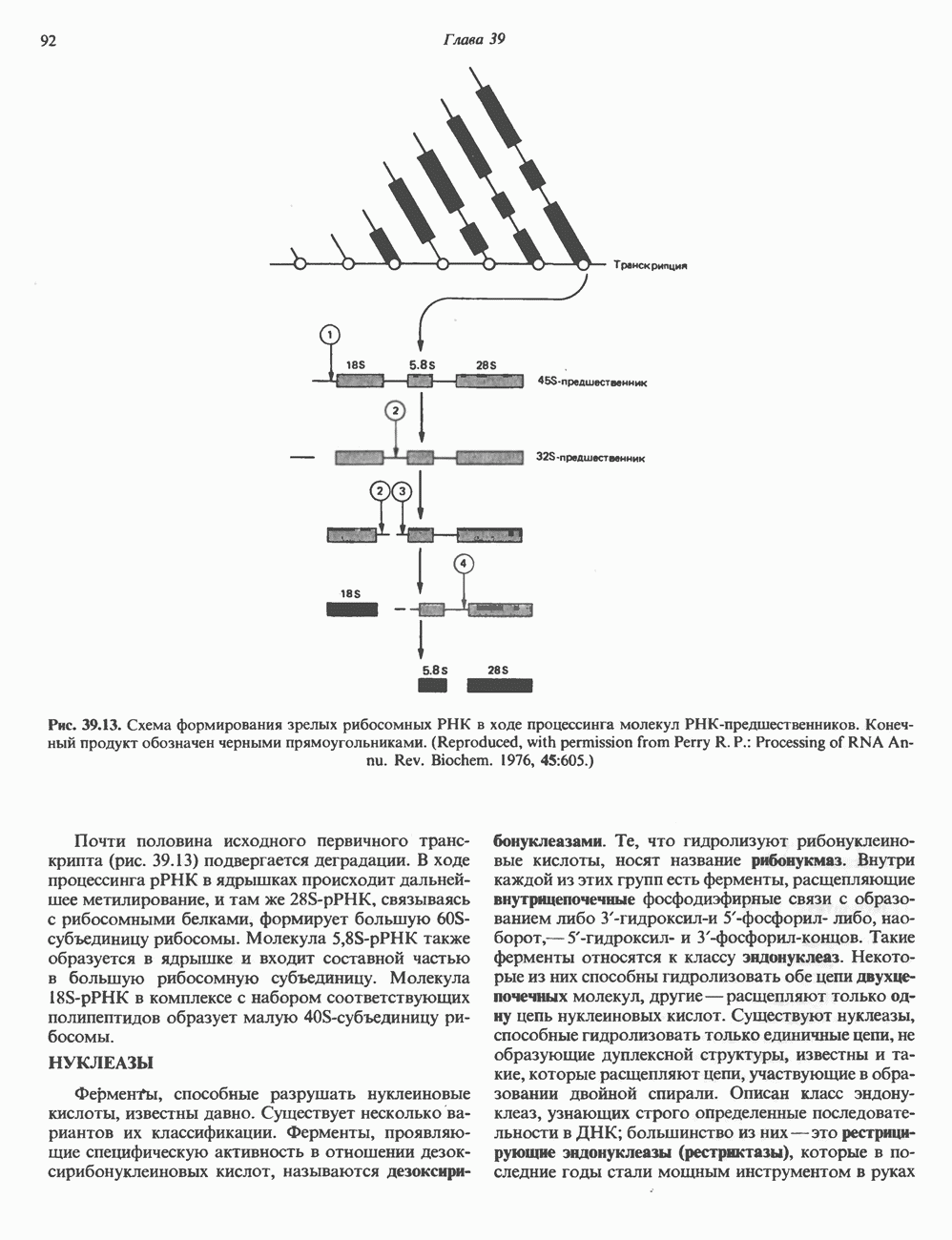
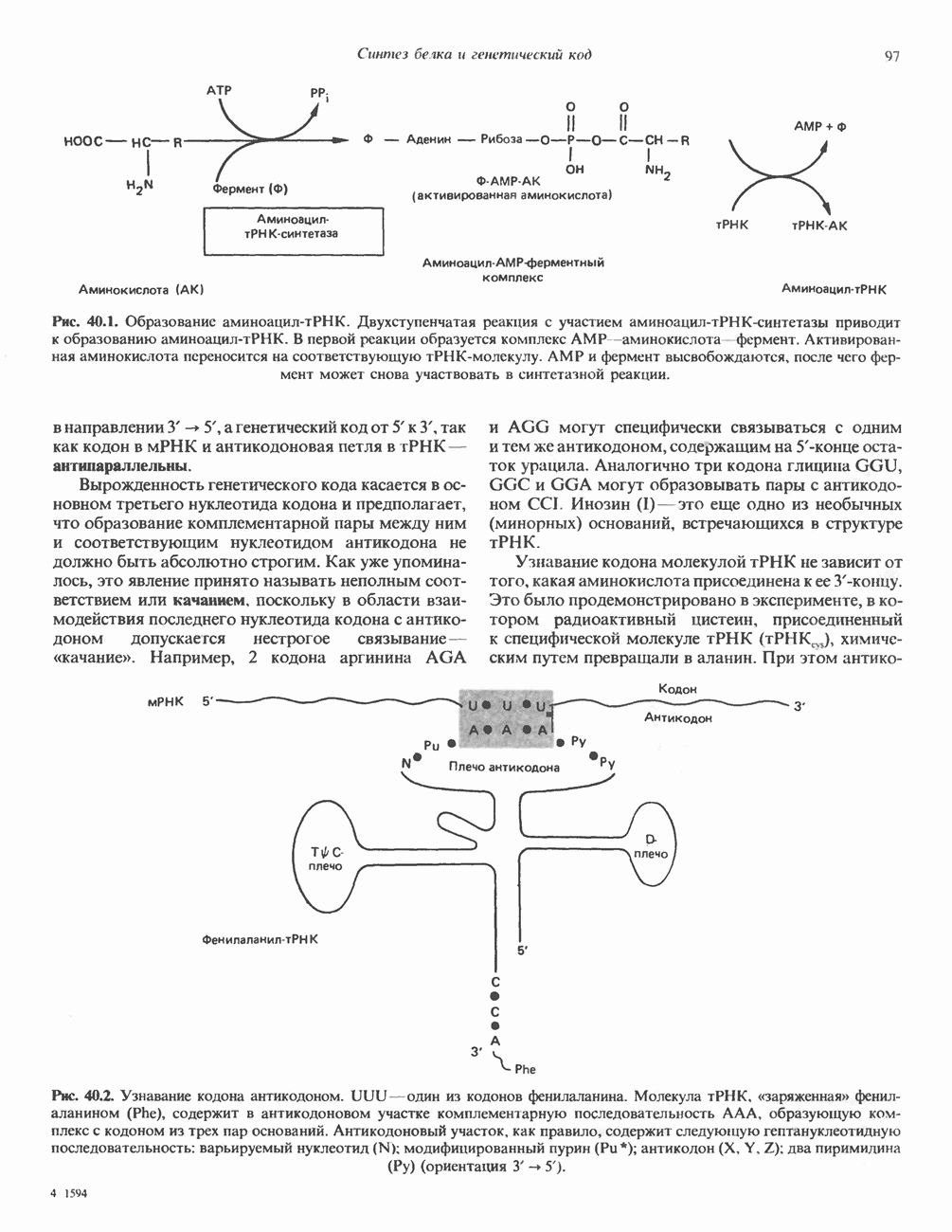
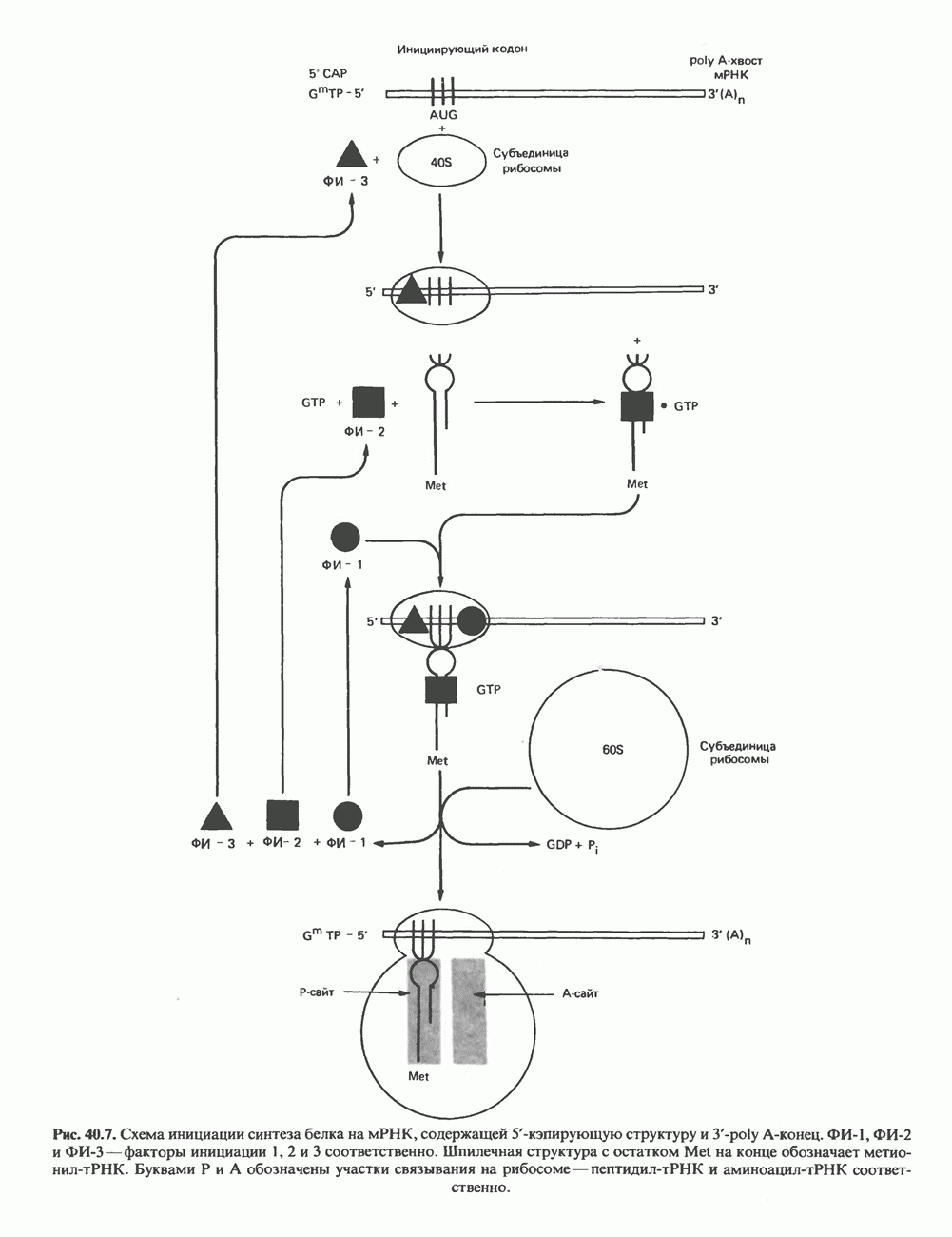
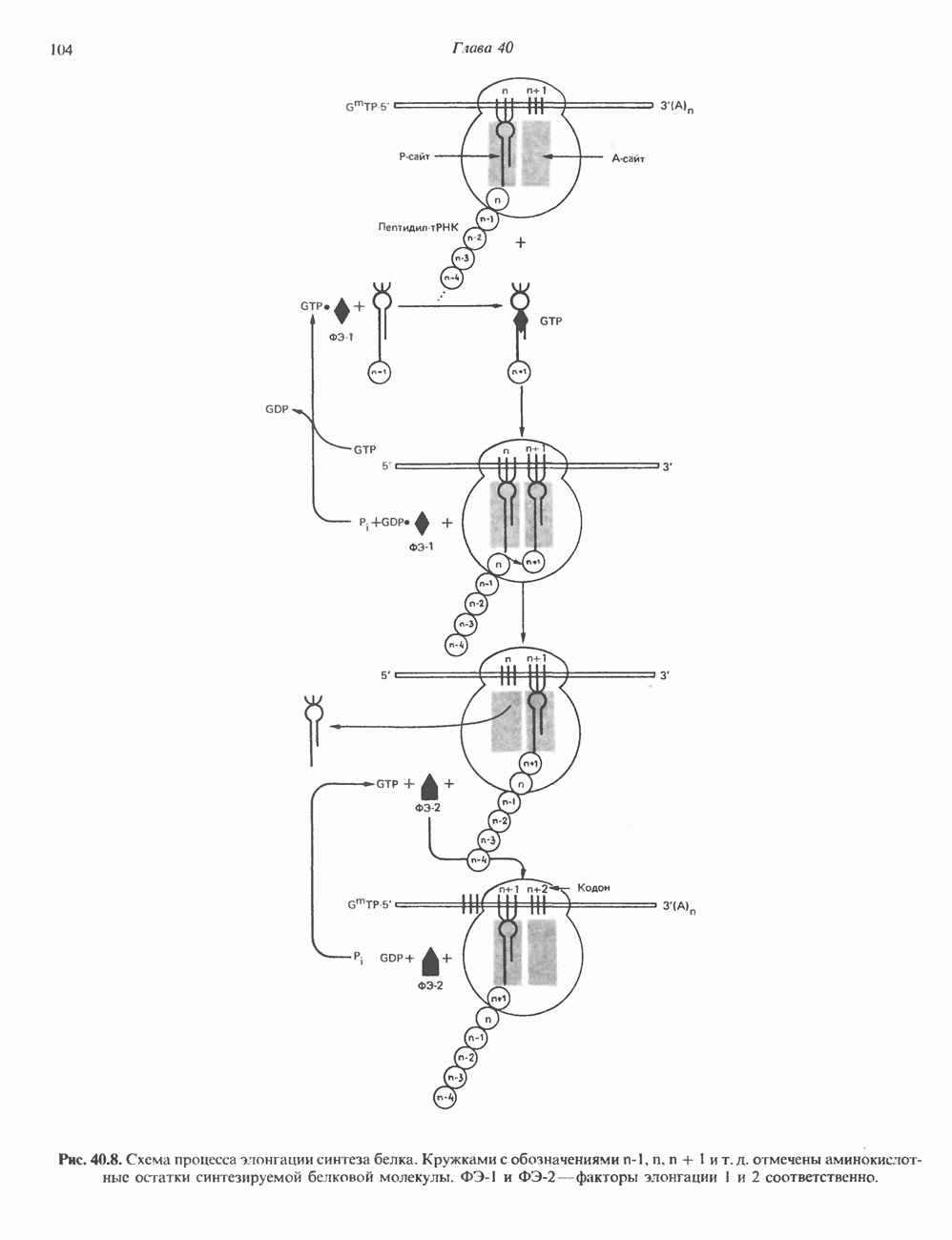
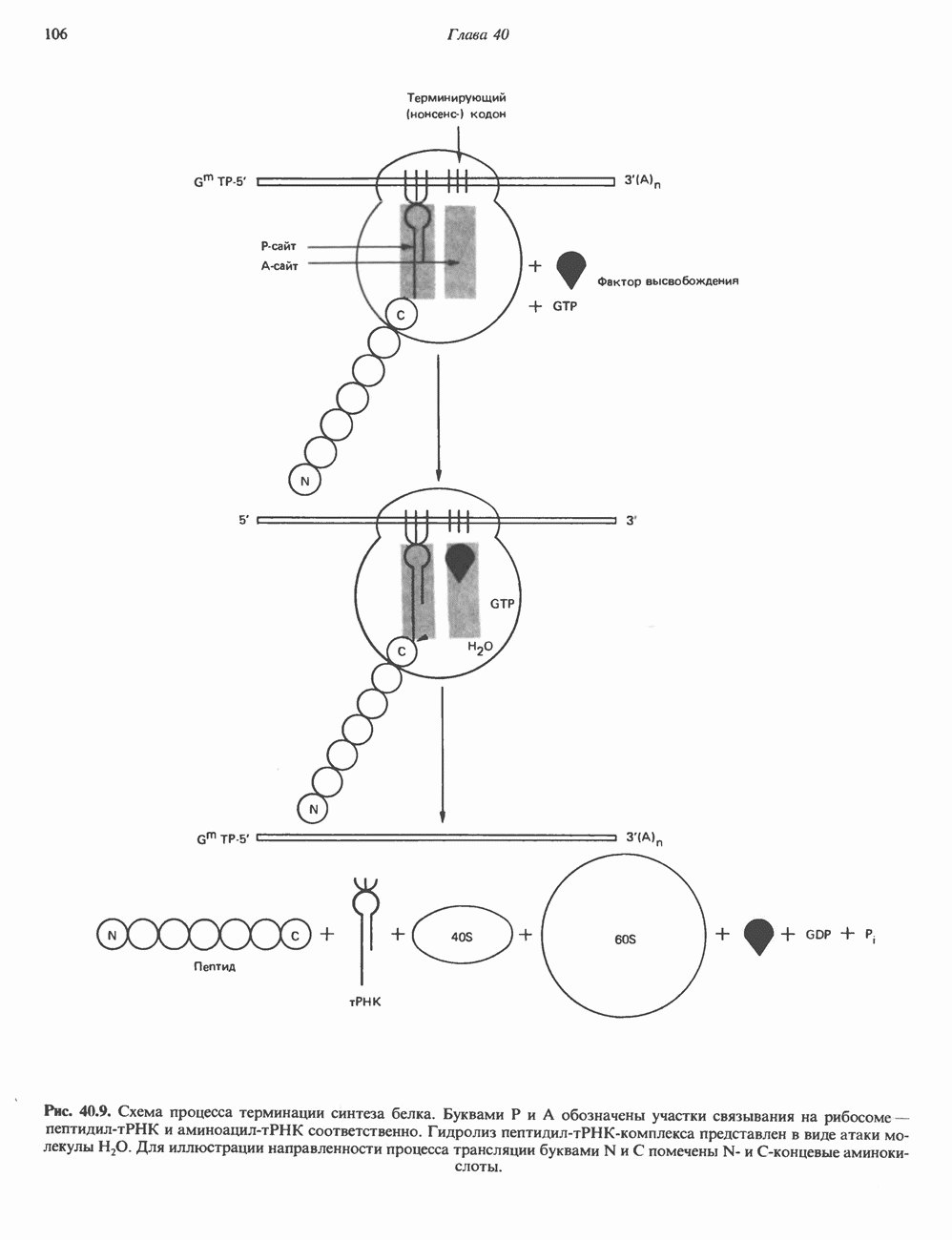
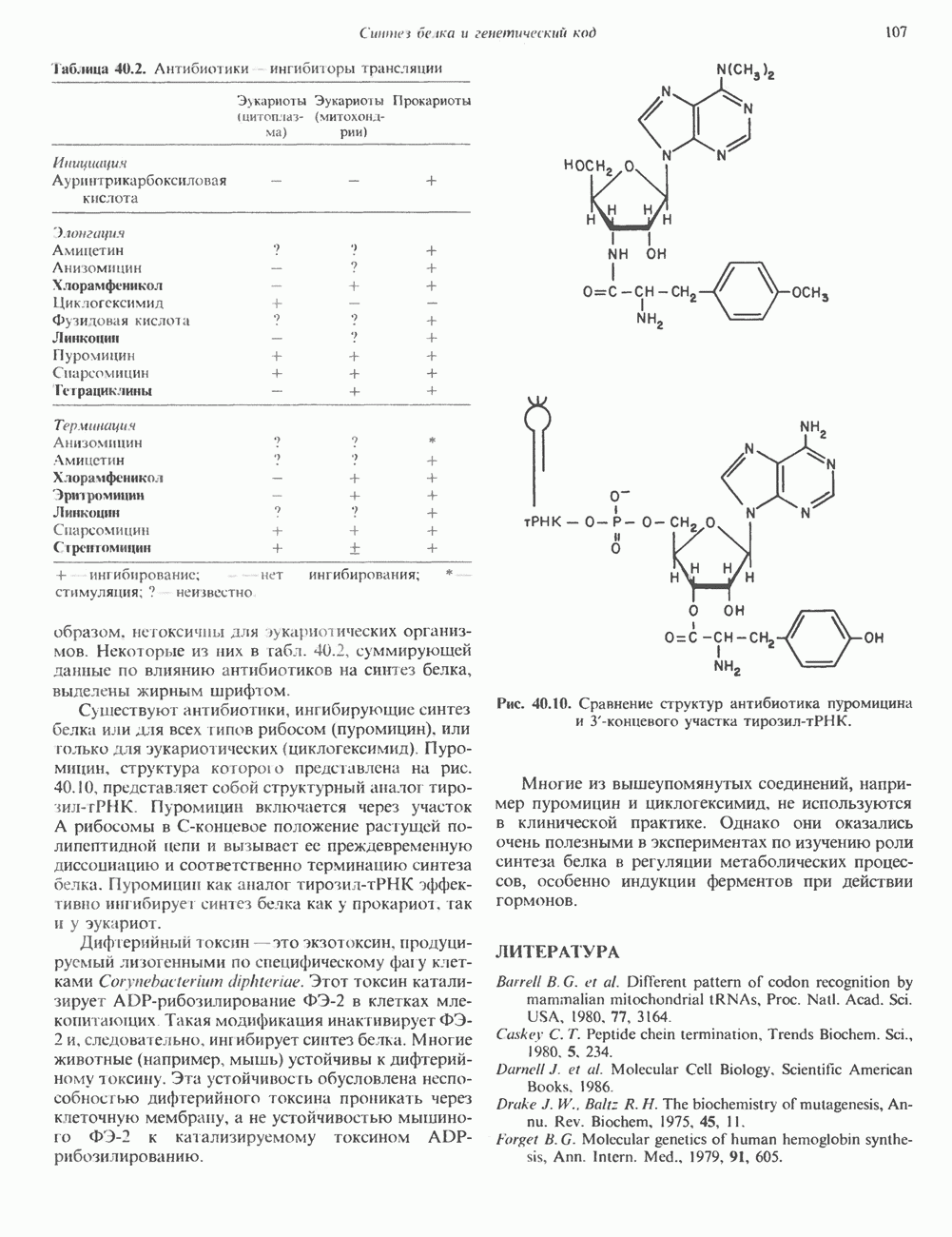
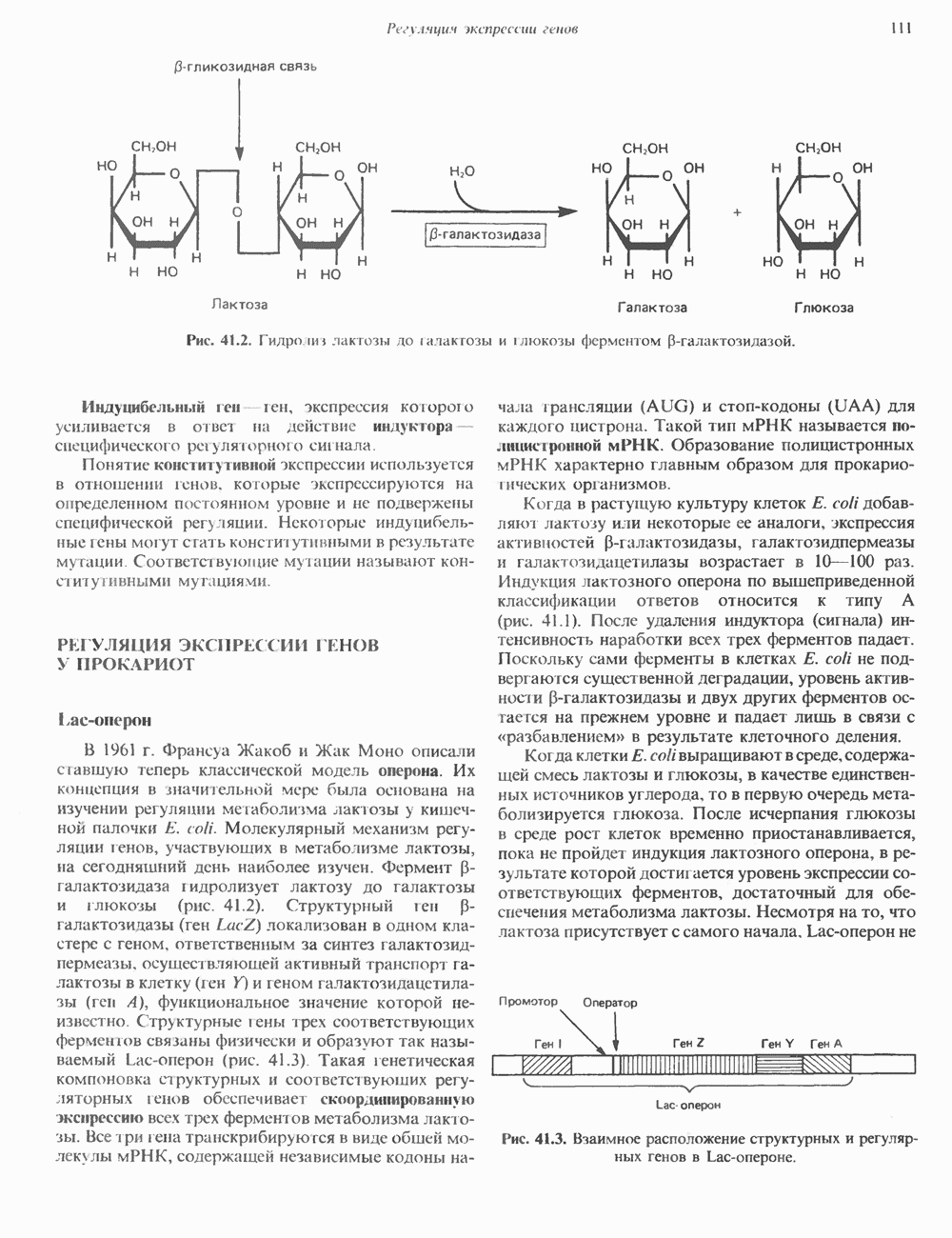
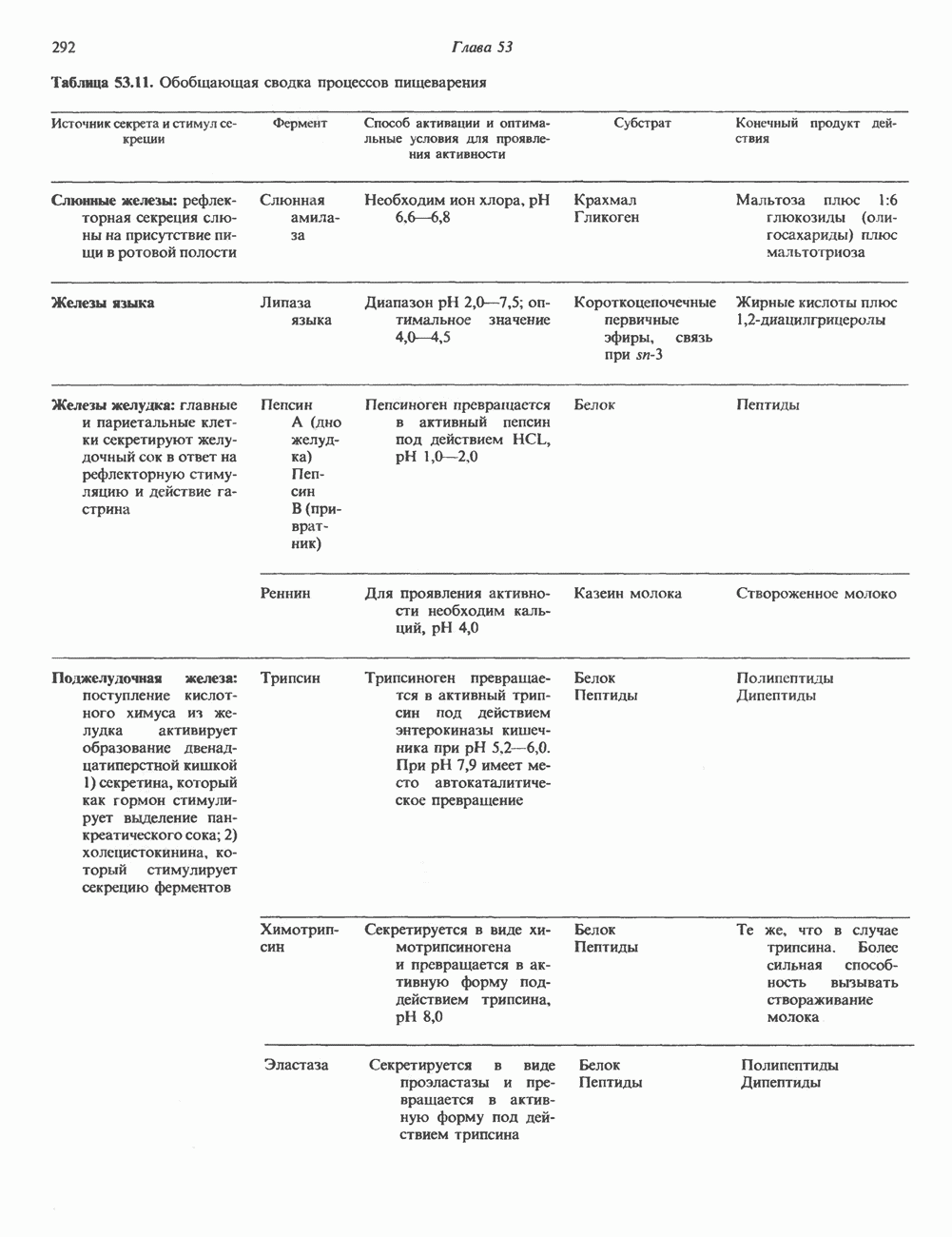
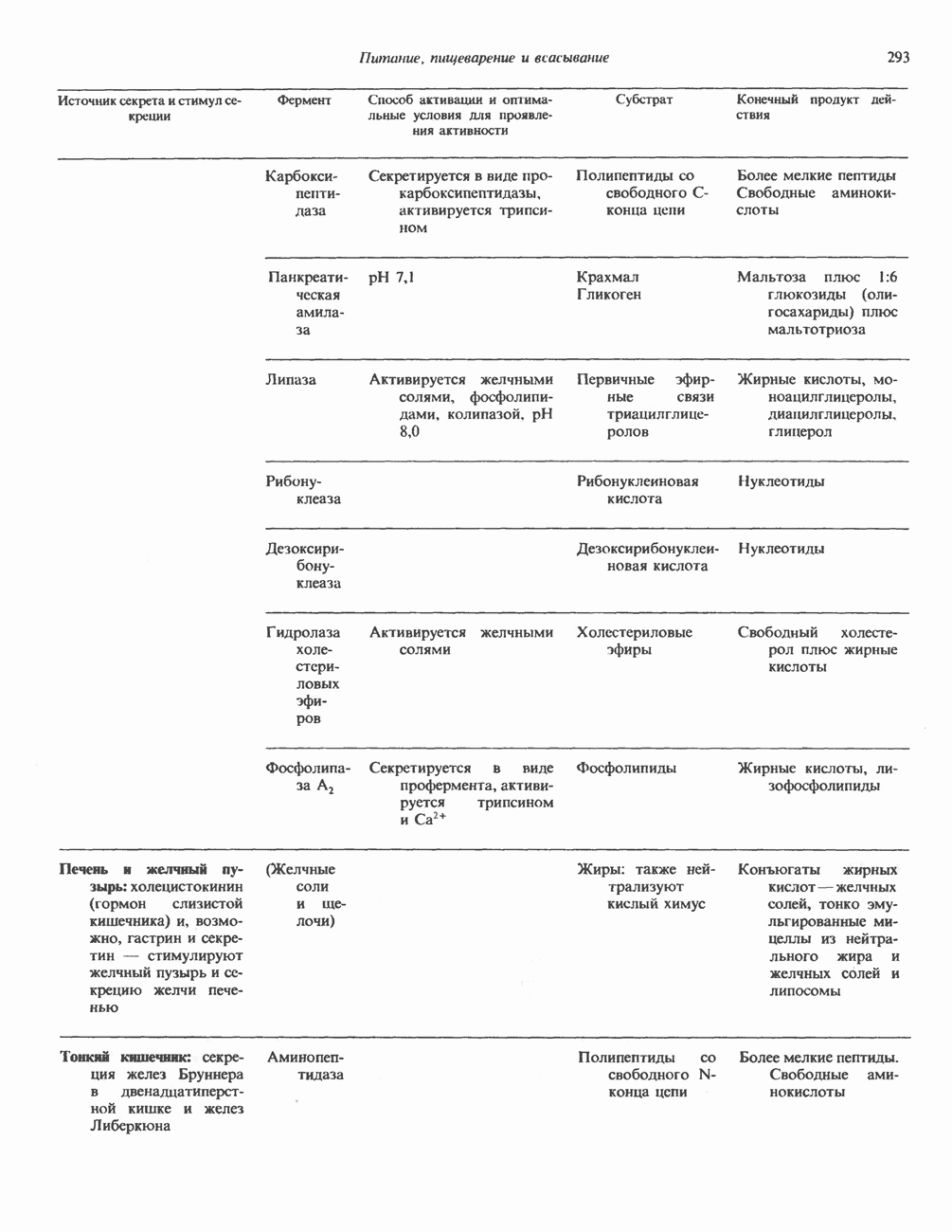
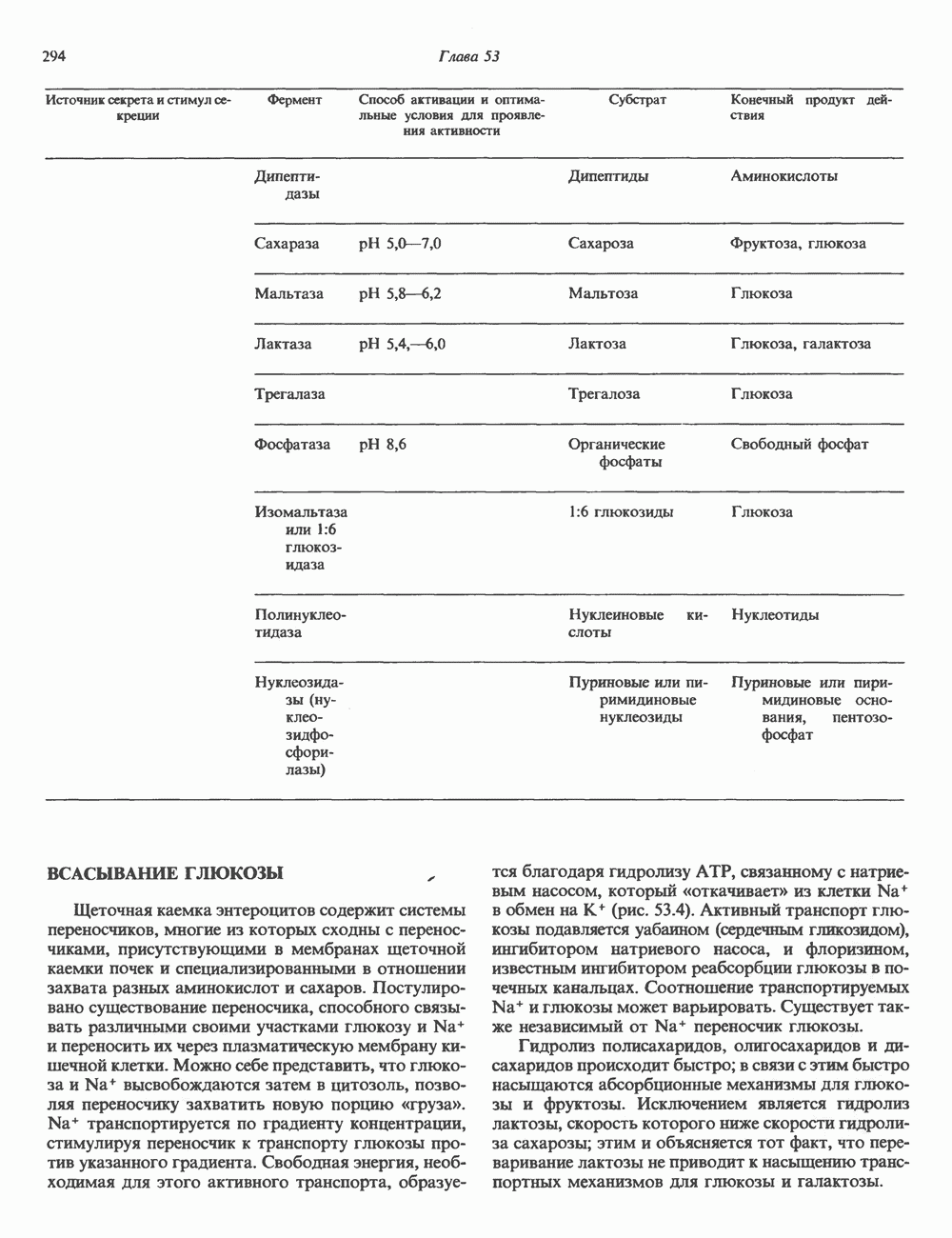
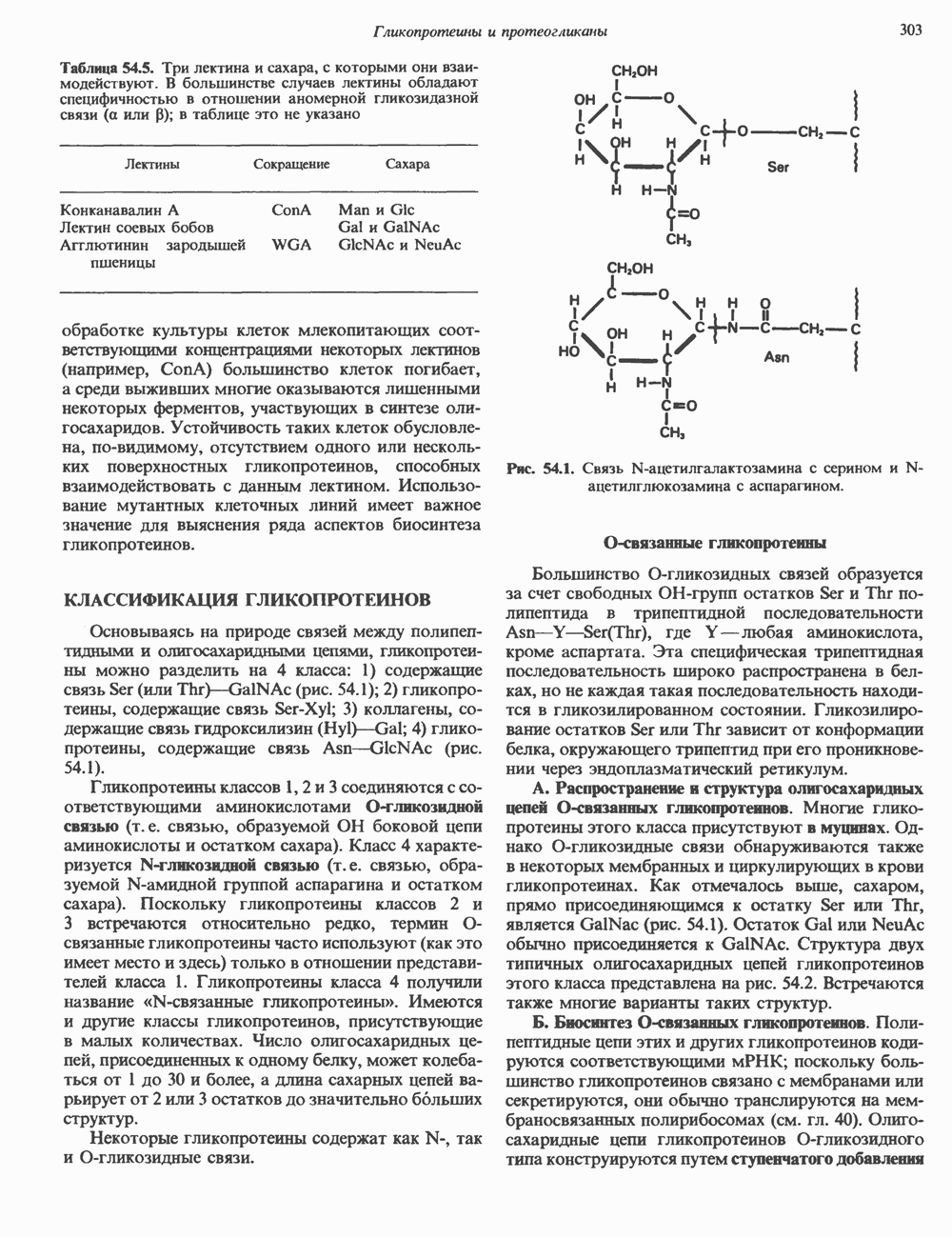
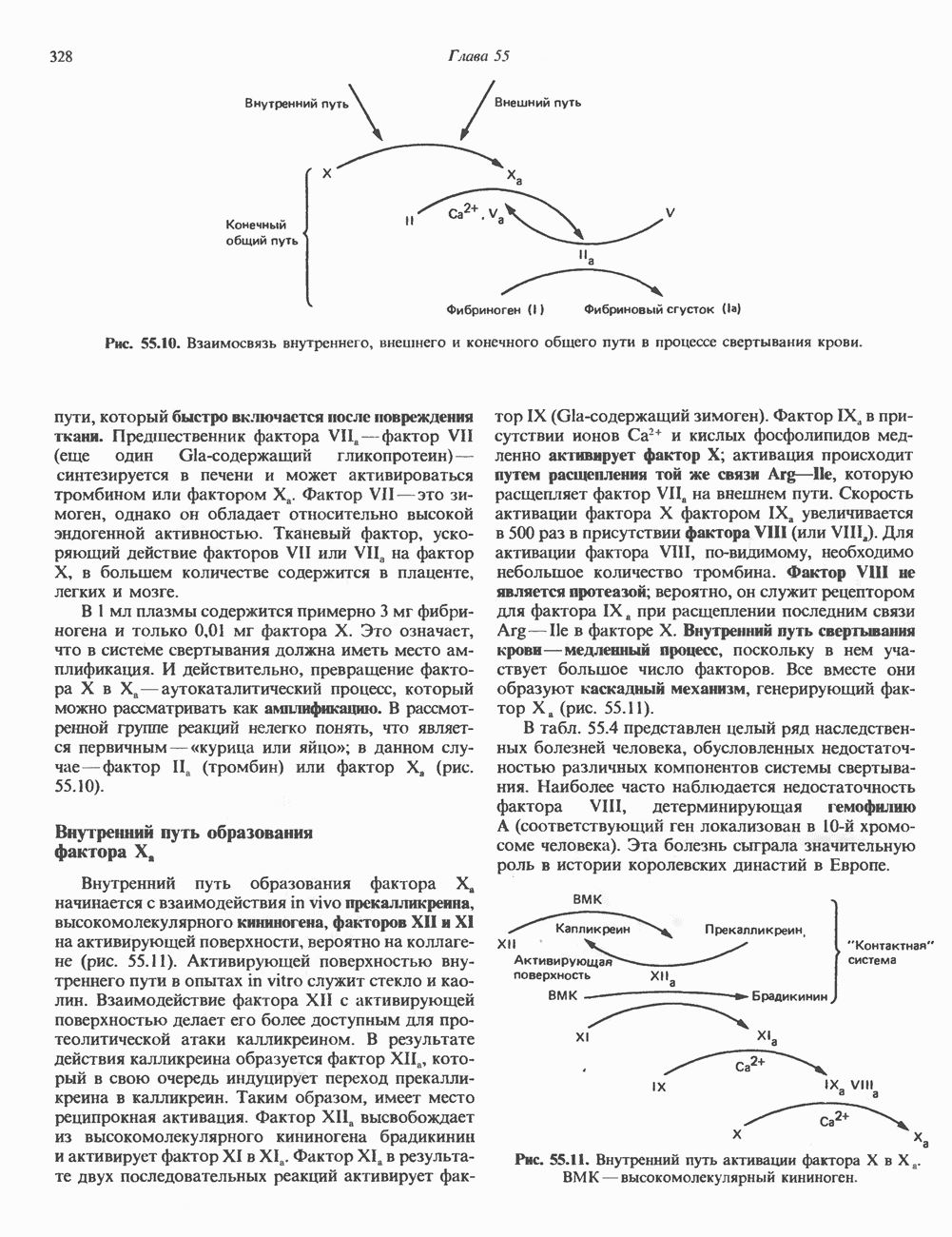
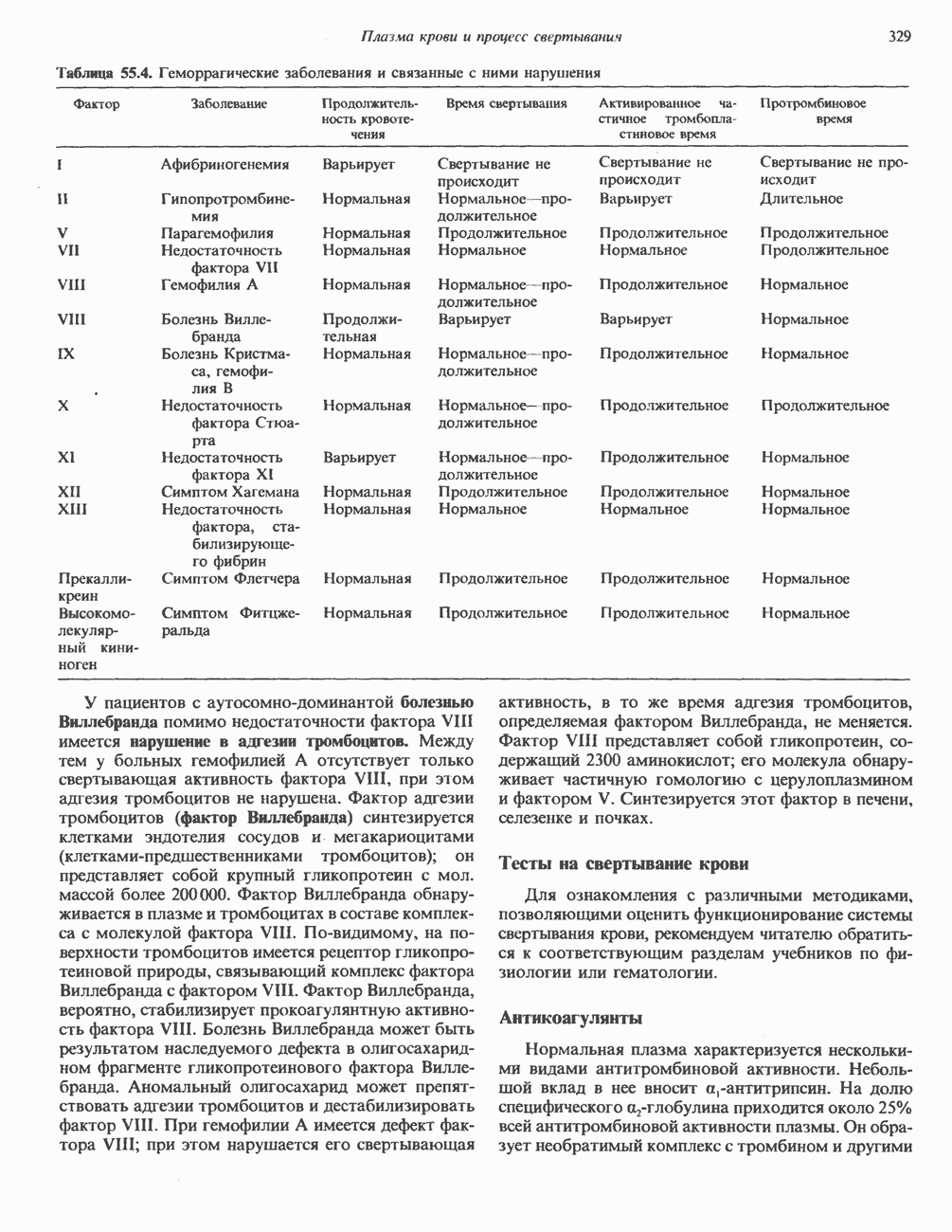
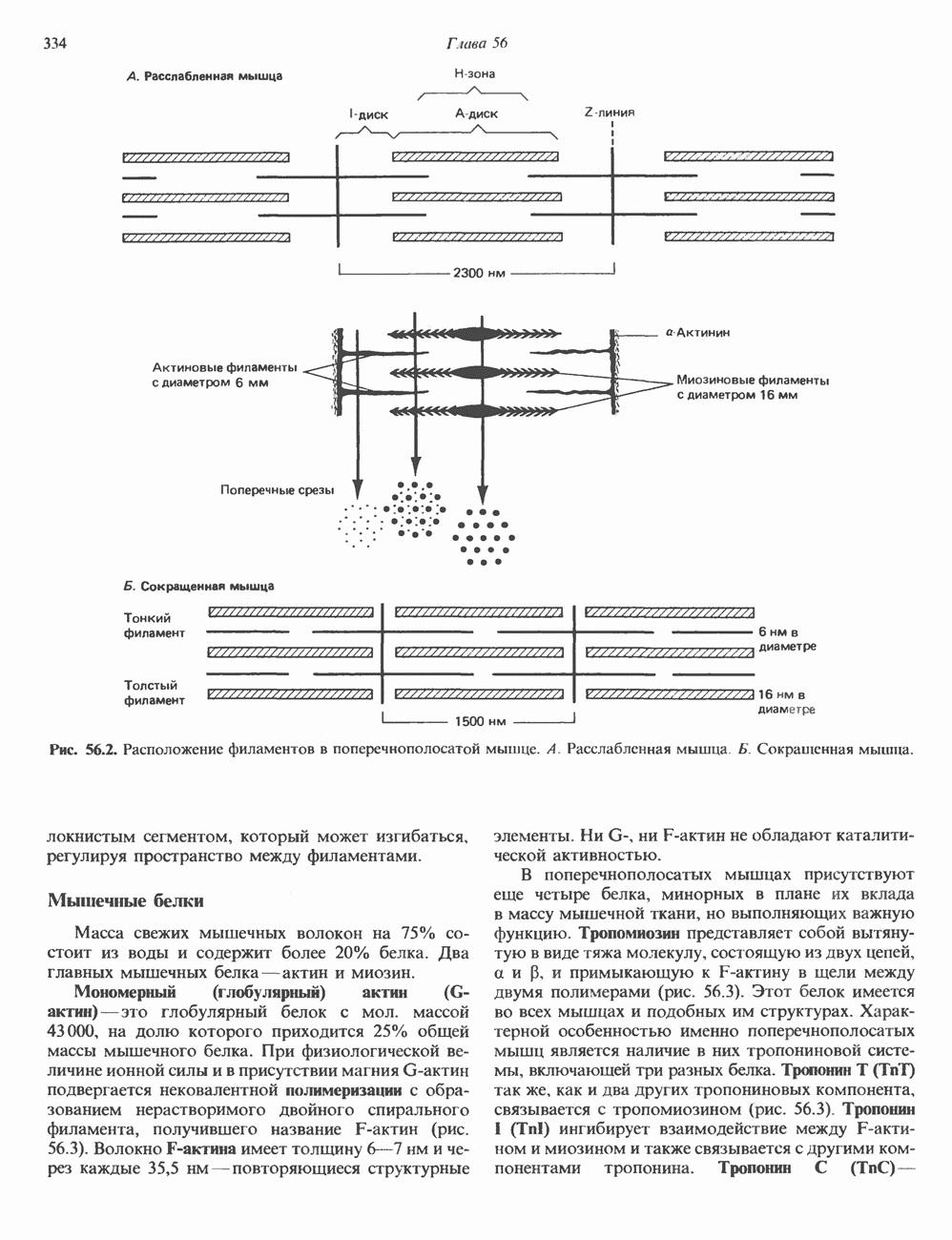
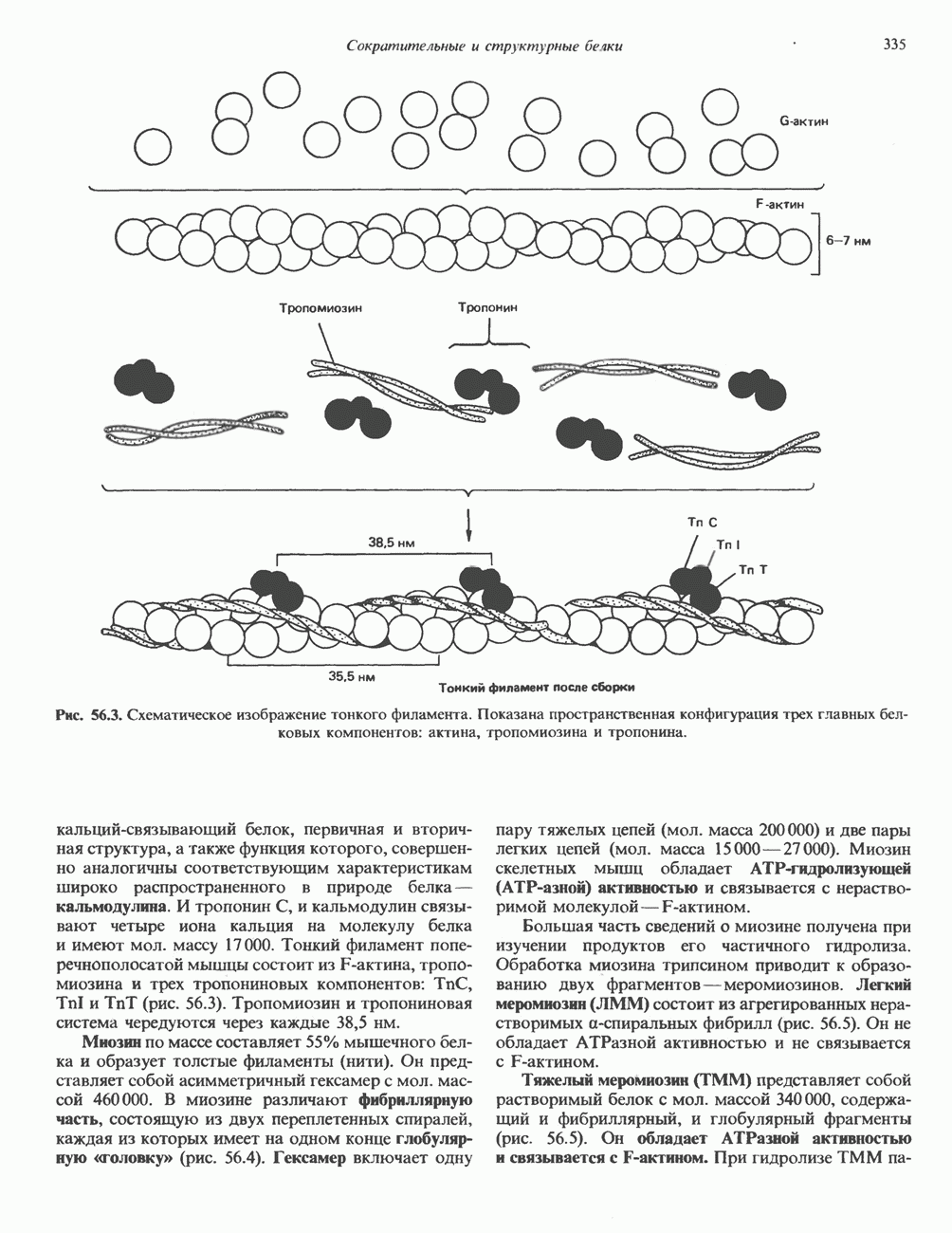
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
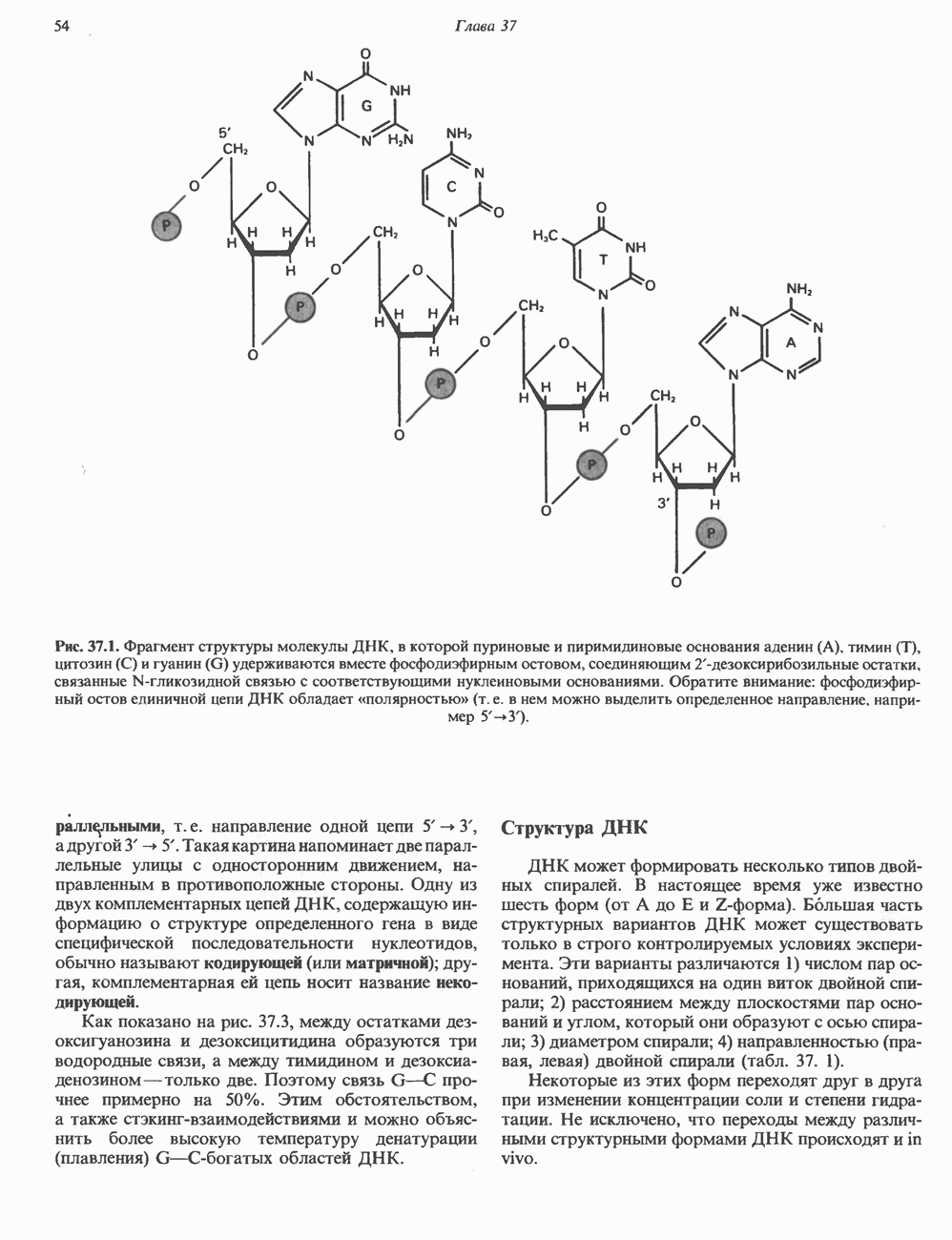
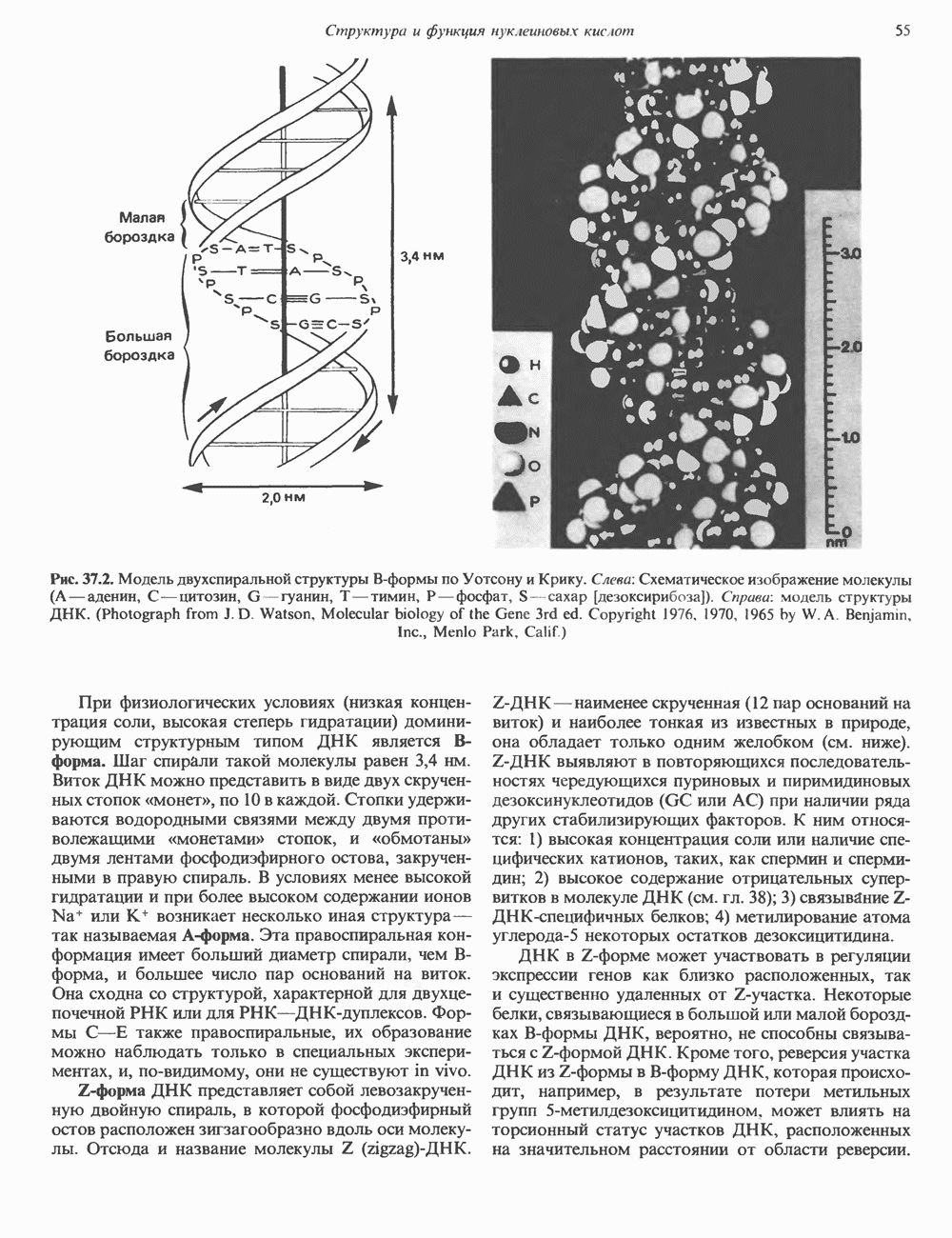
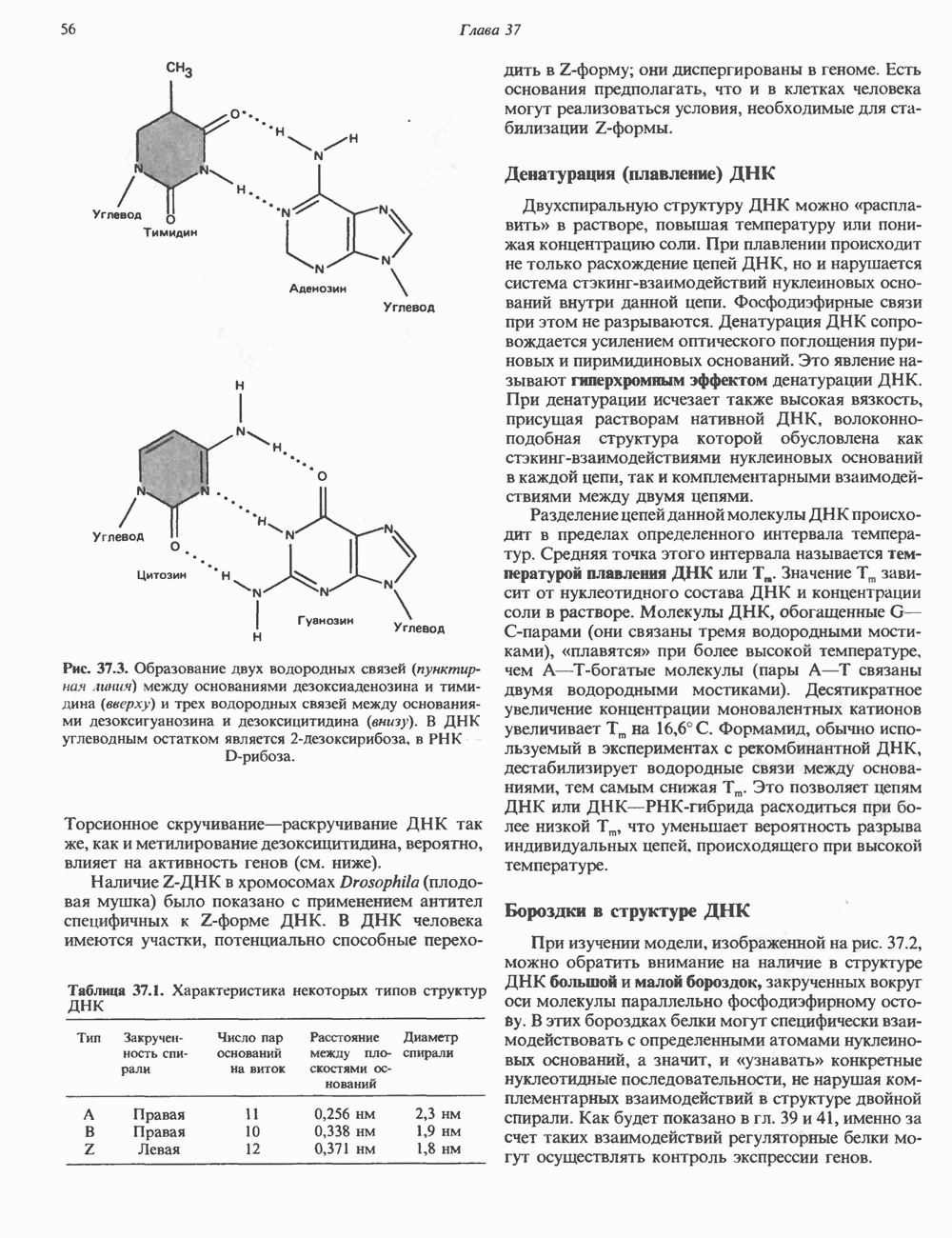
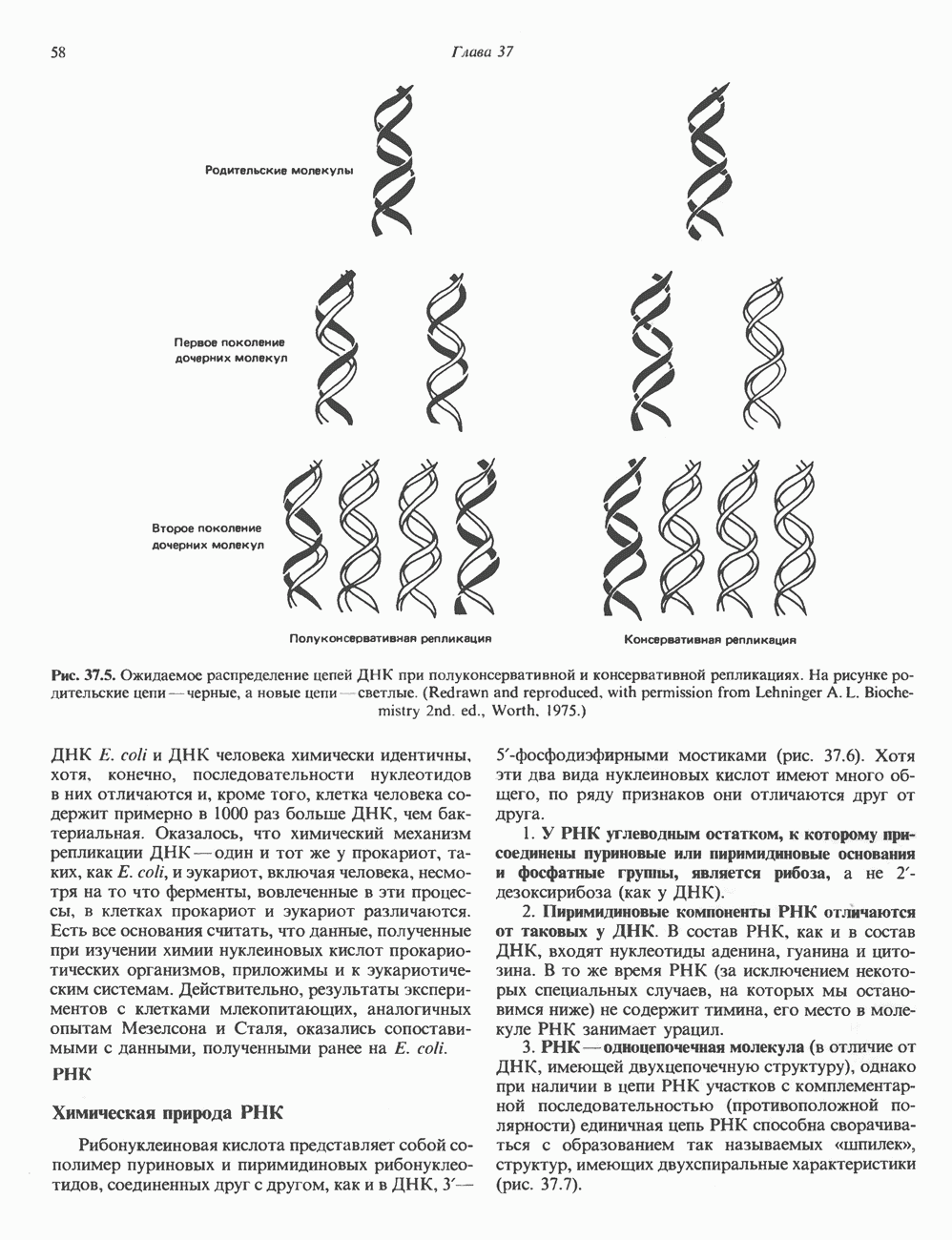
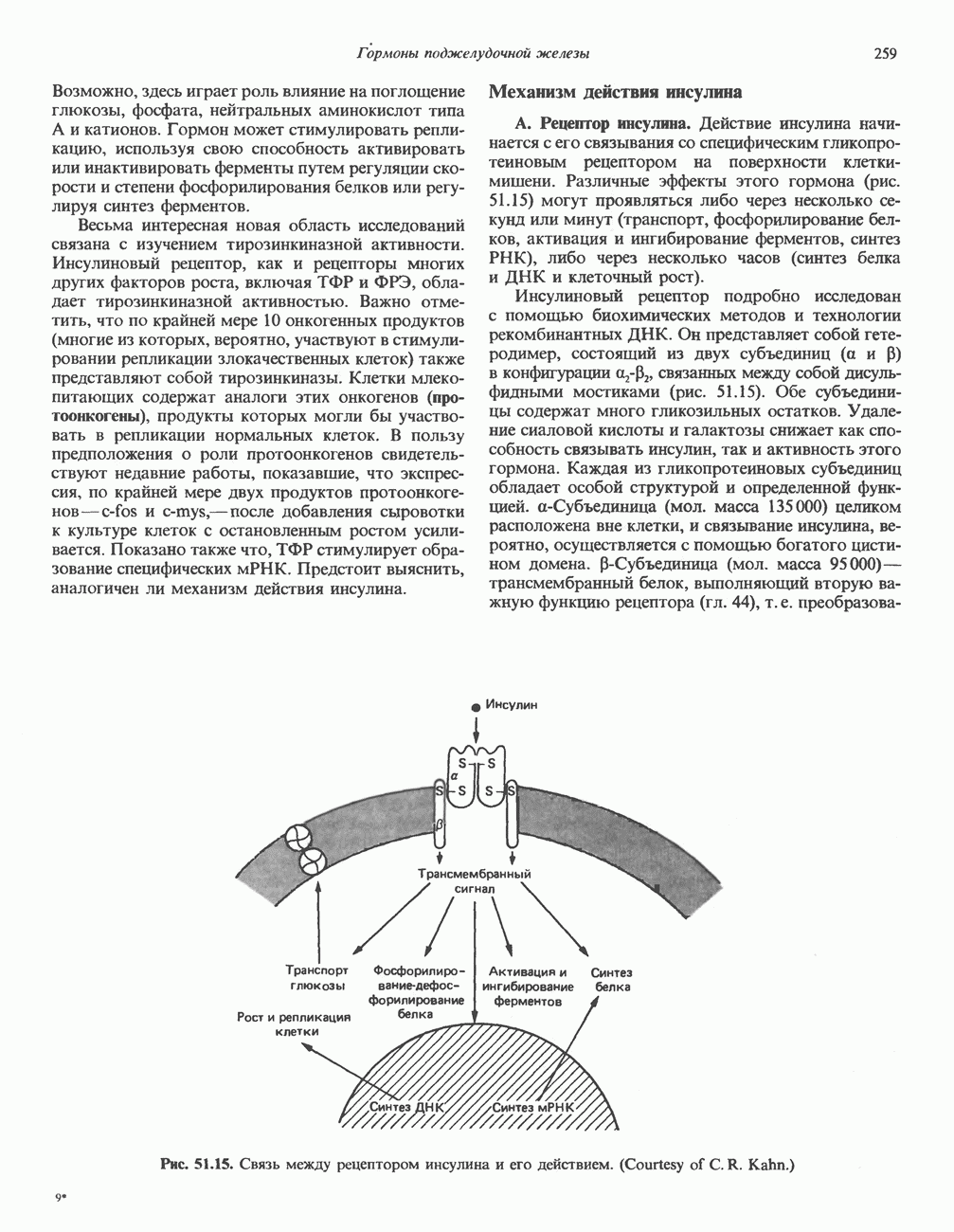
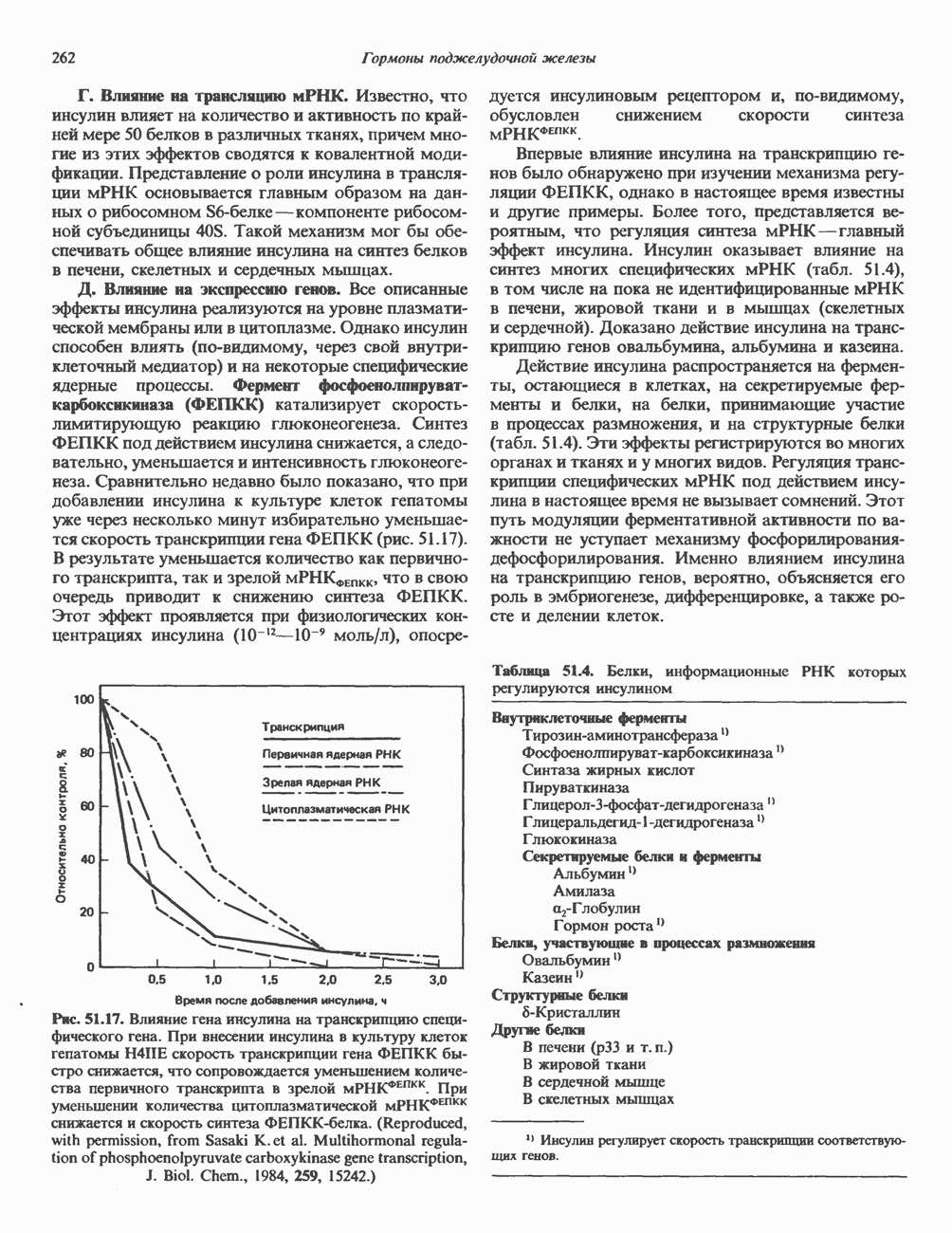
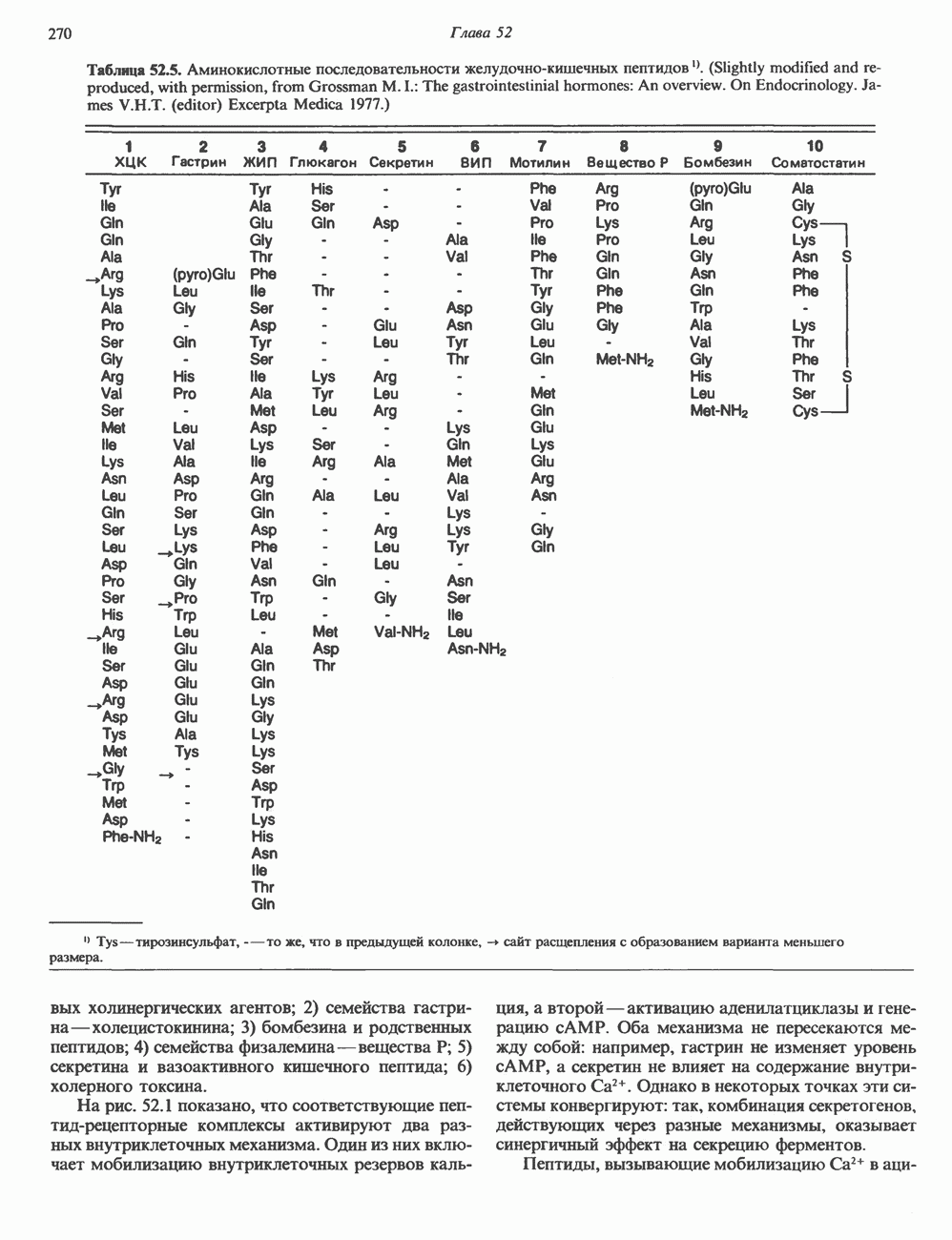
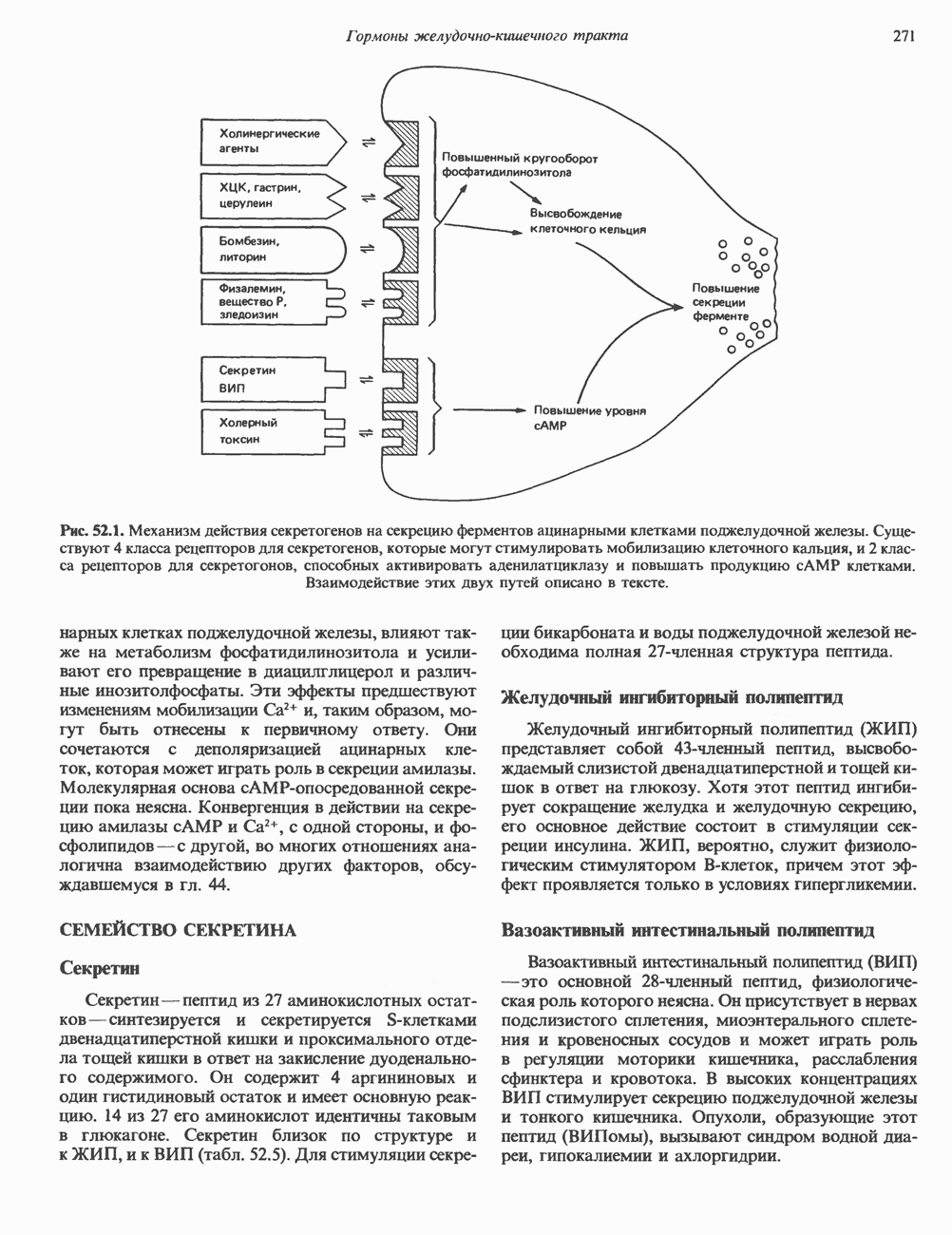
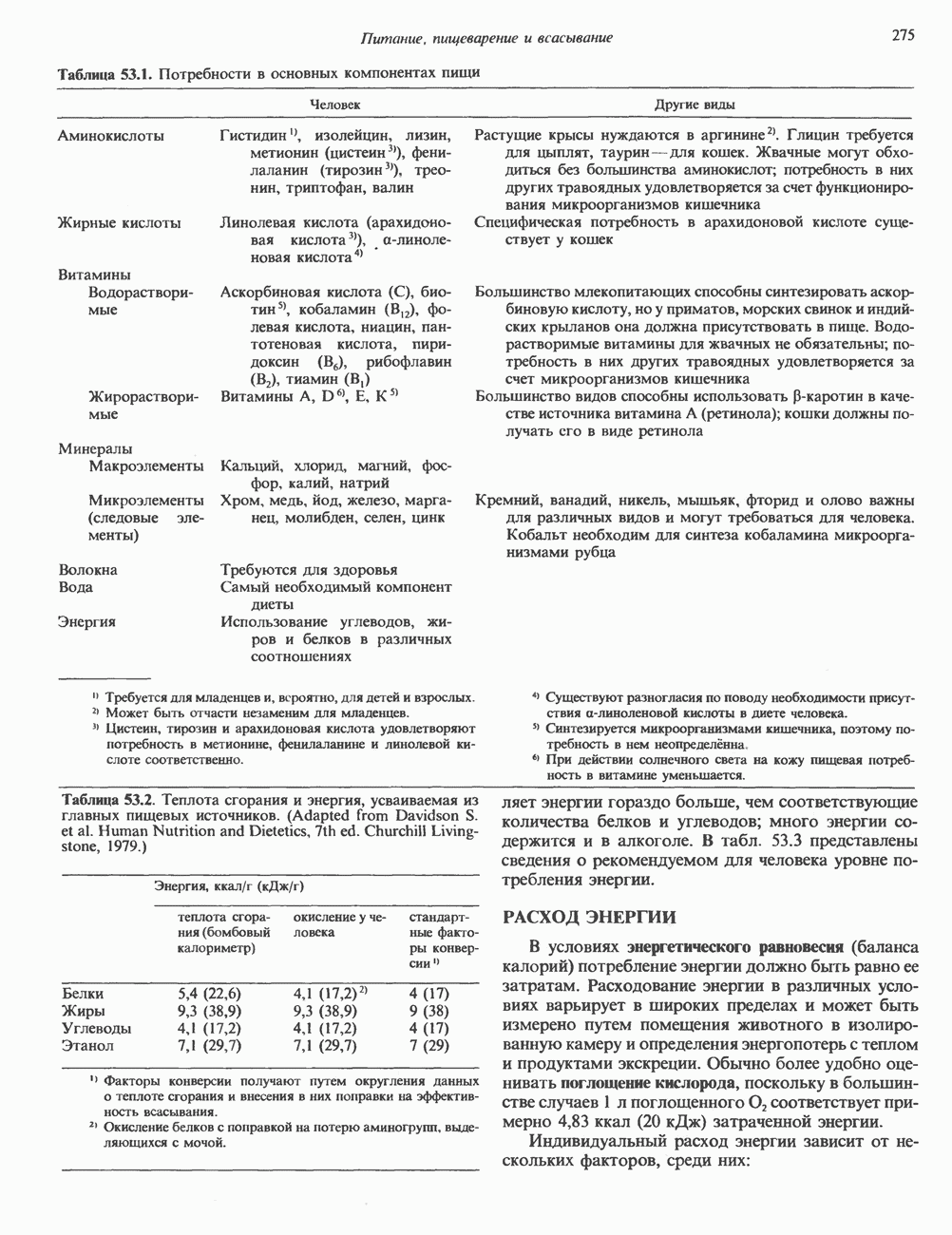
СТРУКТУРА МЕМБРАНМембраны — это сложные структуры, состоящие из липидов, белков и углеводов. Различные мембраны внутри клеток и между ними имеют неодинаковый состав, характеризуемый отношением белки липиды (рис. 42.1). Это неудивительно, если учесть, какие разнообразные функции выполняют мембраны. Мембраны являются асимметричными плоскими замкнутыми структурами, обладающими внешней и внутренней поверхностями. Эти плоские структуры образованы не с помощью ковалентных связей, и тем не менее они термодинамически стабильны и метаболически активны. В мембранах «заякорены» особые белковые молекулы, которые осуществляют функции, специфические для определенных органелл, клеток или организмов. ЛИПИДНЫЙ СОСТАВБольшинство липидов в мембранах млекопитающих представлены фосфолипидами, гликосфинголипидами и холестеролом. ФосфолипидыФосфолипиды в составе мембран подразделяются на две основные группы. Наиболее распространены фосфоглицериды, состоящие из остатка глицерола, к которому присоединены эфирными связями две жирнокислотные молекулы и фосфорилированный спирт (рис. 42.2). Жирные кислоты обычно содержат четное число атомов углерода, в основном 14 или 16. Их углеродные цепочки не разветвлены и могут быть как насыщенными, так и ненасыщенными. Наиболее простым фосфоглицеридом является фосфатидная кислота—1, 2-диацилглицерол-З-фосфат—ключевой интермедиат при образовании
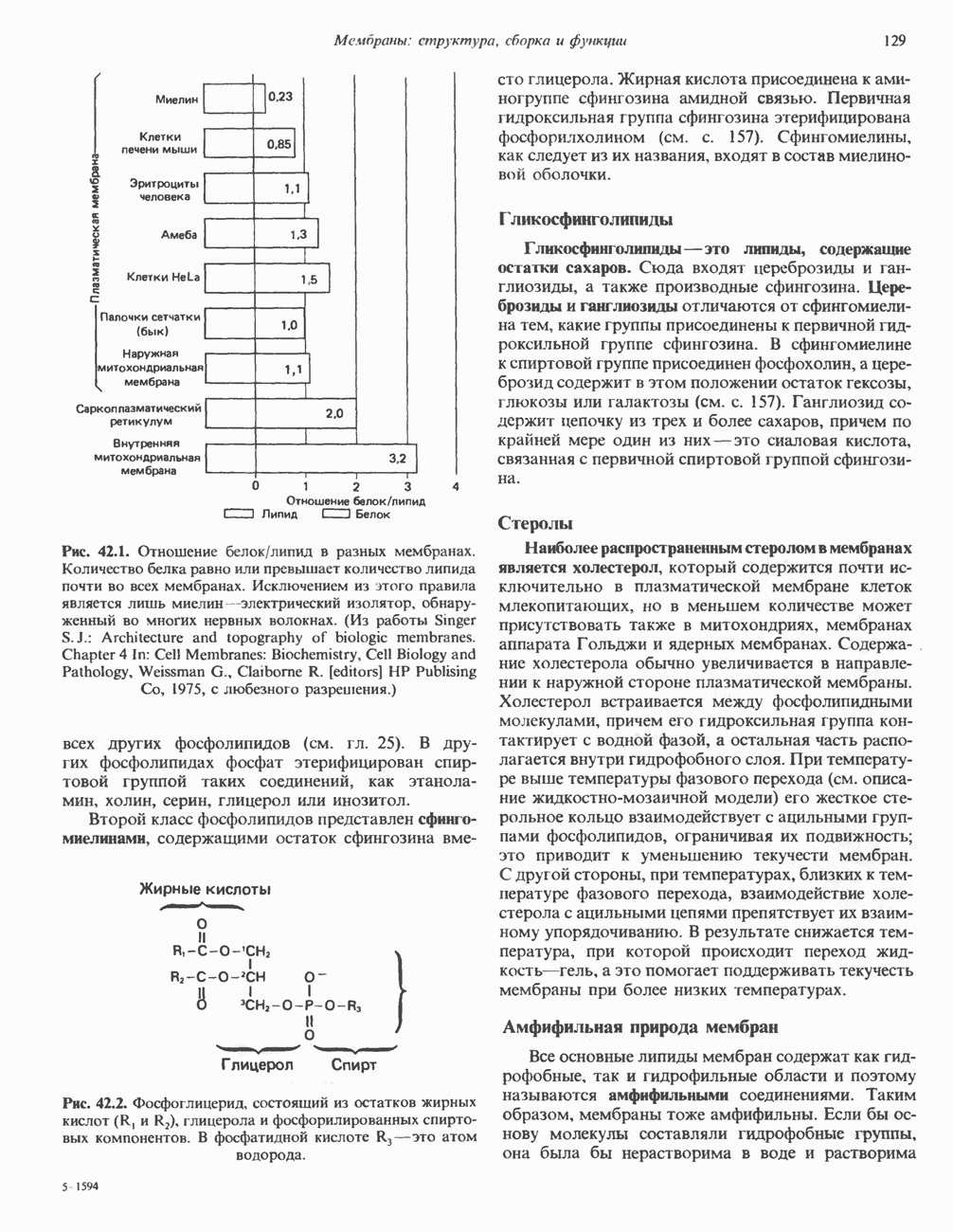
Рис. 42.1. Отношение белок/липид в разных мембранах. Количество белка равно или превышает количество липида почти во всех мембранах. Исключением из этого правила является лишь миелин — электрический изолятор, обнаруженный во многих нервных волокнах. (Из работы Singer S. J.: Architecture and topography of biologic membranes. Chapter 4 In: Cell Membranes: Biochemistry, Cell Biology and Pathology, Weissman G., Claiborne R. [editors] HP Publising Co, 1975, с любезного разрешения.) всех других фосфолипидов (см. гл. 25). В других фосфолипидах фосфат этерифицирован спиртовой группой таких соединений, как этаноламин, холин, серин, глицерол или инозитол. Второй класс фосфолипидов представлен сфингомиелинами, содержащими остаток сфингозина вместо глицерола.
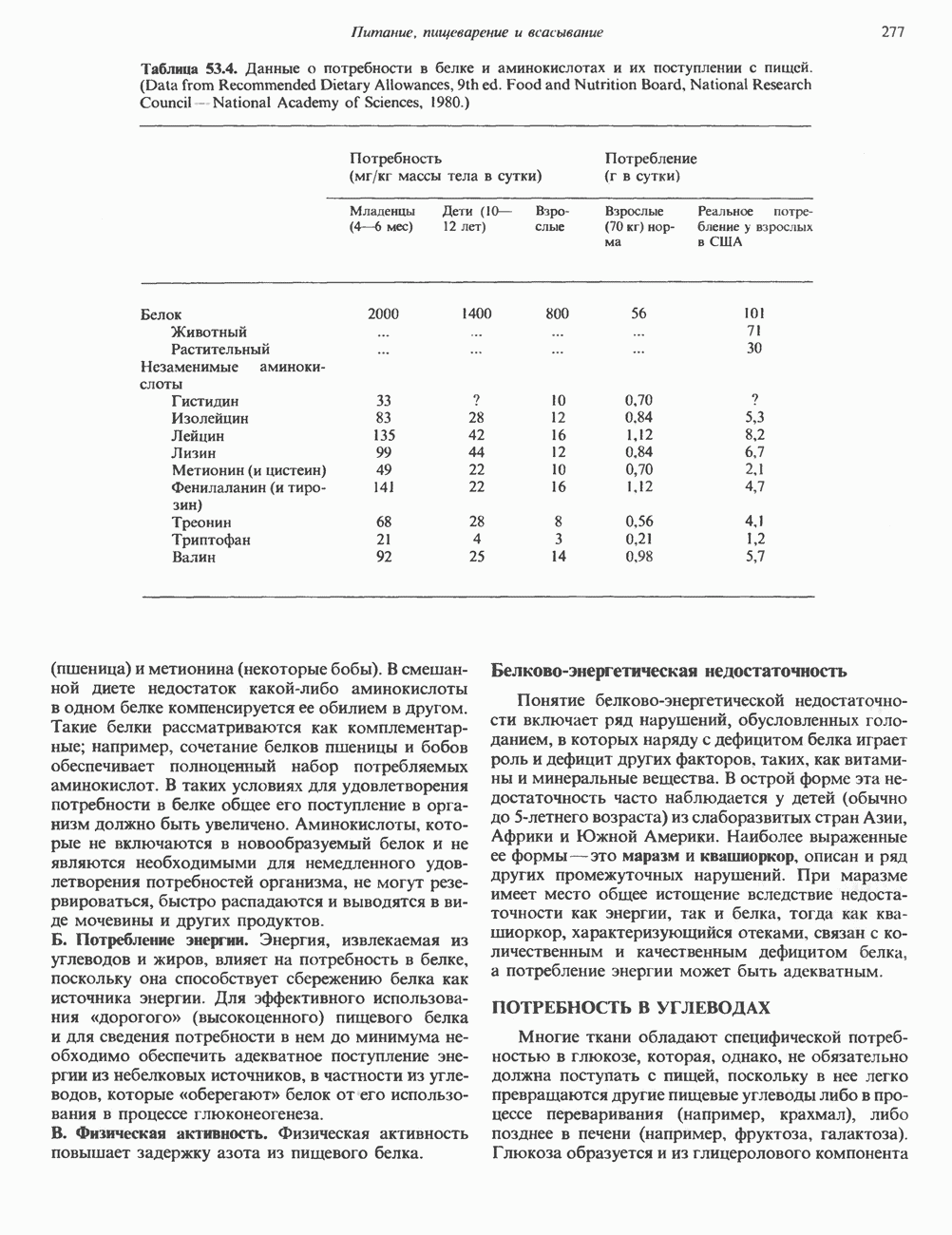
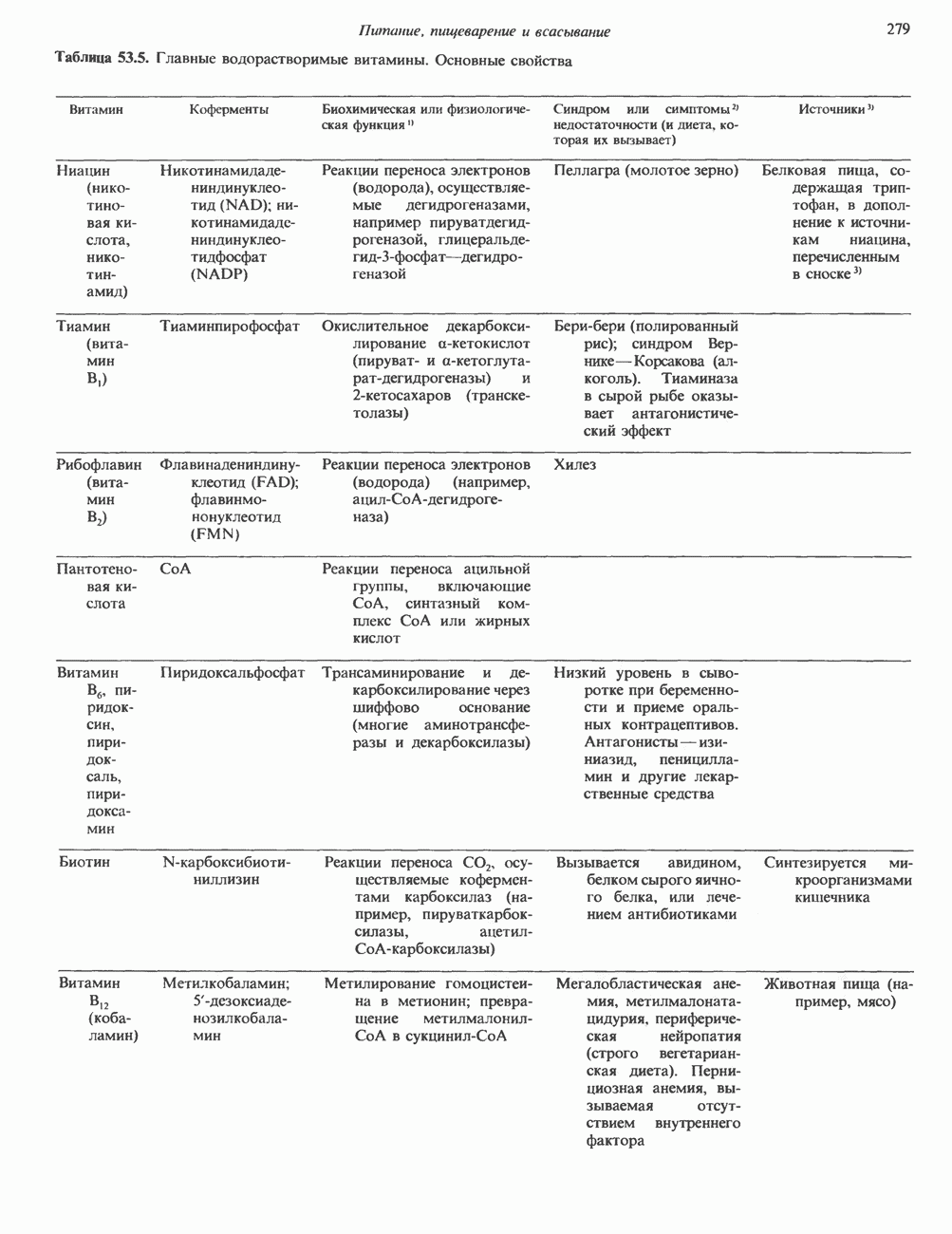
Рис. 42.2. Фосфоглицерид, состоящий из остатков жирных кислот Жирная кислота присоединена к аминогруппе сфингозина амидной связью. Первичная гидроксильная группа сфингозина этерифицирована фосфорилхолином (см. с. 157). Сфингомиелины, как следует из их названия, входят в состав миелиновой оболочки. ГликосфинголипидыГликосфинго липиды — это липиды, содержащие остатки сахаров. Сюда входят цереброзиды и ганглиозиды, а также производные сфингозина. Цереброзиды и ганглиозиды отличаются от сфингомиелина тем, какие группы присоединены к первичной гидроксильной группе сфингозина. В сфингомиелине к спиртовой группе присоединен фосфохолин, а цереброзид содержит в этом положении остаток гексозы, глюкозы или галактозы (см. с. 157). Ганглиозид содержит цепочку из трех и более сахаров, причем по крайней мере один из них — это сиаловая кислота, связанная с первичной спиртовой группой сфингозина. СтеролыНаиболее распространенным стеролом в мембранах является холестерол, который содержится почти исключительно в плазматической мембране клеток млекопитающих, но в меньшем количестве может присутствовать также в митохондриях, мембранах аппарата Гольджи и ядерных мембранах. Содержание холестерола обычно увеличивается в направлении к наружной стороне плазматической мембраны. Холестерол встраивается между фосфолипидными молекулами, причем его гидроксильная группа контактирует с водной фазой, а остальная часть располагается внутри гидрофобного слоя. При температуре выше температуры фазового перехода (см. описание жидкостно-мозаичной модели) его жесткое стерольное кольцо взаимодействует с ацильными группами фосфолипидов, ограничивая их подвижность; это приводит к уменьшению текучести мембран. С друг ой стороны, при температурах, близких к температуре фазового перехода, взаимодействие холестерола с ацильными цепями препятствует их взаимному упорядочиванию. В результате снижается температура, при которой происходит переход жидкость—гель, а это помогает поддерживать текучесть мембраны при более низких температурах. Амфифильная природа мембранВсе основные липиды мембран содержат как гидрофобные, так и гидрофильные области и поэтому называются амфифильными соединениями. Таким образом, мембраны тоже амфифильны. Если бы основу молекулы составляли гидрофобные группы, она была бы нерастворима в воде и растворима
Рис. 42.3. Схематическое представление фосфолипида или другого мембранного липида. Полярная головка гидрофильна, а углеводородные хвосты гидрофобны или липофильны. Жирнокислотные хвосты могут быть насыщенными (S) или ненасыщенными (U); первые обычно присоединены к атому углерода 1 остатка глицерола, а последние — к атому углерода 2. в маслах. Напротив, если бы основу молекулы составляли гидрофильные участки, то она была бы нерастворима в маслах и растворима в воде. Амфифильные мембранные липиды содержат полярную «головку» и неполярные «хвосты» (рис. 42.3). Насыщенные жирнокислотные «хвосты» находятся в вытянутой конформации, а ненасыщенные, находящиеся в мембране в основном в
|
 |
Оглавление
|






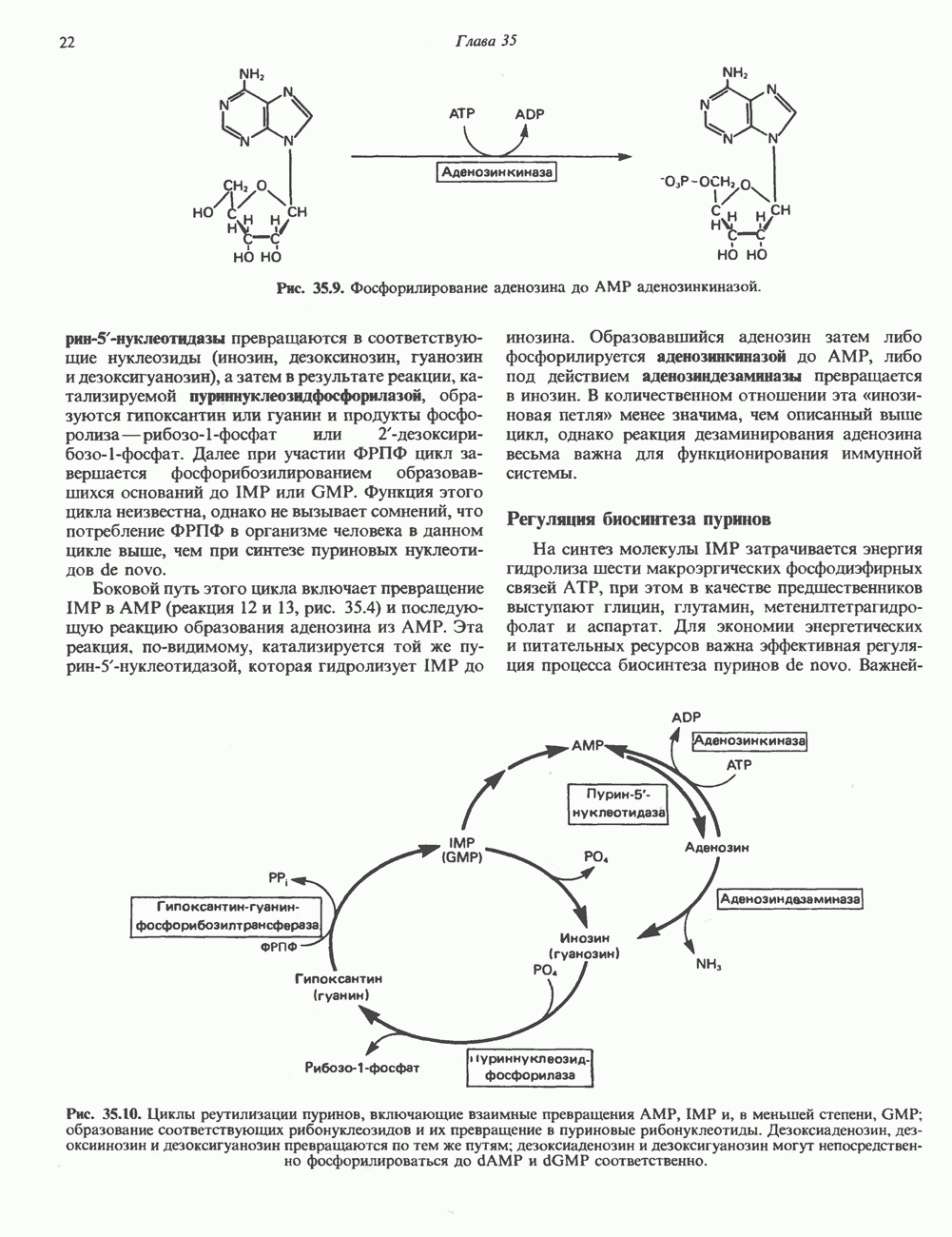
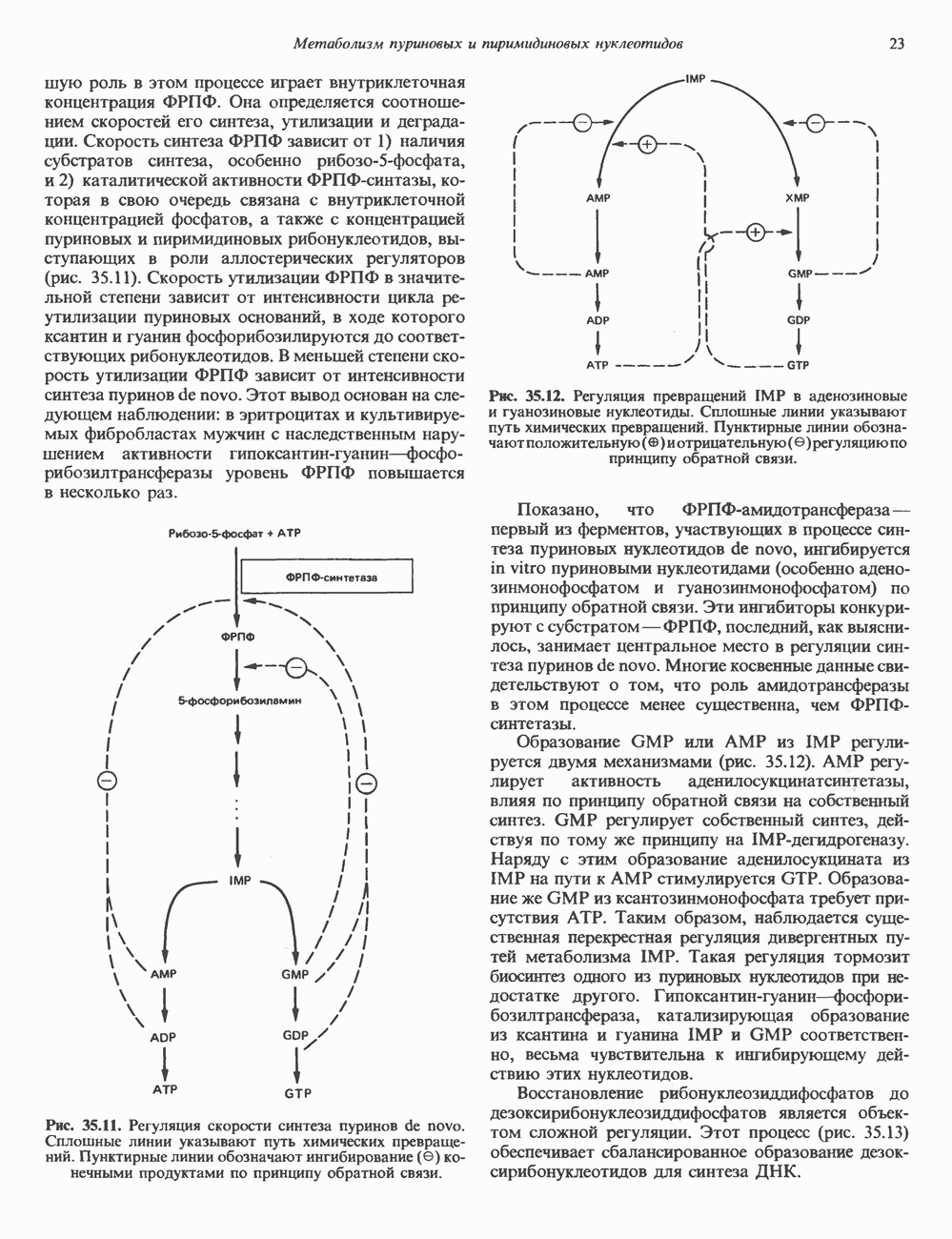
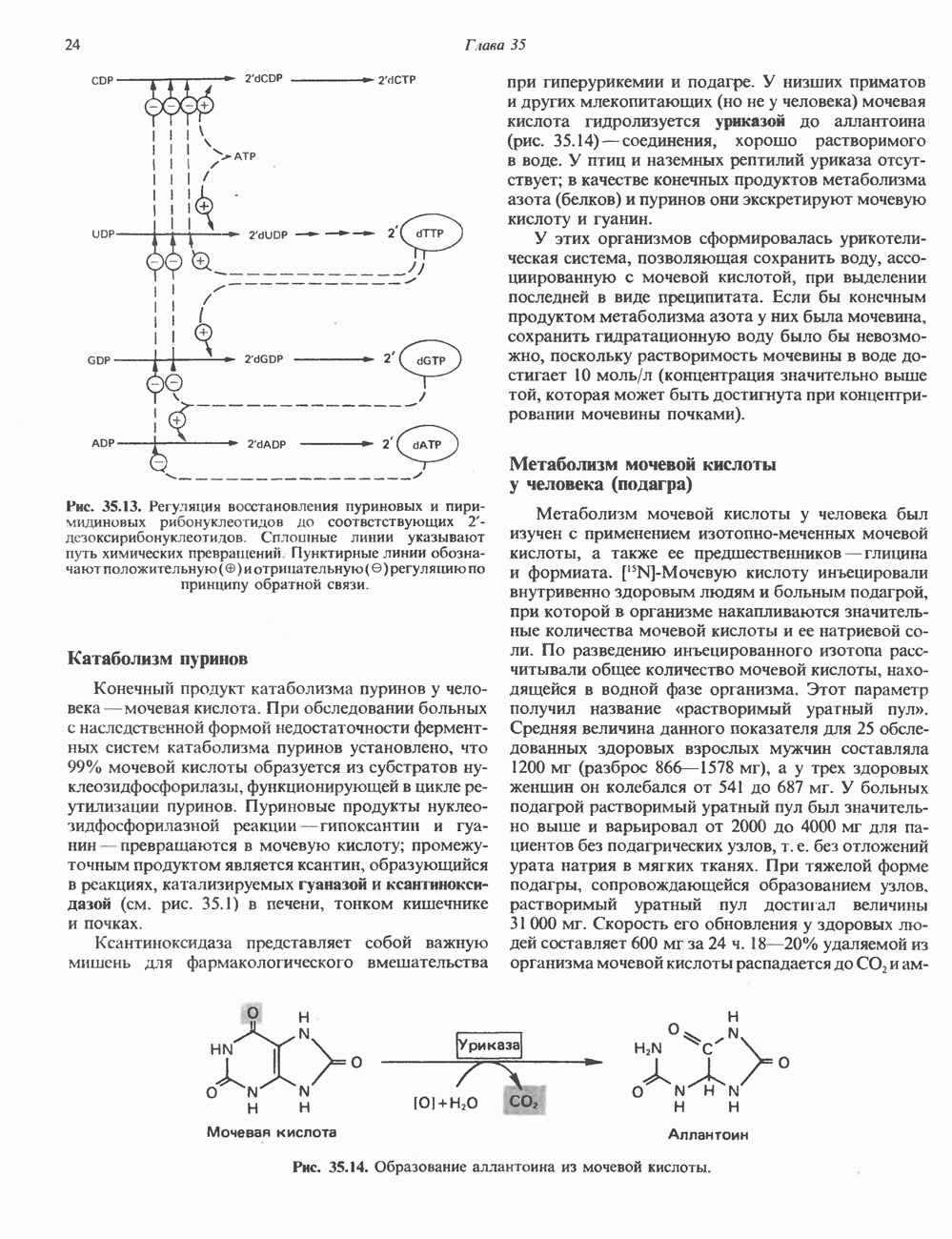

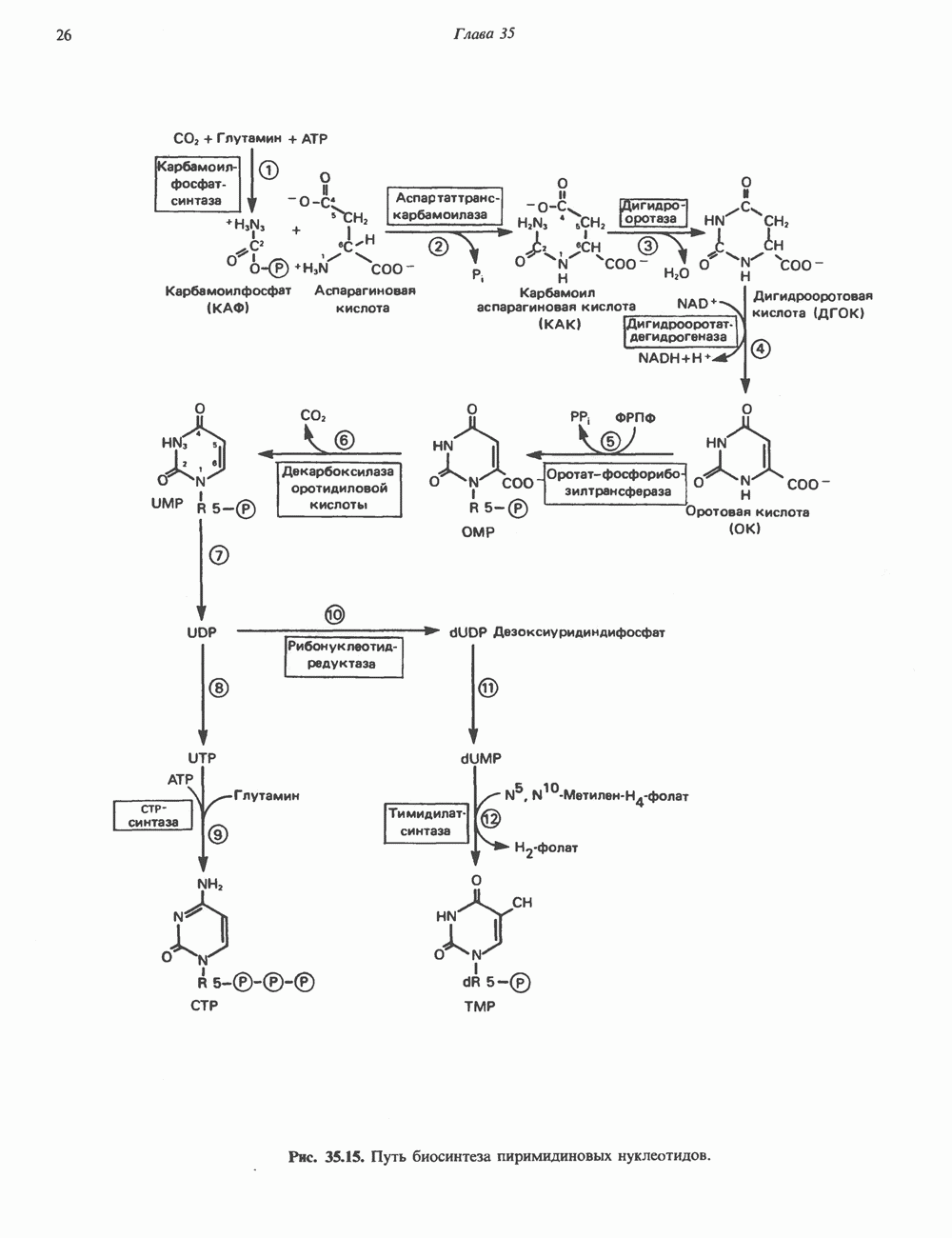












































































































































































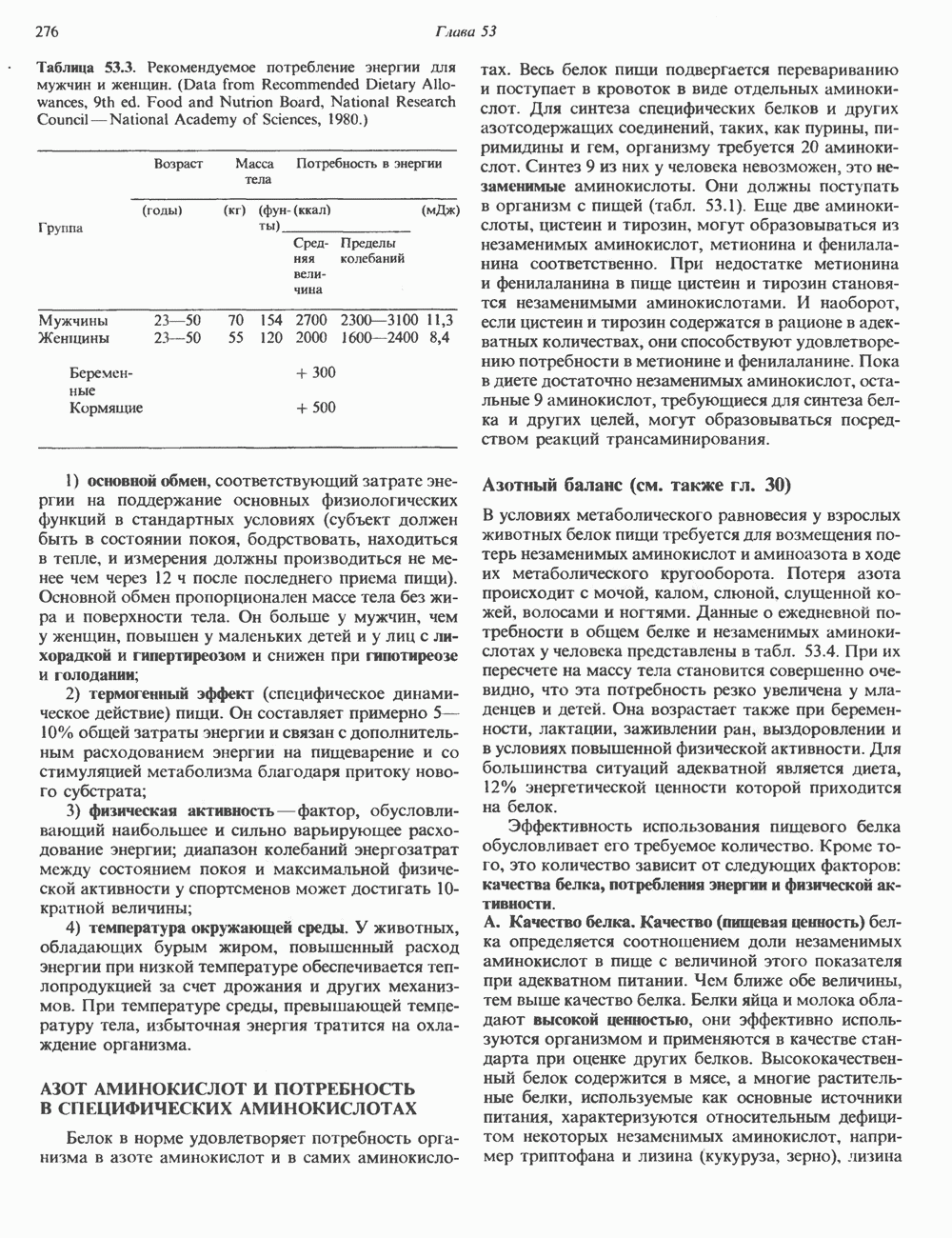





































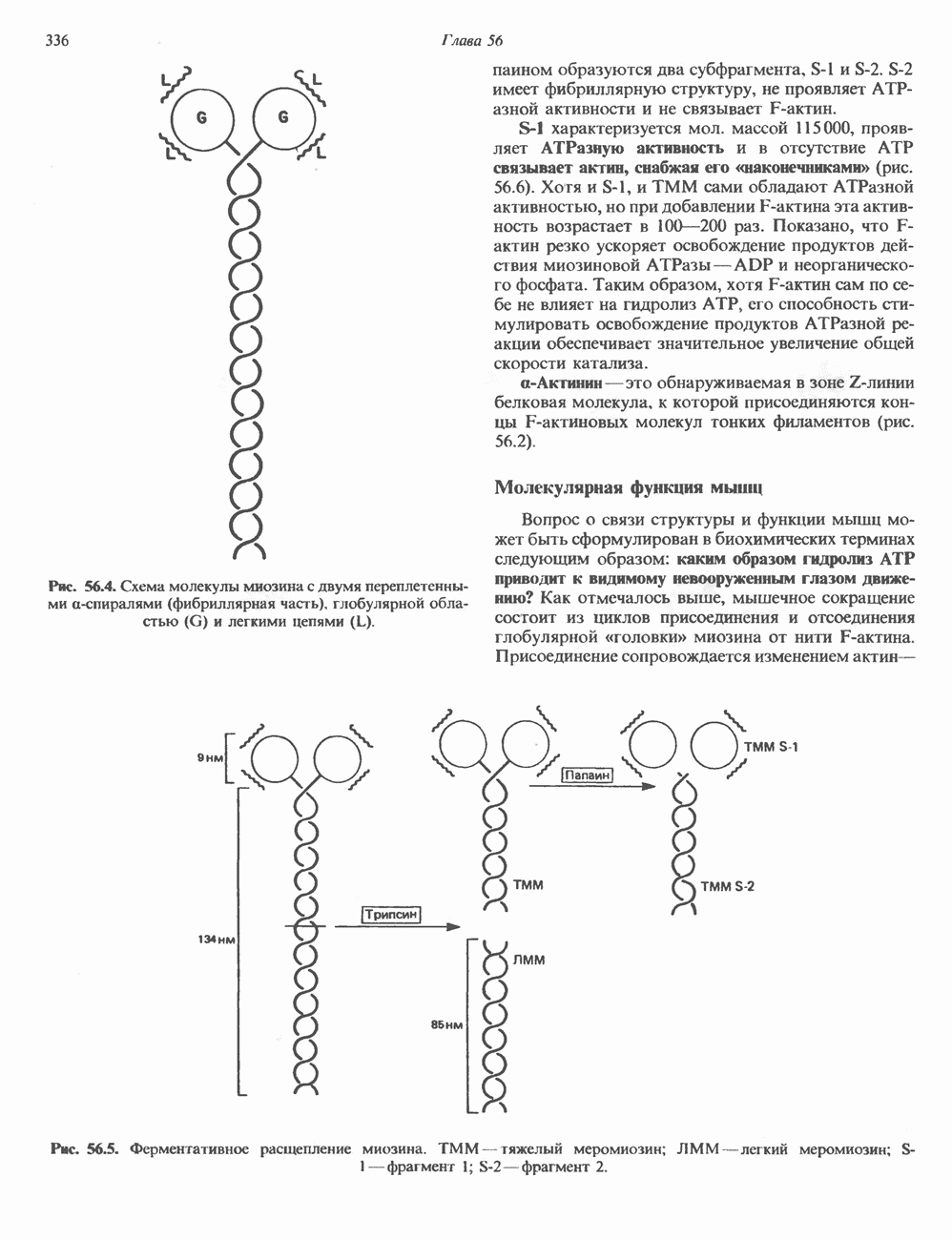


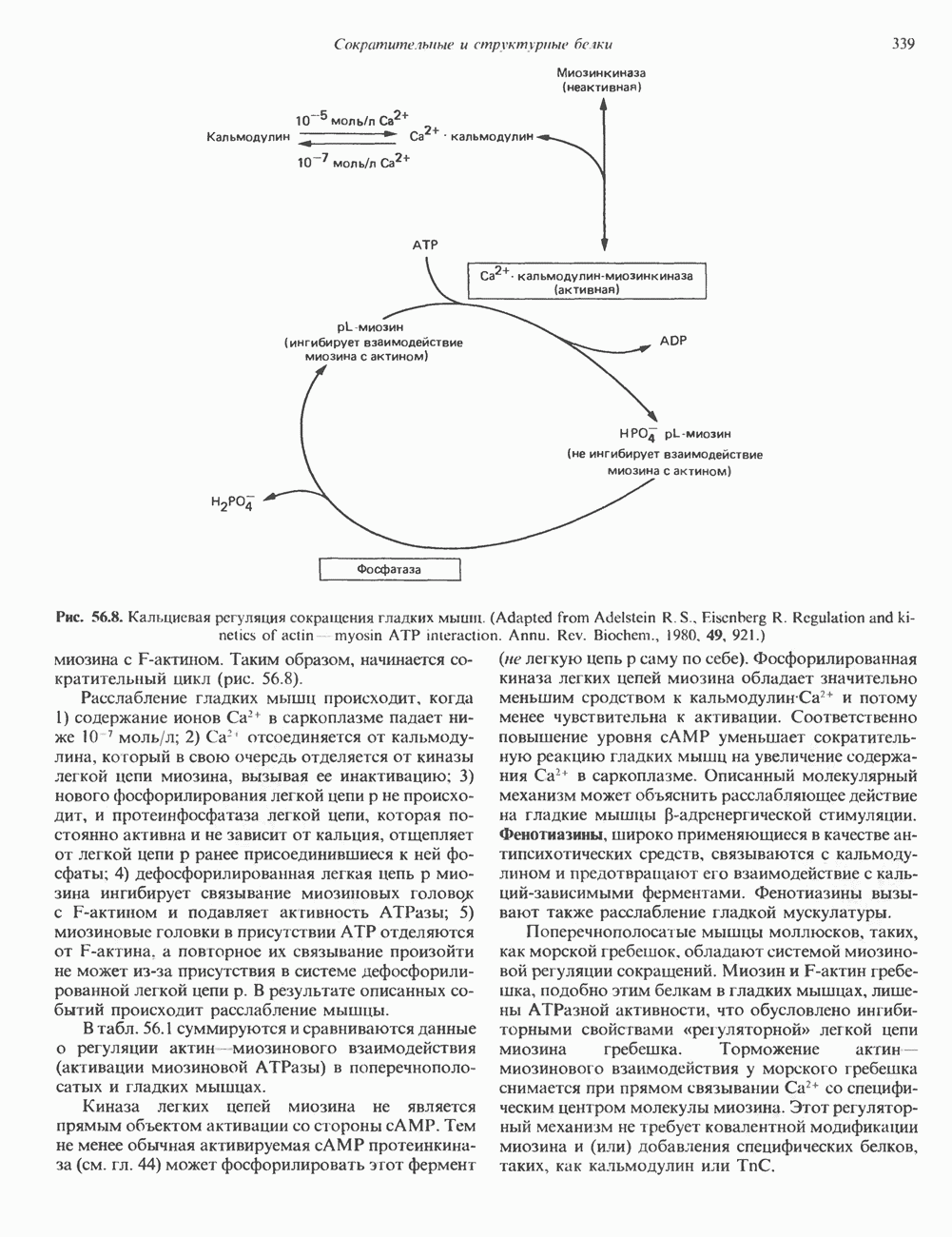



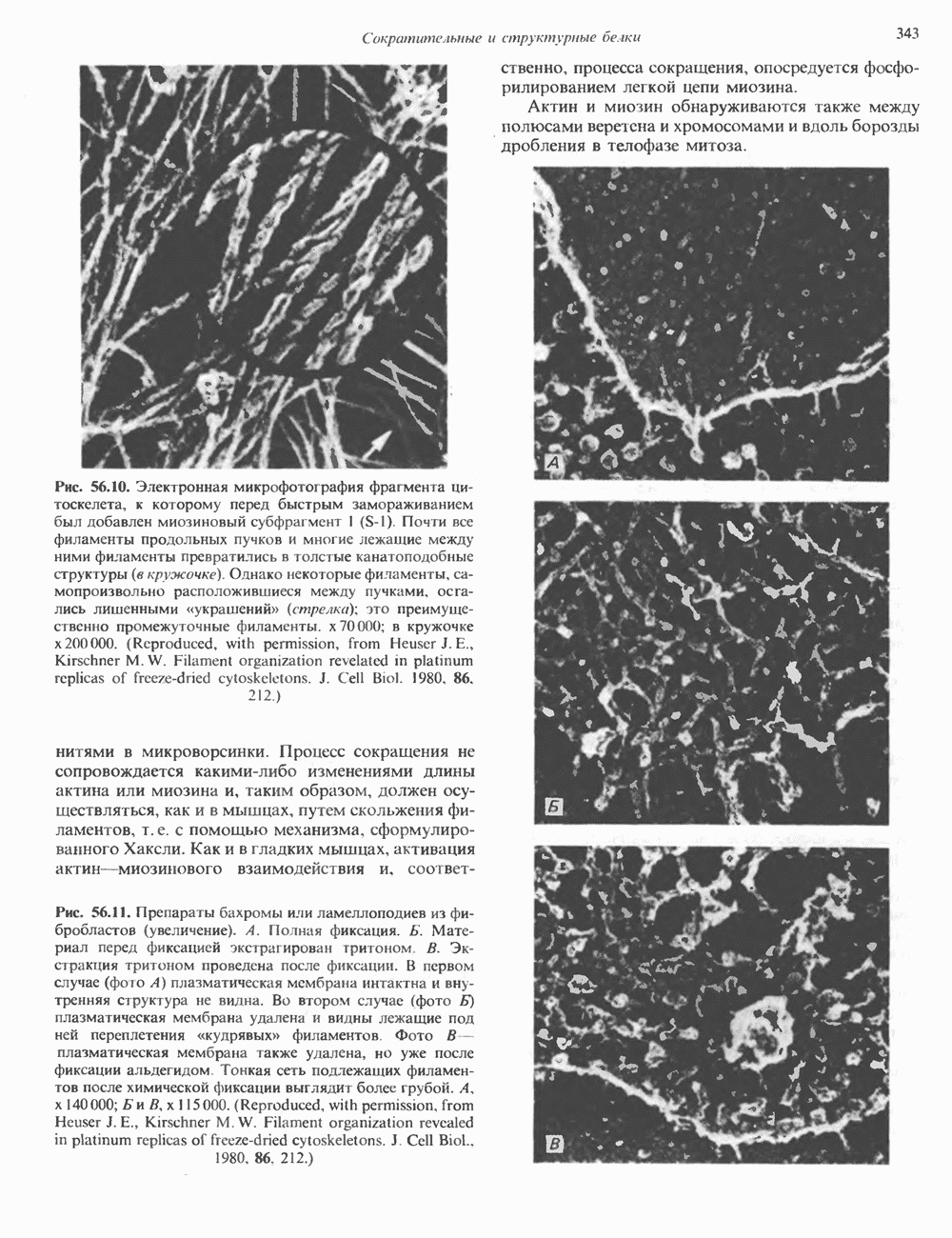
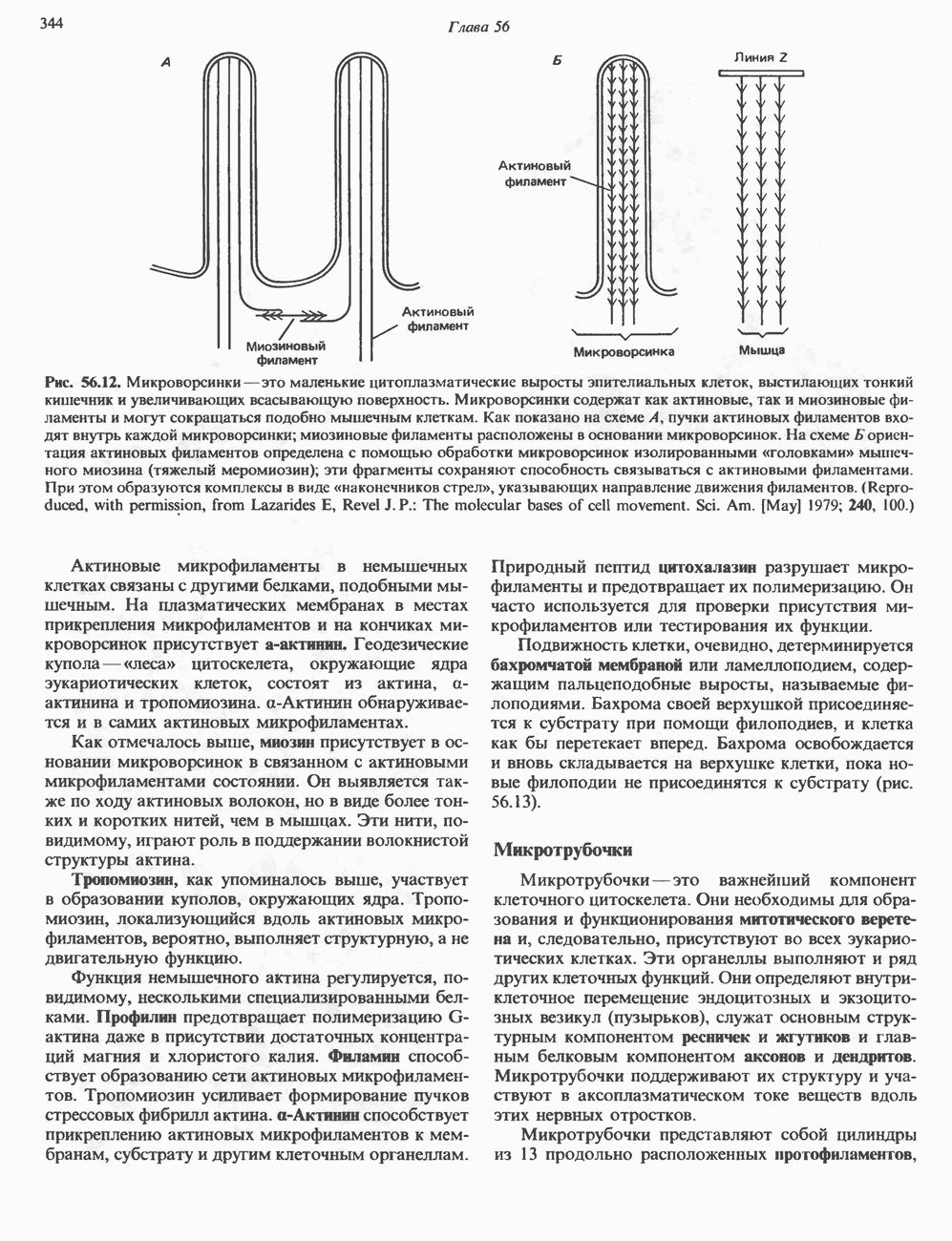








































 глицерола и фосфорилированных спиртовых компонентов. В фосфатидной кислоте
глицерола и фосфорилированных спиртовых компонентов. В фосфатидной кислоте  — это атом водорода.
— это атом водорода.

 -форме, могут иметь изломы. Чем больше таких изломов, тем менее плотна упаковка липидов в мембране и соответственно тем больше ее текучесть. Важную роль в биохимии и быту играют такие амфифильные соединения, как детергенты. Их структура не так уж далека от структуры липидов.
-форме, могут иметь изломы. Чем больше таких изломов, тем менее плотна упаковка липидов в мембране и соответственно тем больше ее текучесть. Важную роль в биохимии и быту играют такие амфифильные соединения, как детергенты. Их структура не так уж далека от структуры липидов.