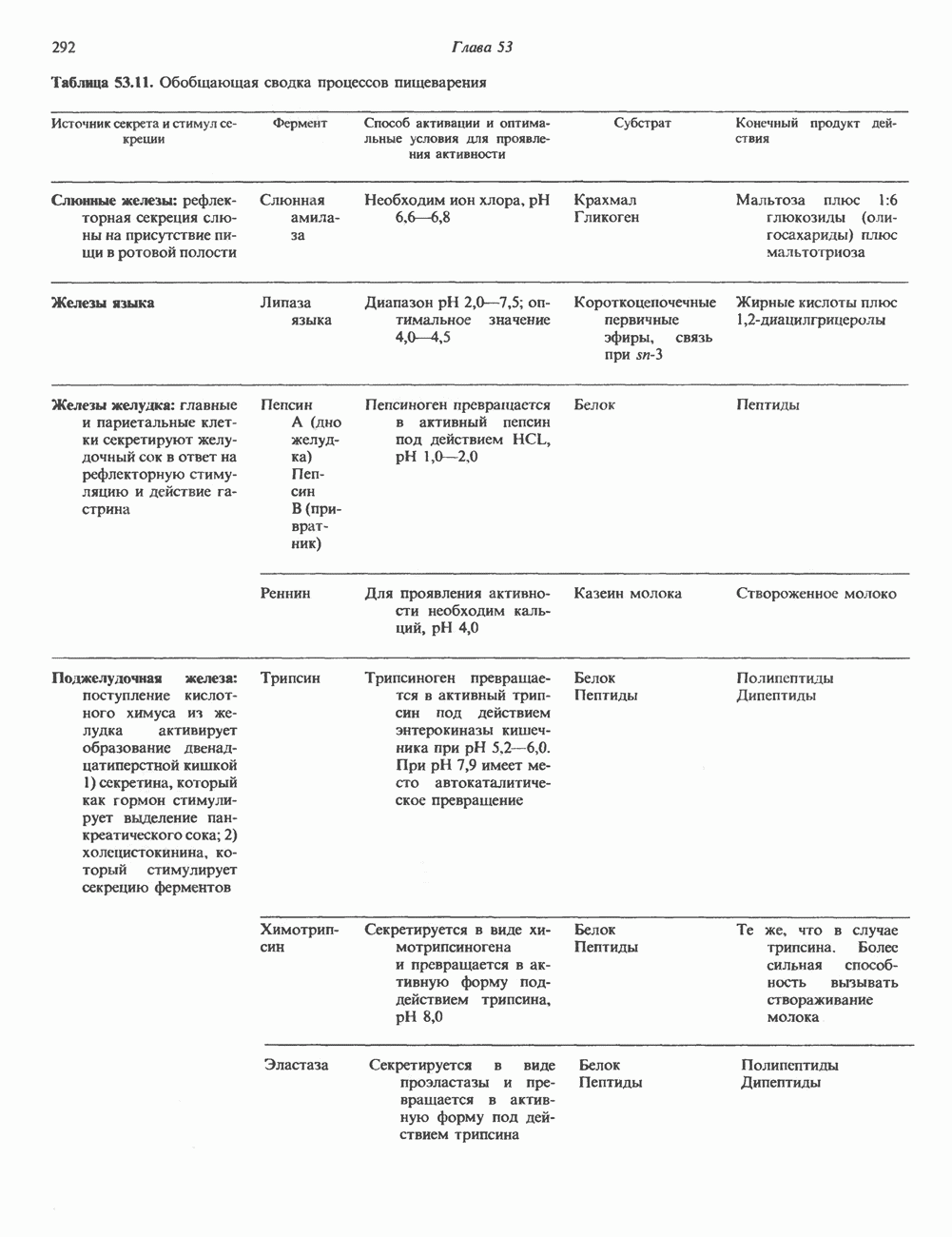
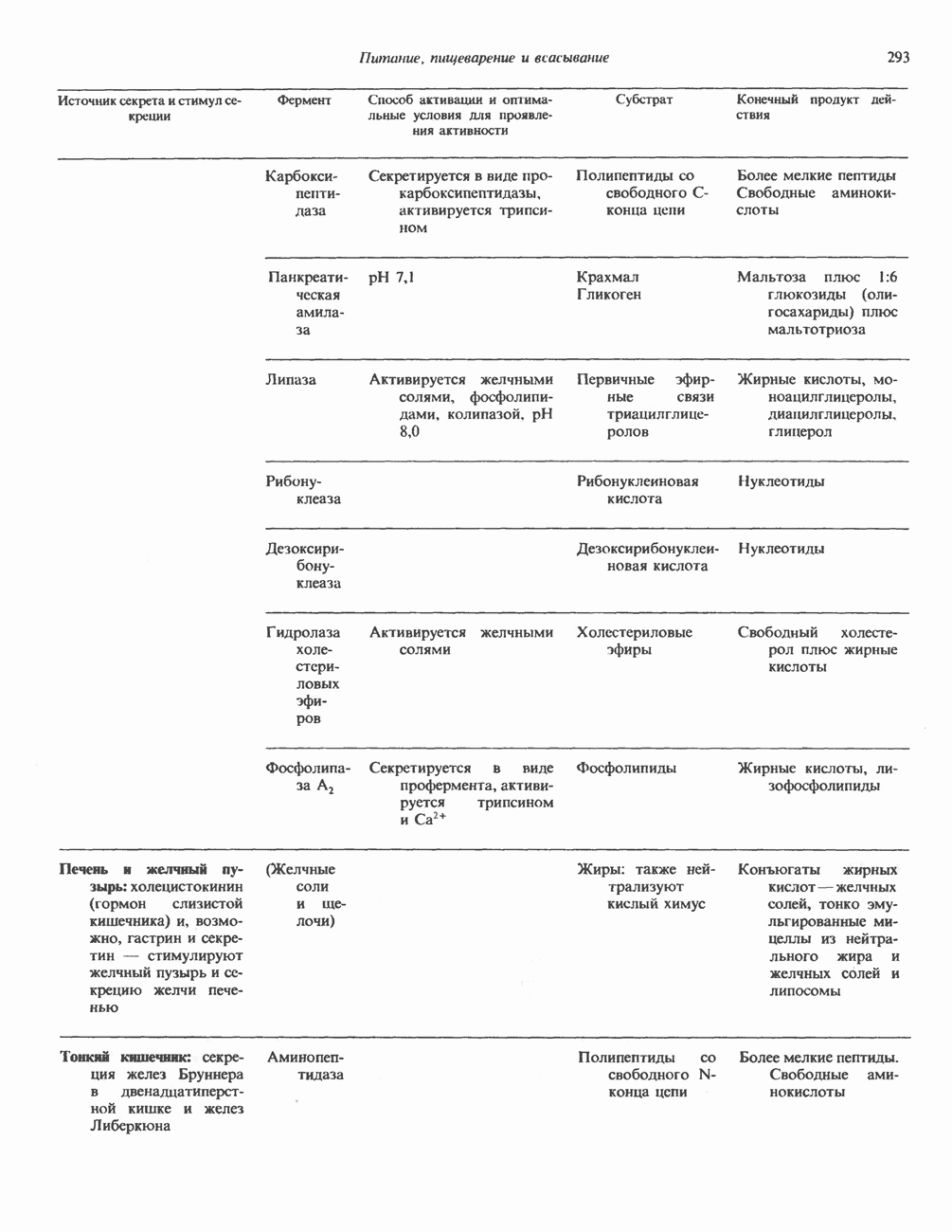
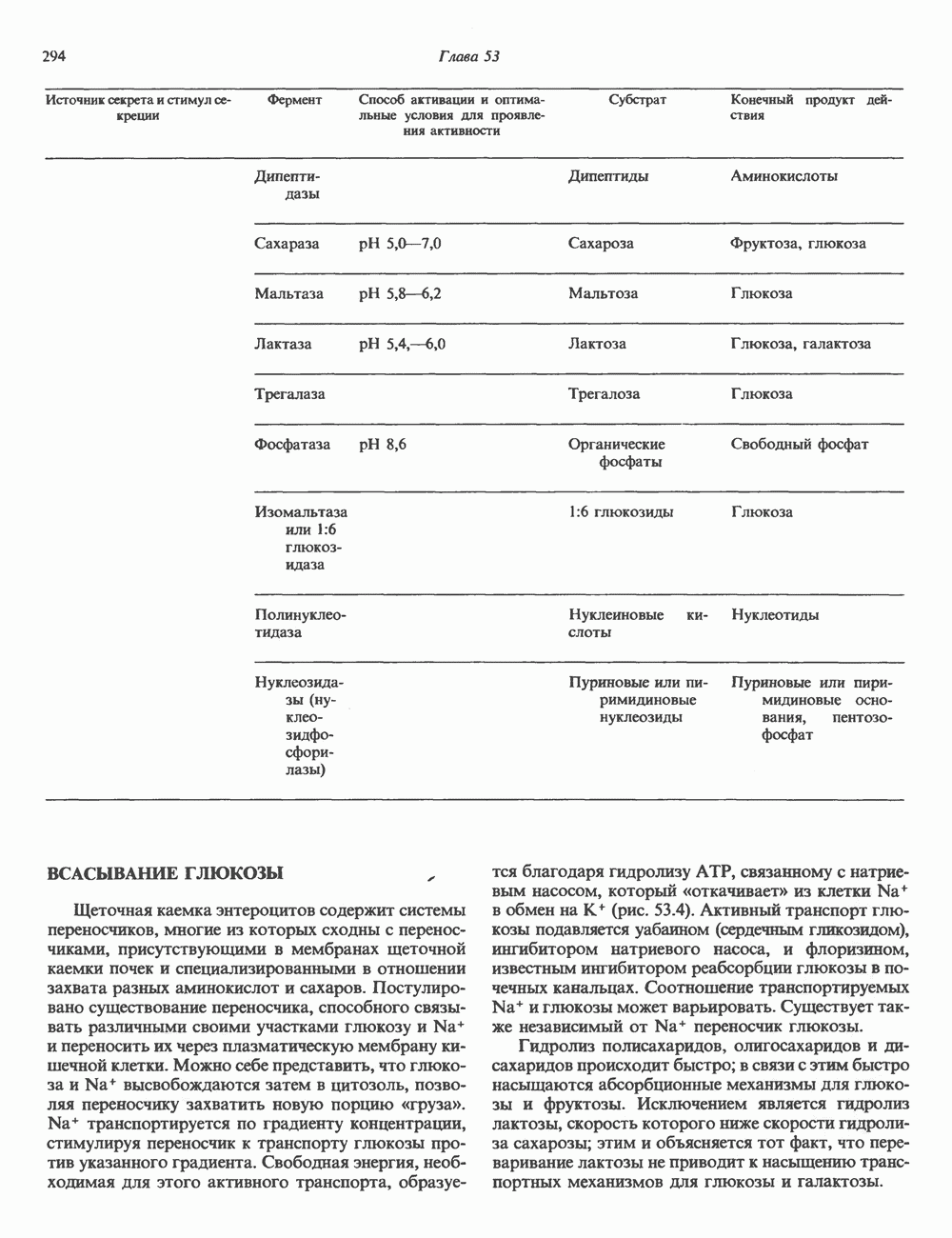
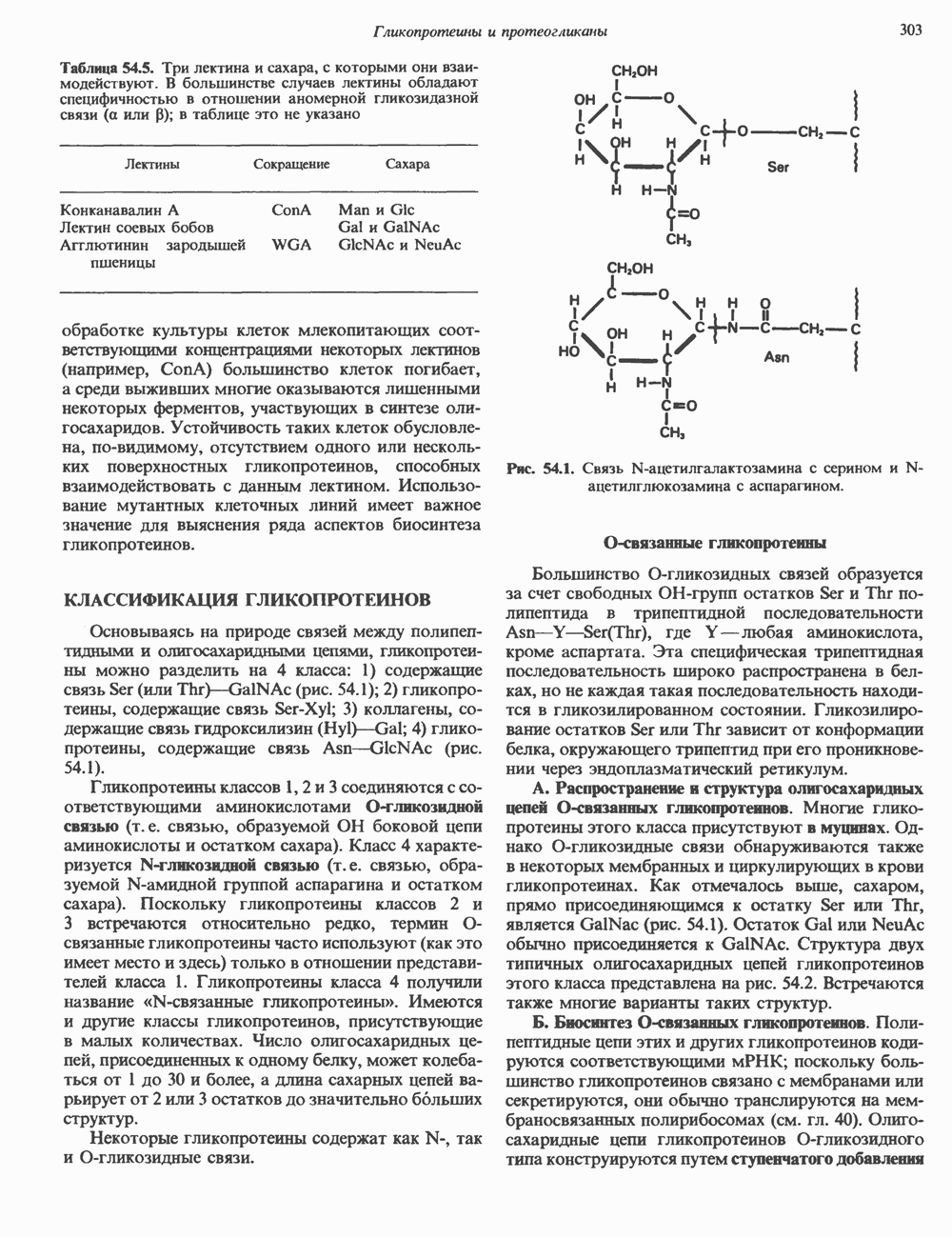
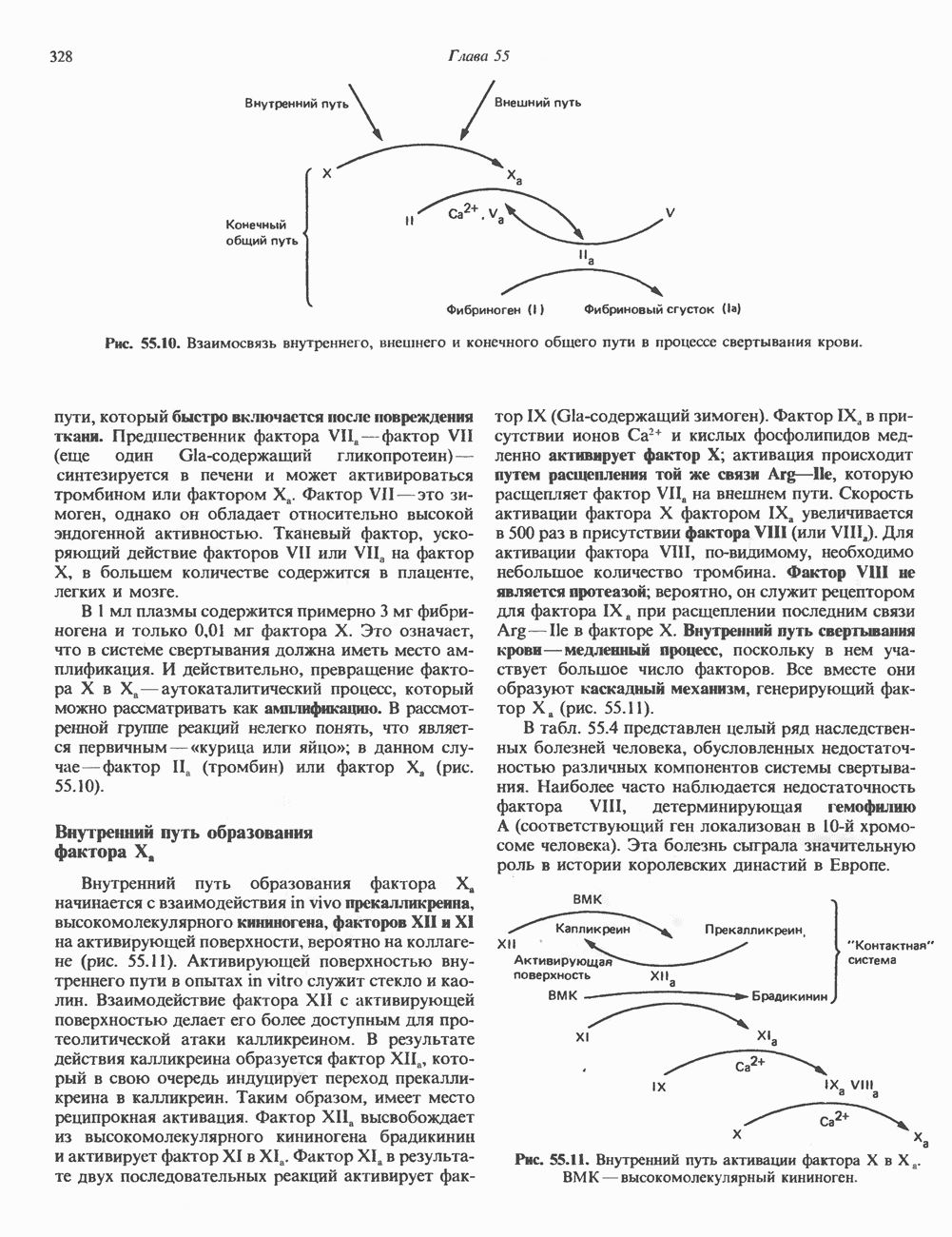
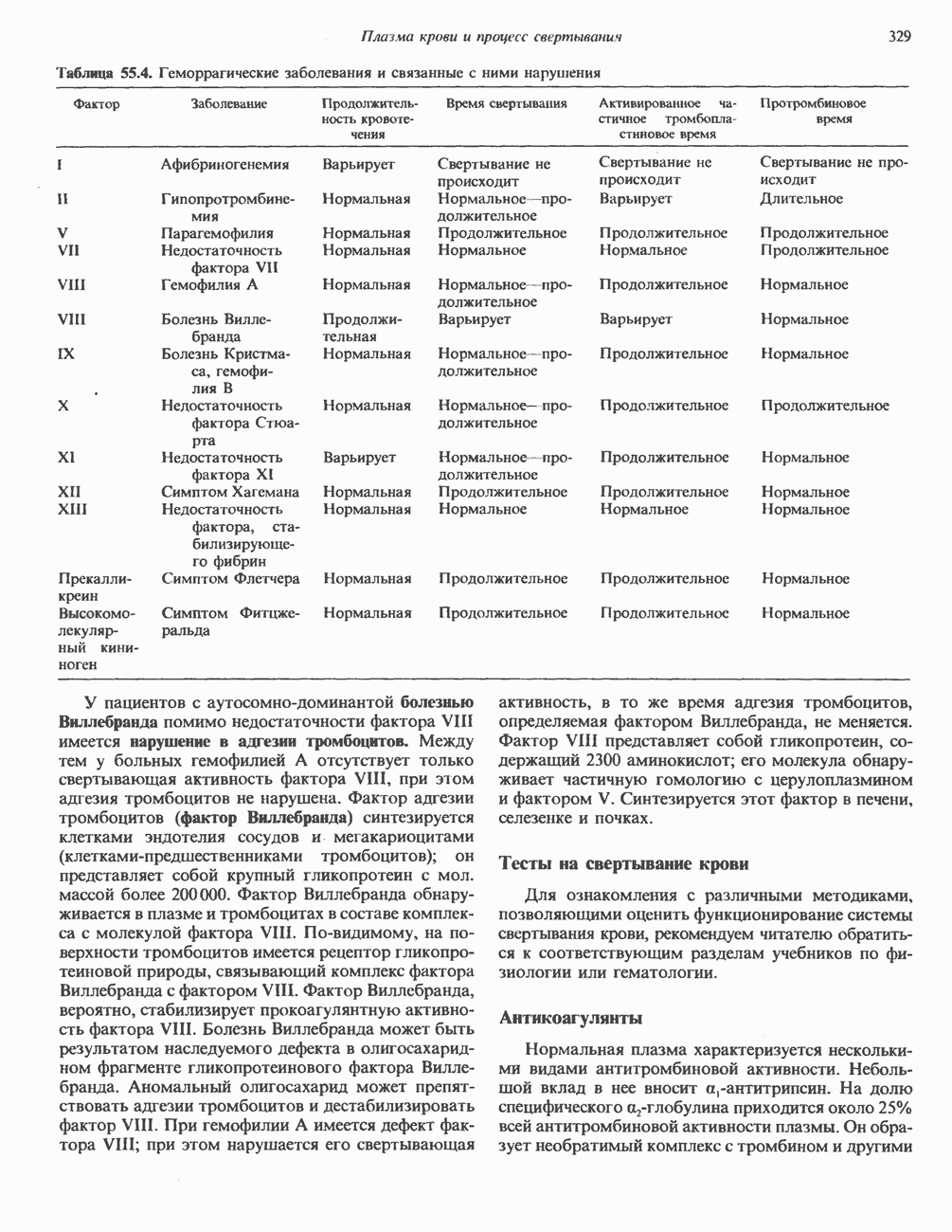
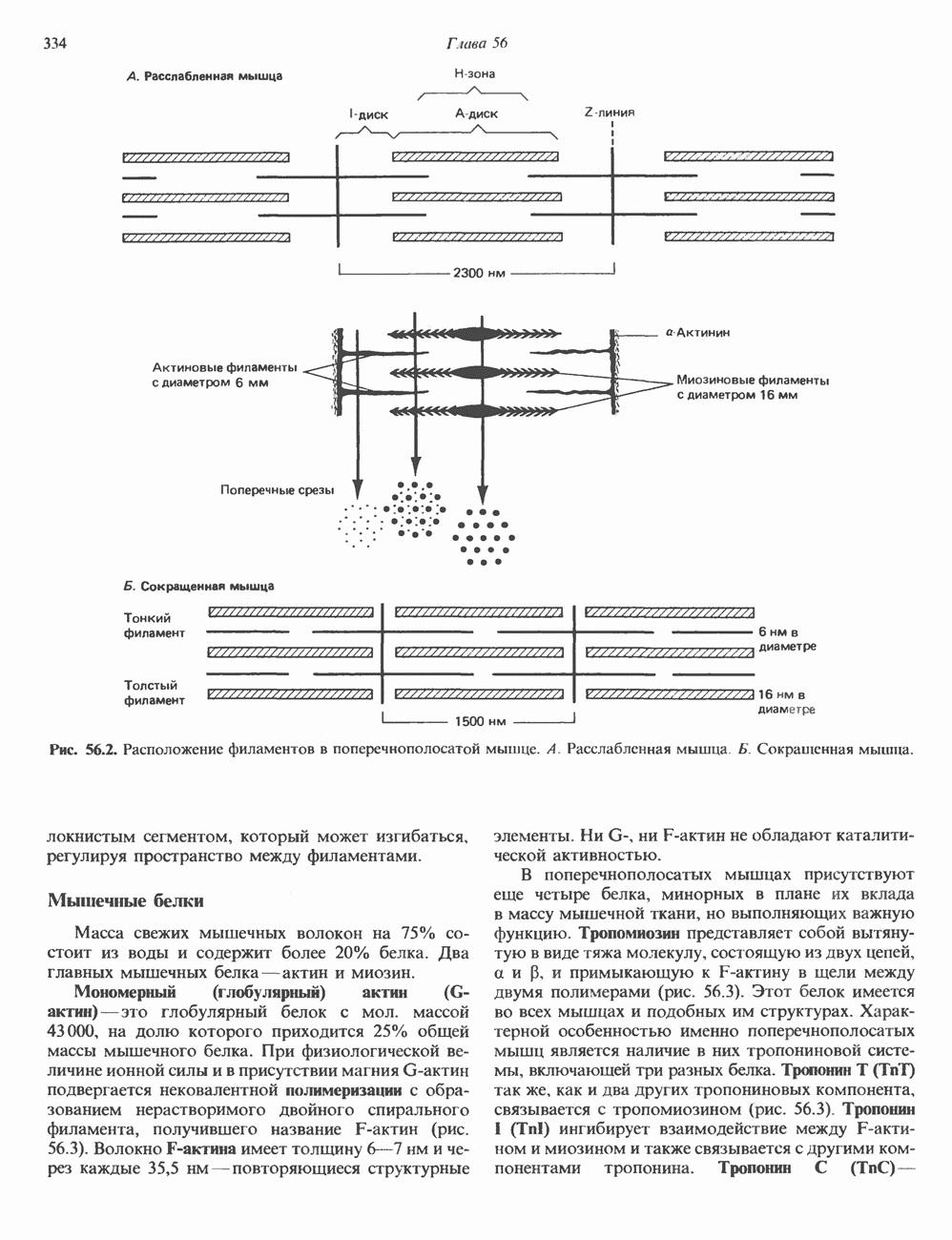
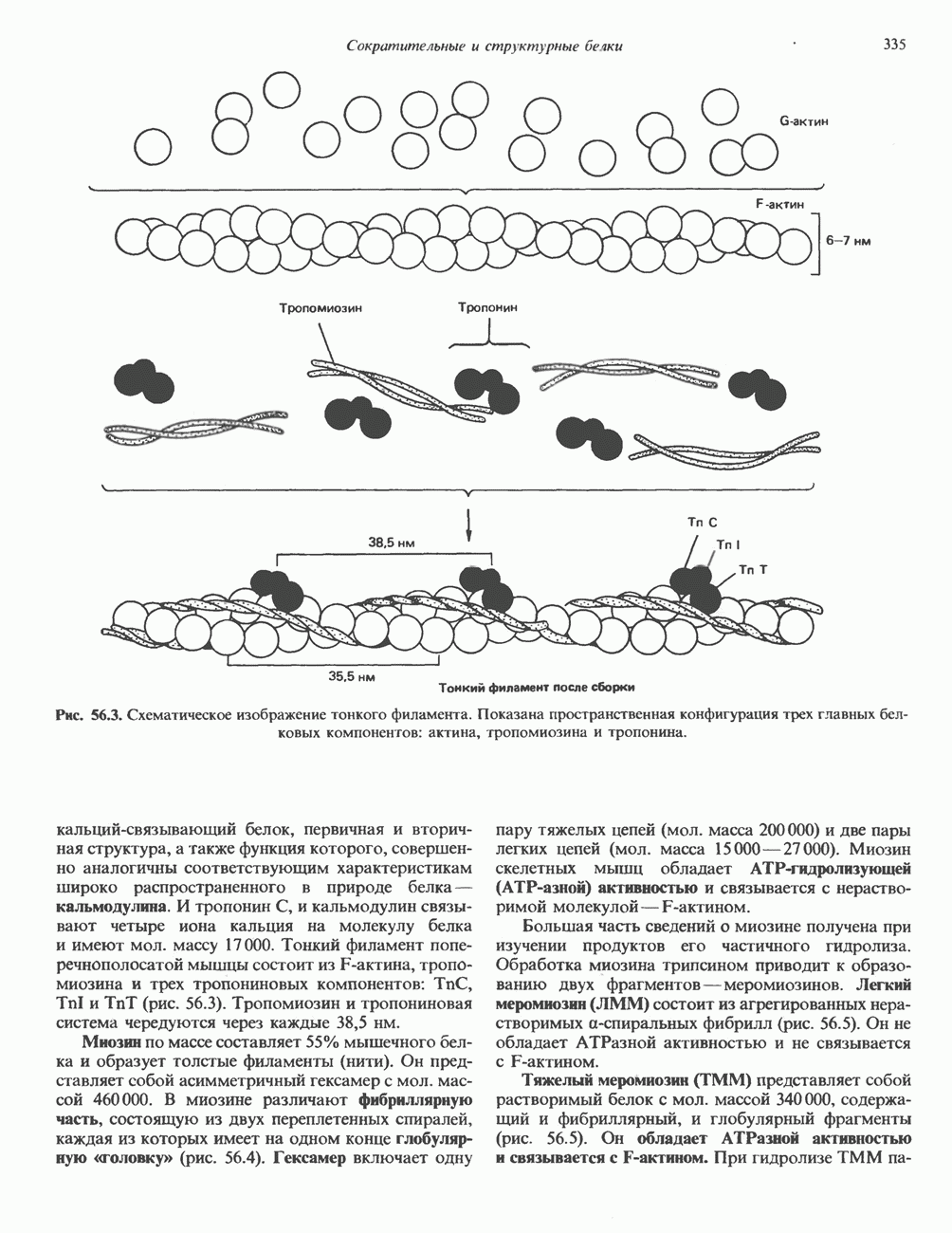
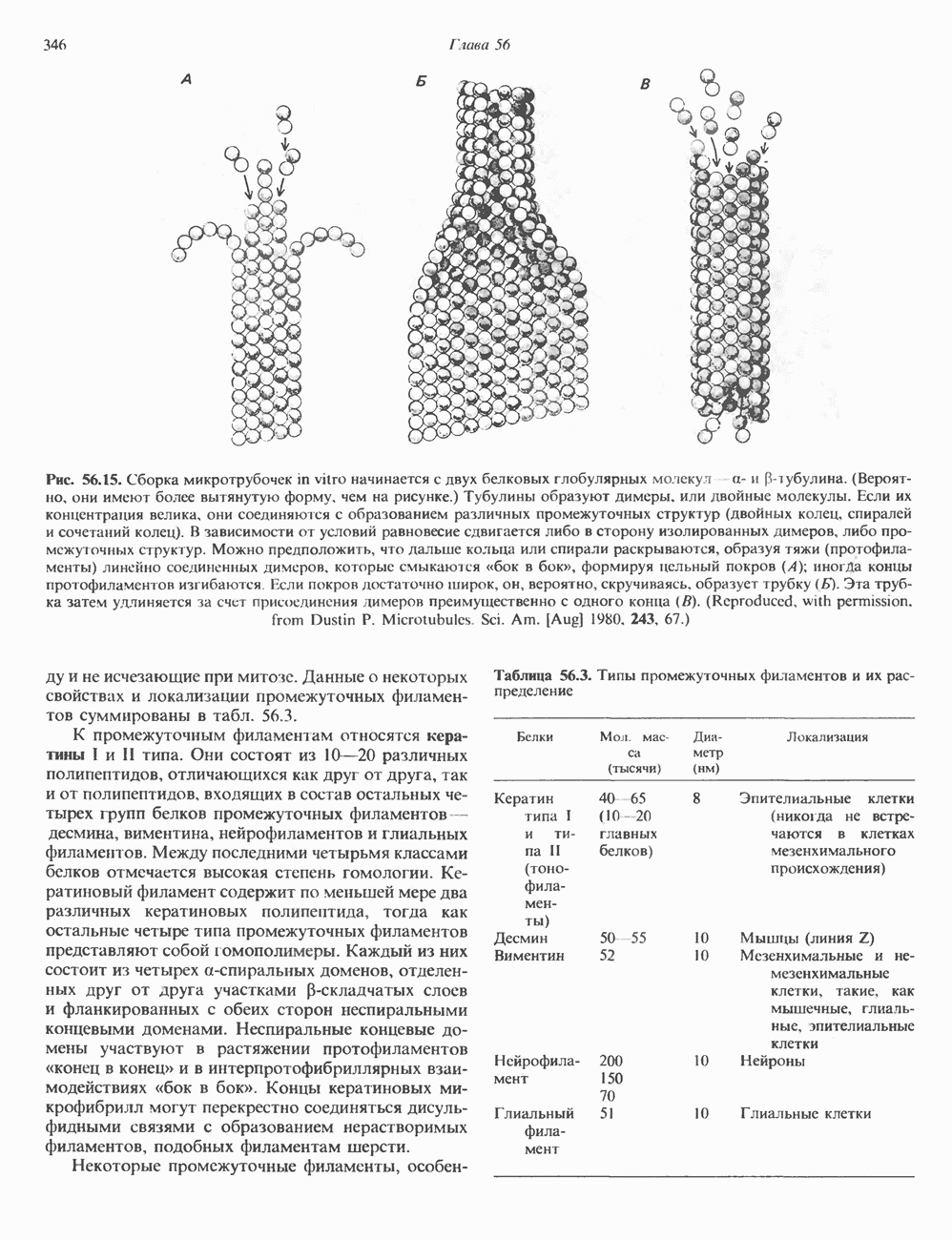
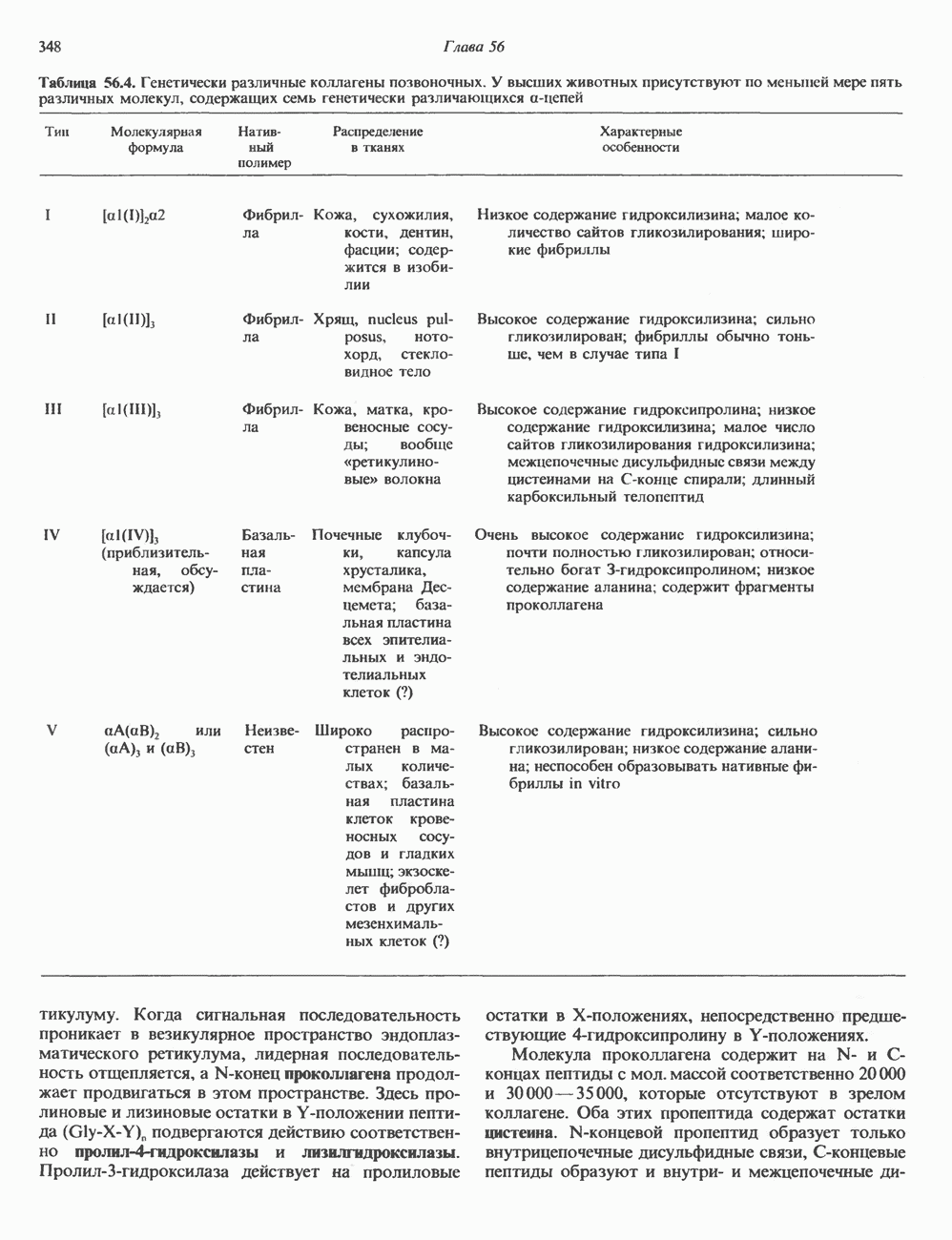
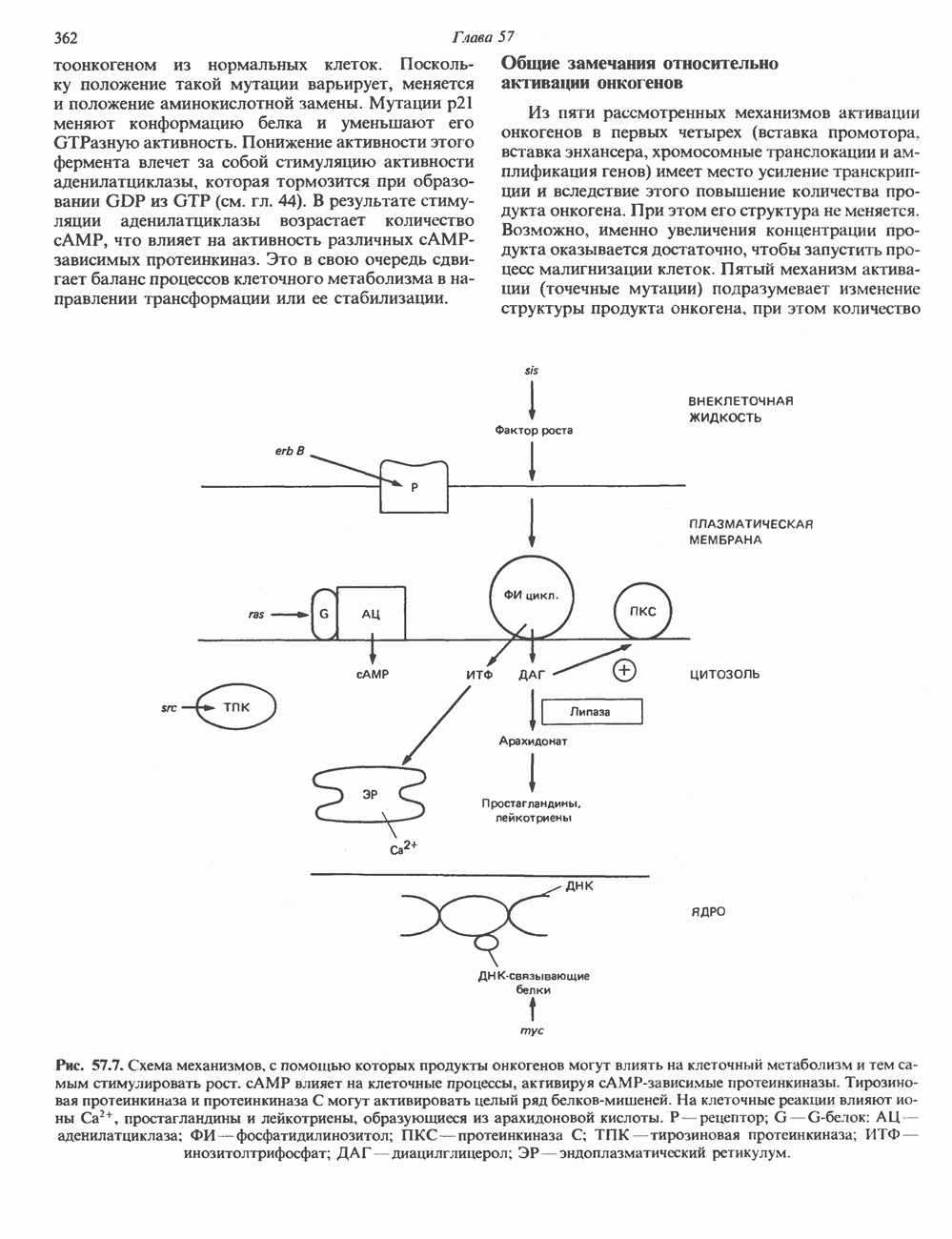
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
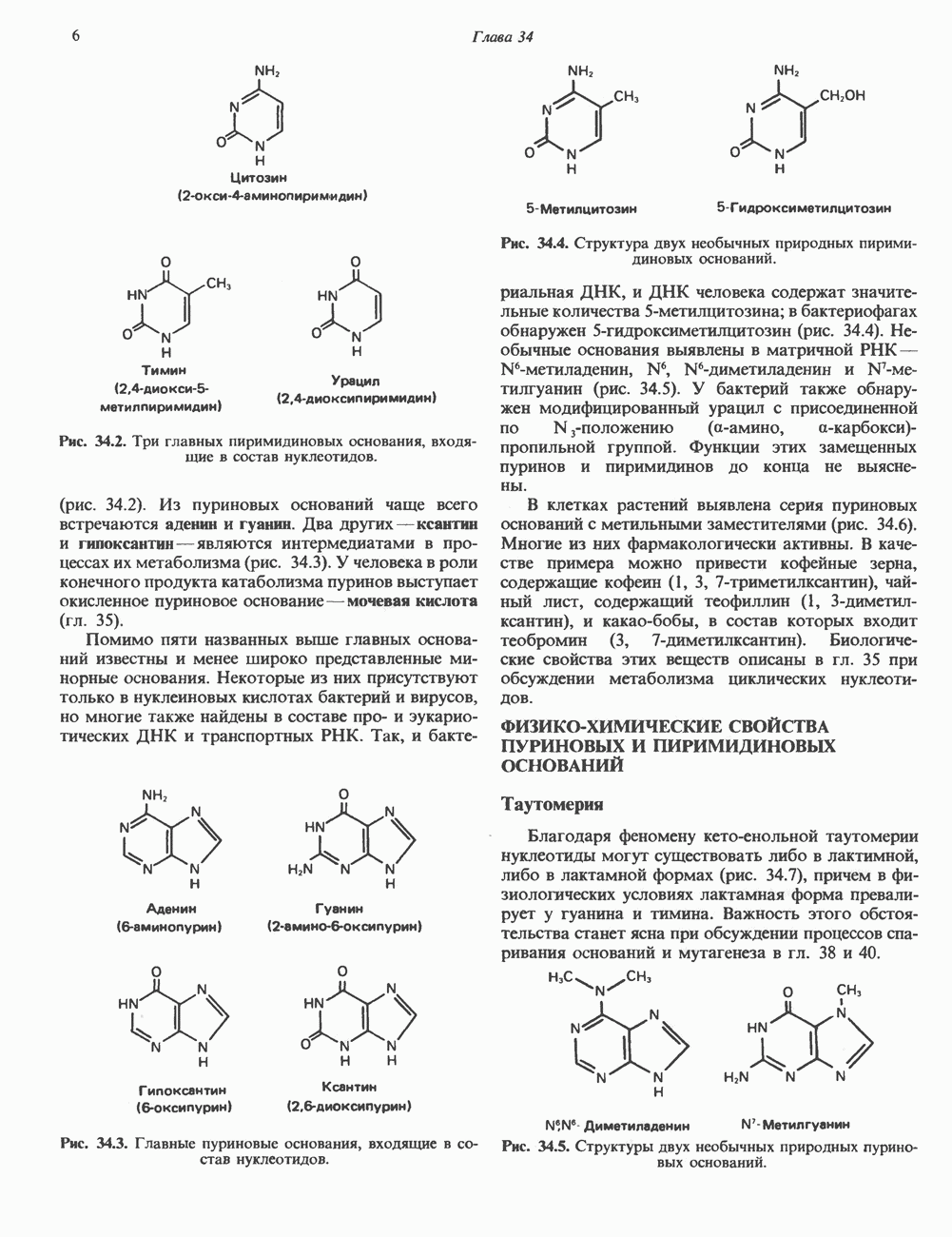
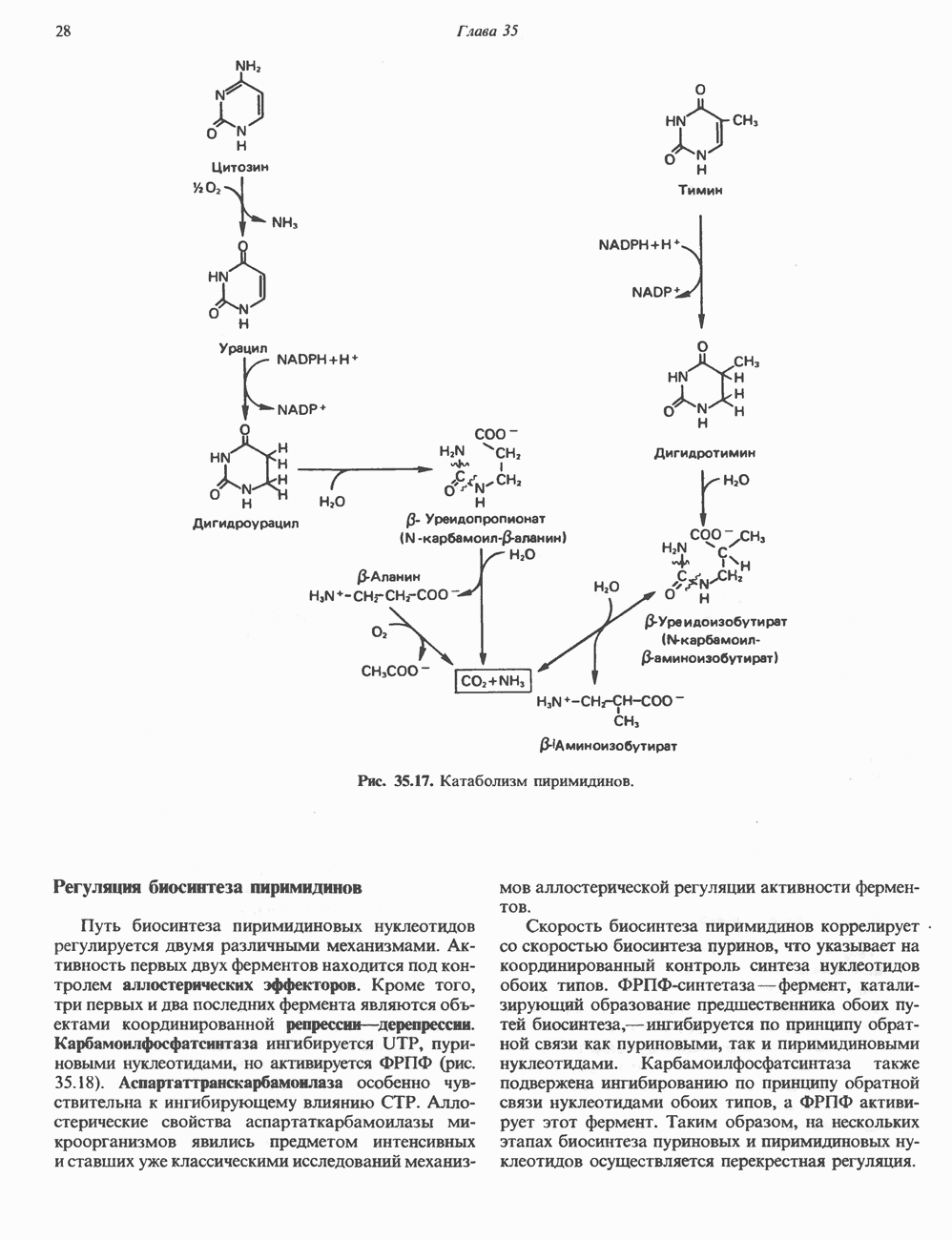
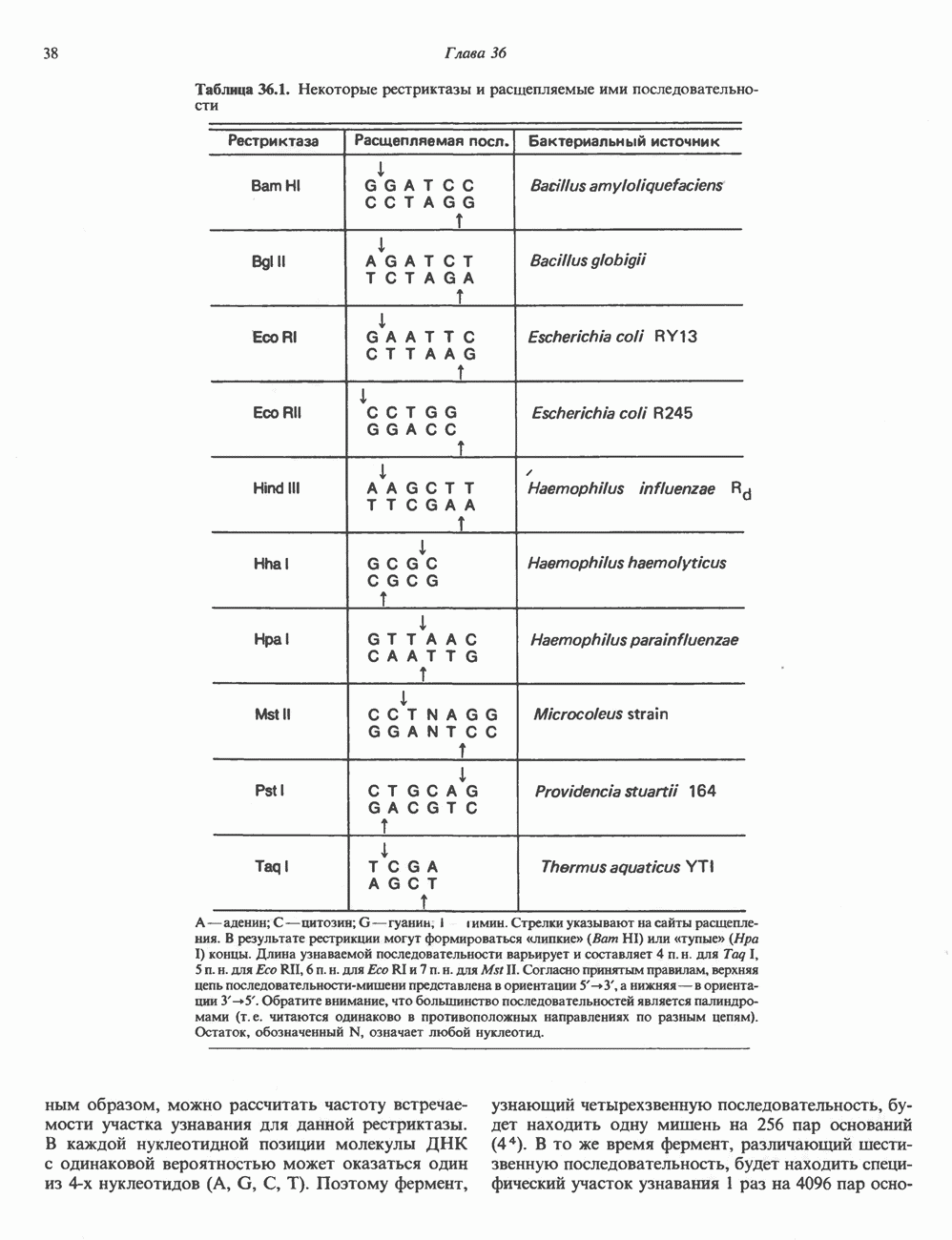
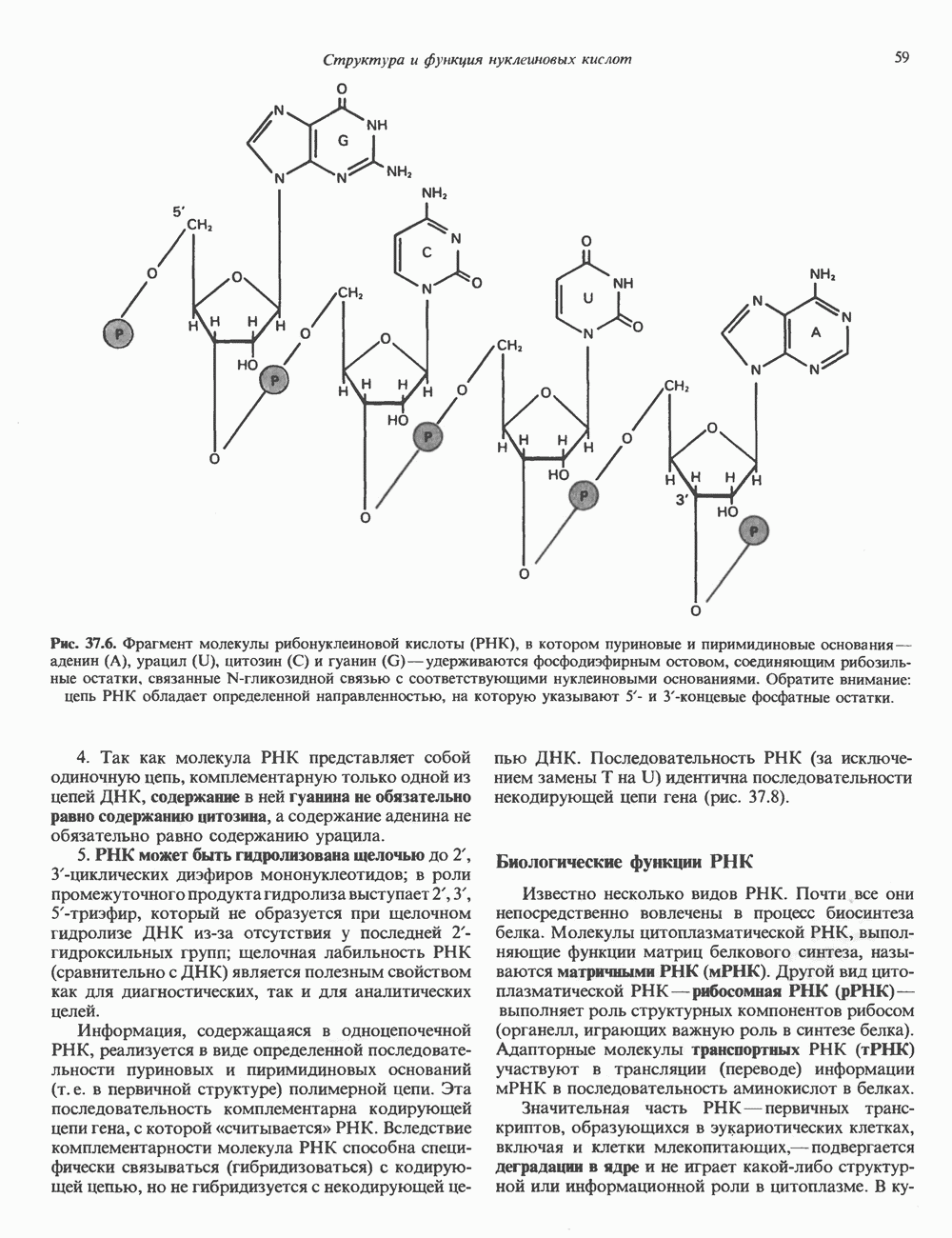
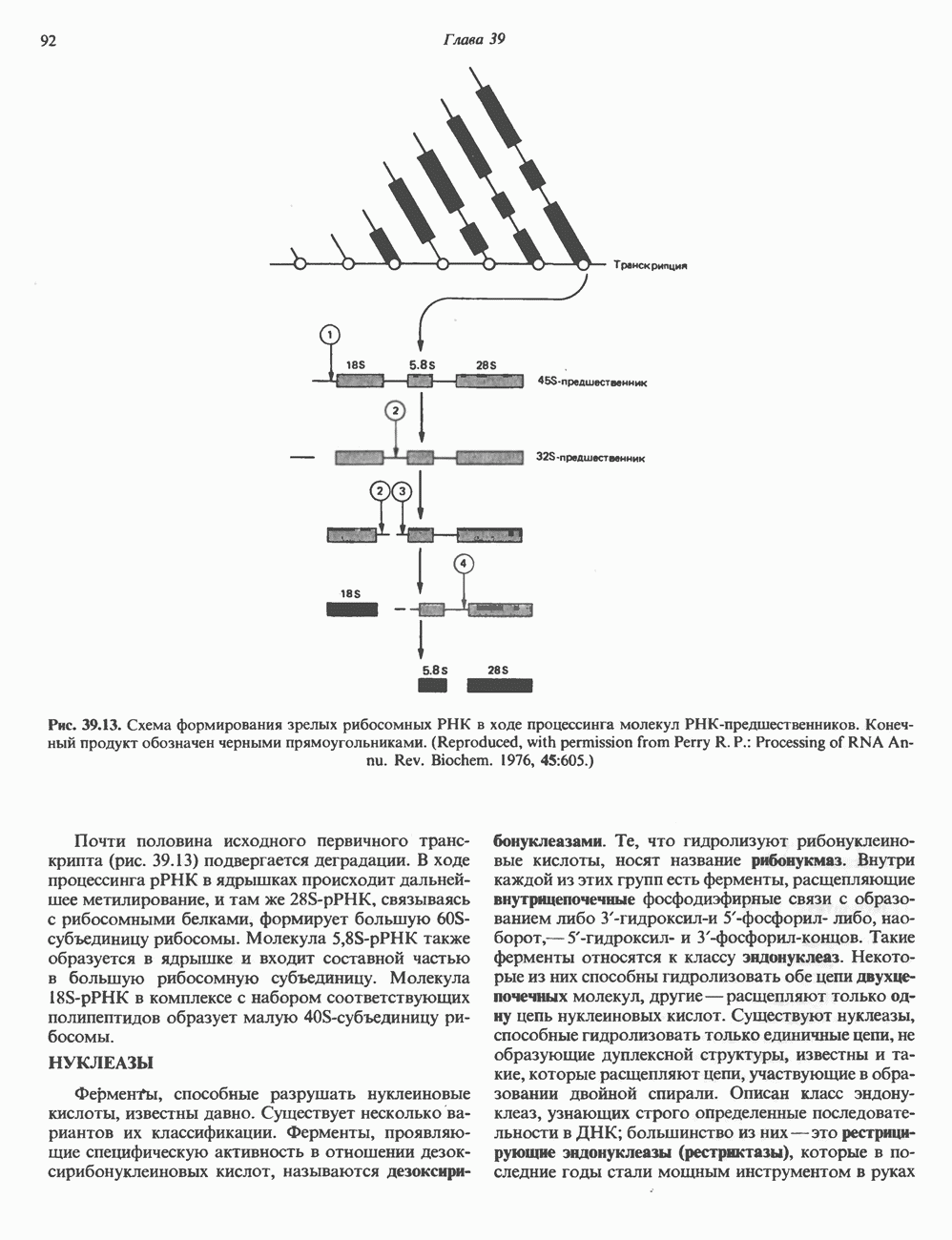
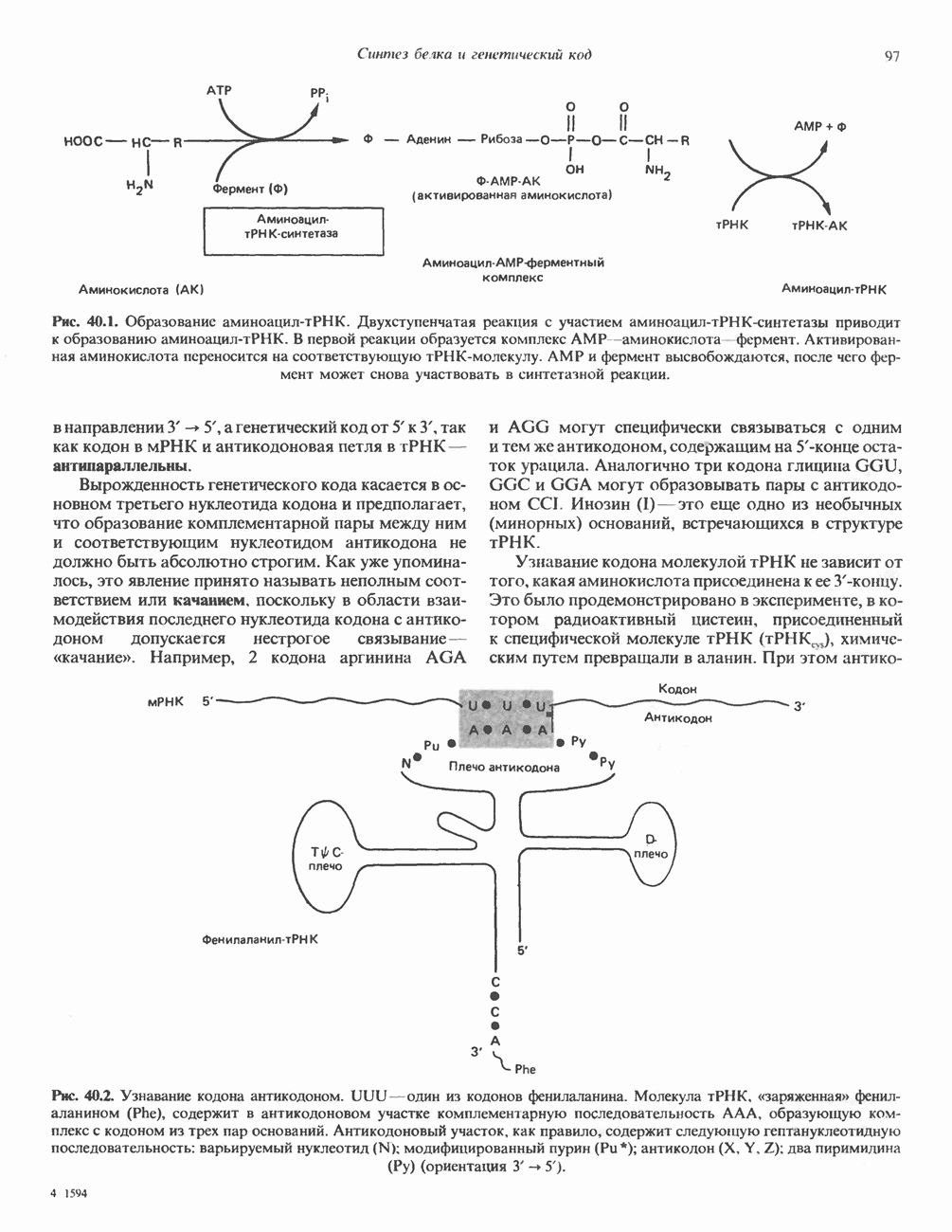
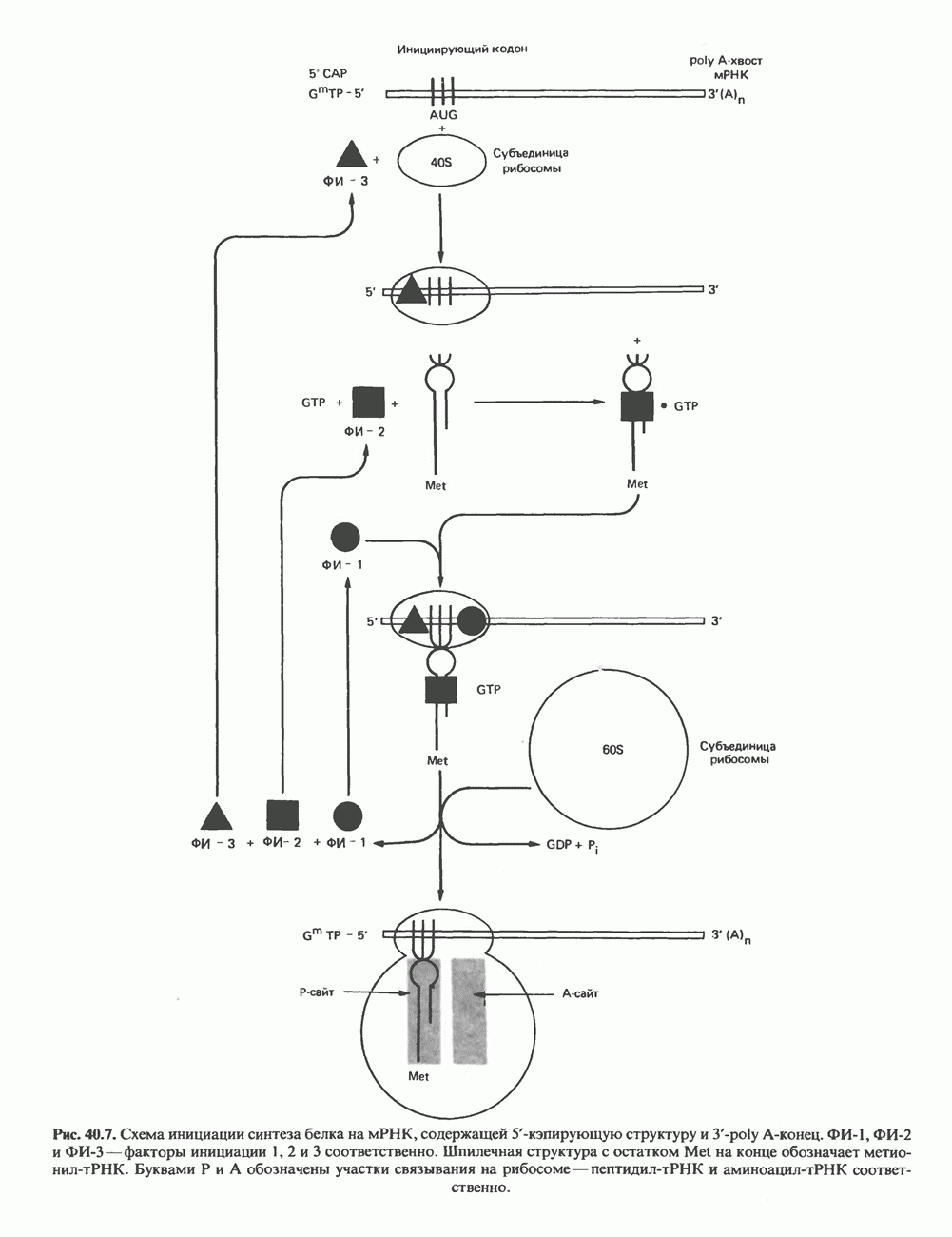
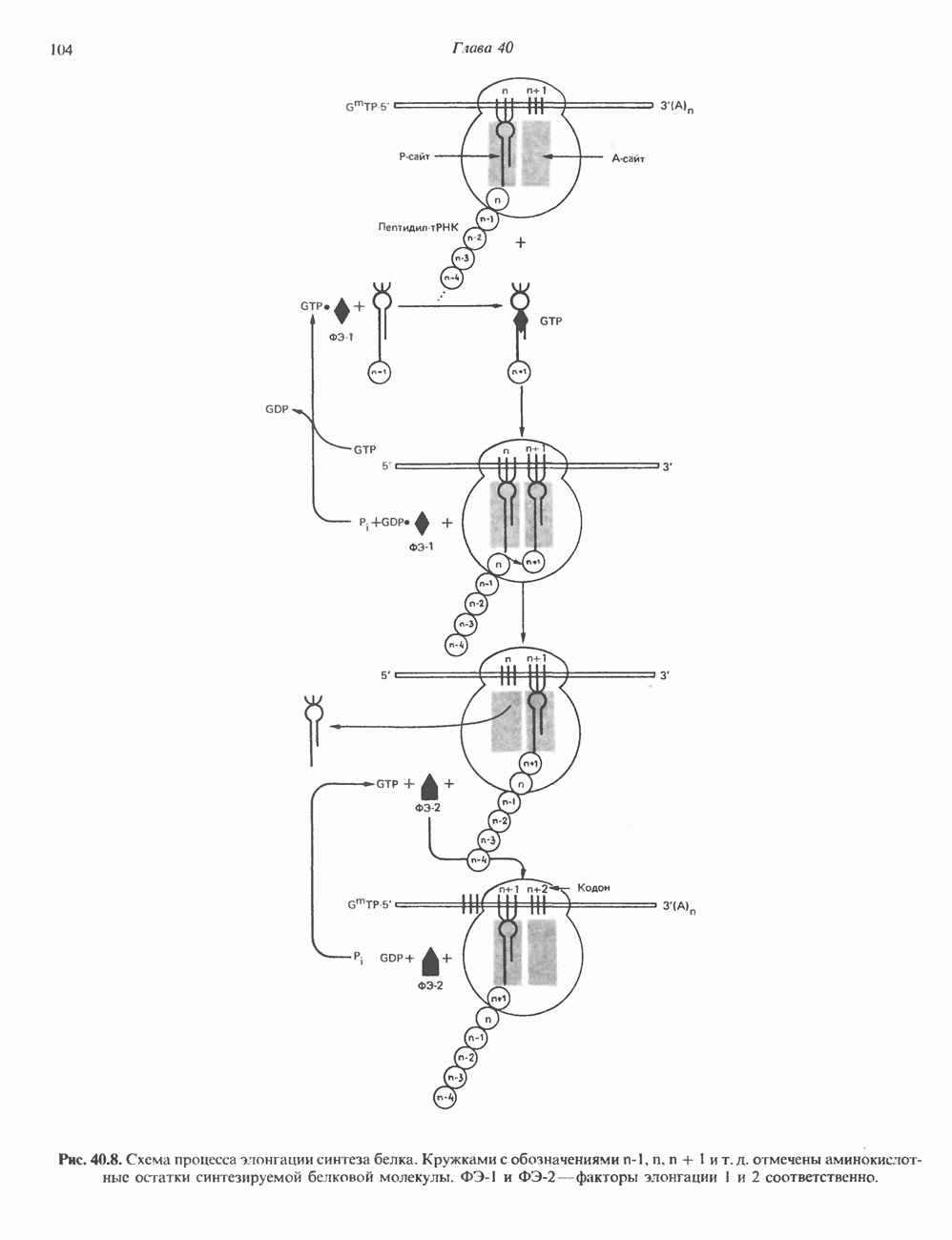
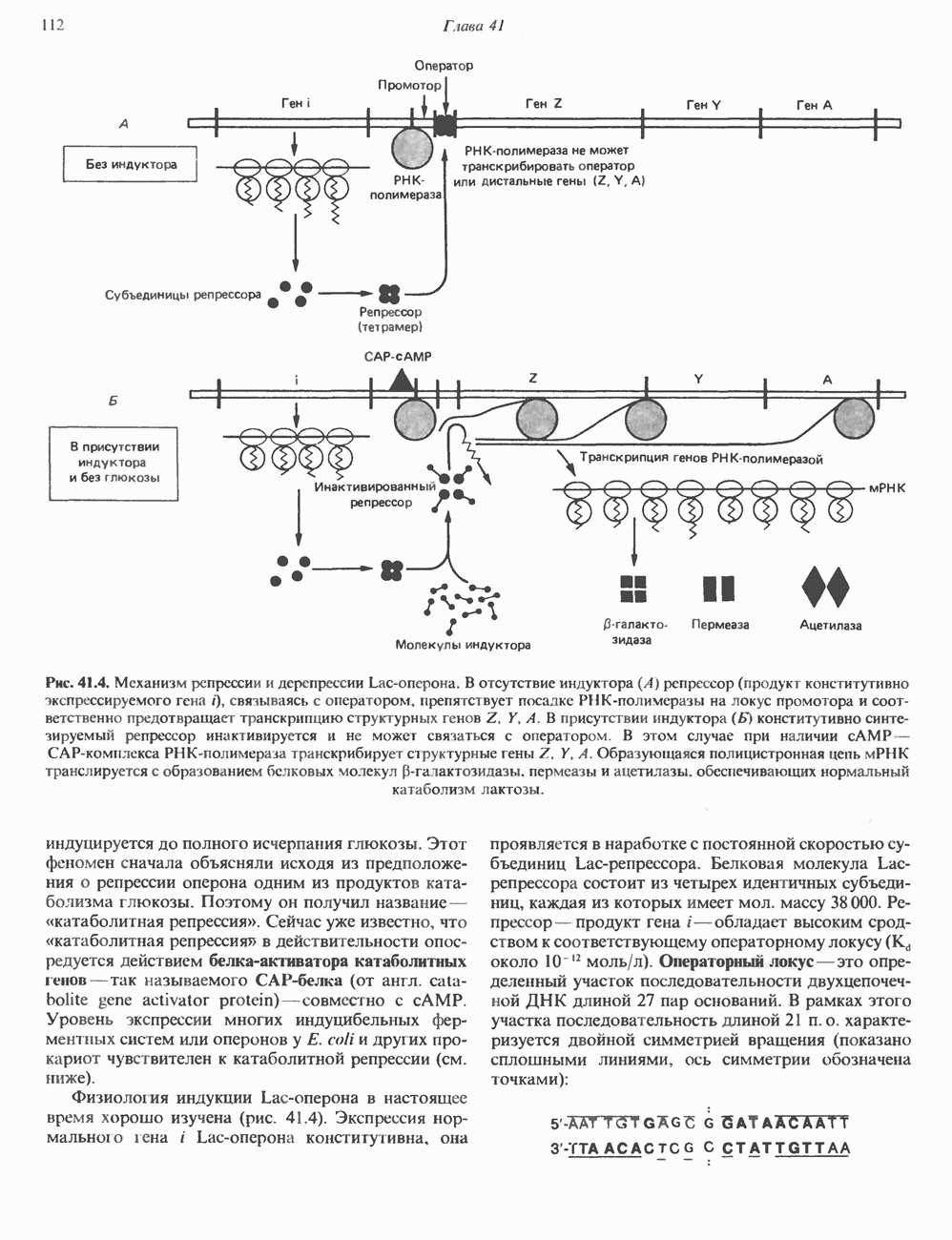
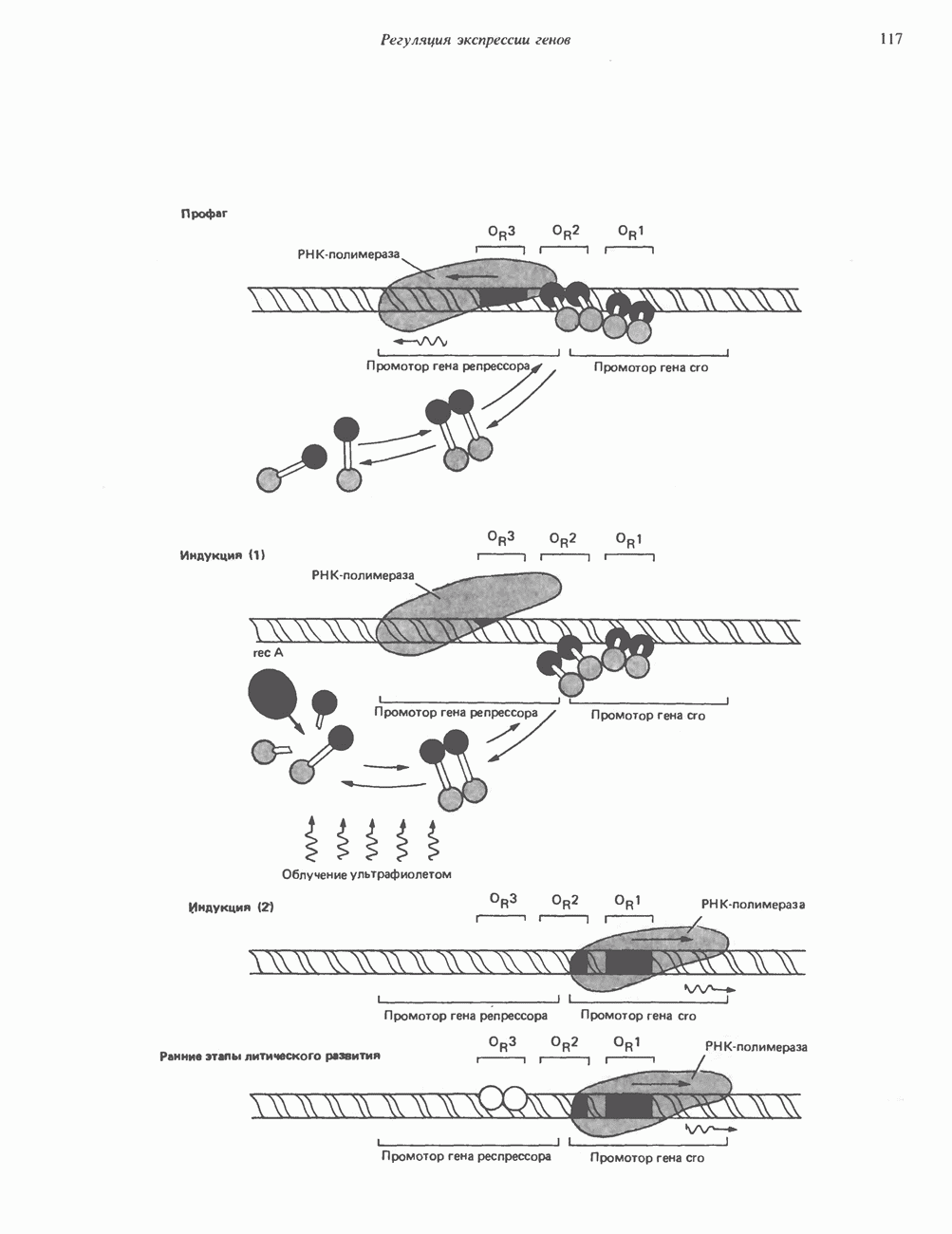
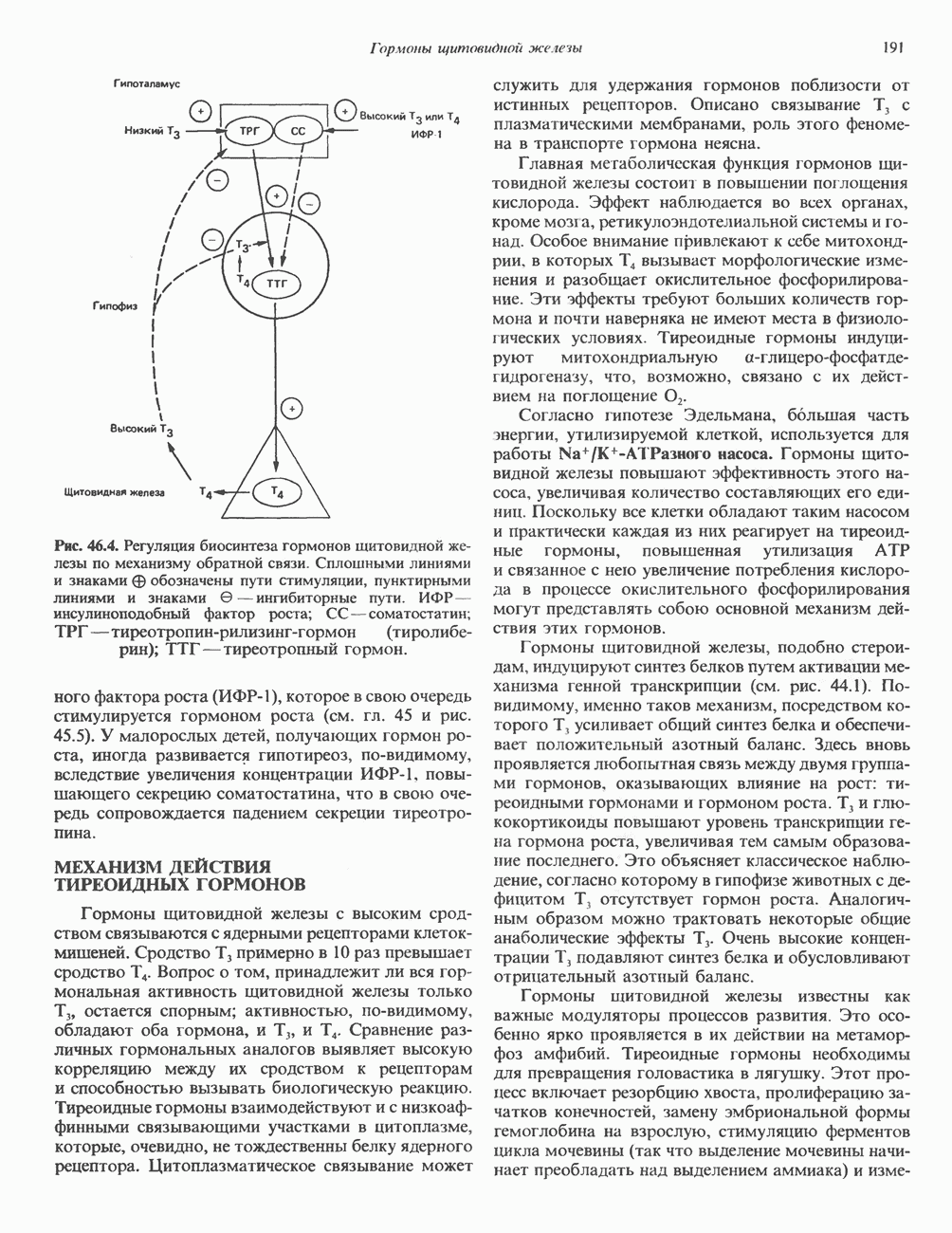
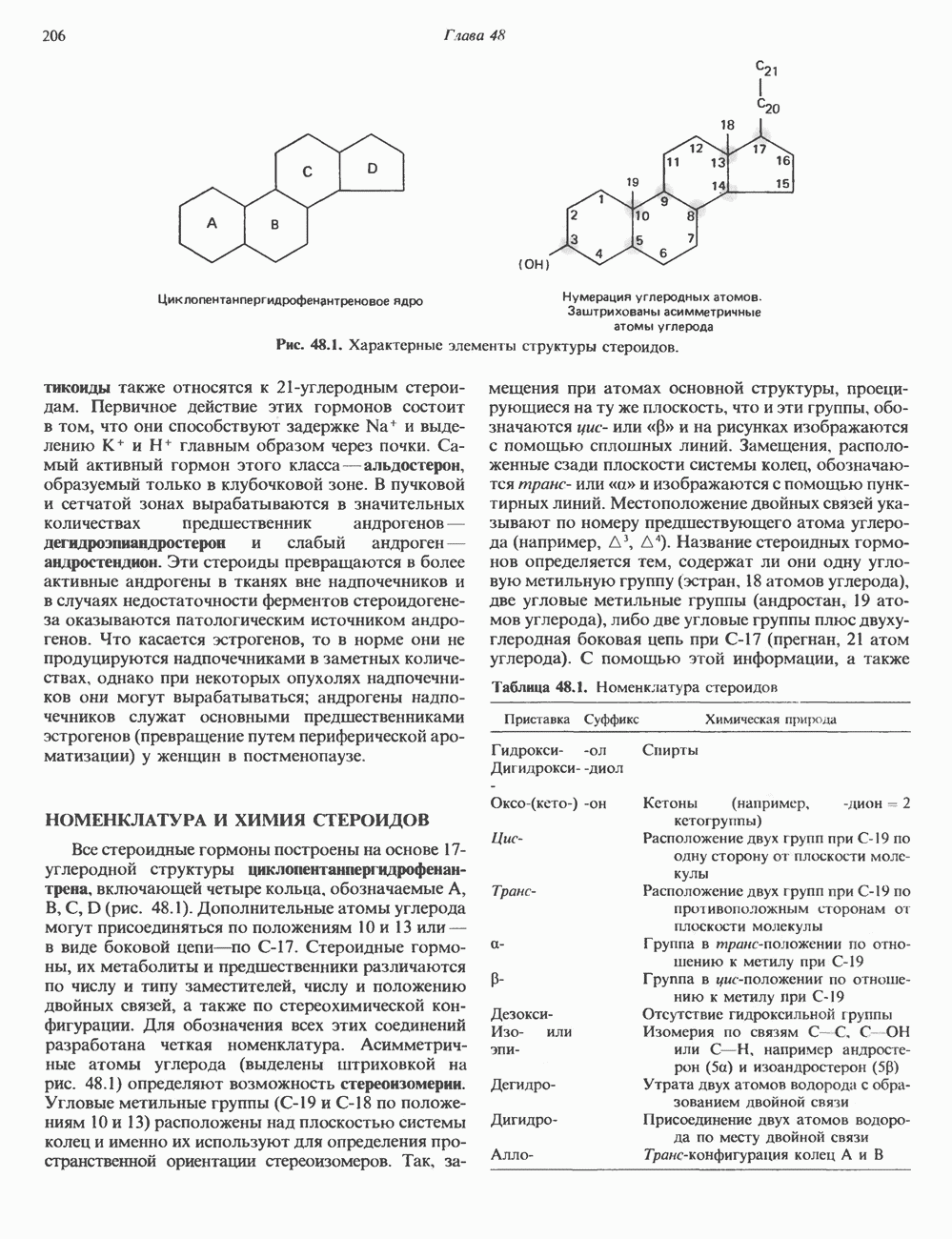
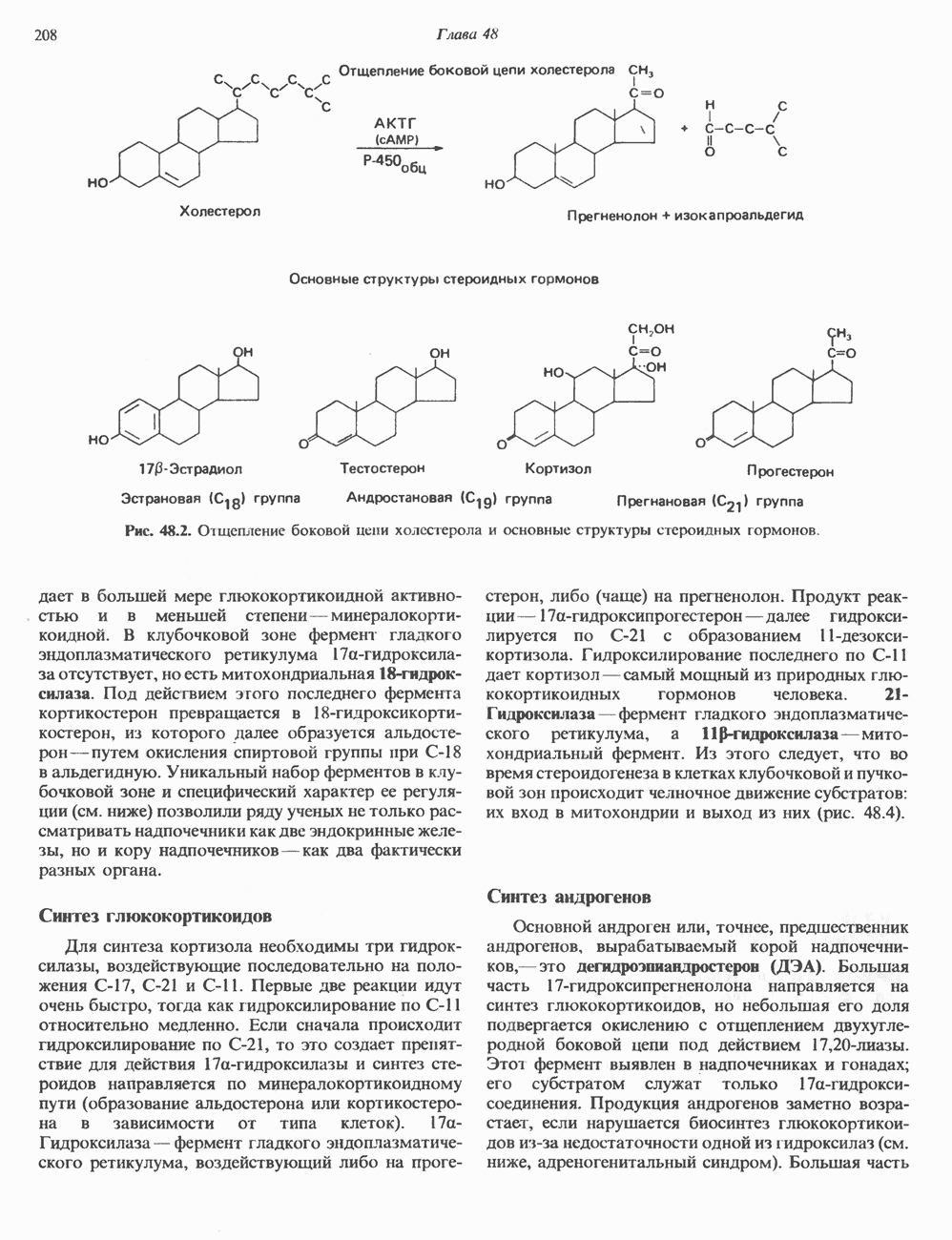
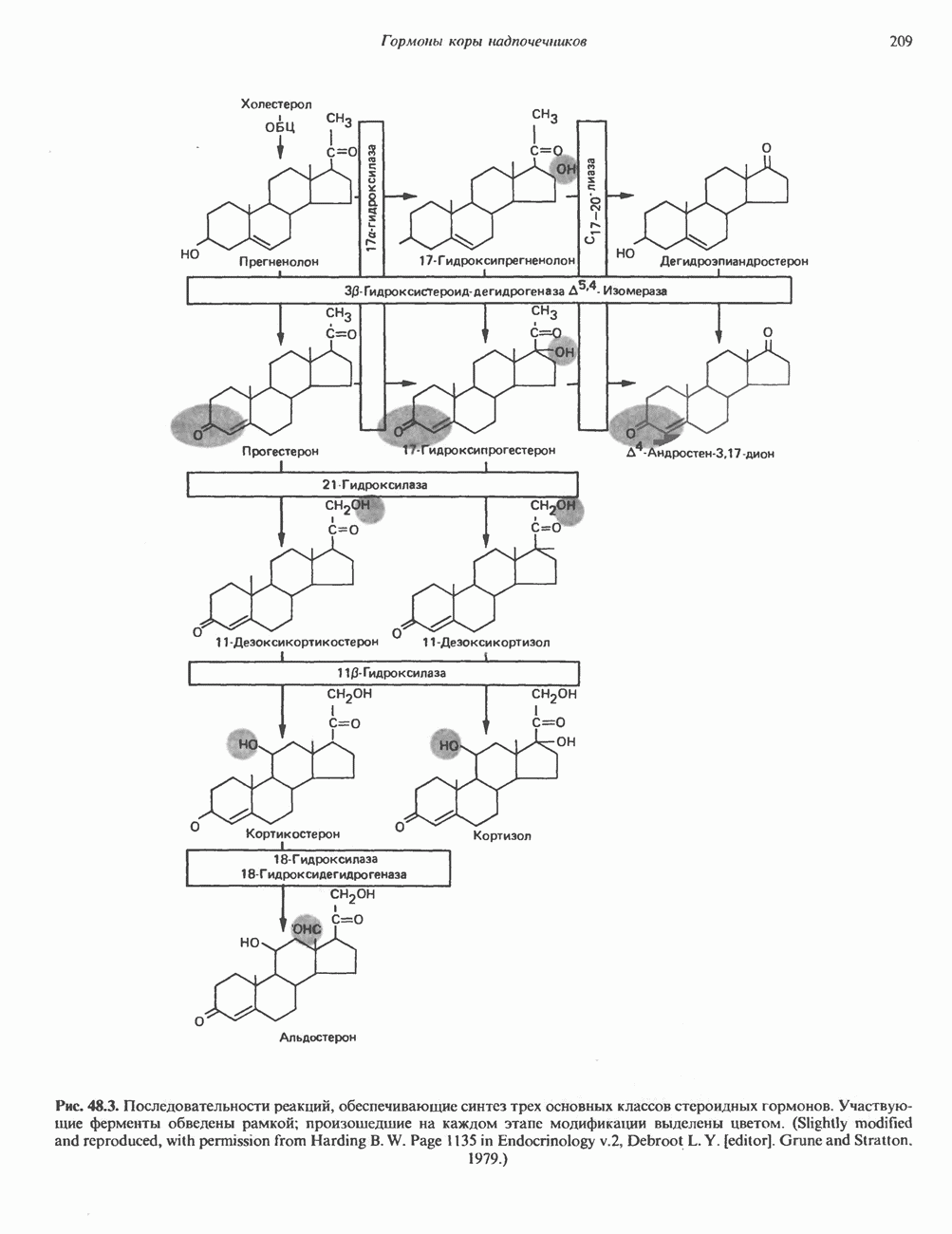
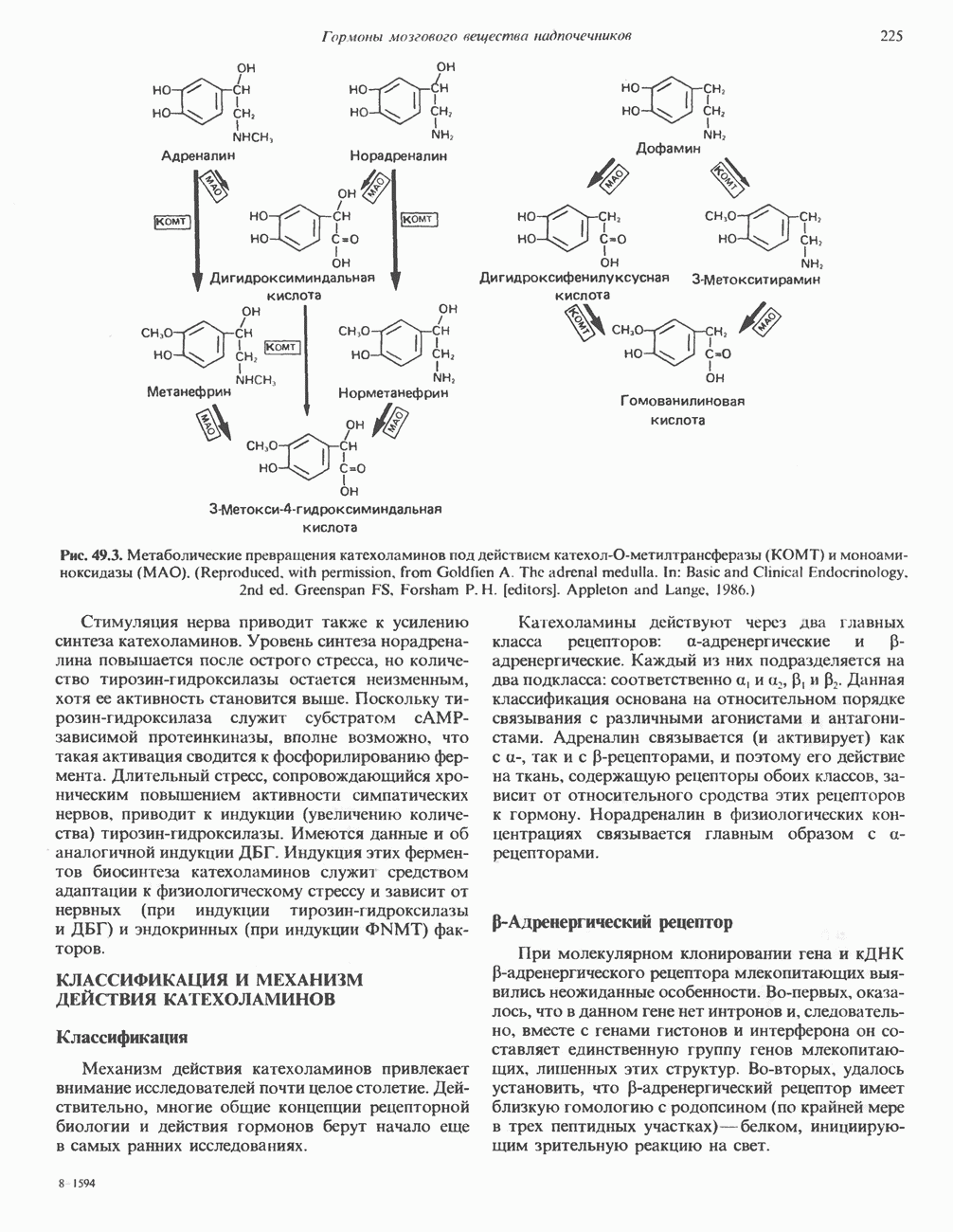
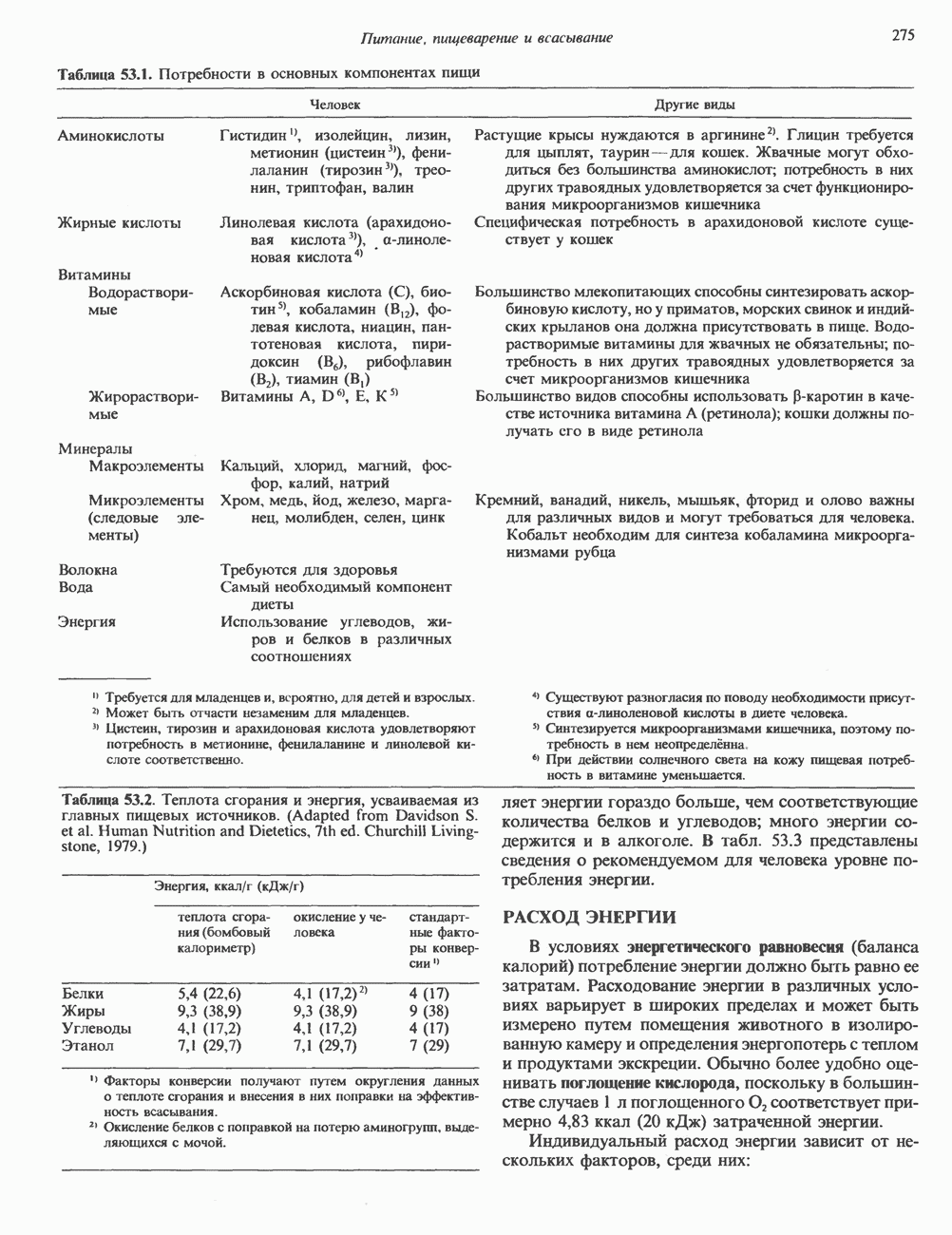
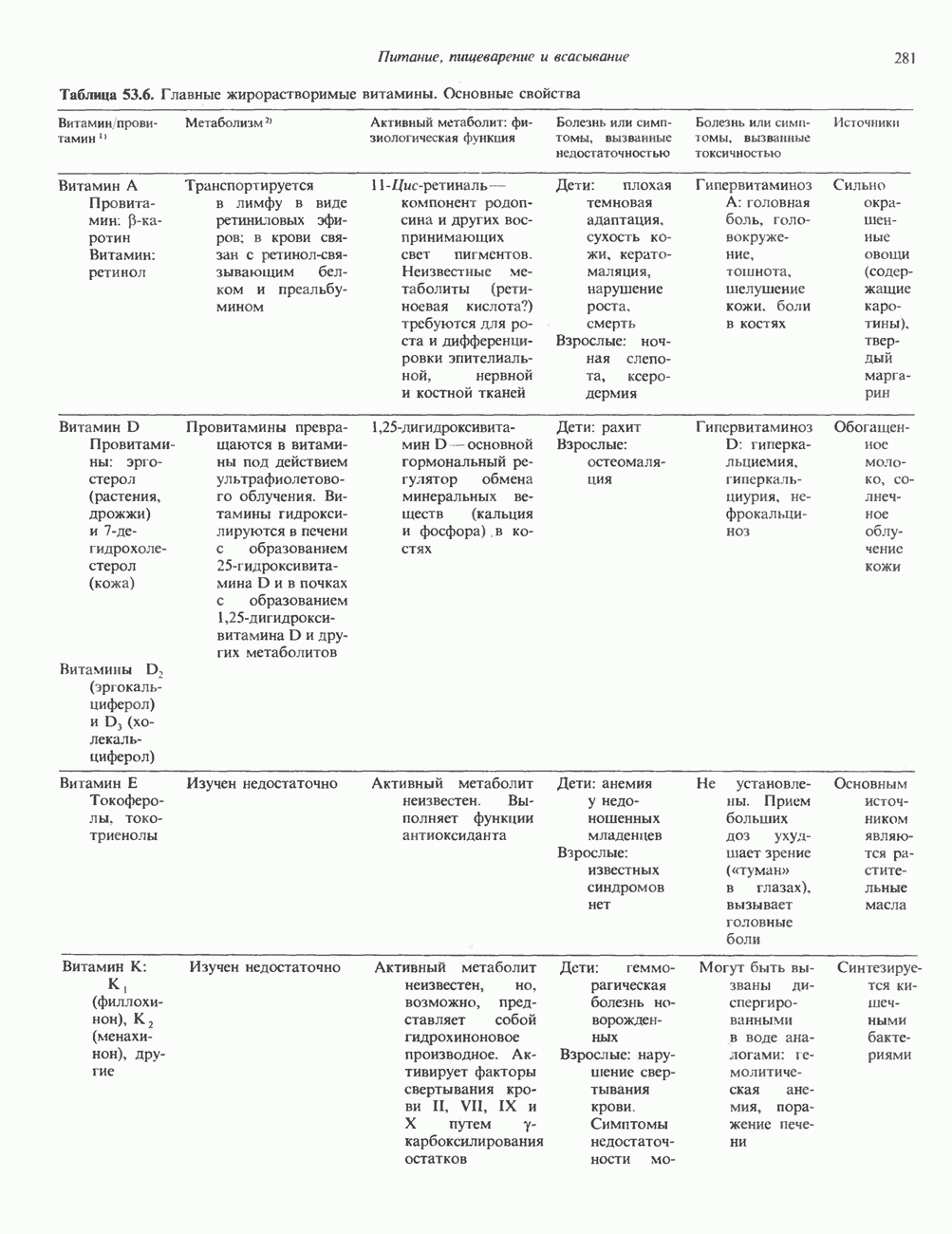
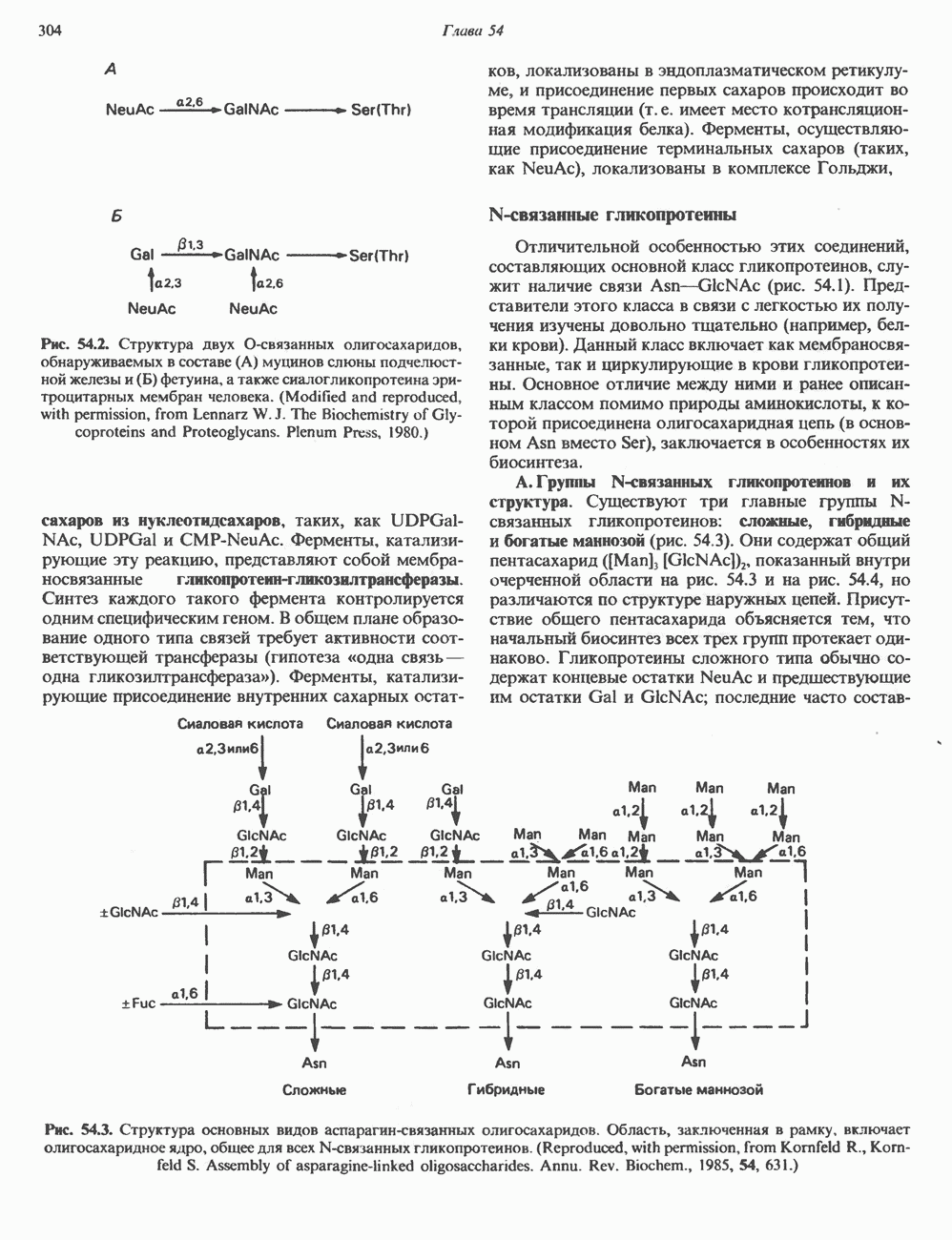
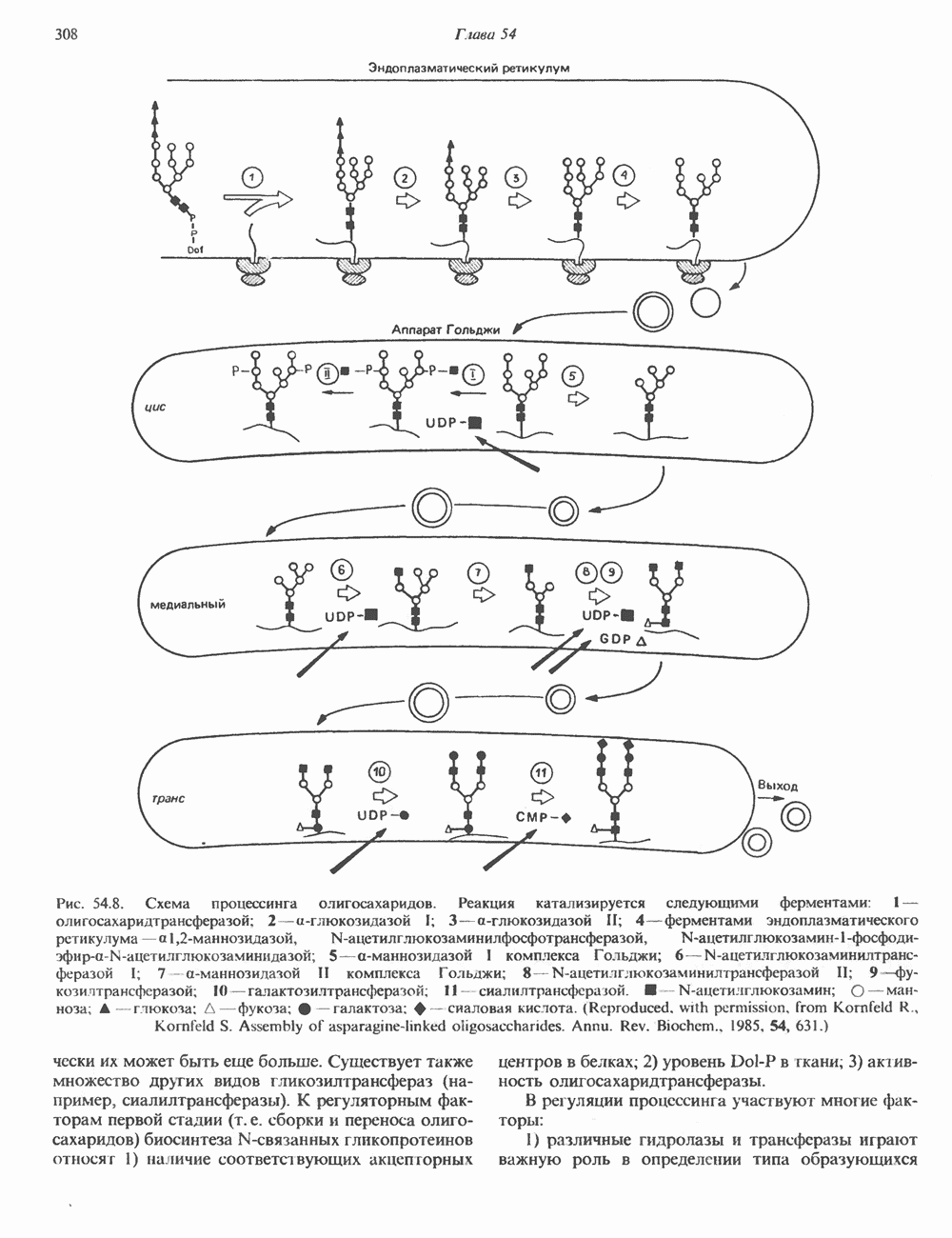
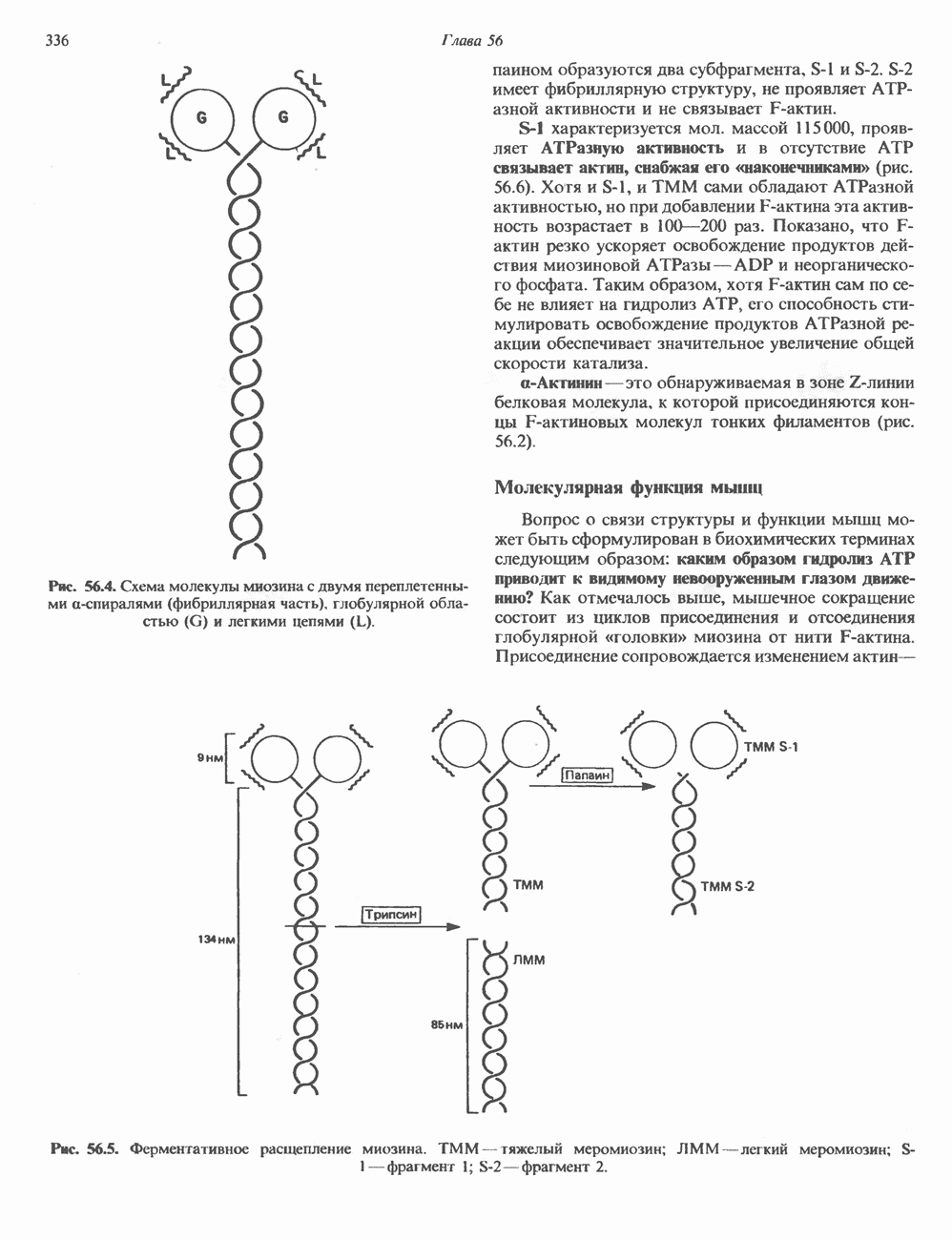
ГИСТОНЫ И НУКЛЕОСОМЫТермином «гистоны» обозначают несколько групп близкородственных основных белков. Н1-гистоны наиболее слабо связаны с хроматином и легко отмываются в солевом растворе. После такой обработки хроматин становится растворимым. Изолированные ядра нуклеосом состоят из гистонов четырех классов: Н2А, Н2В, Н3 и Н4. Структура умеренно богатых лизином гистонов Н2А и Н2В характеризуется значительной консервативностью, еще более консервативна структура гистонов Н3 и Н4 (богатых аргинином). Высокая консервативность структуры гистонов свидетельствует об идентичности функций этих белков у всех эукариот. С-концевая часть их молекулы имеет обычный аминокислотный
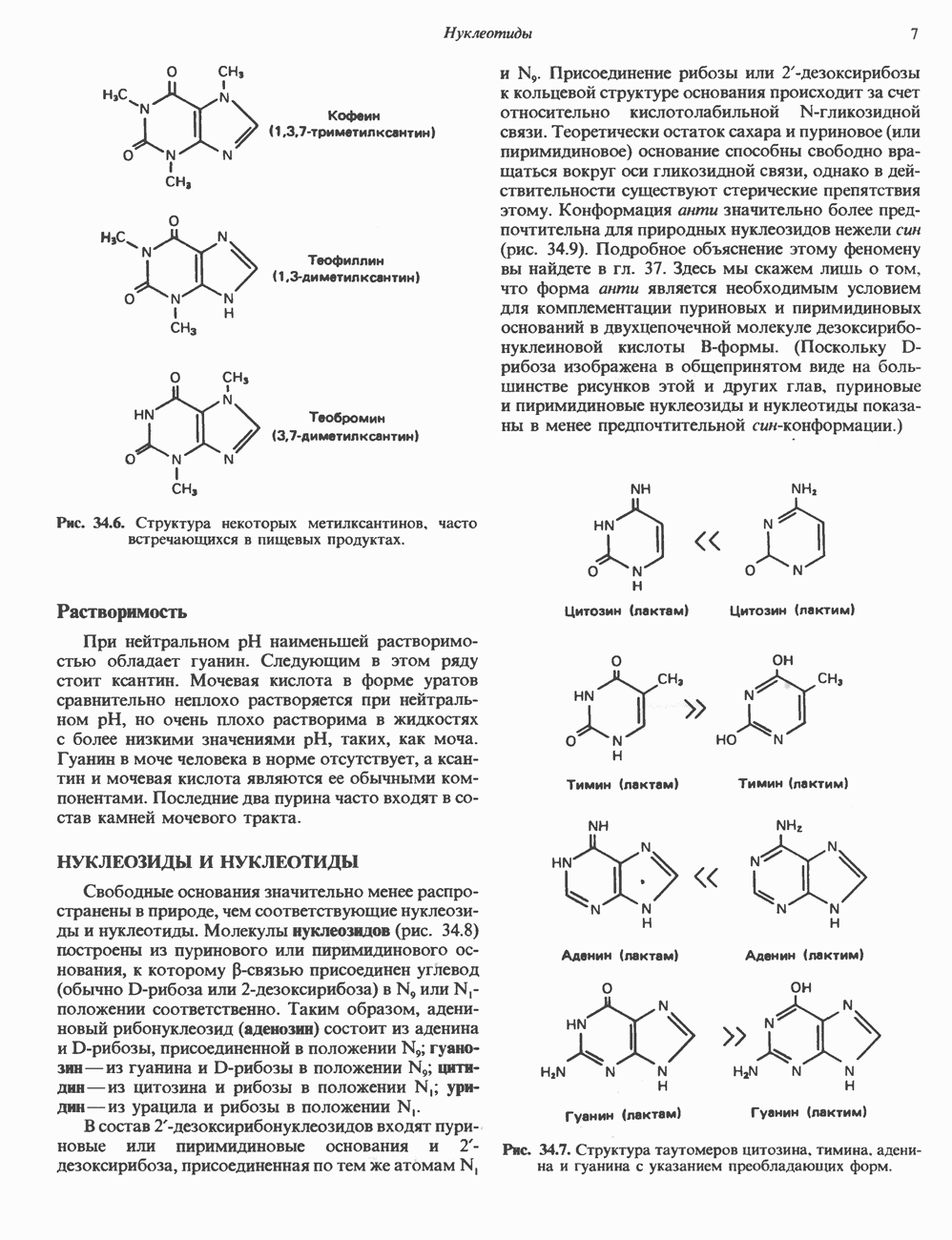
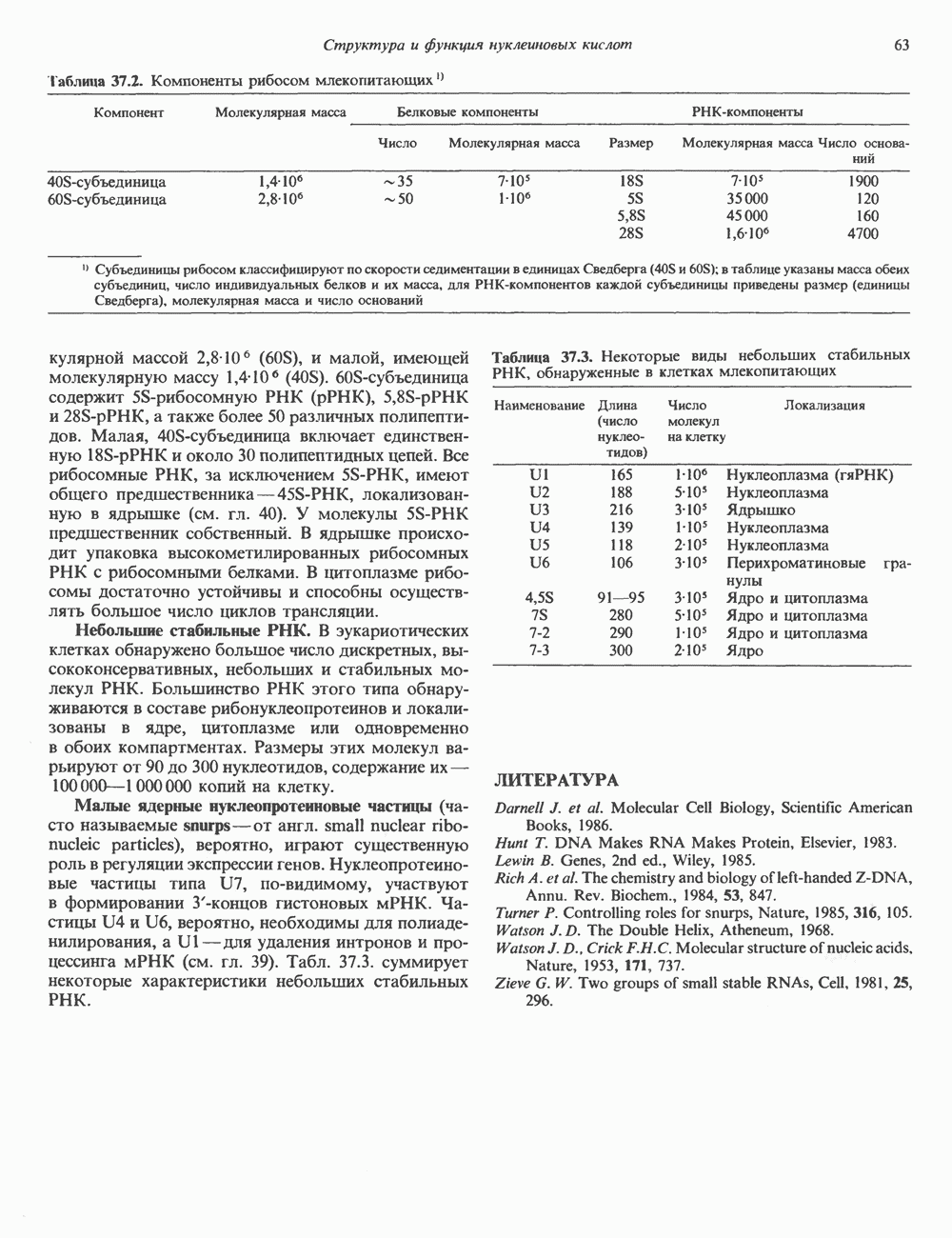
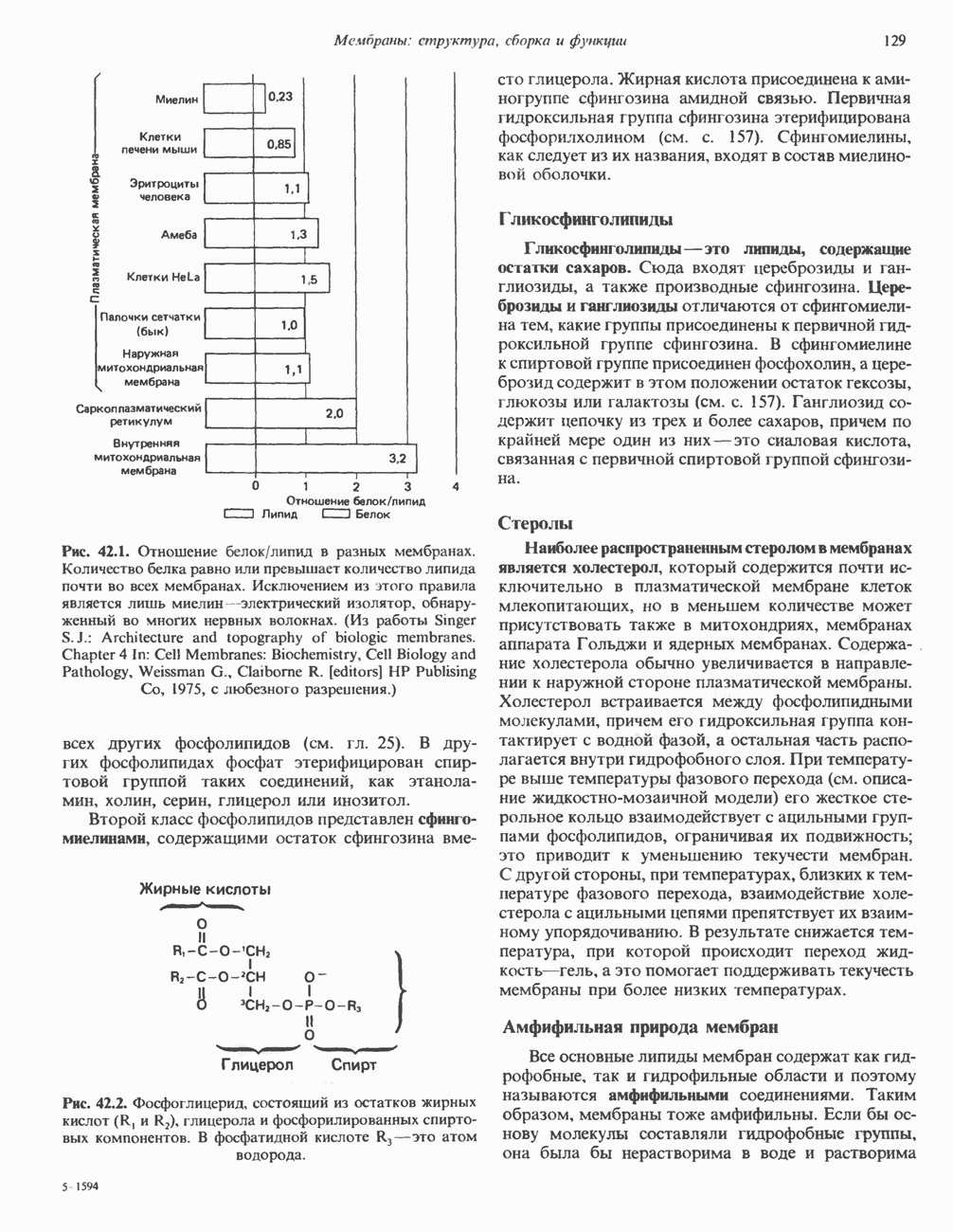
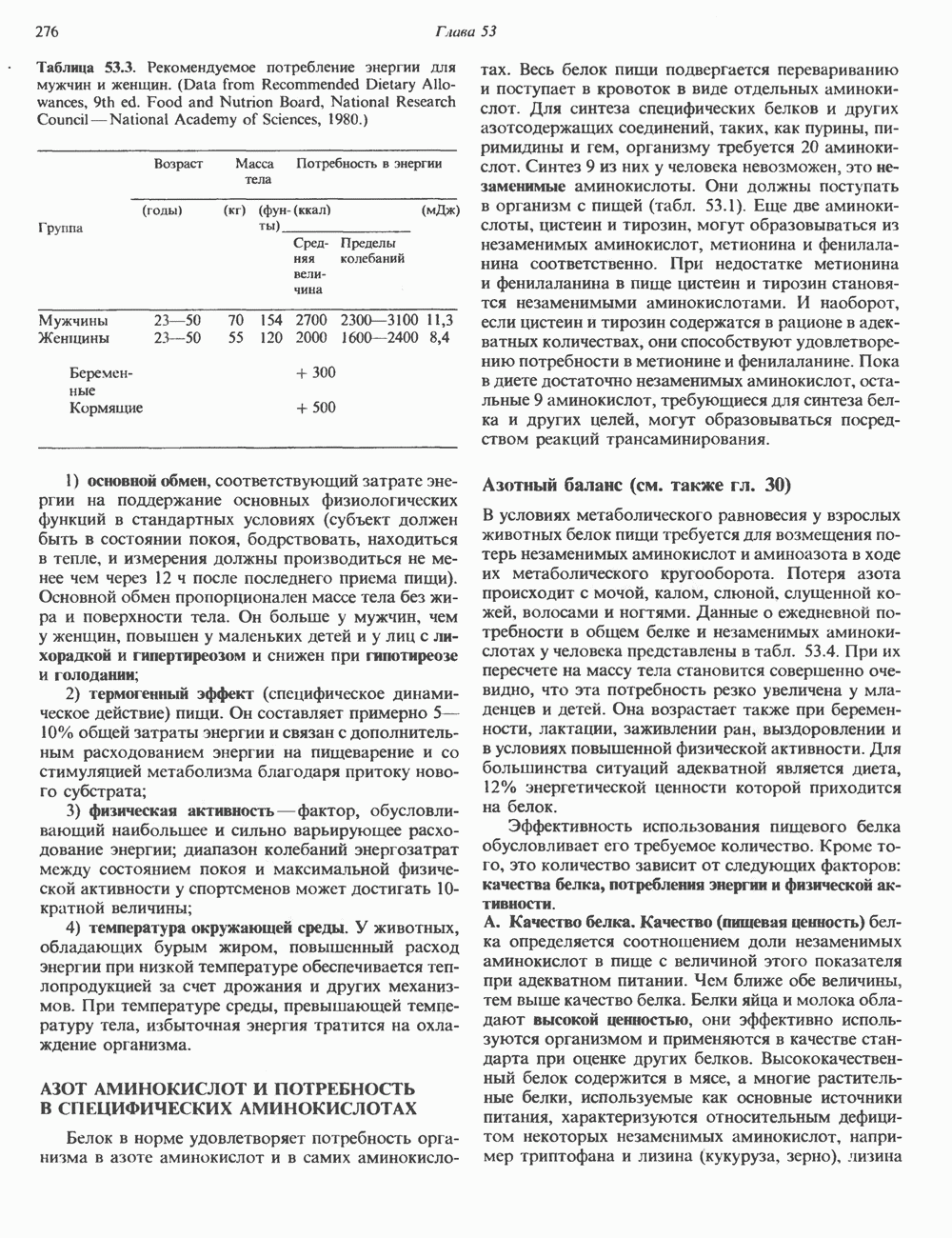
Рис. 38.1. Электронная микрофотография нуклеосом, соединенных ДНК-цепью; белая полоса соответствует 2,5 мкм. (Reproduced with permission, from P. Oudet, M. Gross-Bellard, P. Chambon: Electron microscopic and biochemical evidence that chromatin structure is a repeating unit Cell 1975, 4: 281.) состав, тогда как N-концевая треть молекулы состоит преимущественно из основных аминокислот. Перечисленные выше четыре группы гистонов подвергаются ковалентной модификации пяти типов: ацетилированию, метилированию, фосфорилированию, ADP-рибозилированию и ковалентному связыванию (только Н2А) с убиквитином (ядерным белком). Эти модификации, вероятно, влияют на структуру и функции хроматина (пока данный вопрос изучен недостаточно). Выделенные из хроматина гистоны взаимодействуют между собой. Гистоны НЗ и Н4 агрегируют с образованием тетрамеров, состоящих из двух молекул каждого типа (Н32-Н42). Гистоны Н2А и Н2В образуют либо димеры (Н2А-Н2В), либо олигомерные комплексы [Н2А-Н2В]П. Тетрамер Н32-Н4, не взаимодействует с Н2А-Н2В-димером или олигомером. Гистоны Н1 не связываются в растворе с другими гистонами. Интересно, что смесь Н32-Н42 и Н2А-Н2В с очищенной двухцепочечной ДНК дает картину рентгеновской дифракции, характерную для свежевыделенного хроматина. На электронно-микроскопических фотографиях такого препарата видны вновь образованные нуклеосомы. Более того, оказалось, что образование нуклеосом in vitro из ДНК и гистонов Н2А, Н2В, Н3 и Н4 не зависит от того, из каких организмов или клеток были выделены компоненты смеси. Гистоны Н1 и негистоновые белки для формирования нуклеосомного кора не требуются. В нуклеосомах ДНК суперскручена на поверхности дисковидного гистонового октамера в левостороннюю спираль. Октамер состоит из центрального Тетрамер
Рис. 38.2. Модель структуры нуклеосомы (слева) и нуклеосомного кора (справа), в которой ДНК закручена вокруг белкового цилиндра, содержащего по две молекулы каждого из гистонов Н2А, Н2В, Н3 и Н4. Гистон Н1 (заштрихованная область) расширяет область маскированных участков последовательности ДНК. (Reproduced, with permission, from Laskey R. A. and Eamshaw W.C.: Nucleosome assembly. Nature 1980, 286: 763.) нуклеосомоподобную структуру и, следовательно, играет центральную роль в ее формировании. Два добавочных димера
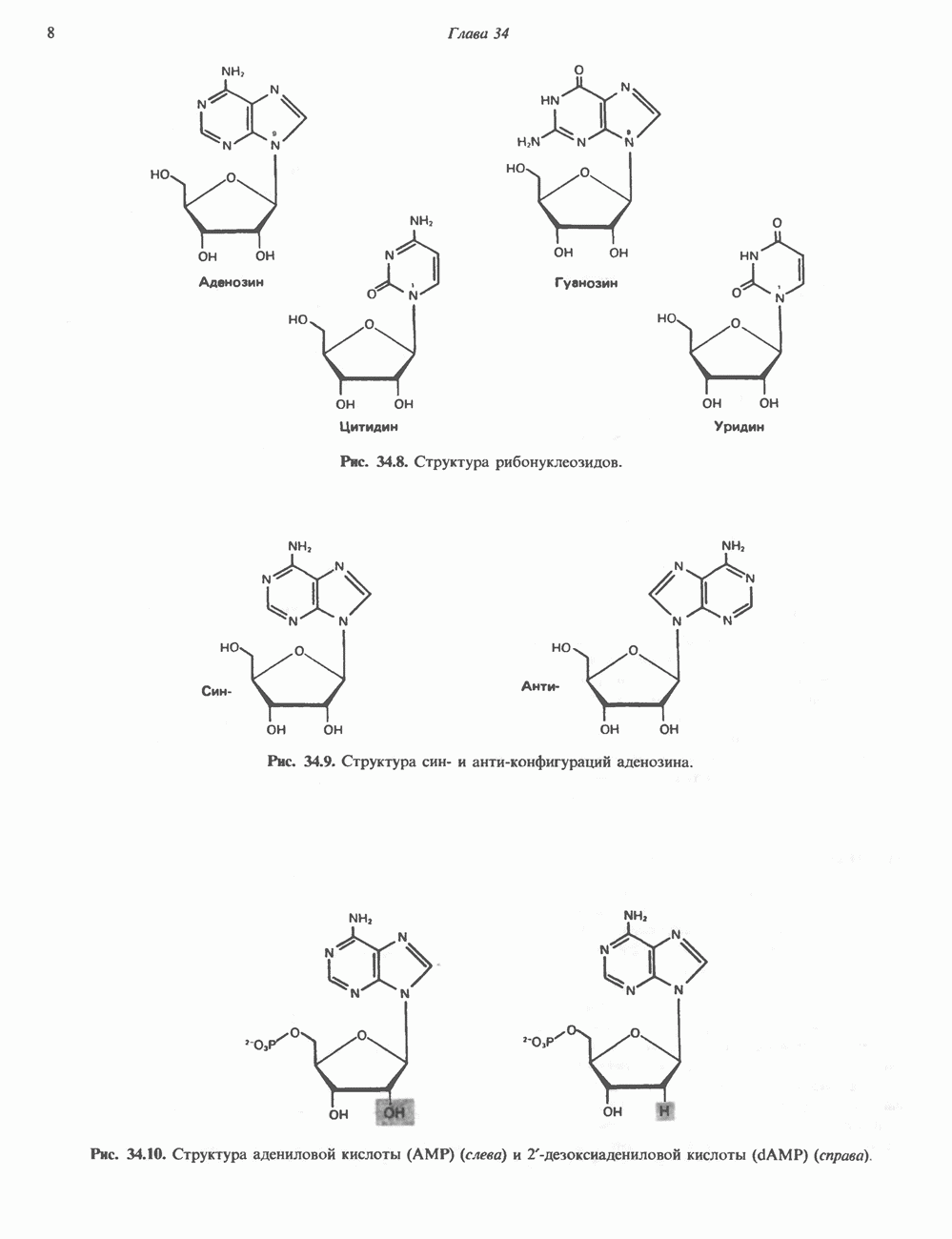
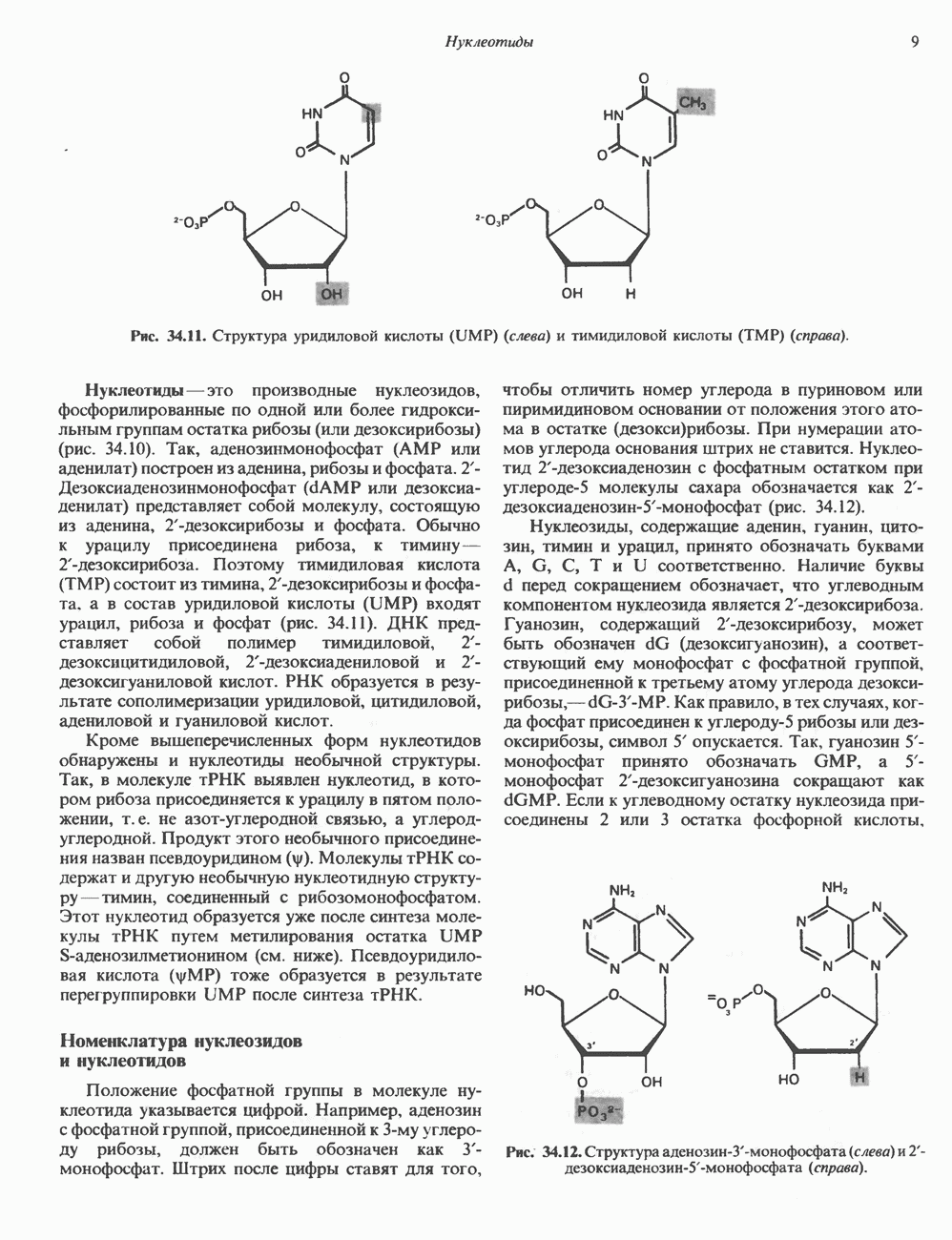
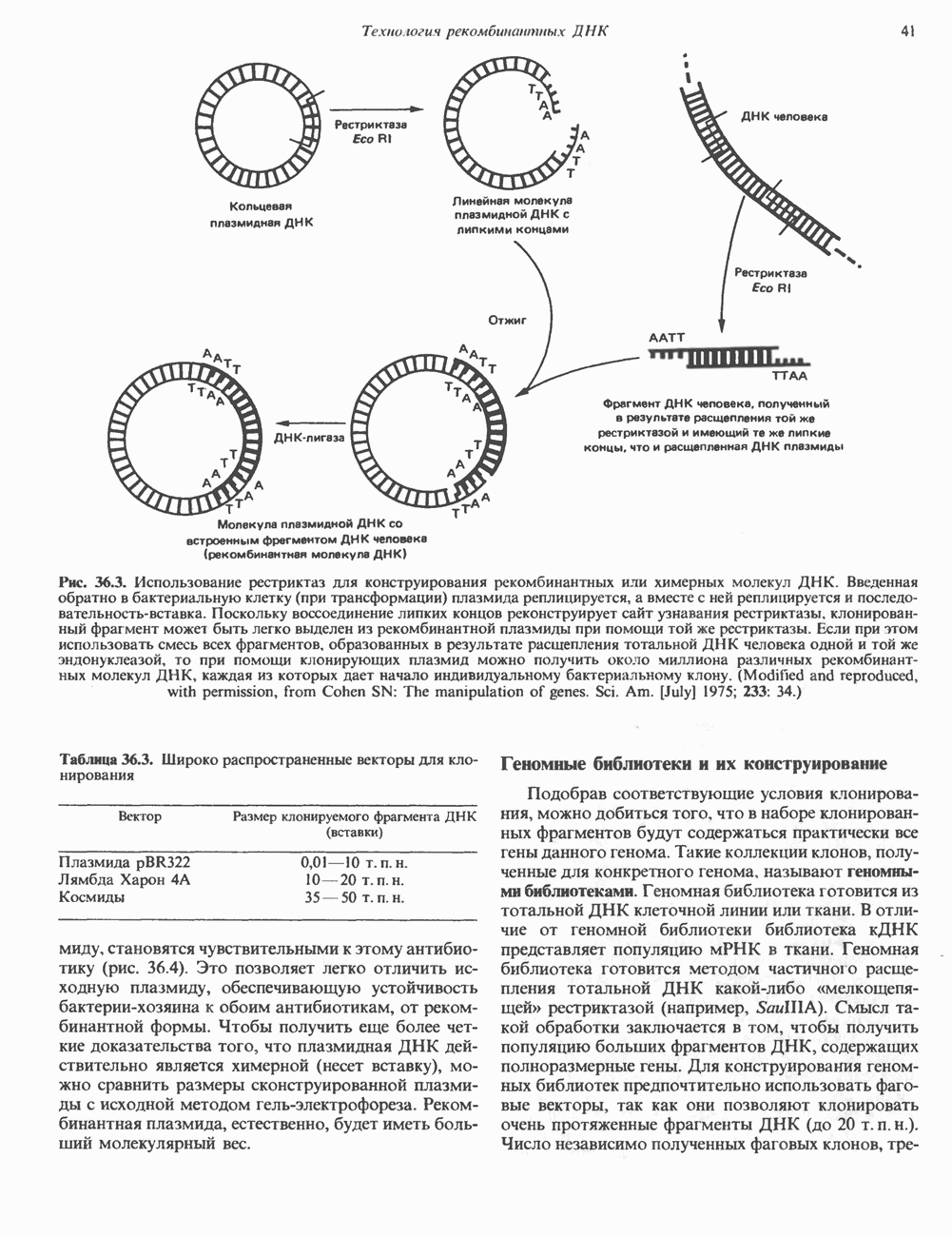
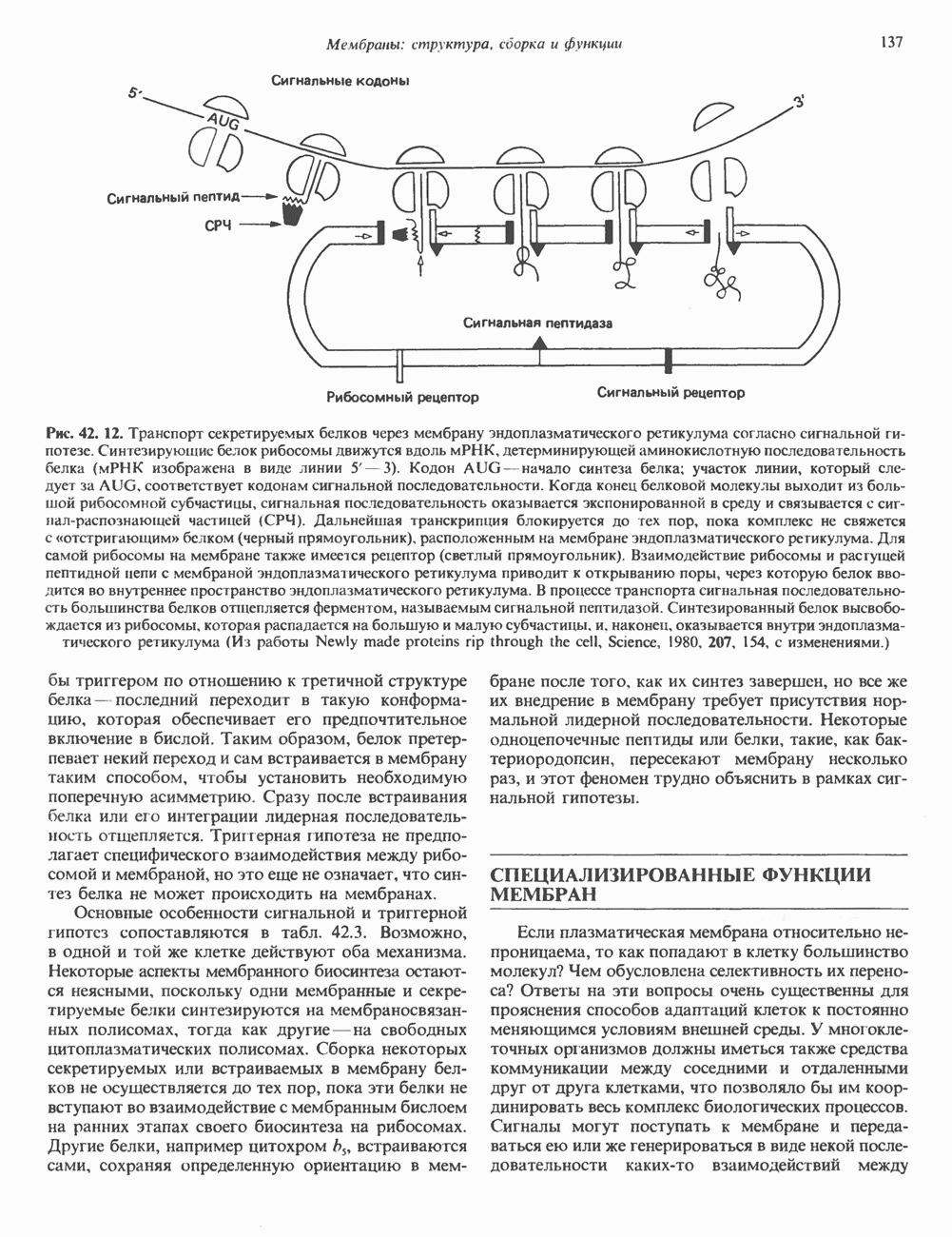
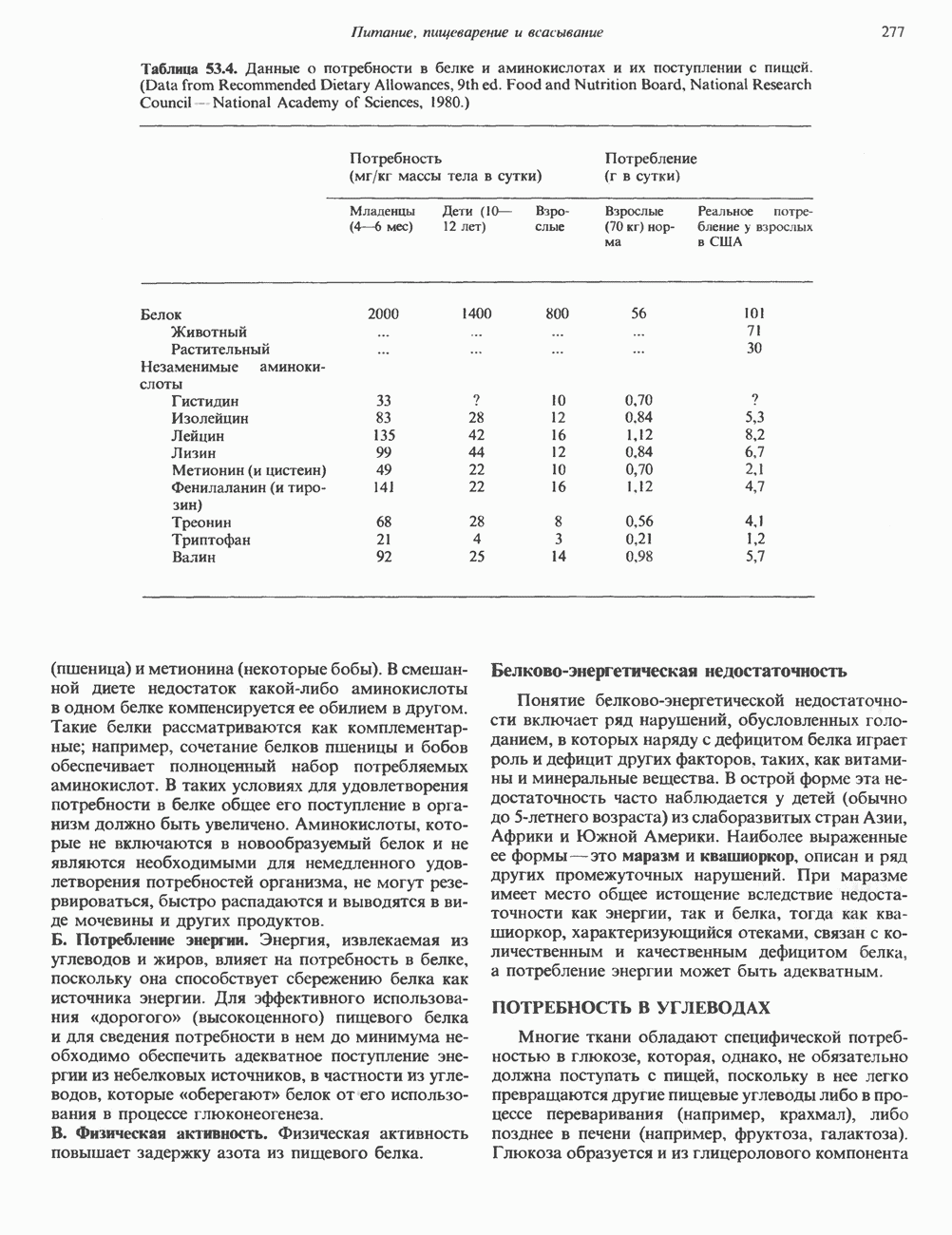
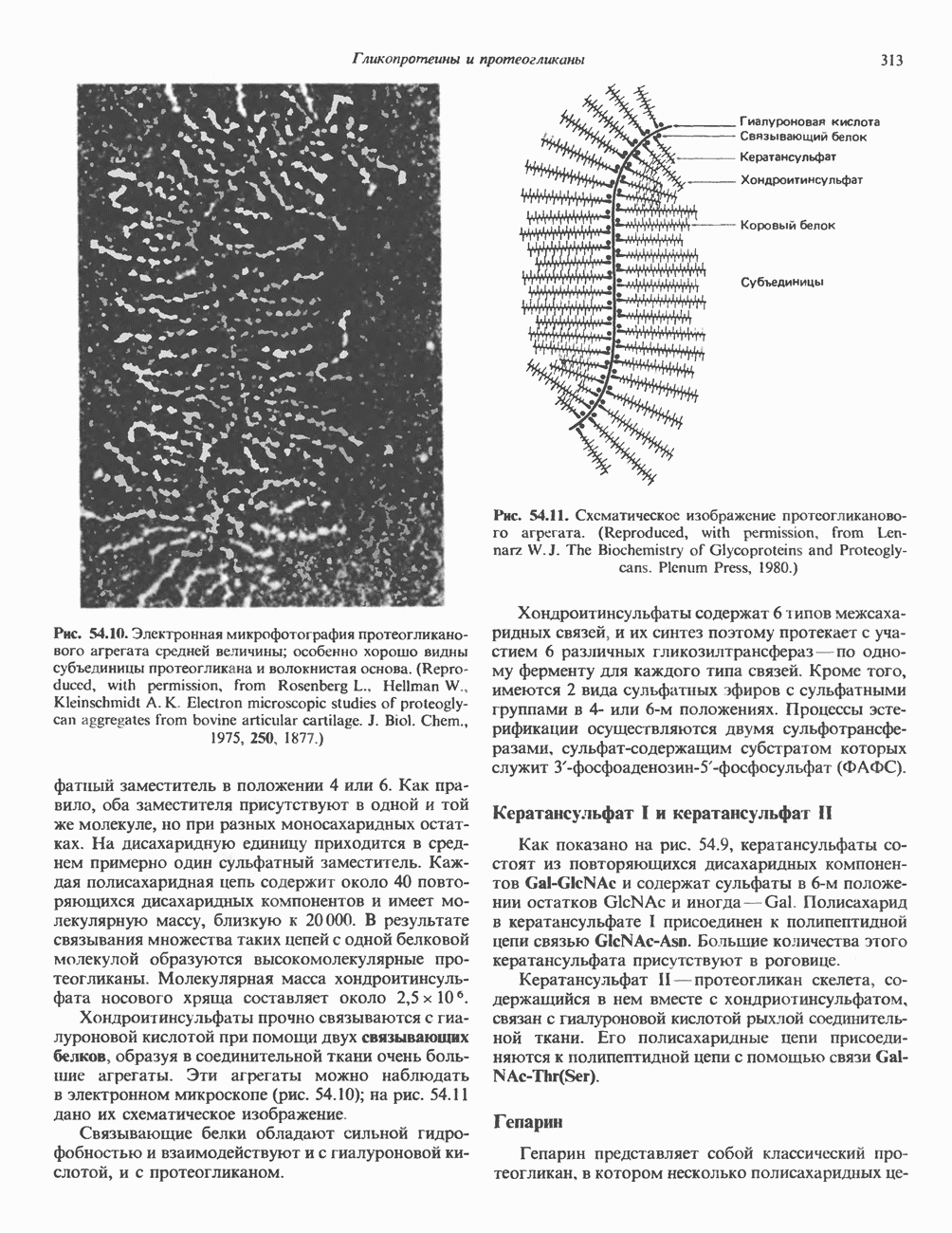
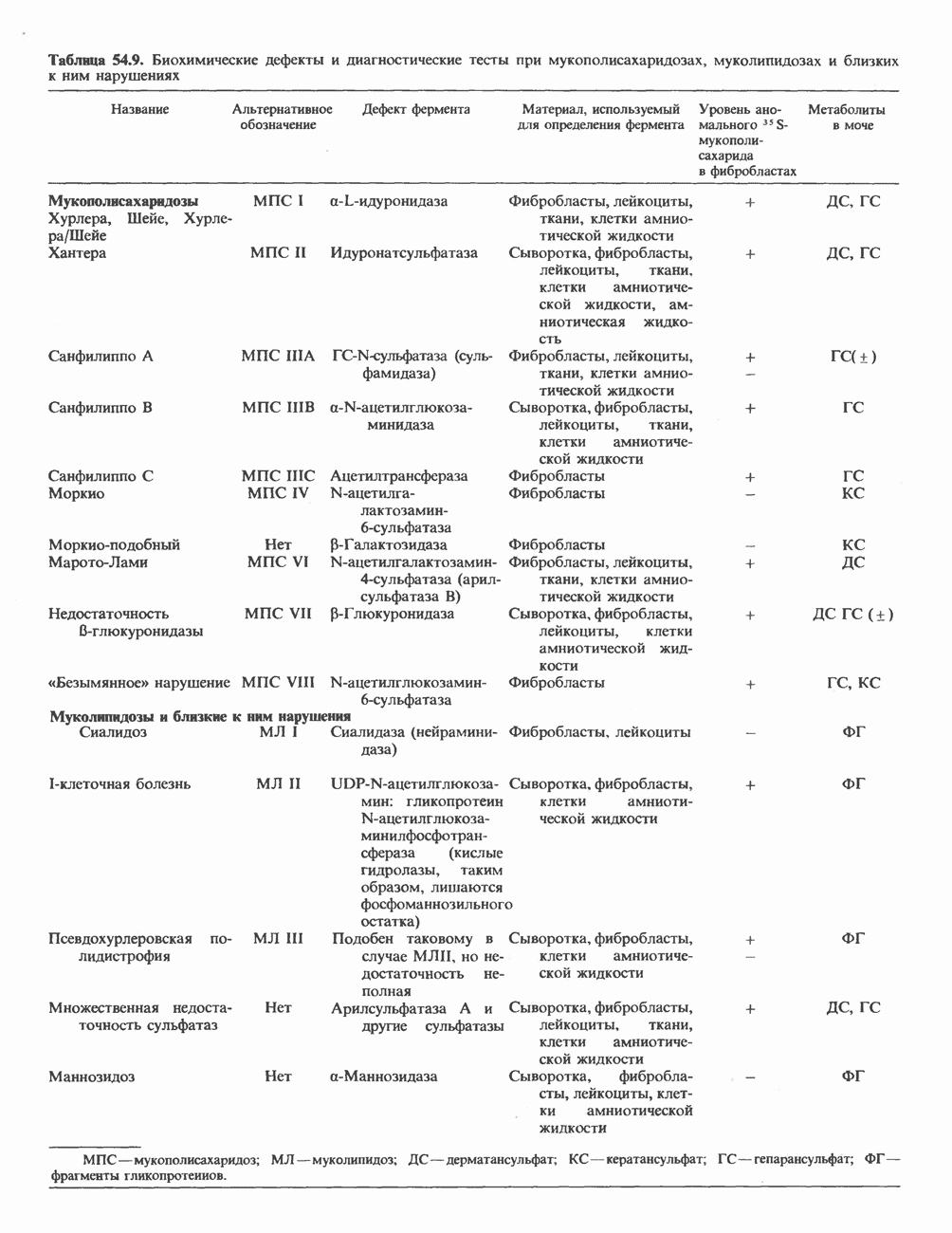
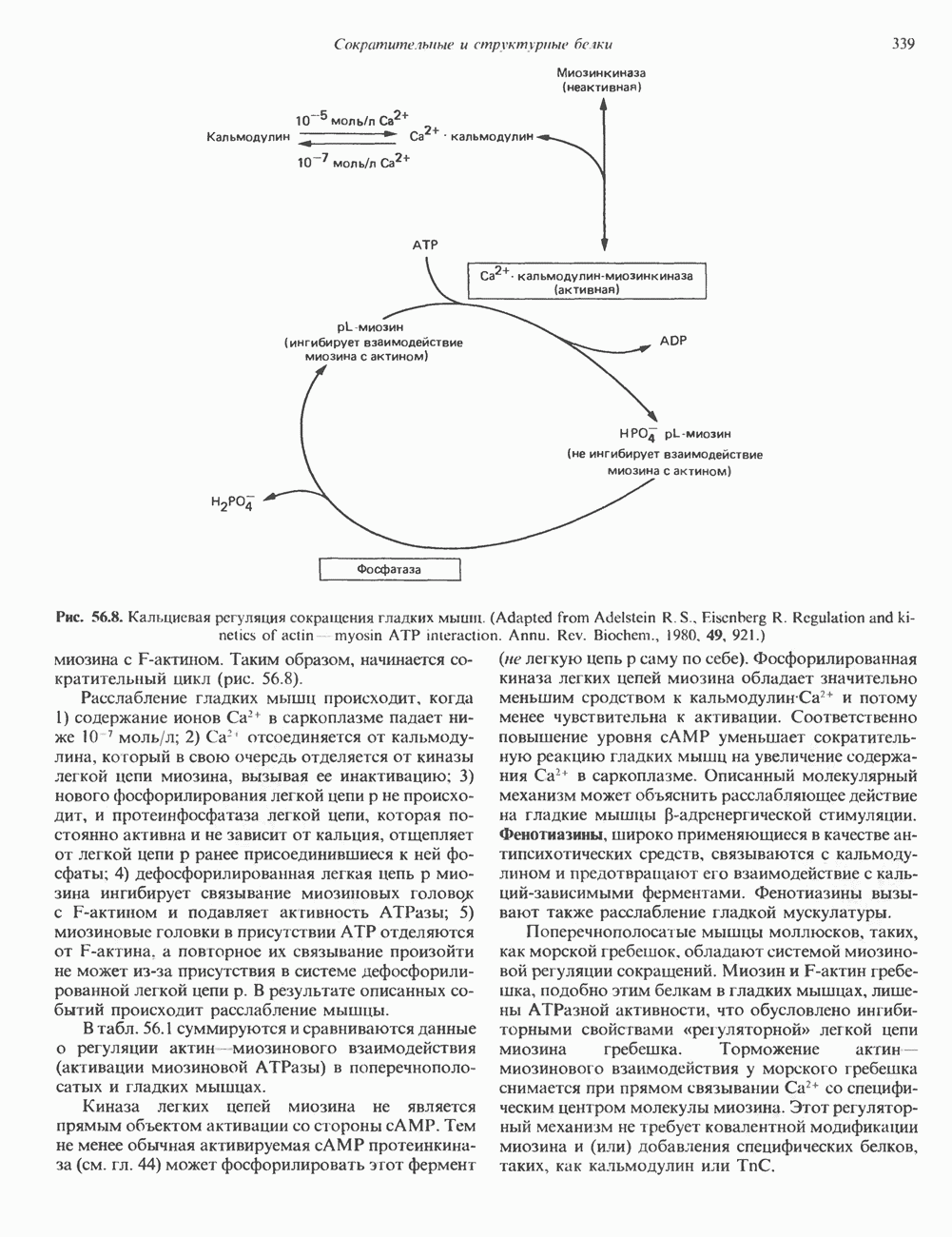
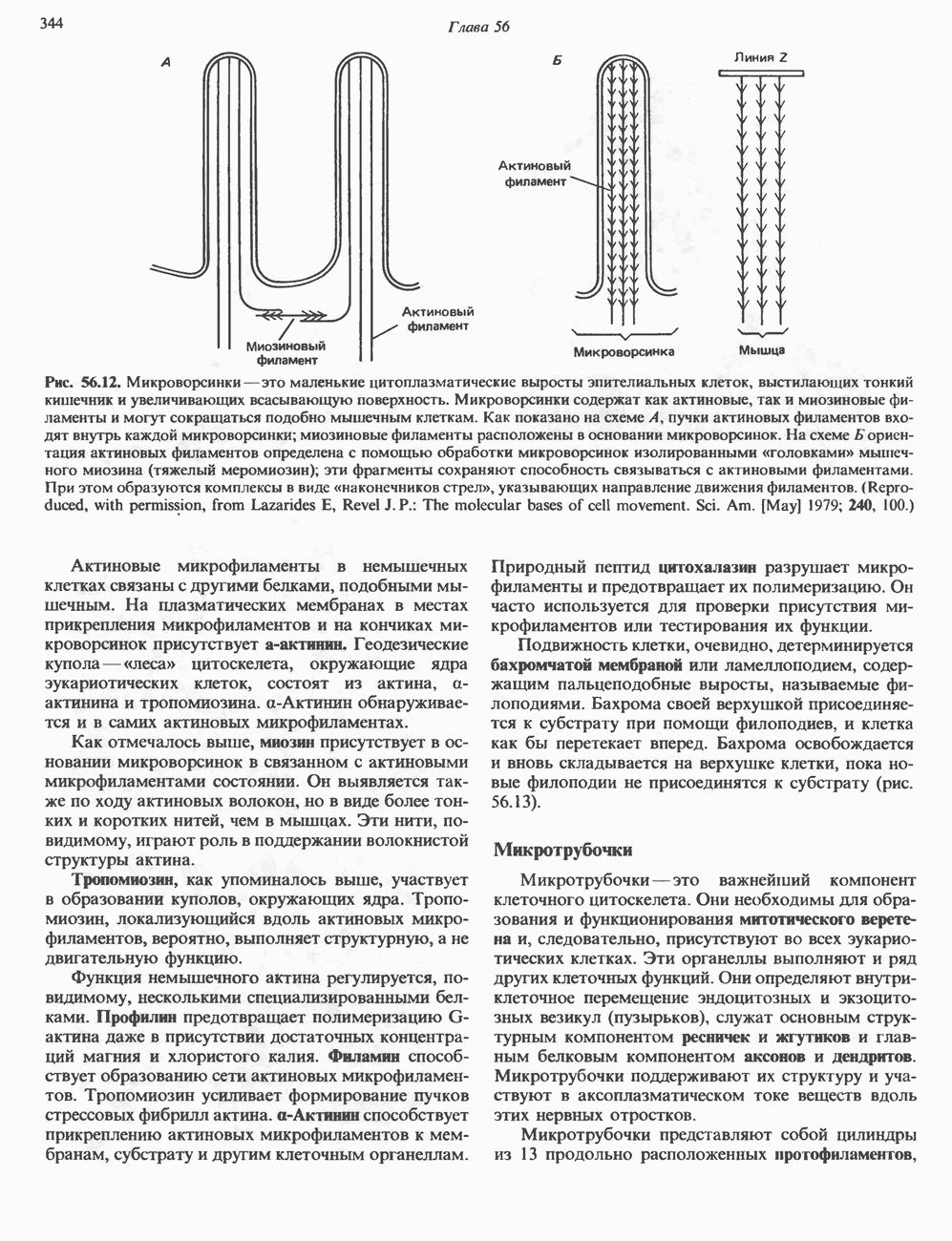
Гистон Н1 связывается с нуклеосомным кором на участке входа и выхода ДНК, «склеивая» 2 оборота, т.е. 166 пар оснований суперспирали ДНК. Так формируется зрелая нуклеосома. В сборке нуклеосомы, вероятно, участвует ядерный белок анионного характера — нуклеоплазмин. Заметим, что гистоны, являясь сильными катионитами, могут неспецифччески связаться с отрицательно заряженной ДНК с образованием солевых мостиков. Ясно, что такое неспецифическое взаимодействие может мешать образованию нуклеосом и проявлению функций хроматина. Нуклеоплазмин — это анионный пентамерный белок, не связывающийся ни с ДНК, ни с хроматином, но способный обратимо соединяться с гистоновым октамером, блокируя способность гистонов к неспецифическому «прилипанию» к отрицательно заряженным структурам, таким, как ДНК. По-видимому, нуклеоплазмин создает в ядре специфическое ионное окружение, способствующее взаимодействию гистонов с ДНК и сборке нуклеосом. После завершения сборки нуклеоплазмин высвобождается из гистонового комплекса. Нуклеоплазмин проявляет избирательность к определенным областям ДНК. Молекулярная основа этого неслучайного распределения, названного фазированием, неизвестна. Возможно, оно связано с относительной физической пластичностью определенных нуклеотидных последовательностей, способных к скручиванию в суперспираль. Упаковка нуклеосом в ядре, по-видимому, зависит от взаимодействия Н1 гистонов с участками двухцепочечной ДНК, соединяющими нуклеосомы. Топология этого взаимодействия, приводящего к образованию межнуклеосомных спейсерных участков, изучена недостаточно полно. Электронная микроскопия хроматина кроме нуклеосом выявила еще две структуры высшего порядка — фибриллы диаметром 10 нм и волокна диаметром 25—30 нм. Дисковидные нуклеосомы (см. выше) имеют диаметр 10 нм и высоту 5 нм. По-видимому, фибриллы толщиной 10 нм состоят из ряда нуклеосом, касающихся друг друга своими краями и ориентированных плоскими поверхностями вдоль оси фибриллы (рис. 38.3). Вероятно, фибриллы тоже скручиваются в спираль, на виток которой приходится 6—7 нуклеосом. В результате образуется хроматиновое волокно диаметром 30 нм (рис. 38.4). Витки такой «суперспирали» должны быть достаточно плоскими, а плоские поверхности нуклеосом последующих витков — параллельными друг другу. В иитерфазных хромосомах хроматиновые волокна организованы в домены или петли, состоящие из 30000—100000 пар оснований и «заякоренные» на внутриядерном поддерживающем матриксе. Распределение участков генома в рамках доменной структуры хроматина, вероятно, не является случайным. Можно предположить, что каждый петлеобразующий домен хроматина содержит как кодирующие, так и некодирующие области генов, соответствующих определенной генетической функции.
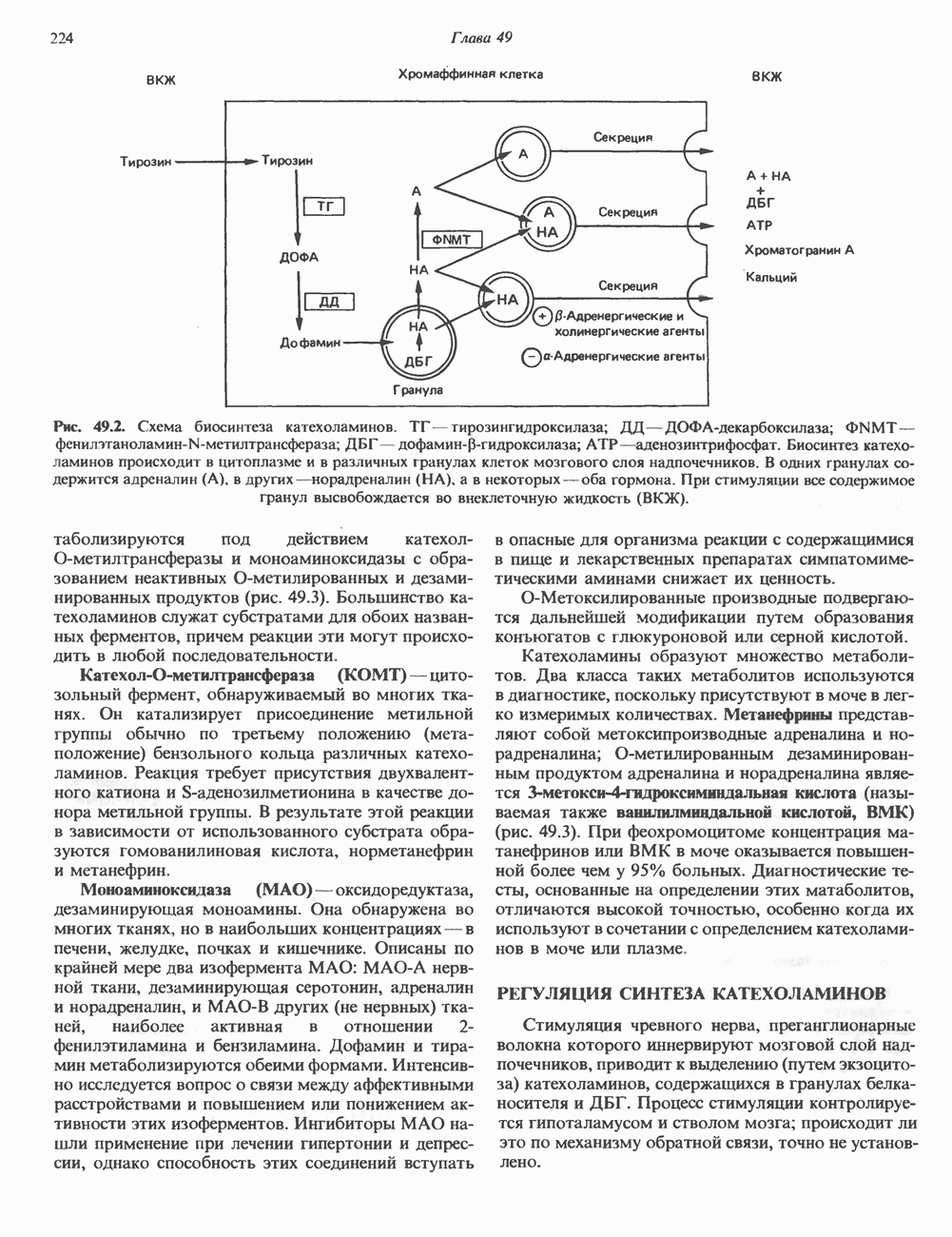
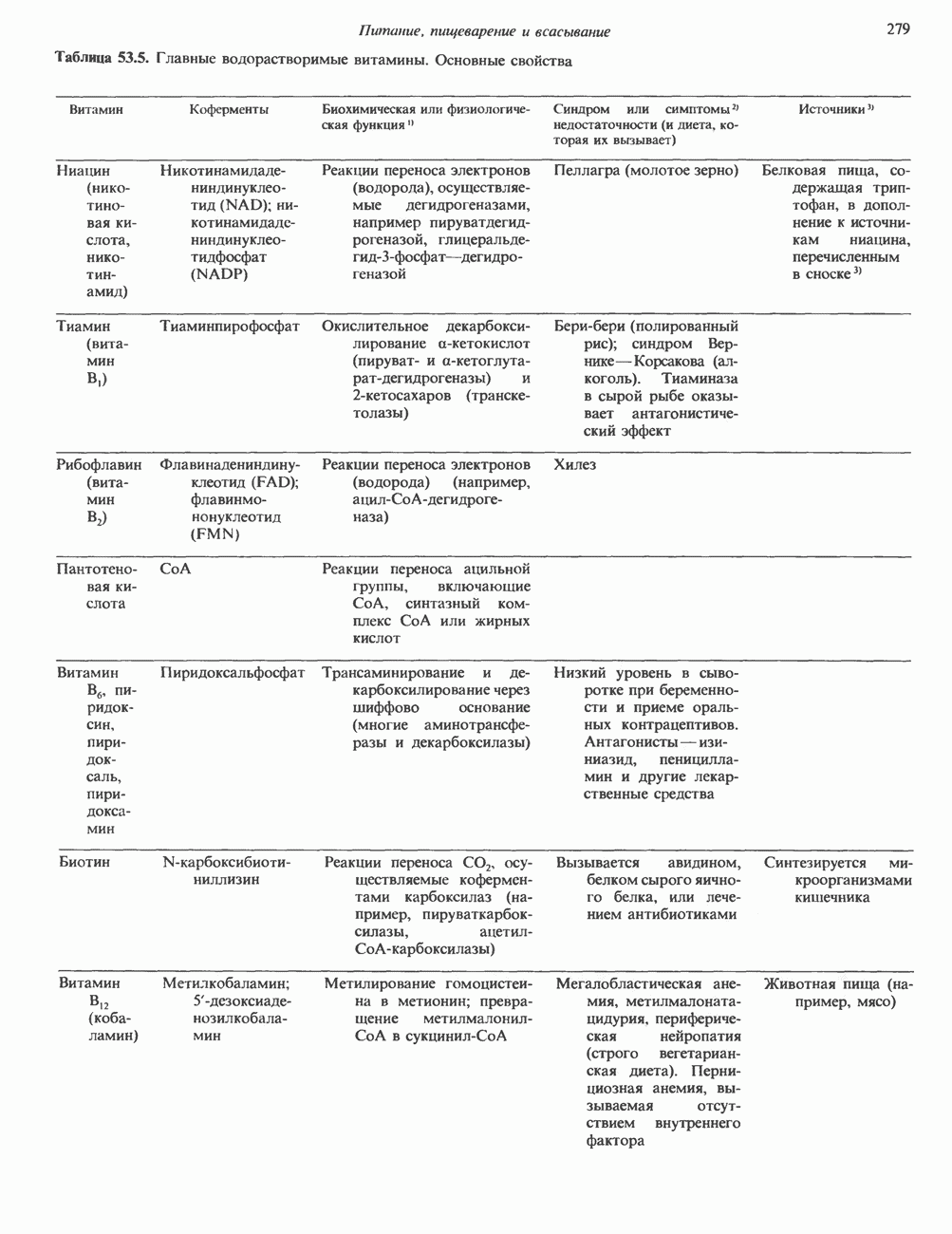
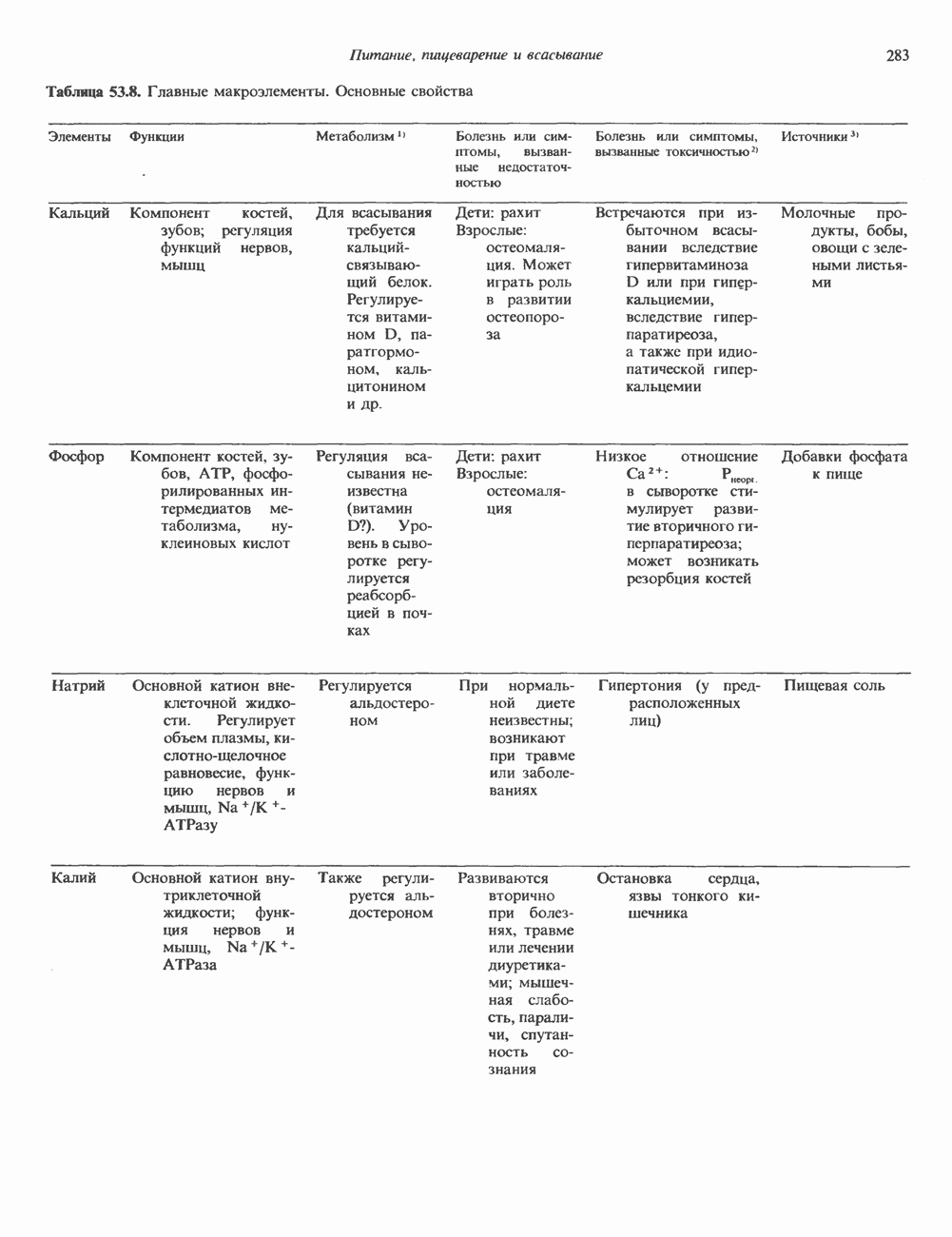
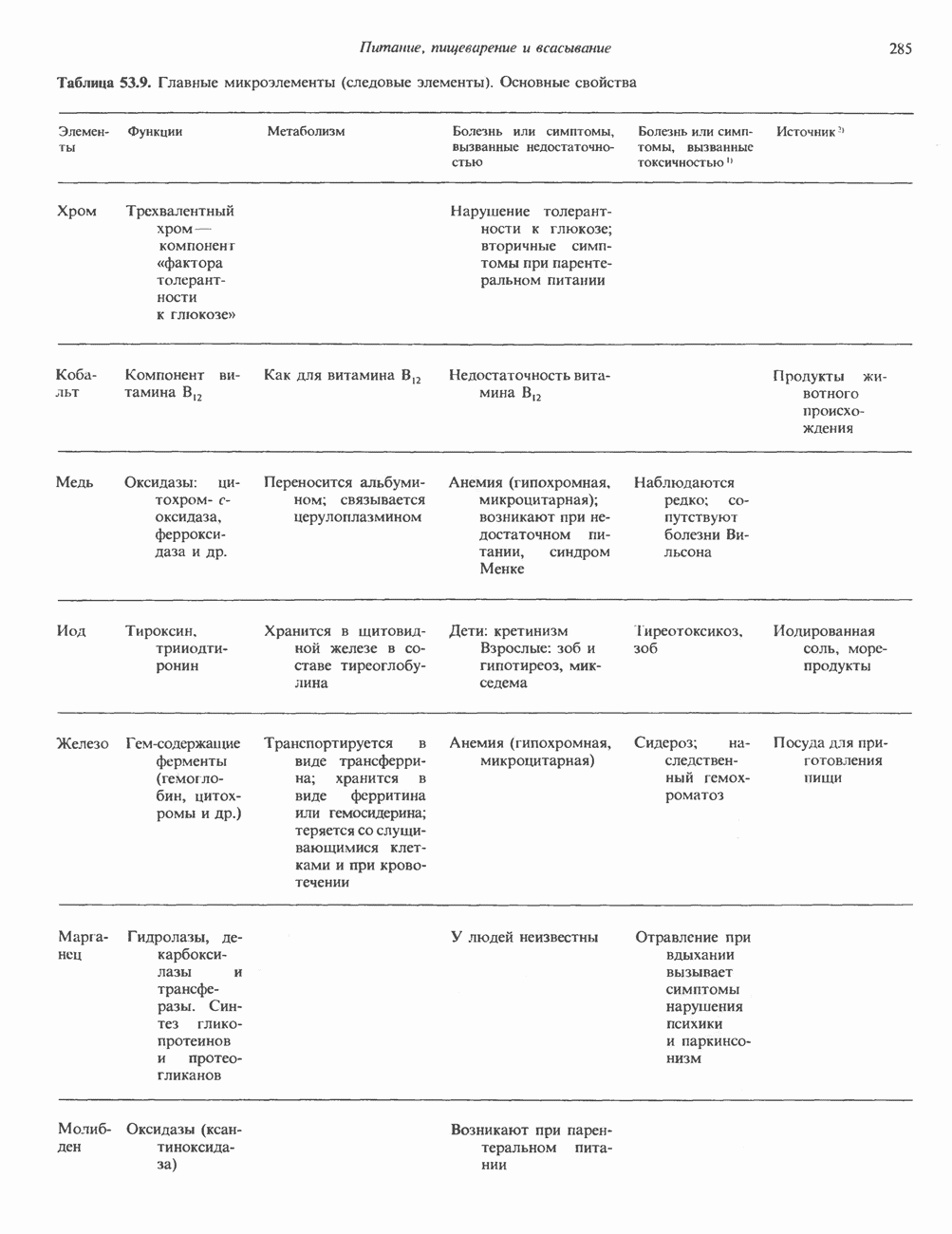
Рис. 38.3. Структура фибриллы хроматина диаметром в 10 нм, состоящей из дискообразных нуклеосом. Положение Н1-гистона не показано.
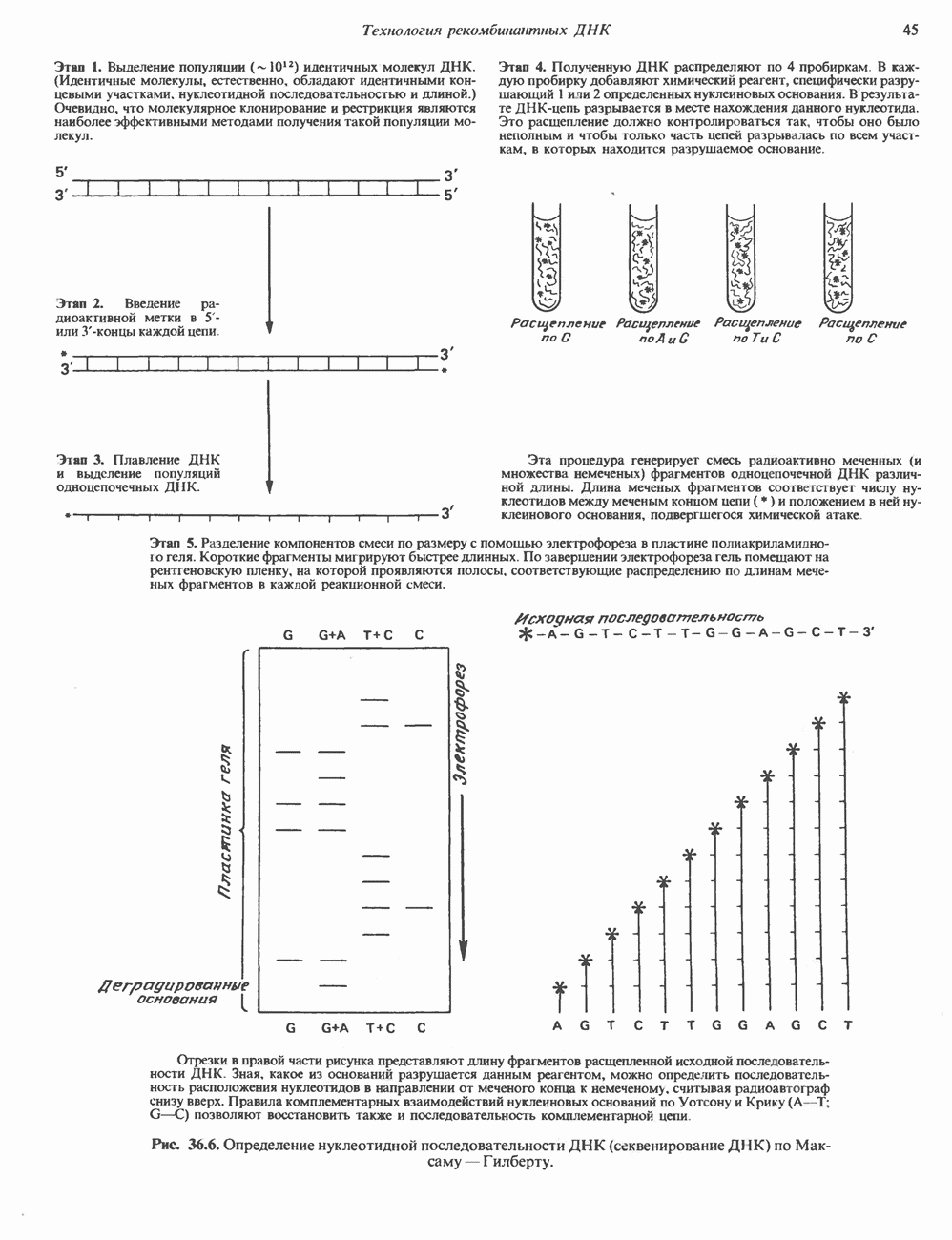
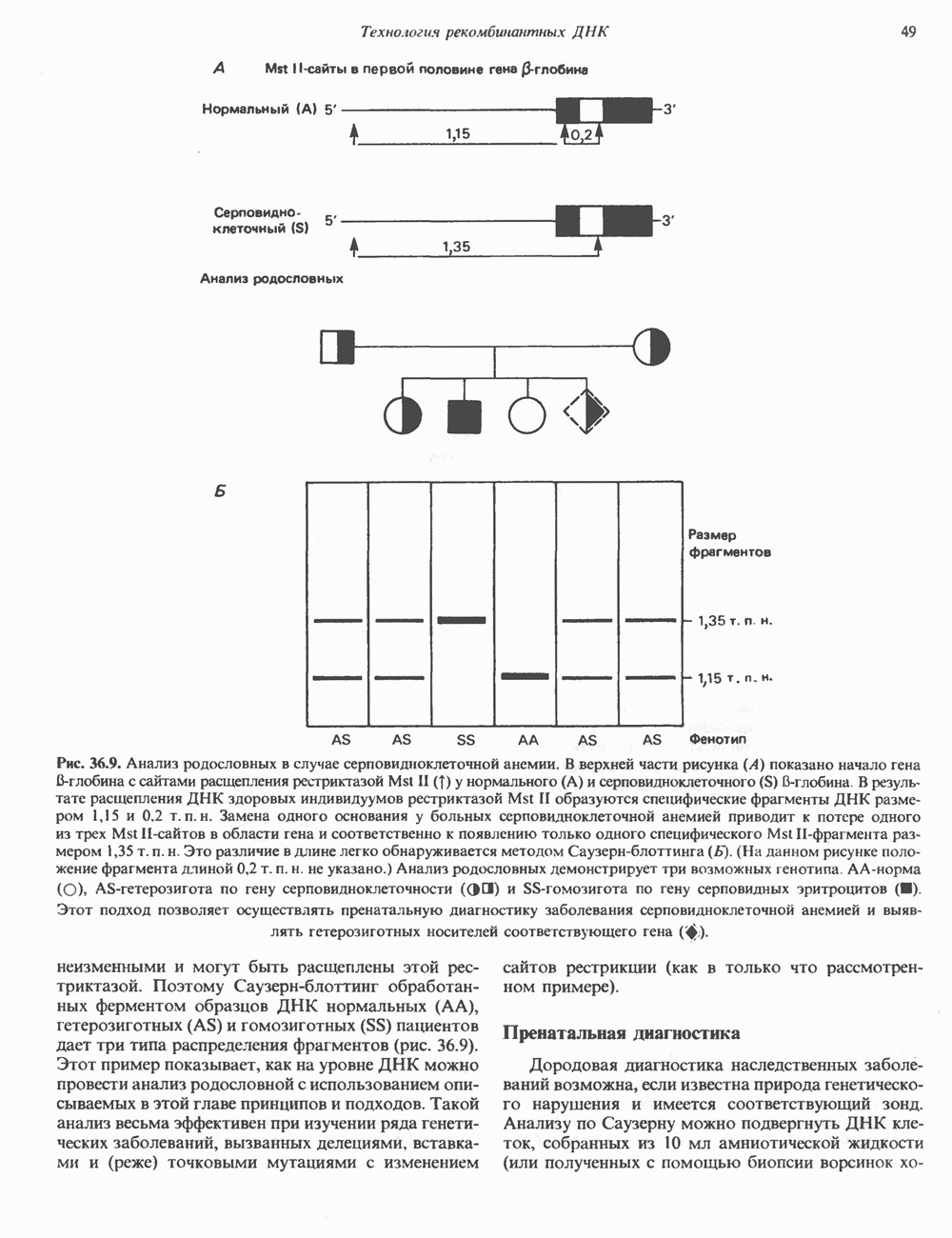
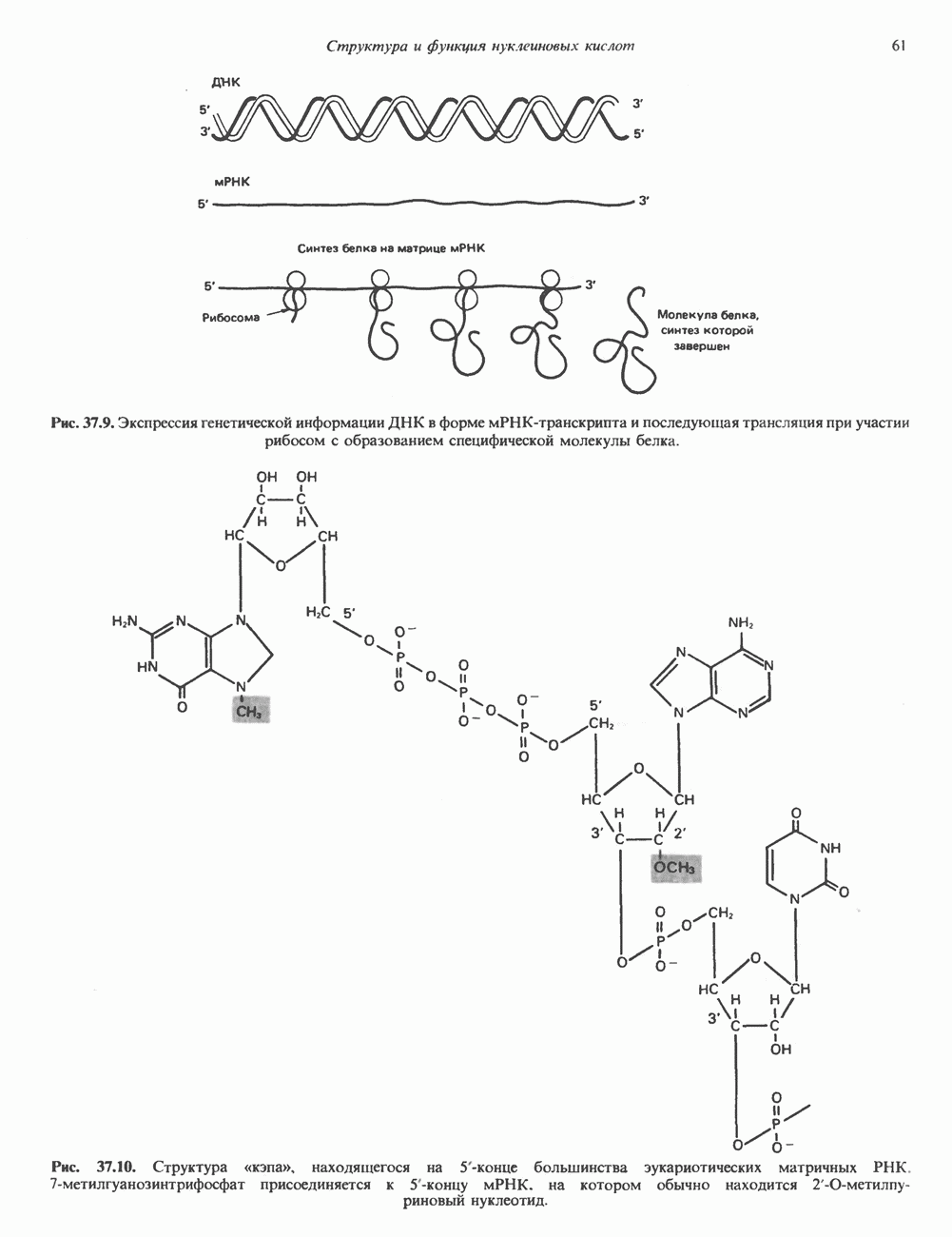
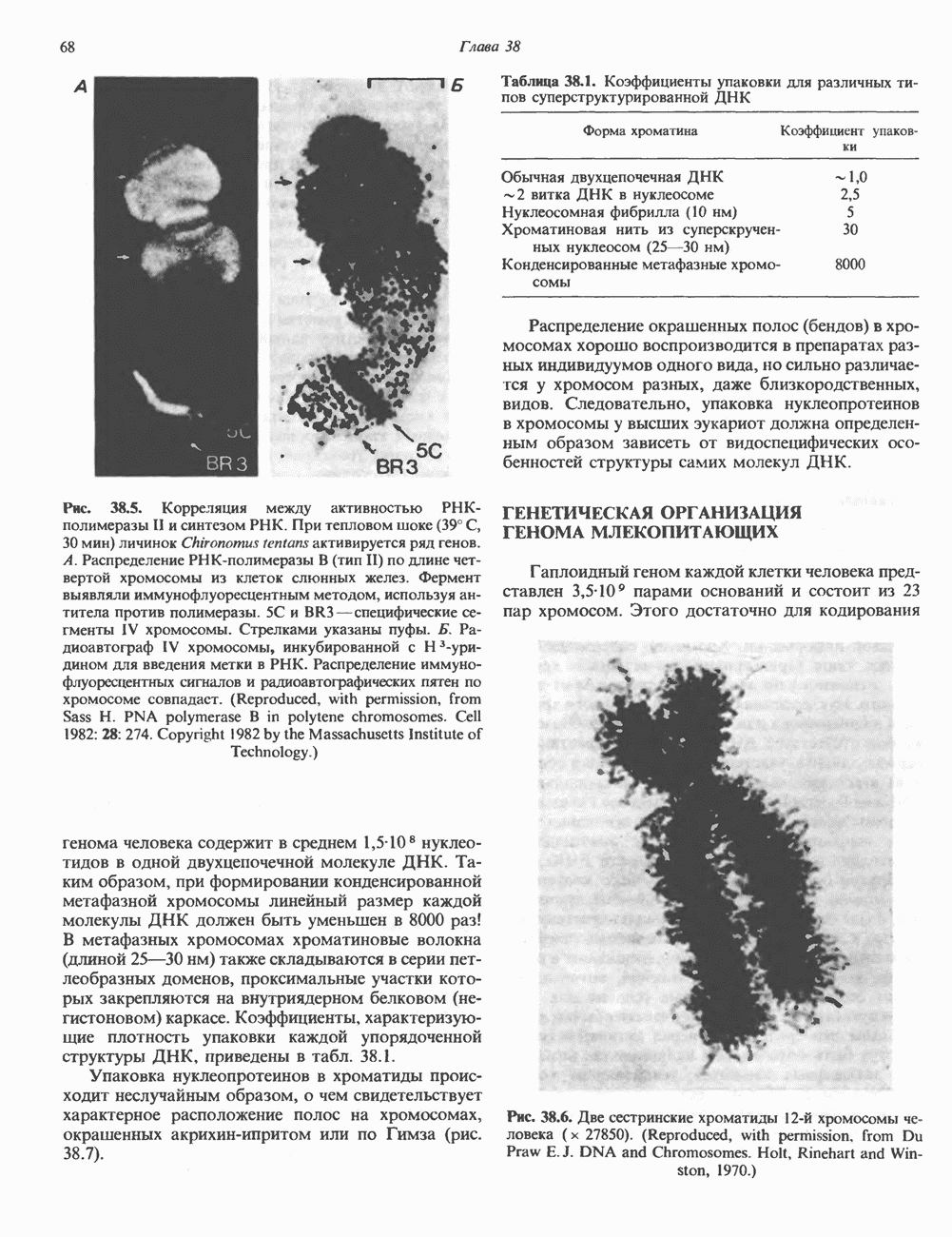
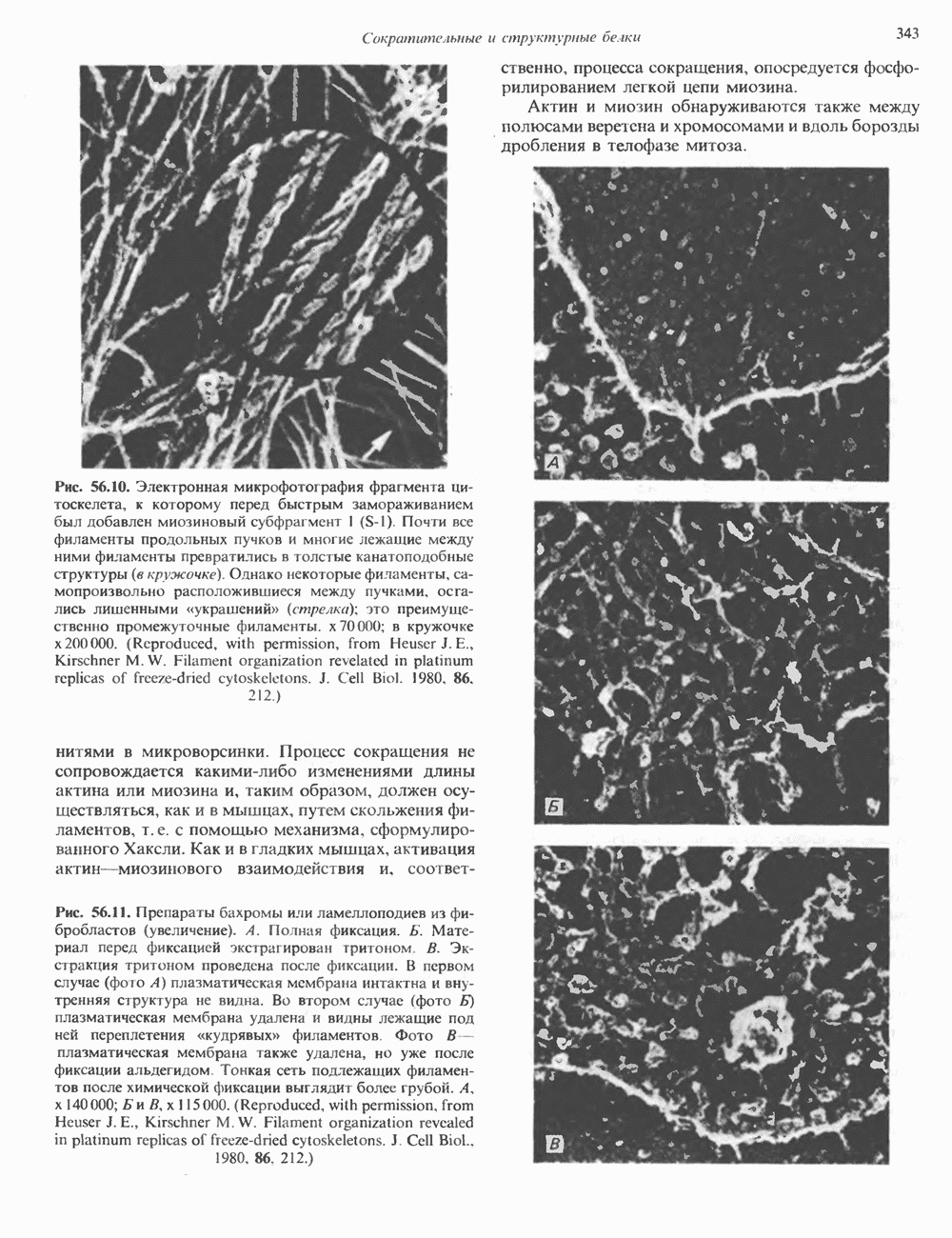
Рис. 38.4. Структура хроматинового волокна диаметром 30 нм, состоящего из суперскрученных фибрилл диаметром 10 нм. Ось волокна направлена перпендикулярно плоскости страницы. Активный хроматинКак правило, каждая клетка многоклеточного организма содержит одну и ту же генетическую информацию в виде одной и той же последовательности ДНК. Из этого следует, что различия между типами клеток данного организма должны объясняться дифференцированной экспрессией общей генетической информации. Хроматин, содержащий активные гены (транскрипционно-активный хроматин), отличается по некоторым признакам от неактивного. Нуклеосомная структура активного хроматина видоизменена или, в особо активных областях, вообще отсутствует. ДНК в активном хроматине содержит длинные участки (около 100000 пар оснований), чувствительные к действию нуклеаз (например, ДНКазы I). Чувствительность к ДНКазе I указывает на возможность транскрипции и в некоторых случаях коррелирует с отсутствием Внутри большой области активного хроматина обнаружены короткие участки (100—300 нуклеотидов) с еще более высокой (на порядок) чувствительностью к ДНКазе I. Эти, так называемые гиперчувствительные сайты, по-видимому, возникают в результате конформационных изменений, которые создают особенно благоприятные условия для действия нуклеазы на ДНК. Такие участки обычно локализованы непосредственно перед активным геном и могут быть обусловлены наличием так называемых энхансерных элементов, усиливающих транскрипцию (см. гл. 39 и 41). Есть основания считать, что во многих случаях транскрипционная активность гена связана с наличием в хроматине гиперчувствительного к ДНКазе сайта, непосредственно прилегающего к началу гена. Вероятно, такие сайты обеспечивают доступность кодирующей цепи для белков, участвующих в процессе транскрипции. Электронная микроскопия интерфазного ядра показывает, что транскрипционно-неактивный хроматин (гетерохроматин) плотно упакован, и потому соответствующие области интенсивно окрашиваются. Участки транскрипционно-активного хроматина (эухроматина) имеют более слабую окраску. В целом в ходе клеточного цикла млекопитающих (см. ниже) эухроматин реплицируется раньше, чем гетерохроматин. Существуют два типа гетерохроматина: конститутивный гетерохроматин и факультативный гетерохроматин. Конститутивный гетерохроматин всегда конденсирован и, следовательно, неактивен. Конститутивный гетерохроматин найден в областях, близких к центромерам и концевым участкам (теломерам) хромосом. Факультативный гетерохроматин временами конденсирован, а временами разуплотнен, активно транскрибируется и таким образом оказывается сходным с эухроматином. Из двух Некоторые клетки насекомых, например Chironomus, содержат гигантские хромосомы, образовавшиеся в результате нерасхождения дочерних хроматид после прохождения Транскрипционно-активные области таких политенных хромосом отличаются особенно отчетливой разуплотненностью — они образуют так называемые «пуфы», в которых, как установлено, локализуются ферменты системы транскрипции и происходит синтез РНК (рис. 38.5). ХромосомыВ метафазе хромосомы млекопитающих обладают двулучевой симметрией второго порядка и состоят из идентичных сестринских хроматид, соединенных в центромере, положение которой характерно для каждой хромосомы (рис. 38.6). Каждая сестринская хроматида содержит одну двухцепочечную молекулу ДНК. В интерфазе упаковка молекулы ДНК менее плотная, чем в метафазе. Метафазные хромосомы транскрипционно-неактивны. Гаплоидный геном человека состоит из
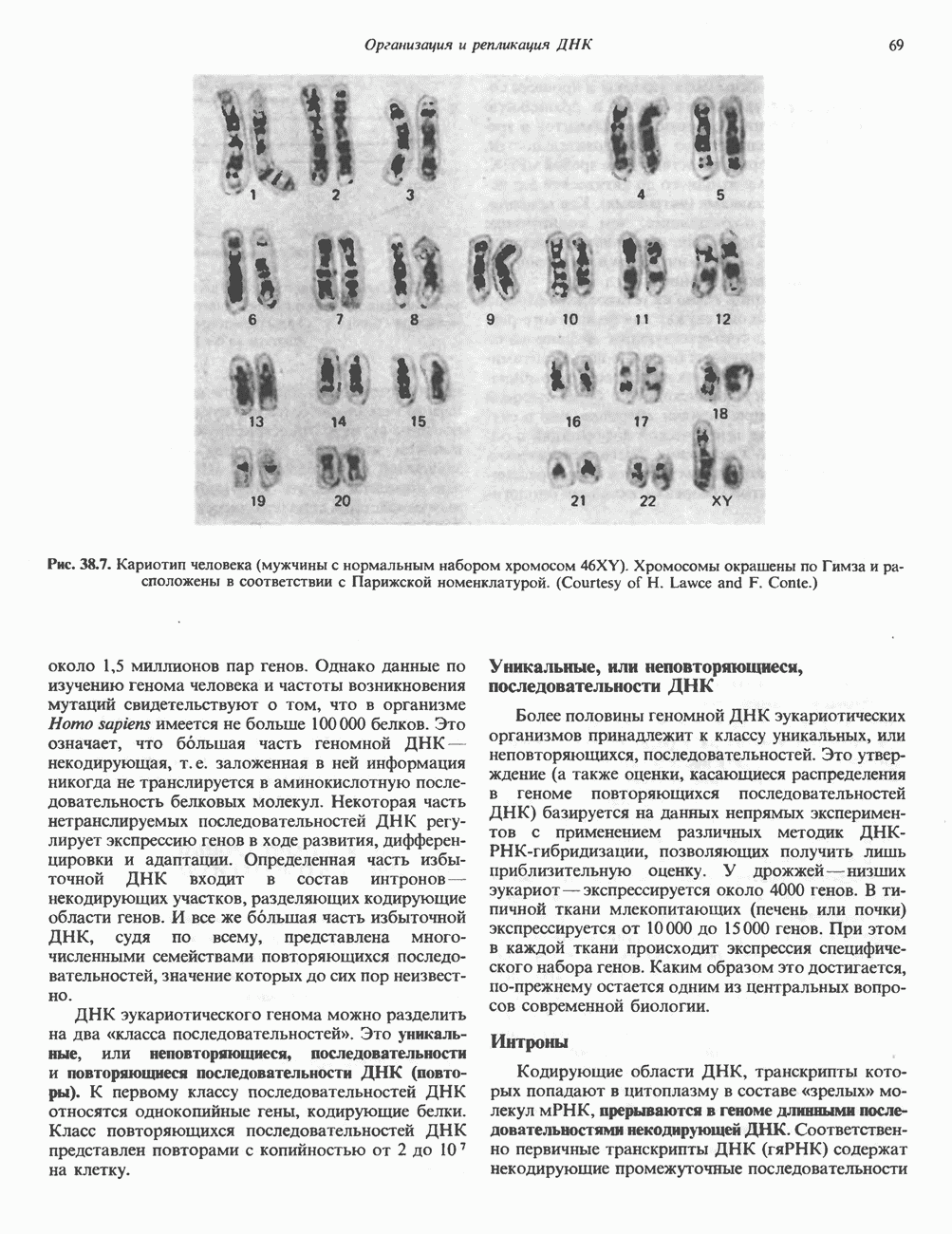
Рис. 38.5. Корреляция между активностью РНК-полимеразы II и синтезом РНК. При тепловом шоке (39° С, 30 мин) личинок Chironomus tentans активируется ряд генов. А. Распределение РНК-полимеразы В (тип И) по длине четвертой хромосомы из клеток слюнных желез. Фермент выявляли иммунофлуоресцентным методом, используя антитела против полимеразы. генома человека содержит в среднем Упаковка нуклеопротеинов в хроматиды происходит неслучайным образом, о чем свидетельствует характерное расположение полос на хромосомах, окрашенных акрихинипритом или по Гимза (рис. 38.7). Таблица 38.1. Коэффициенты упаковки для различных типов суперструктурированной ДНК
Распределение окрашенных полос (бендов) в хромосомах хорошо воспроизводится в препаратах разных индивидуумов одного вида, но сильно различается у хромосом разных, даже близкородственных, видов. Следовательно, упаковка нуклеопротеинов в хромосомы у высших эукариот должна определенным образом зависеть от видоспецифических особенностей структуры самих молекул ДНК.
|
 |
Оглавление
|














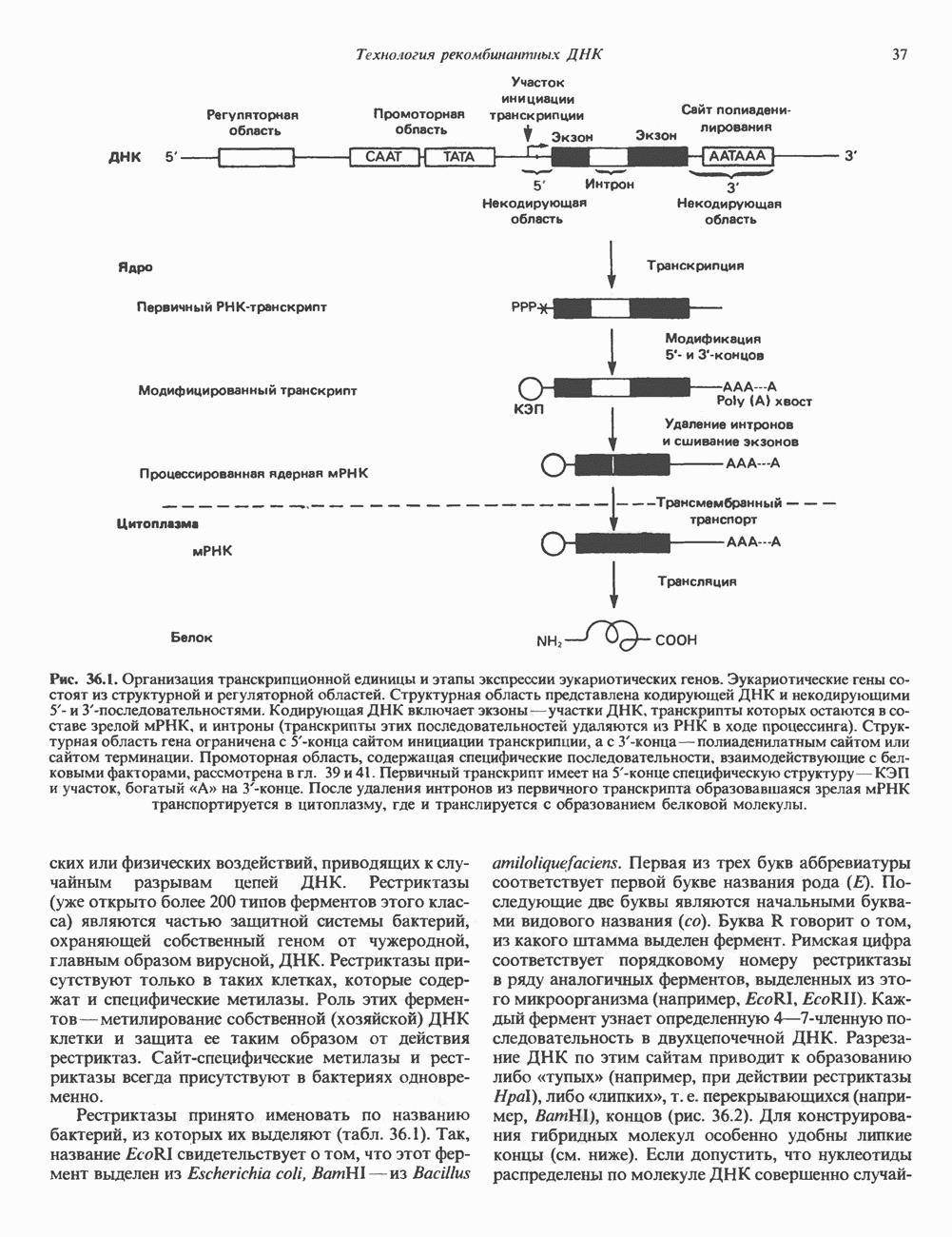












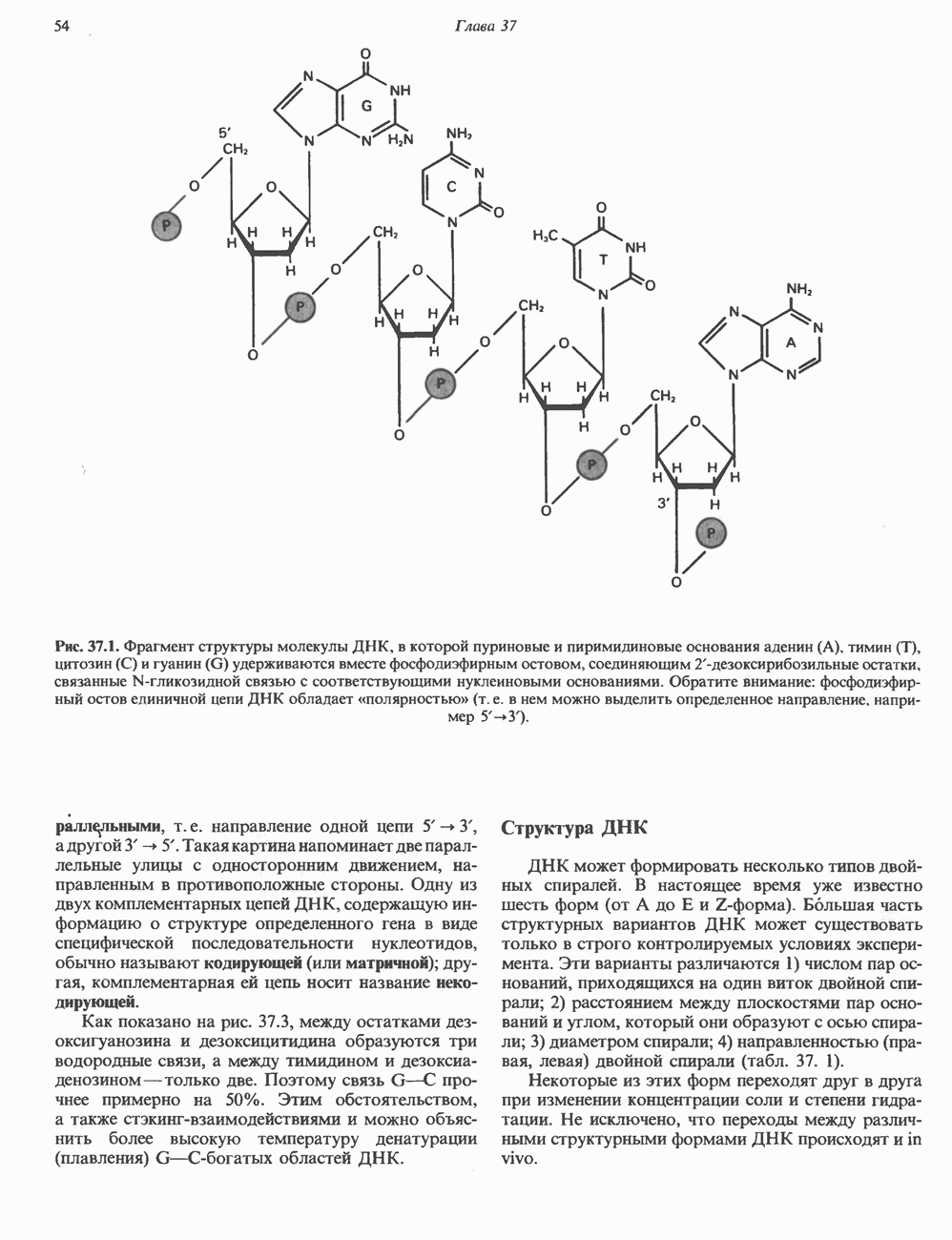
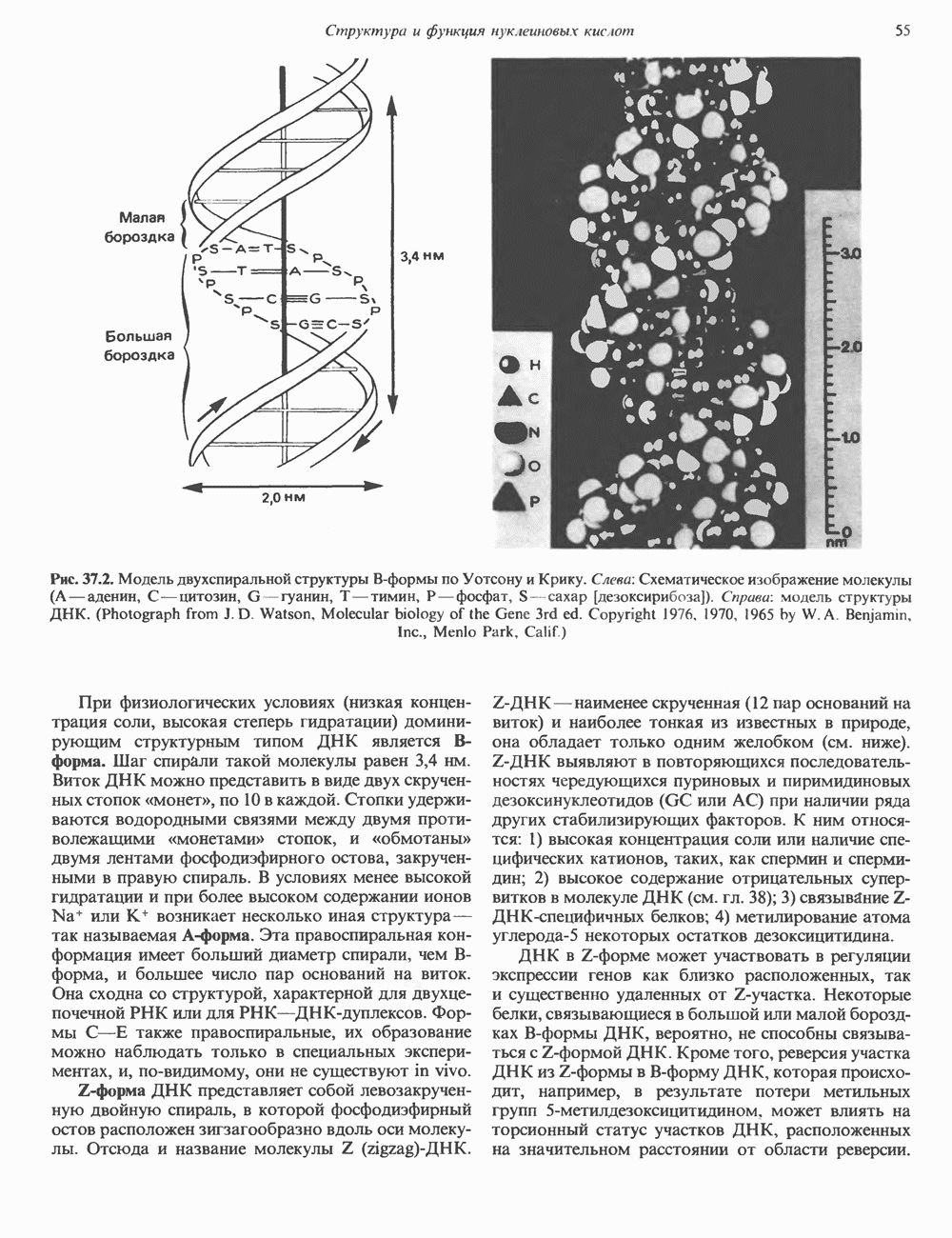
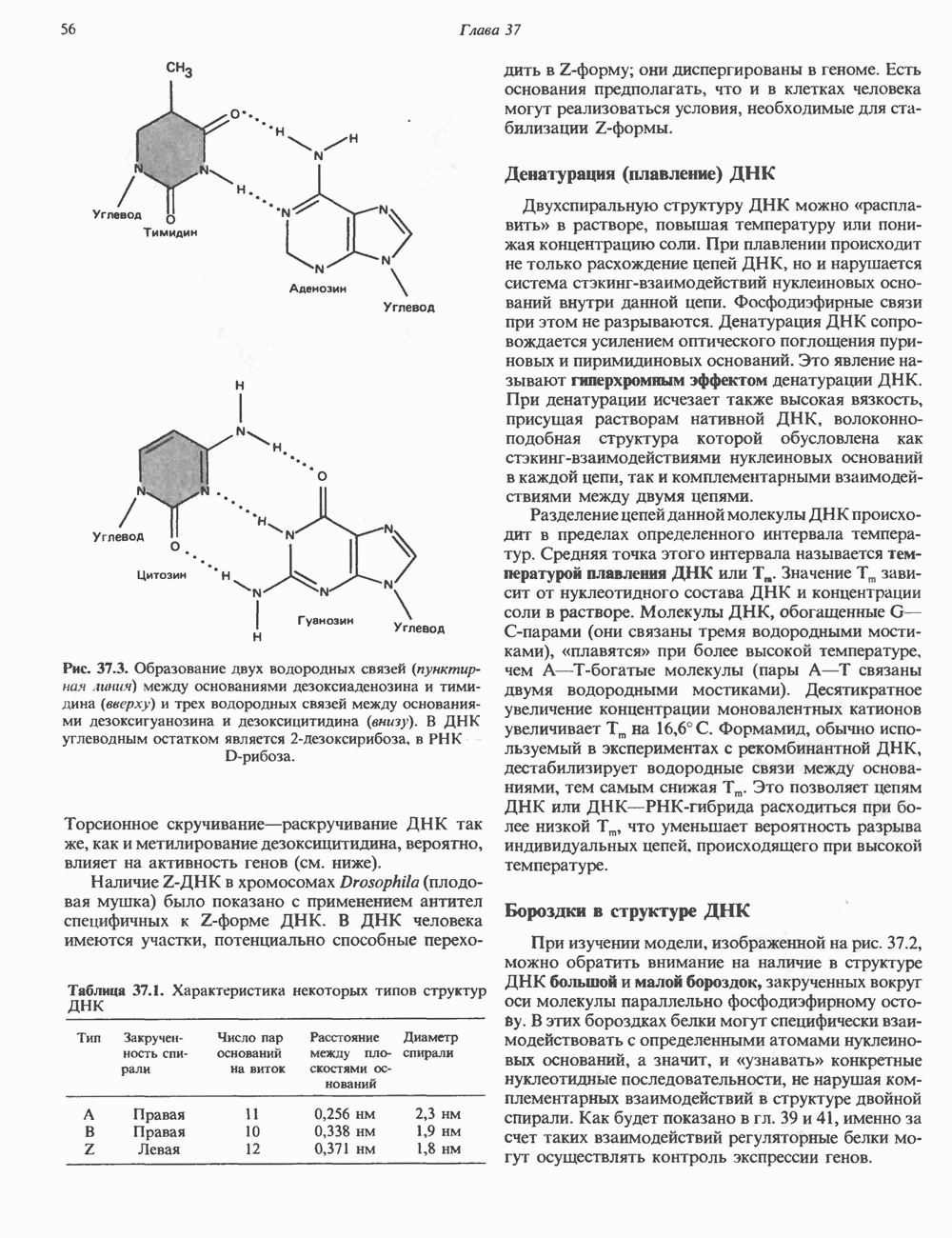

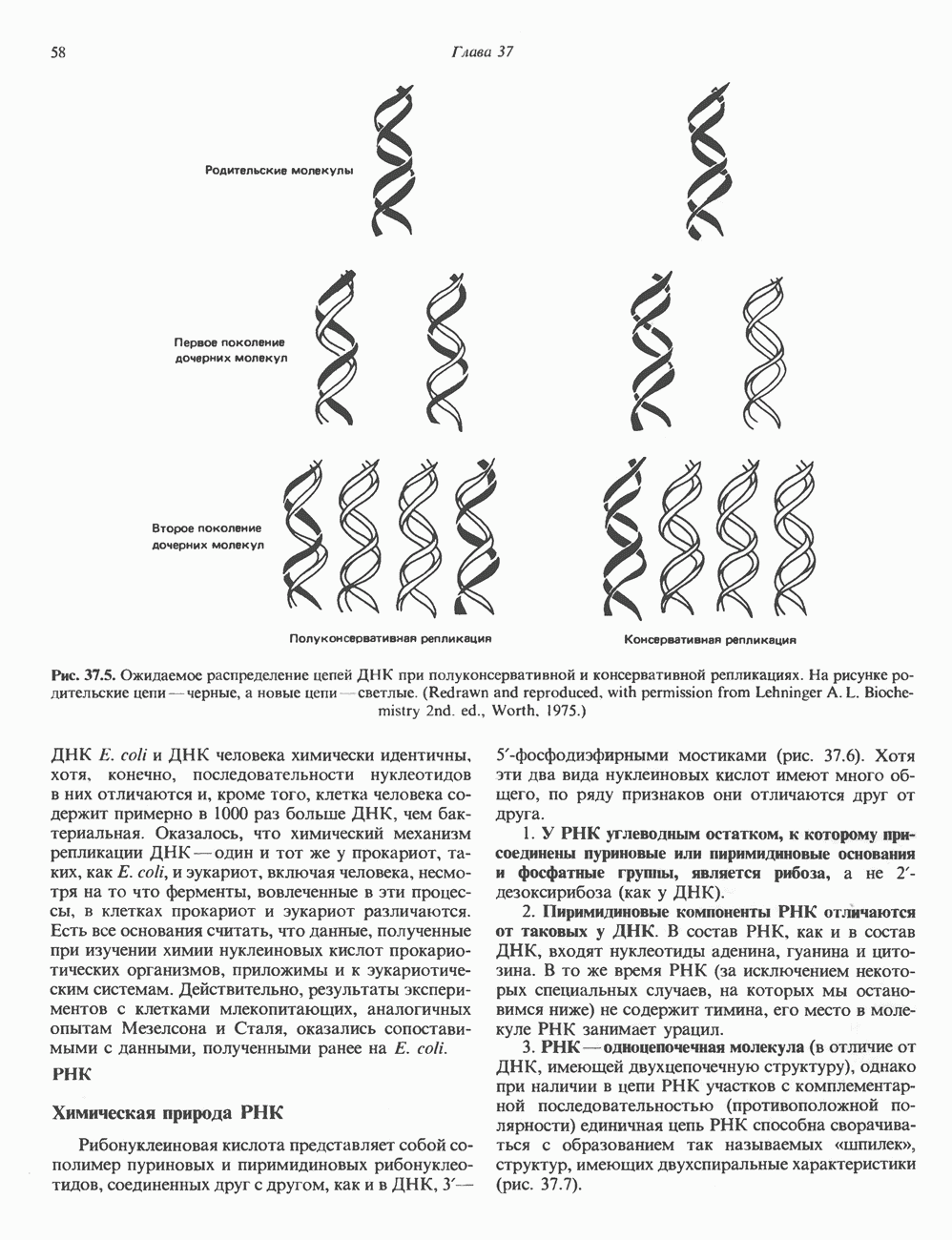















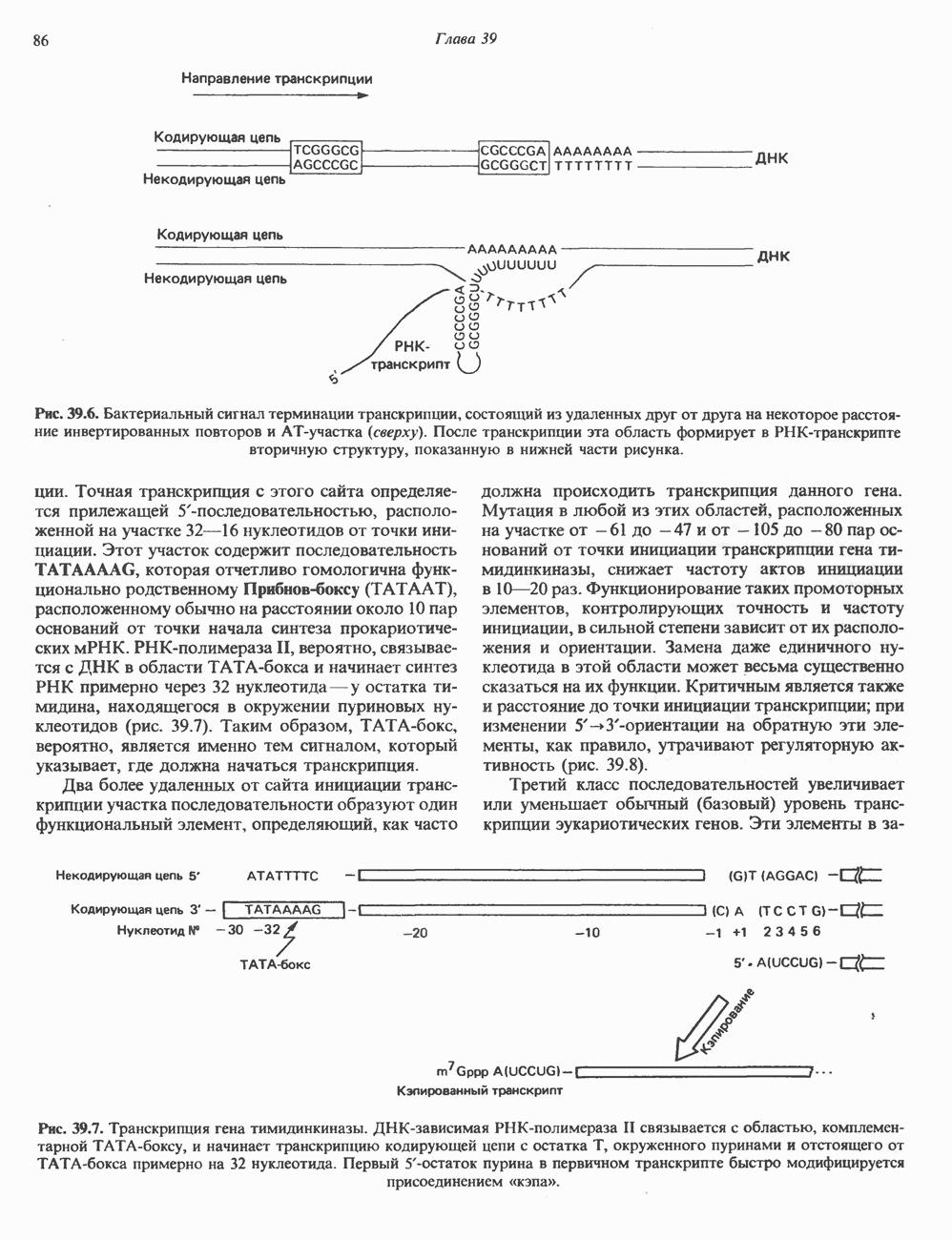
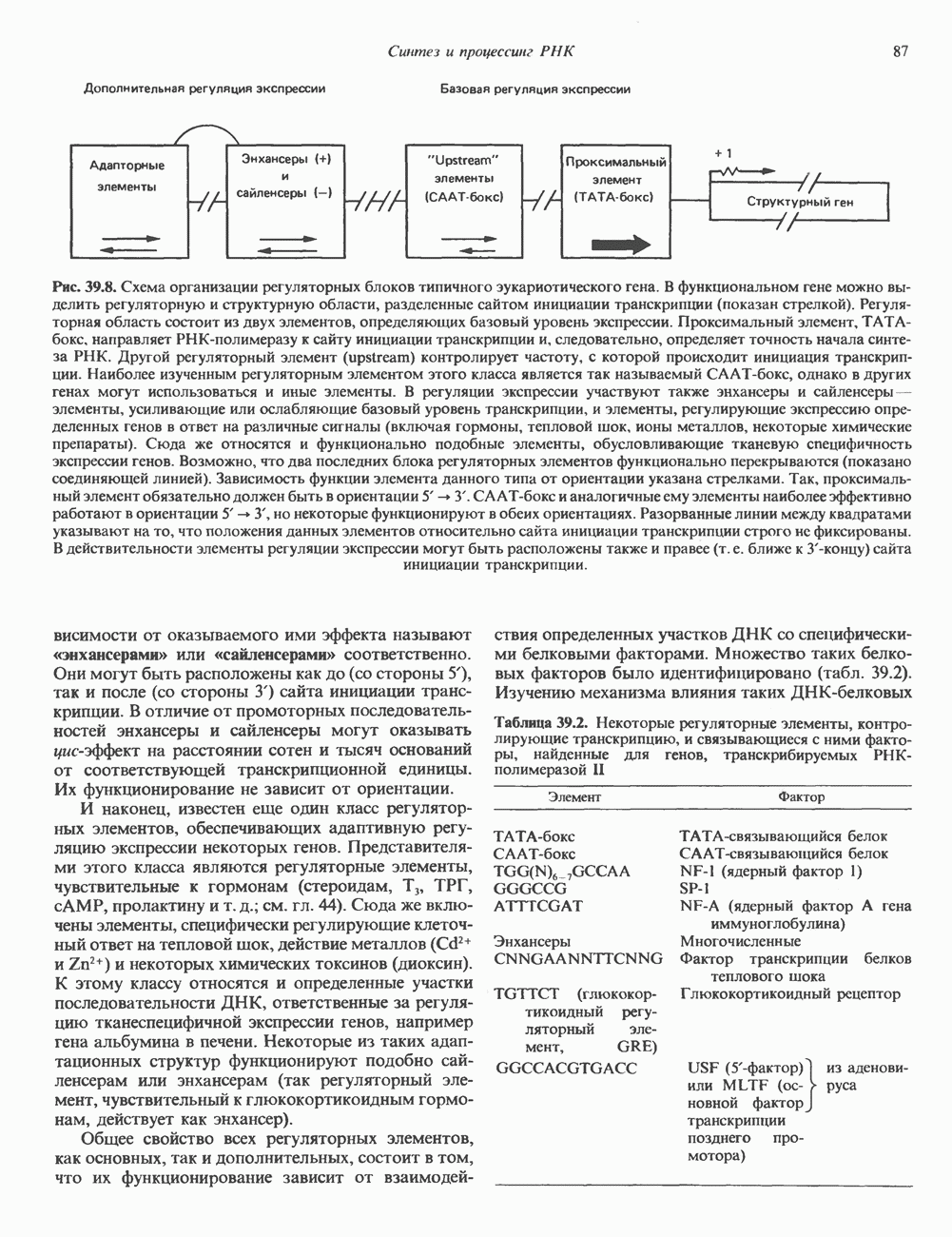

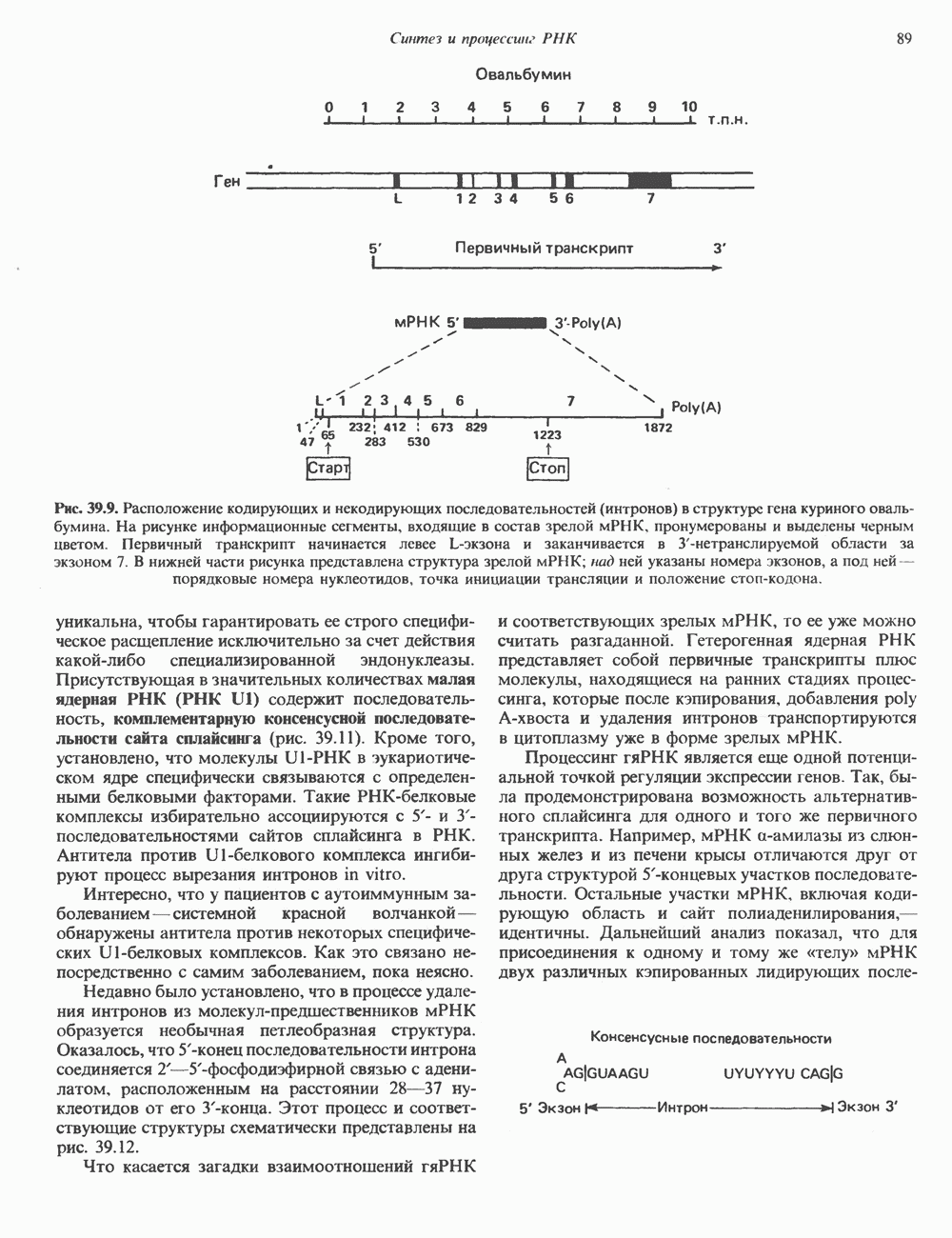
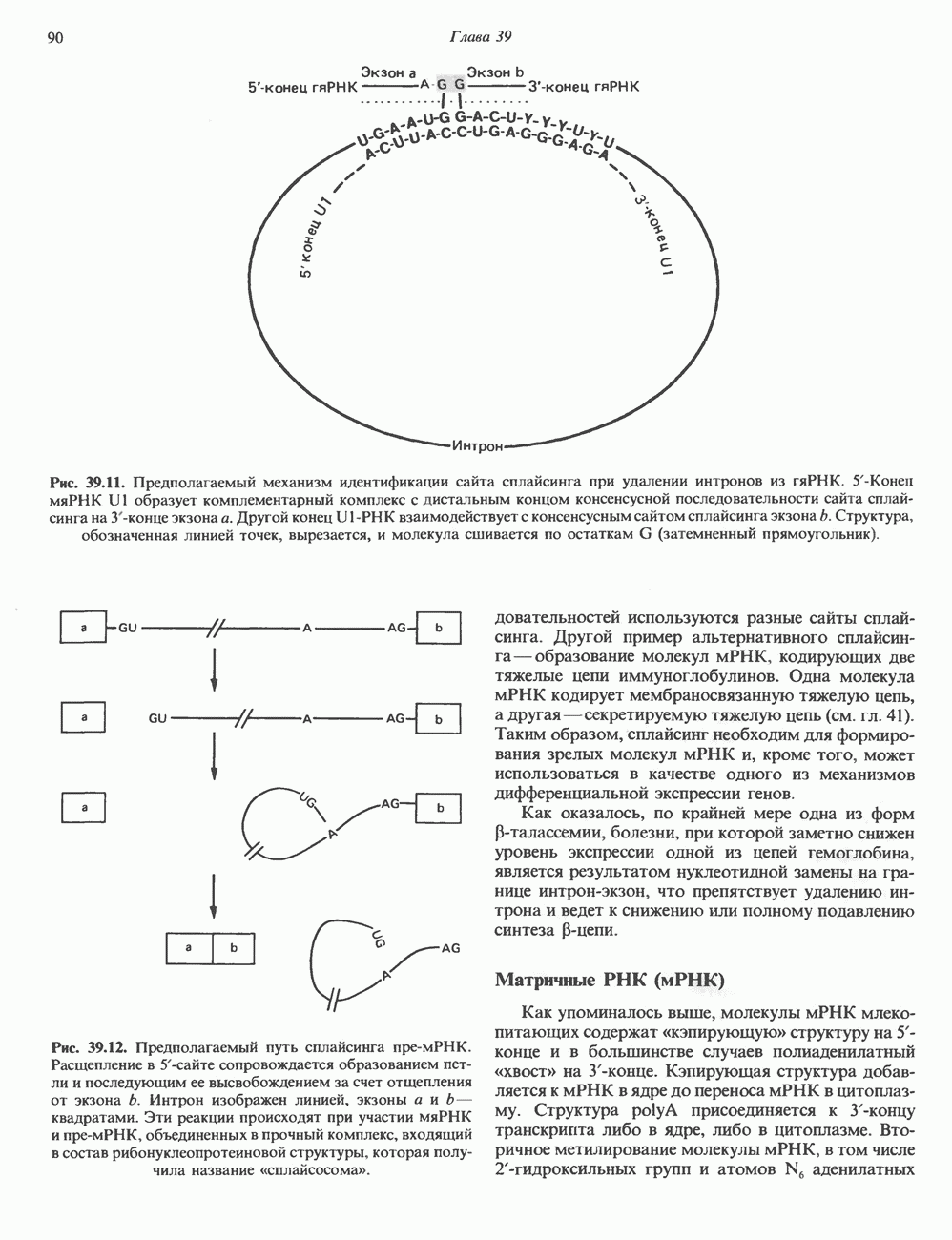

























































































































































































































 и двух
и двух  Гистоновая сердцевина нуклеосомы взаимодействует с внутренней поверхностью суперспирали и не выступает за ее пределы.
Гистоновая сердцевина нуклеосомы взаимодействует с внутренней поверхностью суперспирали и не выступает за ее пределы.
 способен придавать ДНК
способен придавать ДНК

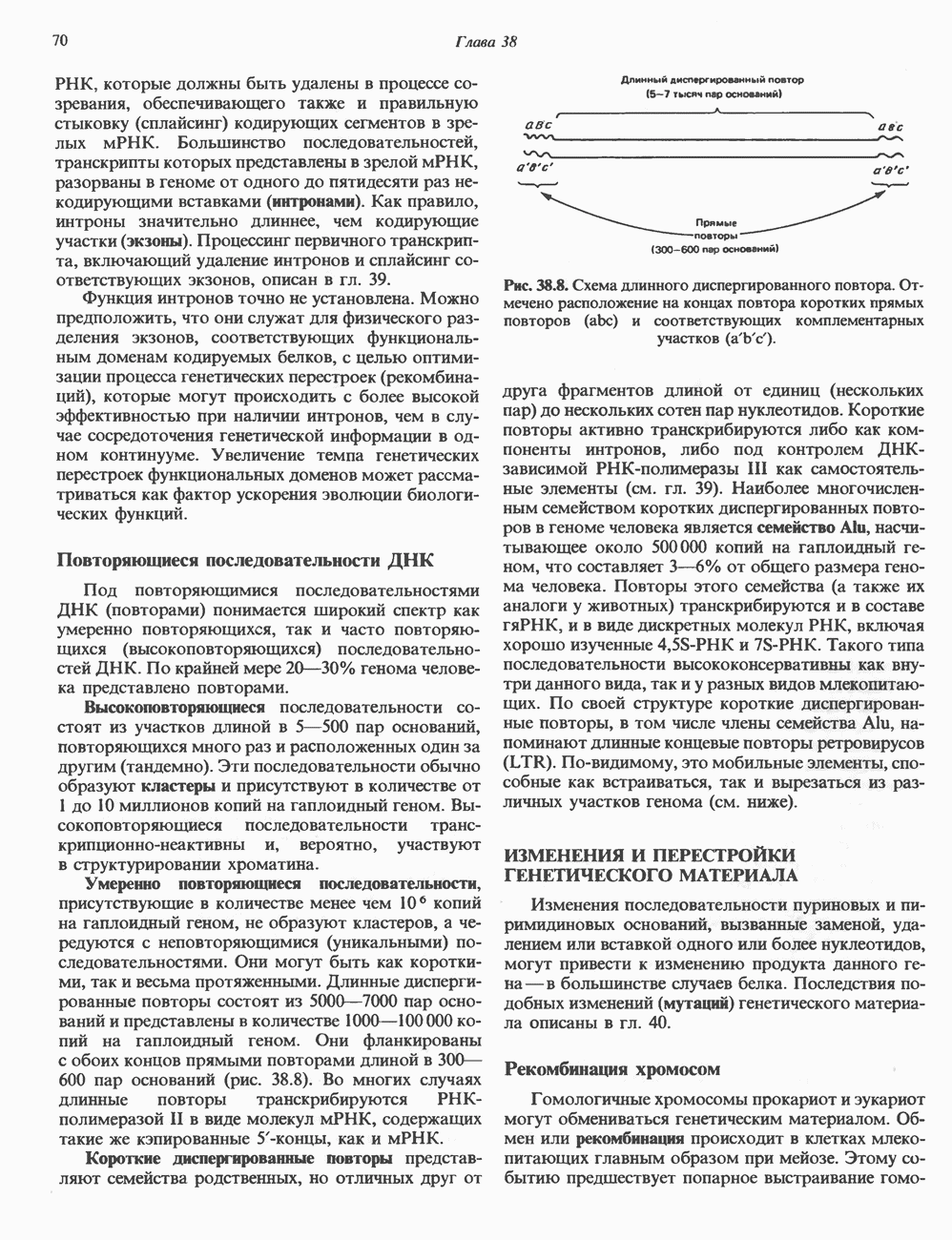
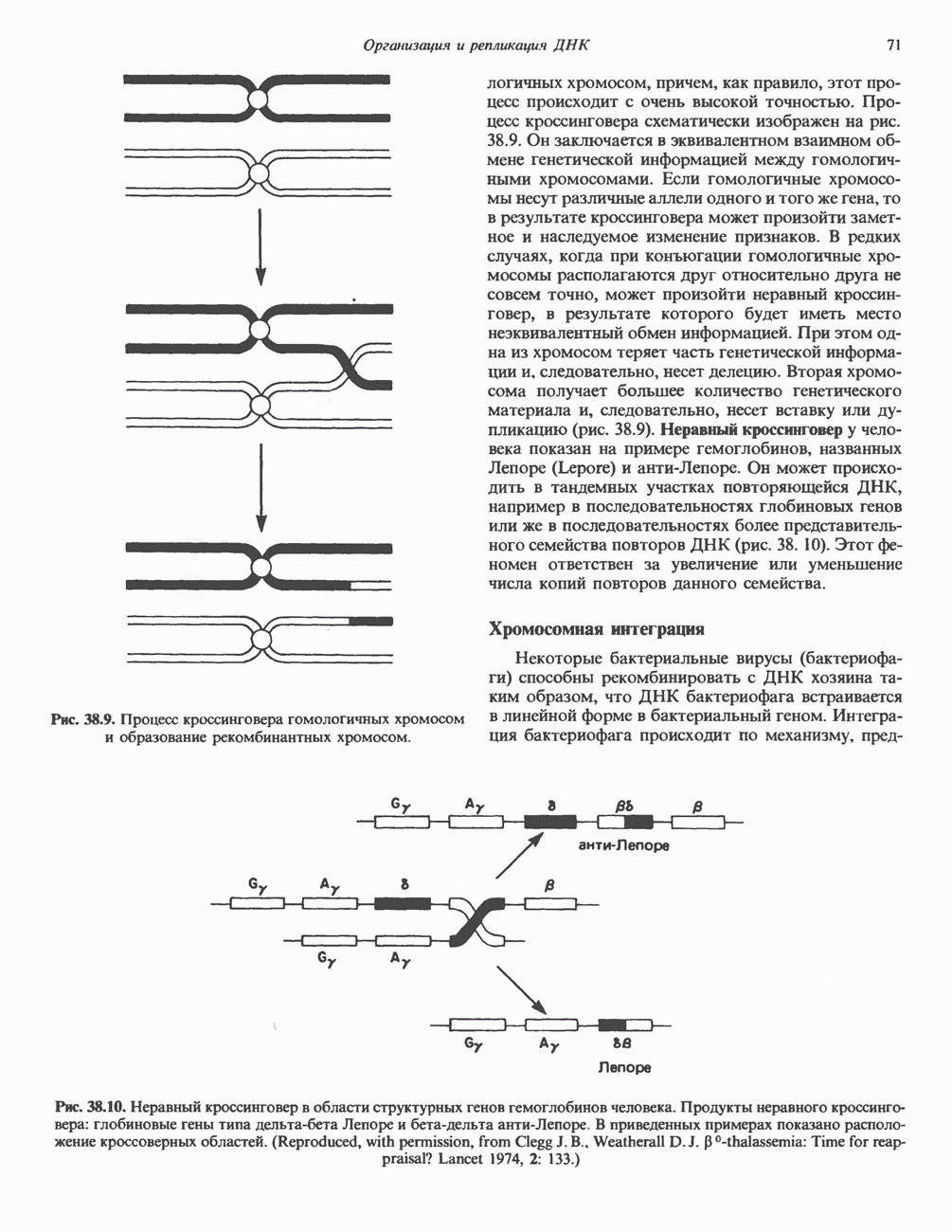
 стабилизируют первичную частицу и прочно соединяют два полувитка ДНК, ранее слабо связанных с Н32-Н42-тетрамером. Таким образом, 1,75 супервитков ДНК закручиваются вокруг гистонового октамера и образуют нуклеосомный кор (или минимальную нуклеосому), который «маскирует» 146 пар оснований ДНК (рис. 38.2). ДНК по ходу спирали вокруг октамера контактирует с гистонами в следующем порядке:
стабилизируют первичную частицу и прочно соединяют два полувитка ДНК, ранее слабо связанных с Н32-Н42-тетрамером. Таким образом, 1,75 супервитков ДНК закручиваются вокруг гистонового октамера и образуют нуклеосомный кор (или минимальную нуклеосому), который «маскирует» 146 пар оснований ДНК (рис. 38.2). ДНК по ходу спирали вокруг октамера контактирует с гистонами в следующем порядке:

 -гистоны, по всей вероятности, стабилизируют структуру волокна, но их расположение так же, как и длина спейсерных участков ДНК, точно не определены. Вероятно, нуклеосомы способны формировать еще ряд компактных суперструктур. Для того чтобы образовалась митотическая хромосома нормального размера, волокно диаметром 30 нм должно подвергнуться дополнительной компактизации с уменьшением результирующей длины еще в 100 раз (см. ниже).
-гистоны, по всей вероятности, стабилизируют структуру волокна, но их расположение так же, как и длина спейсерных участков ДНК, точно не определены. Вероятно, нуклеосомы способны формировать еще ряд компактных суперструктур. Для того чтобы образовалась митотическая хромосома нормального размера, волокно диаметром 30 нм должно подвергнуться дополнительной компактизации с уменьшением результирующей длины еще в 100 раз (см. ниже).


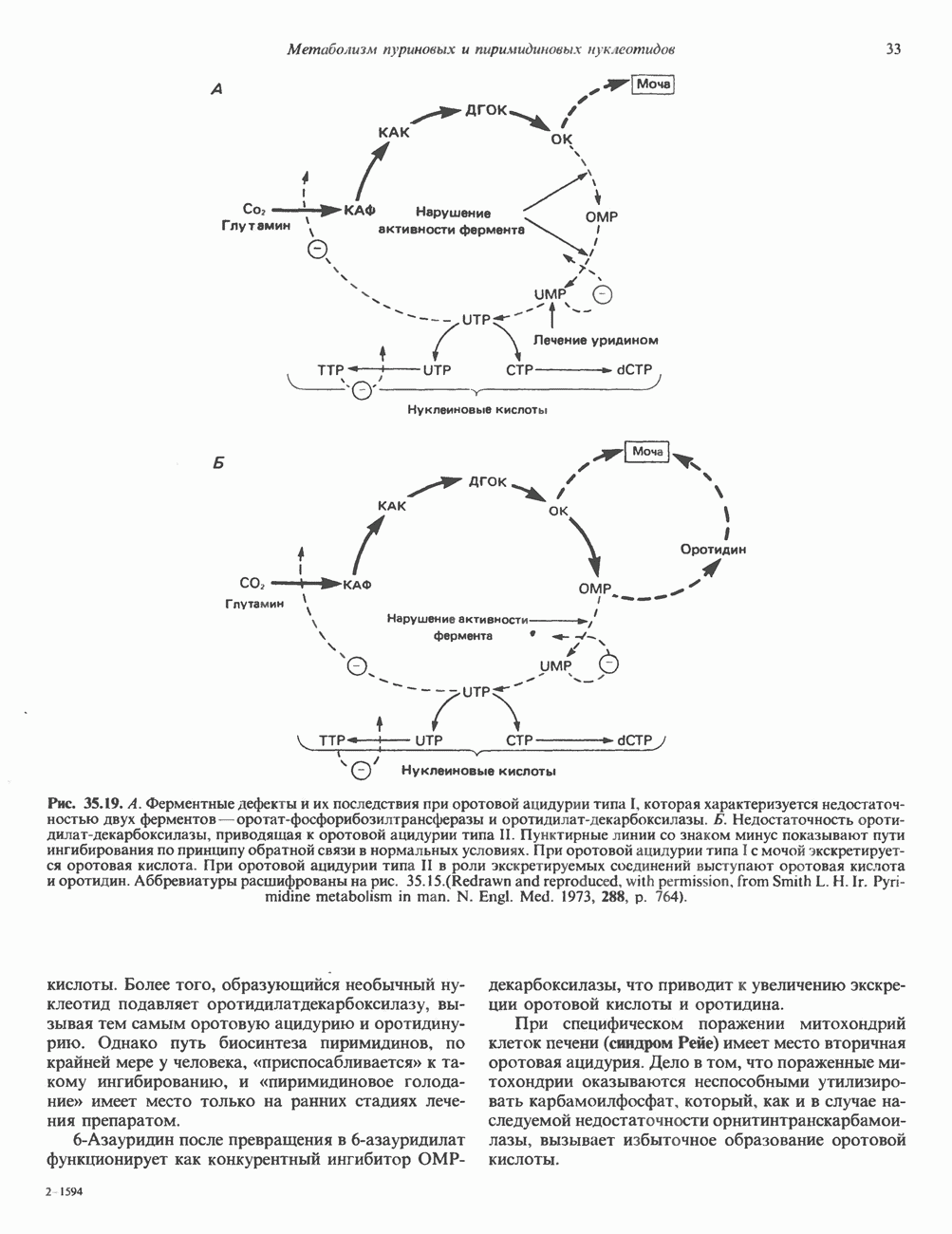
 -метилдезок-сицитидина в соответствующей области ДНК.
-метилдезок-сицитидина в соответствующей области ДНК.
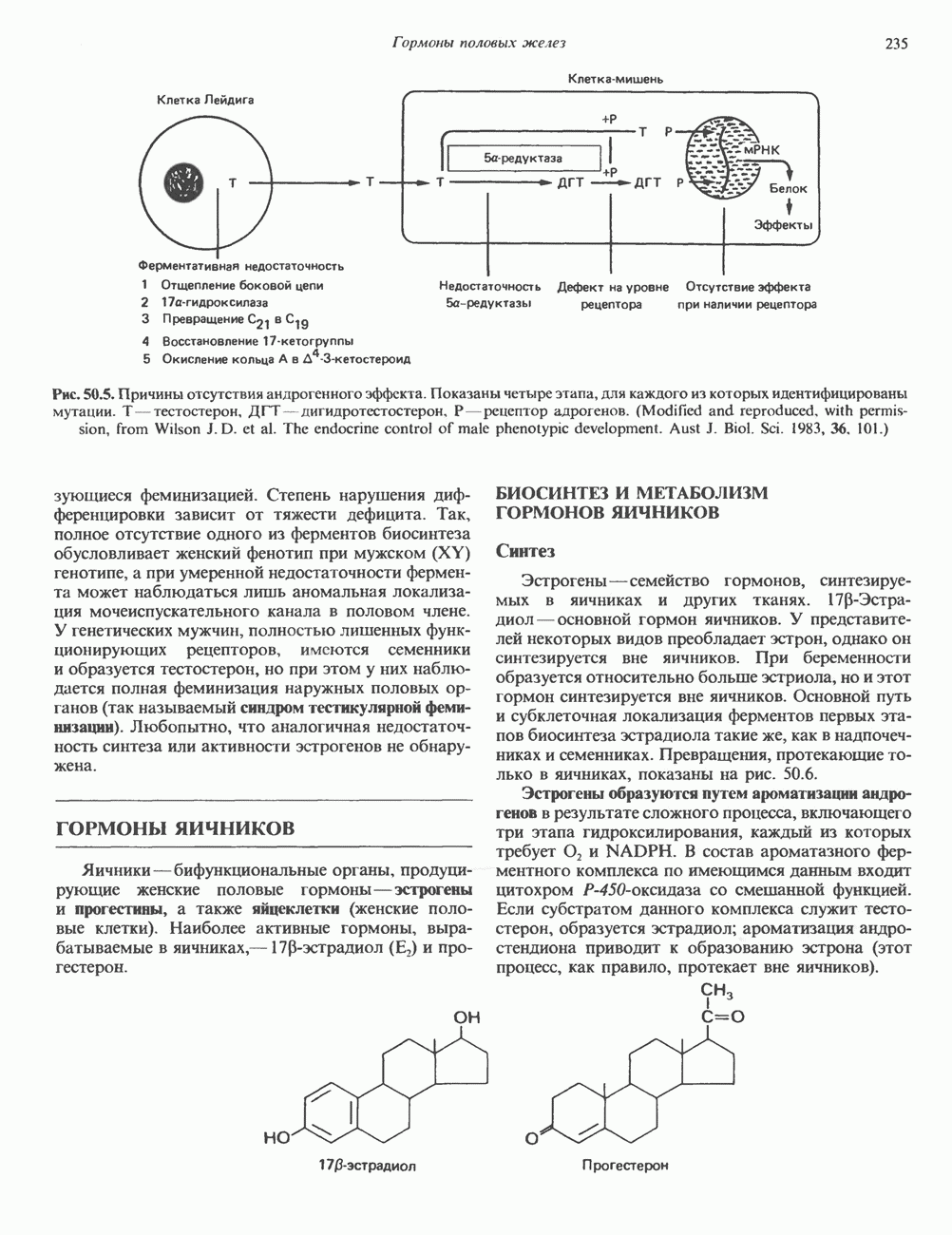
 -хромосом самок млекопитающих — одна практически полностью транскрипционно-неактивна, т. е. проявляет свойства гетерохроматина. Однако при гаметогенезе и на ранних стадиях эмбриогенеза гетерохроматиновая Х-хромосома становится транскрипционно-активной и, следовательно, проявляет свойства факультативного гетерохроматина.
-хромосом самок млекопитающих — одна практически полностью транскрипционно-неактивна, т. е. проявляет свойства гетерохроматина. Однако при гаметогенезе и на ранних стадиях эмбриогенеза гетерохроматиновая Х-хромосома становится транскрипционно-активной и, следовательно, проявляет свойства факультативного гетерохроматина.
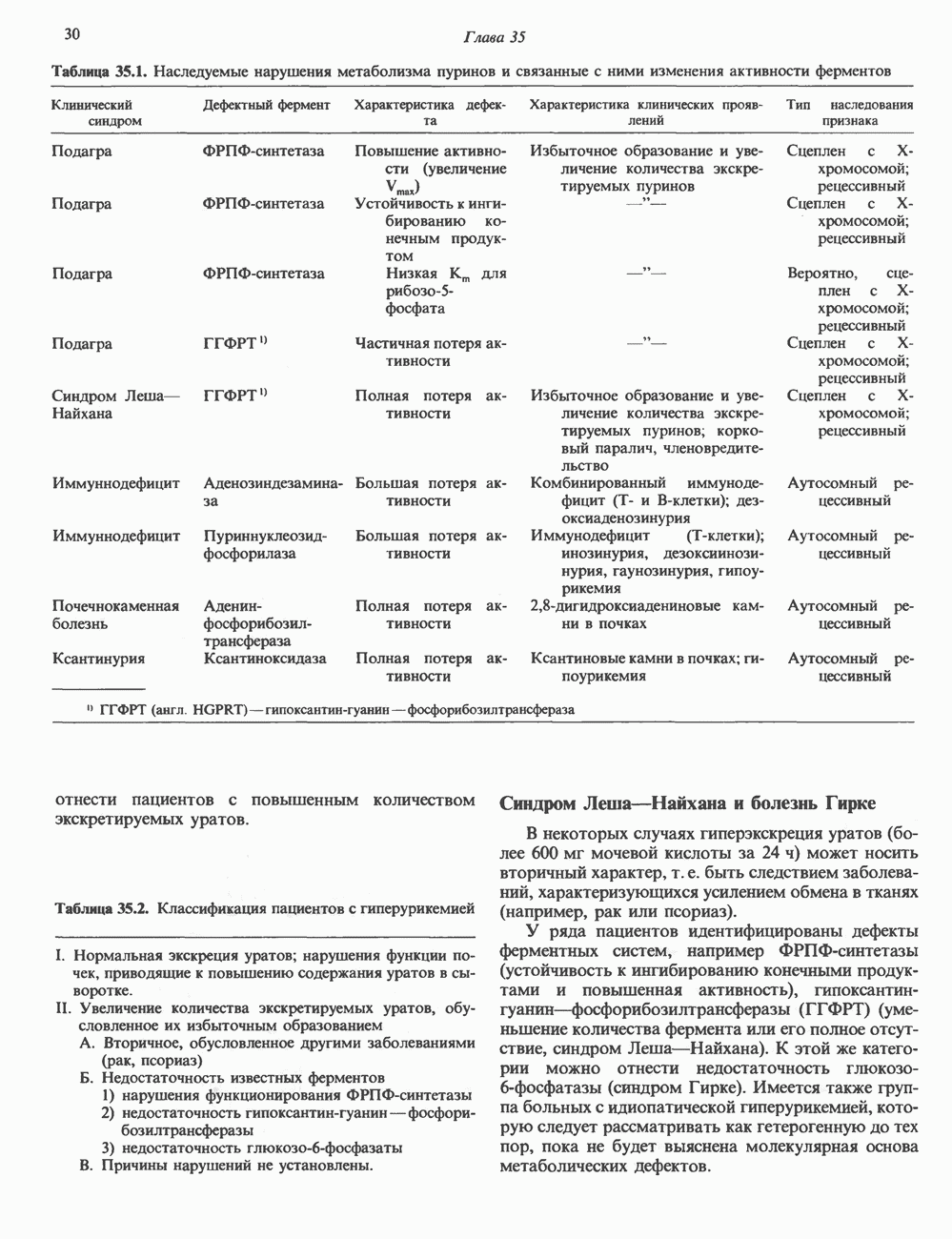
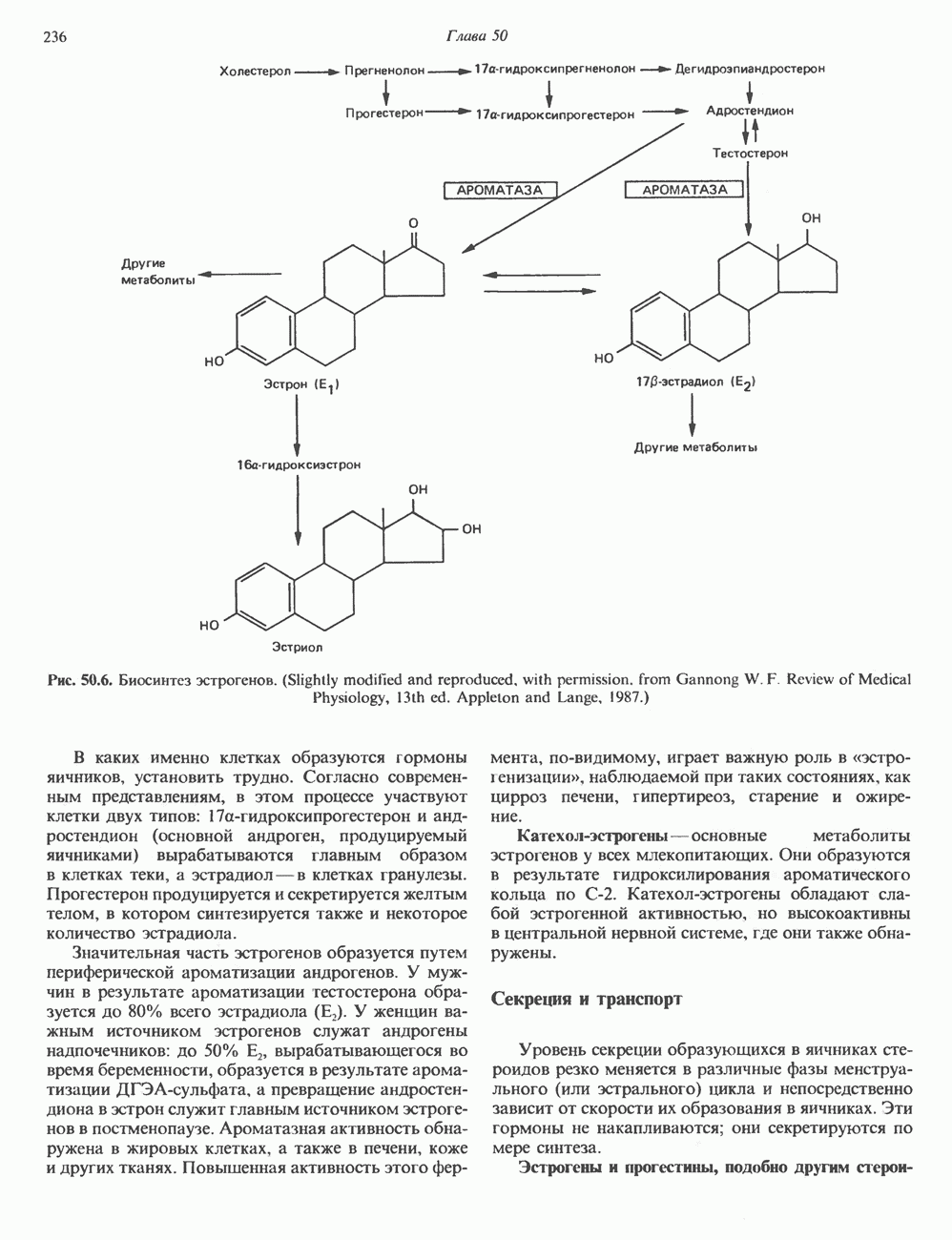
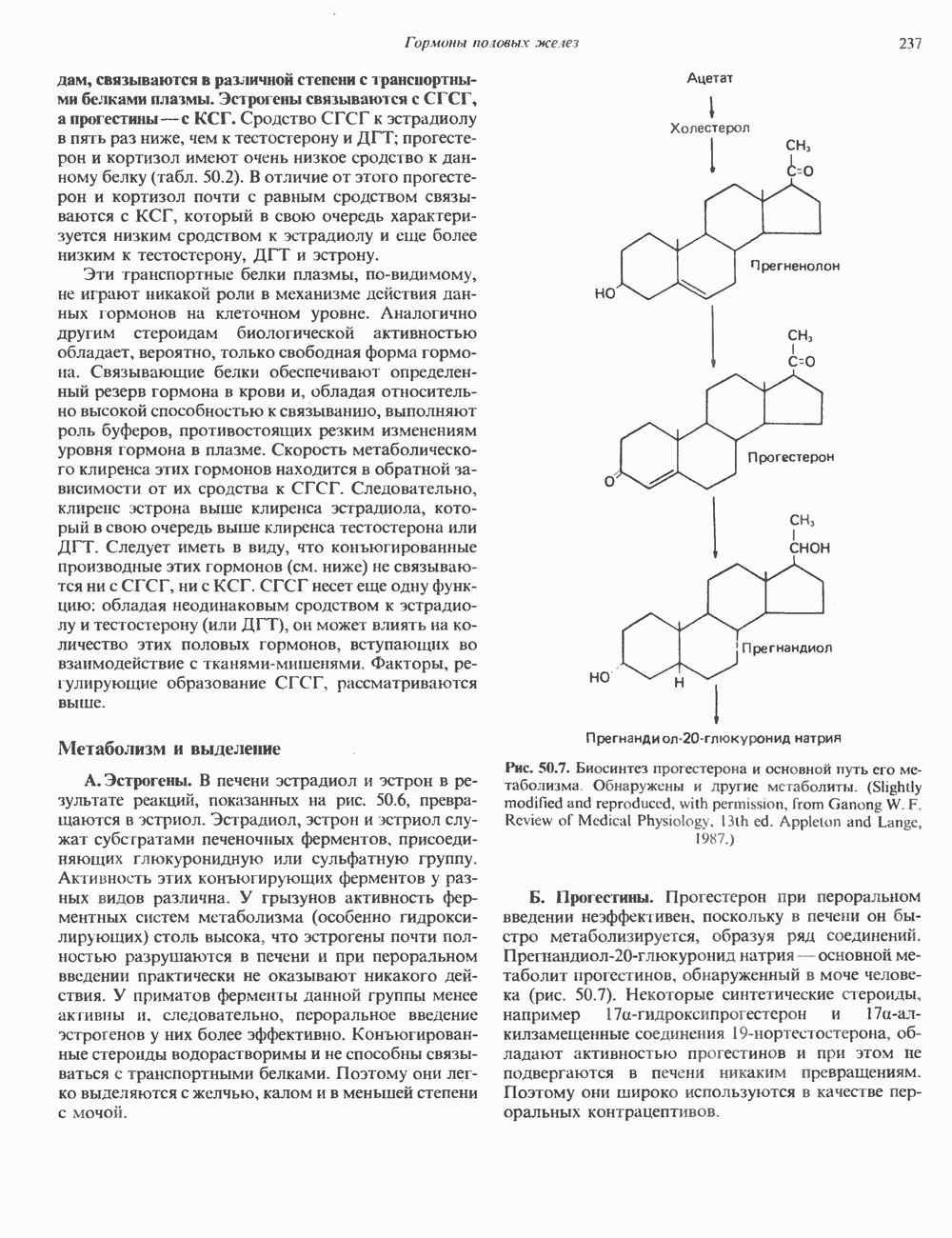
 циклов репликации. Копии ДНК, лежащие рядом в точном соответствии с локализованными на них генами, образуют хромосому с четко выраженным распределением полос — конденсированного и менее плотного хроматина.
циклов репликации. Копии ДНК, лежащие рядом в точном соответствии с локализованными на них генами, образуют хромосому с четко выраженным распределением полос — конденсированного и менее плотного хроматина.
 пар оснований и примерно из
пар оснований и примерно из  нуклеосом. Следовательно, каждая из 23 хроматид гаплоидного
нуклеосом. Следовательно, каждая из 23 хроматид гаплоидного

 и
и  -специфические сегменты IV хромосомы. Стрелками указаны пуфы. Б. Радиоавтограф IV хромосомы, инкубированной с
-специфические сегменты IV хромосомы. Стрелками указаны пуфы. Б. Радиоавтограф IV хромосомы, инкубированной с  -ури-дином для введения метки в РНК. Распределение иммуно-флуоресцентных сигналов и радиоавтографических пятен по хромосоме совпадает. (Reproduced, with permission, from Sass H. PNA polymerase В in polytene chromosomes. Cell 1982: 28: 274. Copyright 1982 by the Massachusetts Institute of Technology.)
-ури-дином для введения метки в РНК. Распределение иммуно-флуоресцентных сигналов и радиоавтографических пятен по хромосоме совпадает. (Reproduced, with permission, from Sass H. PNA polymerase В in polytene chromosomes. Cell 1982: 28: 274. Copyright 1982 by the Massachusetts Institute of Technology.)
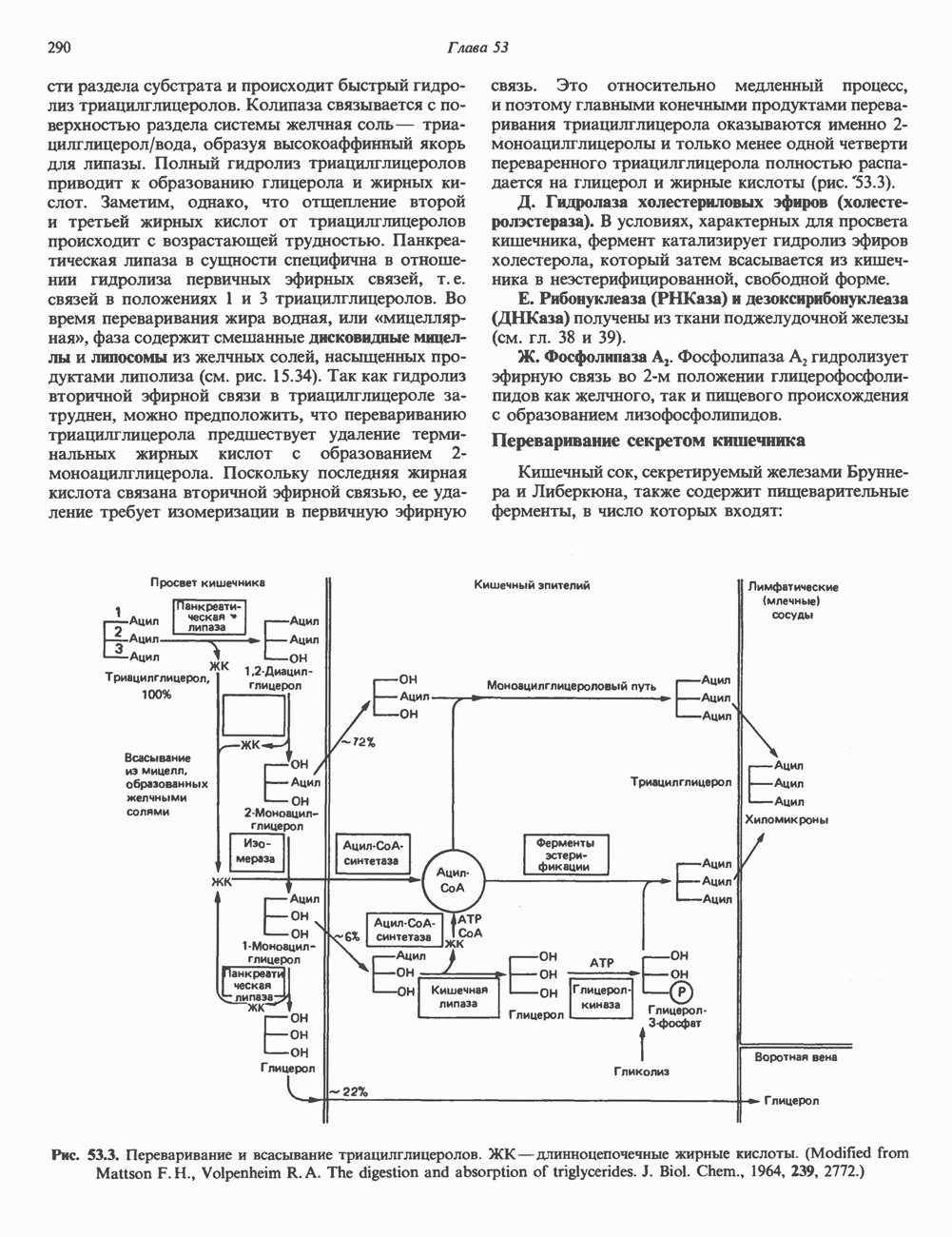
 нуклеотидов в одной двухцепочечной молекуле ДНК. Таким образом, при формировании конденсированной метафазной хромосомы линейный размер каждой молекулы ДНК должен быть уменьшен в 8000 раз! В метафазных хромосомах хроматиновые волокна (длиной 25—30 нм) также складываются в серии петлеобразных доменов, проксимальные участки которых закрепляются на внутриядерном белковом (не-гистоновом) каркасе. Коэффициенты, характеризующие плотность упаковки каждой упорядоченной структуры ДНК, приведены в табл. 38.1.
нуклеотидов в одной двухцепочечной молекуле ДНК. Таким образом, при формировании конденсированной метафазной хромосомы линейный размер каждой молекулы ДНК должен быть уменьшен в 8000 раз! В метафазных хромосомах хроматиновые волокна (длиной 25—30 нм) также складываются в серии петлеобразных доменов, проксимальные участки которых закрепляются на внутриядерном белковом (не-гистоновом) каркасе. Коэффициенты, характеризующие плотность упаковки каждой упорядоченной структуры ДНК, приведены в табл. 38.1.
