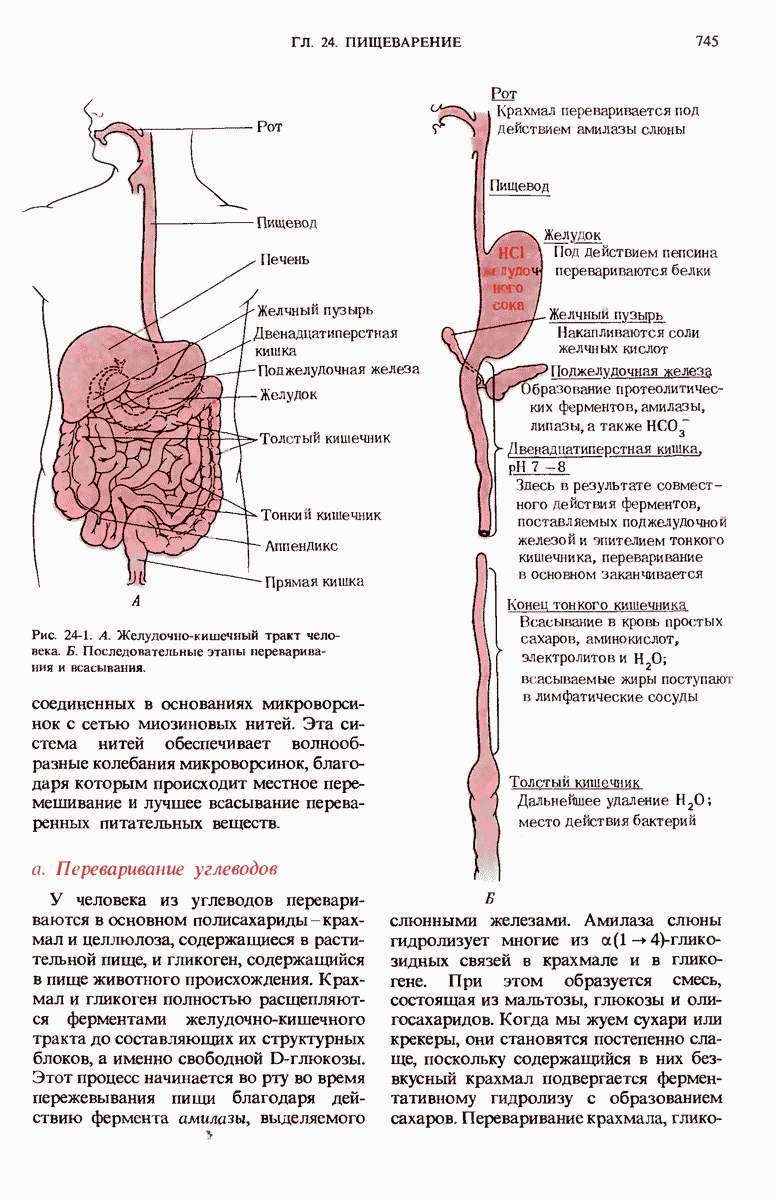
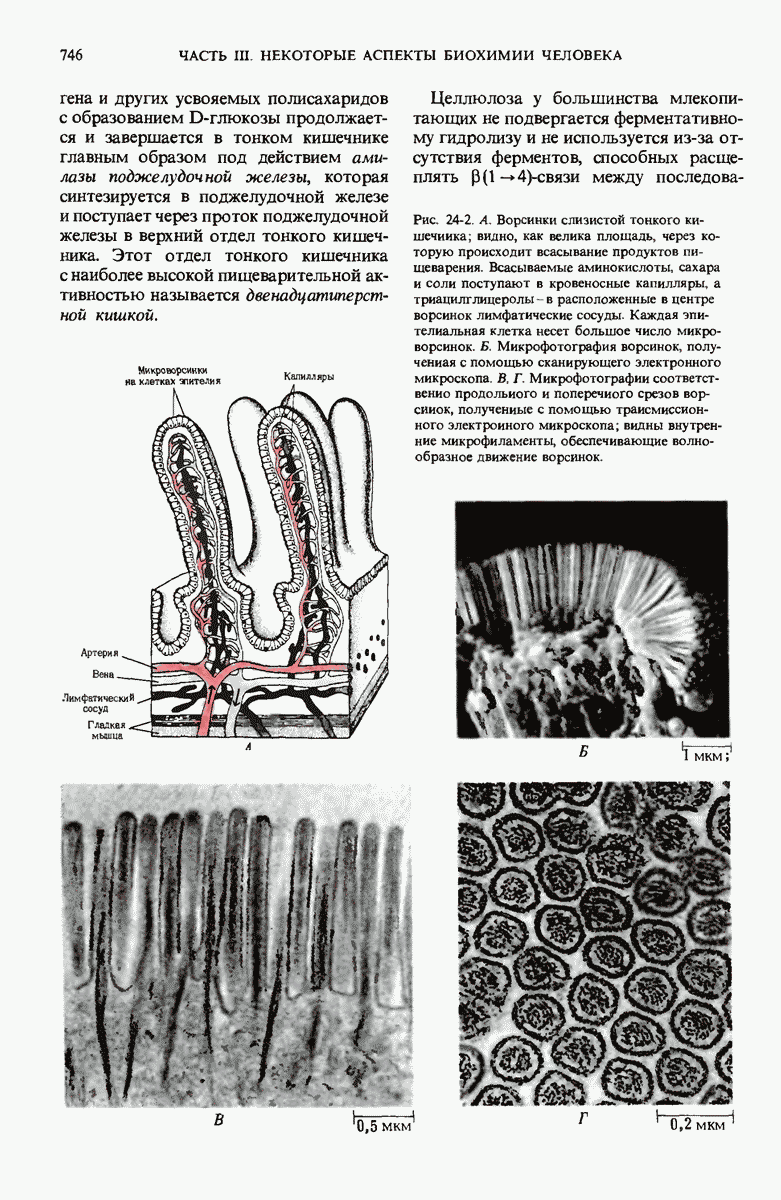
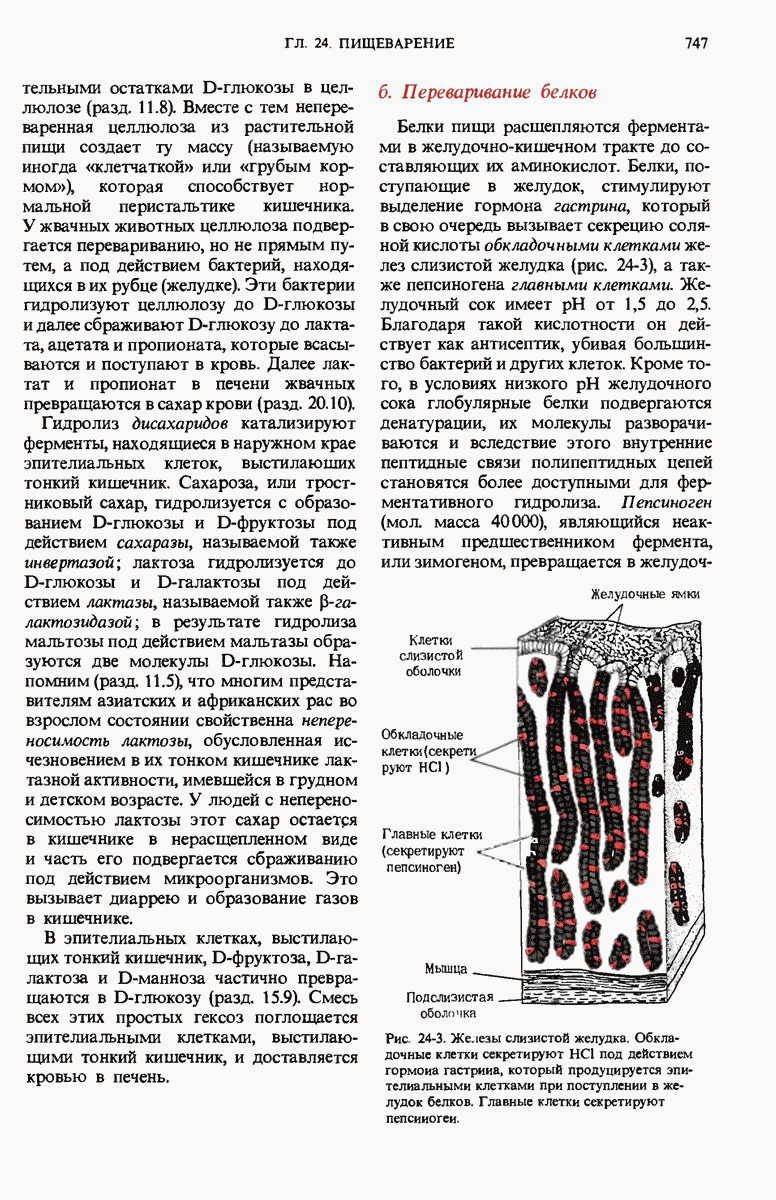
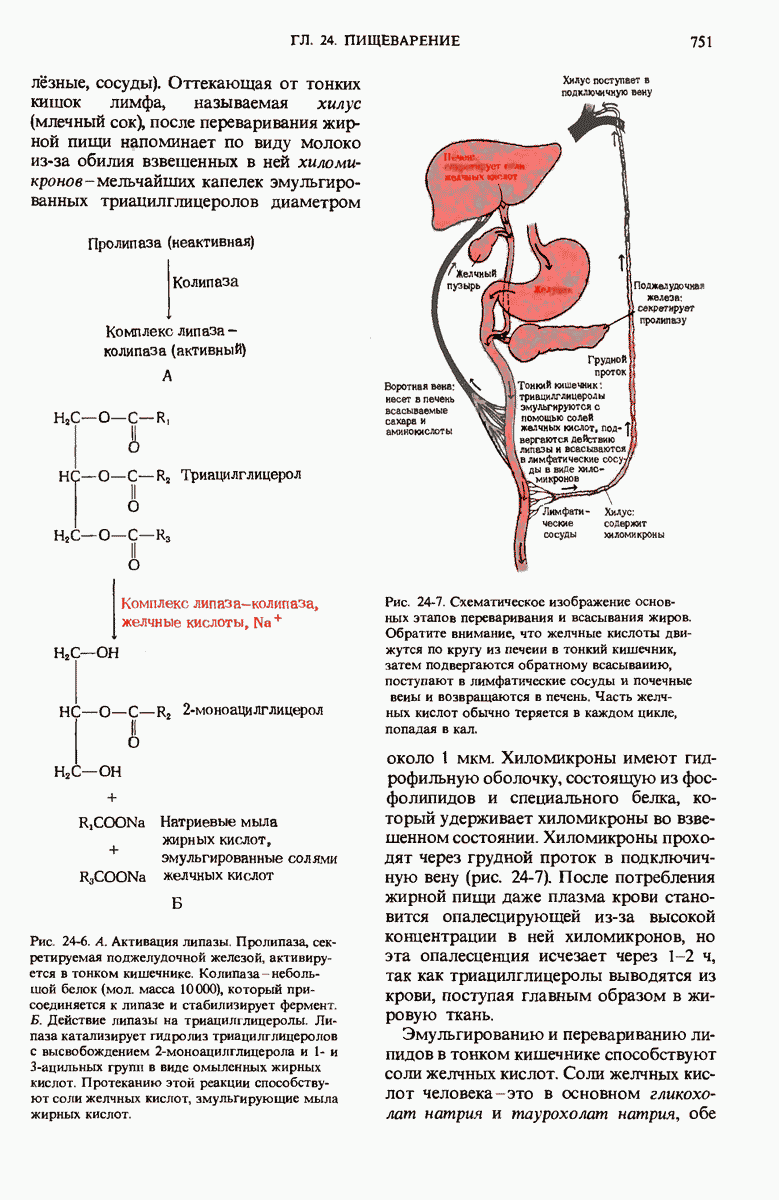
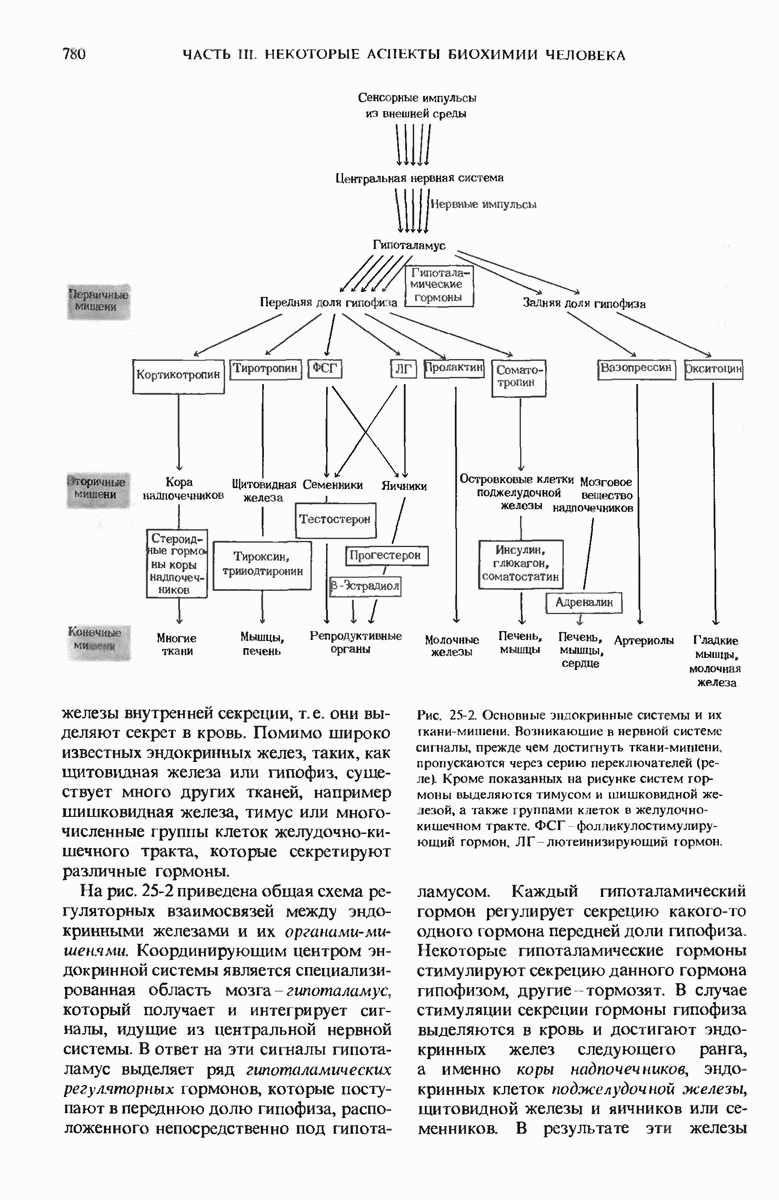
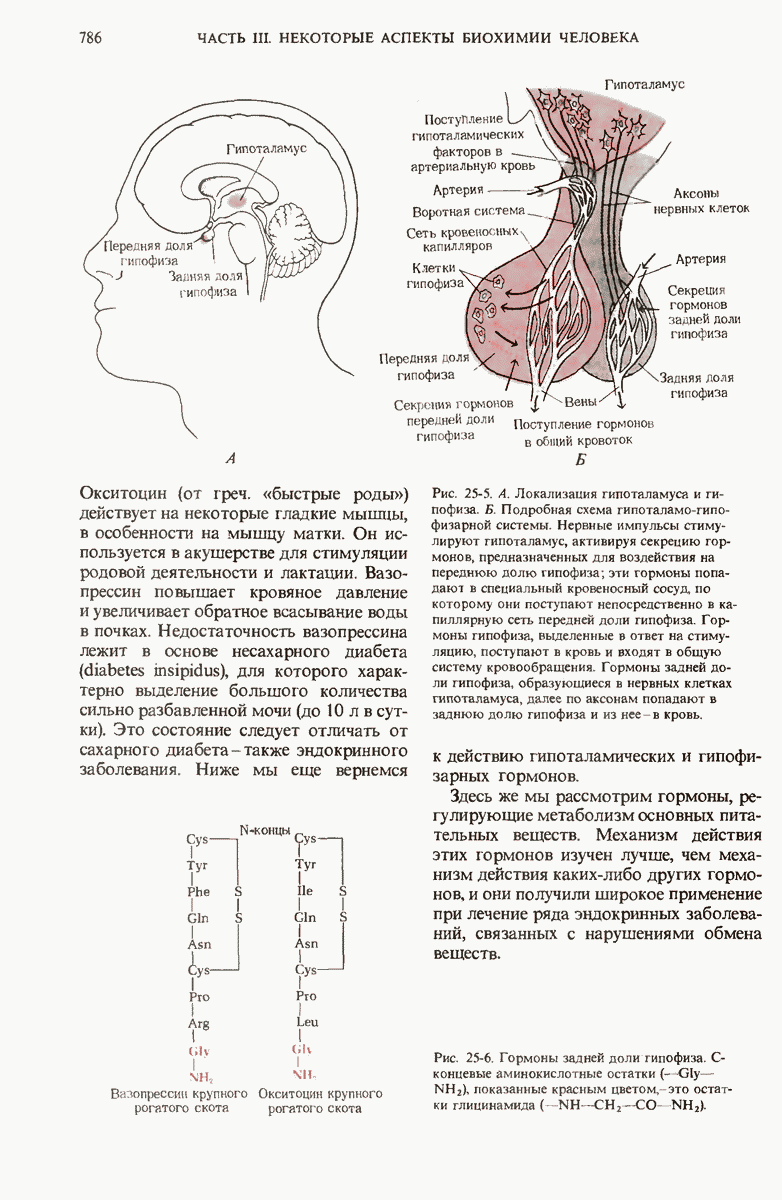
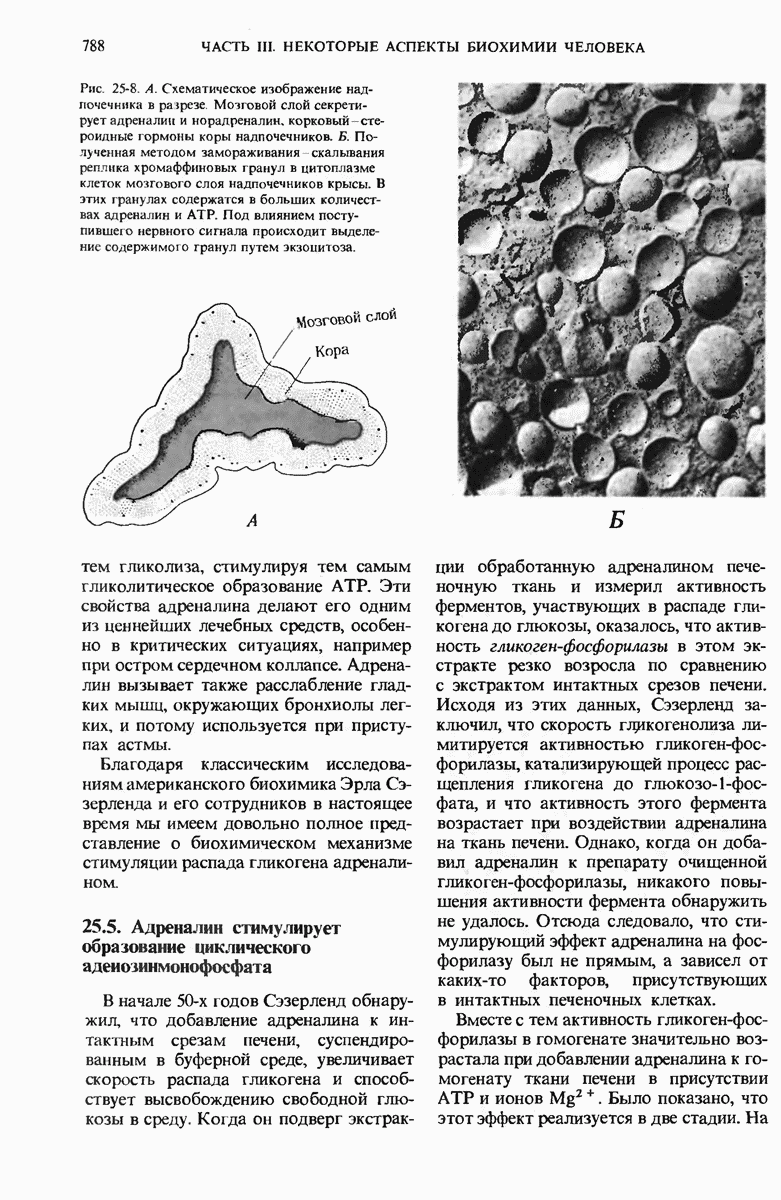
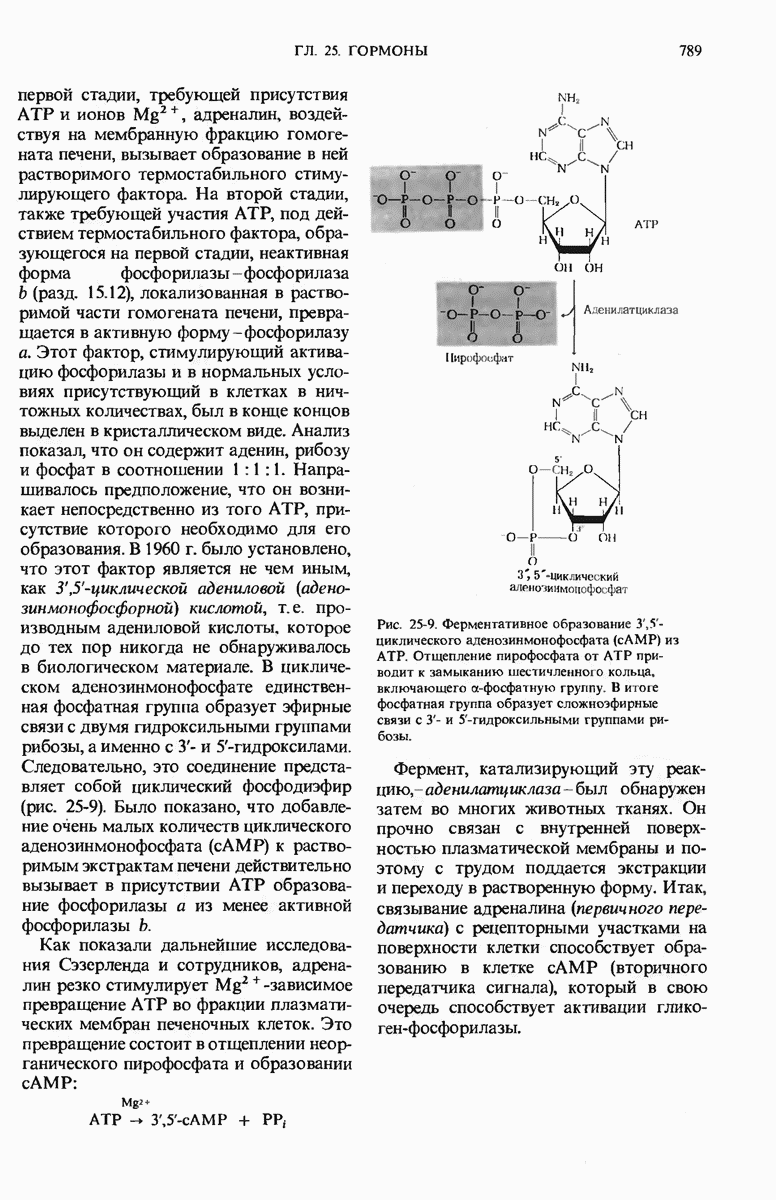
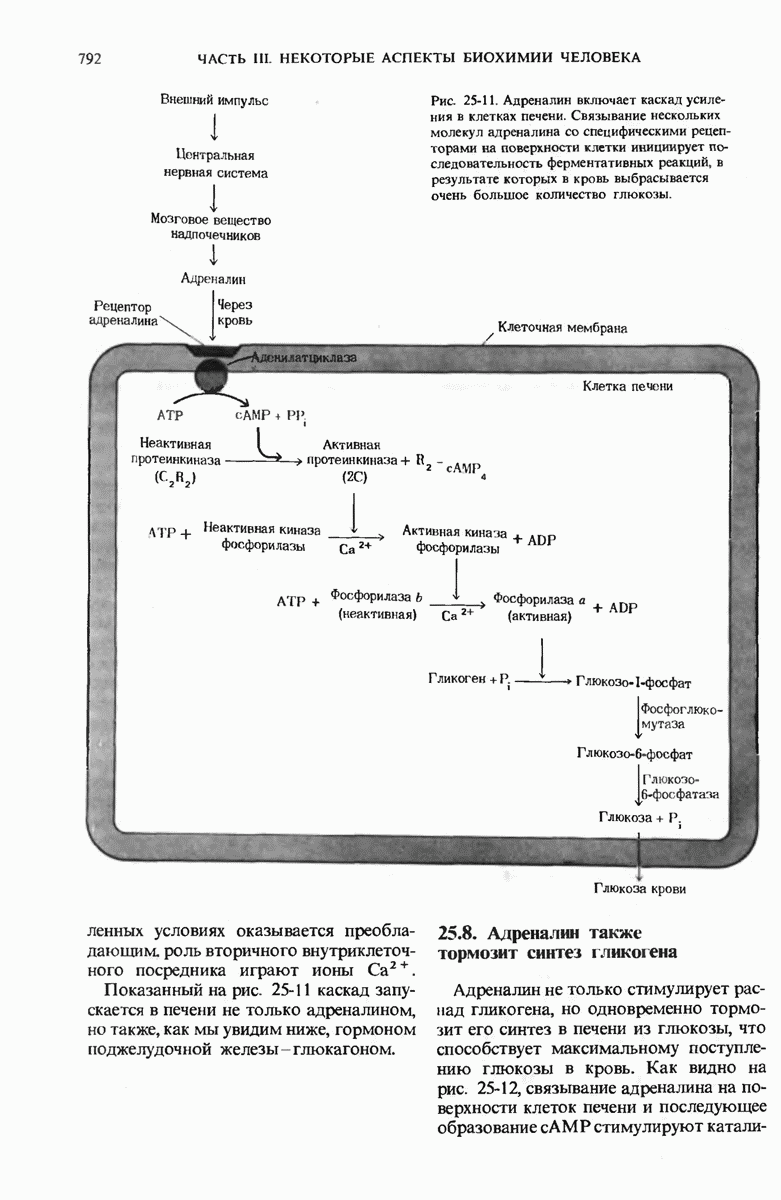
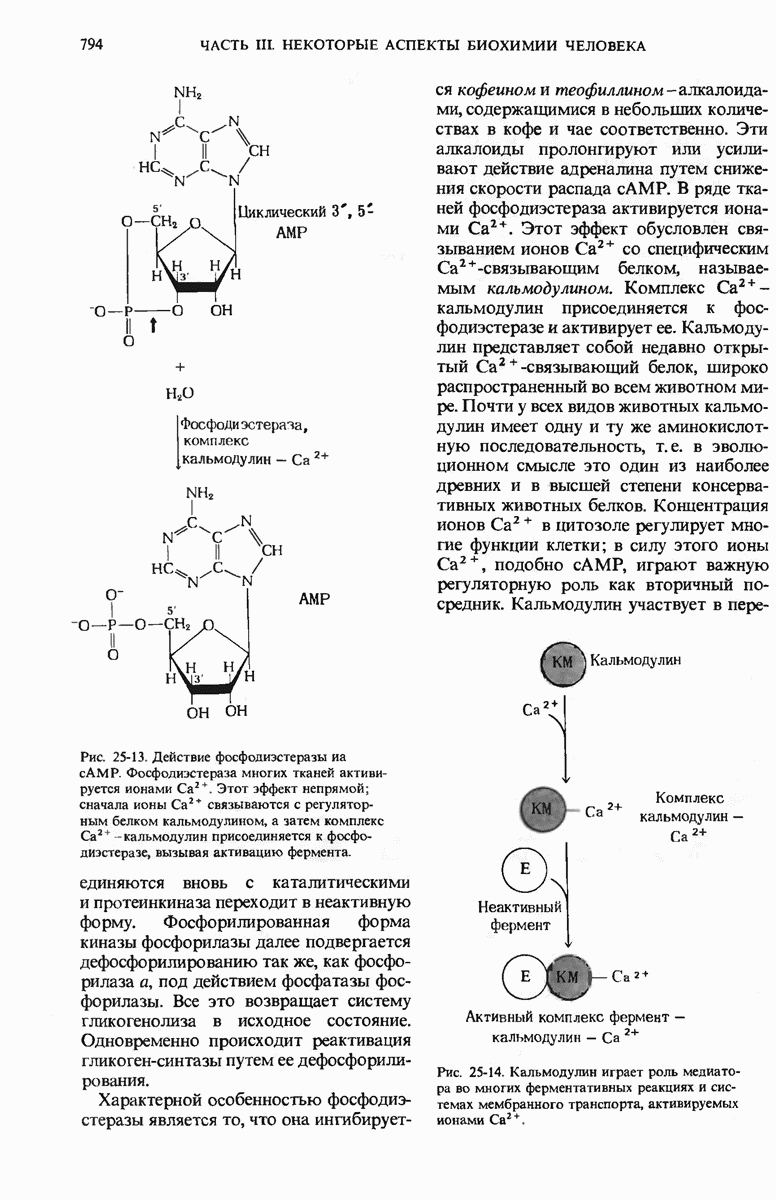
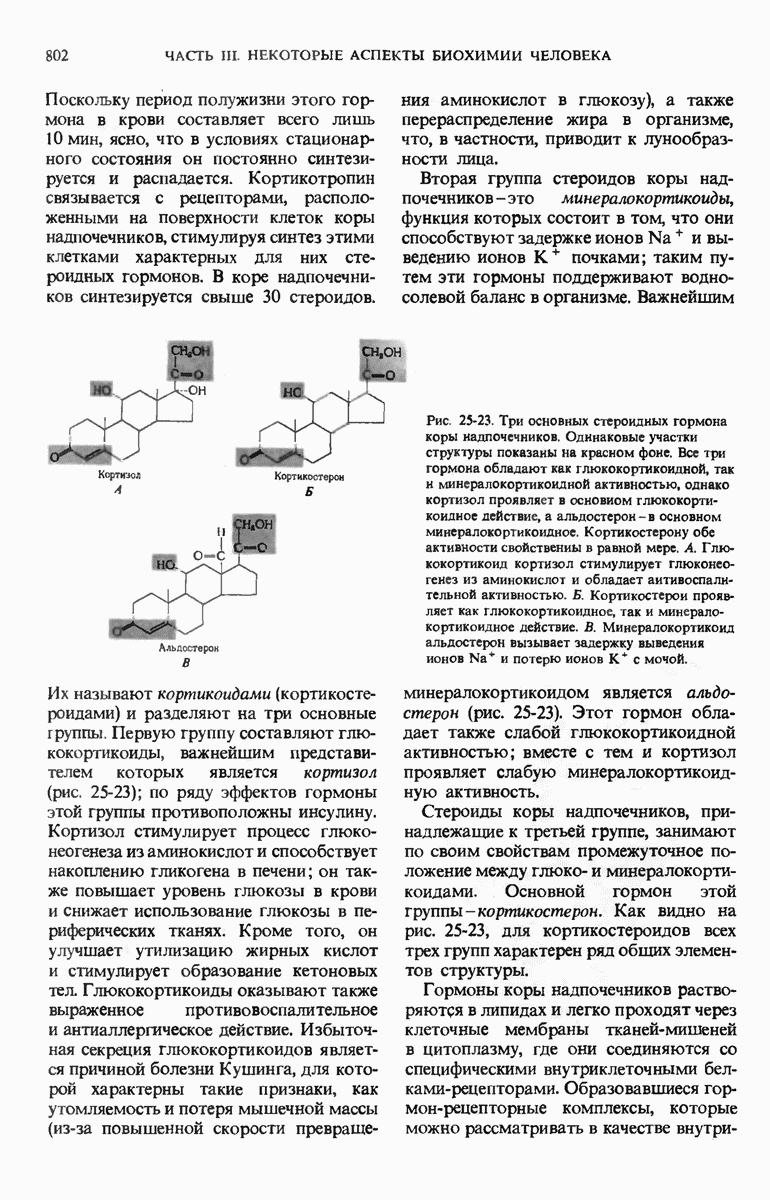
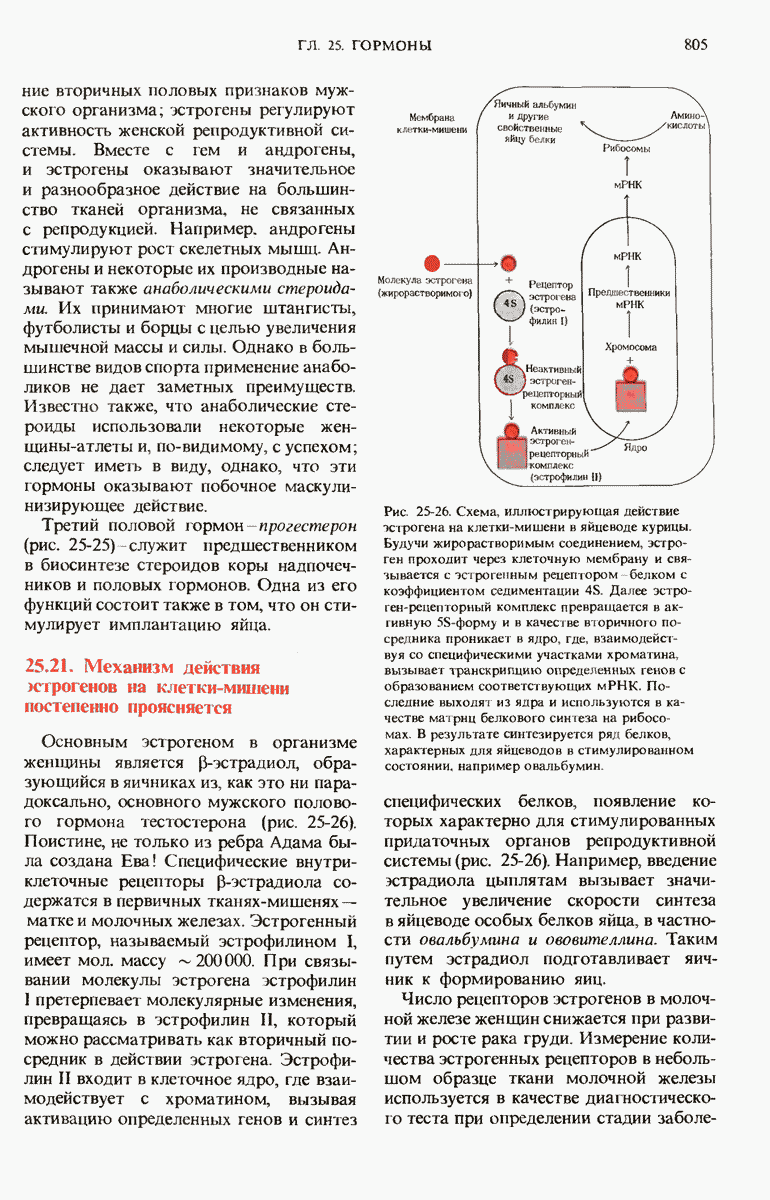
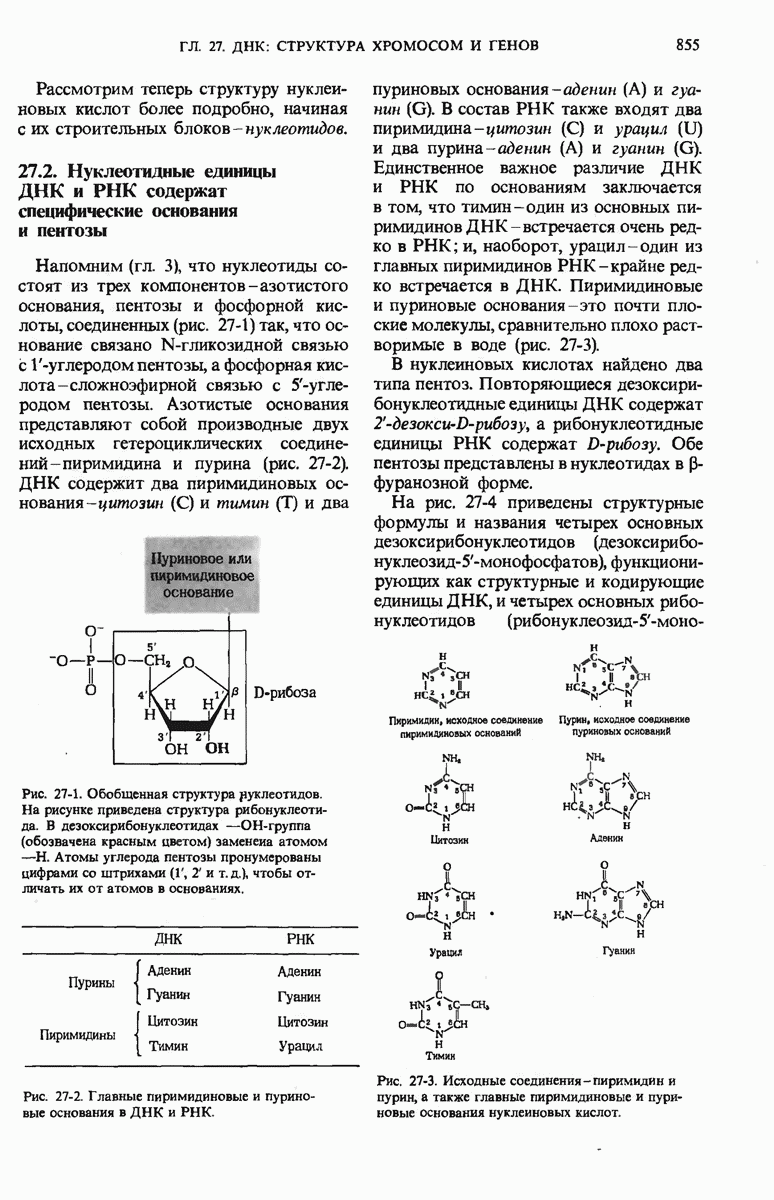
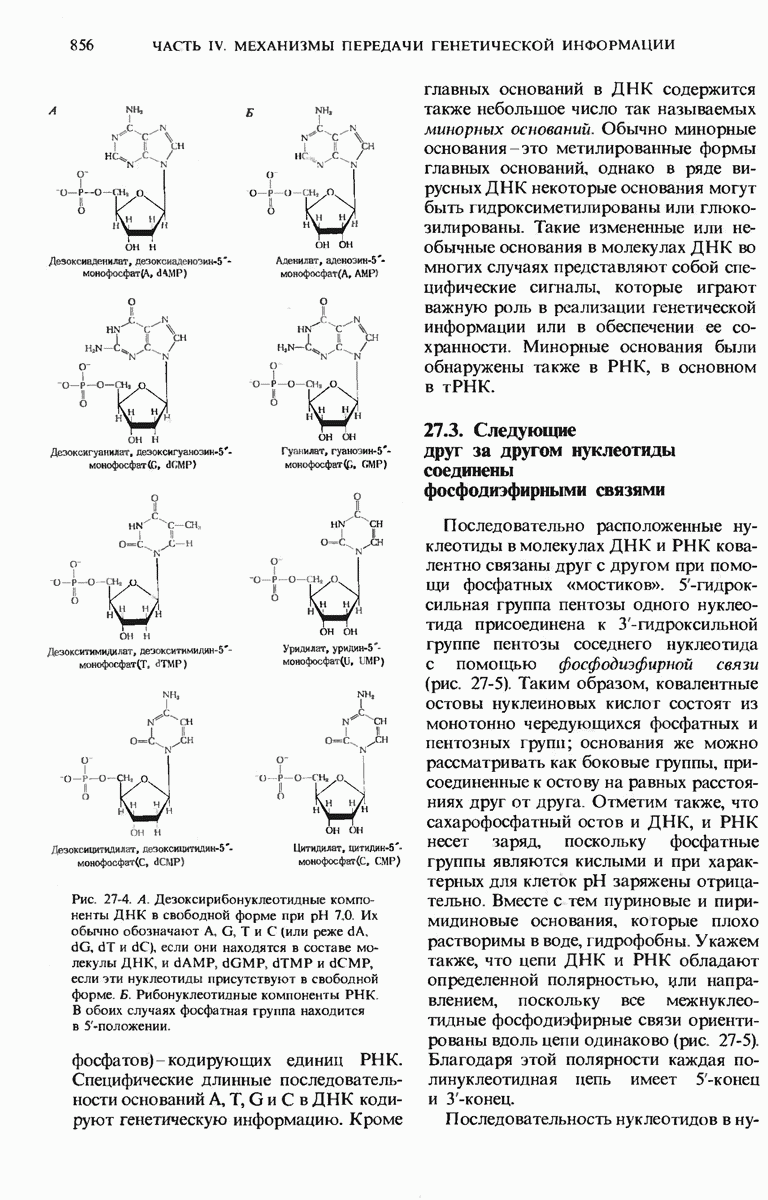
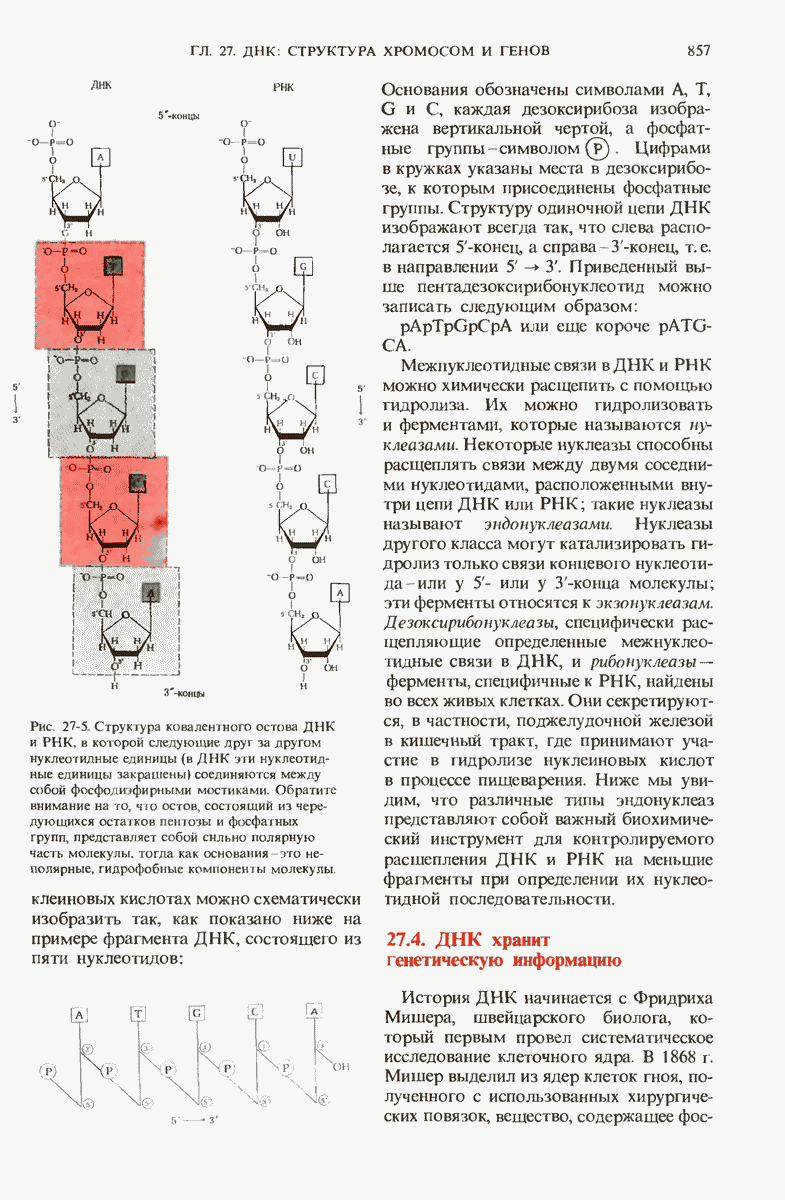
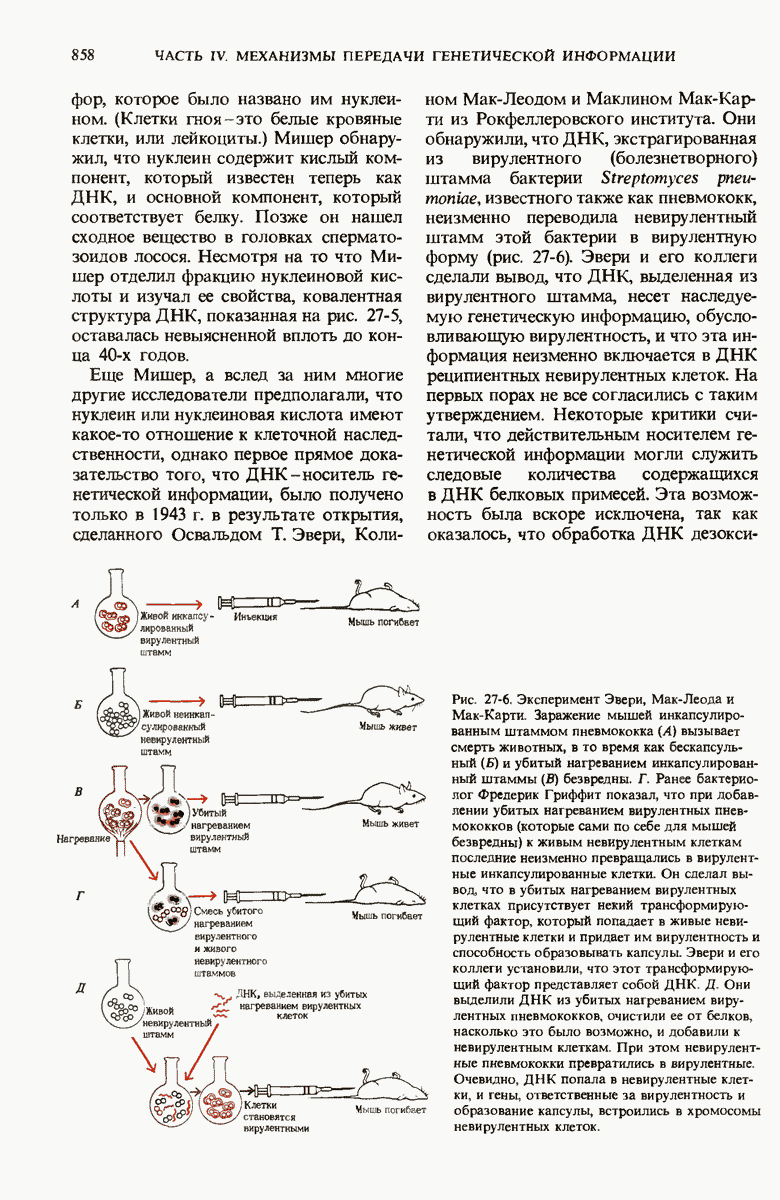
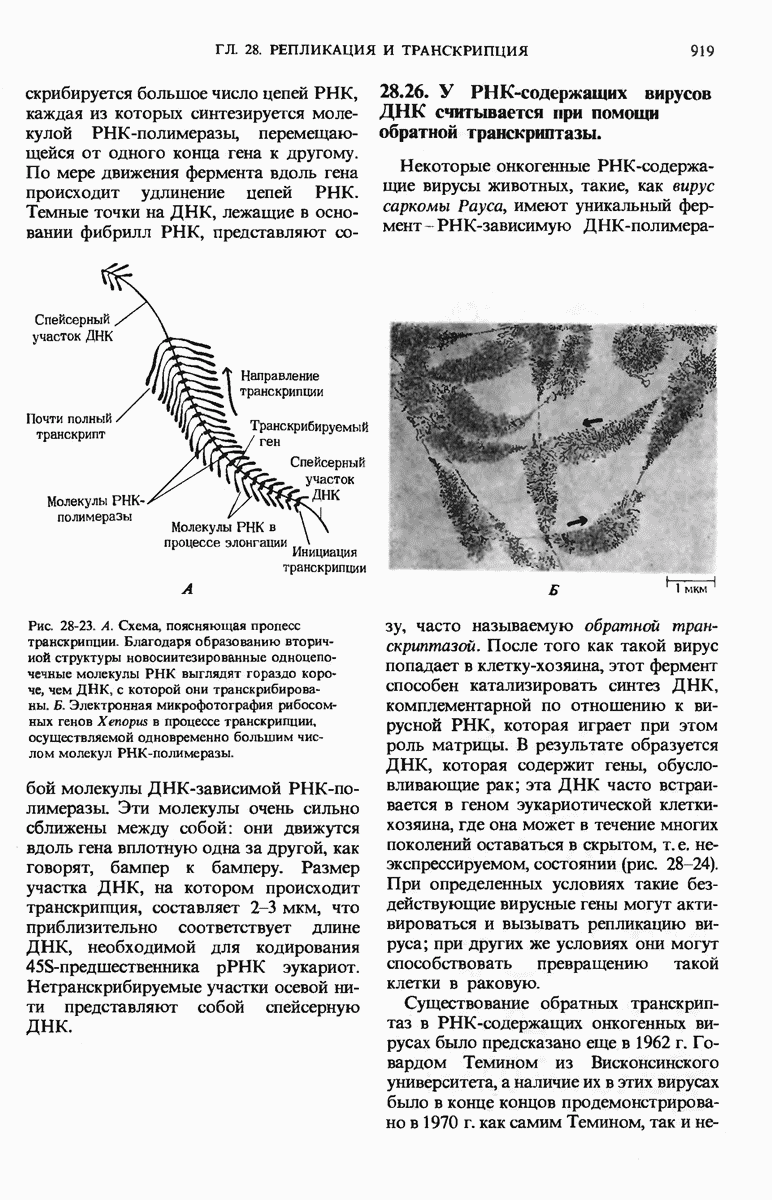
Краткое содержание главы
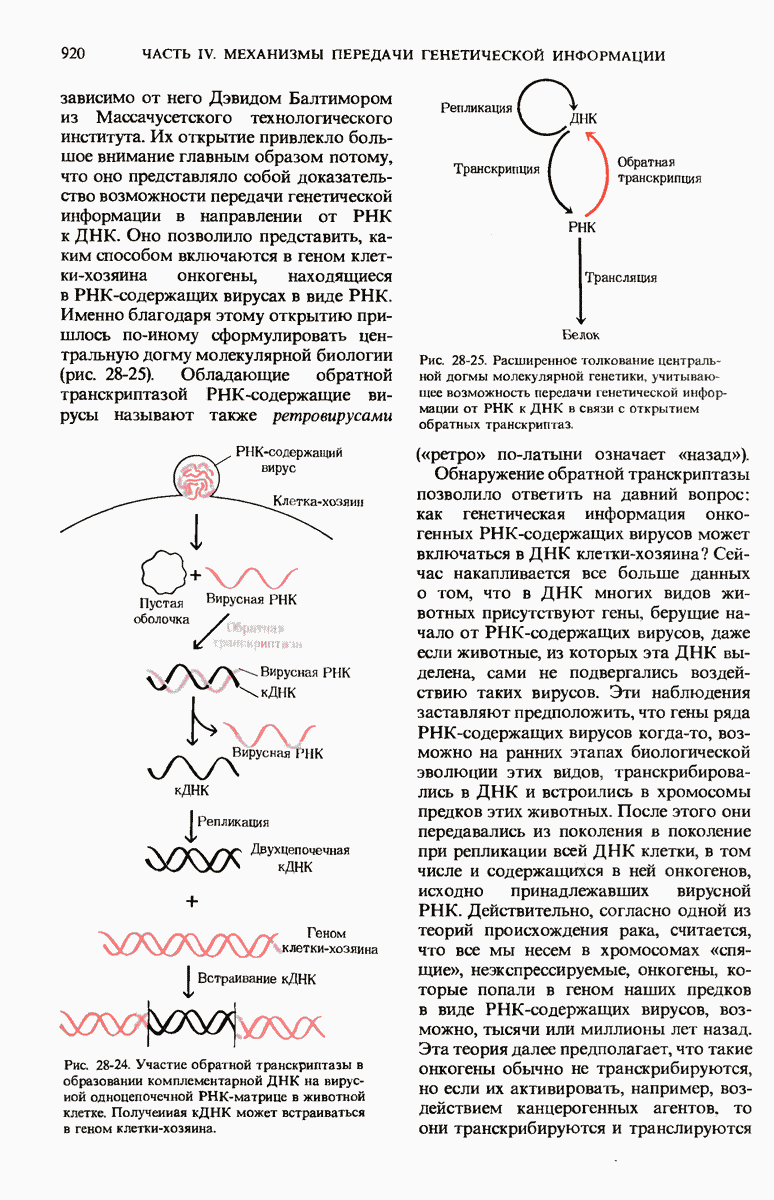
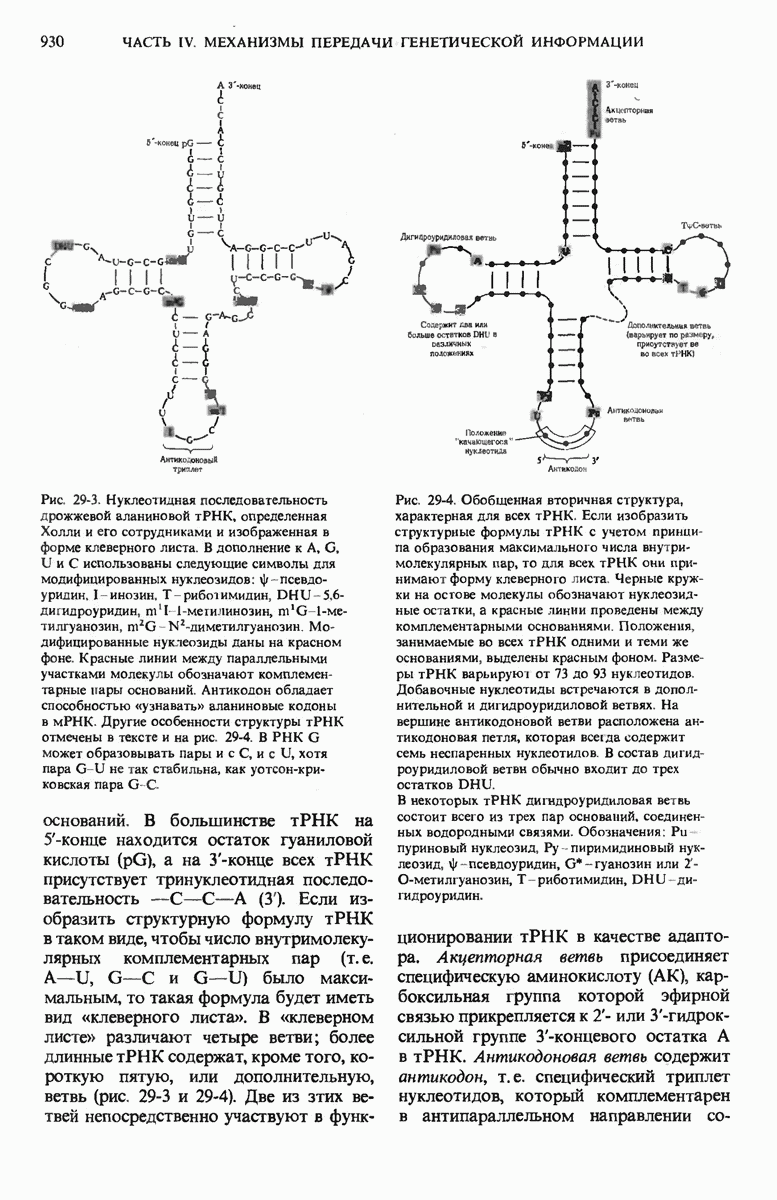
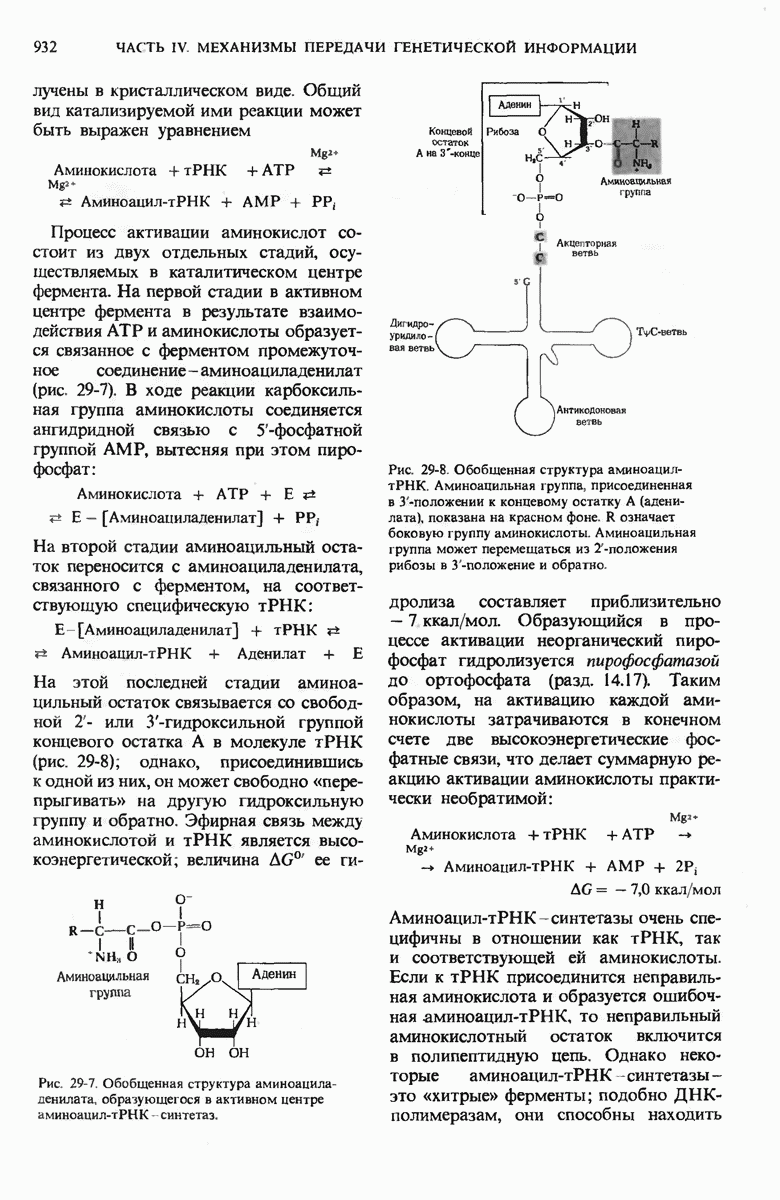
Чтобы принять участие в синтезе белка, сначала аминокислоты активируются в цитозоле с помощью специфических аминоацил-тРНК-синтетаз. Эти ферменты катализируют образование эфирной связи между аминоацильным остатком и соответствующей тРНК, сопровождающееся расщеплением АТР до АМР и пирофосфата. тРНК содержат от 73 до 93 нуклеотидных остатков, часть из которых включает модифицированные основания. Молекулы тРНк имеют в своем составе акцепторную ветвь с концевой последовательностью (3) -С-С- (к которой эфирной связью присоединяется аминокислота), антикодоновую ветвь,  -ветвь и дигидроуридиловую ветвь; некоторые тРНК содержат пятую добавочную ветвь.
-ветвь и дигидроуридиловую ветвь; некоторые тРНК содержат пятую добавочную ветвь.

Рис. 29-28. Структура промотор-операторной области lас-оперона Е. coli. Показана нуклеотидная последовательность обеих цепей ДНК. начиная с последних 15 оснований регуляторного  гена и кончая первыми девятью основаниями гена г. Видно, что промотор перекрывает оператор. Участок связывания комплекса САР-сАМР состоит приблизительно из 38 оснований, а участок первоначального связывания РНК-полимеразы-приблизительно из 40 оснований. Участок связывания lac-репрессора в операторе содержит около 28 пар оснований и характеризуется симметрией второго порядка.
гена и кончая первыми девятью основаниями гена г. Видно, что промотор перекрывает оператор. Участок связывания комплекса САР-сАМР состоит приблизительно из 38 оснований, а участок первоначального связывания РНК-полимеразы-приблизительно из 40 оснований. Участок связывания lac-репрессора в операторе содержит около 28 пар оснований и характеризуется симметрией второго порядка.
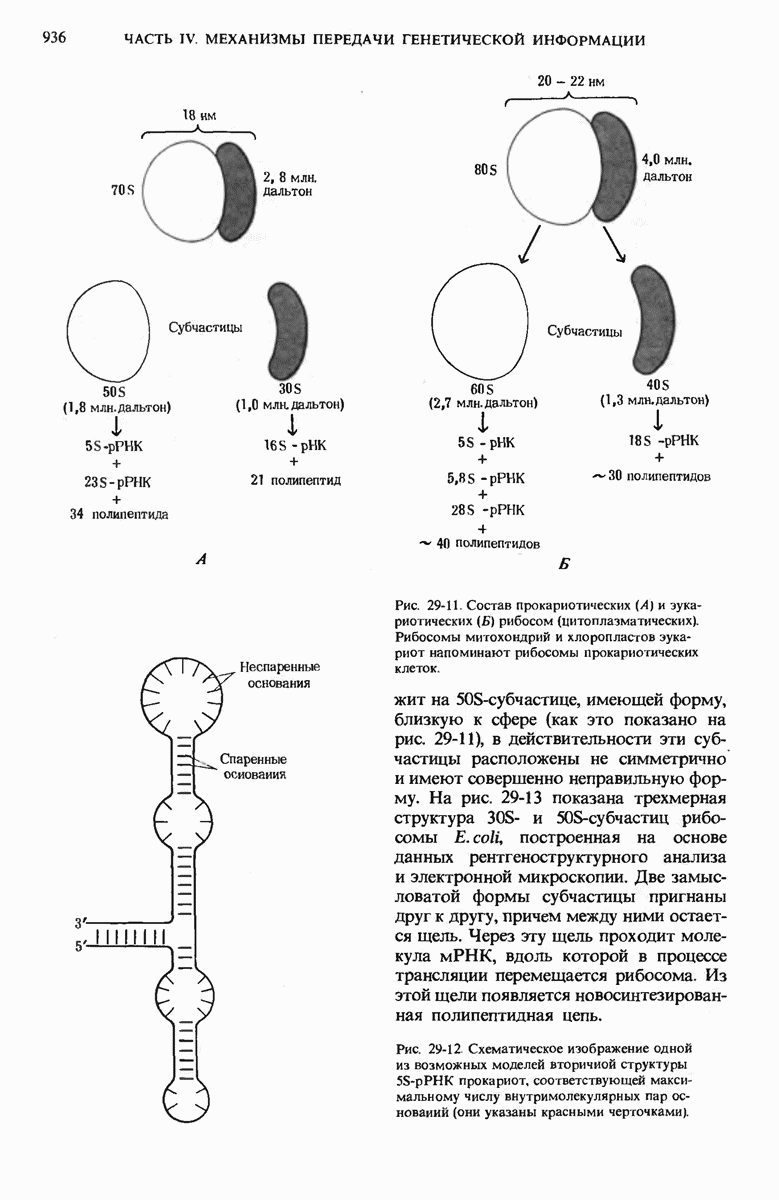
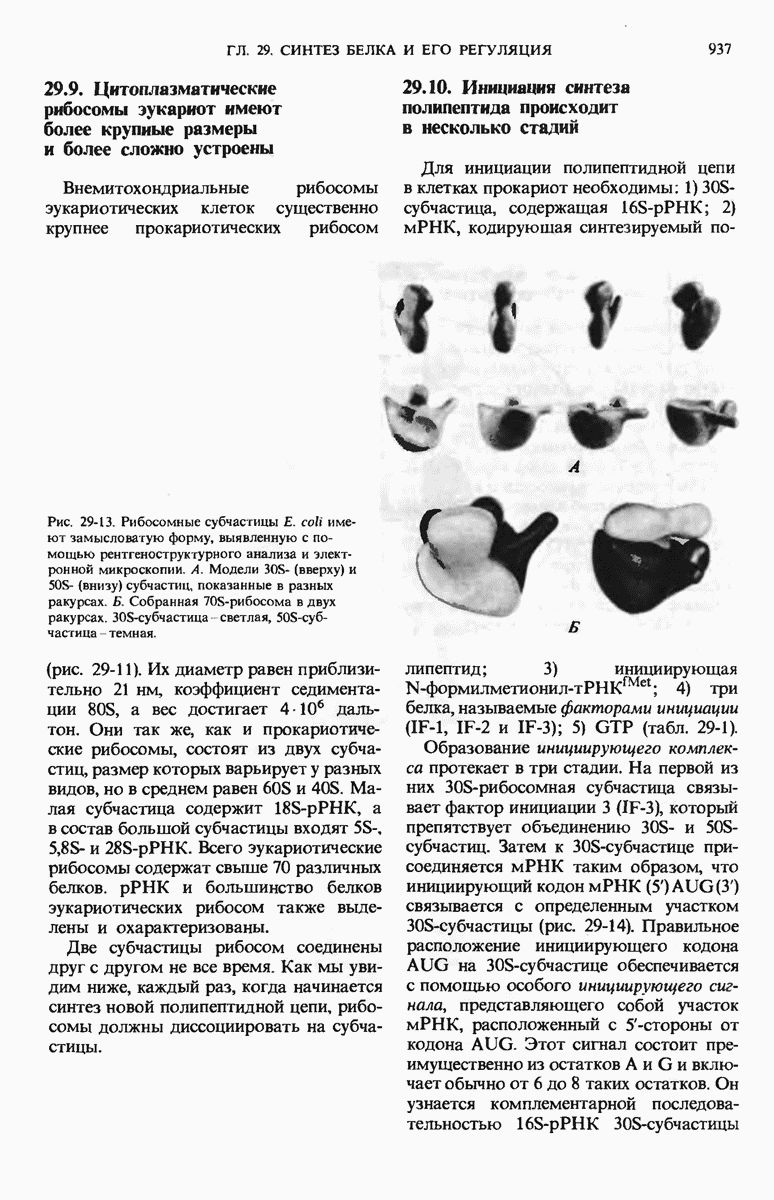
Тройка нуклеотидов в тРНК (триплет), образующая антикодон, отвечает за специфичность взаимодействия аминоацил-тРНК с комплементарным кодоновым триплетом в мРНК. Рост полипептидной цепи на рибосомах начинается с N-концевой аминокислоты и продолжается в результате последовательного добавления новых остатков к С-концу. Прокариоты содержат 70S-рибосомы, состоящие из большой 50S- и малой  -субчастиц. Эукариотические рибосомы значительно крупнее и содержат больше белков, чем прокариотические.
-субчастиц. Эукариотические рибосомы значительно крупнее и содержат больше белков, чем прокариотические.
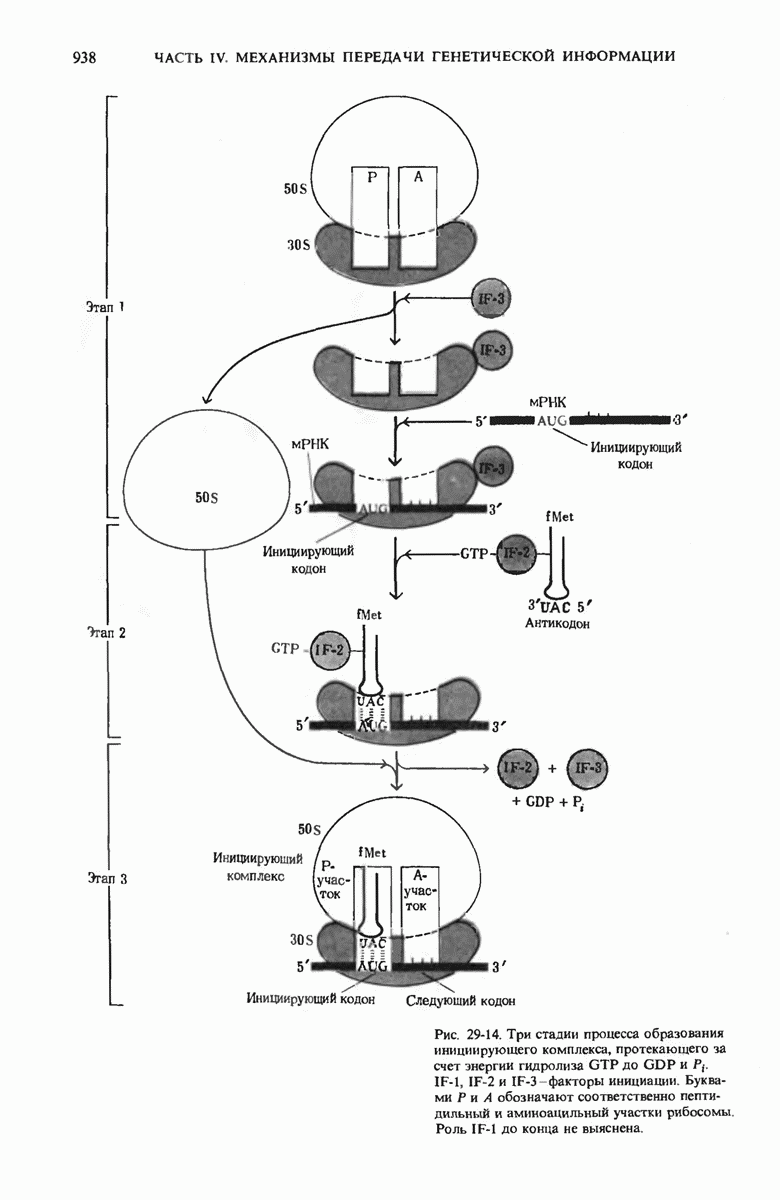
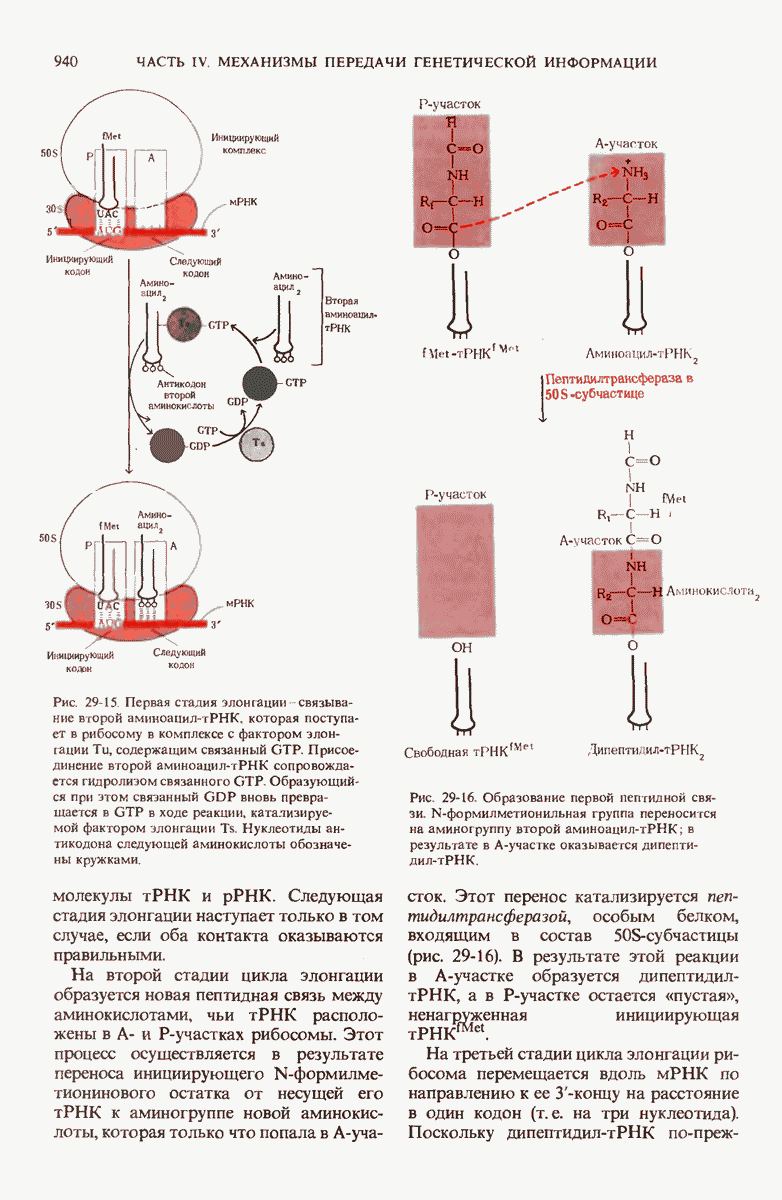
У бактерий инициирующим N-концевым остатком во всех белках служит N-формилметионил-тРНК. Она образует комплекс с фактором инициации  -рибосомной субчастицей, мРНК и GTP; этот сложный комплекс взаимодействует с 50S-субчастицей, формируя инициирующий комплекс с одновременным расщеплением GTP до GDP и отделением IF-2. На следующих за этим этапах элонгации необходимо присутствие GTP и трех факторов элонгации, участвующих в присоединении поступающей аминоацил-тРНК к аминоацилсвязывающему участку рибосомы (А-участку).
-рибосомной субчастицей, мРНК и GTP; этот сложный комплекс взаимодействует с 50S-субчастицей, формируя инициирующий комплекс с одновременным расщеплением GTP до GDP и отделением IF-2. На следующих за этим этапах элонгации необходимо присутствие GTP и трех факторов элонгации, участвующих в присоединении поступающей аминоацил-тРНК к аминоацилсвязывающему участку рибосомы (А-участку).
В ходе пептидилтрансферазной реакции остаток формилметионина переносится на аминогруппу вновь поступившей аминоацил-тРНК. Удлиненная таким образом пептидил-тРНК перемещается с аминоацильного на пепти-дильный участок рибосомы; этот процесс требует гидролиза GTP. После многократного повторения таких циклов элонгации происходит терминация полипептидной цепи, осуществляемая с помощью так называемых рилизинг-факторов. Синтез белка происходит в полирибосомах комплексах, состоящих из нескольких или большого числа рибосом, прикрепленных к молекуле мРНК; каждая из рибосом считывает мРНК и синтезирует белок независимо. На образование каждой пептидной связи расходуются по меньшей мере четыре высокоэнергетические фосфатные связи; вероятно, это необходимо для гарантии точности трансляции.
Кодоны для аминокислот представляют собой специфические тройки нуклеотидов (триплеты). Нуклеотидная последовательность в кодонах была установлена в результате экспериментов с использованием синтетических мРНК известного нуклеотидного состава и известной нуклеотидной последовательности. В аминокислотном коде почти каждой аминокислоте соответствует несколько кодовых слов. Третья буква каждого кодона гораздо менее специфична, чем первые две: про нее говорят, что она «качается». Стандартные слова генетического кода, вероятно, универсальны для всех организмов, правда в митохондриях человека найдены кодоны, значение которых отличается от универсального. Инициирующая аминокислота N-формилметионин кодируется кодоном AUG, причем для ее взаимодействия с этим кодоном необходимо наличие с 5'-стороны от AUG инициирующего сигнала с повышенным содержанием А и G. Триплеты UAA, UGA и UAG не кодируют никакую аминокислоту, они служат сигналами терминации полипептидной цепи. В некоторых вирусных ДНК одна и та же нуклеотидная последовательность может кодировать два разных белка, мРНК для которых транскрибируются с использованием разных рамок считывания.
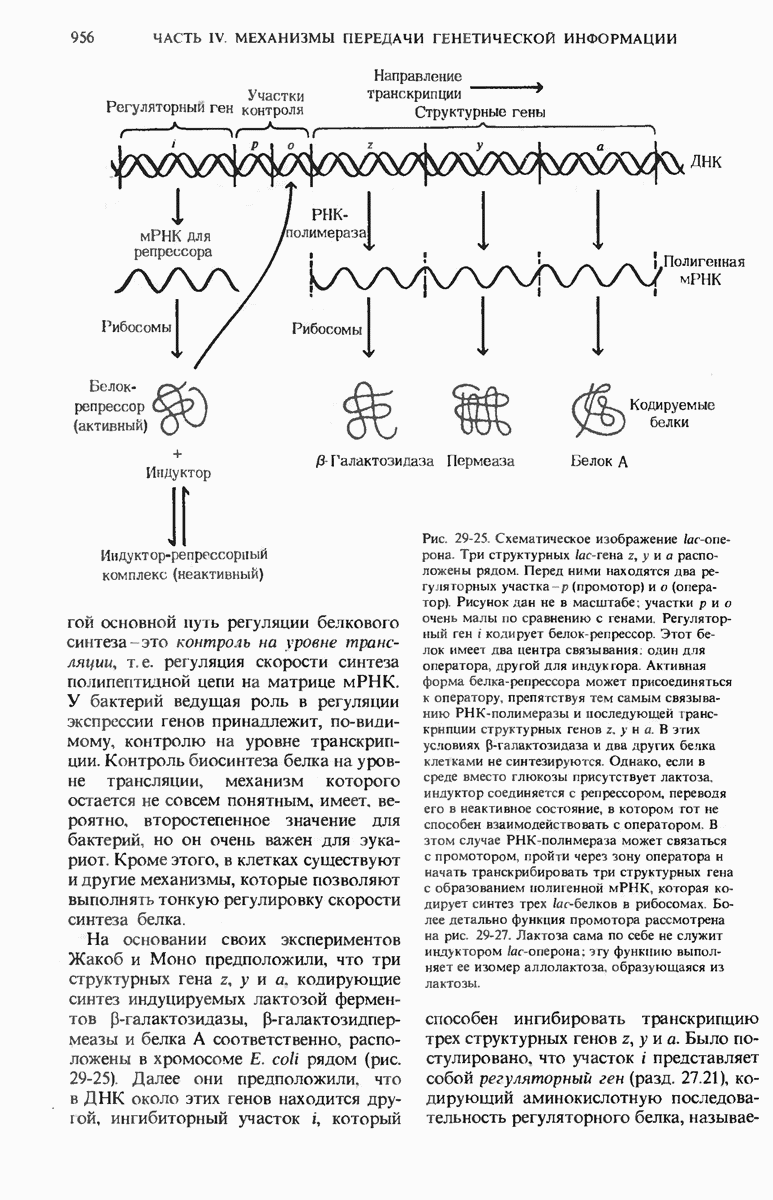
Синтез белка у прокариот регулируется главным образом на уровне транскрипции ДНК, т. е. на уровне образования мРНК. Транскрипция группы метаболически связанных между собой генов регулируется путем присоединения (или отделения) особого белка - репрессора к операторному участку ДНК. Оператор и группа связанных друг с другом генов вместе составляют оперон. Транскрипция такой группы генов может индуцироваться специфическим питательным субстратом, например лактозой. Лактоза может связывать репрессор и вызывать тем самым его отделение от оператора. Благодаря этому разрешается транскрипция генов, кодирующих белки, необходимые клетке для использования лактозы в качестве источника углерода и энергии. Некоторые опероны имеют также промоторный участок, содержащий регуляторную часть - так называемый САР-участок; последний предназначен для связывания комплекса, образованного белком, активируюшим катаболитный ген (САР), и сАМР. Этот комплекс, формирующийся при отсутствии в среде глюкозы, дает возможность РНК-полимеразе присоединиться к месту инициации транскрипции генов, ответственных за катаболизм лактозы.
ЛИТЕРАТУРА
Активация аминокислот
Rich A., Kim S. Н. The Three-Dimensional Structure of Transfer RNA, Sci. Am., 238, 52-62, January 1978.
Schimmel P. R. Understanding the Recognition of Transfer RNAs by Aminoacyl Transfer Synthetases, Adv. Enzymol., 49, 187-222 (1979).
Рибосомы
Nomura M. Assembly of Bacterial Ribosomes, Science, 179, 864-873 (1973).
Wittman H.G. Structure and Function of E. coli Ribosomes, Fed. Proc., 36, 2025-2080 (1977).
Этапы инициации, элонгации и терминации белкового синтеза
Weissbach Н„ Pestka S. (eds.). Molecular Mechanisms of Protein Biosynthesis, Academic, New York, 1977. Обзорные статьи по различным вопросам белкового синтеза.
Генетический код
Barrett В. G., Air G. М., Hutchinson С. А., III. Overlapping Genes in Bacteriophage фХ174, Nature, 264, 34-40 (1976).
Crick F. H. C. The Genetic Code III, Sci. Am., 215, 55-62, October 1966.
Fiddes J. C. The Nucleotide Sequence of a Viral DNA, Sci. Am., 237, 54-67, December 1977.
Hall B. D. Mitochondria Spring Surprises, Nature, 282, 129-130 (1979). Исключения из принципа универсальности генетического кода.
Nirenberg М. The Genetic Code II, Sci. Am., 208, 80-94, March 1963. Описание первых экспериментов по расшифровке генетического кода.
Регуляция экспрессии генов
Brown D.D. Gene Expression in Eukaryotes, Science, 211, 667-674 (1981).
Lewin B. Gene Regulation II, 2nd ed., Wiley, New York, 1980. Книга содержит большое количество информации по проблеме регуляции у эукариот.
Maniatis Т., Ptashne М. A DNA Operator-Repressor System, Sci. Am., 234, 64-76, January 1976.
O'Malley B.W. et al. The Ovalbumin Gene: Organization, Structure, Transcription, and Regulation, Recent Progr. Horm. Res., 35, 1-42 (1979).
Pastan I. Cyclic AMP, Sci. Am., 227, 97-105, August 1972.
Специальные вопросы
Palade G. Intracellular Aspects of the Process of Protein Synthesis, Science, 189, 347-357 (1975).
Yarus M. Accuracy of Translation, Progr. Nucleic Acid Res., 23, 195-225 (1979).