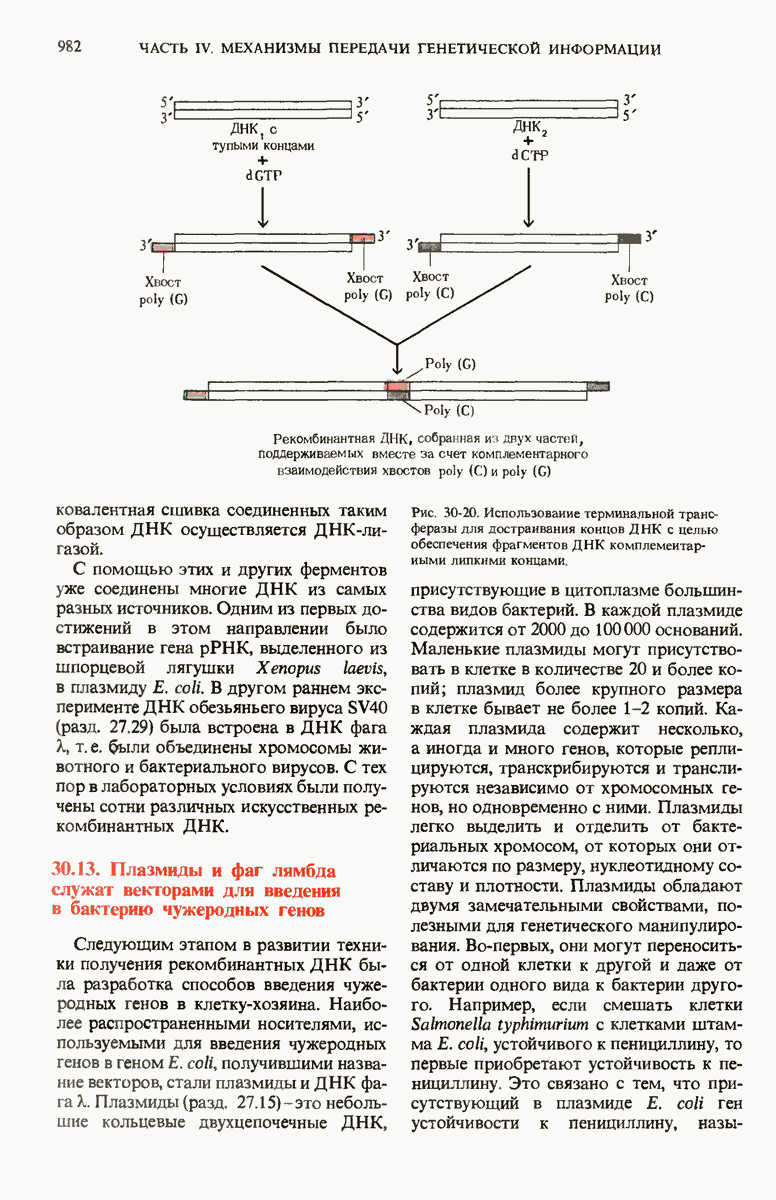
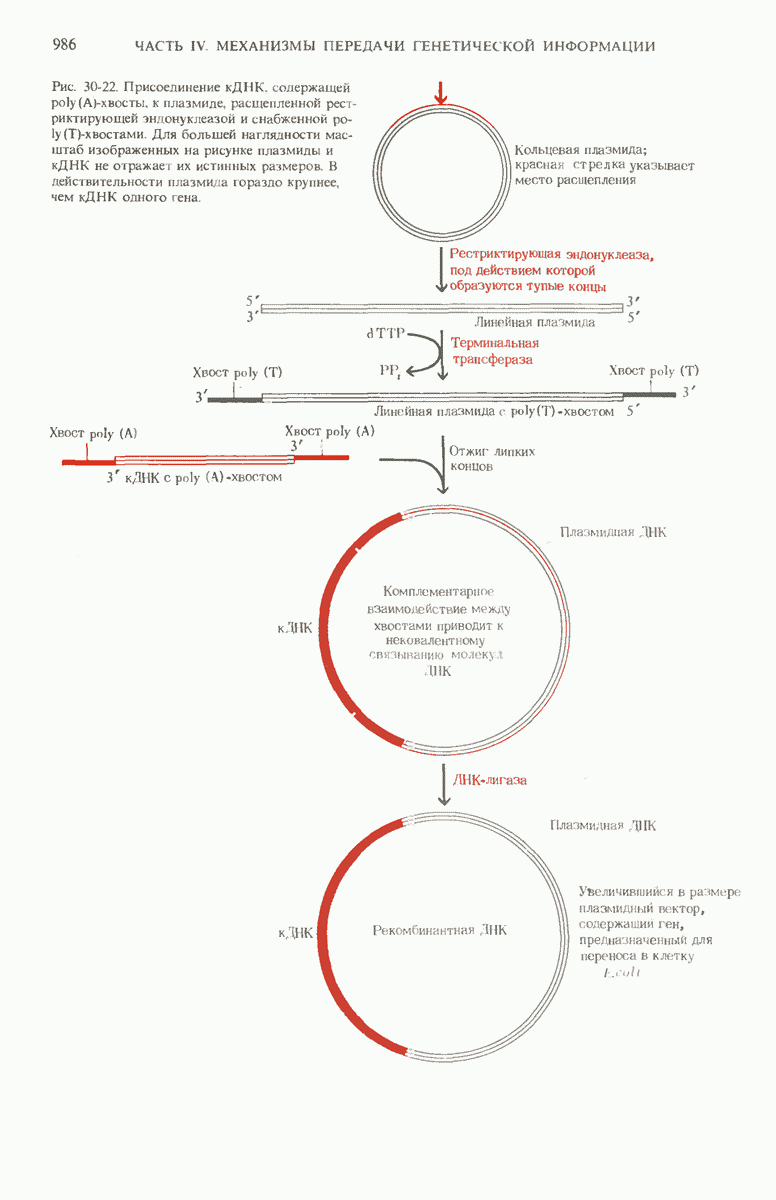
Краткое содержание главы
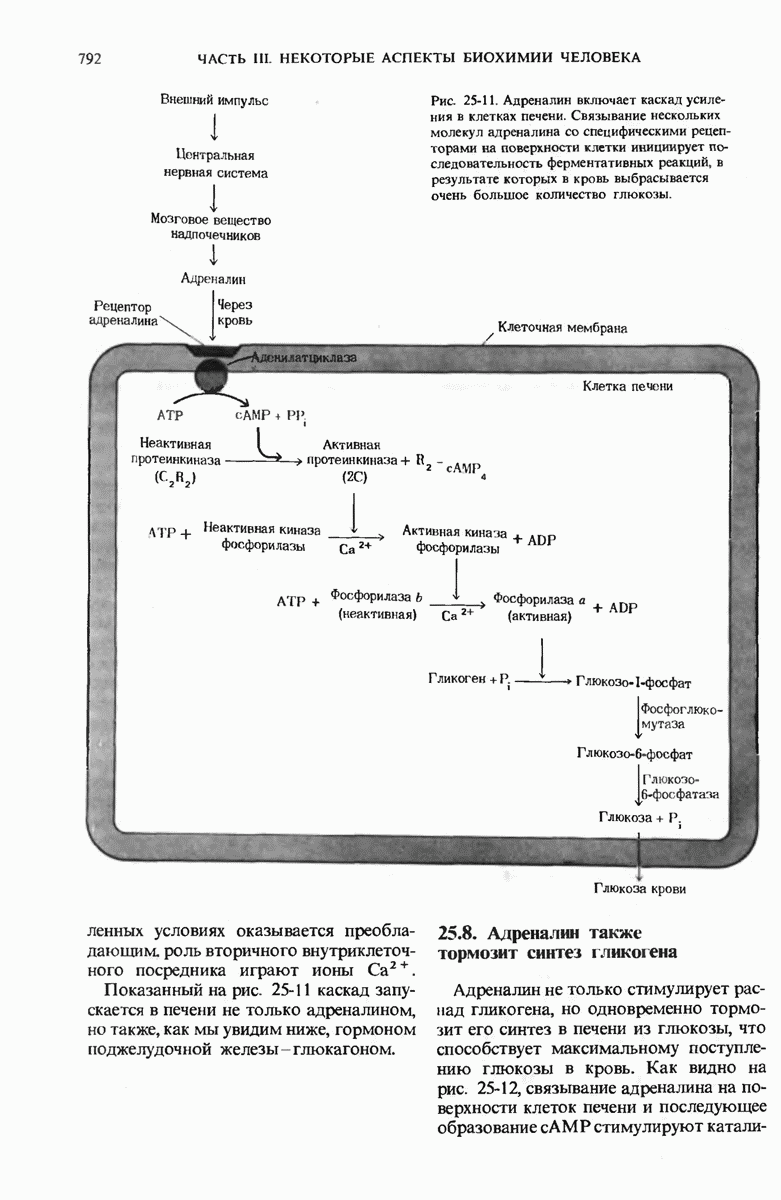
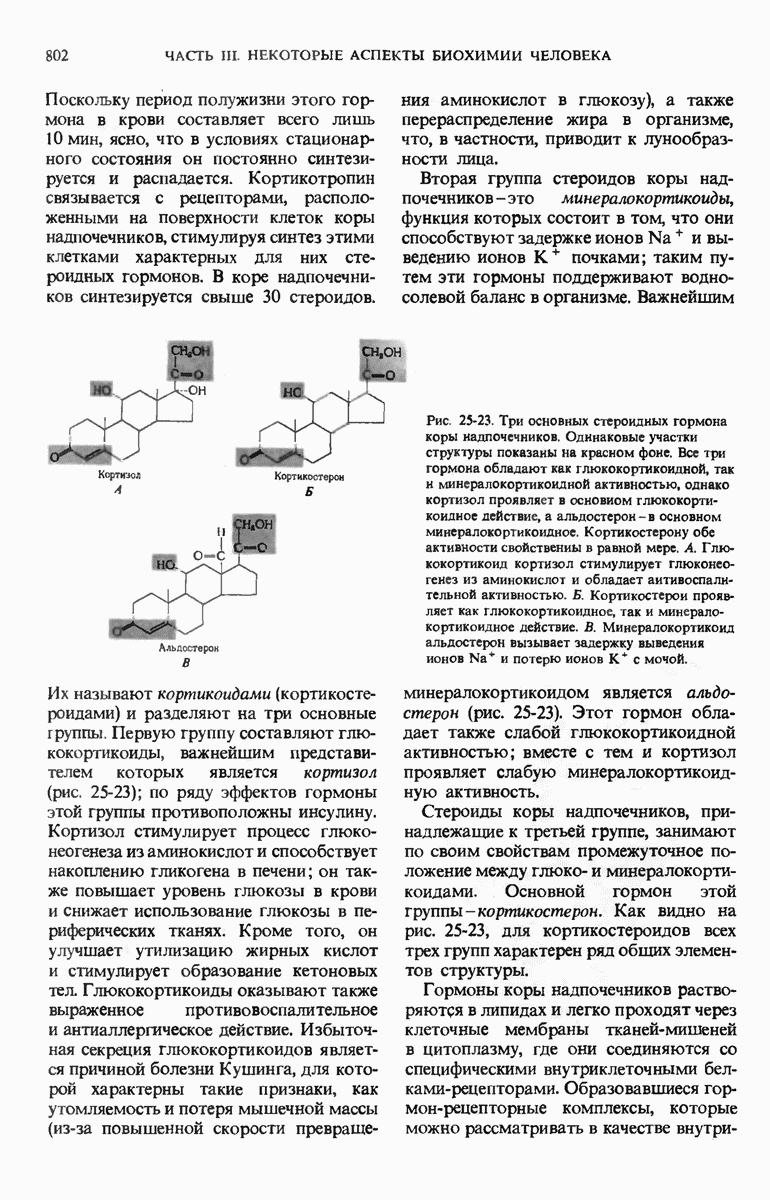
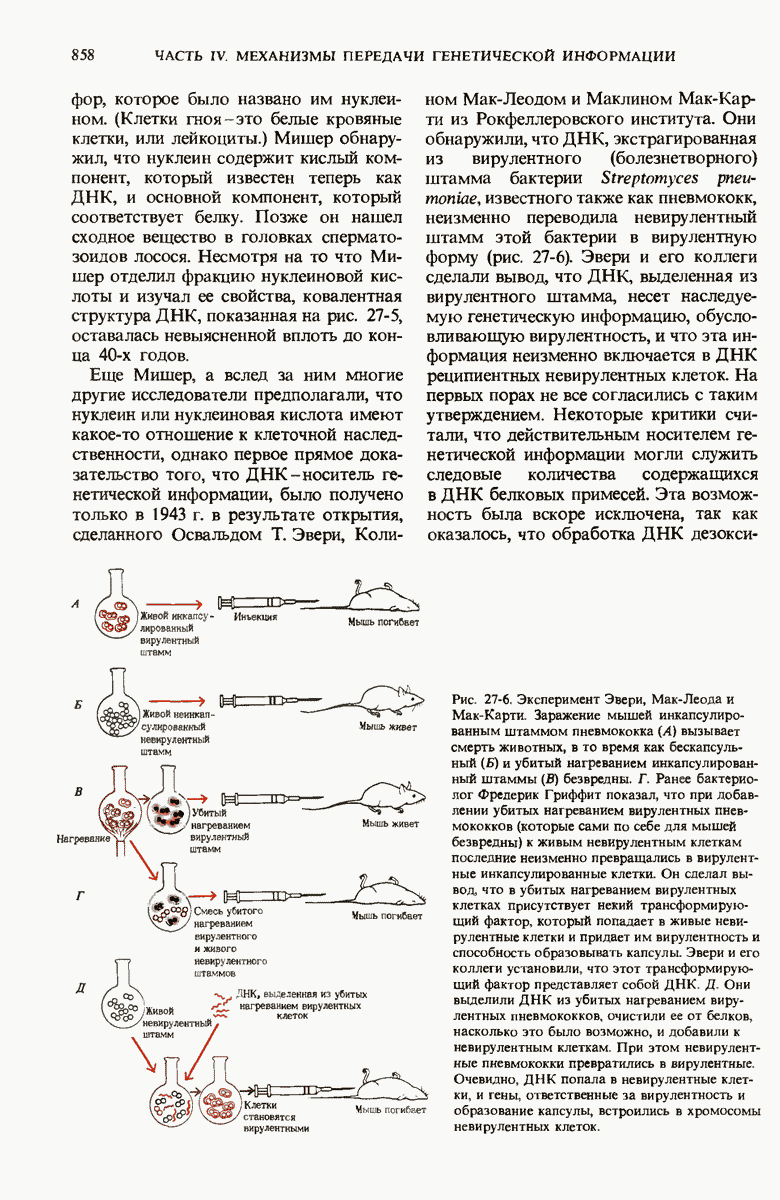
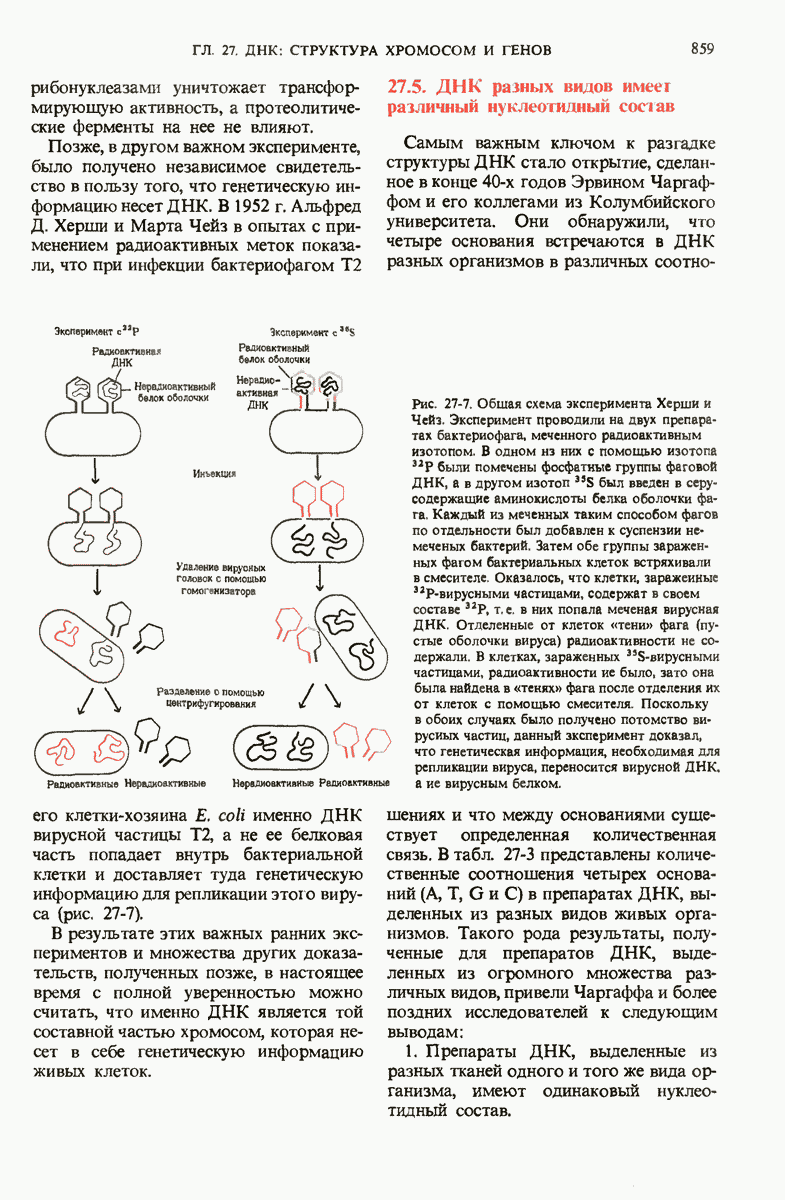
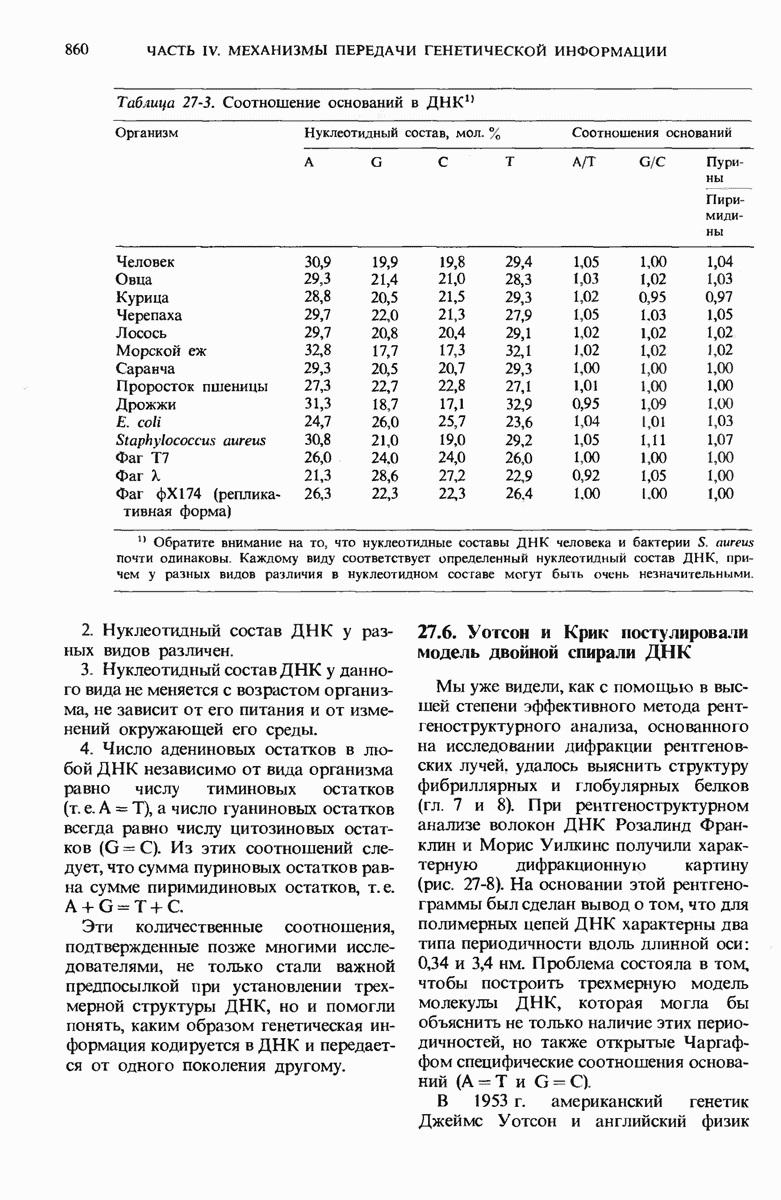
Роль ДНК как носителя генетической информации подтверждается целым рядом фактов. Эксперимент Эвери, МакЛеода и Мак-Карти показал, что ДНК, выделенная из одного штамма бактерий, способна проникнуть в клетки другого штамма и трансформировать их, придавая им некоторые наследуемые признаки донора. Опыт Херши и Чейз продемонстрировал, что именно ДНК бактериофага, а не его белковая оболочка несет генетическое сообщение для репликации вируса в клетке-хозяине. Все соматические клетки организма данного вида содержат ДНГ с одинаковым нуклеотидным составом, который не зависит ни от питания, ни от условий окружающей среды. Хотя нуклеотидный состав ДНК у разных ввдов различен, в двухцепочечных ДНК всех видов число остатков аденина всегда равно числу остатков тимина, а число гуаниновых остатков всегда равно числу цитозиновых остатков.
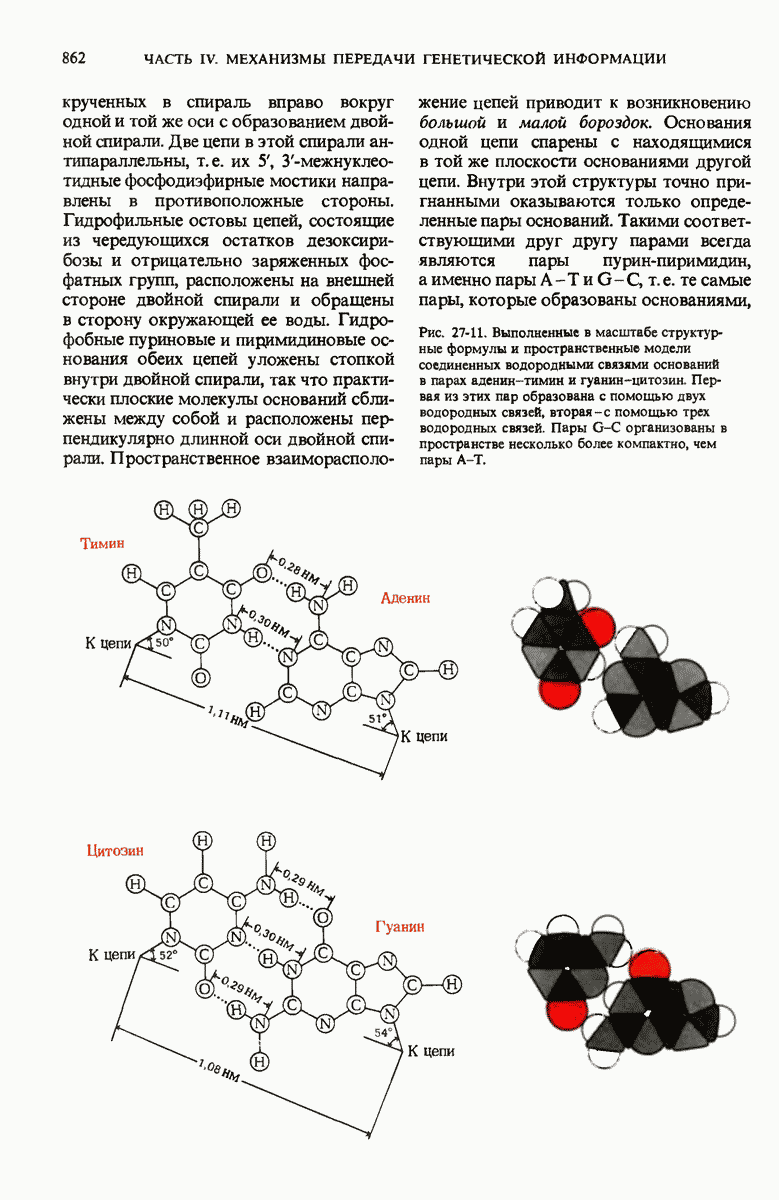
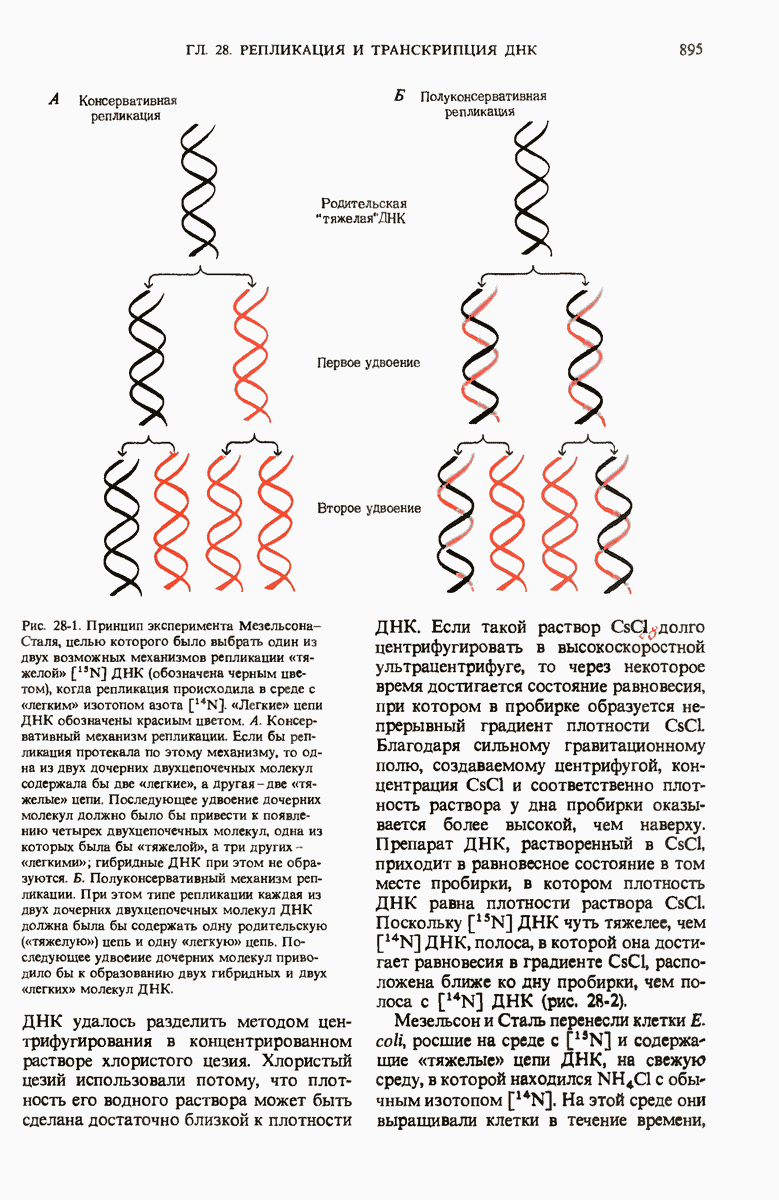
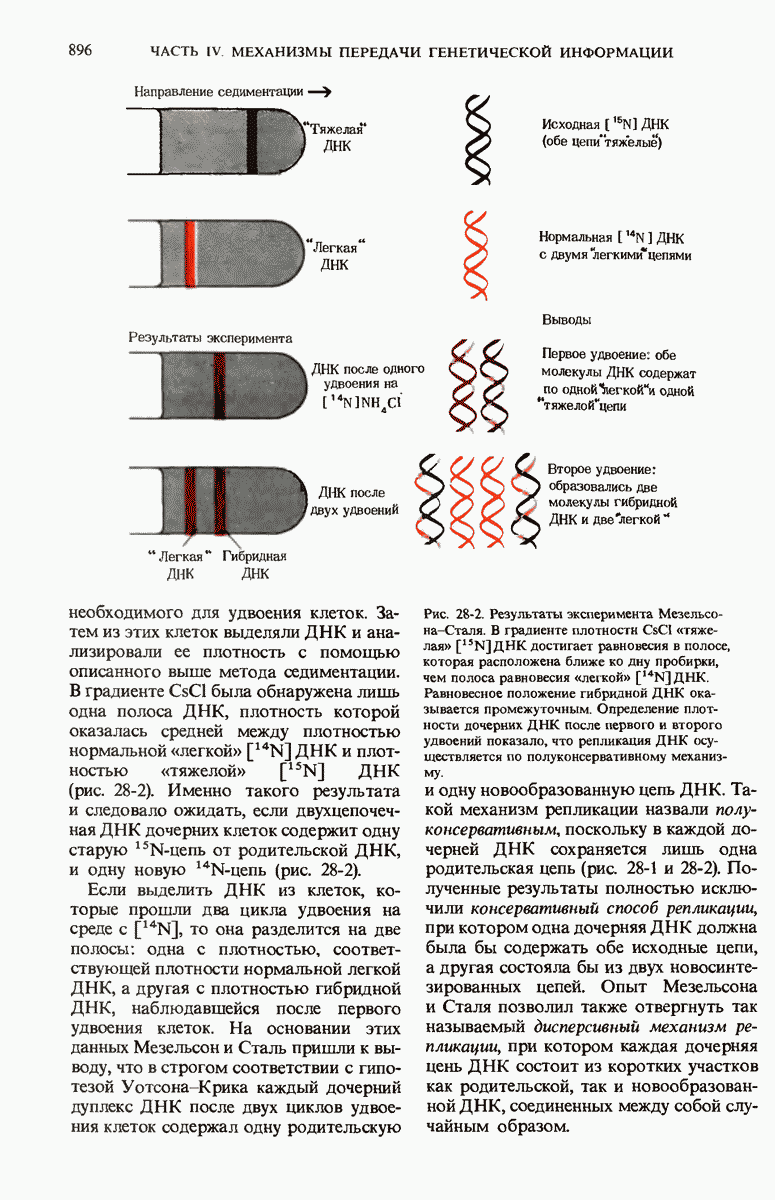
На основании рентгеноструктурного анализа волокон ДНК и принципа комплементарности оснований в ДНК Уотсон и Крик пришли к заключению, что нативная ДНК состоит из двух антипараллельных цепей, скрученных в двойную спираль. Комплементарные основания А—Т и G—С образуют с помощью водородных связей пары внутри спирали, а гидрофильный сахарофосфатный остов располагается с наружной стороны макромолекулы. Пары оснований плотно уложены в стопку перпендикулярно длинной оси на расстоянии 0,34 нм друг от друга; на один полный виток двойной спирали приходится приблизительно 10 нуклеотидных пар оснований. Комплементарность цепей в двойной спирали позволяет понять механизм их точной репликации.
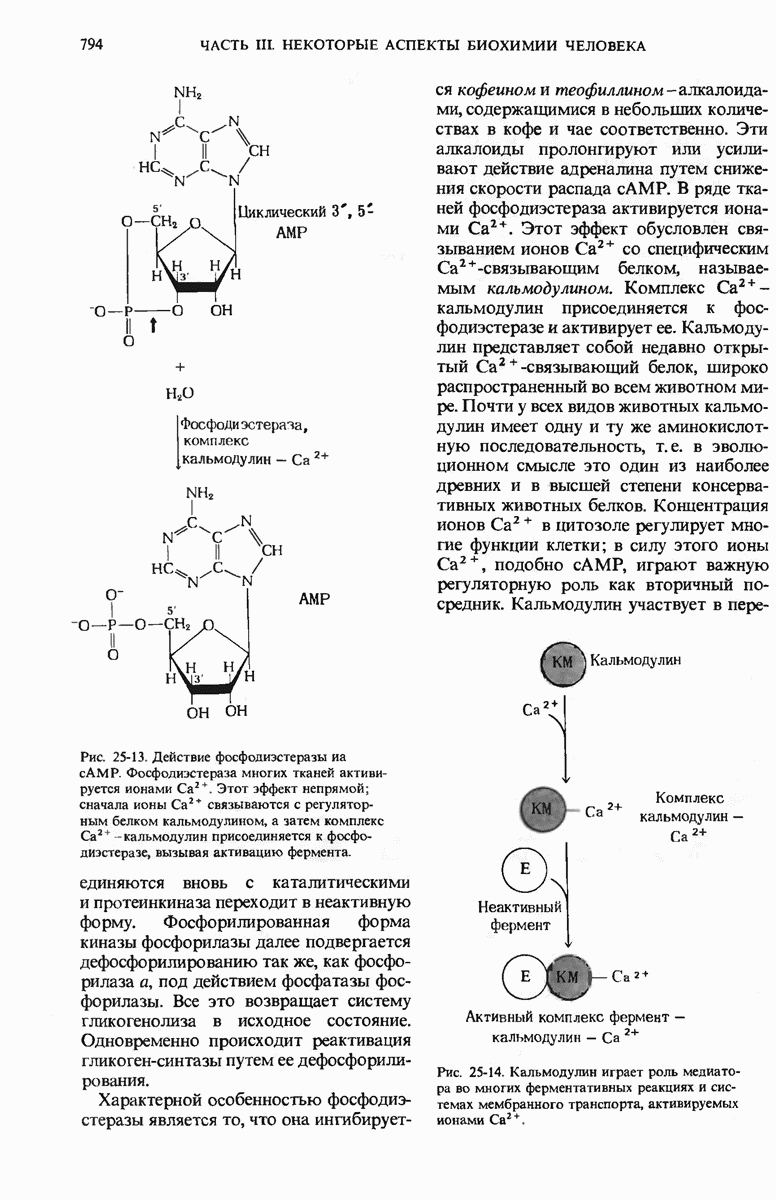
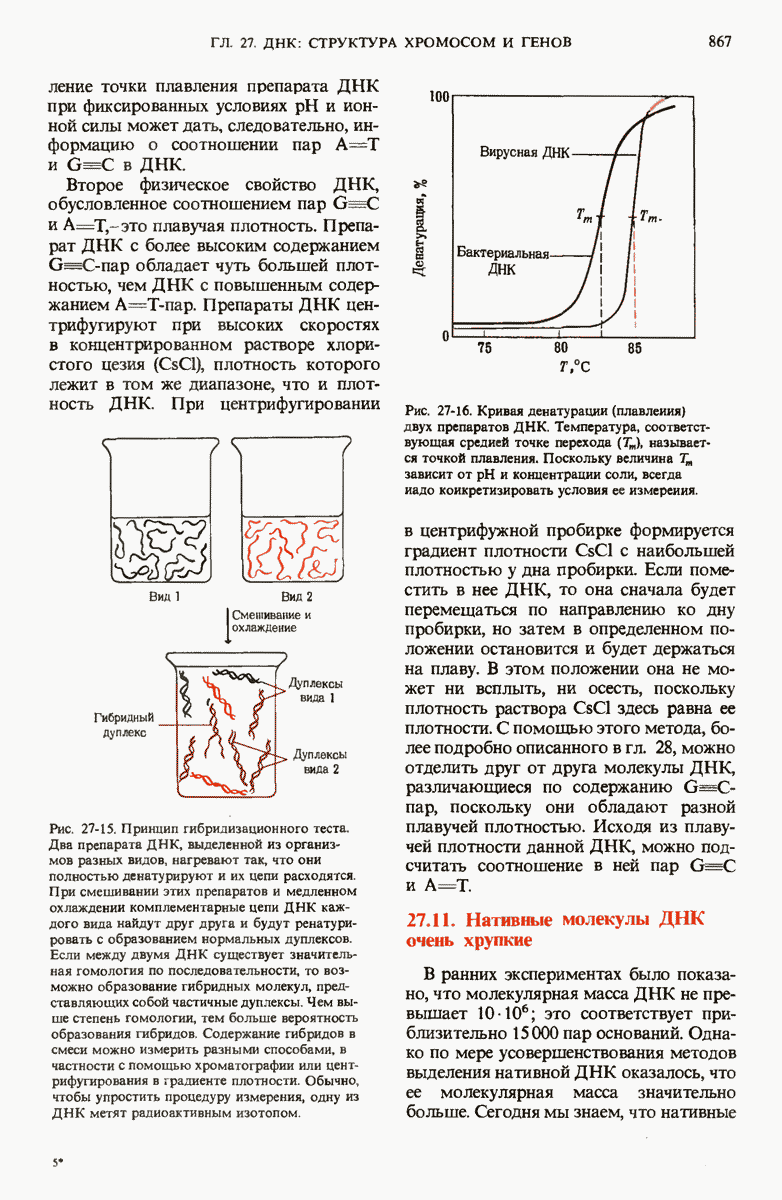
При нагревании или при экстремальных значениях pH нативная ДНК обратимо расплетается и ее цепи разделяются. Поскольку пары оснований G=C более стабильны, чем пары А=Т, точка плавления ДНК, богатой парами G=C, выше точки плавления ДНК с большим содержанием пар А=Т. Денатурированная одноцепочечная ДНК из одного вида может образовать гибридный дуплекс с денатурированной одноцепочечной ДНК из другого вида при условии, что нуклеотидные последовательности этих двух цепей имеют хотя бы некоторое сходство (гомологию). На основании эффективности образования таких гибридов судят о возможном родстве разных видов и о гомологии между ДНК и РНК.
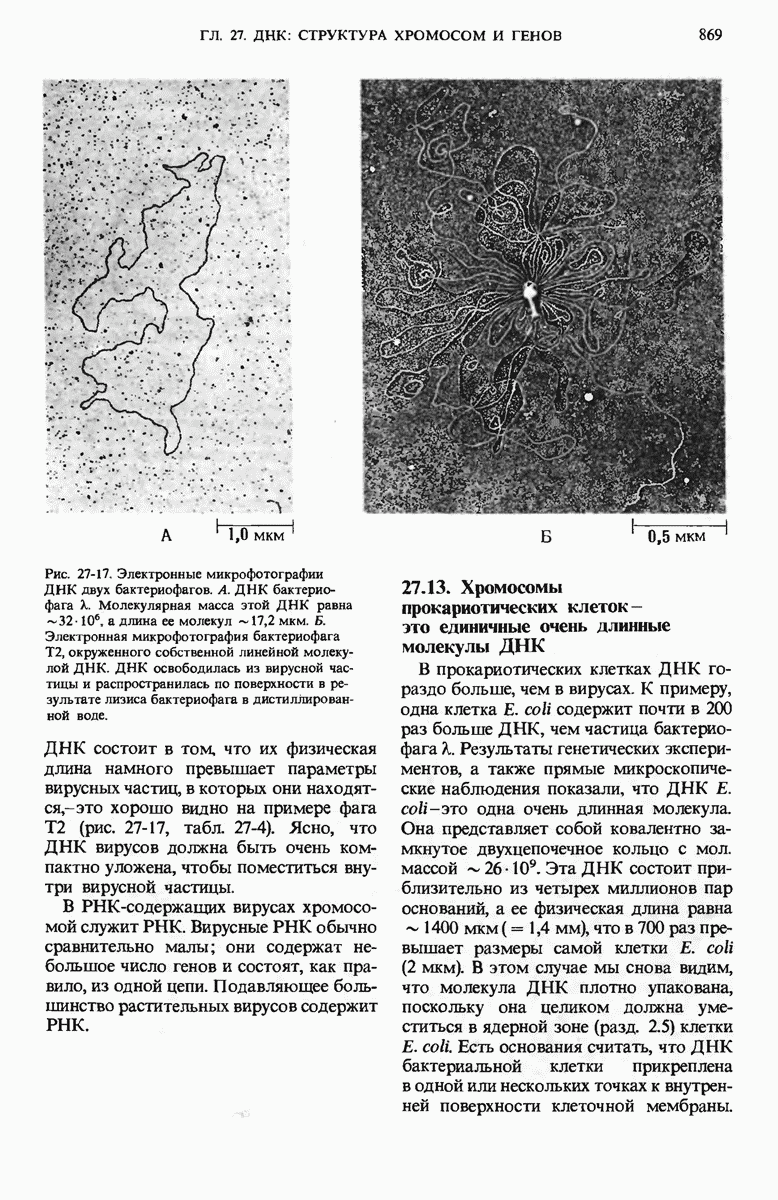
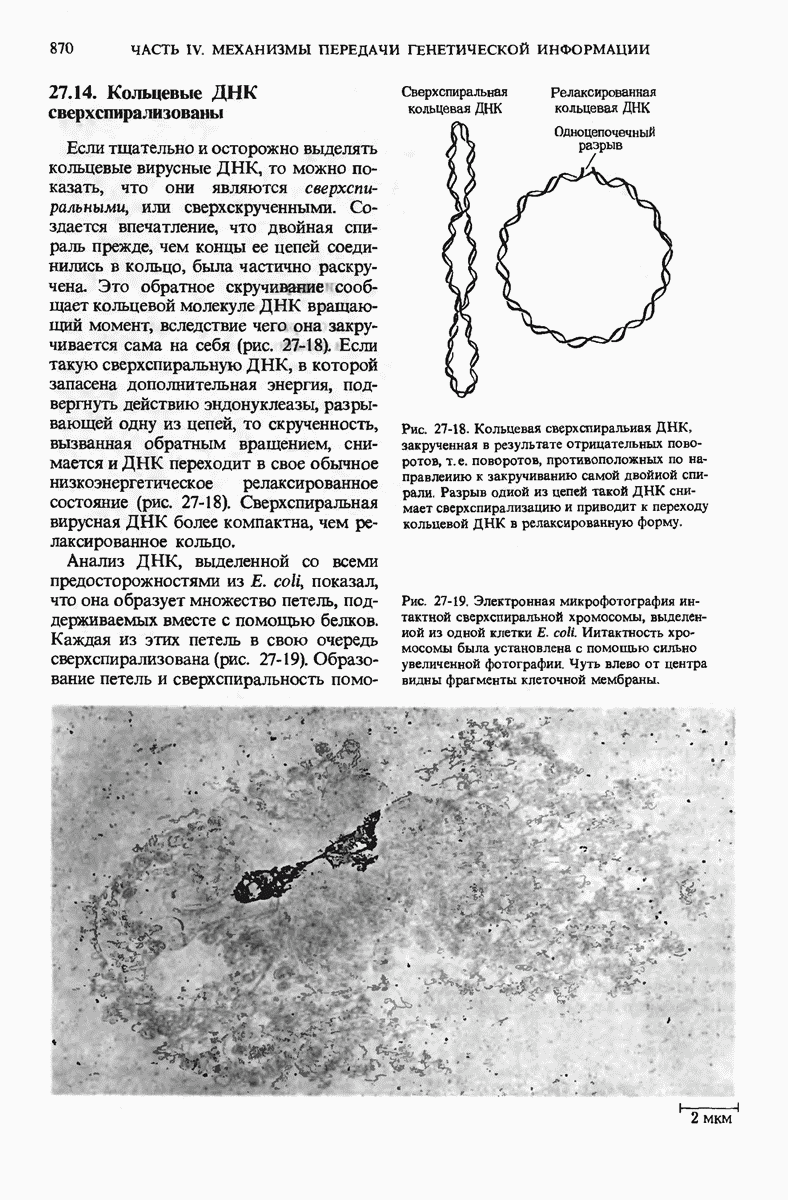
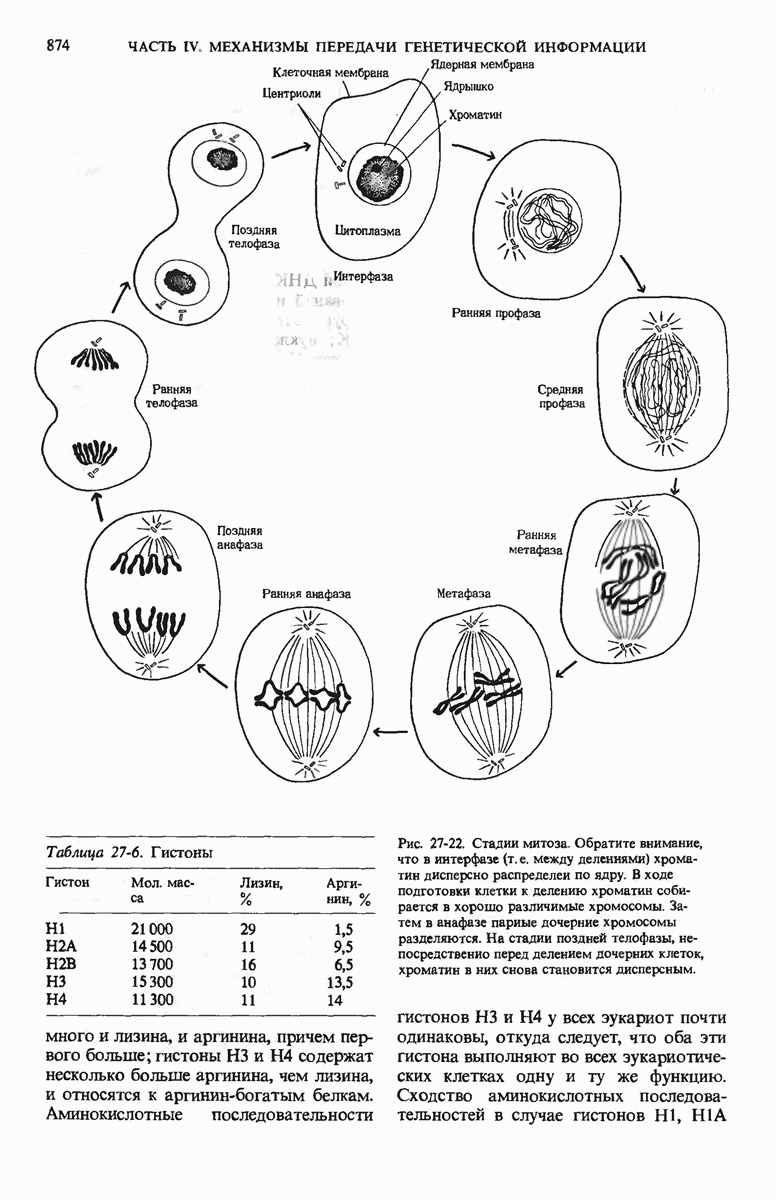
У ДНК-содержащих вирусов бактерий единственная двухцепочечная молекула ДНК может быть либо замкнутой в кольцо, либо линейной; некоторые вирусные ДНК, например ДНК фХ174, - это одноцепочечные молекулы. Вирусные ДНК сверхспирализованы, что облегчает их плотную упаковку внутри вириона. Единственная хромосома бактерий представляет собой значительно больший по размеру, ковалентно замкнутый кольцевой дуплекс. Бактериальная ДНК свернута с образованием большого числа петель, каждая из которых сверхспирализована. Эукариотические клетки содержат много хромосом, причем каждая хромосома представляет собой одну очень длинную линейную молекулу ДНК, в 4-100 раз превосходящую по длине единственную хромосому прокариот. Эукариотическая ДНК обматывает белковые частицы, состоящие из нескольких молекул основных белков - гистонов, которые располагаются вдоль ДНК через определенные интервалы. Такие комплексы участков ДНК и гистонов называются нуклеосомами.
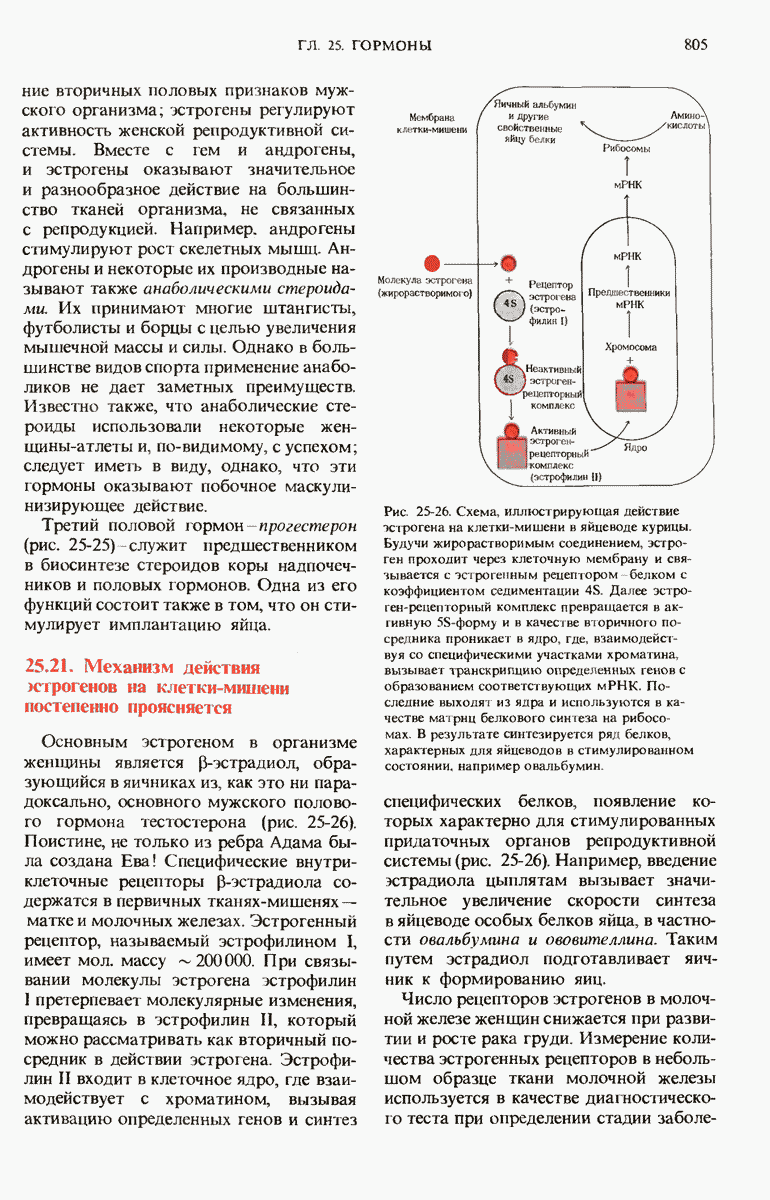
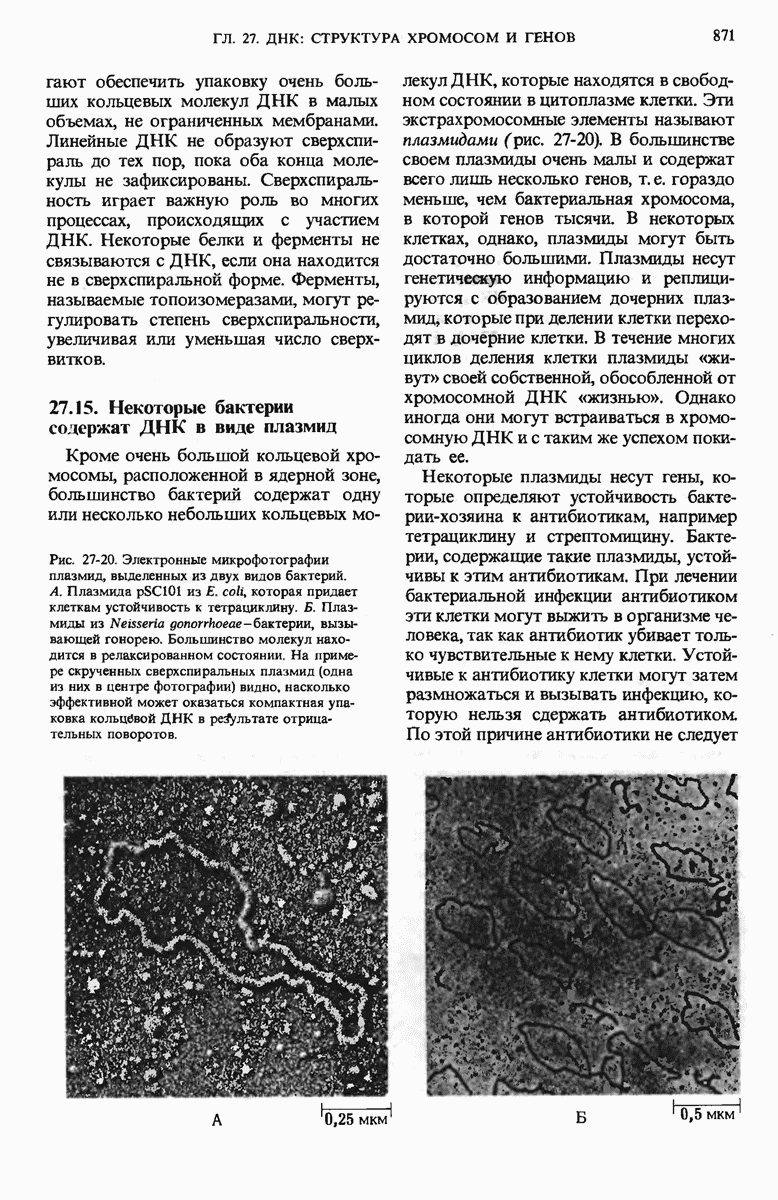
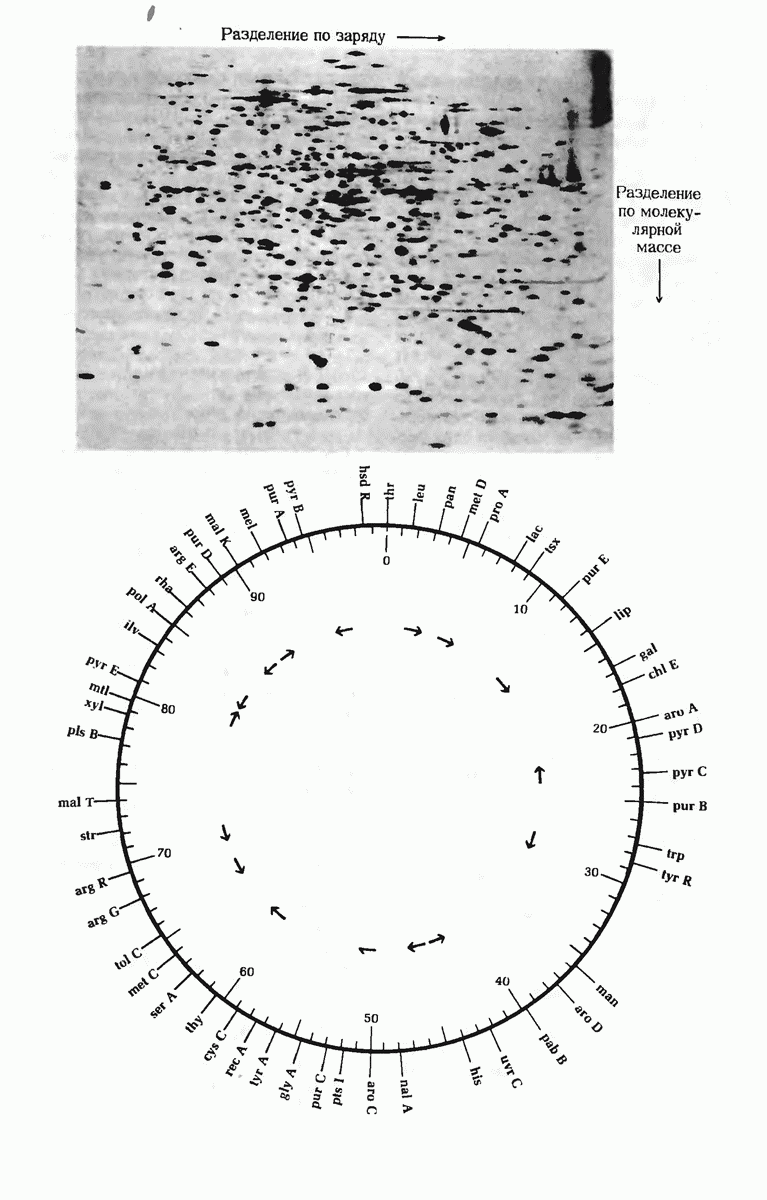
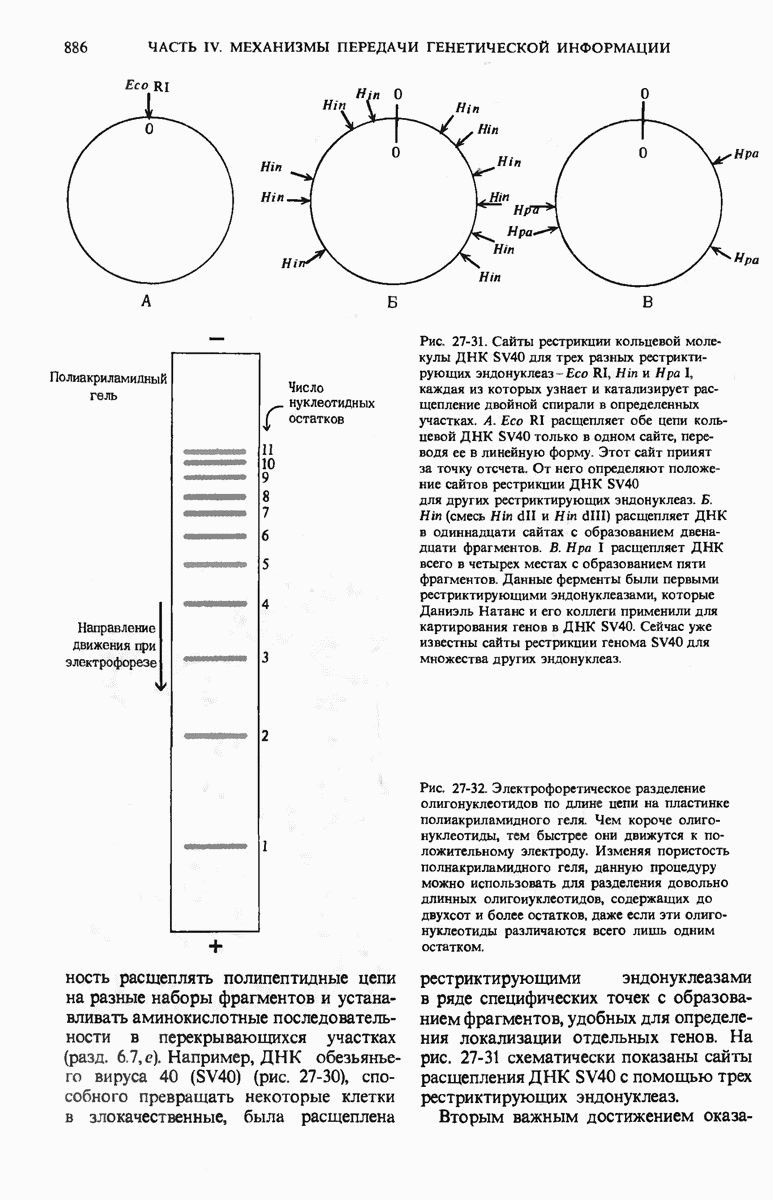
Структурные гены - это участки ДНК, которые кодируют полипептидные цепи, тРНК и рРНК. Вирусные ДНК содержат сравнительно небольшое число генов в отличие от ДНК Е. coli, содержащей более 3000 генов. К настоящему времени расположение многих из них в кольцевой хромосоме уже установлено. Бактерии защищают свою собственную ДНК путем метилирования некоторых оснований, расположенных в определенных местах молекулы, с помощью модифицирующих метилаз.
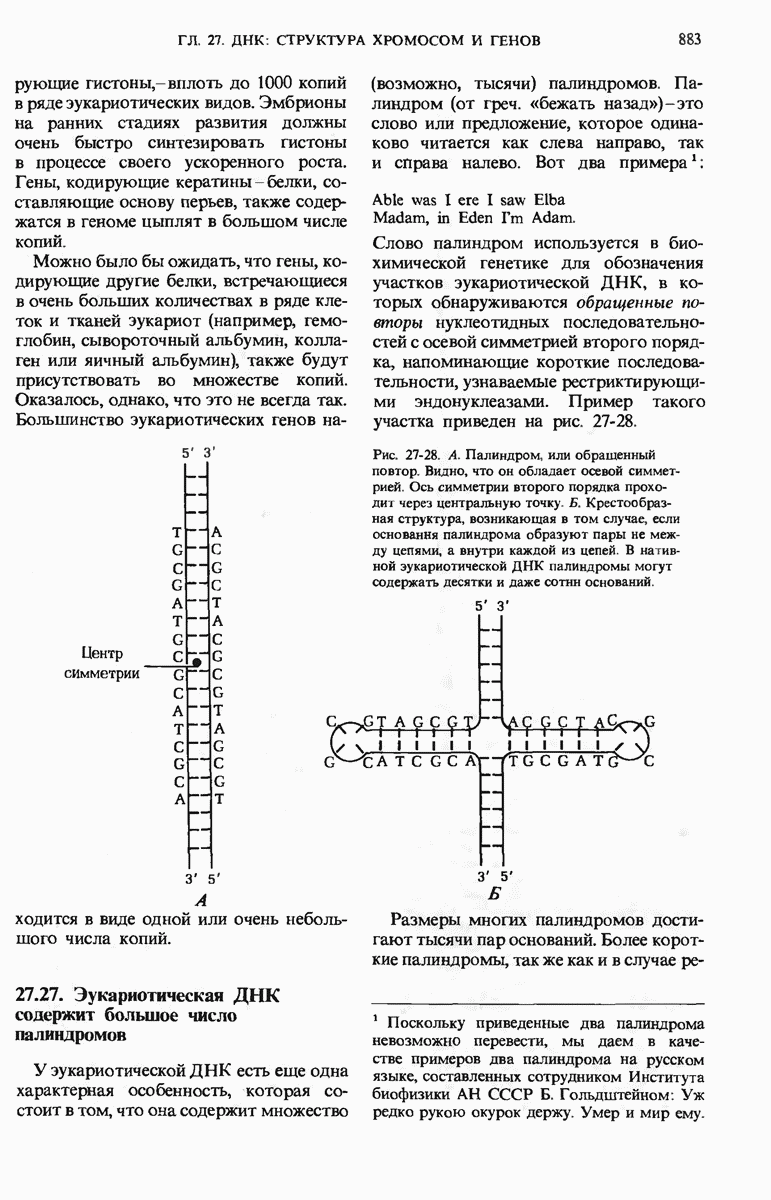
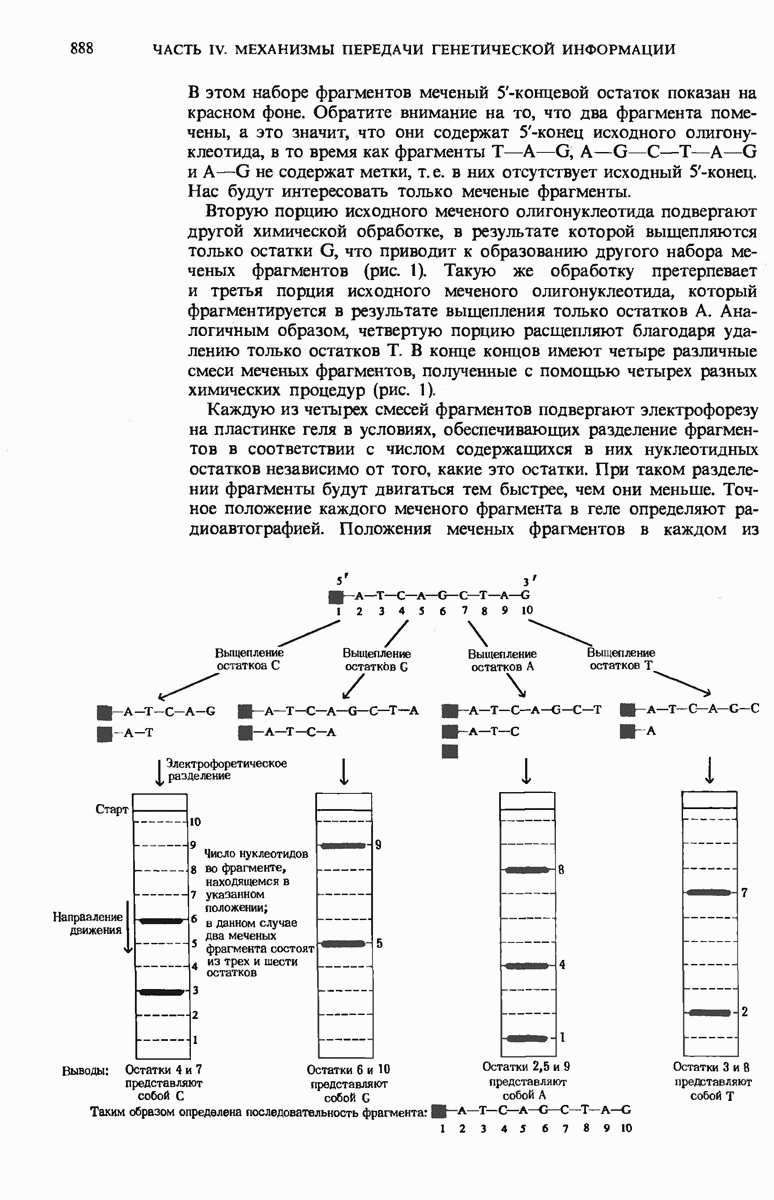
При этом чужеродная ДНК, не имеющая этих «опознавательных» метальных групп, разрушается рестриктирующими эндонуклеазами. В эукариотических ДНК присутствует большое число высокоповторяющихся коротких последовательностей, меньшее число более длинных умеренных повторов, которые, как полагают, играют регуляторную роль, и ряд уникальных (неповторяющихся) участков, представляющих собой, видимо, структурные гены. Эукариотические гены содержат вставочные нетранслируемые нуклеотидные последовательности, называемые интронами, которые вставлены между транслируемыми участками, называемыми экзонами. С помощью недавно разработанных методов были определены нуклеотидные последовательности ряда генов и вирусных ДНК.
ЛИТЕРАТУРА
Общие вопросы
Ayala F., Kiger J. Modern Genetics, Benjamin-Cummings, Menlo Park, Calif., 1980. Прекрасное описание основ генетики.
Komberg A. DNA Replication, Freeman, San Francisco, 1980. Всеобъемлющая и авторитетная монография.
Упаковка
Bauer W. R., Crick F. H. C., White J. H. Super-coiled DNA, Sci. Am., 243, 118-133, July 1980.
Плазмиды
Novick R.P. Plasmids, Sci. Am., 243, 102-127, Decemberg 1980.
Ферменты рестрикции-модификации Nathans D. Restriction Endonucleases, Simian 40, and the New Genetics, Science, 206, 903-909 (1979).
Smith H. Nucleotide Sequence Specificity of Restriction Endonucleases, Science, 205, 455-462 (1979).
Выделение и синтез генов
Khorana Н. G. Total Synthesis of a Gene, Science, 203, 614-625 (1979).
Повторяющиеся последовательности Britten R.J., Kohne D.E. Repeated Segments of DNA, Sci. Am., 222, 24-31, April 1970.
Davidson E. H., Britten R. J. Possible Role of Repetitive Sequences, Science, 204, 1052-1059 (1979).
Вставочные последовательности
Catterall J. F, colleagues. The Chick Ovomucoid Gene Contains at Least Six Intervening Sequences, Science, 204, 264-271 (1979).
Crick F. Split Genes and RNA Splicing, Science, 204, 264-271 (1979).
Хроматин и нуклеосомы
Komberg R. D., Klug A. The Nucleosome, Sci. Am., 244, 52-78, February 1981.
Olins D.E., Olins A.L. Nucleosomes: The Quantum Beyond DNA, Am. Sci., 66, 704-711 (1978).
Нуклеотидная последовательность ДНК
Fiddes J. С. The Nucleotide Sequence of a Viral DNA, Sci. Am., 237, 55-67, December 1977. Прекрасное описание того, как была расшифрована нуклеотидная последовательность ДНК фХ174.
Махат А. М., Gilbert W. A New Method for Sequencing DNA, Proc. Natl. Acad. Sci. US, 74, 560-564 (1977).
Reddy V. B., colleagues. The Genome of Simian Virus 40, Science, 200, 494-502 (1978).
Sanger F. Determination of Nucleotide Sequences in DNA, Biosci. Repts., 1, 3-18 (1981).
Исторические вопросы
Gaims J., Stent G., Watson J. D. (eds.). Phage and the Origins of Molecular Biology, Cold Spring Harbor Laboratory, New York, 1966. Рассказ о начальных этапах развития молекулярной генетики и о некоторых людях, которые способствовали ее расцвету.
Judson Н. The Eighth Day of Creation, Simon and Schuster, New York, 1979.
Olby R. The Path to the Double Helix, University of Washington Press, Seattle, 1974.
Watson J. D. The Double Helix, Atheneum, 1968. Рассказ об истории открытия двойной спирали ДНК.