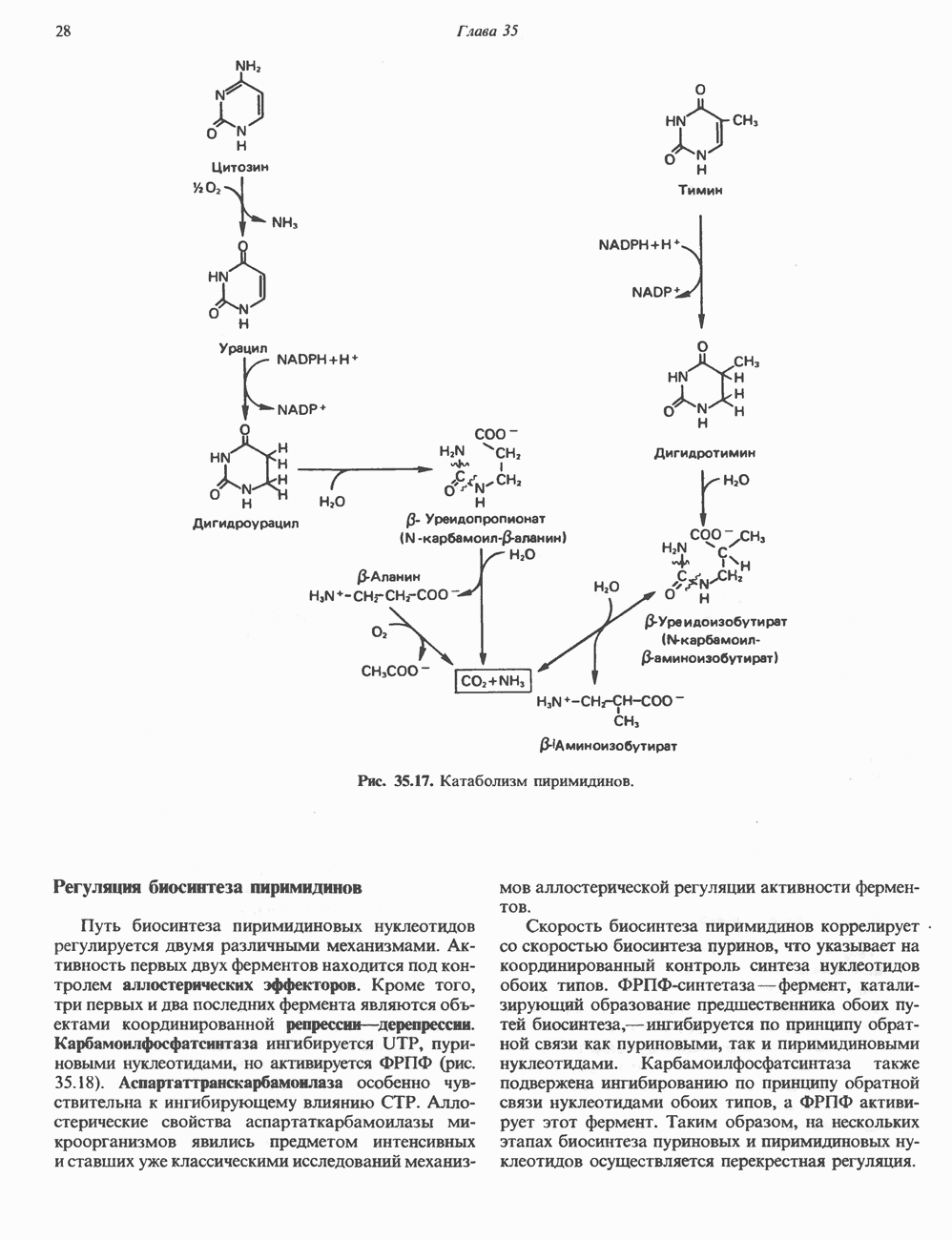
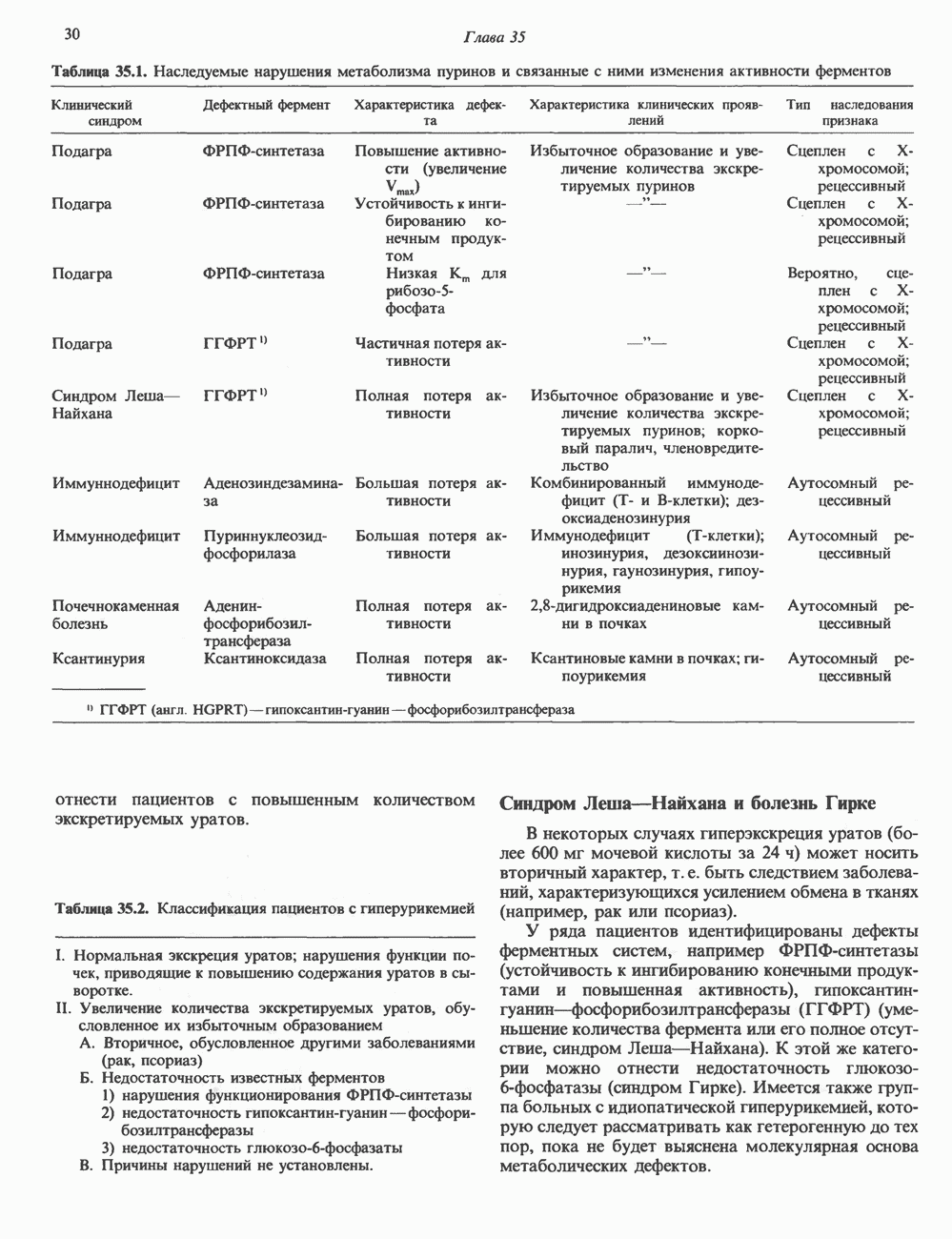
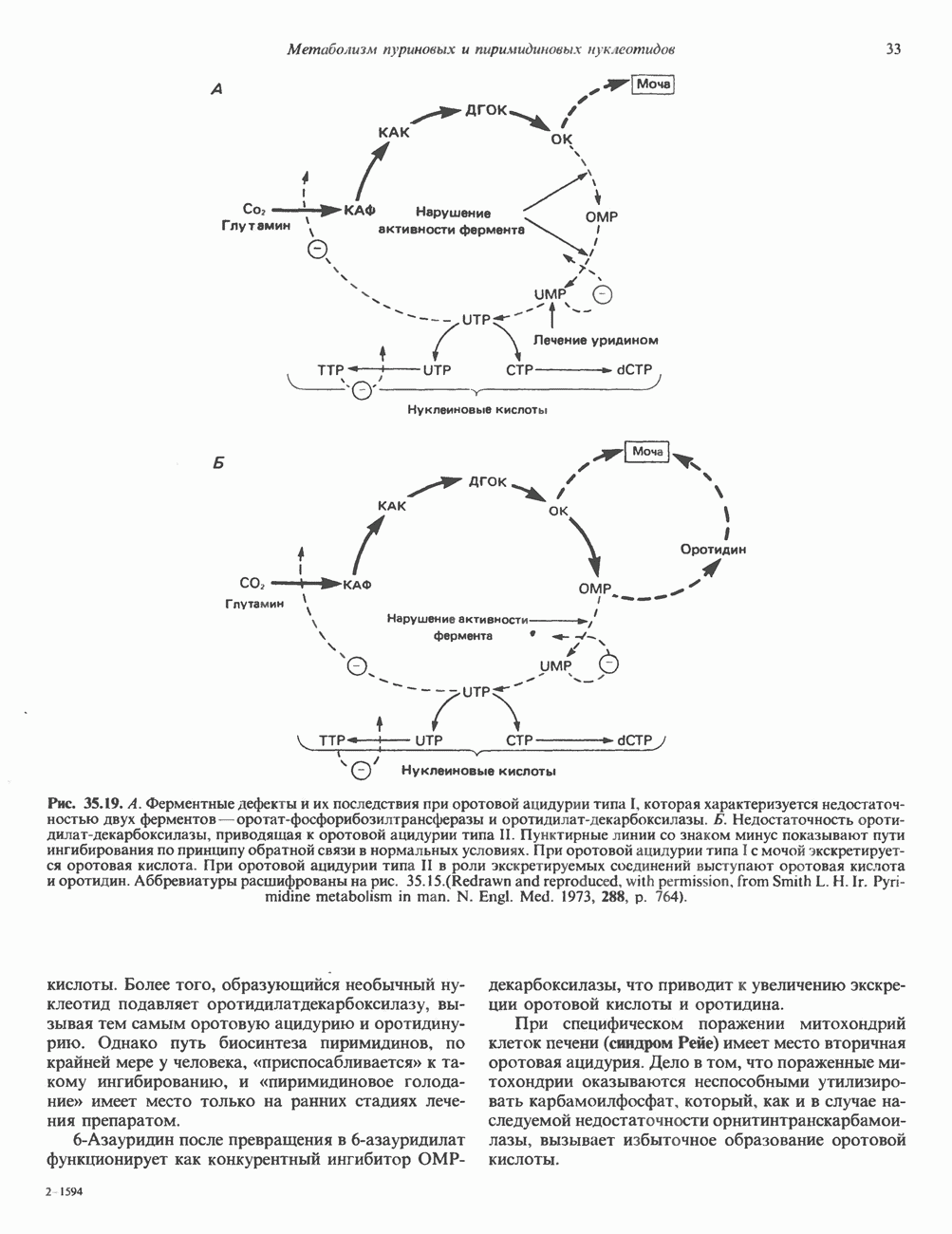
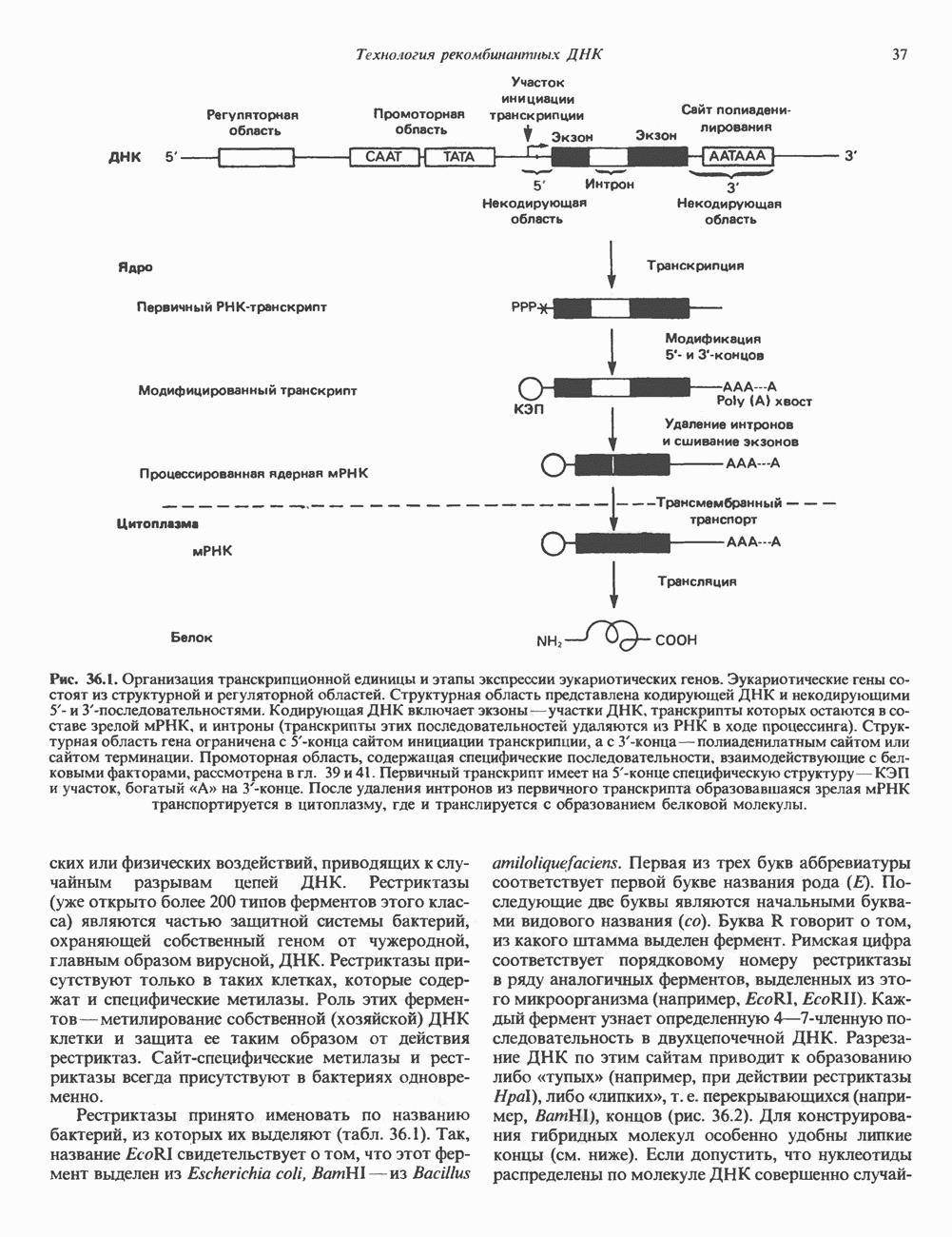
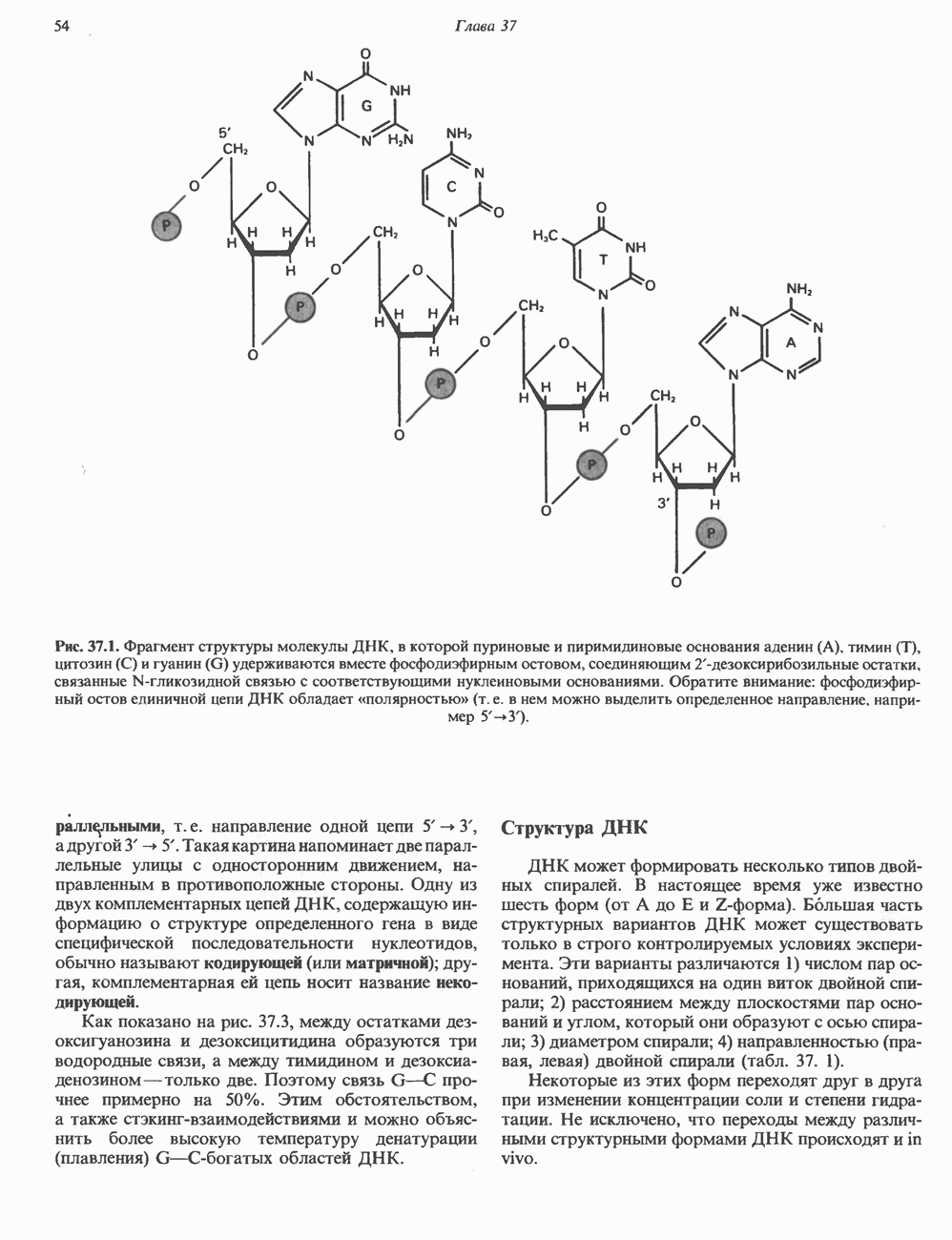
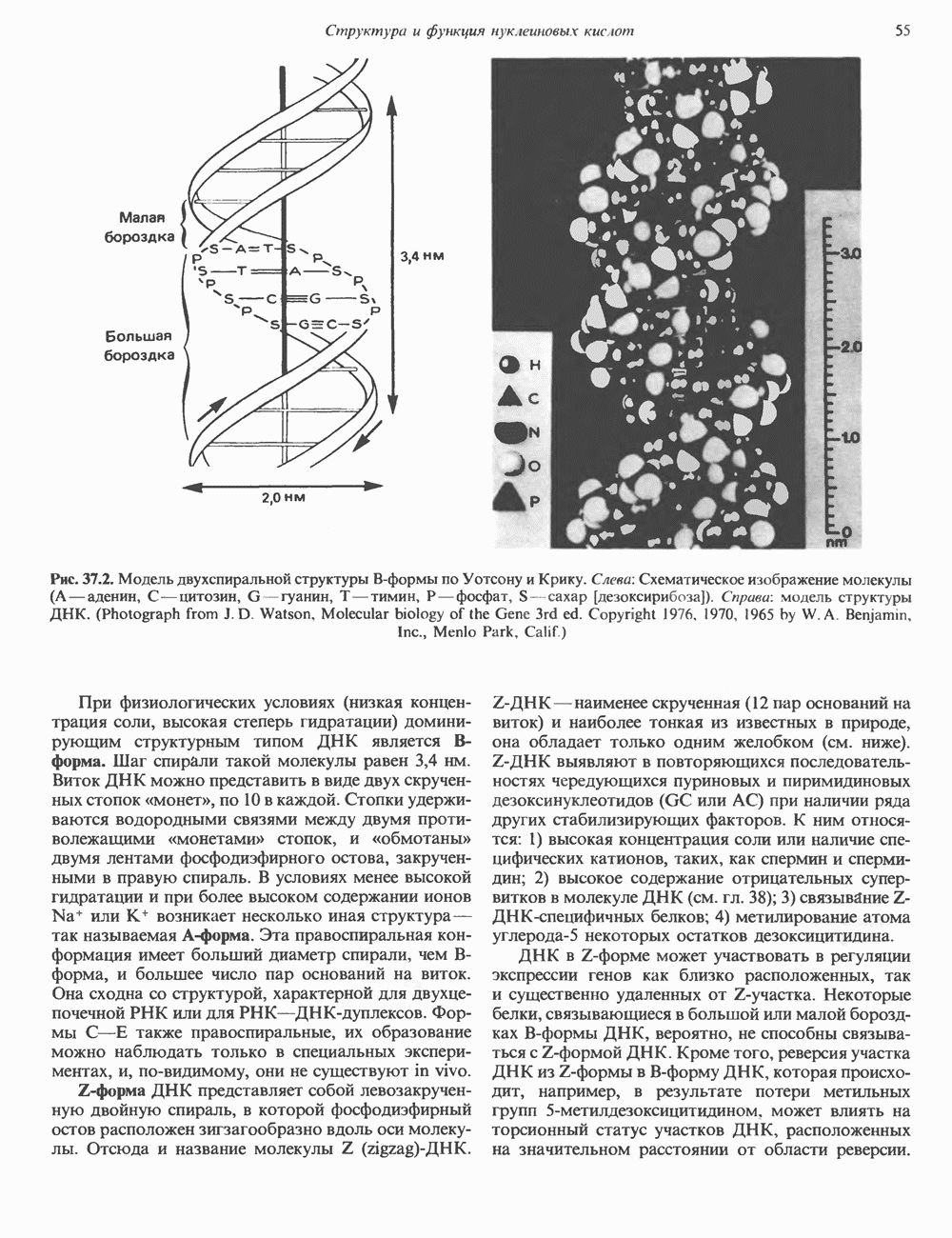
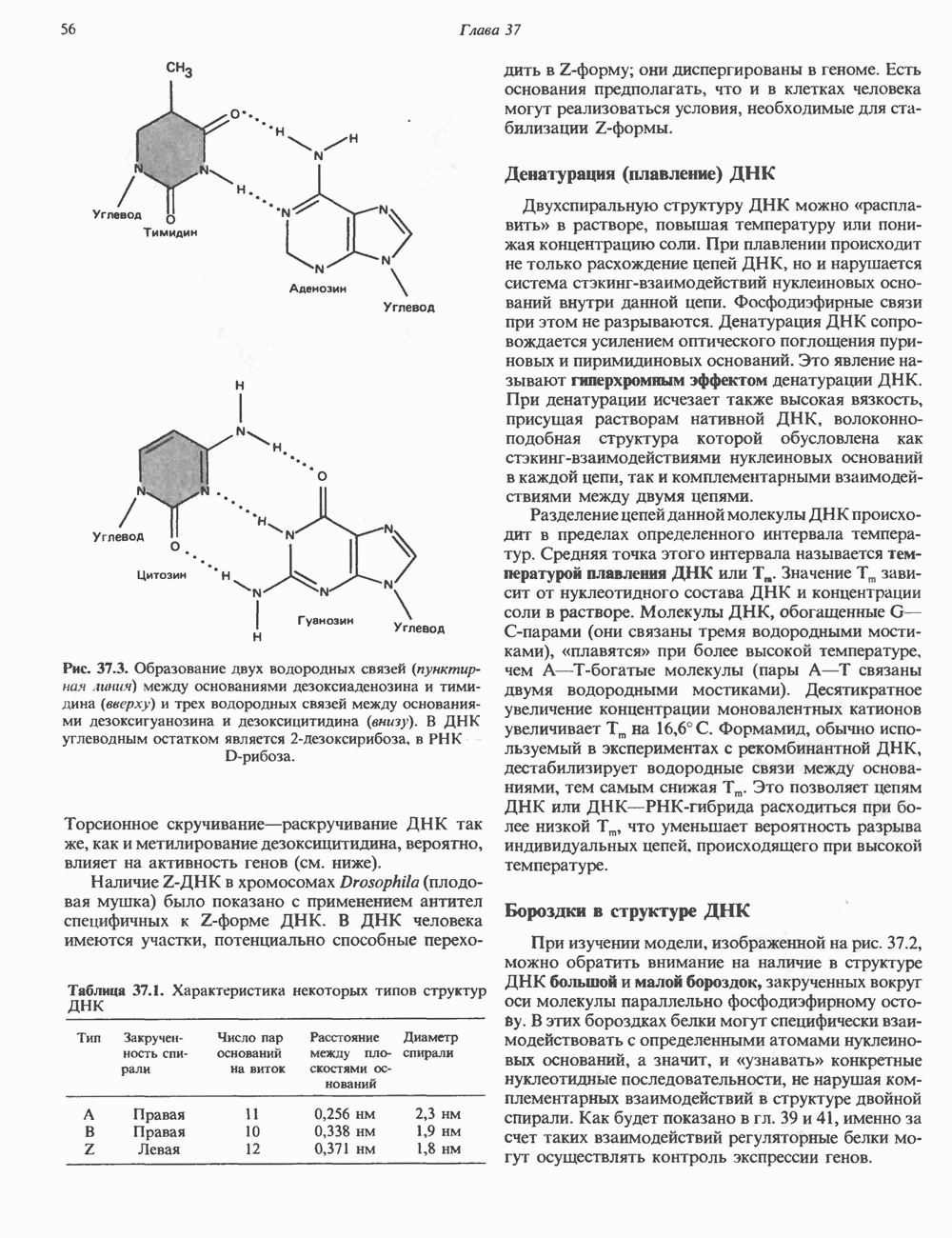
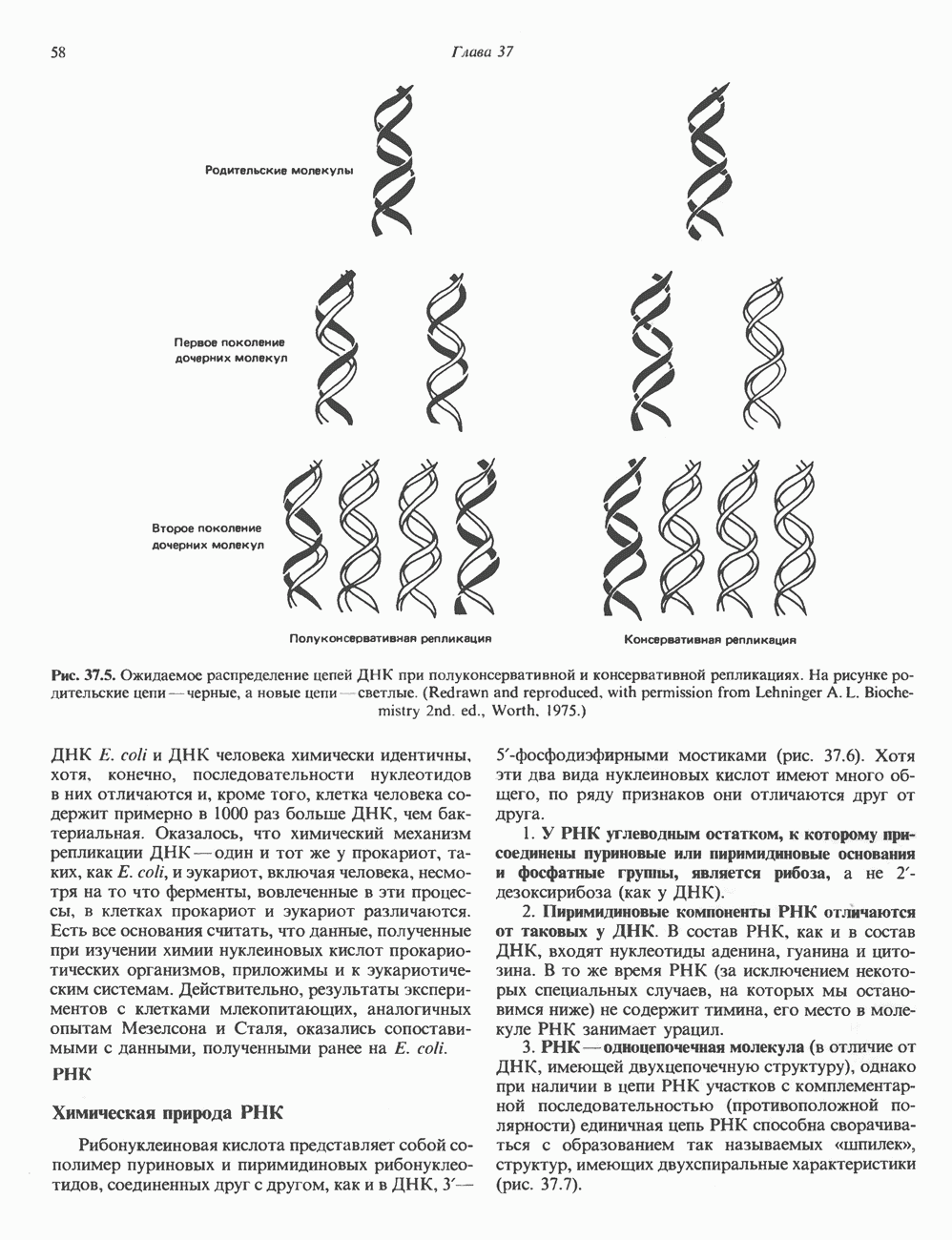
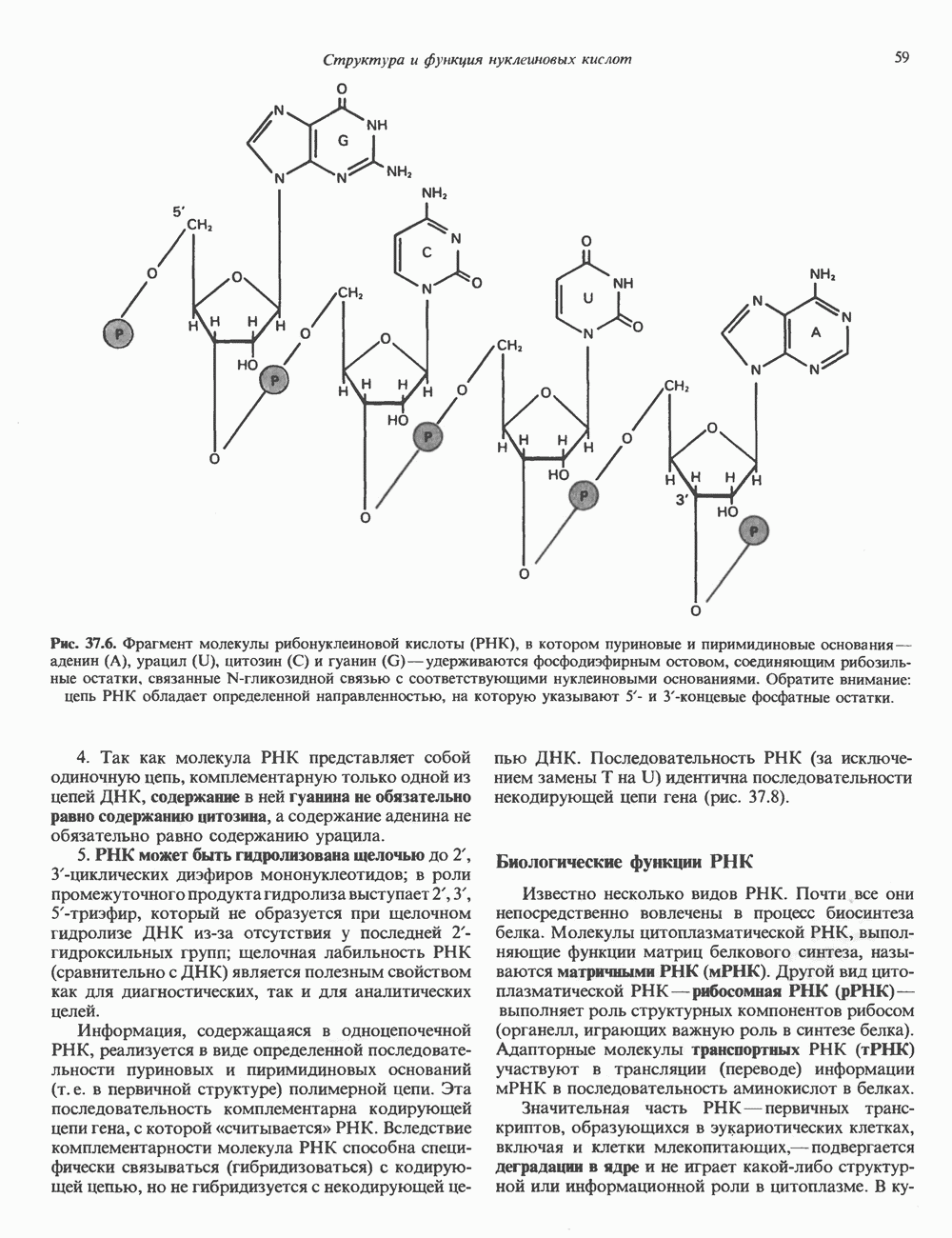
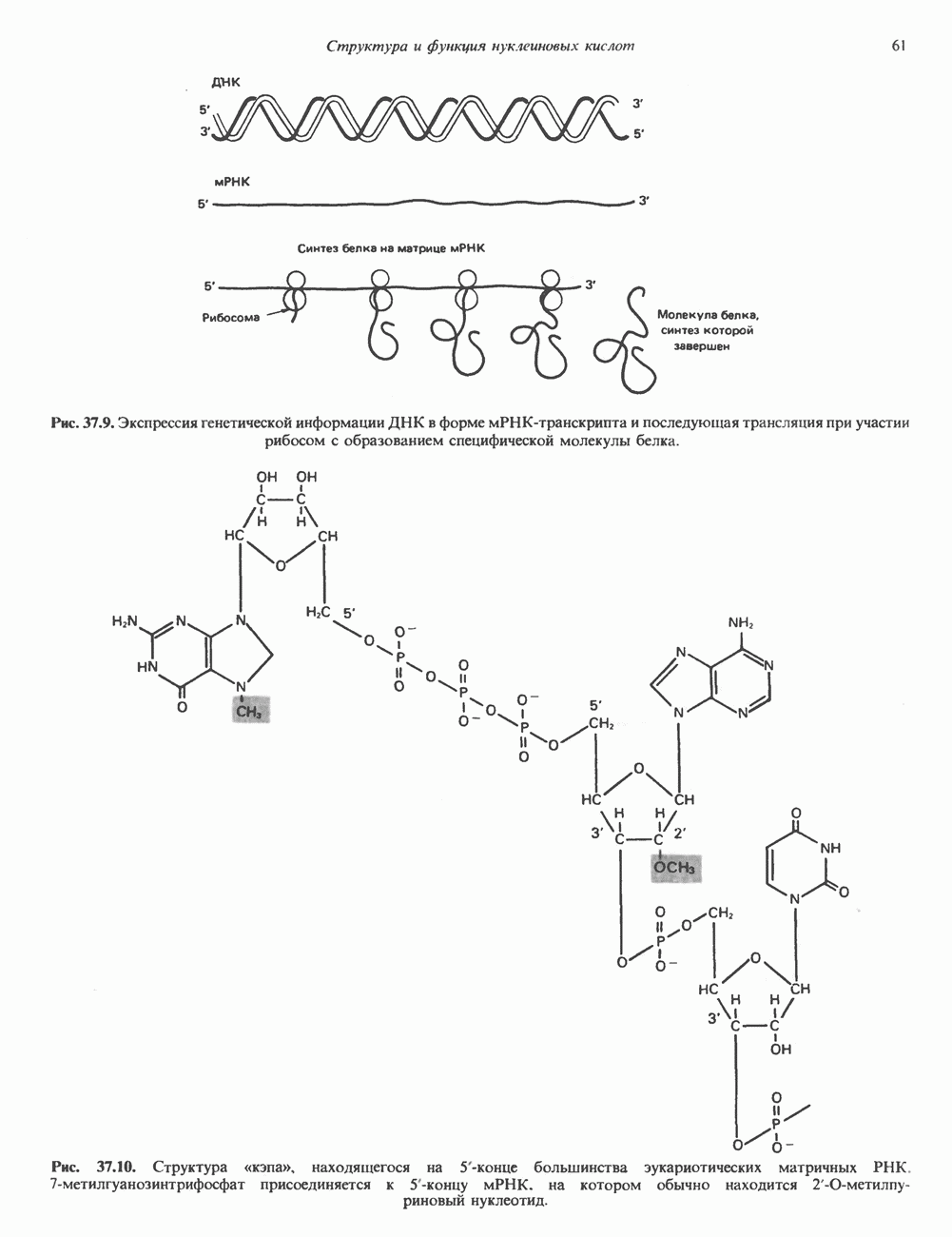
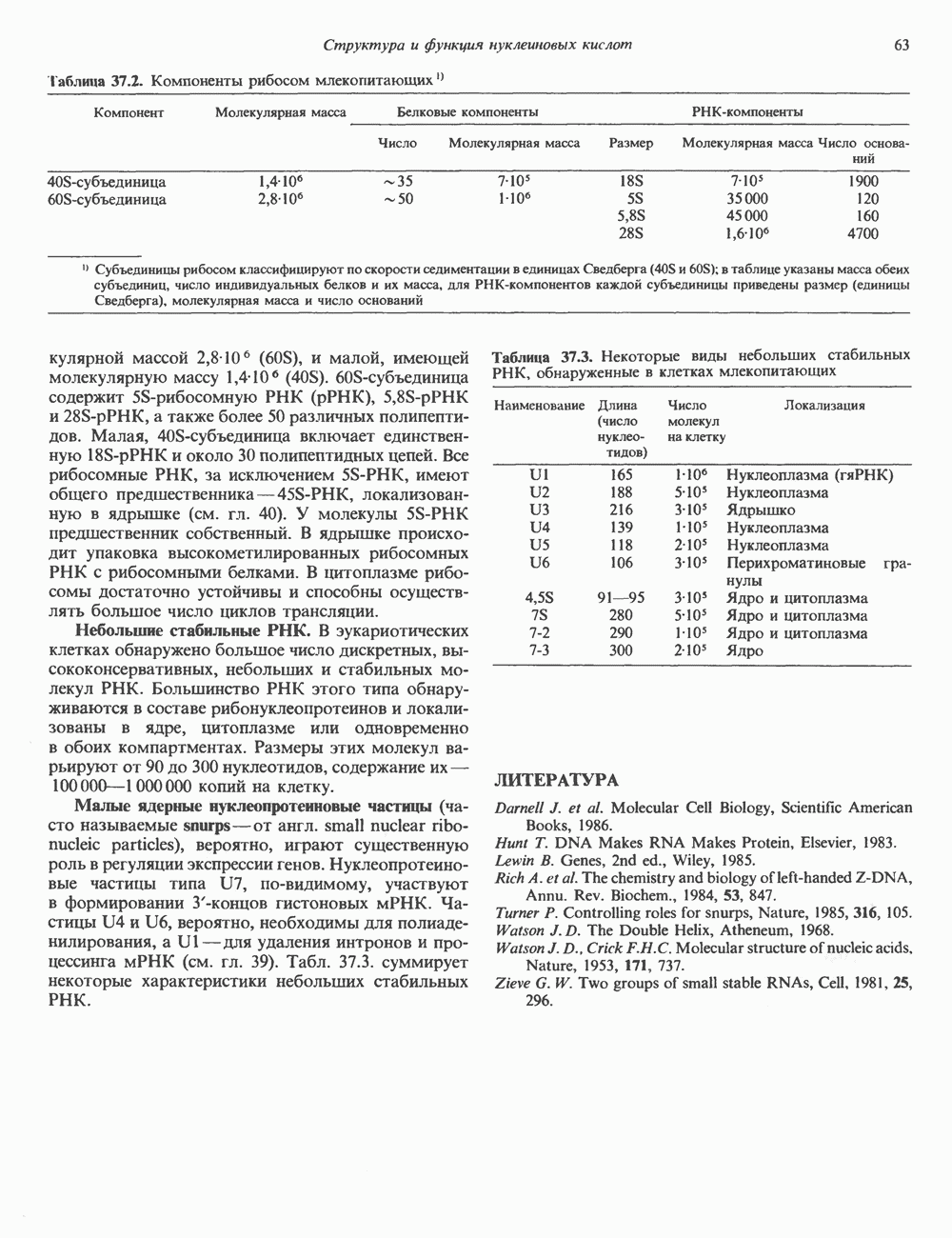
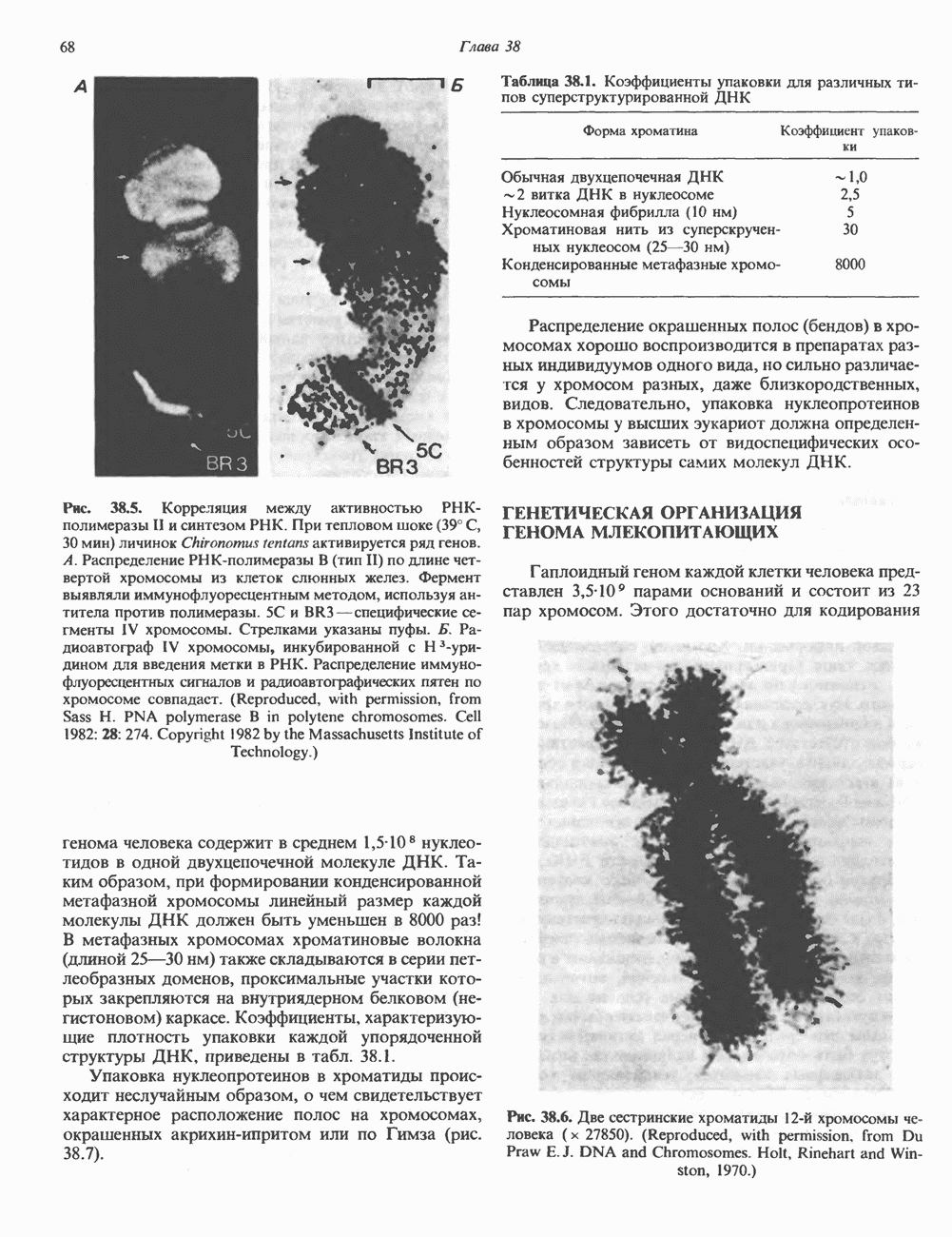
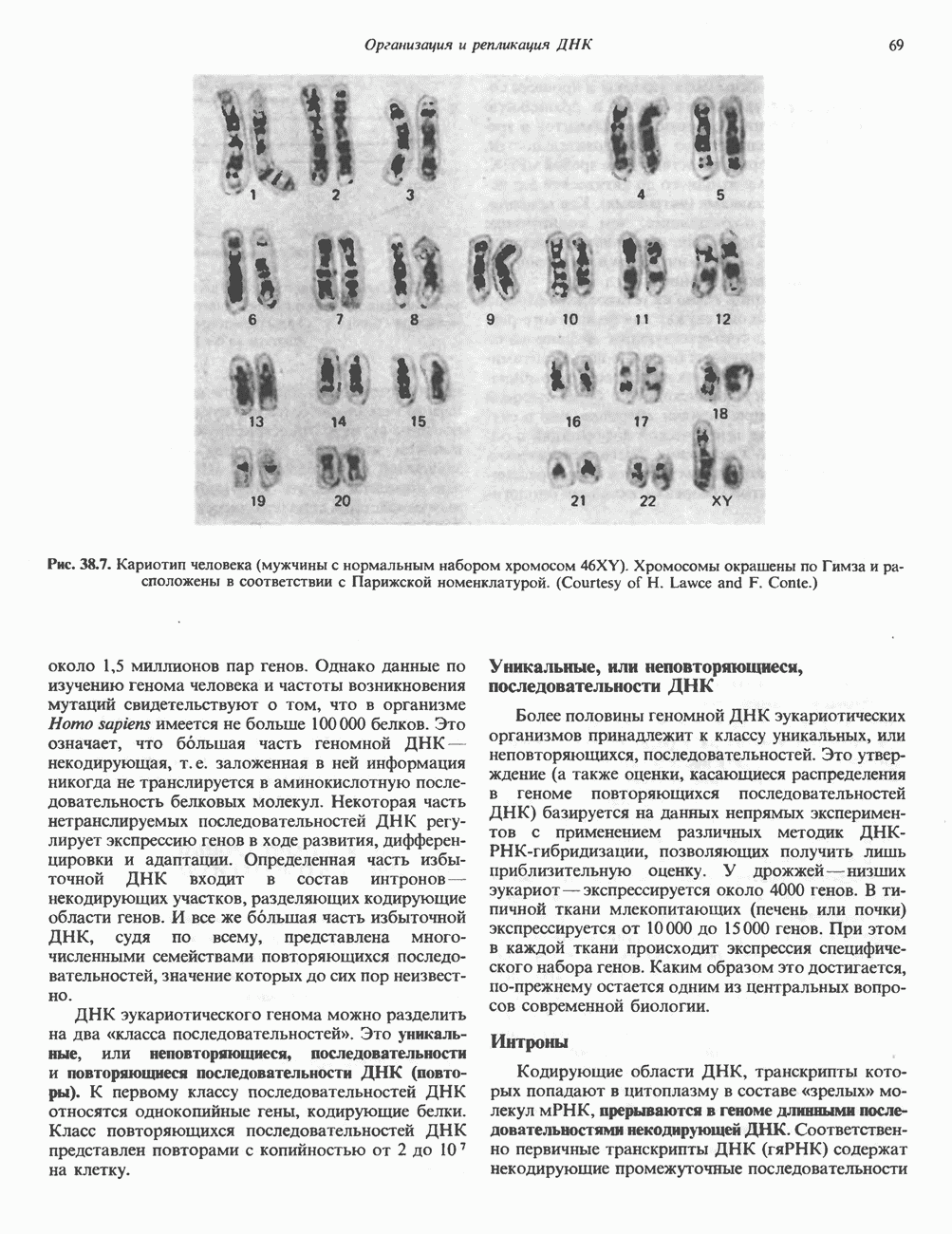
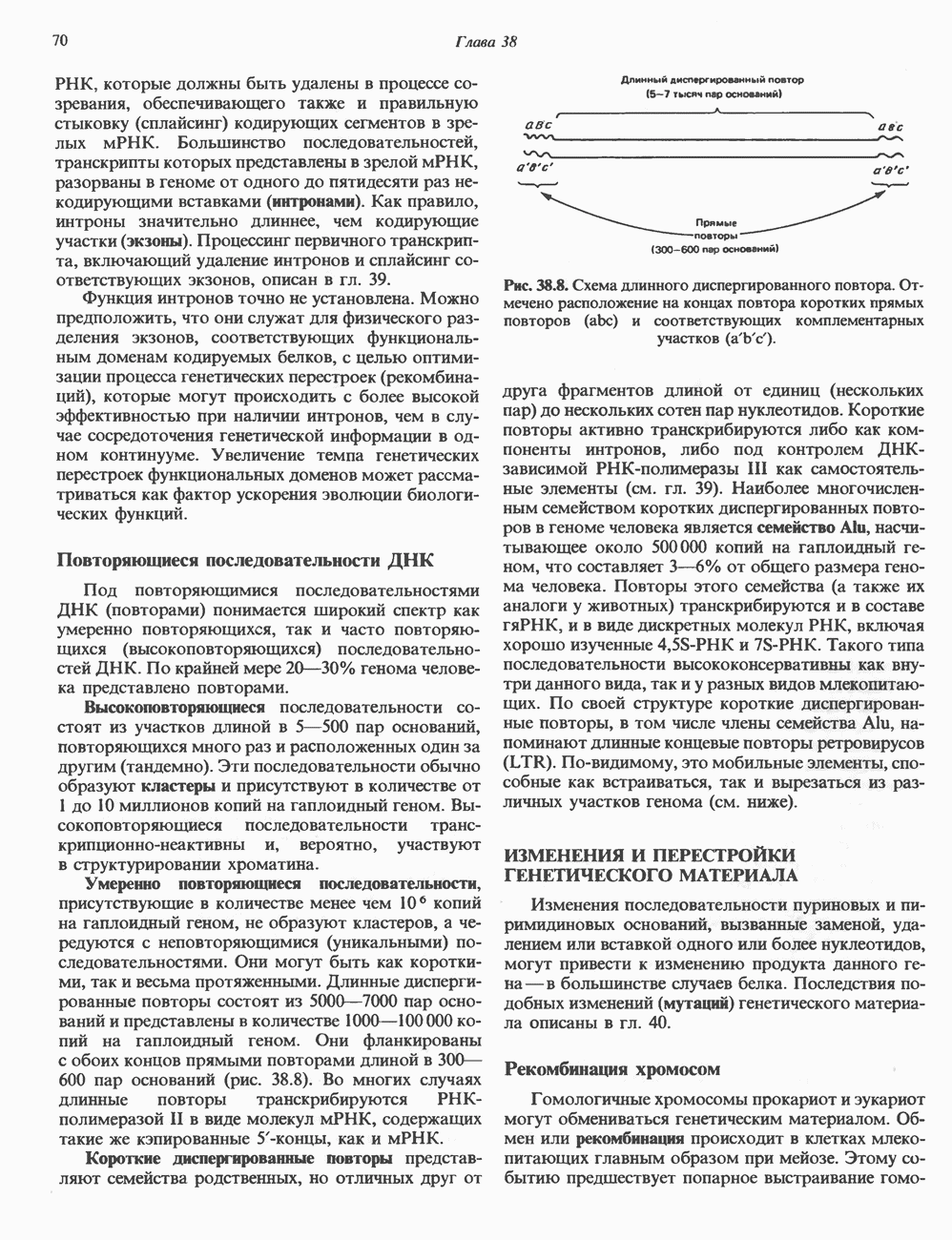
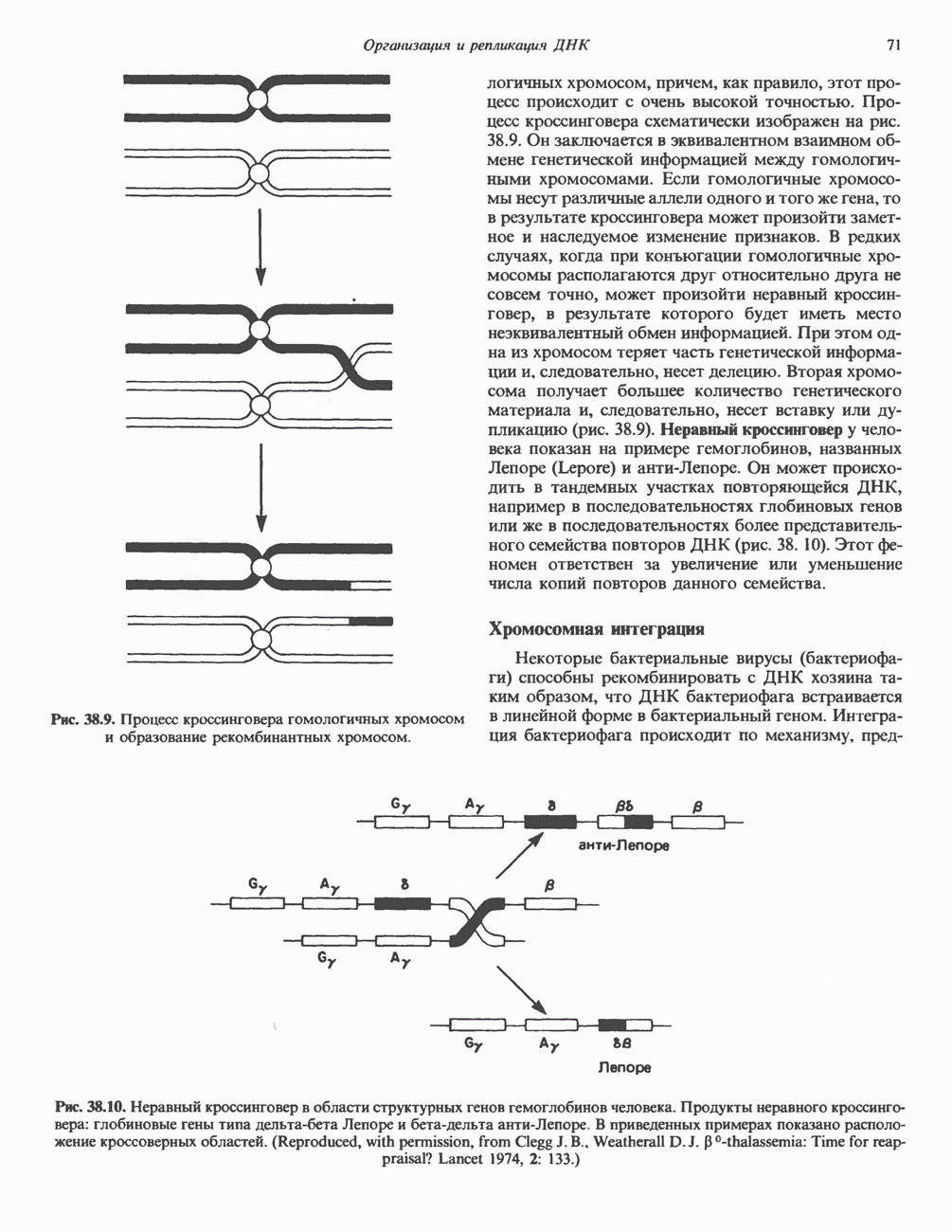
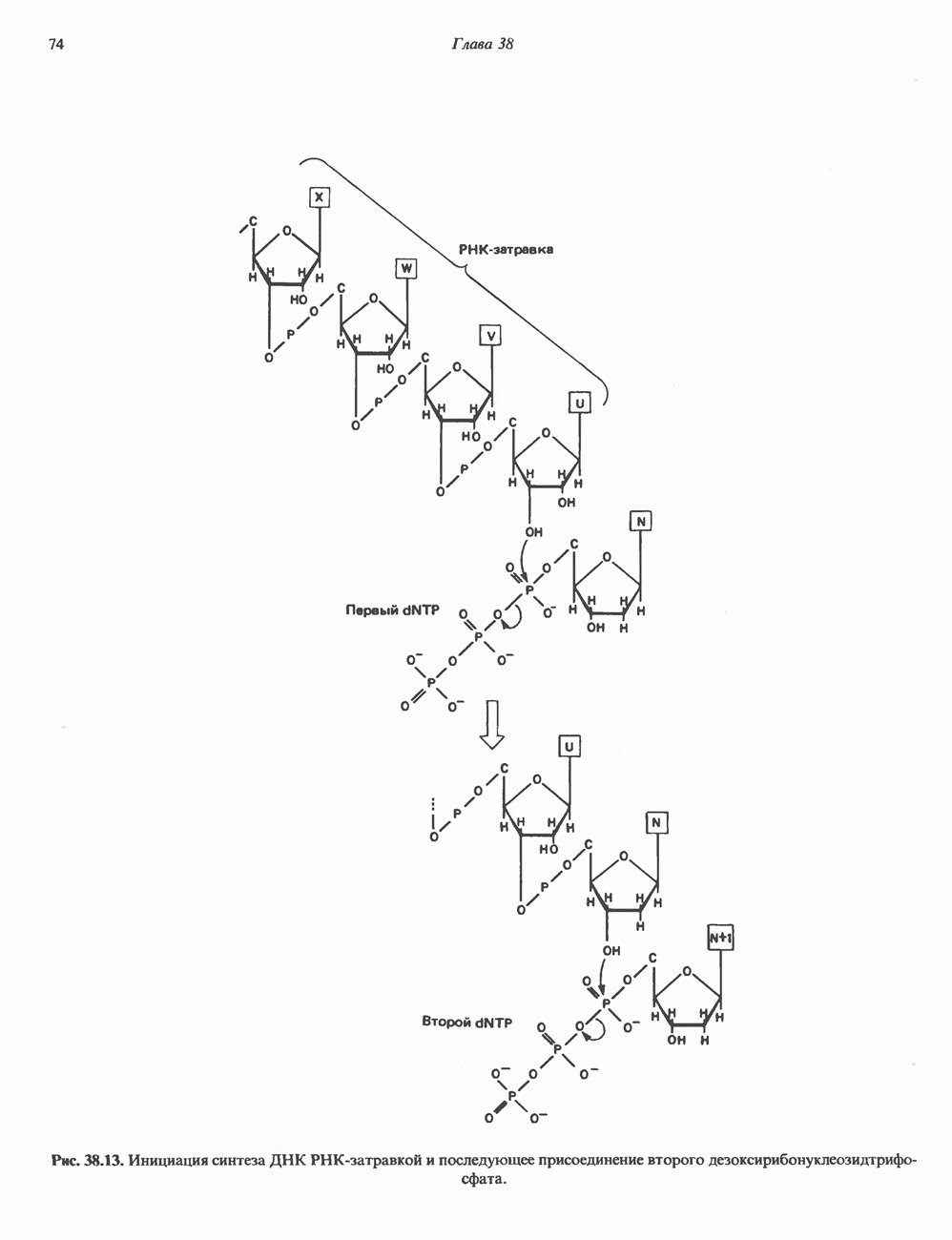
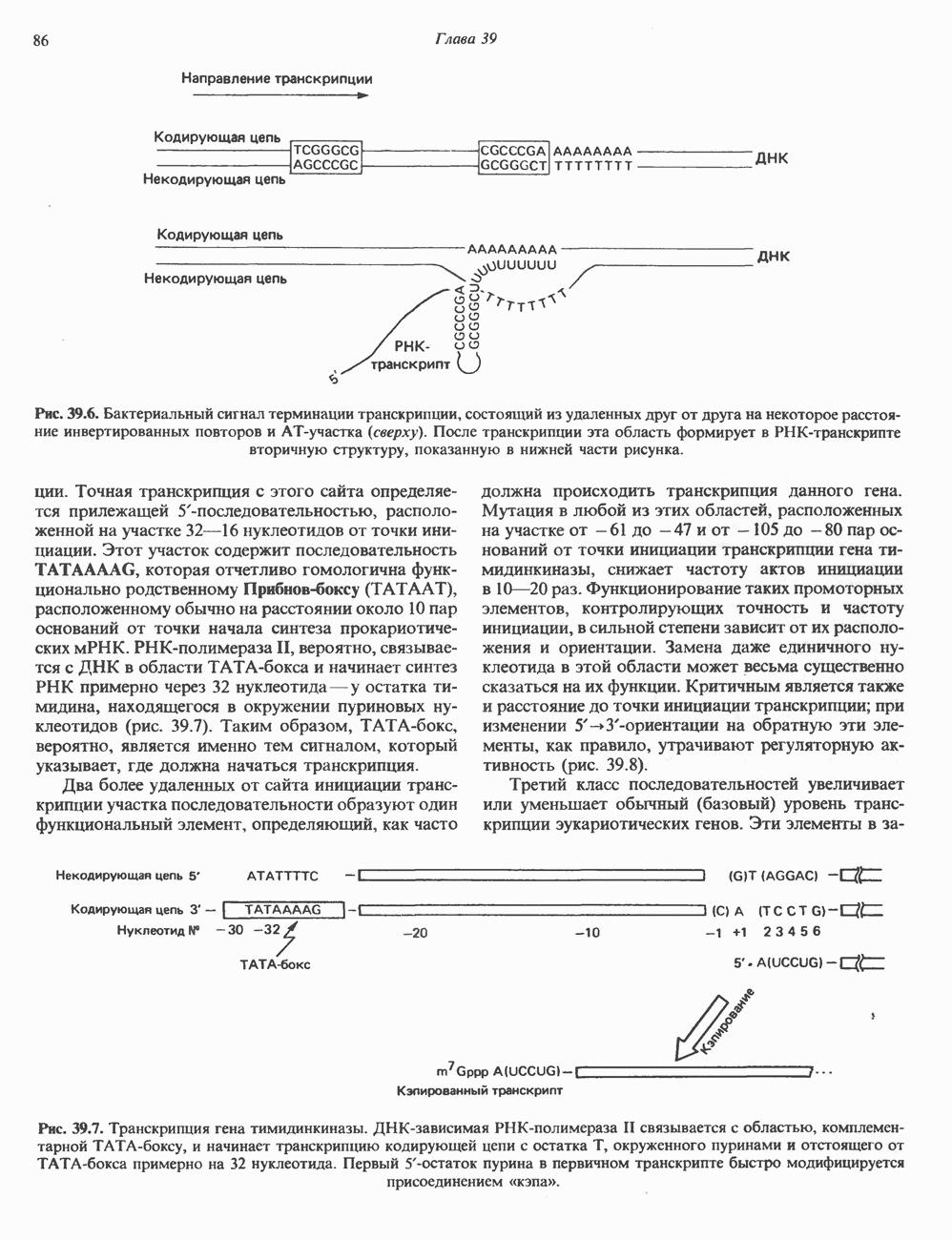
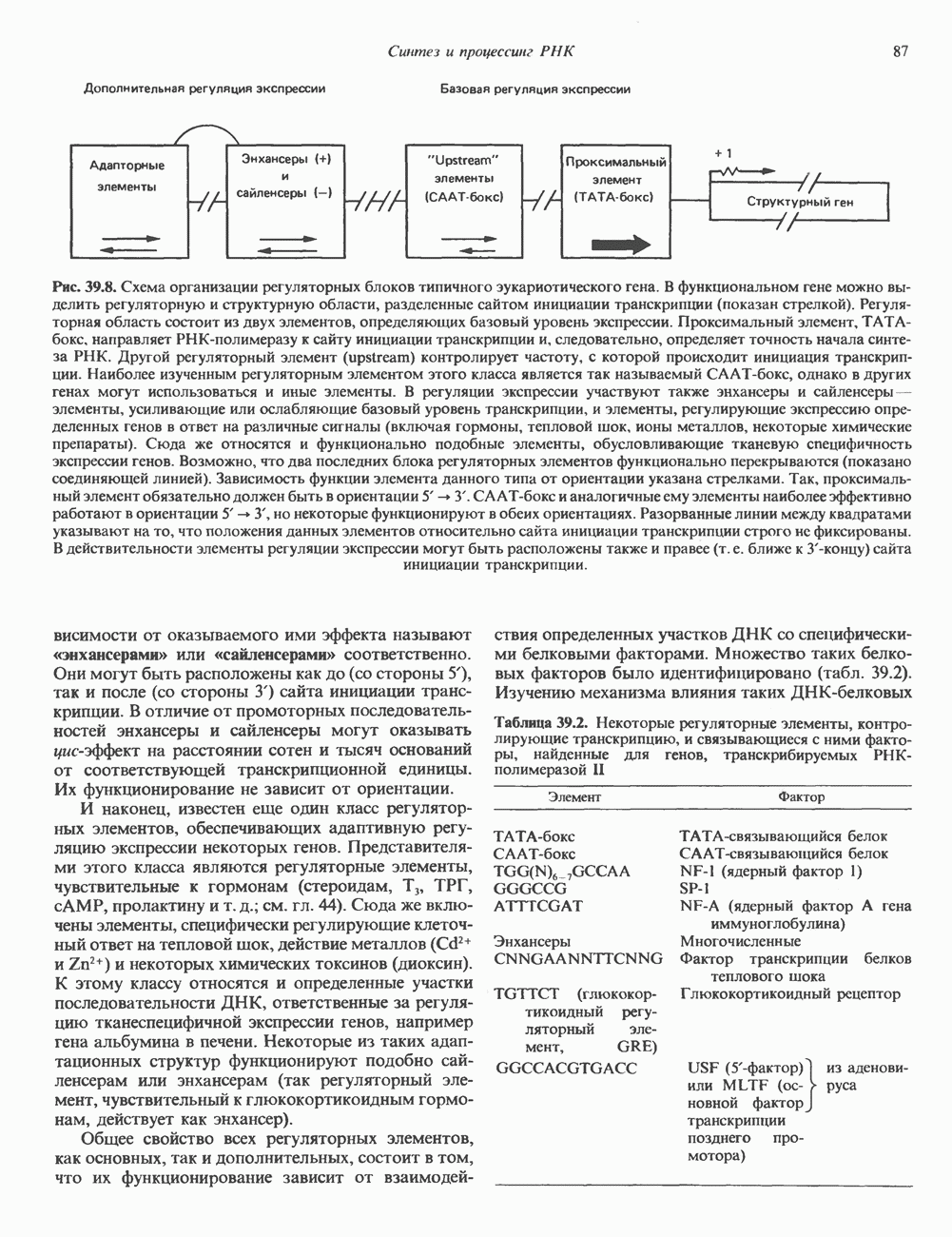
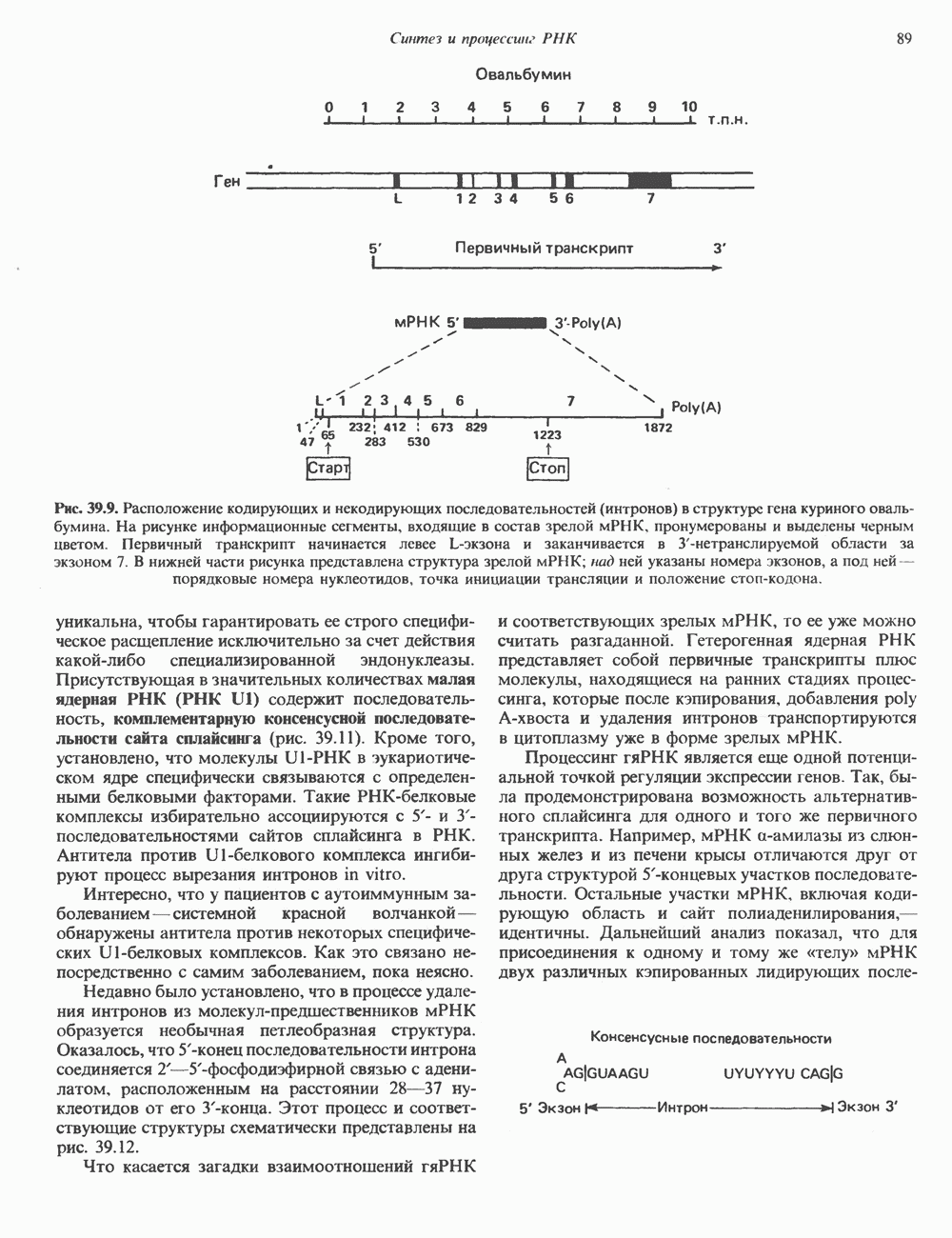
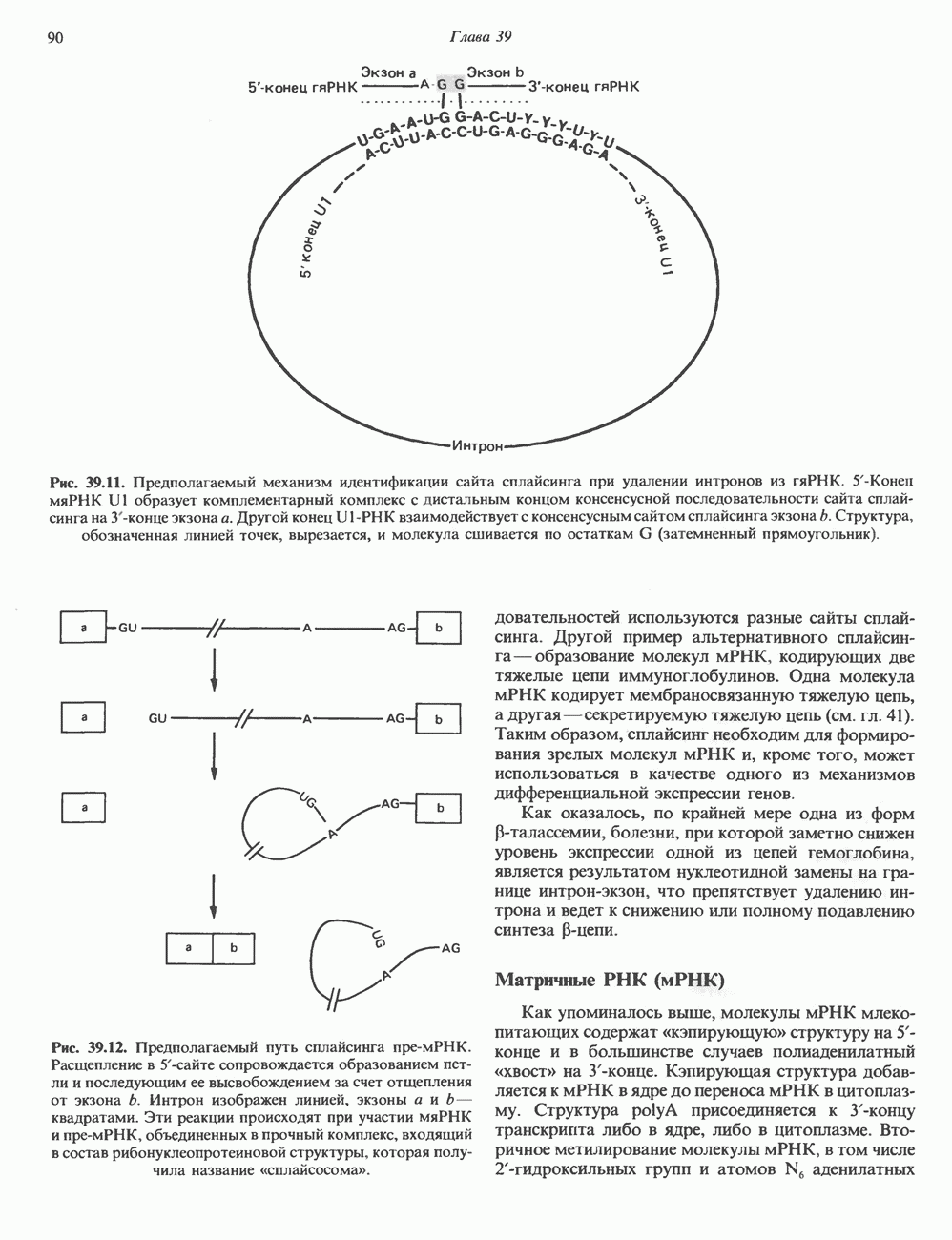
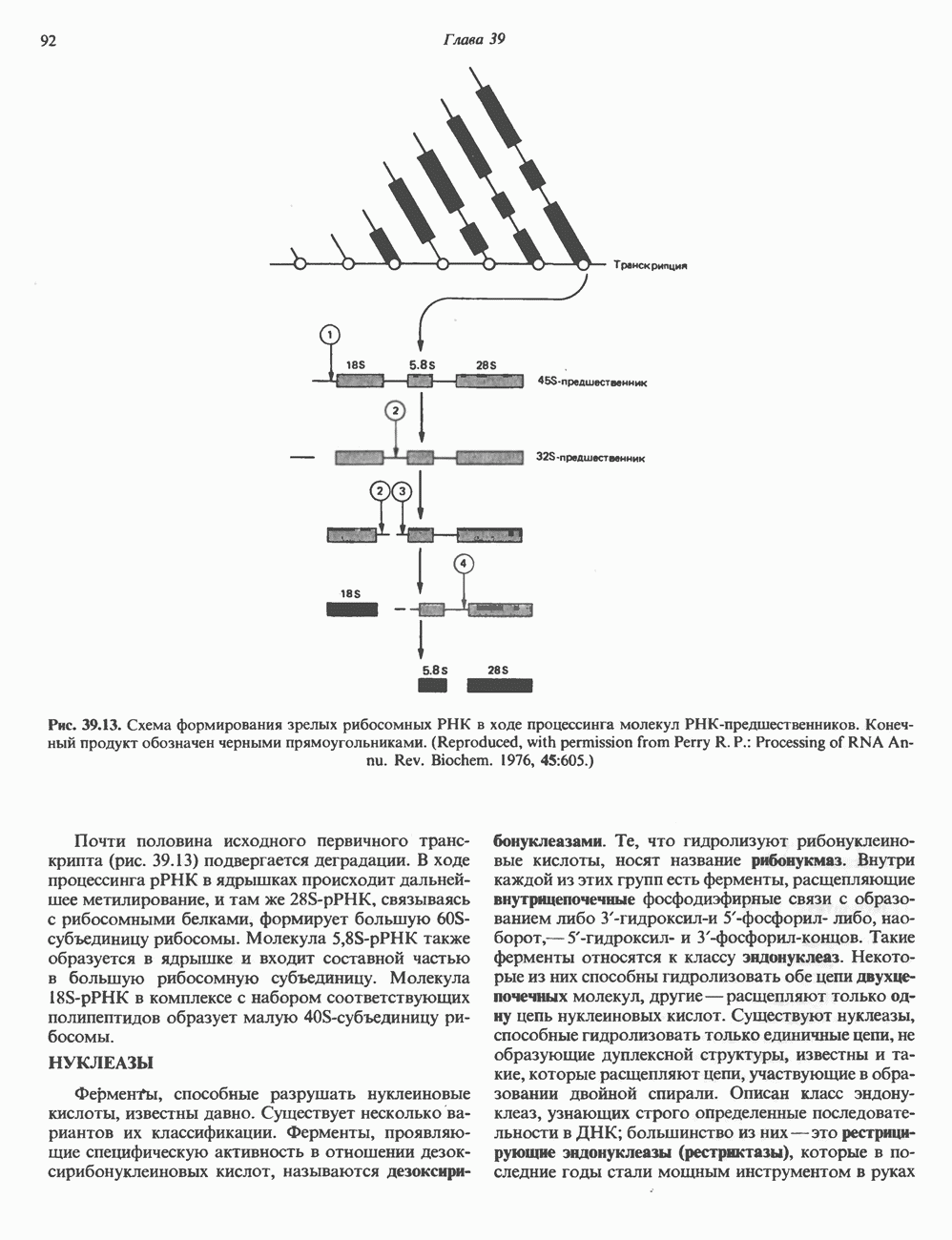
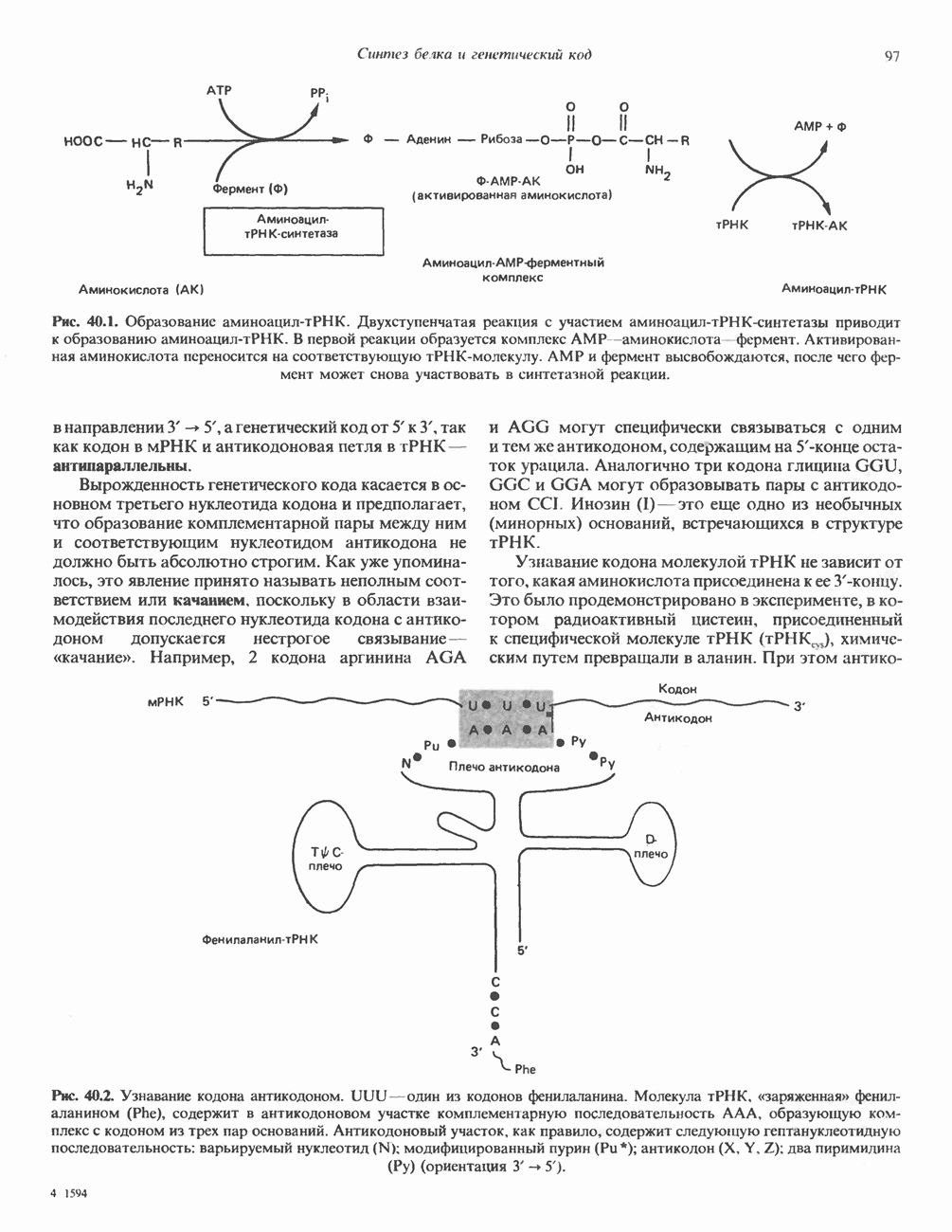
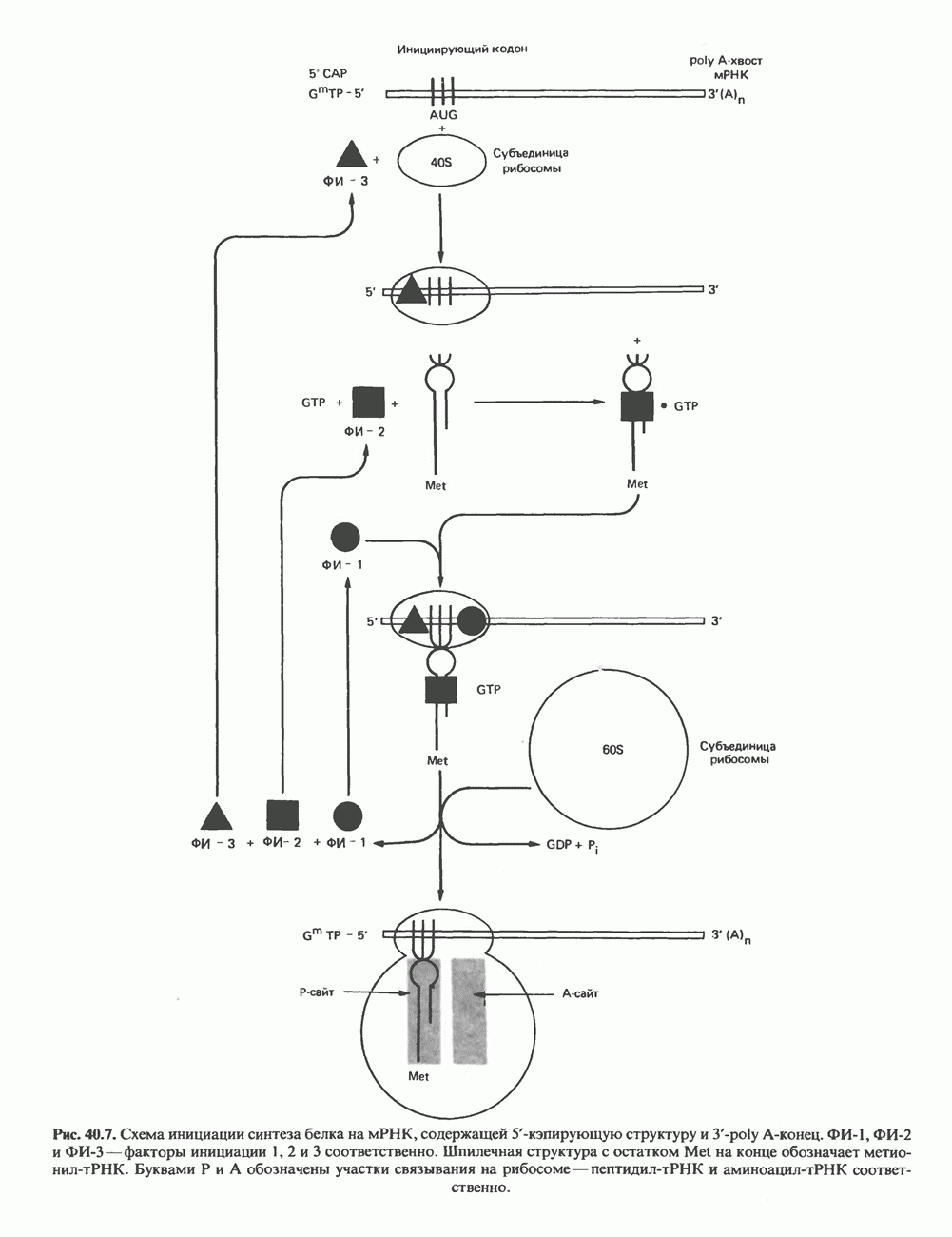
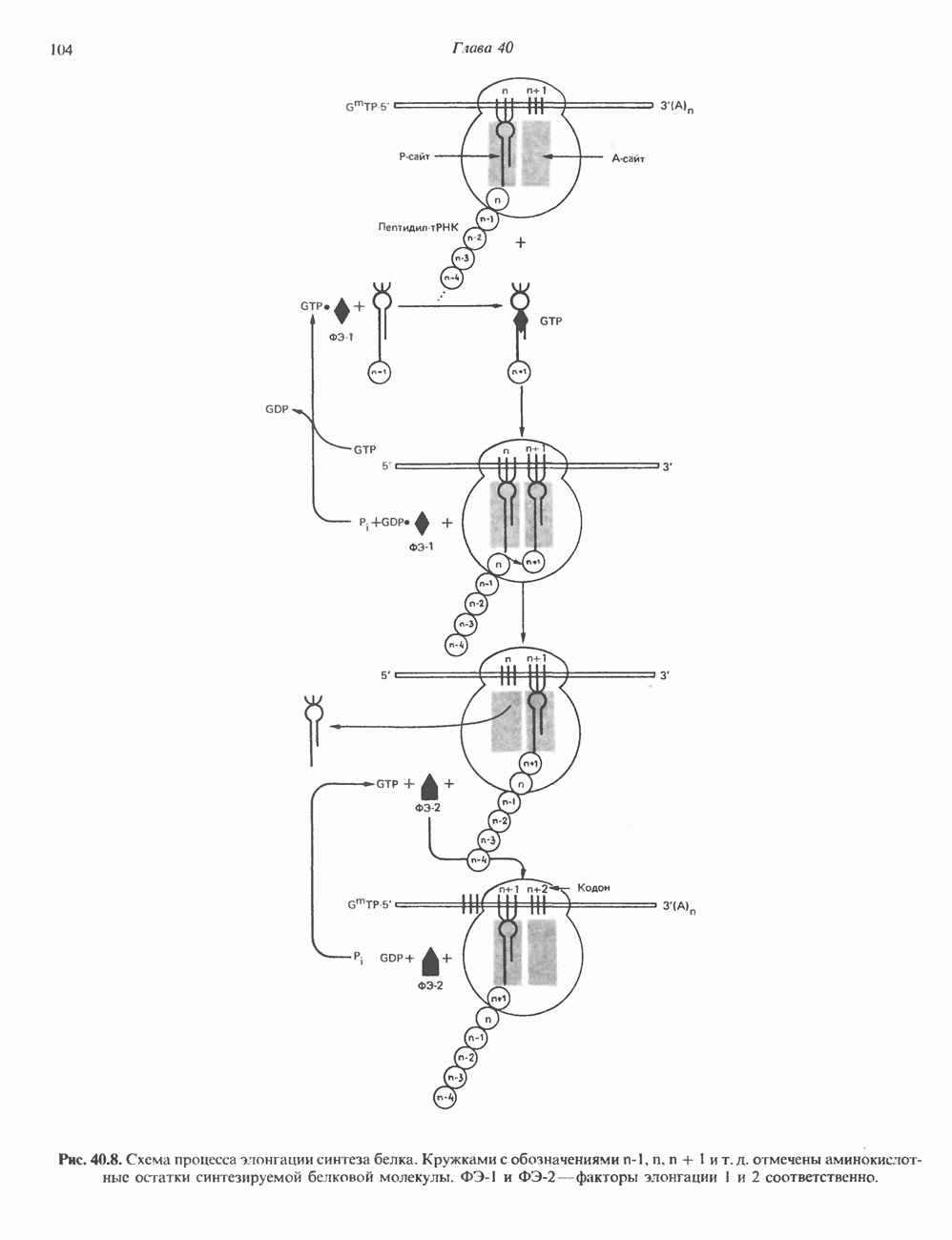
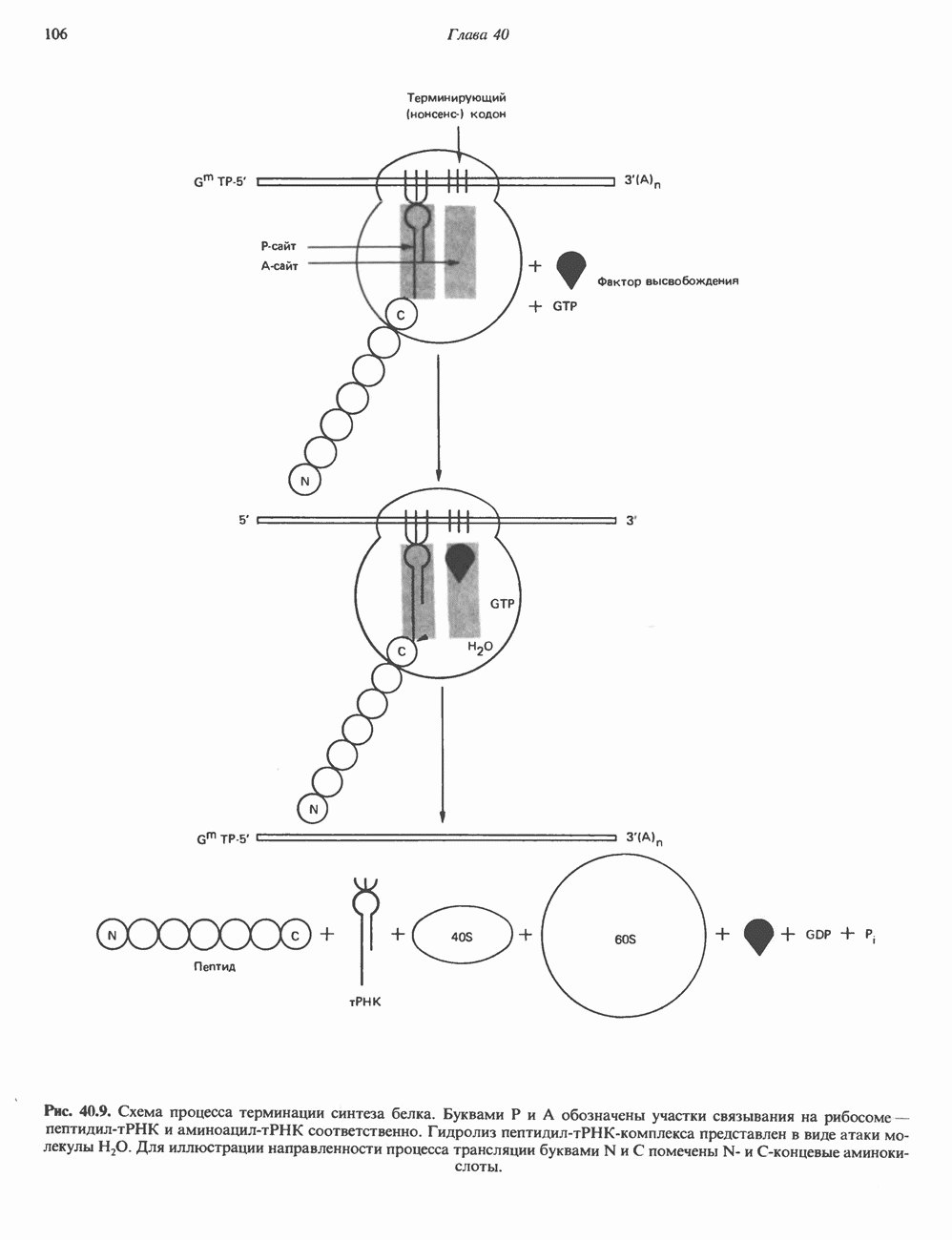
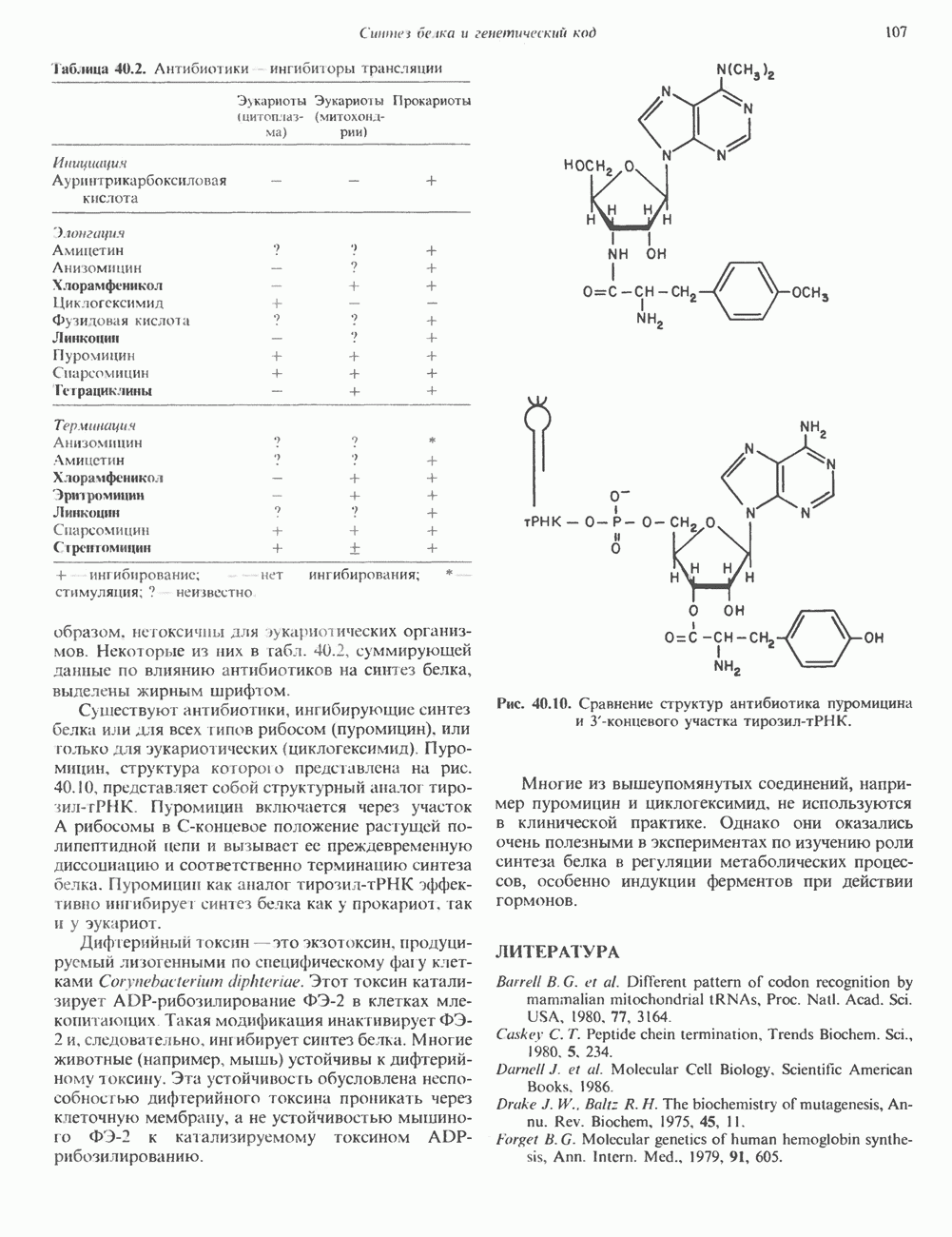
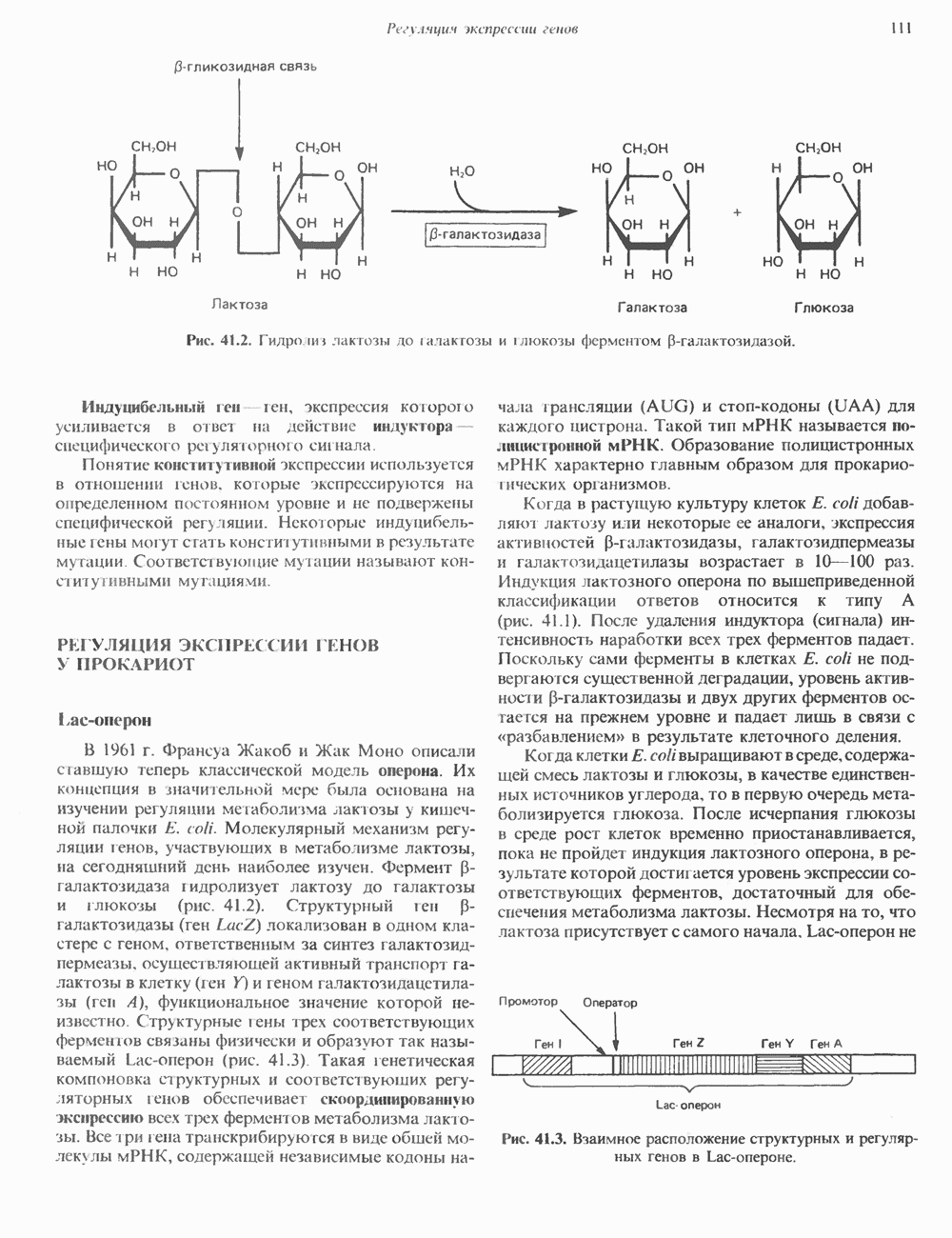
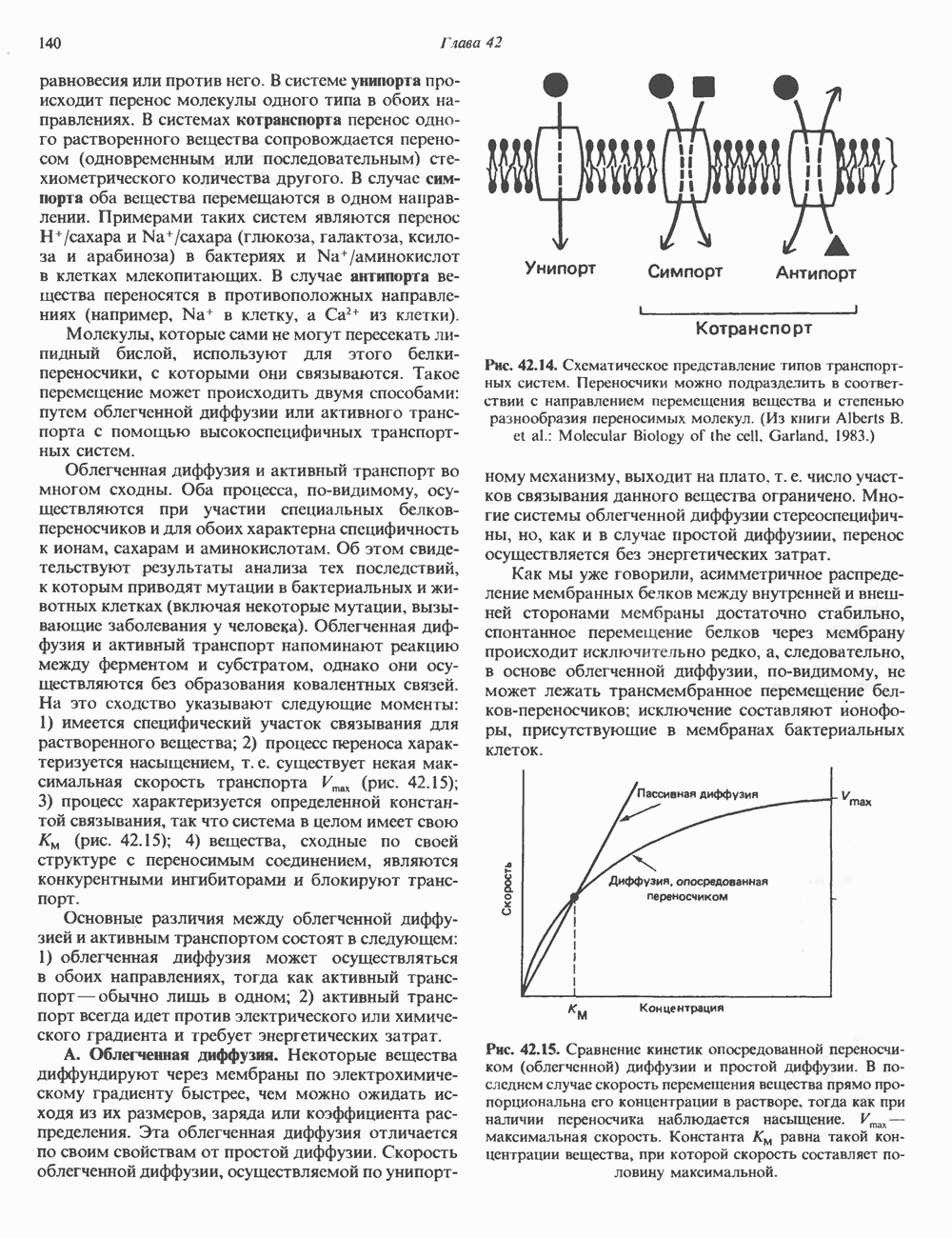
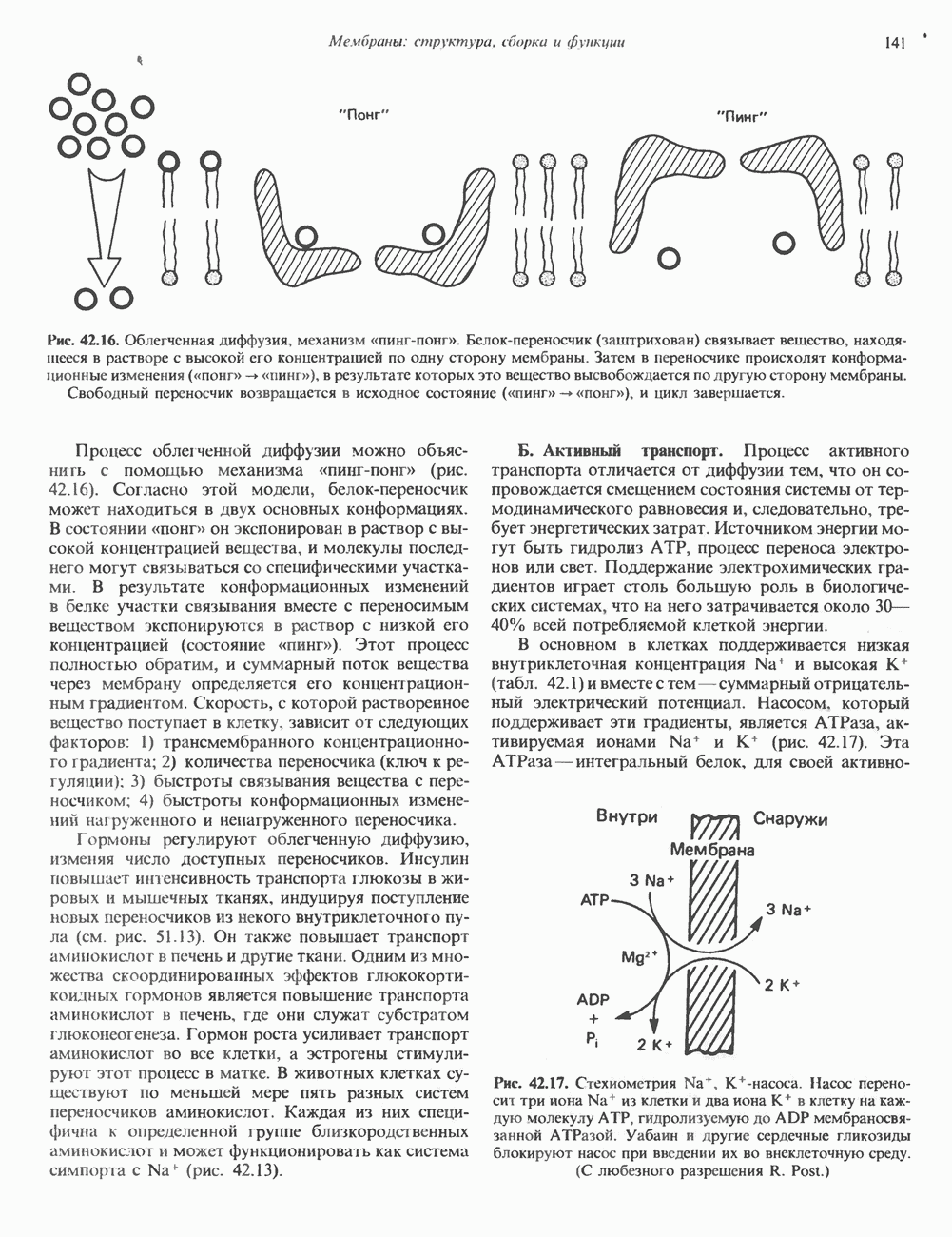
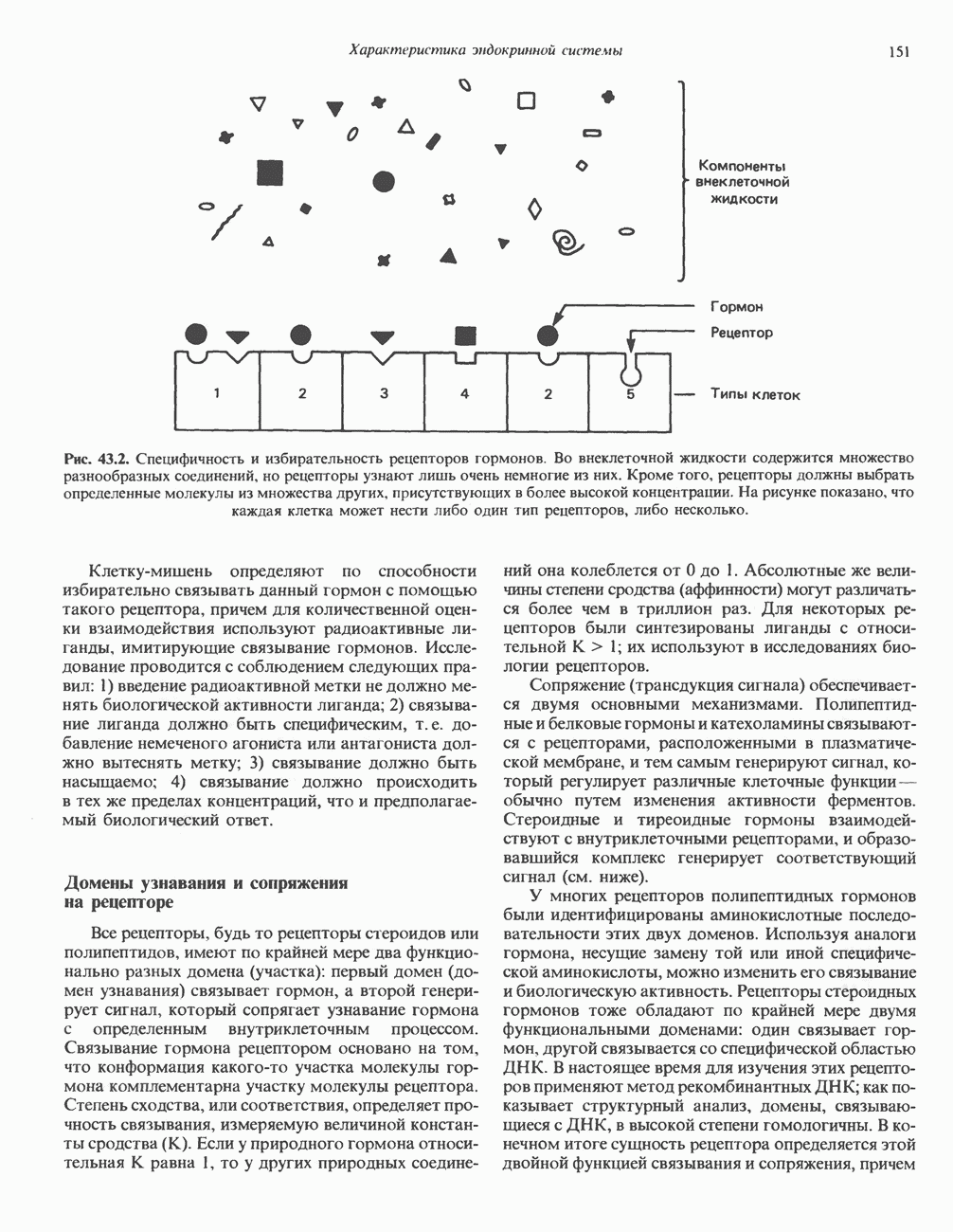
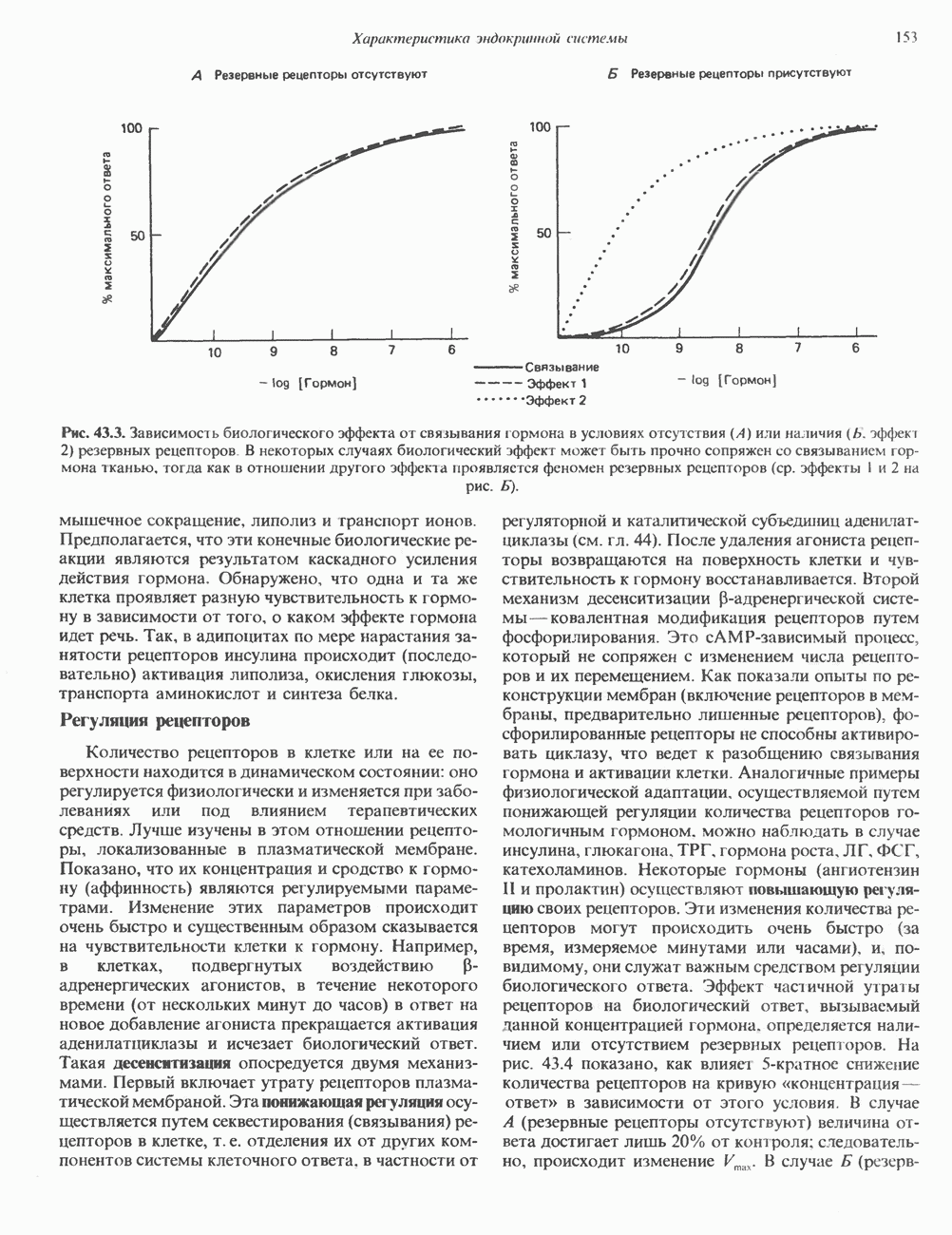
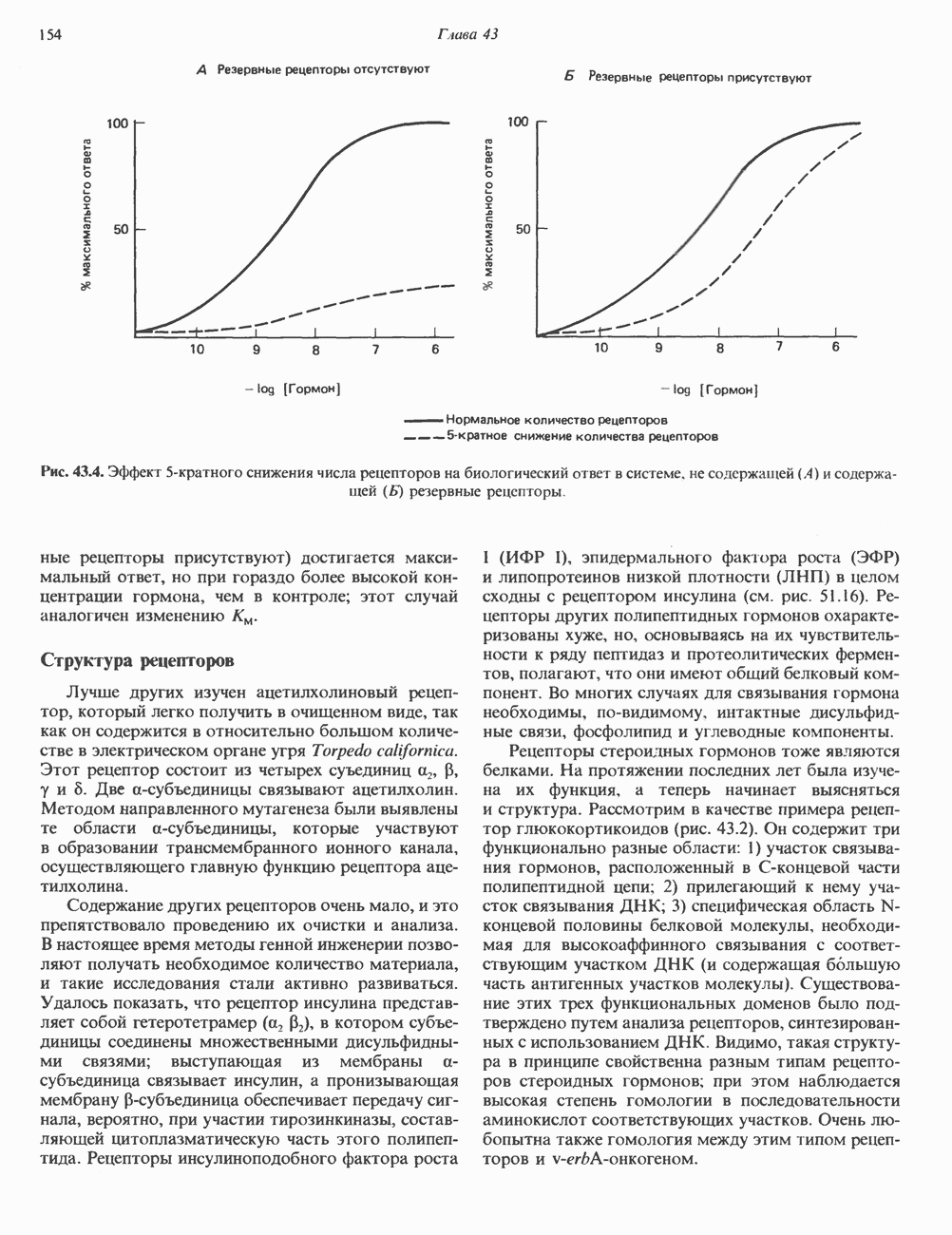
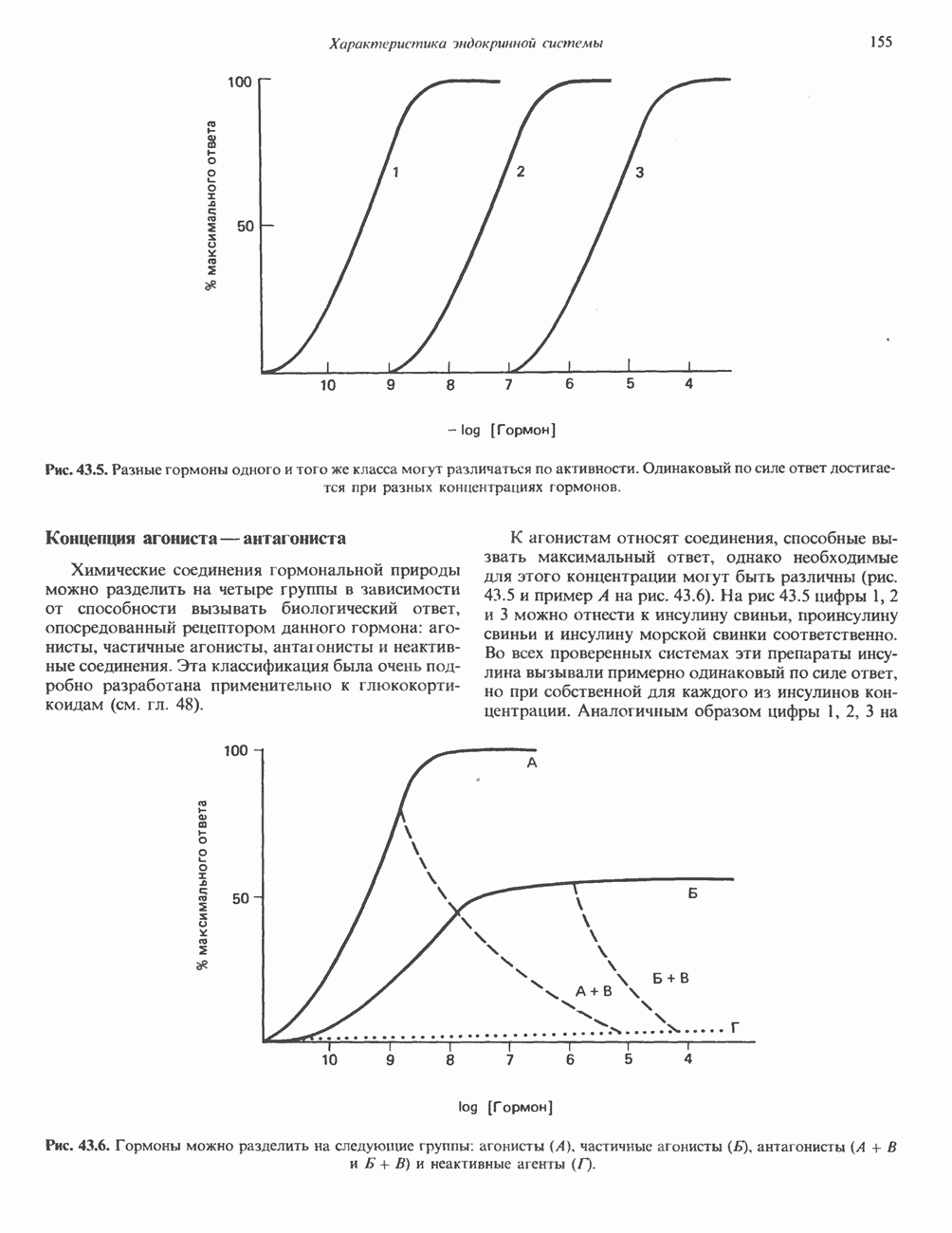
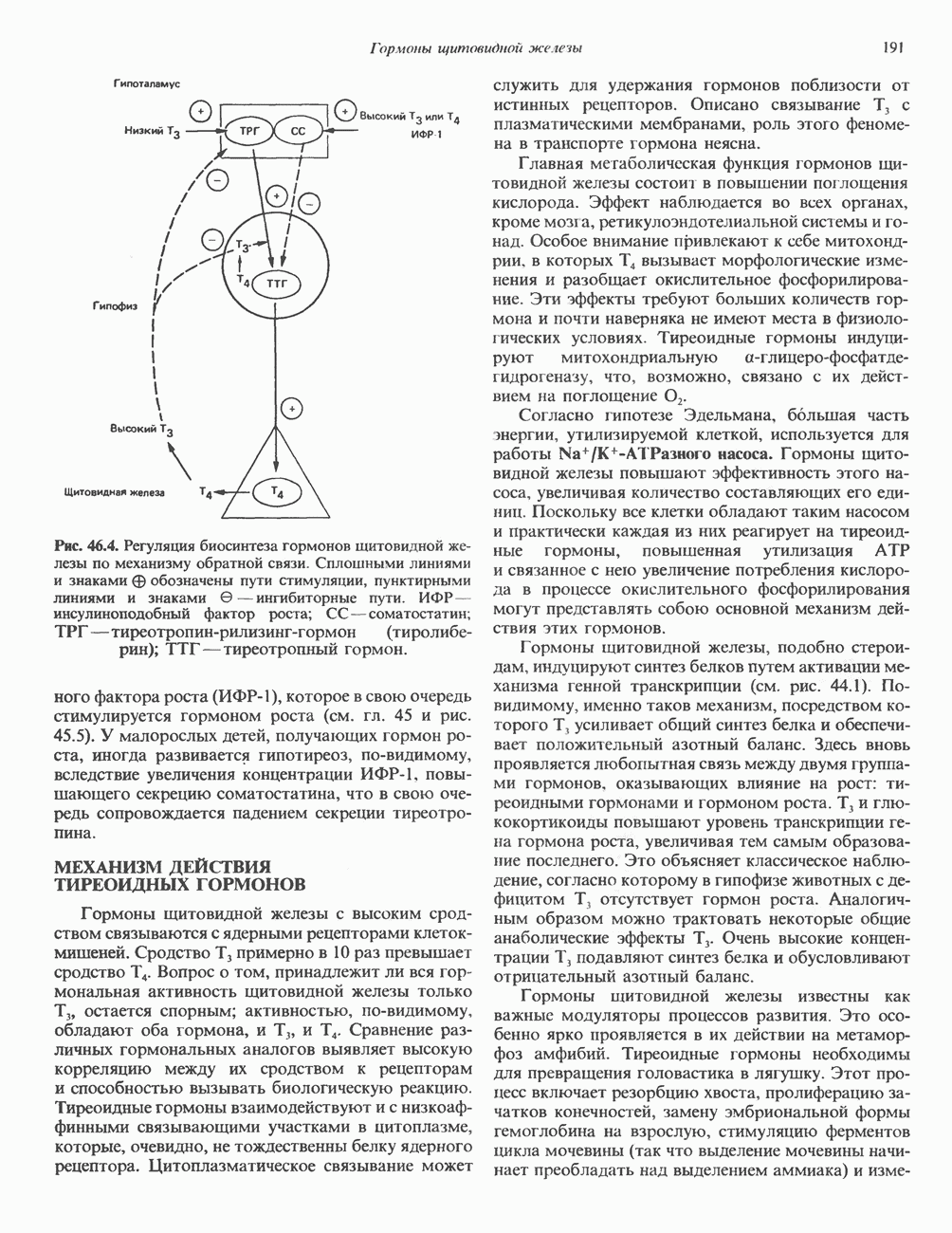
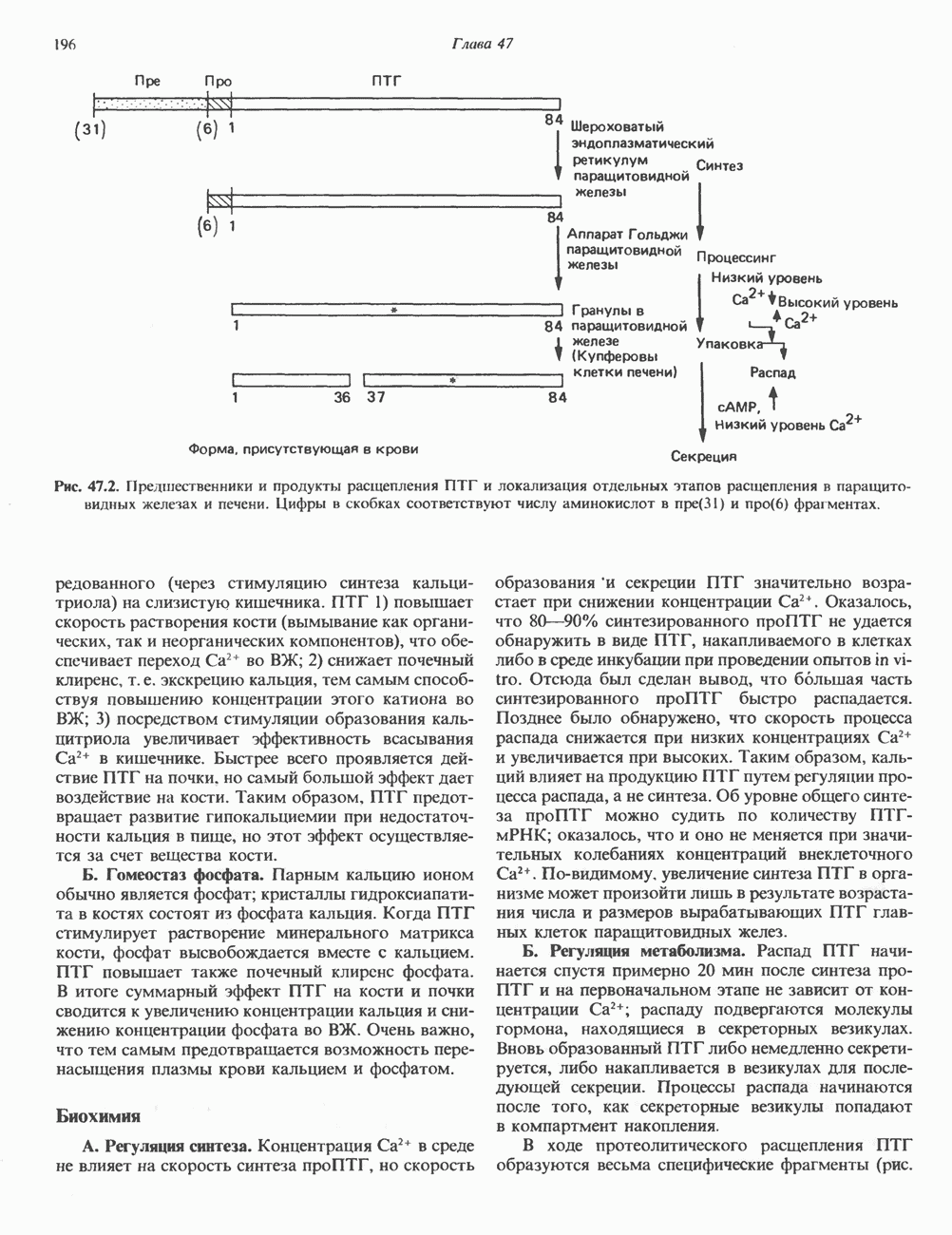
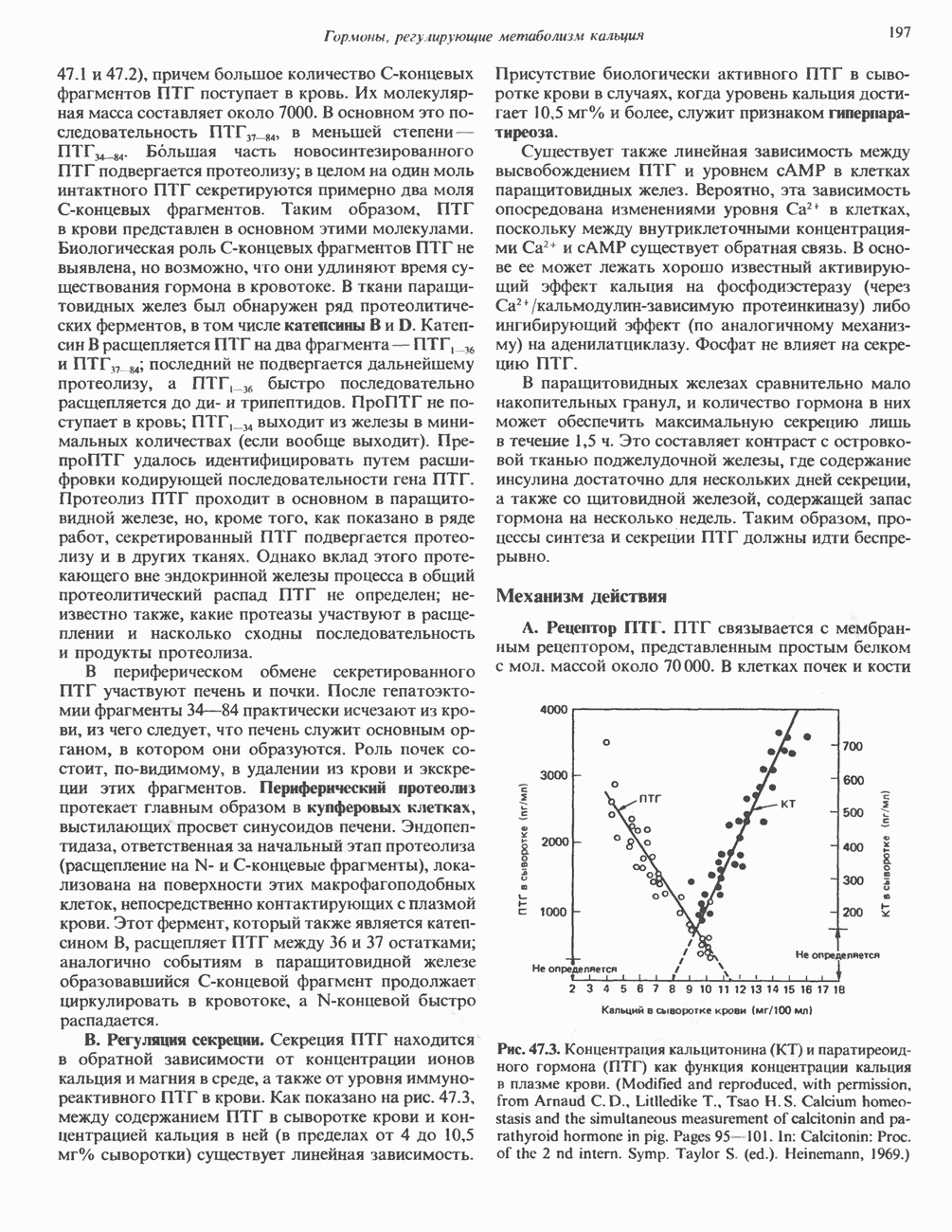
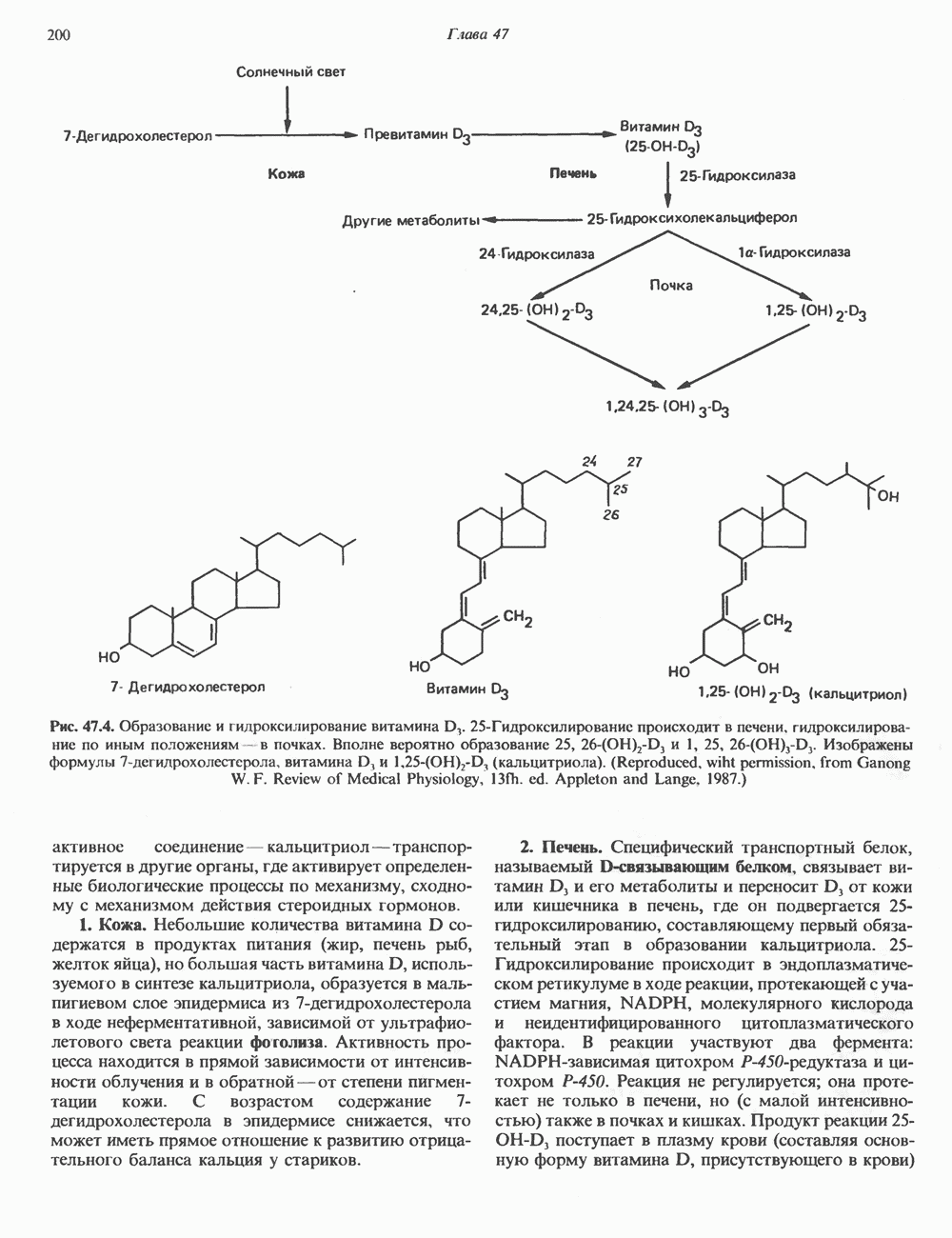
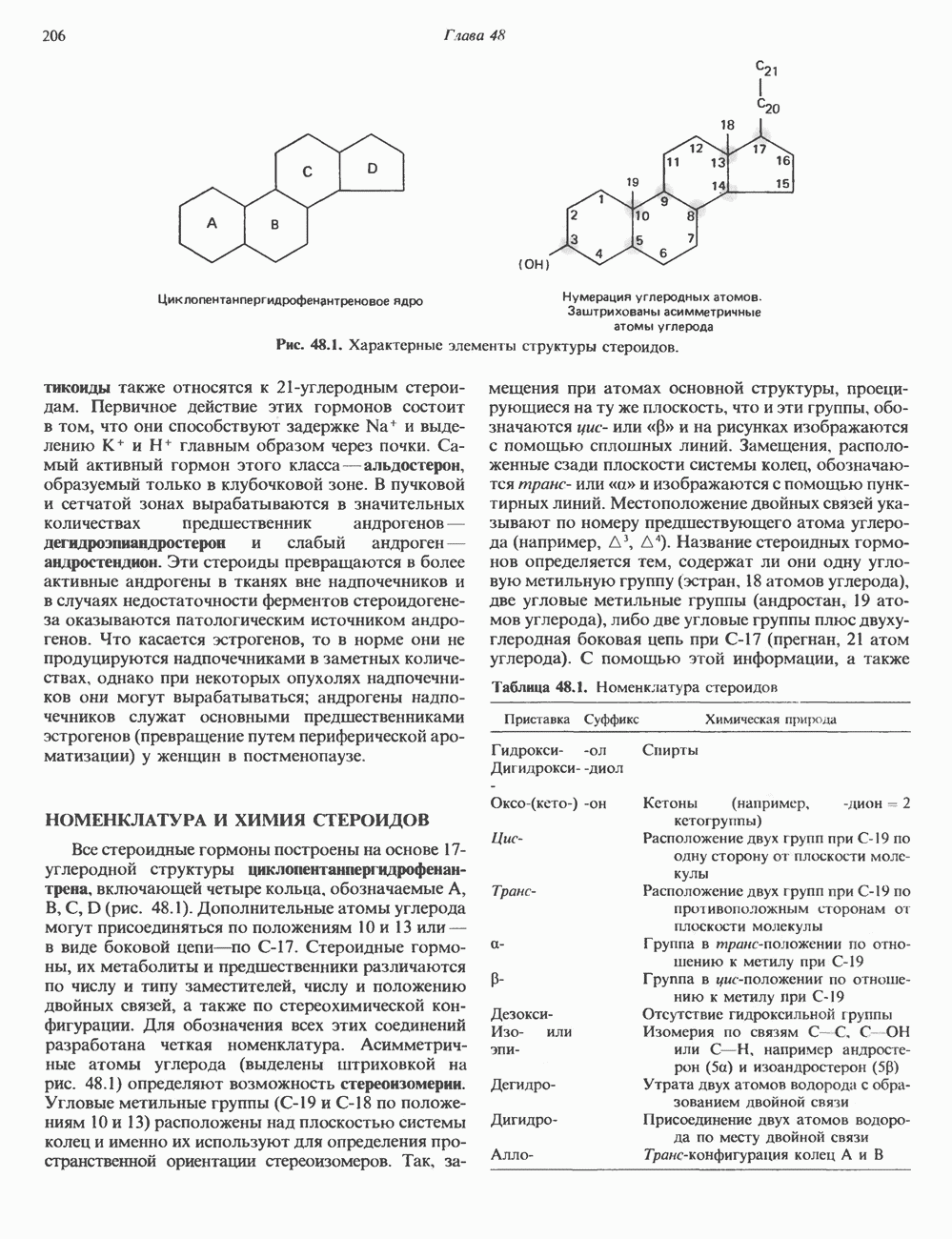
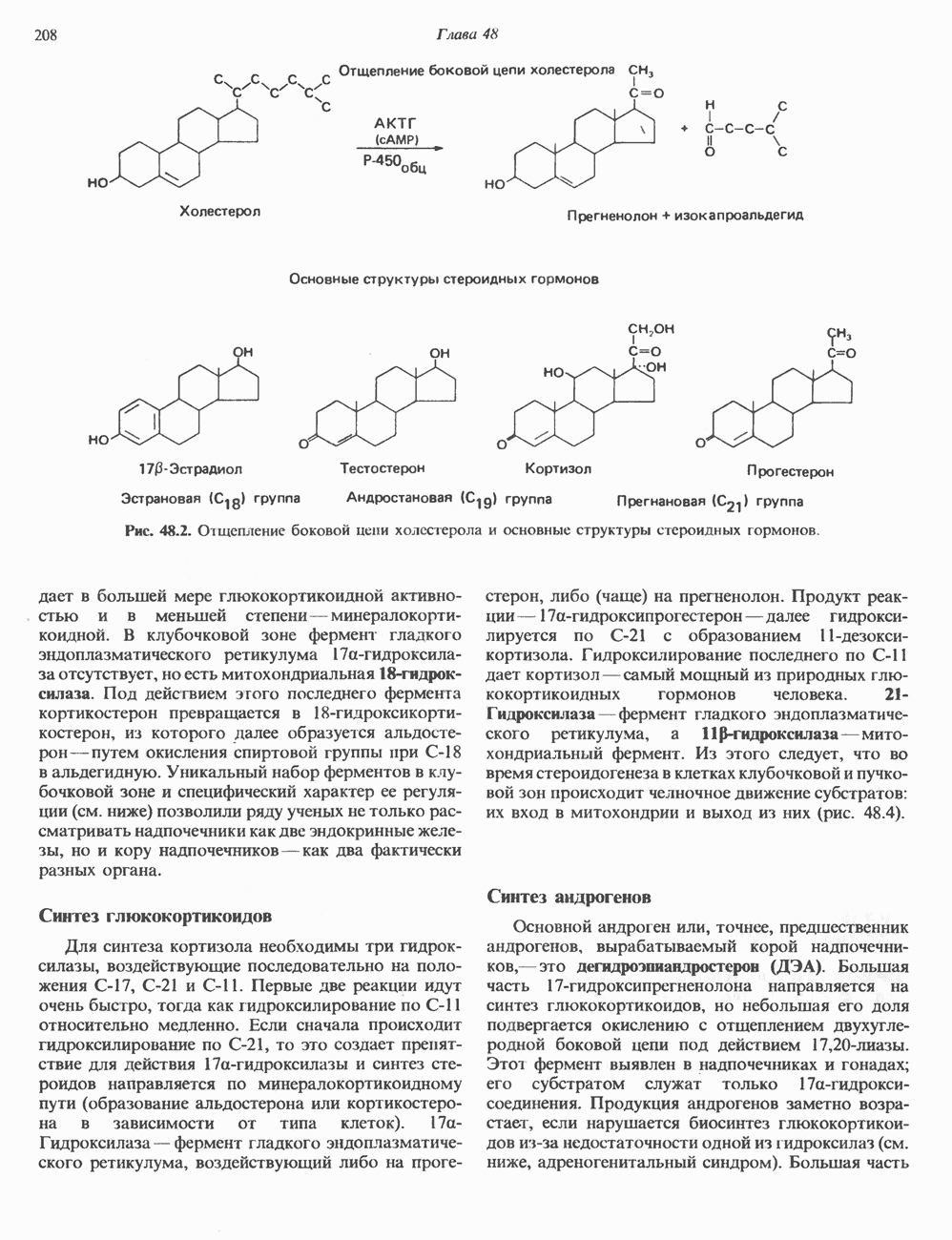
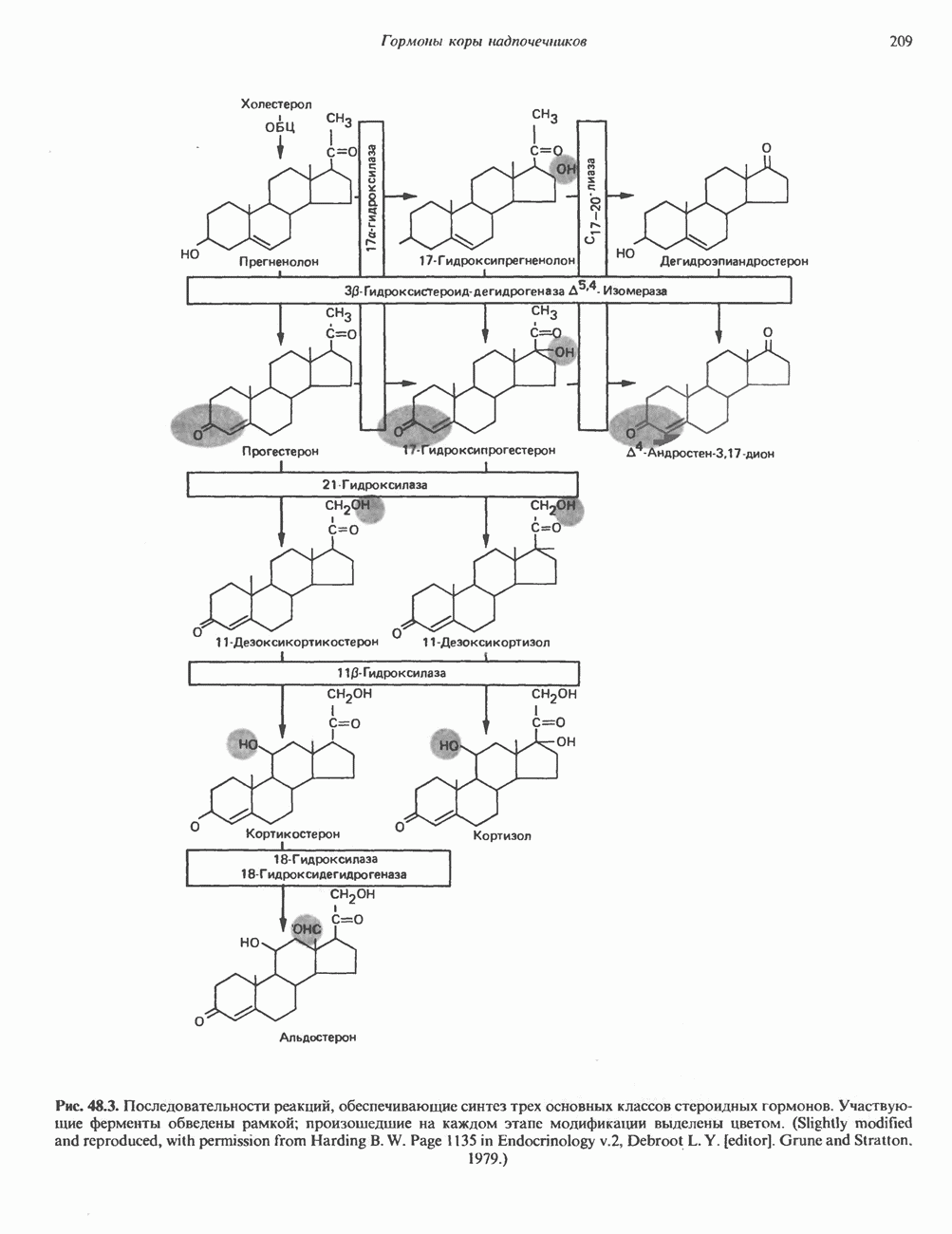
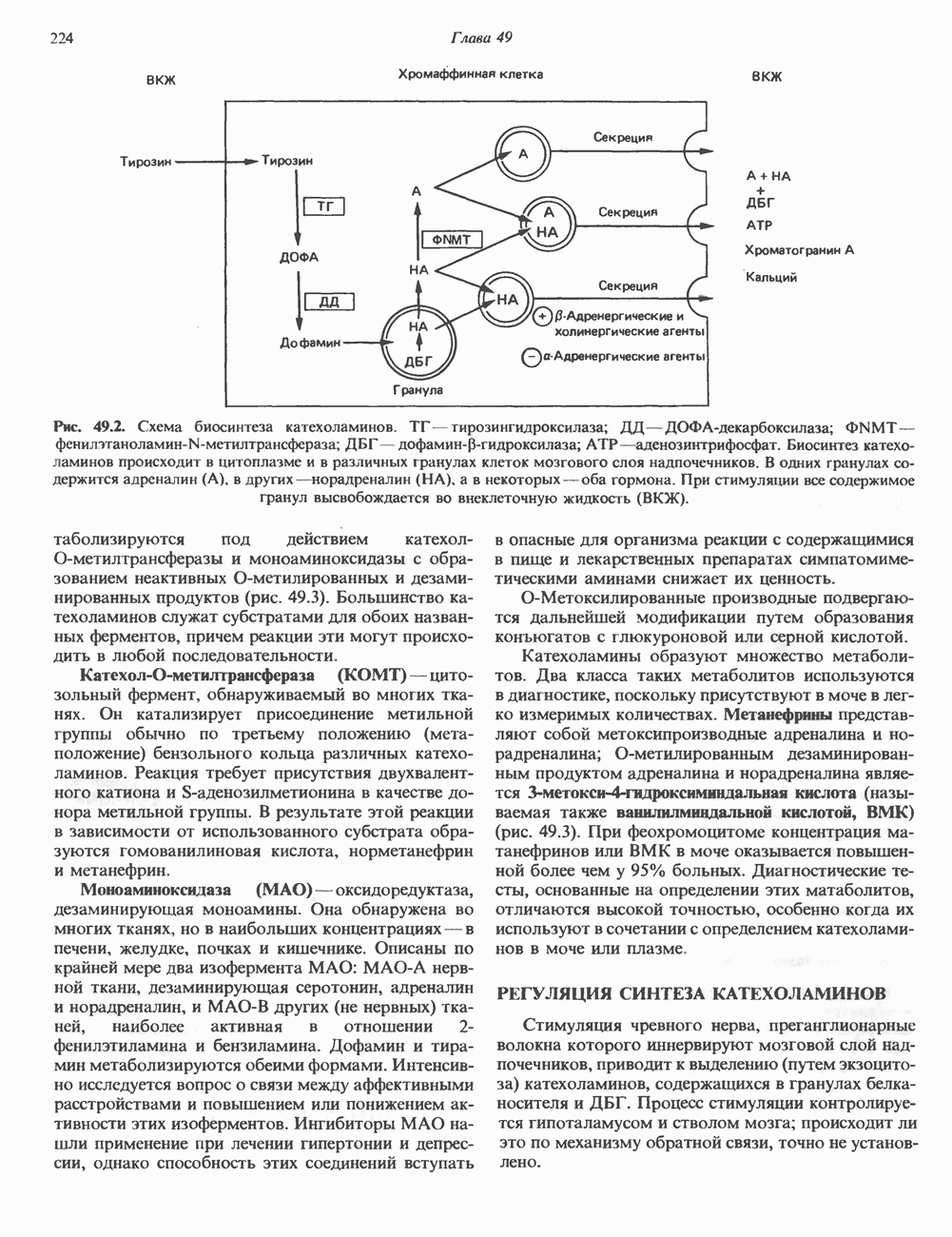
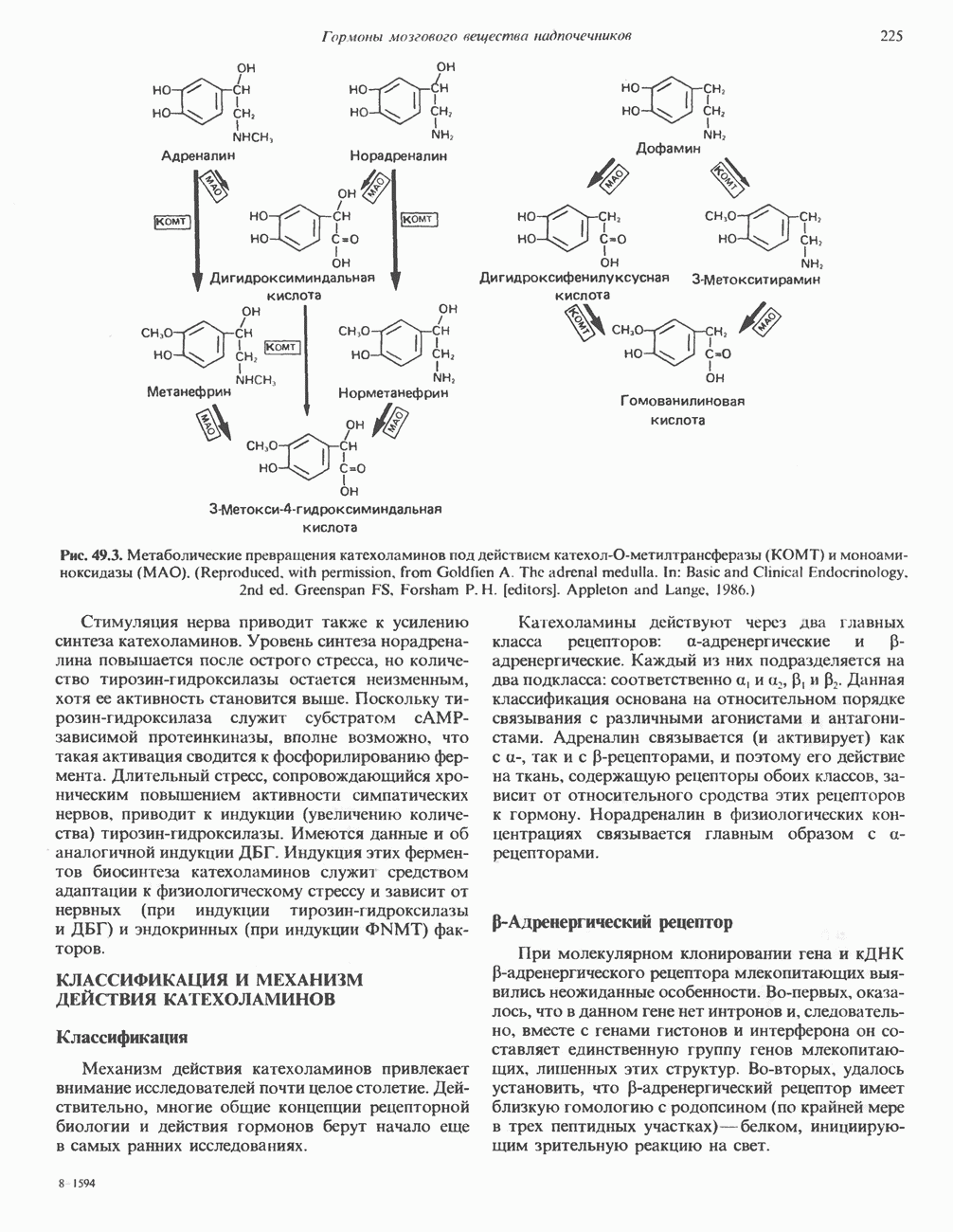
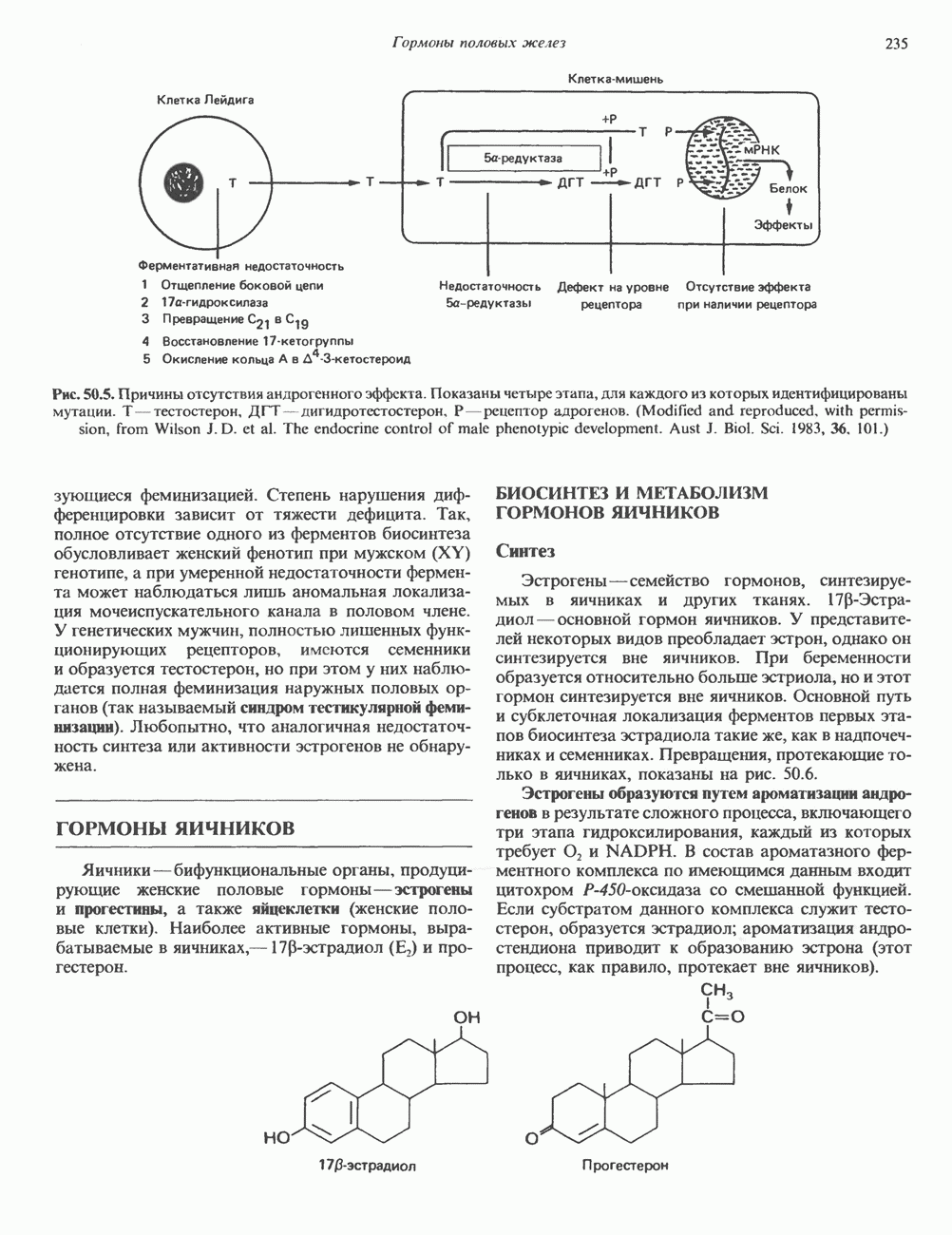
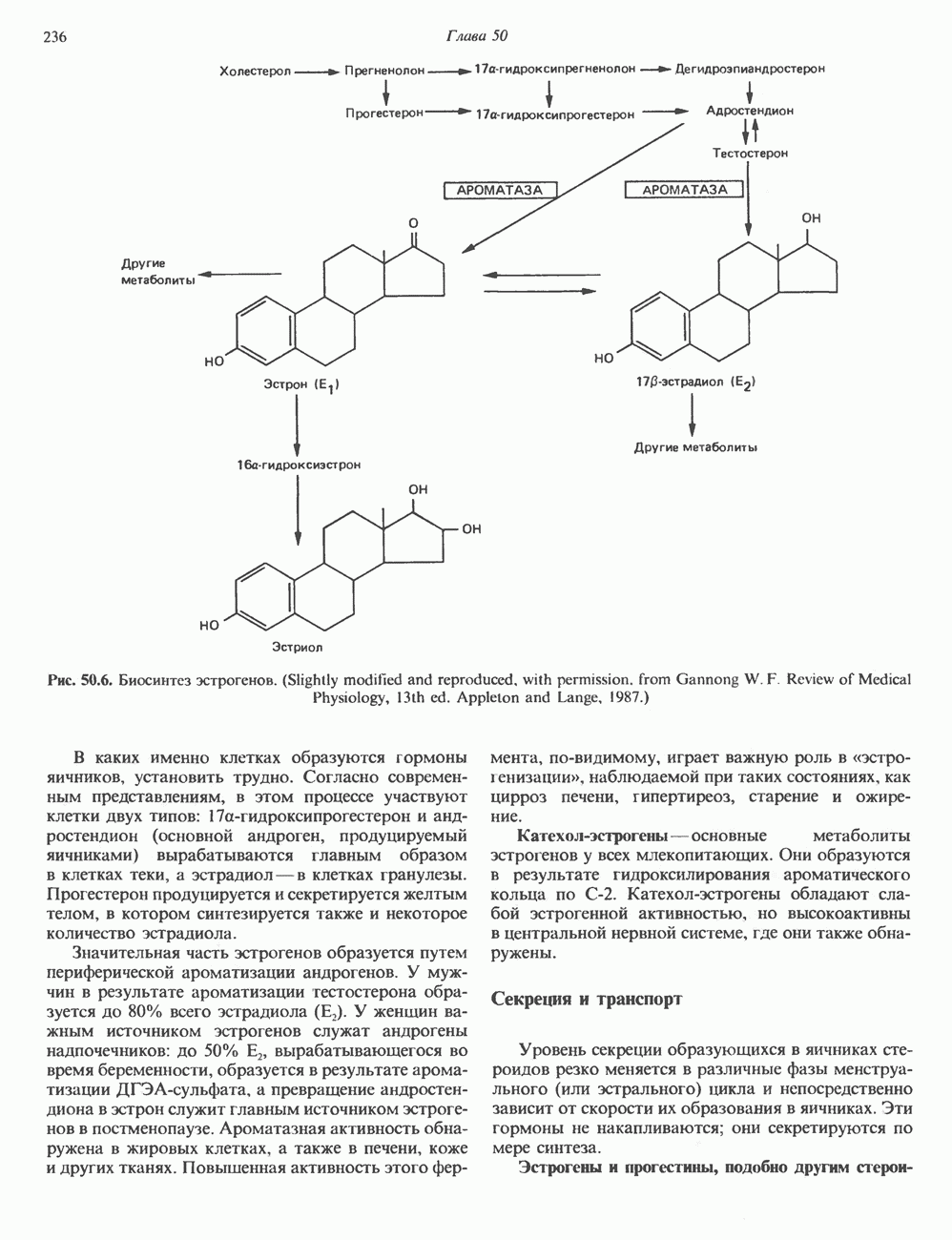
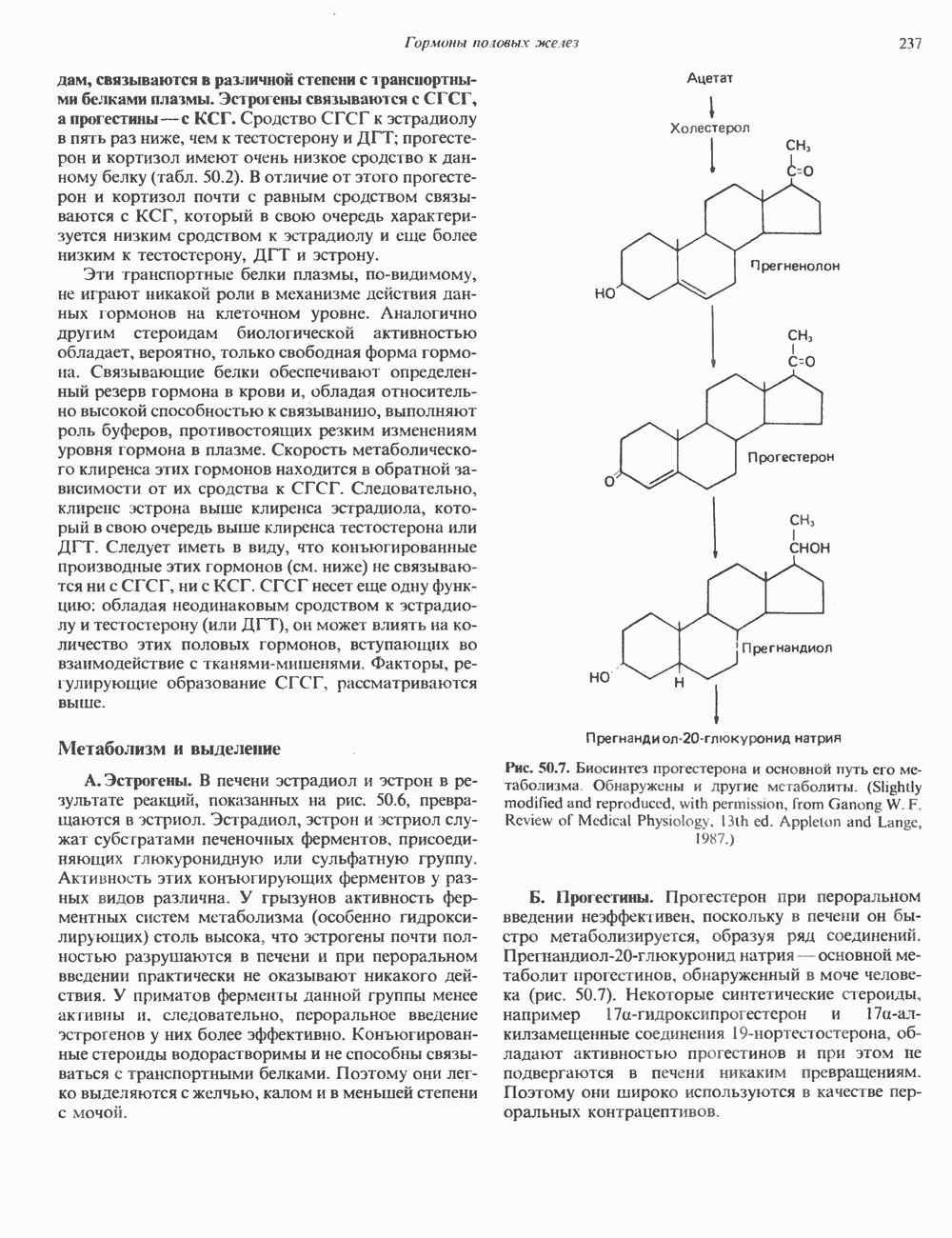
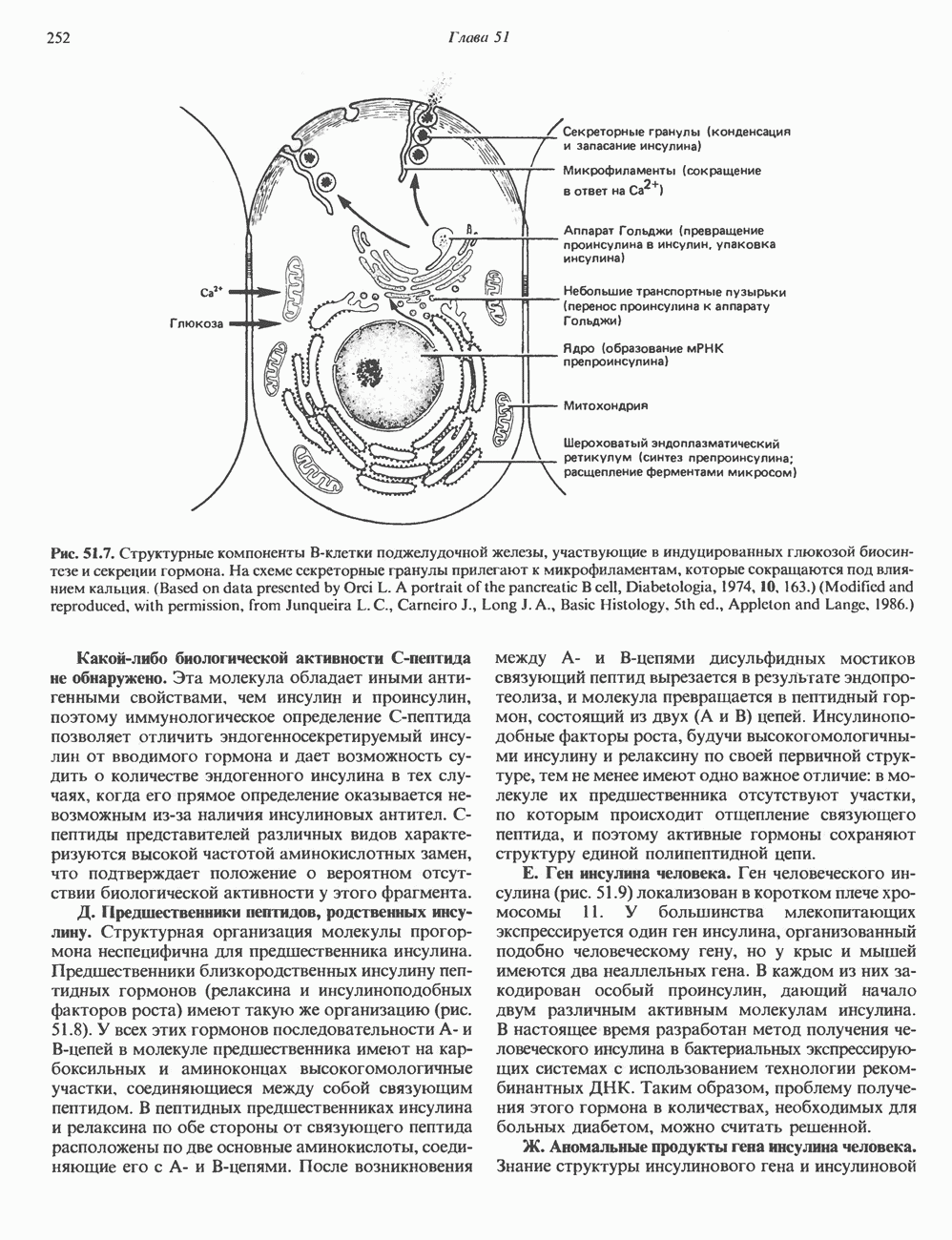
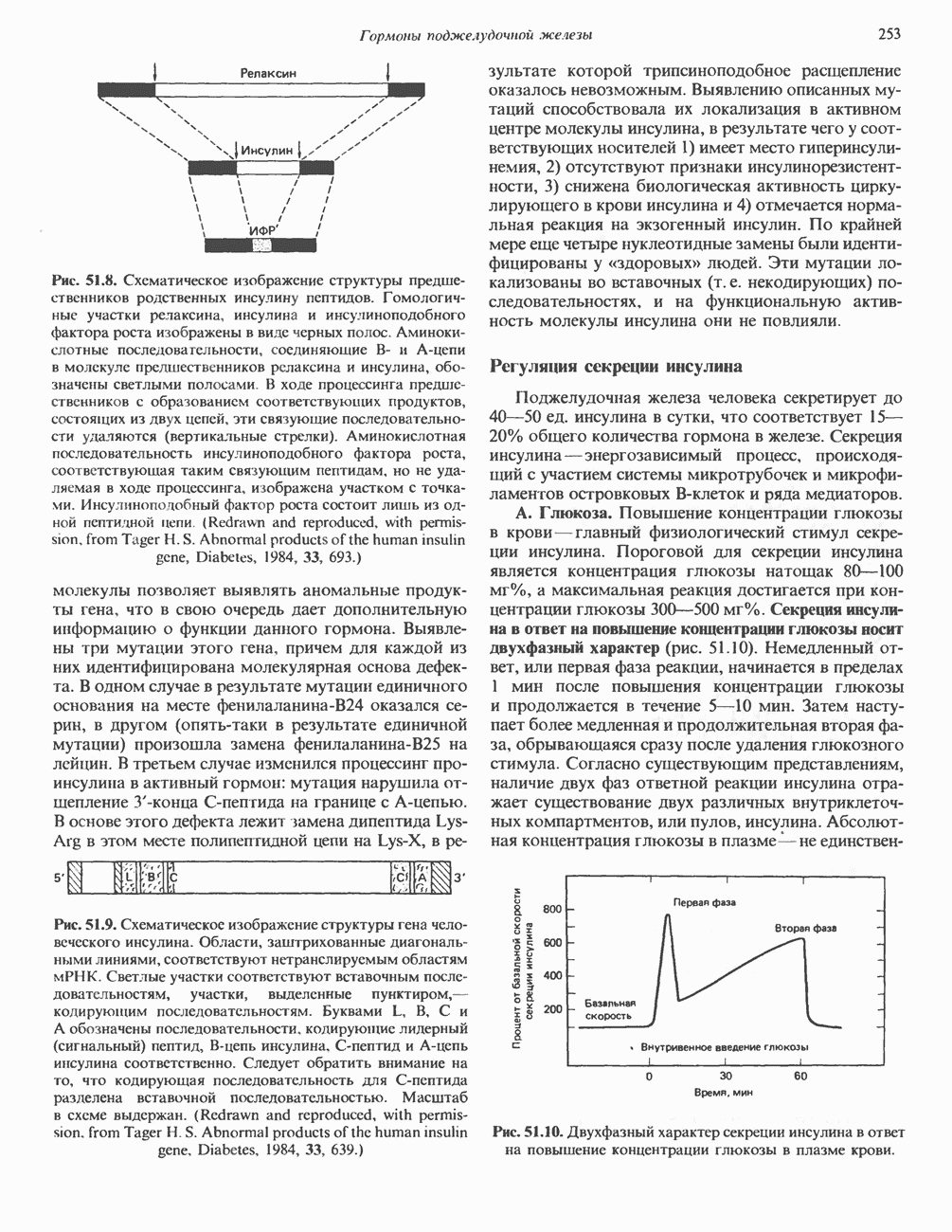
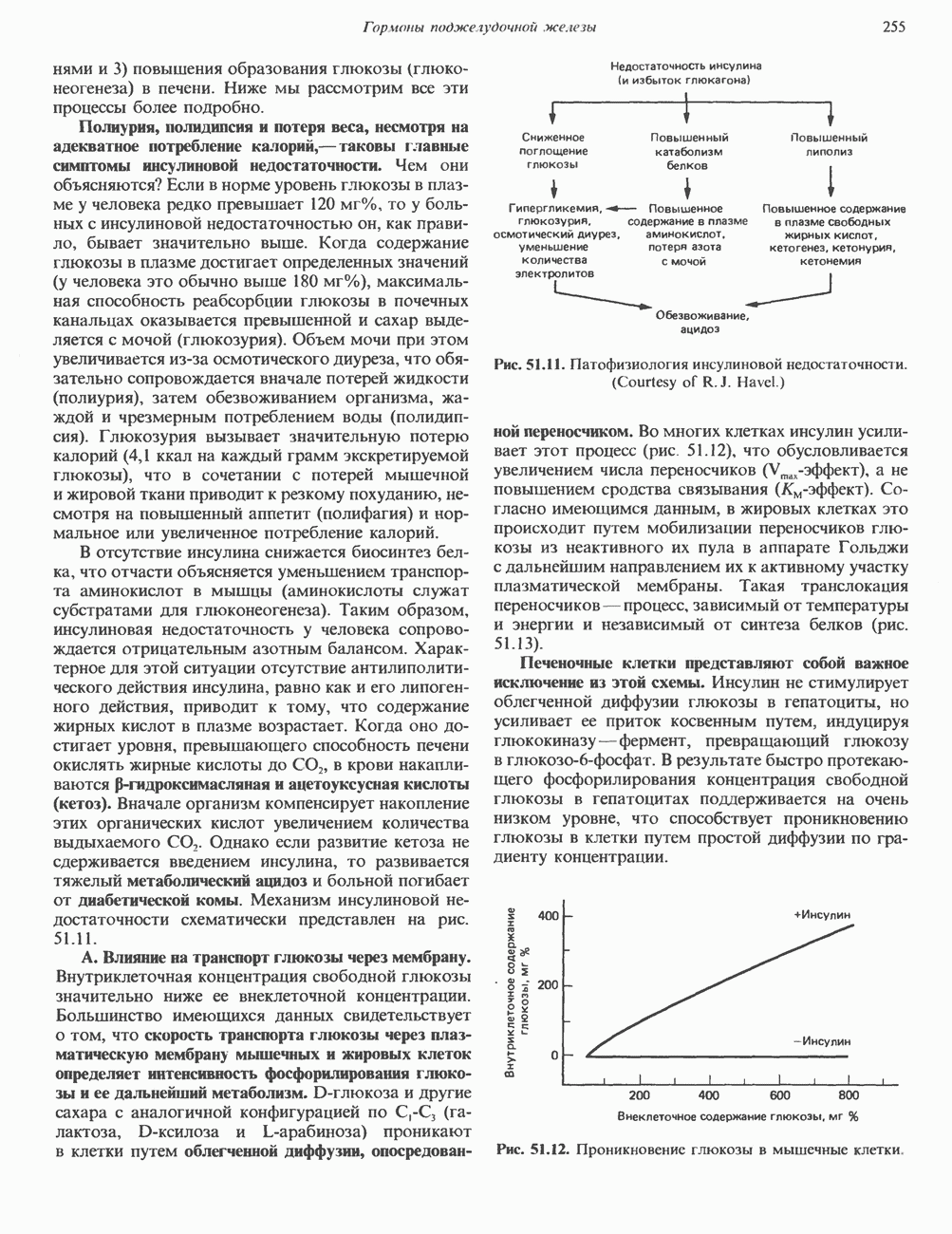
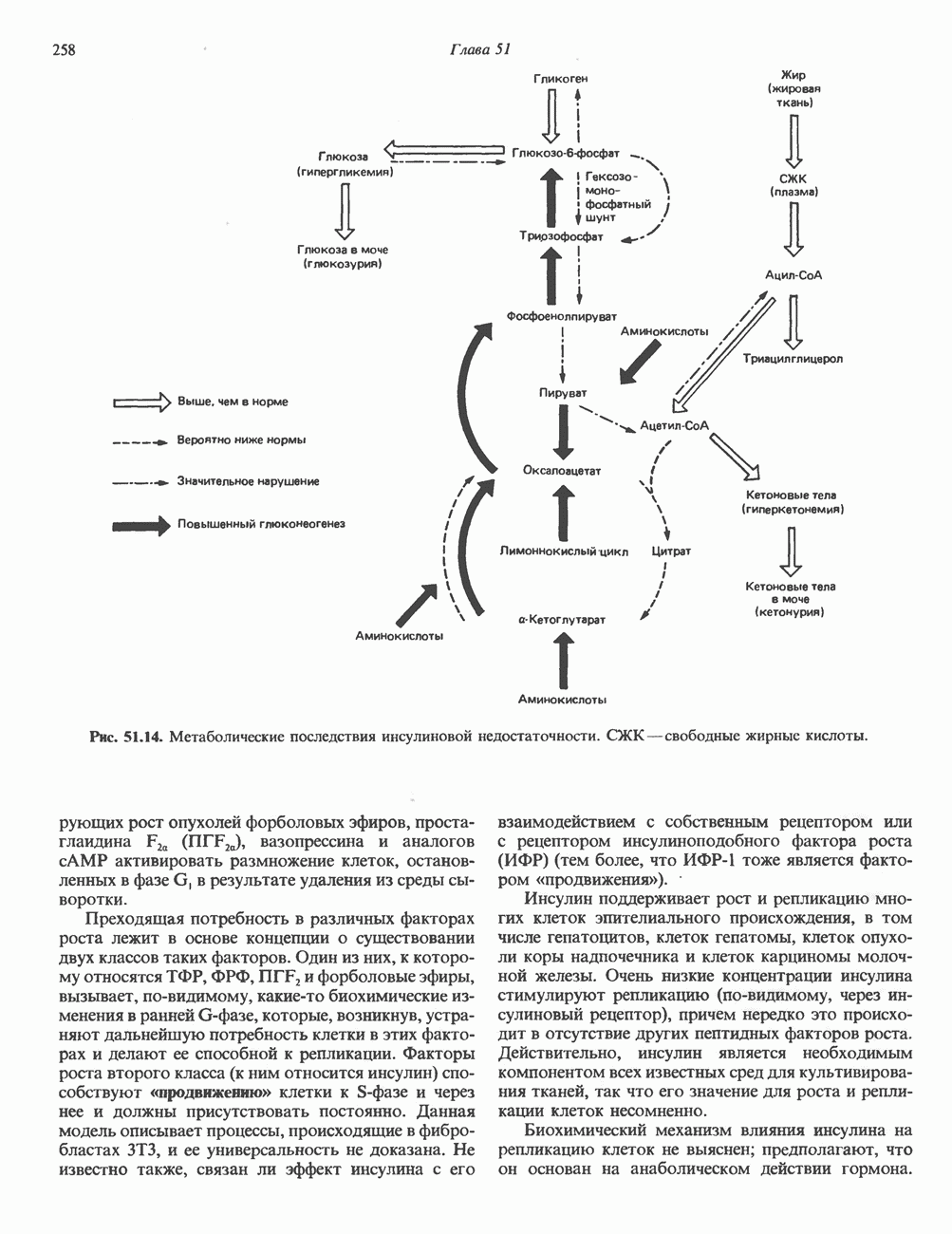
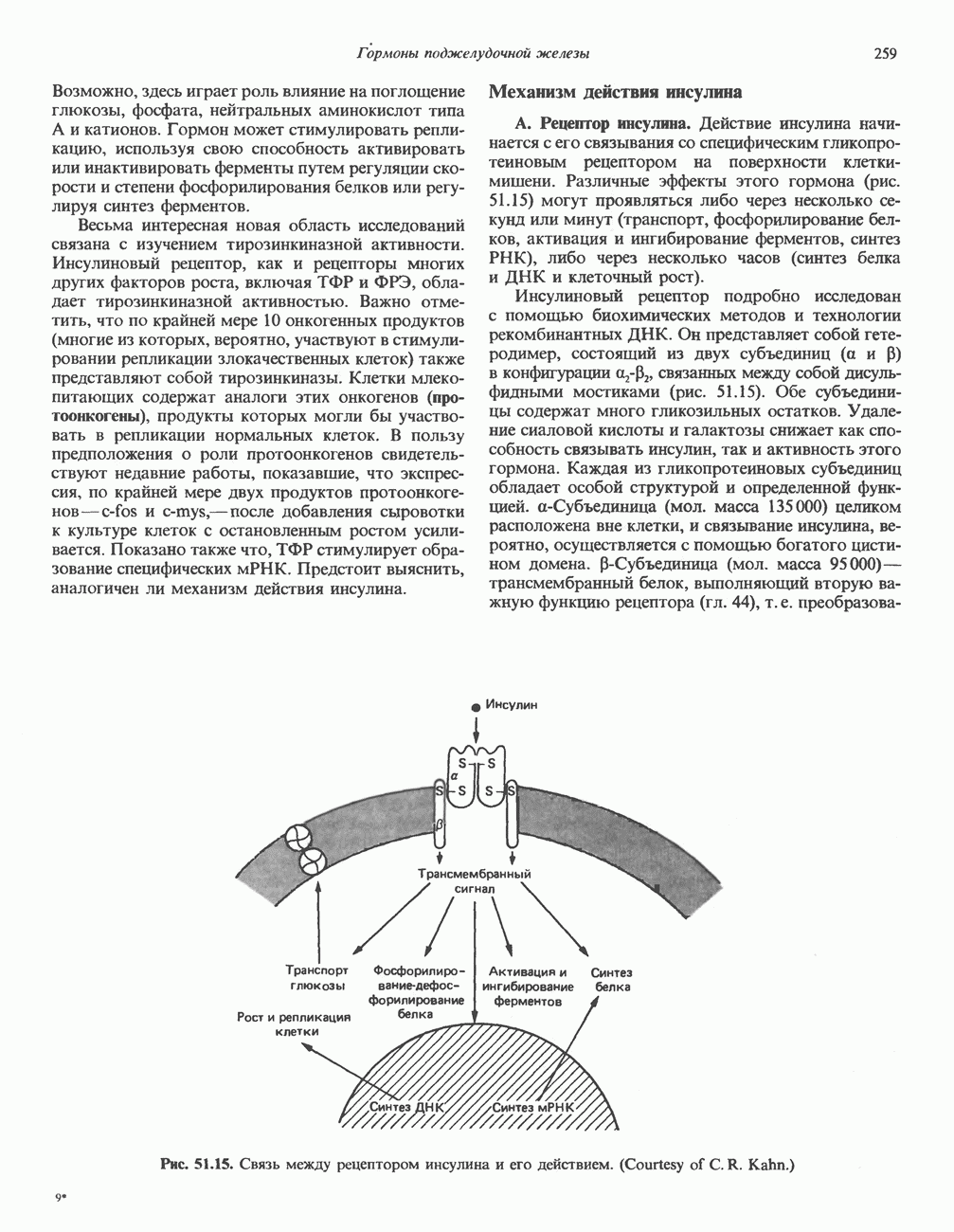
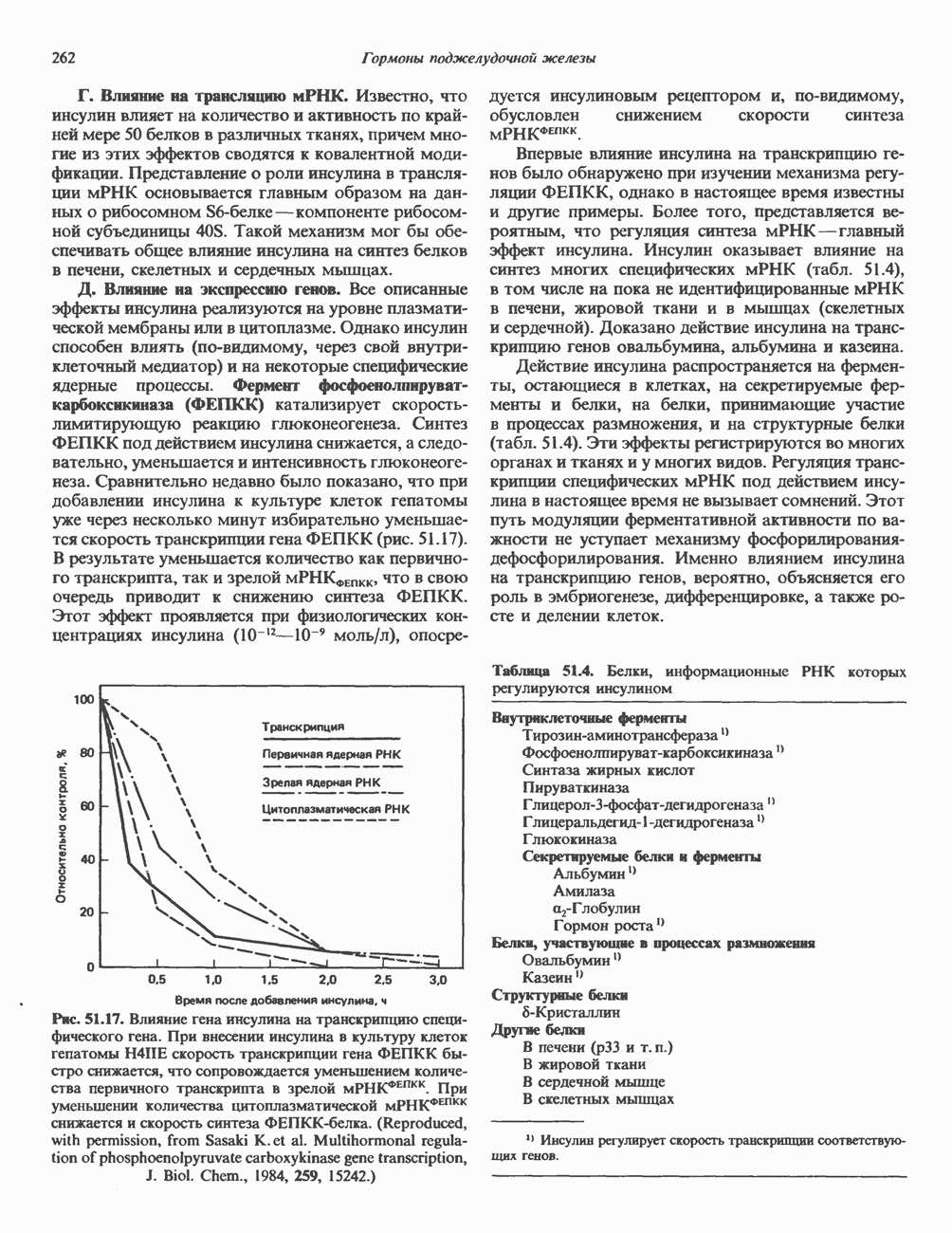
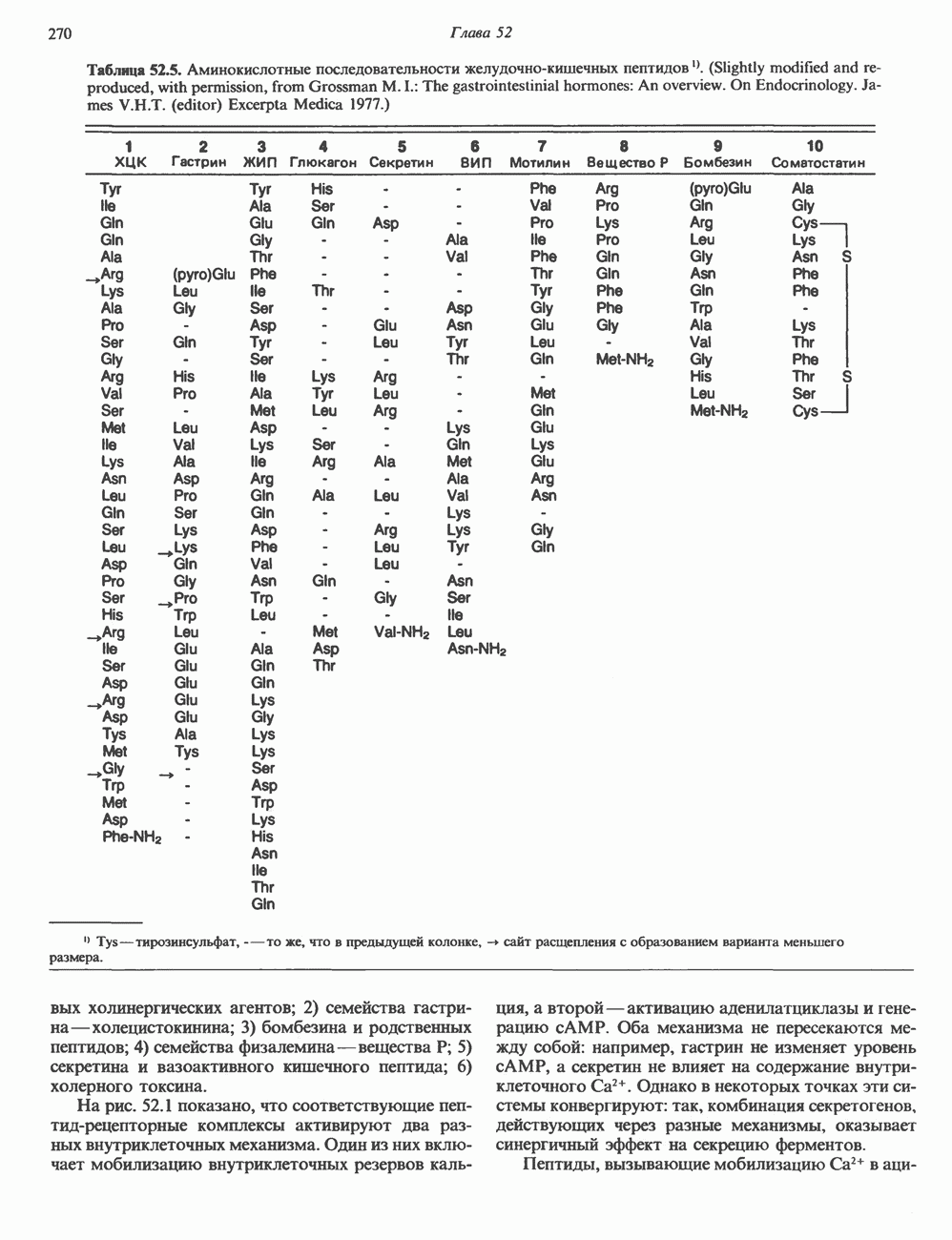
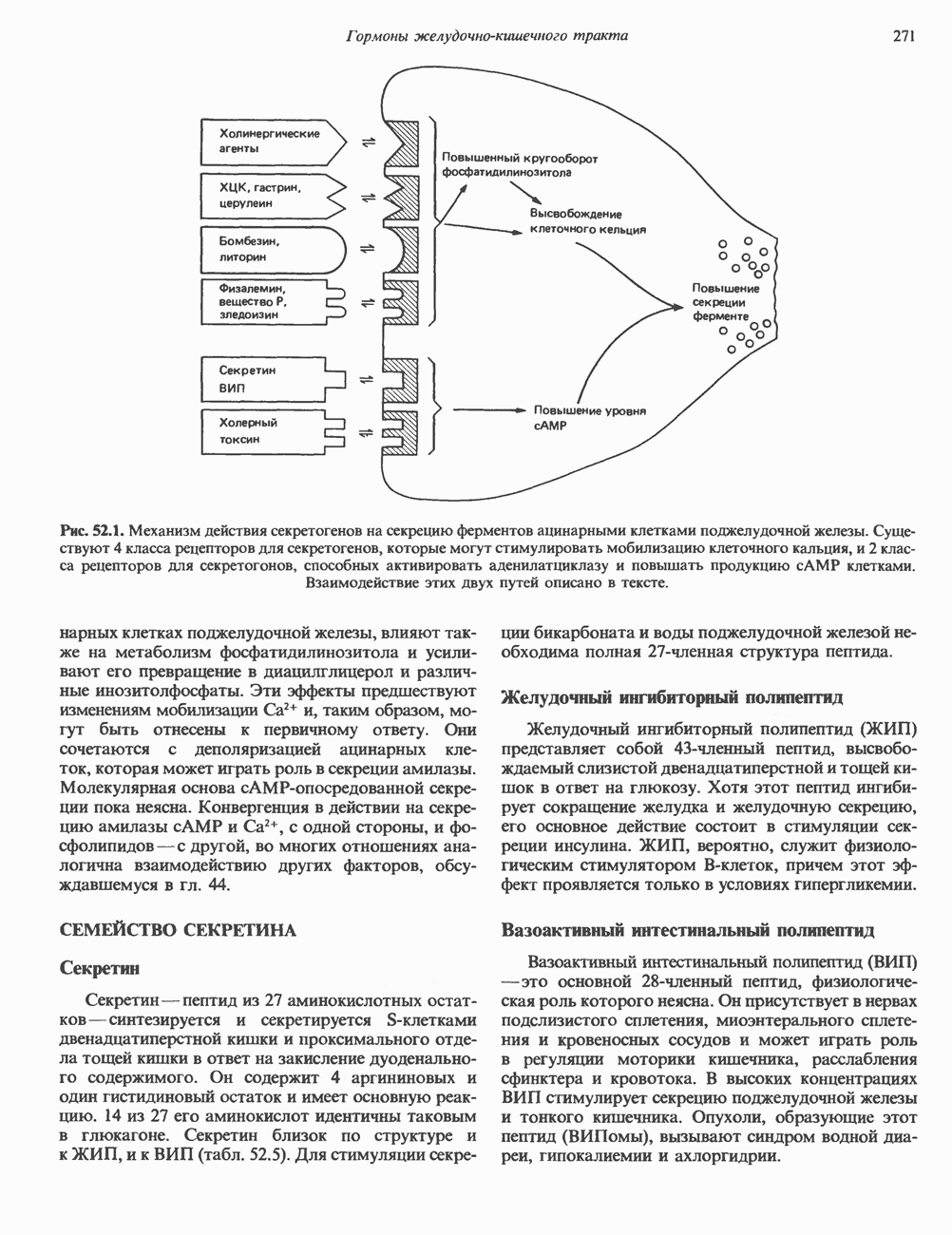
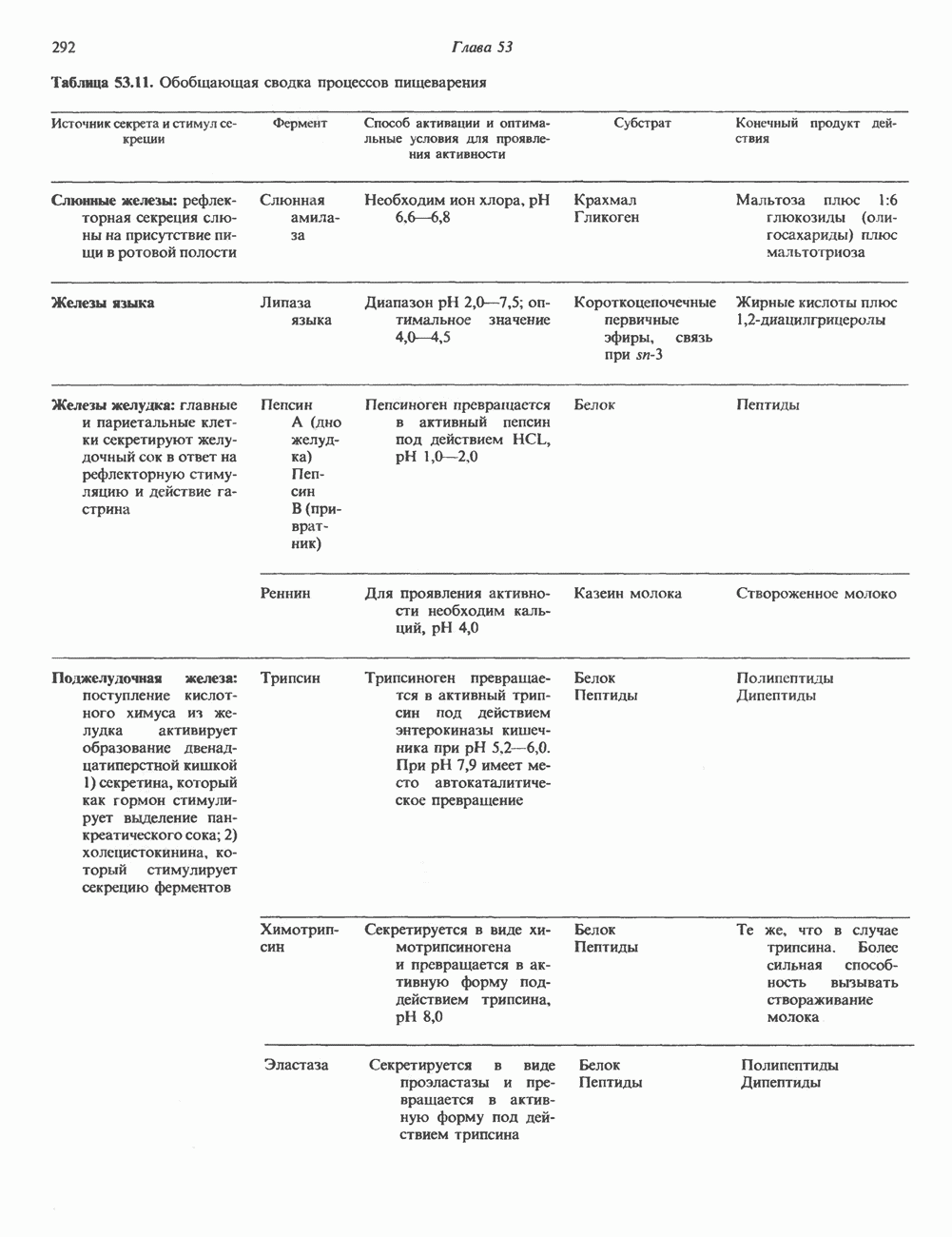
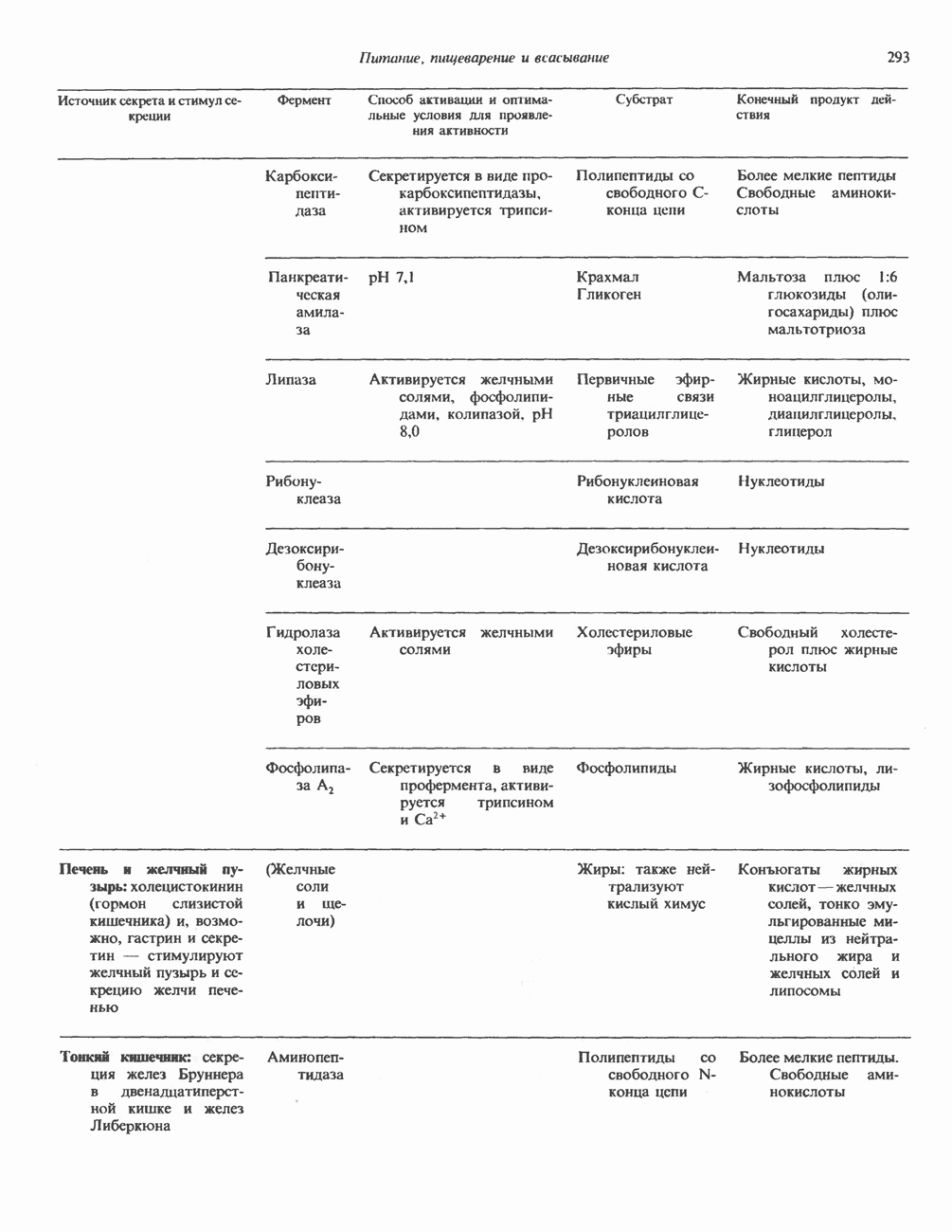
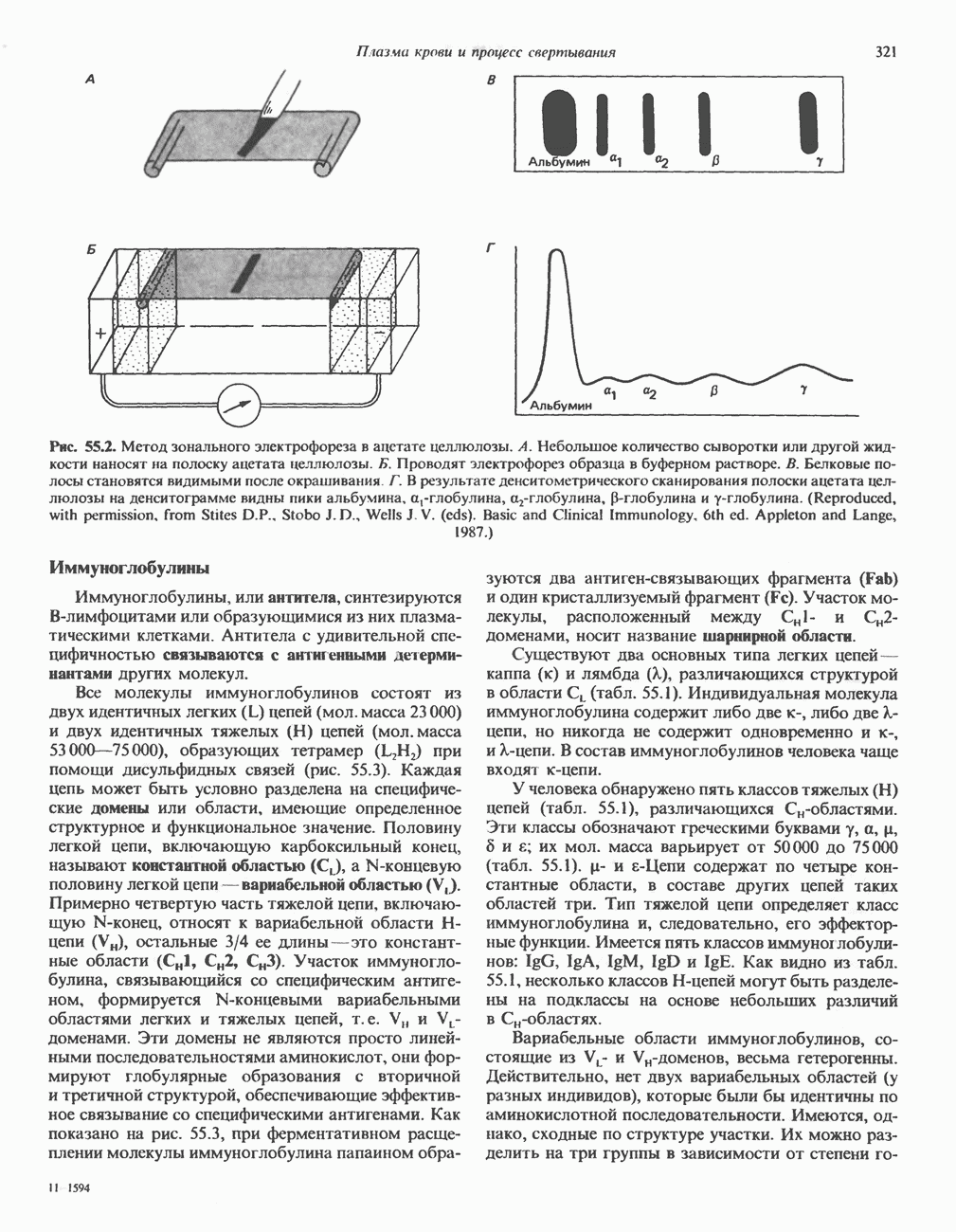
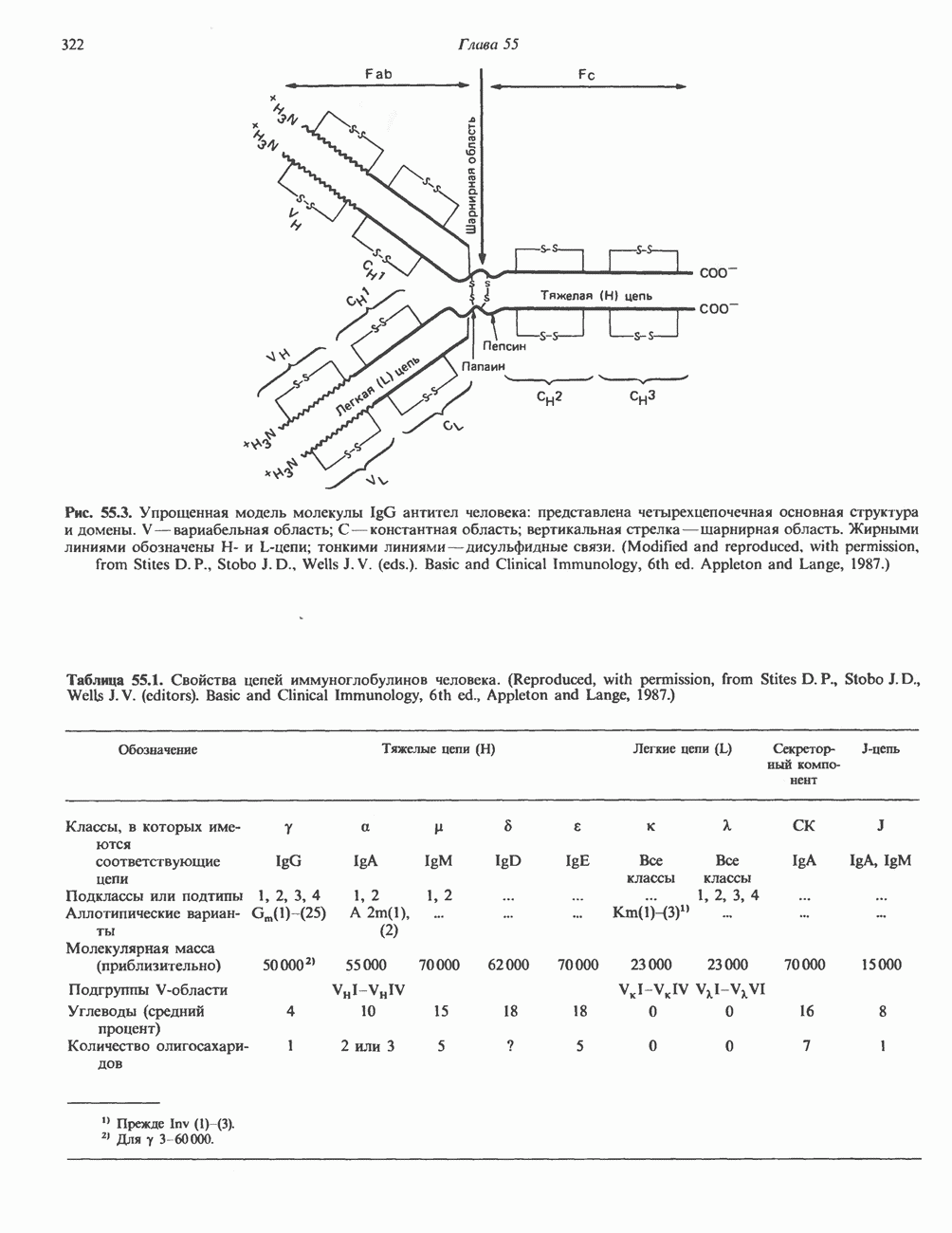
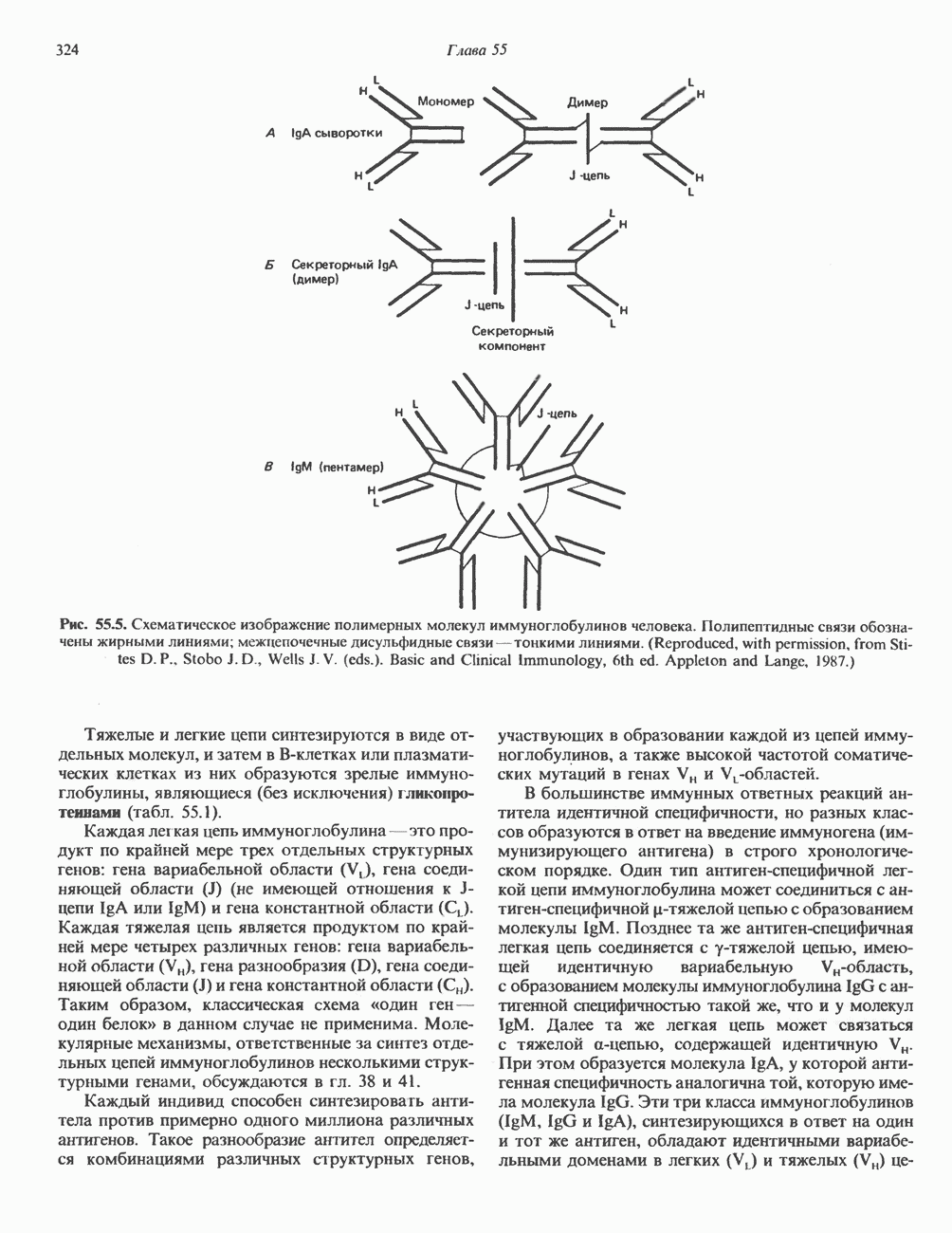
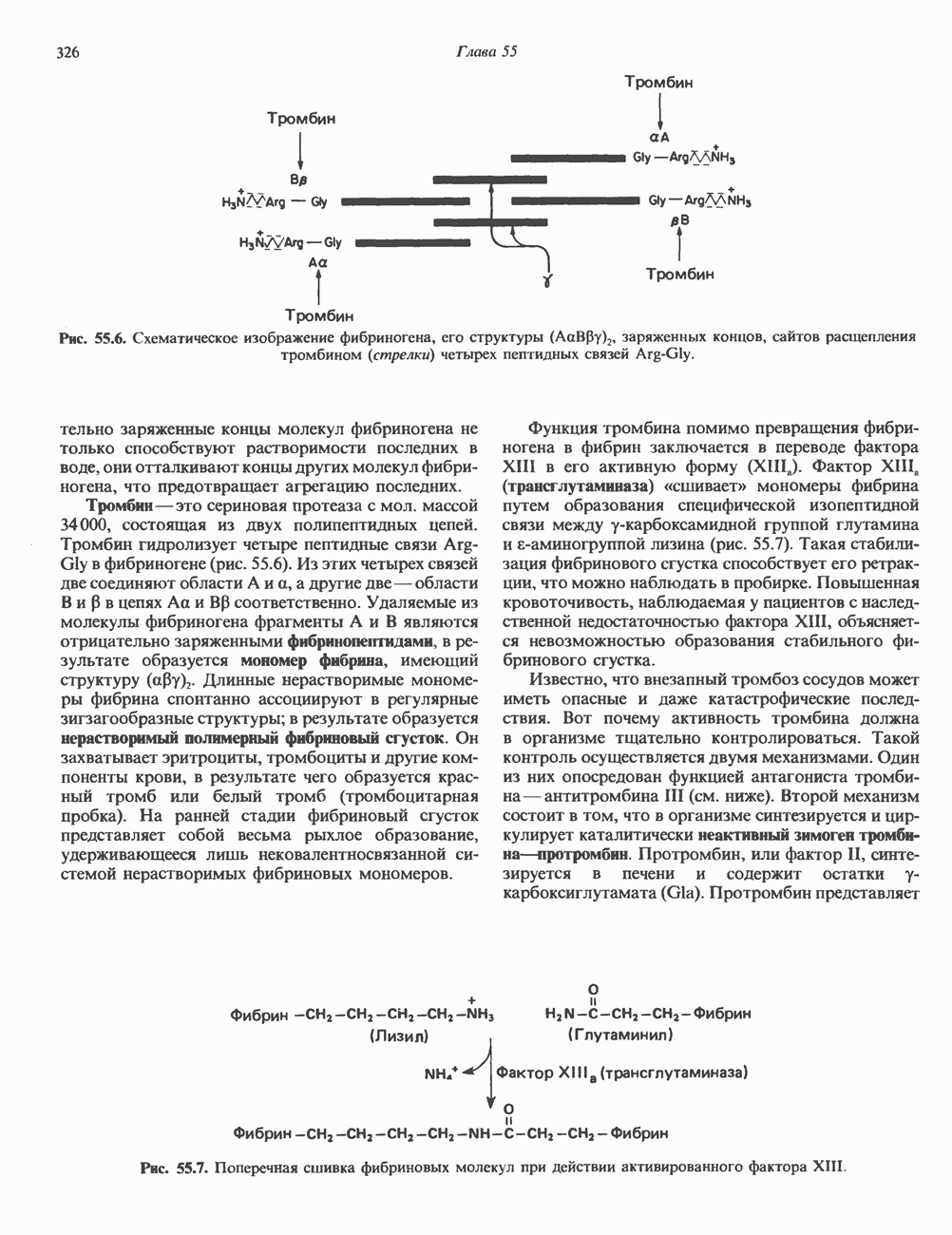
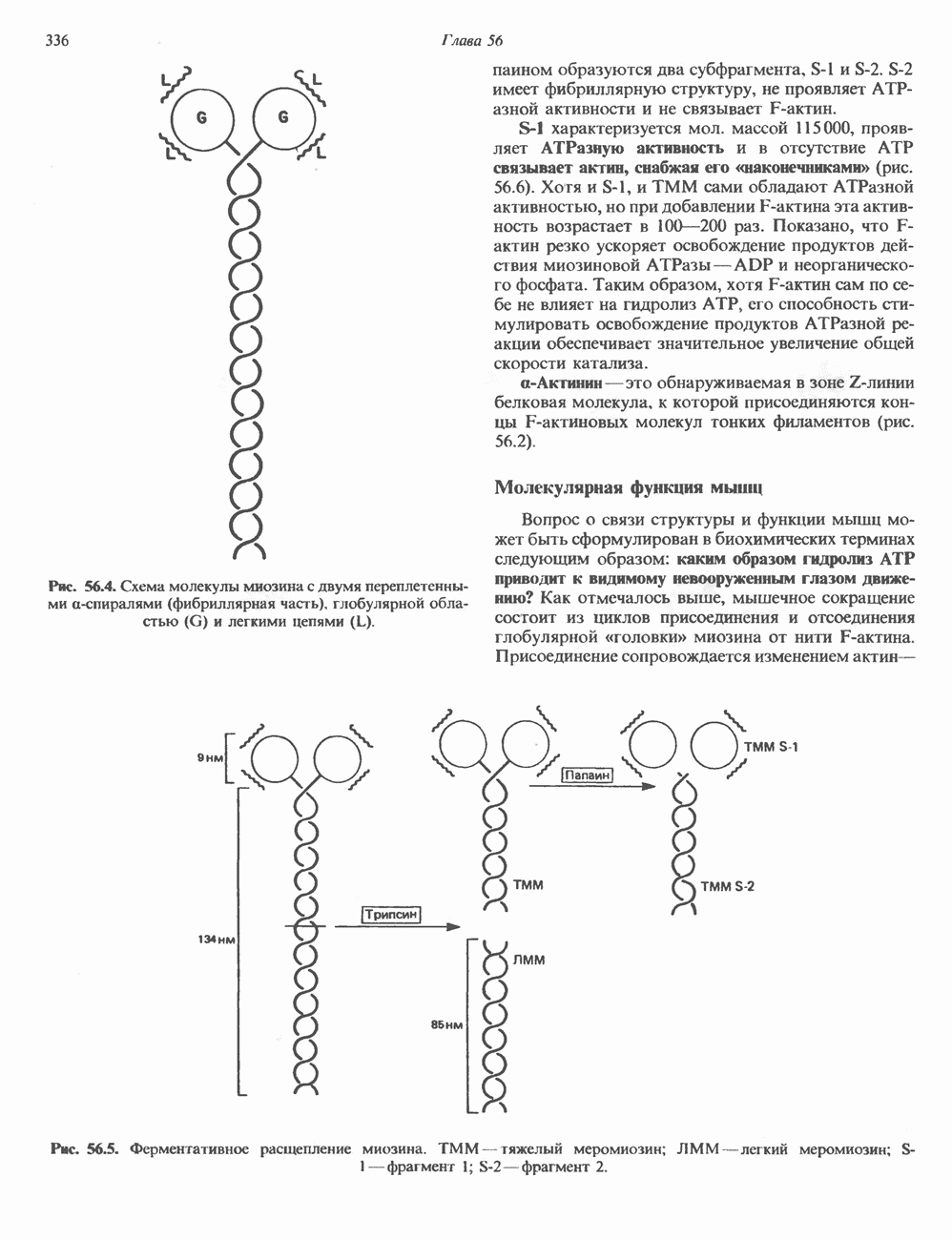
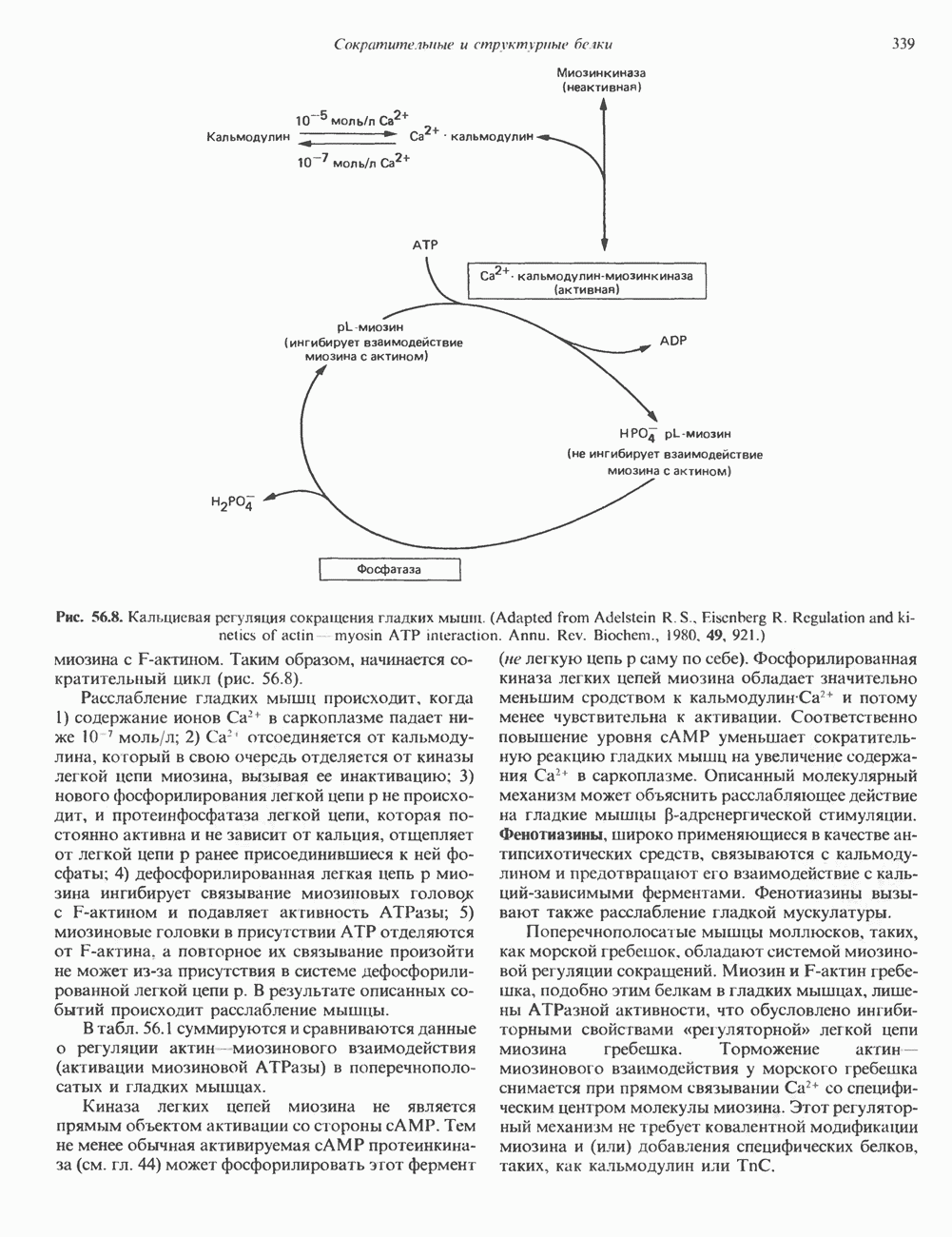
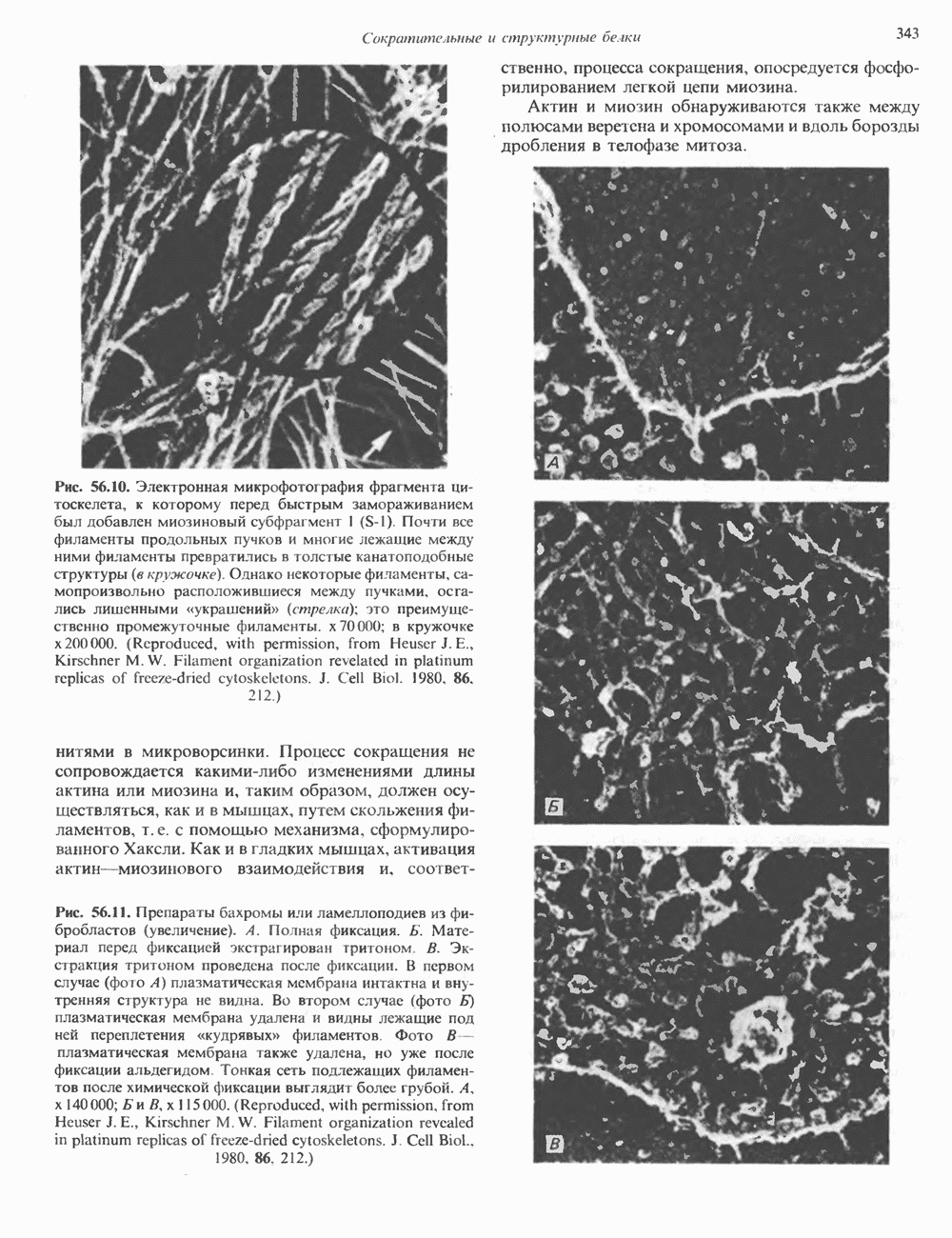
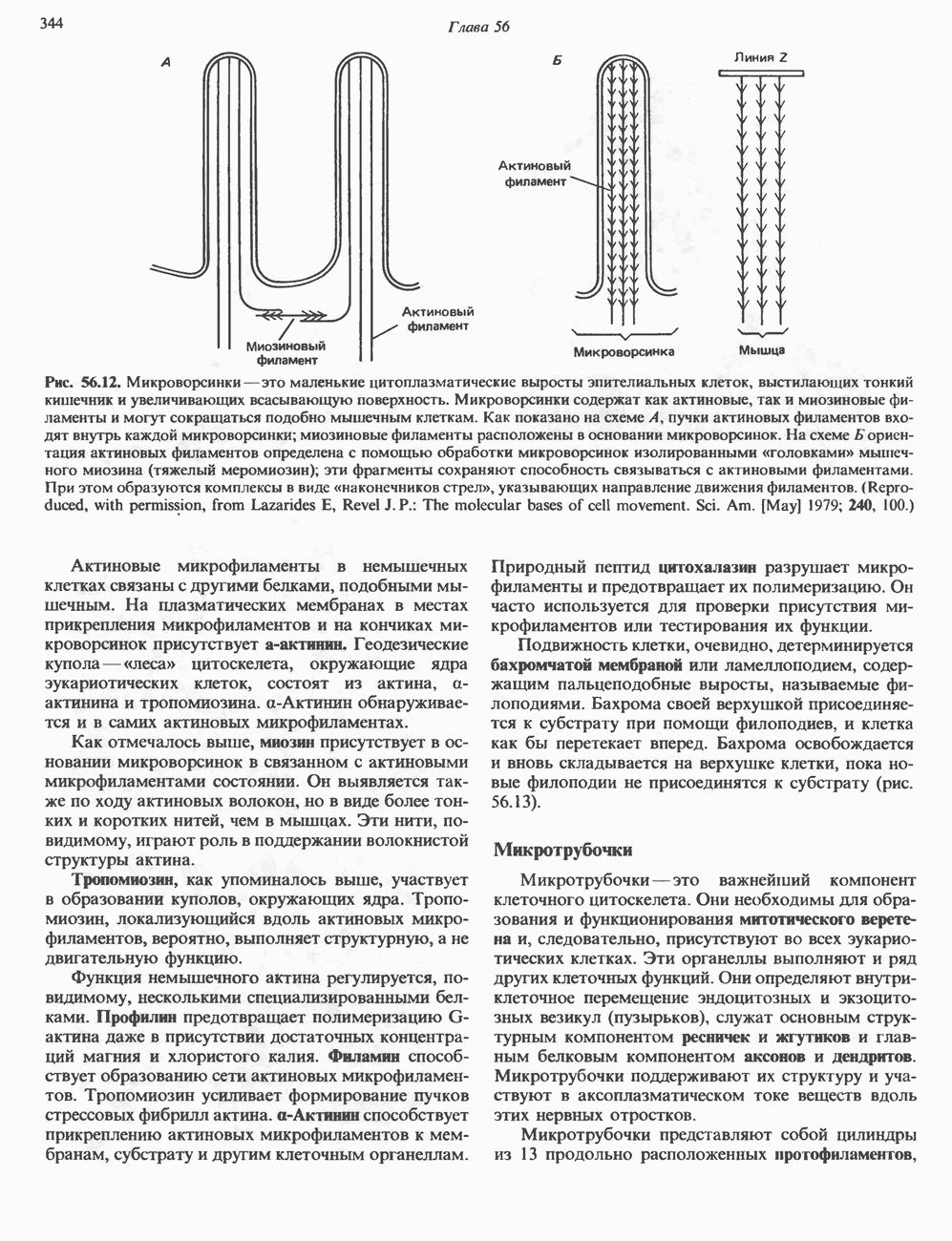
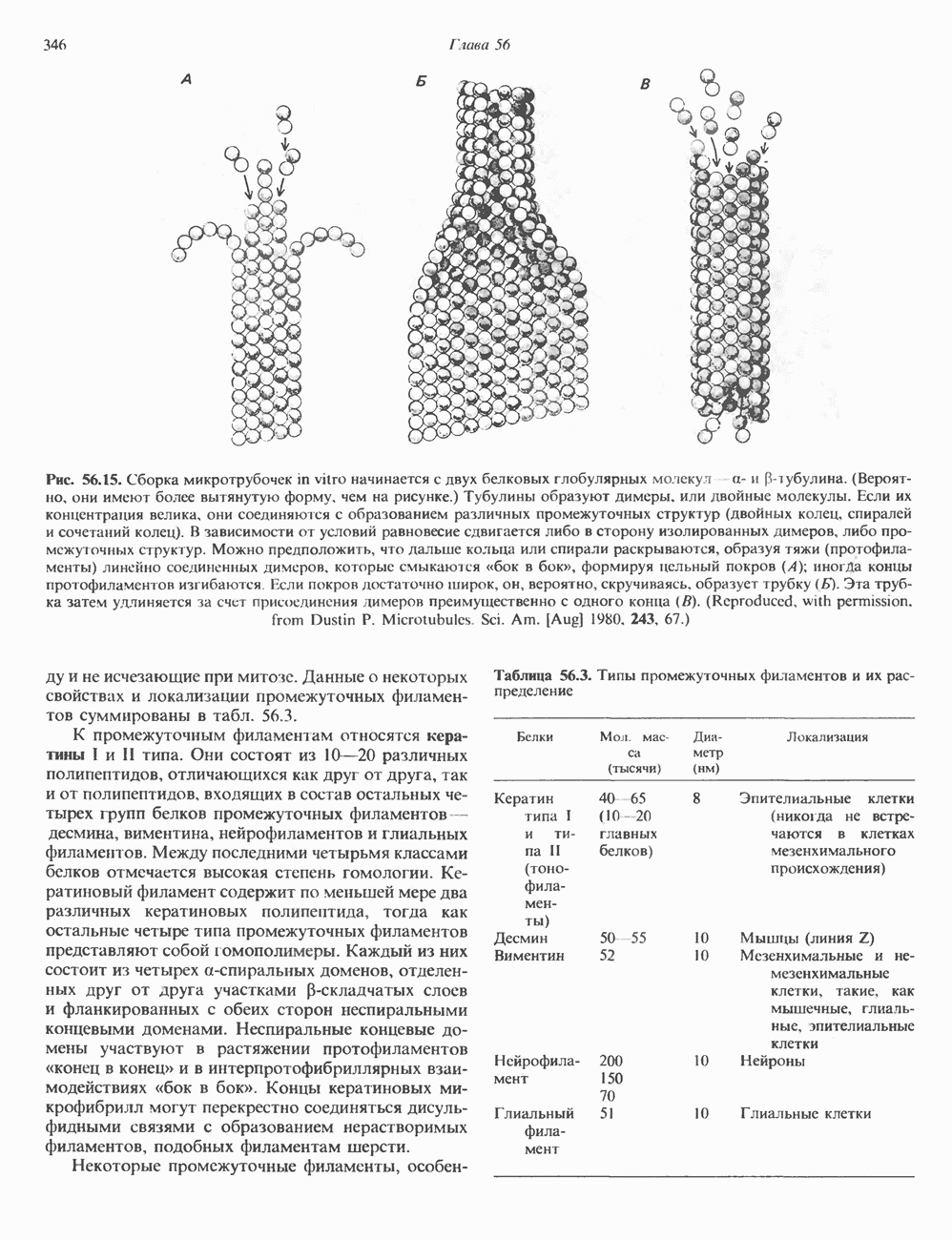
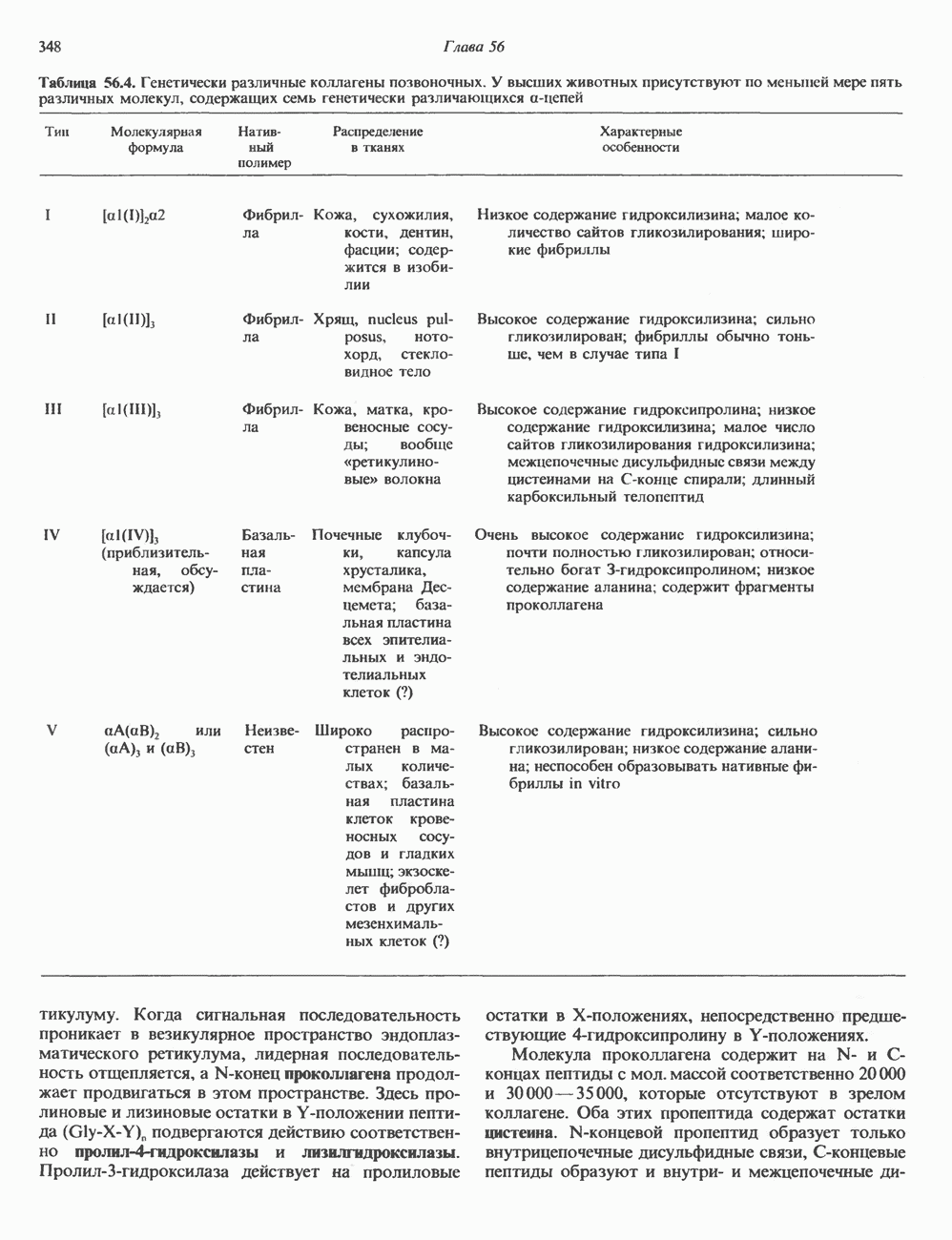
КЛАССИФИКАЦИЯ И МЕХАНИЗМ ДЕЙСТВИЯ КАТЕХОЛАМИНОВ
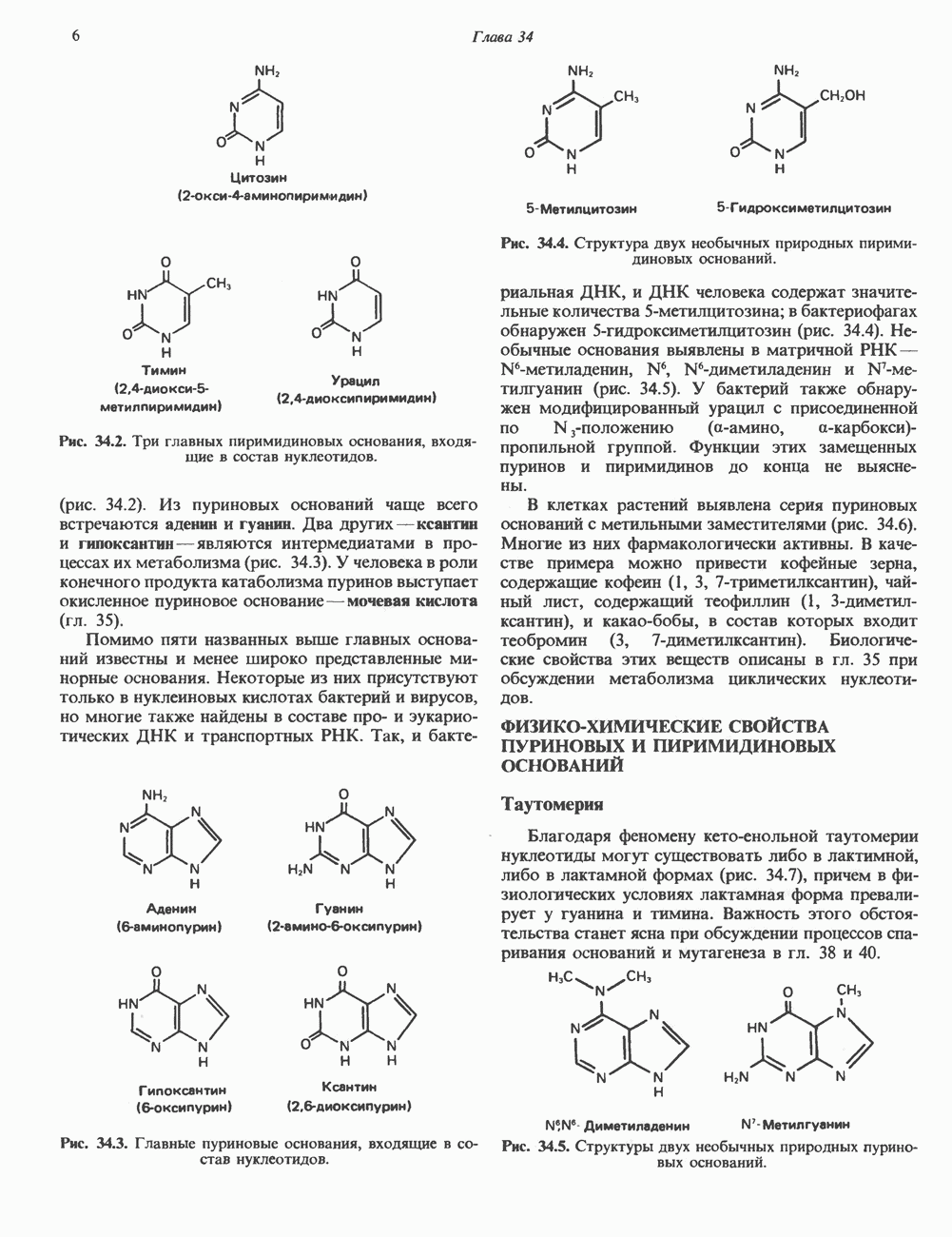
Классификация
Механизм действия катехоламинов привлекает внимание исследователей почти целое столетие. Действительно, многие общие концепции рецепторной биологии и действия гормонов берут начало еще в самых ранних исследованиях.
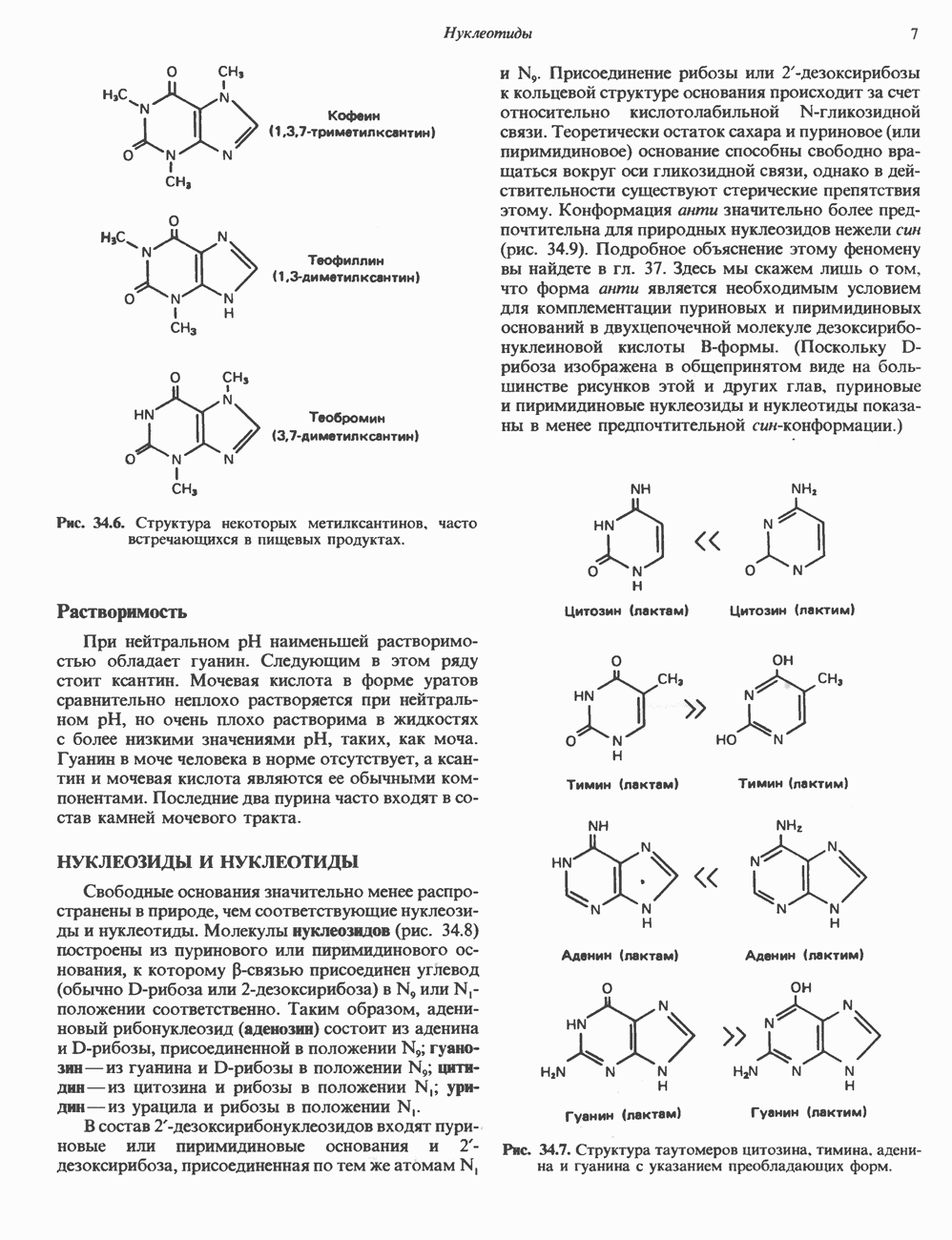
Катехоламины действуют через два главных класса рецепторов: а-адренергические и  -адренергические. Каждый из них подразделяется на два подкласса: соответственно
-адренергические. Каждый из них подразделяется на два подкласса: соответственно  и
и  . Данная классификация основана на относительном порядке связывания с различными агонистами и антагонистами. Адреналин связывается (и активирует) как с
. Данная классификация основана на относительном порядке связывания с различными агонистами и антагонистами. Адреналин связывается (и активирует) как с  , так и с
, так и с  -рецепторами, и поэтому его действие на ткань, содержащую рецепторы обоих классов, зависит от относительного сродства этих рецепторов к гормону. Норадреналин в физиологических концентрациях связывается главным образом с а-рецепторами.
-рецепторами, и поэтому его действие на ткань, содержащую рецепторы обоих классов, зависит от относительного сродства этих рецепторов к гормону. Норадреналин в физиологических концентрациях связывается главным образом с а-рецепторами.
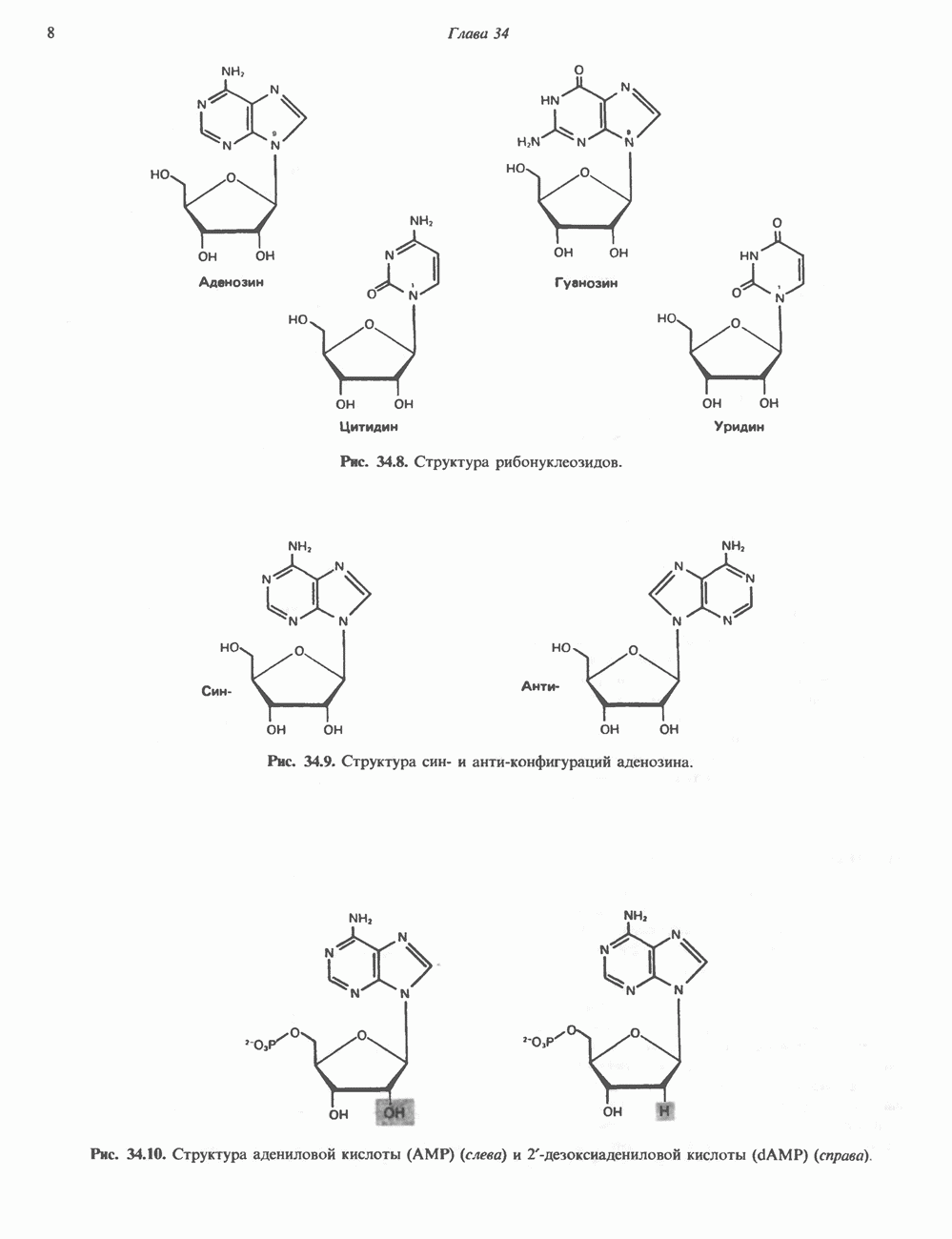
b-Адренергический рецептор
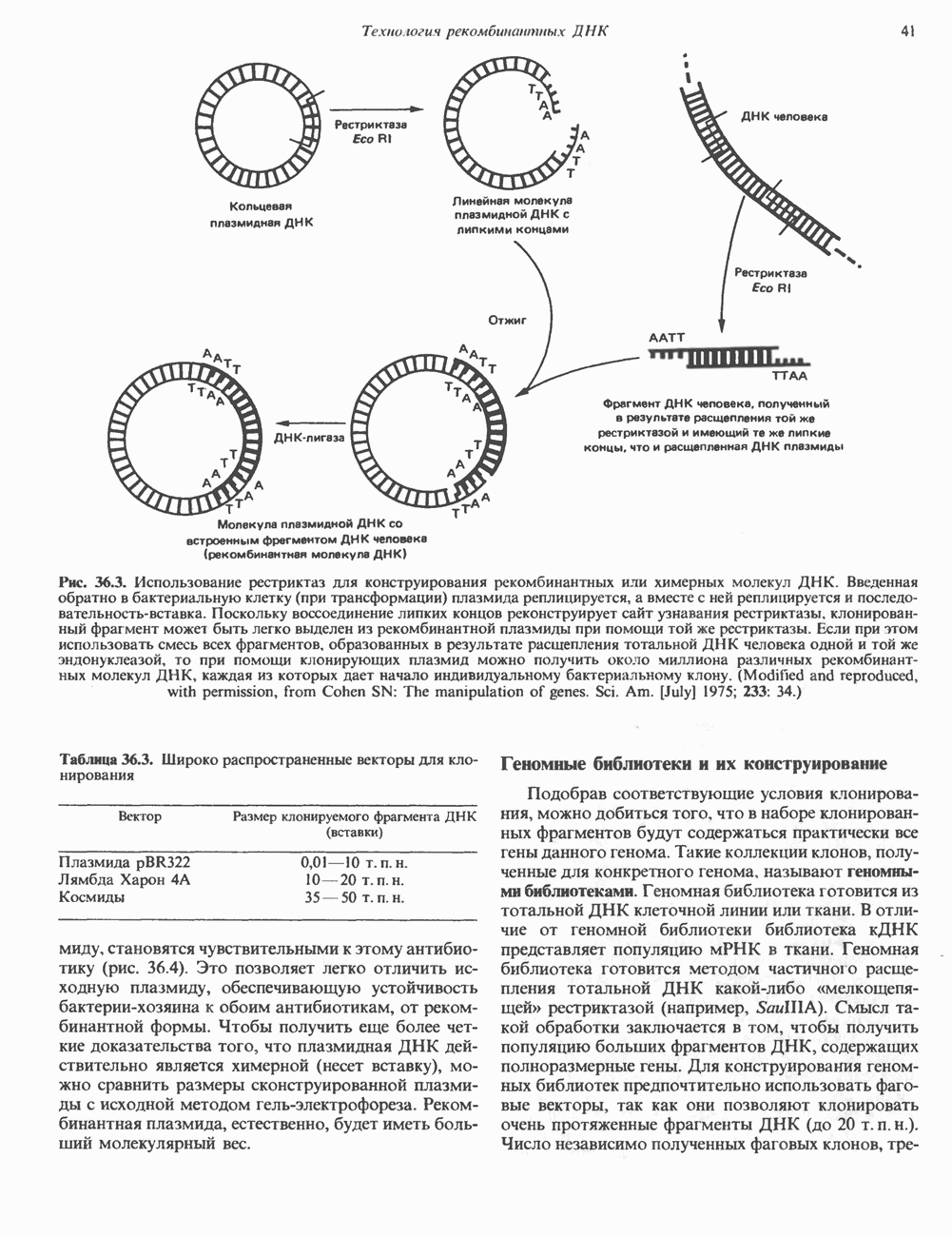
При молекулярном клонировании гена и кДНК  -адренергического рецептора млекопитающих выявились неожиданные особенности. Во-первых, оказалось, что в данном гене нет интронов и, следовательно, вместе с генами гистонов и интерферона он составляет единственную группу генов млекопитающих, лишенных этих структур. Во-вторых, удалось установить, что
-адренергического рецептора млекопитающих выявились неожиданные особенности. Во-первых, оказалось, что в данном гене нет интронов и, следовательно, вместе с генами гистонов и интерферона он составляет единственную группу генов млекопитающих, лишенных этих структур. Во-вторых, удалось установить, что  -адренергический рецептор имеет близкую гомологию с родопсином (по крайней мере в трех пептидных участках) - белком, инициирующим зрительную реакцию на свет.
-адренергический рецептор имеет близкую гомологию с родопсином (по крайней мере в трех пептидных участках) - белком, инициирующим зрительную реакцию на свет.
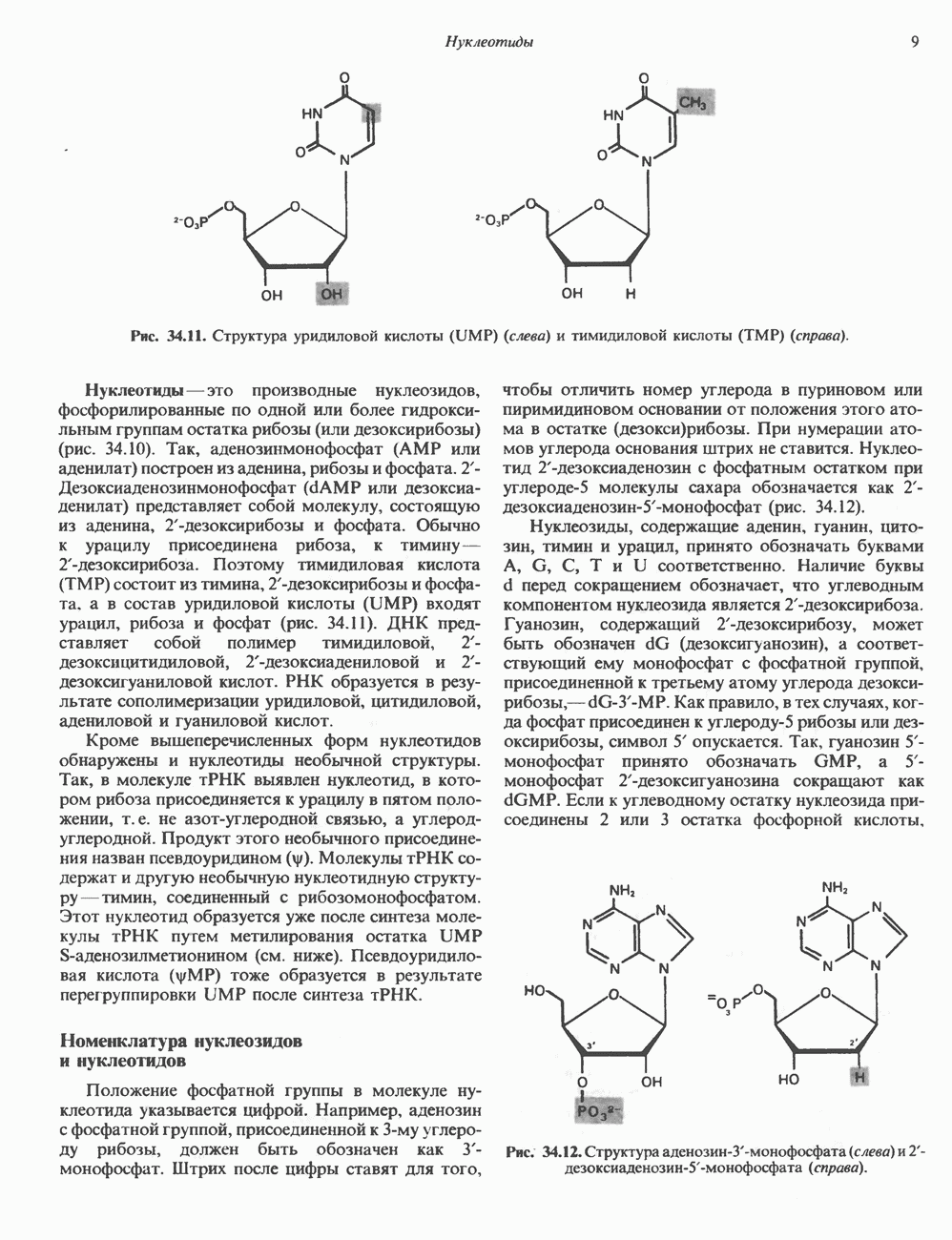
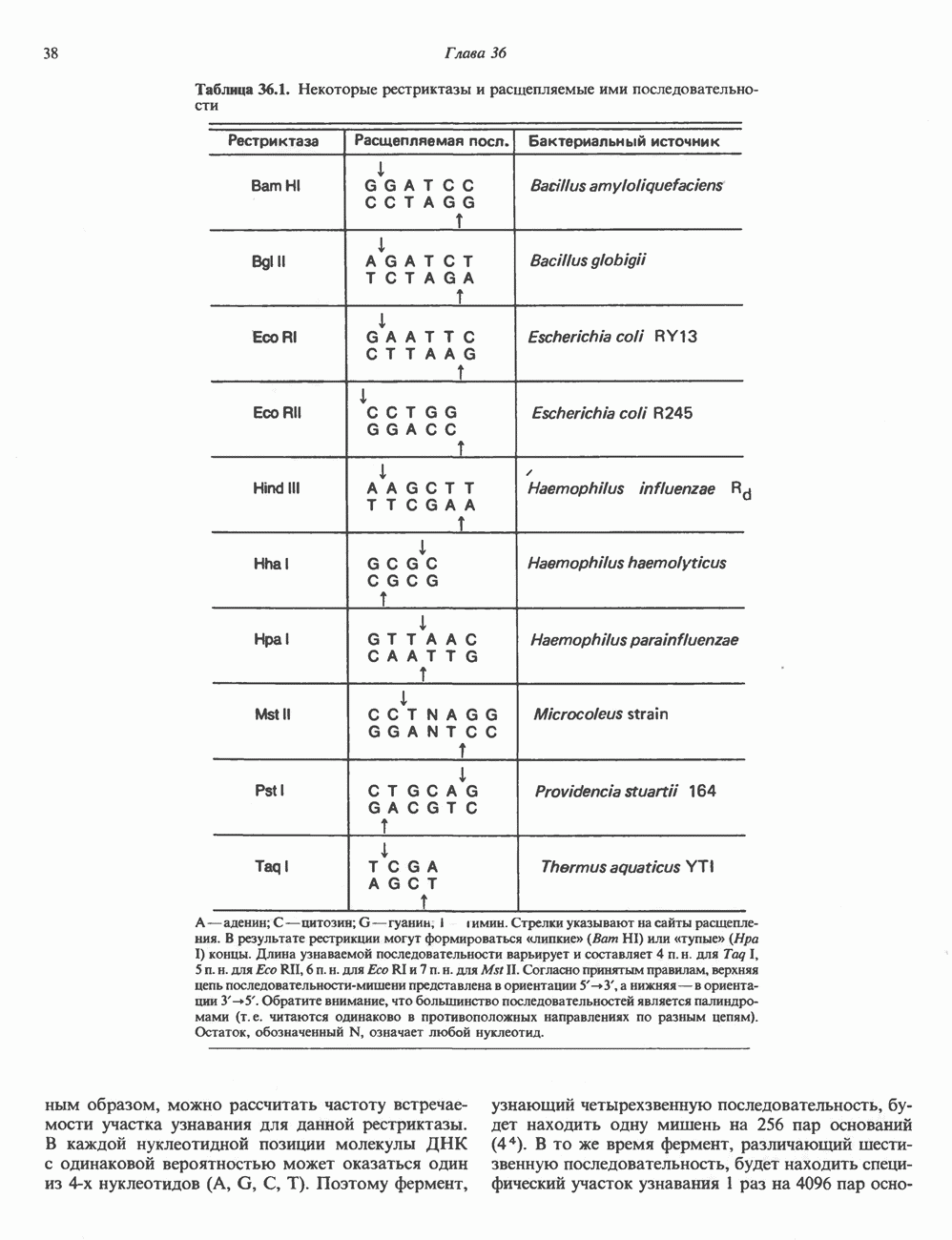
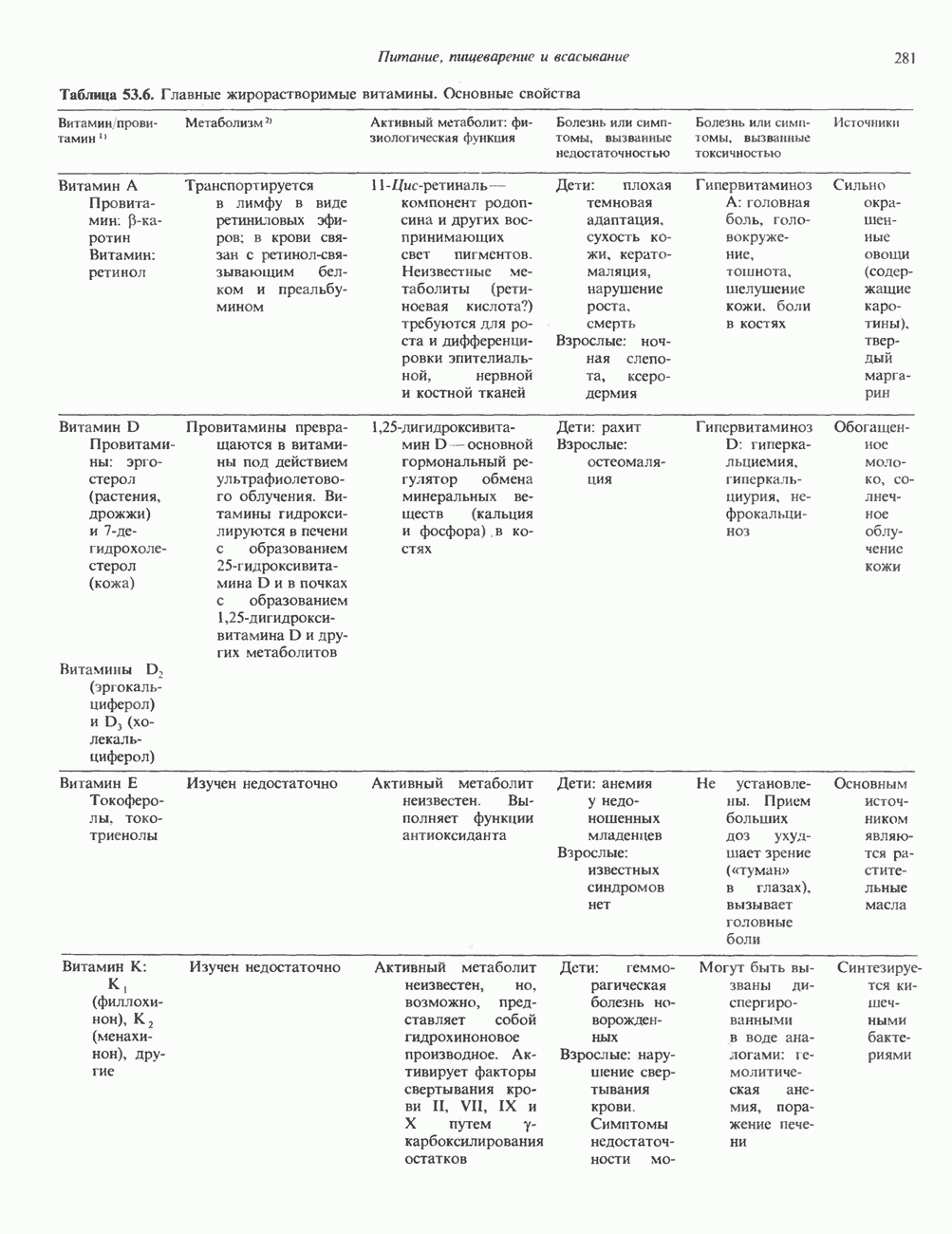
Таблица 49.2. Эффекты, опосредуемые различными адренергическими рецепторами

Механизм действия
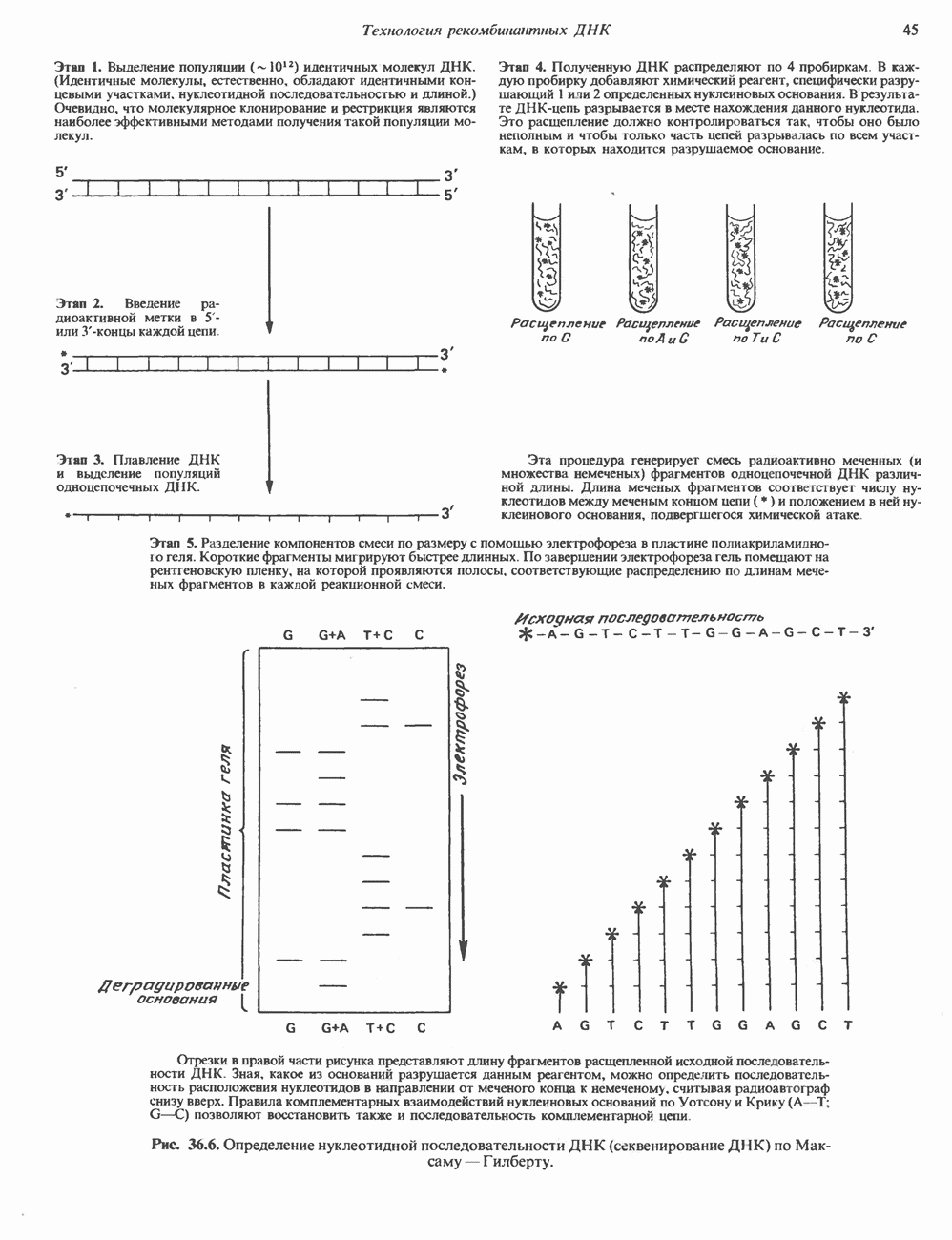
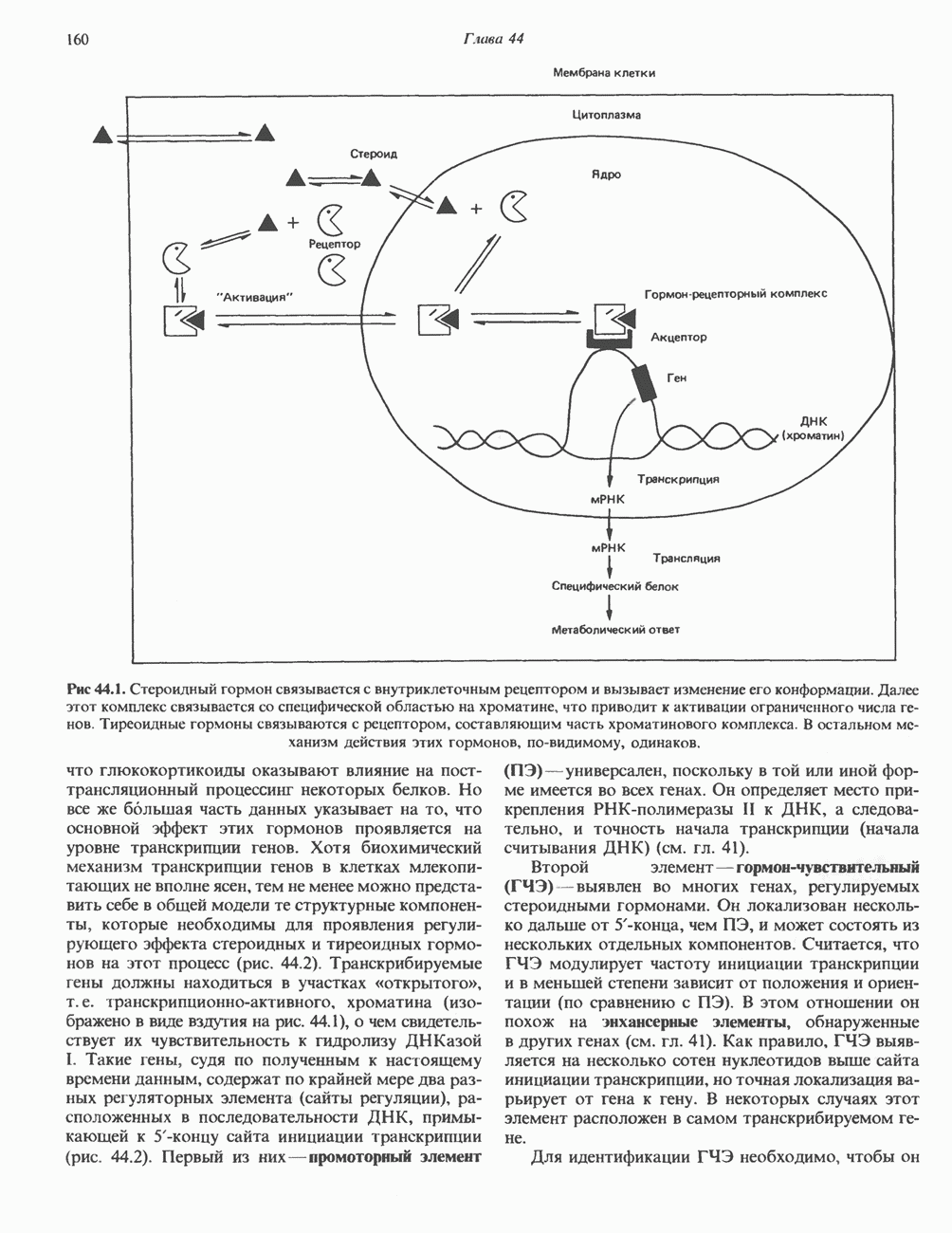
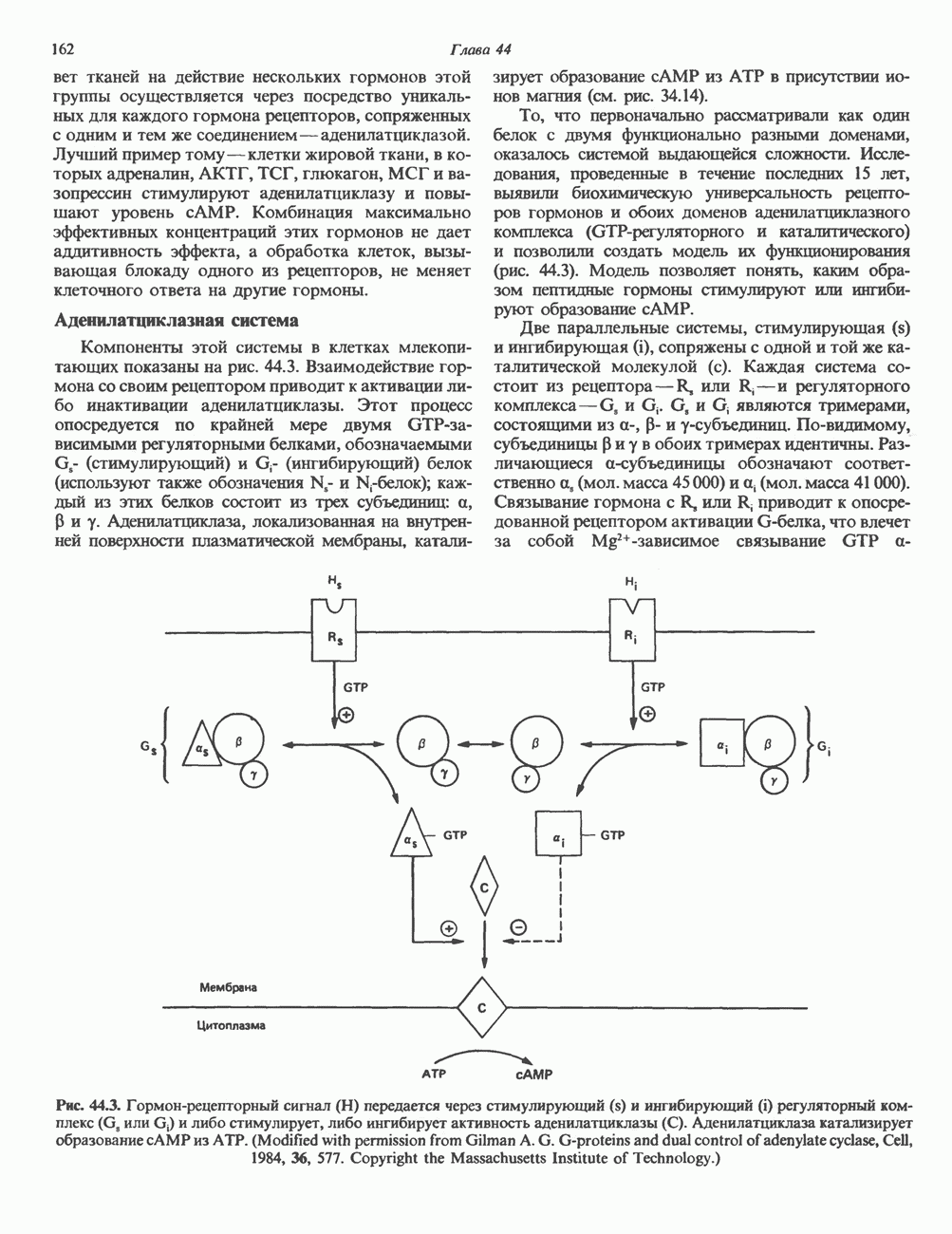
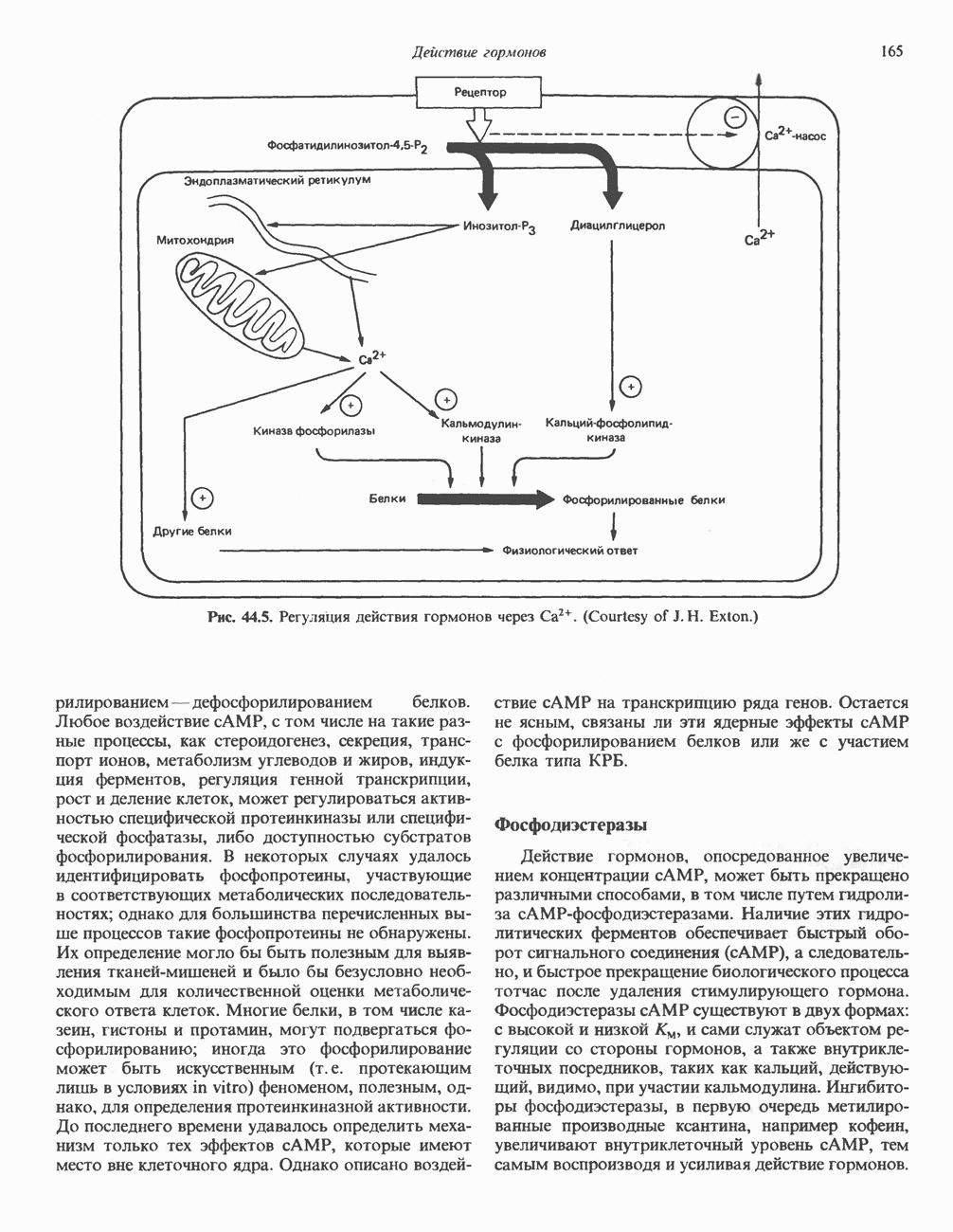
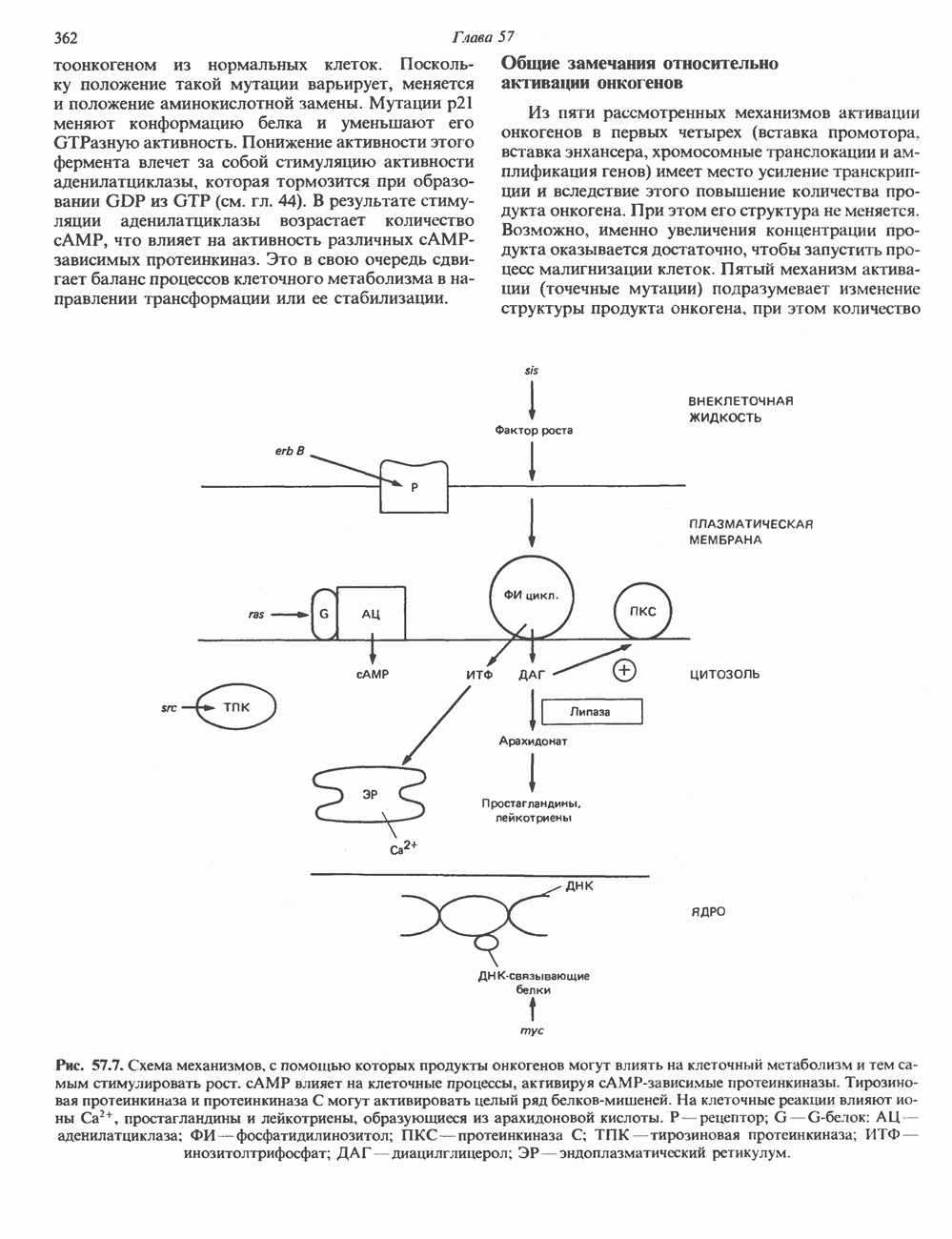
Рецепторы трех из этих подгрупп сопряжены с аденилатциклазной системой. Гормоны, связывающиеся с р,- и Р2-рецепторами, активируют аденилат-циклазу, тогда как гормоны, ассоциированные с а2-рецепторами, ингибируют ее (см. рис. 44.3 и табл. 44.3). Связывание катехоламинов индуцирует конденсирование рецептора с G-белком, связывающим загем GTP. Это либо стимулирует (Gs), либо ингибирует (GJ аденилатциклазу, что в результате приводит к усилению или подавлению синтеза с AM Р. Реакция выключается, когда GTPa3a, связанная с а-субъединицей G-белка, гидролизует GTP (см. рис. 44.2). а,-Рецепторы участвуют в процессах, ведущих к изменению внутриклеточной концентрации кальция или к изменению метаболизма фосфатидилинозитида (либо к тому и другому). Не исключено, что для этой реакции необходим особый G-белковый комплекс.
Между рецептором катехоламинов и системой зрительной реакции существует функциональное сходство. При световой стимуляции происходит сопряжение родопсина с трансдуцином — G-белковым комплексом, а-субъединица которого также связывает GTP. Активированный G-белок в свою очередь стимулирует фосфодиэстеразу, гидролизующую cGMP. В результате ионные каналы в мембране клеток колбочек сетчатки закрываются и возникает зрительная реакция. Она выключается, когда ассоциированная с а-субъединицей GTPa3a гидролизует связанный GTP. Неполный перечень биохимических и физиологических эффектов, опосредованных различными адренергическими рецепторами, приведен в табл. 49.2.
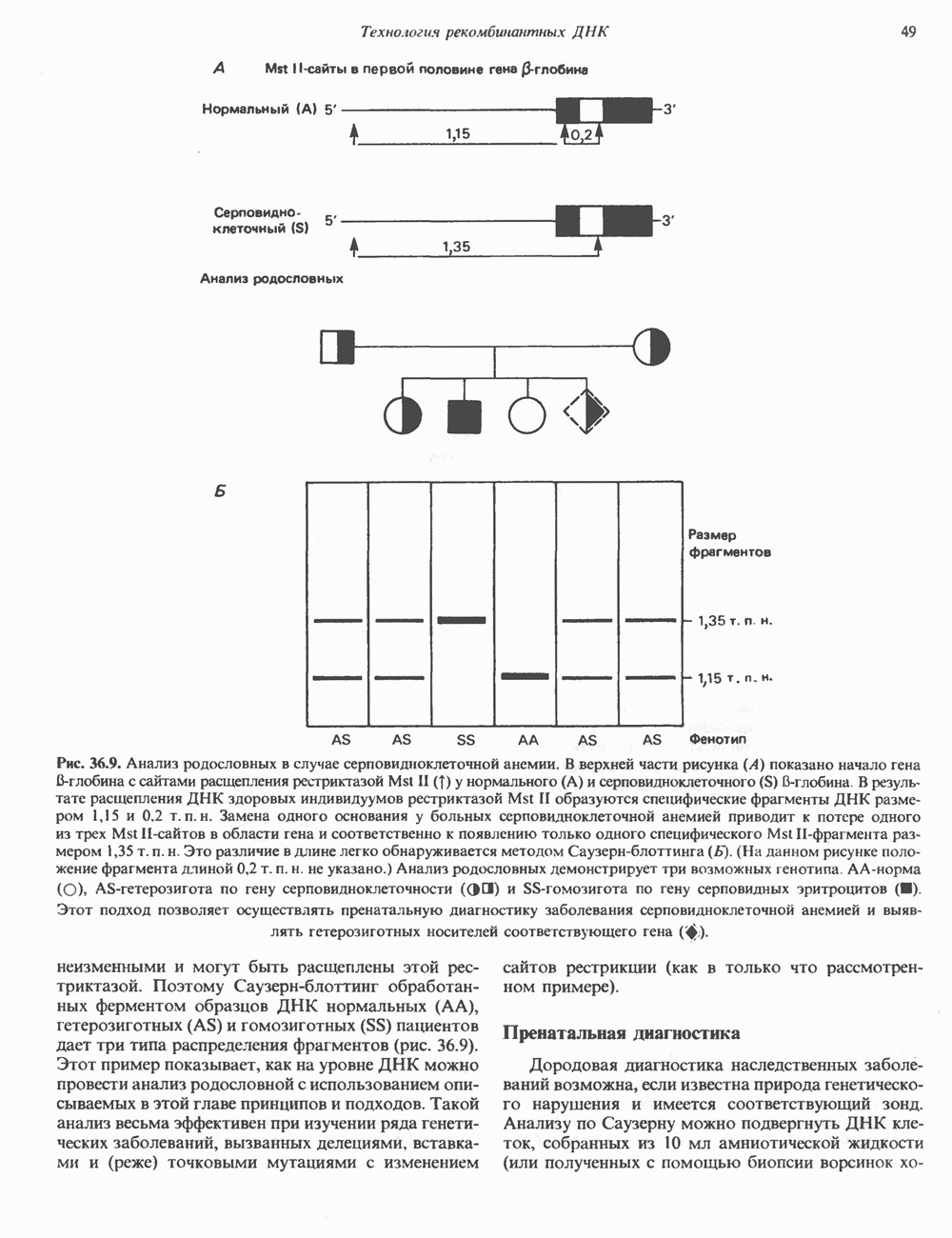
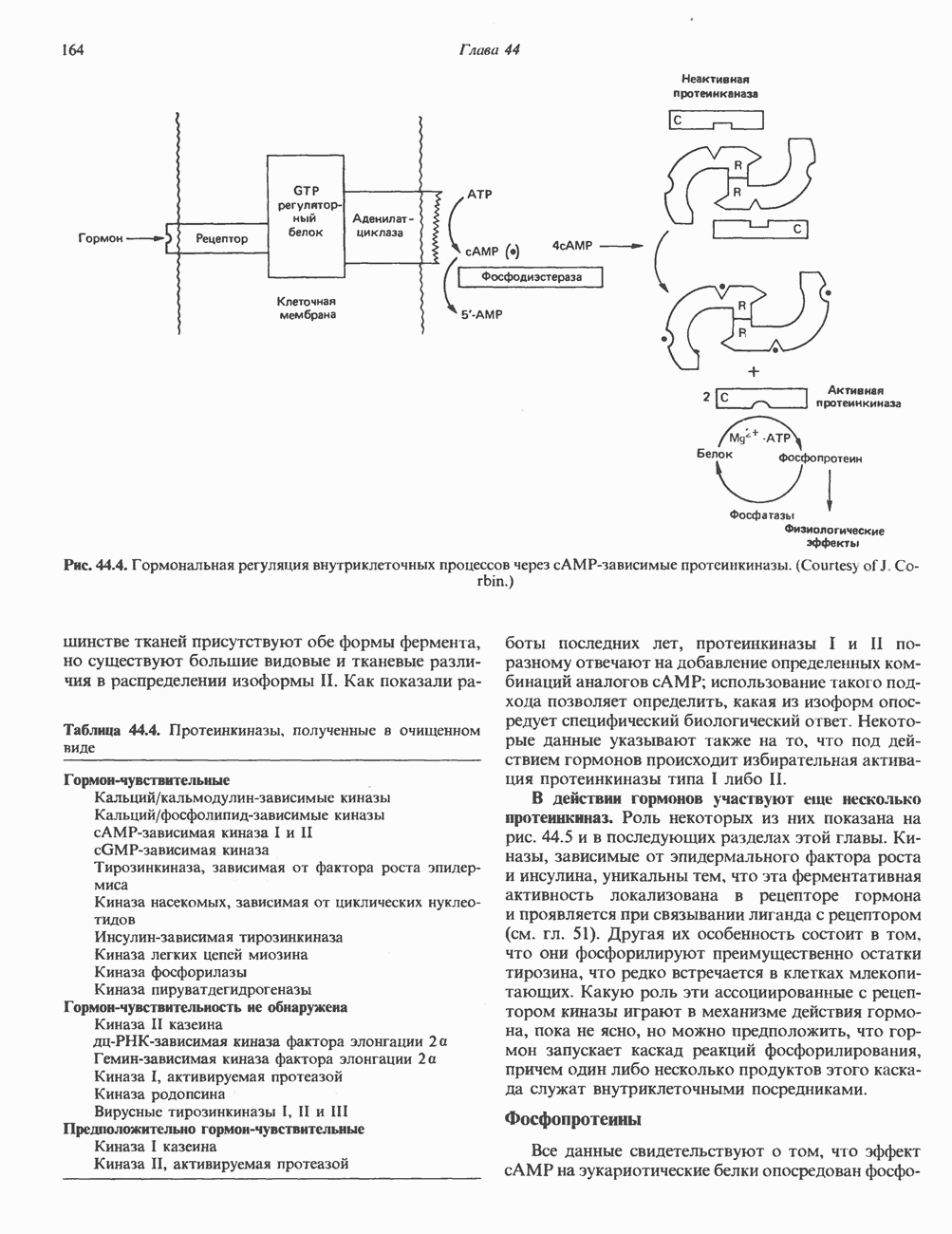
Активация фосфопротеинов сАМР-зависимой протеинкиназой (см. рис. 44.4) обусловливает многие биохимические эффекты адреналина. В мышцах и в меньшей степени в печени адреналин стимулирует гликогенолиз путем активации протеинкиназы, которая в свою очередь активирует фосфорилазный каскад (см. рис. 19.7). Фосфорилирование гликоген-синтазы, напротив, ослабляет синтез гликогена. Действуя на сердце, адреналин увеличивает минутный объем в результате повышения силы (инотропный эффект) и частоты (хронотропный эффект) сокращений, что также связано с увеличением содержания сАМР. В жировой ткани адреналин повышает содержание сАМР, под действием которого чувствительная к гормонам липаза превращается в активную (фо-сфорилированную) форму. Этот фермент усиливает липолиз и высвобождение жирных кислот в кровь. Жирные кислоты используются в качестве источника энергии в мышцах и, кроме того, могут активировать глюконеогенез в печени.