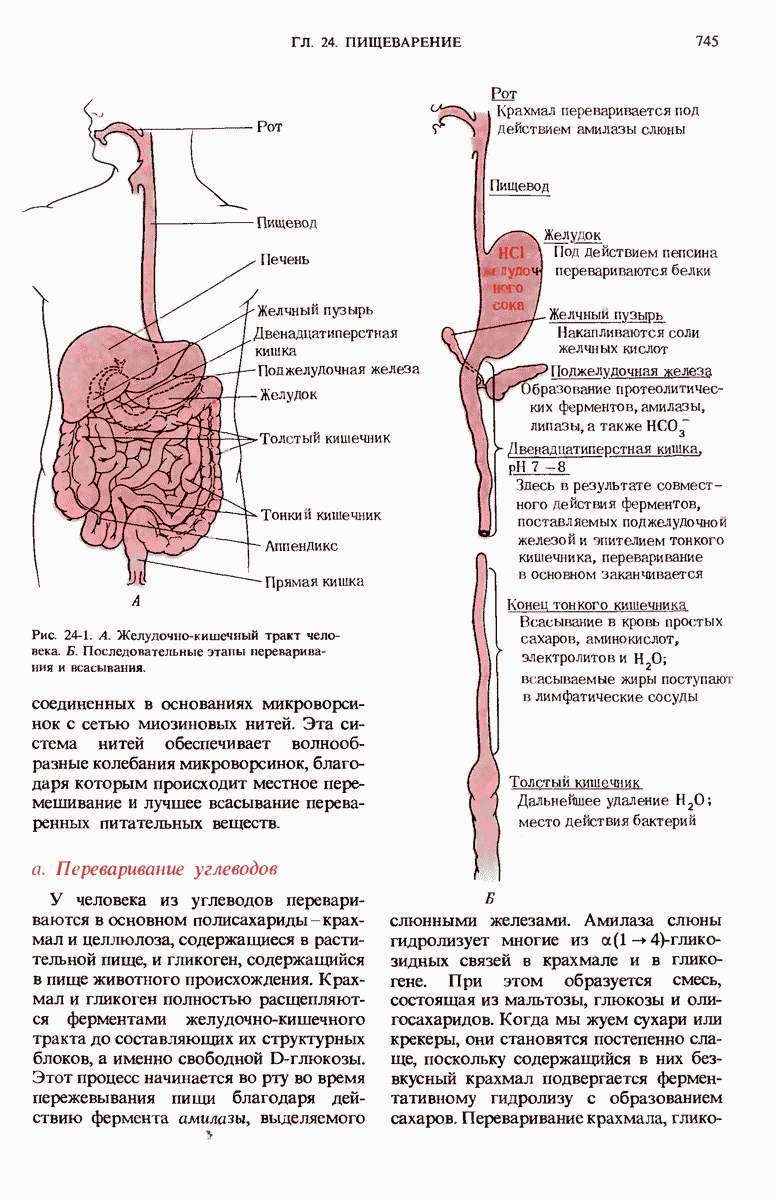
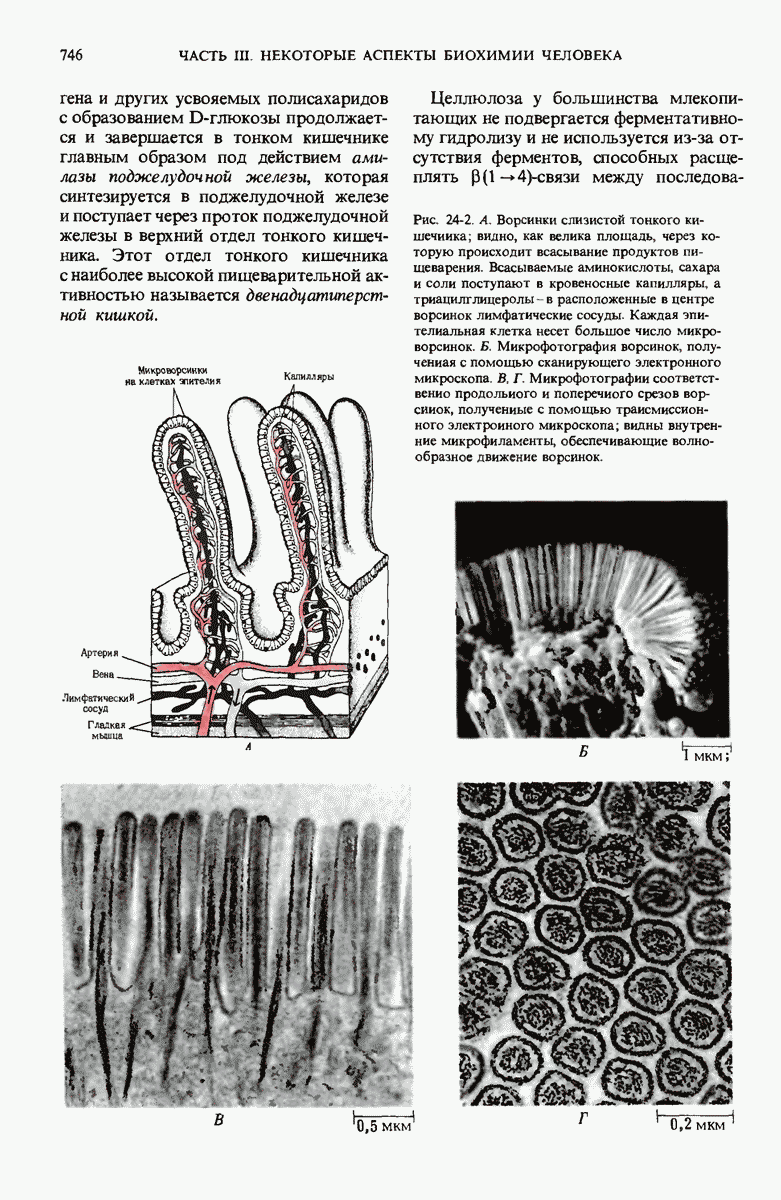
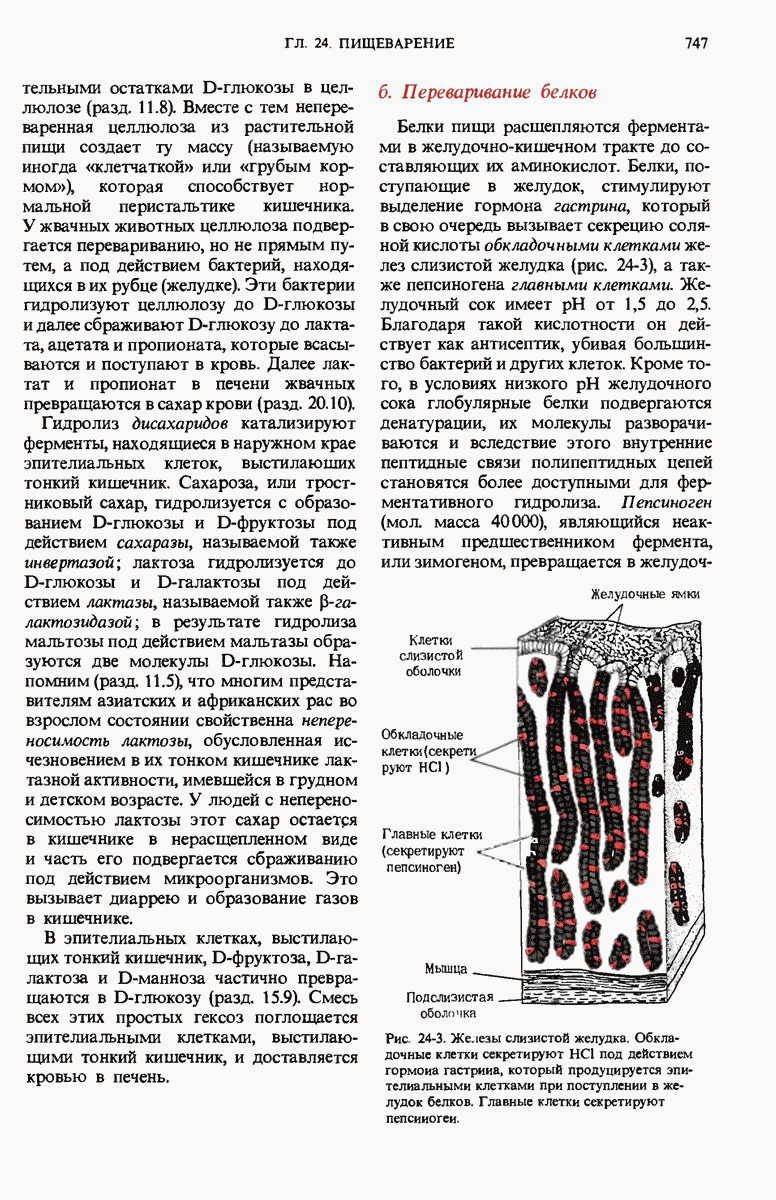
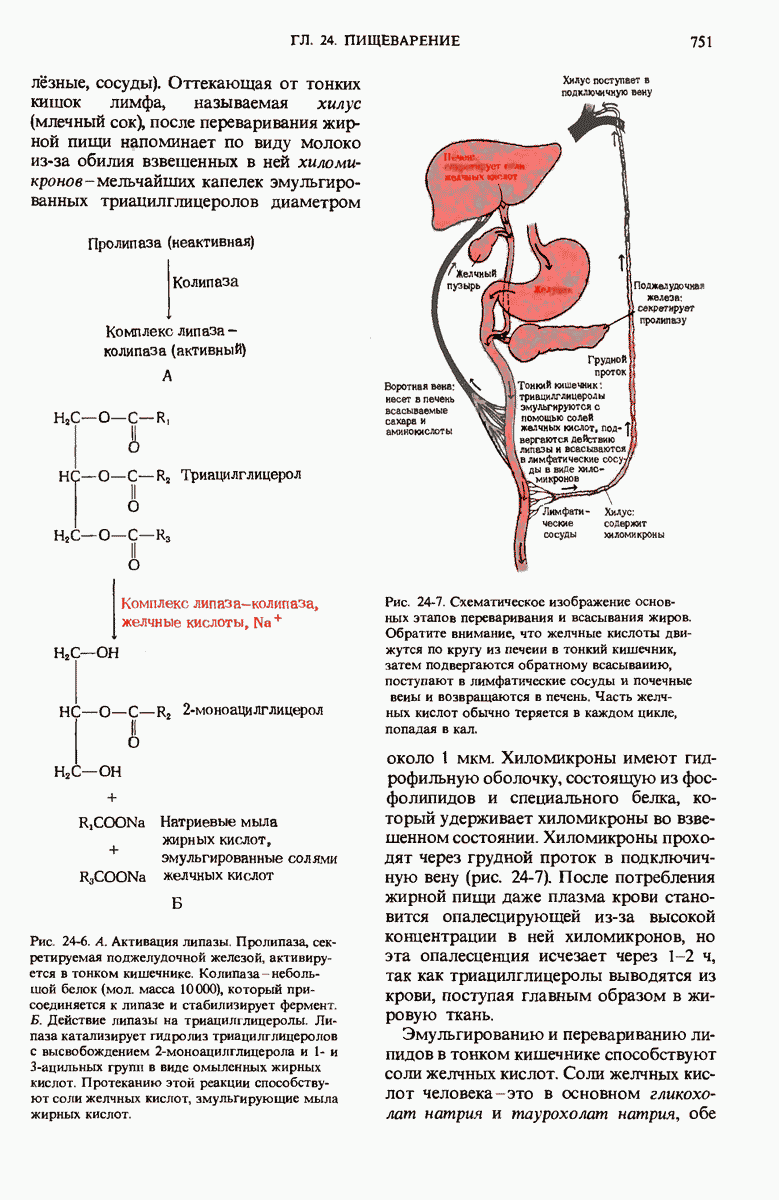
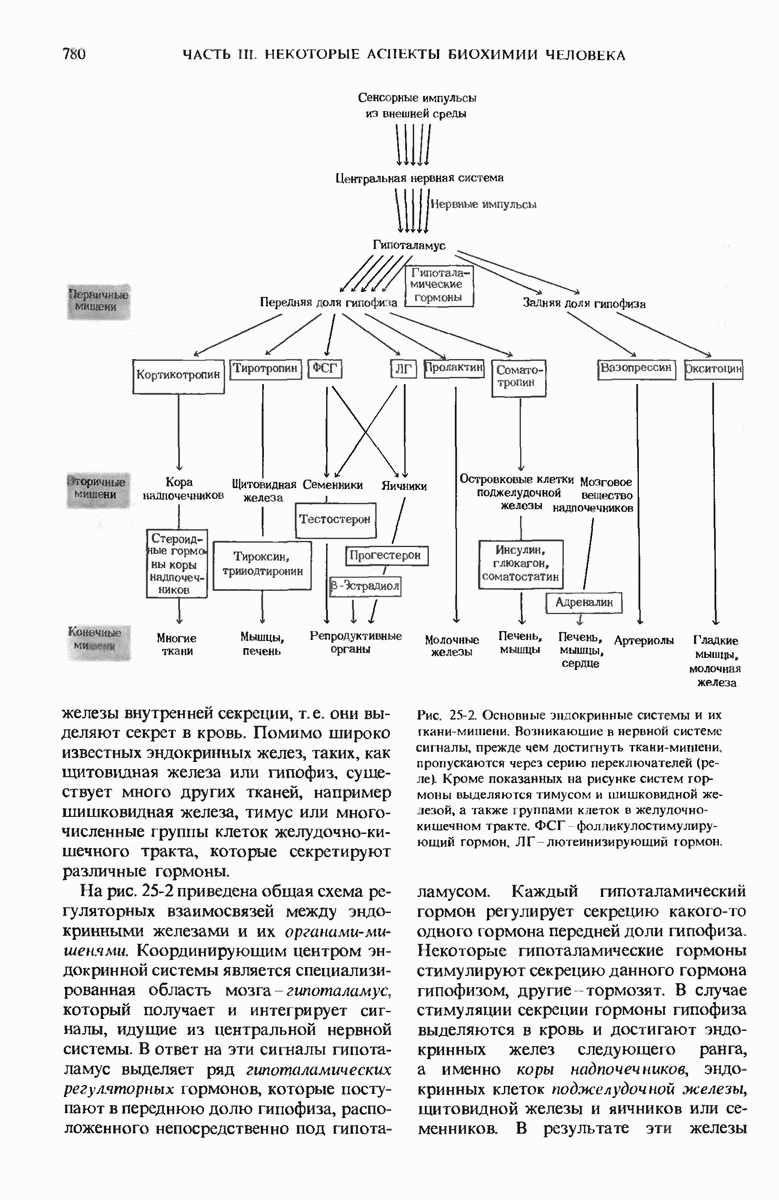
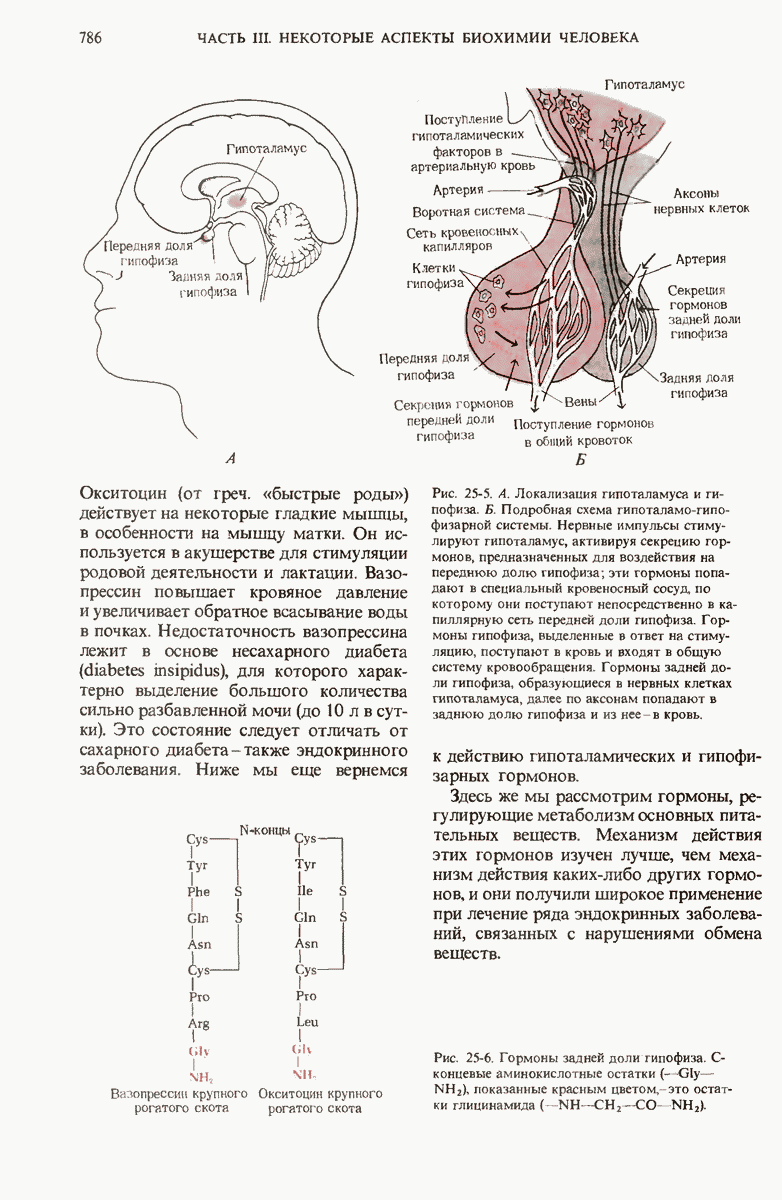
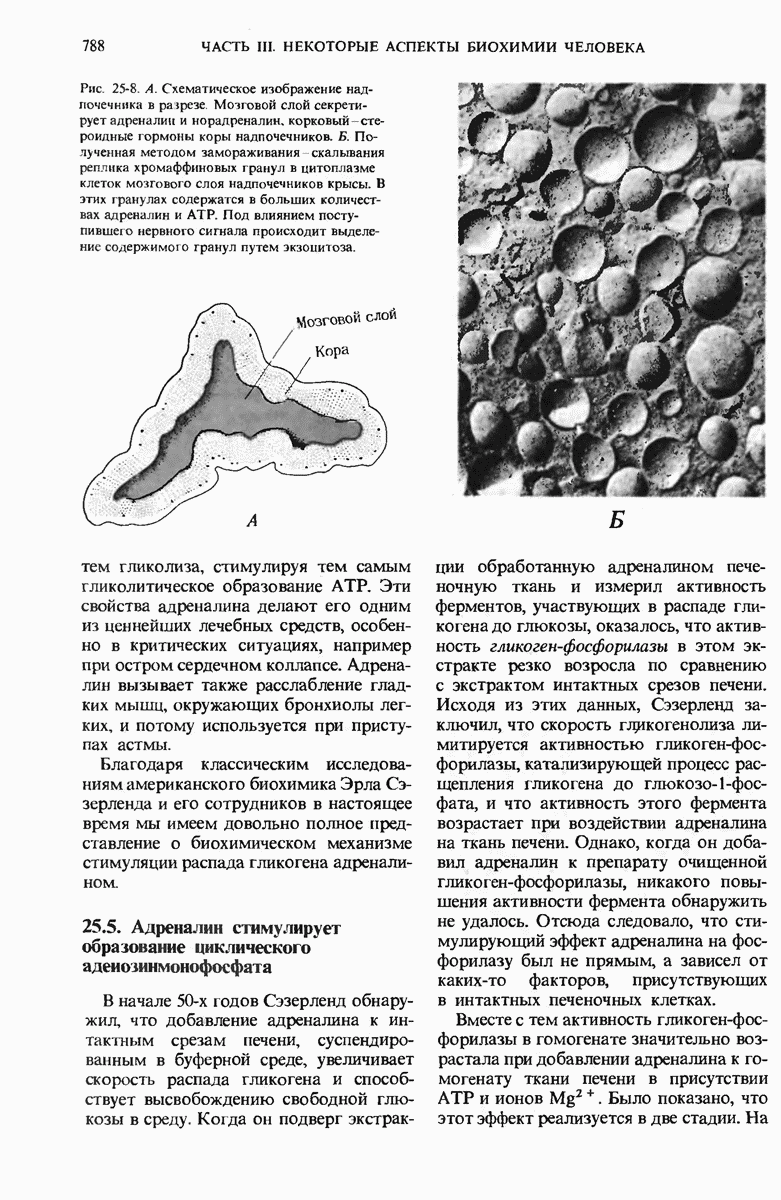
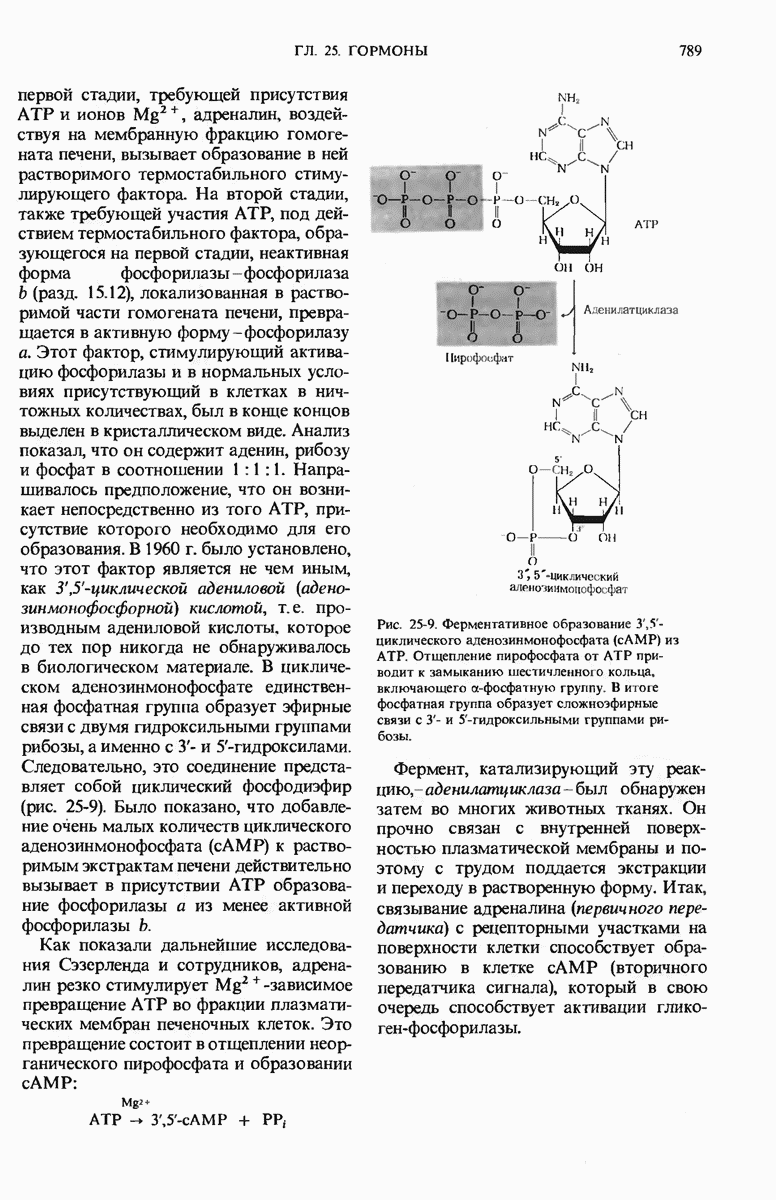
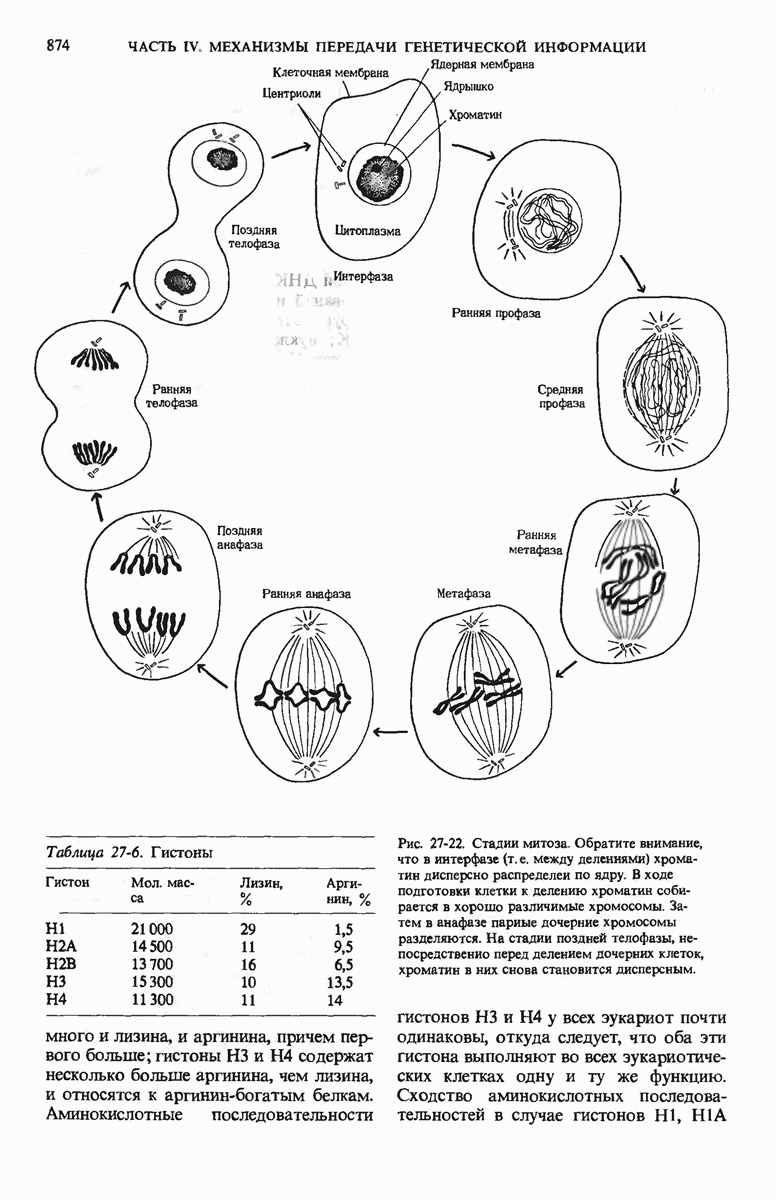
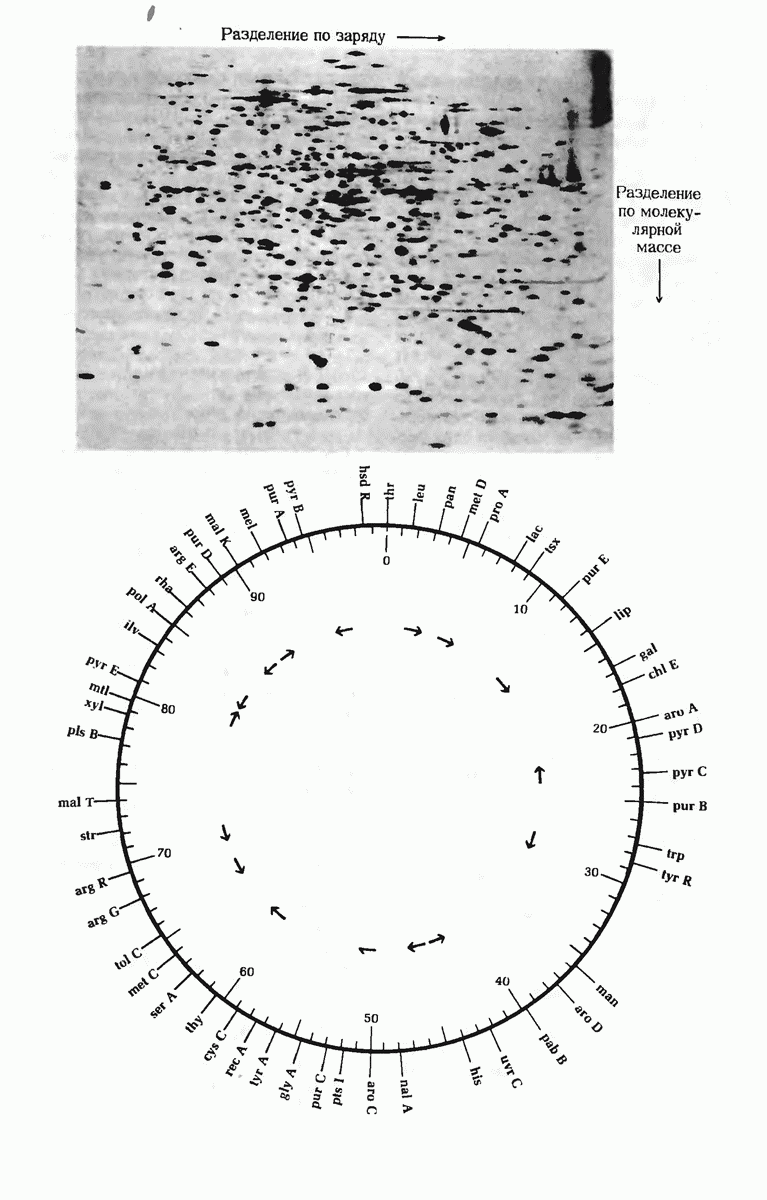
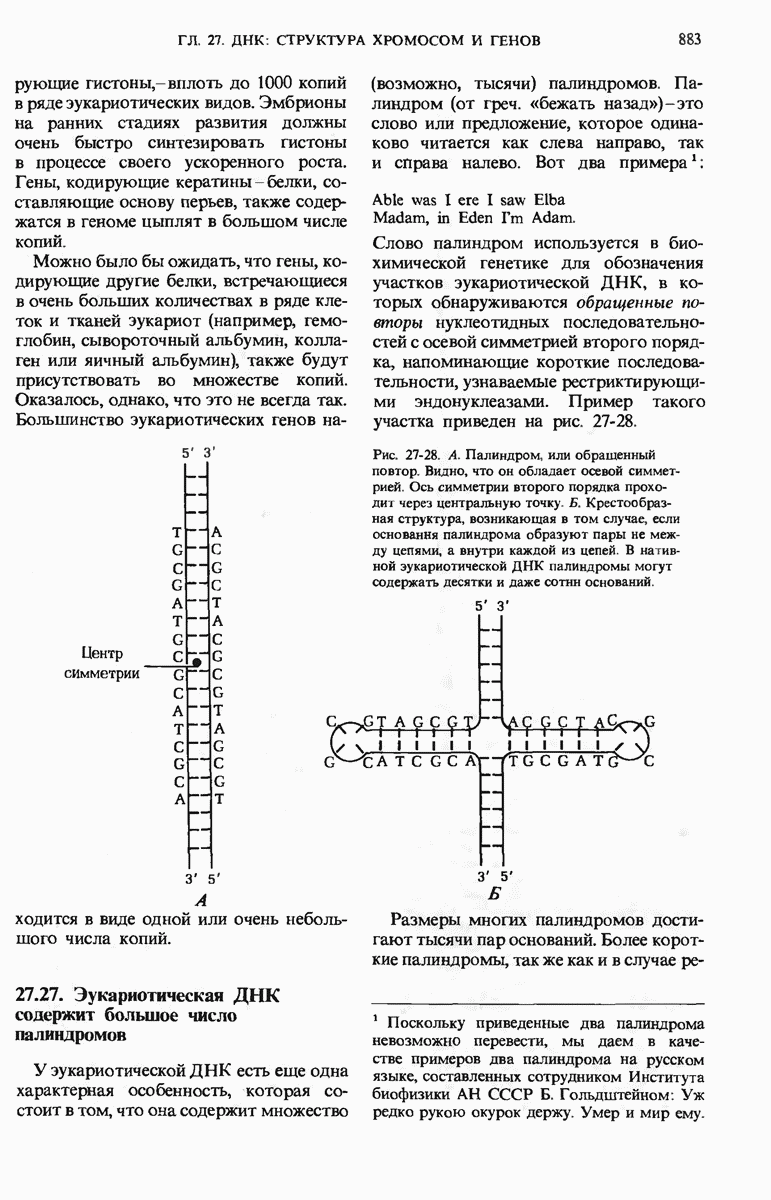
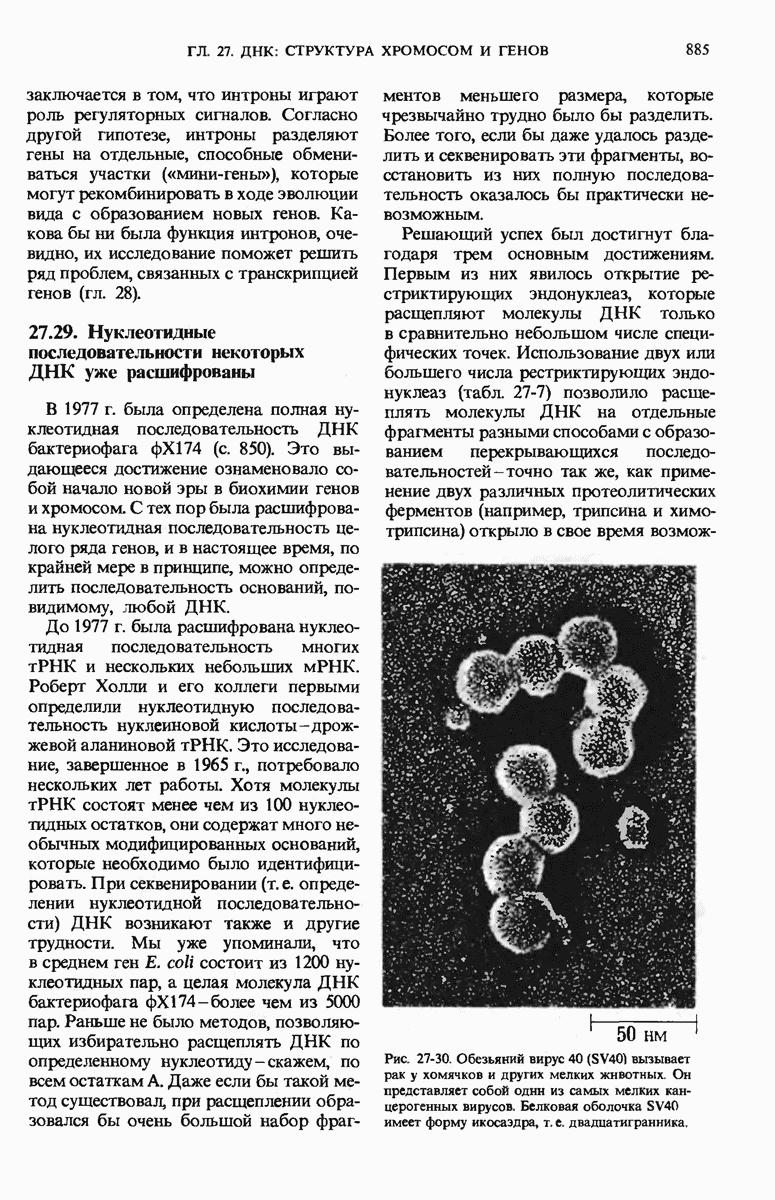
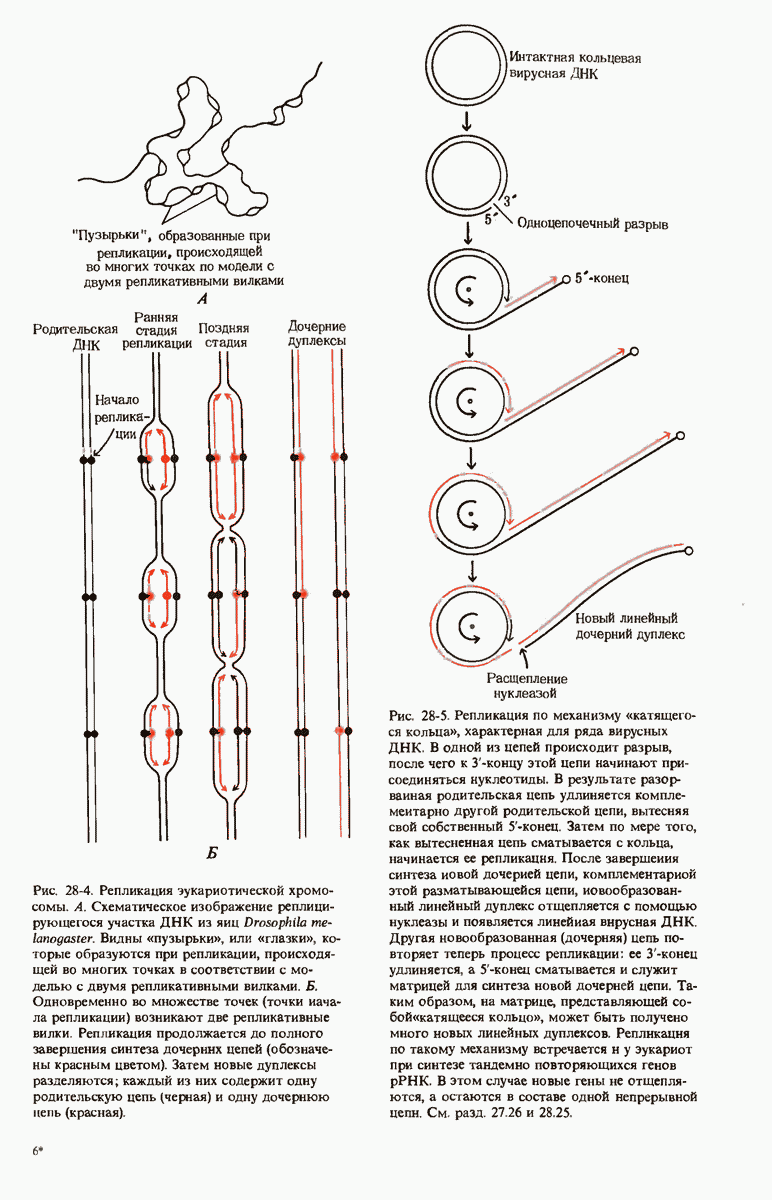
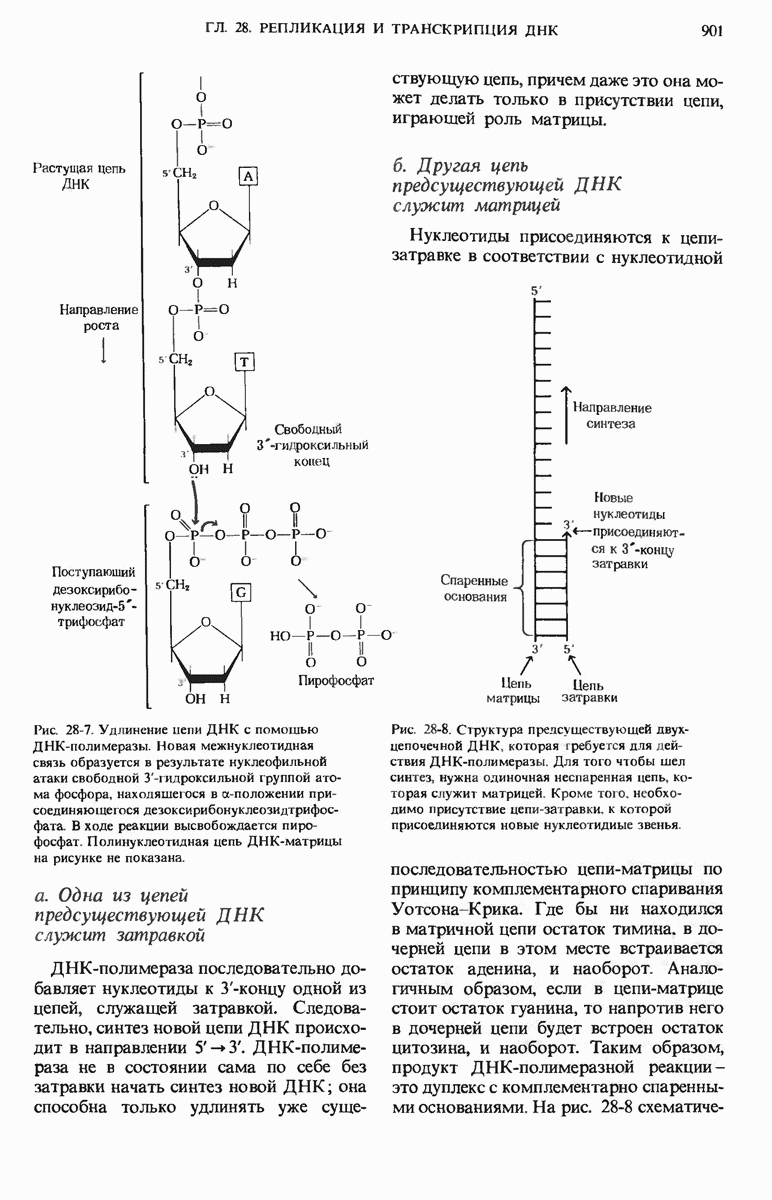
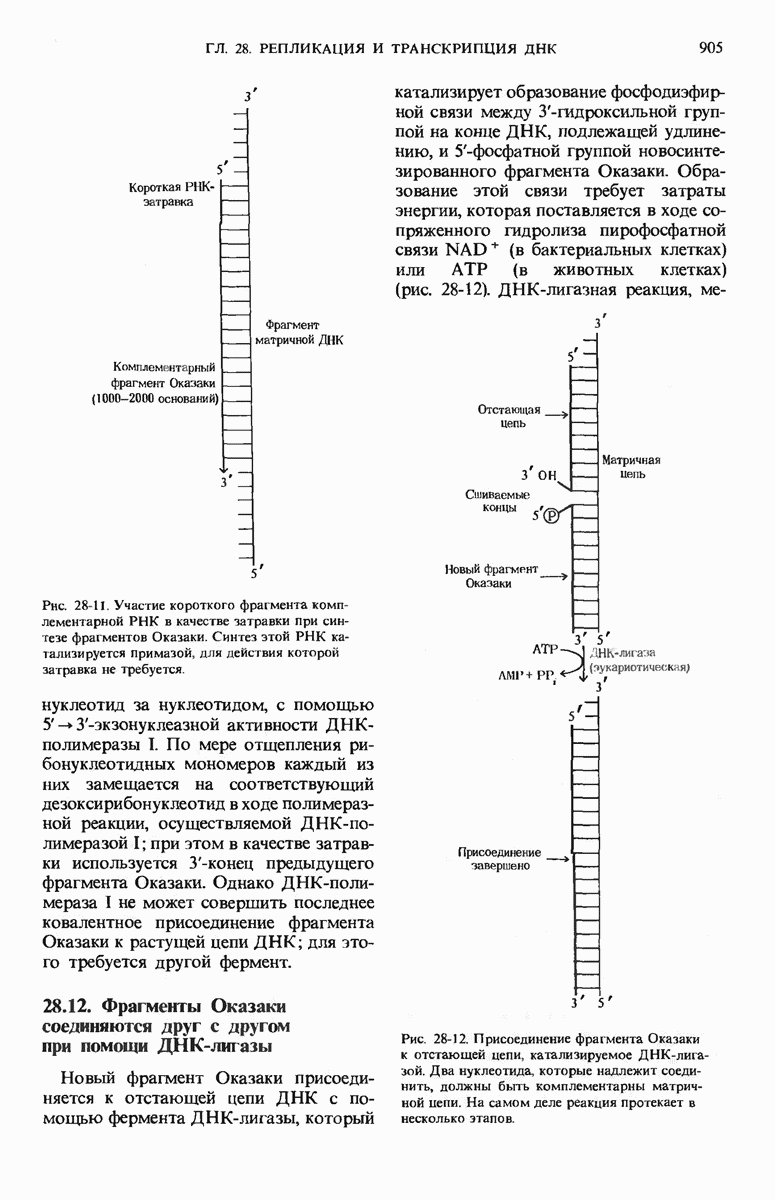
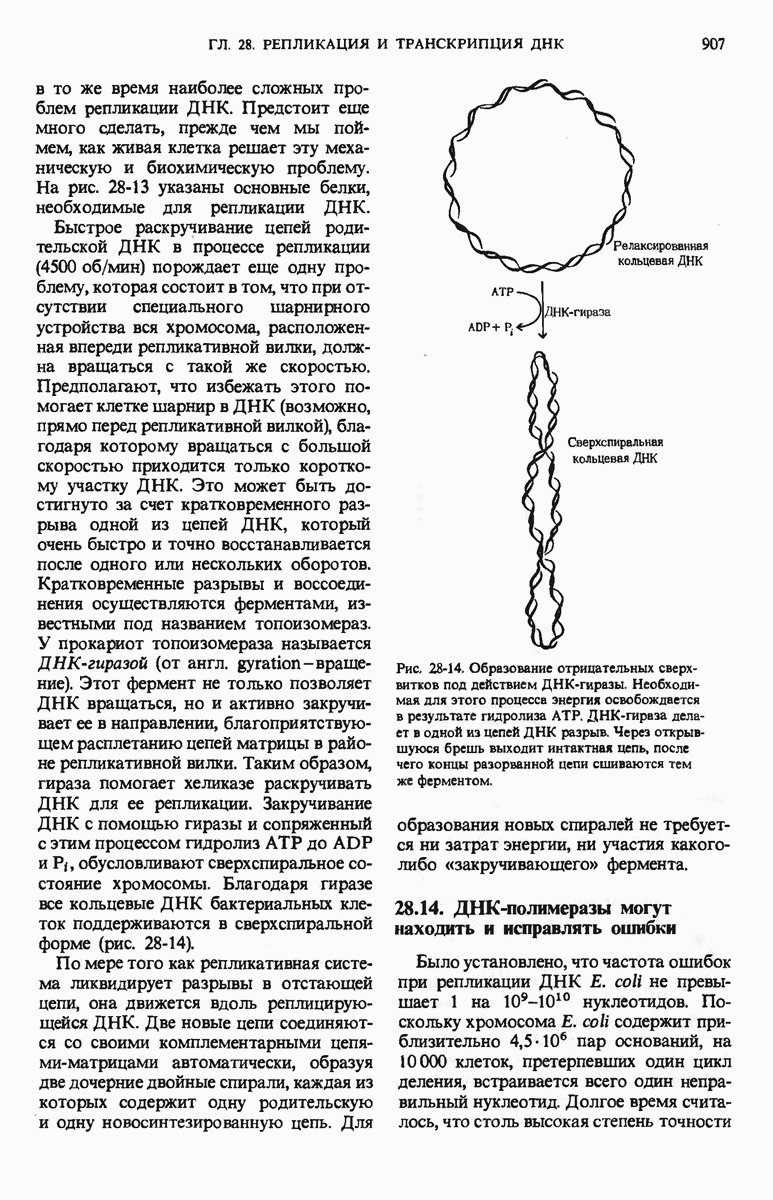
Краткое содержание главы
Как в прокариотических, так и в эукариотических клетках содержатся ферментные системы, способные исправлять ошибки репликации и различные формы повреждения ДНК, вызываемые гидролизом или действием внешних мутагенов - ультрафиолетового излучения, ионизирующей радиации, а также дезаминирующих и алкилирующих агентов.
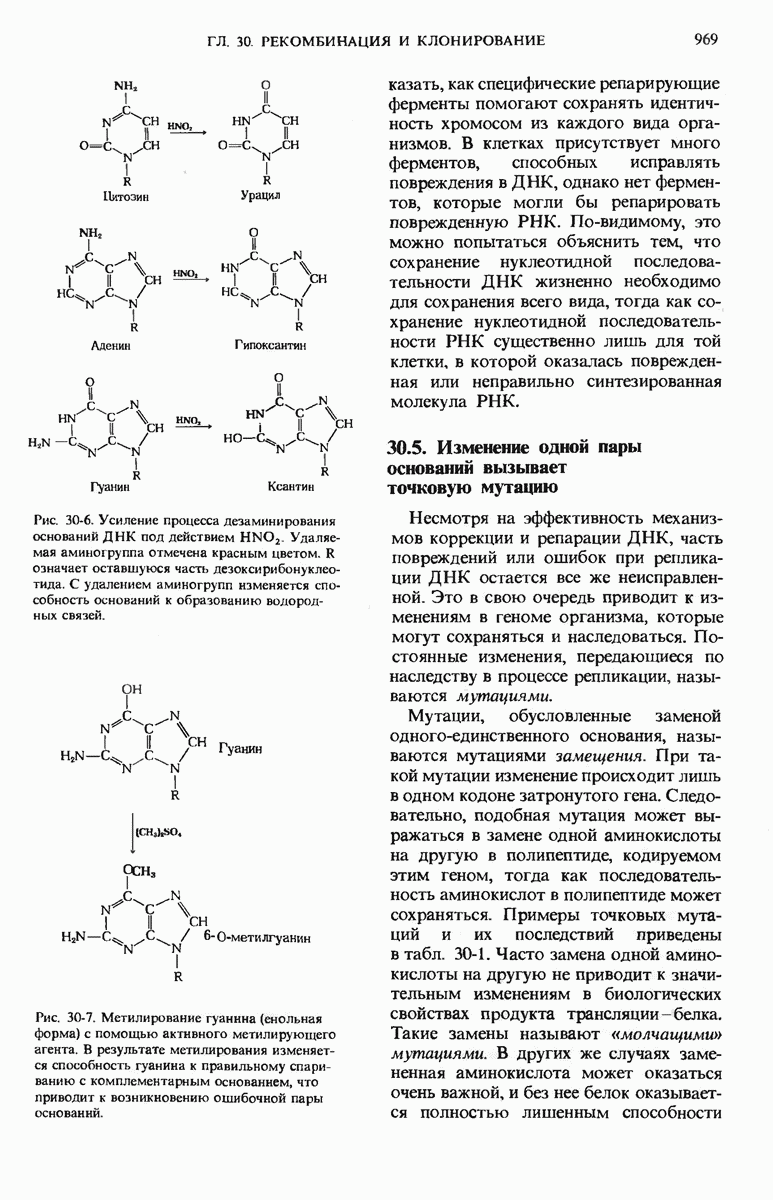
Неисправленные с помощью механизма реперации повреждения ДНК приводят к наследуемым мутациям, которые могут быть летальными, неполностью подавляющими функцию, «молчащими» или даже благоприятными в зависимости от места и природы повреждения. Один из типов мутаций обусловлен встраиванием в ДНК неправильного основания, что приводит к изменению одного кодона и в большинстве случаев к замене одной аминокислоты в белковом продукте. Другой важный тип мутаций - это так называемые мутации со сдвигом рамки, которые представляют собой потерю или приобретение одного или нескольких нуклеотидов. Такие мутации вызывают сдвиг рамки считывания кодонов, в результате чего образуется белковый продукт, аминокислотная последовательность которого искажена, начиная с того места, где произошло выпадение или добавление нуклеотида. Мутации обычно возникают беспорядочно, однако вероятность их появления сильно возрастает под действием мутагенов, к которым относятся дезаминирующие и алкилирующие агенты, а также другие вещества, способные изменять структуру оснований. Почти все канцерогенные соединения являются мутагенами, что может быть проверено с помощью простых тестов, основанных на выращивании бактерий.
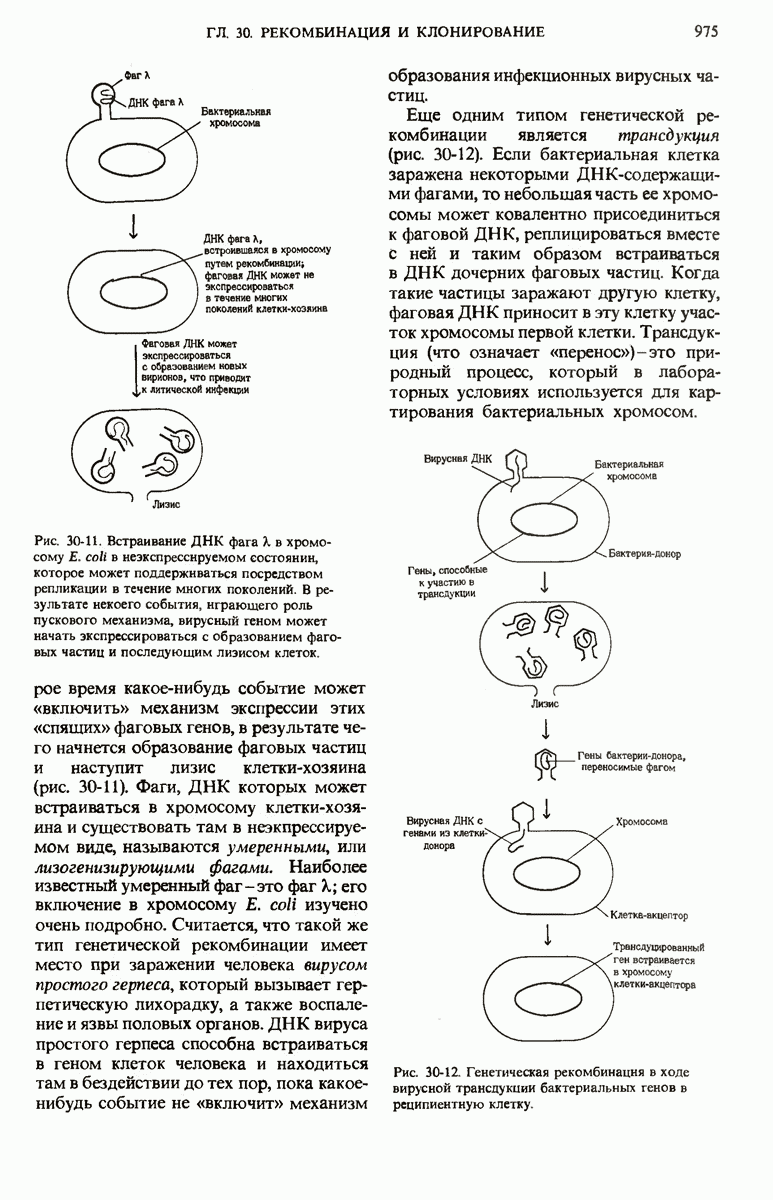
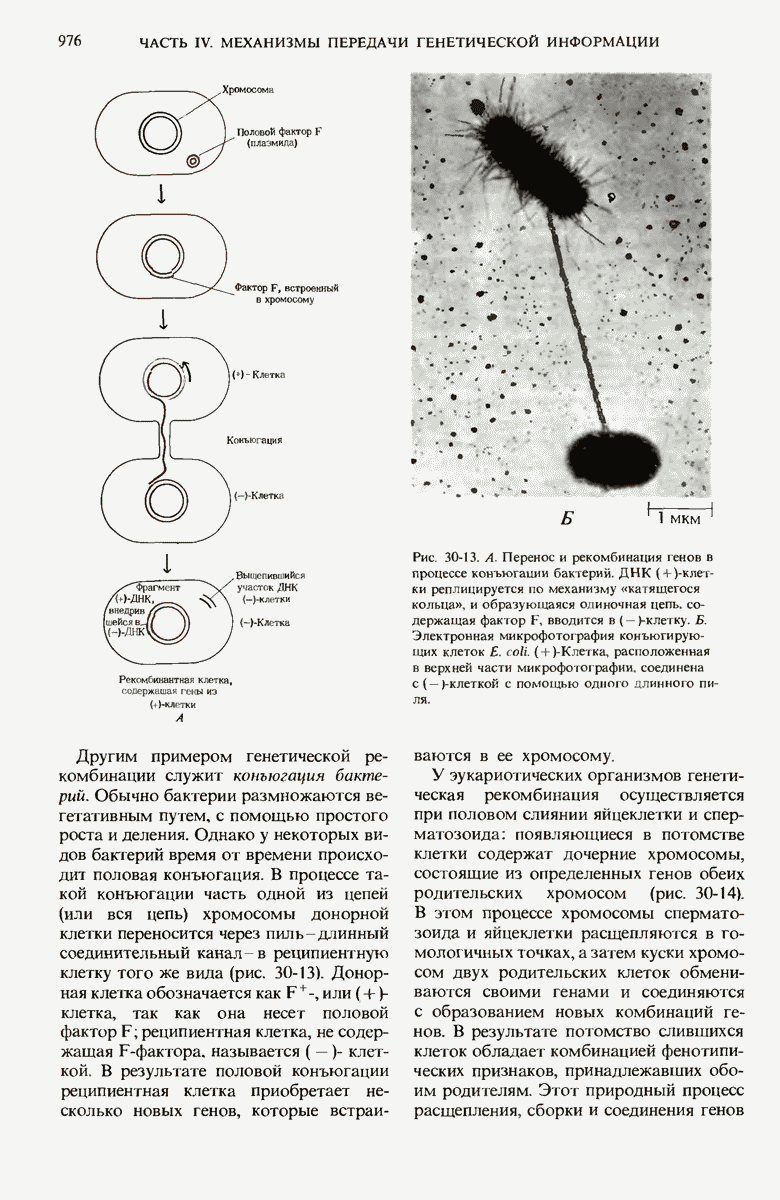
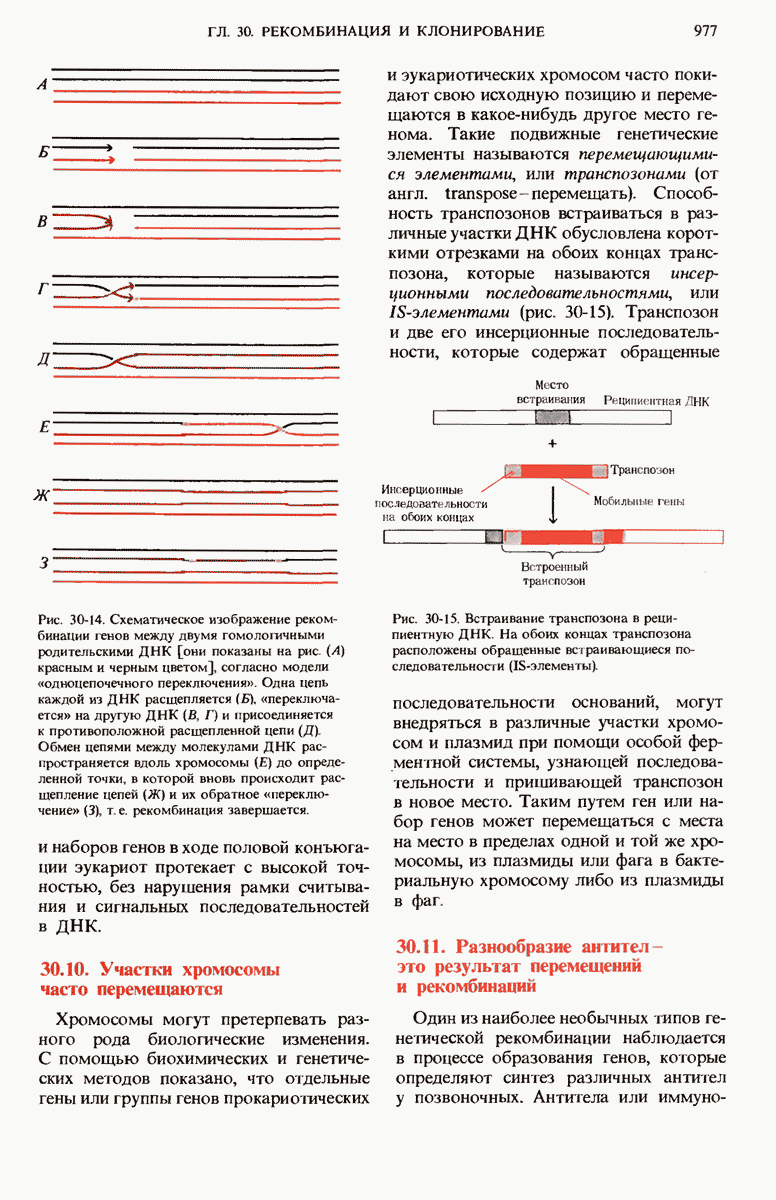
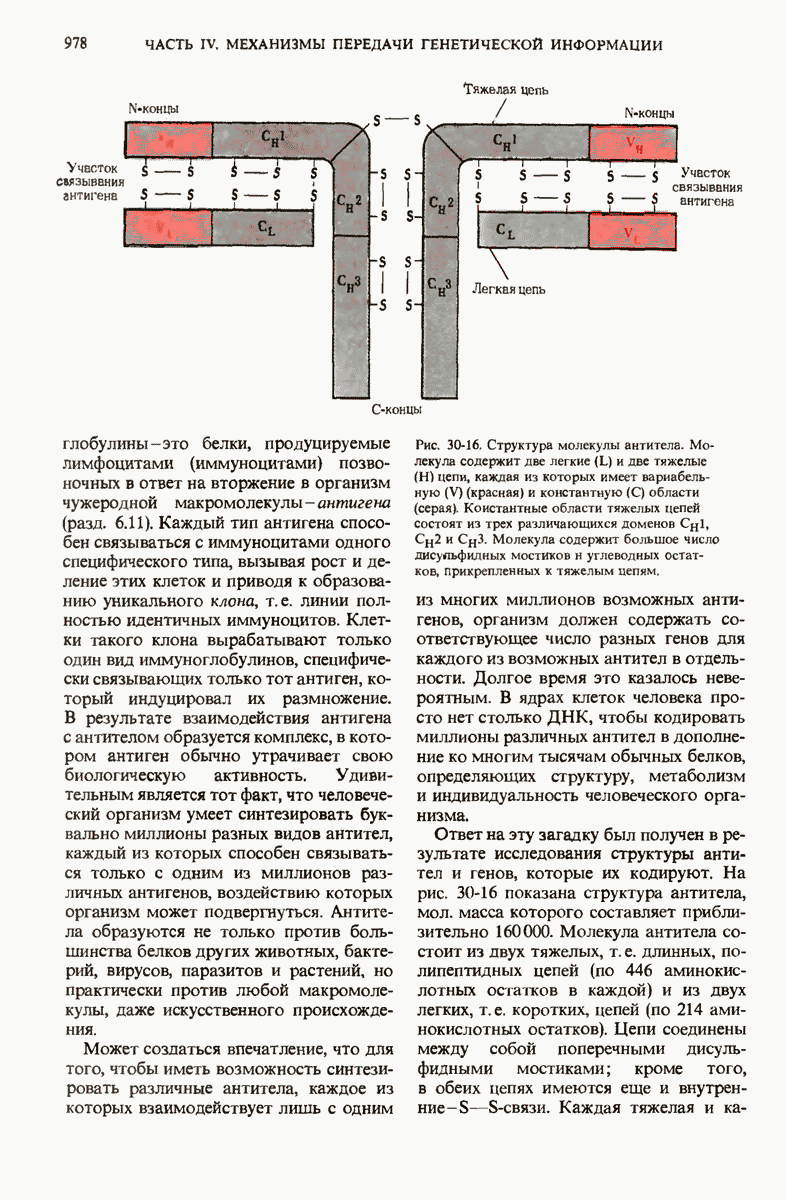
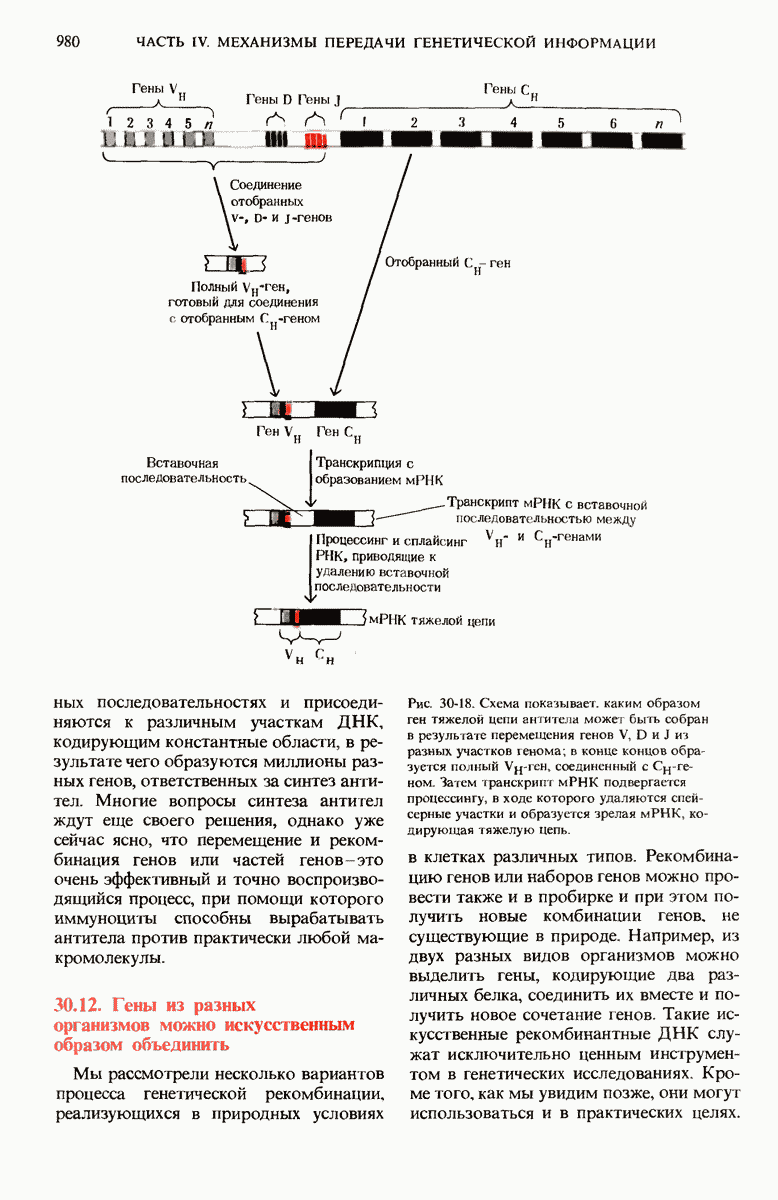
В нормальных природных условиях гены и наборы генов претерпевают рекомбинацию в ходе таких биологических процессов, как трансформация бактерий, вирусная трансдукция, конъюгация бактерий и обмен генов при слиянии половых эукариотических клеток. Гены и группы генов могут также перемещаться с одного места на другое как в пределах одной и той же хромосомы, так и между разными хромосомами. Например, белки-антитела, которые вырабатываются клетками крови (иммуноцитами) позвоночных против миллионов самых разных, не содержащихся в организме макромолекул (антигенов), кодируются последовательностями ДНК, возникающими в результате перемещения и перегруппировки сравнительно небольшого числа разных типов генов. При транскрипции с таких последовательностей могут синтезироваться самые разные мРНК для миллионов разных антител, каждый из которых способен специфически взаимодействовать с одним и только одним типом молекул антигена.
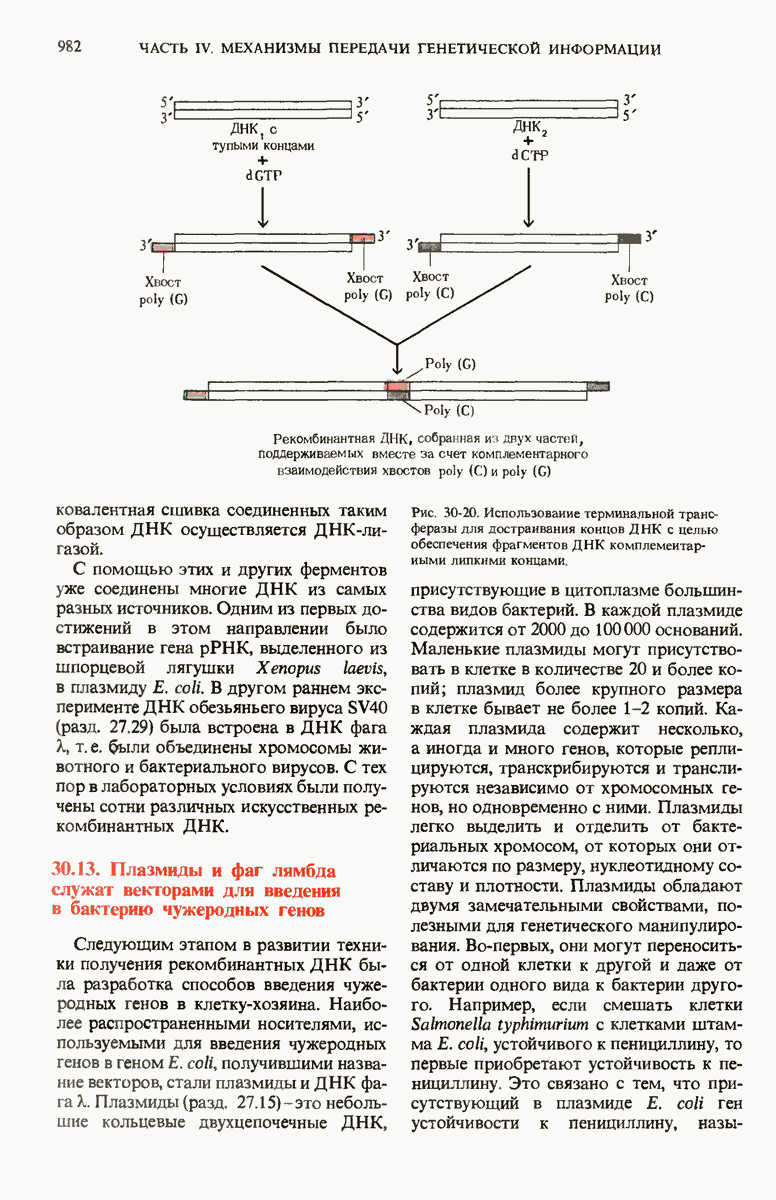
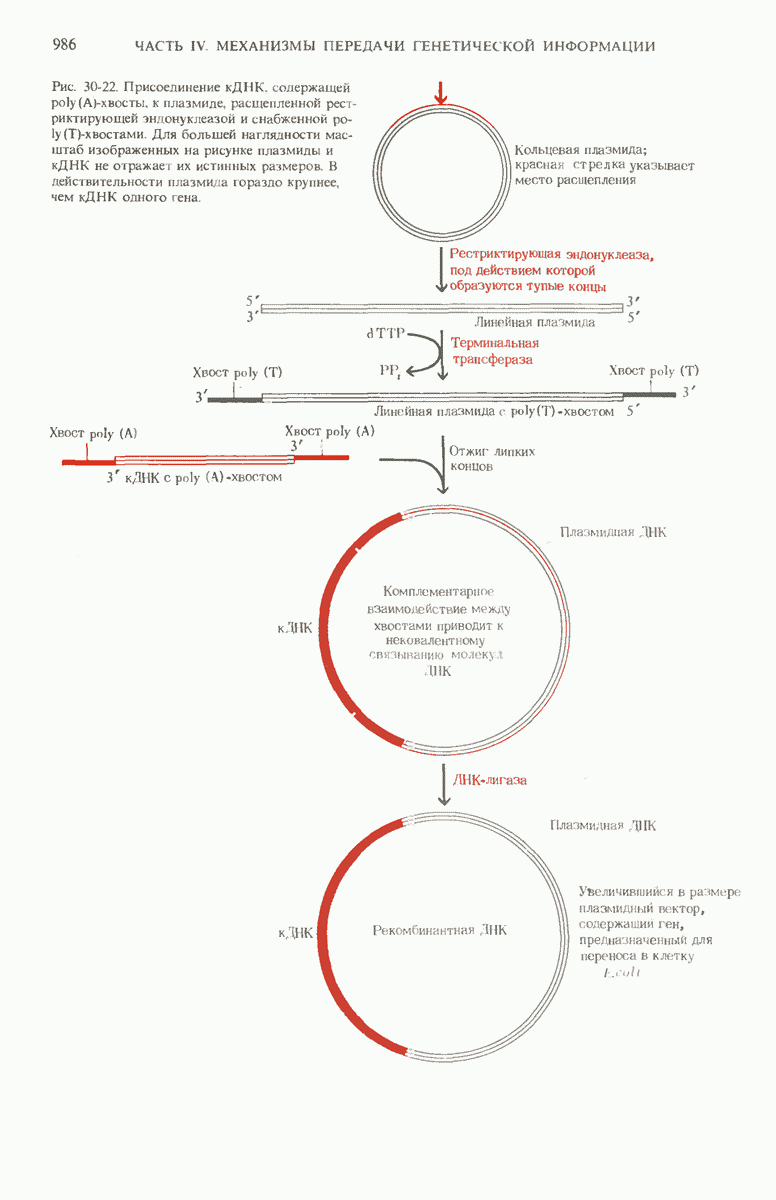
Новые комбинации генов можно создать и искусственным путем в лабораторных условиях с помощью таких ферментов, как рестриктирующие эндонуклеазы, ДНК-лигаза и терминальная трансфераза. Чтобы встроить чужеродный ген в геном клеток Е. coli, этот ген сначала пришивают к вектору, которым служит либо плазмида Е. coli, либо ДНК фага X. Полученная рекомбинантная ДНК может затем попасть в клетку Е. coli, ковалентно включиться в ее хромосому и в составе этой хромосомы реплицироваться. Если новый ген, содержащийся в рекомбинантной ДНК, обладает подходящими сигнальными последовательностями, указывающими начало и конец транскрипции, то такой ген будет транскрибироваться и транслироваться с образованием соответствующего белкового продукта. Многие гены из животных клеток уже были введены в бактерии, а бактериальные гены в свою очередь были встроены в эукариотические клетки. С помощью бактерий, в геномы которых введены соответствующие гены, можно получать применяемые в медицине белковые препараты-инсулин, гормон роста и интерфероны.
ЛИТЕРАТУРА
Репарация, мутации и канцерогены
Ames В. N. Identifying Environmental Chemicals Causing Mutations and Cancer, Science, 204, 587-593 (1979).
Cairns J. The Cancer Problem, Sci. Am., 233, 64-78, November 1975. Вдумчивый анализ проблемы, вделанный молекулярным биологом.
Devoret R. Bacterial Tests for Potential Carcinogens, Sci. Am., 241,40-49, August 1979.
Drake J. M. The Molecular Basis of Mutation, Holden-Day, San Francisco, 1970.
Hanawalt P. et al. DNA Repair in Bacteria and Mammalian Cells, Annu. Rev. Biochem., 48, 783-836 (1979). Подробный обзор современного состояния проблемы.
Рекомбинация в природе
Ayala F. J., Kiger J. A., Jr. Modern Genetics, Benjamin/Cummings, Menlo Park, Calif., 1980. Дано описание бактериальной трансформации, вирусной трансдукции и бактериальной конъюгации.
Campbell А. М. How Viruses Insert Their DNA into the DNA of the Host Cell, Sci. Am., 235, 102-113, December 1976.
Cohen S.N., Shapiro J.A. Transposable Genetic Elements, Sci. Am., 242, 40-49, February 1980.
Антитела
Hood L. E„ Weissman I. L., Wood W. B. Immunology, Benjamin/Cummings, Menlo Park, Calif., 1978.
Milstein C. Monoclonal Antibodies, Sci. Am., 243, 66-74, October 1980. Описана техника получения антител одного вида в больших количествах в культуре животных клеток.
Клонирование ДНК и генетическая инженерия
Abelson J., Butz Е. (eds). Recombinant DNA, Science, vol. 209 (1980). Этот том целиком состоит из важных научно-исследователь-ских статей и содержит ценный словарь терминов.
Anderson W. В., Diacumakos Е. G. Genetic Engineering in Mammalian Cells, Sci. Am., 245, 106-121, July 1981.
Derobertis E. М., Gurdon J. B. Gene Transplantation and the Analysis of Development, Sci. Am., 241, 74-82, December 1979.
Grobstein, Clifford. The Recombinant-DNA Debate, Sci. Am., 237, 22-33, July 1977.
Industrial Microbiology and the Advent of Genetic Engineering, Freeman, San Francisco, 1981. Сборник статей, появившихся в сентябрьском номере журнала Scientific American, который посвящен генетической инженерии и использованию микроорганизмов в промышленности; особое внимание уделено генетическим изменениям и клонированию.
Setlow J. К., Hollaender A. (eds.). Genetic Engineering: Principles and Methods, Plenum, New York, 1979. Сборник статей, написанных специалистами.
Watson J.D., Tooze J. The DNA Story: A Documentary History of Gene Cloning, Freeman, San Francisco, 1981.
Интерфероны
Derynck R., Content J., Declercq E„ Volckaert G., Tavemier J.. Devos R., Fiers W. Isolation and Structure of a Human Fibroblast Interferon Gene, Nature, 285, 542-547 (1980).