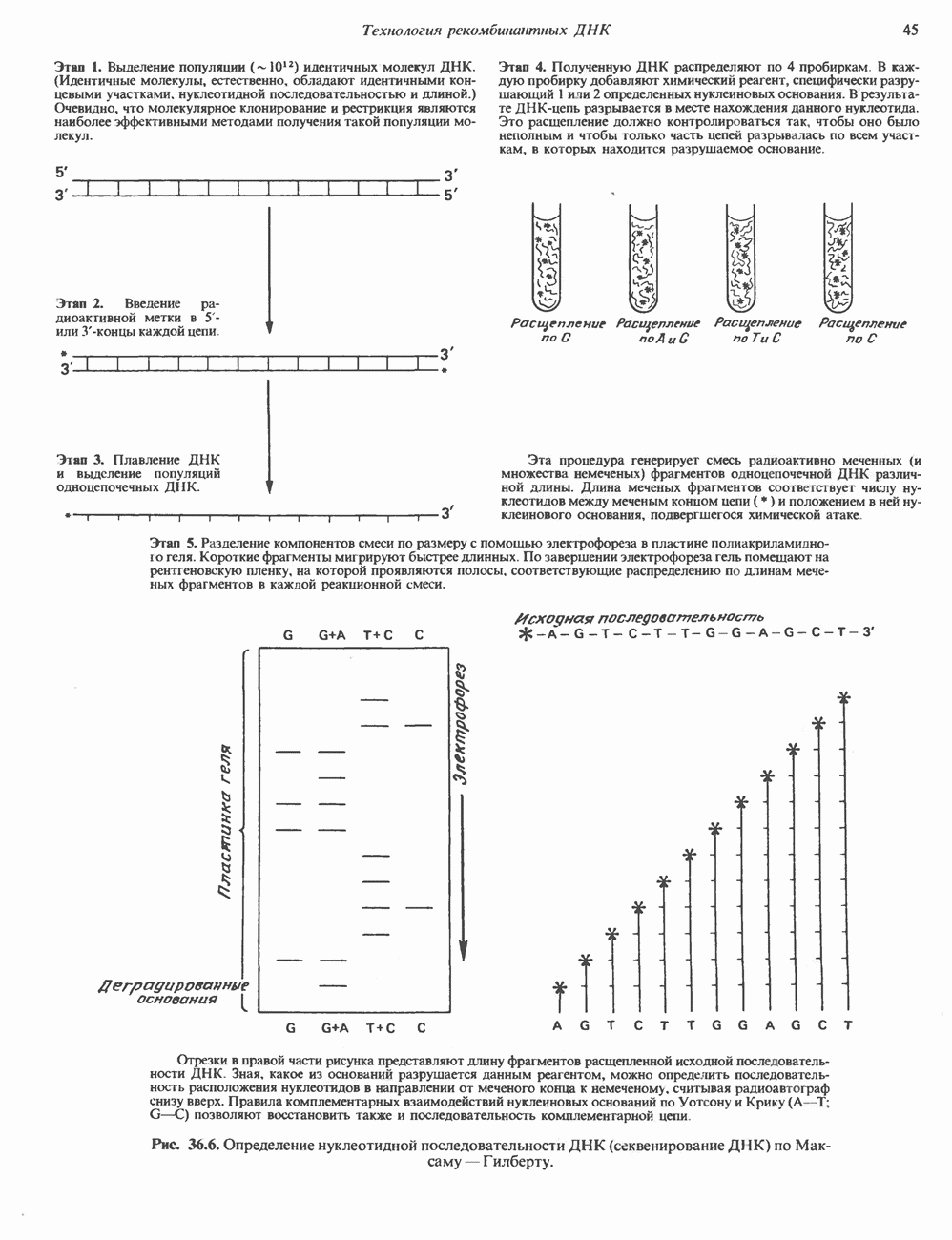
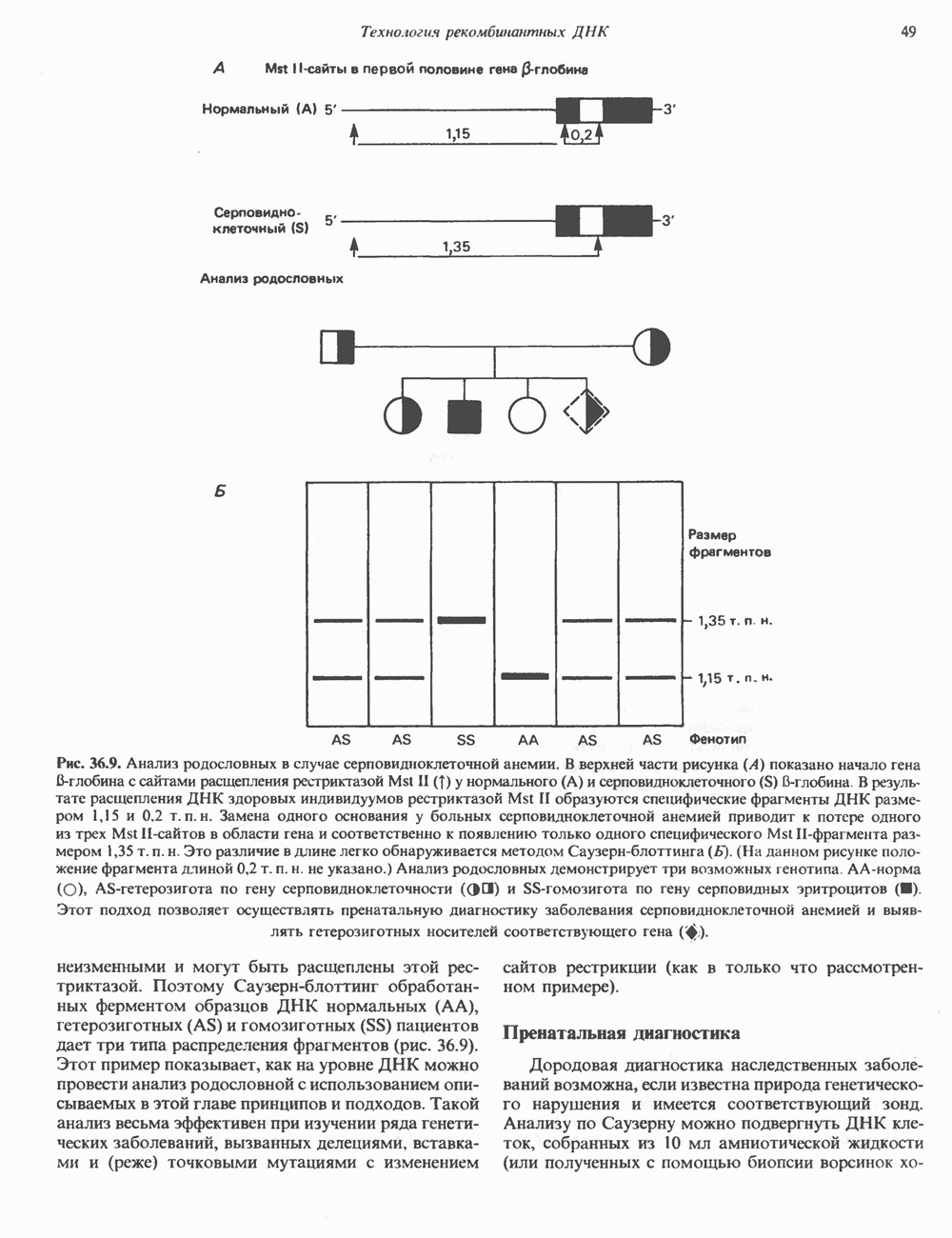
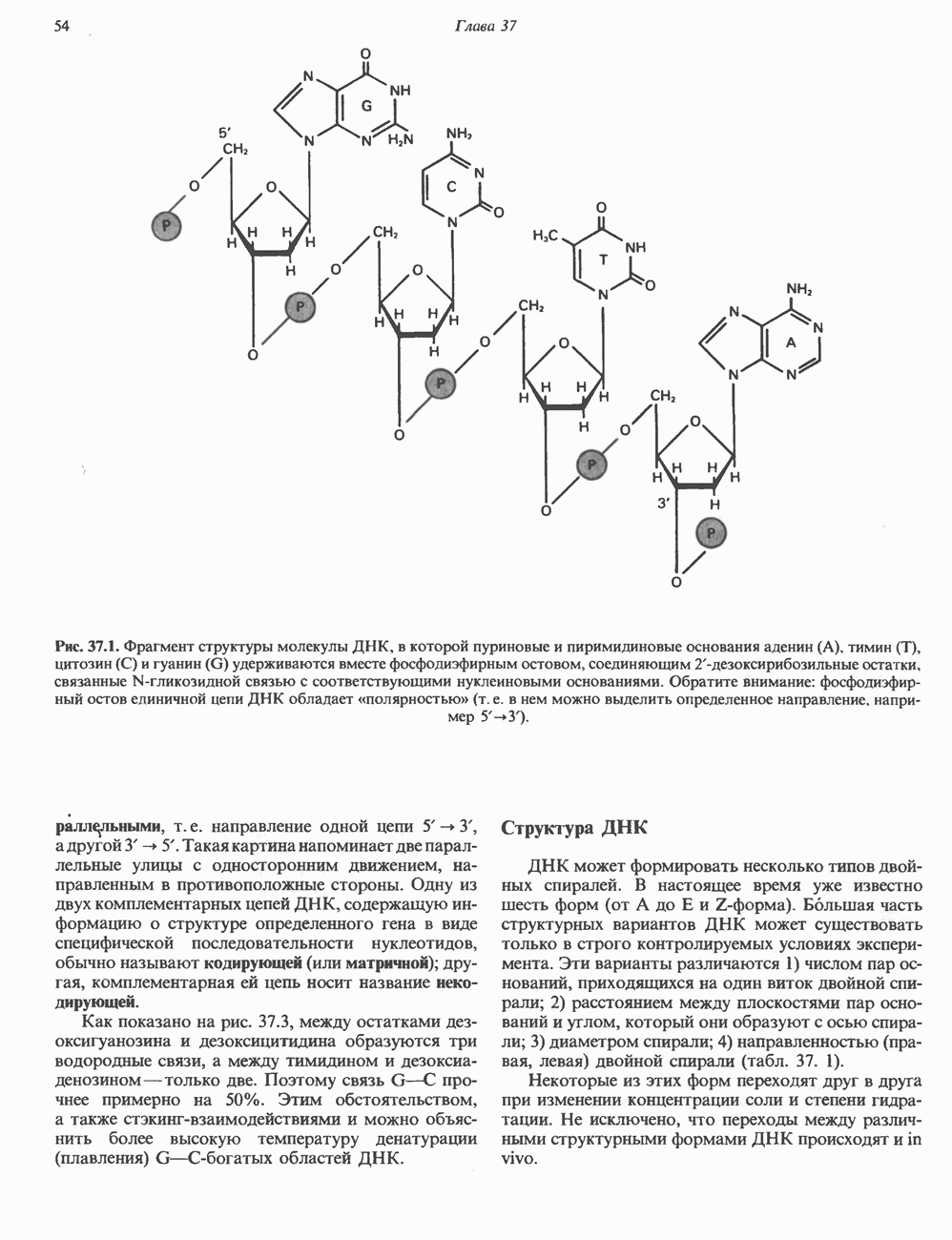
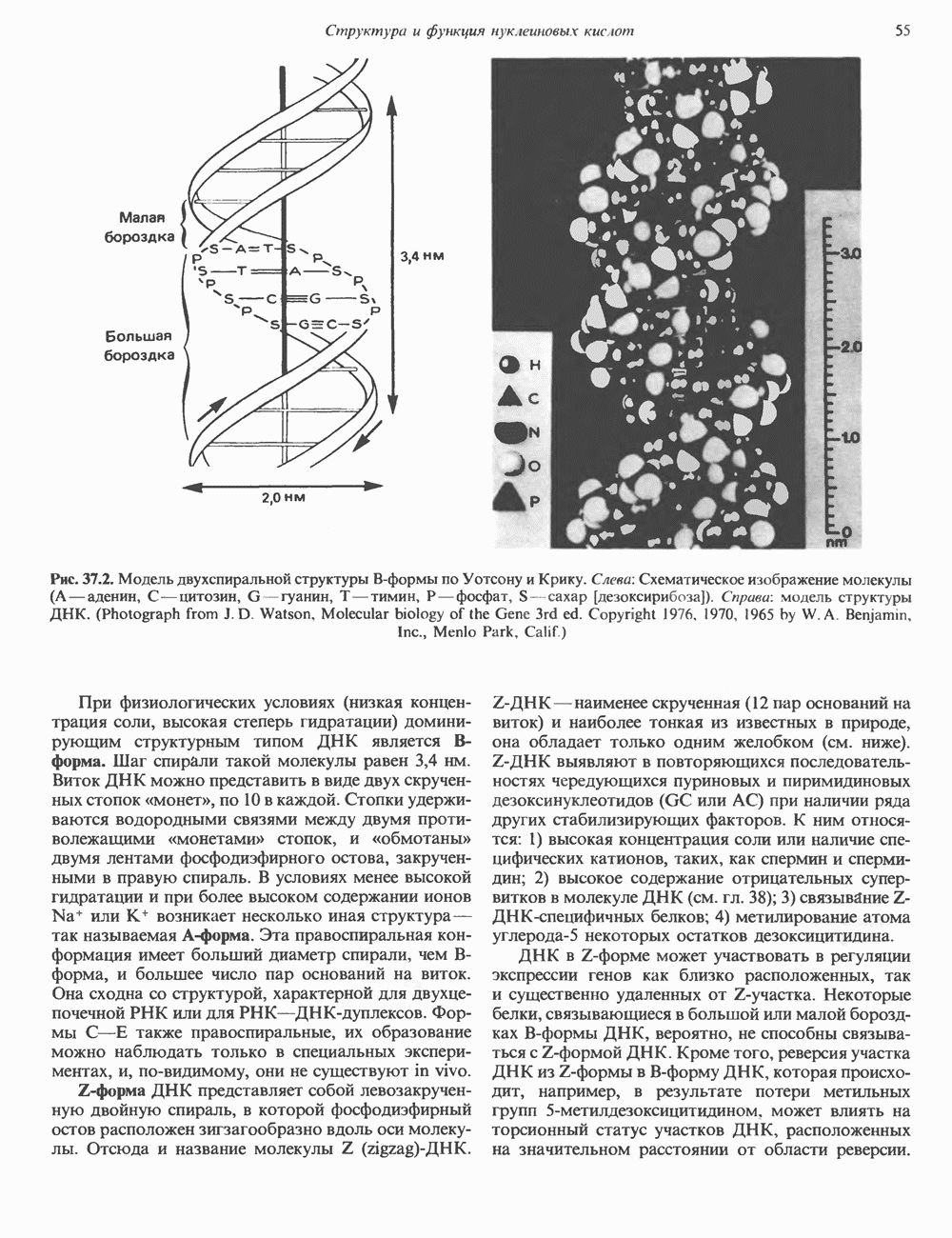
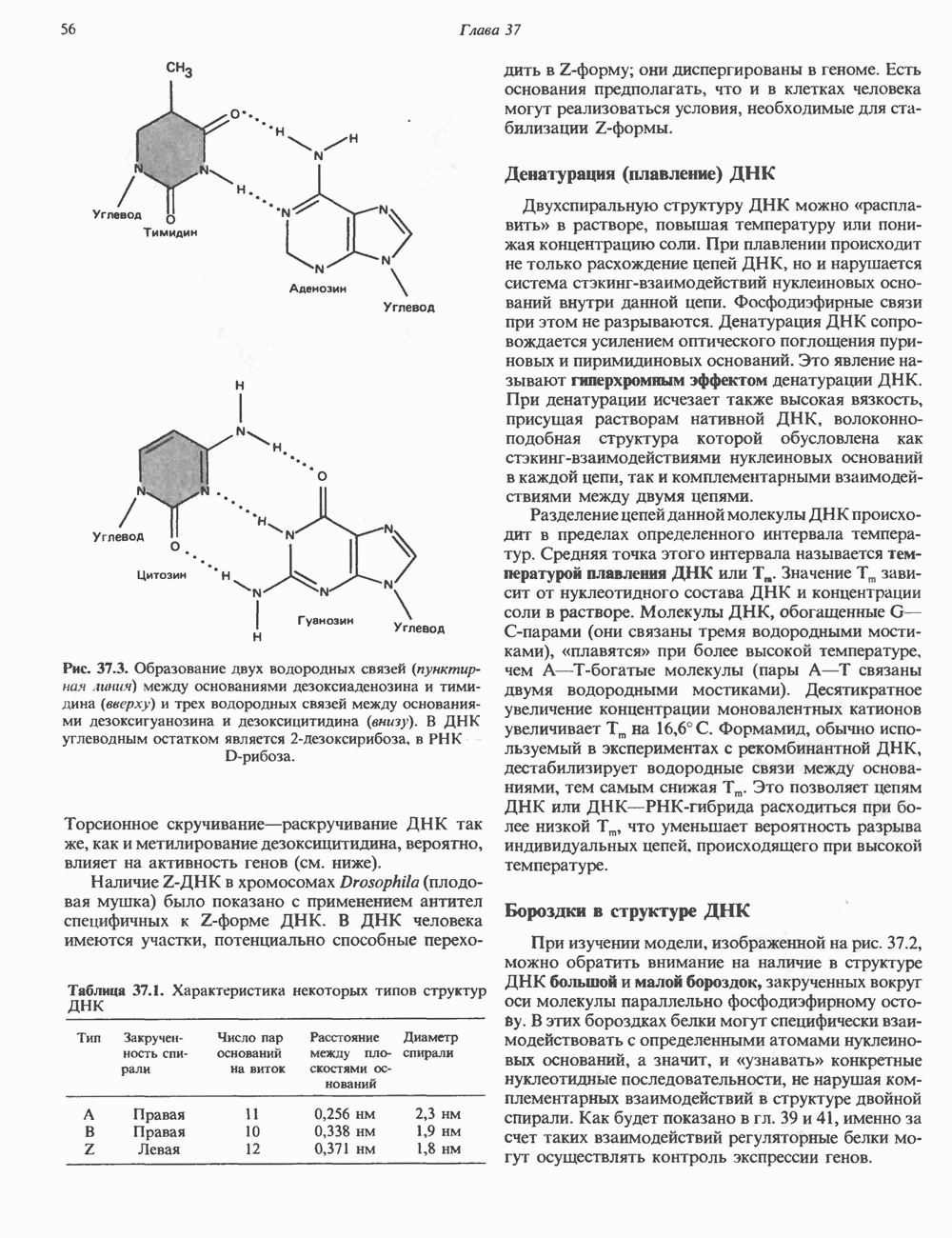
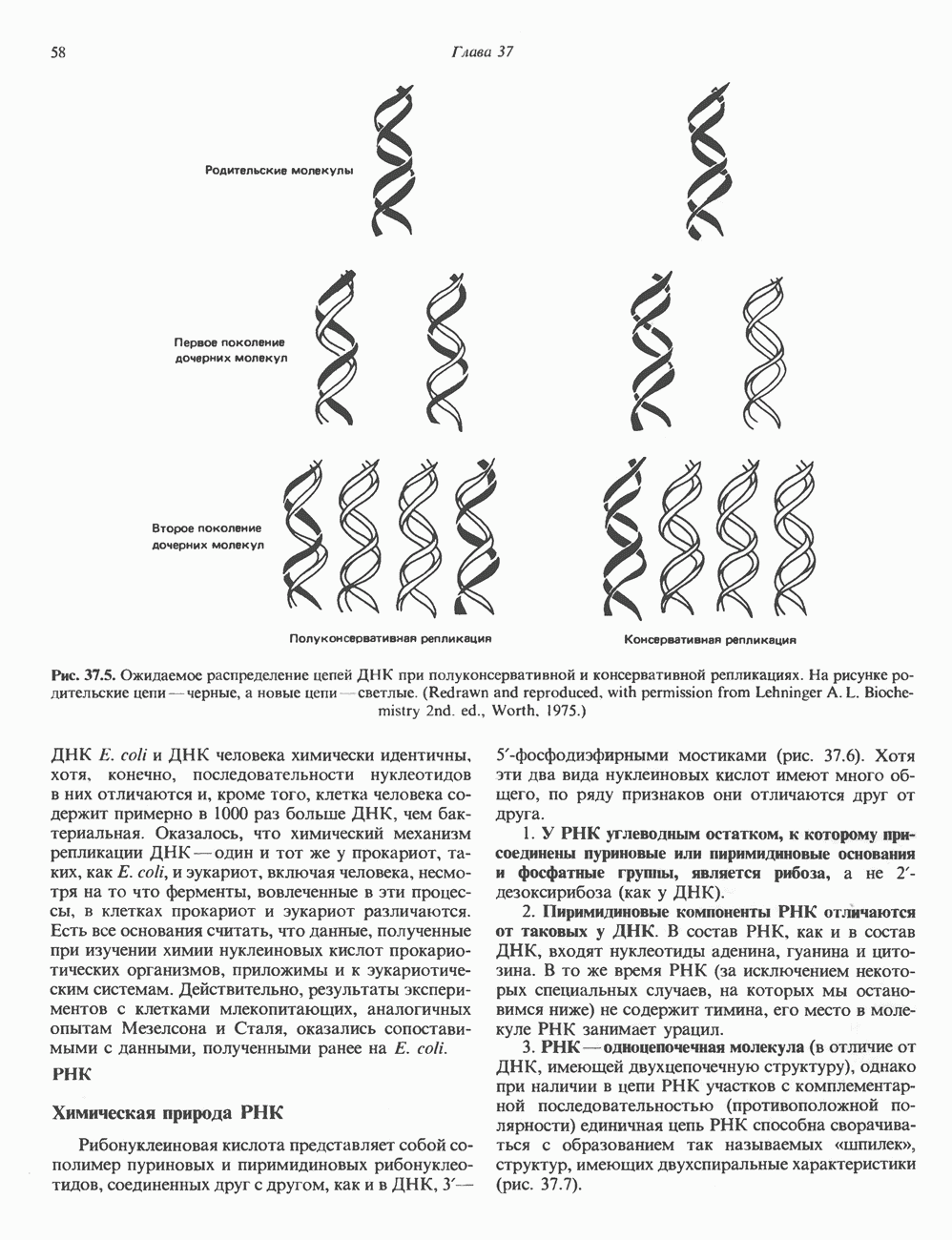
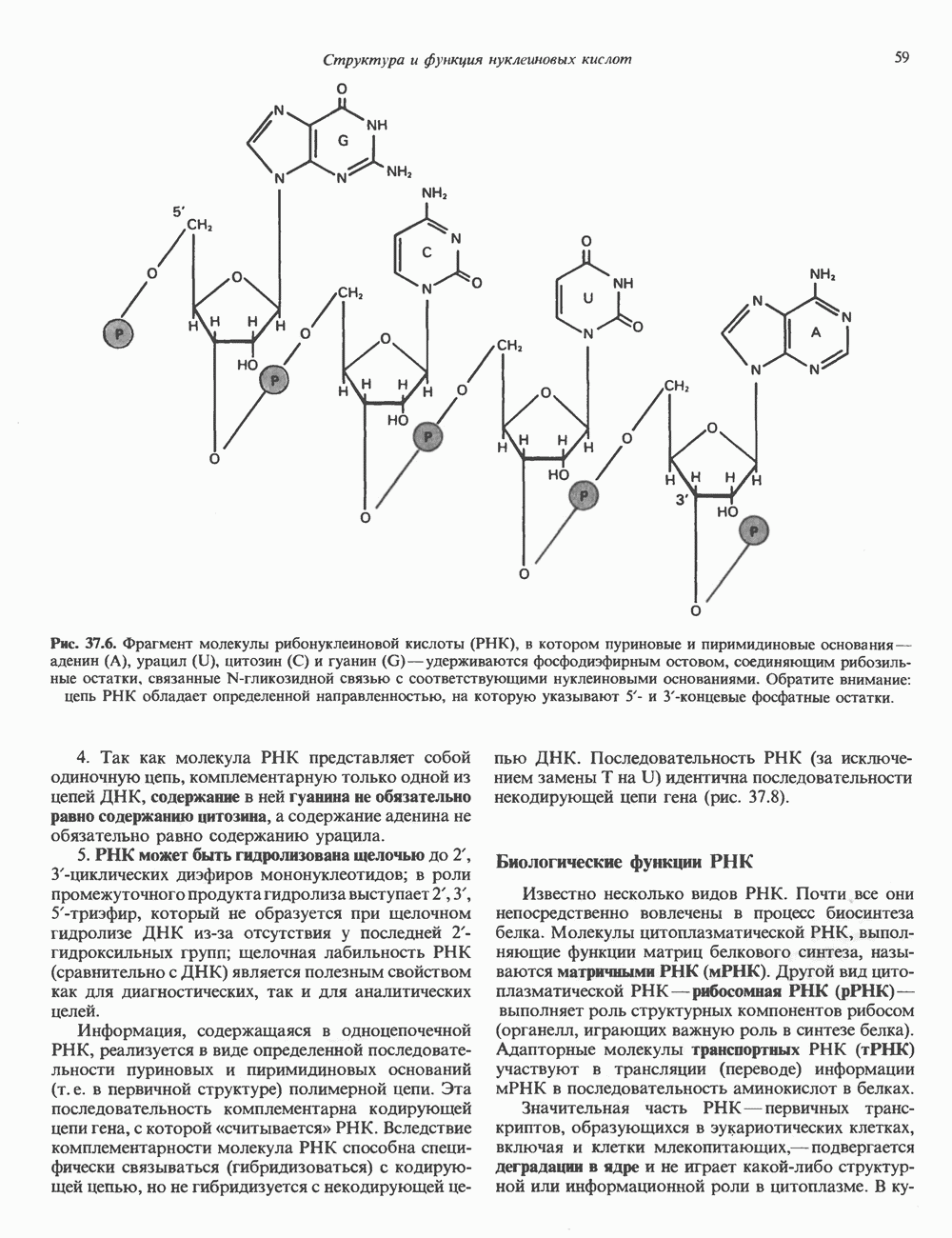
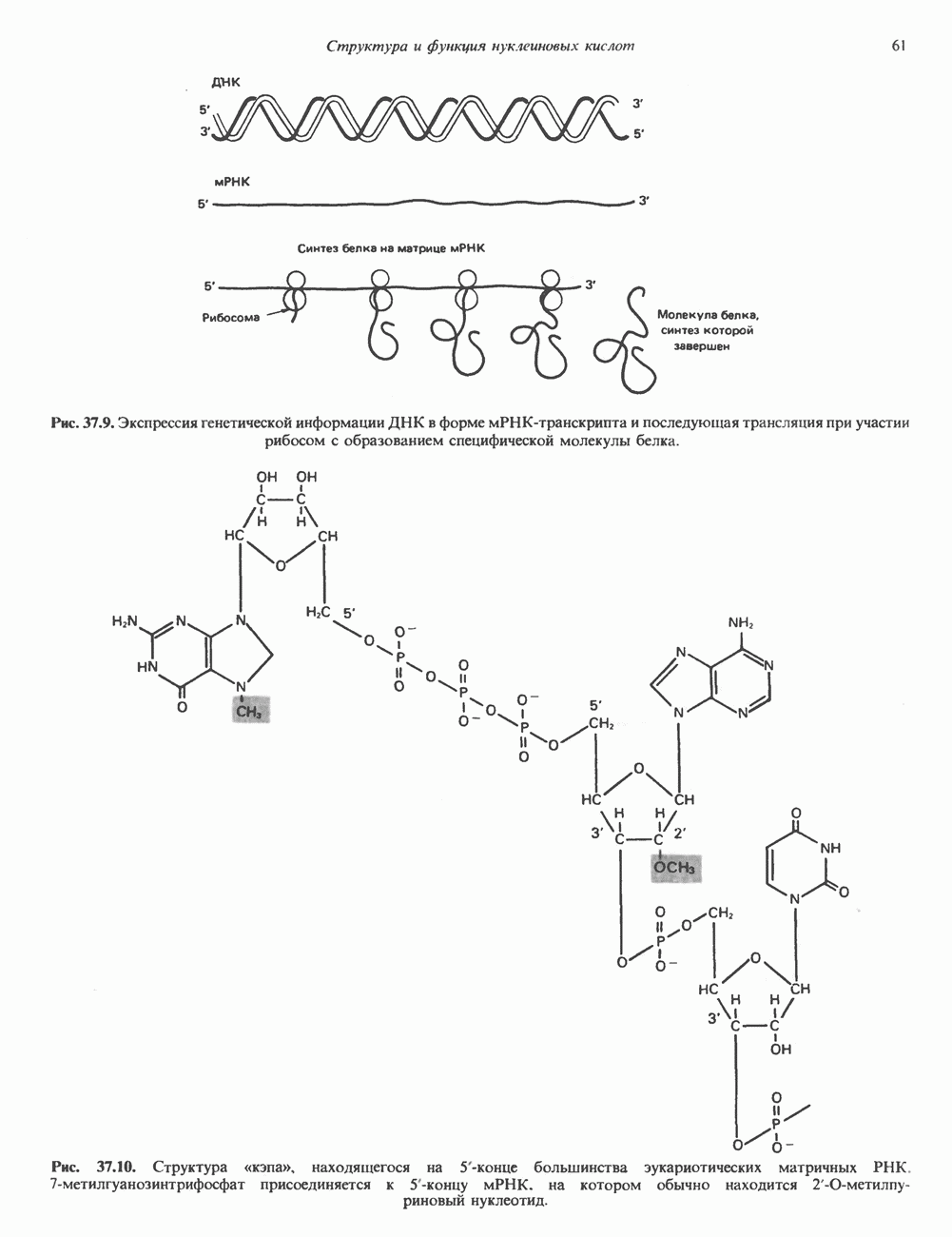
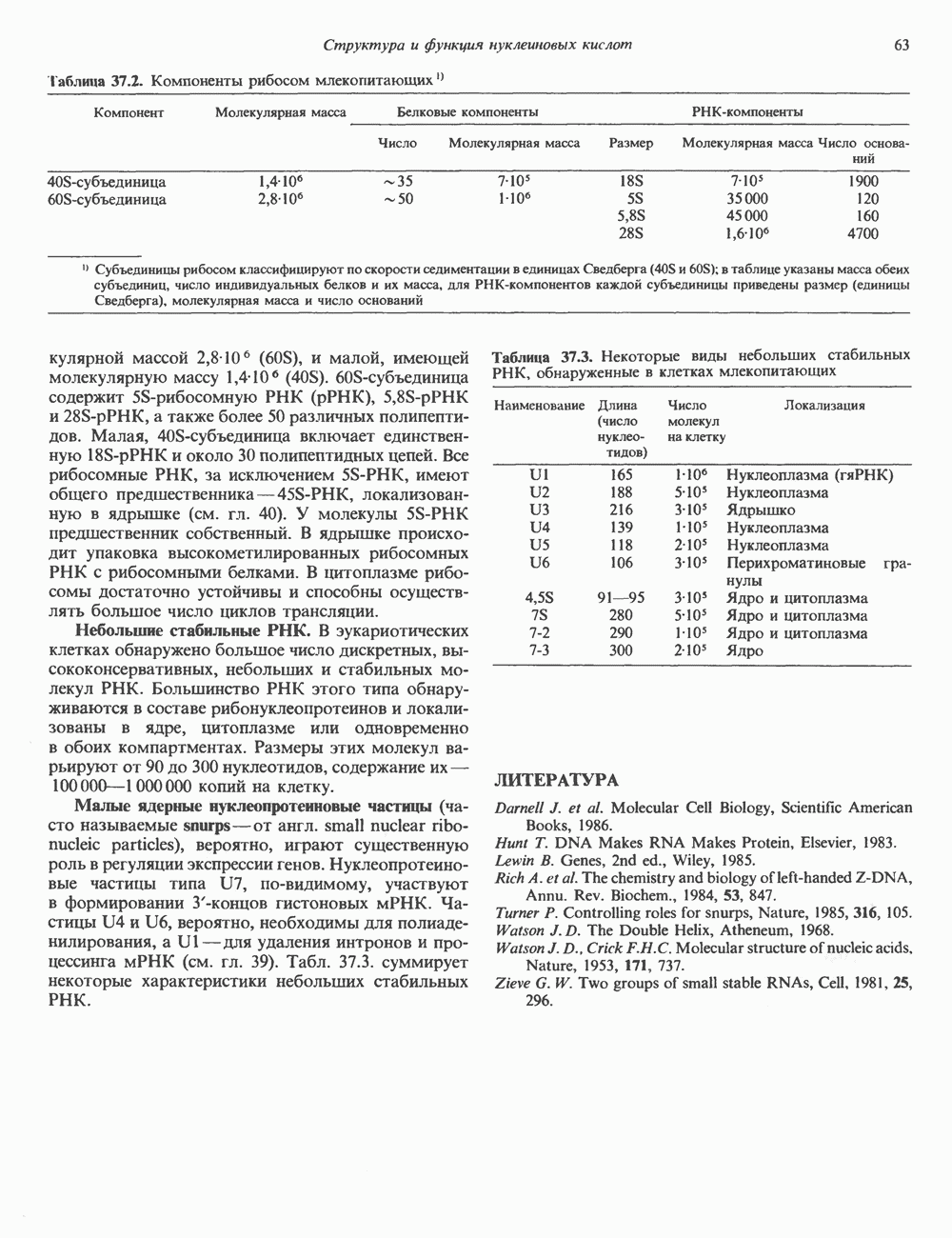
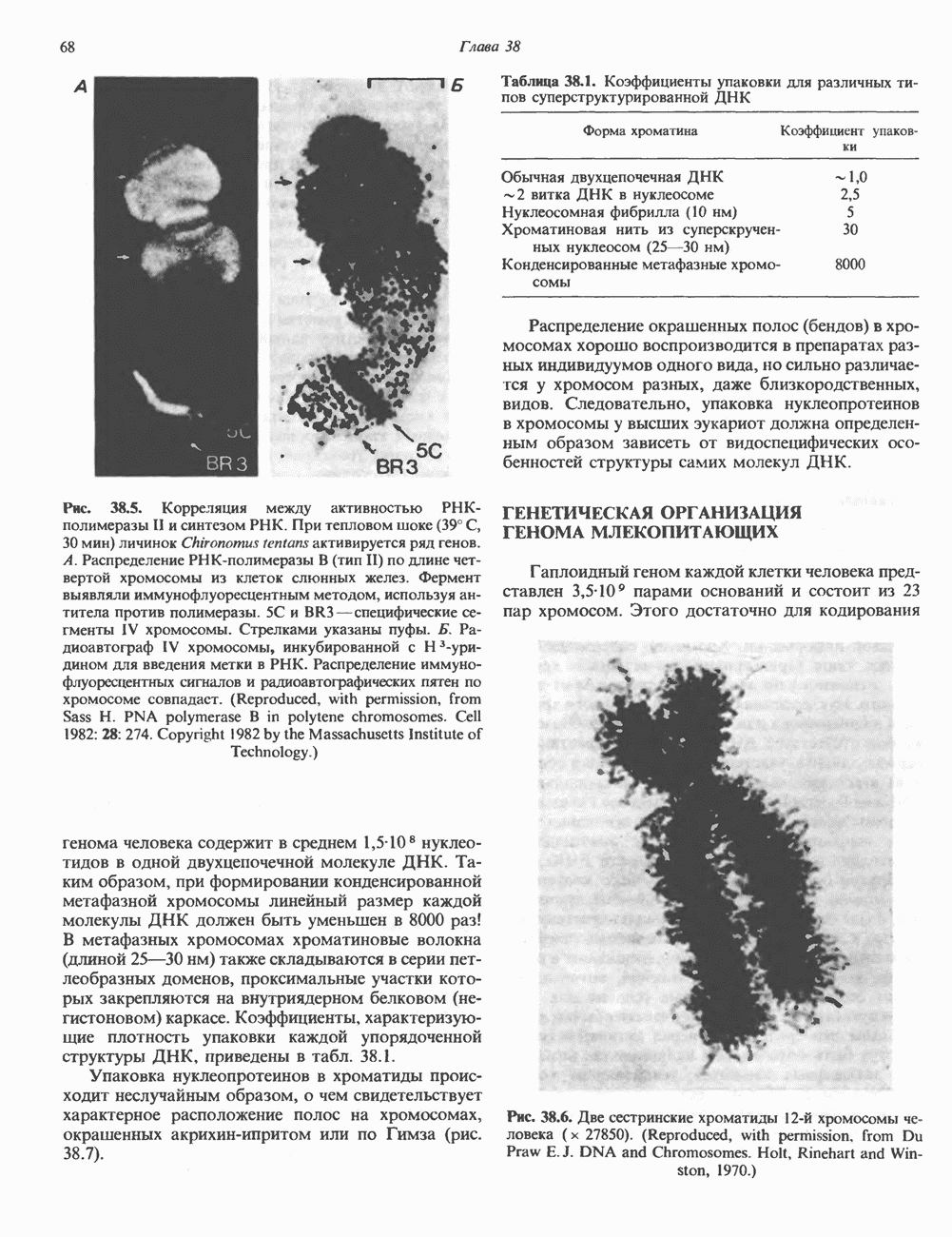
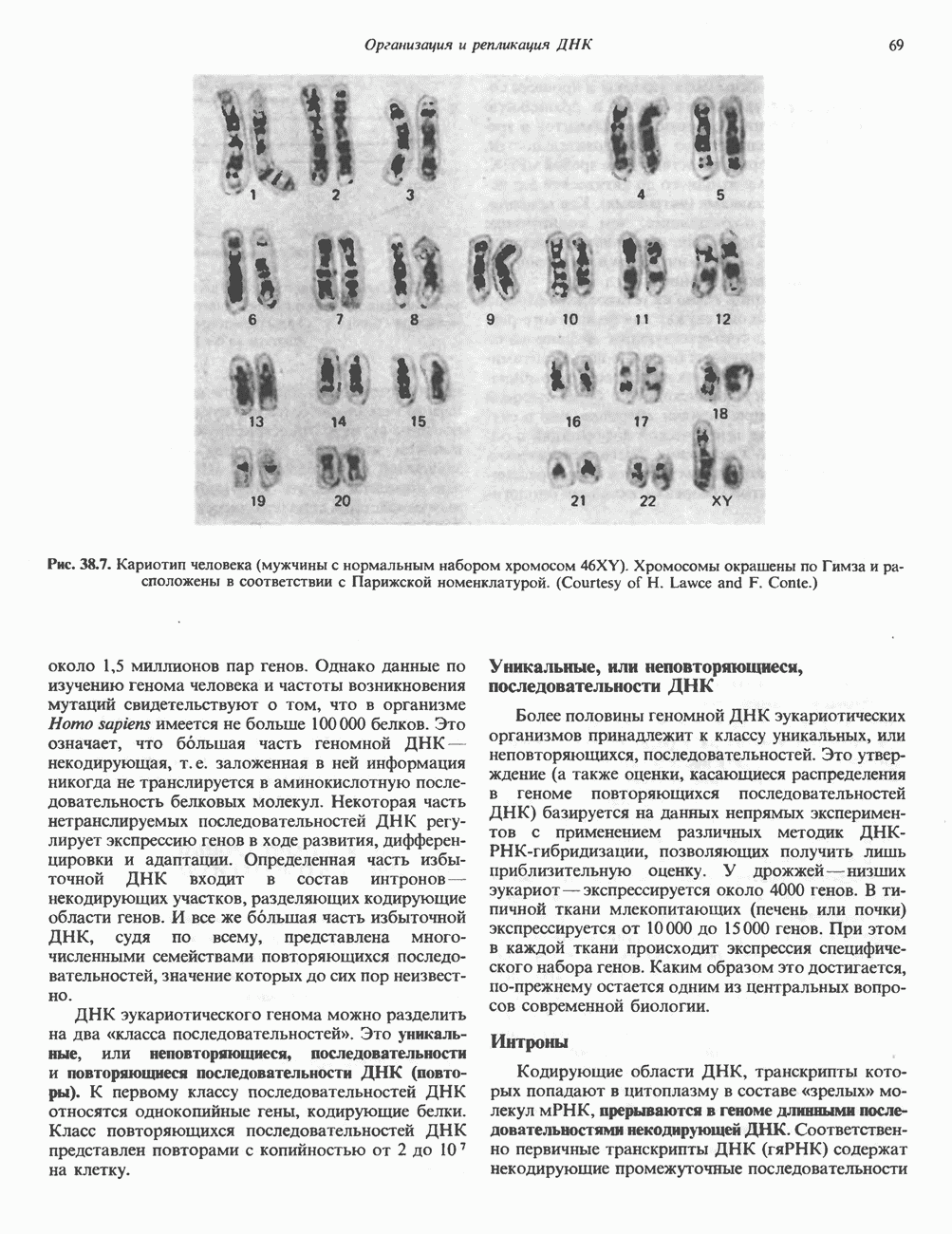
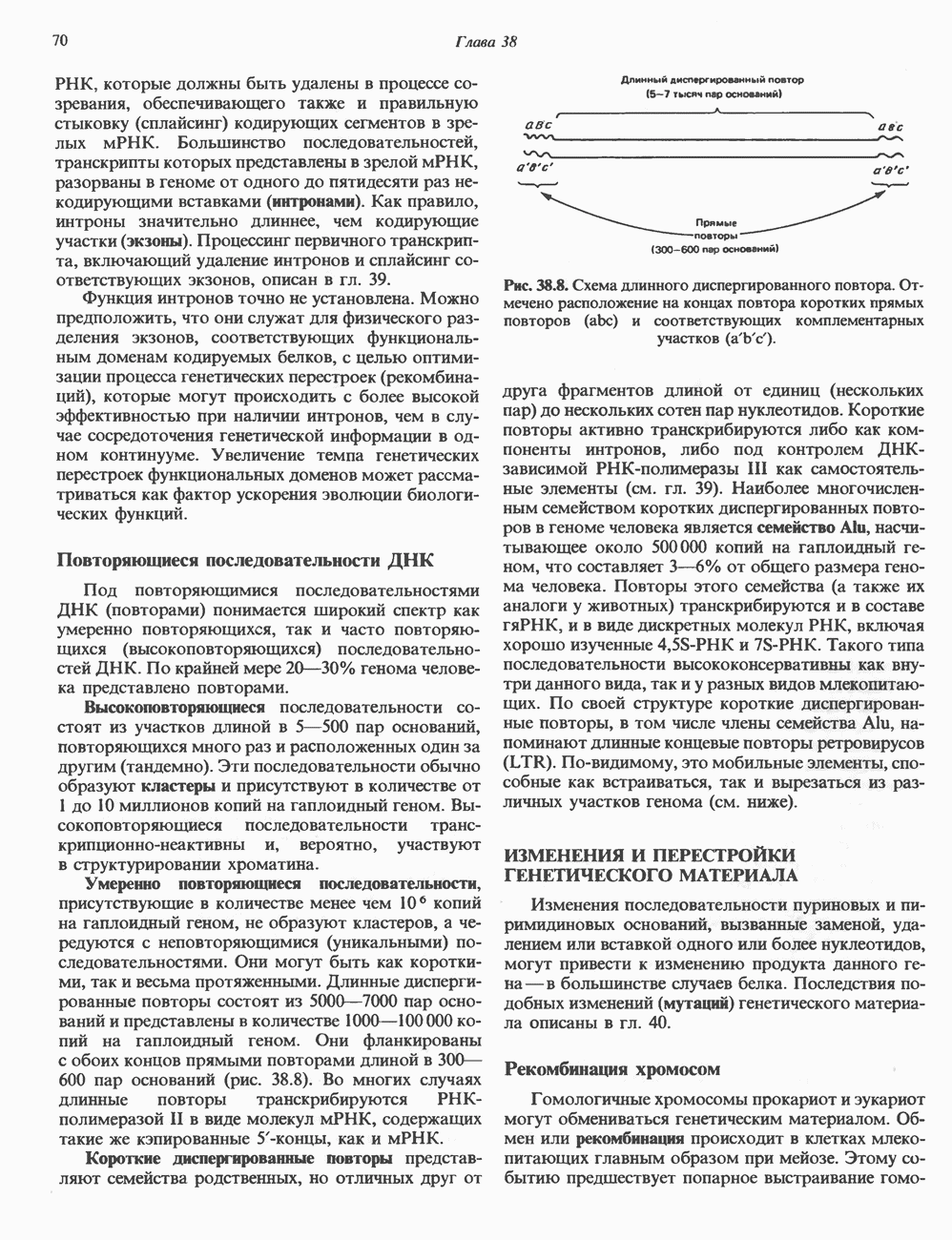
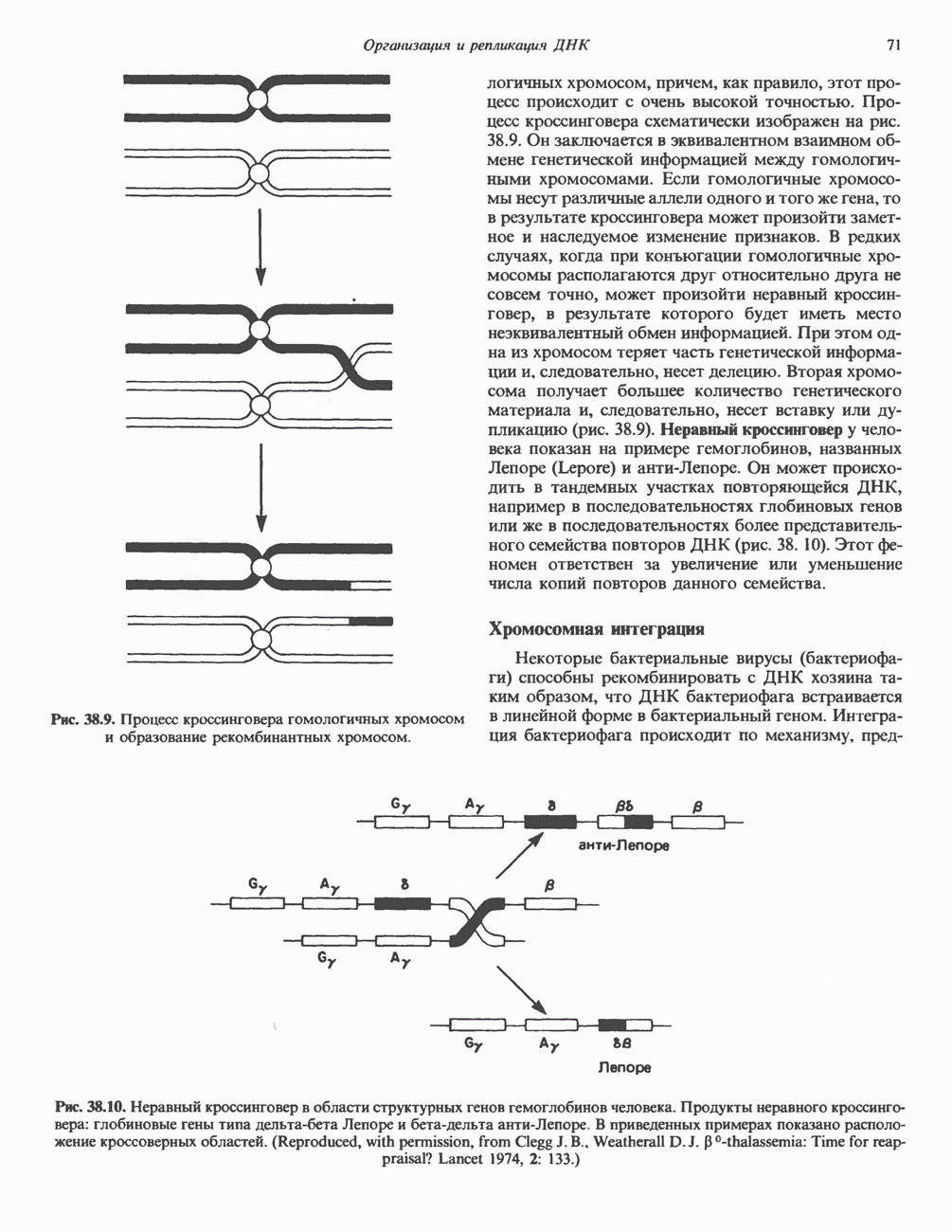
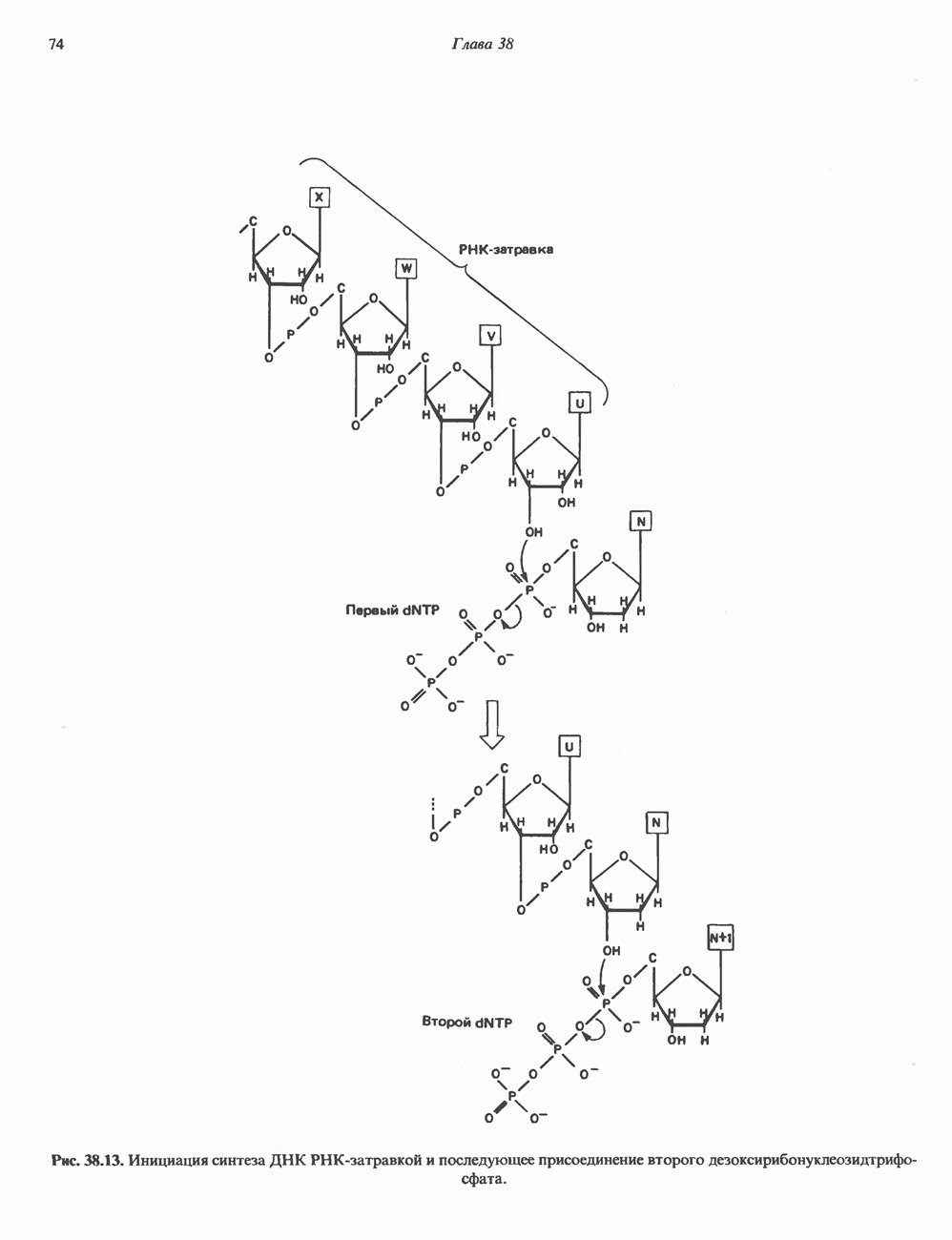
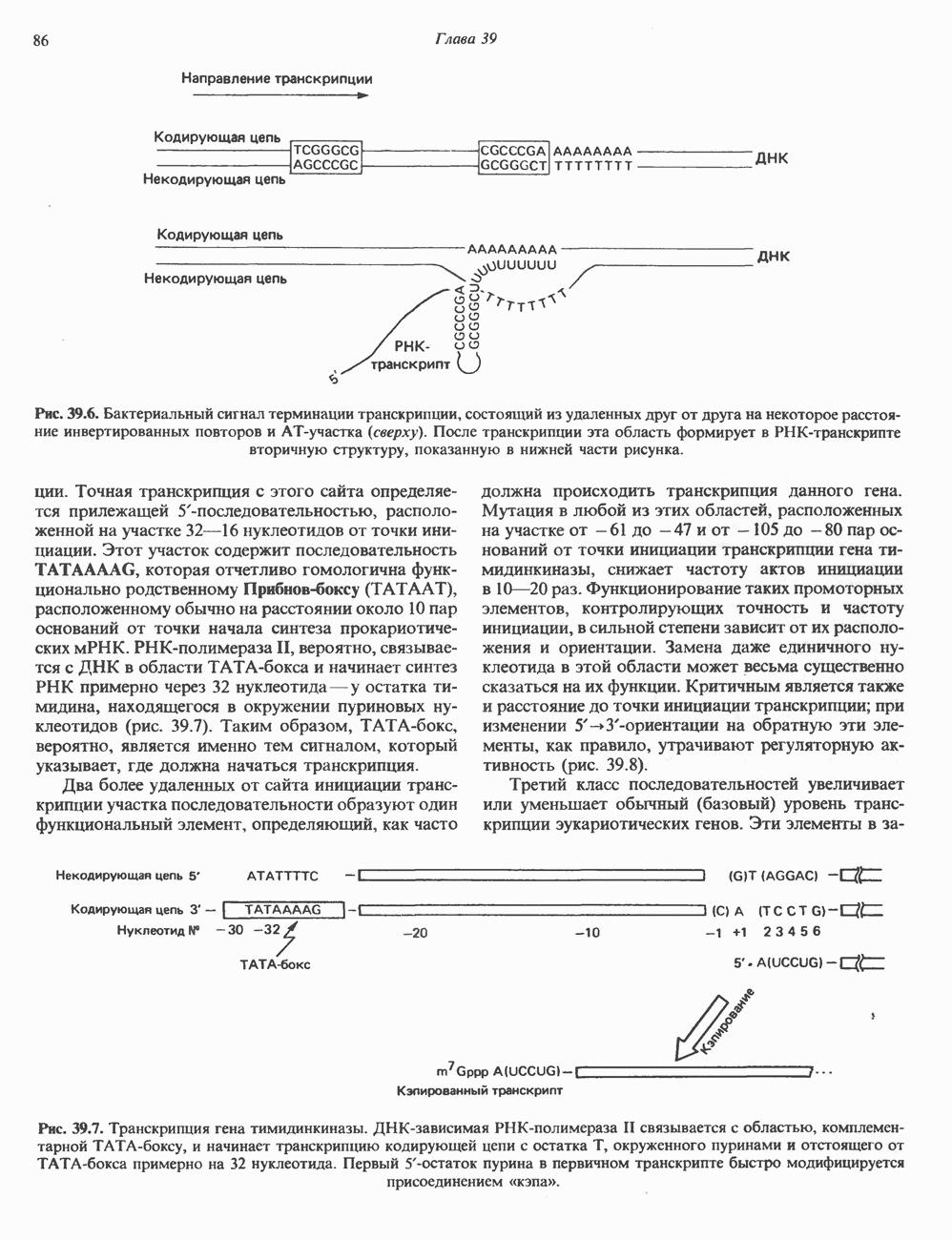
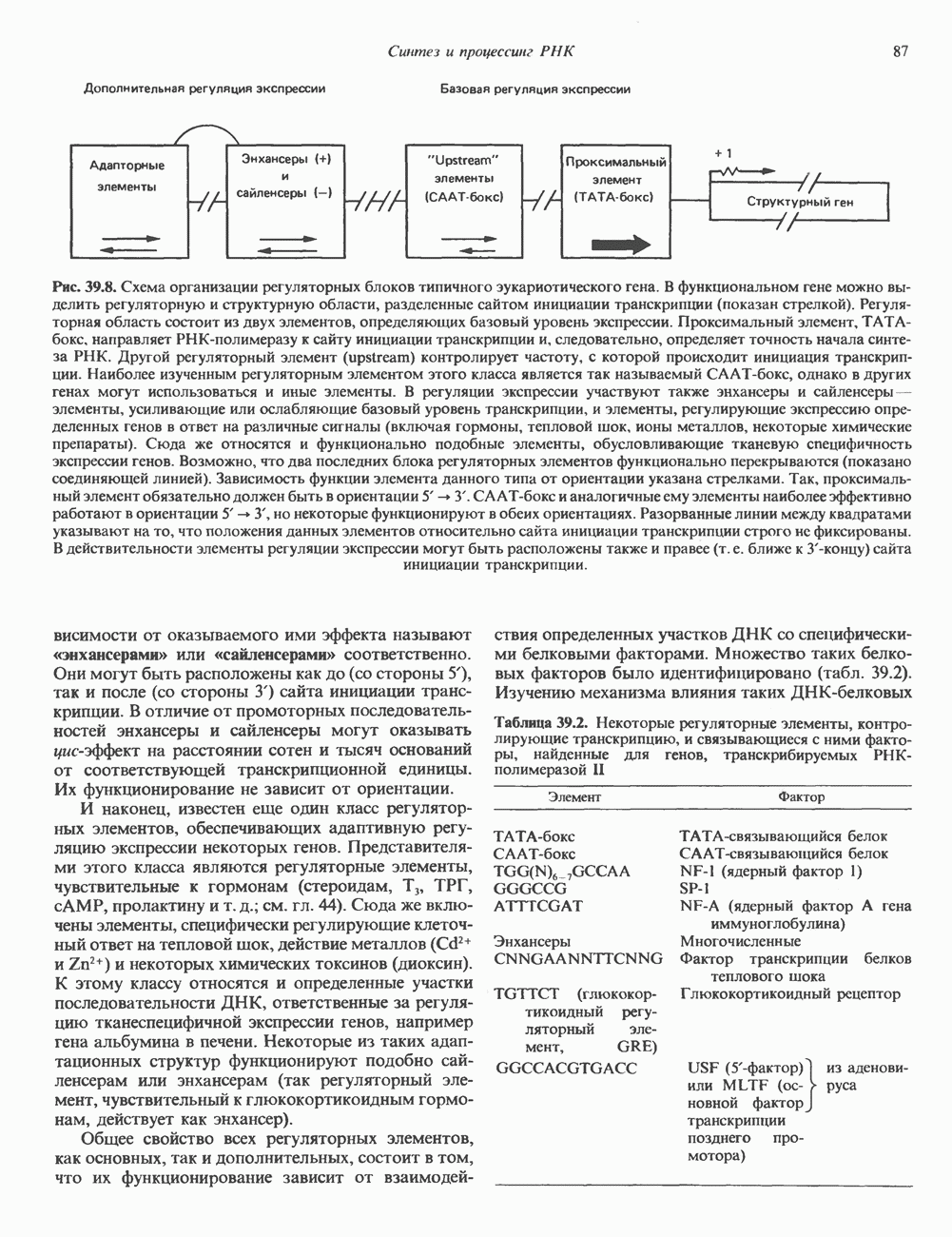
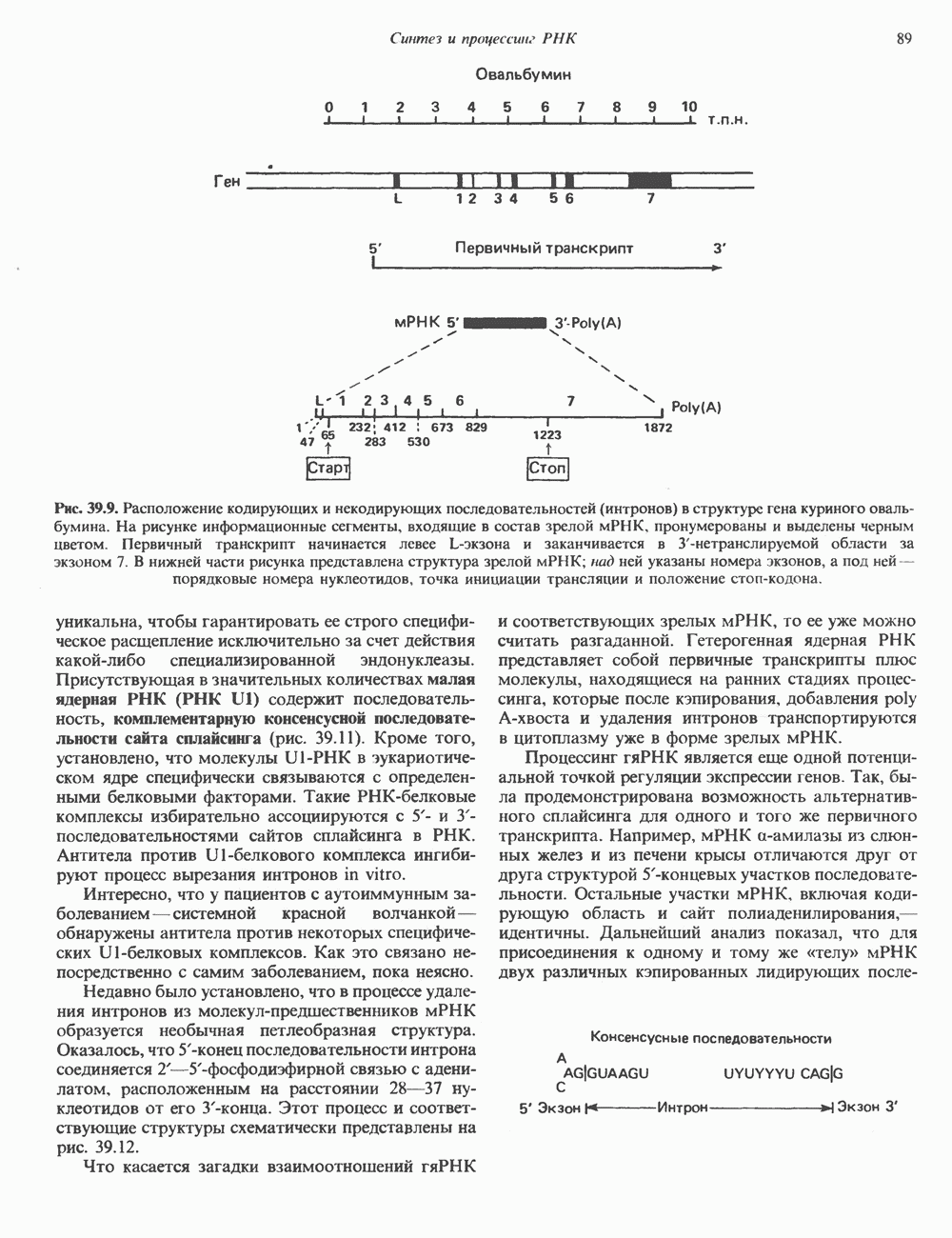
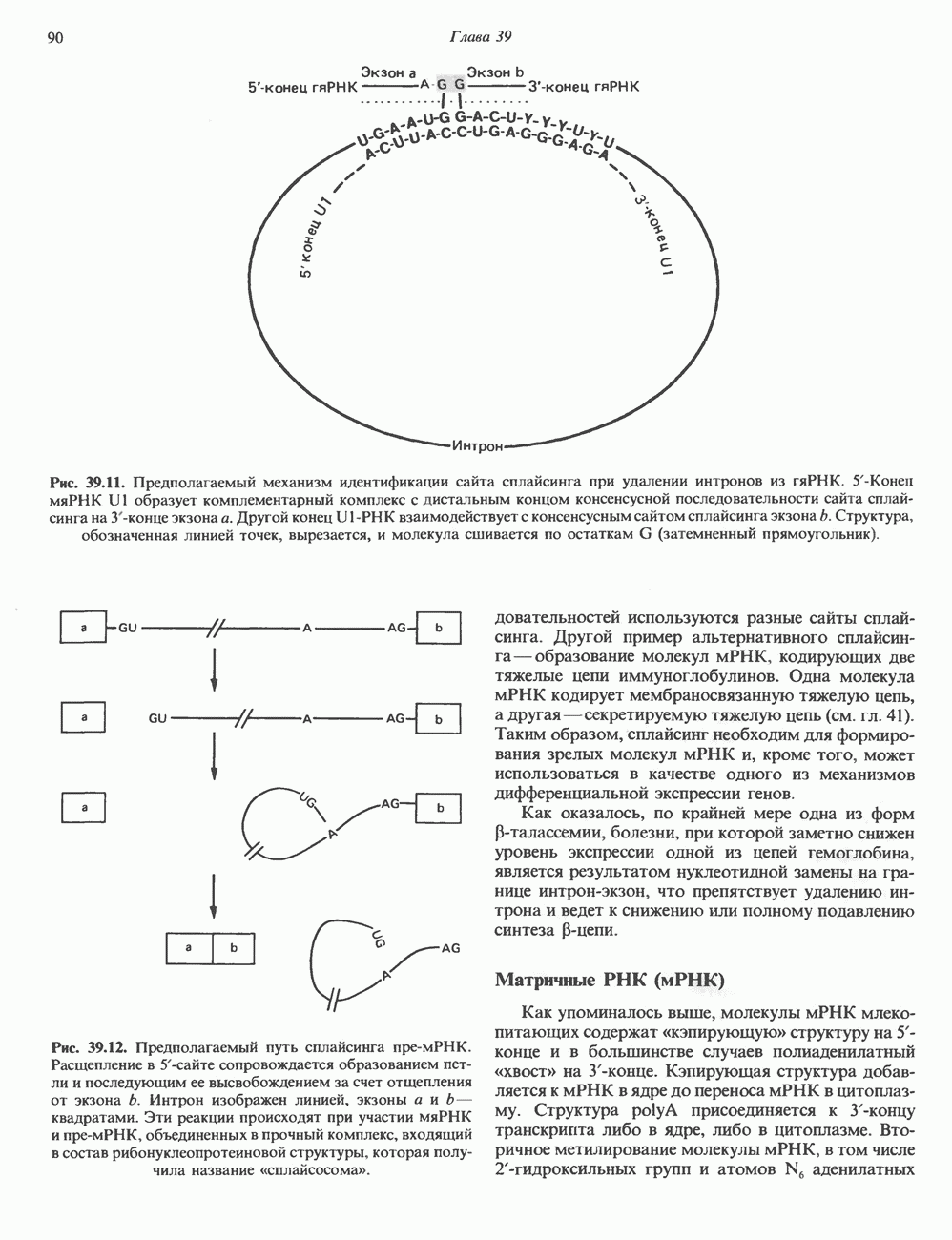
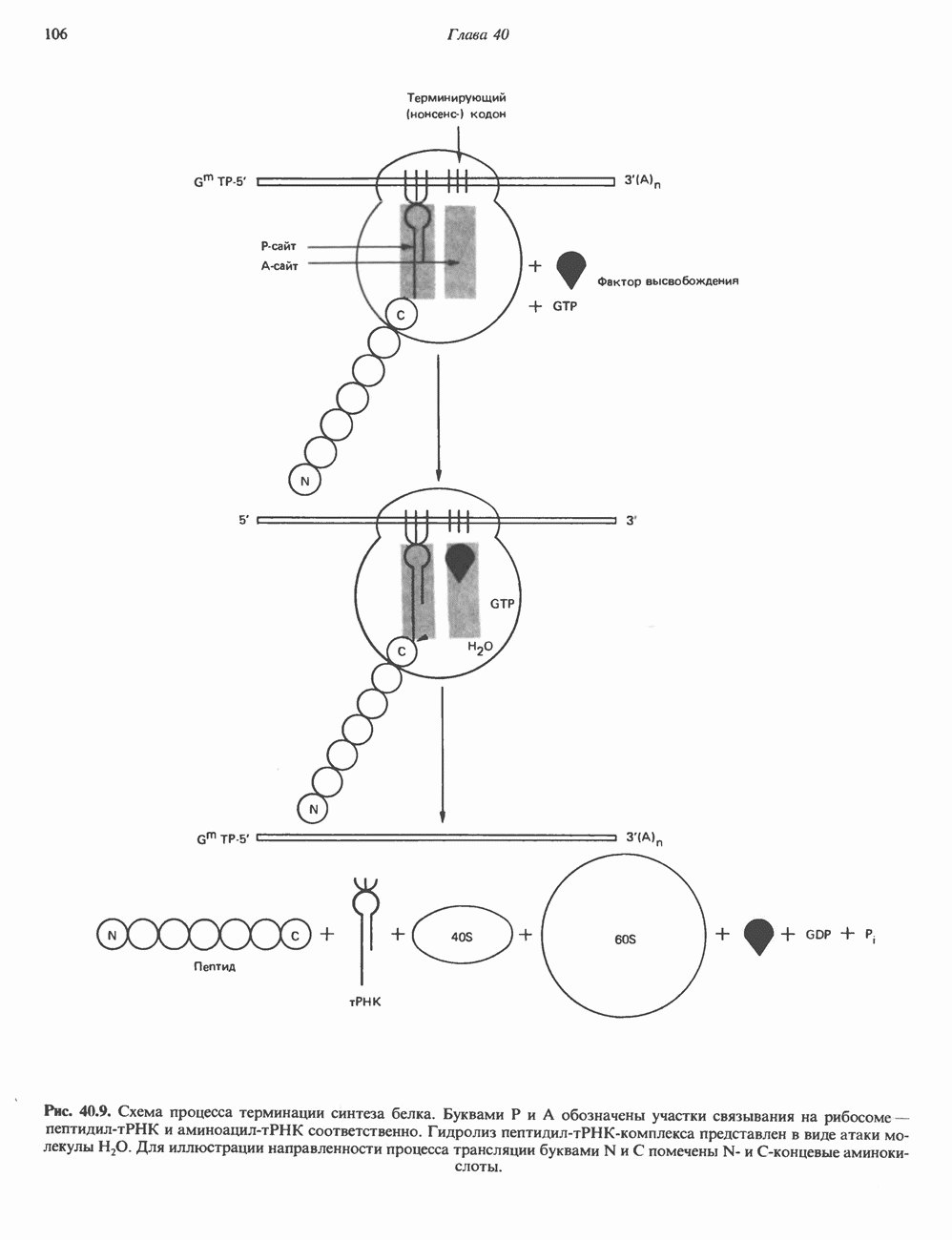
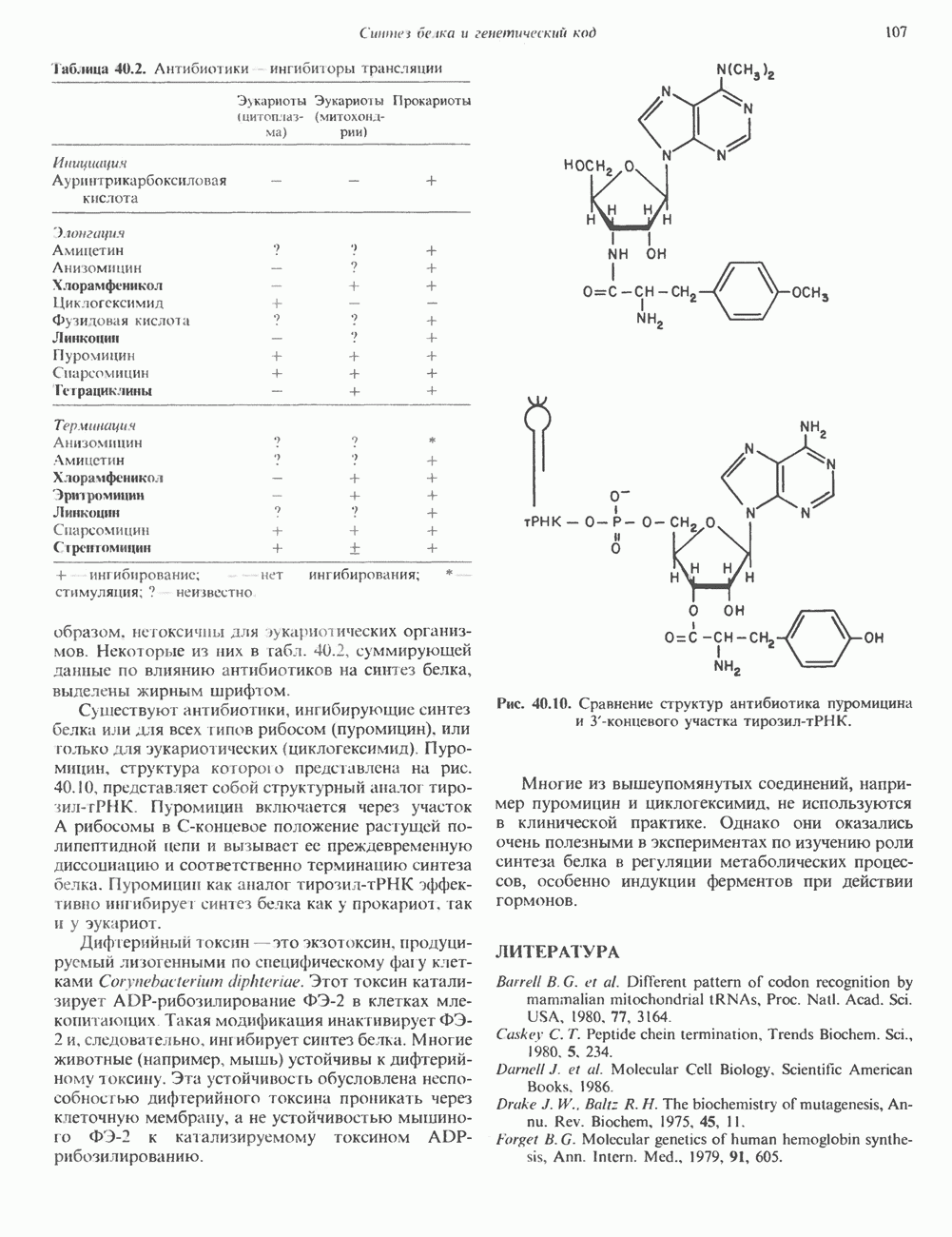
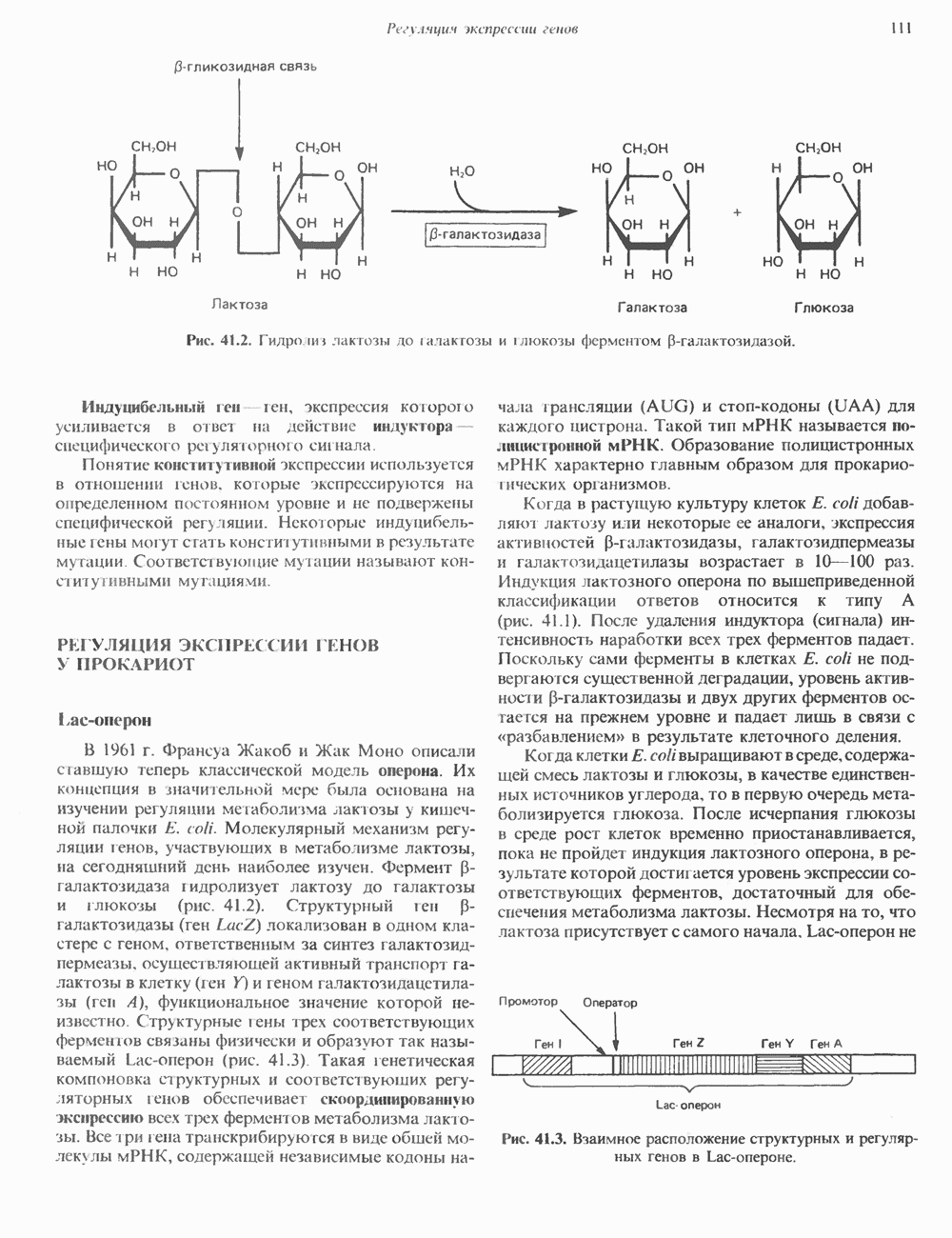
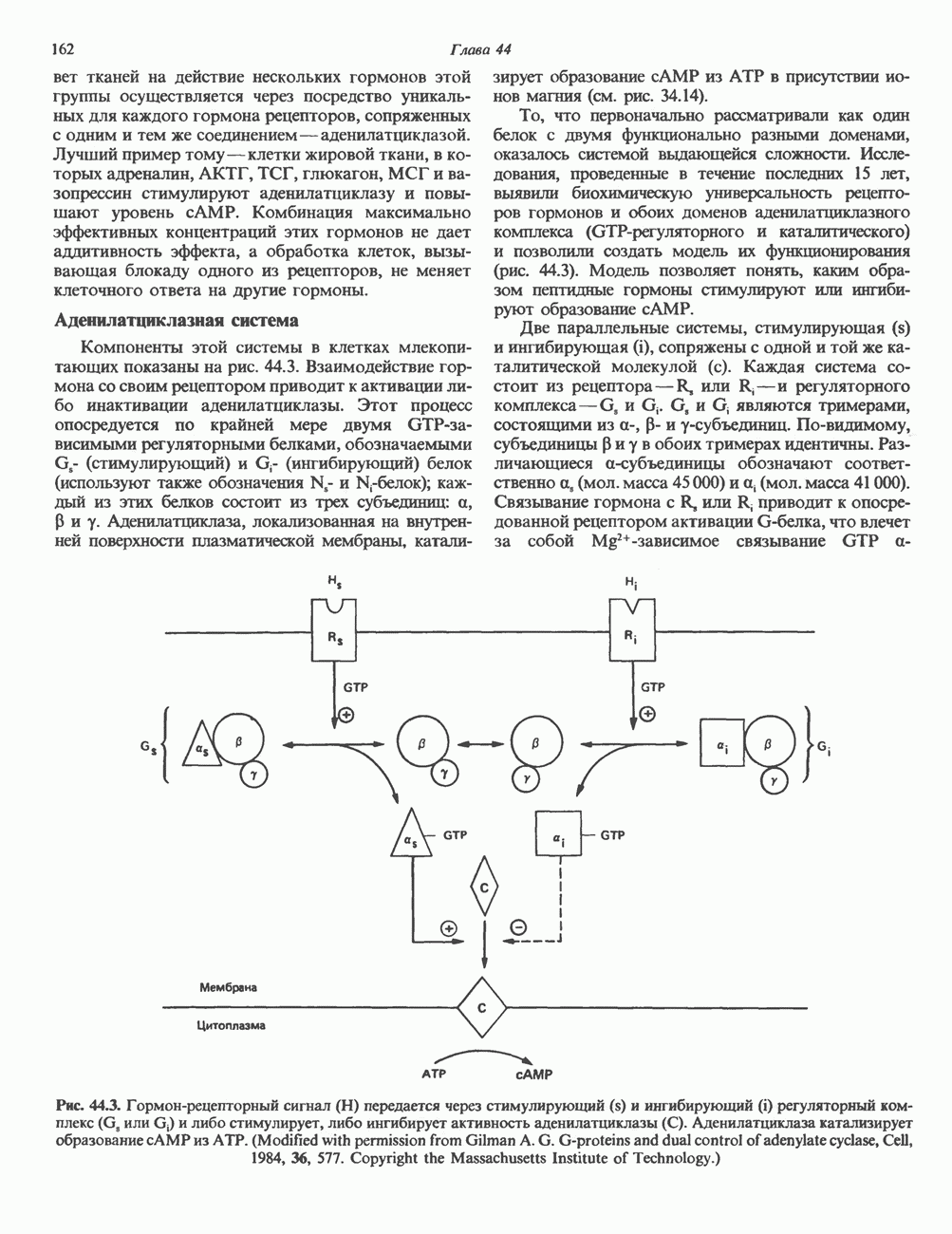
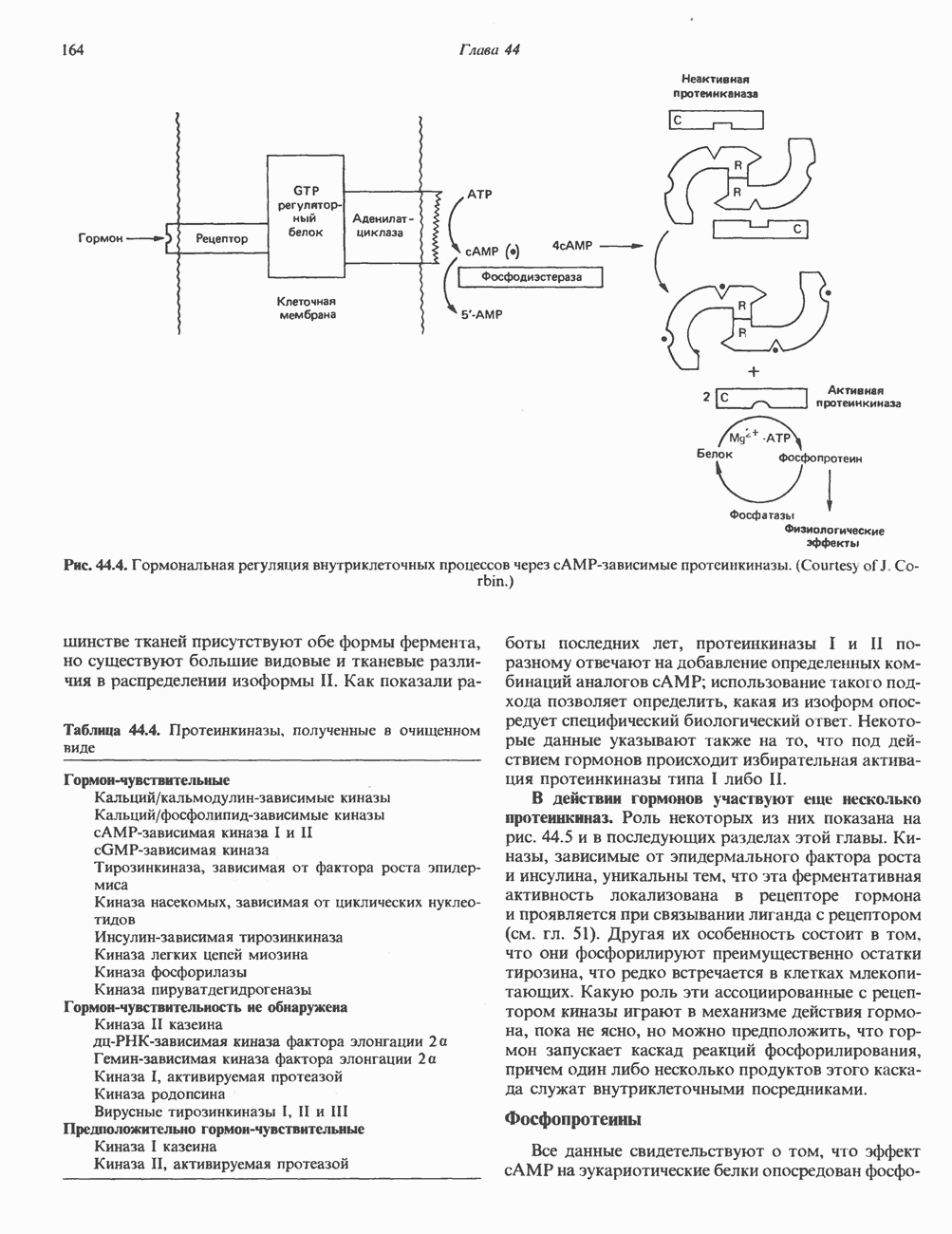
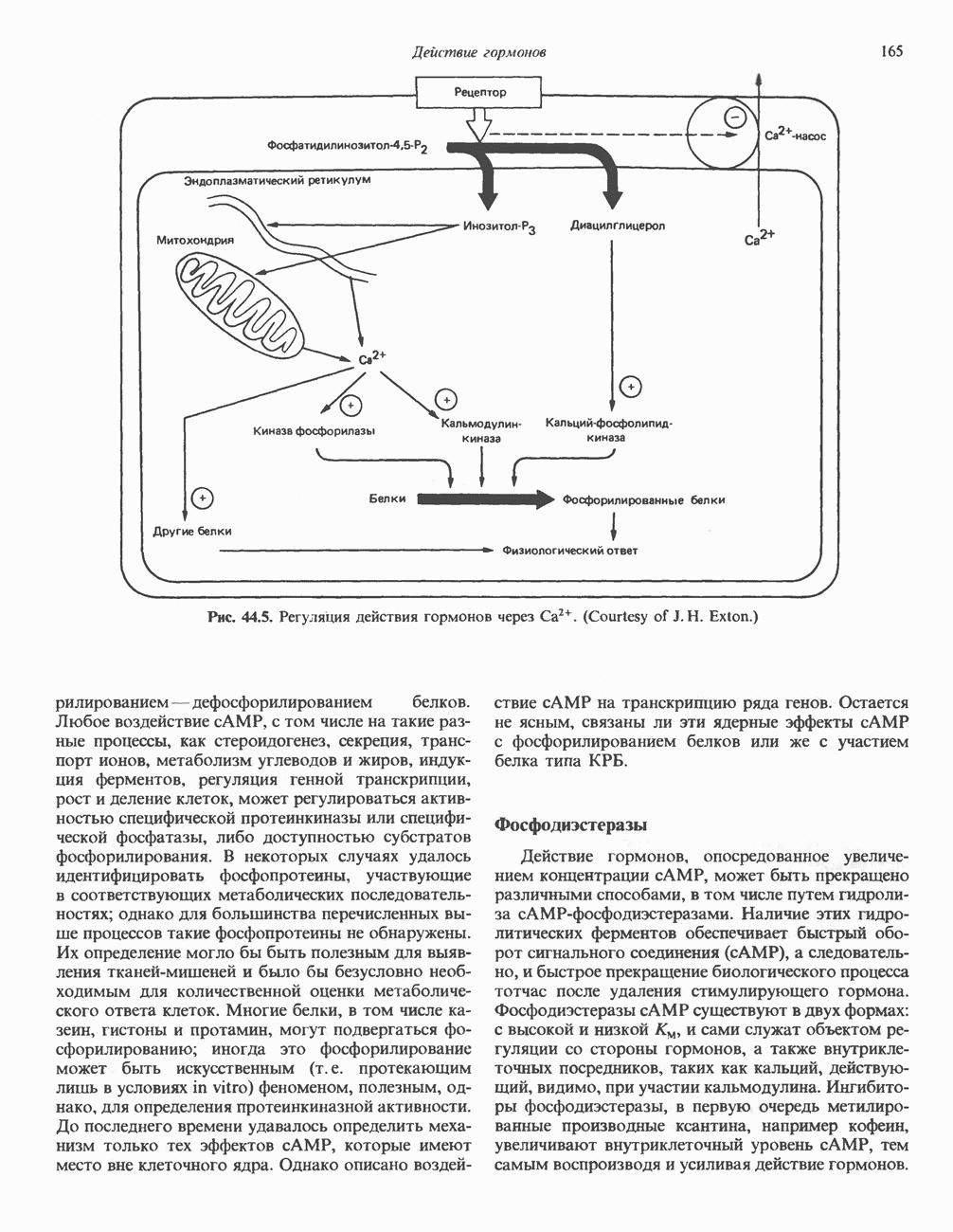
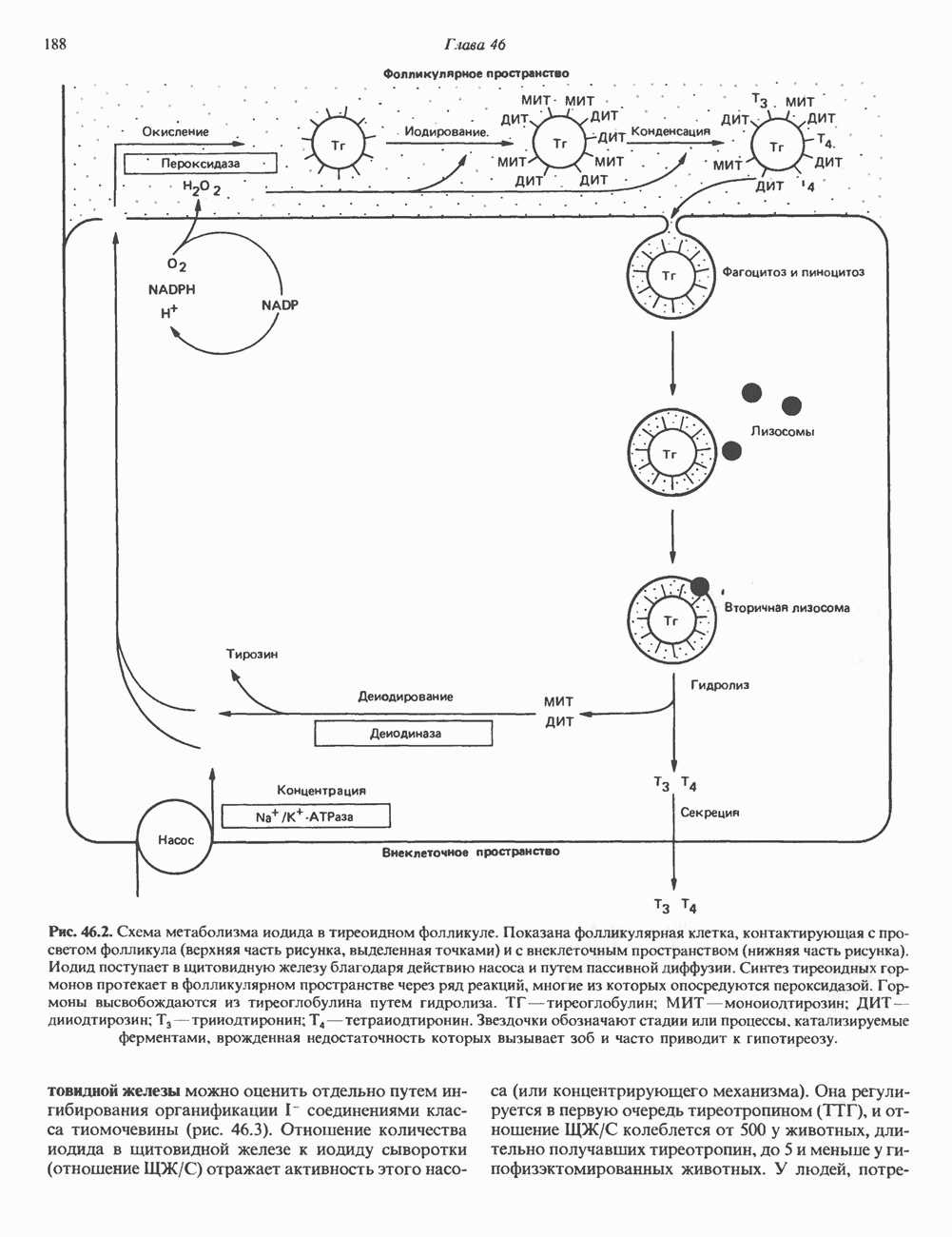
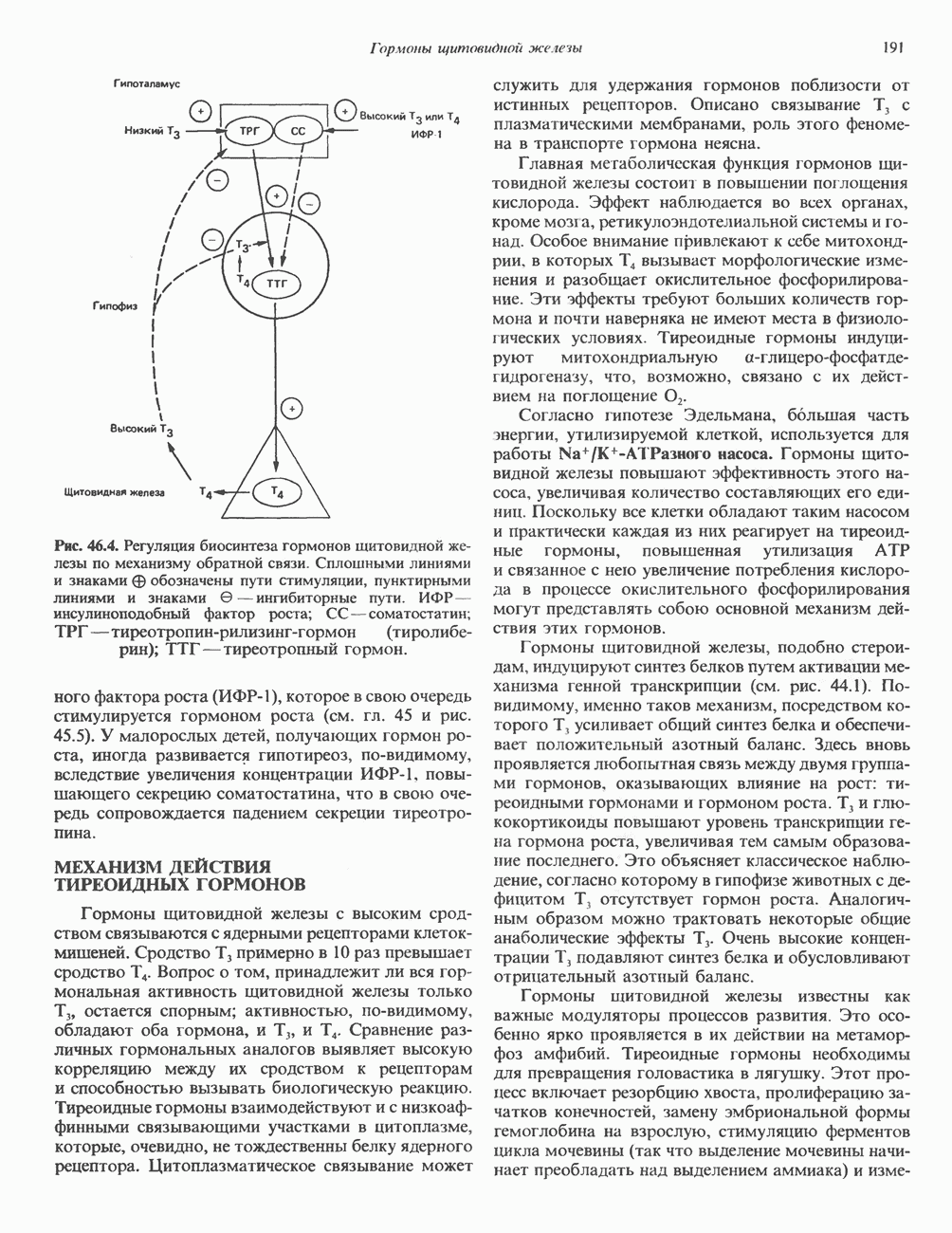
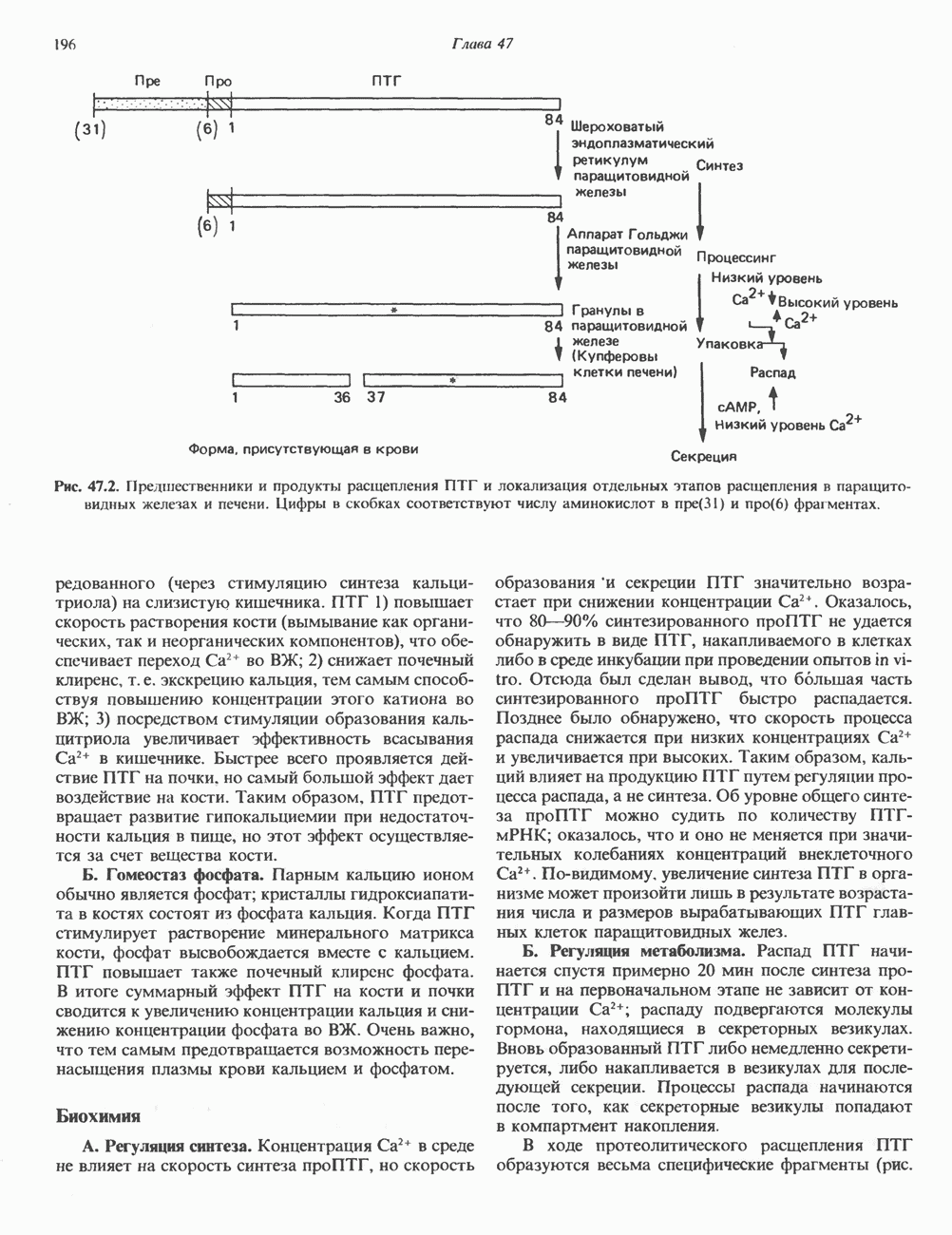
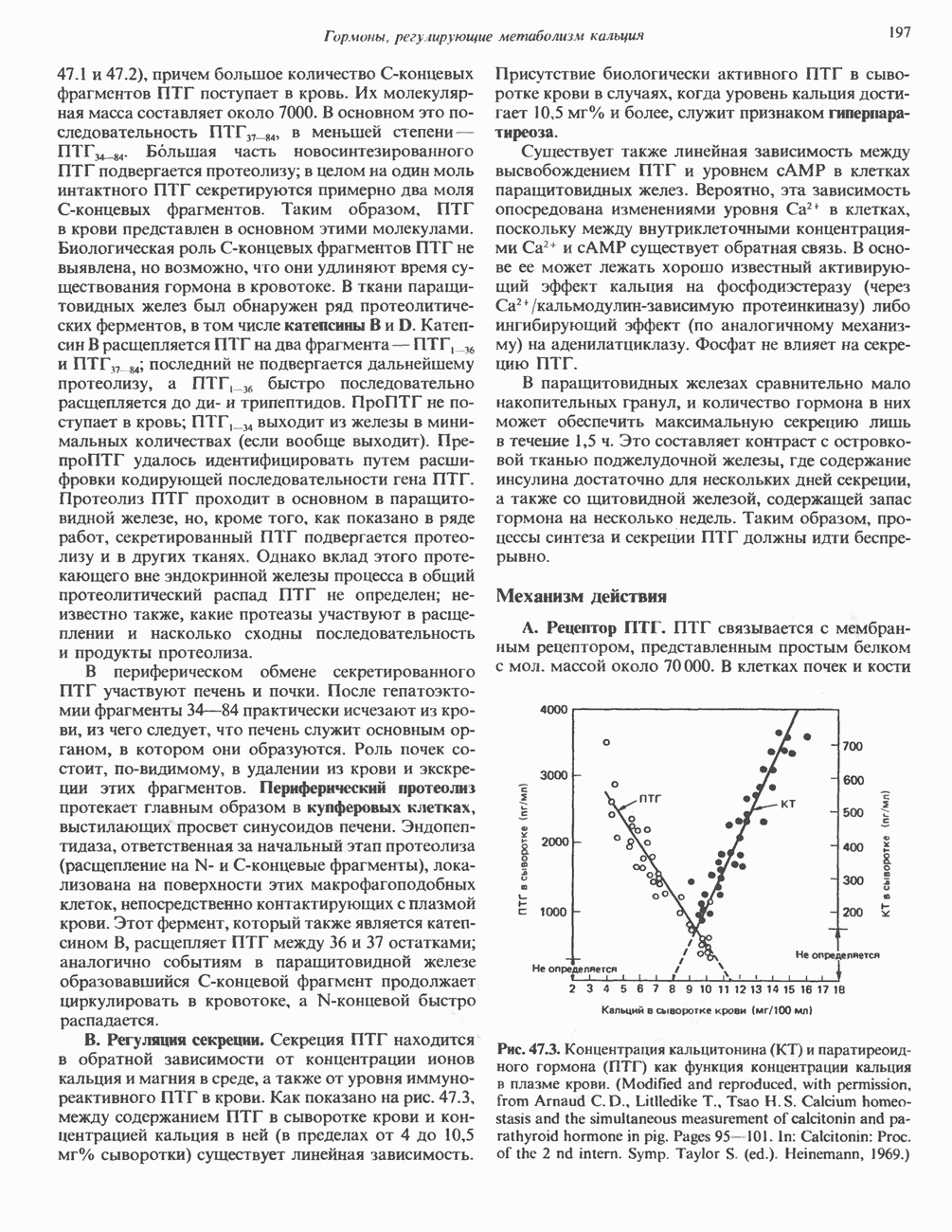
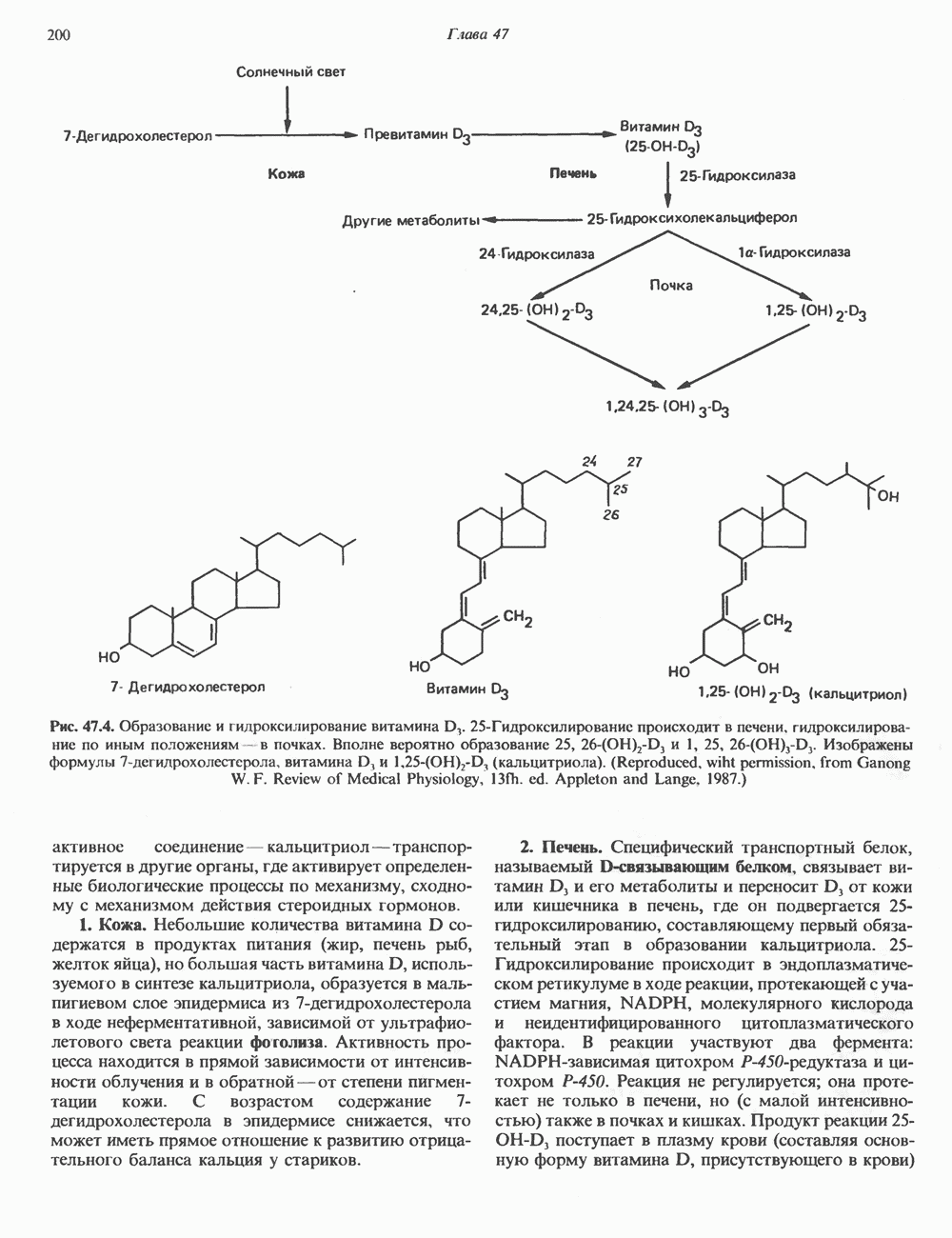
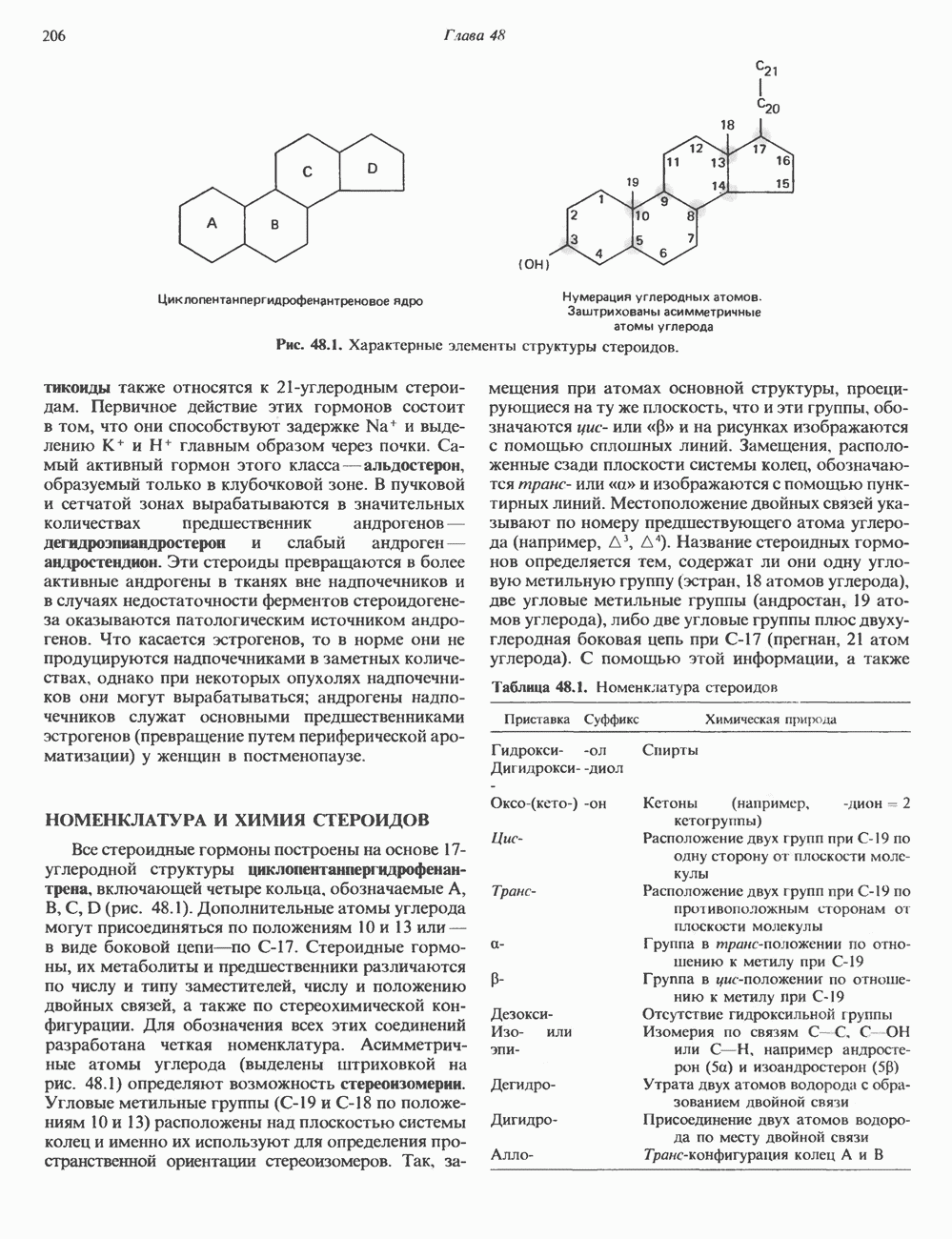
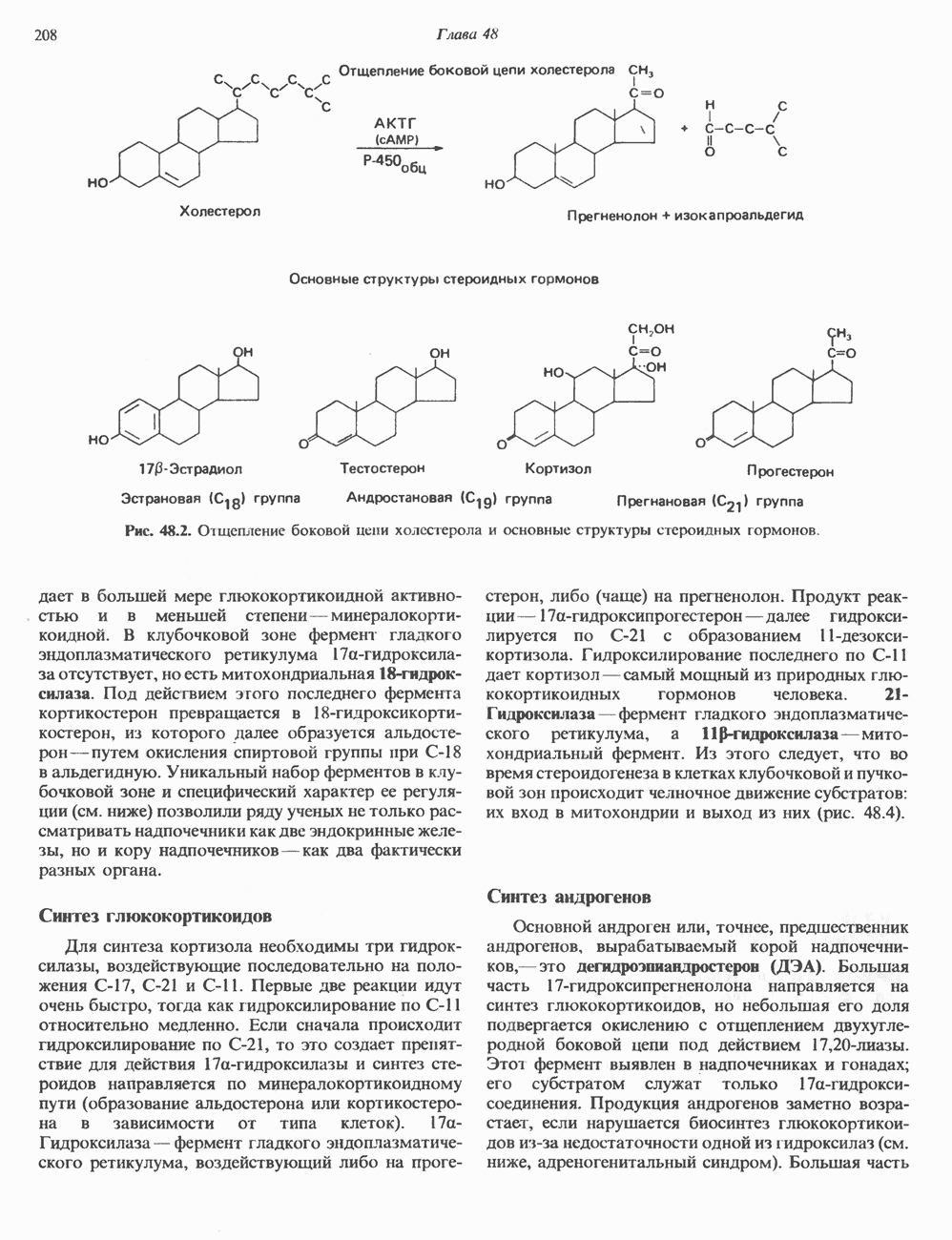
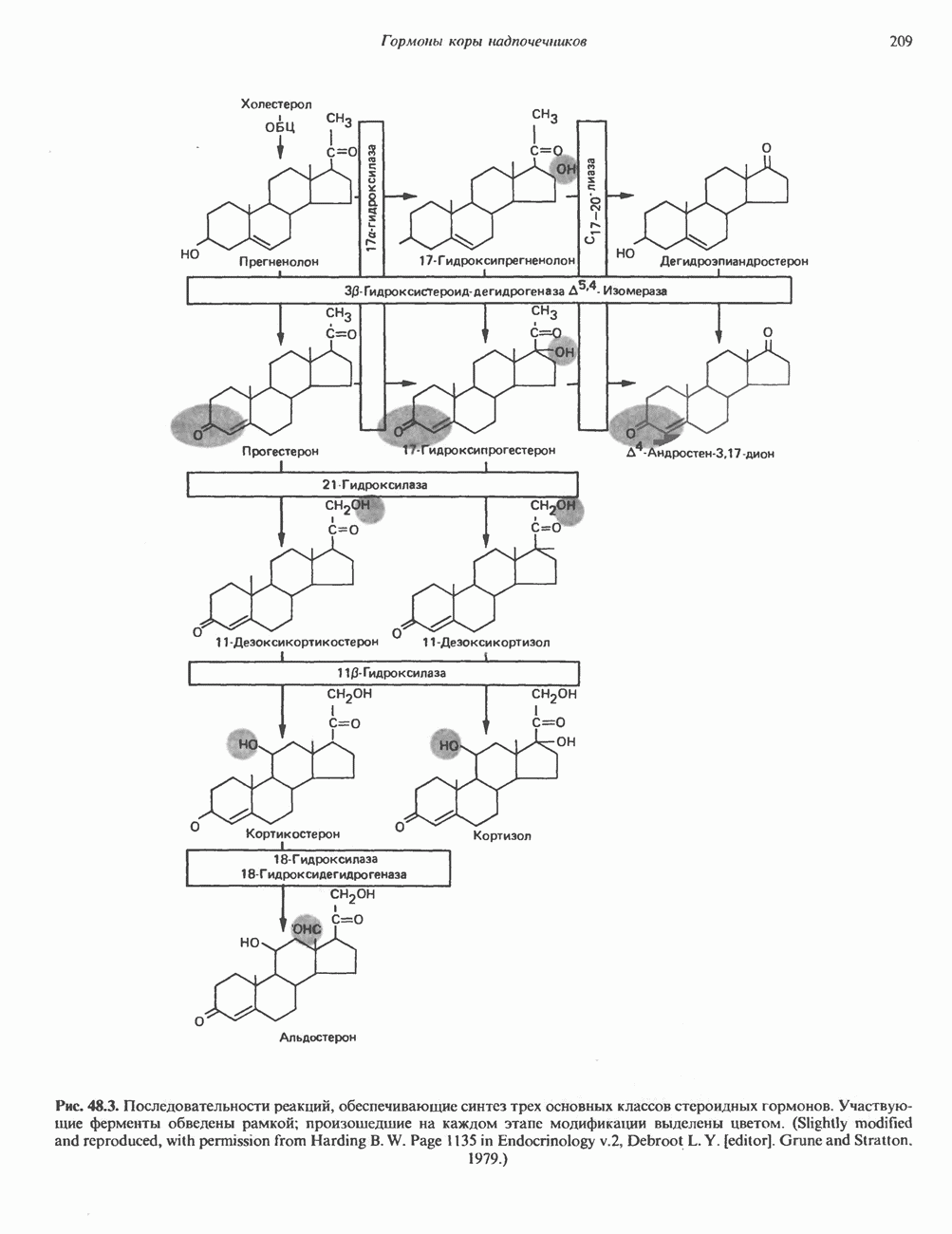
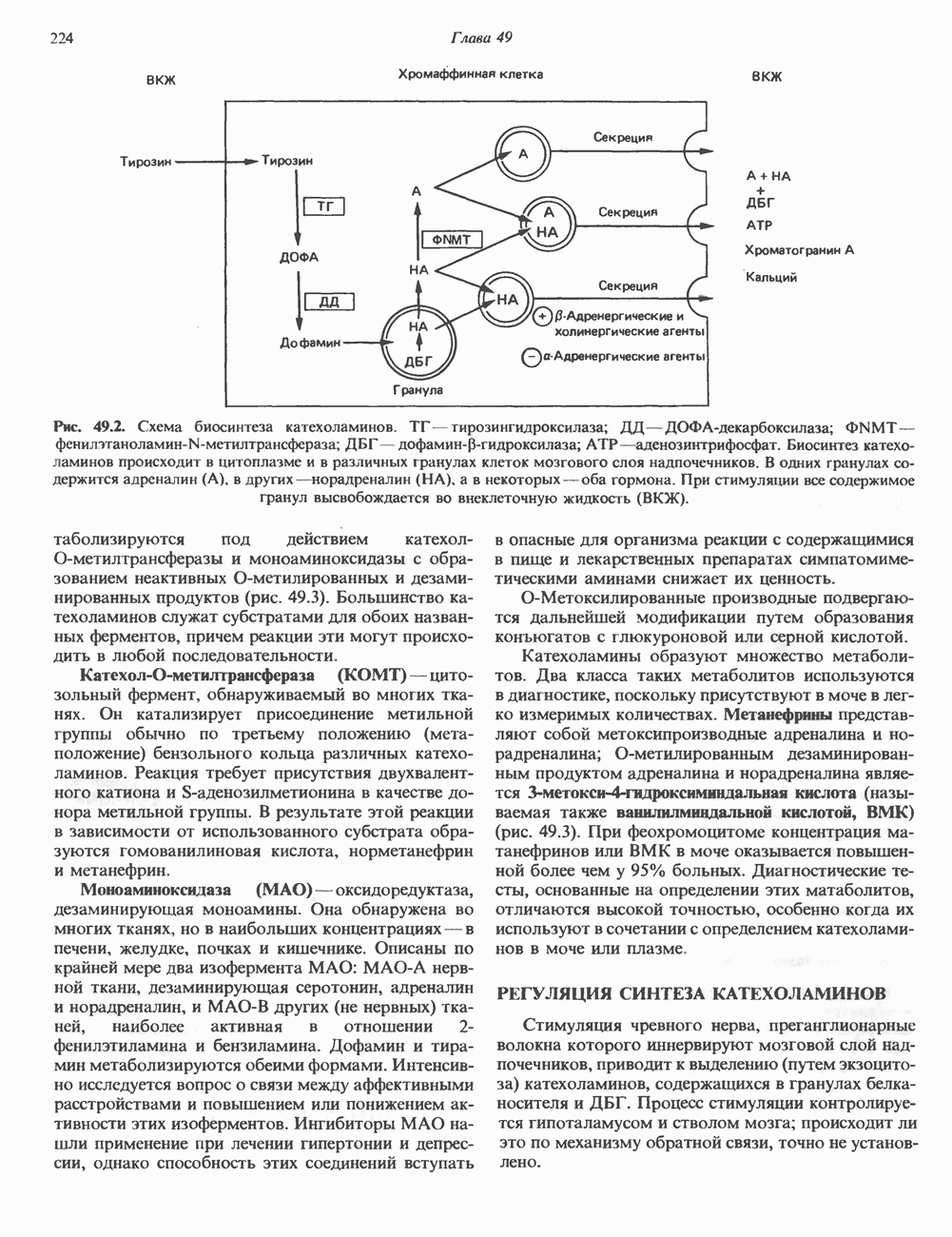
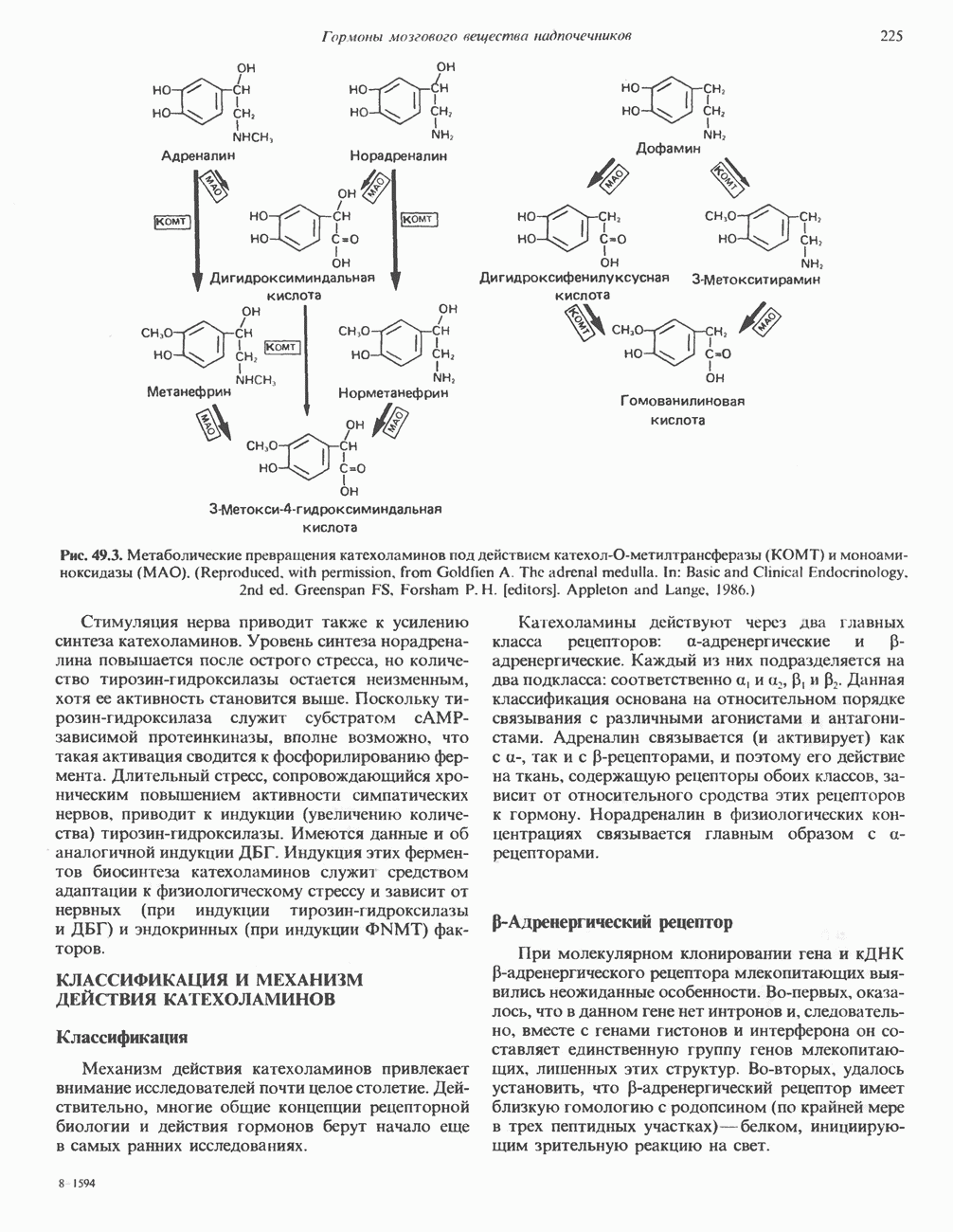
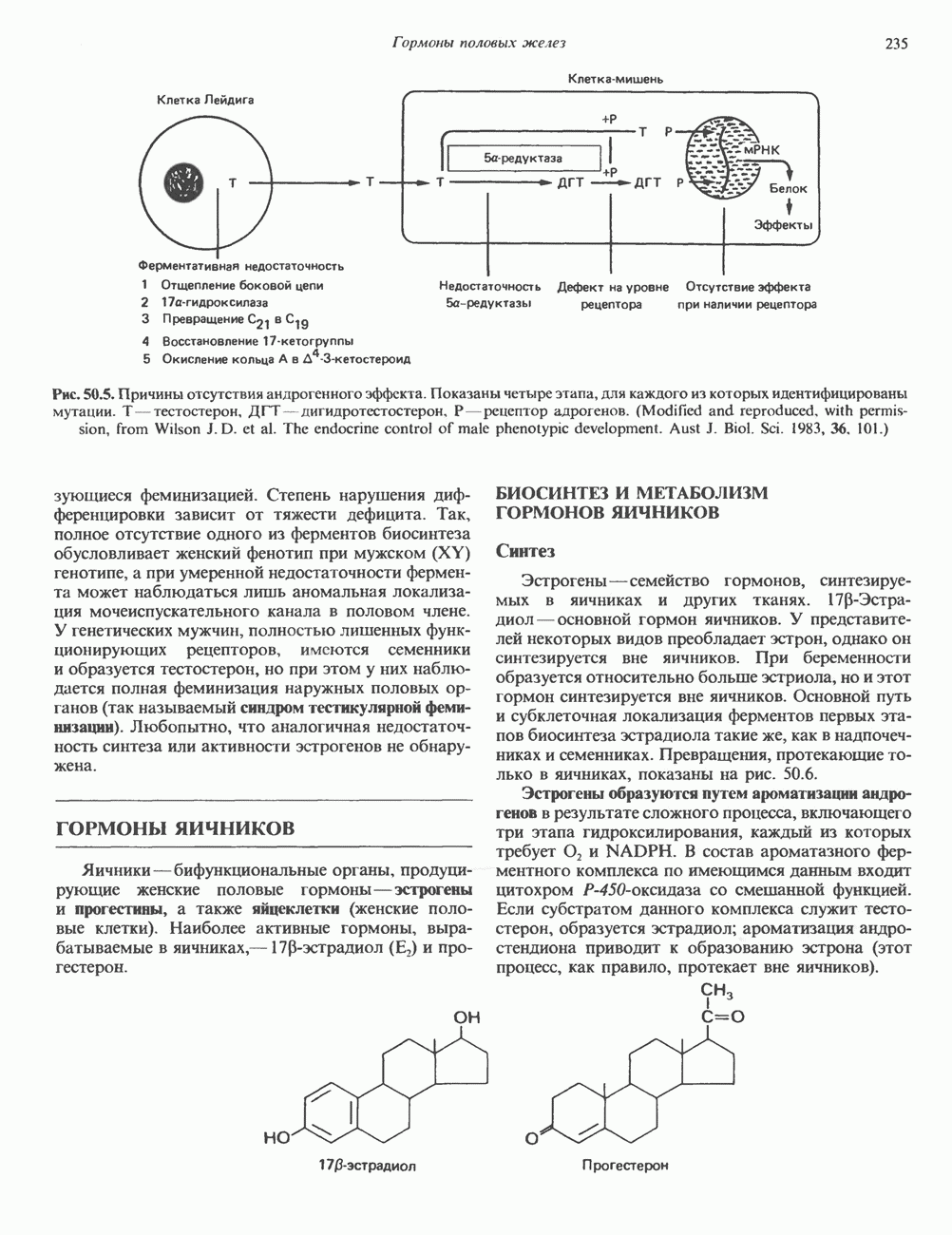
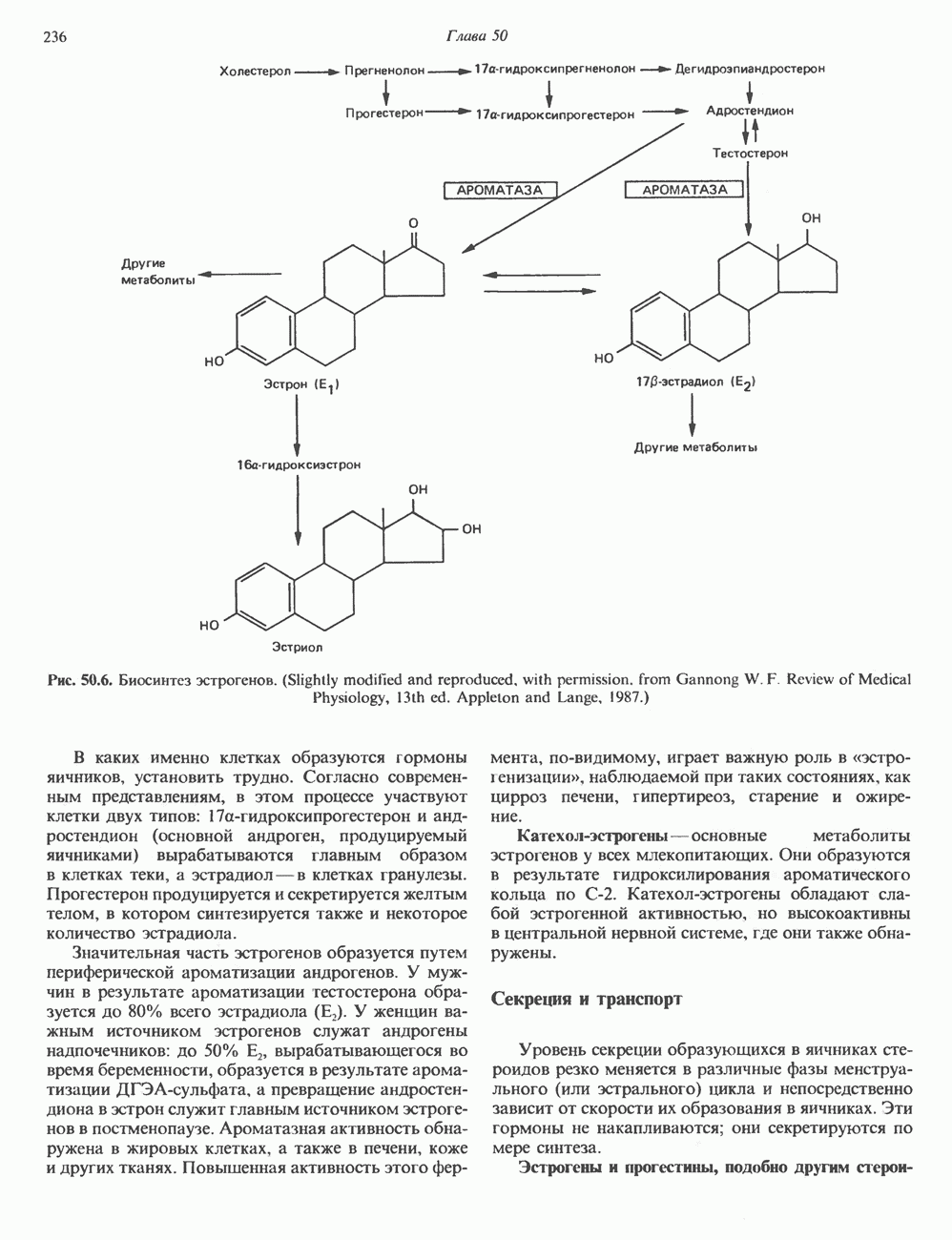
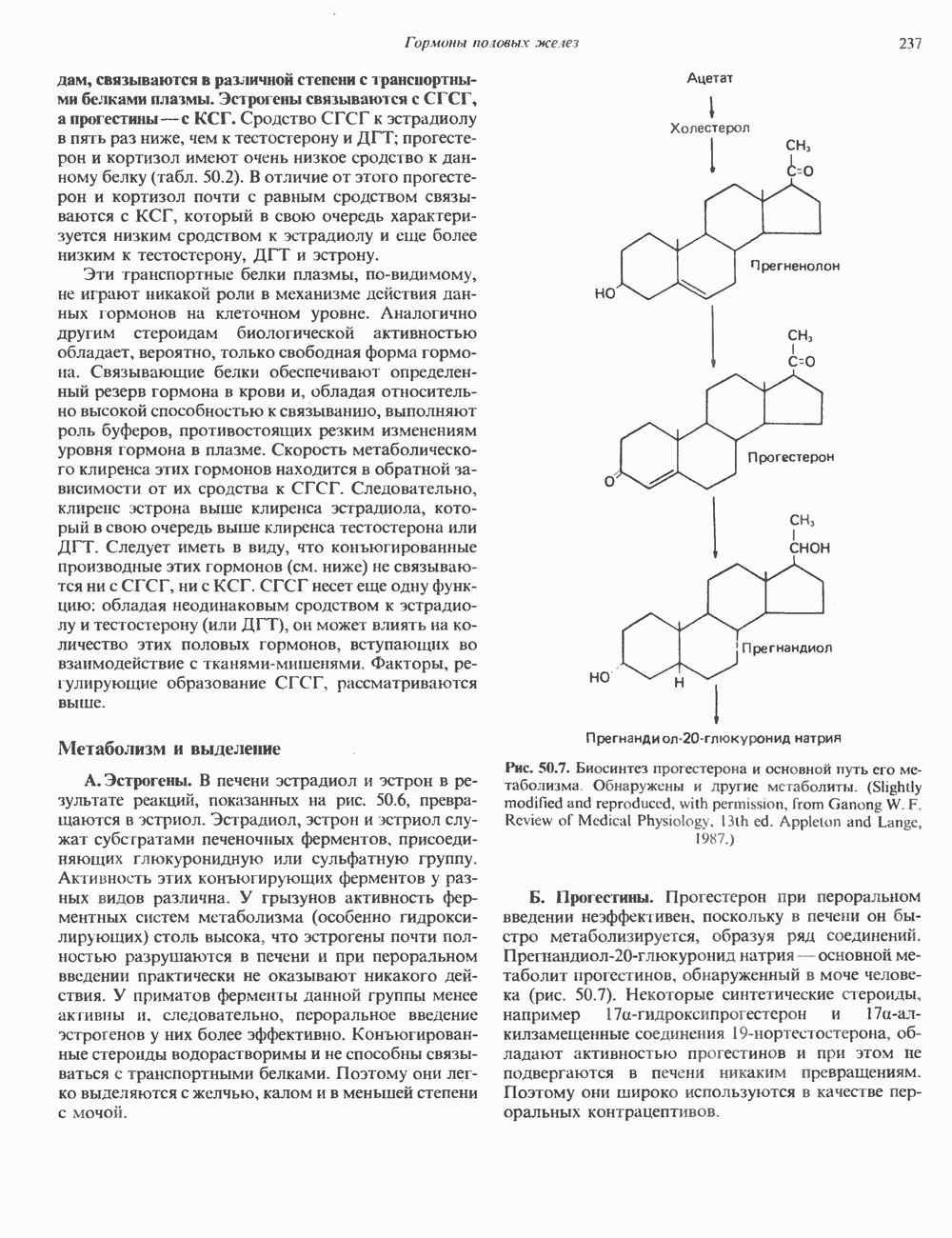
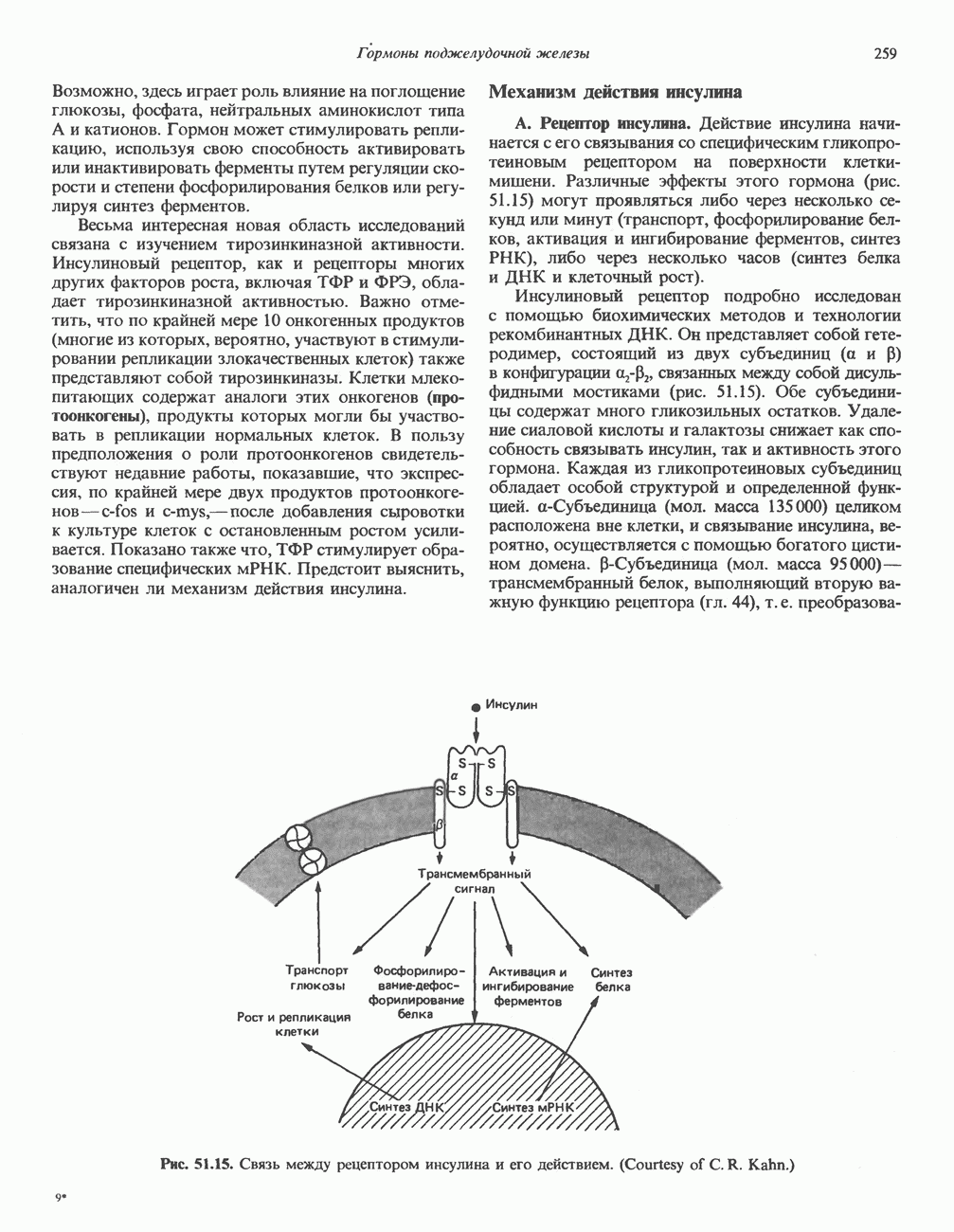
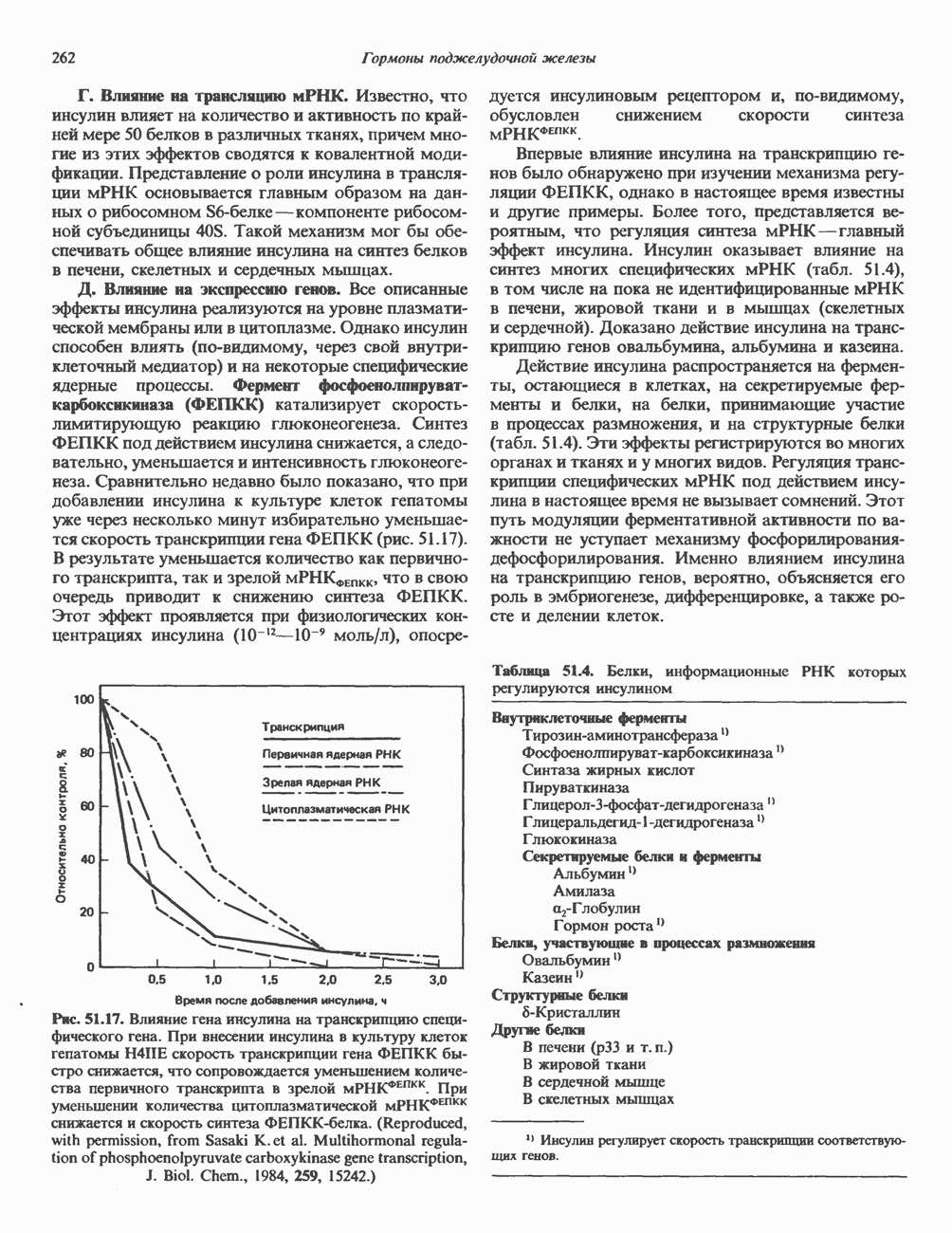
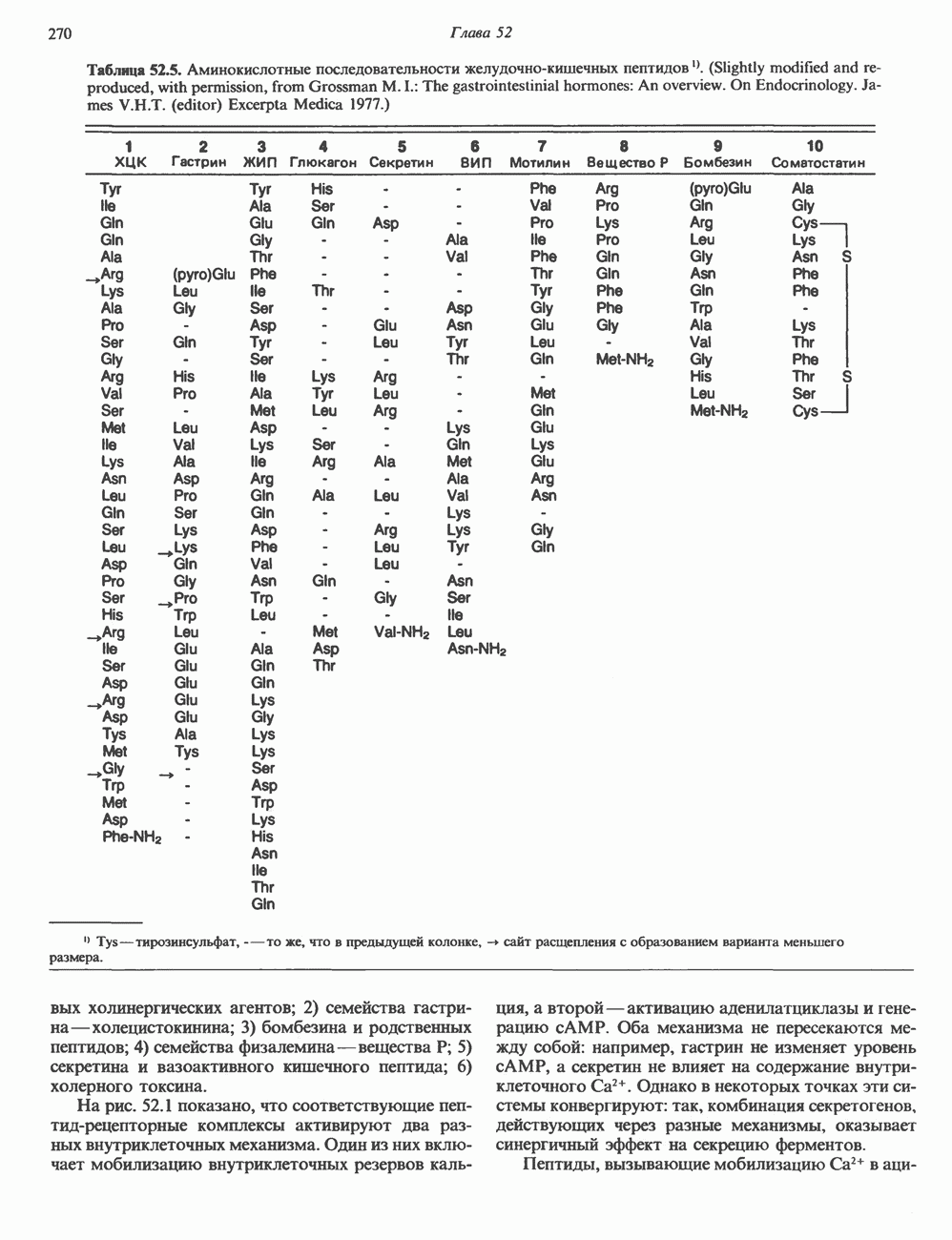
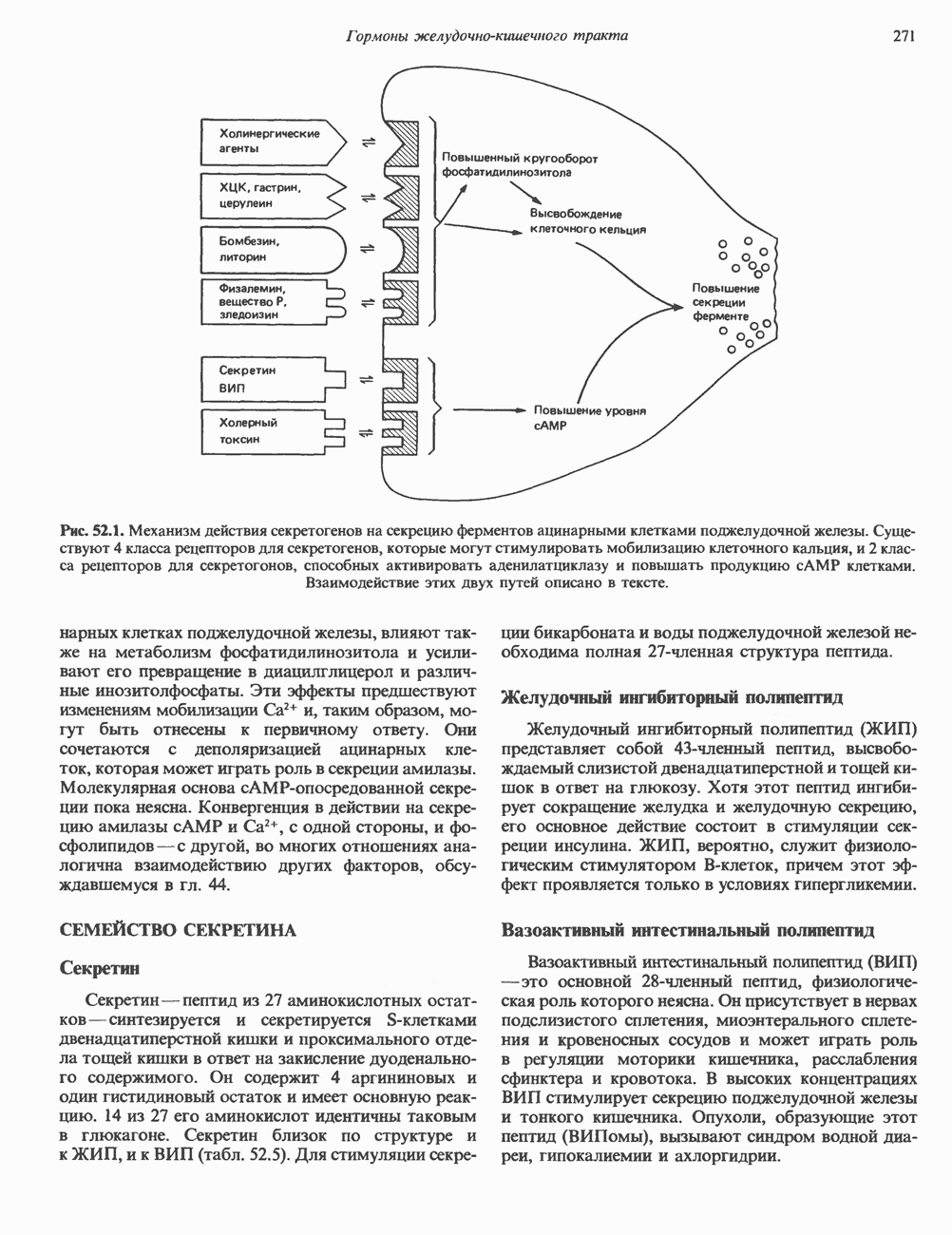
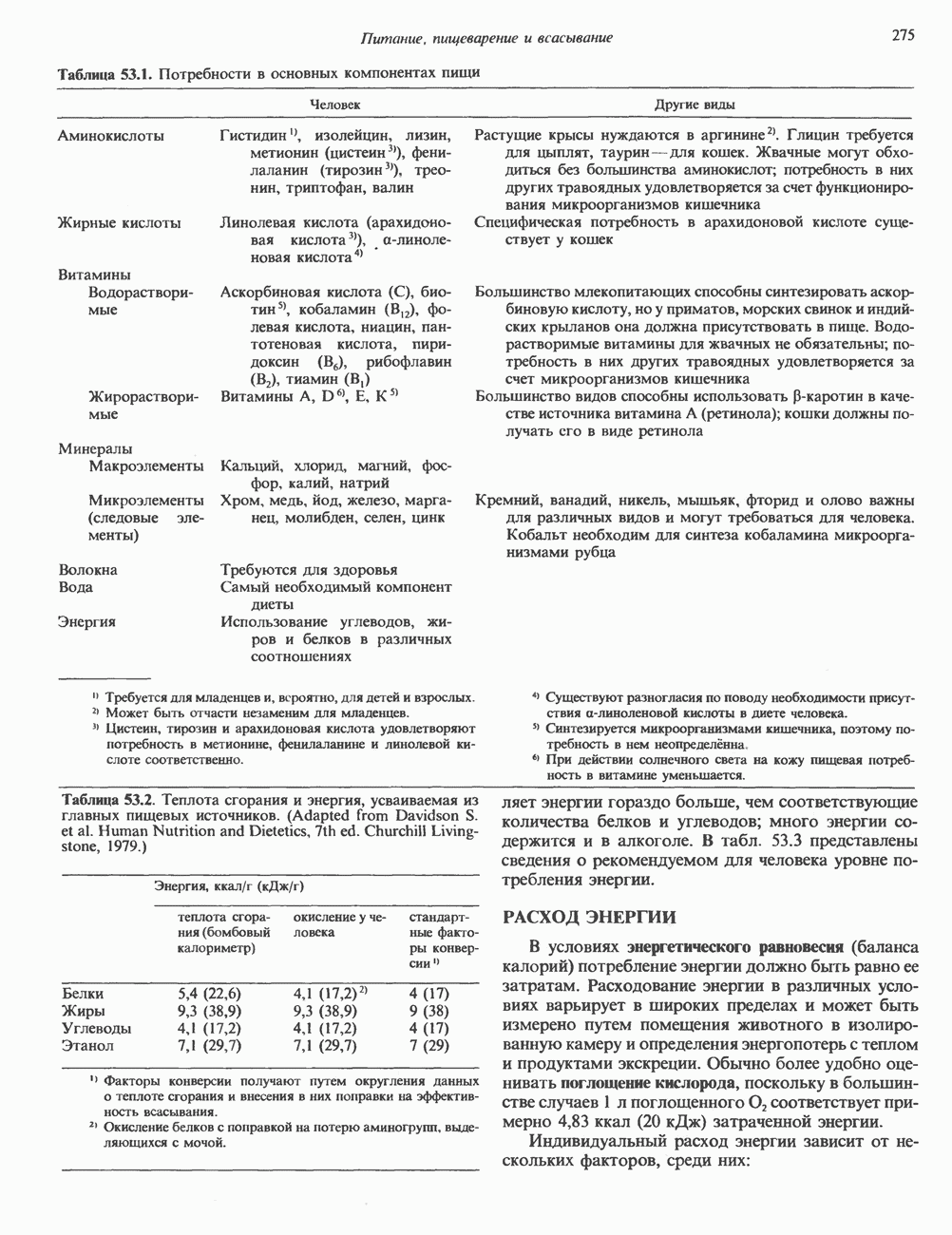
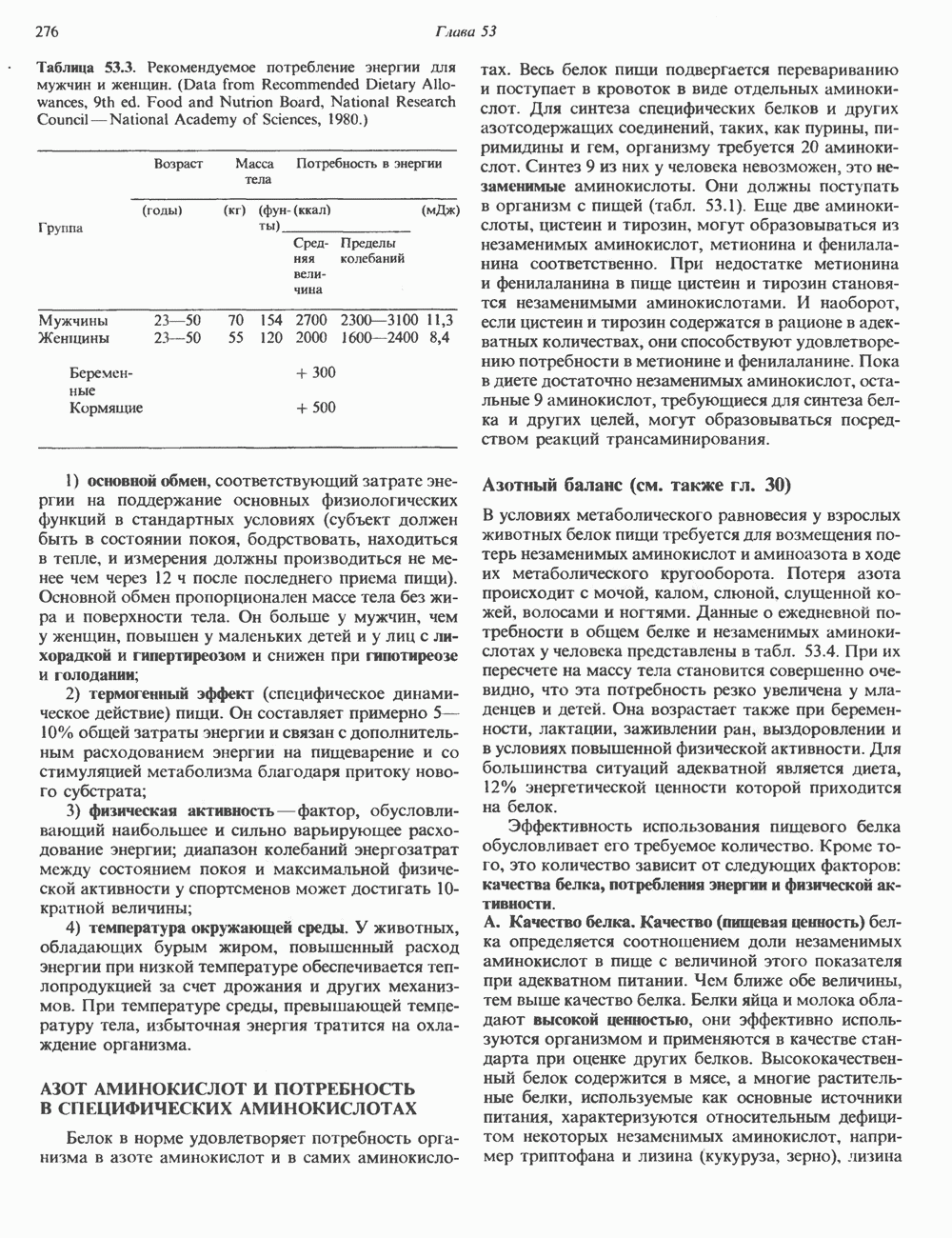
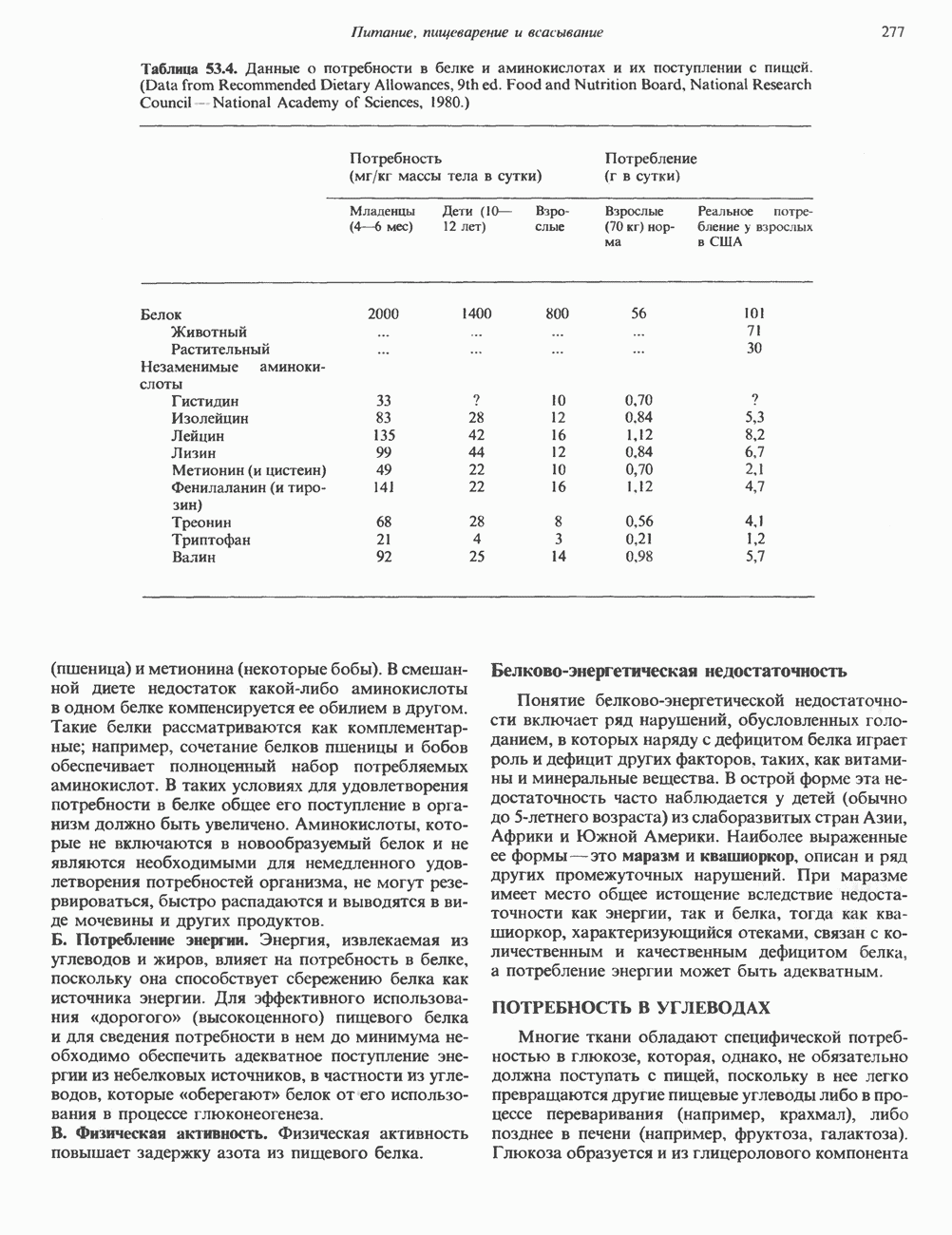
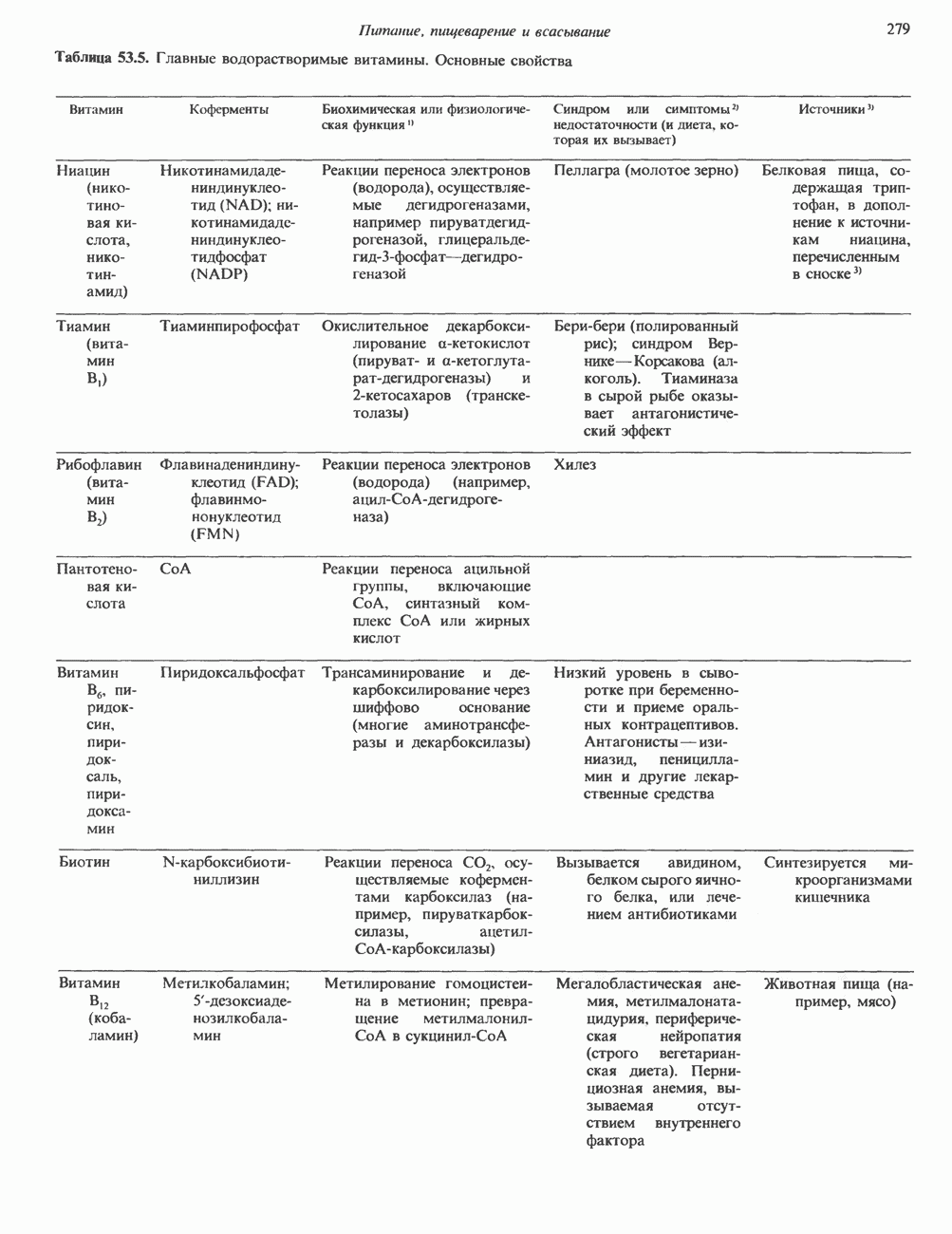
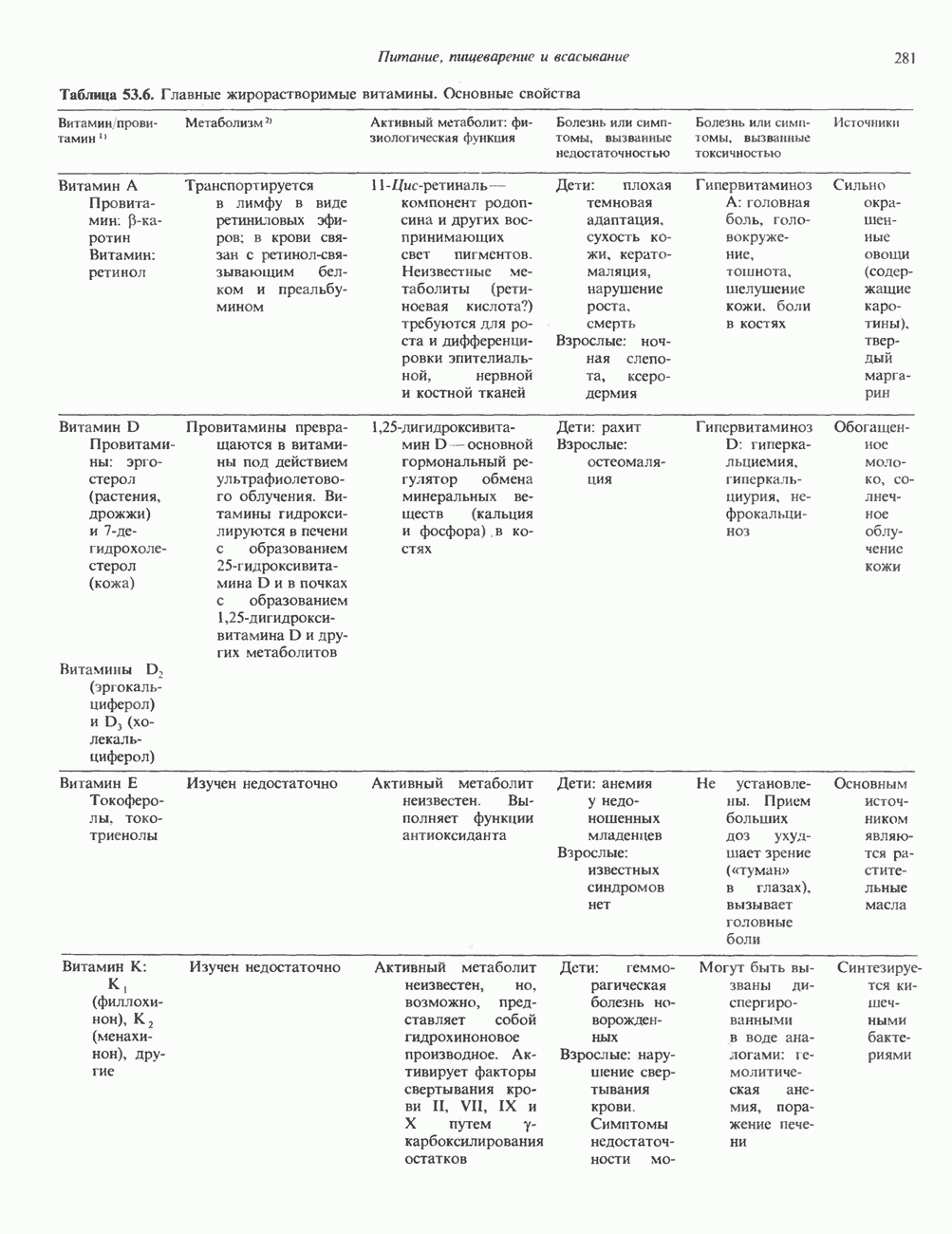
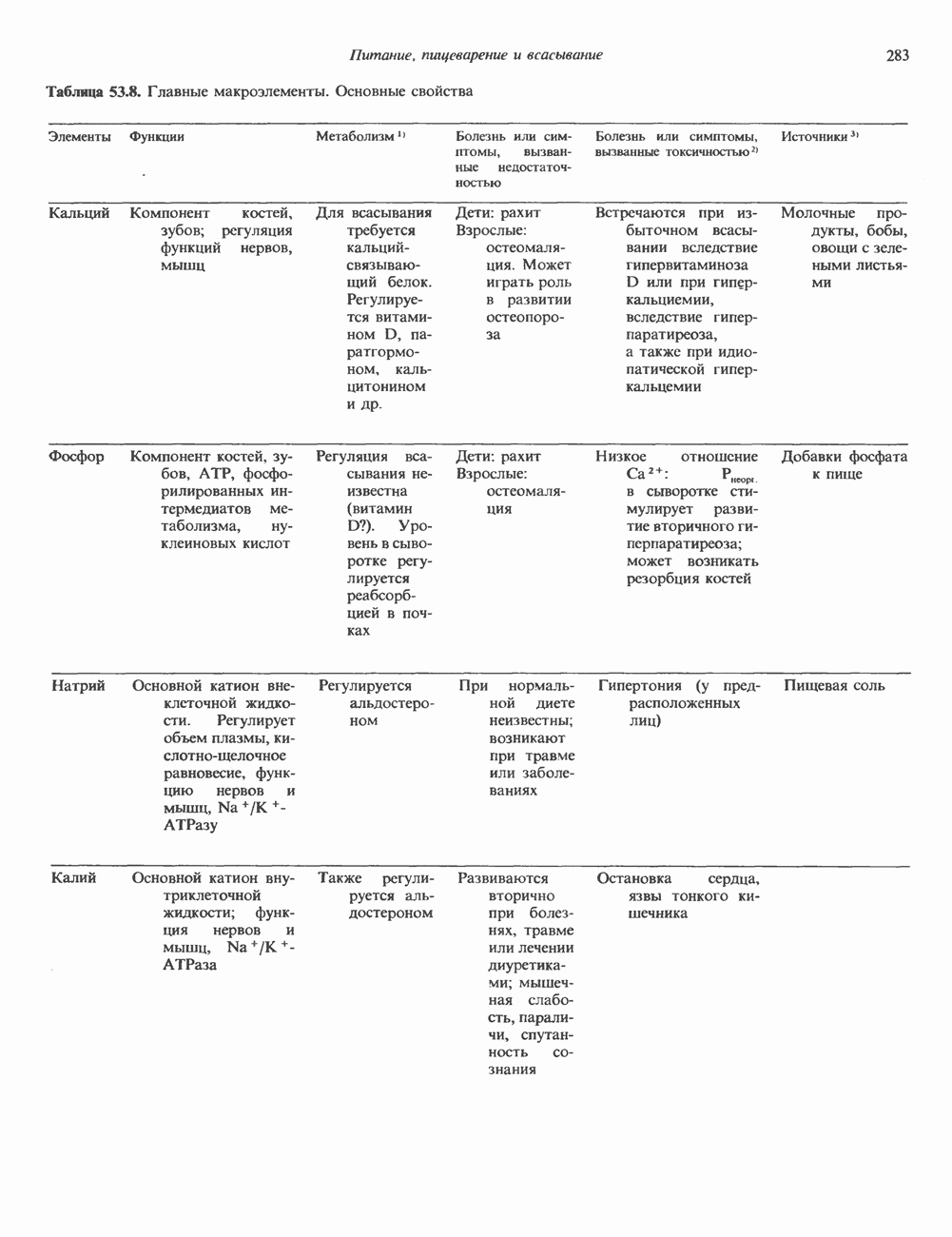
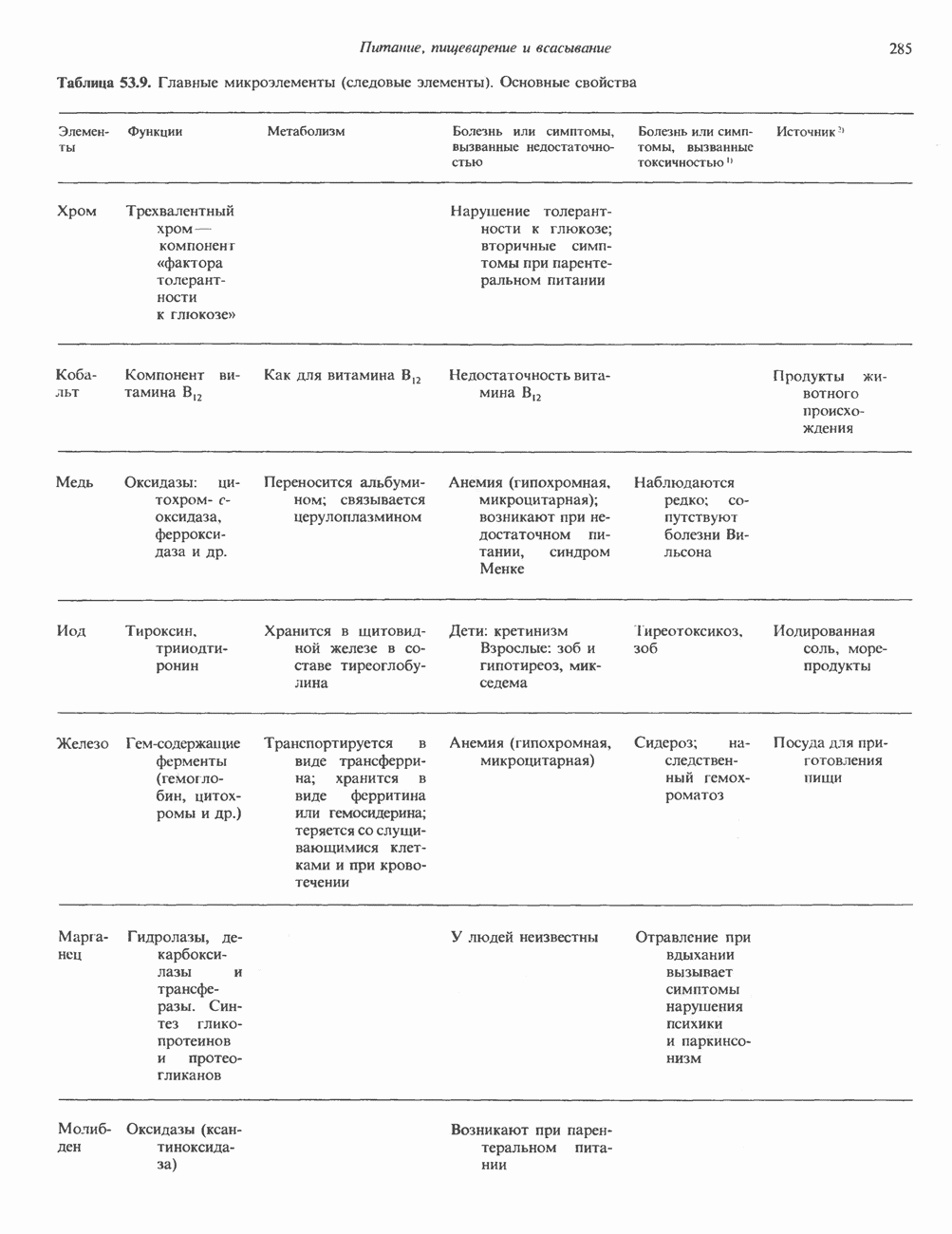
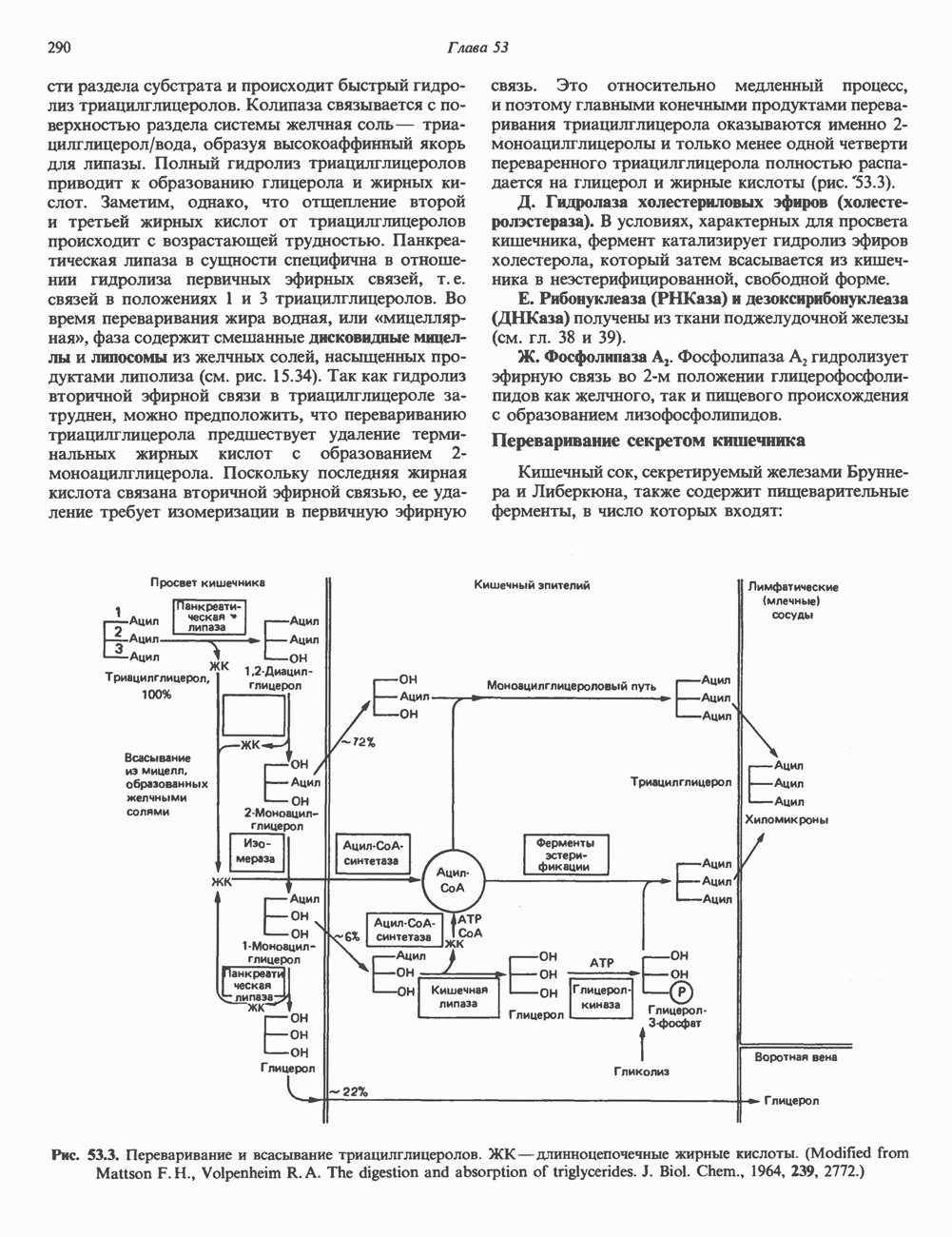
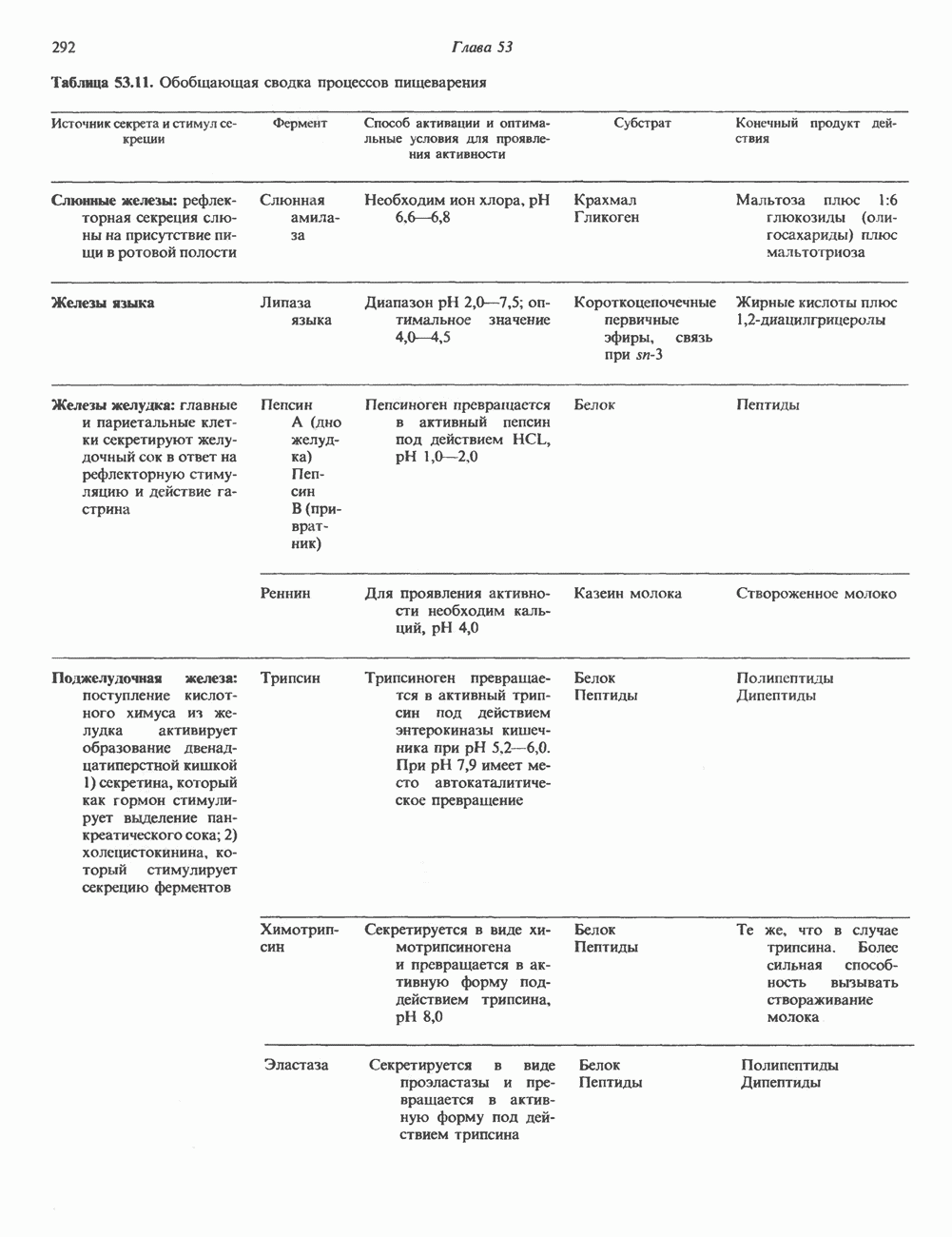
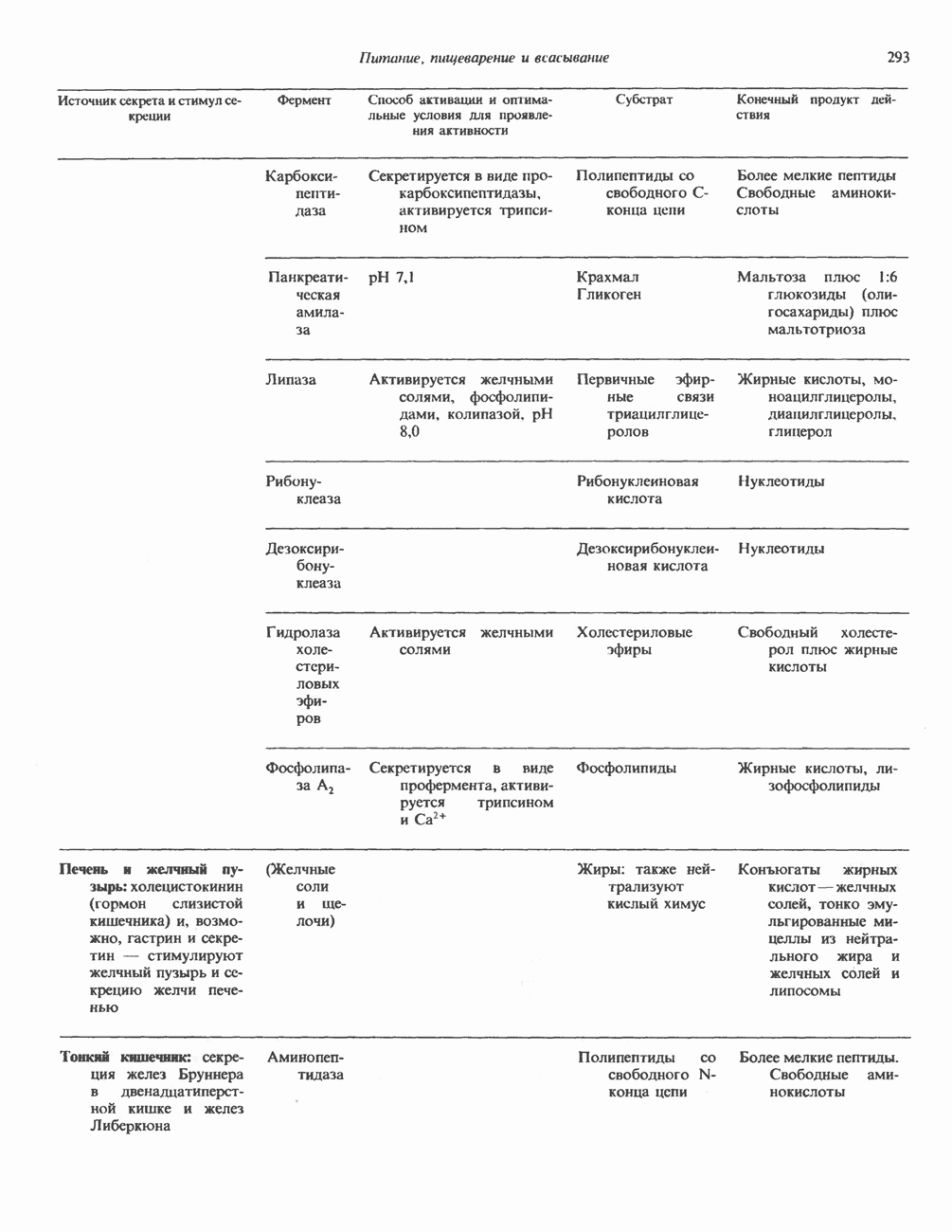
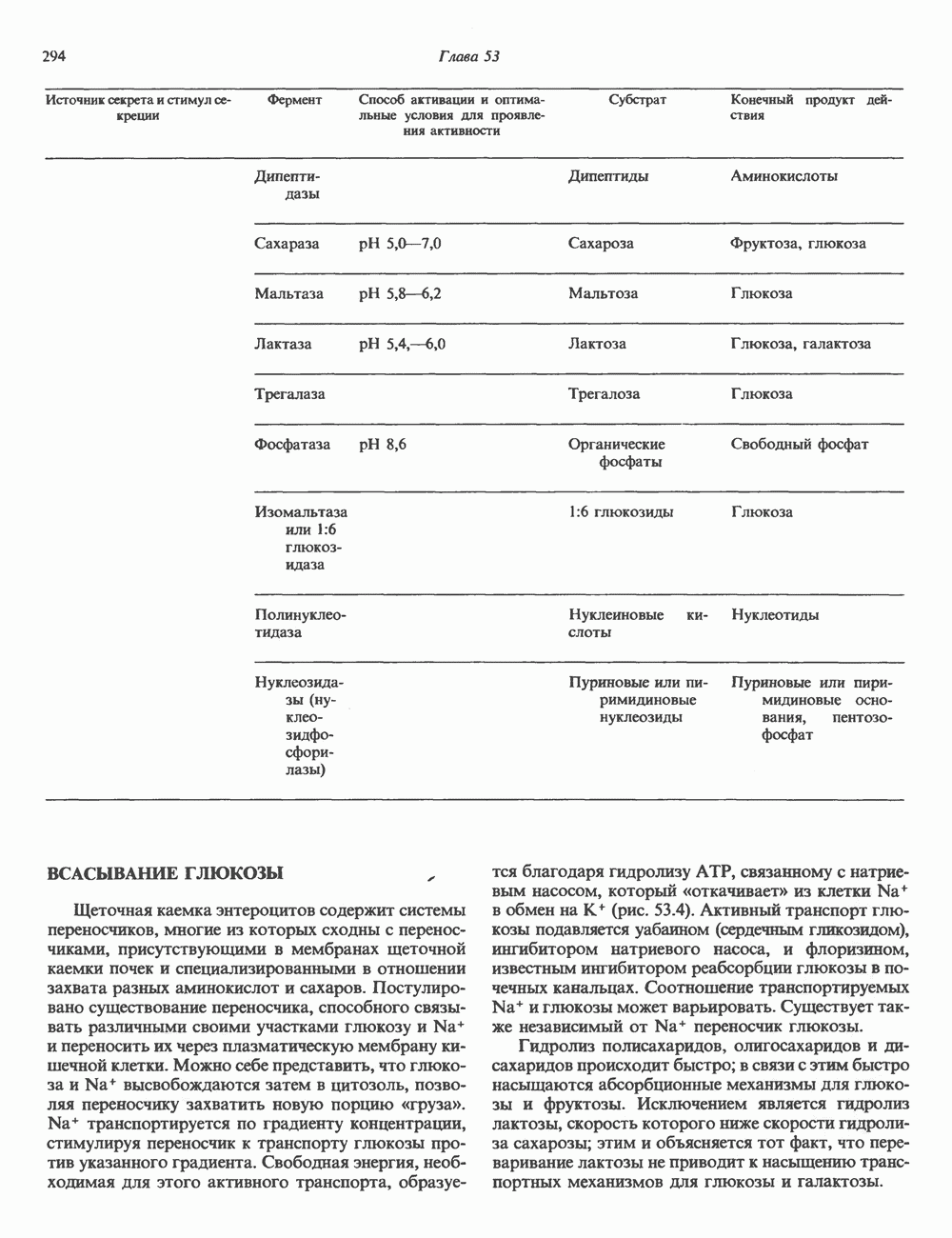
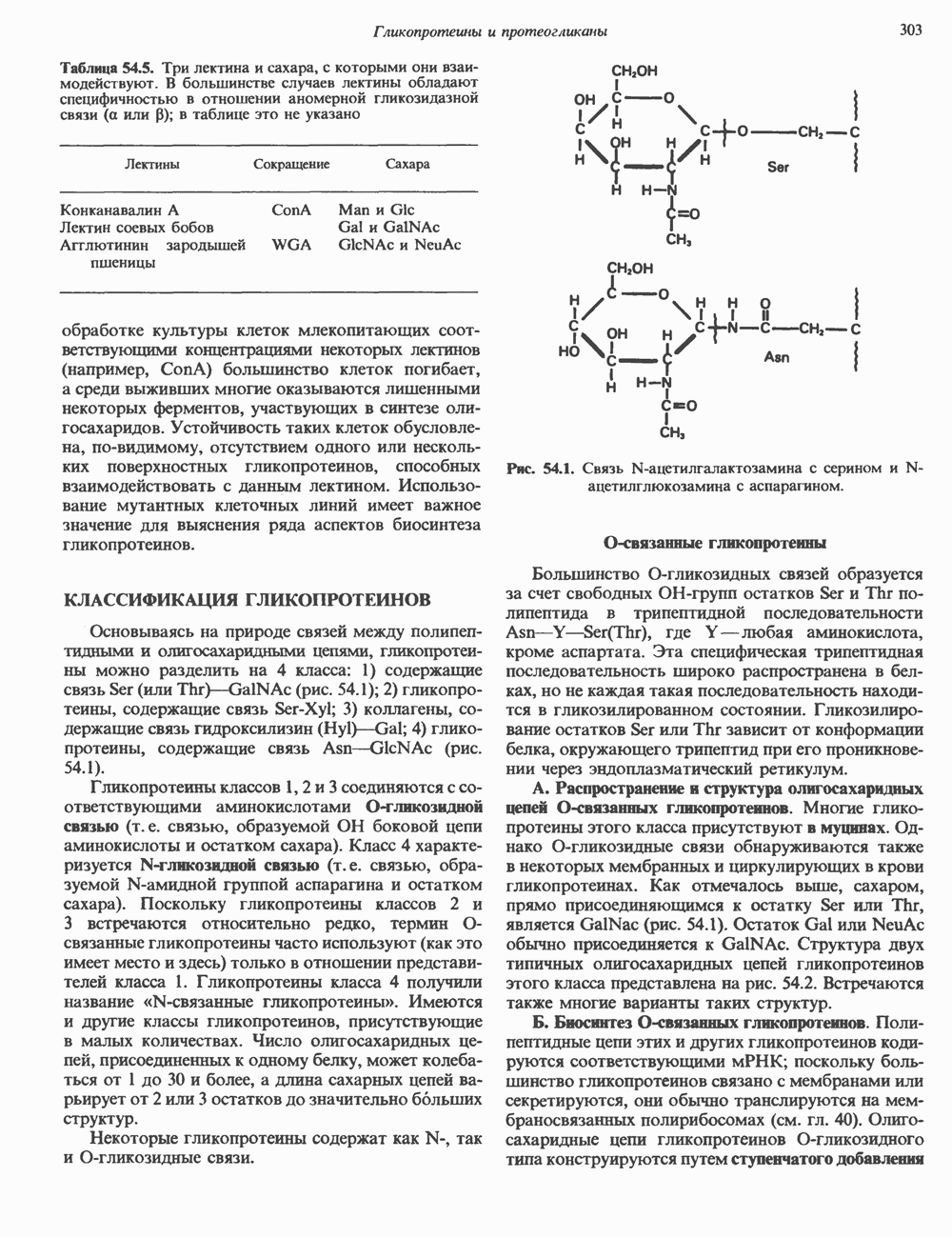
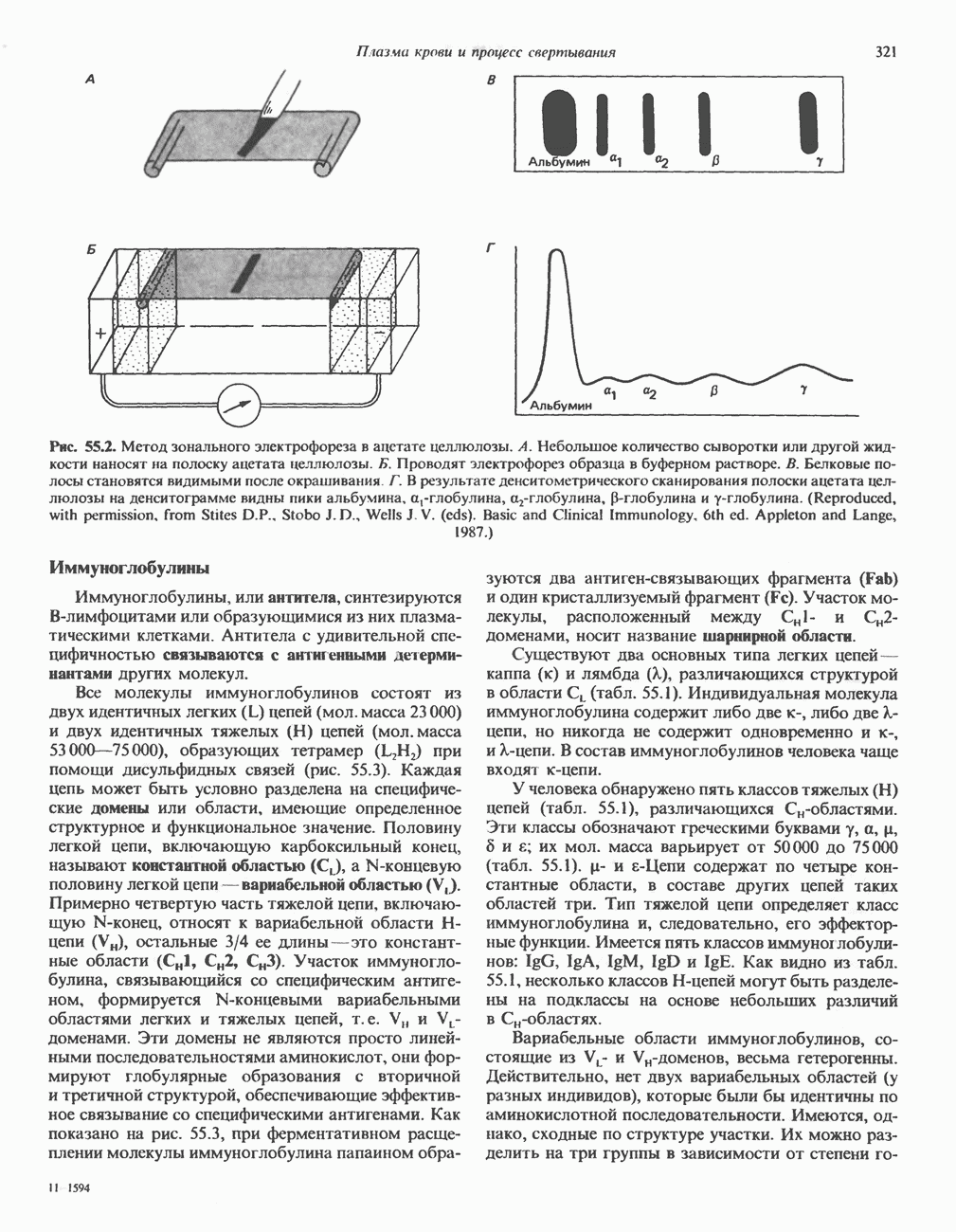
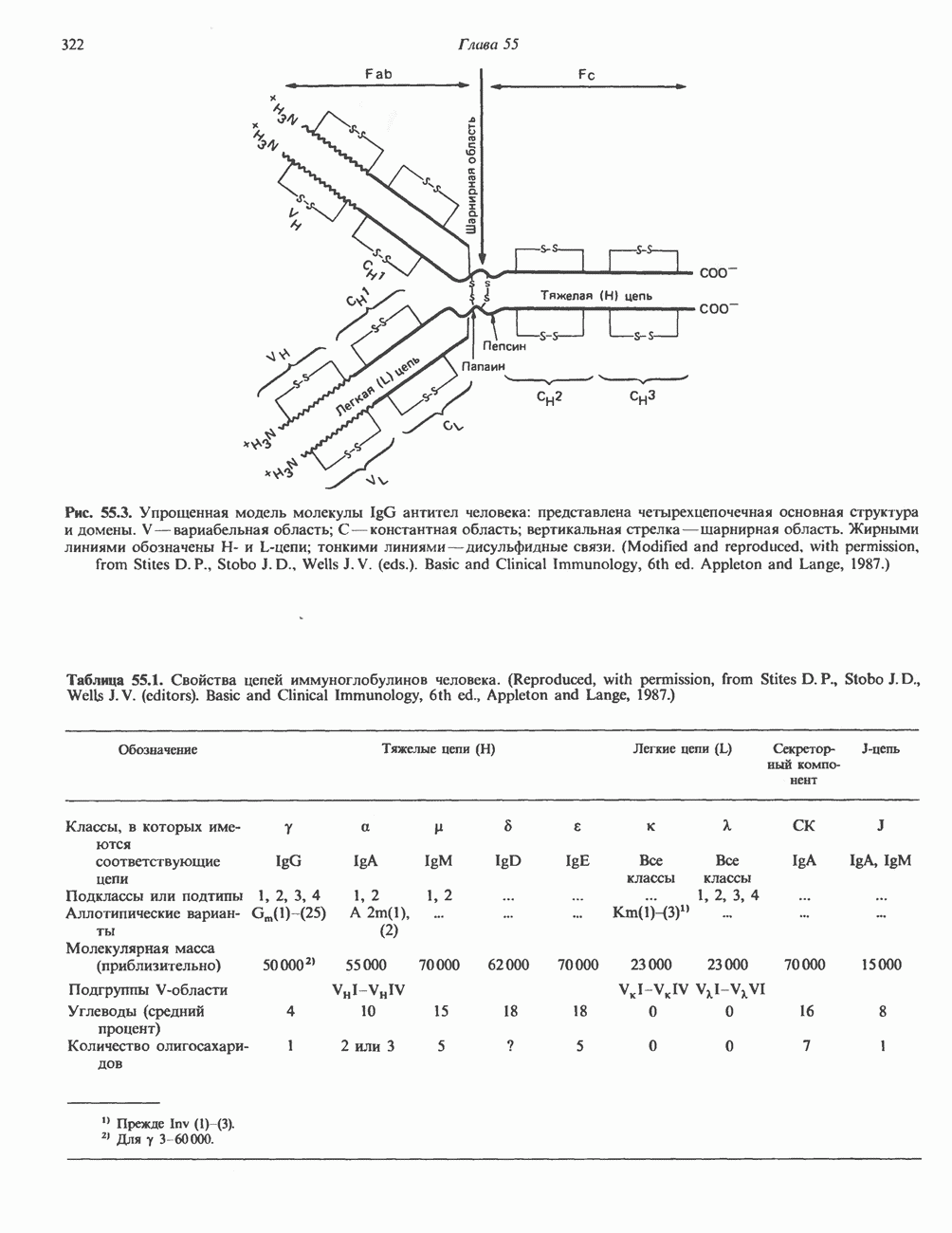
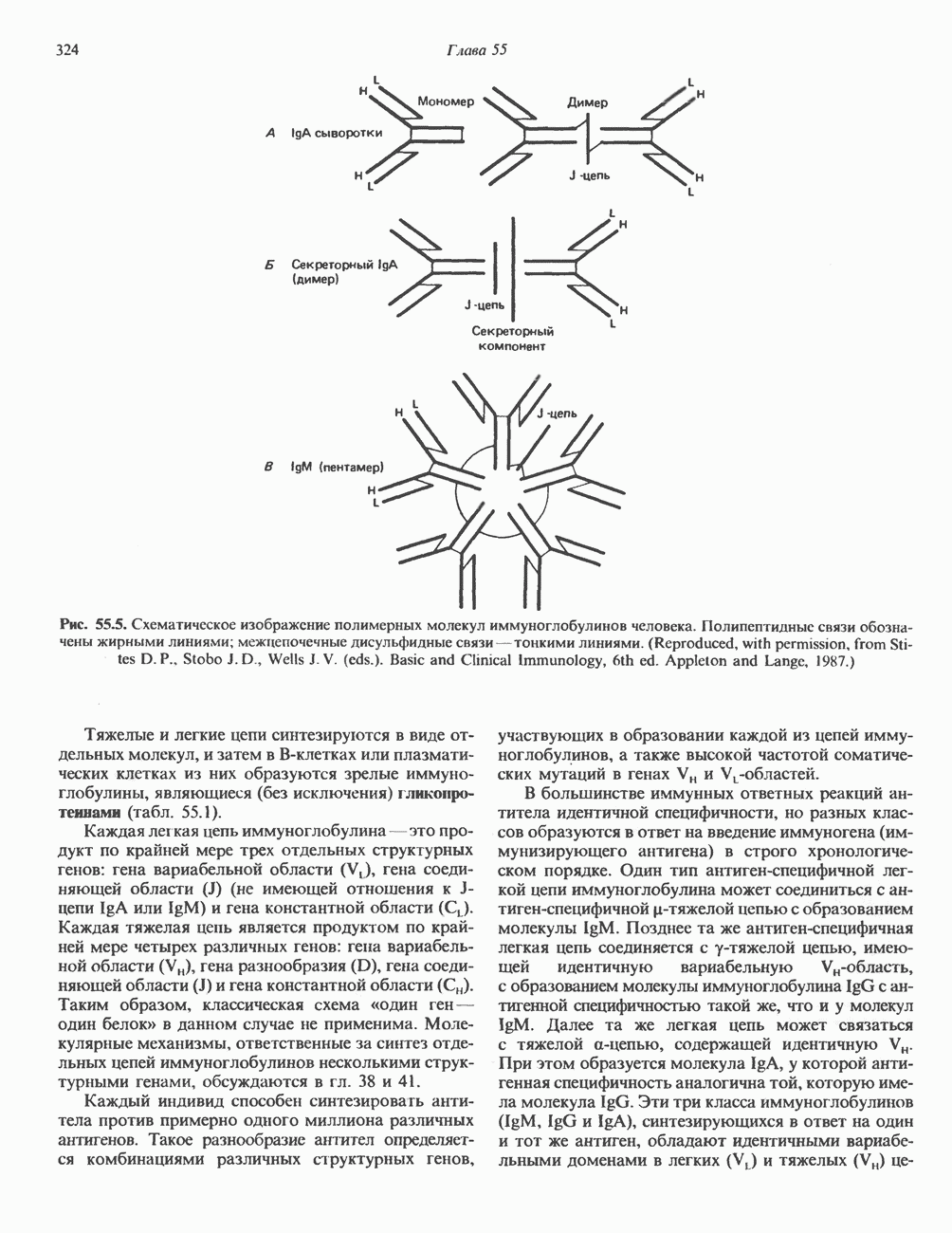
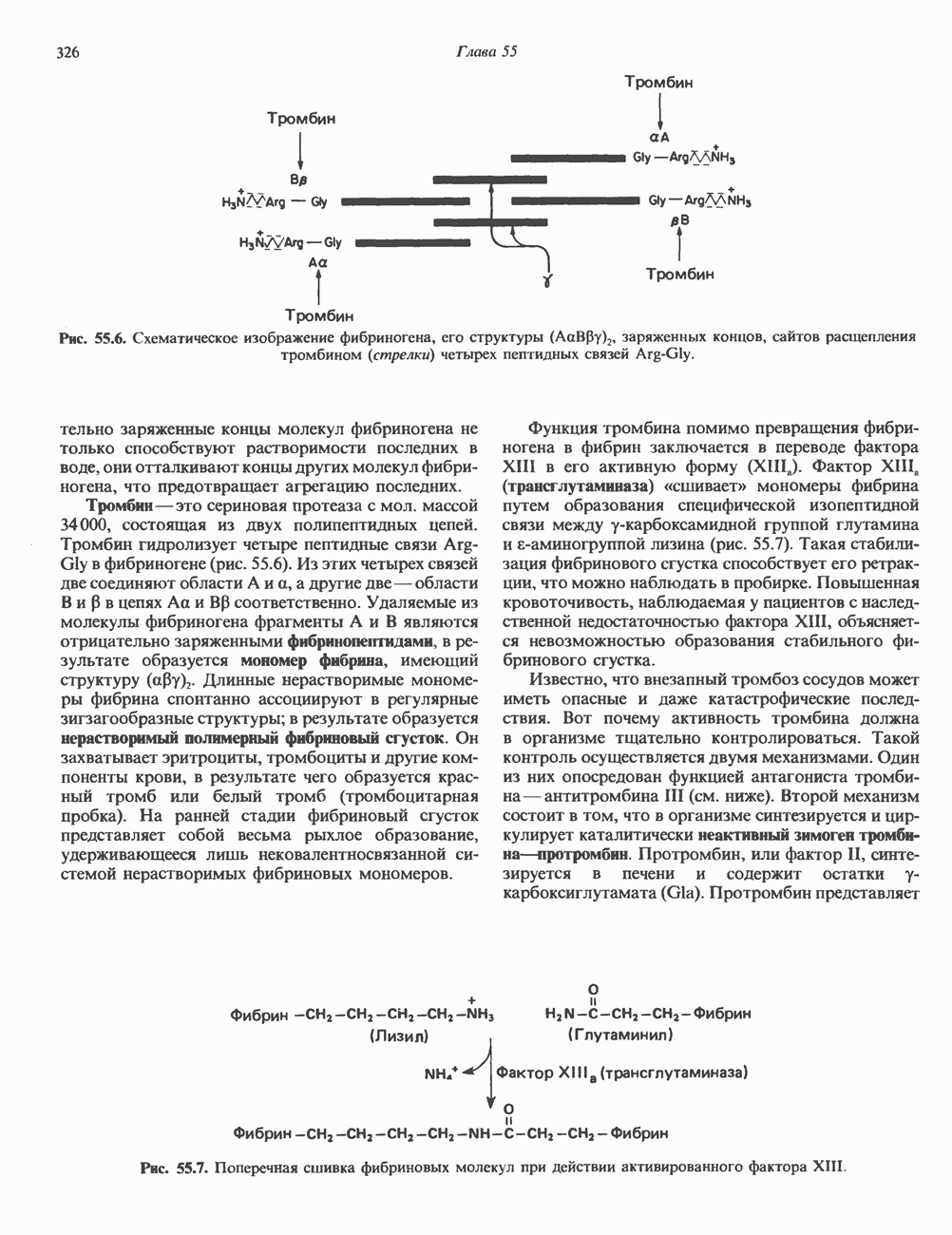
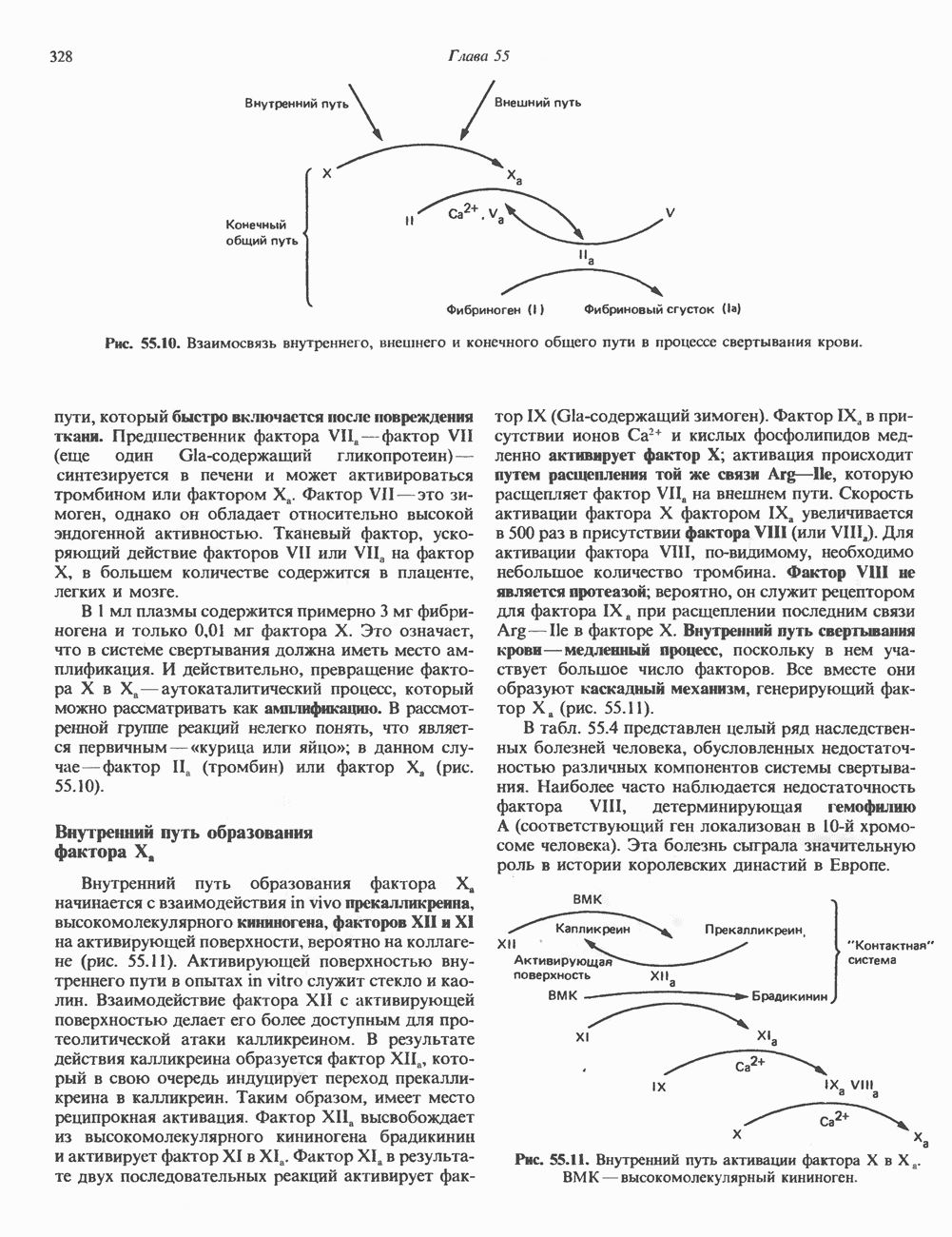
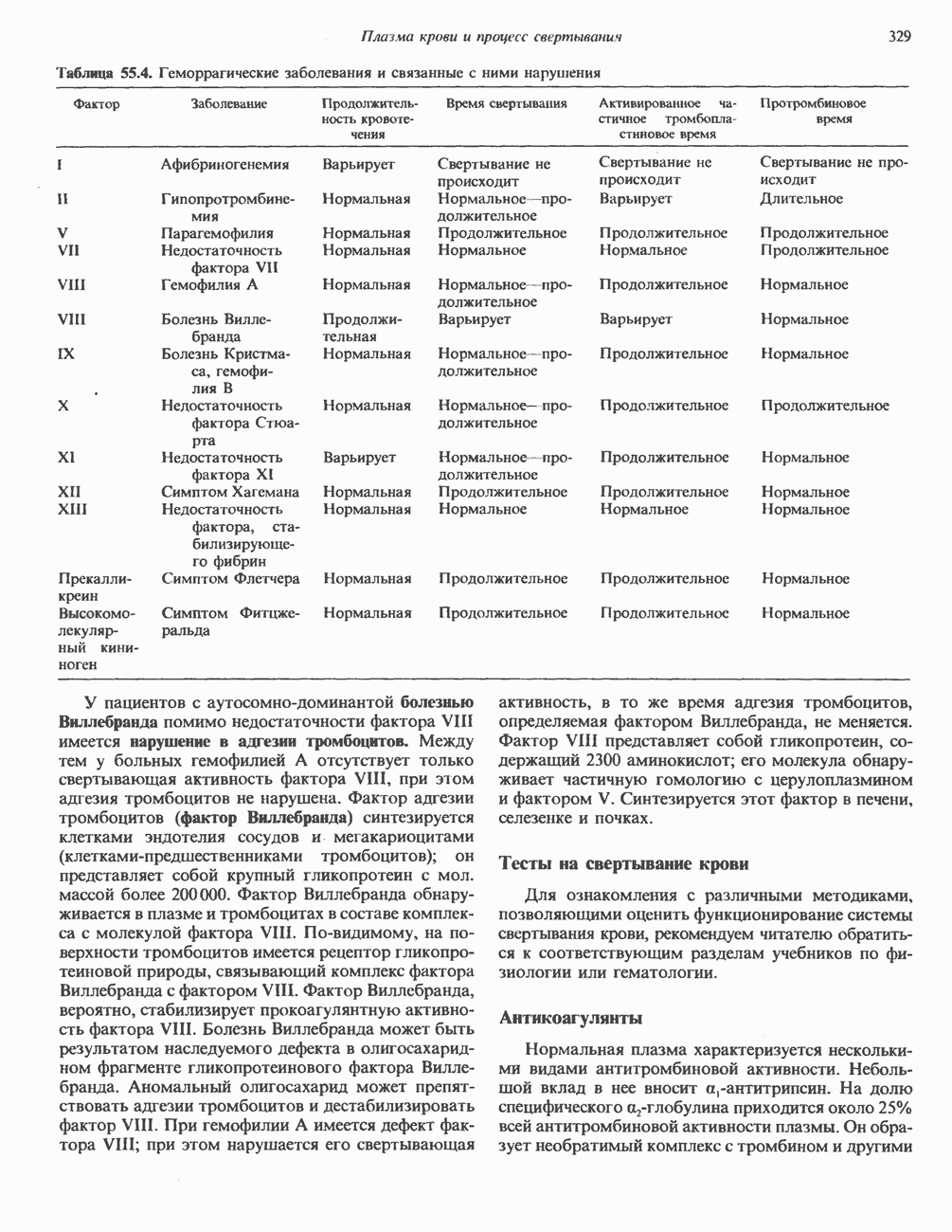
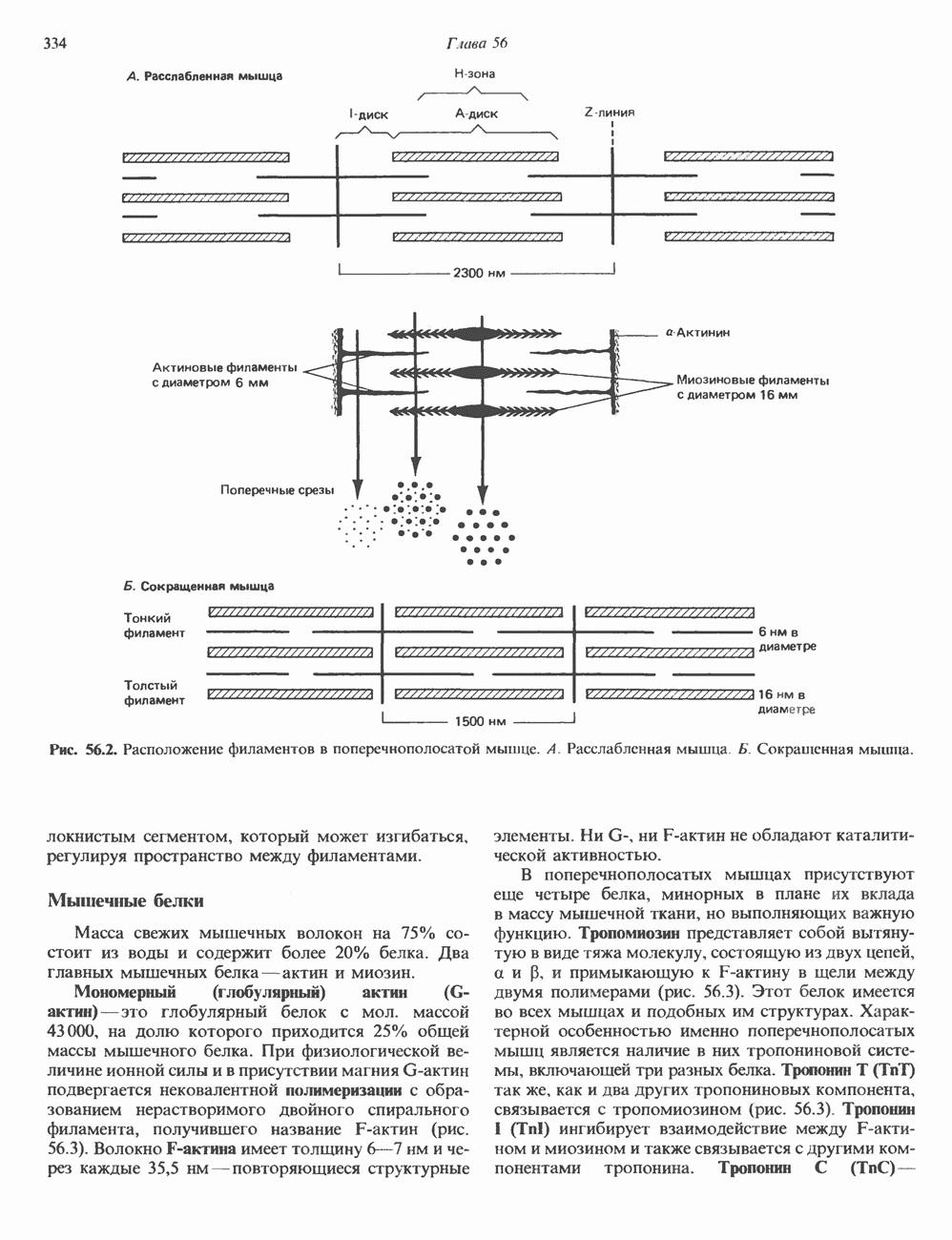
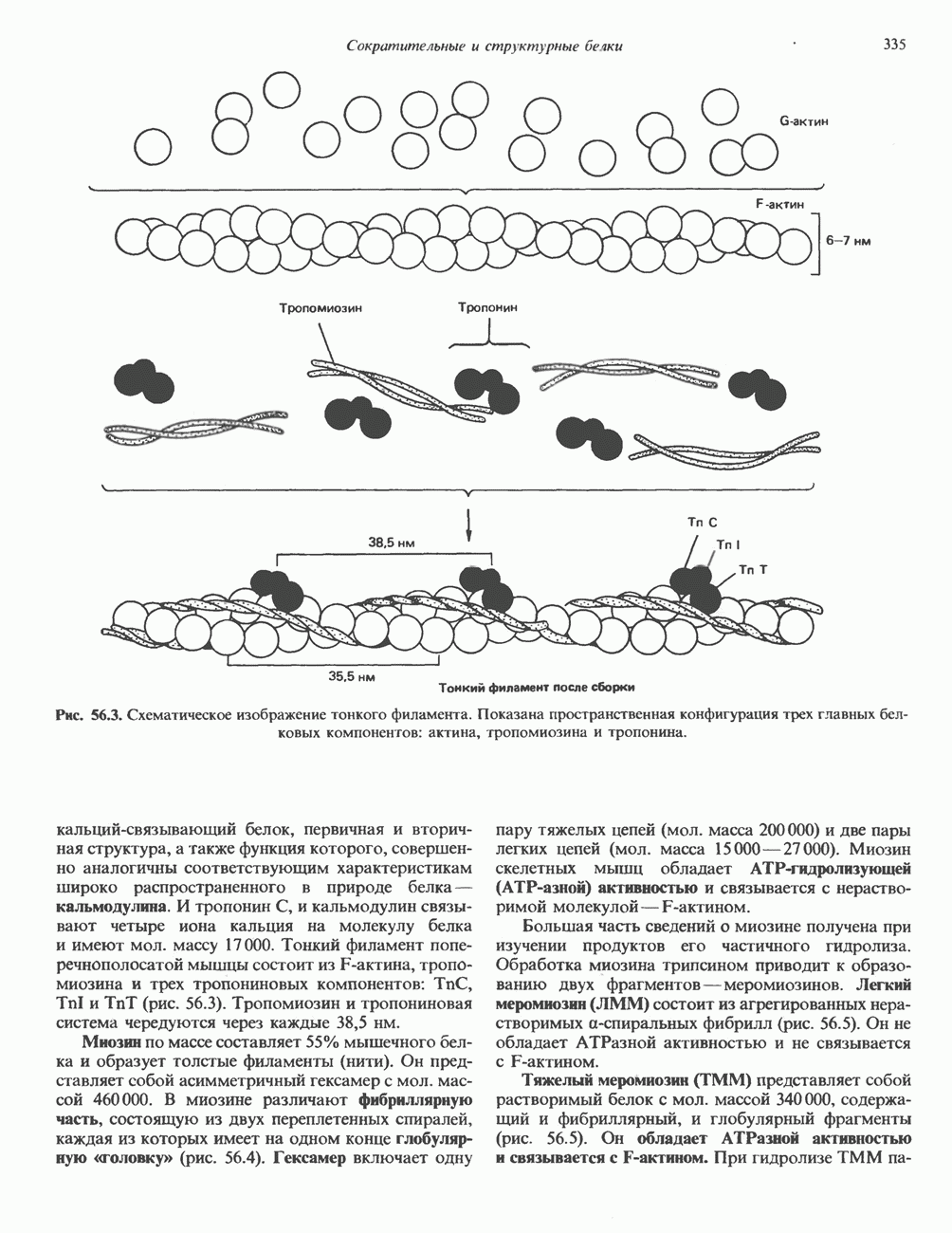
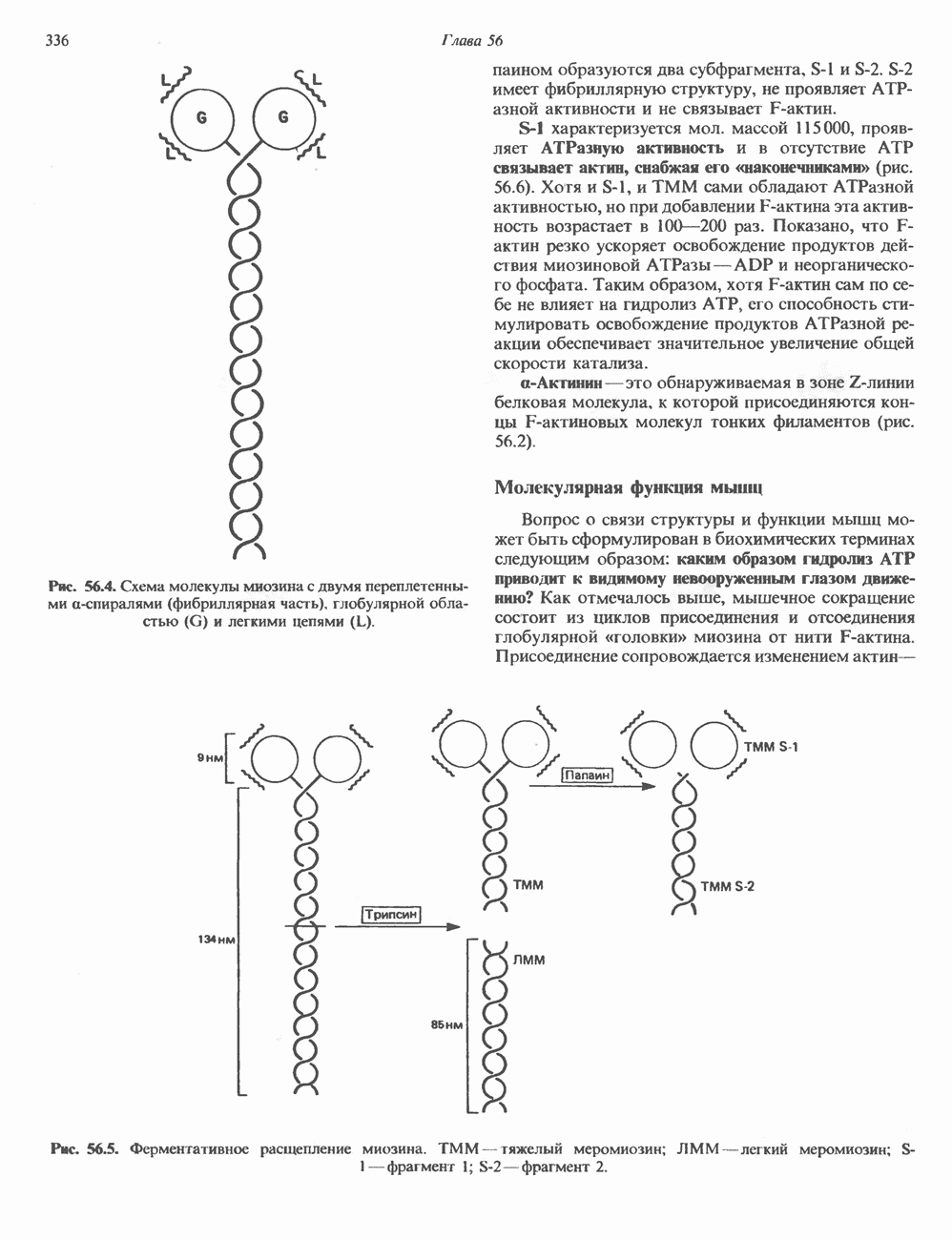
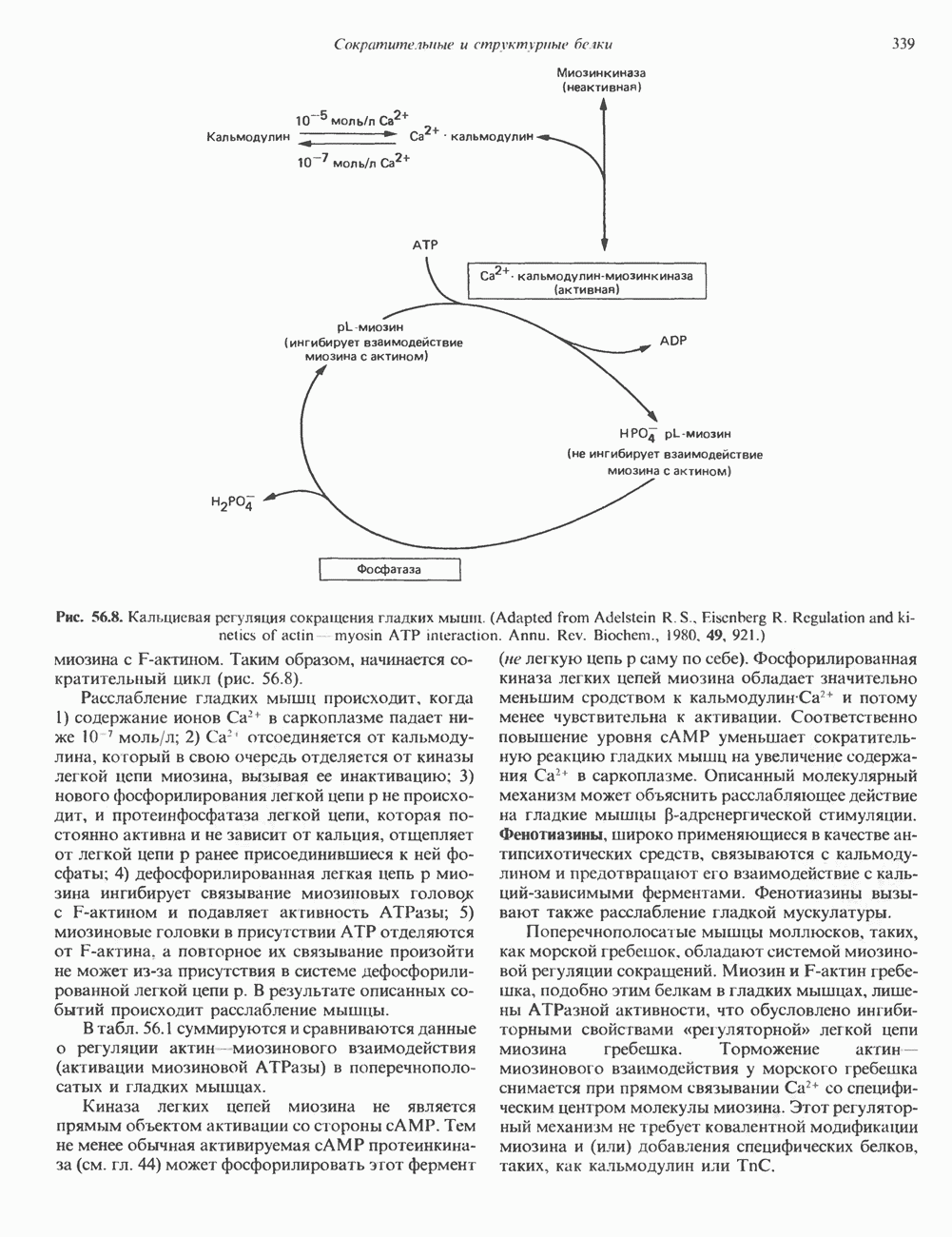
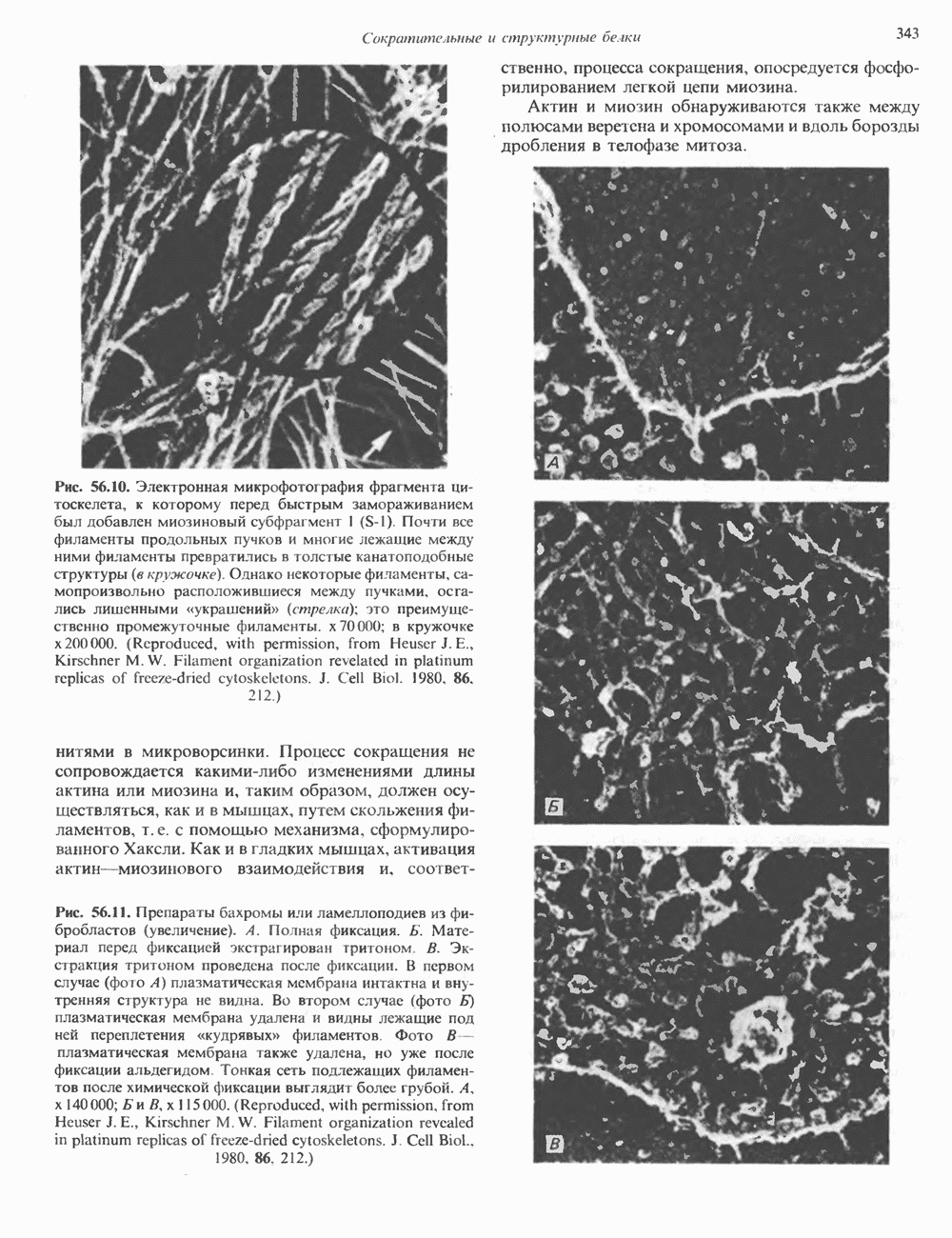
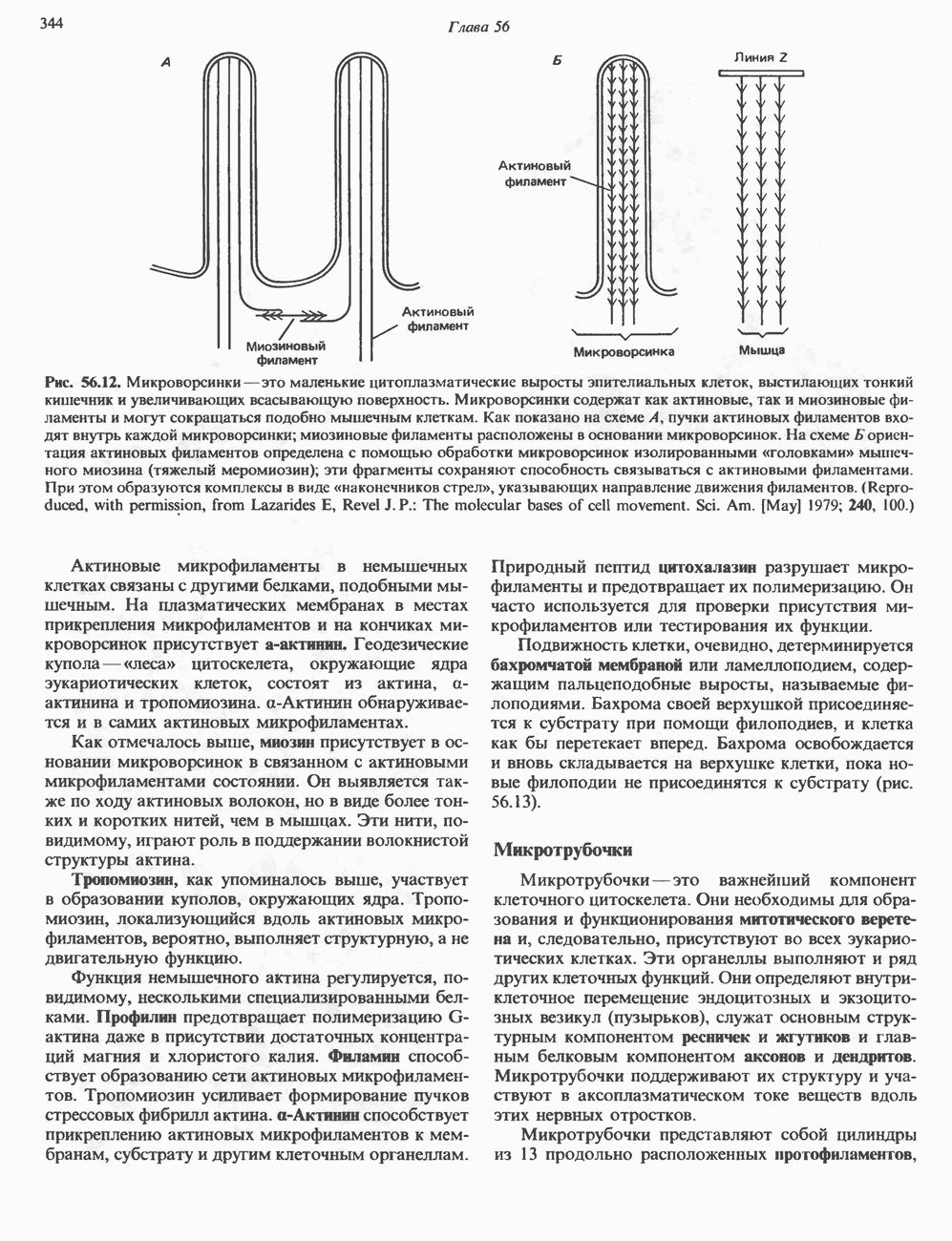
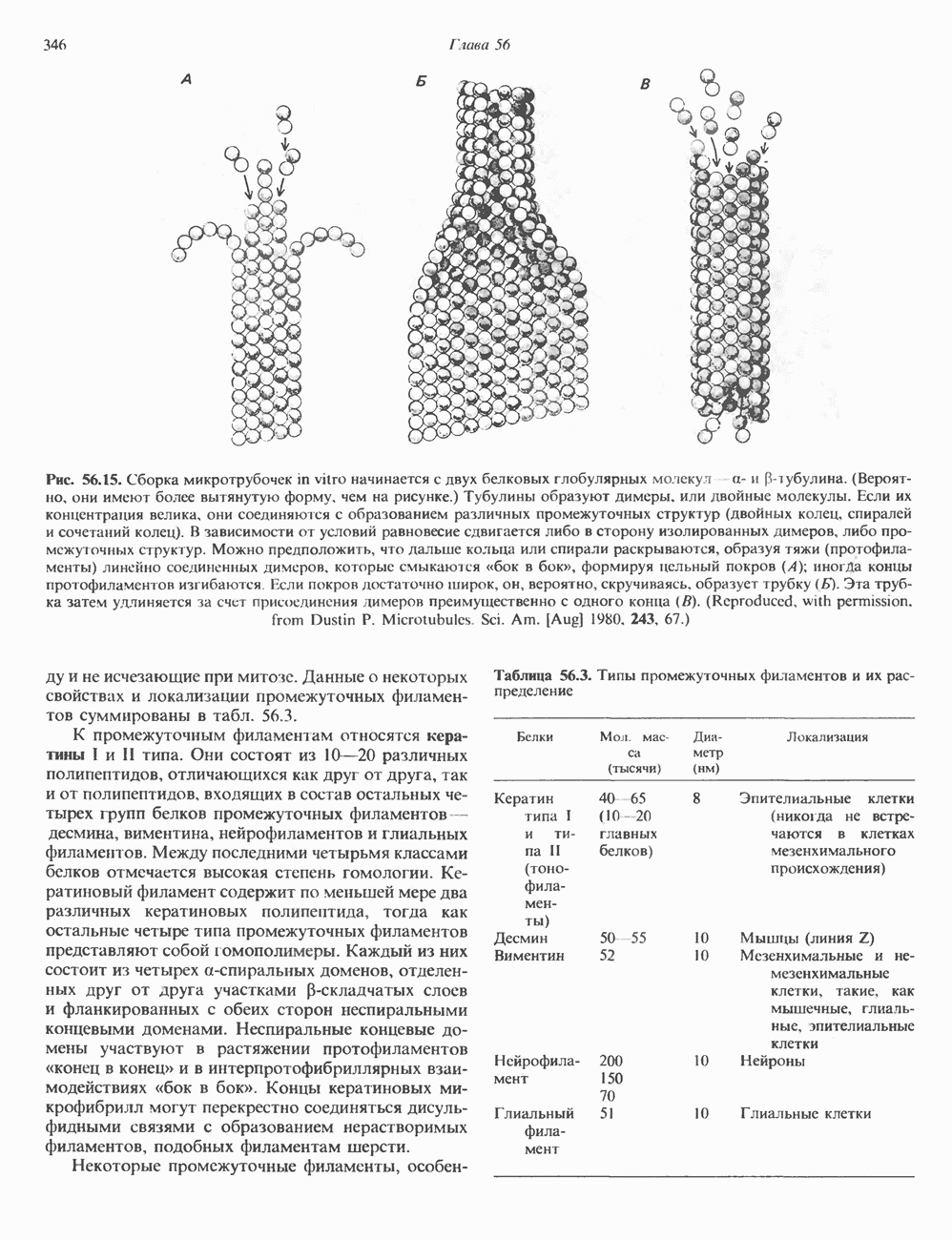
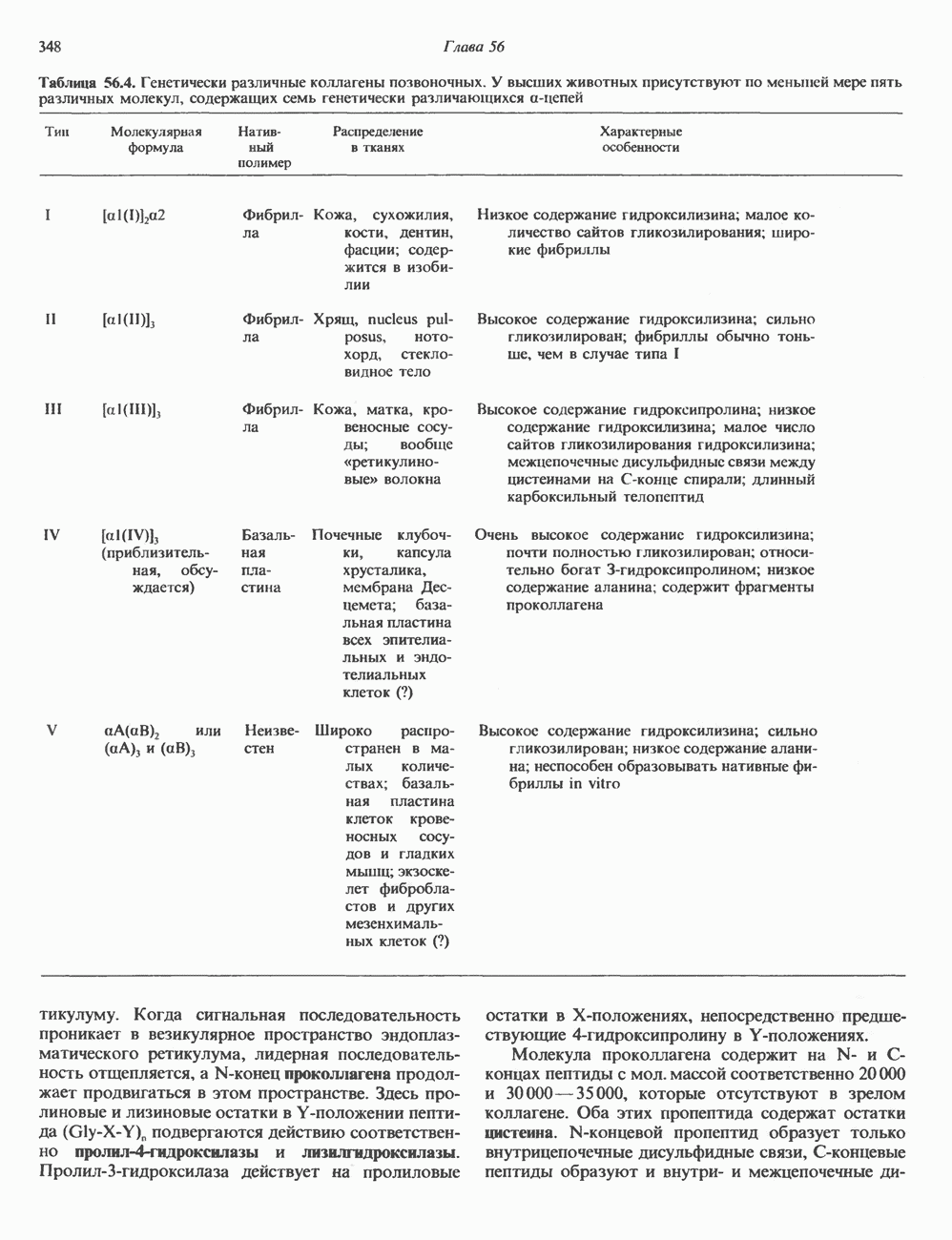
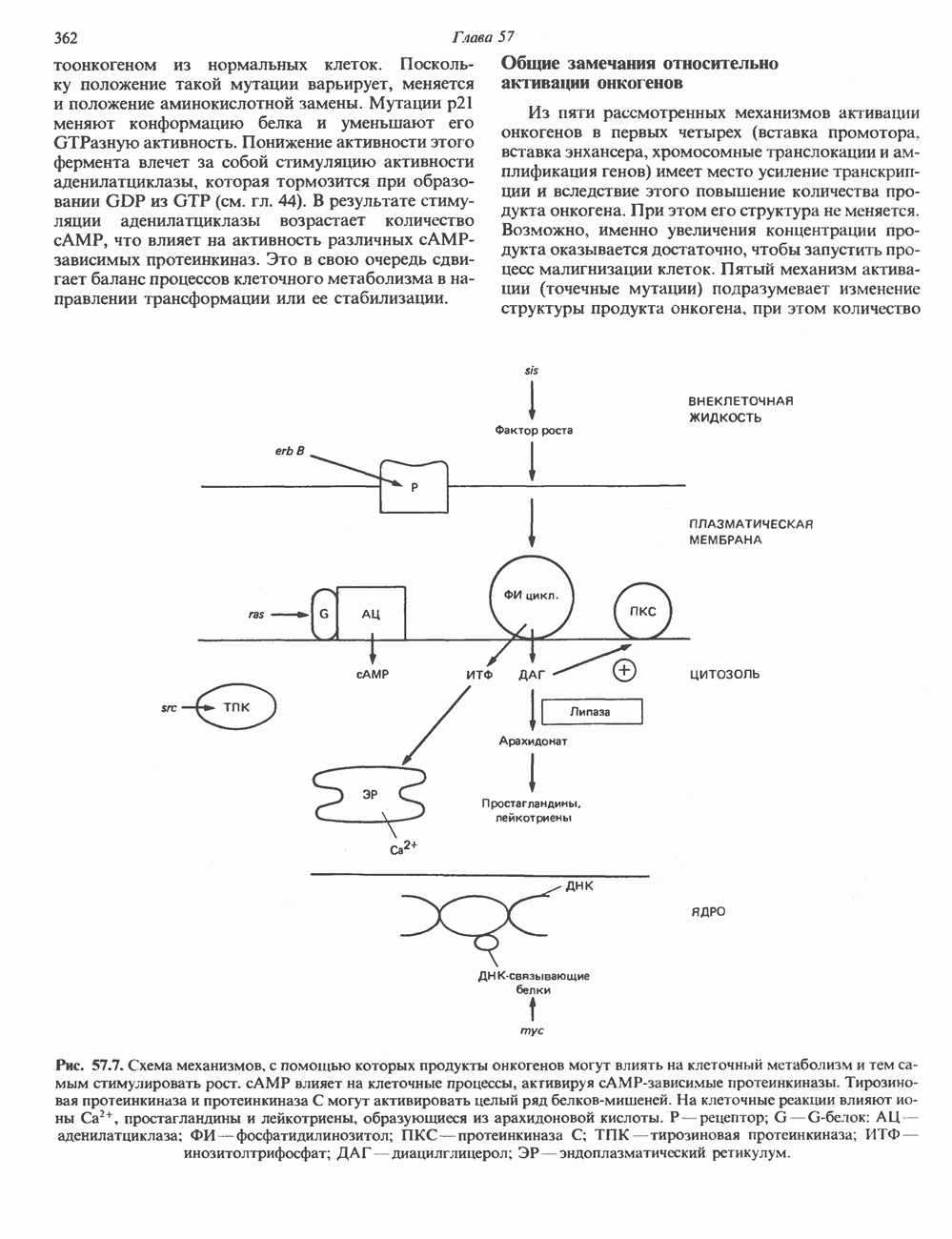
2. АНТИДИУРЕТИЧЕСКИЙ ГОРМОН (АДГ; ВАЗОПРЕССИН)
Регуляция секреции
Нервные импульсы, вызывающие секрецию АДГ, являются результатом действия ряда различных стимулирующих факторов. Главный физиологический стимул — это повышение осмоляльности плазмы. Его эффект опосредуется осморецепторами, локализованными в гипоталамусе, и барорецепторами, находящимися в сердце и других отделах сосудистой системы. Гемодилюция (снижение осмоляльности) оказывает противоположное действие. К другим стимулам относятся эмоциональный и физический стресс и воздействие фармакологических агентов, в том числе ацетилхолина, никотина и морфина. В большинстве случаев усиление секреции сочетается с повышением синтеза АДГ и нейрофизина II, поскольку при этом не происходит истощения резервов гормона. Адреналин и агенты, вызывающие увеличение объема плазмы, подавляют секрецию АДГ; аналогичным эффектом обладает этанол.
Механизм действия
Наиболее важные в физиологическом плане клетки-мишени для АДГ у млекопитающих — клетки дистальных извитых канальцев и собирательных трубочек почки. Эти протоки пересекают мозговое вещество почек, где градиент осмоляльности внеклеточных растворенных веществ в 4 раза выше, чем в плазме. Клетки этих протоков относительно непроницаемы для воды, так что в отсутствие АДГ моча не концентрируется и может выделяться в количествах, превышающих  в сутки. АДГ увеличивает проницаемость клеток для воды и способствует поддержанию осмотического равновесия между мочой собирательных трубочек и гипертоническим содержимым интерстициального пространства, благодаря чему объем мочи сохраняется в пределах
в сутки. АДГ увеличивает проницаемость клеток для воды и способствует поддержанию осмотического равновесия между мочой собирательных трубочек и гипертоническим содержимым интерстициального пространства, благодаря чему объем мочи сохраняется в пределах  в сутки. На слизистых (мочевых) мембранах эпителиальных клеток этих структур присутствуют рецепторы АДГ, которые связаны с аденилатциклазой; считают, что действие АДГ на почечные канальцы опосредуется сАМР. Описанное физиологическое действие послужило основанием для того, чтобы назвать гормон «антидиуретическим». сАМР и ингибиторы фосфодиэстеразы имитируют эффекты АДГ. В
в сутки. На слизистых (мочевых) мембранах эпителиальных клеток этих структур присутствуют рецепторы АДГ, которые связаны с аденилатциклазой; считают, что действие АДГ на почечные канальцы опосредуется сАМР. Описанное физиологическое действие послужило основанием для того, чтобы назвать гормон «антидиуретическим». сАМР и ингибиторы фосфодиэстеразы имитируют эффекты АДГ. В
условиях in vivo повышение уровня кальция в среде, омывающей слизистую поверхность канальцев, тормозит действие АДГ на перемещение воды (очевидно, путем ингибирования аденилатциклазы, поскольку эффект самого сАМР при этом не уменьшается). Описанный механизм может отчасти обусловливать повышенный диурез, характерный для больных с гиперкальциемией.
Патофизиология
Нарушения секреции или действия АДГ приводят к несахарному диабету, который характеризуется выделением больших объемов разбавленной мочи. Первичный несахарный диабет, связанный с дефицитом АДГ, обычно развивается при повреждении гипоталамо-гипофизарного тракта вследствие перелома основания черепа, опухоли или инфекции: однако он может иметь и наследственную природу. При наследственном нефрогенном несахарном диабете секреция АДГ остается нормальной, но клетки-мишени утрачивают способность реагировать на гормон, вероятно, из-за нарушения его рецепции (см. табл. 43.2). Этот наследственный дефект отличается от приобретенного нефрогенного несахарного диабета, который чаще всего возникает при терапевтическом введении лития больным с маниакально-депрессивным психозом. Синдром неадекватной секреции АДГ связан обычно с эктопическим образованием гормона различными опухолями (обычно опухолями легких), но может также наблюдаться и при болезнях мозга, легочных инфекциях или гипотиреозе. Неадекватной такая секреция считается потому, что продукция АДГ происходит с нормальной или повышенной скоростью в условиях гипоосмоляльности, и это вызывает устойчивую и прогрессивную гипонатриемию с выделением гипертонической мочи.
ЛИТЕРАТУРА
Гормоны передней доли гипофиза
Douglass J., Cive/li О., Herbert Е. Polyprotein gene expression: Generation of diversity of neuroendocrine peptides, Annu. Rev. Biochem., 1984, 53, 665.
Frantz A.G. Prolactin, N. Engl. J. Med., 1978, 298, 201.
Krieger D. T. The multiple faces of pro-opiomelanocortin, a prototype precursor molecule, Clin. Res., 1983, 3, 342.
Krulich L. Central neurotransmitters and the selection of prolactin. GH, LH, and TSH, Annu. Rev. Physiol., 1979, 41, 603.
Nikolics K. et al. A prolactin-inhibiting factor with the precursor for human gonadotropin-releasing hormone, Nature, 1986. 316, 511.
Pierce J.G.. Parsons T.F. Glycoprotein hormones: Structure and function, Annu. Rev. Biochem., 1981, 50, 465.
Seeburg P. The human growth hormone gene family: Structure and evolution of the chromosomal locus. Nucleic Acids Res., 1983, 11, 3939.
Гормоны задней доли гипофиза
Chord I. T. The posterior pituitary gland, Clin. Endocrinol., 1975, 4, 89.
Robertson G. L. Regulation of vasopressin function in health and disease, Resent Prog. Horm. Res.. 1977, 33, 333.
Гормоны гипоталамуса
Imura H. et al. Effect of CNS peptides on hypothalamic regulation of pituitary secretion, Adv. Biochem. Psychopharma-col, 1981. 28, 557.
Labrie F. et al. Mechanism of action of hypothalamic hormones in the adenohypophysis, Annu. Rev. Physiol., 1979, 41, 555.
Reichlin S. Systems for the study of regulation of neuropeptide secretion. In: Neurosecretion and Brain Peptides: Implications for Brain Function and Neurological Disease, Martin J. B., Reichlin S., Bick K. L. (eds.), Raven Press, 1981.