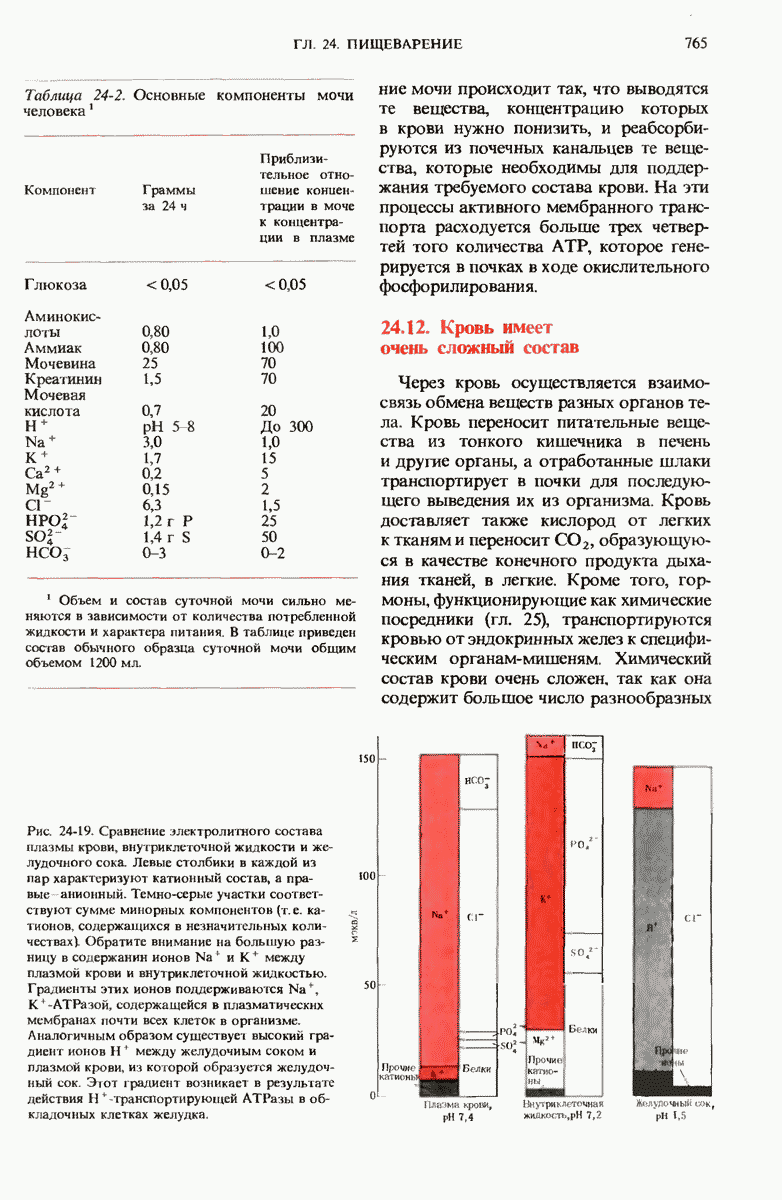
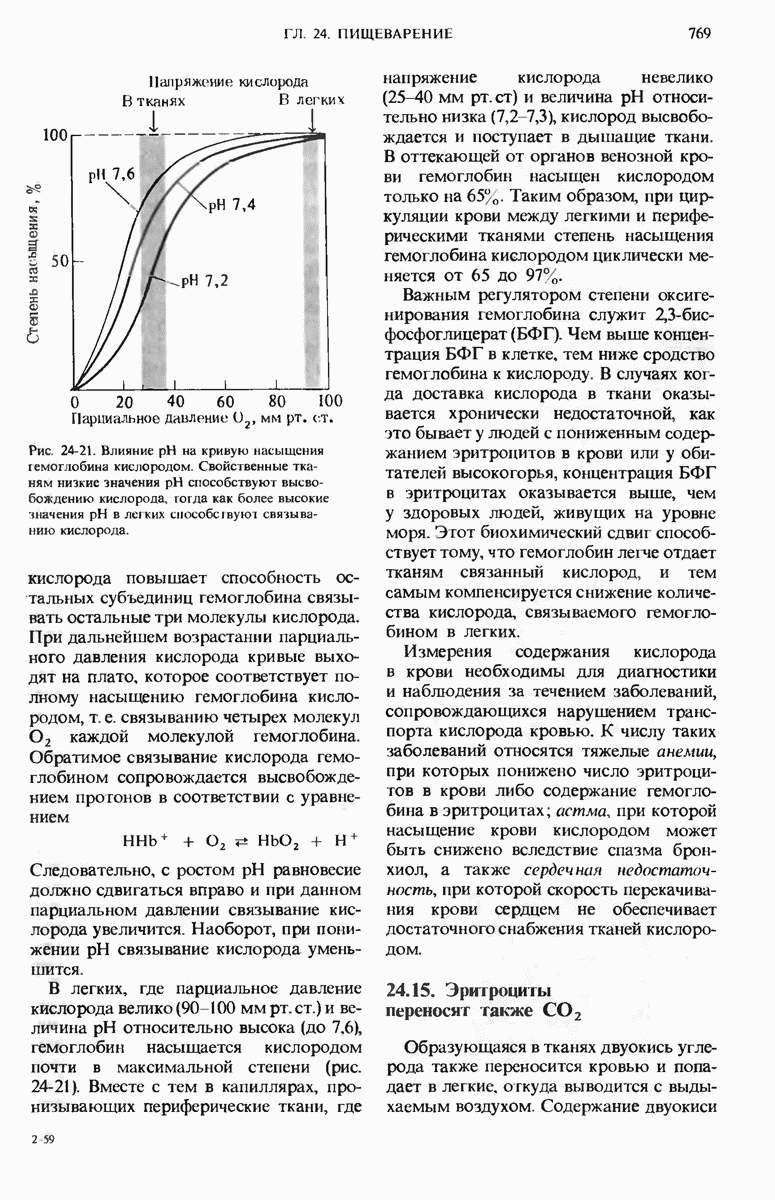
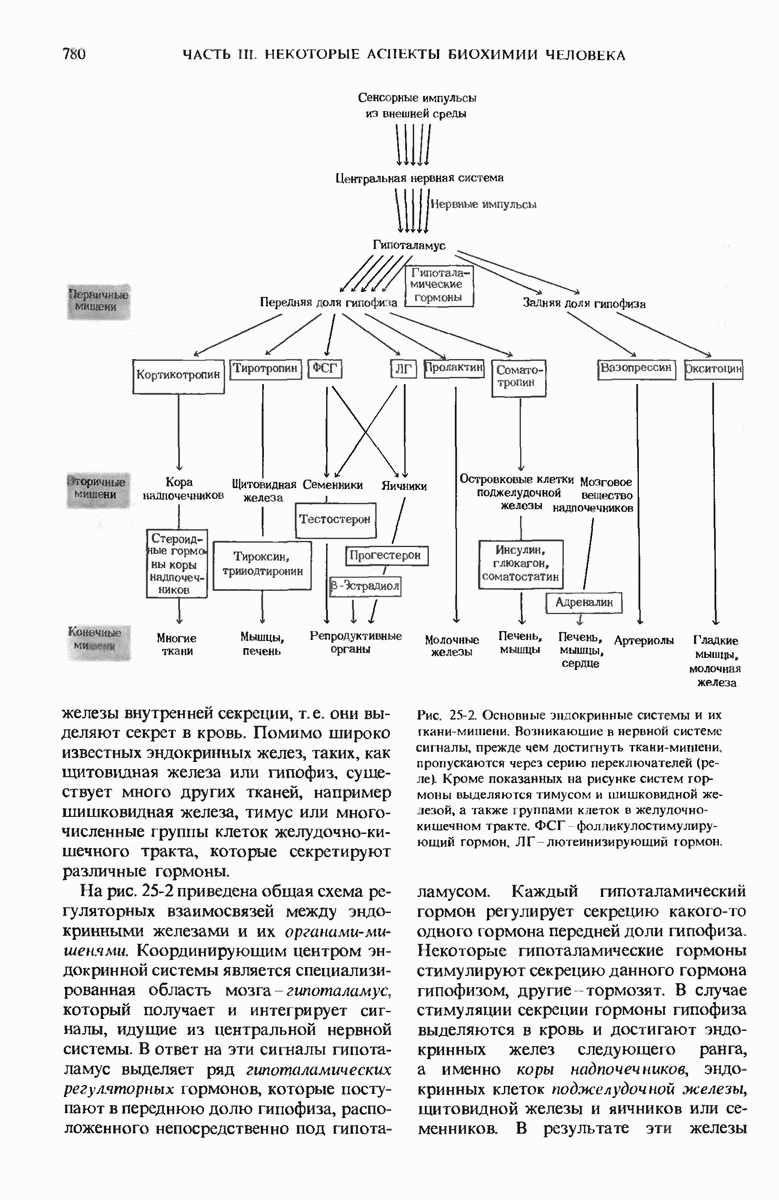
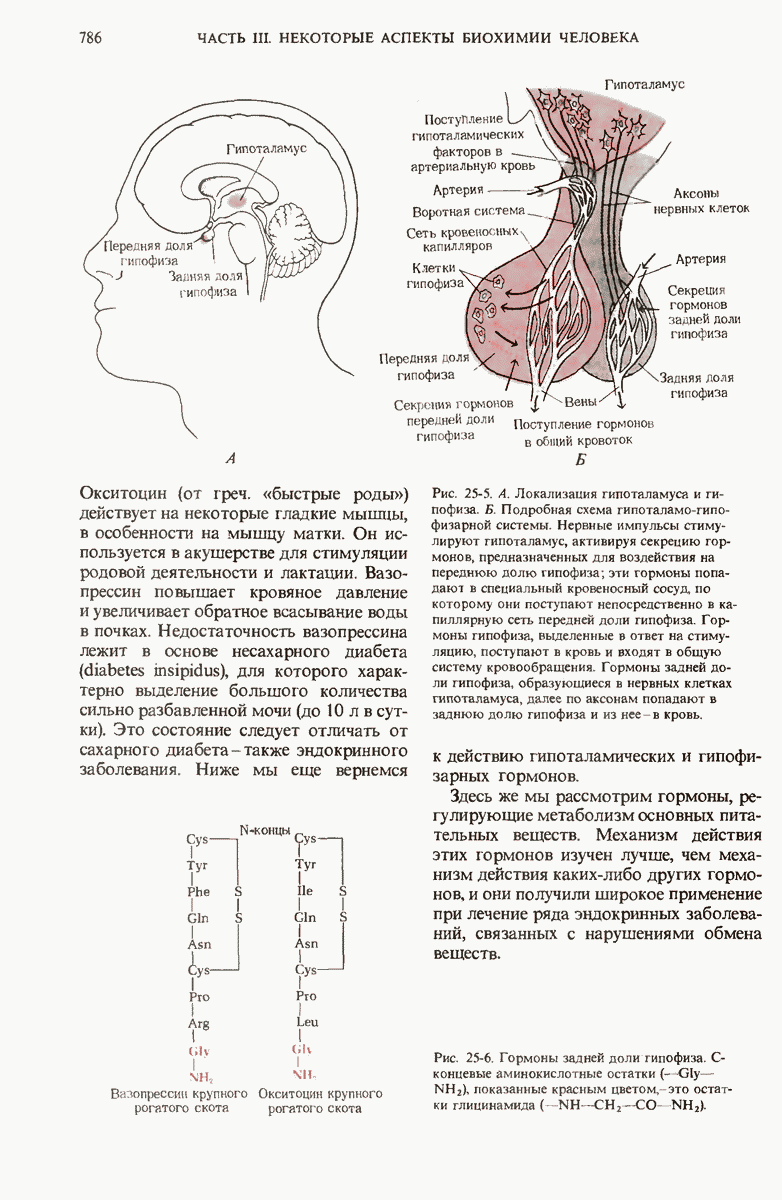
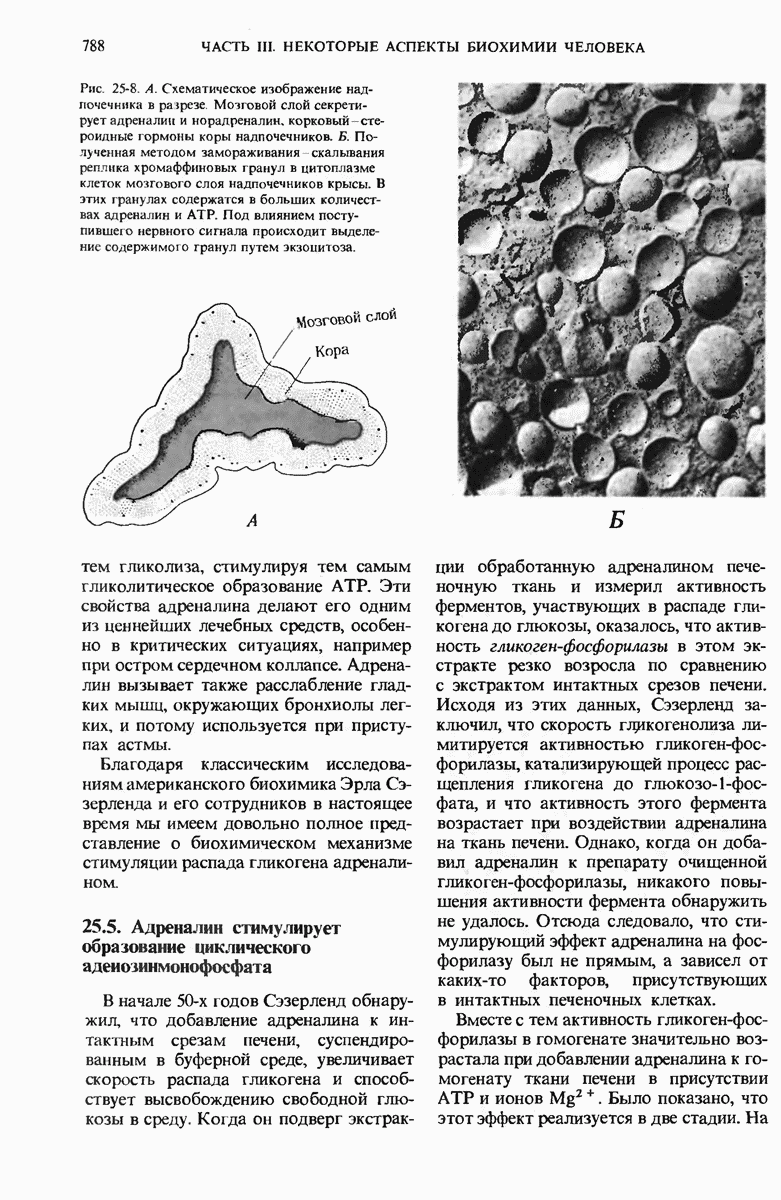
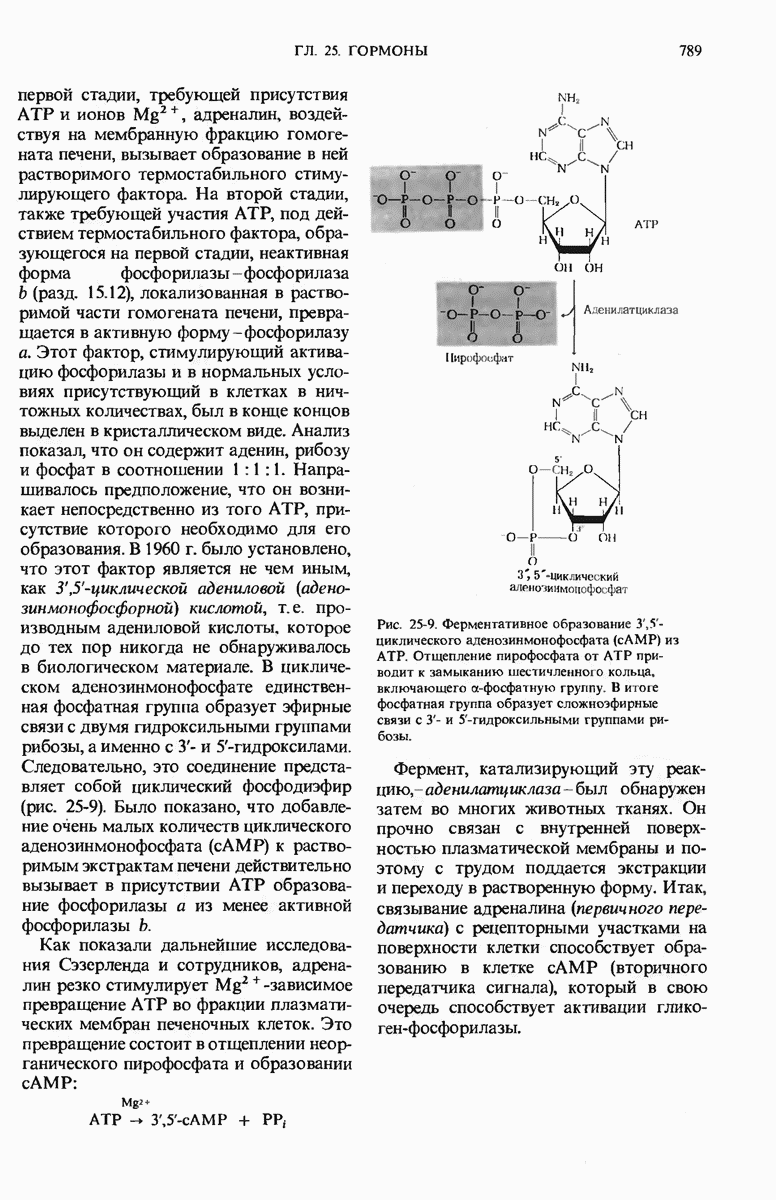
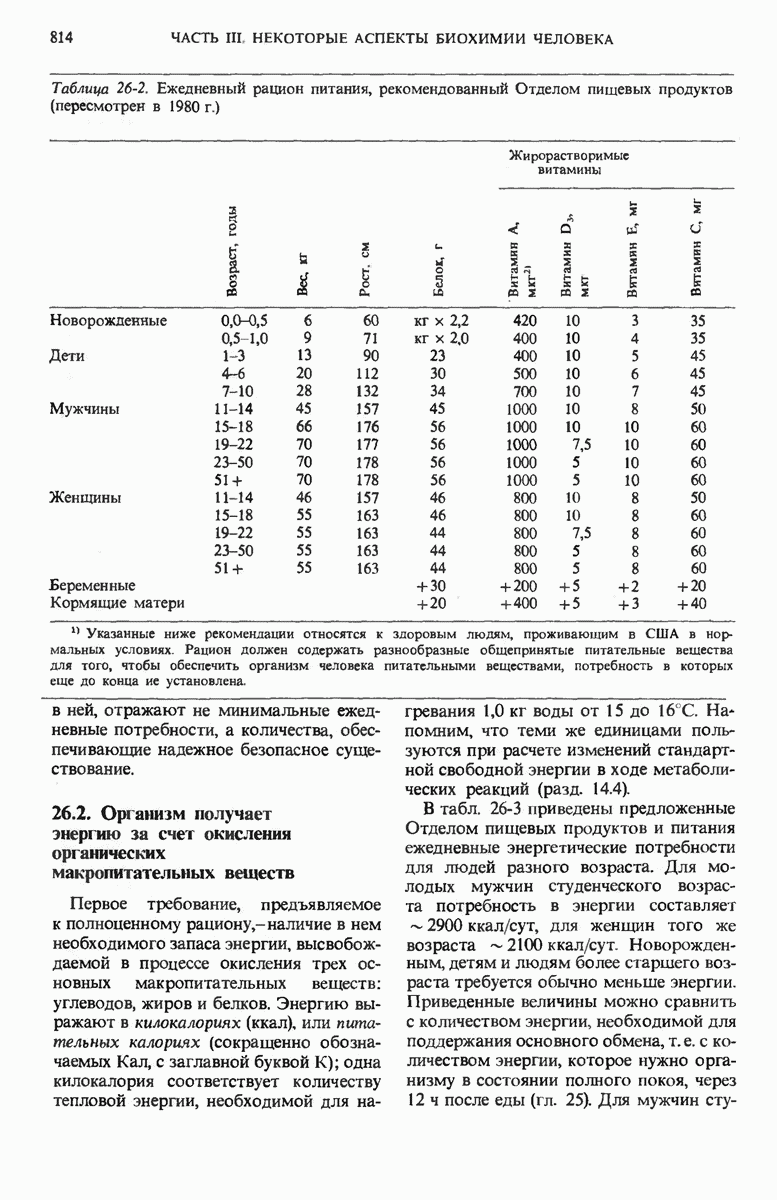
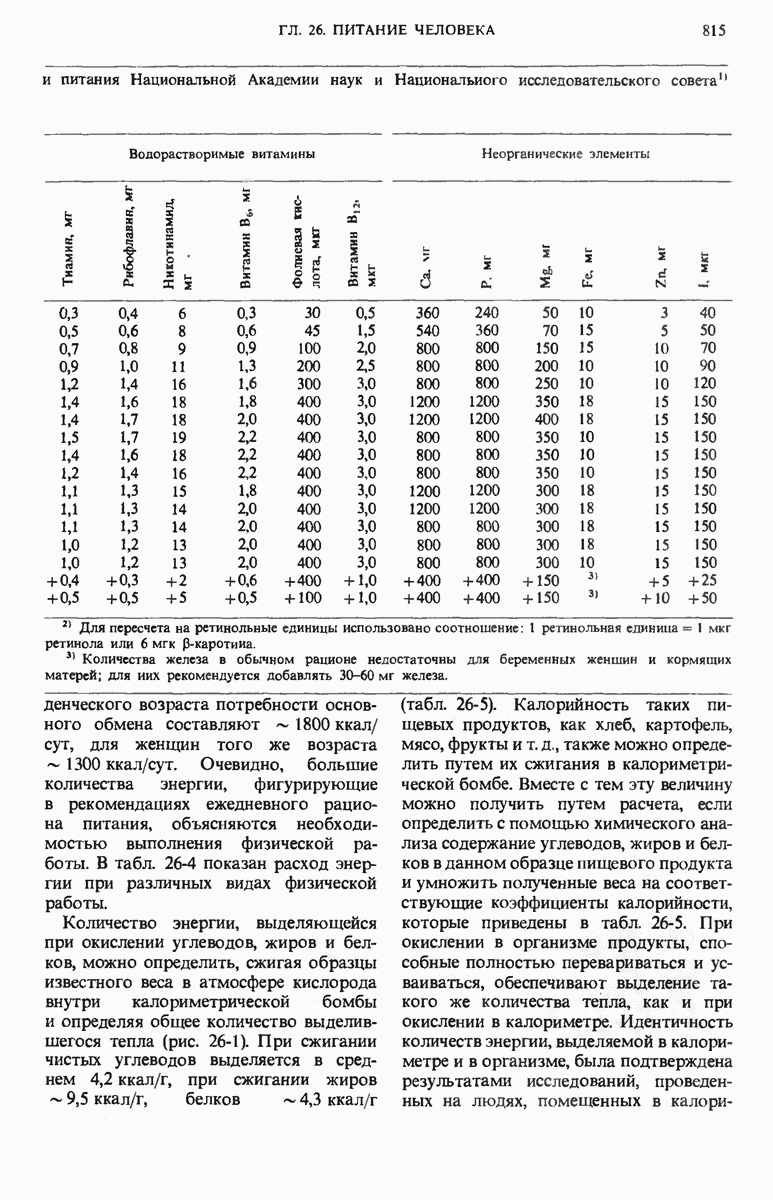
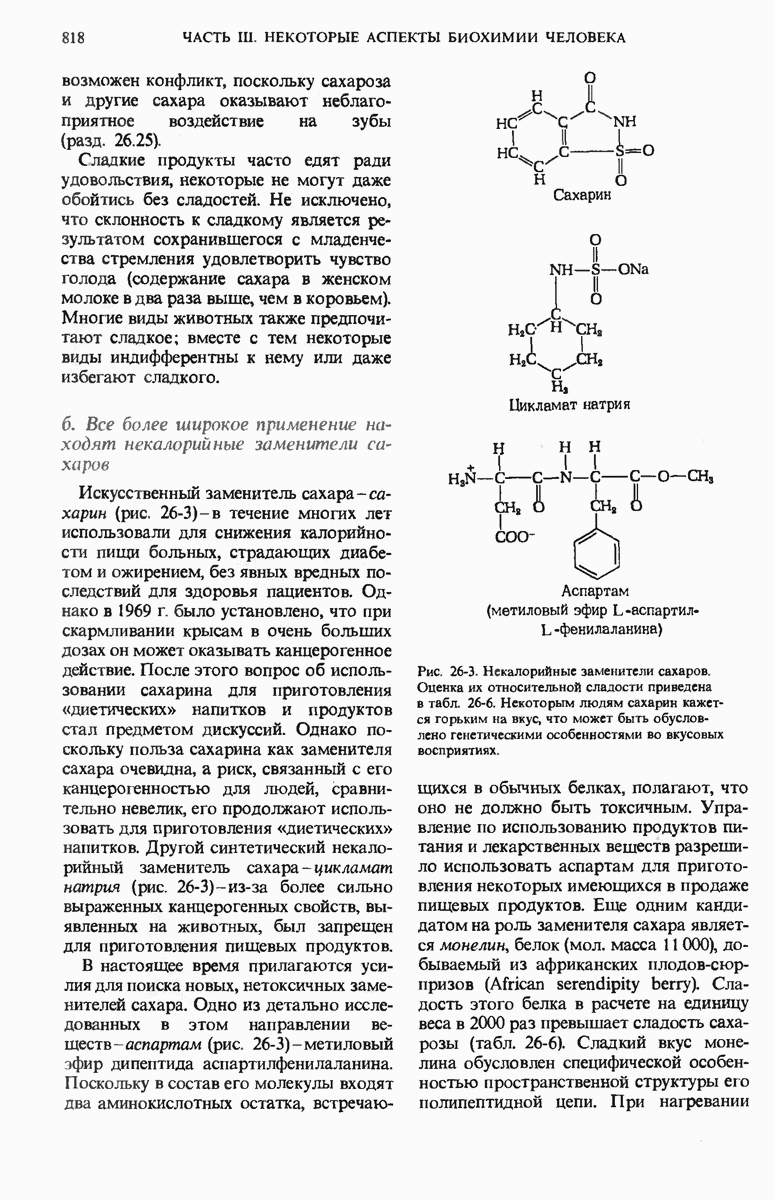
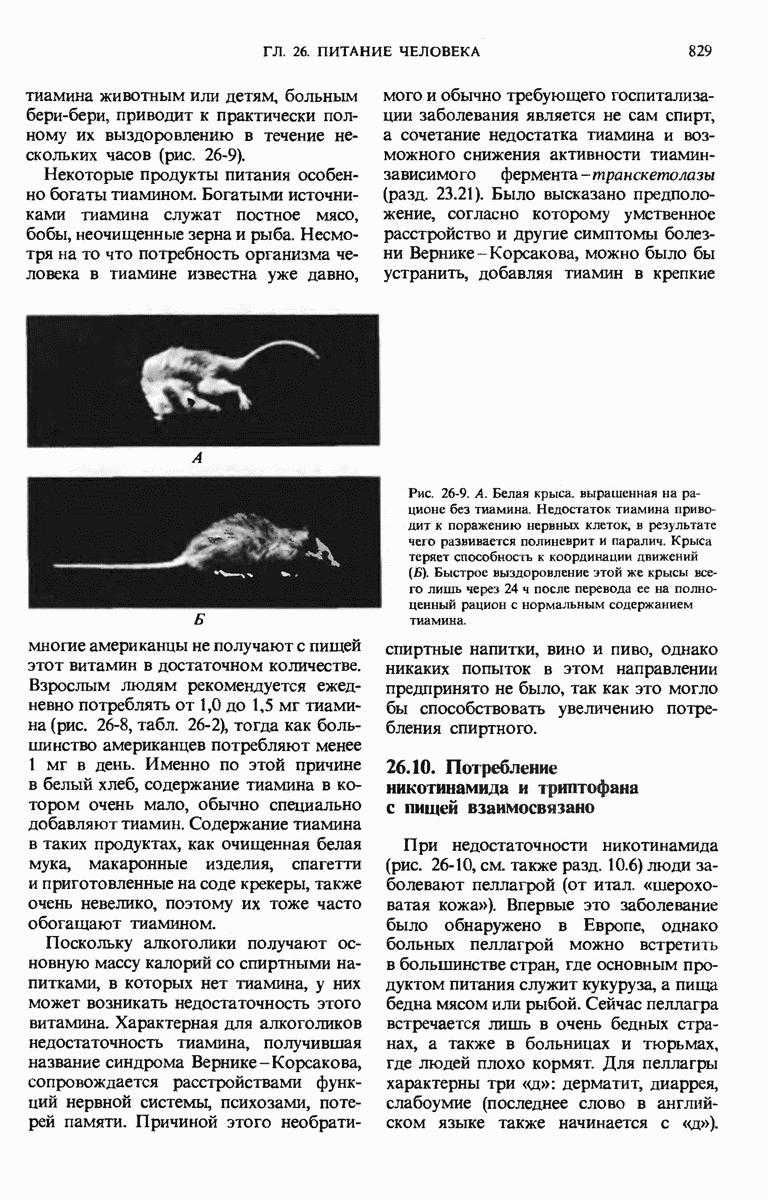
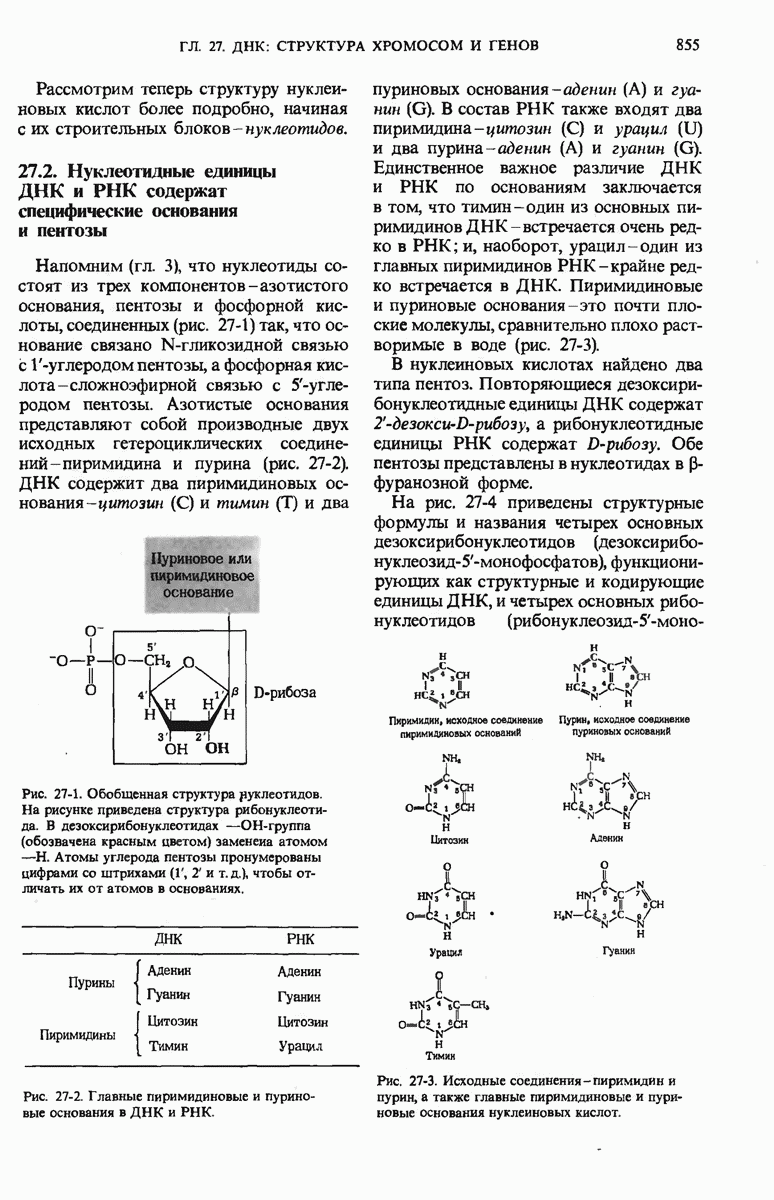
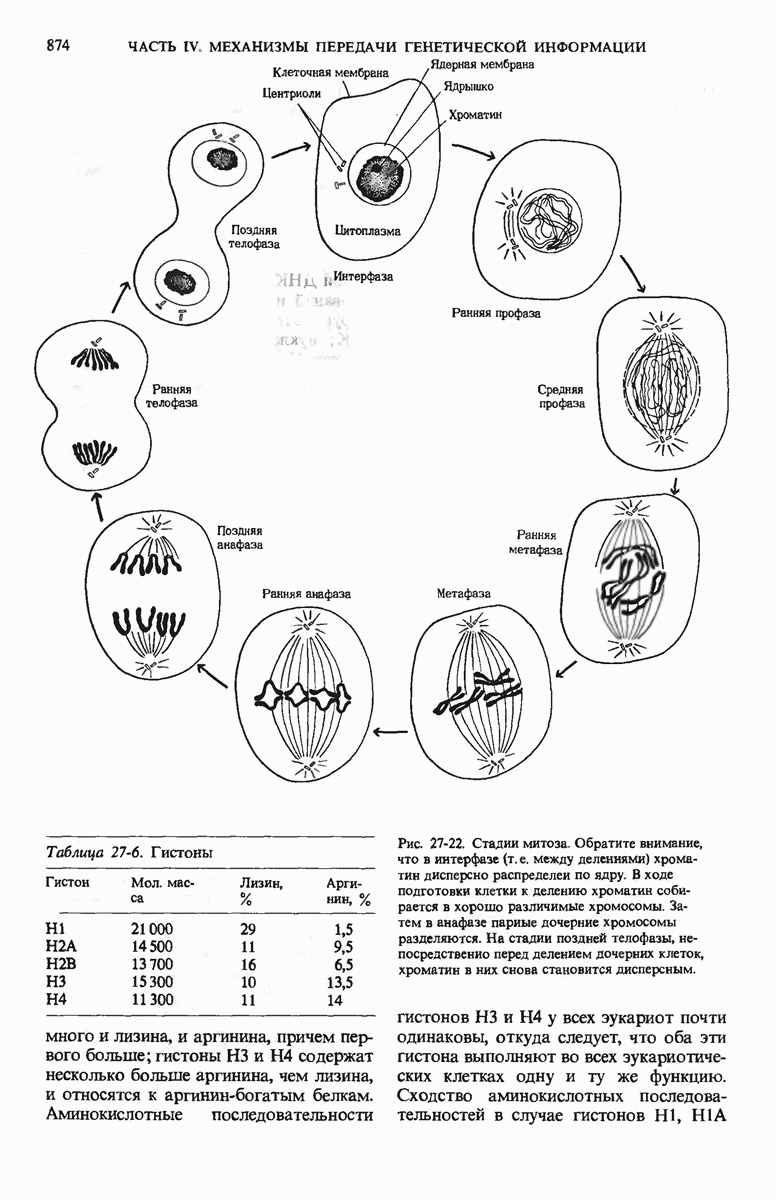
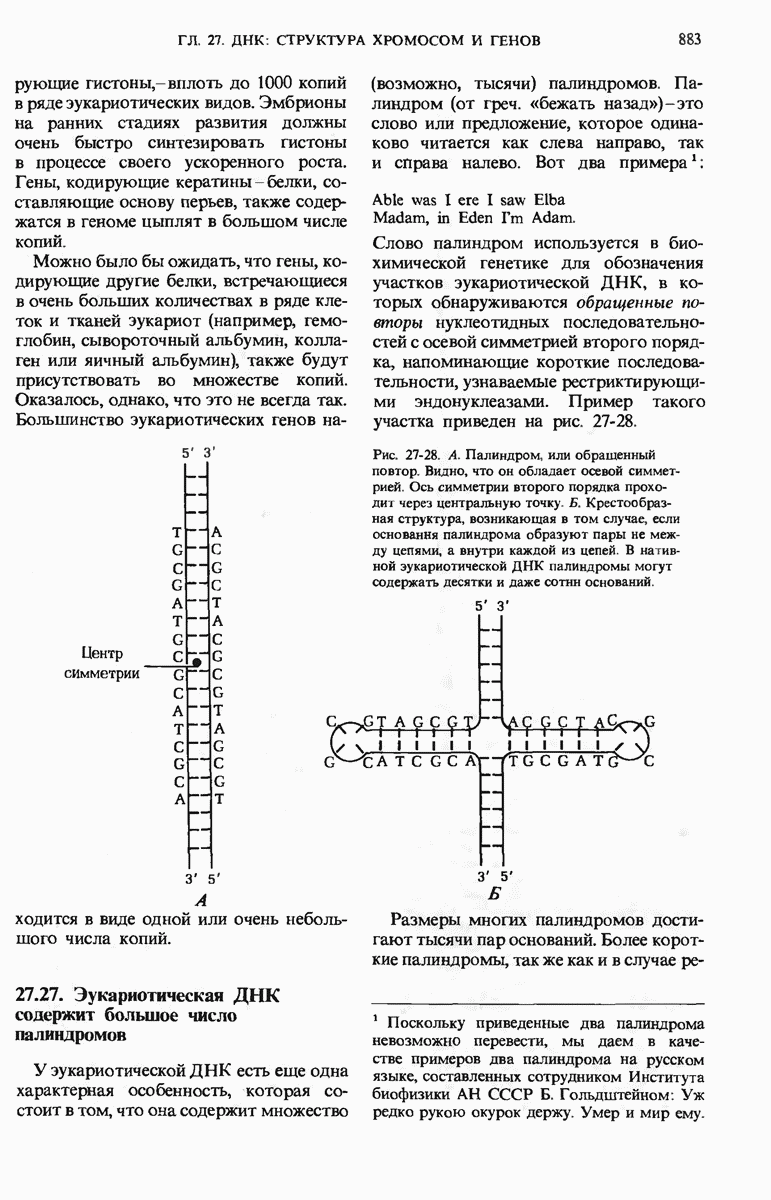
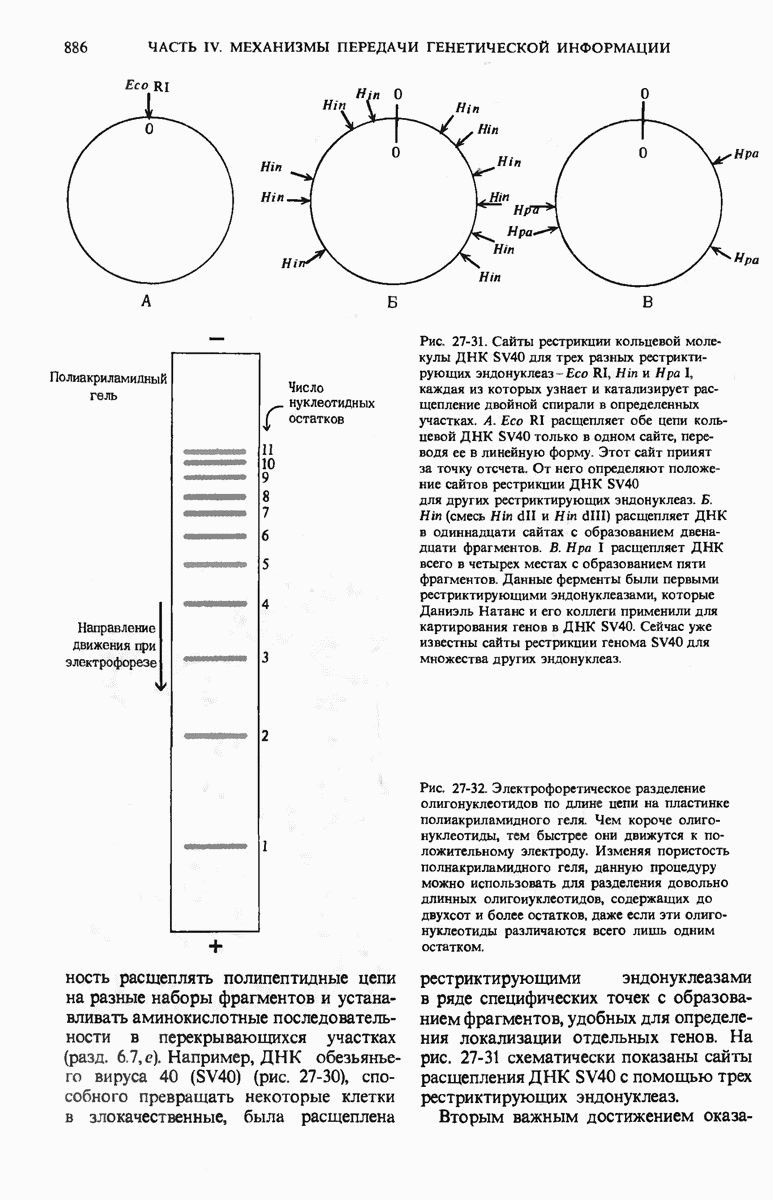
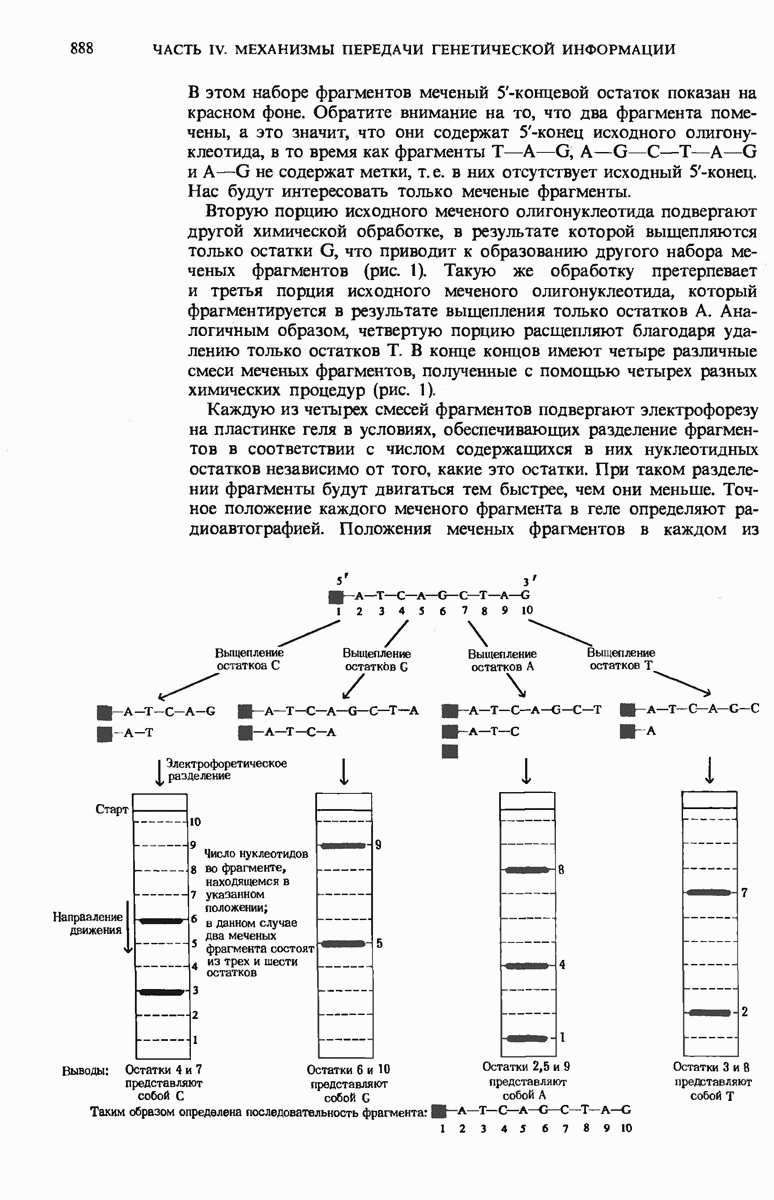
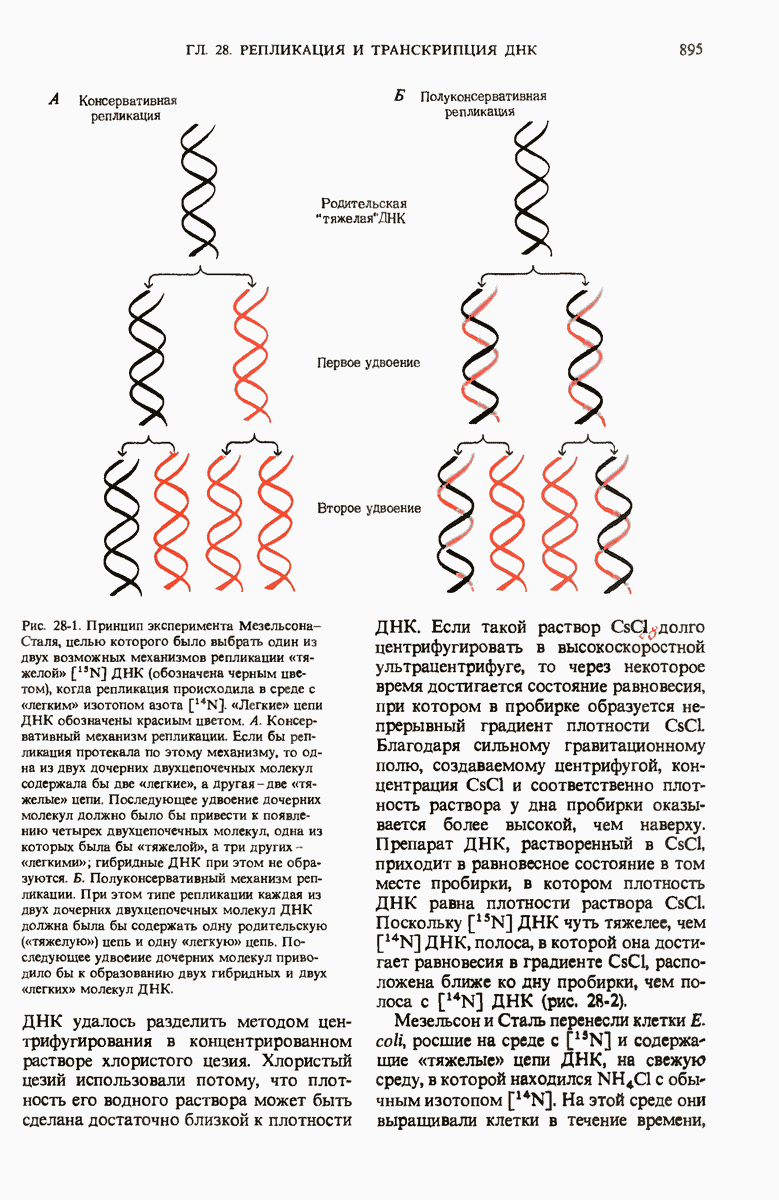
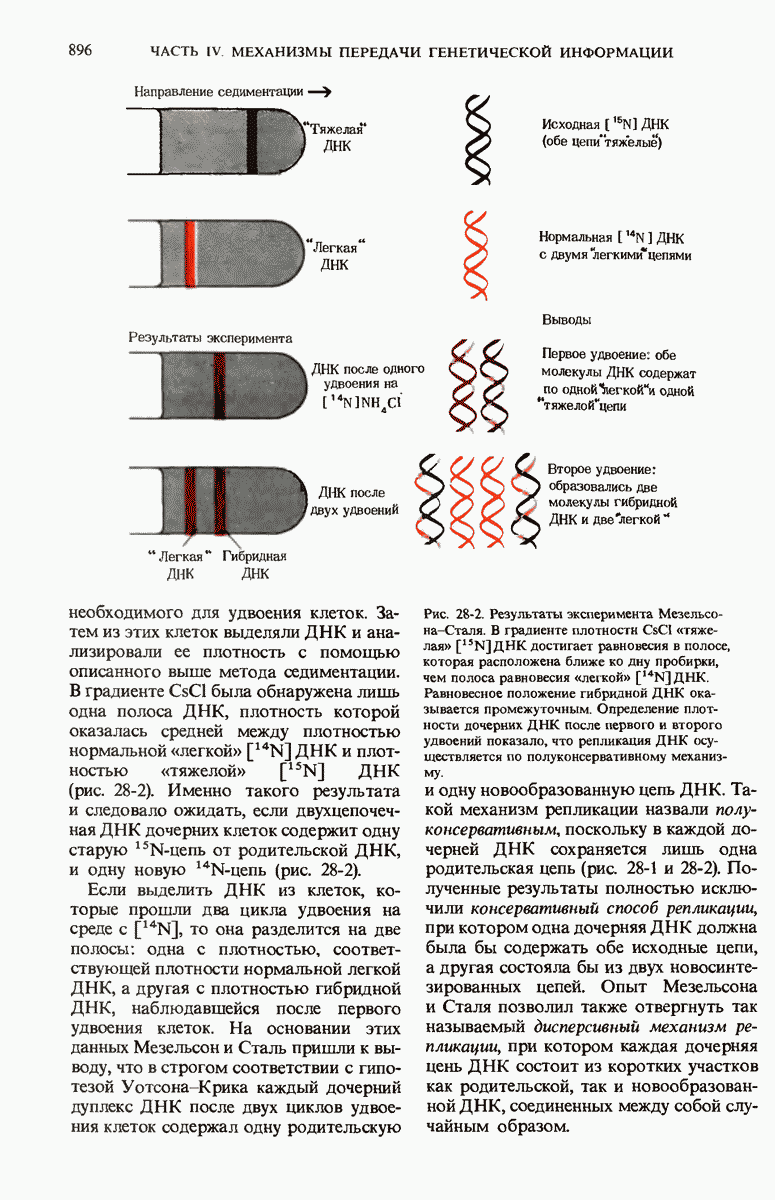
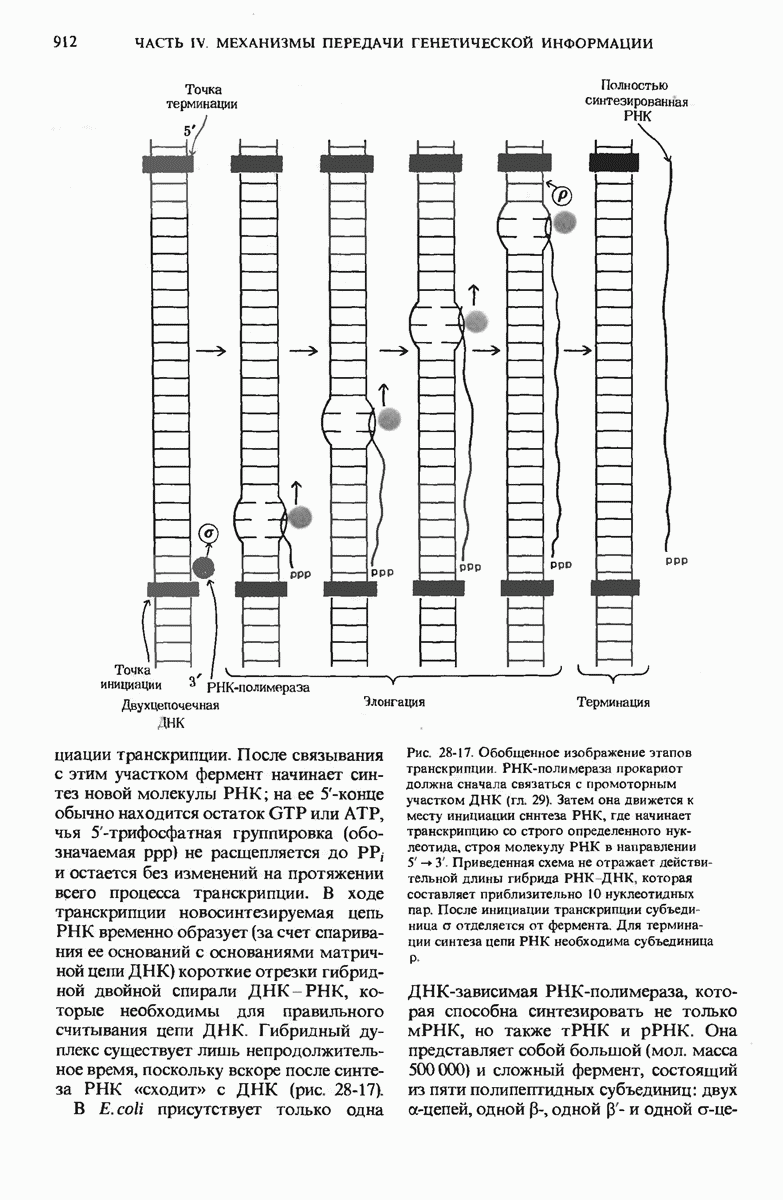
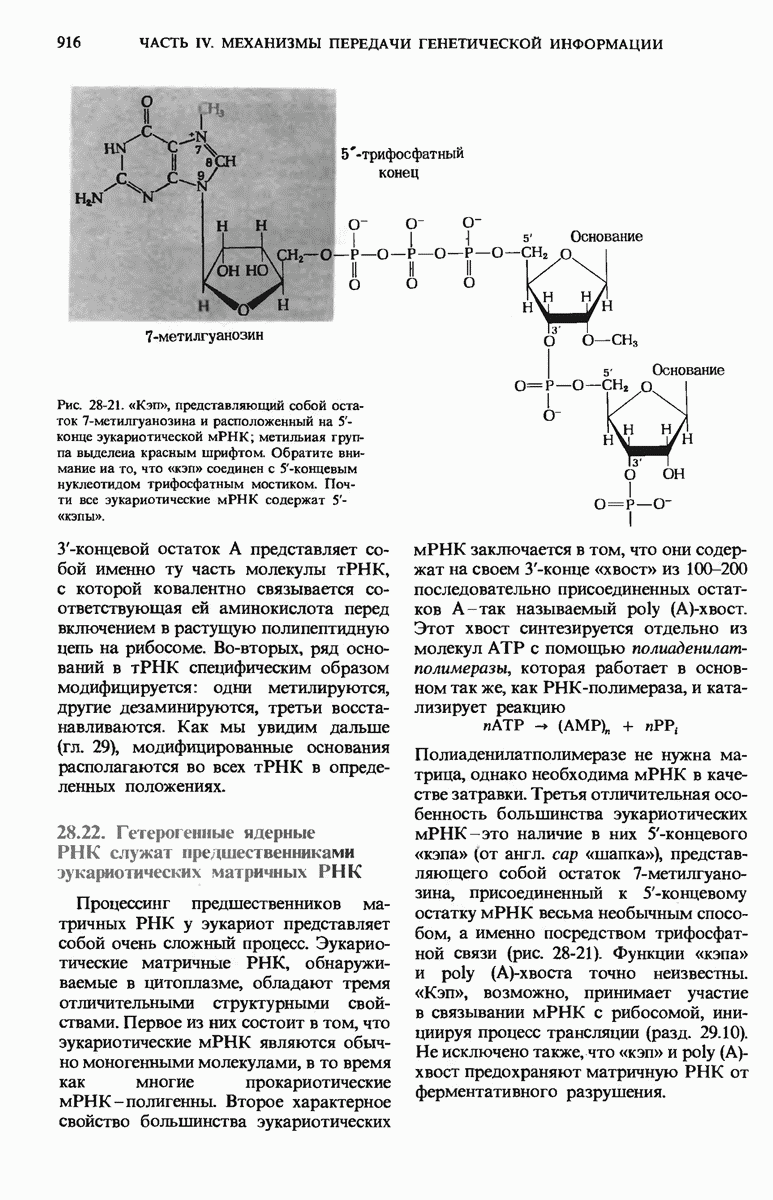
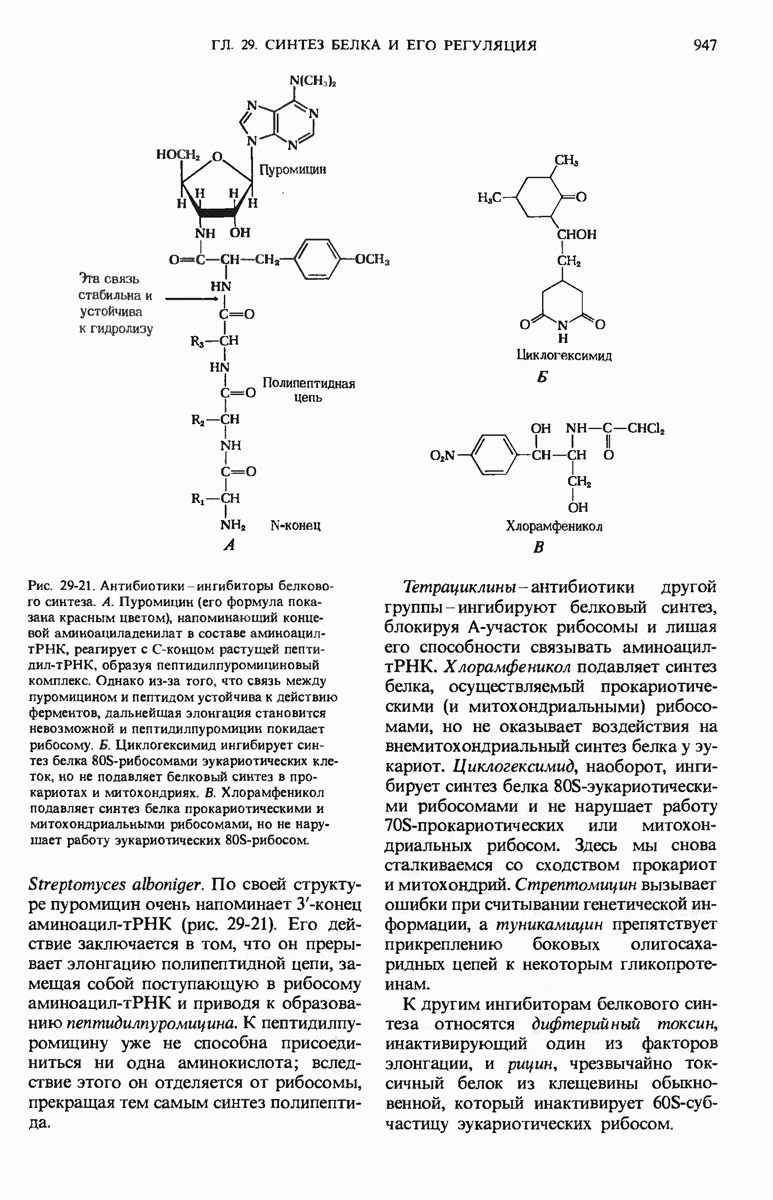
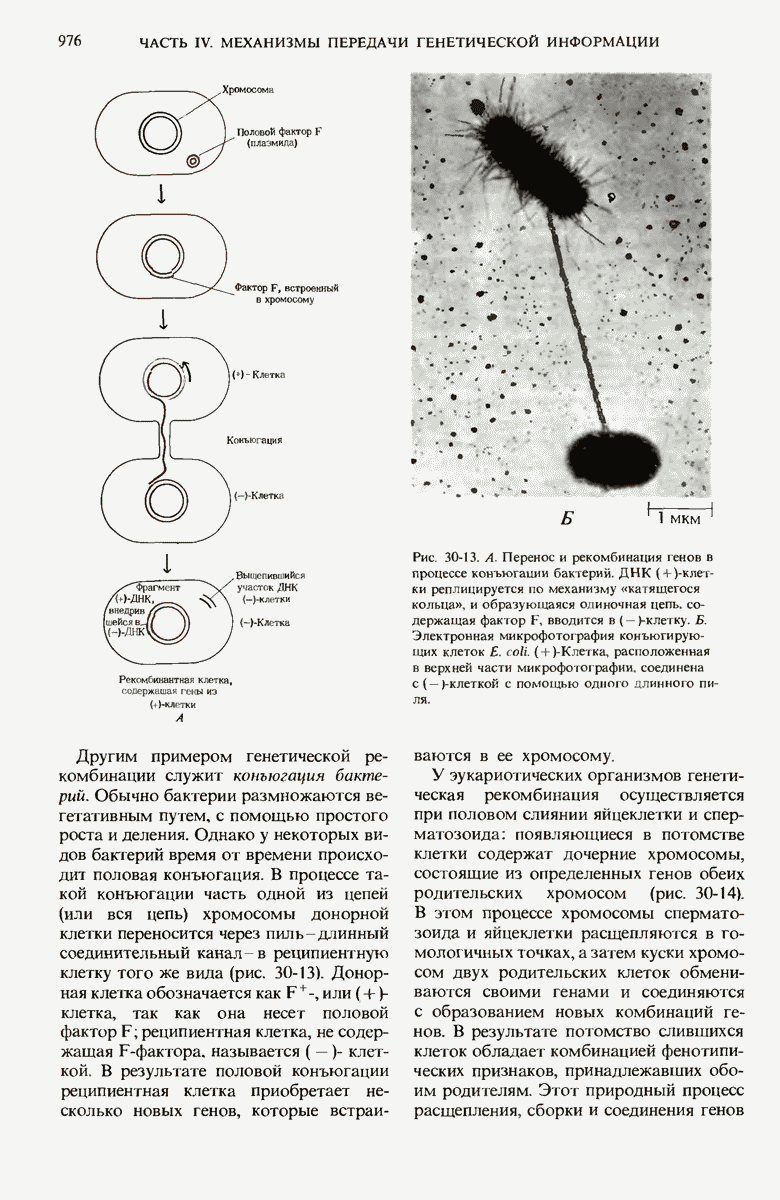
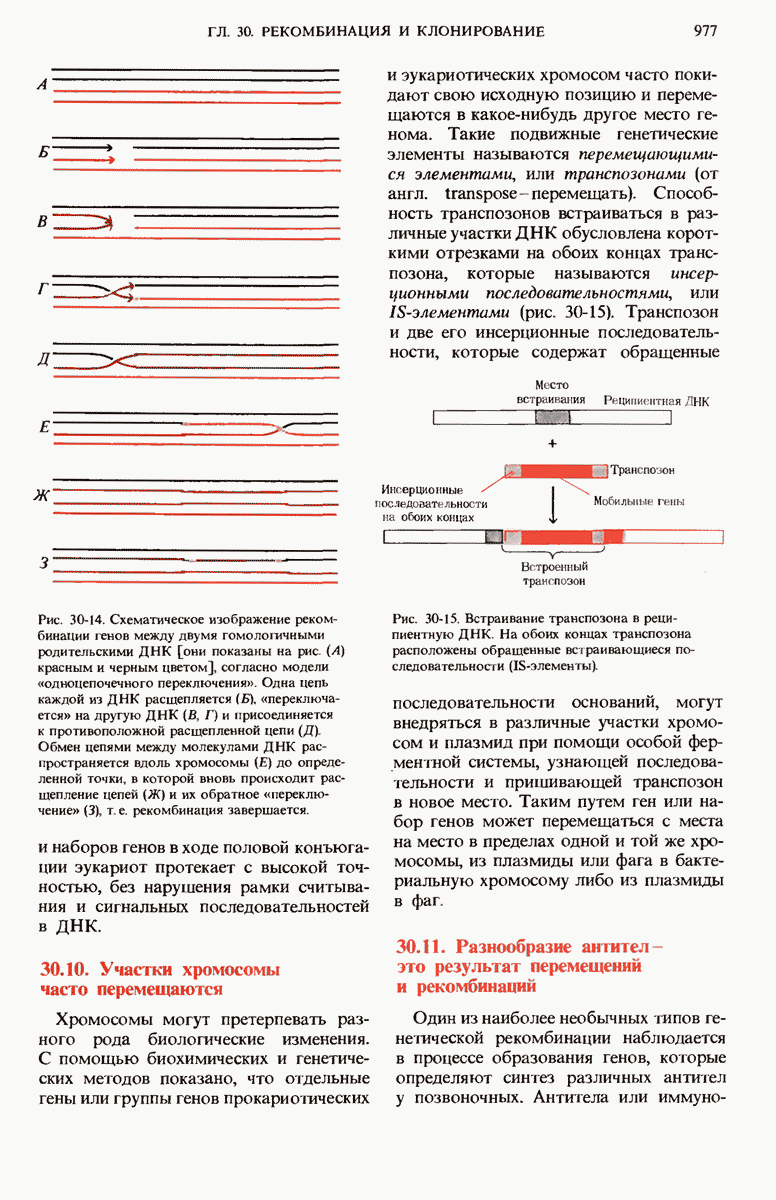
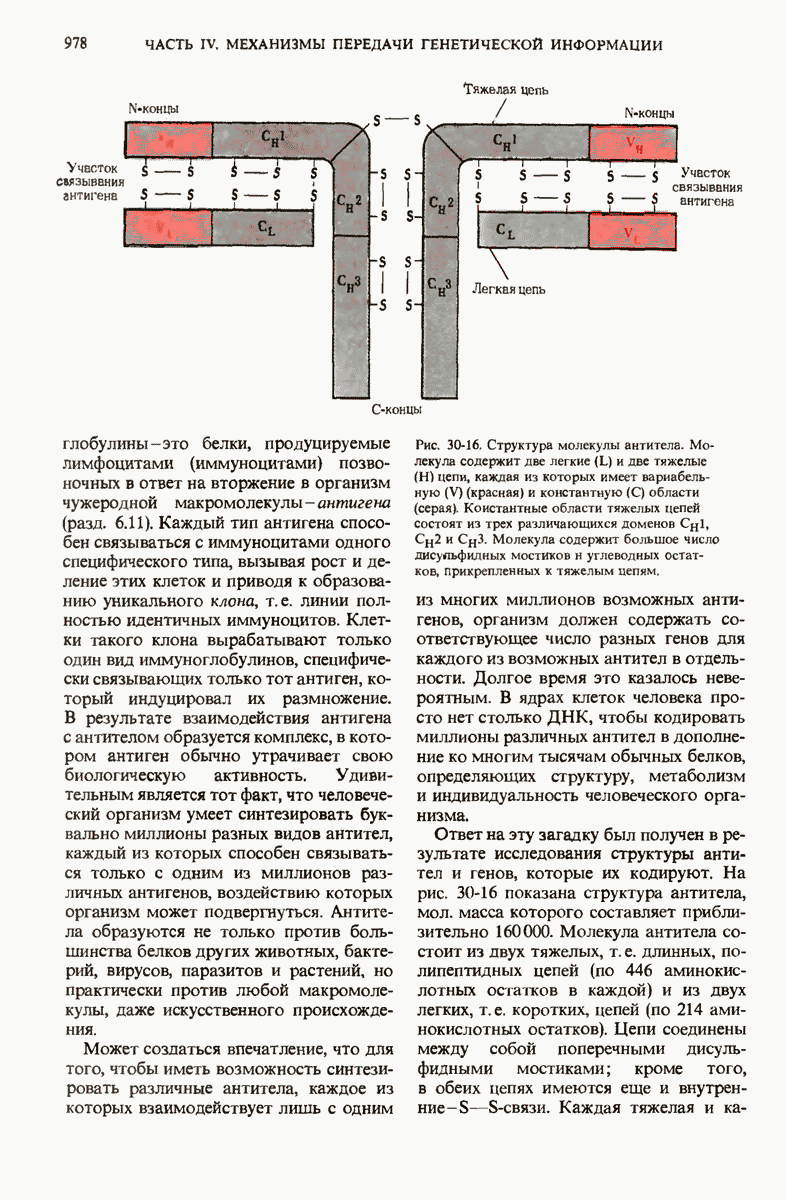
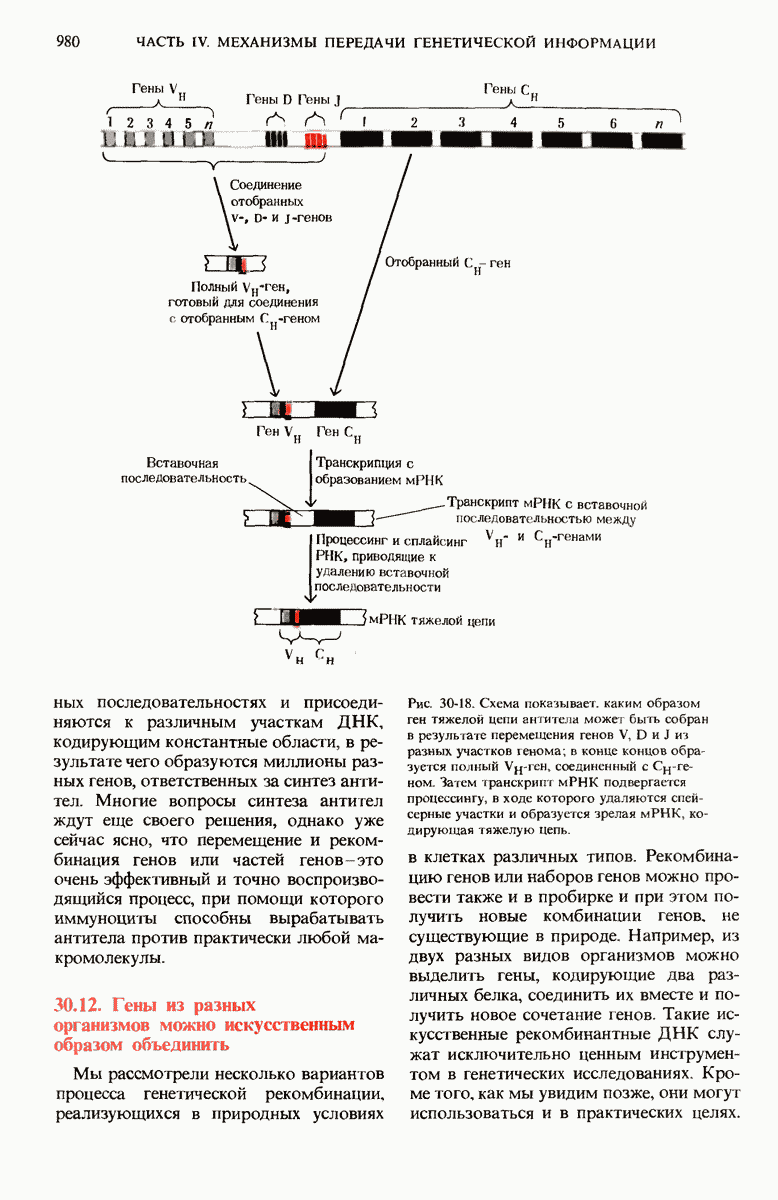
29.15. Полипептидные цепи претерпевают сворачивание и процессинг
Как мы видели в гл. 7 и 8, белок остается биологически неактивным до тех пор, пока он не свернется с образованием присущей ему нативной конформации, которая определяется аминокислотной последовательностью. В какой-то момент во время синтеза полипептидной цепи или после его завершения - белок самопроизвольно принимает свою нативную конформацию (разд. 8.6 и 8.7), т.е. линейная или одномерная генетическая информация, содержавшаяся в матричной РНК, преобразуется в специфическую трехмерную структуру новосинтезированного полипептида.


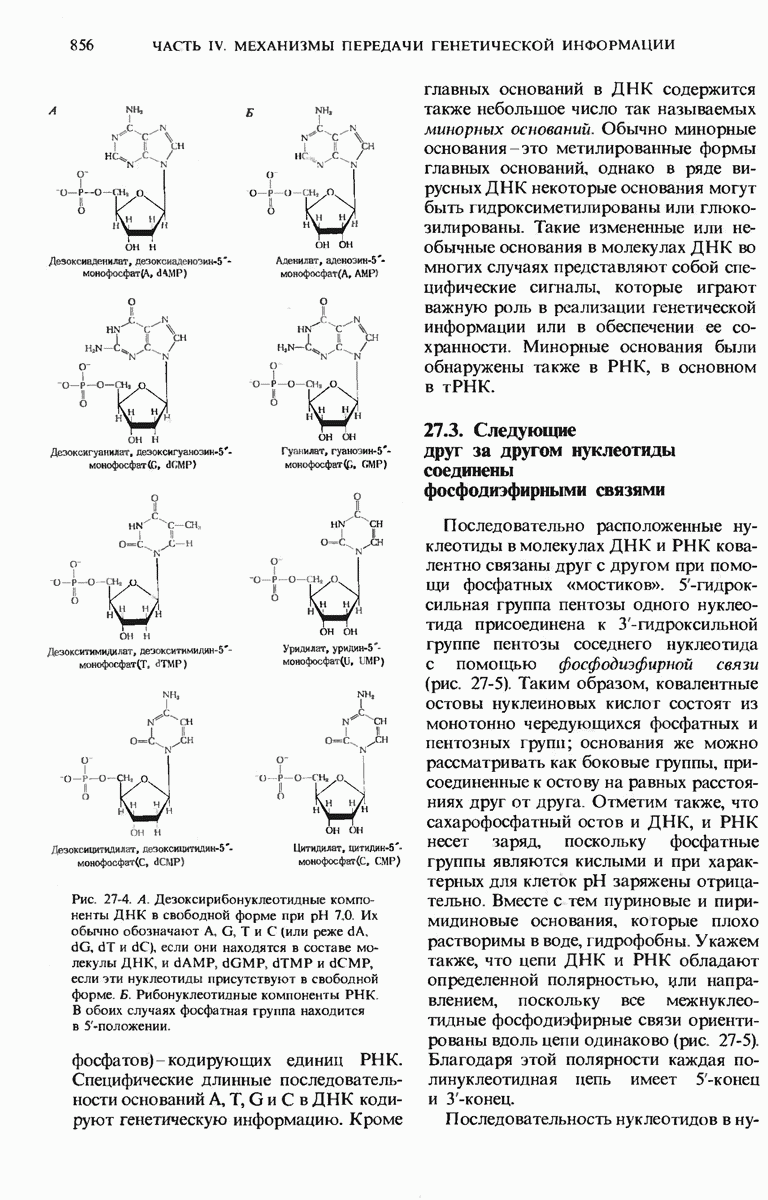
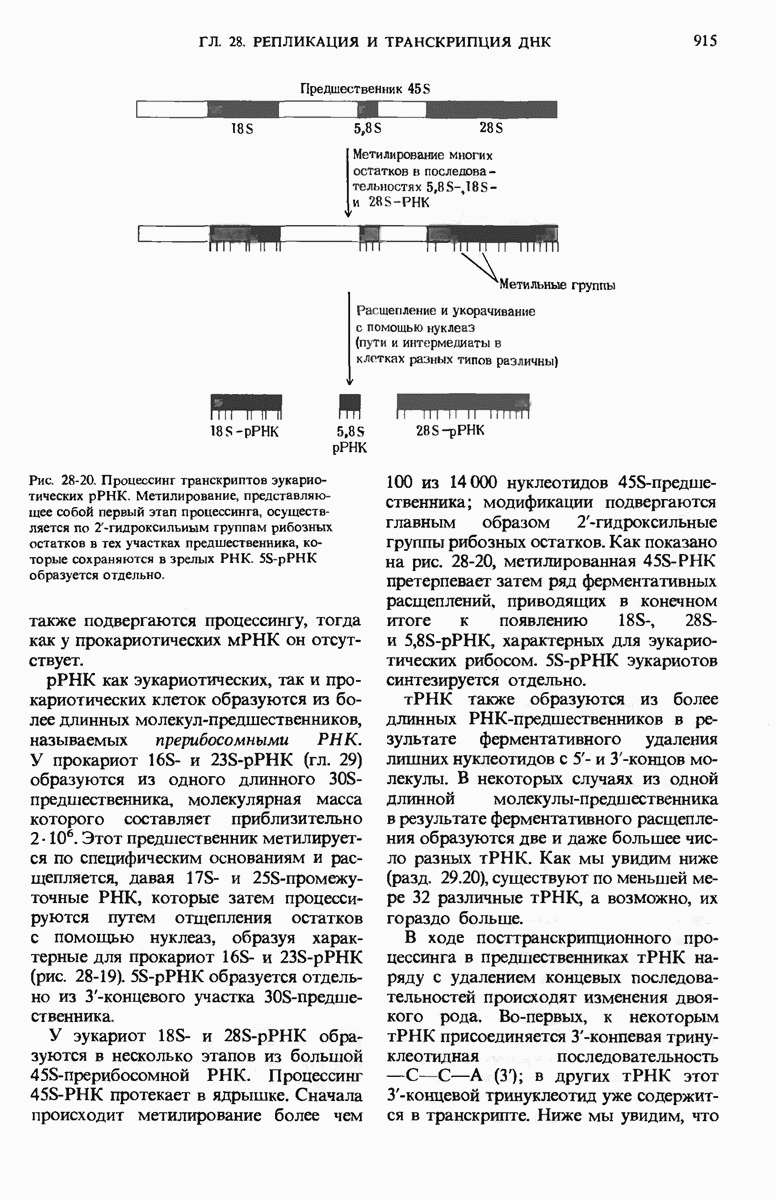
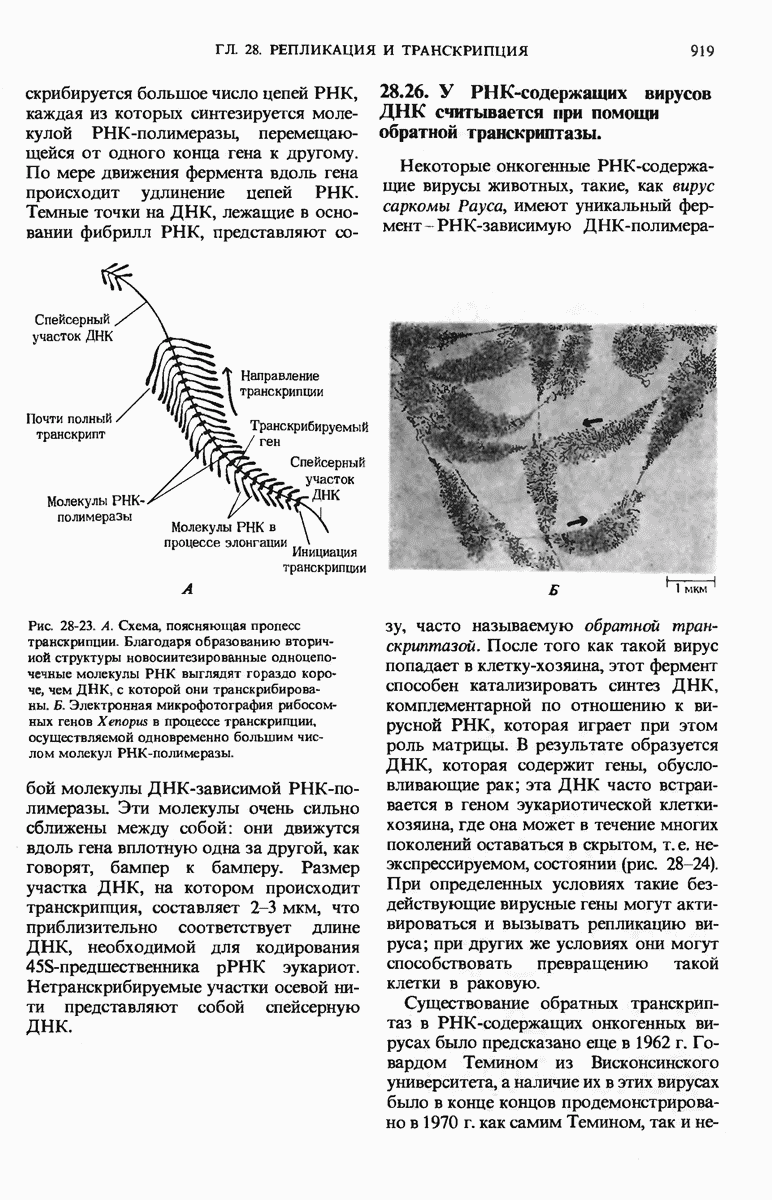
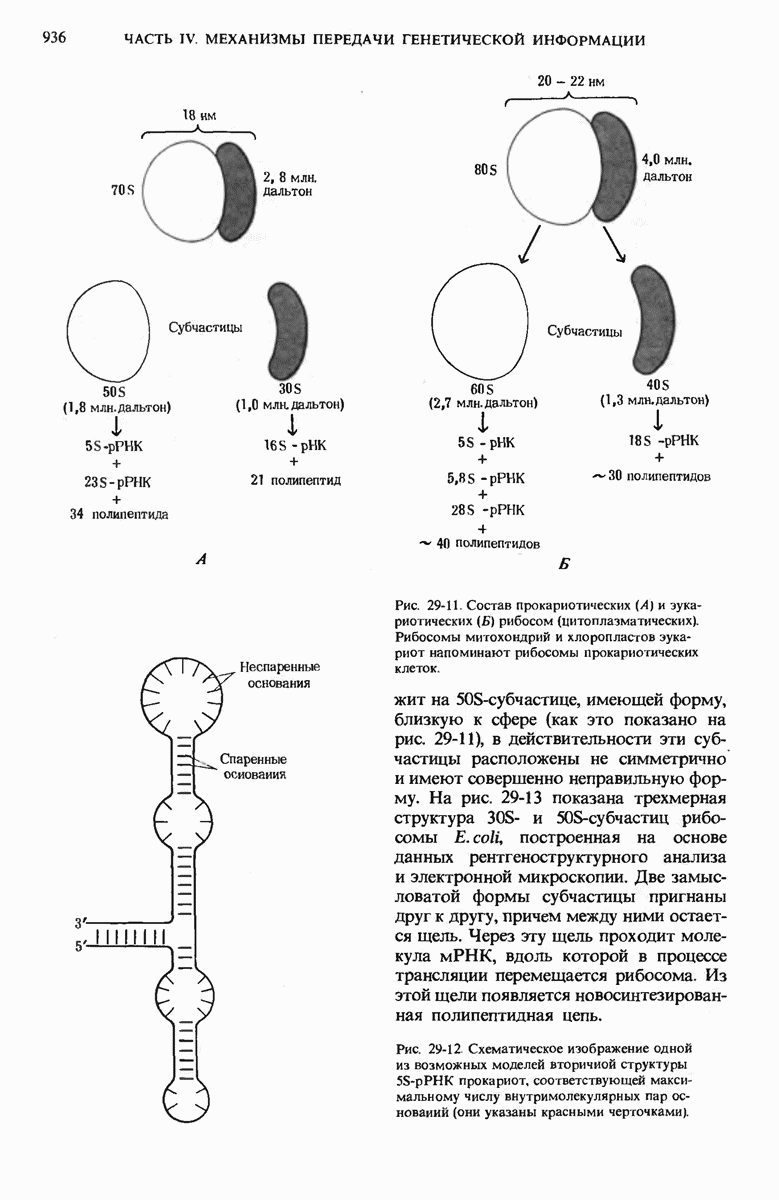
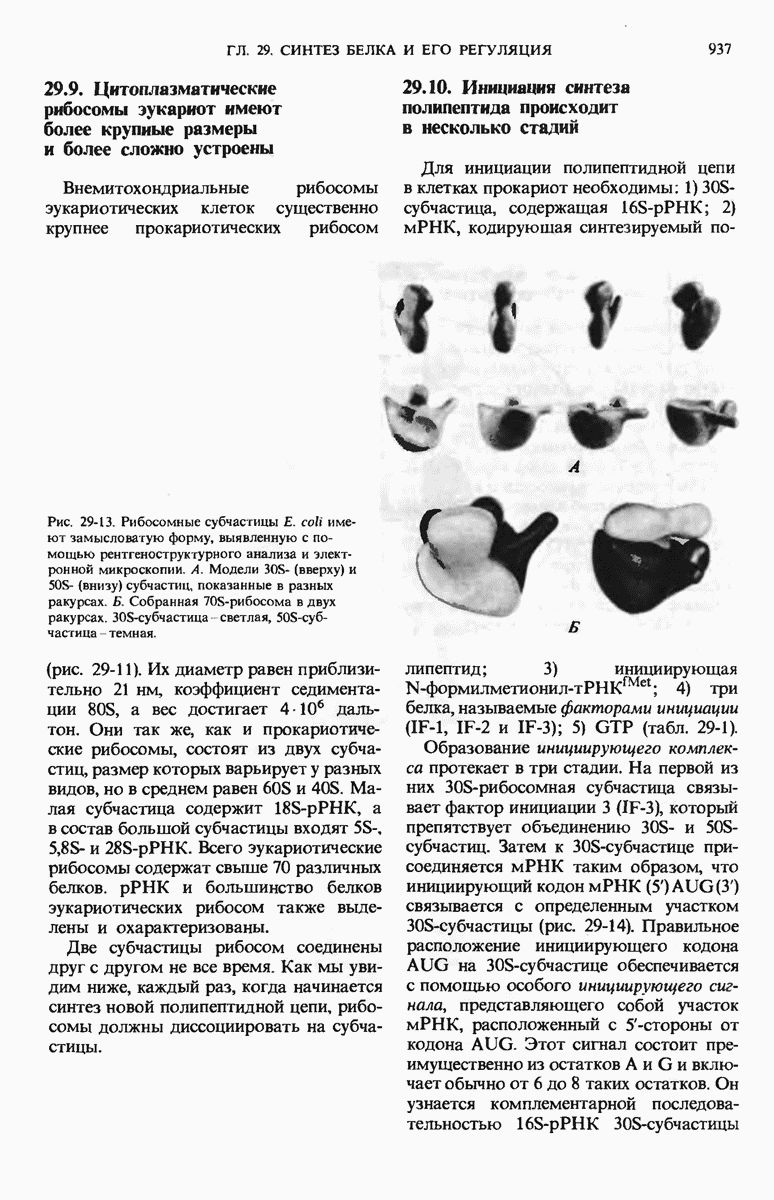
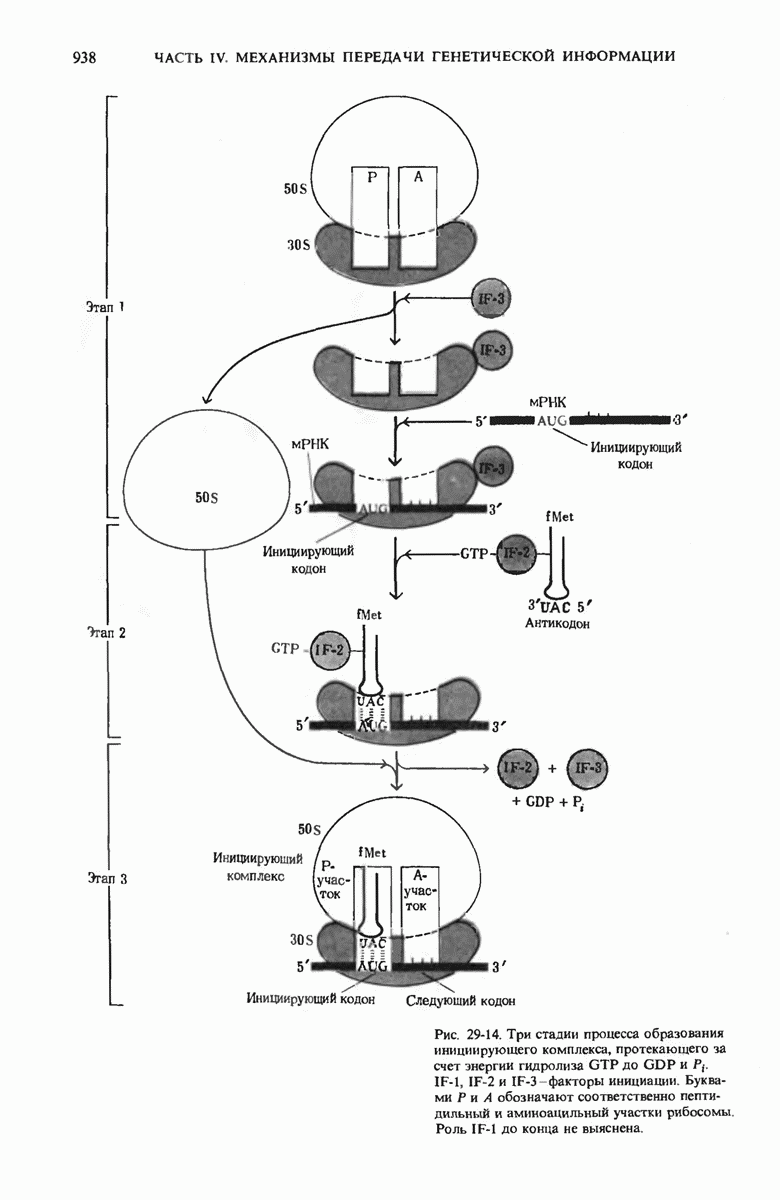
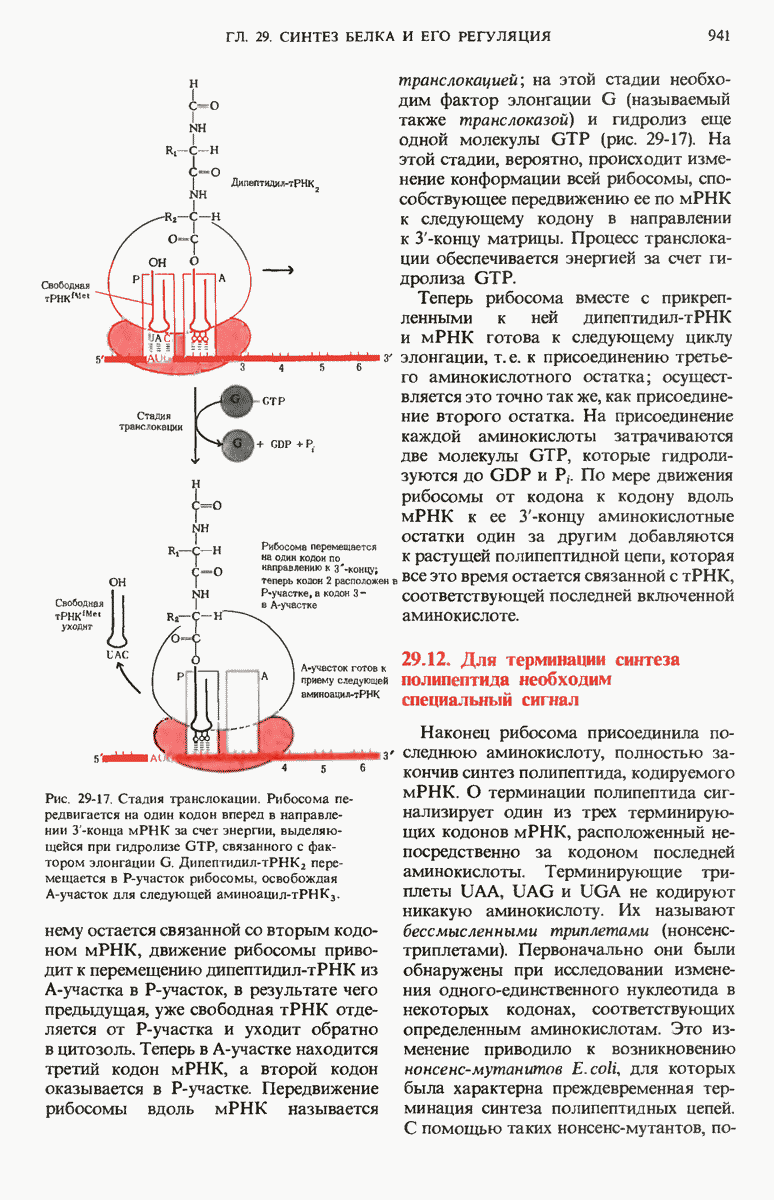
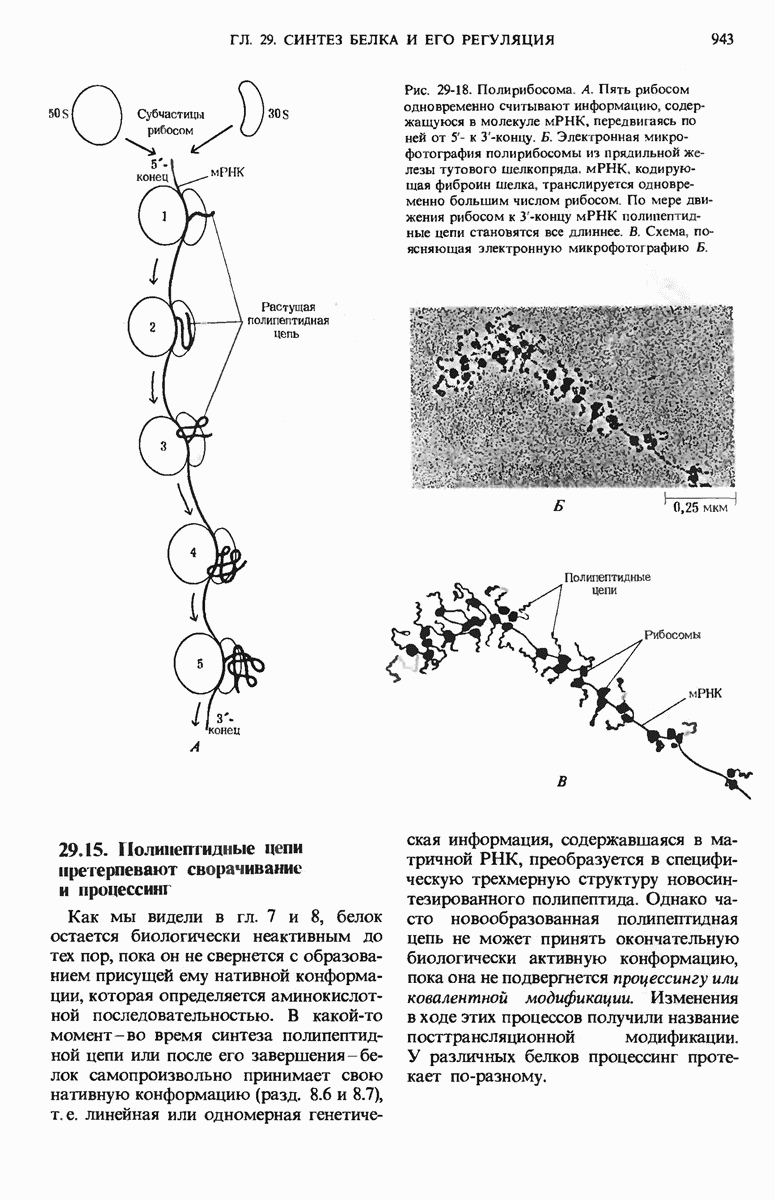
Рис. 29-18. Полирибосома. А. Пять рибосом одновременно считывают информацию, содержащуюся в молекуле мРНК, передвигаясь по ней от 5- к 3-концу. Б. Электронная микрофотография полирибосомы из прядильной железы тутового шелкопряда. мРНК, кодирующая фиброин шелка, транслируется одновременно большим числом рибосом. По мере движения рибосом к 3-концу мРНК полипептидные цепи становятся все длиннее. В. Схема, поясняющая электронную микрофотографию Б.
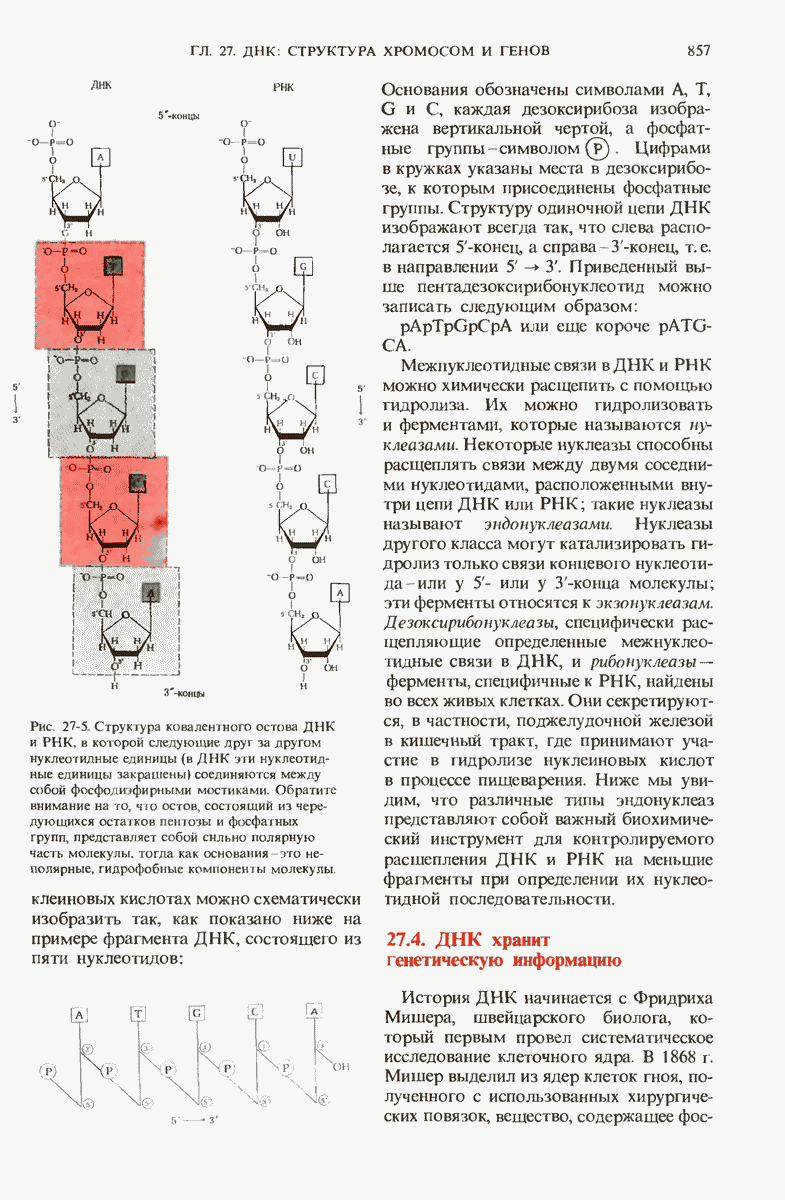
Однако часто новообразованная полипептидная цепь не может принять окончательную биологически активную конформацию, пока она не подвергнется процессингу или ковалентной модификации. Изменения в ходе этих процессов получили название посттрансляционной модификации. У различных белков процессинг протекает по-разному.

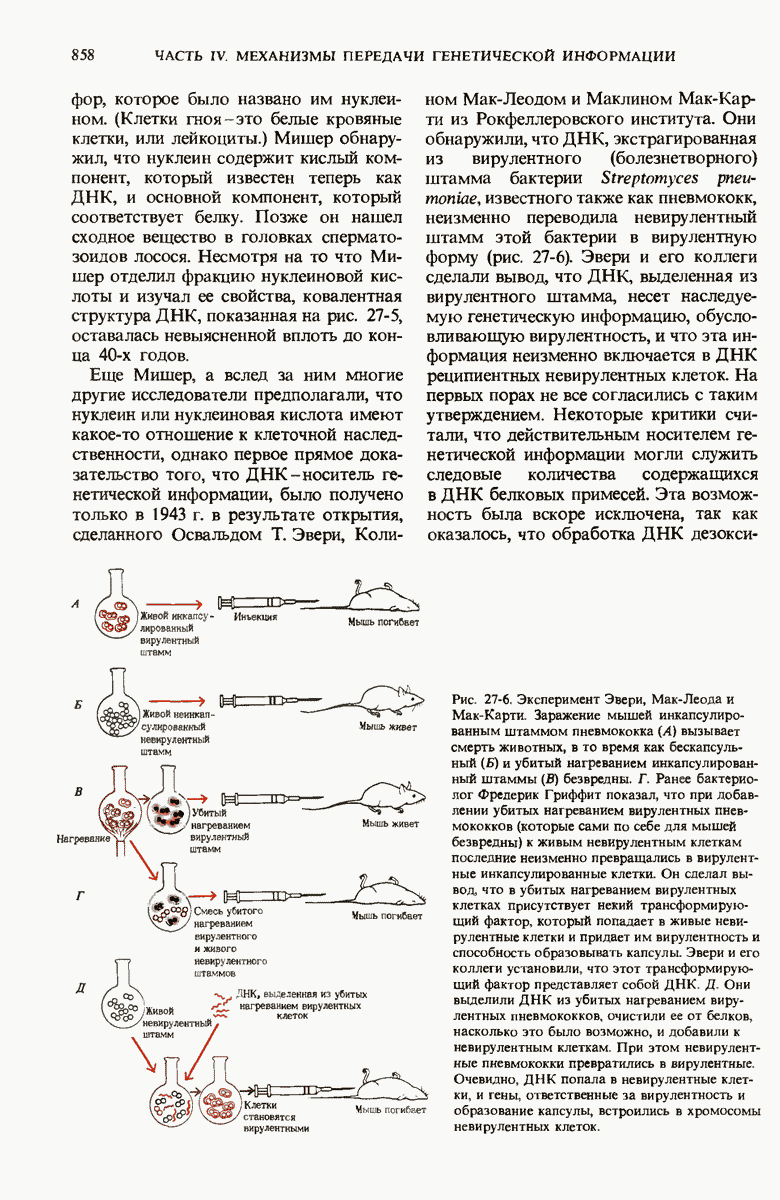
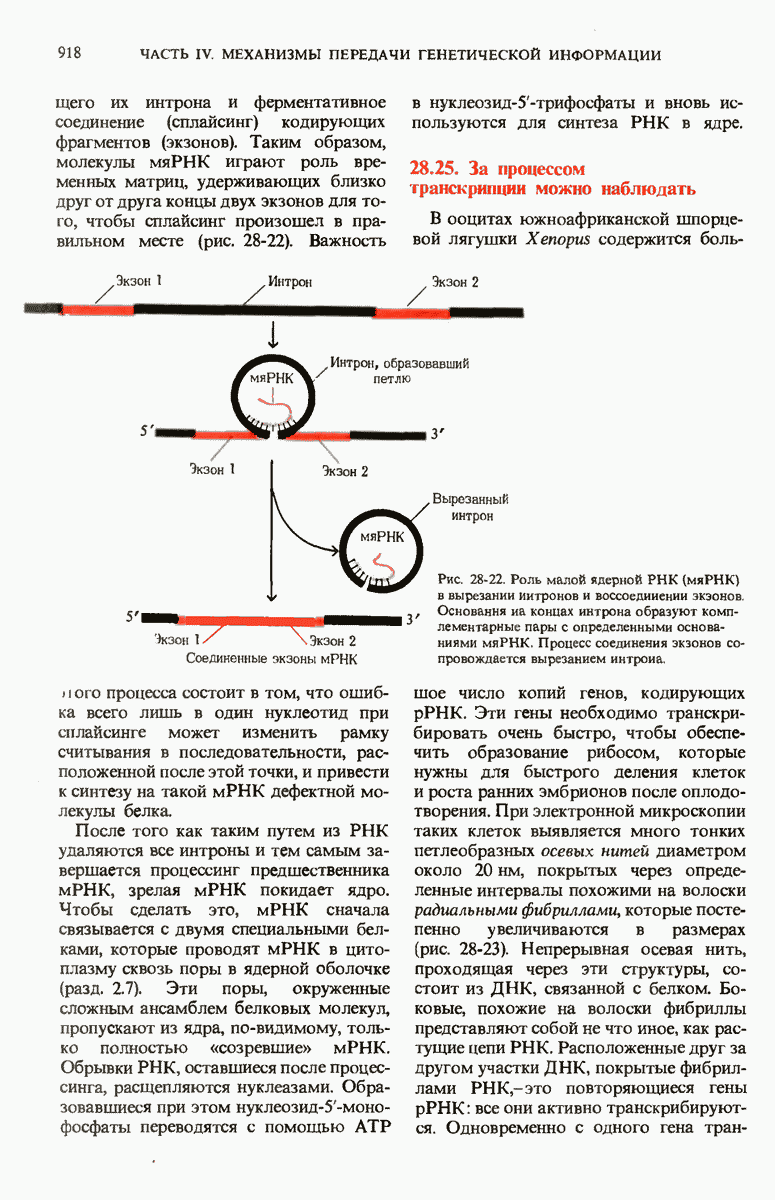
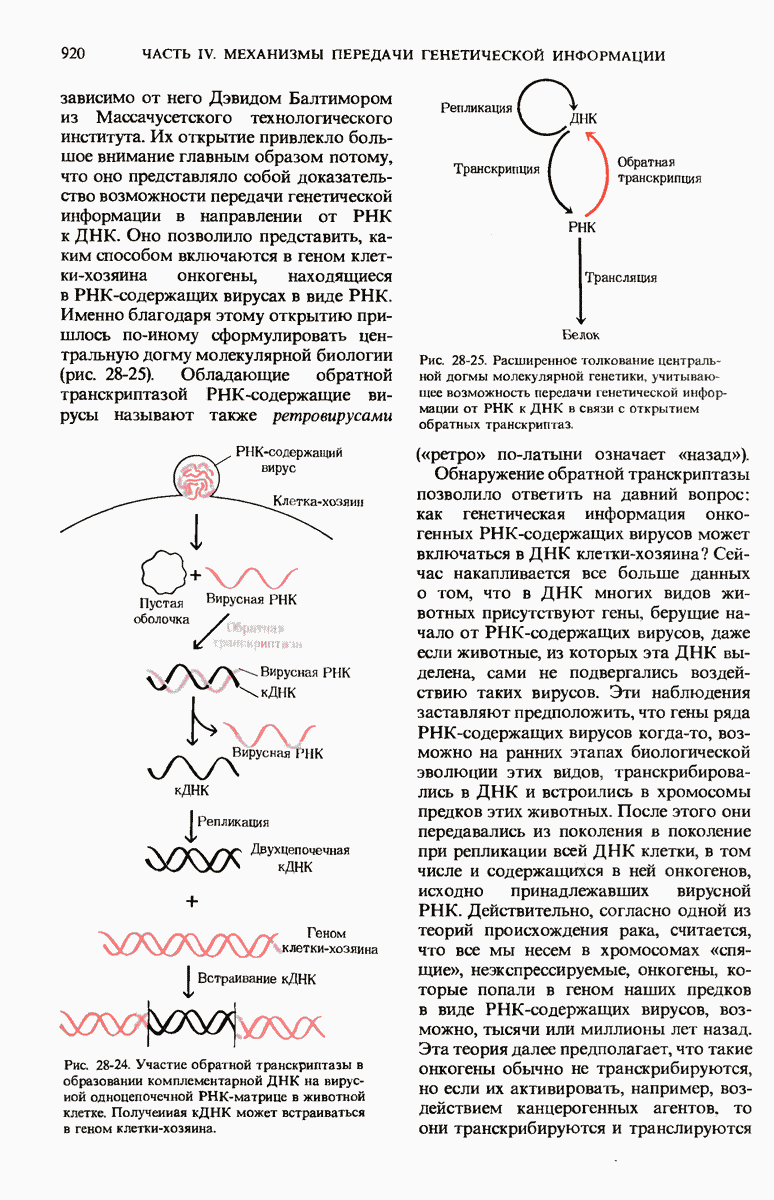
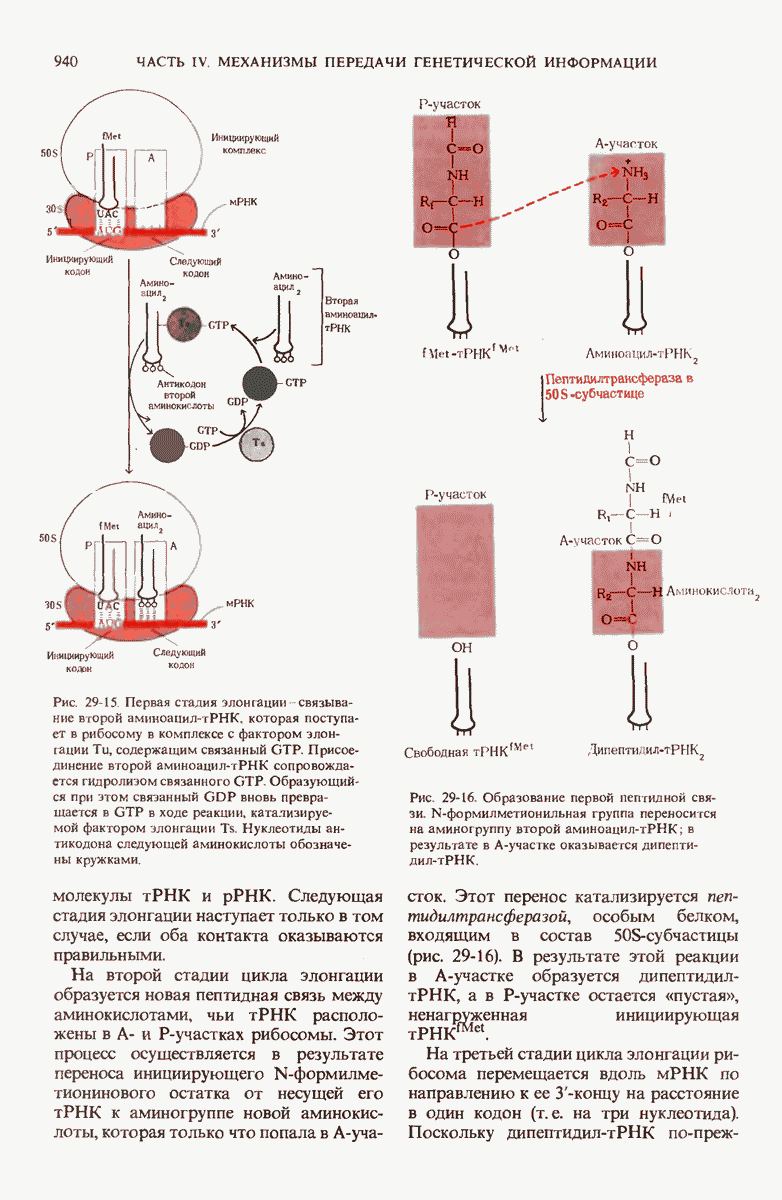
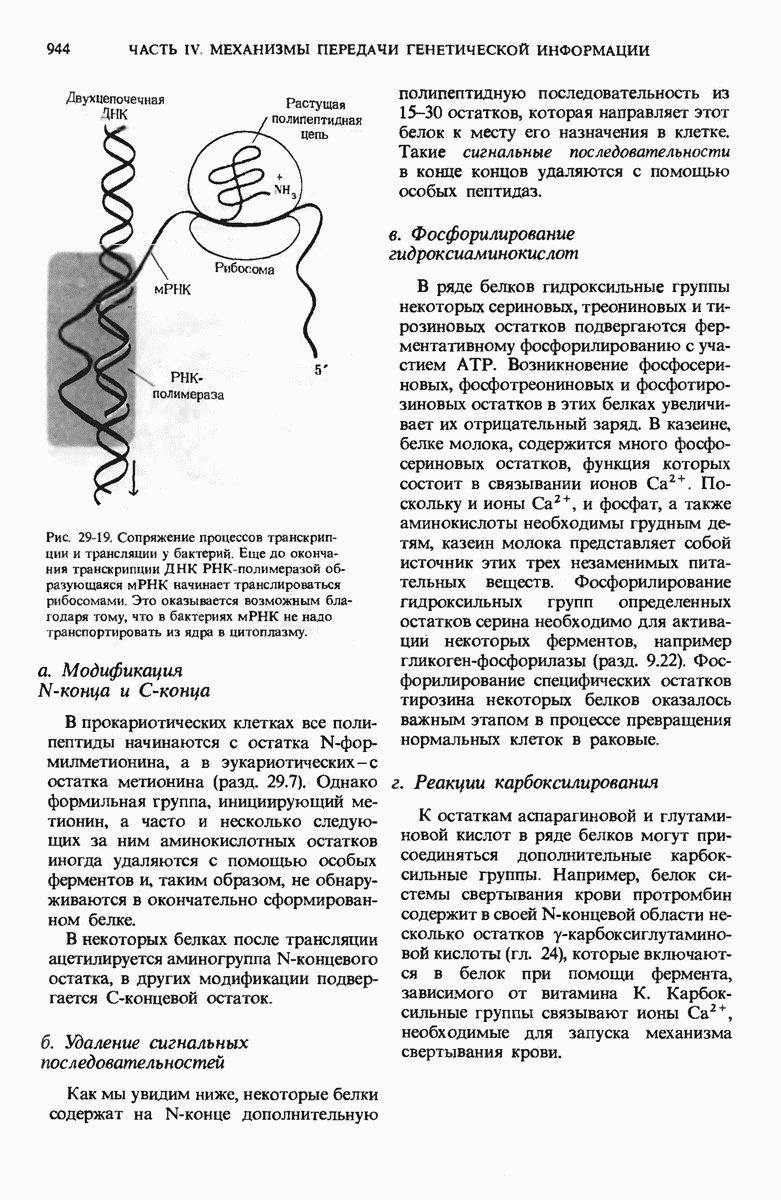
Рис. 29-19. Сопряжение процессов транскрипции и трансляции у бактерий. Еще до окончания транскрипции ДНК РНК-полимеразой образующаяся мРНК начинает транслироваться рибосомами. Это оказывается возможным благодаря тому, что в бактериях мРНК не надо транспортировать из ядра в цитоплазму.
а. Модификация N-конца и С-конца
В прокариотических клетках все полипептиды начинаются с остатка N-формилметионина, а в эукариотических - с остатка метионина (разд. 29.7). Однако формильная группа, инициирующий метионин, а часто и несколько следующих за ним аминокислотных остатков иногда удаляются с помощью особых ферментов и, таким образом, не обнаруживаются в окончательно сформированном белке.
В некоторых белках после трансляции ацетилируется аминогруппа N-концевого остатка, в других модификации подвергается С-концевой остаток.
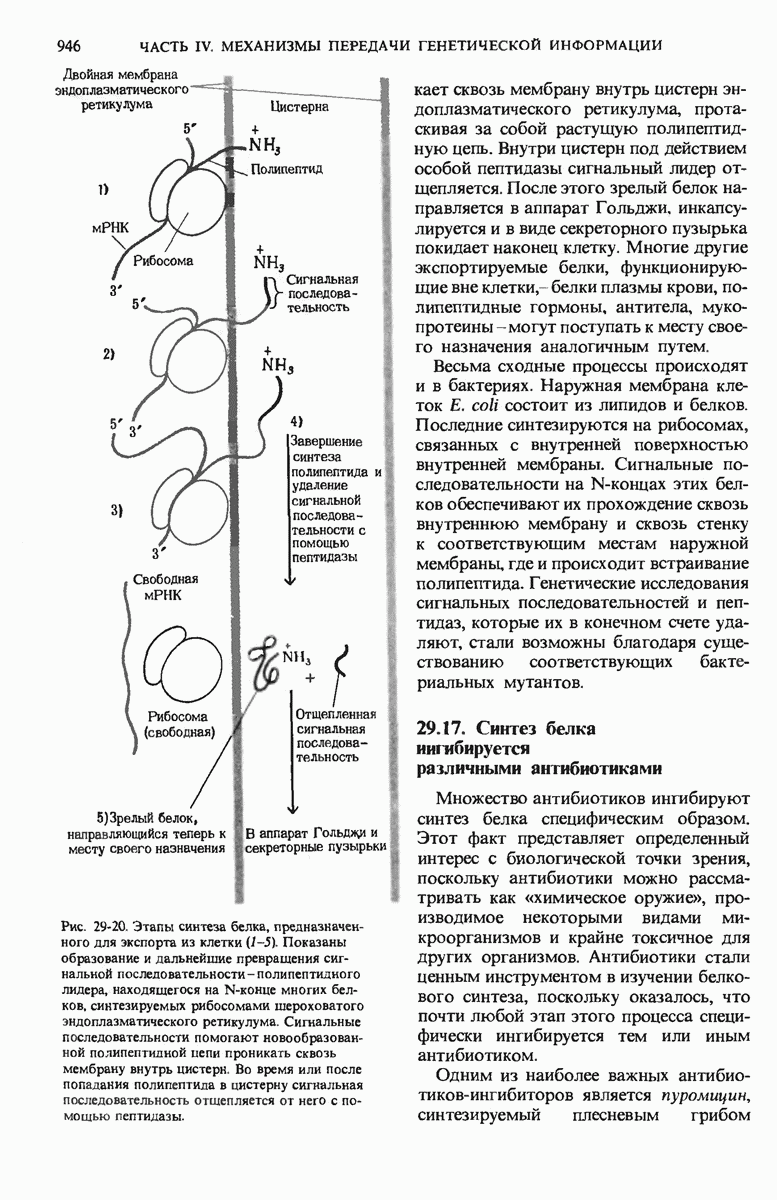
б. Удаление сигнальных последовательностей
Как мы увидим ниже, некоторые белки содержат на N-конце дополнительную полипептидную последовательность из 15-30 остатков, которая направляет этот белок к месту его назначения в клетке. Такие сигнальные последовательности в конце концов удаляются с помощью особых пептидаз.
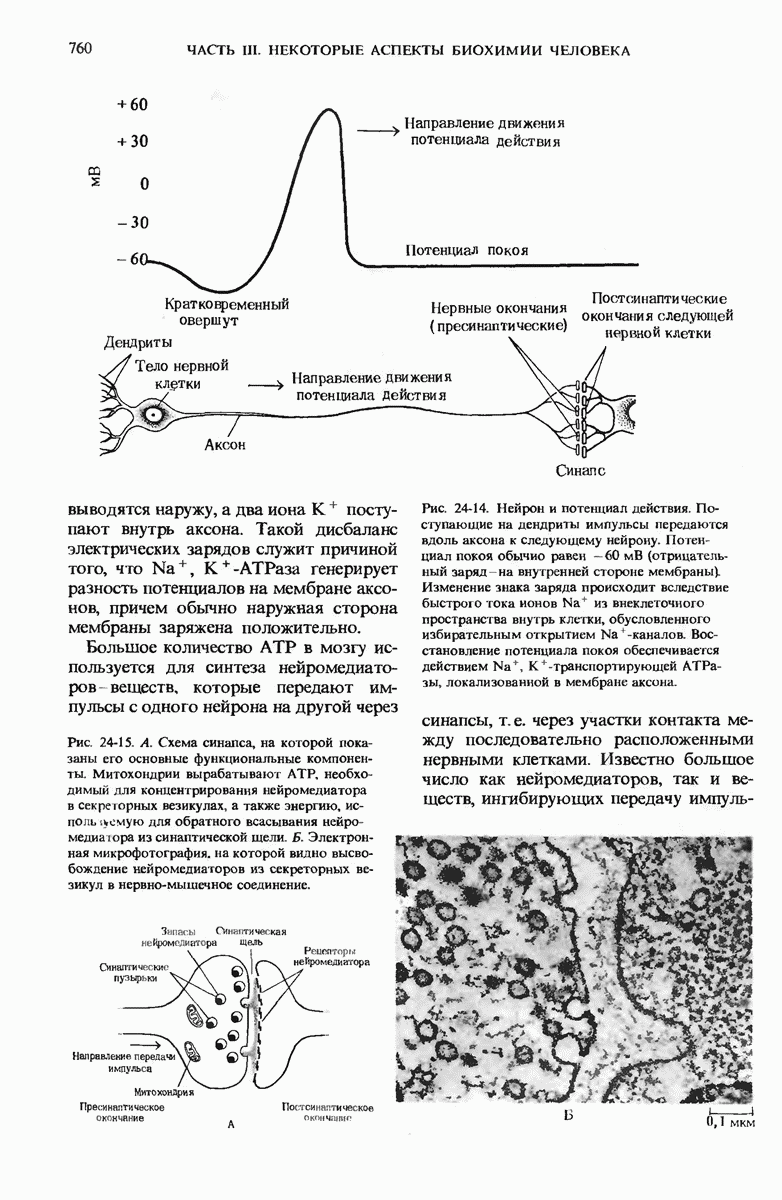
в. Фосфорилирование гидроксиаминокислот
В ряде белков гидроксильные группы некоторых сериновых, треониновых и тирозиновых остатков подвергаются ферментативному фосфорилированию с участием АТР. Возникновение фосфосериновых, фосфотреониновых и фосфотирозиновых остатков в этих белках увеличивает их отрицательный заряд. В казеине, белке молока, содержится много фосфосериновых остатков, функция которых состоит в связывании ионов  . Поскольку и ионы
. Поскольку и ионы  , и фосфат, а также аминокислоты необходимы грудным детям, казеин молока представляет собой источник этих трех незаменимых питательных веществ. Фосфорилирование гидроксильных групп определенных остатков серина необходимо для активаций некоторых ферментов, например гликоген-фосфорилазы (разд. 9.22). Фосфорилирование специфических остатков тирозина некоторых белков оказалось важным этапом в процессе превращения нормальных клеток в раковые.
, и фосфат, а также аминокислоты необходимы грудным детям, казеин молока представляет собой источник этих трех незаменимых питательных веществ. Фосфорилирование гидроксильных групп определенных остатков серина необходимо для активаций некоторых ферментов, например гликоген-фосфорилазы (разд. 9.22). Фосфорилирование специфических остатков тирозина некоторых белков оказалось важным этапом в процессе превращения нормальных клеток в раковые.
г. Реакции карбоксилирования
К остаткам аспарагиновой и глутаминовой кислот в ряде белков могут присоединяться дополнительные карбоксильные группы. Например, белок системы свертывания крови протромбин содержит в своей N-концевой области несколько остатков  -карбоксиглутаминовой кислоты (гл. 24), которые включаются в белок при помощи фермента, зависимого от витамина К. Карбоксильные группы связывают ионы
-карбоксиглутаминовой кислоты (гл. 24), которые включаются в белок при помощи фермента, зависимого от витамина К. Карбоксильные группы связывают ионы  необходимые для запуска механизма свертывания крови.
необходимые для запуска механизма свертывания крови.
д. Метилирование групп
В ряде белков определенные остатки лизина подвергаются ферментативному метилированию. Остатки монометил и диметиллизина обнаруживаются в некоторых мышечных белках и в цитохроме с. В других белках метилированию подвергаются карбоксильные группы ряда остатков глутаминовой кислоты, что приводит к нейтрализации их отрицательных зарядов.
е. Присоединение боковых углеводных цепей
Боковые углеводные цепи гликопротеинов ковалентно присоединяются к полипептиду во время или после синтеза последнего. В одних гликопротеинах боковая углеводная цепь прикрепляется с помощью ферментов к остаткам аспарагиновой кислоты, в других - серина или треонина. Многие белки, которые работают вне клетки, а также «смазочные» протеогликаны, покрывающие слизистые оболочки, содержат боковые олигосахаридные цепи.
ж. Добавление простетических групп
В состав многих ферментов входят обязательные для их активности ковалентно связанные с белком простетические группы; они гакже присоединяются к полипептидной цепи после того, как та покидает рибосому. Примерами таких простетических групп могут служить молекула биотина, ковалентно связанная с ацетил-СоА-карбоксилазой (разд. 21.2), и гемовая группа цитохрома с (разд. 17.10).
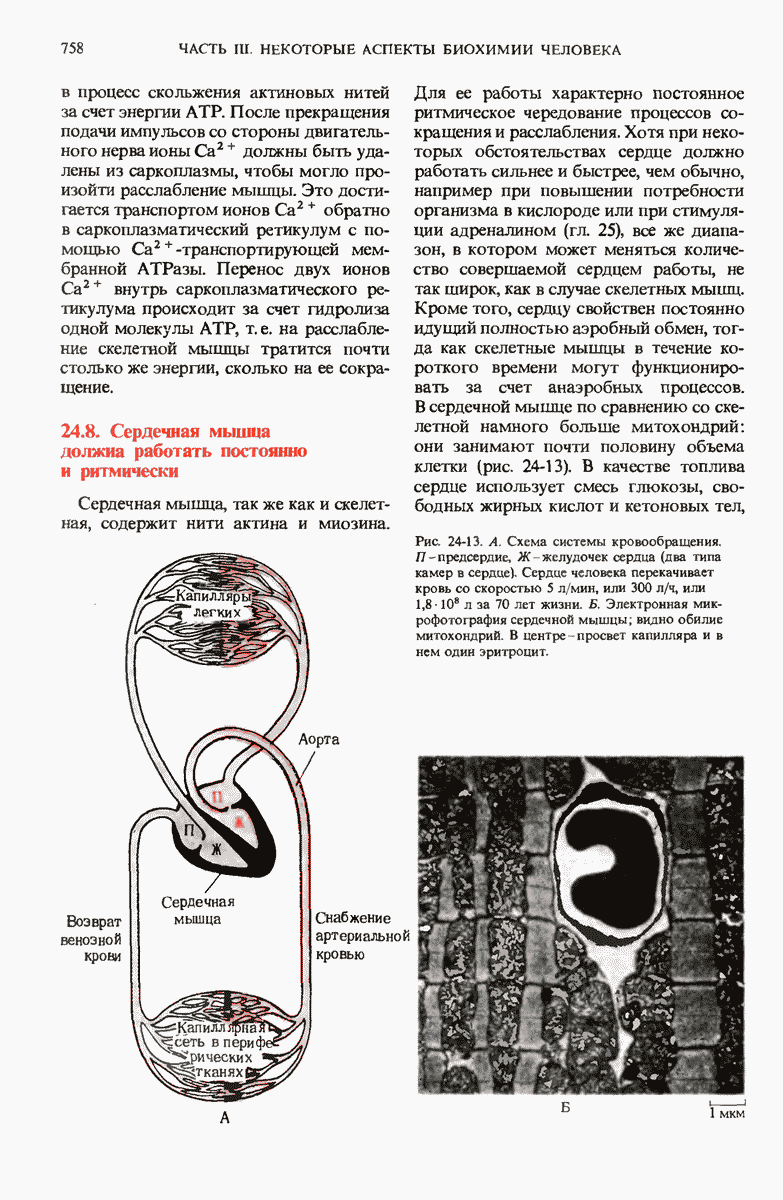
з. Образование дисульфидных мостиков
Во многих белках, предназначенных для выхода из эукариотической клетки, в процессе формирования их нативной конформации появляются поперечные сшивки в результате ферментативного образования дисульфидных мостиков между остатками цистина; эти мостики соединяют друг с другом две полипептидные цепи или две части одной цепи (разд. 6.8). Поперечные дисулъфидные мостики помогают уберечь нативную конформацию белковой молекулы от денатурации.