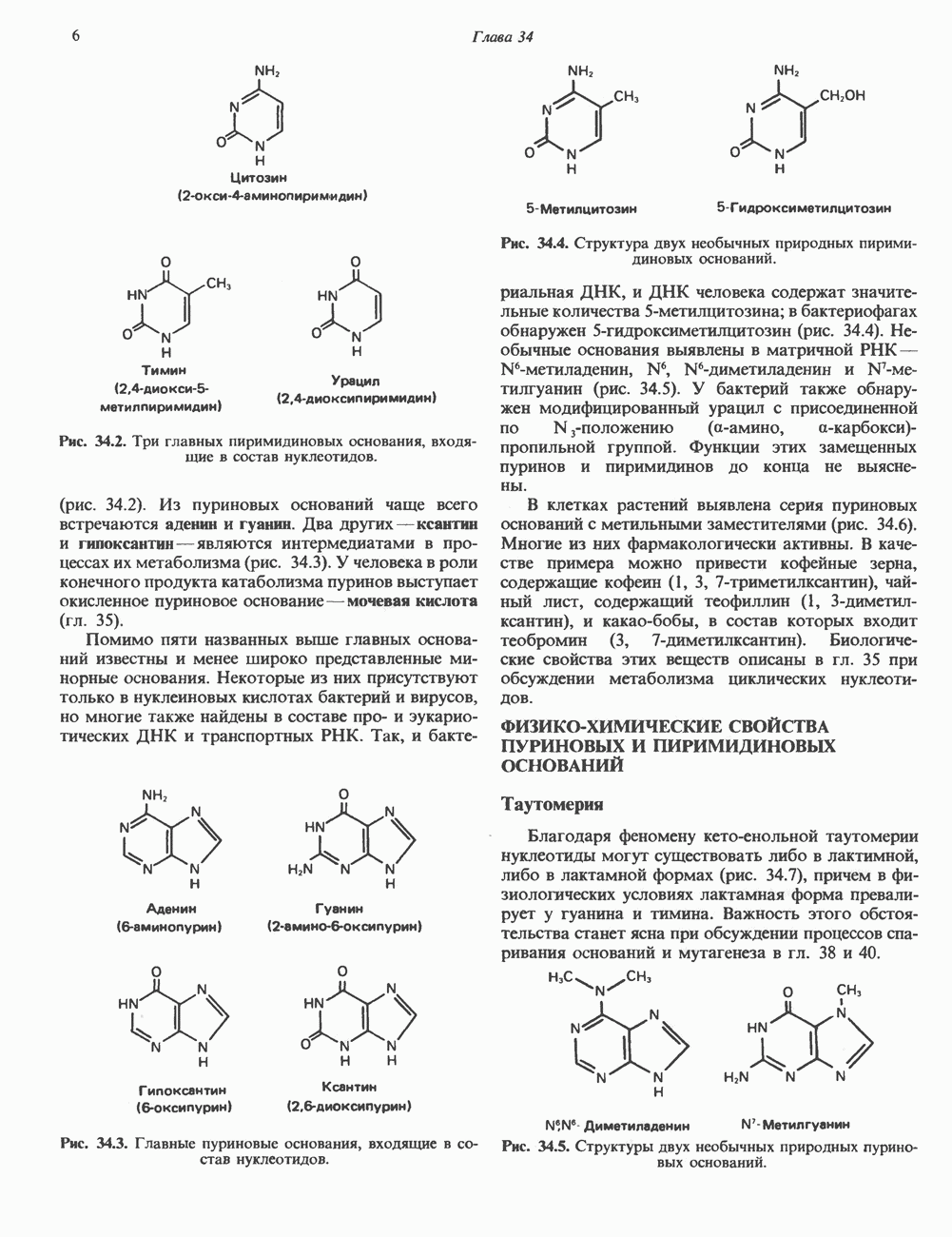
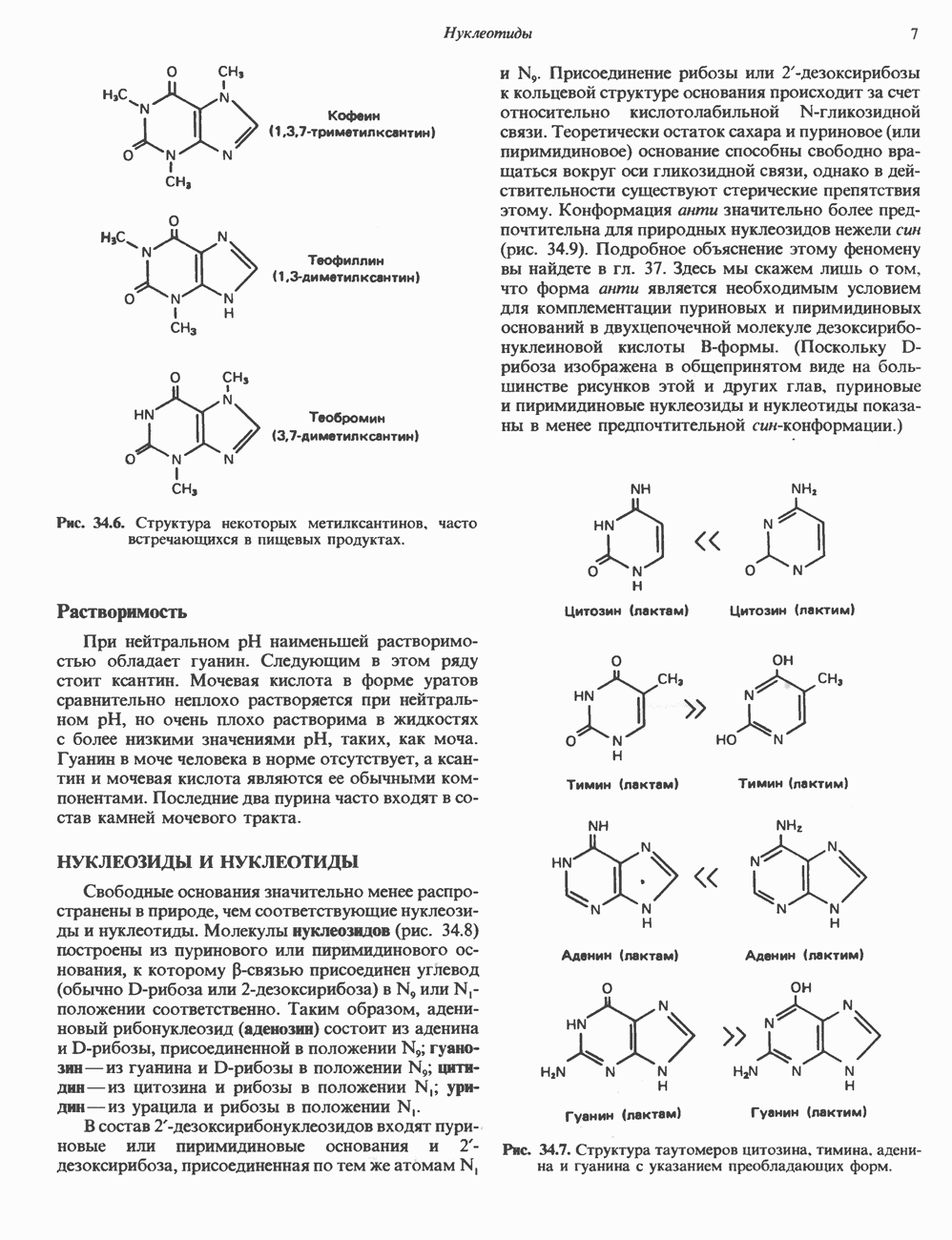
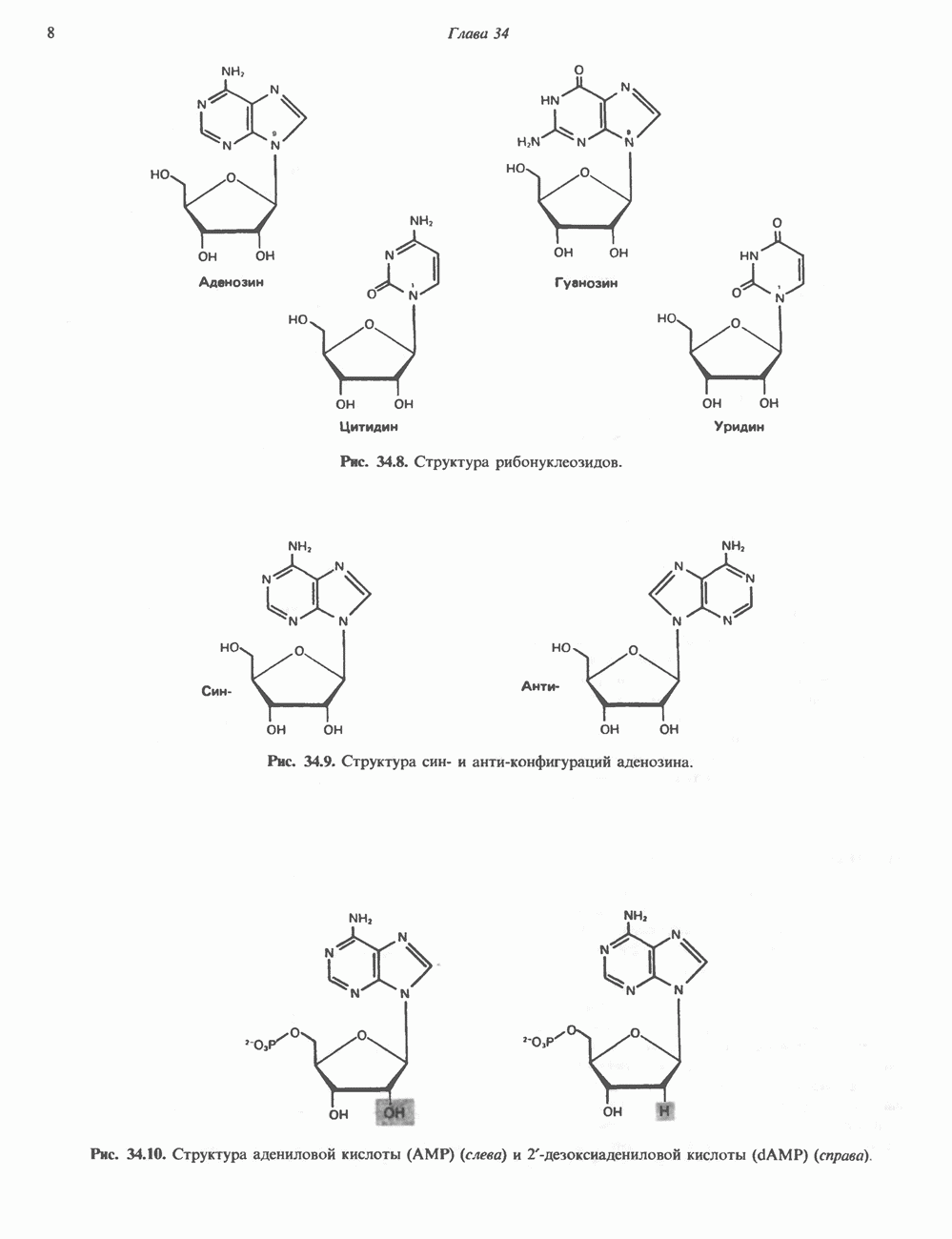
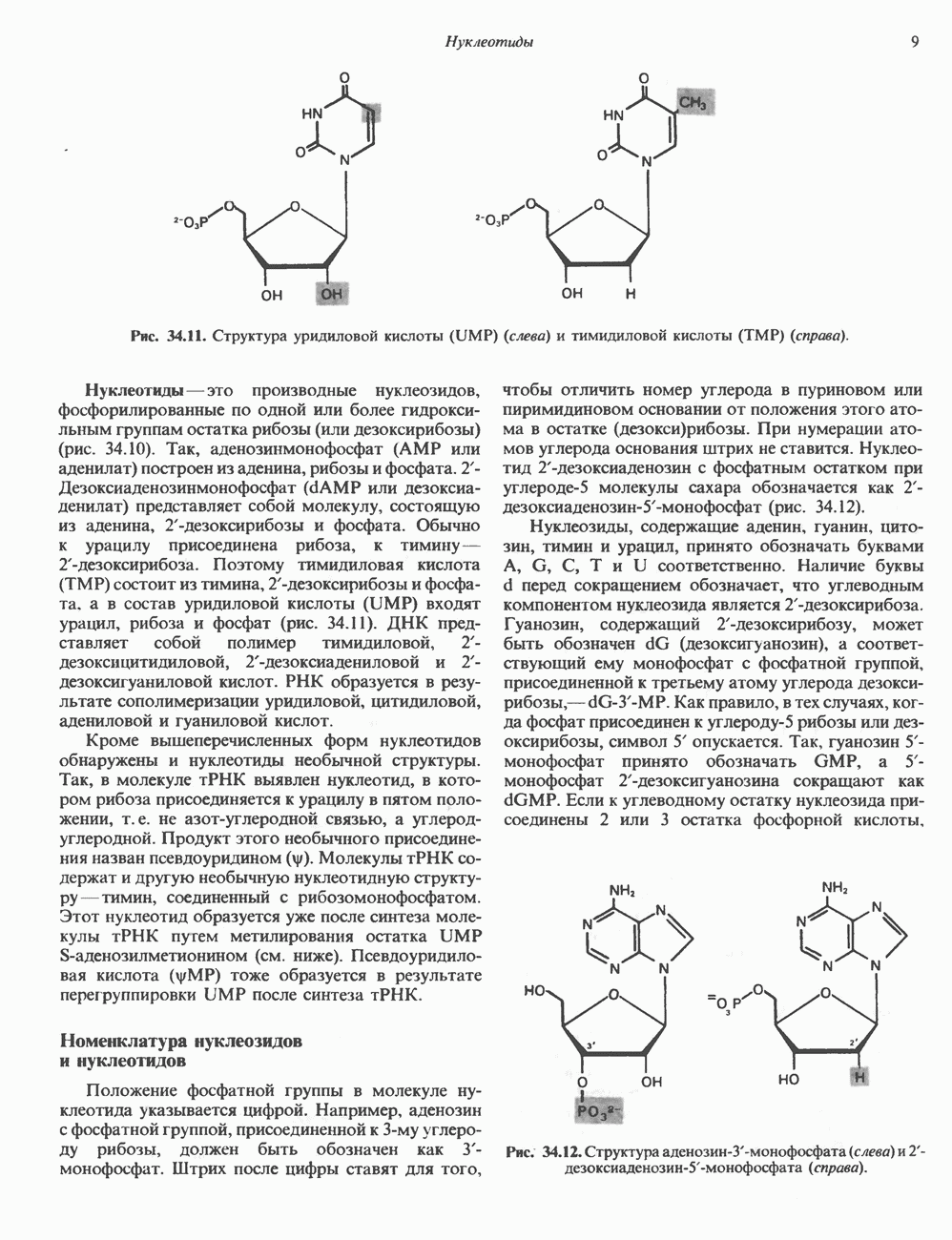
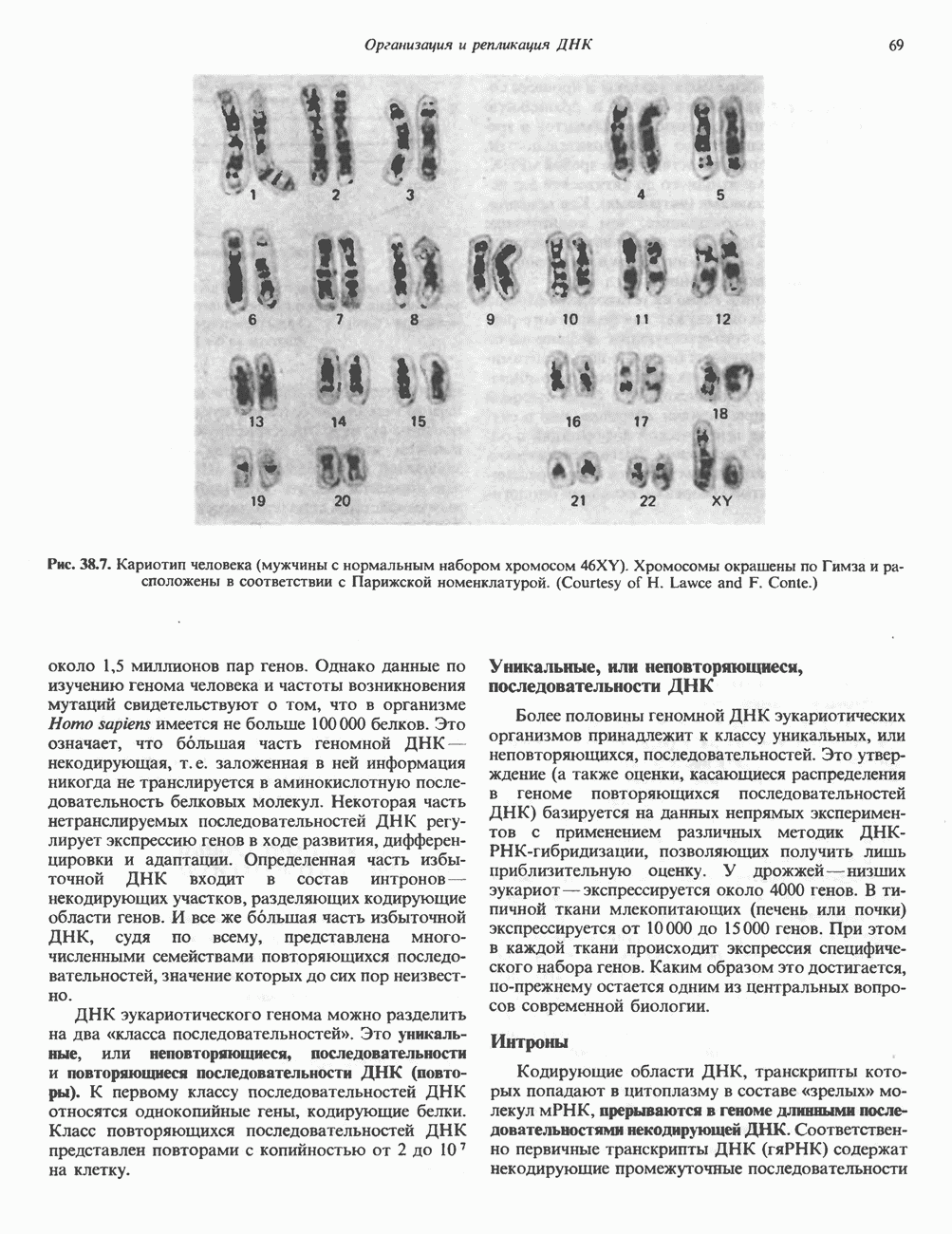
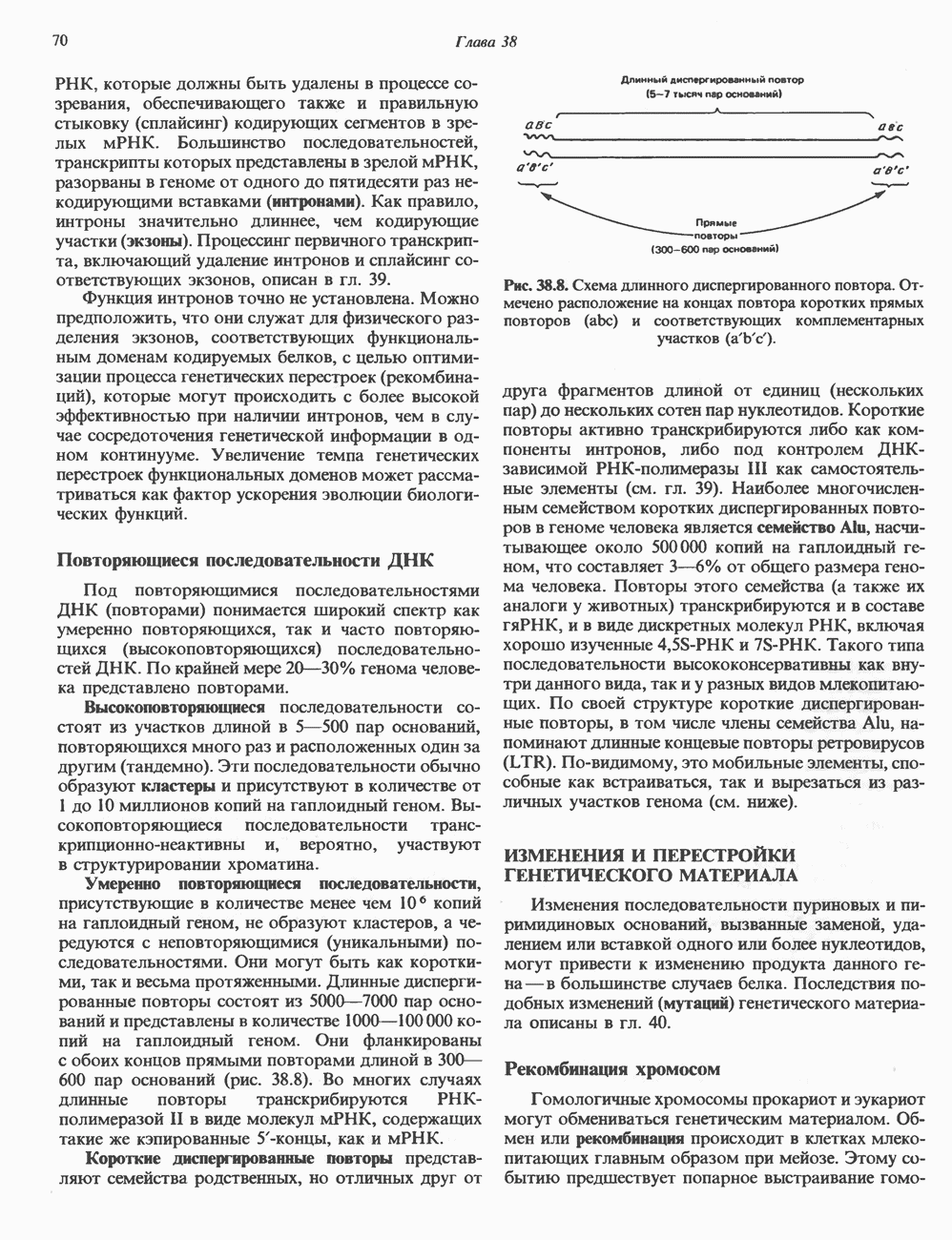
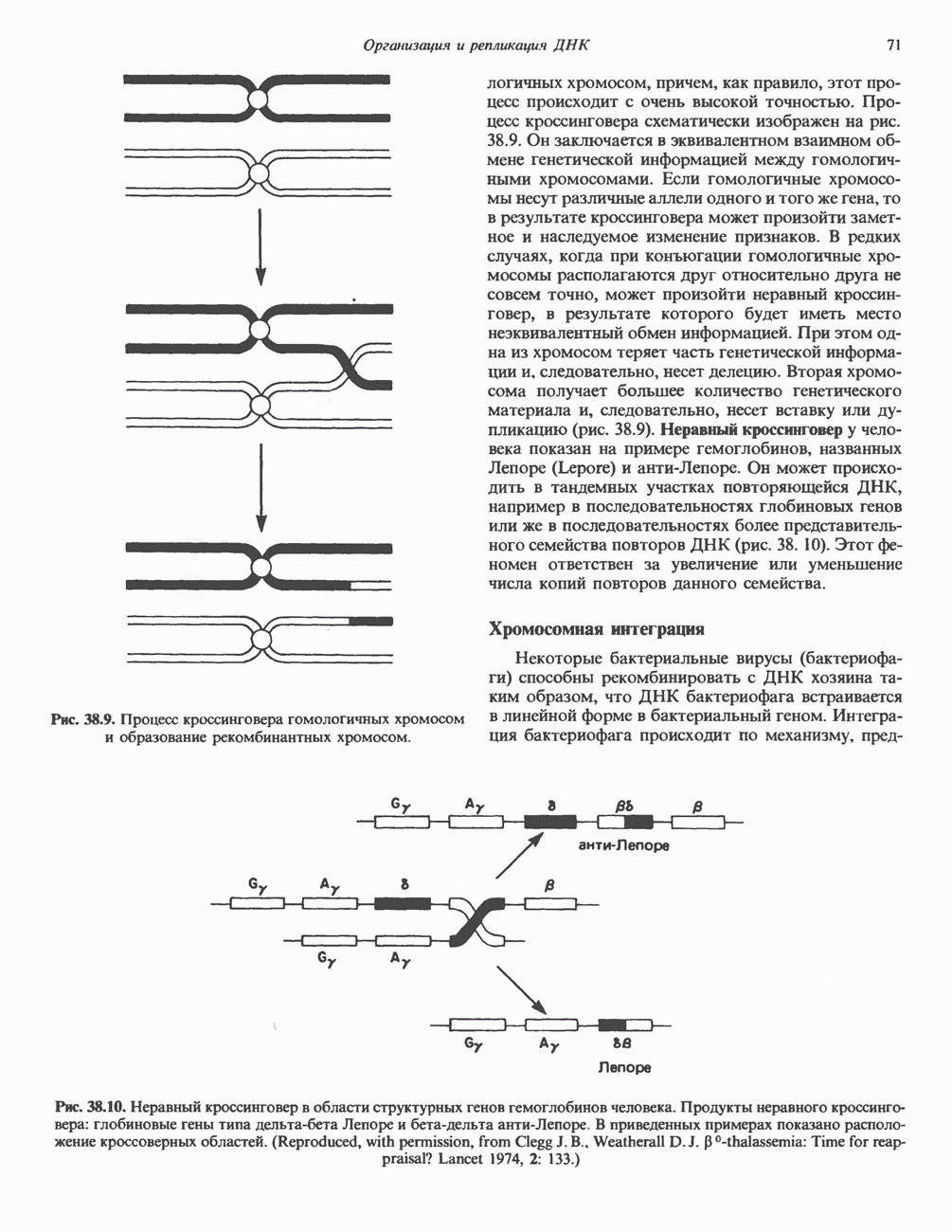
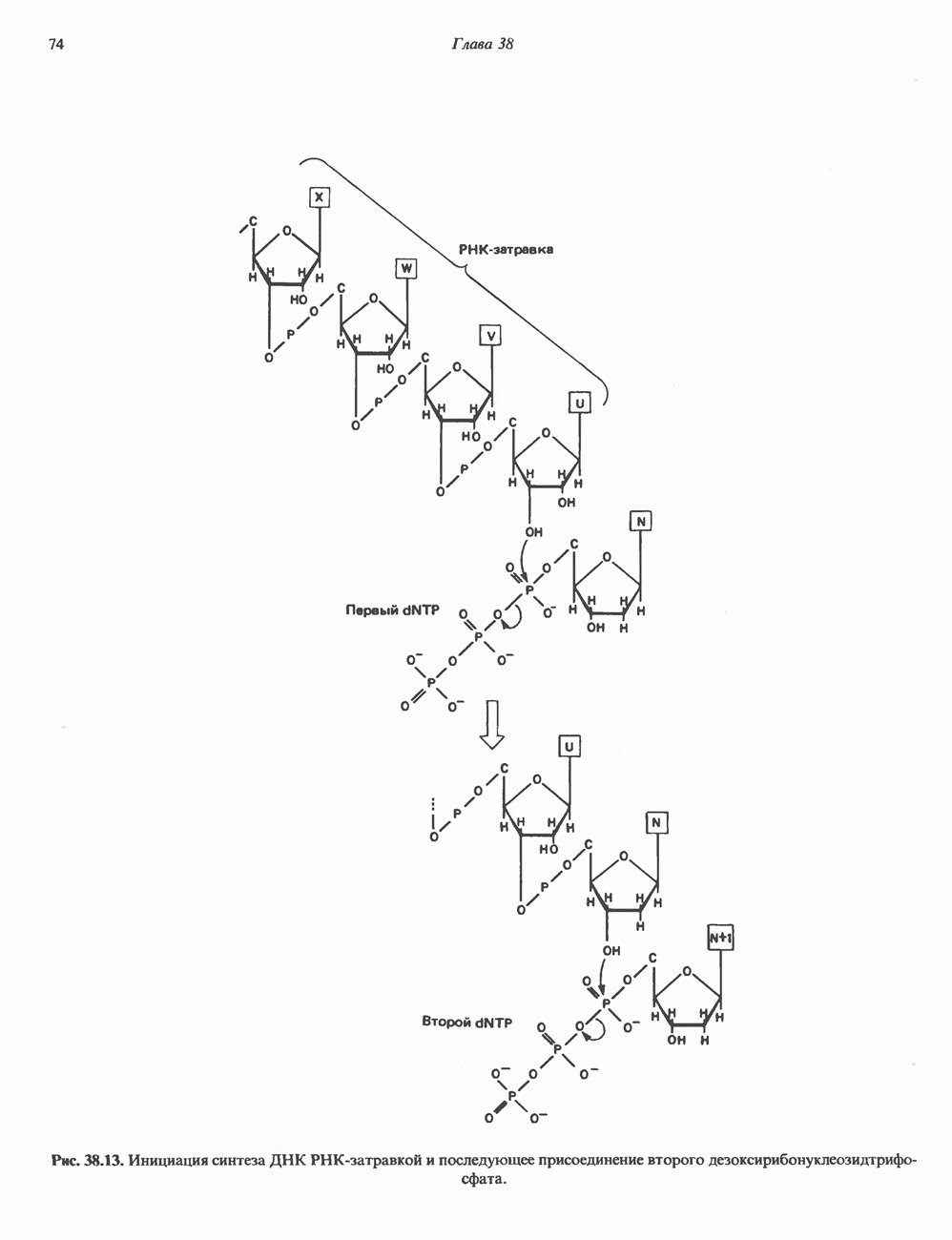
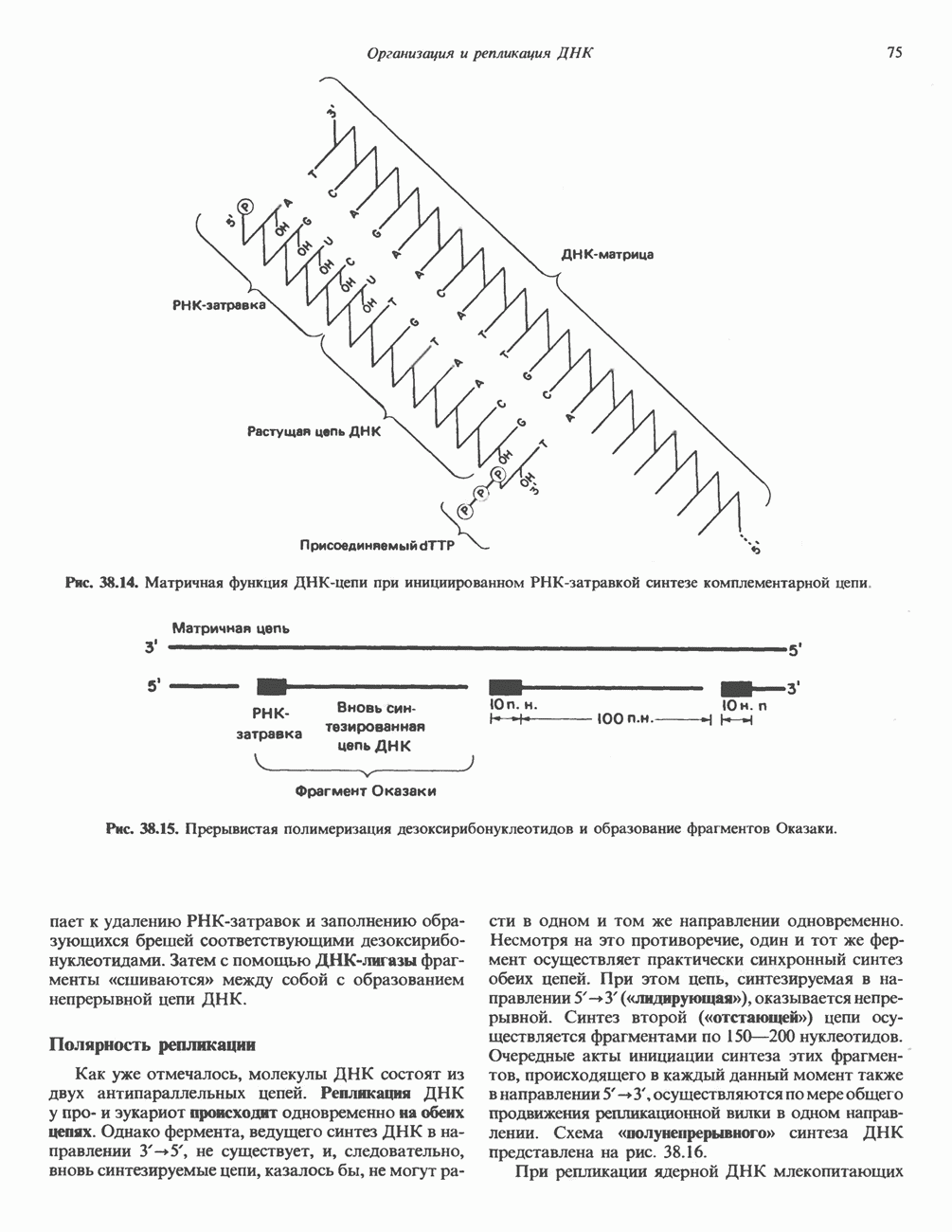
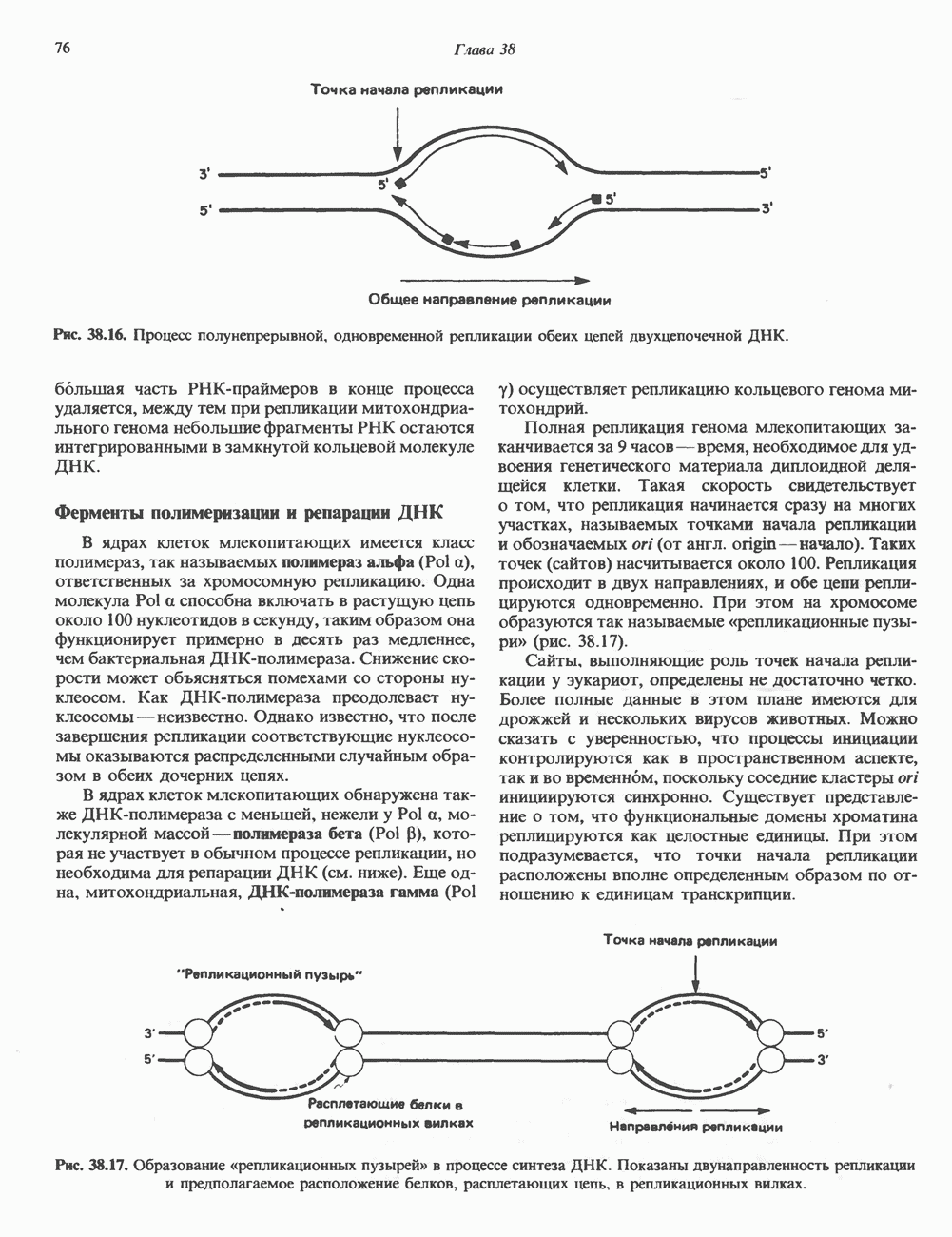
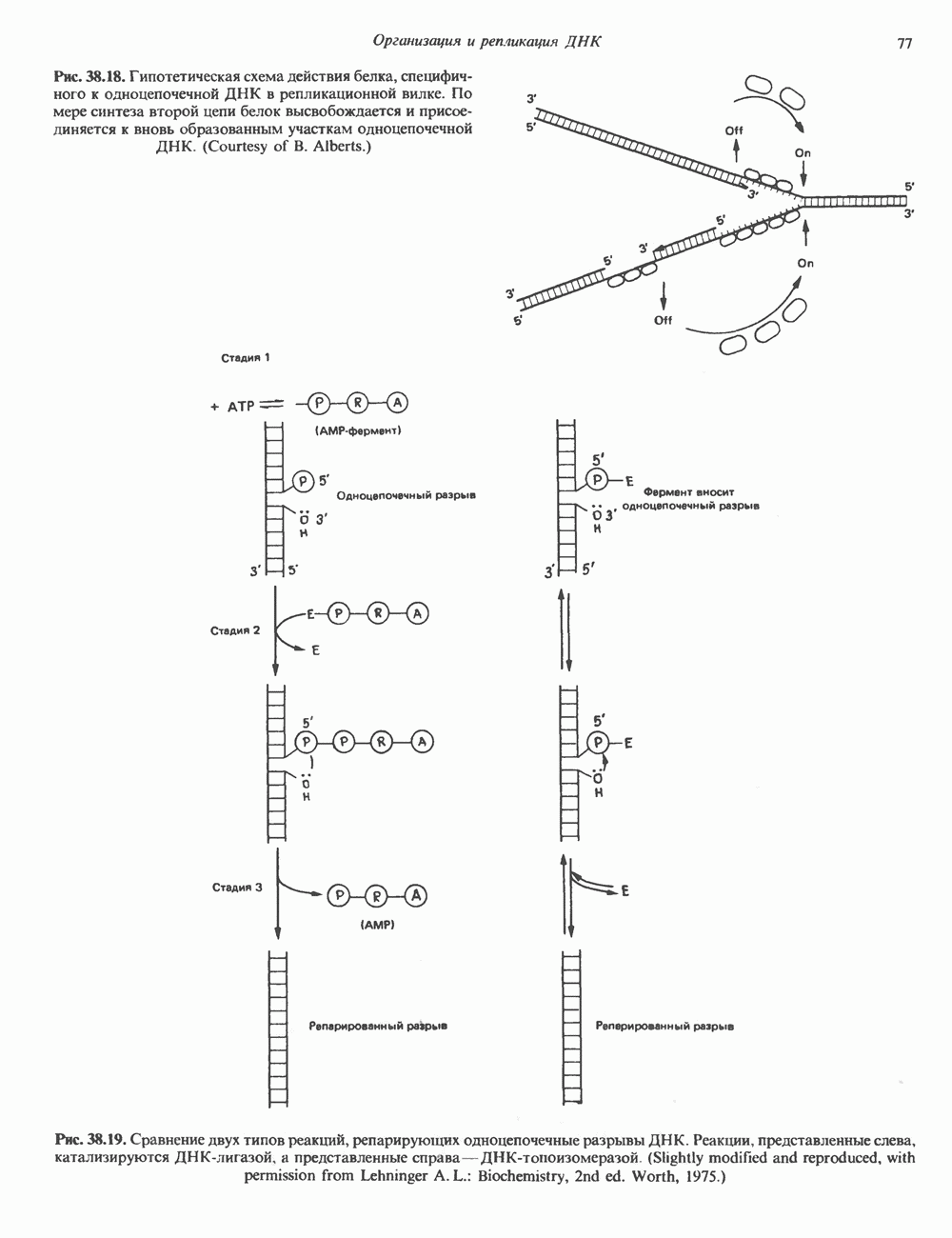
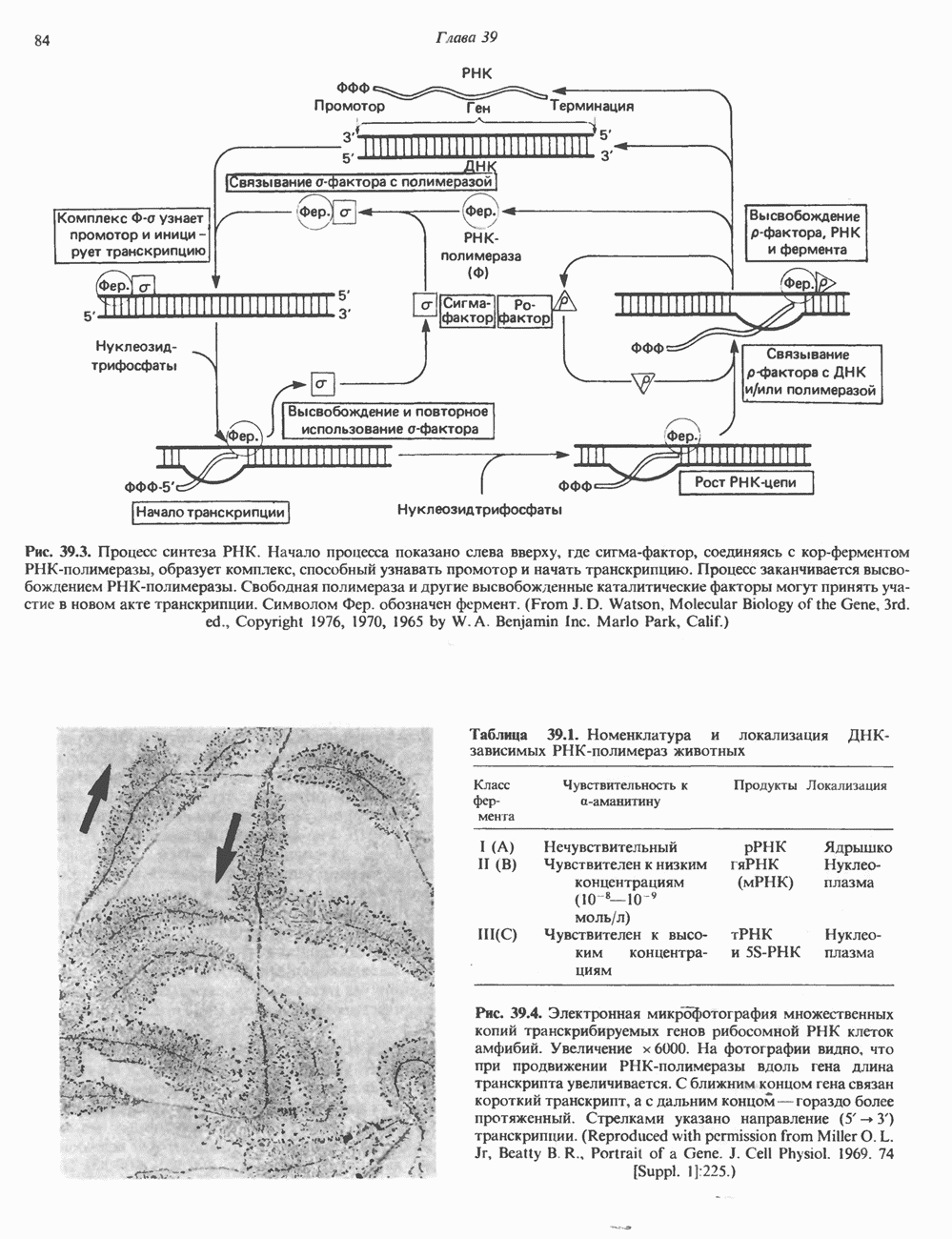
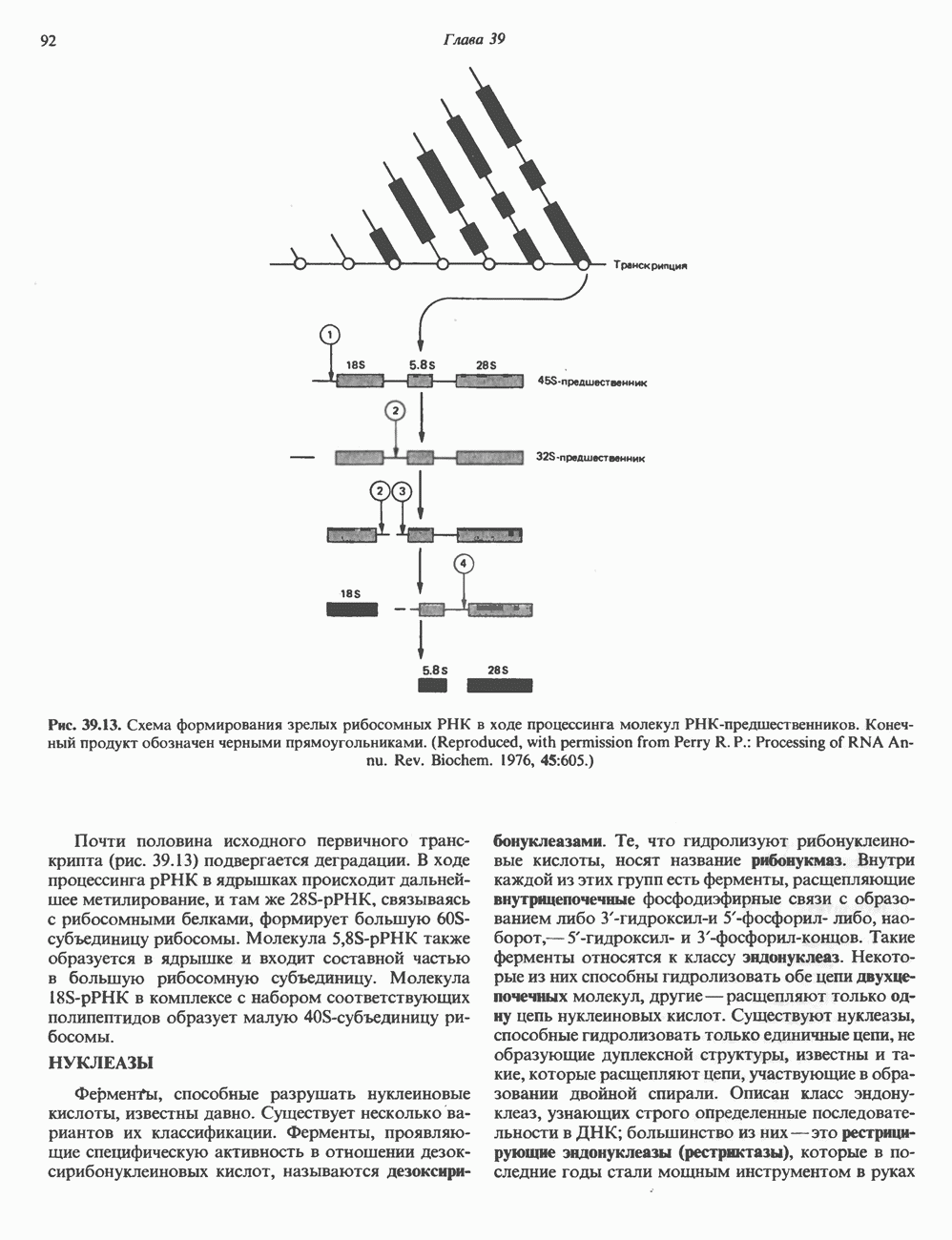
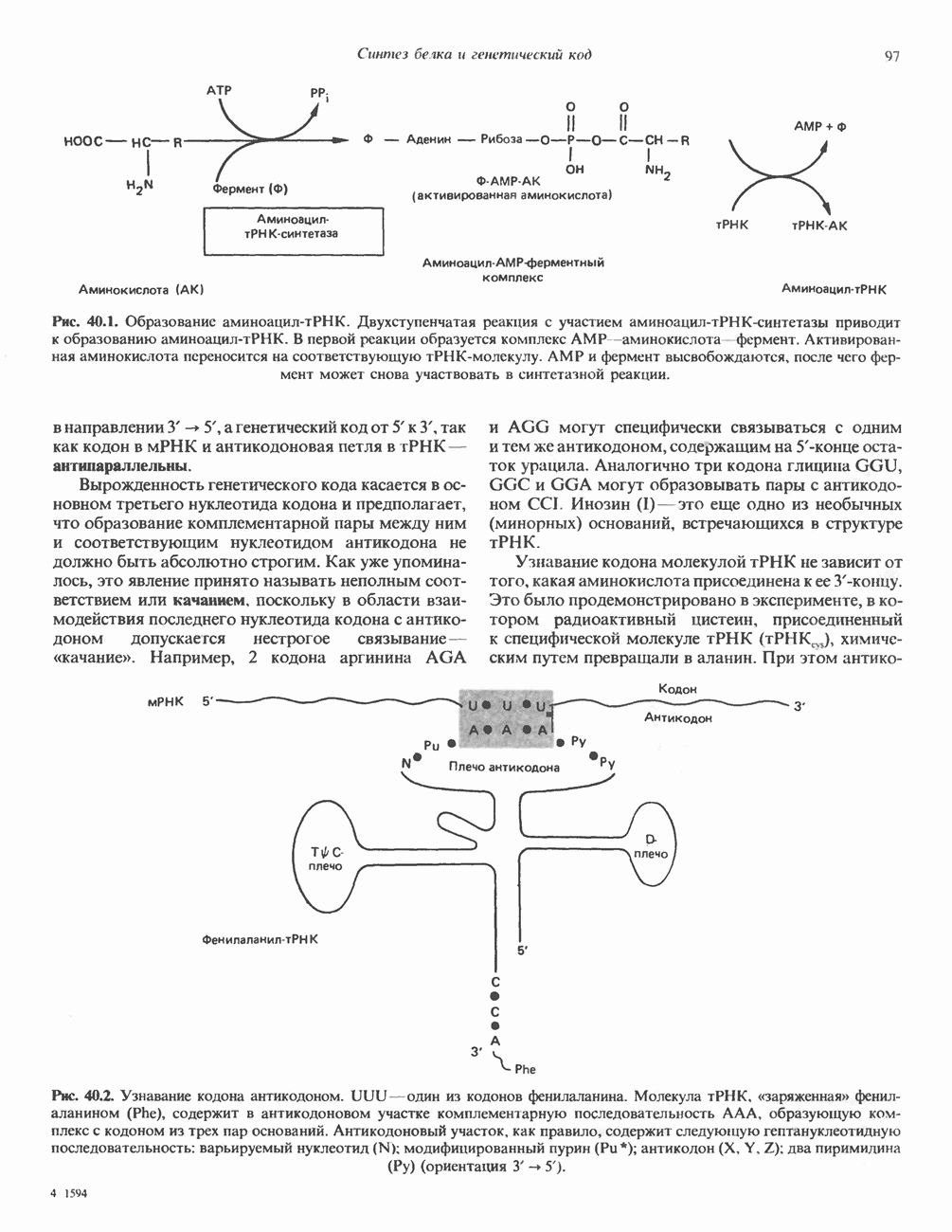
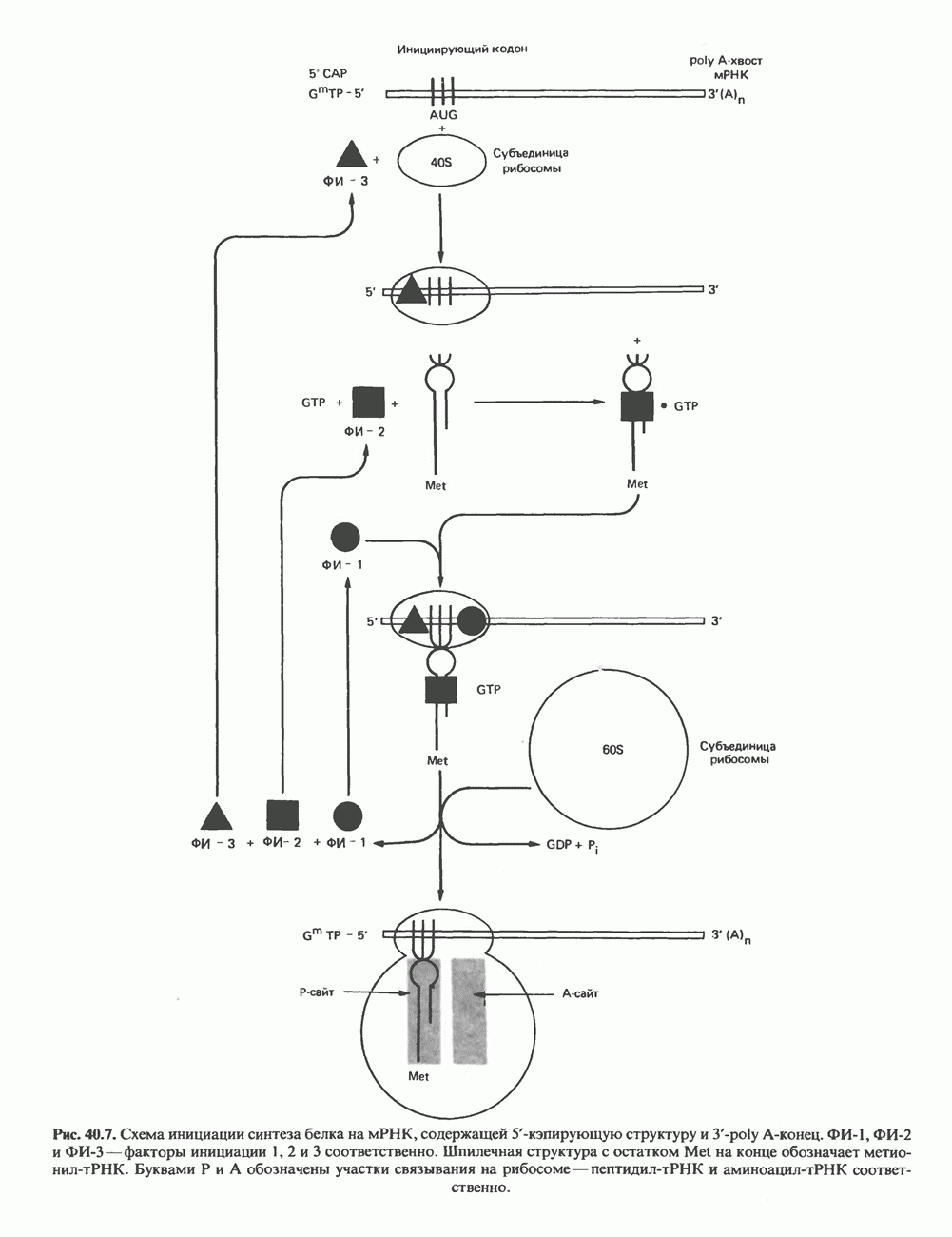
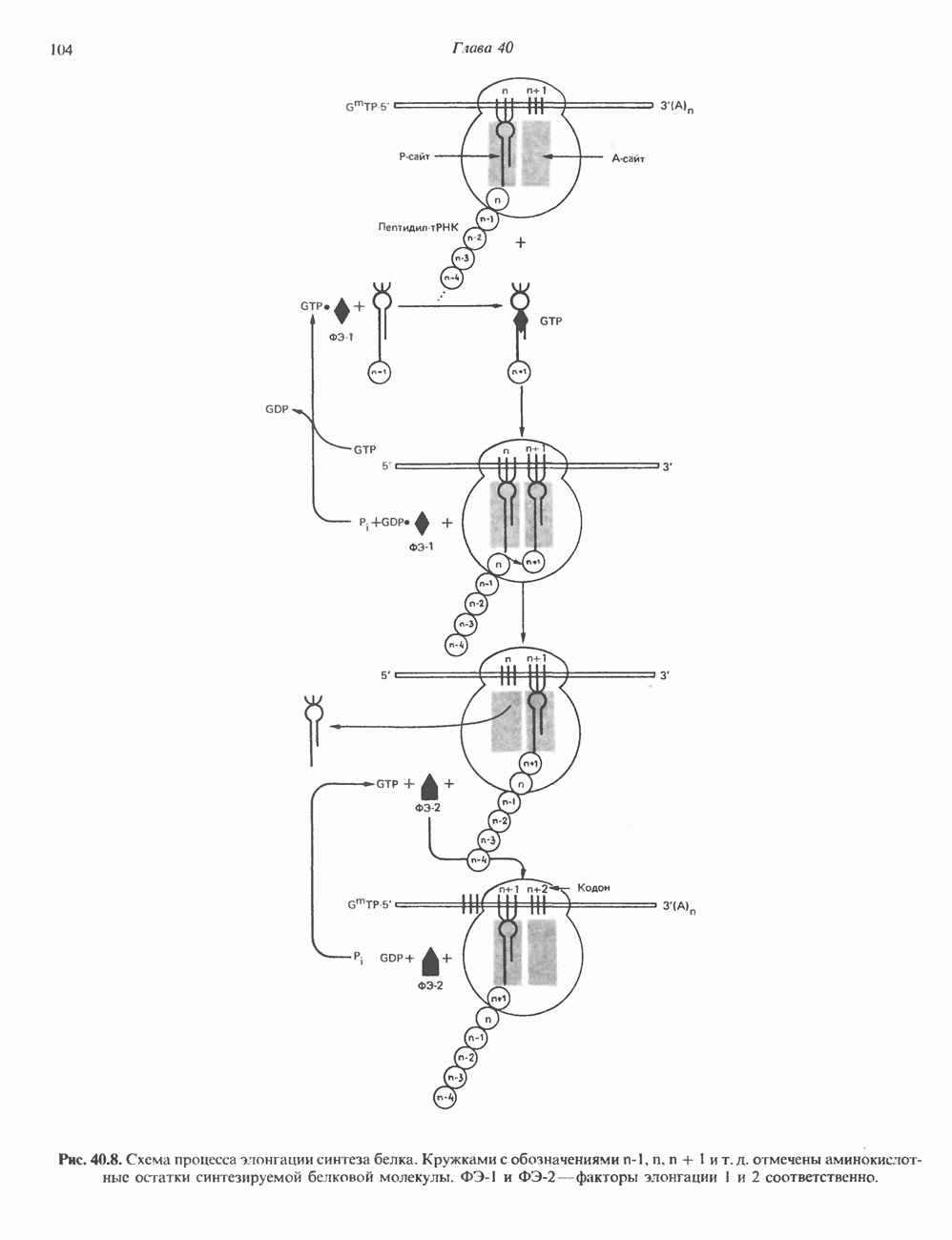
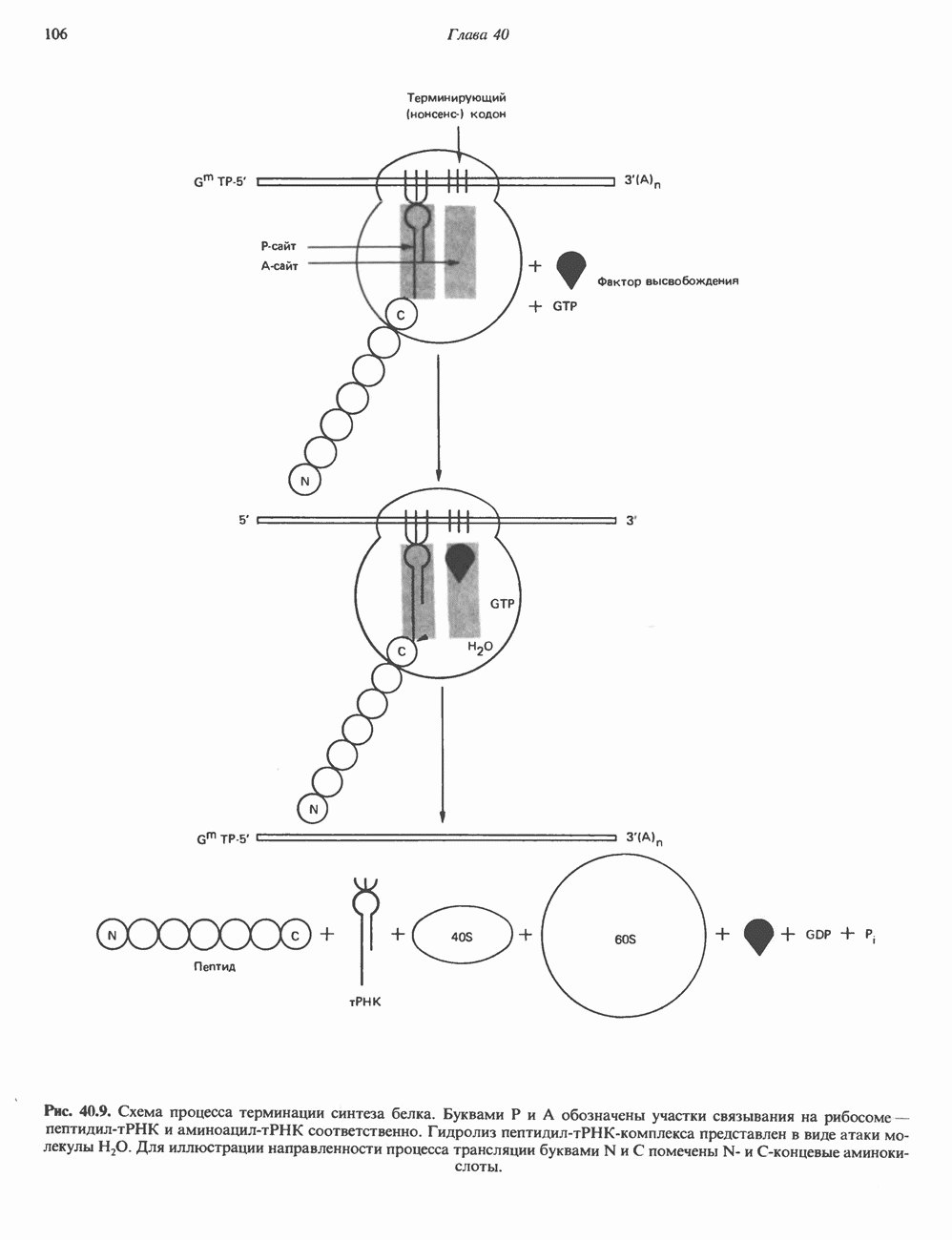
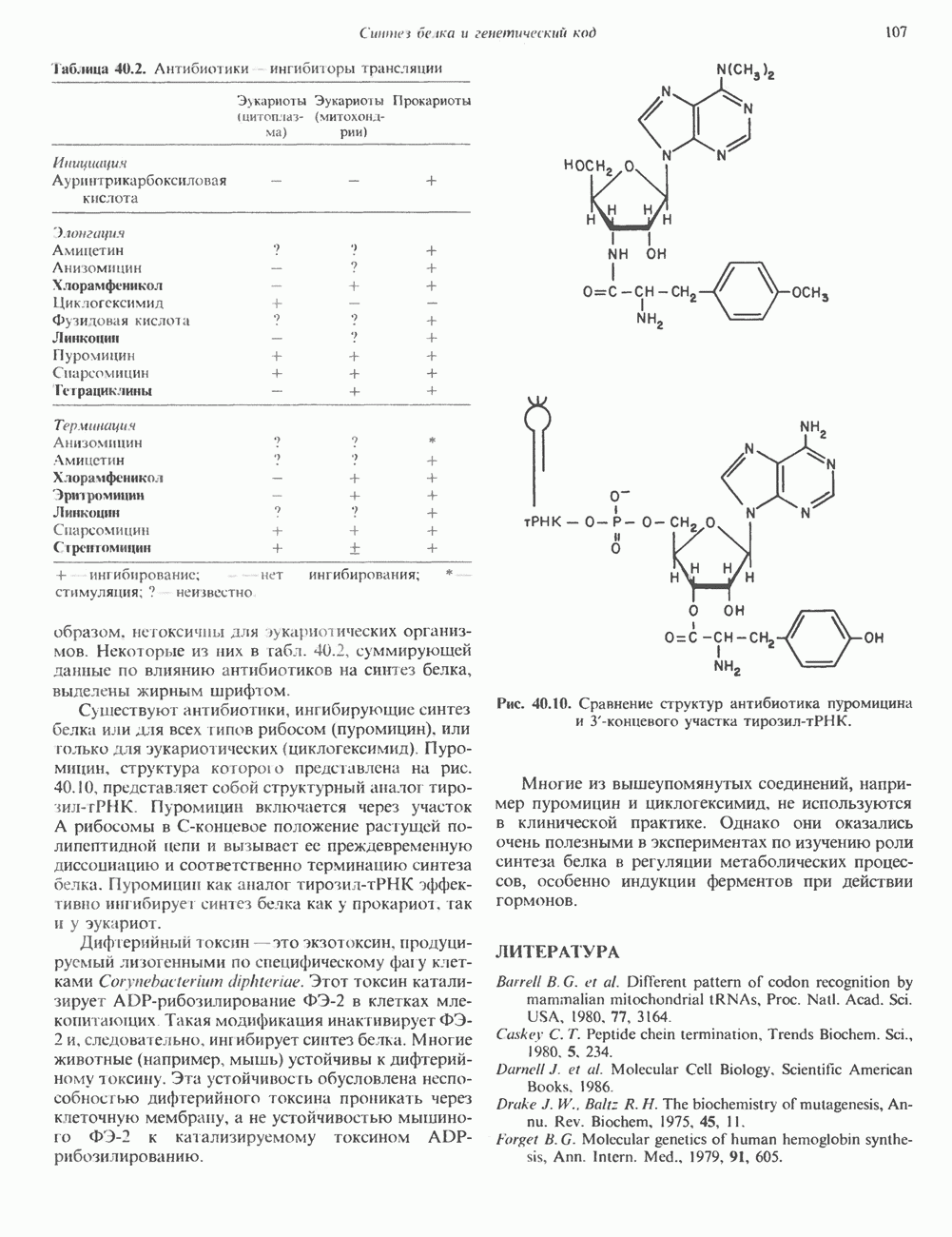
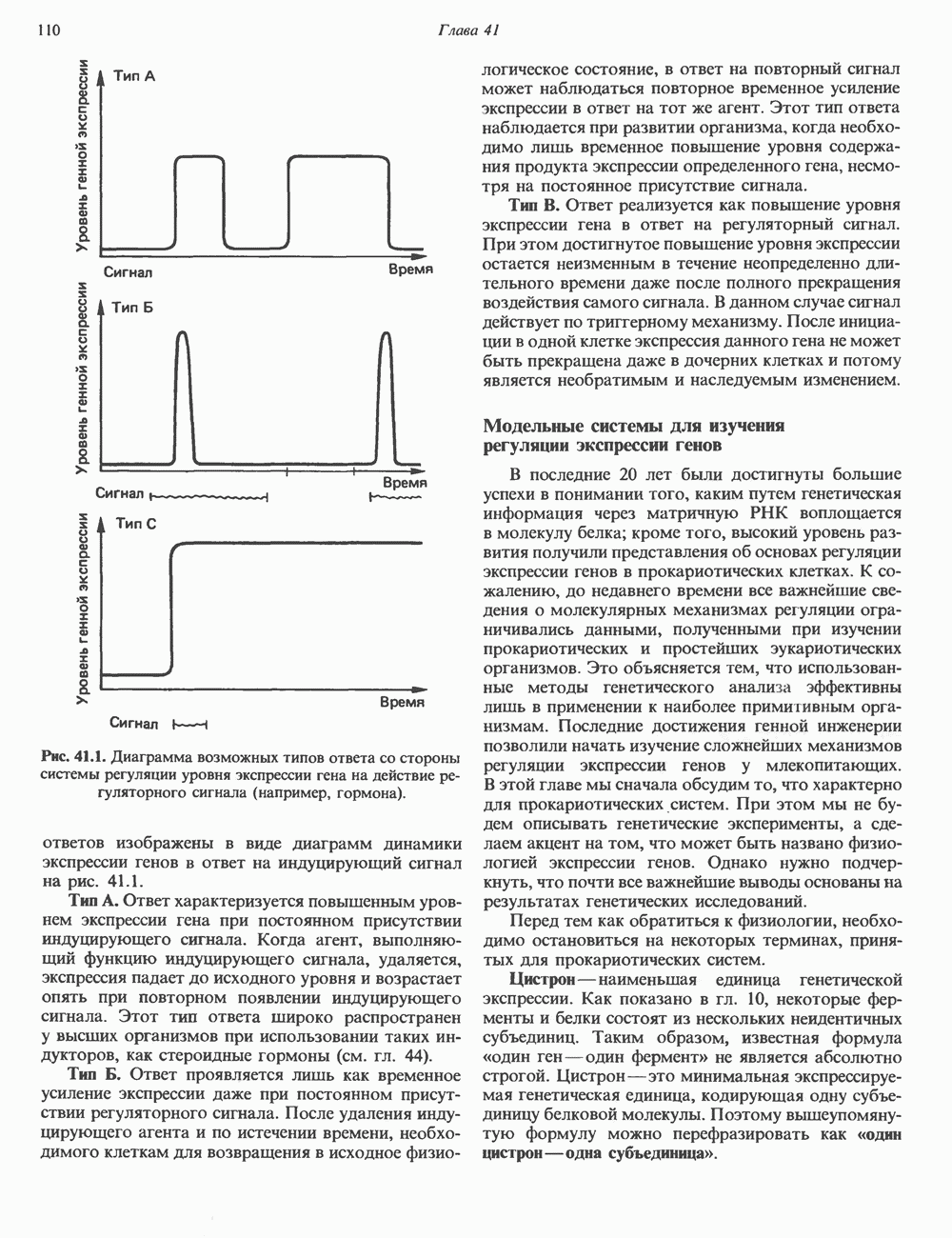
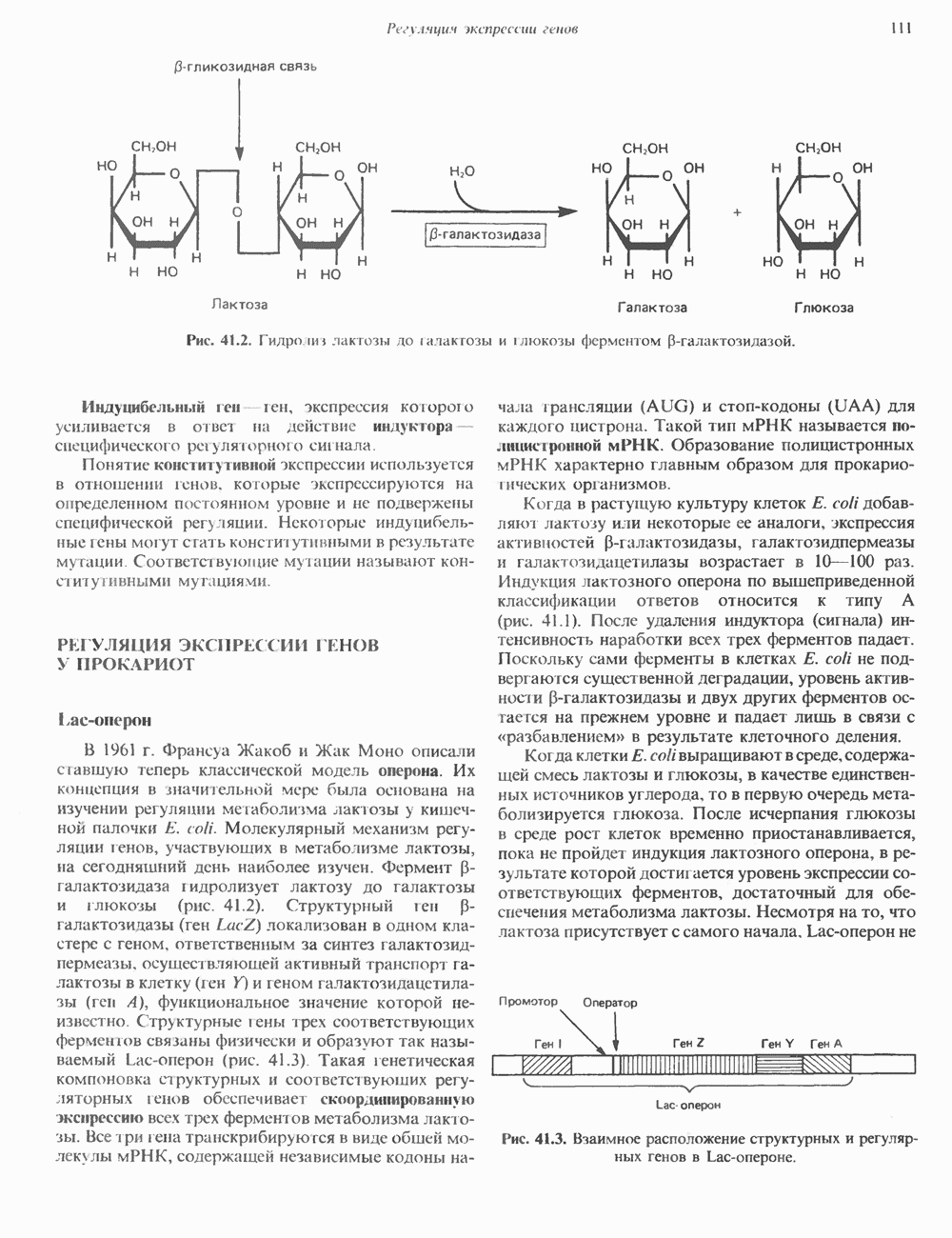
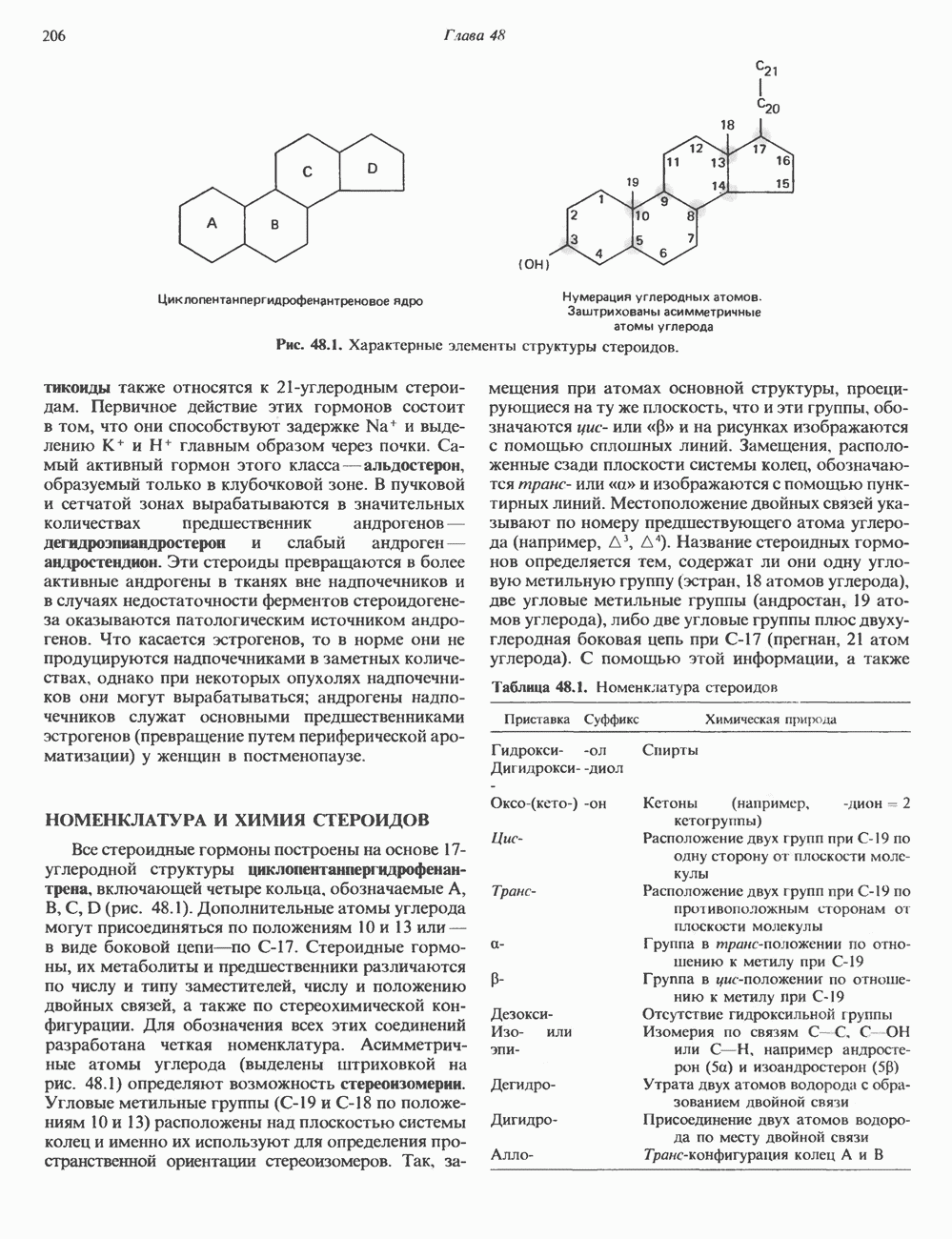
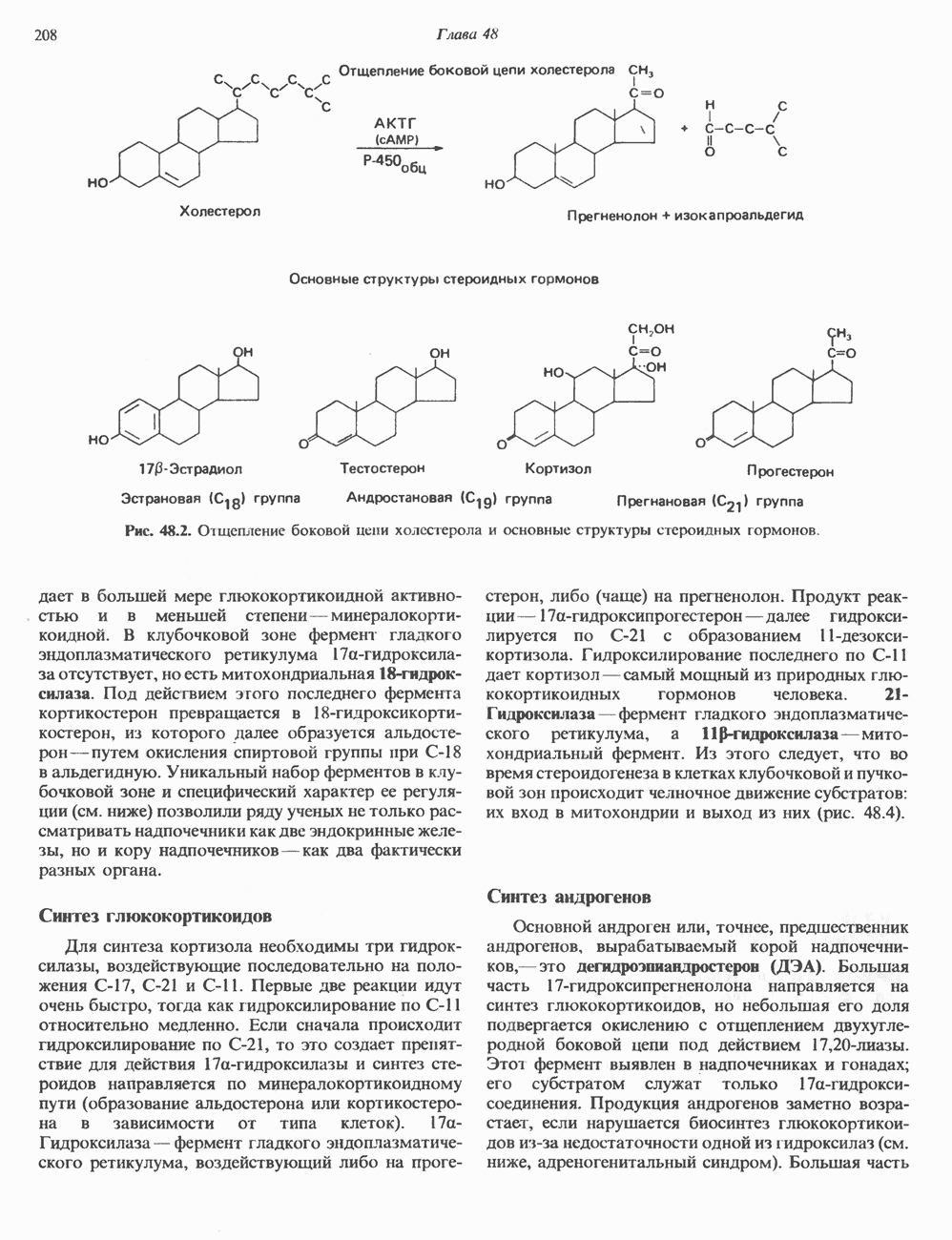
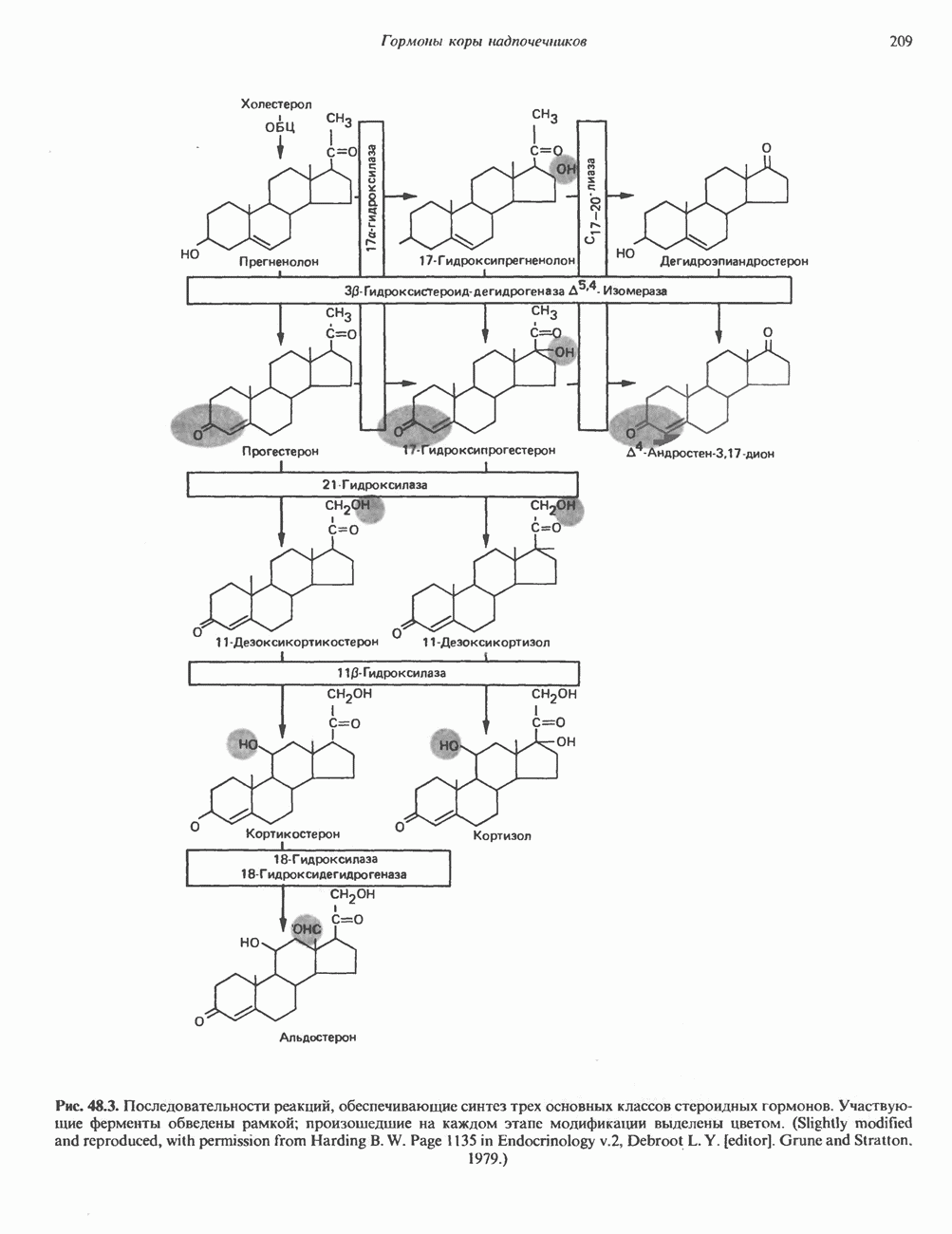
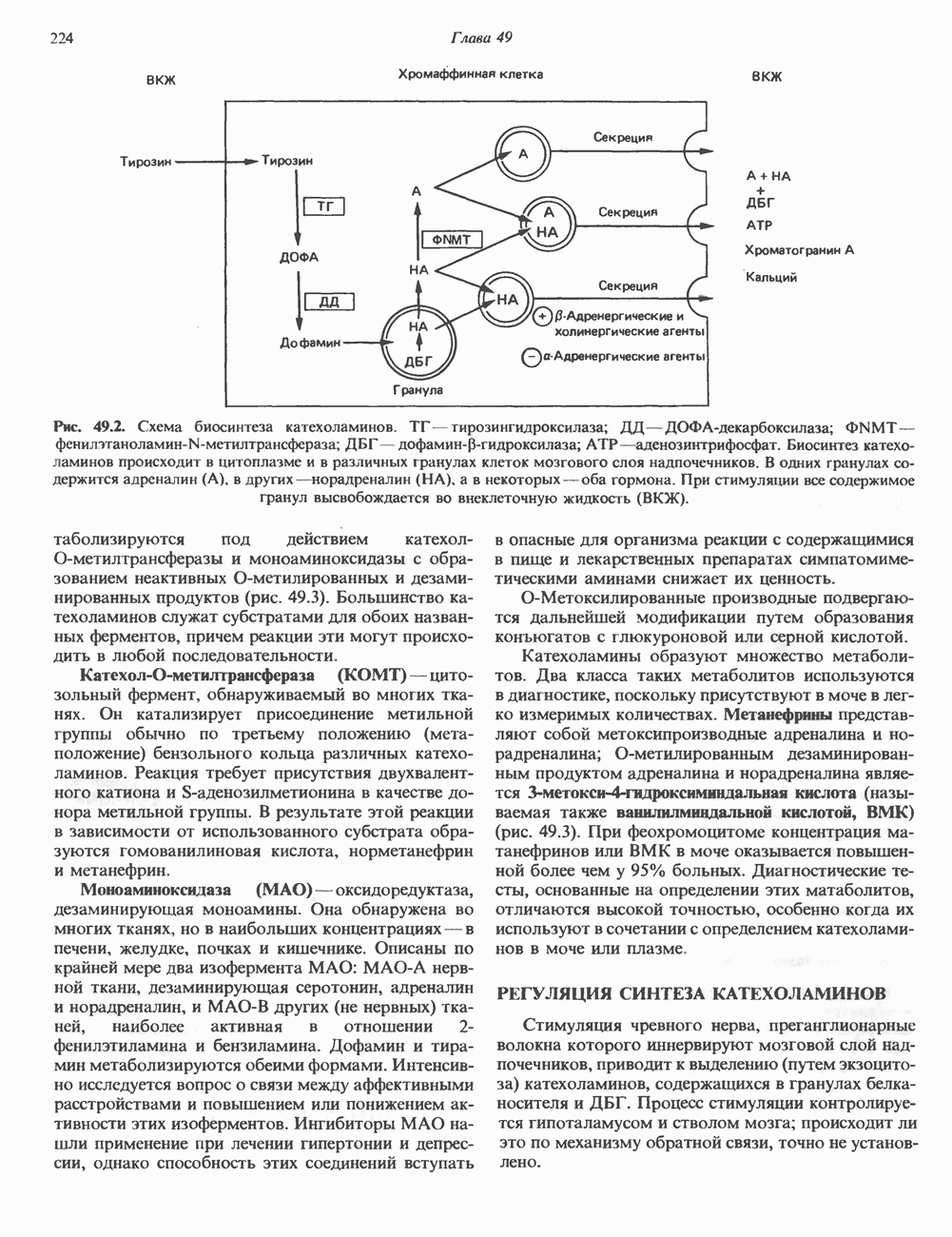
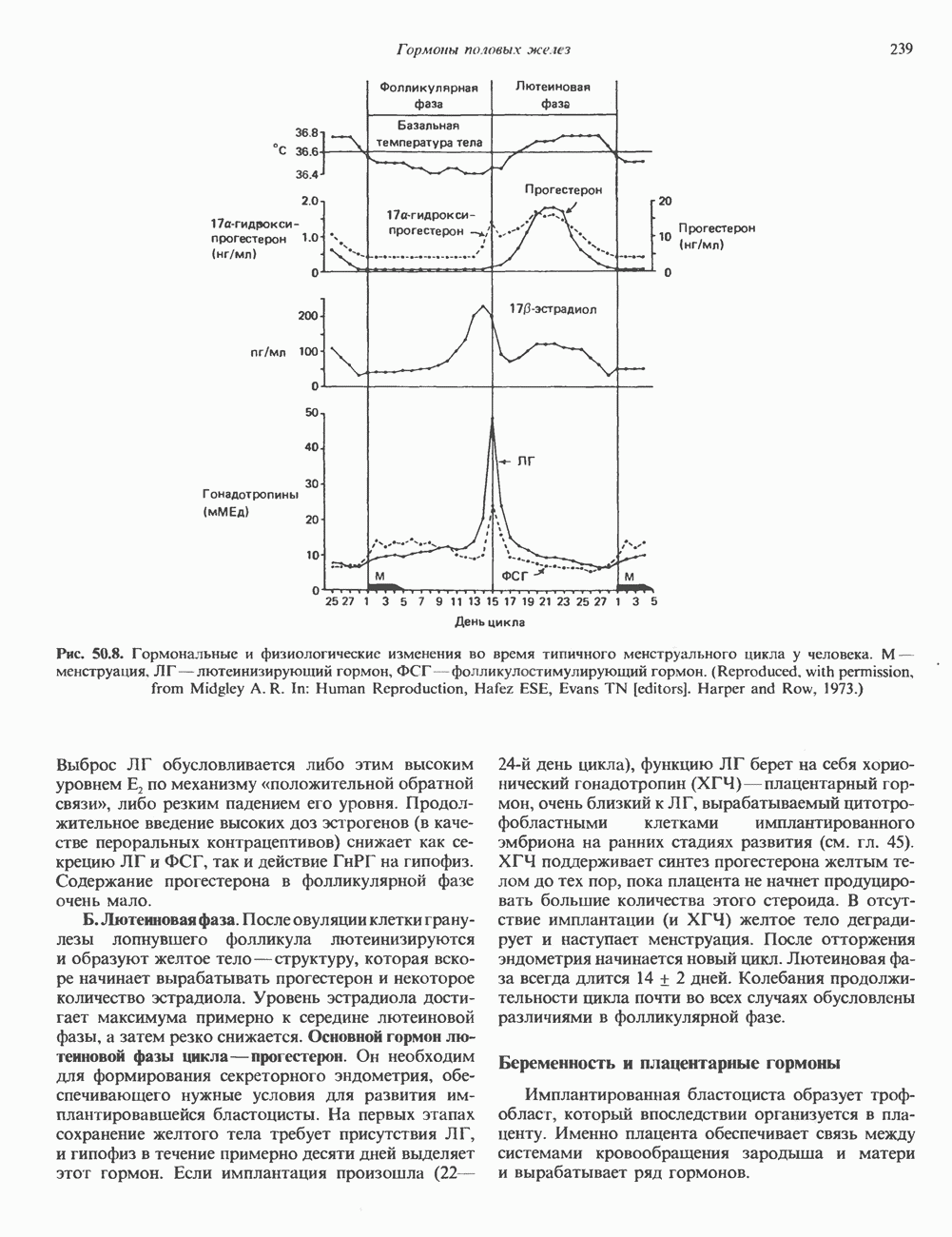
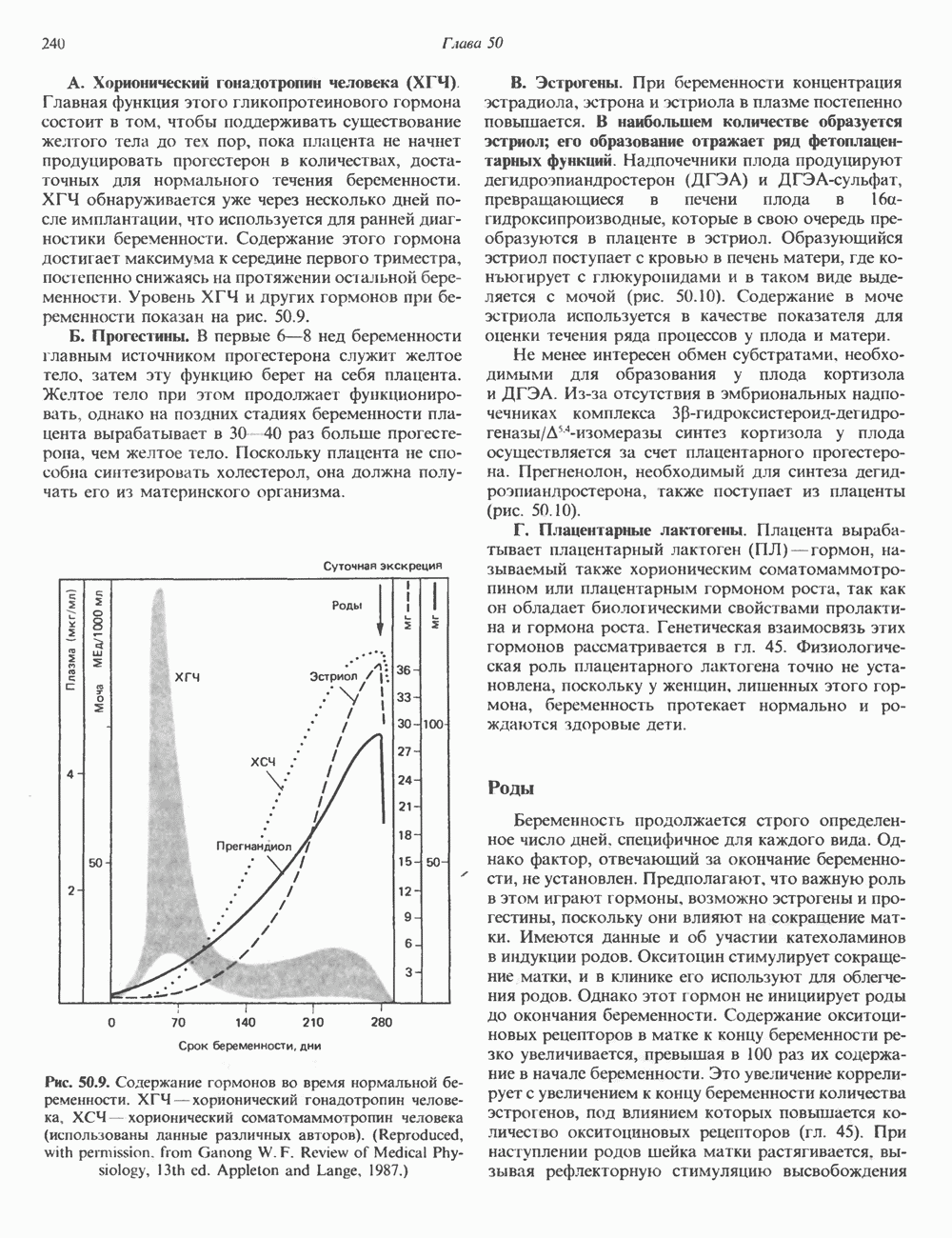
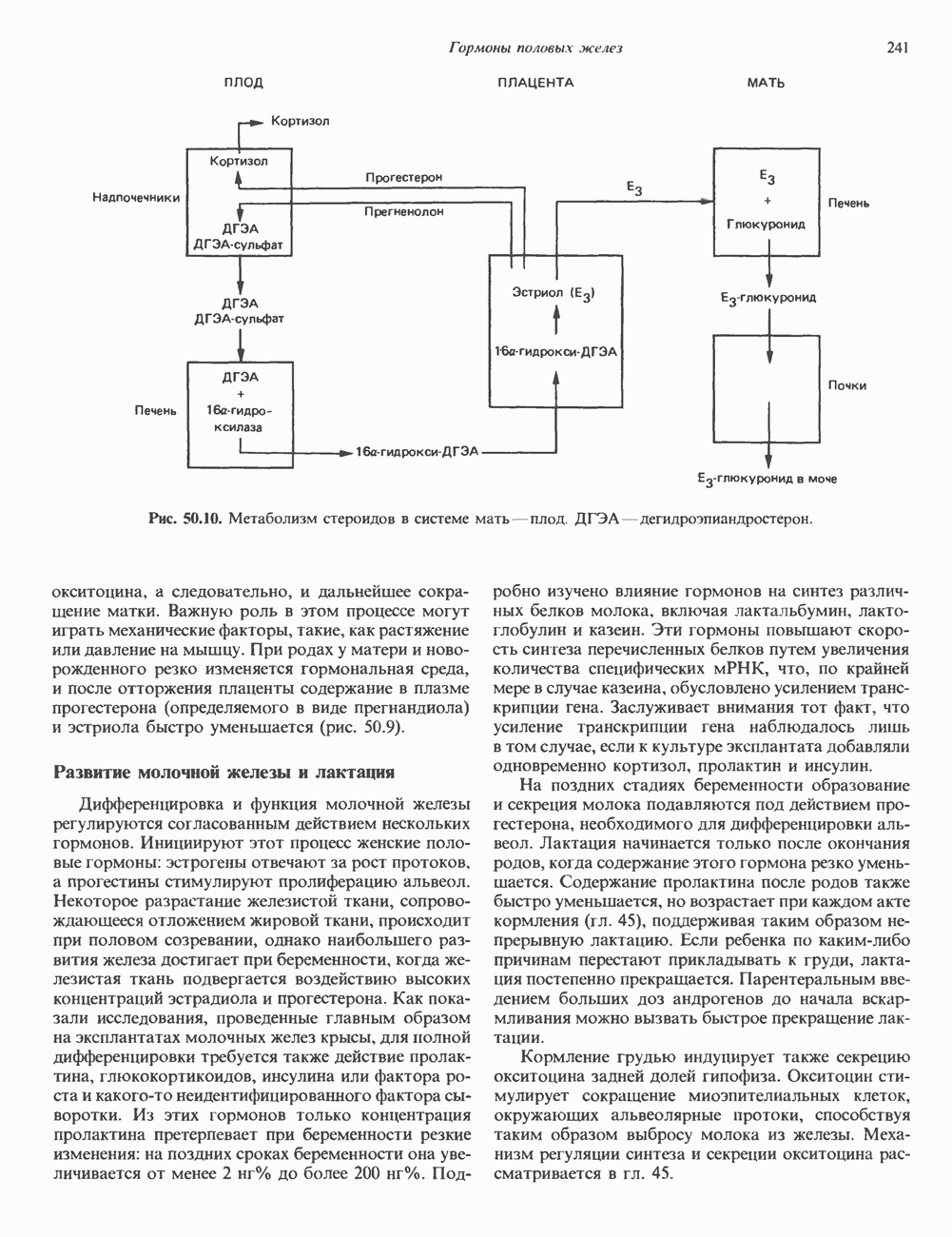
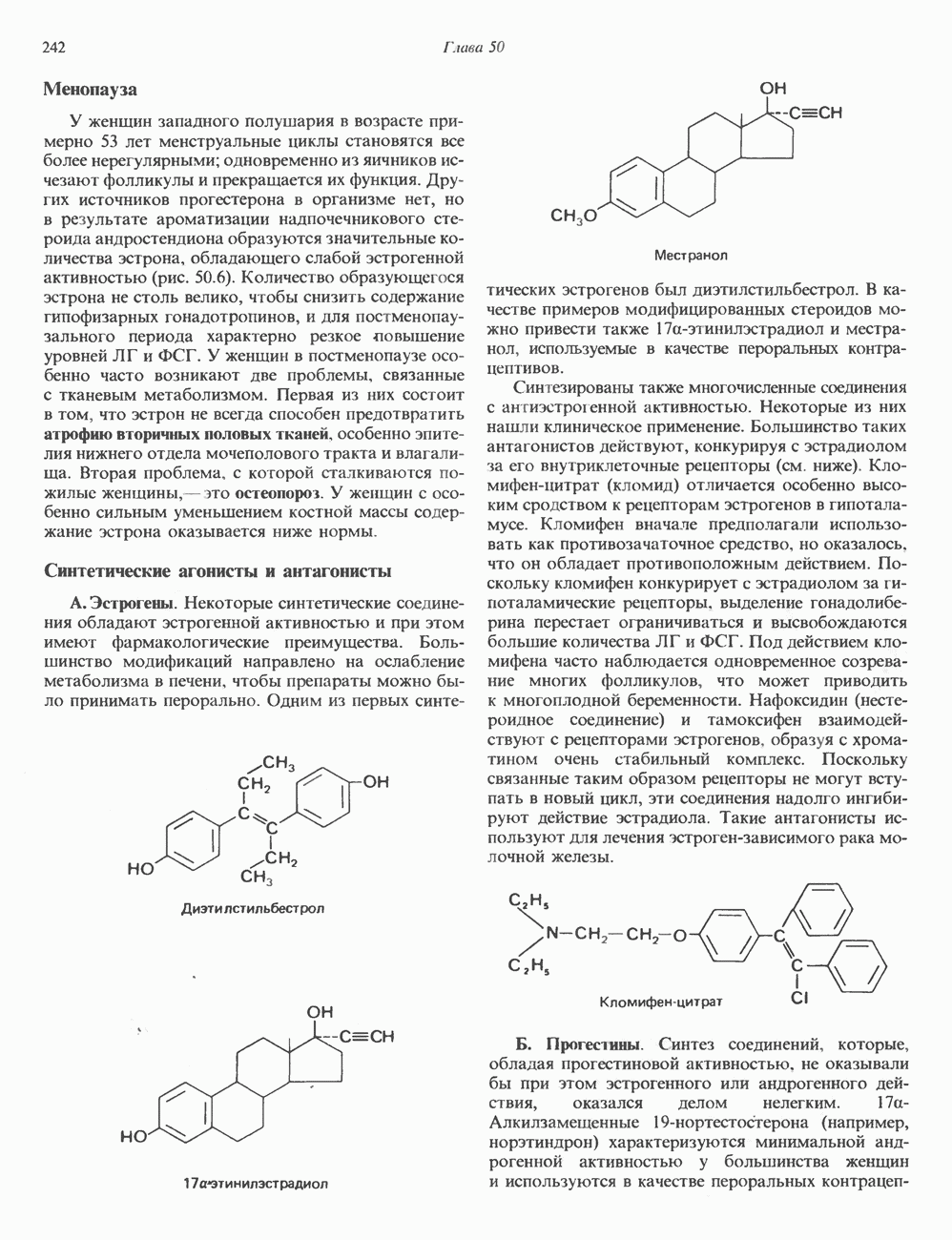
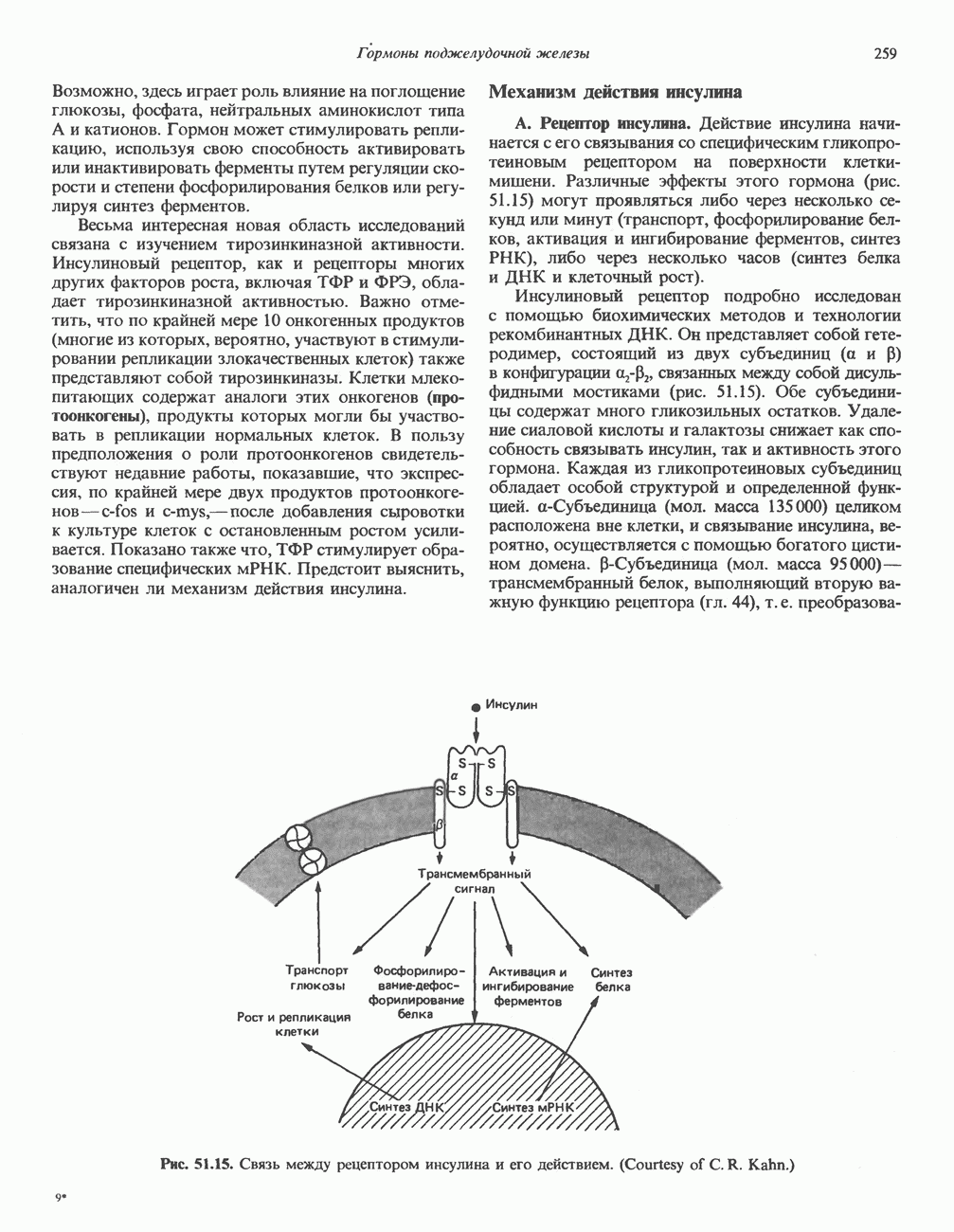
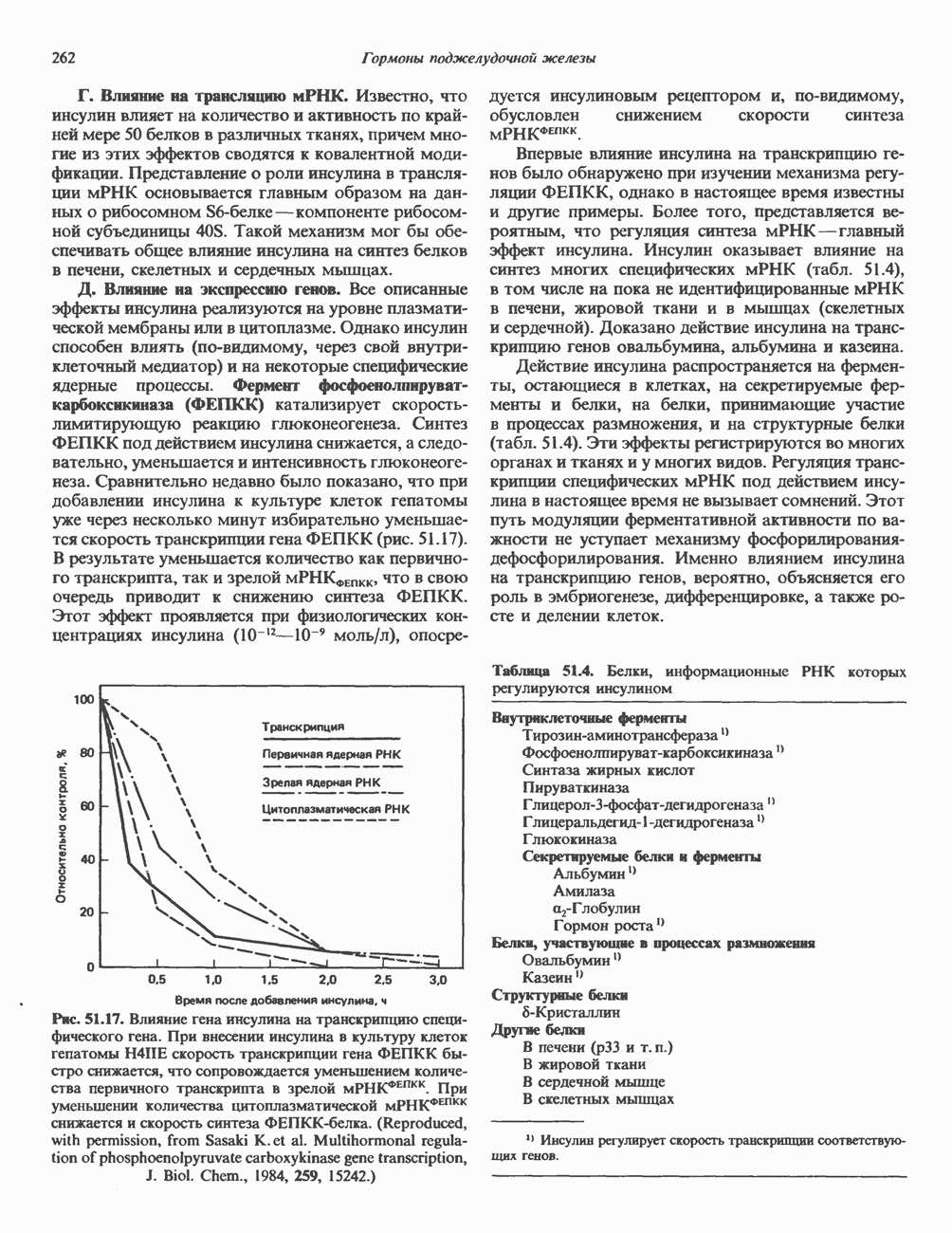
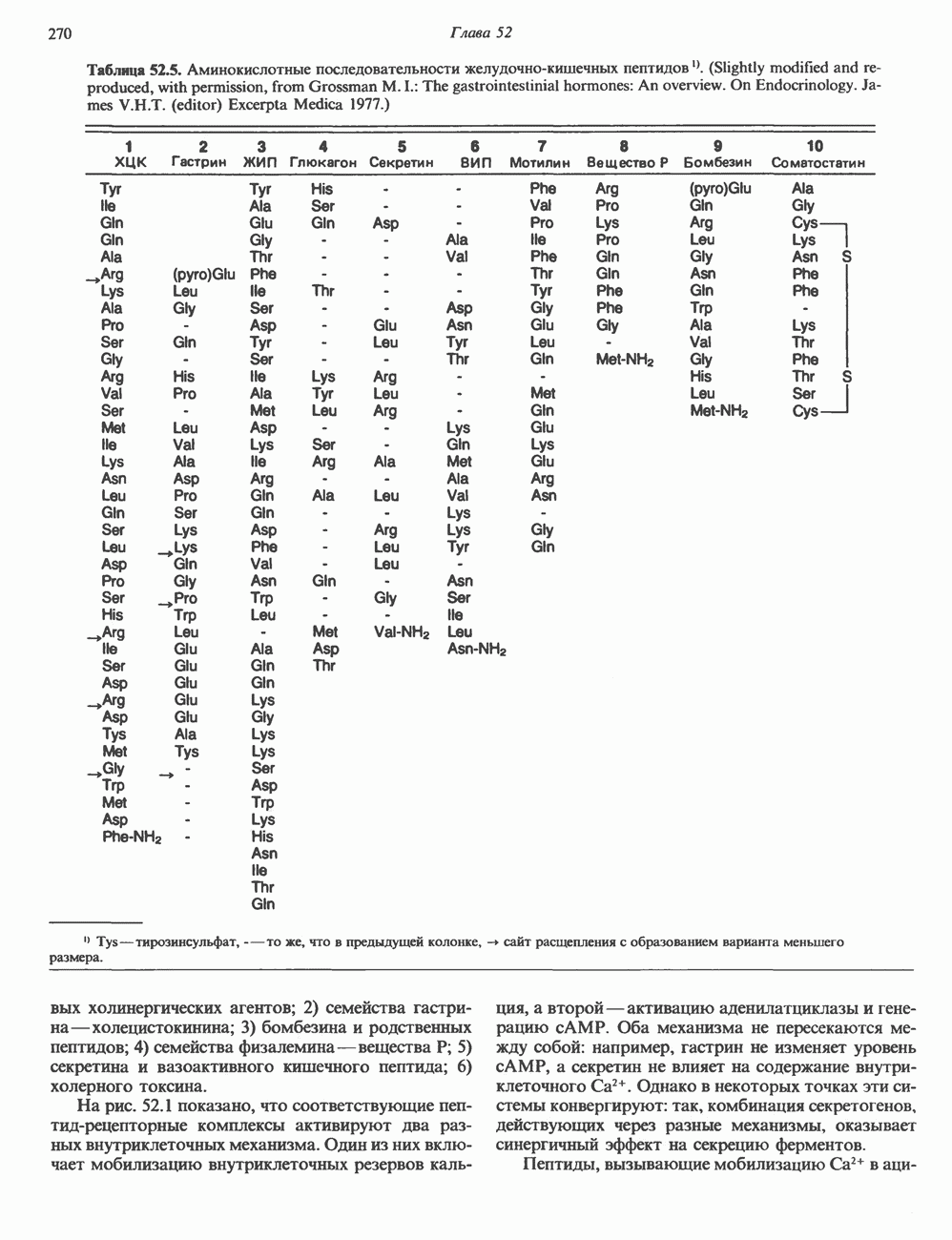
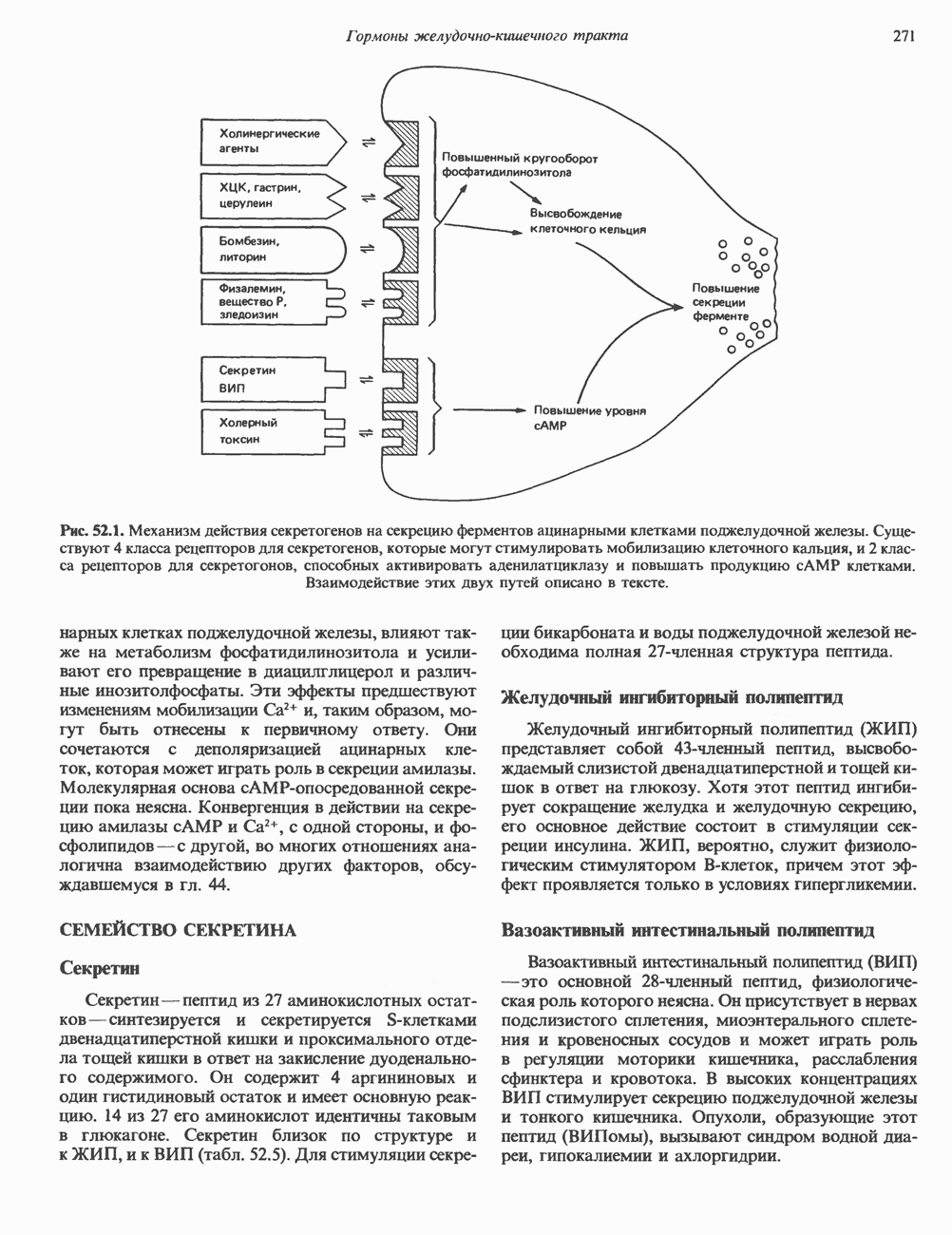
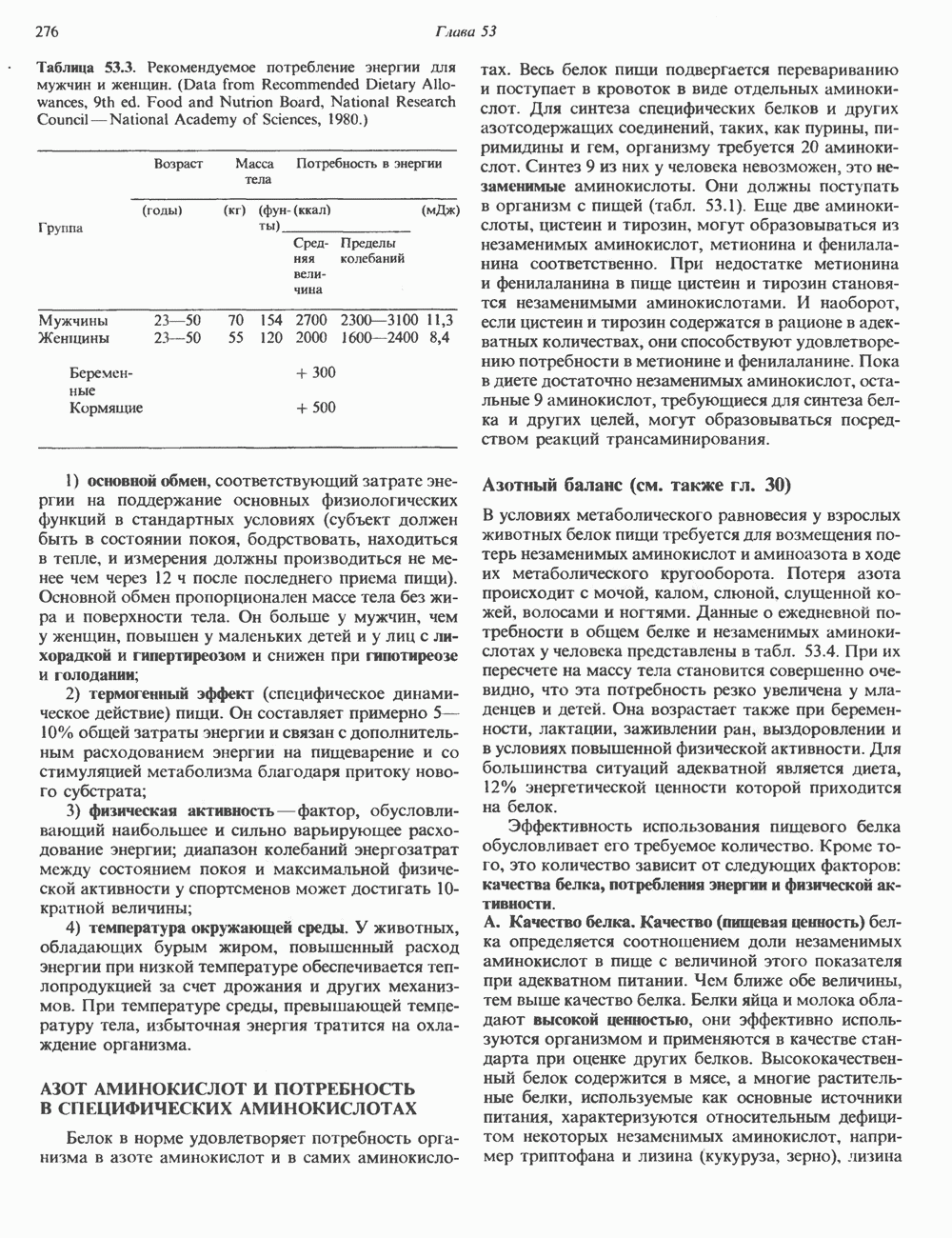
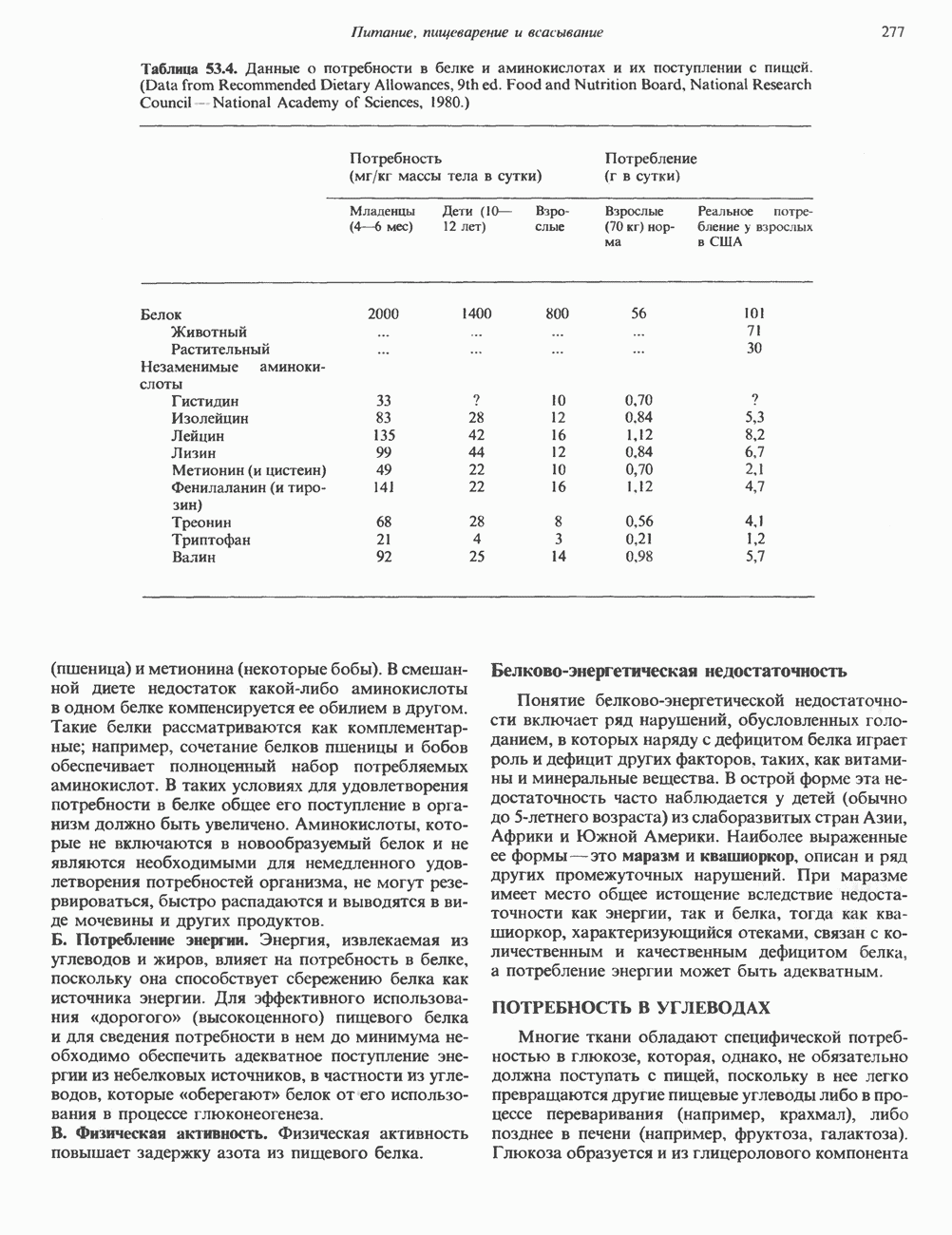
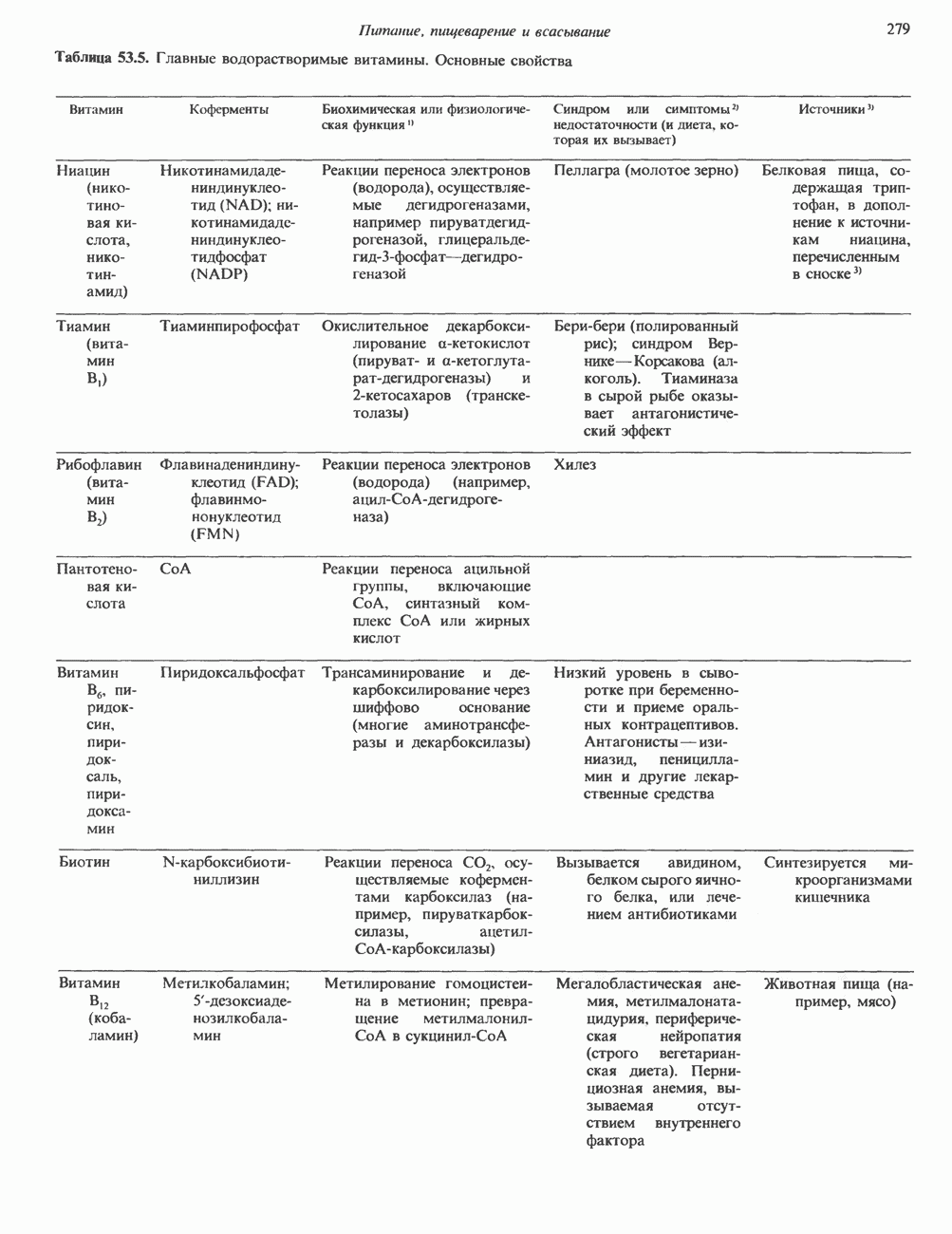
Пред.
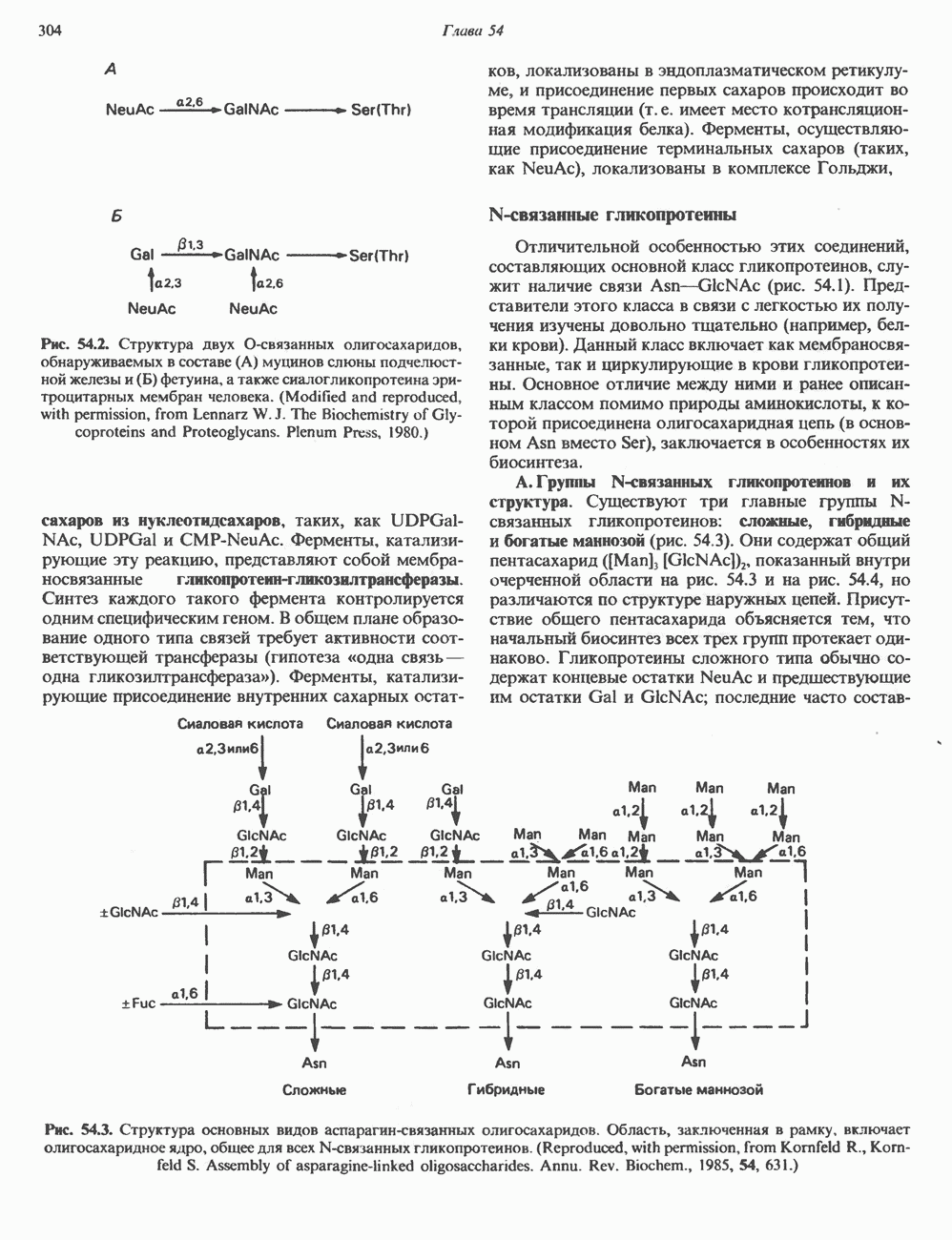
След.
Макеты страниц
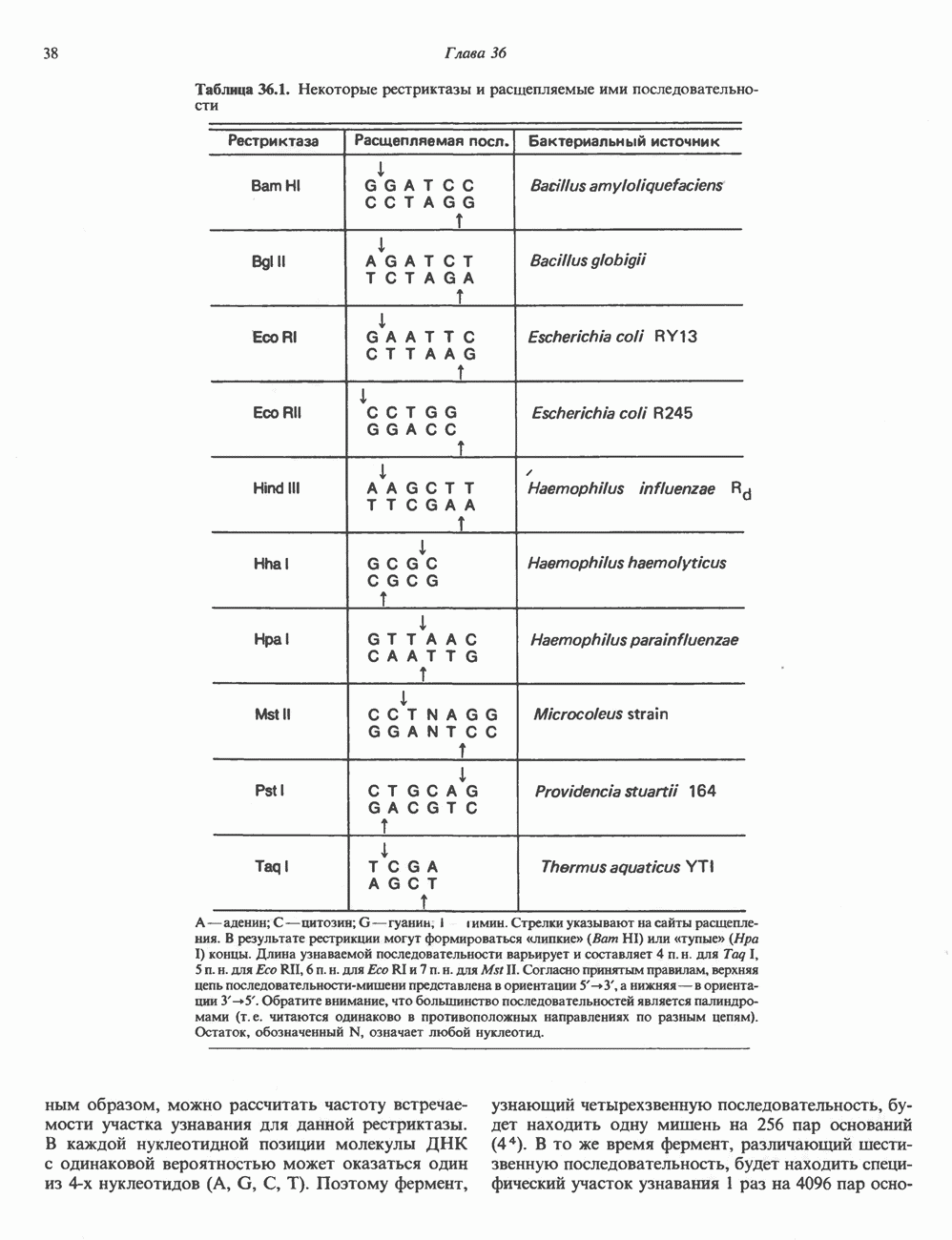
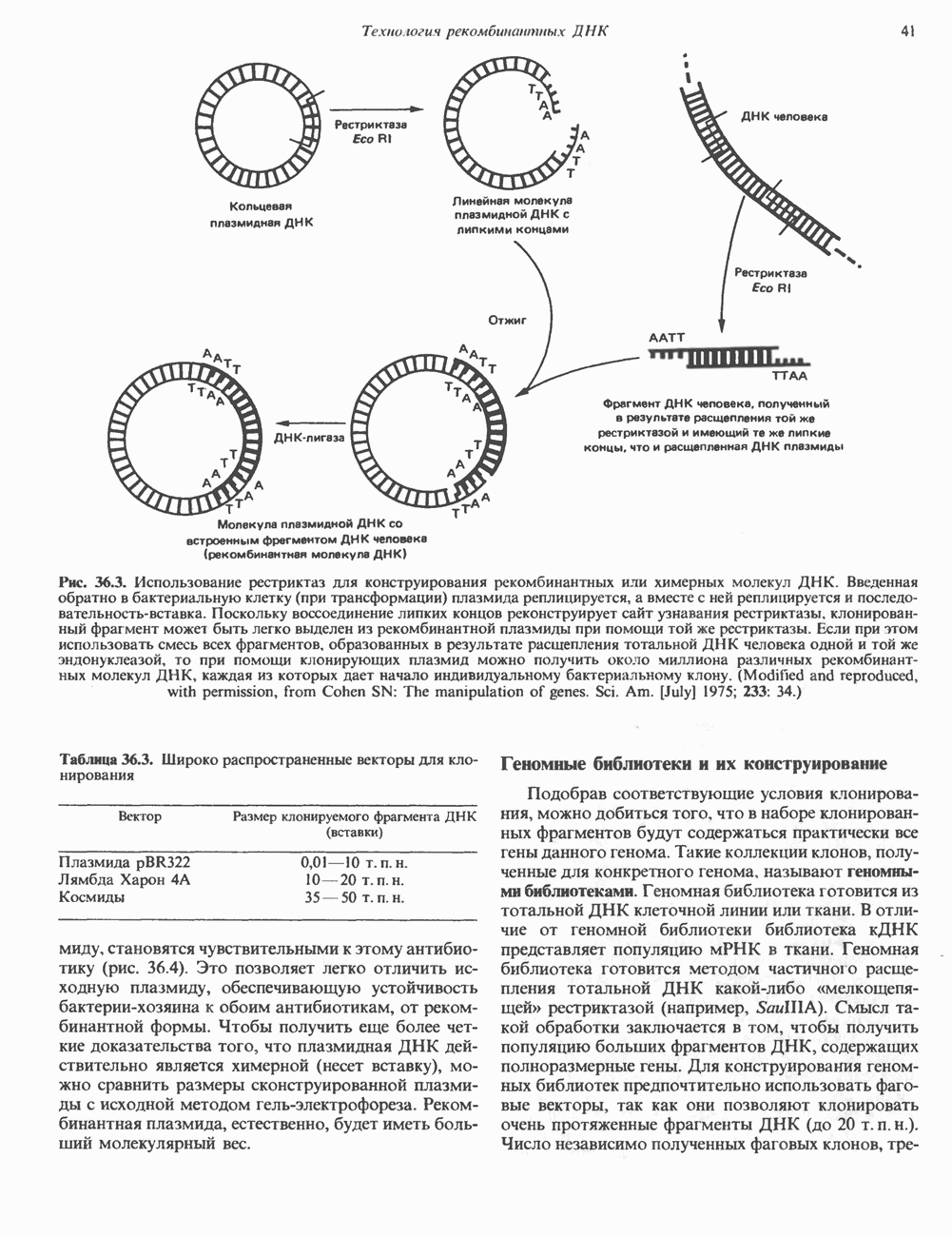
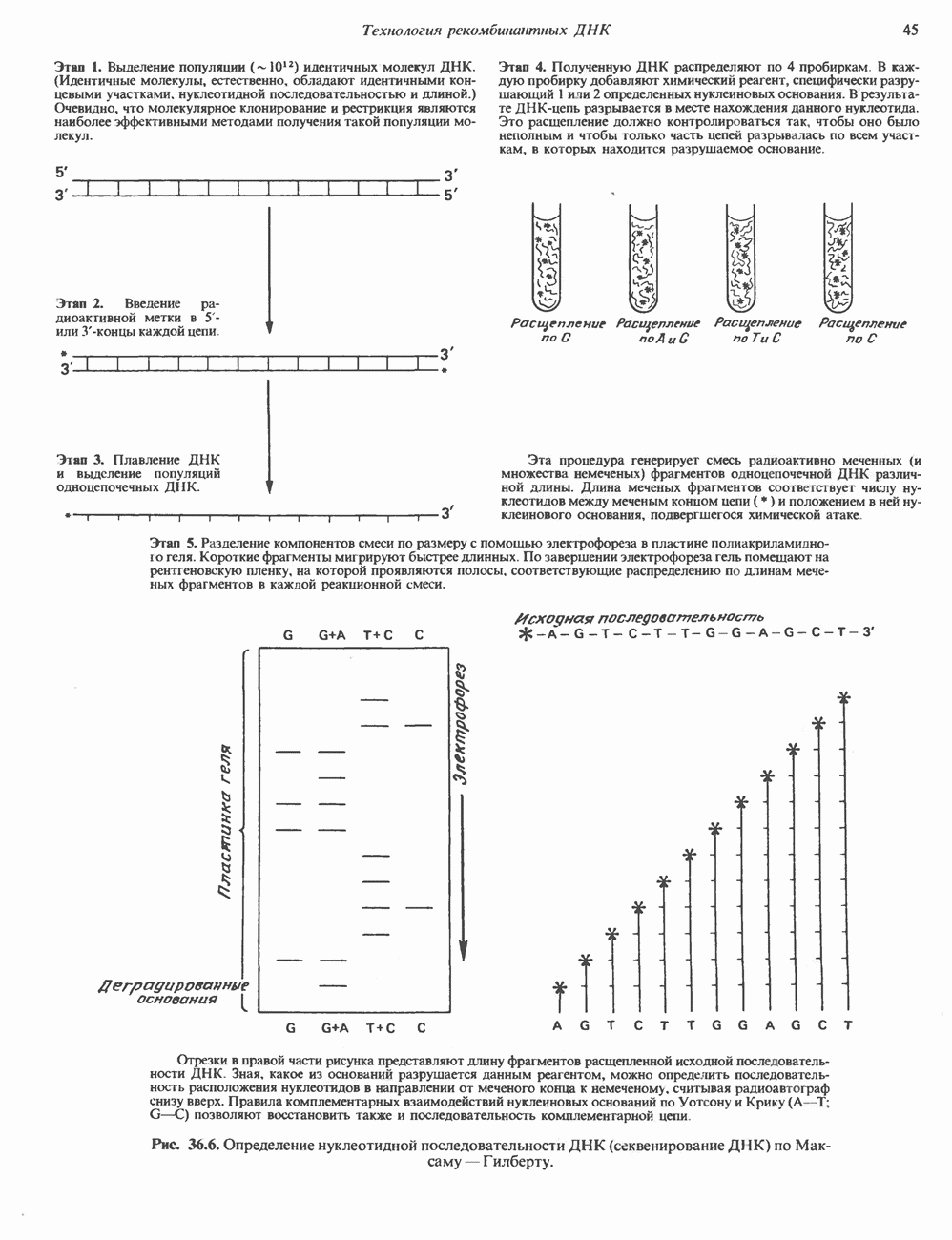
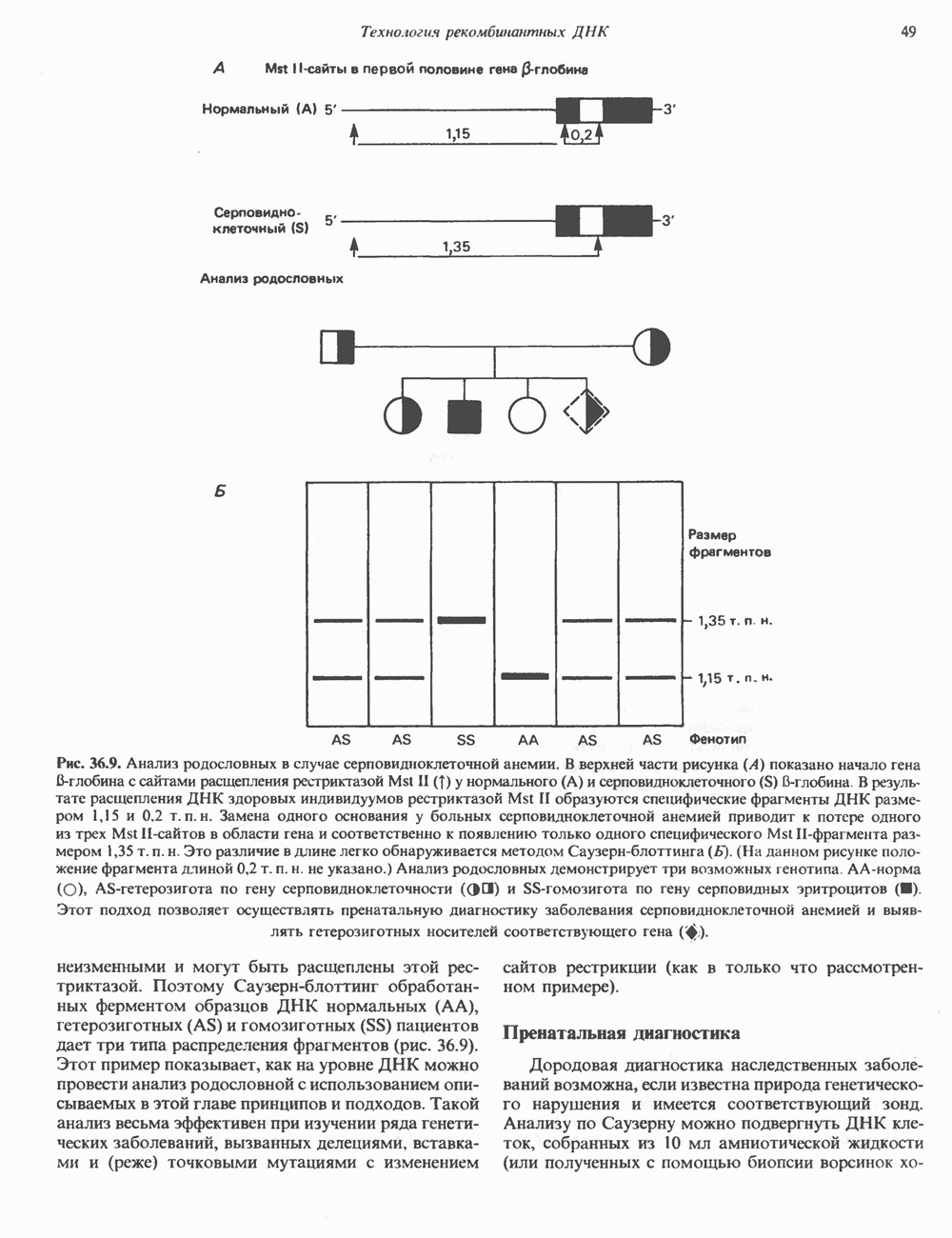
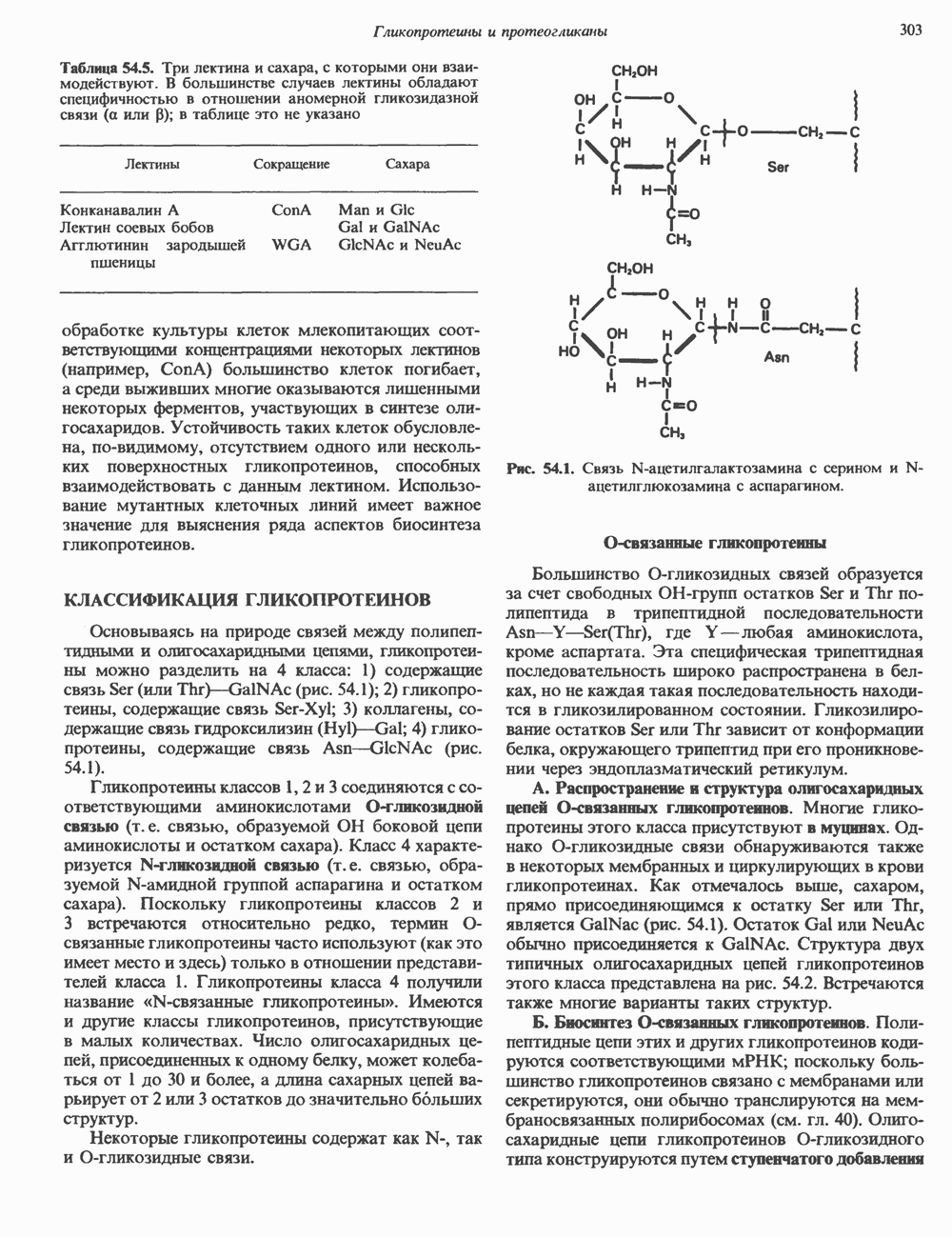
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
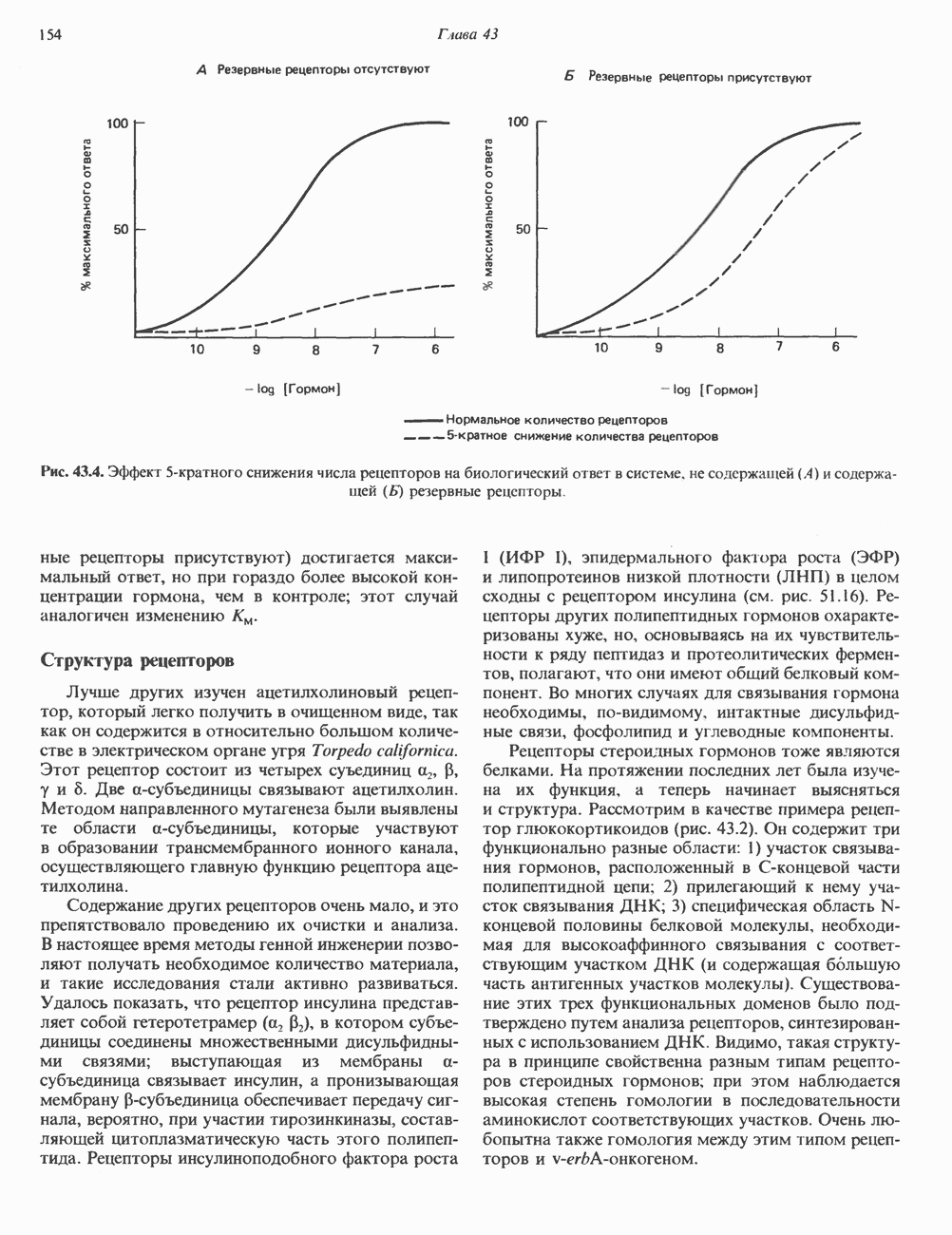
ZADANIA.TO
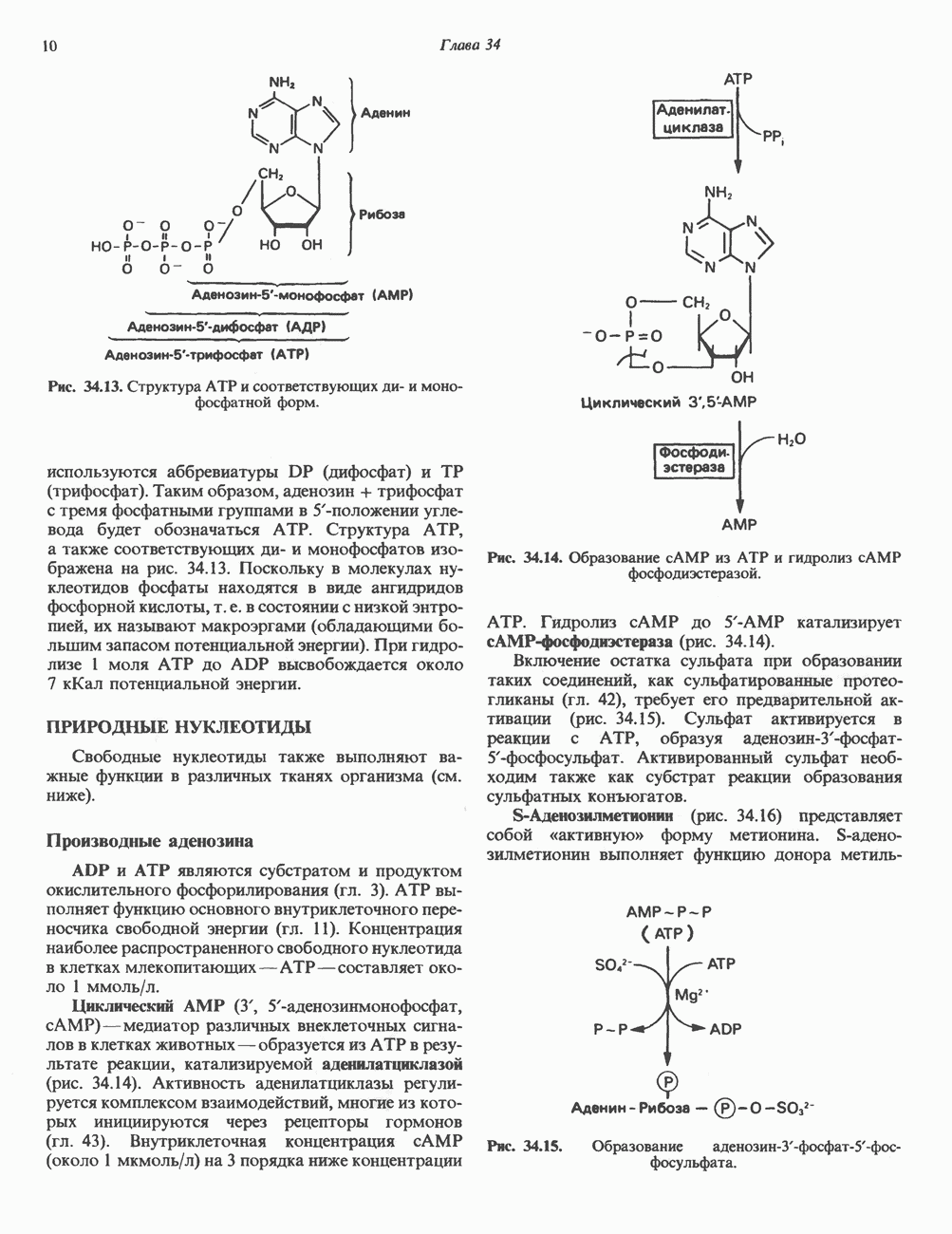
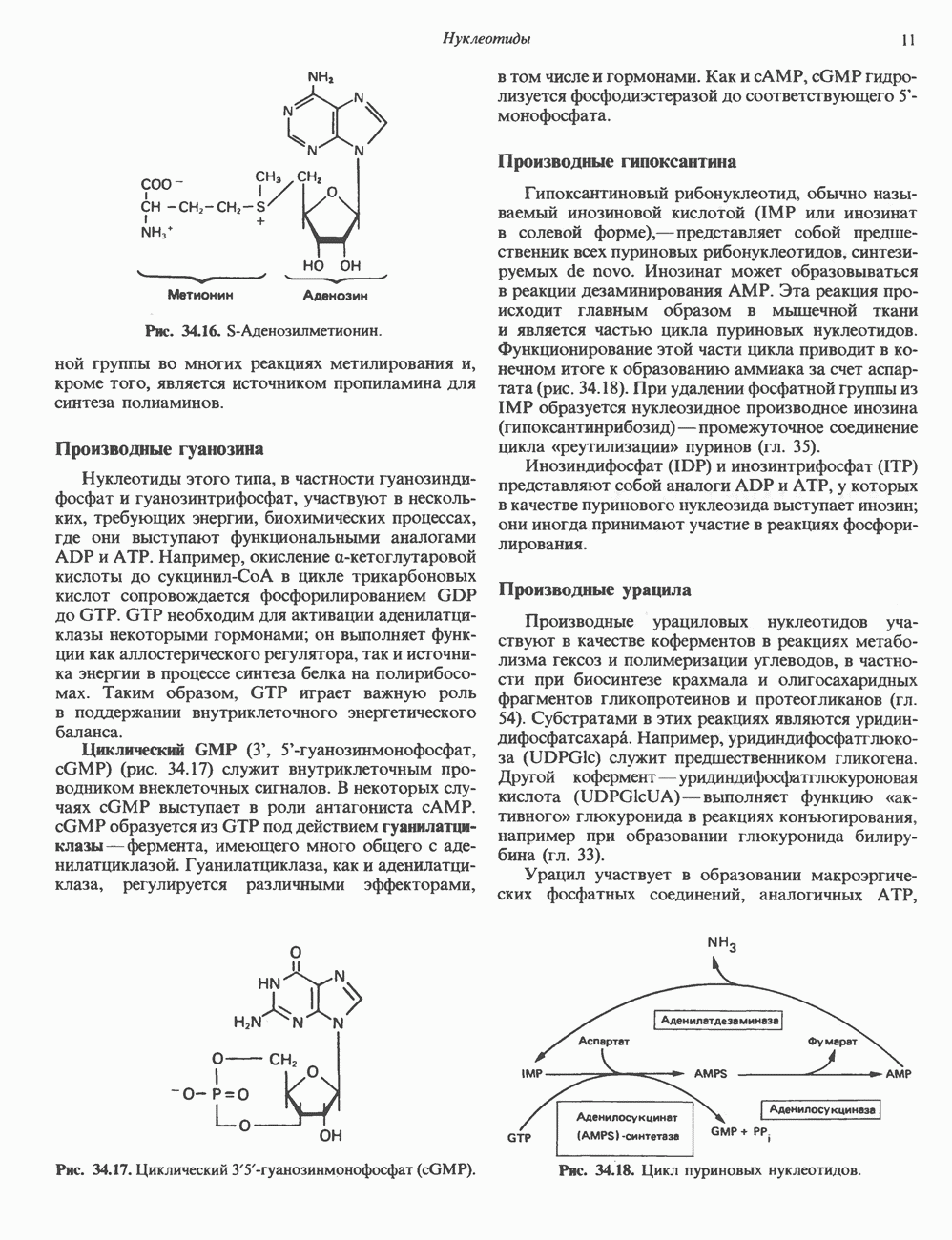
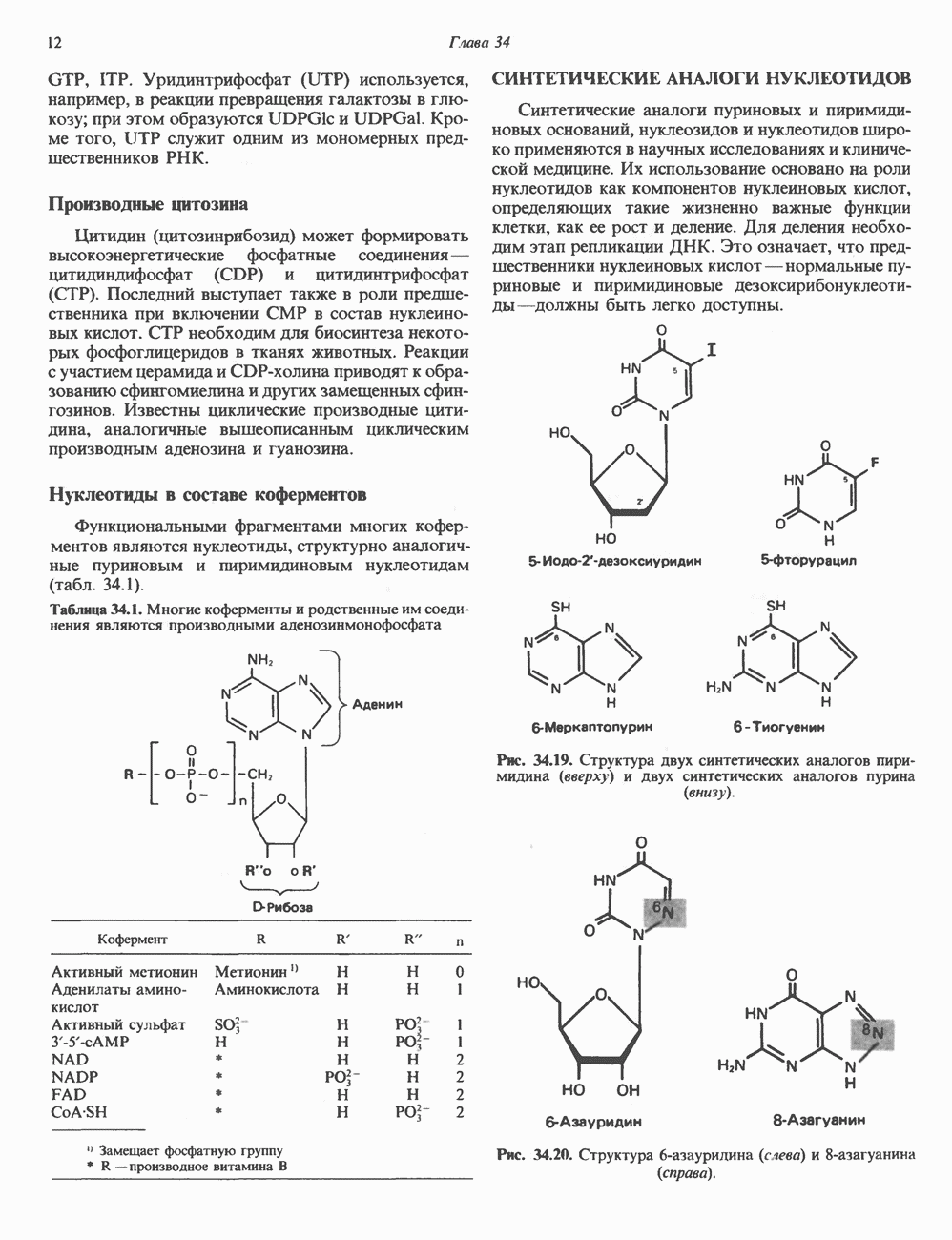
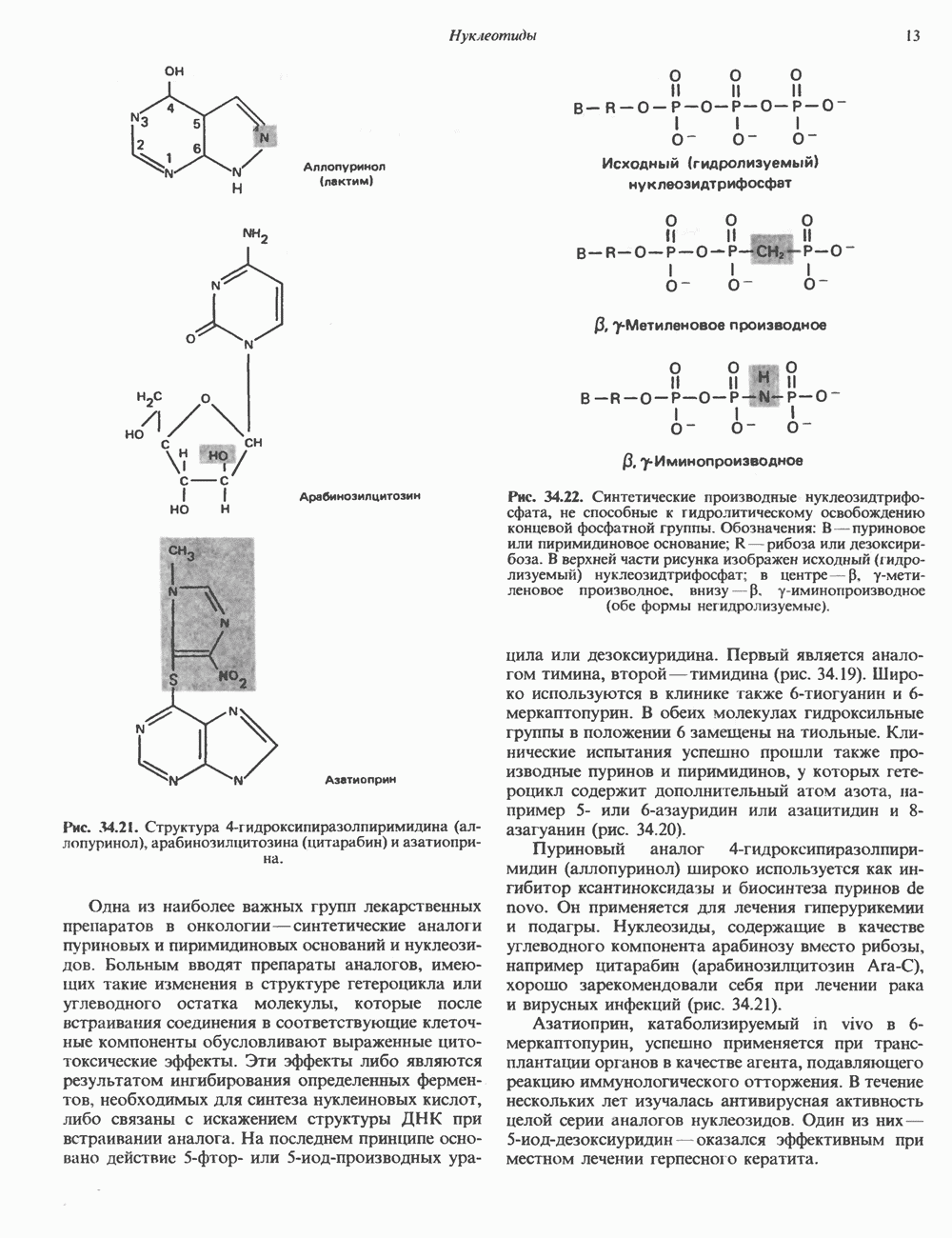
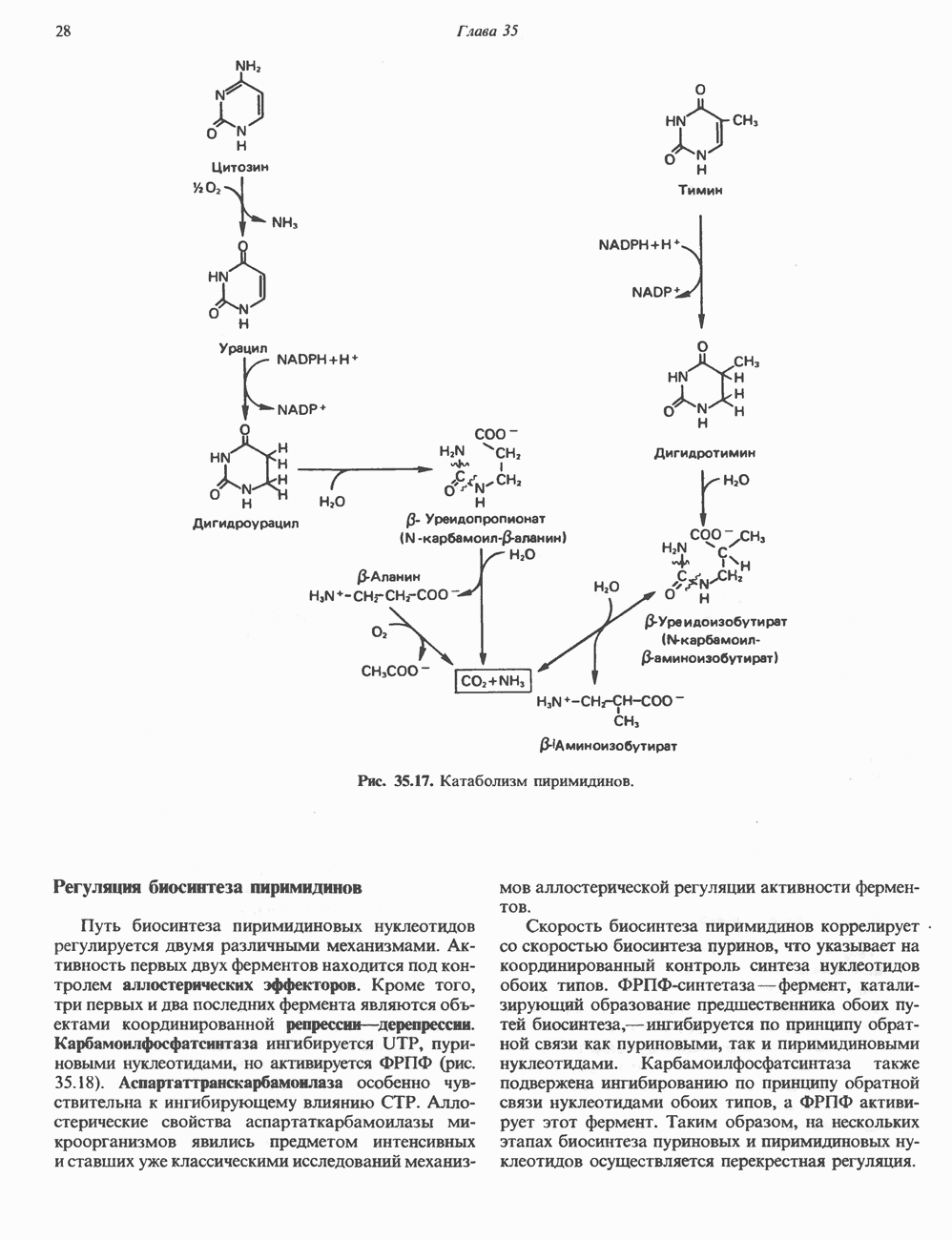
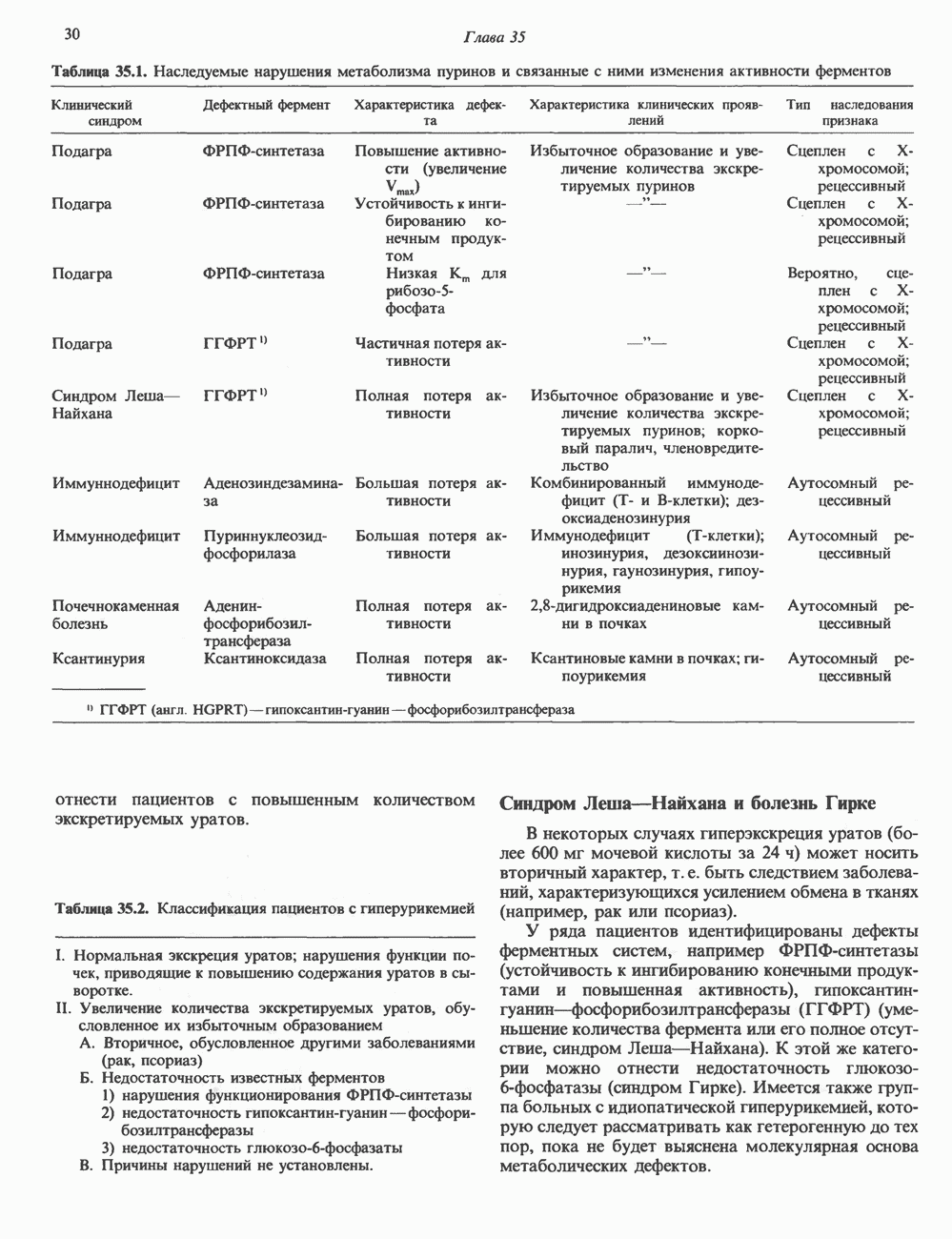
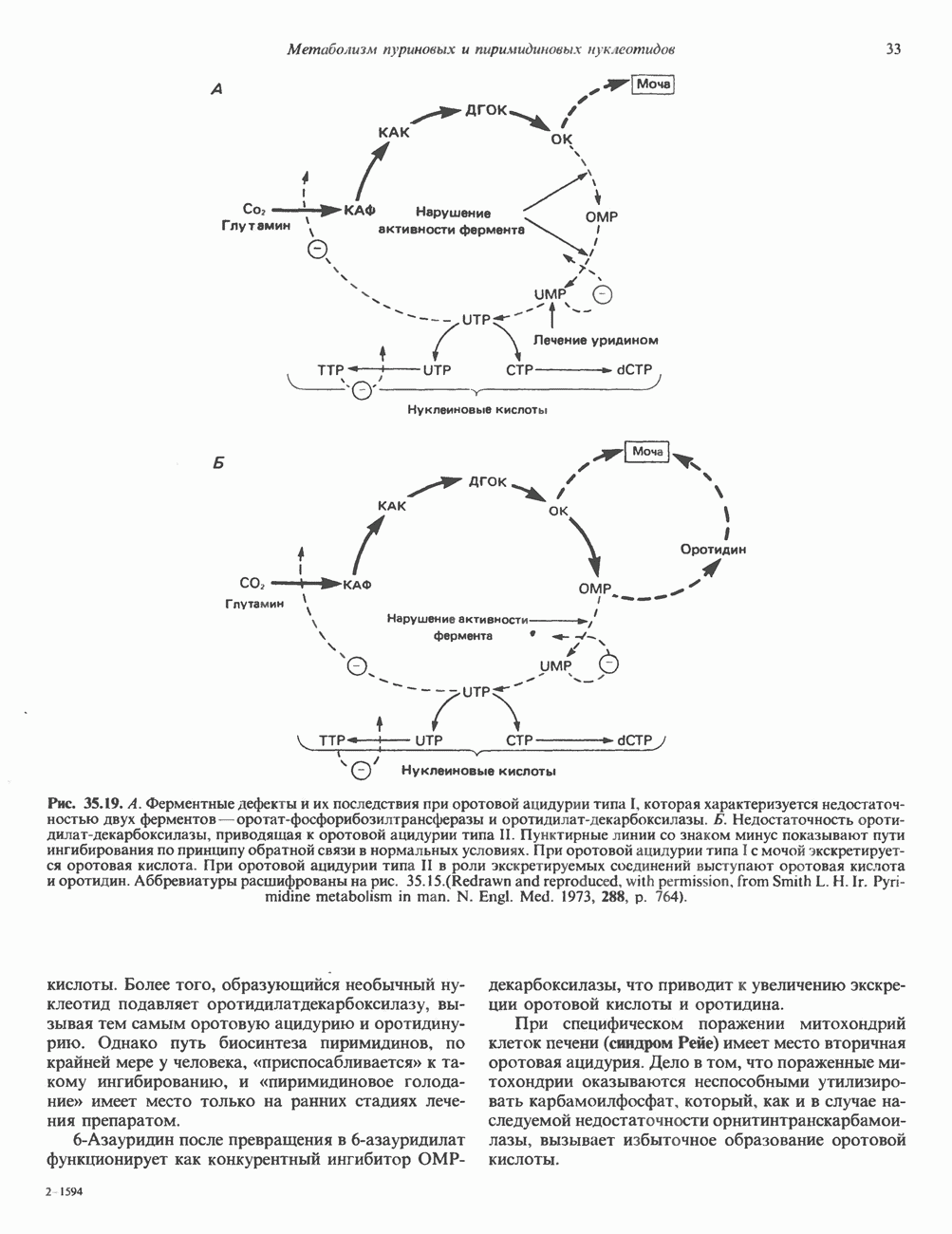
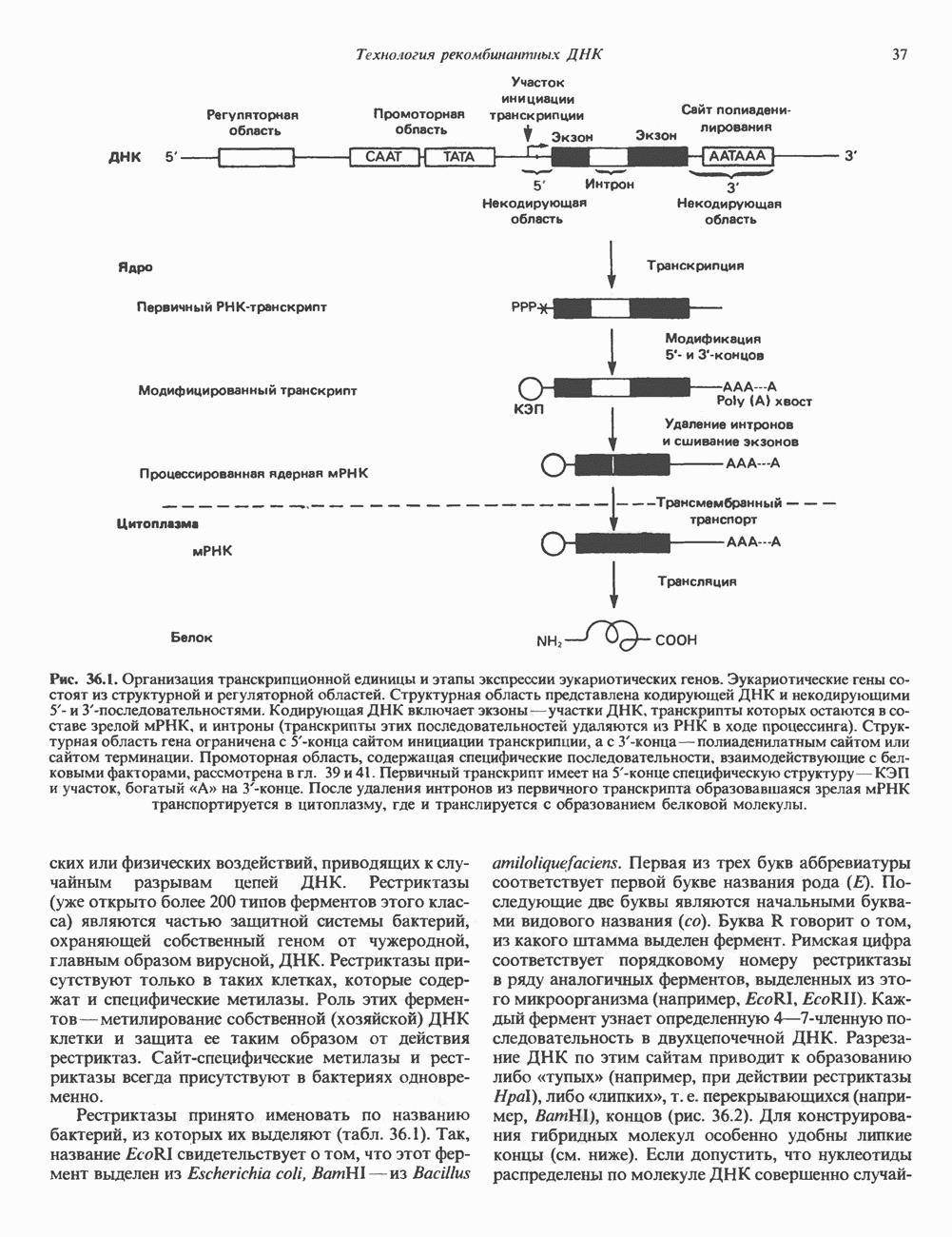
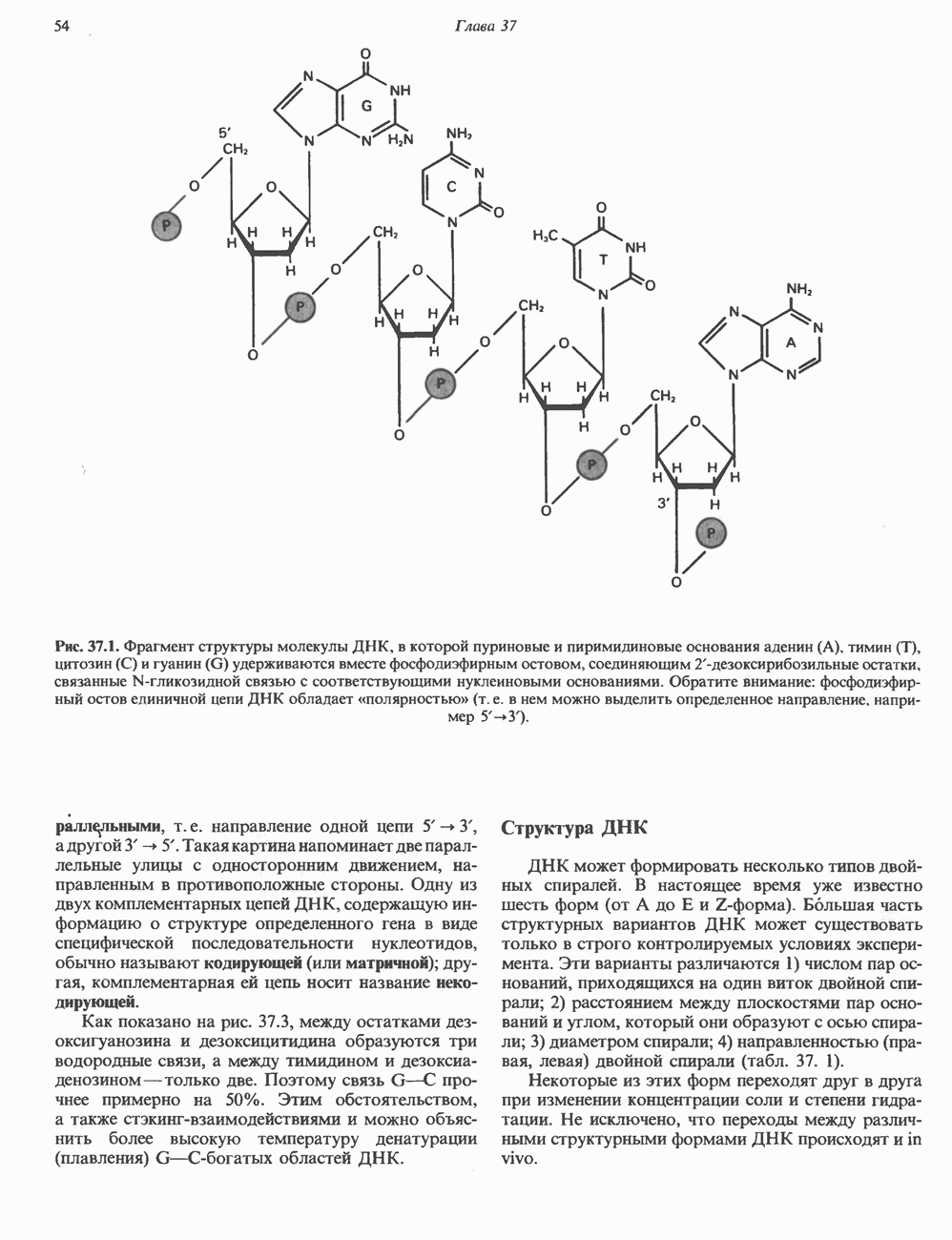
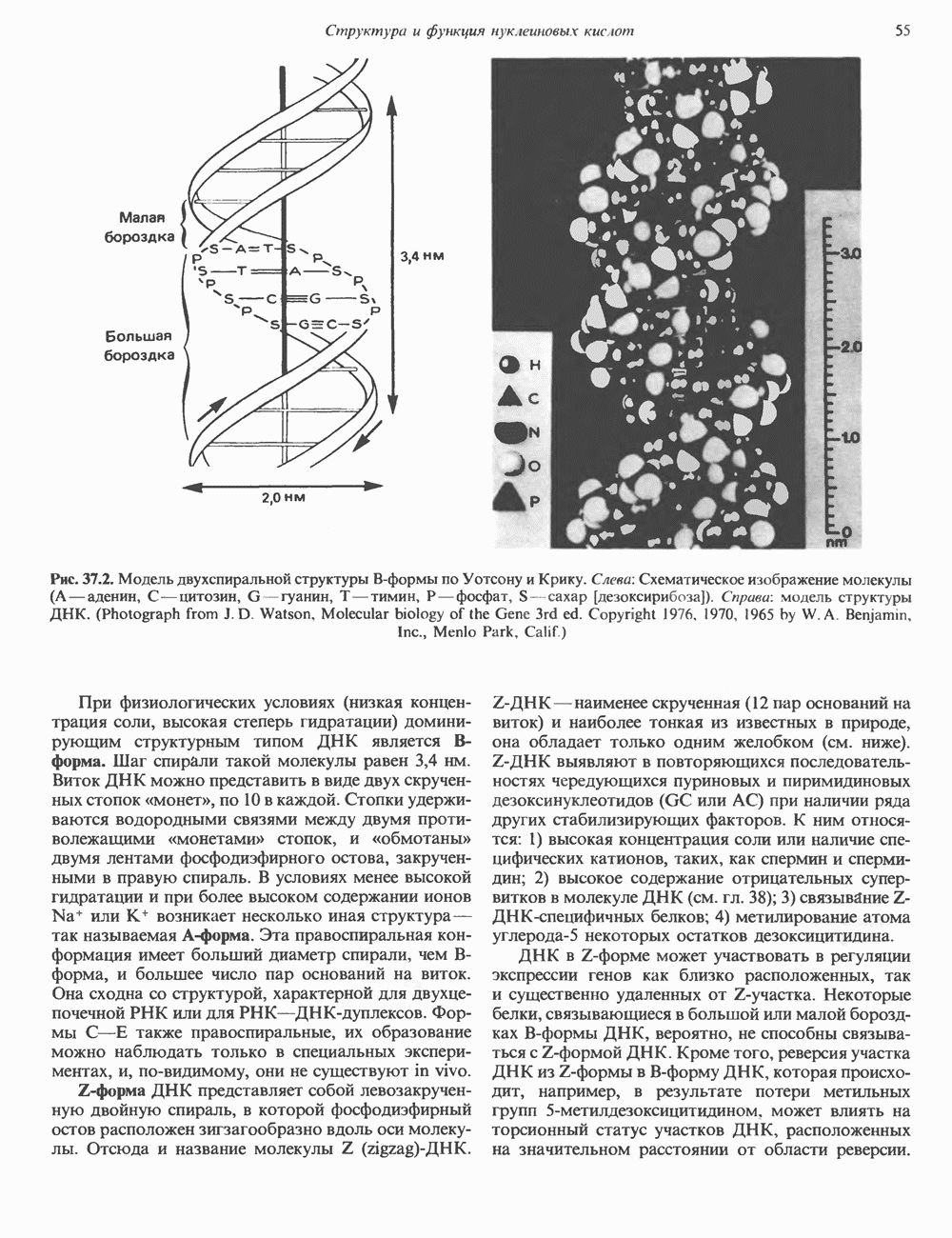
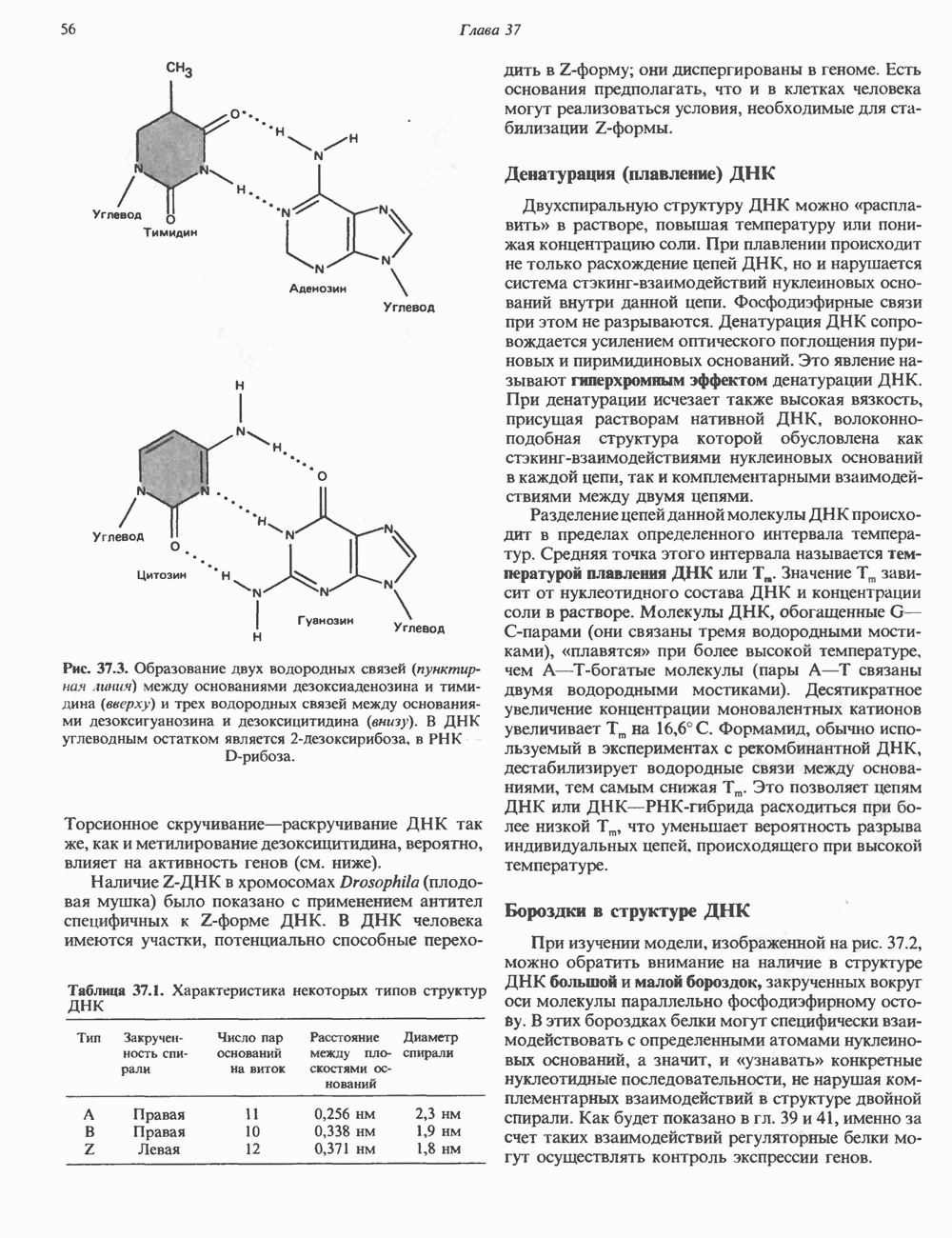
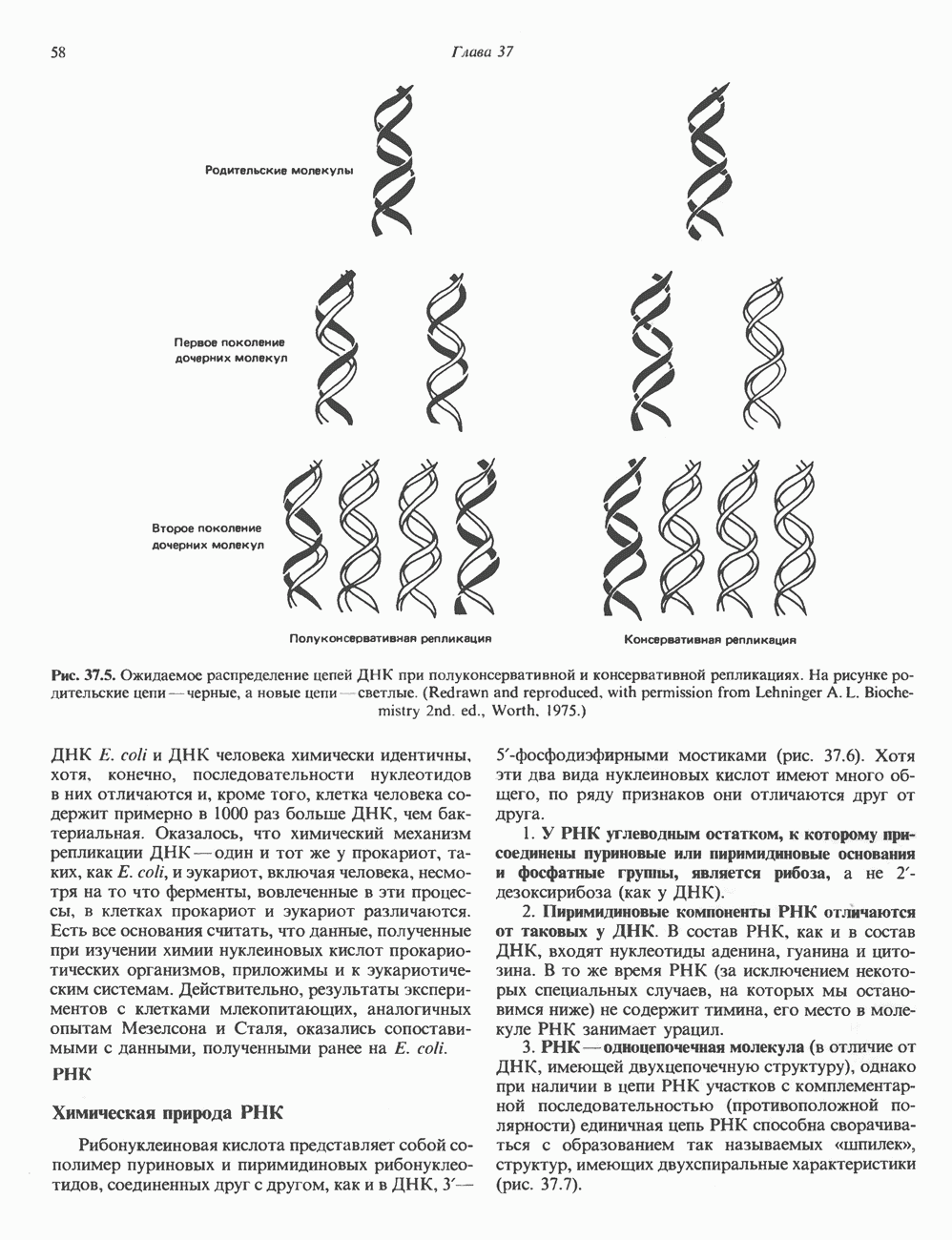
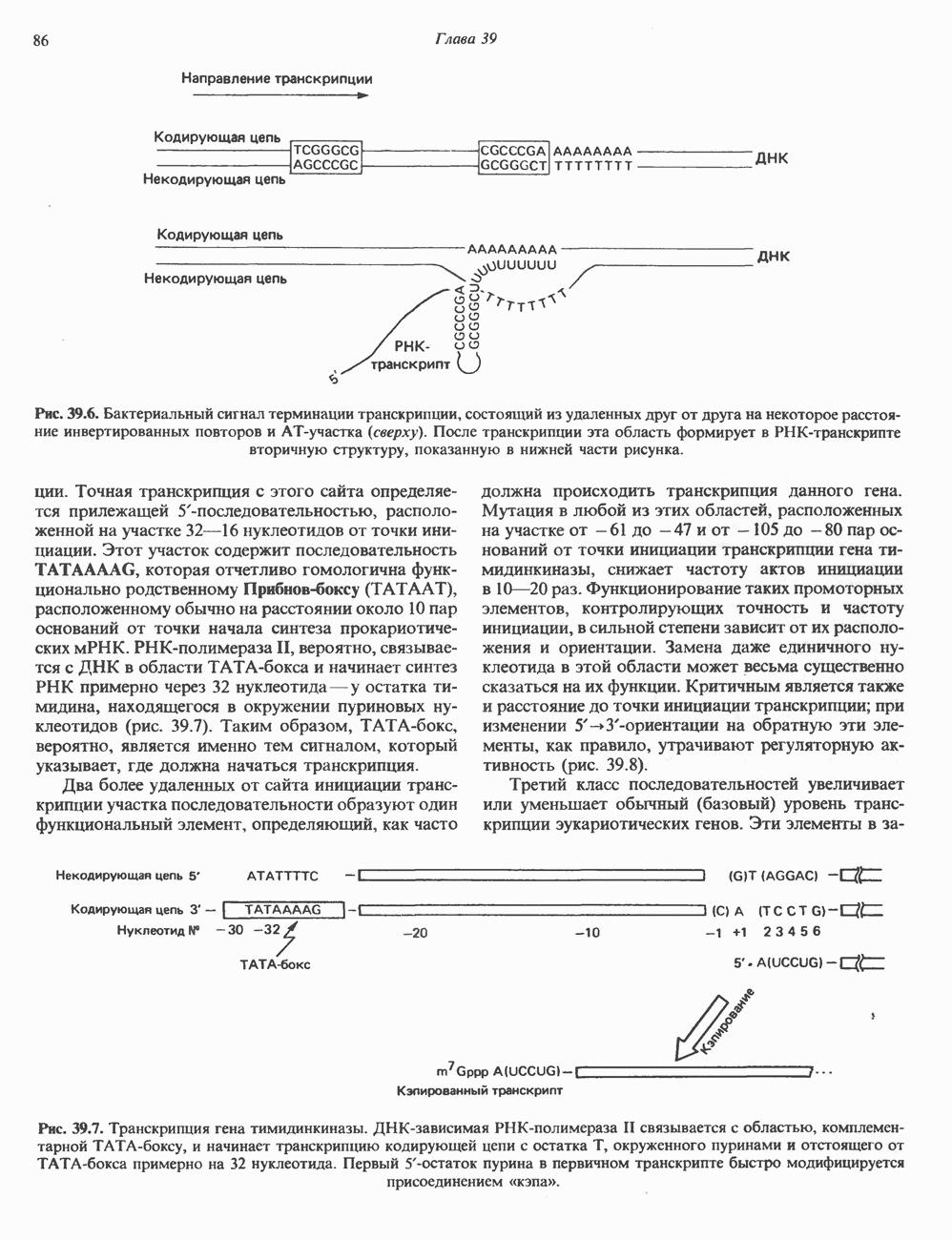
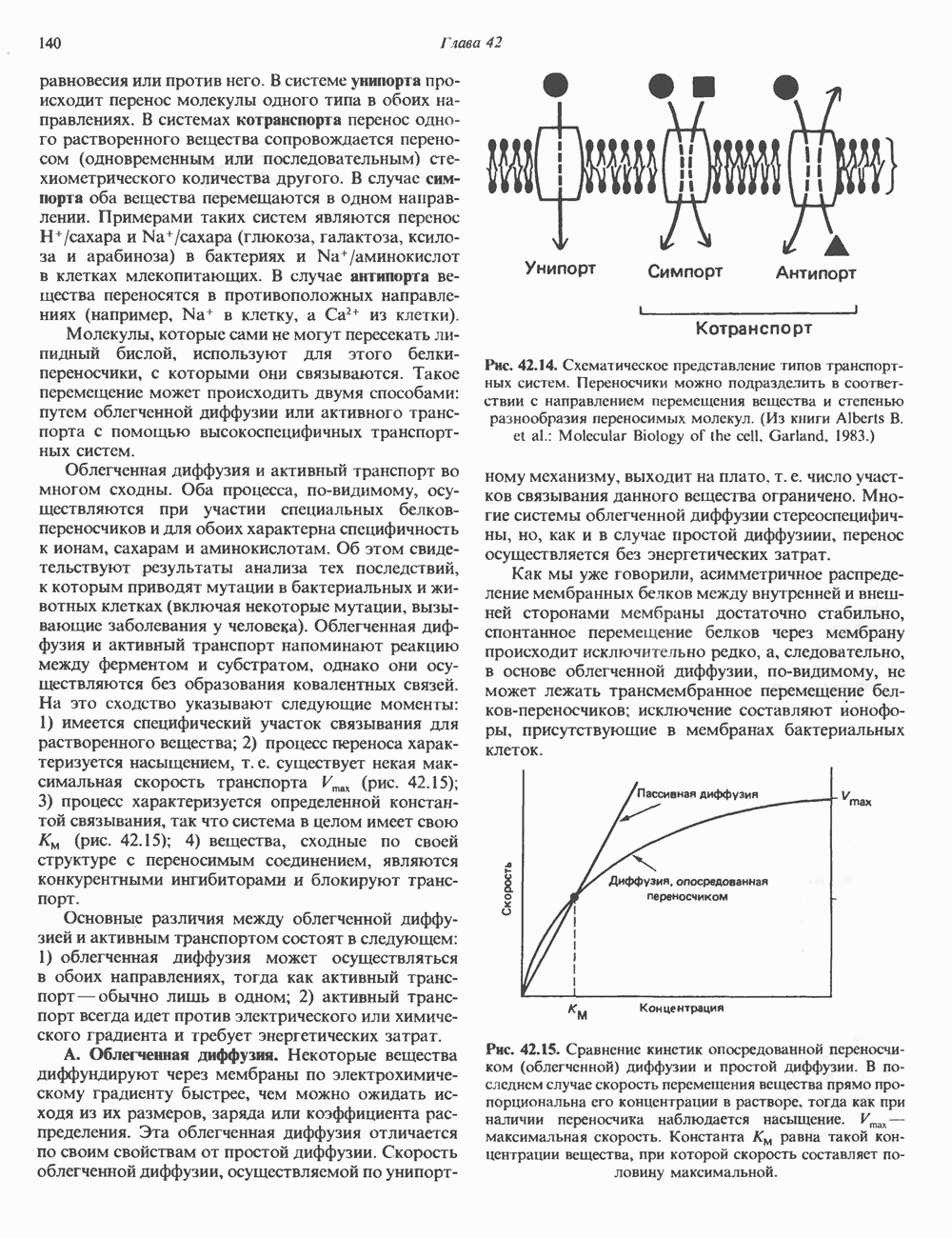
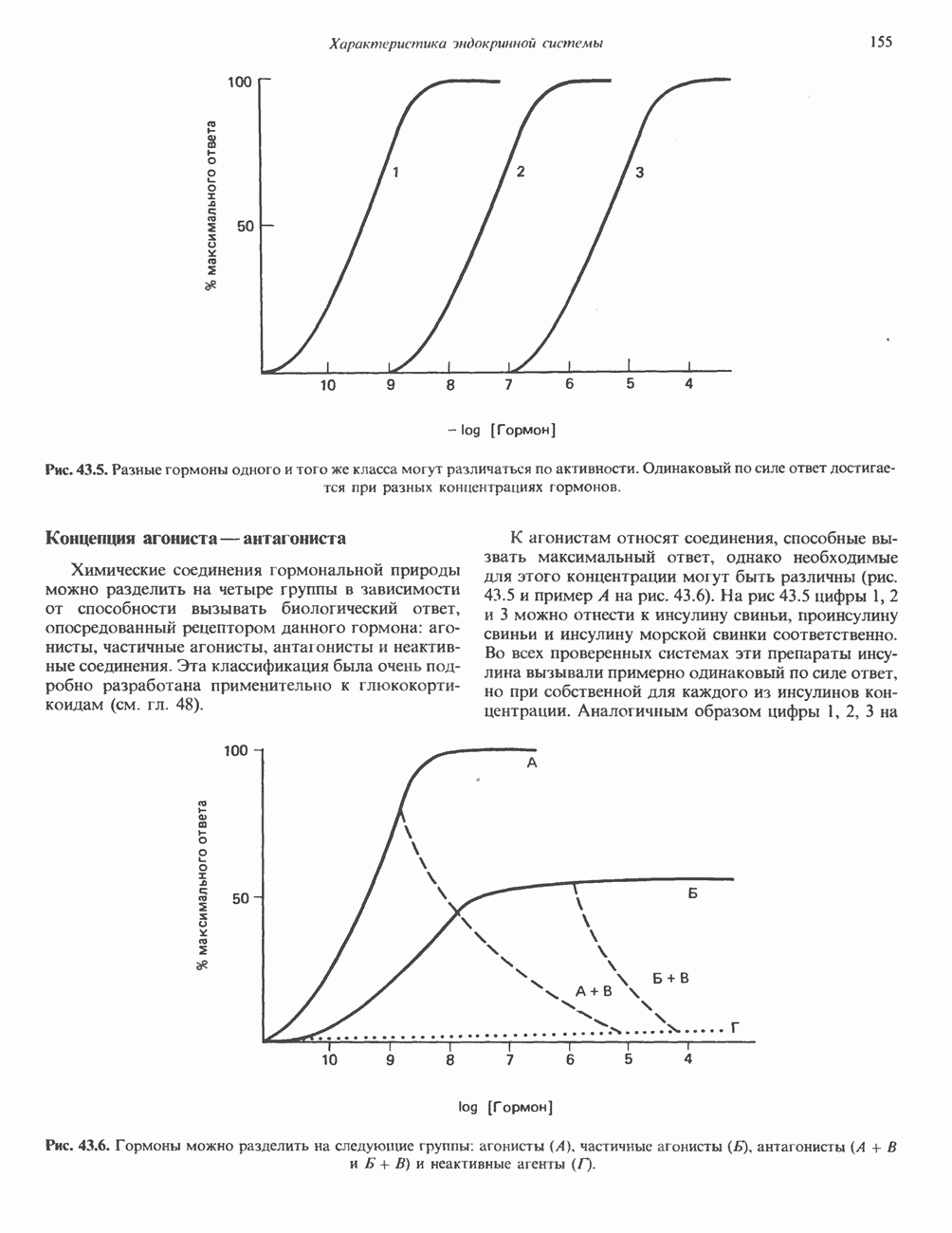
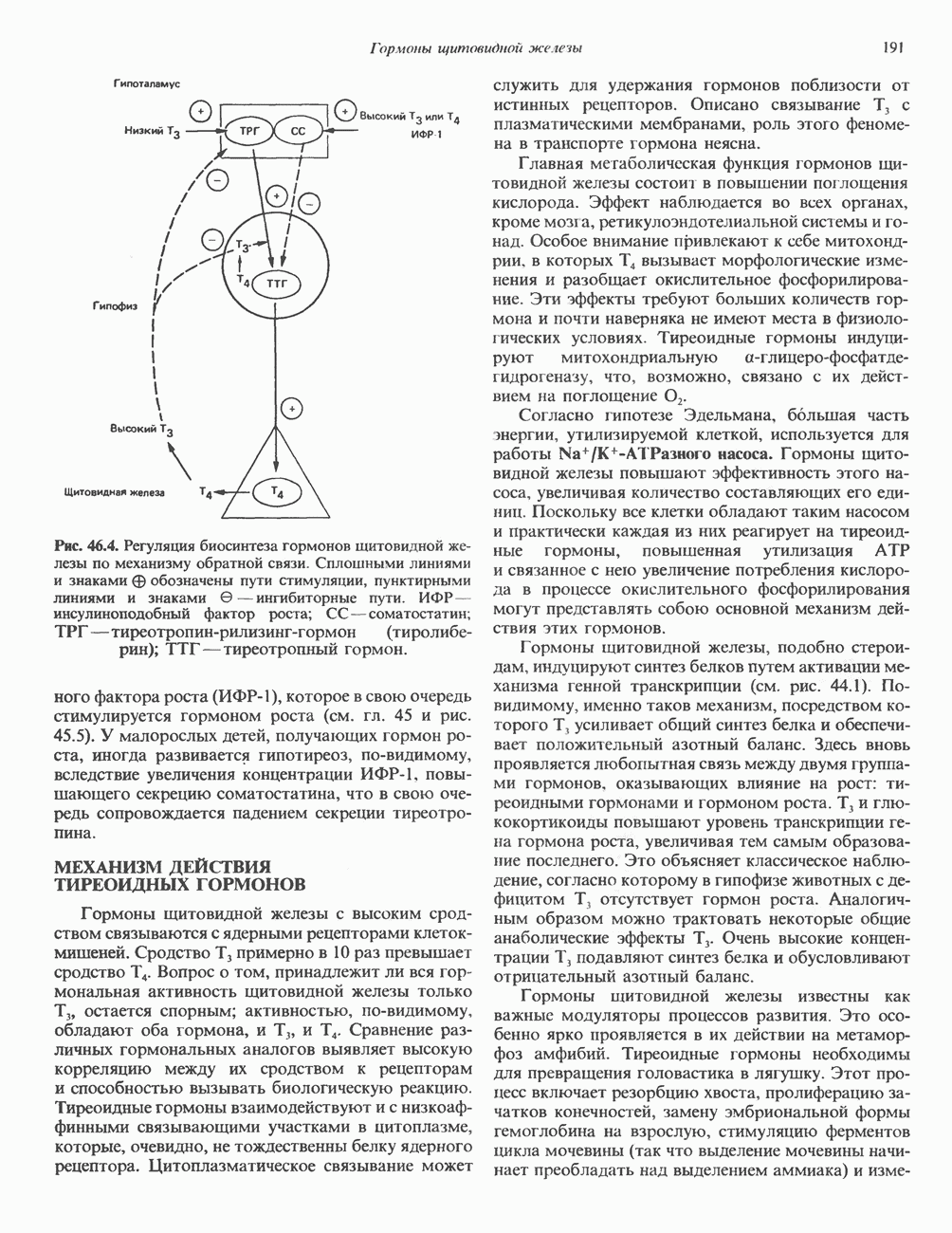
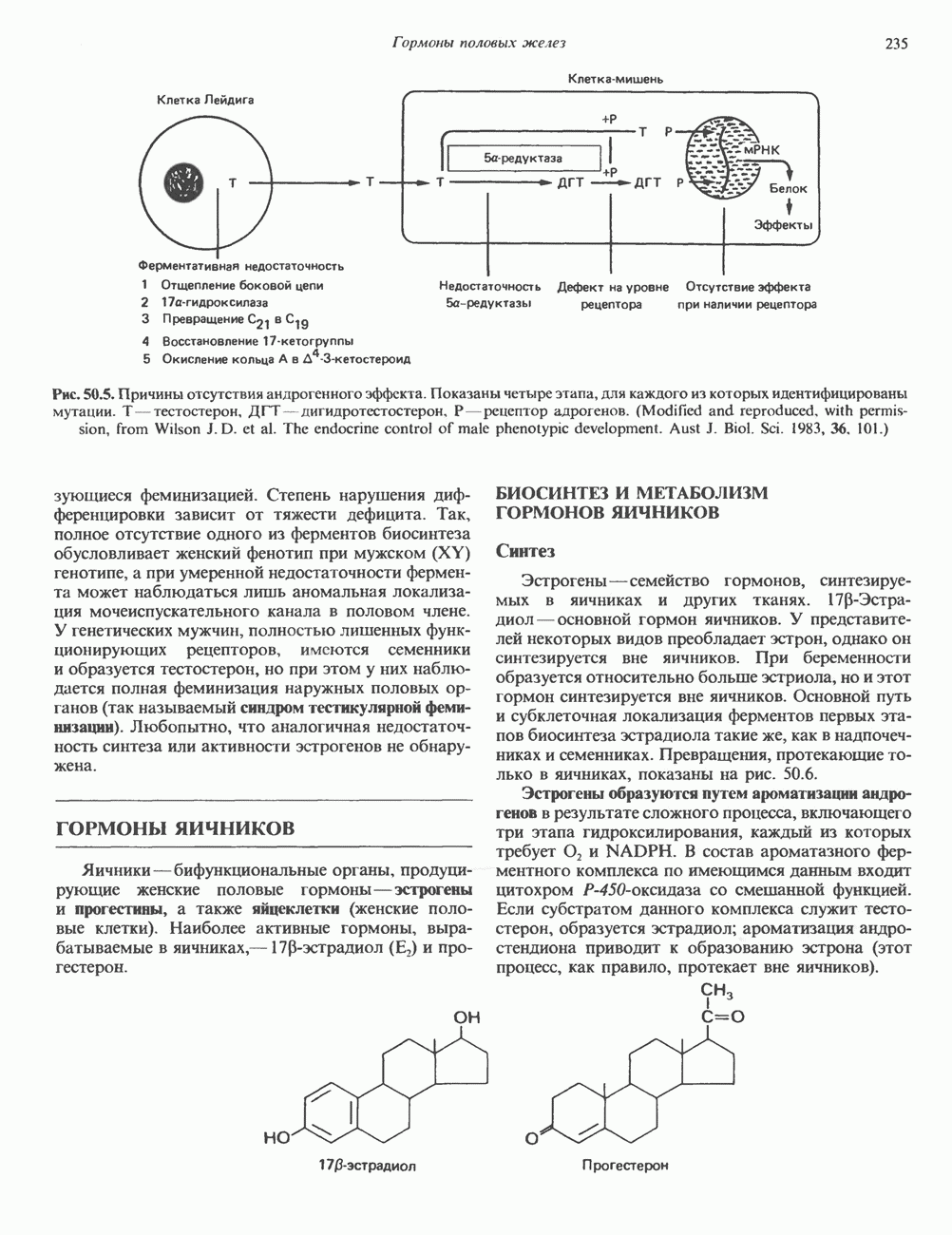
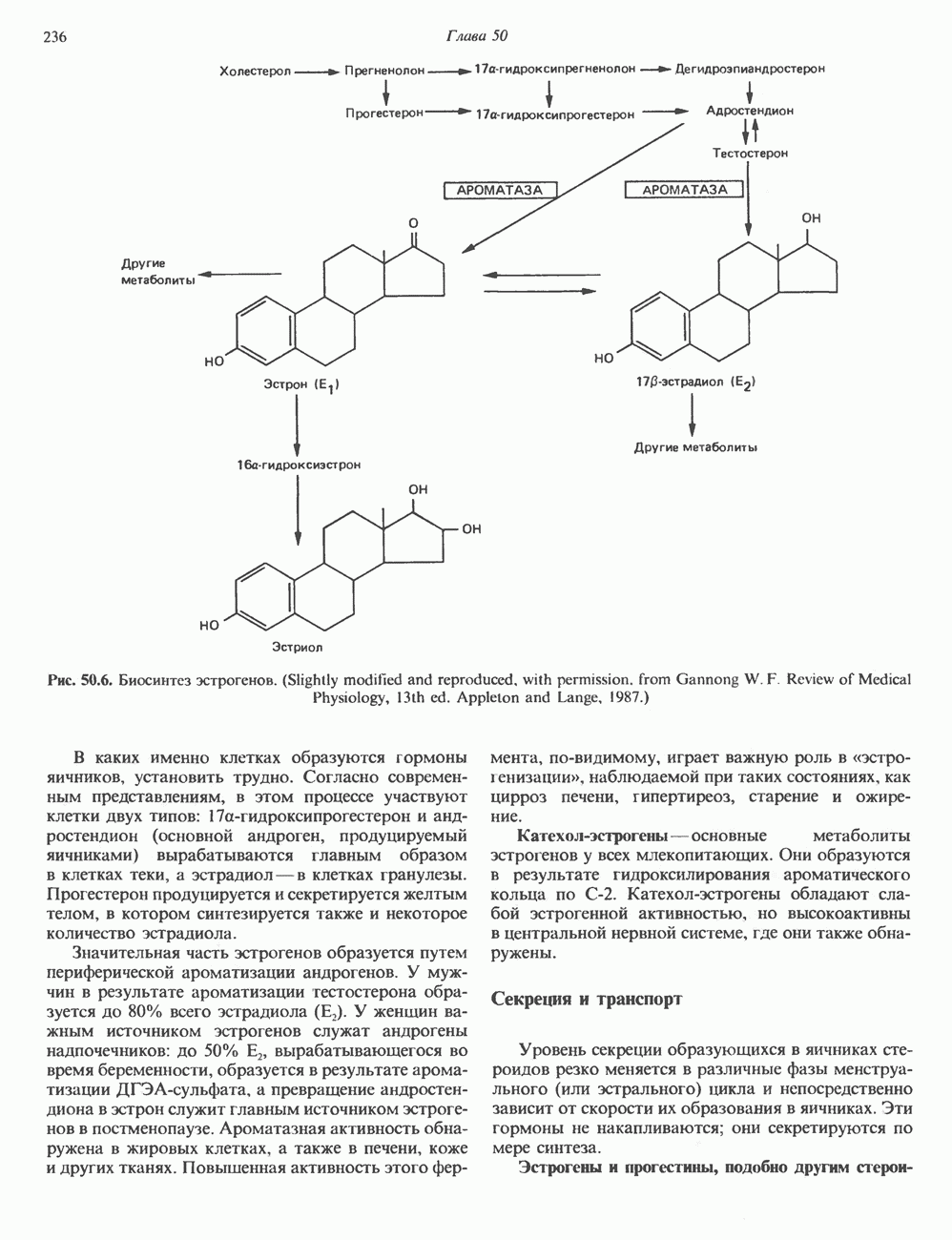
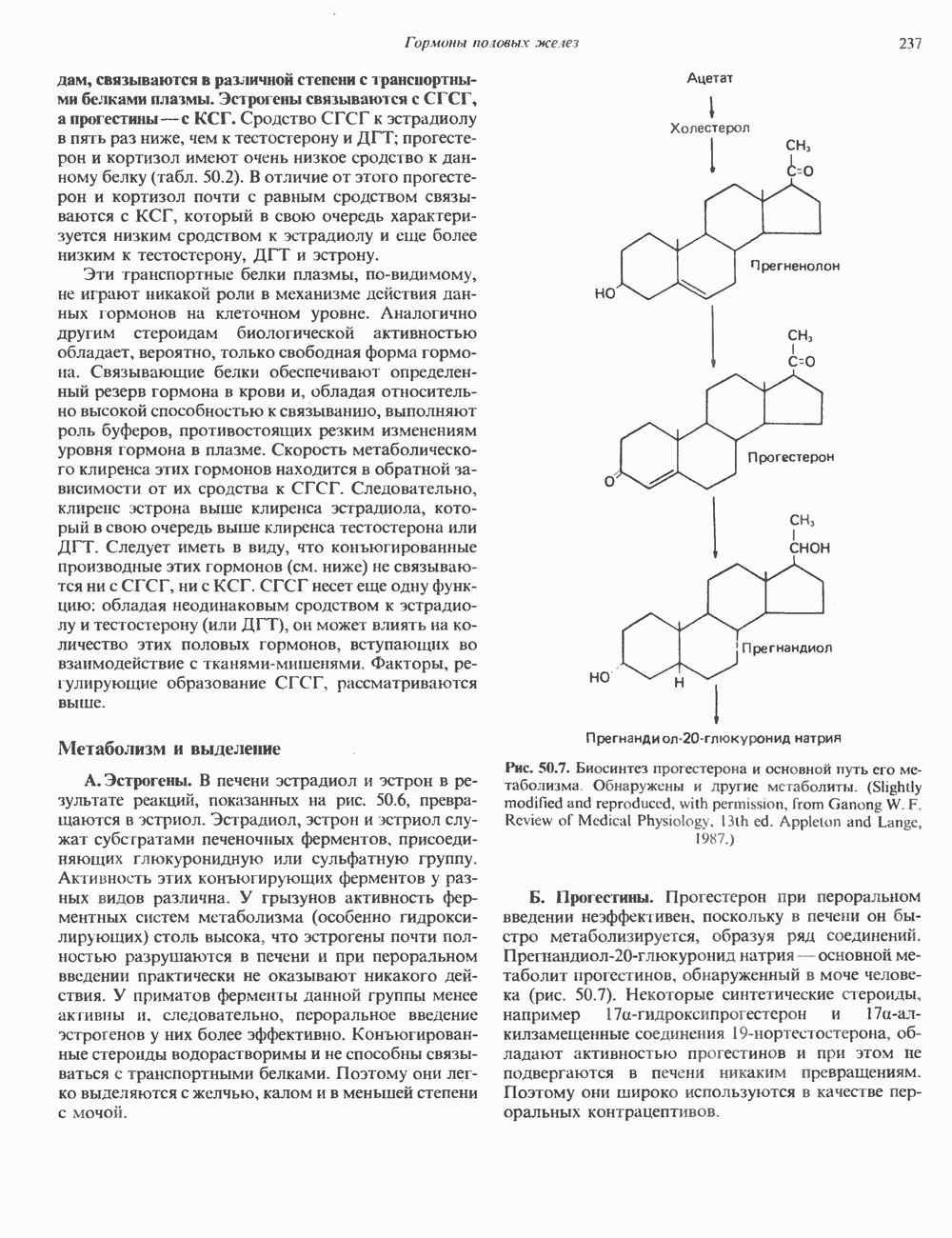
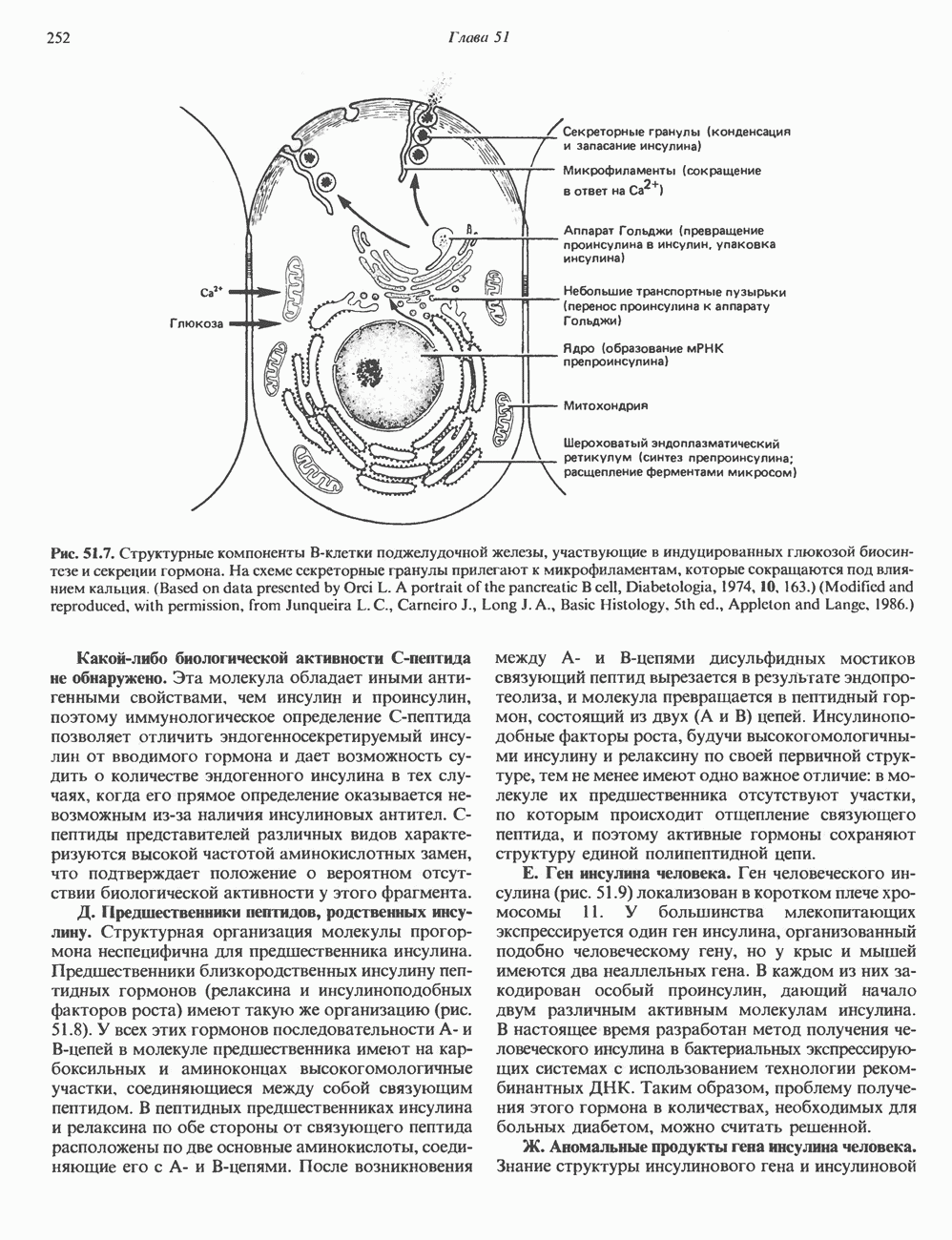
КЛЕТОЧНАЯ ПОДВИЖНОСТЬ И ЦИТОСКЕЛЕТСовершенно очевидно, что механическую работу, включающую амебоидные движения, морфогенез, деление, эндоцитоз, экзоцитоз, внутриклеточный транспорт и изменение формы, выполняют и немышечные клетки. Эти клеточные функции осуществляются обширной внутриклеточной сетью волокнистых структур, образующих цитоскелет. Клеточная цитоплазма — это не просто мешок с жидкостью, как думали раньше. Практически все эукариотические клетки содержат три типа волокнистых структур: актиновые филаменты (нити) (7-9,5 нм в диаметре), микротрубочки (25 нм) и промежуточные нити (10—12 нм). Каждый из этих типов можно отличить с помощью специфических биохимических и электронно-микроскопических методик. Немышечный актинБелок G-актин, выделенный из немышечных клеток, имеет мол. массу около 43 000 и, подобно мышечному актину (а-актину), содержит N-метилгистидиловые остатки. В присутствии магния и хлористого калия этот актин спонтанно полимеризуется с образованием двуспиральных F-актиновых нитей, подобных мышечным филаментам. В немышечных клетках имеется по меньшей мере два типа актина: Р-актин и Микрофиламенты плотно упакованы в виде петлистой сети, располагающейся под ведущим краем или «бахромой» подвижной клетки (рис. 56.11). Актиновые микрофиламенты обнаруживаются во всех клеточных микроотростках, таких, как филоподии и микроворсинки. Например, микроворсинки клеток слизистой кишечника содержат 20—30 актиновых микрофиламентов, расположенных продольно, как показано на рис. 56.12. Эти микрофиламенты, декорированные миозиновым фрагментом S-1, проявляют однородную полярность (рис. 56.12). В основании микроворсинок находятся миозиновые филаменты, способные втягиваться вместе с актиновыми
Рис. 56.10. Электронная микрофотография фрагмента цитоскелета, к которому перед быстрым замораживанием был добавлен миозиновый субфрагмент нитями в микроворсинки. Процесс сокращения не сопровождается какими-либо изменениями длины актина или миозина и, таким образом, должен осуществляться, как и в мышцах, путем скольжения филаментов, т. е. с помощью механизма, сформулированного Хаксли. Как и в гладких мышцах, активация актин—миозинового взаимодействия и, соответственно, процесса сокращения, опосредуется фосфорилированием легкой цепи миозина.
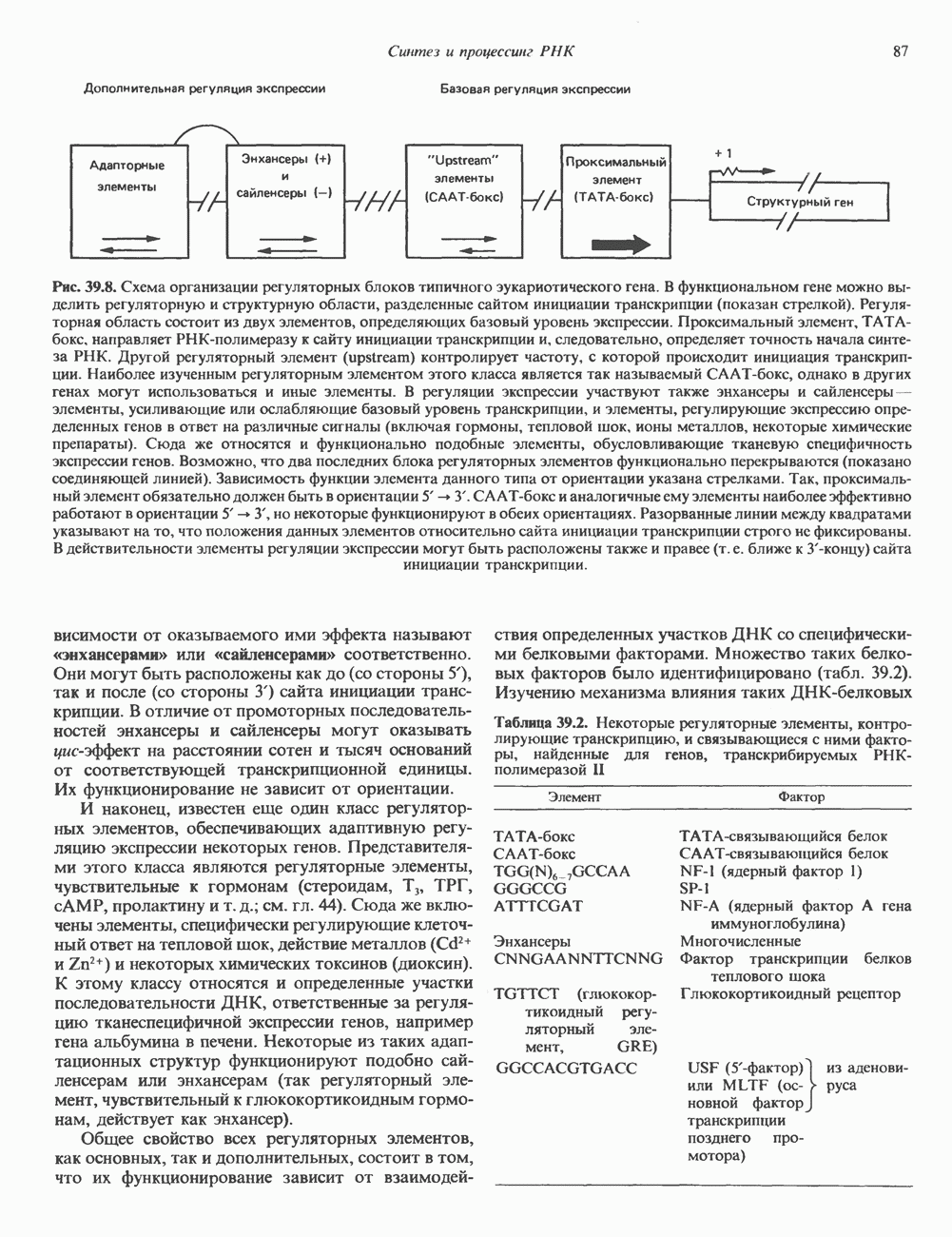
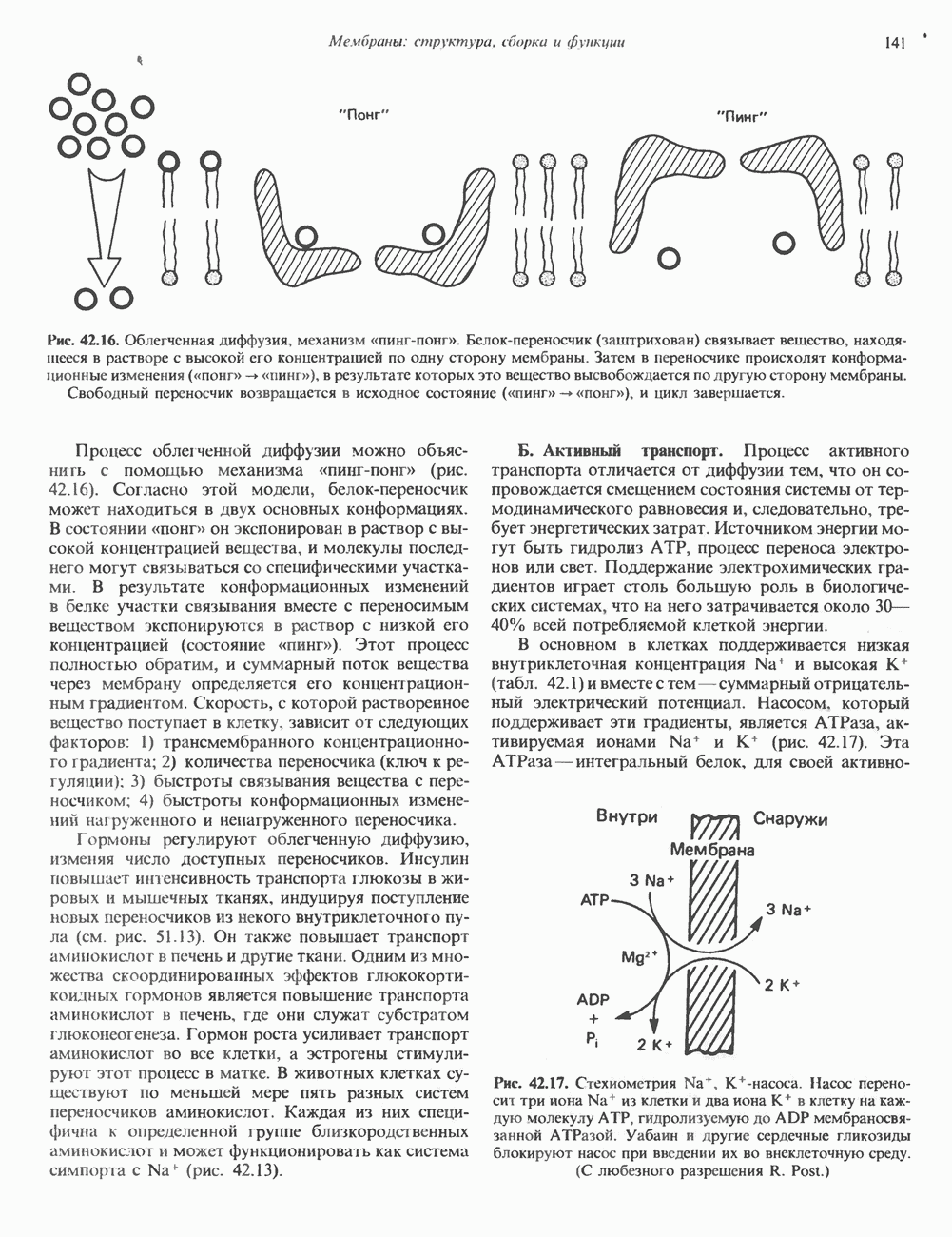
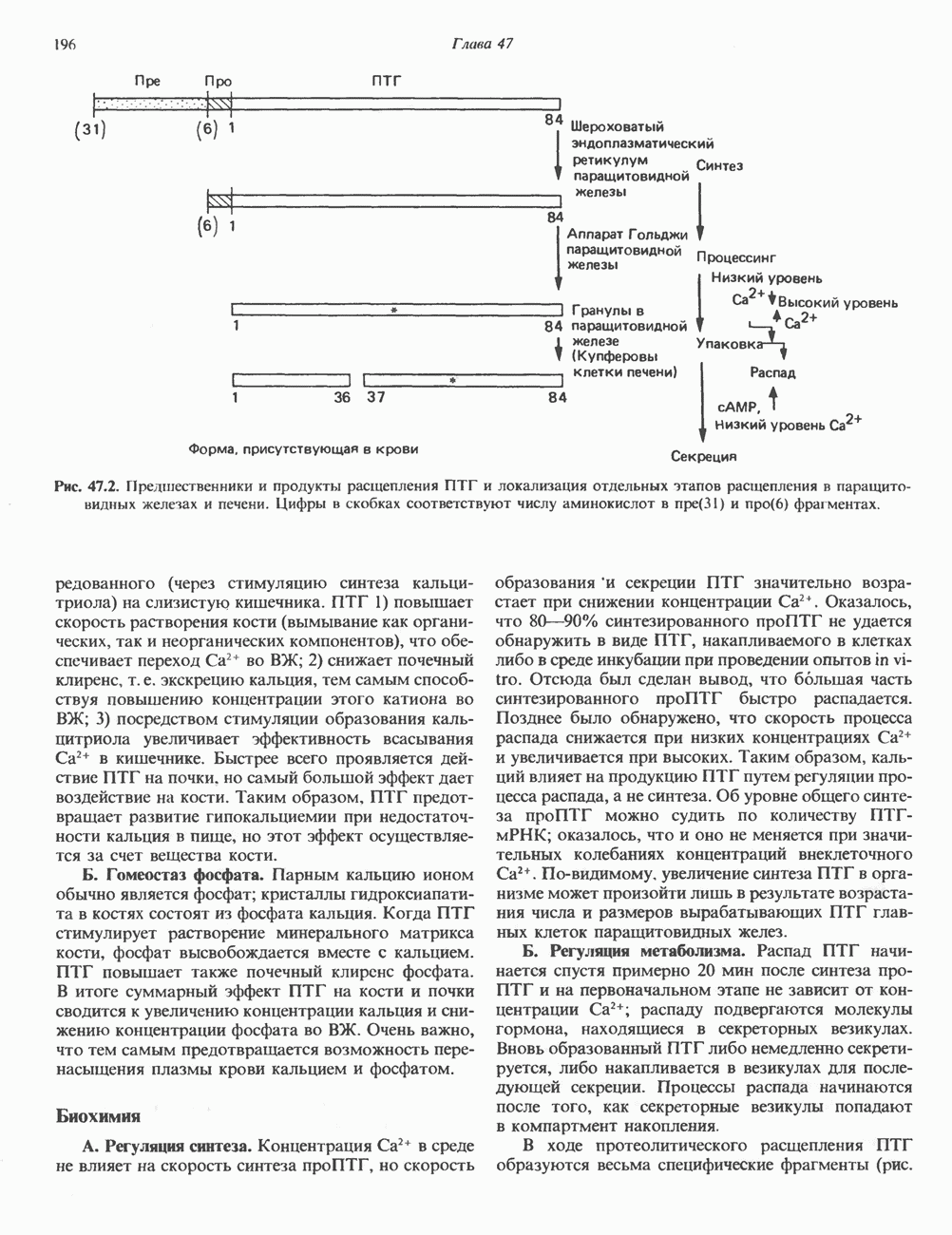
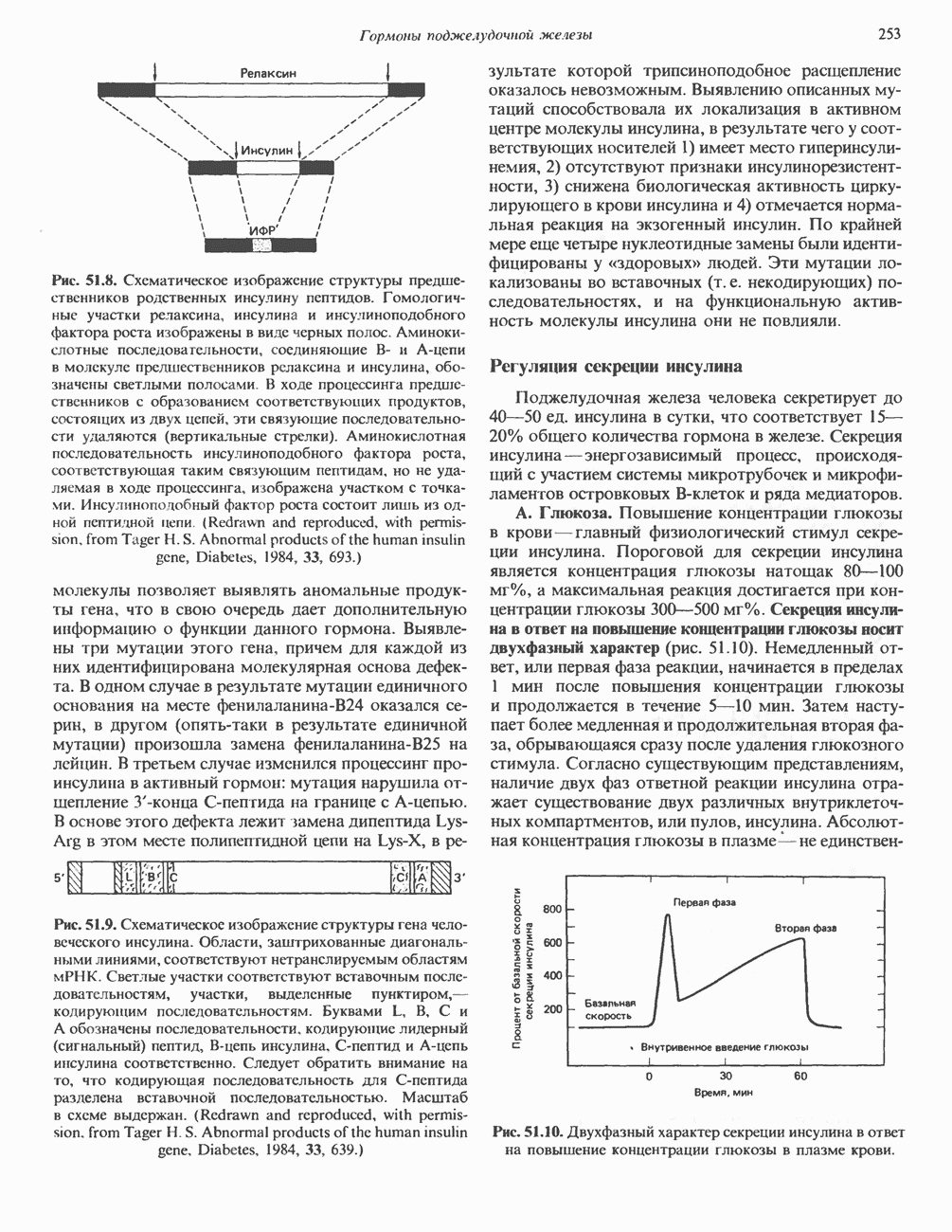
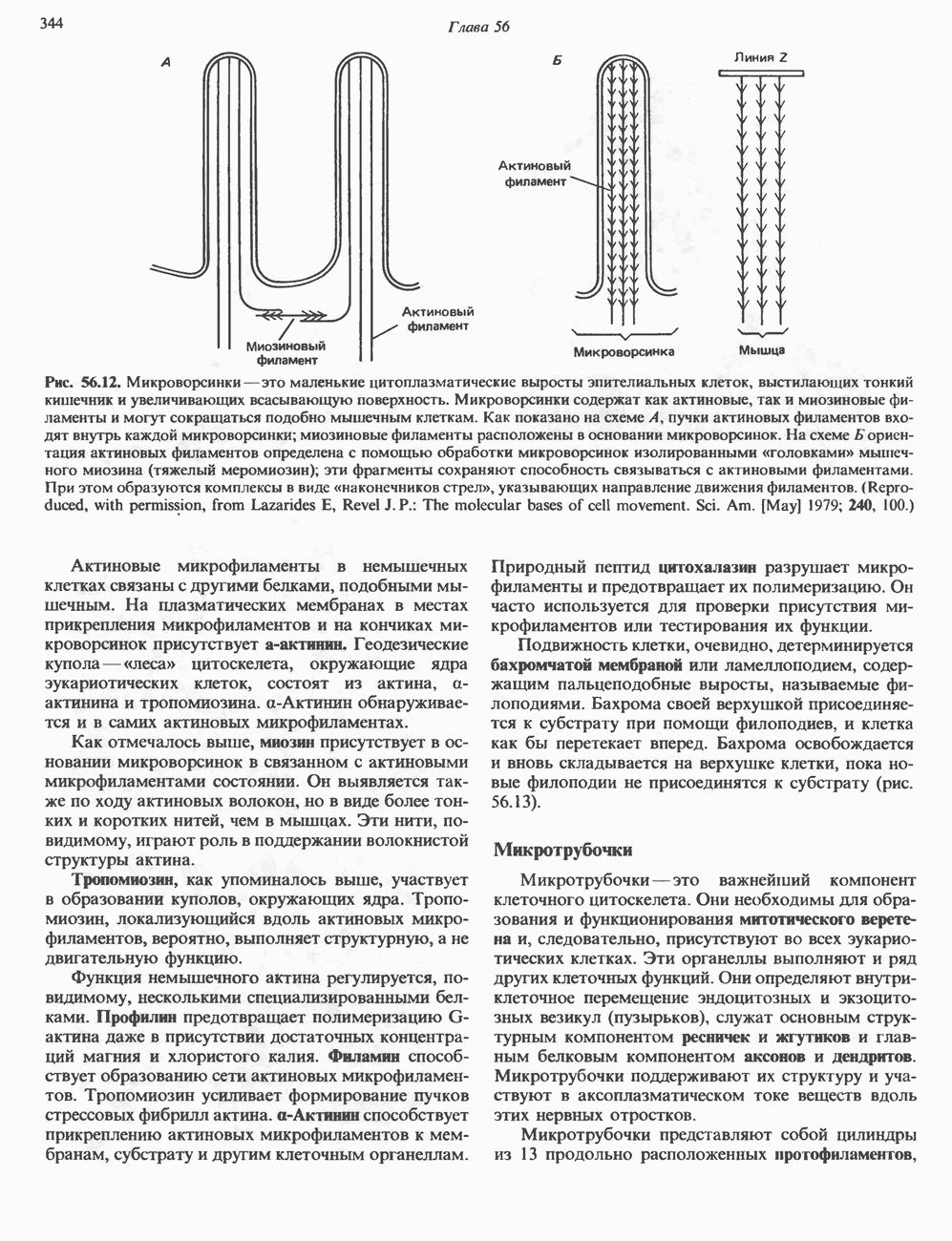
Рис. 56.11. Препараты бахромы или ламеллоподиев из фибробластов (увеличение). А. Полная фиксация. Б. Материал перед фиксацией экстрагирован тритоном В. Экстракция тритоном проведена после фиксации. В первом случае (фото А) плазматическая мембрана интактна и внутренняя структура не видна. Во втором случае (фото Б) плазматическая мембрана удалена и видны лежащие под ней переплетения «кудрявых» филаментов. Фото В — плазматическая мембрана также удалена, но уже после фиксации альдегидом. Тонкая сеть подлежащих филаментов после химической фиксации выглядит более грубой. А, х 140 000- Б и В, х 115000. (Reproduced, with permission, from Heuser J. E., Kirschner M. W. Filament organ nation revealed in platinum replicas of freeze-dried cytoskeletons. J. Cell Biol.. 1980, 86. 212.) Актин и миозин обнаруживаются также между полюсами веретена и хромосомами и вдоль борозды дробления в телофазе митоза.
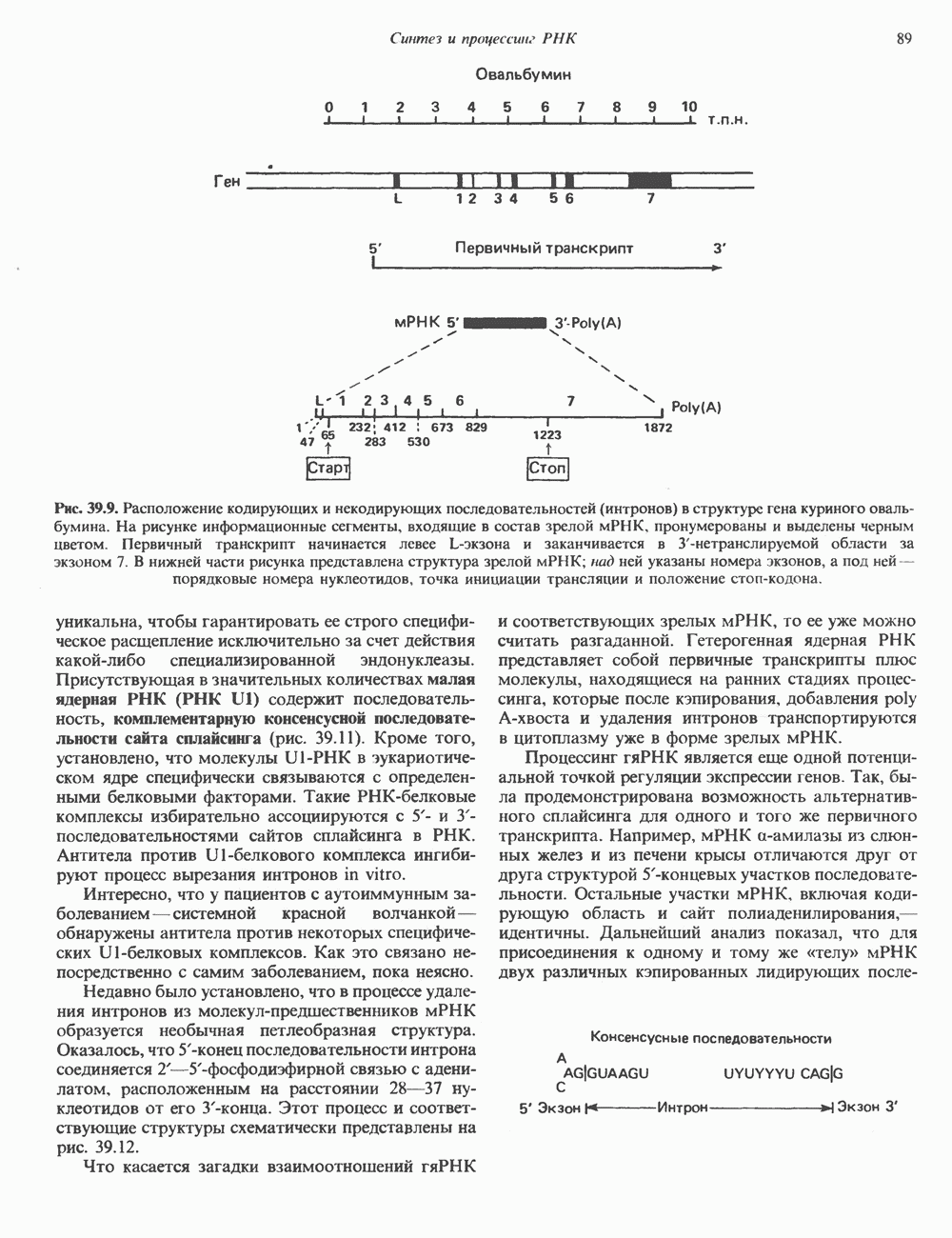
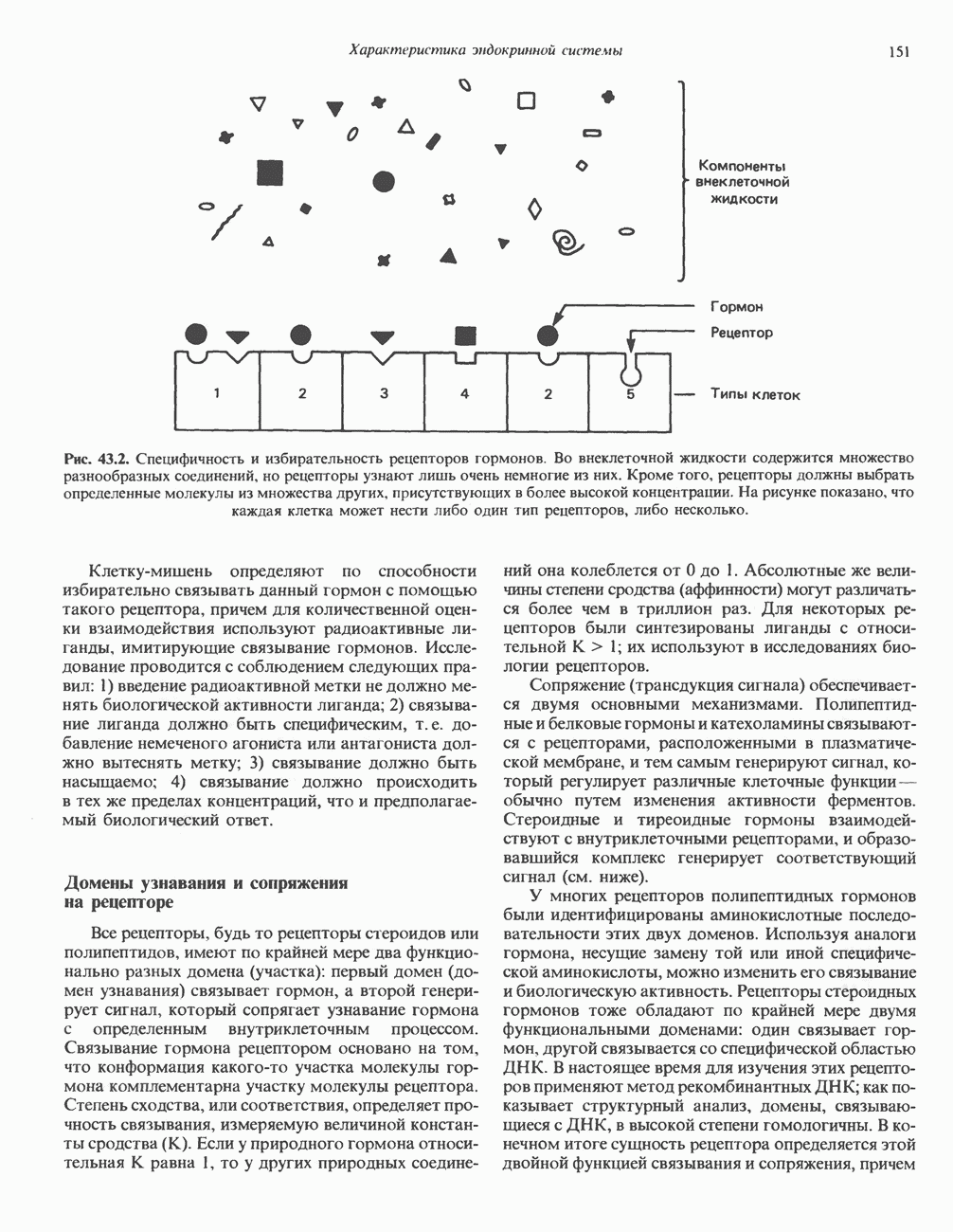
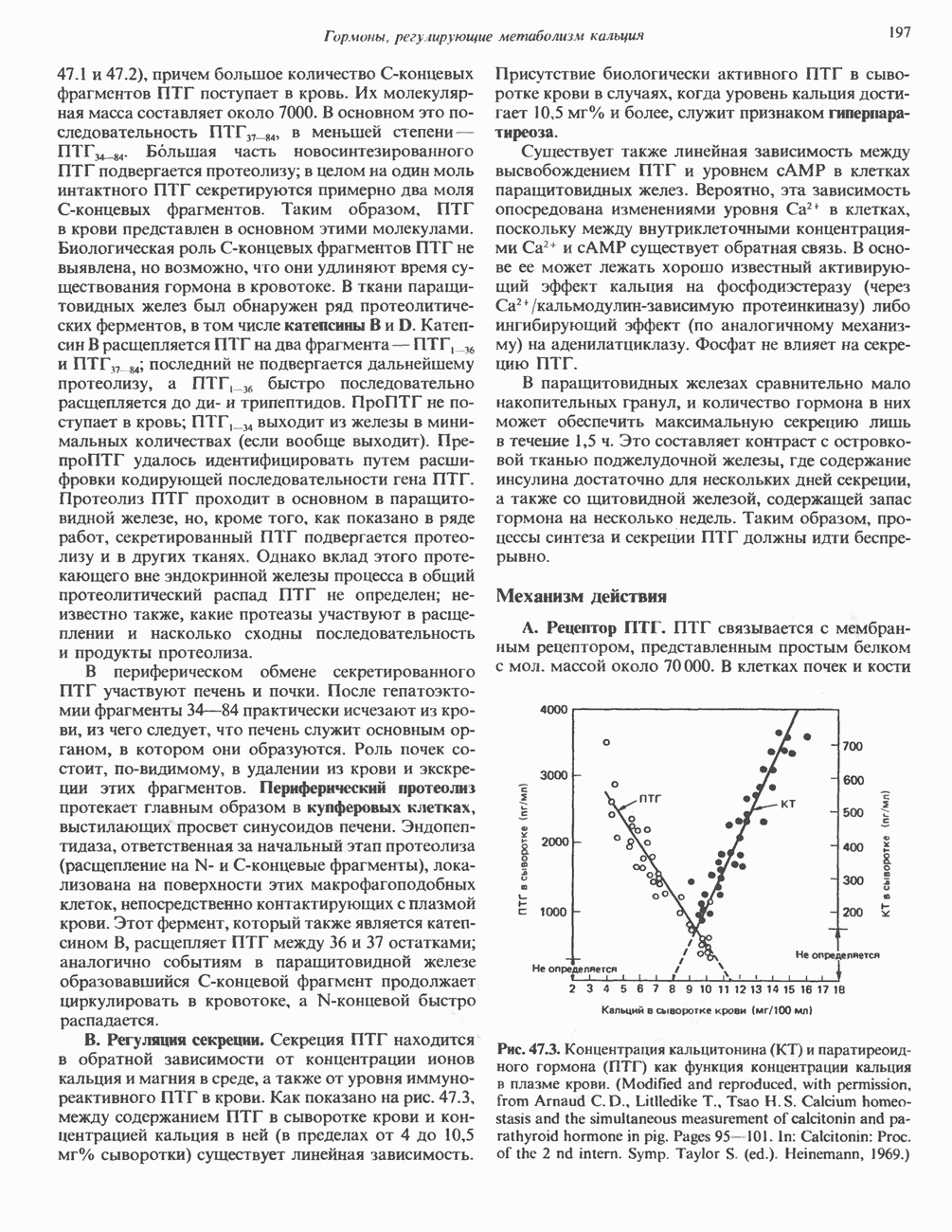
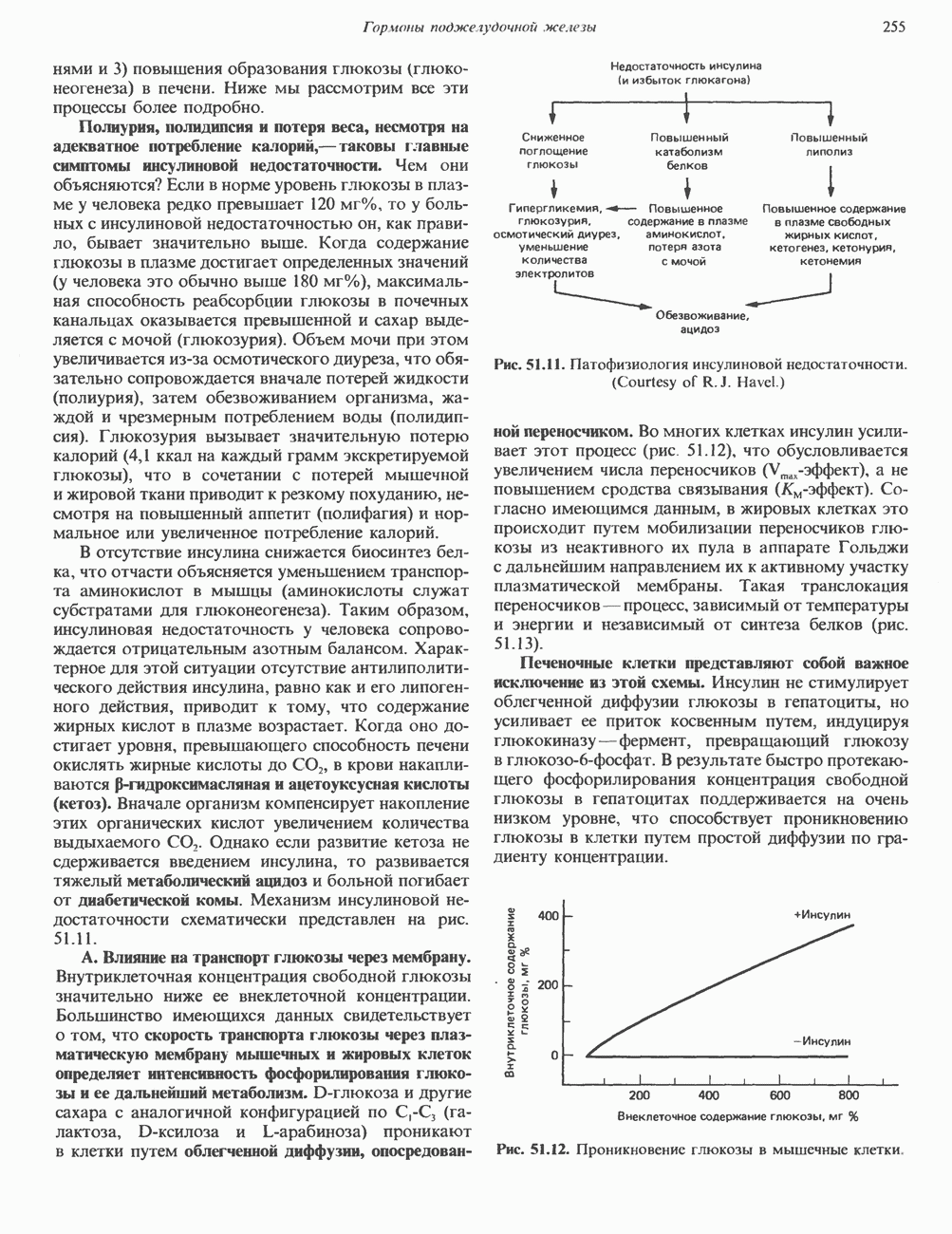
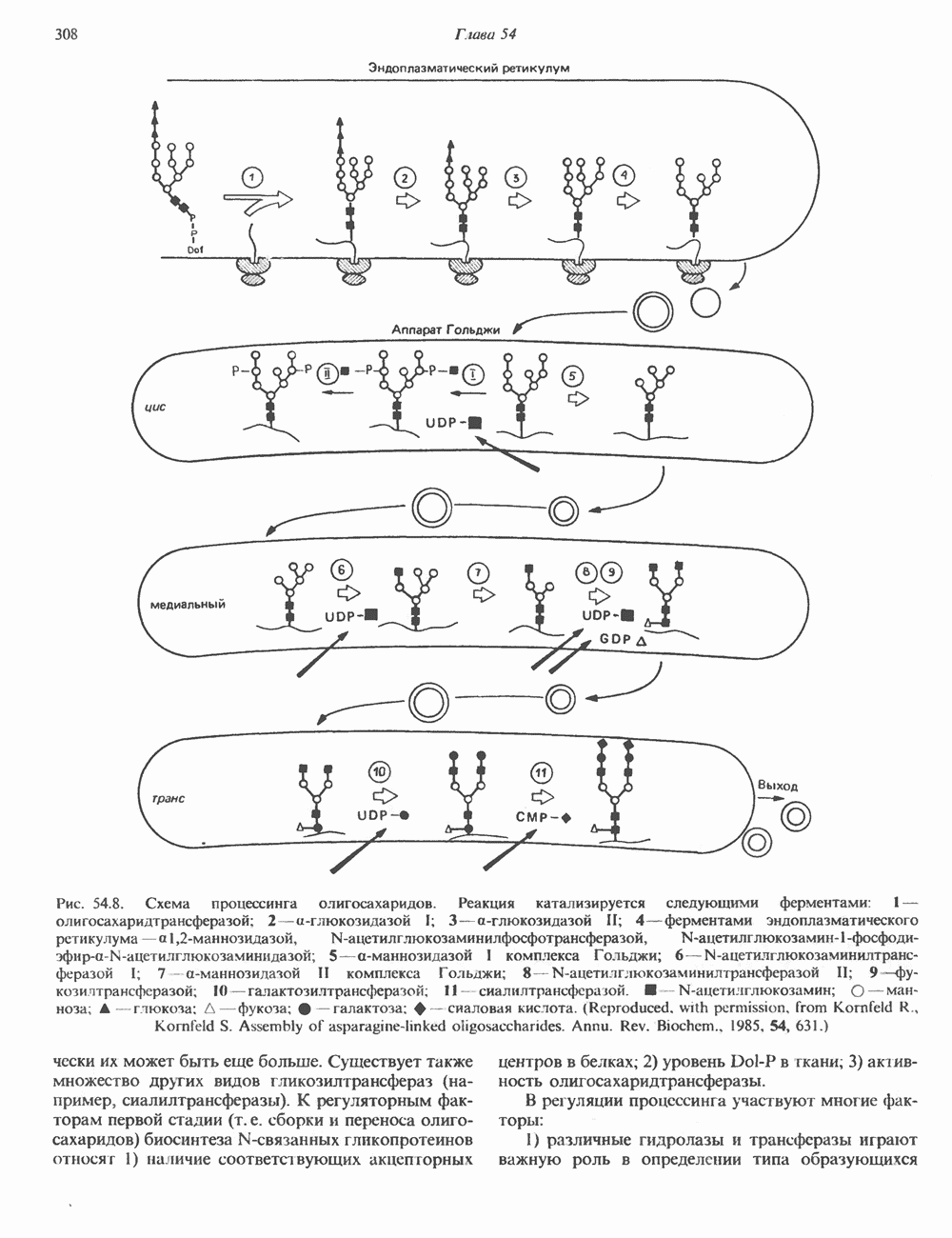
Рис. 56.12. Микроворсинки — это маленькие цитоплазматические выросты эпителиальных клеток, выстилающих тонкий кишечник и увеличивающих всасывающую поверхность. Микроворсинки содержат как актиновые, так и миозиновые филаменты и могут сокращаться подобно мышечным клеткам. Как показано на схеме А, пучки актиновых филаментов входят внутрь каждой микроворсинки; миозиновые филаменты расположены в основании микроворсинок. На схеме Б ориентация актиновых филаментов определена с помощью обработки микроворсинок изолированными «головками» мышечного миозина (тяжелый меромиозин); эти фрагменты сохраняют способность связываться с актиновыми филаментами. При этом образуются комплексы в виде «наконечников стрел», указывающих направление движения филаментов. (Reproduced, with permission, from Lazarides E, Revel J.P.: The molecular bases of cell movement. Sci. Am. [May] 1979; 240, 100.) Актиновые микрофиламенты в немышечных клетках связаны с другими белками, подобными мышечным. На плазматических мембранах в местах прикрепления микрофиламентов и на кончиках микроворсинок присутствует а-актинин. Геодезические купола — «леса» цитоскелета, окружающие ядра эукариотических клеток, состоят из актина, а-актинина и тропомиозина. а-Актинин обнаруживается и в самих актиновых микрофиламентах. Как отмечалось выше, миозин присутствует в основании микроворсинок в связанном с актиновыми микрофиламентами состоянии. Он выявляется также по ходу актиновых волокон, но в виде более тонких и коротких нитей, чем в мышцах. Эти нити, по-видимому, играют роль в поддержании волокнистой структуры актина. Тропомиозин, как упоминалось выше, участвует в образовании куполов, окружающих ядра. Тропомиозин, локализующийся вдоль актиновых микрофиламентов, вероятно, выполняет структурную, а не двигательную функцию. Функция немышечного актина регулируется, по-видимому, несколькими специализированными белками. Профилин предотвращает полимеризацию G-актина даже в присутствии достаточных концентраций магния и хлористого калия. Филамин способствует образованию сети актиновых микрофиламентов. Тропомиозин усиливает формирование пучков стрессовых фибрилл актина. а-Актинин способствует прикреплению актиновых микрофиламентов к мембранам, субстрату и другим клеточным органеллам. Природный пептид цитохалазин разрушает микрофиламенты и предотвращает их полимеризацию. Он часто используется для проверки присутствия микрофиламентов или тестирования их функции. Подвижность клетки, очевидно, детерминируется бахромчатой мембраной или ламеллоподием, содержащим пальцеподобные выросты, называемые филоподиями. Бахрома своей верхушкой присоединяется к субстрату при помощи филоподиев, и клетка как бы перетекает вперед. Бахрома освобождается и вновь складывается на верхушке клетки, пока новые филоподии не присоединятся к субстрату (рис. 56.13). МикротрубочкиМикротрубочки — это важнейший компонент клеточного цитоскелета. Они необходимы для образования и функционирования митотического веретена и, следовательно, присутствуют во всех эукариотических клетках. Эти органеллы выполняют и ряд других клеточных функций. Они определяют внутриклеточное перемещение эндоцитозных и экзоцитозных везикул (пузырьков), служат основным структурным компонентом ресничек и жгутиков и главным белковым компонентом аксонов и дендритов. Микротрубочки поддерживают их структуру и участвуют в аксоплазматическом токе веществ вдоль этих нервных отростков. Микротрубочки представляют собой цилиндры из 13 продольно расположенных протофиламентов,
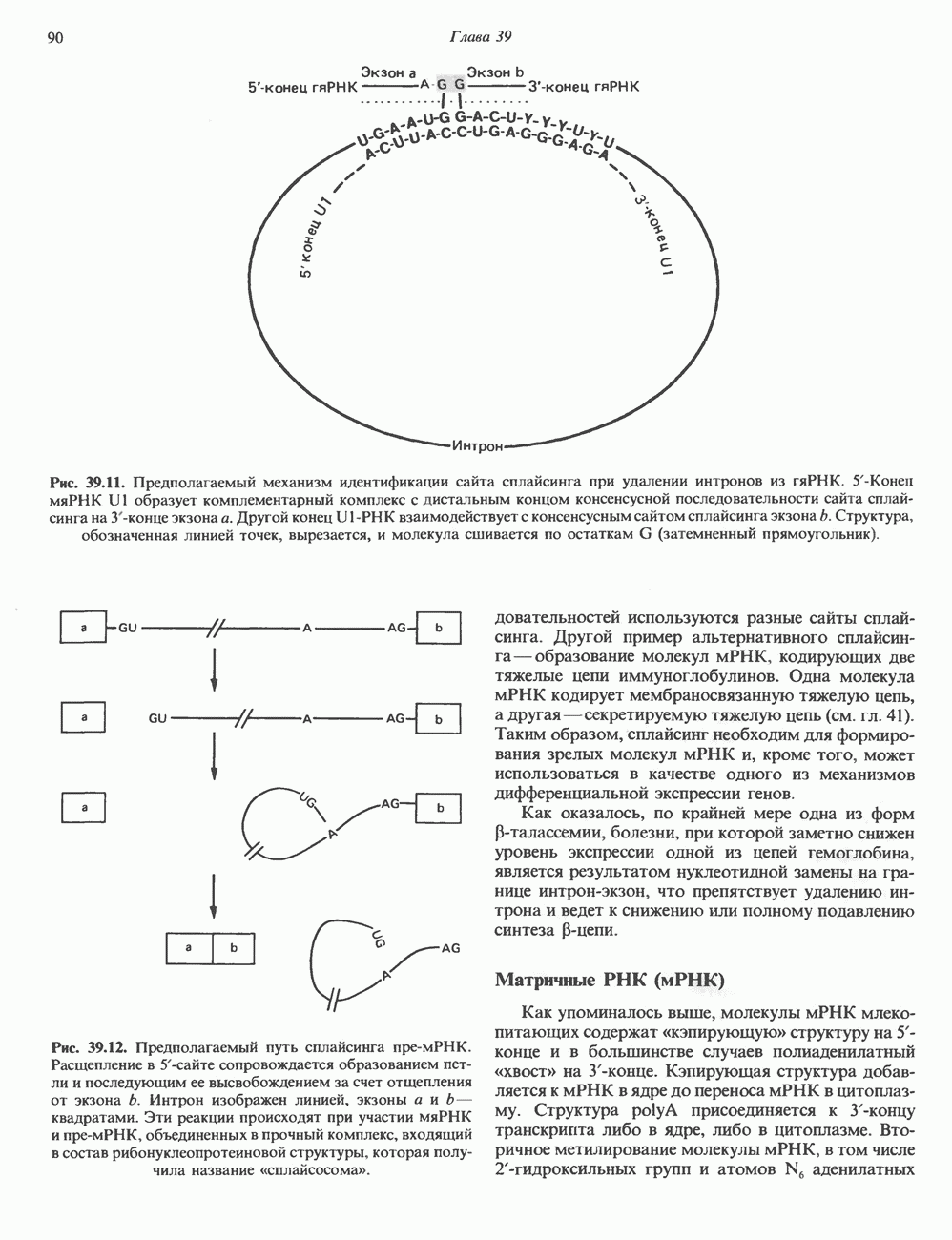
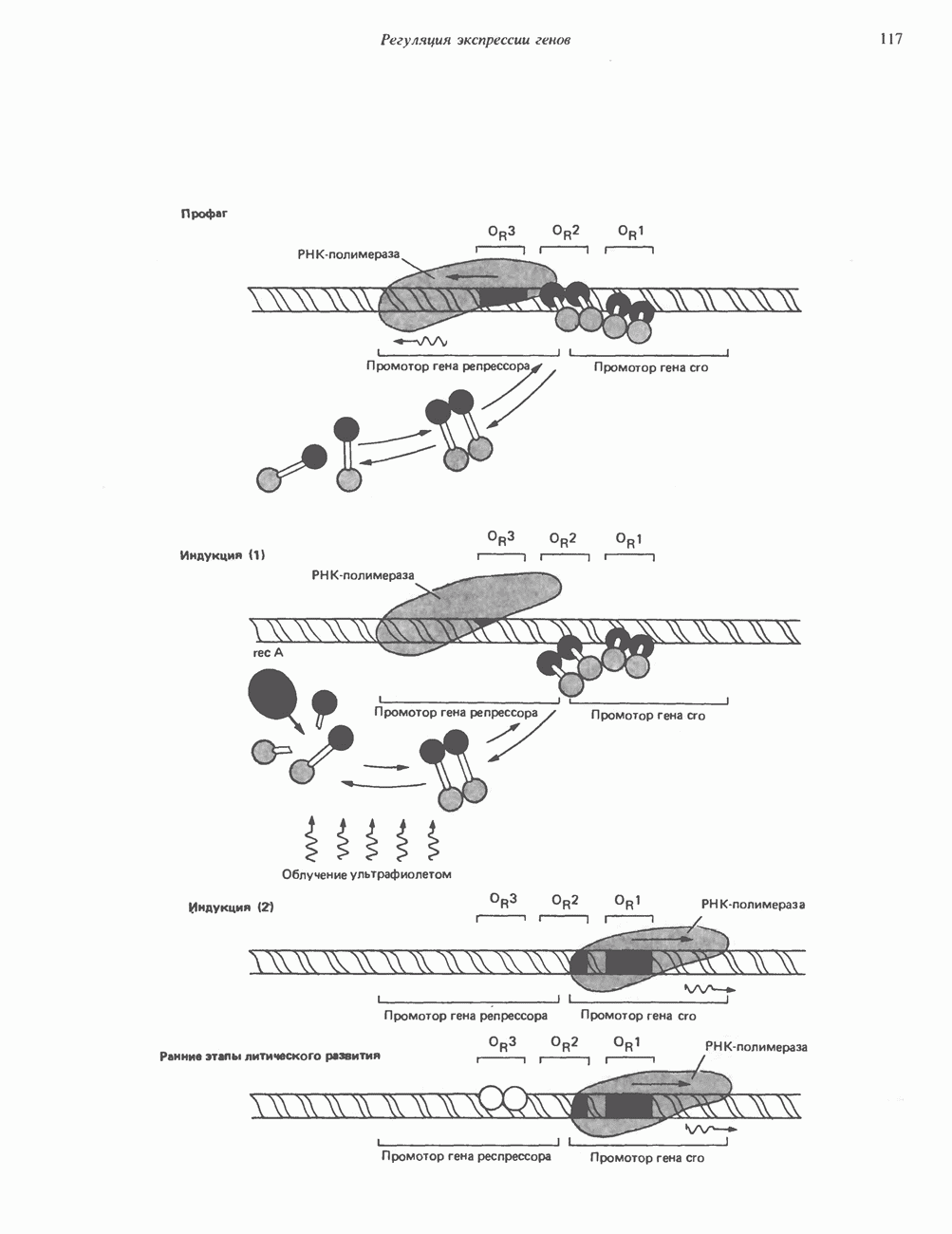
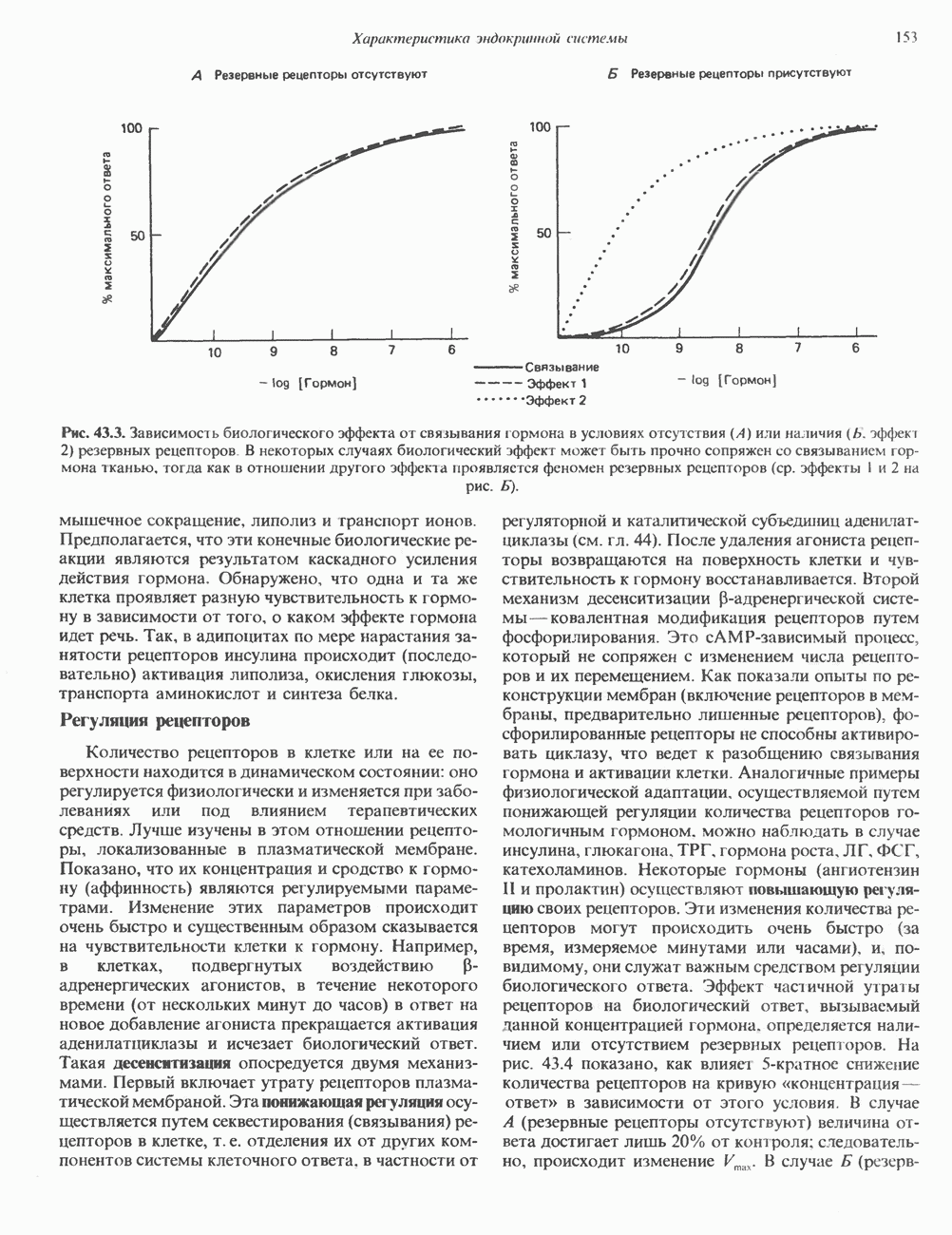
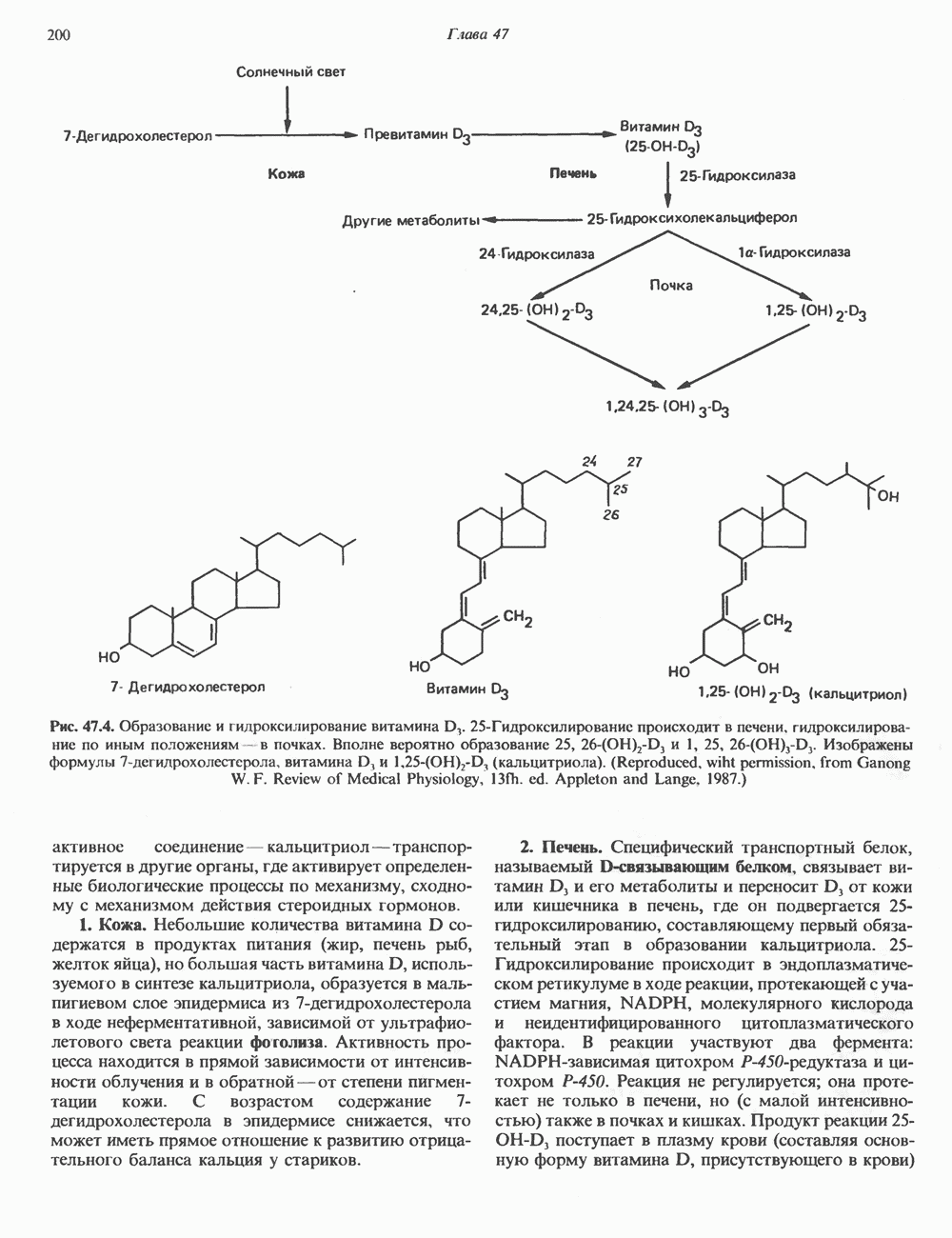
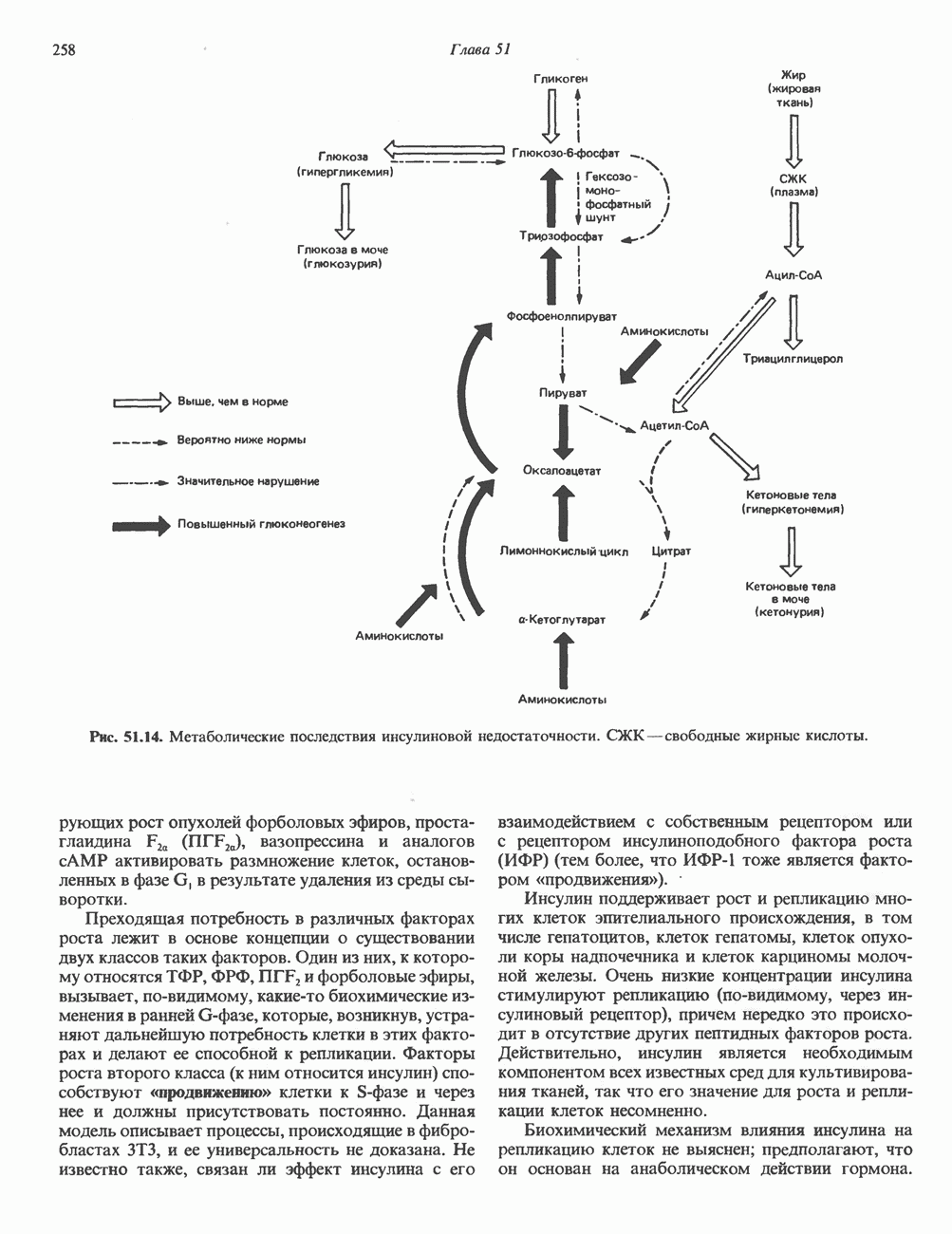
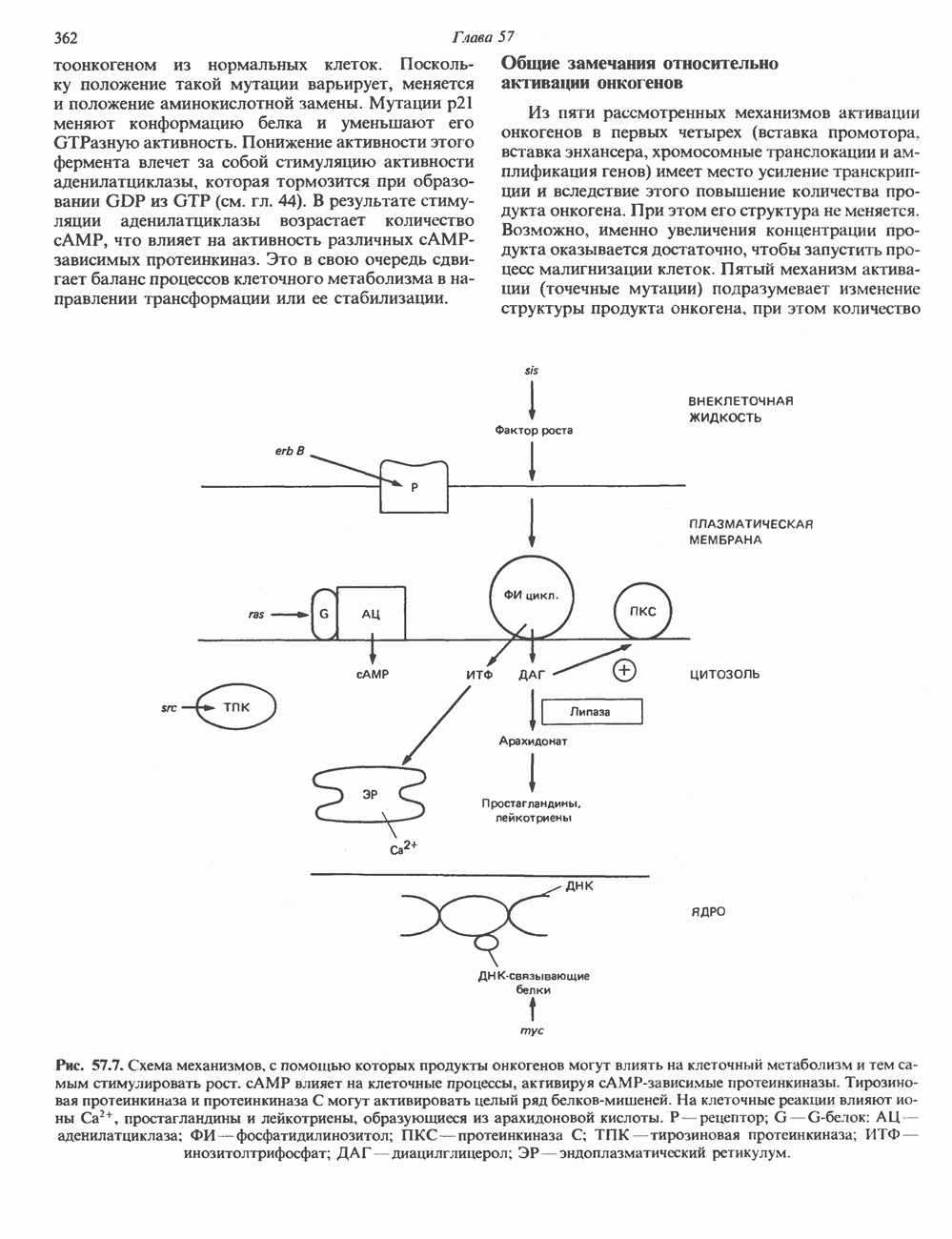
Рис. 56.13. Отдельная клетка тканевой культуры. Структура внизу справа — это бахрома или ламеллоподий, который указывает на ведущий край клетки. Клетка изображена под косым углом, как она движется сквозь субстрат, образуя с помощью бахромы новые точки прикрепления. (Reproduced, with permission, from Lazarides E., Revel J.P.: The molecular basis of cell movement. Sci. Am. [May] 1979; 240, 100.) каждый из которых состоит из димеров а- и р-тубулина (рис. 56.14). а-Тубулин (мол. масса 53000) и Р-тубулин (мол. масса 55 000) являются близкородственными белками. Тубулиновые димеры собираются в протофиламенты, затем в тяжи и, наконец, в цилиндры, как показано на рис. 56.15. Для сборки тубулина в микротрубочки требуются две молекулы GTP на димер тубулина. Два белка, получившие название «высокомолекулярный белок» (ВМБ) и «тау-белок», способствуют образованию микротрубочек, но не обязательны для их сборки. В сборке микротрубочек могут участвовать также кальмодулин и реакции фосфорилирования. Многие известные алкалоиды способны предотвращать сборку микротрубочек. К ним относятся колхицин и его производное демекольцин (применяющийся для лечения острого подагрического артрита), винбластин (алкалоид Vinca, используемый для лечения рака) и гризеофульвин (противогрибковое средство). Микротрубочки «растут» в одном направлении от специфических центров (центриолей) внутри клетки. На каждой хроматиде хромосомы (см. гл. 37) имеется кинетохор, откуда начинается рост микротрубочек. Многие нарушения в делении хромосом являются результатом аномалий в структуре или функции кинетохоров. Движение хромосом в анафазе митоза зависит от микротрубочек, но молекулярный механизм этого эффекта пока не расшифрован.
Рис. 56.14. Электронная микрофотография микротрубочки с деформированной вследствие быстрого замораживания структурой. В левой половине рисунка видна наружная поверхность микротрубочки с пучками протофиламентов, отстоящими друг от друга на 55 нм. Справа микротрубочка лопнула, так что видны ее внутренние стенки и расположенные под углом полосы, разделенные промежутками в 40 нм. Считают, что ретикулум, окружающий микротрубочку, состоит из неполимеризованного тубулина и связанных с микротрубочкой белков. (Reproduced, with permission, from Heuser J. E., Kirschner M. W. Filament organization revealed in platinum replicas of frase-dried cytoskeletons. J. Cell Biol. 1980, 86, 212.) В основании жгутиков и ресничек эукариотических клеток расположена структура, называемая базальным телом; она идентична центриоле и функционирует как центр формирования девятипарной структуры микротрубочек в жгутиках и ресничках. Эти органеллы специализированы для движения. Каждый член пары (дублета) имеет со своим партнером общую стенку из трех протофиламентов; пары соединены между собой гибким белком, нексином. Движение осуществляется путем скольжения дублетов относительно друг друга, что вызывает волнообразные изгибы ресничек. С одним из дублетов ресничек связан большой белок динеин, обладающий АТРазной активностью, которая необходима для скольжения пар микротрубочек. Промежуточные филаментыНедавние исследования продемонстрировали существование внутриклеточной системы волокнистых филаментов. Их диаметр колеблется от 8 до 10 нм, т.е. по толщине они занимают промежуточное положение между микрофиламентами (диаметр 6 нм) и микротрубочками (диаметр 23 нм). Эти филаменты делятся на 6 основных групп, которые имеют общие антигенные детерминанты. Промежуточные филаменты представляют собой прочные относительно стабильные компоненты цитоскелета, не подверженные быстрой сборке и распаду
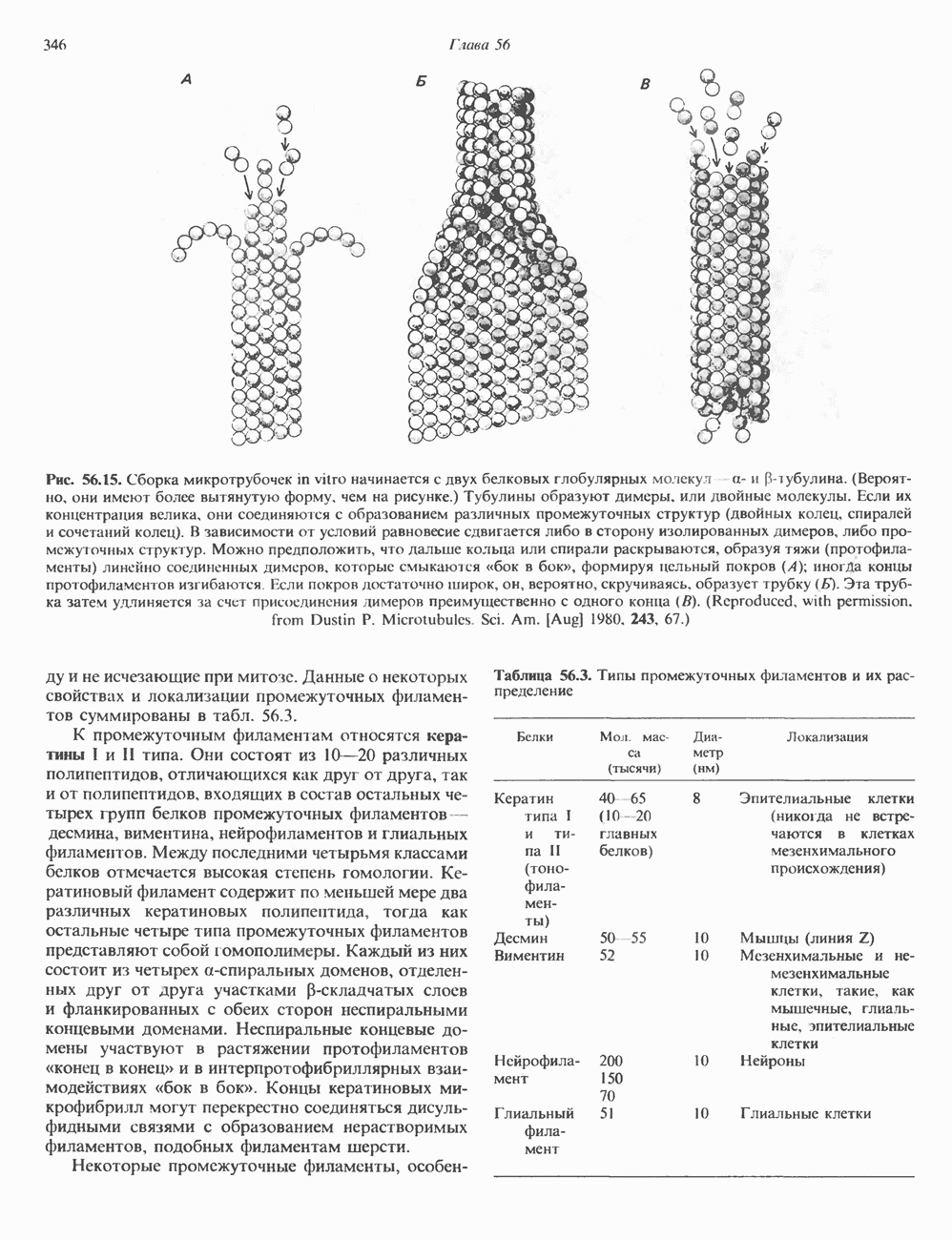
Рис. 56.15. Сборка микротрубочек in vitro начинается с двух белковых глобулярных мотекул а- и Р тубулина. (Вероятно, они имеют более вытянутую форму, чем на рисунке.) Тубулины образуют димеры, или двойные молекулы. Если их концентрация велика, они соединяются с образованием различных промежуточных структур (двойных колец, спиралей и сочетаний колец). В зависимости от условий равновесие сдвигается либо в сторону изолированных димеров, либо промежуточных структур. Можно предположить, что дальше кольца или спирали раскрываются, образуя тяжи (протофила-менты) линейно соединенных димеров, которые смыкаются «бок в бок», формируя цельный покров (А); иногда концы протофиламентов изгибаются. Гели покров достаточно широк, он, вероятно, скручиваясь, образует трубку (Б). Эта трубка затем удлиняется за счет присоединения димеров преимущественно с одного конца (5). (Reproduced, w th permission. from Dustin P. Microtubules. Sci. Am. [Aug] 1980, 243 67.) и не исчезающие при митозе. Данные о некоторых свойствах и локализации промежуточных филаментов суммированы в табл. 56.3. К промежуточным филаментам относятся кератины I и II типа. Они состоят из 10—20 различных полипептидов, отличающихся как друг от друга, так и от полипептидов, входящих в состав остальных четырех групп белков промежуточных филаментов — десмина, виментина, нейрофиламентов и глиальных филаментов. Между последними четырьмя классами белков отмечается высокая степень гомологии. Кератиновый филамент содержит по меньшей мере два различных кератиновых полипептида, тогда как остальные четыре типа промежуточных филаментов представляют собой гомополимеры. Каждый из них состоит из четырех а-спиральных доменов, отделенных друг от друга участками Р-складчатых слоев и фланкированных с обеих сторон неспиральными концевыми доменами. Неспиральные концевые домены участвуют в растяжении протофиламентов «конец в конец» и в интерпротофибриллярных взаимодействиях «бок в бок». Концы кератиновых микрофибрилл могут перекрестно соединяться дисульфидными связями с образованием нерастворимых филаментов, подобных филаментам шерсти. Некоторые промежуточные филаменты, особенно Таблица 56.3. Типы промежуточных филаментов и их распределение
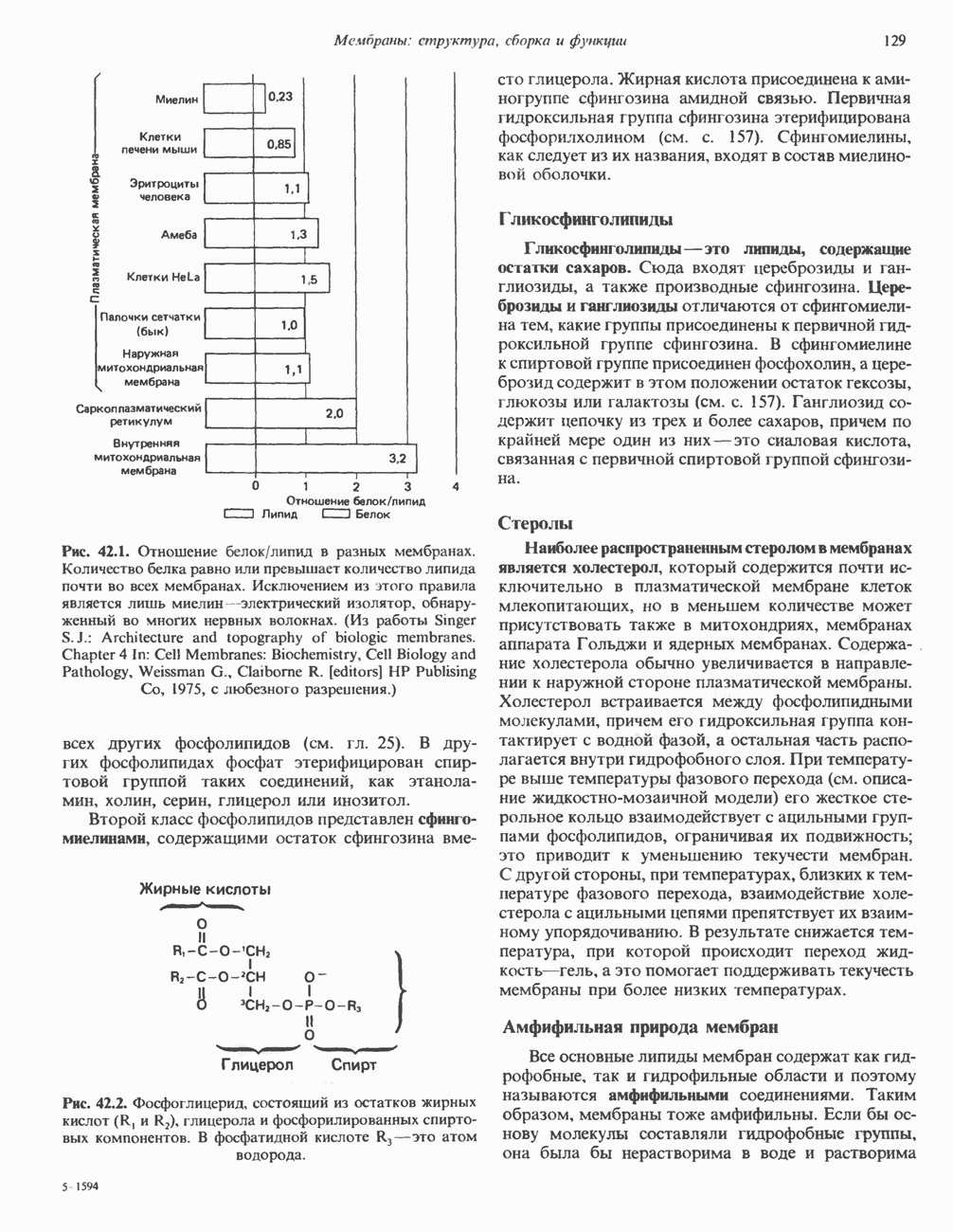
мышечного и мезенхимального происхождения, присутствуют в очень многих тканях. КОЛЛАГЕН Коллаген, главная макромолекула соединительной ткани, является наиболее распространенным белком в животном мире. Он обеспечивает внеклеточный каркас у всех многоклеточных и присутствует практически в любой животной ткани. В тканях млекопитающих существует не менее пяти различных типов коллагена: они образуют семейство молекул со многими общими свойствами. Наиболее характерная особенность коллагеновых молекул — это их трехспиральная структура. Каждая субъединица, или Коллагеновые фибриллы образуют продольные зигзагообразные структуры длиной чуть меньше Еще одна характерная особенность молекулы коллагена заключается в том, что каждый третий остаток тройной спирали a-цепи представляет собой глицин. Глицин — это единственная аминокислота, размер которой позволяет ей поместиться под центральным ядром тройной спиральной молекулы. Таким образом, центральное ядро этой молекулы состоит из глициновых остатков всех трех а-субъединиц. Эту повторяющуюся структуру можно представить как В коллагене млекопитающих около 100 X-положений занято пролином и около 100 Y-положений-4-гидроксипролином. Эти «жесткие» аминокислоты ограничивают вращение полипептид-ного стержня и таким образом увеличивают стабильность тройной спирали. Остатки гидроксипролина придают структуре дополнительную стабильность за счет образования большего количества внутримолекулярных водородных связей (для этого используются окружающие молекулы воды). В некоторых Х-положениях коллагена содержится также
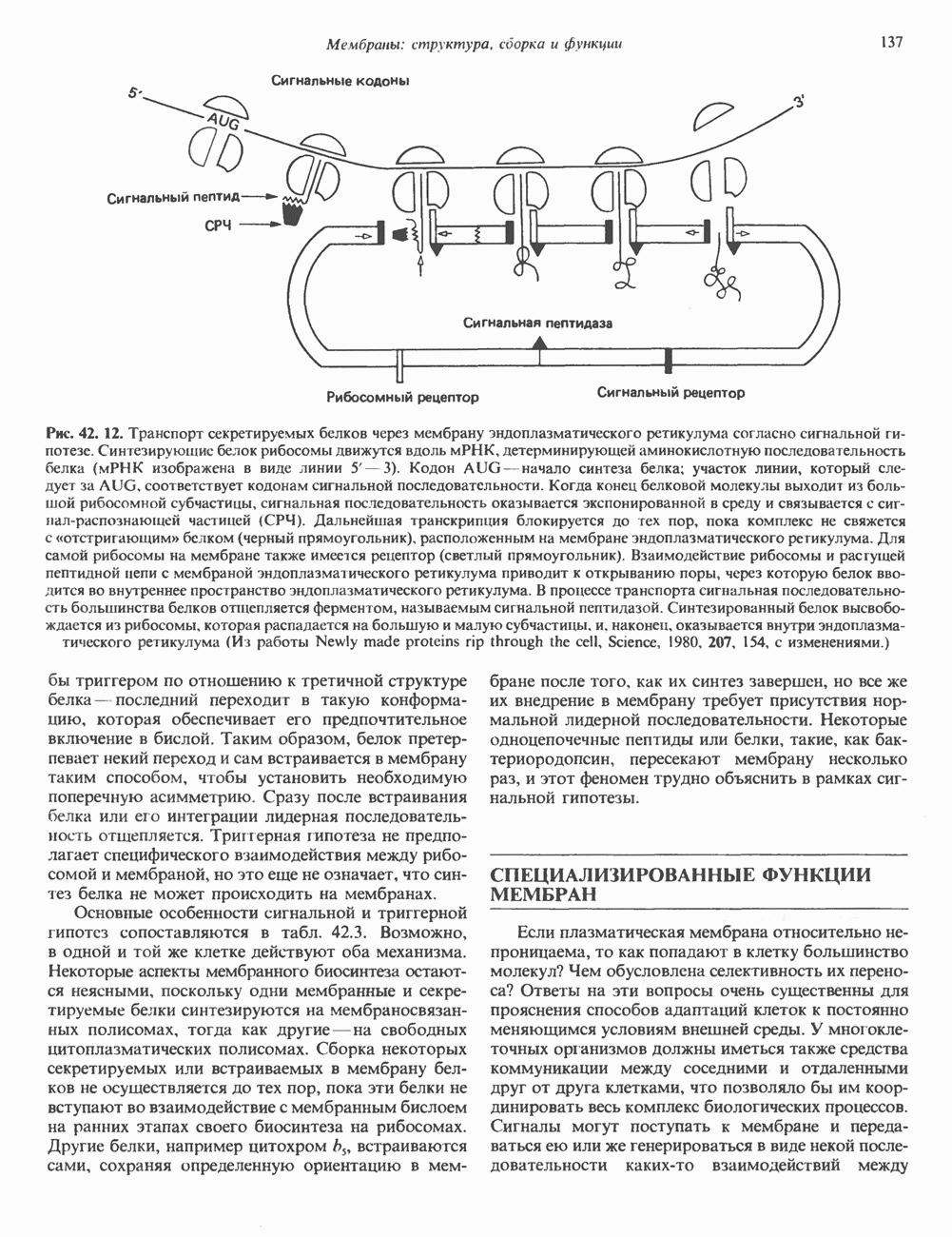
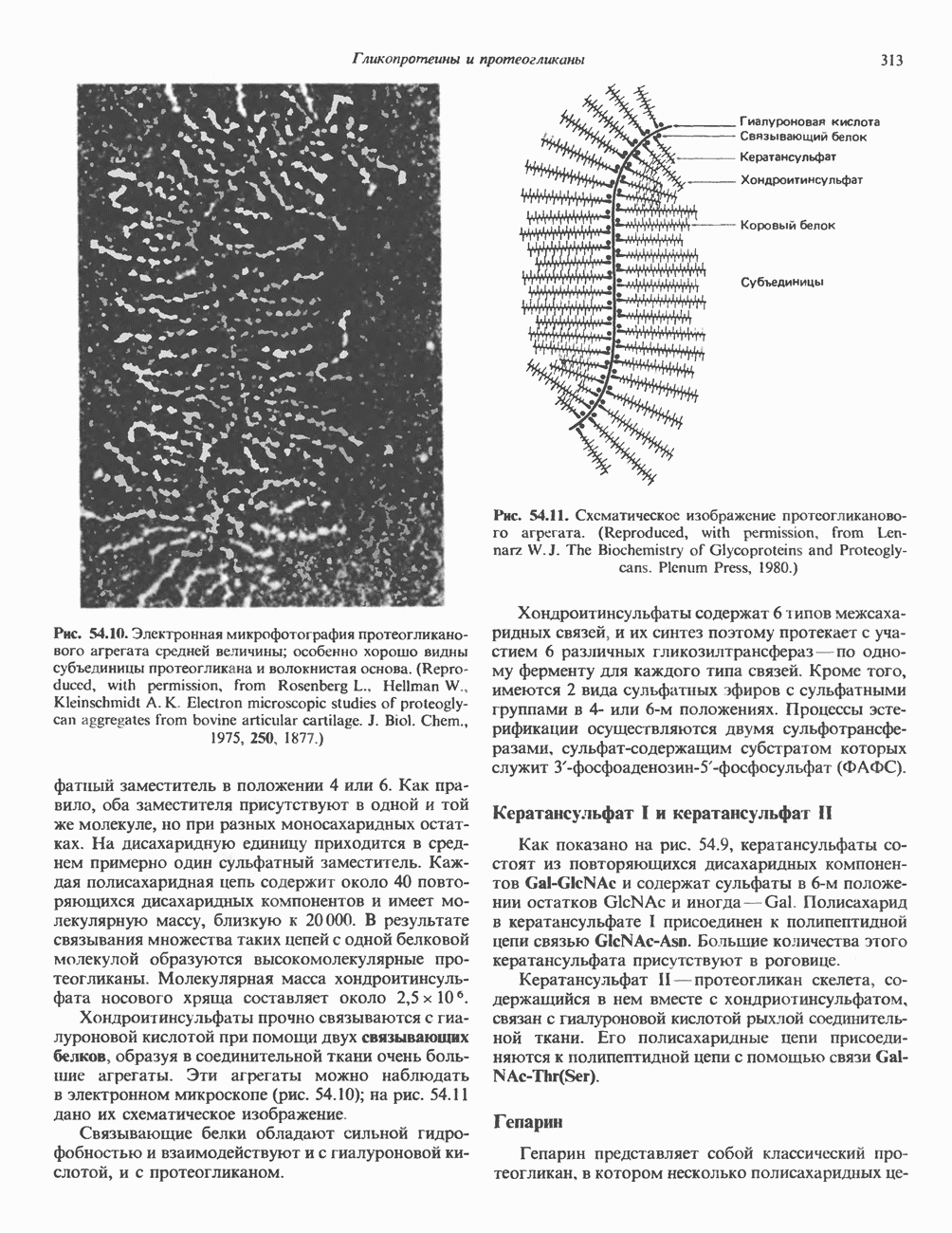
Рис. 56.16. Особенности молекулярной структуры коллагена от первичной последовательности до фибрилл. (Slightly modified and reproduced with permission, from Eyre D. R. Collagen. Molecular diversity in the body’protein scaffold. Science 1980, 207, 1315.) Тройная спираль коллагена стабилизируется многочисленными межценочечными поперечными сшивками Между лизиловыми и гидроксилизиловыми остатками. Химическая природа этих перекрестных связей рассматривается ниже. Зрелый коллаген представляет собой гликопротеин, содержащий сахариды, связанные с остатками гидроксилизина О-гликозидной связью. Данные о коллагенах позвоночных, их распределении в тканях и отличительных особенностях суммированы в табл. 56.4. Синтез коллагенаКоллаген — внеклеточный белок, но он синтезируется в виде внутриклеточной молекулы-предшественника, которая перед образованием фибрилл зрелого коллагена подвергается посттрансляционной модификации. Подобно всем секретируемым белкам, предшественник коллагена претерпевает процессинг в ходе прохождения через эндоплазматический ретикулум и комплекс Гольджи до появления во внеклеточном пространстве. Наиболее ранним предшественником коллагена является препроколлаген, который содержит на N-конце лидерную, или сигнальную, последовательность примерно из 100 аминокислот. Препроколлаген образуется на рибосомах, прикрепленных к эндоплазматическому Таблица 56.4. Генетически различные коллагены позвоночных. У высших животных присутствуют по меньшей мере пять различных молекул, содержащих семь генетически различающихся а-цепей (см. скан) ретикулуму. Когда сигнальная последовательность проникает в везикулярное пространство эндоплазматического ретикулума, лидерная последовательность отщепляется, а Молекула проколлагена содержит на N- и Суконцах пептиды с мол. массой соответственно 20 000 и 30000—35000, которые отсутствуют в зрелом коллагене. Оба этих пропептида содержат остатки цистеина. N-концевой пропептид образует только внутрицепочечные дисульфидные связи, С-концевые пептиды образуют и внутри- и межцепочечные сульфидные связи. Вслед за образованием этих связей молекулы проколлагена собираются в тройную спираль. После формирования тройной спирали дальнейшее гидроксилирование пролиловых и лизиловых остатков становится невозможным. Гликозилтрансфераза, которая переносит глюкозу или галактозу на остатки гидроксилизина, также может действовать только на a-цепи проколлагена, не имеющие еще спиральной конфигурации. После завершения этого внутриклеточного процессинга молекулы гликозилированного проколлагена продвигаются к наружной поверхности клетки через комплекс Гольджи. Внеклеточные аминопротеаза и карбоксипротеаза проколлагена удаляют соответственно аминоконцевой и карбоксиконцевой про пептиды. Вновь образованные молекулы коллагена содержат примерно 1000 аминокислот на цепь и спонтанно собираются в коллагеновые фибриллы, неотличимые от зрелых фибрилл, присутствующих в тканях. Однако эти фибриллы не обладают силой напряжения зрелых коллагеновых фибрилл до тех пор, пока не произойдет перекрестное связывание их рядом ковалентных связей. Внеклеточный медь-содержащий фермент, лизилоксидаза, осуществляет окислительное дезаминирование е-аминогрупп некоторых лизиловых и гидроксилизиловых остатков коллагена с образованием реактивных альдегидов. Альдегиды могут формировать Шиффовы основания с е-аминогруппами других лизинов, гидроксилизинов или даже гликозилированных гидроксилизинов. Эти Шиффовы основания подвергаются химической перегруппировке, обеспечивая стабильные ковалентные сшивки. Альдегидный компонент, образующийся из гидроксилизина, формирует более стабильную перекрестную связь, чем альдегид из лизилового остатка. Альдольные мостики также обеспечивают перекрестное связывание (сшивку) молекул. Данные о внутриклеточном и внеклеточном процессинге молекулы-предшественника коллагена суммированы в табл. 56.5. Таблица 56.5. Порядок и локализация этапов процессинга предшественника коллагена (содержащего повторяющуюся структуру [Gly-X-Y]n
Клетки, секретирующие коллаген, секретируют также фибронектнн, большой гликопротеин, присутствующий на клеточной поверхности, во внеклеточном матриксе и в крови. Фибронектин связывается с агрегирующимися проколлагеновыми фибриллами и меняет кинетику образования фибрилл в перицеллюлярном матриксе. С фибронектином и проколлагеном в этом матриксе ассоциированы протеогликаны гепарансульфат и хондроитинсульфат (см. гл. 54). Хрящ—это внеклеточный матрикс, в котором коллаген обусловливает силу напряжения, а протеогликаны — выраженную эластичность.
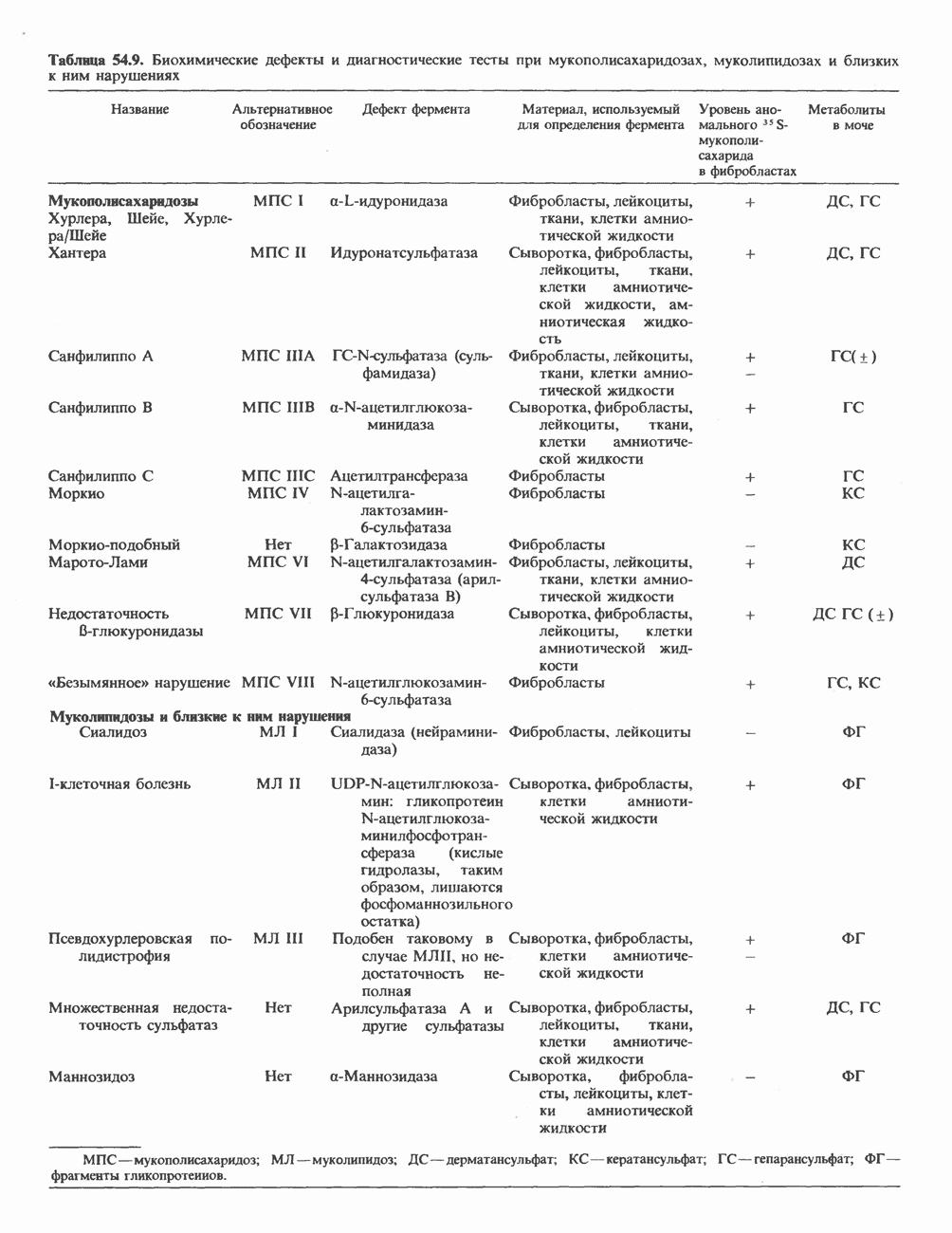
Рис. 56.17. Сайты мутаций в молекуле проколлагена типа I. СЭД—синдром Элерса — Данлоса; СМ—синдром Марфа-на. НО — несовершенный остеогенез. (Reproduced, with permission, from Prockop D. J., Kivirikko K.I. Heritable diseases of collagen. N. Engl. J. Med. 1984, 311, 376.) Таблица 56.6. Молекулярные дефекты, лежащие в основе четырех наследственных коллагенозов. (Reproduced, with permission, from Prockop D.J., Kivirikko К. I. Heritable diseases of collagen. N. Engl. J. Med., 1984, 311, 376.) (см. скан) Наследственные дефекты коллагена и его сборкиНаследственные болезни, обусловленные аномалиями коллагена, носят название коллагенозов. Наиболее известные среди них: несовершенный остеогенез, синдром Марфана, синдром Элерса — Данлоса и синдром Менке (синдром курчавых волос). Первоначально эти болезни диагностировали только на основании фенотипа пробандов, но но мере накопления знаний о структуре и функции коллагена становились понятными молекулярные механизмы, лежащие в основе этих синдромов. Многие клинические симптомы объясняются нарушением процессинга предшественников коллагена из-за аномалий самого предшественника или дефектов в ферментной системе процессинга. Понимание молекулярной природы нарушения позволяет предсказать характер наследования патологии (рецессивный или доминантный). В табл. 56.6 суммированы молекулярные дефекты и их последствия при некоторых наследственных коллагенозах человека, а на рис. 56.17 указаны дефектные сайты в молекуле проколлагена типа I. ЛИТЕРАТУРАAdelstein R. S., Eisenberg R. Regulation and kinetics of actin — myosin ATP interaction, Annu. Rev. Biochem., 1980, 49, 921. Adelstein R. S. et al. Phosphorylation of muscle contractile proteins, Fed. Proc., 1980, 39, 1544. Barany М.. Barany K. Phosphorylation of the myofibrillar proteins, Annu. Rev. Physiol., 1980 42, 275. Bornstein P., Sage H. Structurally distinct collagen types, Annu. Rev. Biochem., 1980, 49, 957. Caplan A. Cartilage, Sci. Am. (Oct.), 1984. 250, 84. Clark М., Spudich J. A. Nonmuscle contractile proteins: The role of actin and myosin in cell motility and shape determination, Annu. Rev. Biochem., 1977, 46, 797. DeCrombrugghe B., Past an I. Structure and regulation of a collagen gene. Trends Biochem. Sci., 1982, 7, 11. Dustin P. Microtubules, Sci. Am. (Aug.), 1980, 243, 67. Eyre D. R. et al. Cross-linking in collagen and elastin, Annu. Rev. Biochem., 1984, 53, 717. Fuchs E., Hanukoglu I. Unraveling the structure of intermediate filaments. Cell, 1983, 34, 332. Heuser J. E., Kirschner M. W. Filament organization revealed in platinum replicas of freeze-dried cytoskelctons. J. Cell Biol., 1980, 86, 212. Kleinman H.K., Klebe R.J., Martin G.R. Role of collagenous matrices in the adhesion and growth of cells, J. Cell Biol., 1981, 88, 473. Lcizarides E. Intermediate filaments: A chemically heterogeneous. developmentally regulated class of proteins. Annu. Rev. Biochem., 1982. 51, 219. Lazarides E., Revel J. P. The molecular basis of cell movement, Sci. Am. (May), 1979, 240. 100. Murray J. М., Weber A. The cooperative action of muscle proteins, Sci. Am. (Feb.), 1974, 230, 59. Prockop D.J., Kivirikko К. I. Heritable diseases of collagen, N. Engl. J. Med., 1984. 311. 376. Sandberg L. B., Soskel N. Т., Leslie J. G. Elastin structure, biosynthesis, and relation to disease states, N. Engl, J. Med., 1981, 304, 566.
|
 |
Оглавление
|



























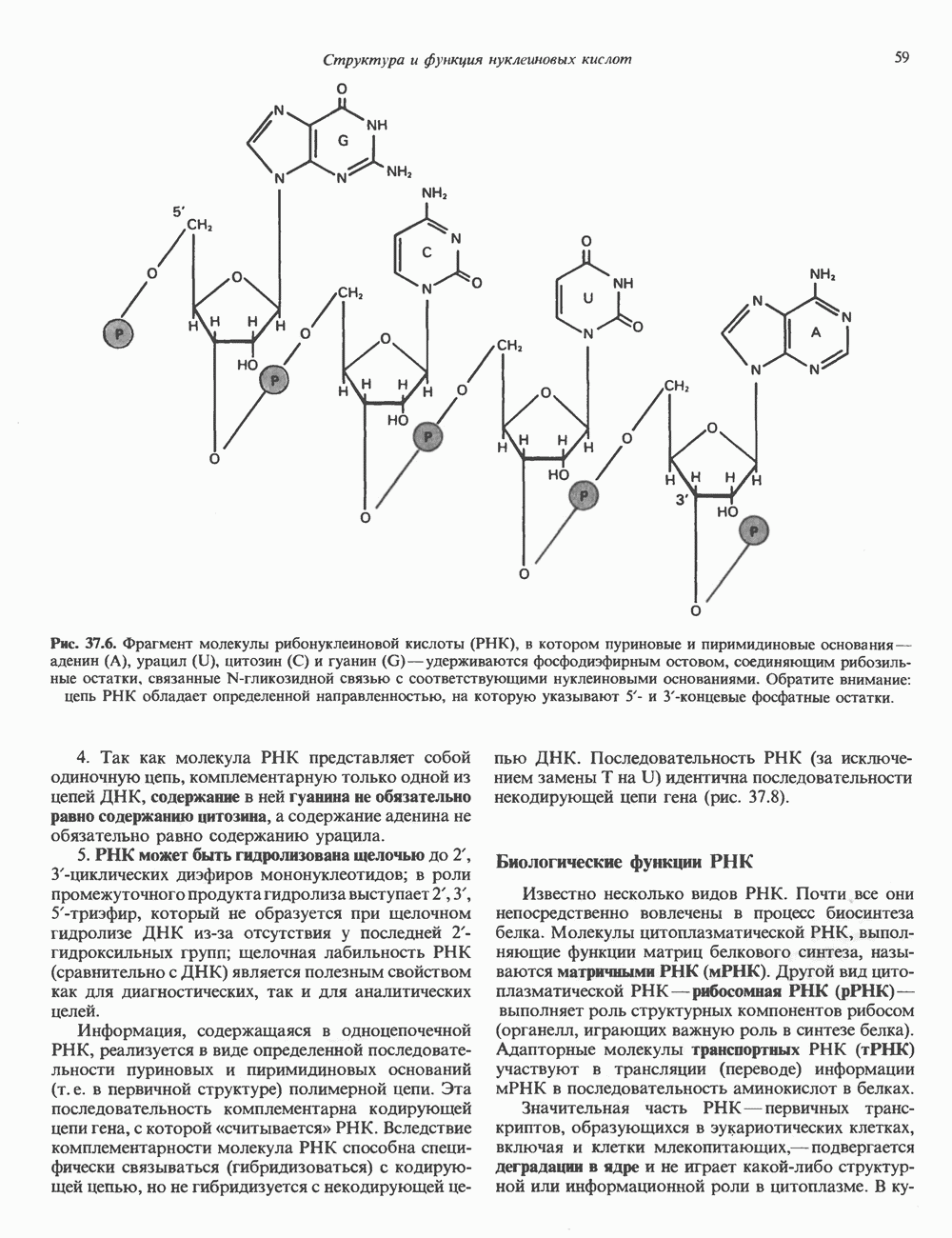





























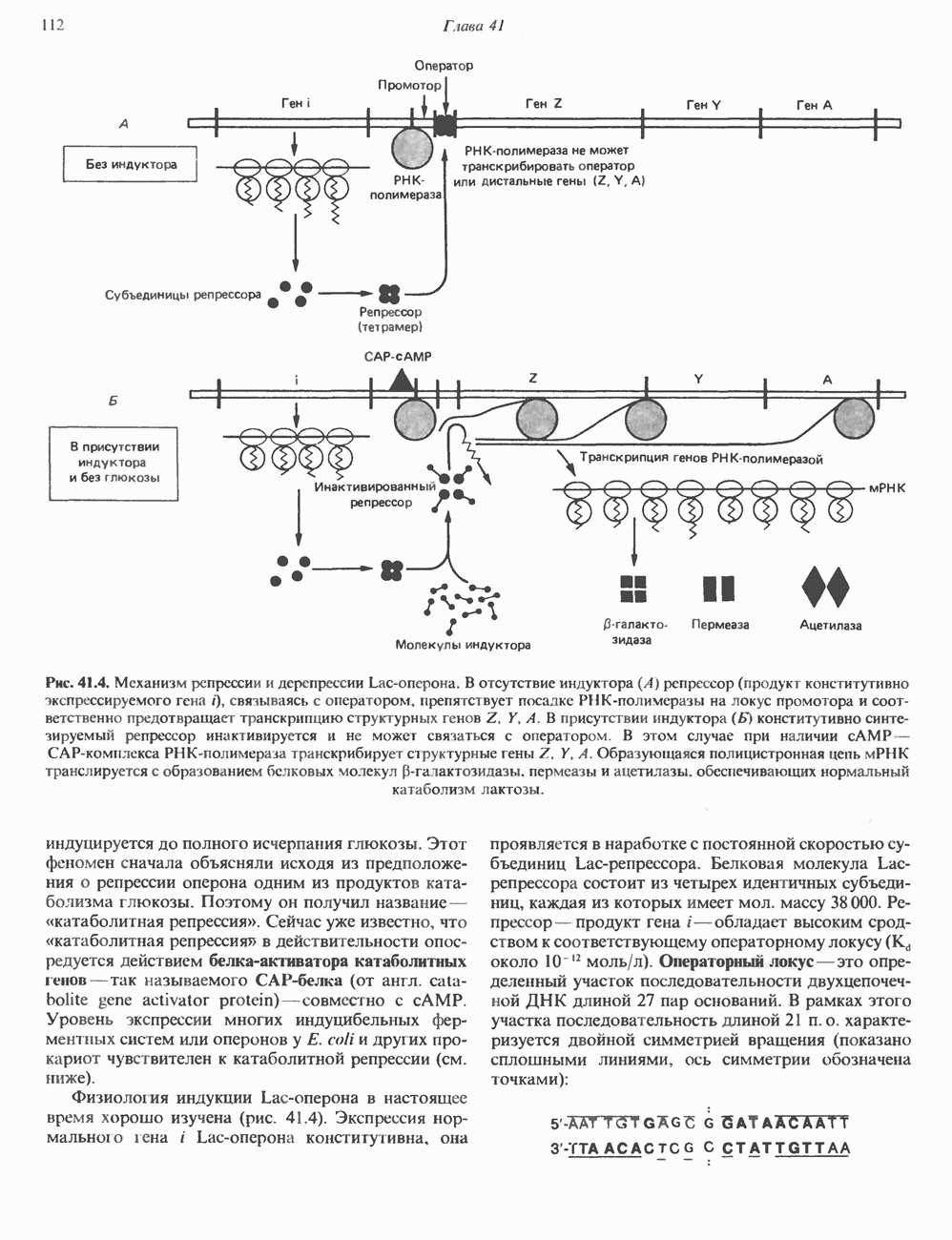

























































































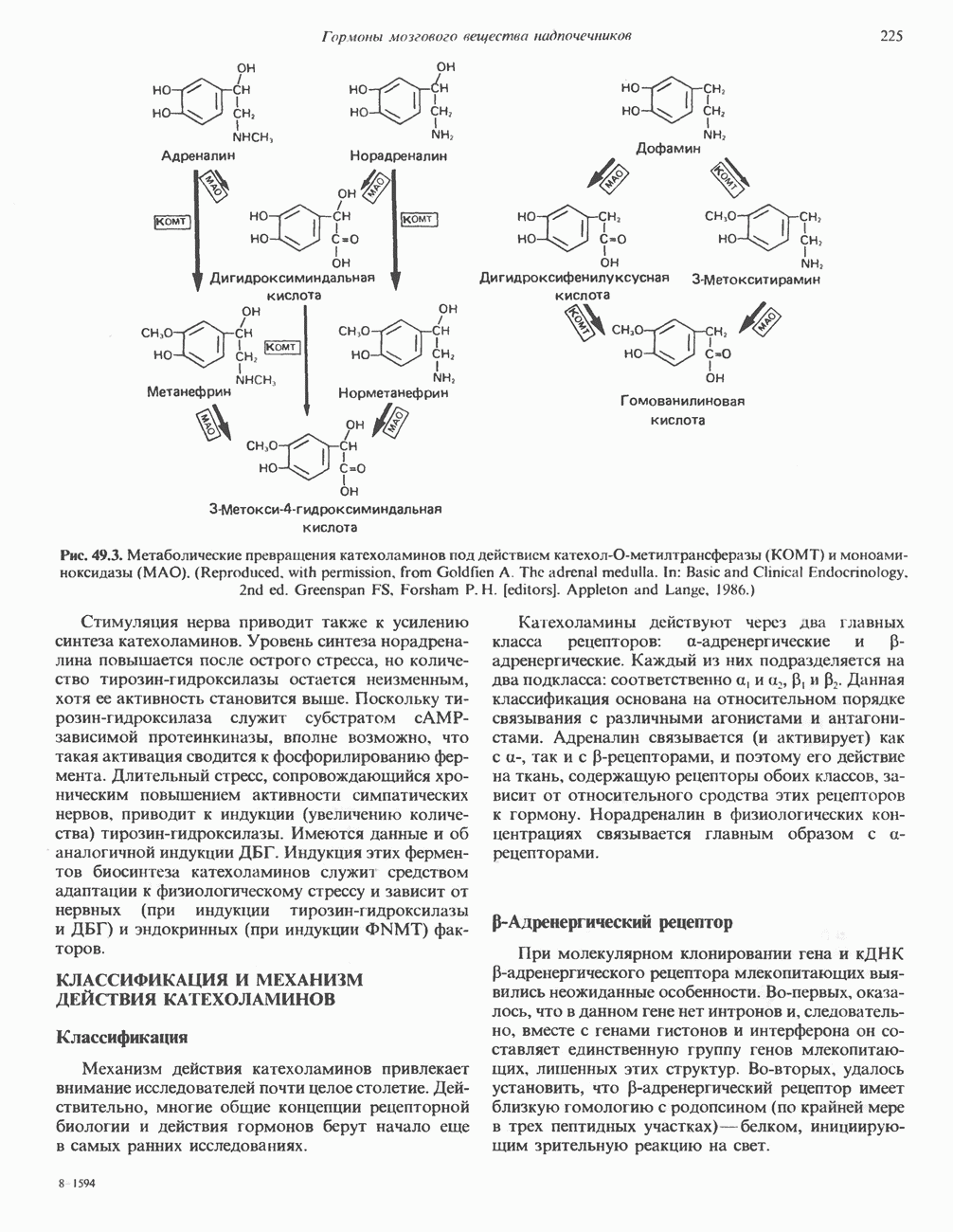







































































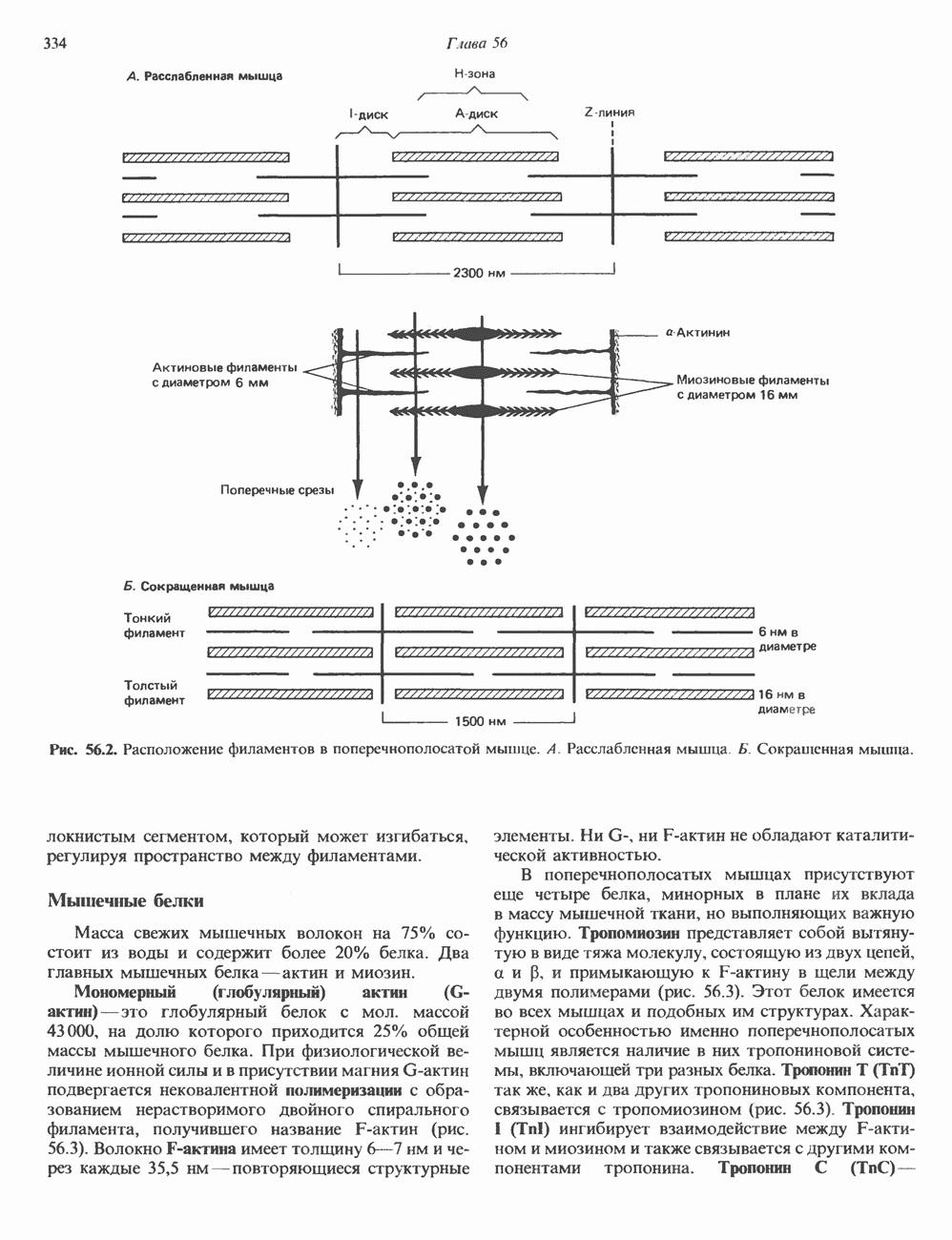
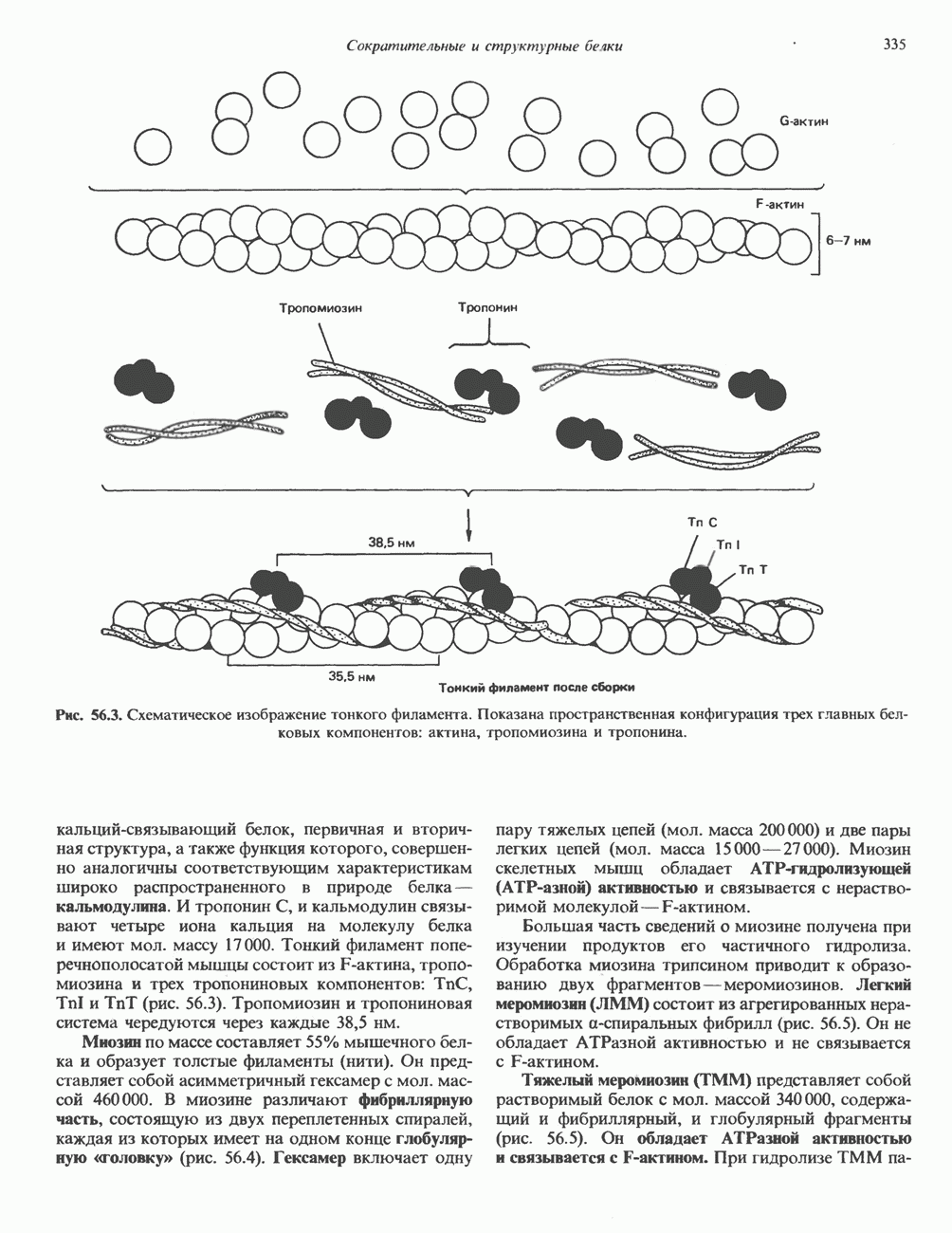





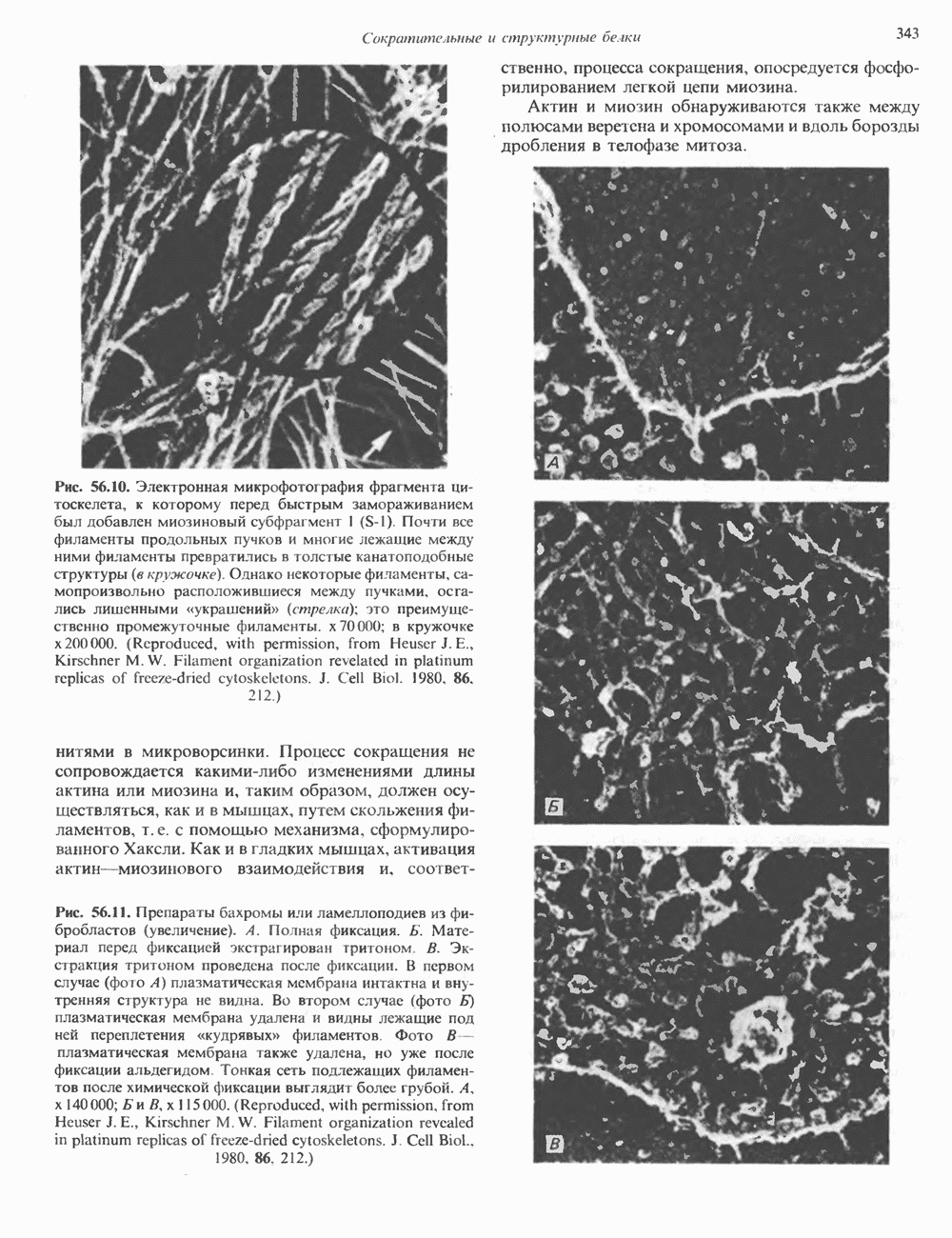






































 -актин. Оба они могут присутствовать в одной и той же клетке и даже, вероятно, сополимеризоваться в одну и ту же нить. В цитоплазме клеток актин образует микрофиламенты диаметром 7-9,5 нм. которые часто имеют вид пучков спутанных сетей. Пучки микрофиламентов располагаются под плазматической мембраной покоящихся клеток и называются «стрессовыми» фибриллами. Эти фибриллы имеют структуру двойной спирали и декорированы миозиновым фрагментом S-1 (рис. 56.10). Стрессовые фибриллы исчезают при увеличении клеточной подвижности, а также при злокачественной трансформации клеток химическими агентами или онкогенными вирусами.
-актин. Оба они могут присутствовать в одной и той же клетке и даже, вероятно, сополимеризоваться в одну и ту же нить. В цитоплазме клеток актин образует микрофиламенты диаметром 7-9,5 нм. которые часто имеют вид пучков спутанных сетей. Пучки микрофиламентов располагаются под плазматической мембраной покоящихся клеток и называются «стрессовыми» фибриллами. Эти фибриллы имеют структуру двойной спирали и декорированы миозиновым фрагментом S-1 (рис. 56.10). Стрессовые фибриллы исчезают при увеличении клеточной подвижности, а также при злокачественной трансформации клеток химическими агентами или онкогенными вирусами.

 Почти все филаменты продольных пучков и многие лежащие между ними филаменты превратились в толстые канатоподобные структуры (в кружочке). Однако некоторые филаменты, самопроизвольно расположившиеся между пучками, остались лишенными «украшений» (стрелка) - это преимущественно промежуточные филаменты. х 70 000; в кружочке х 200000. (Reproduced, with permission, from Heuser J. E., Kirschner M. W. Filament organization revelated in platinum replicas of freeze-dried cytoskelctons. J. Cell Biol. 1980, 86. 212.)
Почти все филаменты продольных пучков и многие лежащие между ними филаменты превратились в толстые канатоподобные структуры (в кружочке). Однако некоторые филаменты, самопроизвольно расположившиеся между пучками, остались лишенными «украшений» (стрелка) - это преимущественно промежуточные филаменты. х 70 000; в кружочке х 200000. (Reproduced, with permission, from Heuser J. E., Kirschner M. W. Filament organization revelated in platinum replicas of freeze-dried cytoskelctons. J. Cell Biol. 1980, 86. 212.)






 -цепь, представляет собой левозакрученную спираль, у которой на виток приходится по три аминокислотных остатка. Три такие левые спирали закручиваются далее в правую суперспираль. В результате формируется жесткая палочковидная молекула с диаметром 1,4 нм и длиной около 300 нм. Затем эти молекулы коллагена ассоциируют в фибриллы (рис. 56.16).
-цепь, представляет собой левозакрученную спираль, у которой на виток приходится по три аминокислотных остатка. Три такие левые спирали закручиваются далее в правую суперспираль. В результате формируется жесткая палочковидная молекула с диаметром 1,4 нм и длиной около 300 нм. Затем эти молекулы коллагена ассоциируют в фибриллы (рис. 56.16).
 длины тройной спирали. Между концом одной тройной спирали и началом следующей имеется пространство, которое может служить местом отложения кристаллов гидроксилапатита при образовании кости. Диаметр коллагеновых фибрилл колеблется от 10 до 100 нм, и под микроскопом они видны в виде пучков в экстрацеллюлярном матриксе соединительной ткани.
длины тройной спирали. Между концом одной тройной спирали и началом следующей имеется пространство, которое может служить местом отложения кристаллов гидроксилапатита при образовании кости. Диаметр коллагеновых фибрилл колеблется от 10 до 100 нм, и под микроскопом они видны в виде пучков в экстрацеллюлярном матриксе соединительной ткани.
 где X и Y-любые аминокислоты, кроме глицина.
где X и Y-любые аминокислоты, кроме глицина.
 -гидроксипролин, а в Y-положениях-5-гидроксилизин.
-гидроксипролин, а в Y-положениях-5-гидроксилизин.

 -конец проколлагена продолжает продвигаться в этом пространстве. Здесь пролиновые и лизиновые остатки в Y-положении пептида
-конец проколлагена продолжает продвигаться в этом пространстве. Здесь пролиновые и лизиновые остатки в Y-положении пептида  подвергаются действию соответственно пролил-4-гидроксилазы и лнзилгидрокснлазы. Пролил-З-гидроксилаза действует на пролиловые остатки в Х-положениях, непосредственно предшествующие 4-гидроксипролину в Y-положениях.
подвергаются действию соответственно пролил-4-гидроксилазы и лнзилгидрокснлазы. Пролил-З-гидроксилаза действует на пролиловые остатки в Х-положениях, непосредственно предшествующие 4-гидроксипролину в Y-положениях.

