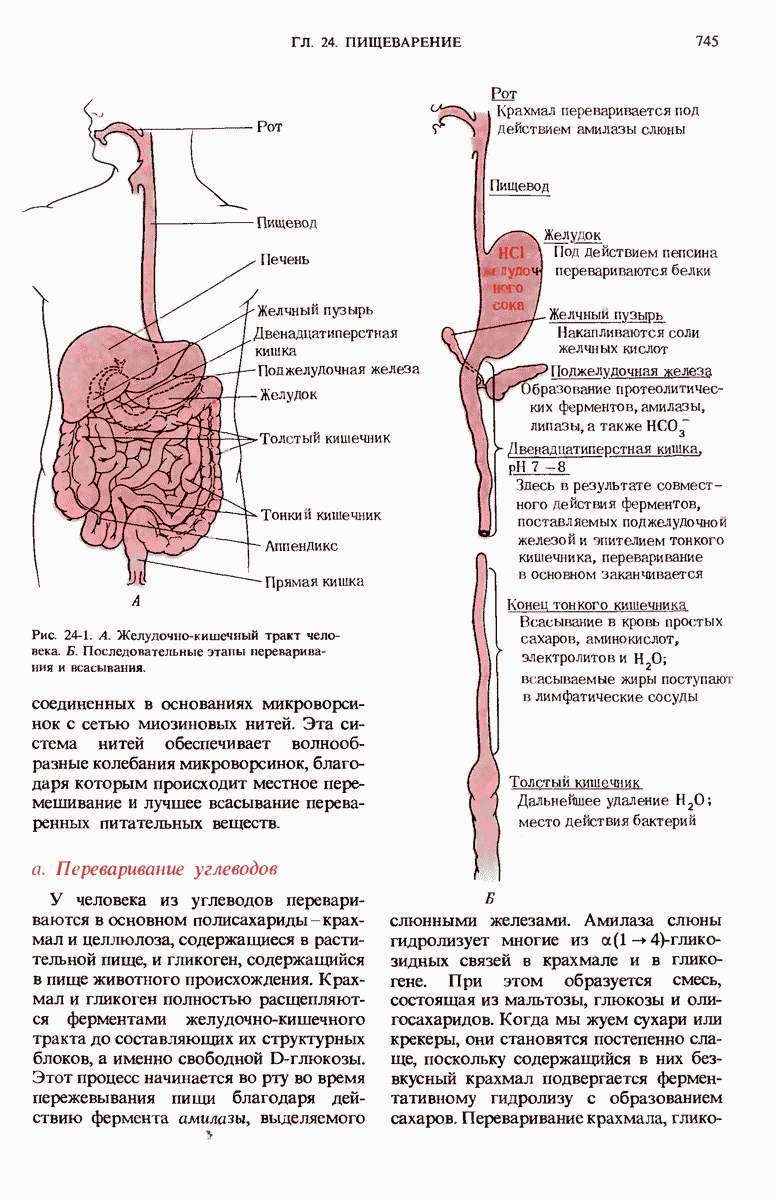
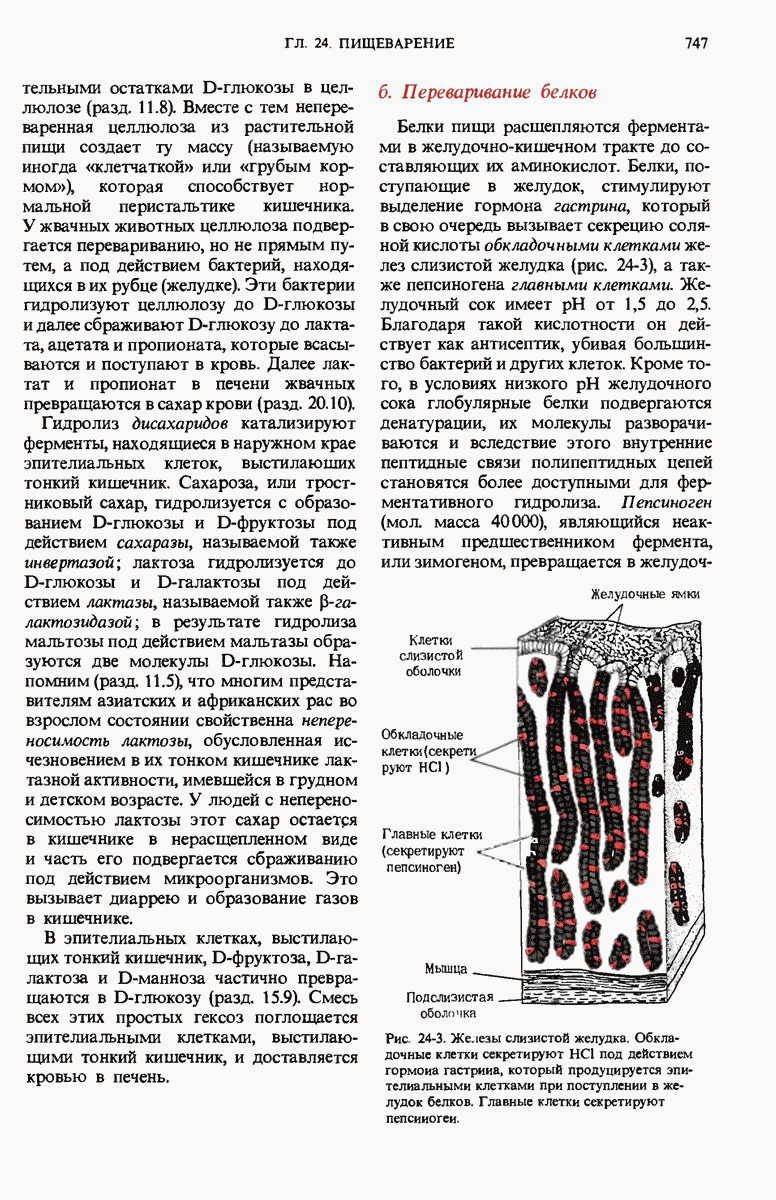
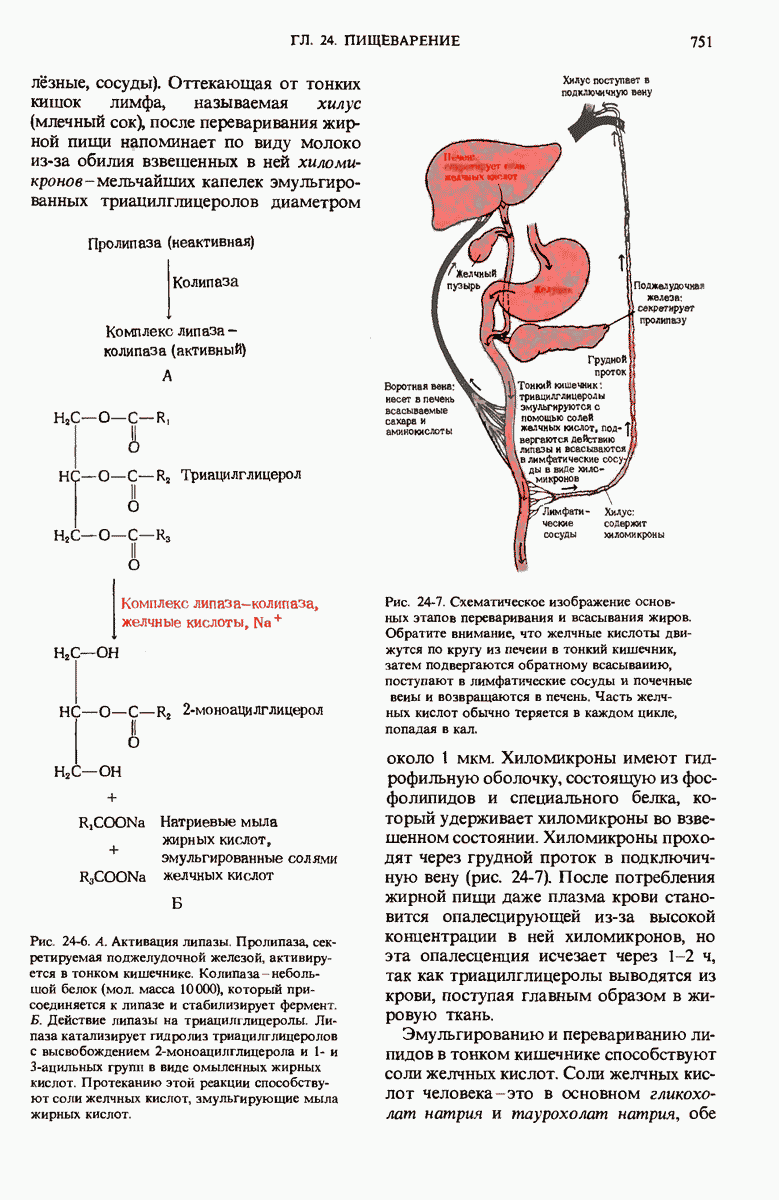
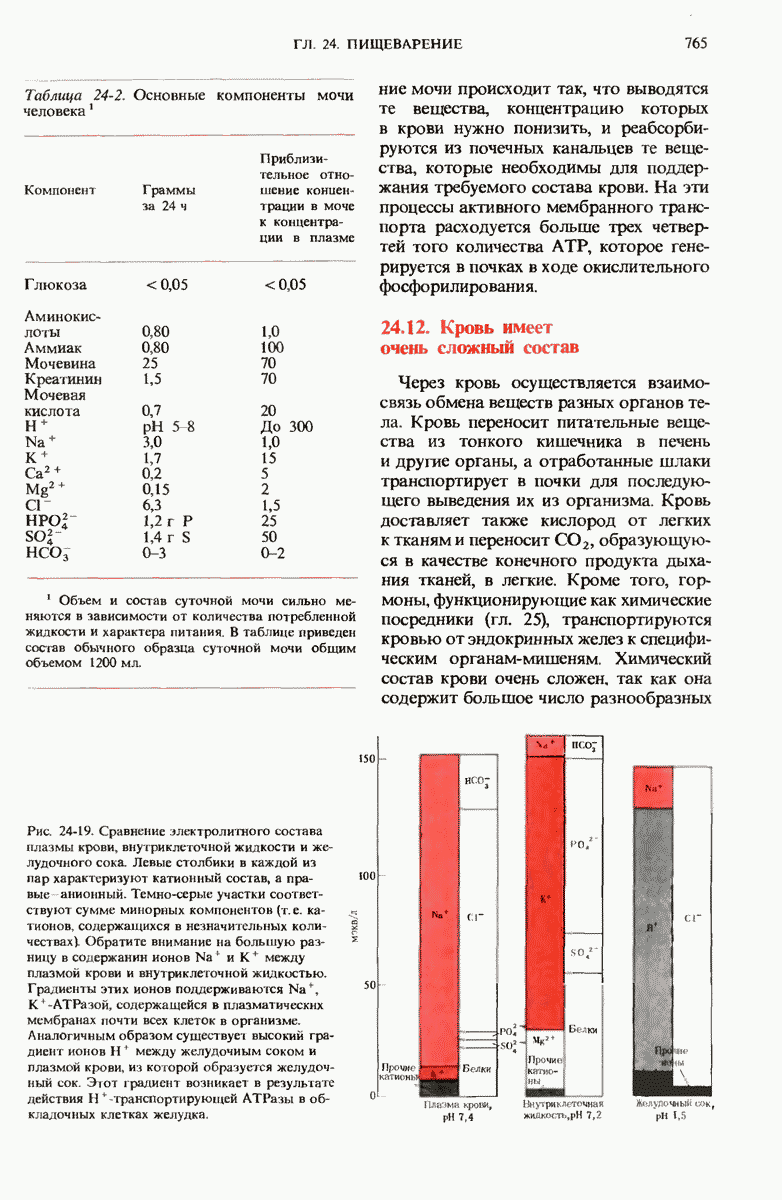
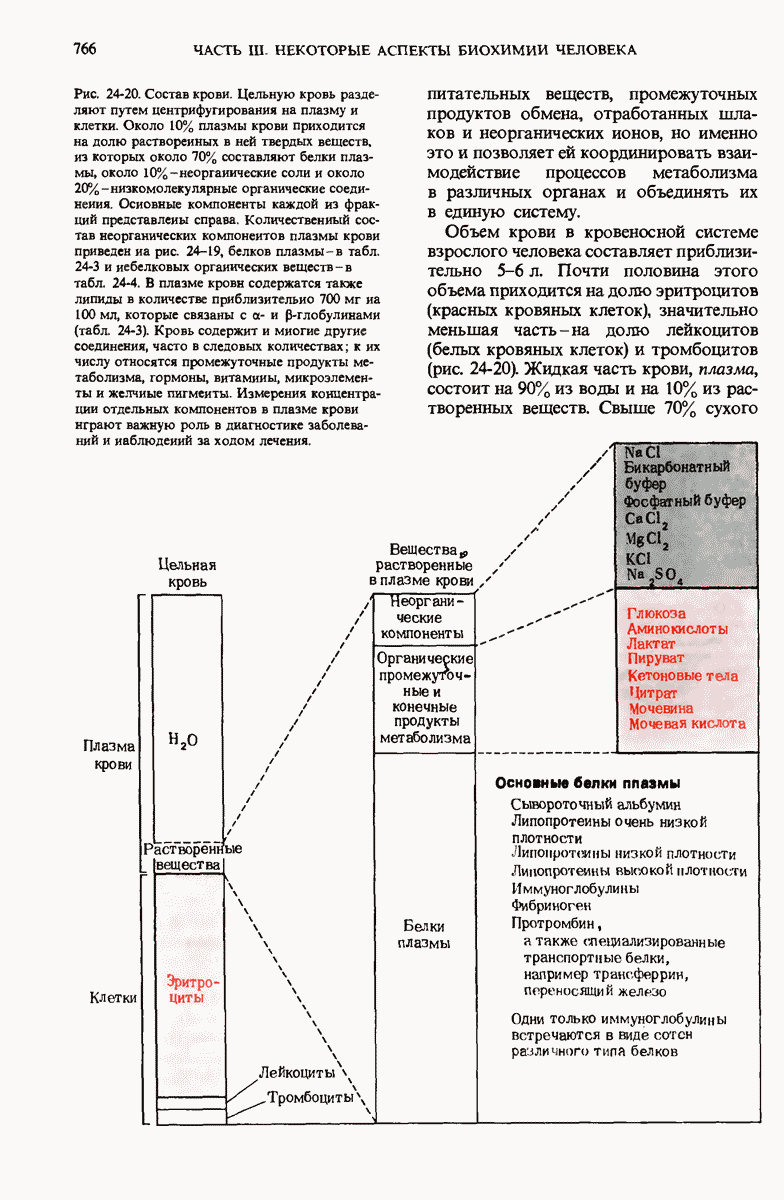
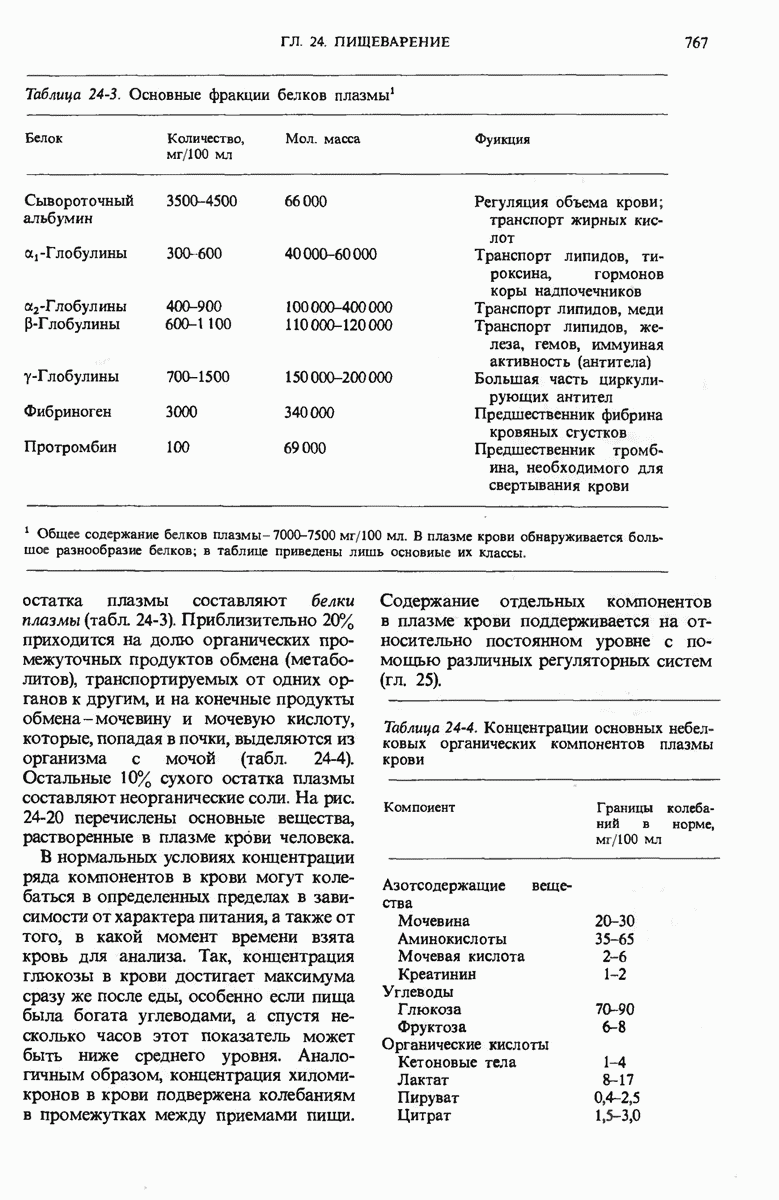
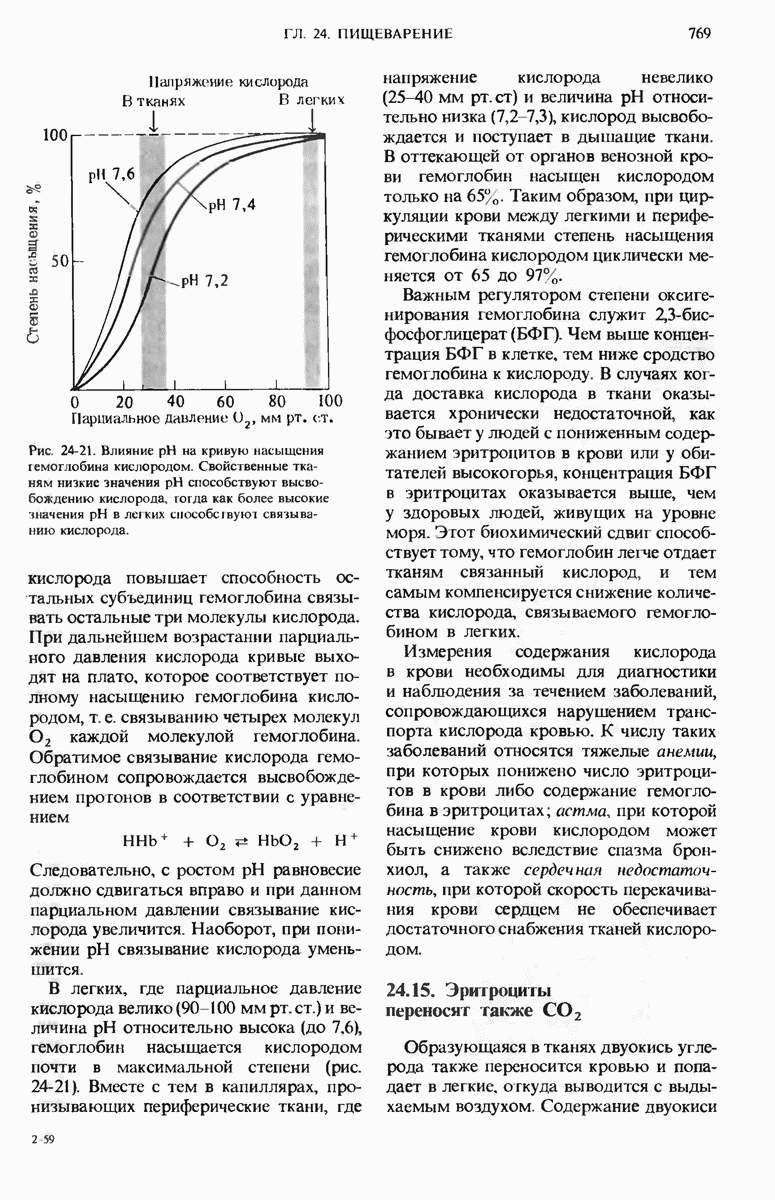
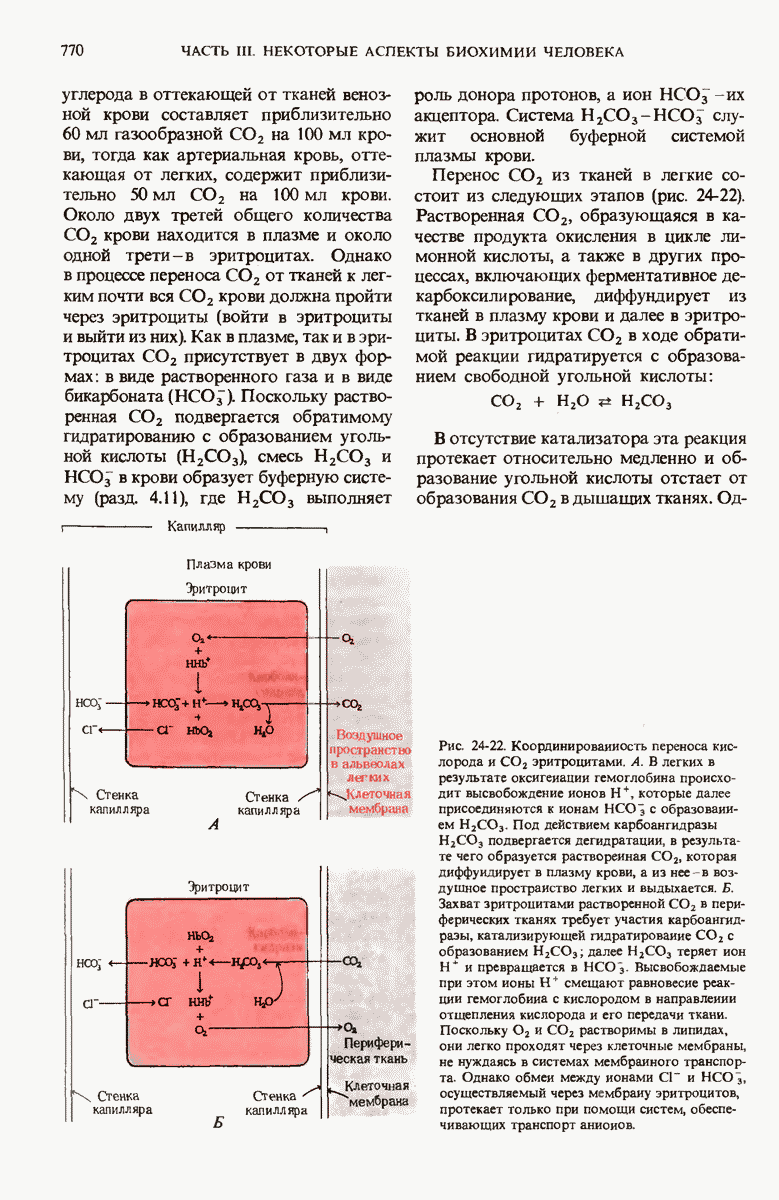
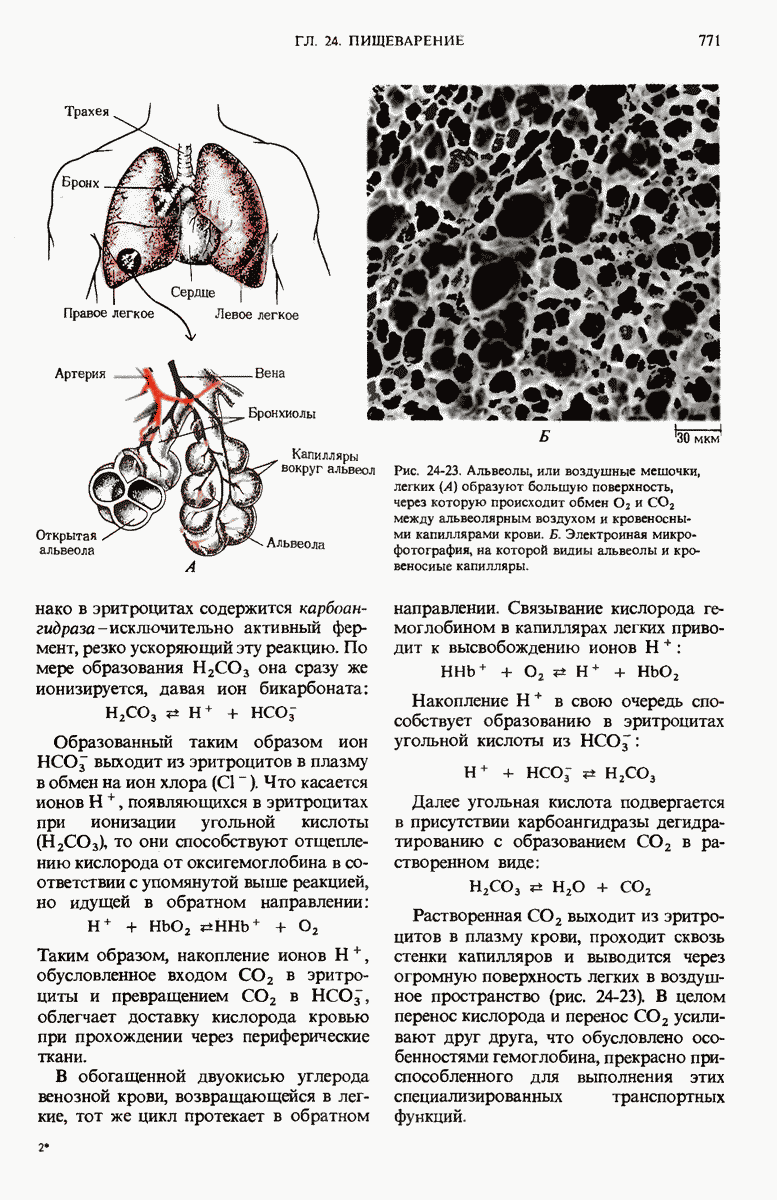
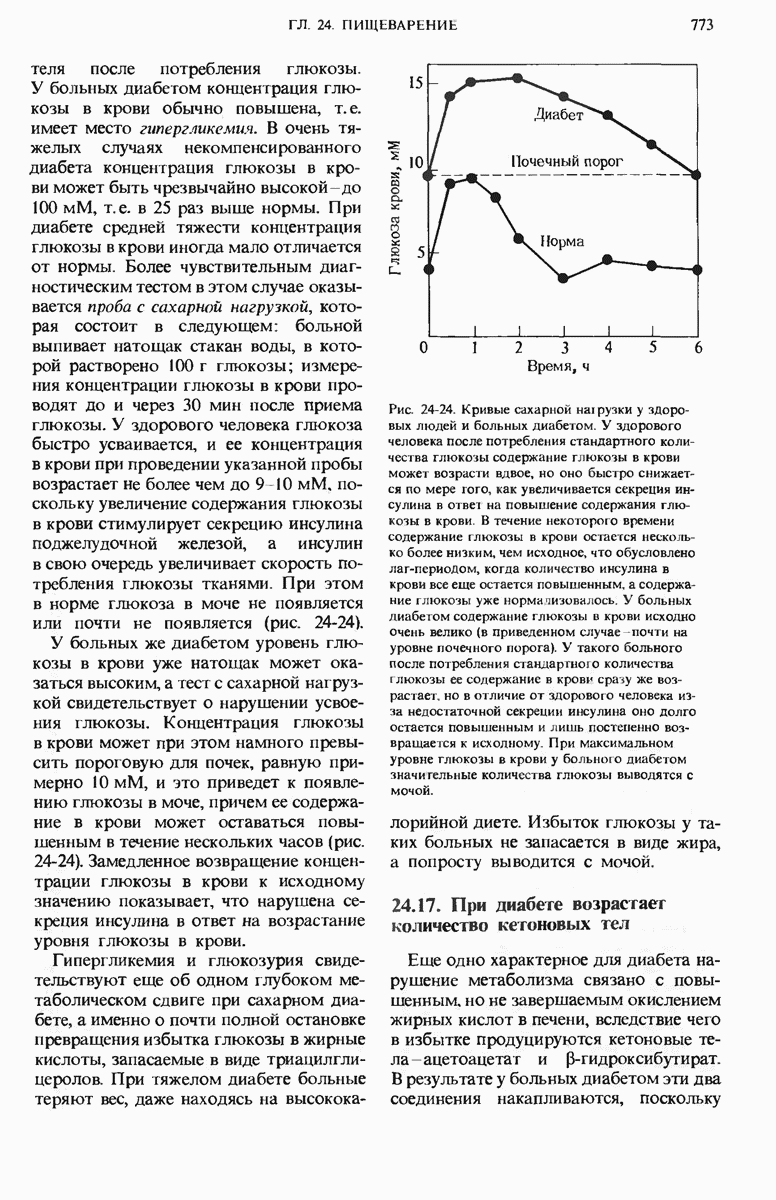
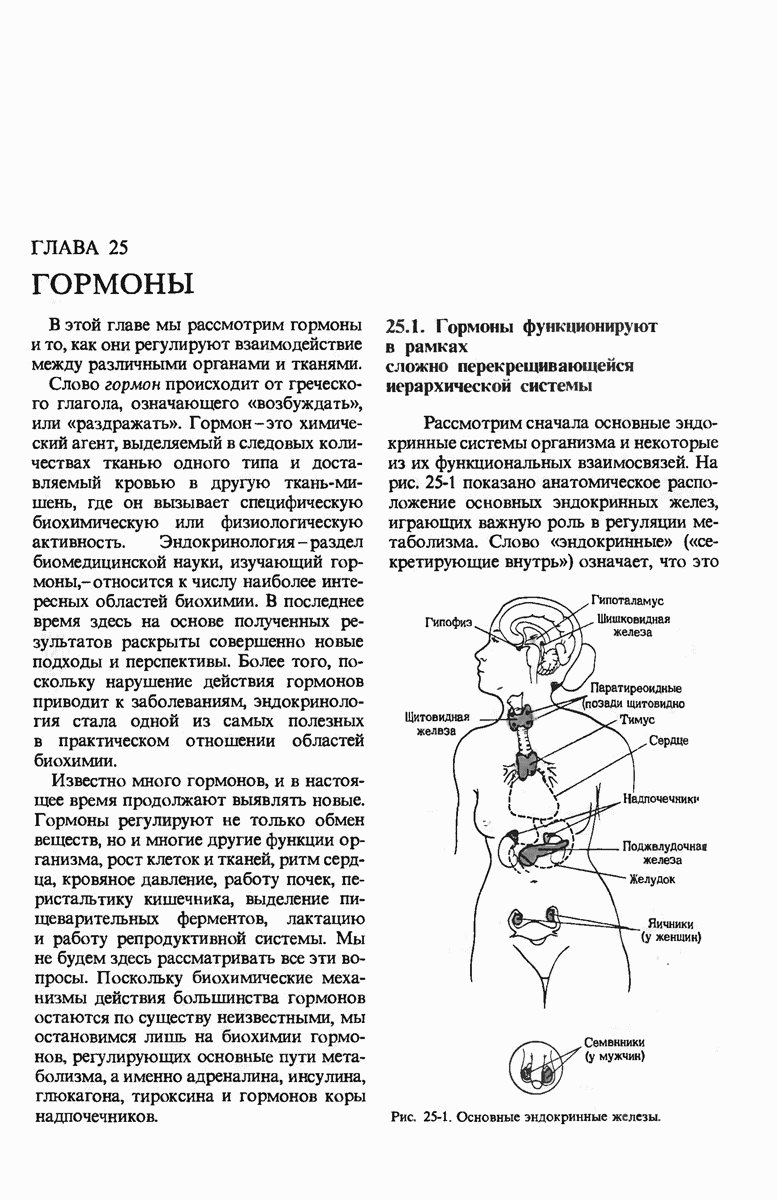
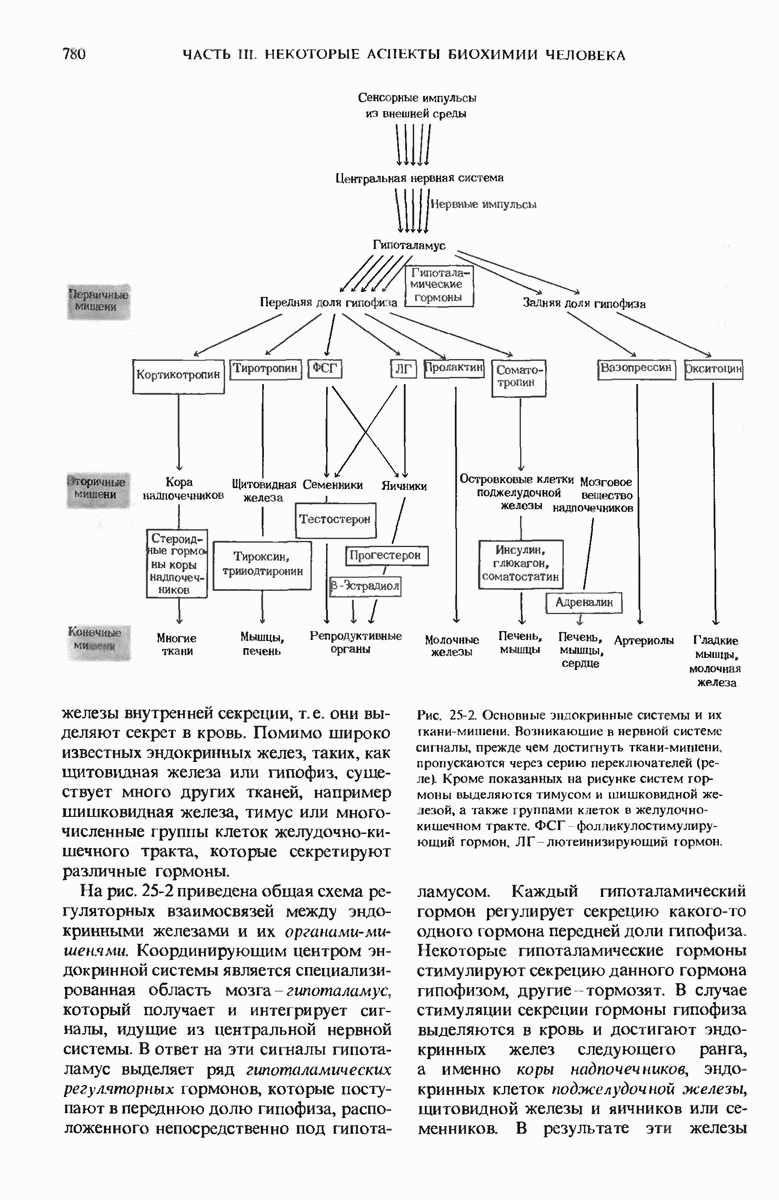
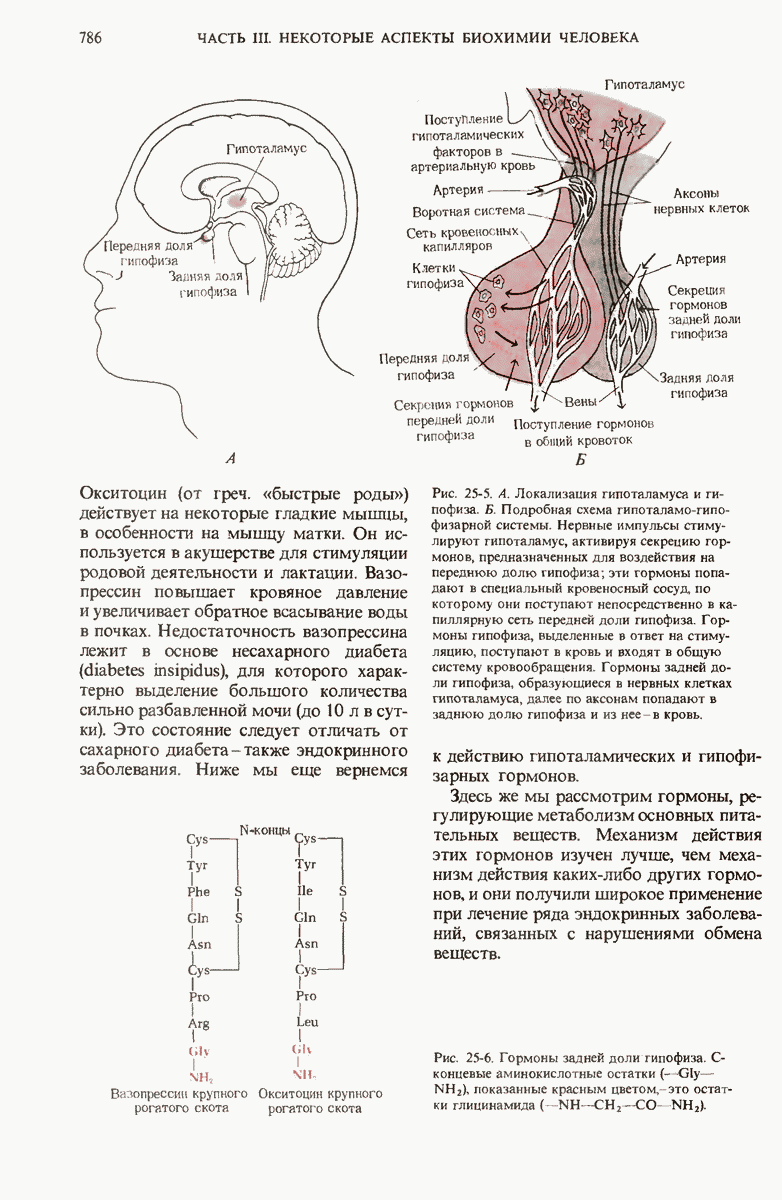
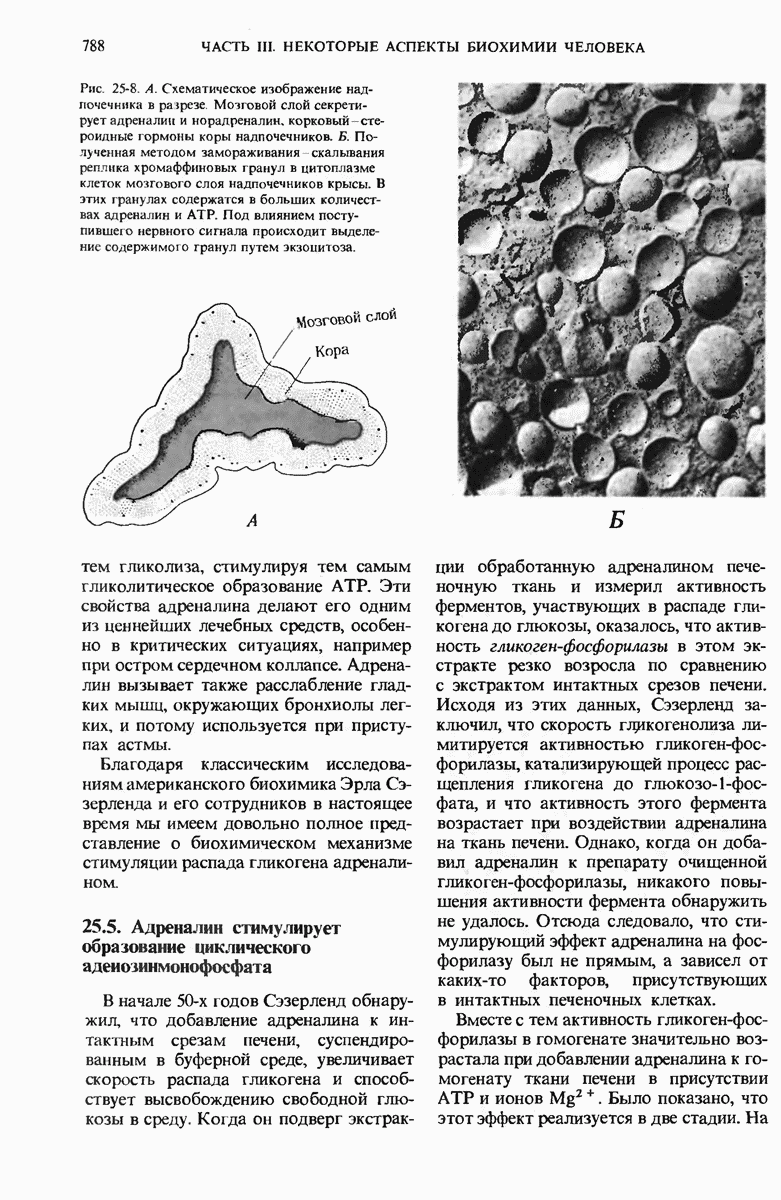
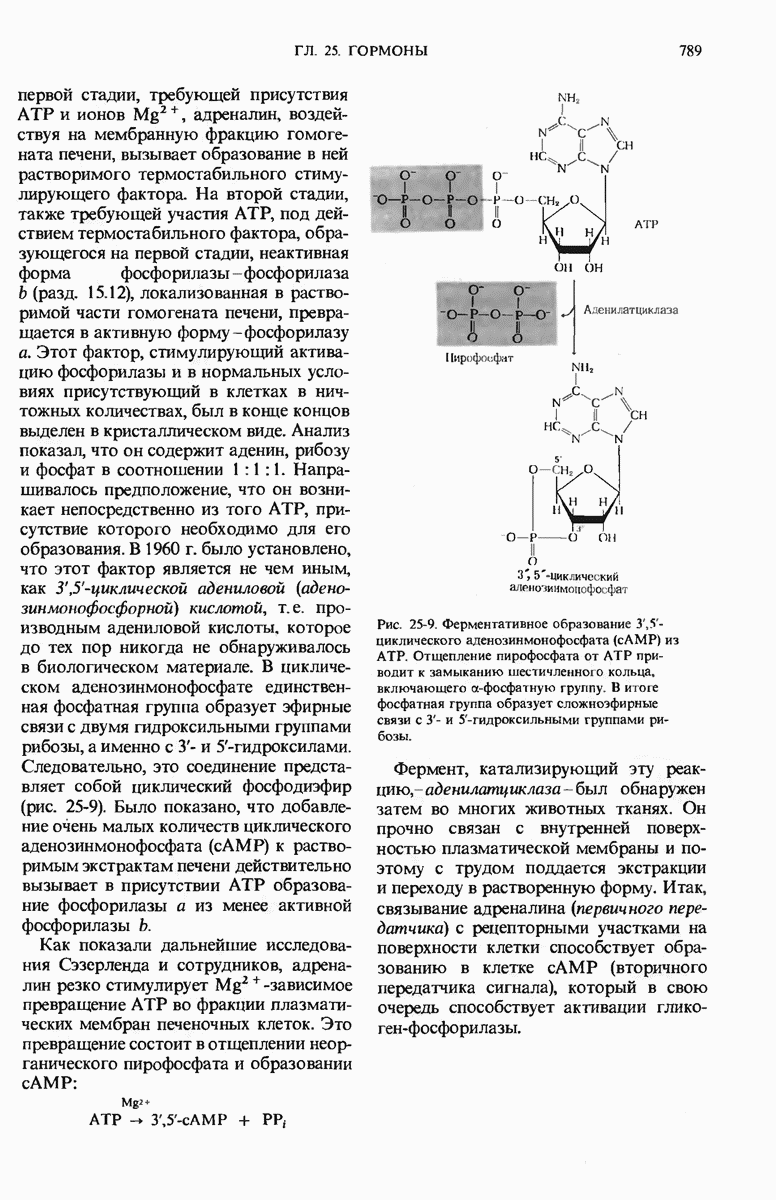
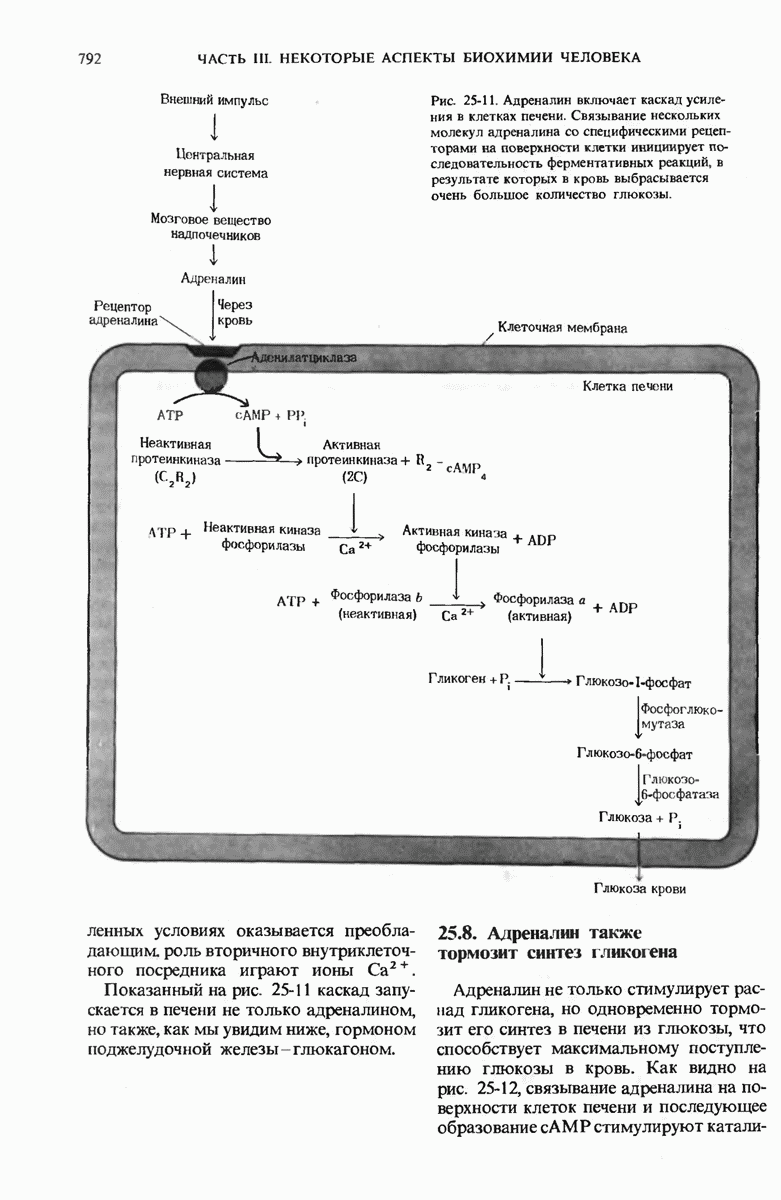
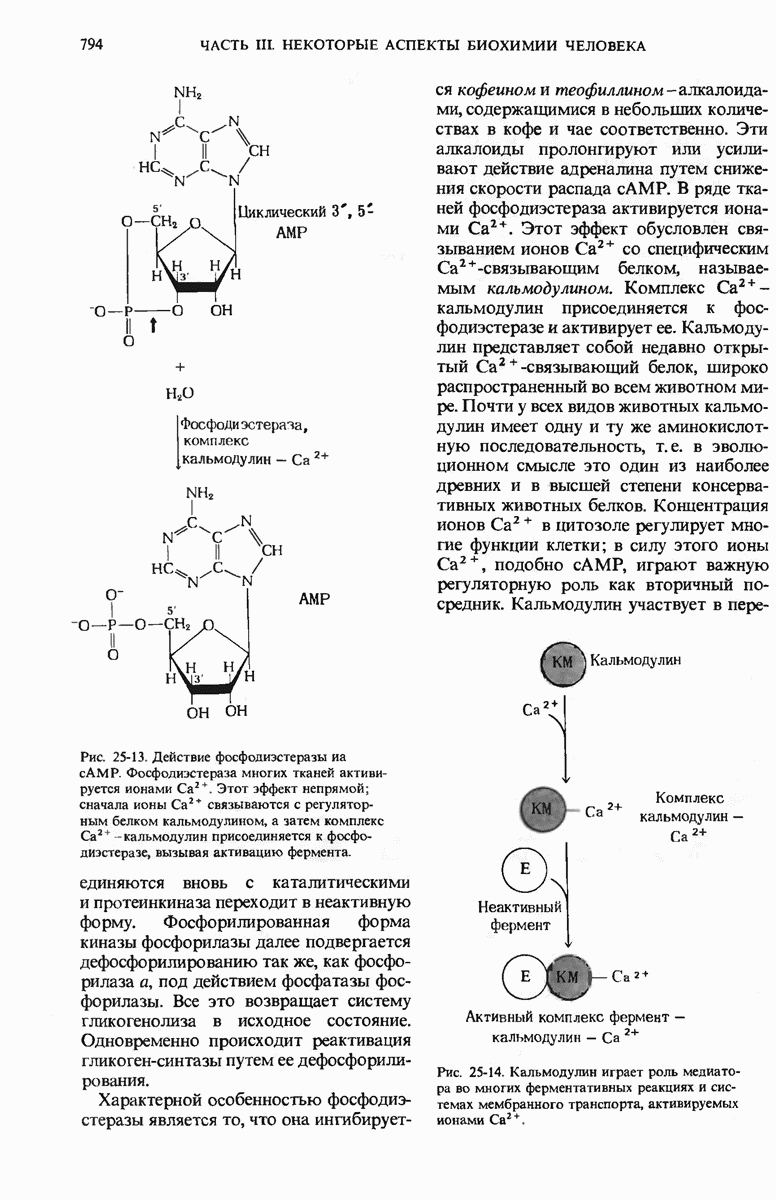
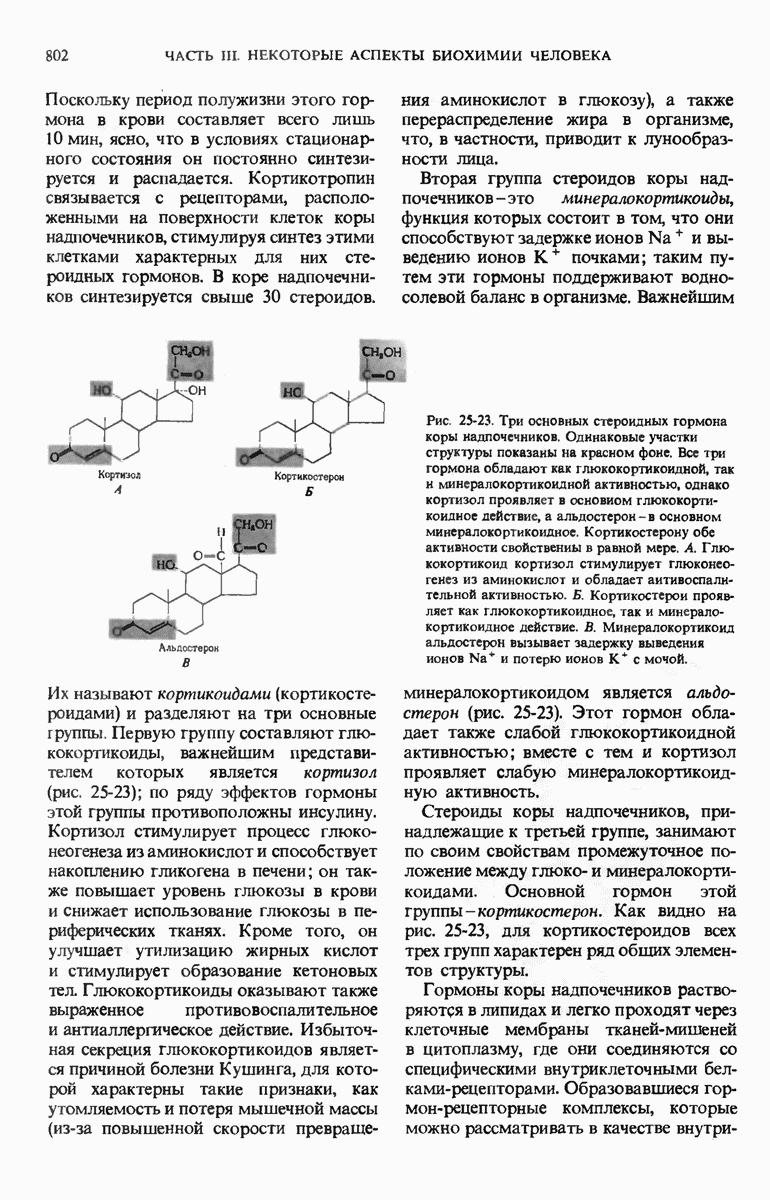
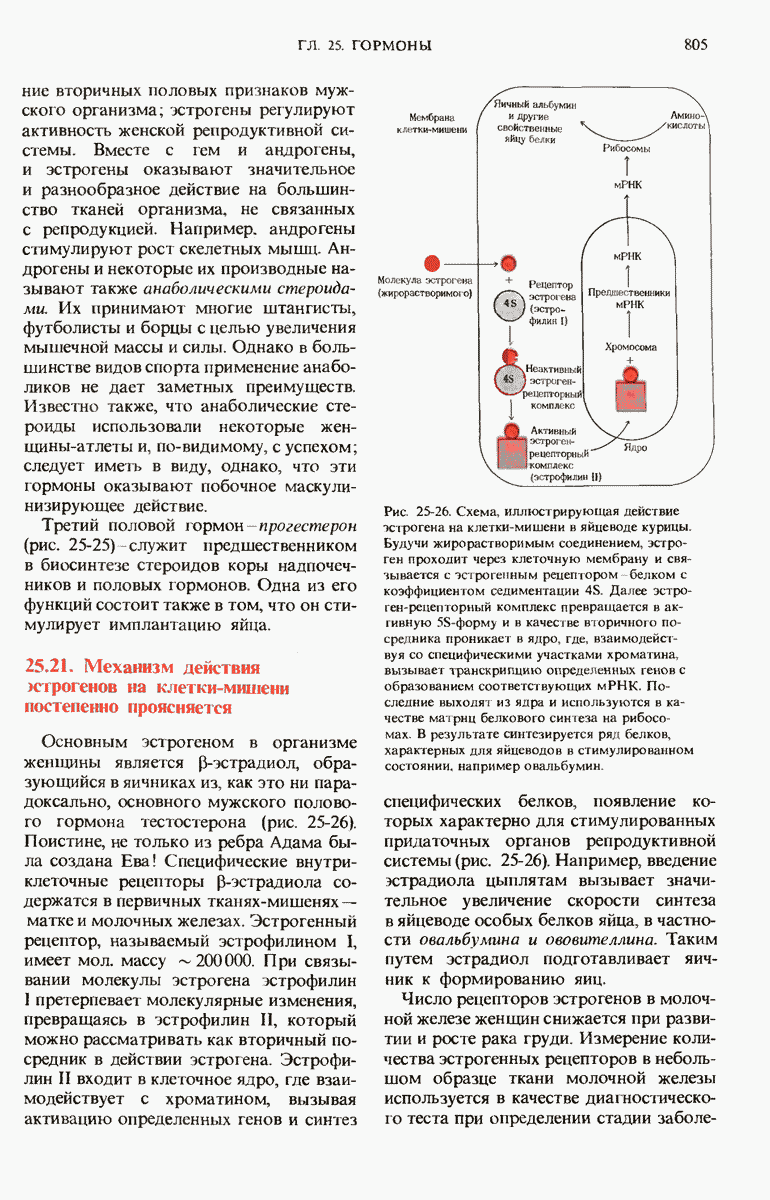
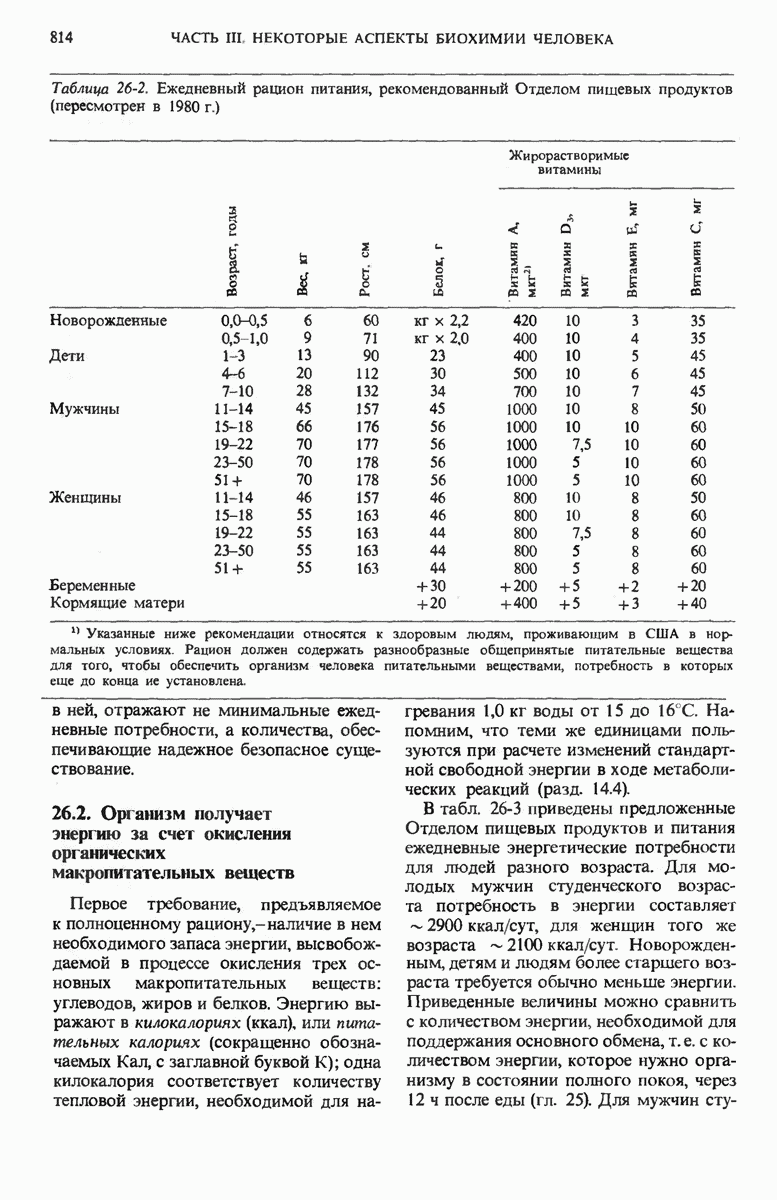
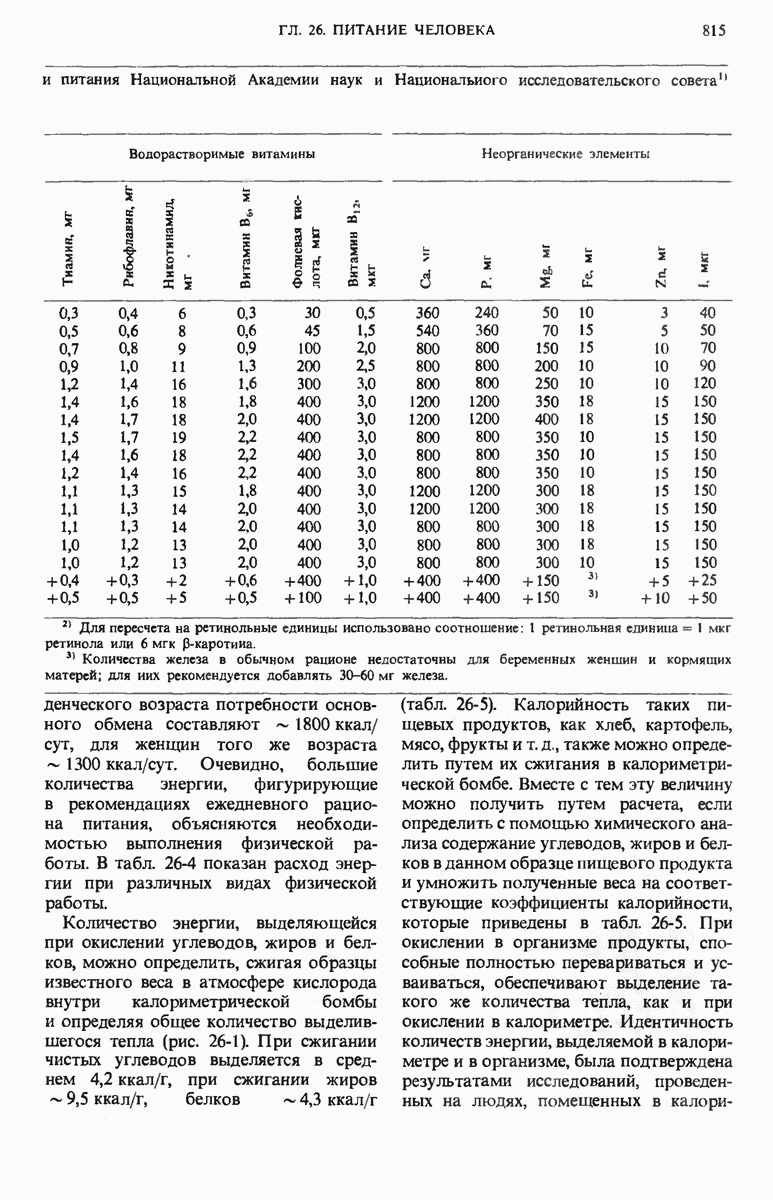
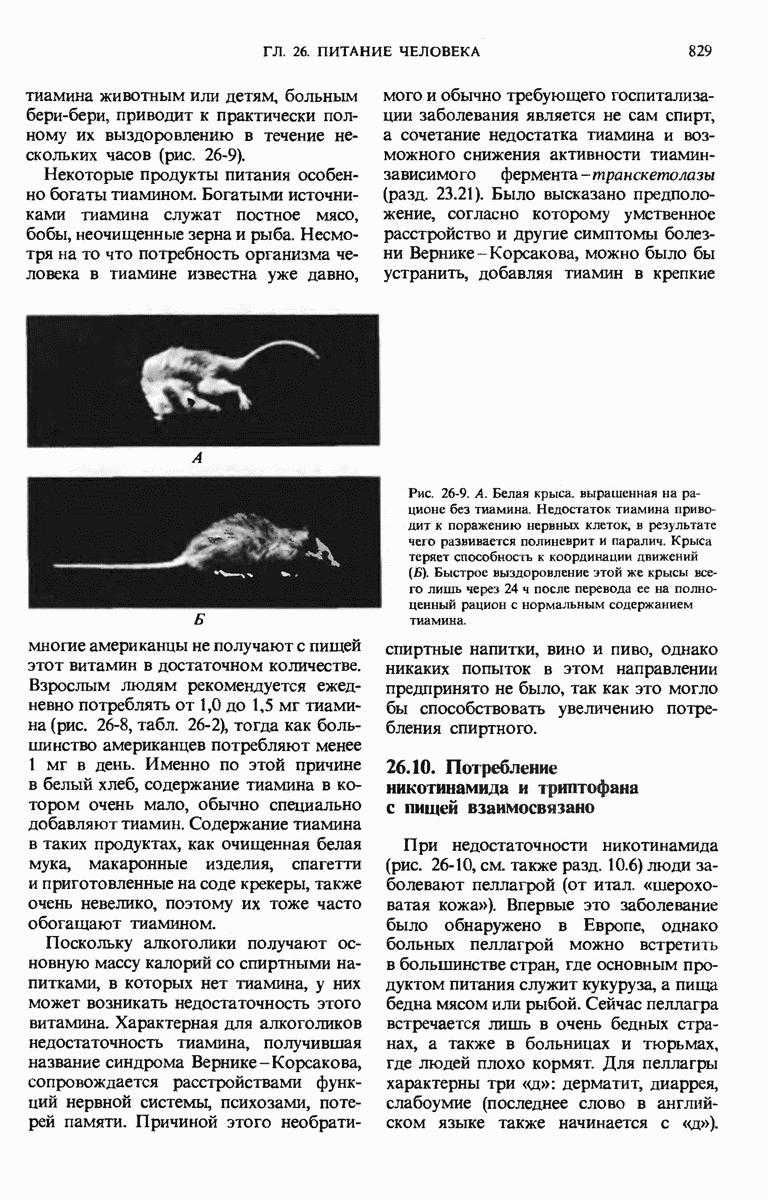
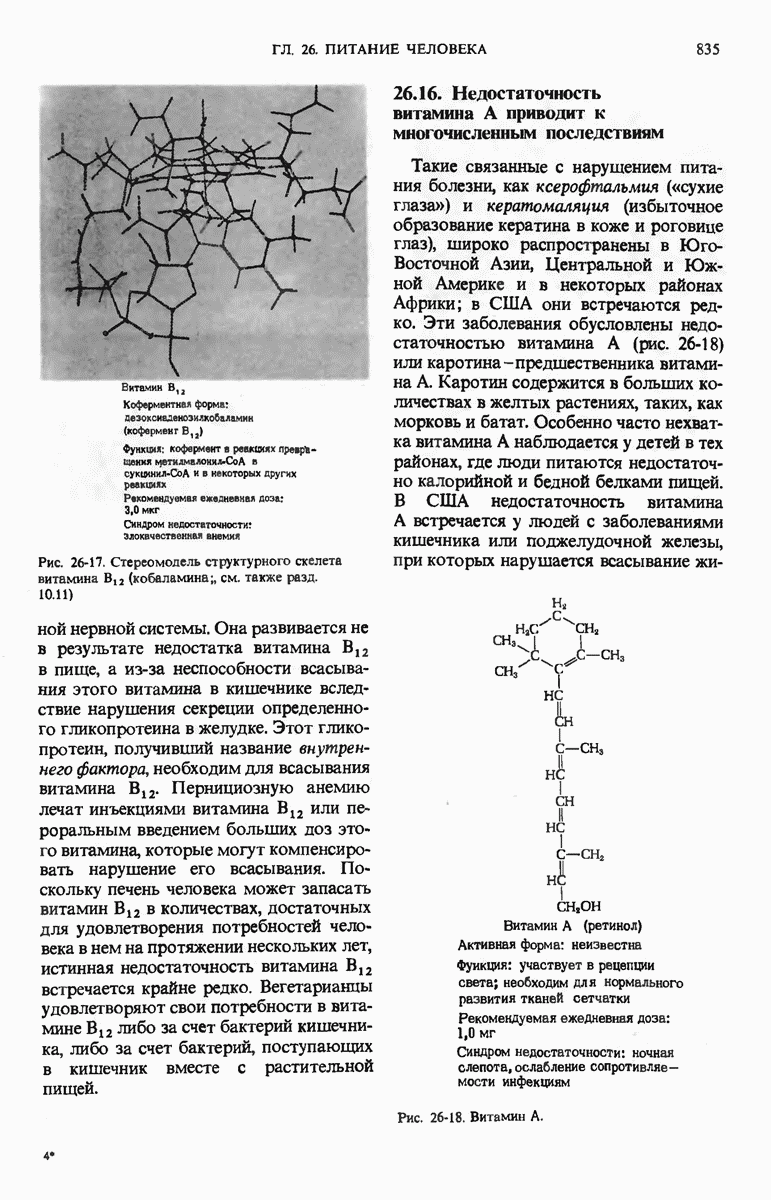
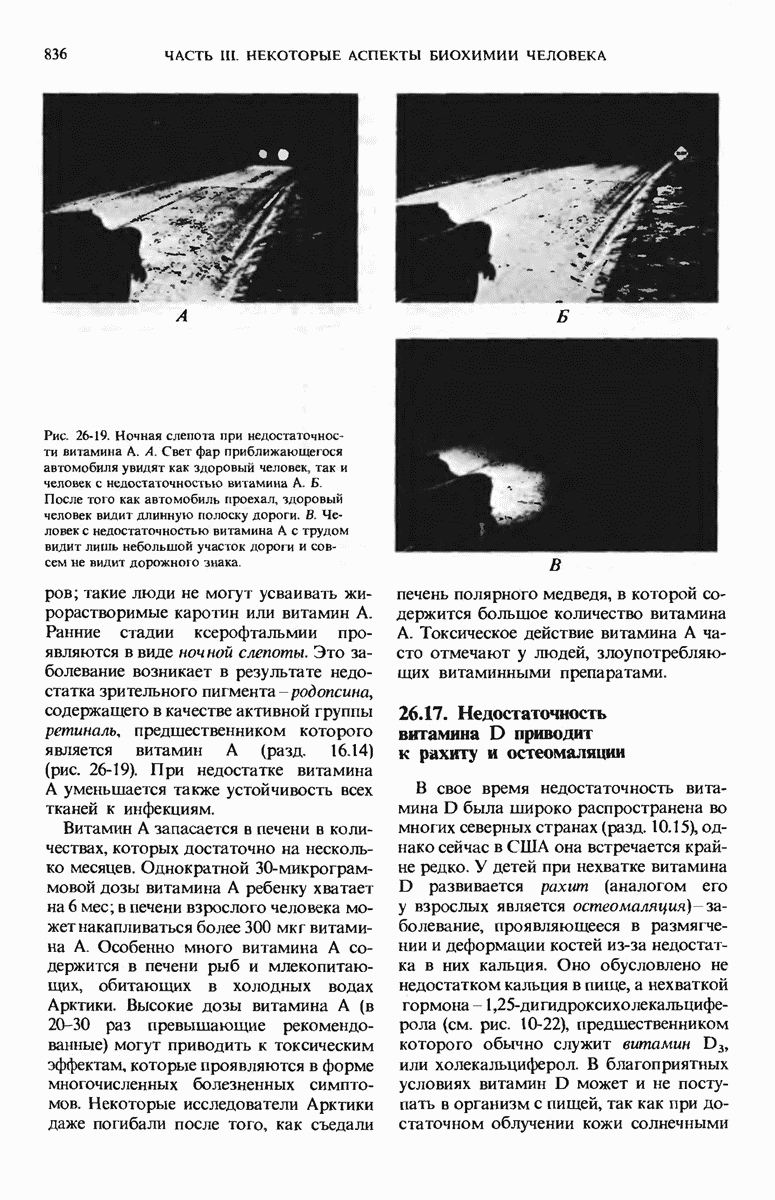
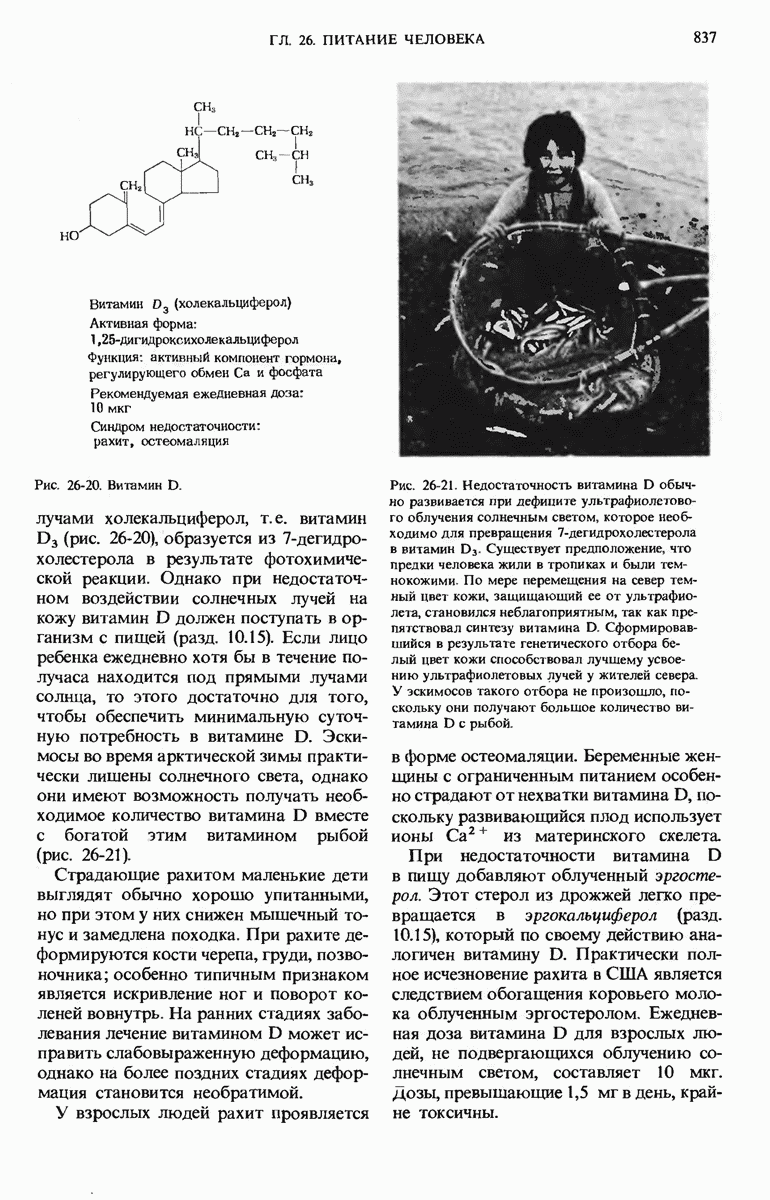
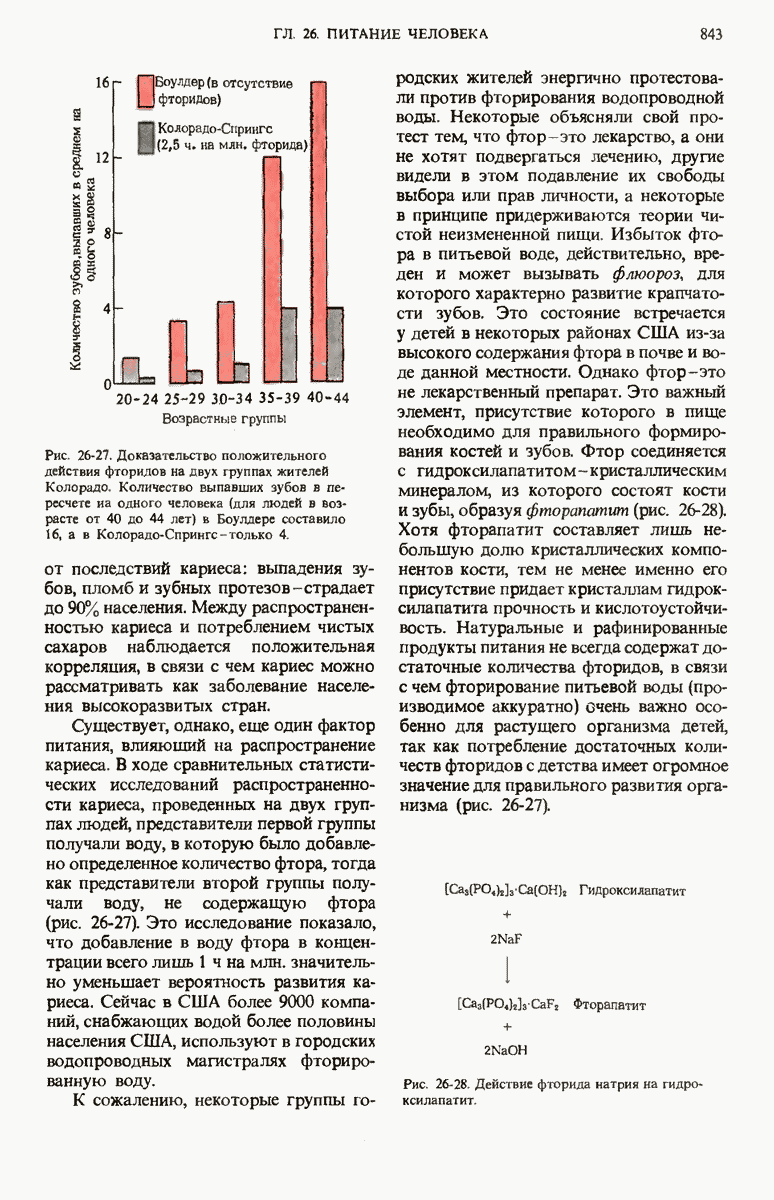
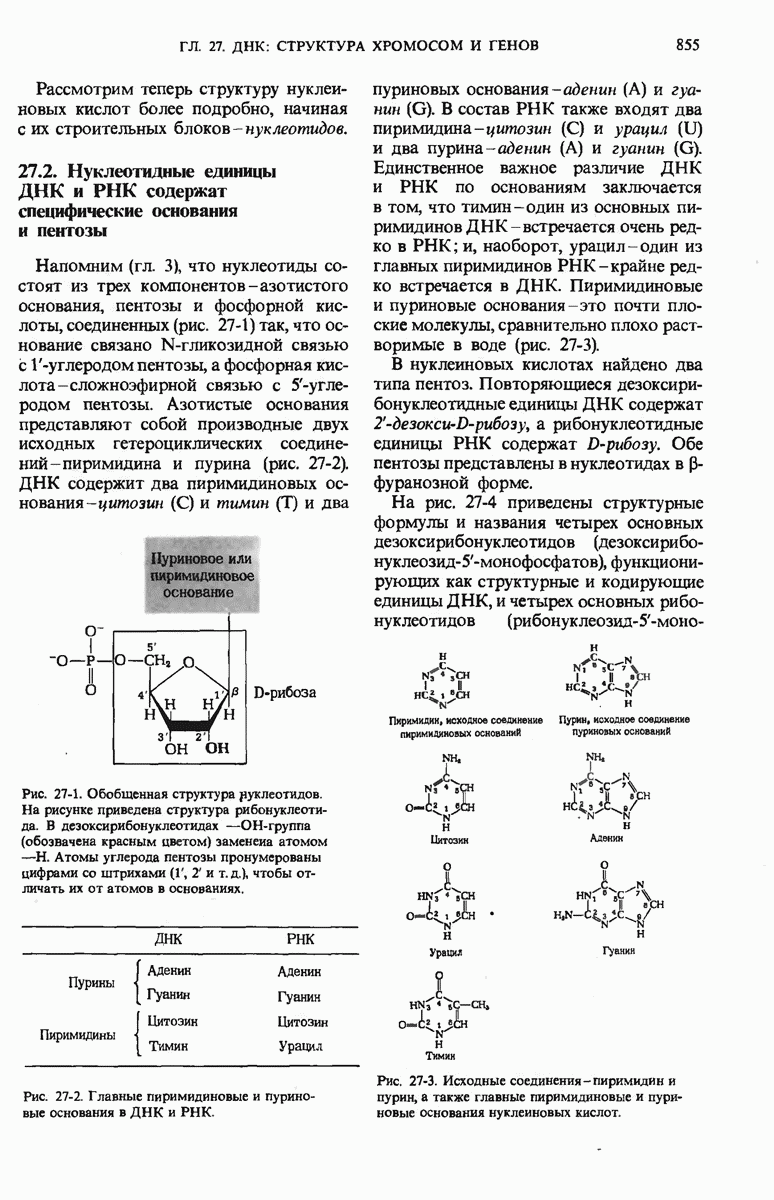
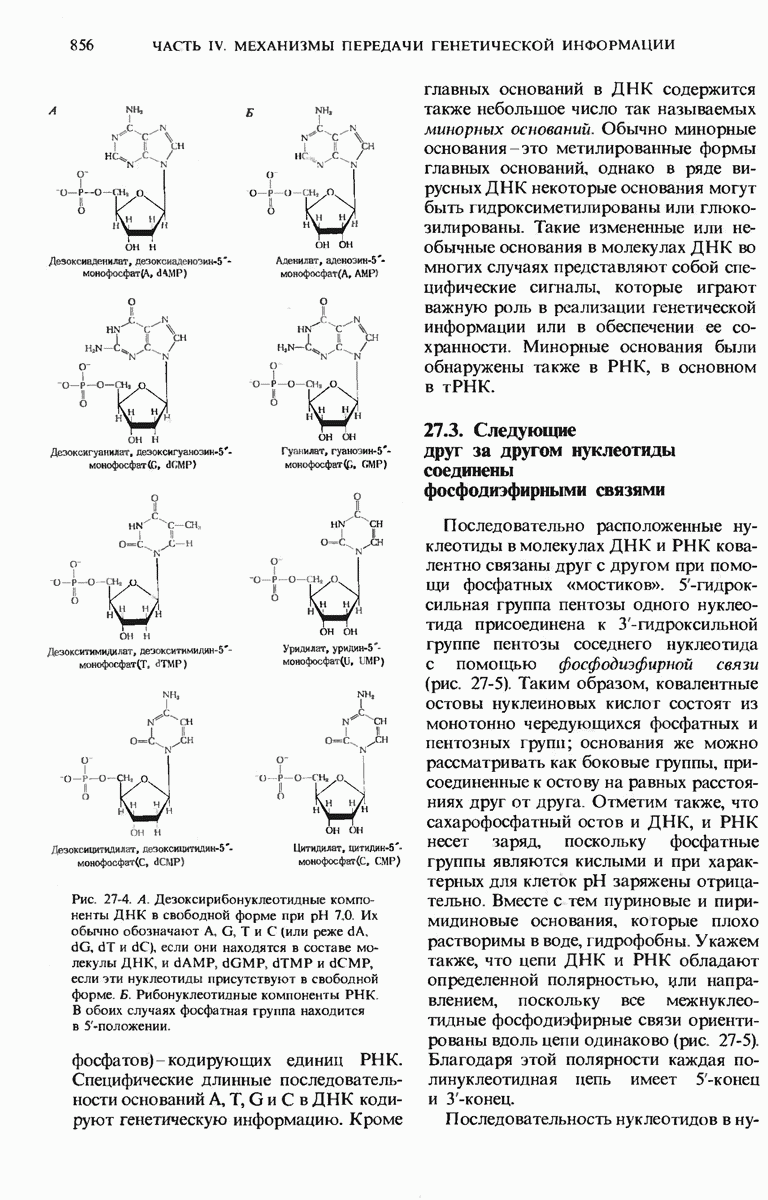
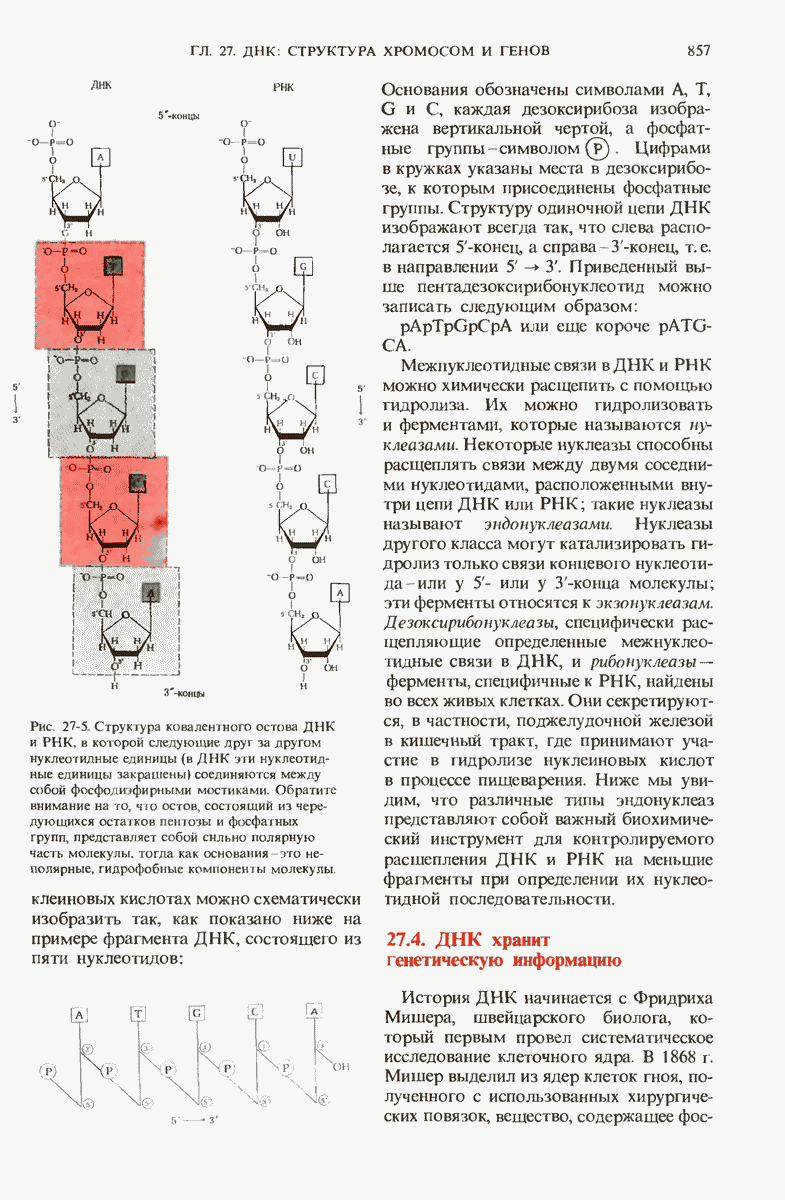
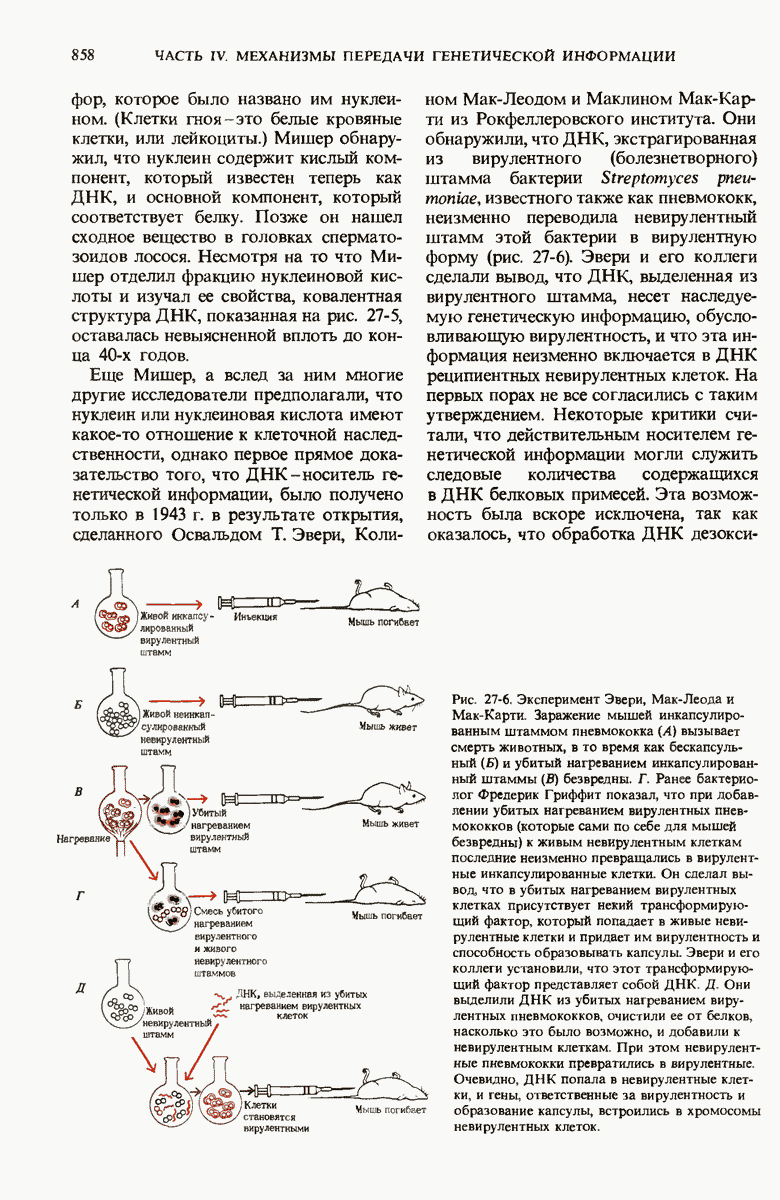
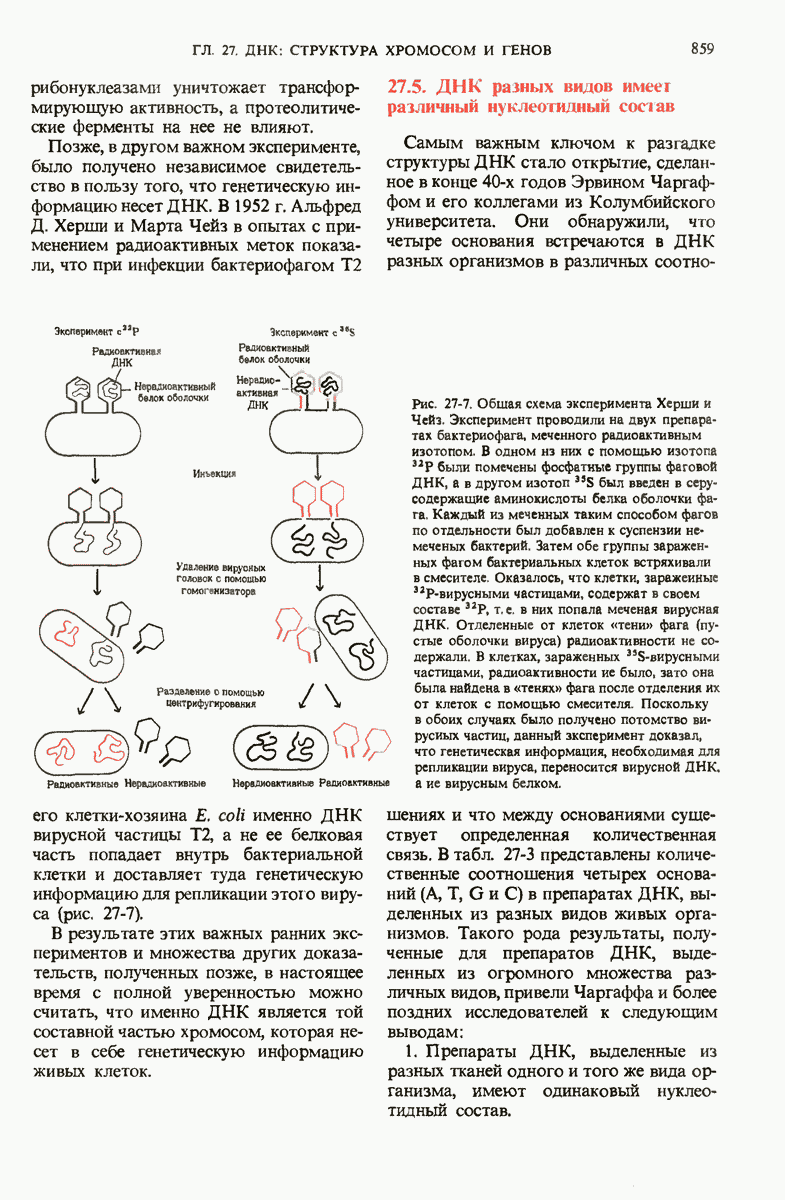
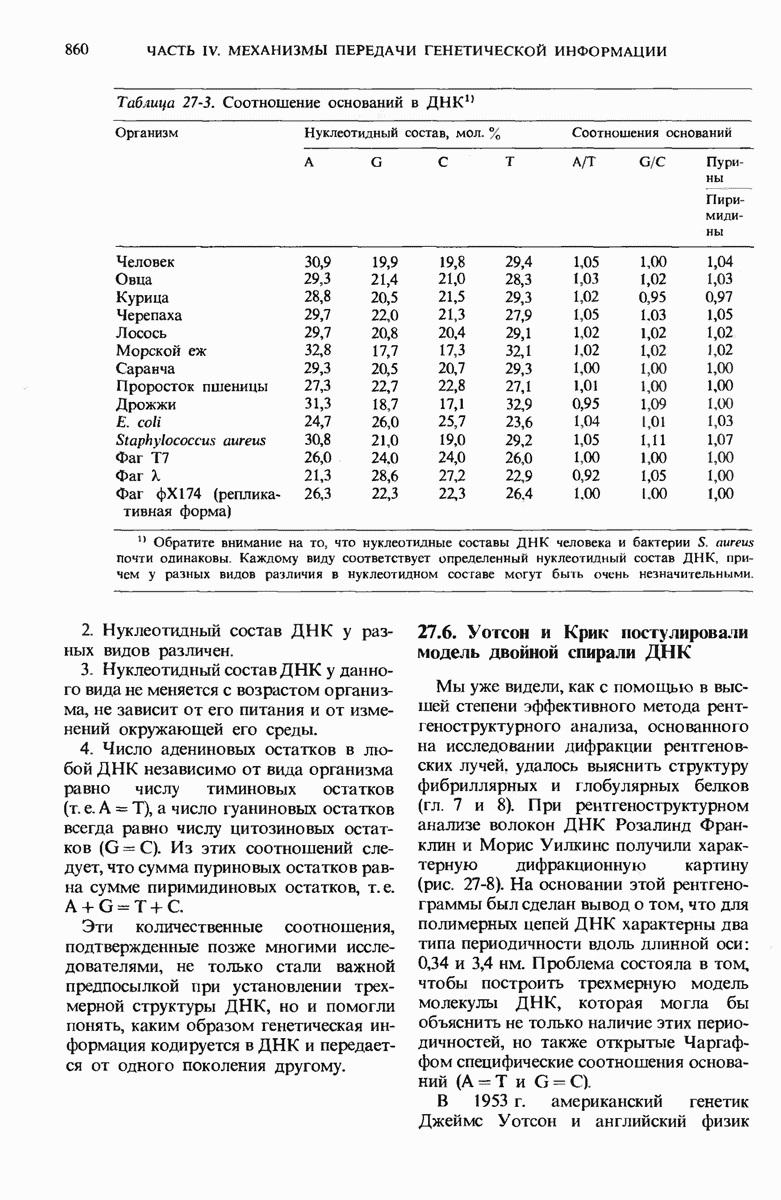
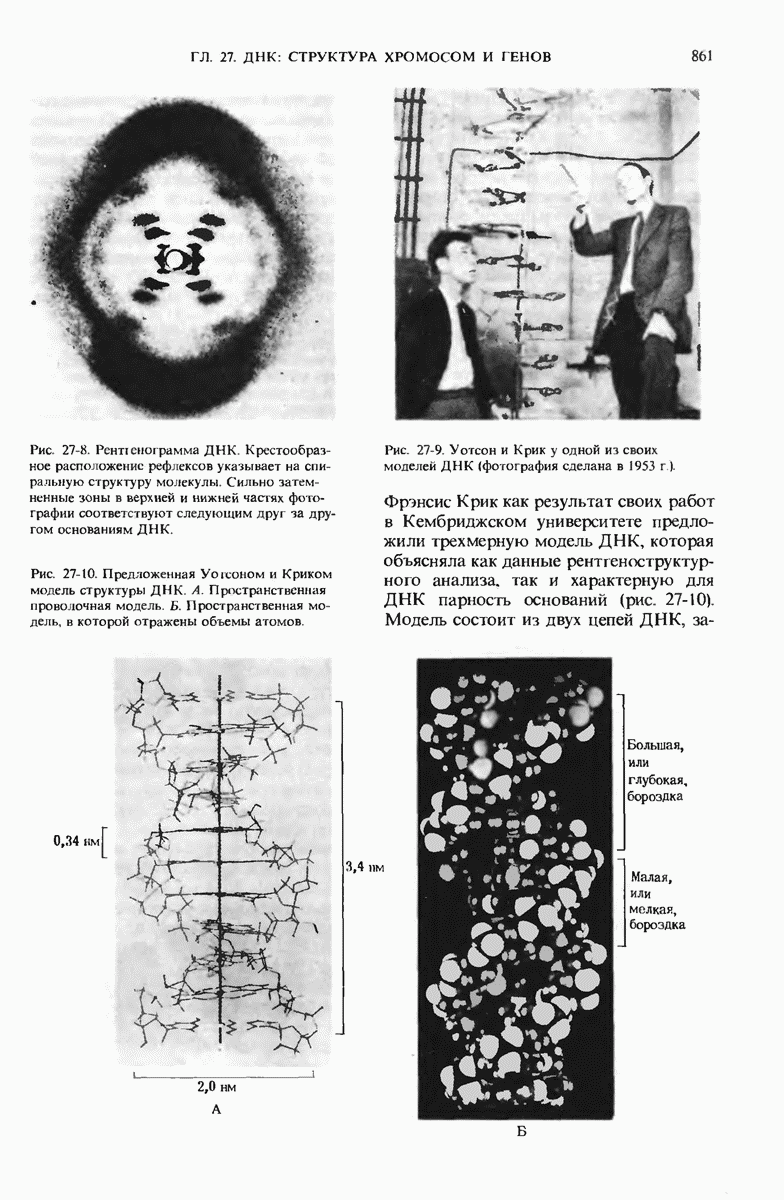
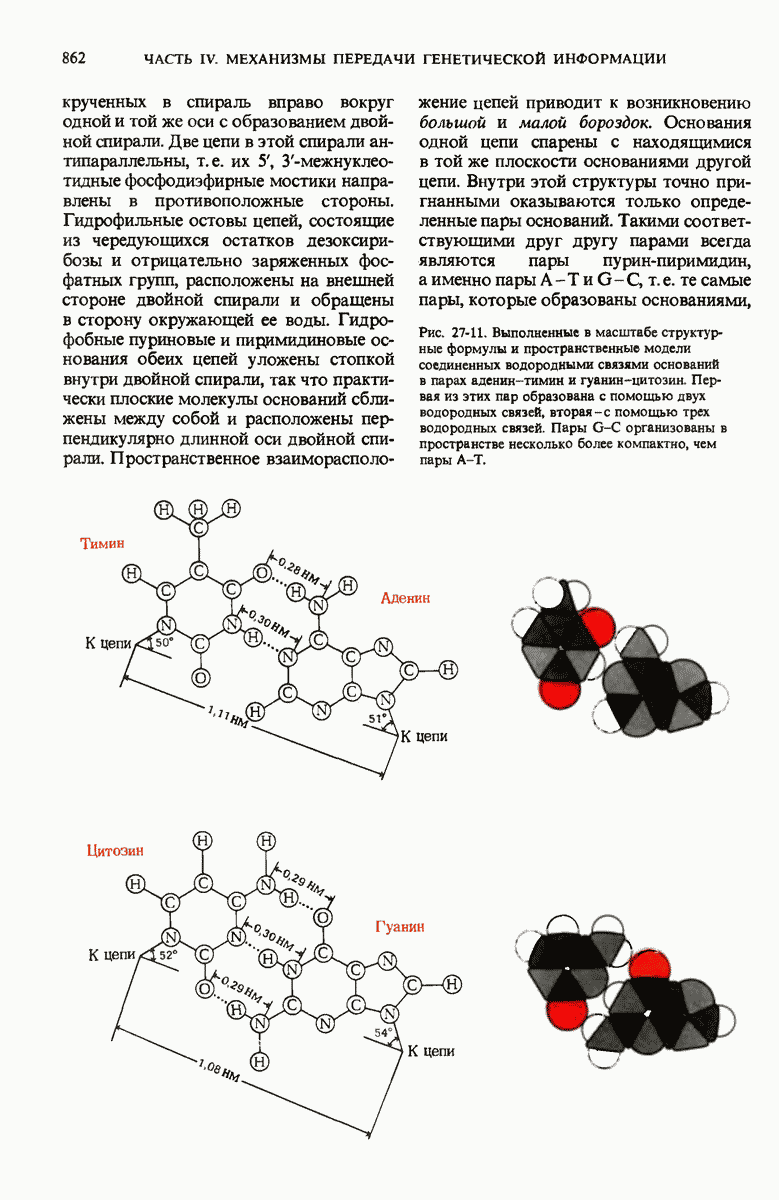
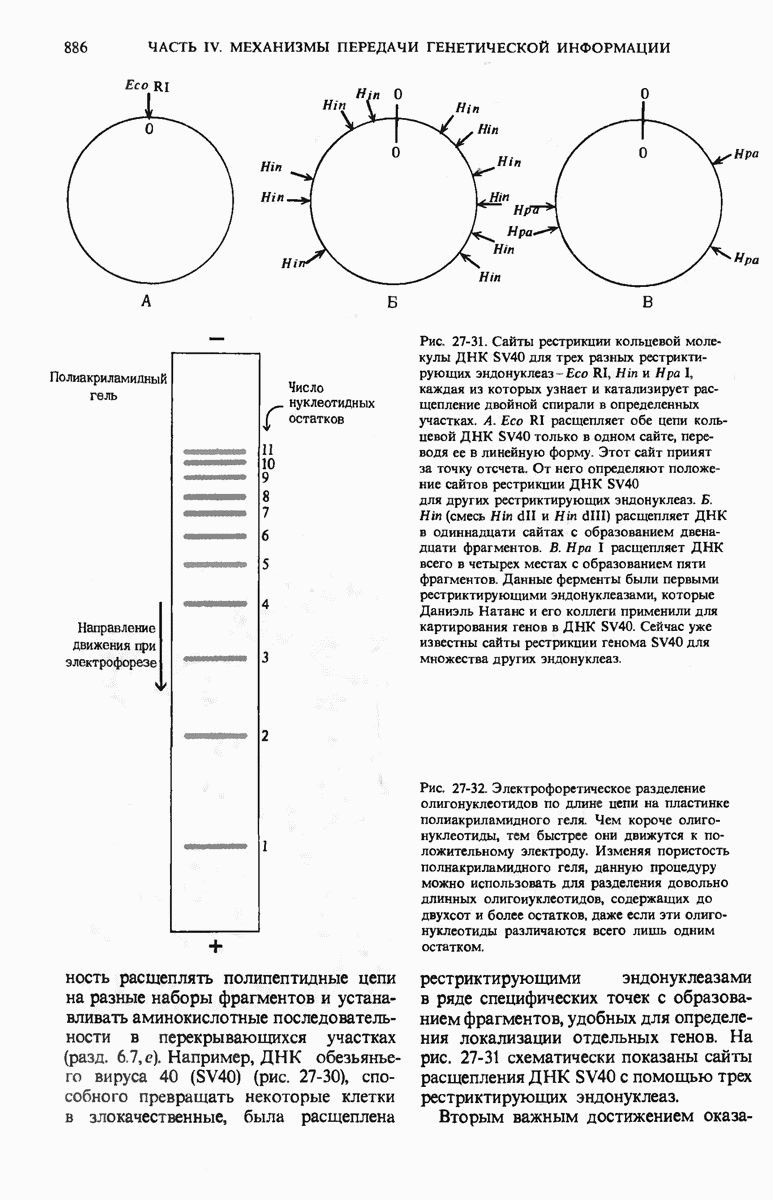
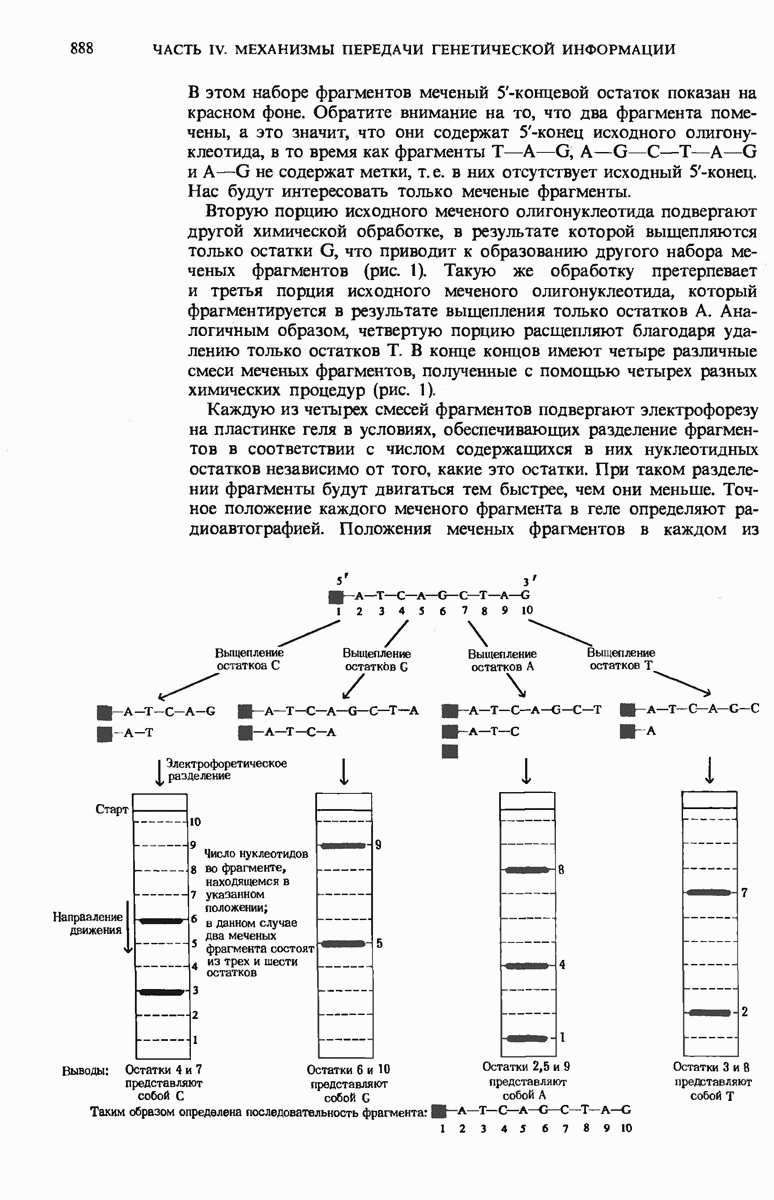
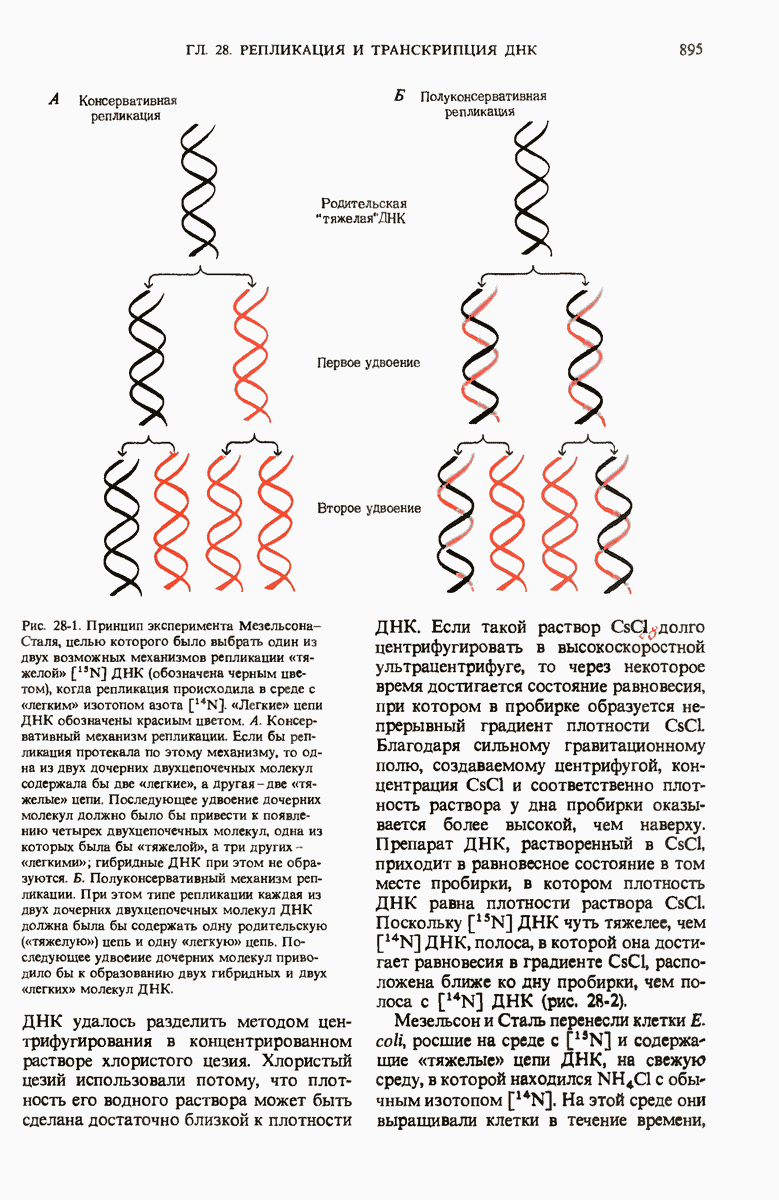
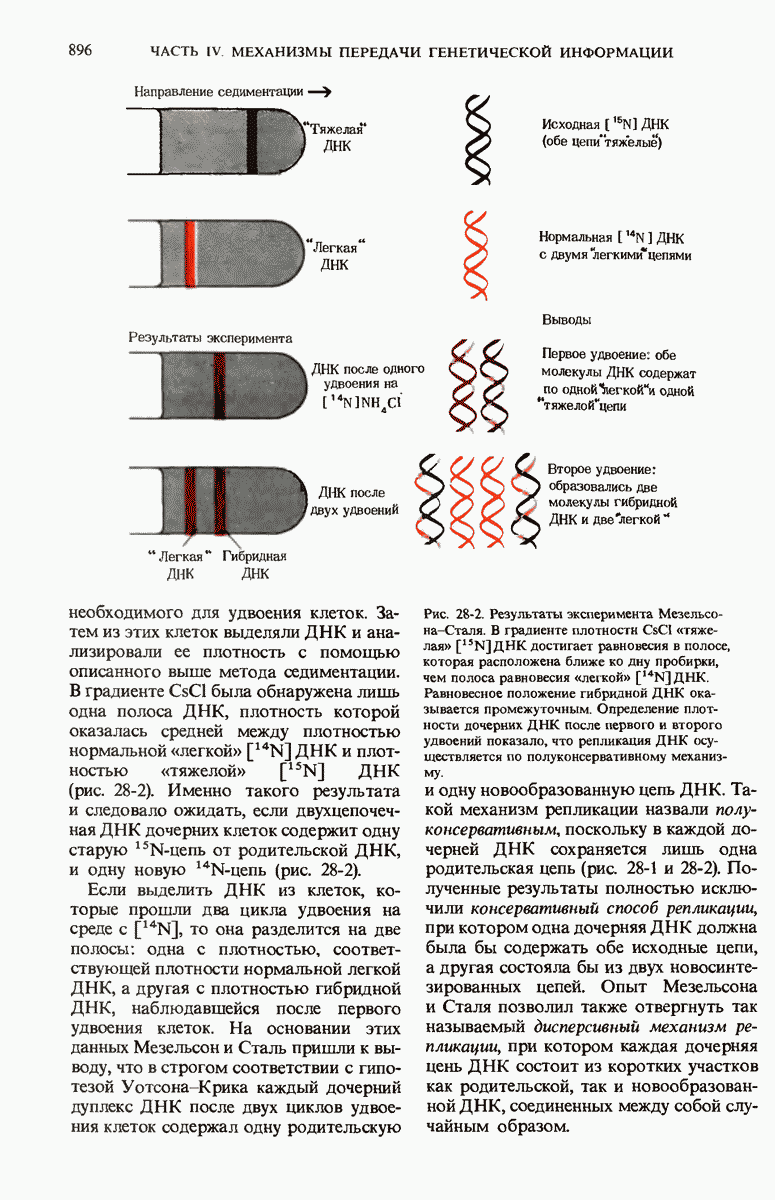
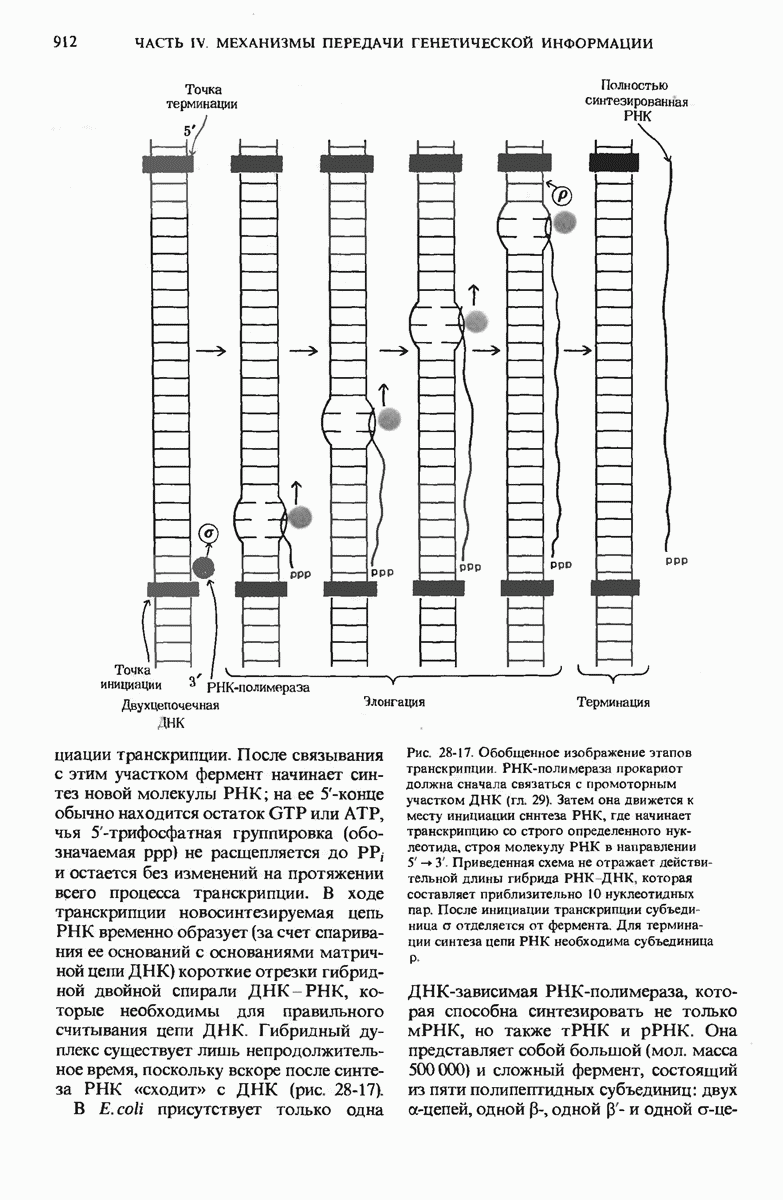
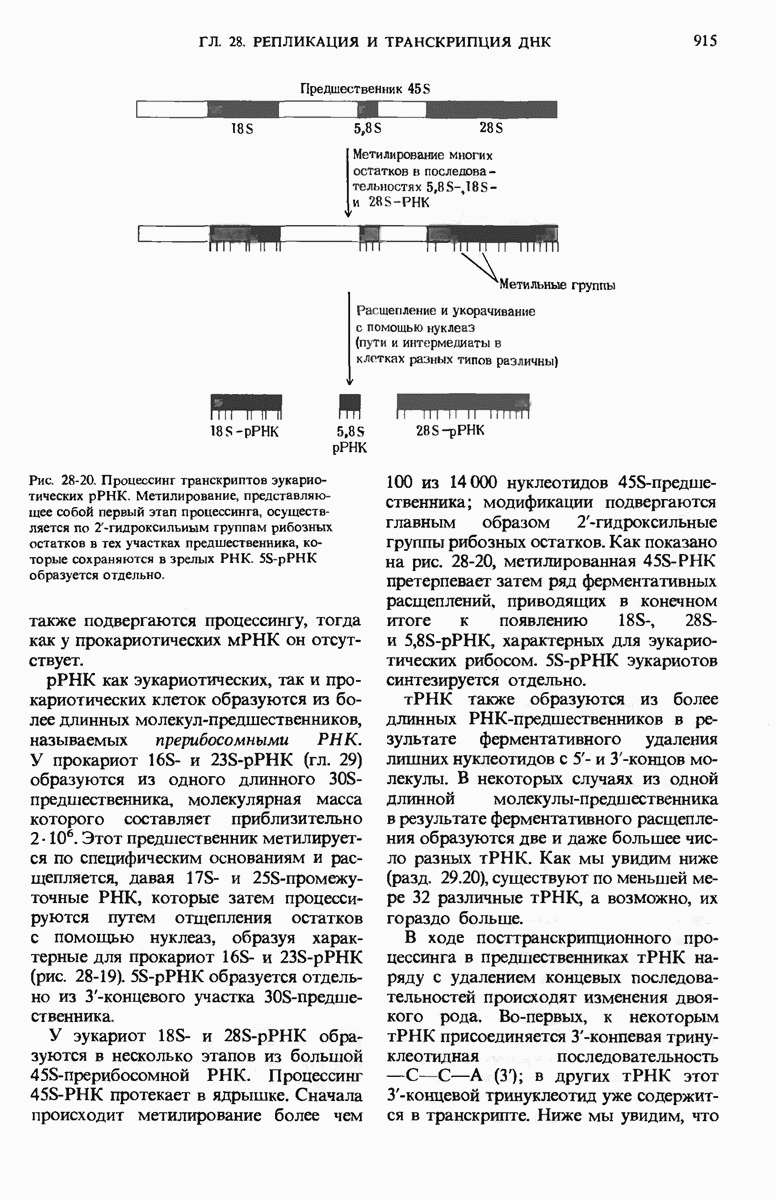
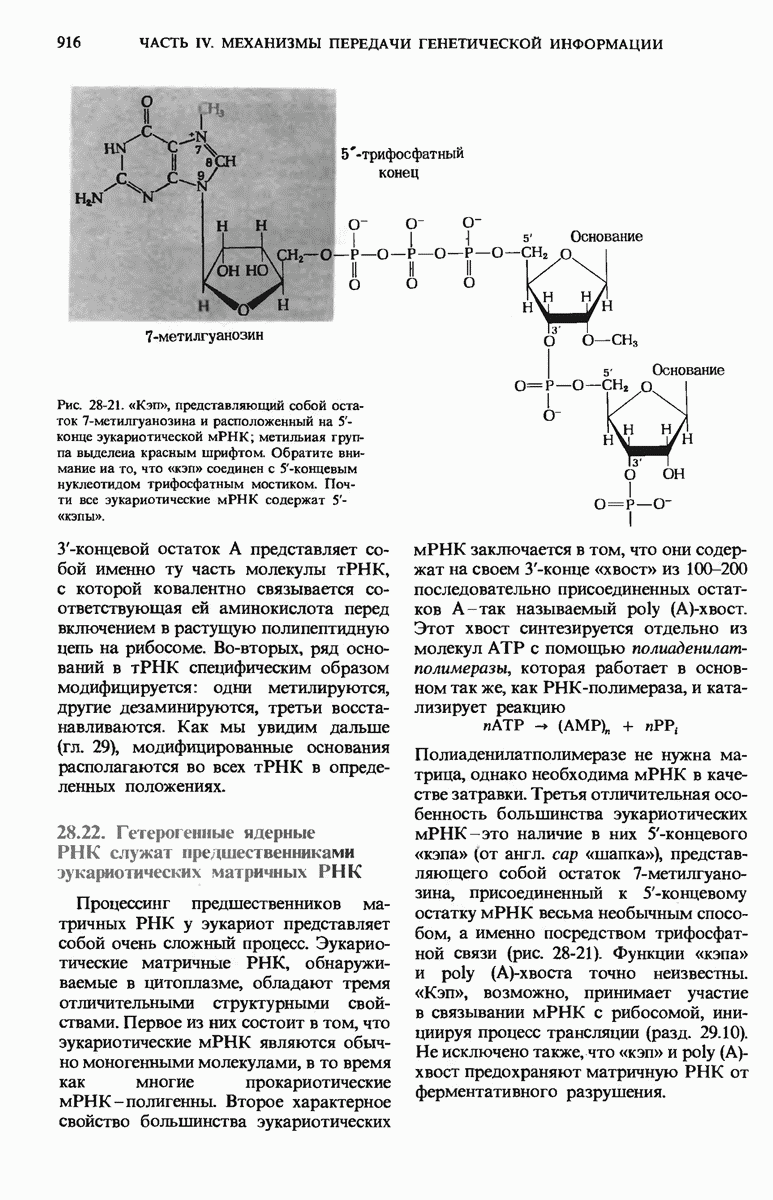
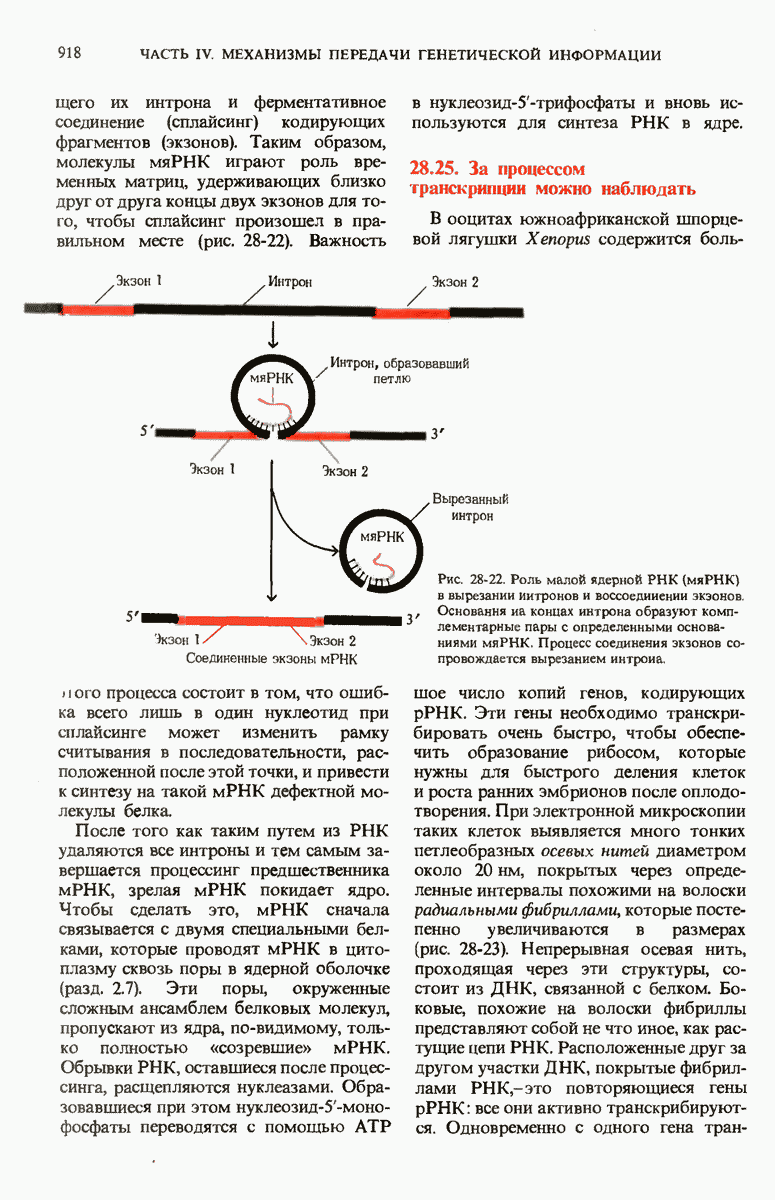
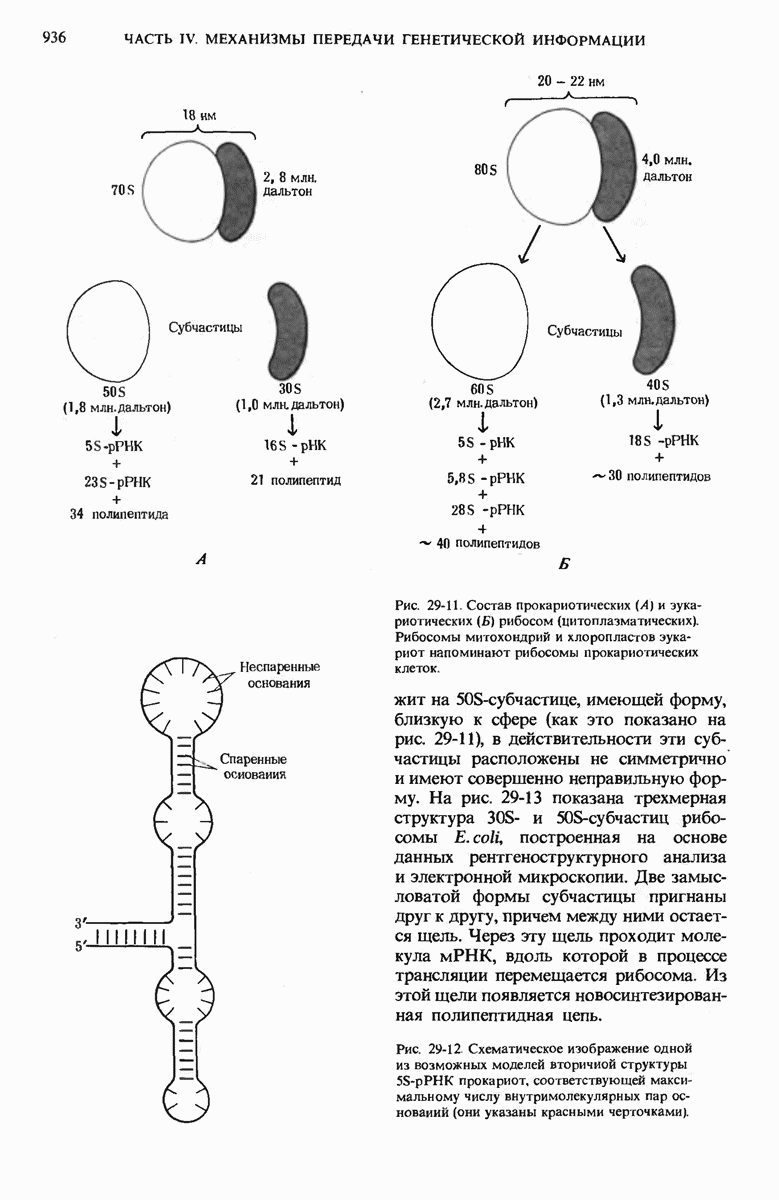
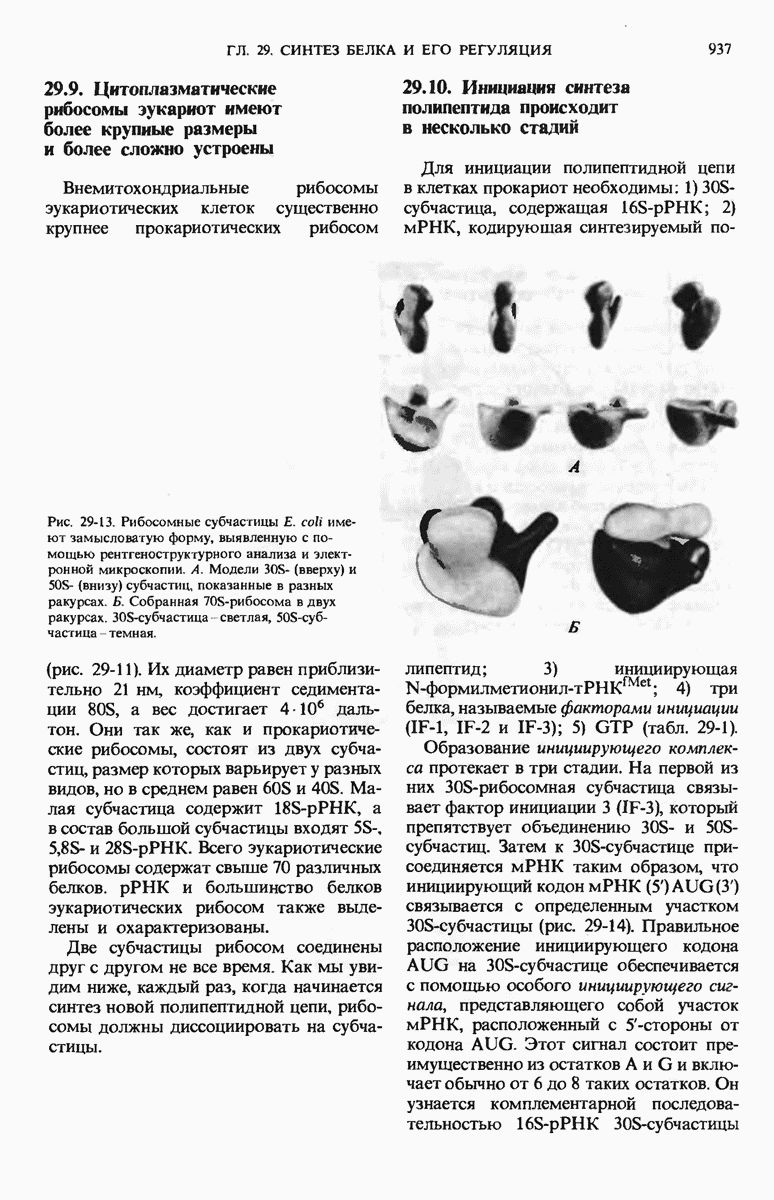
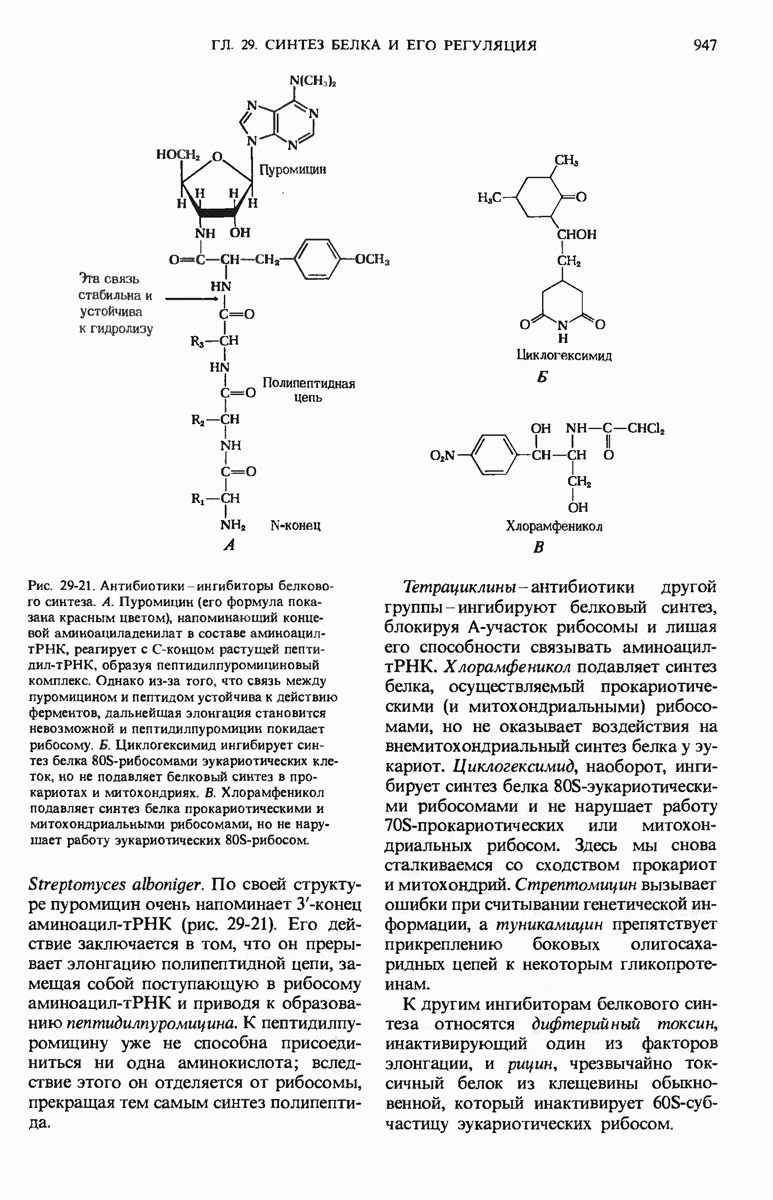
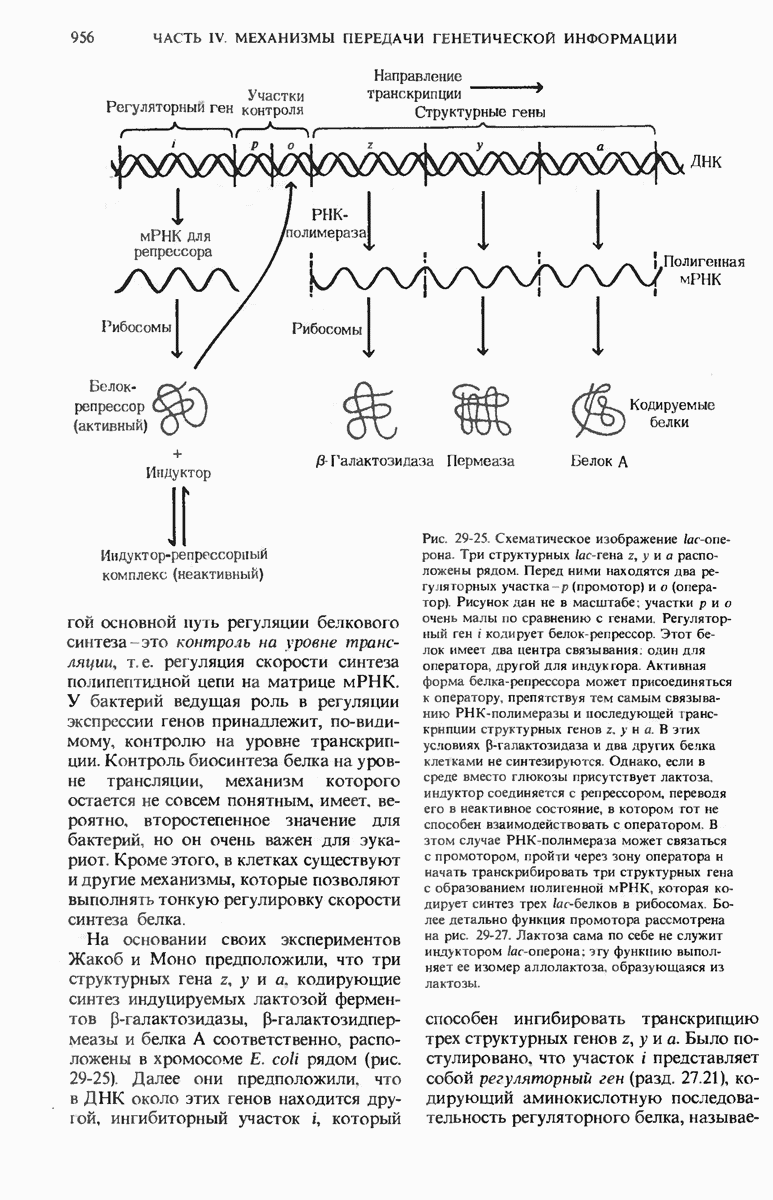
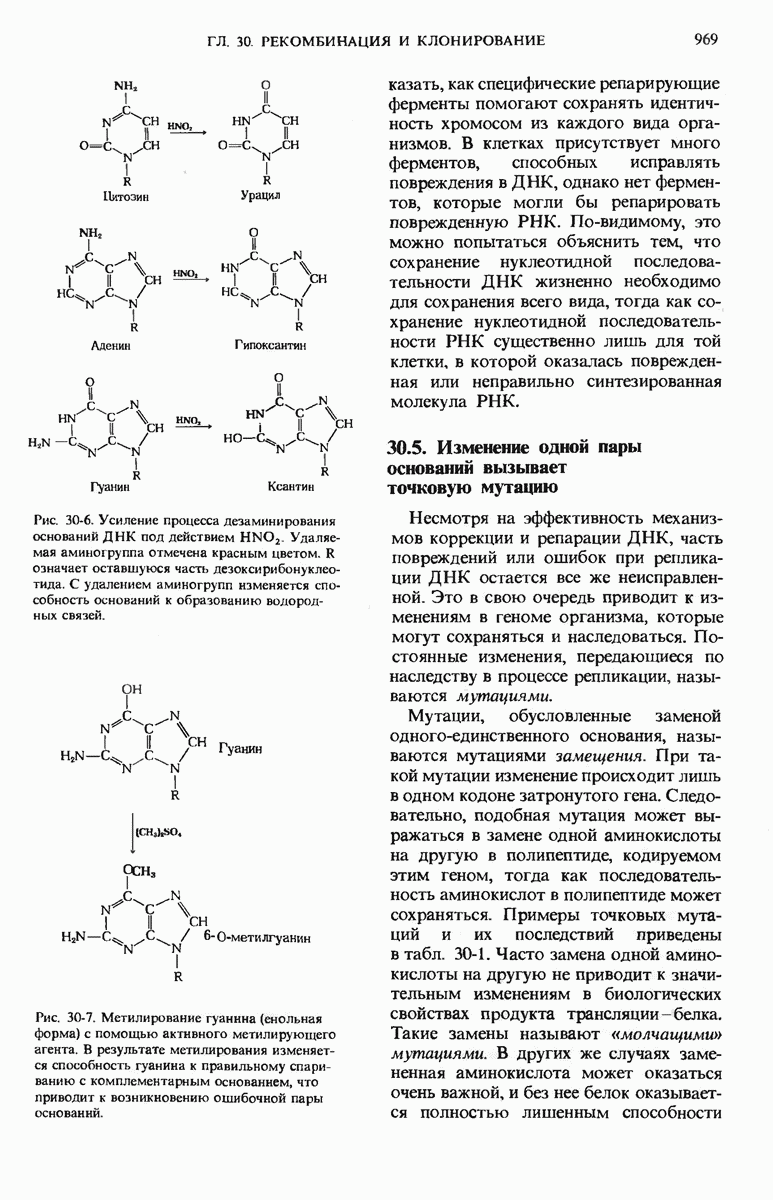
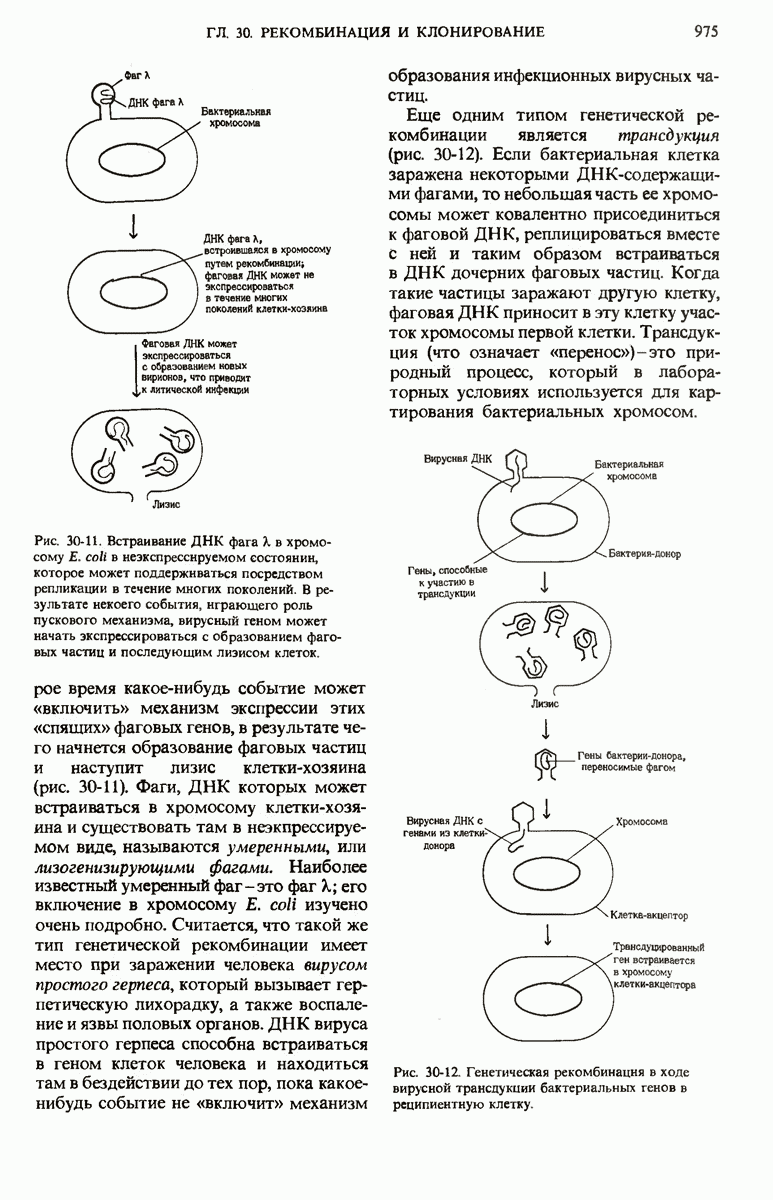
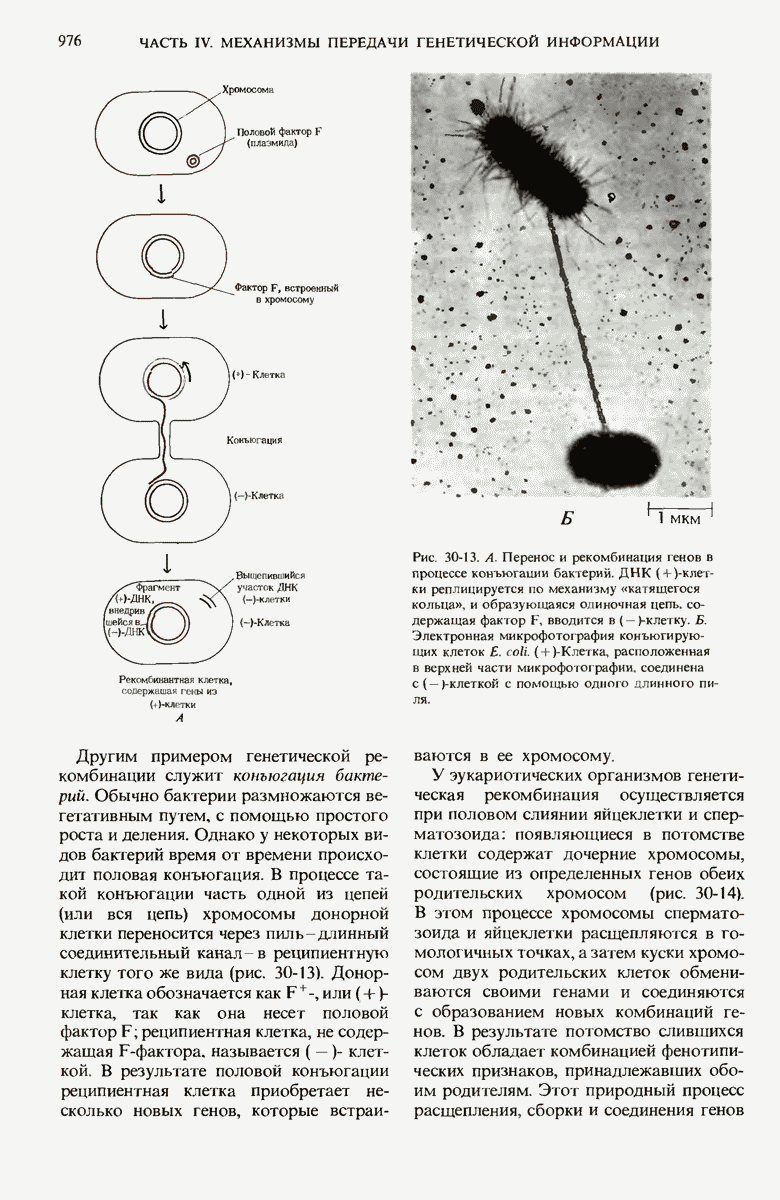
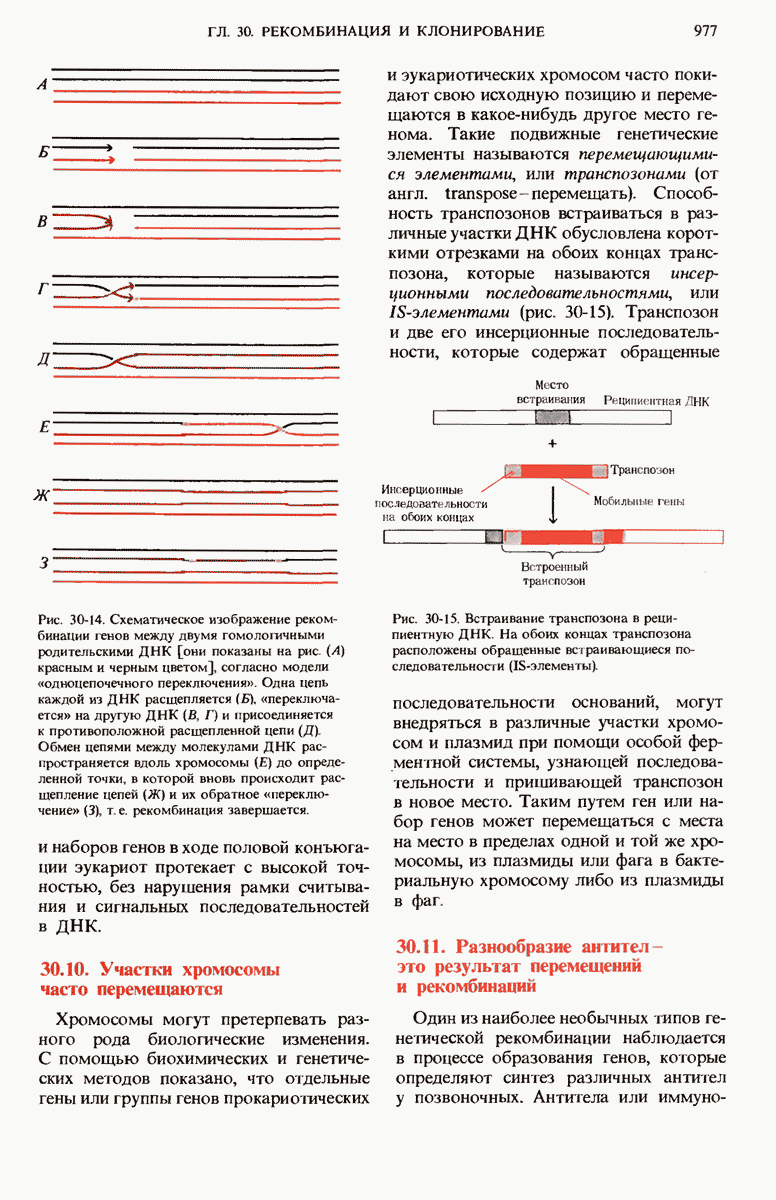
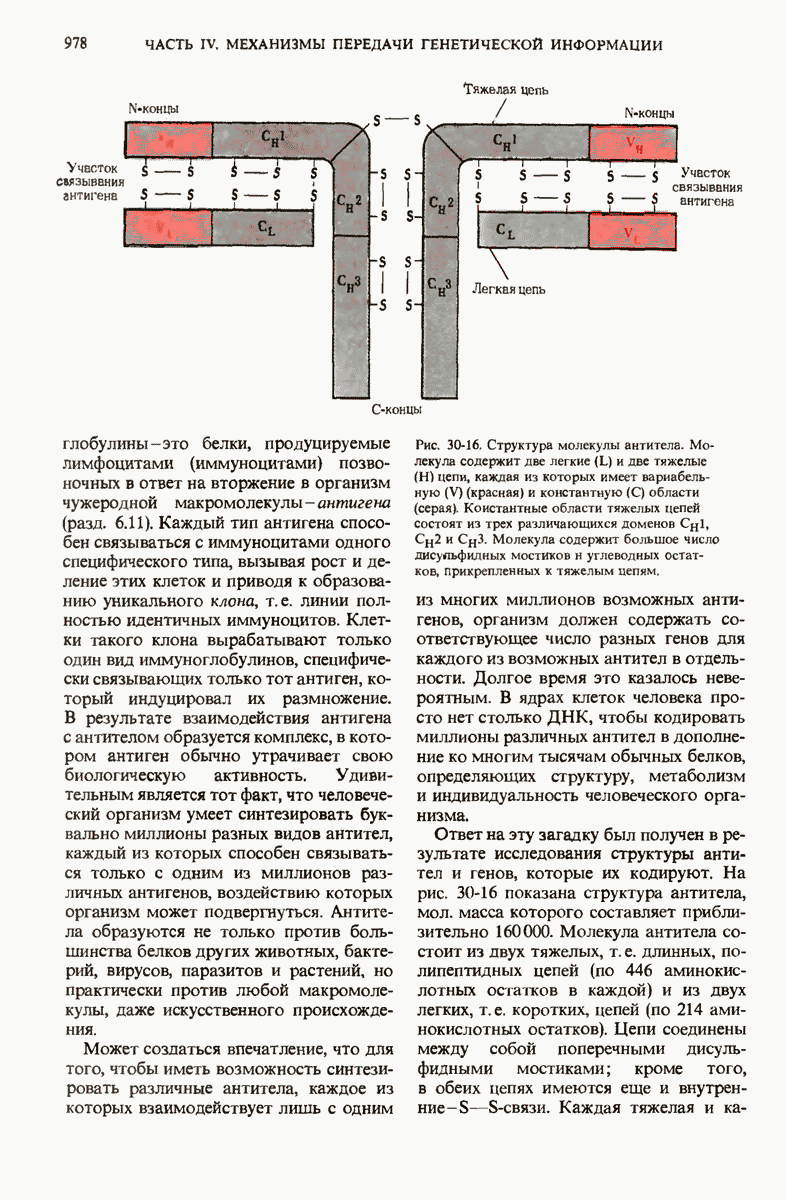
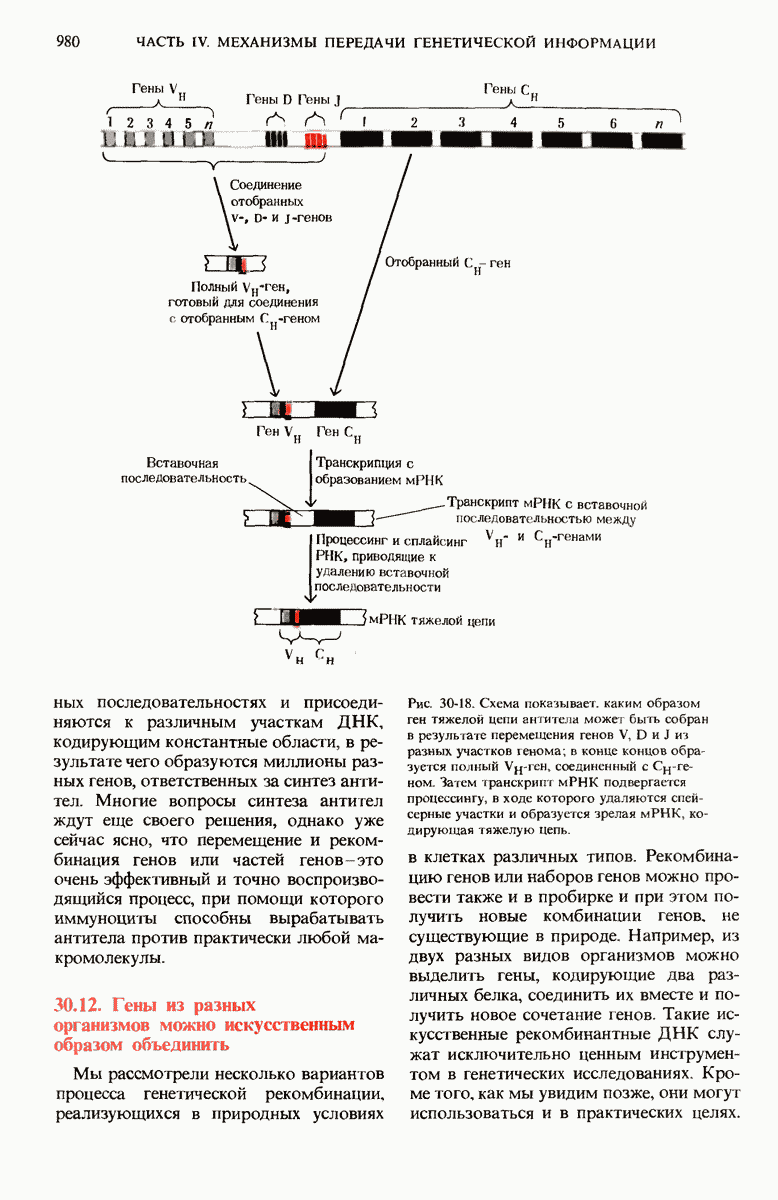
28.4. Иногда ДНК реплицируется по механизму «катящегося кольца»
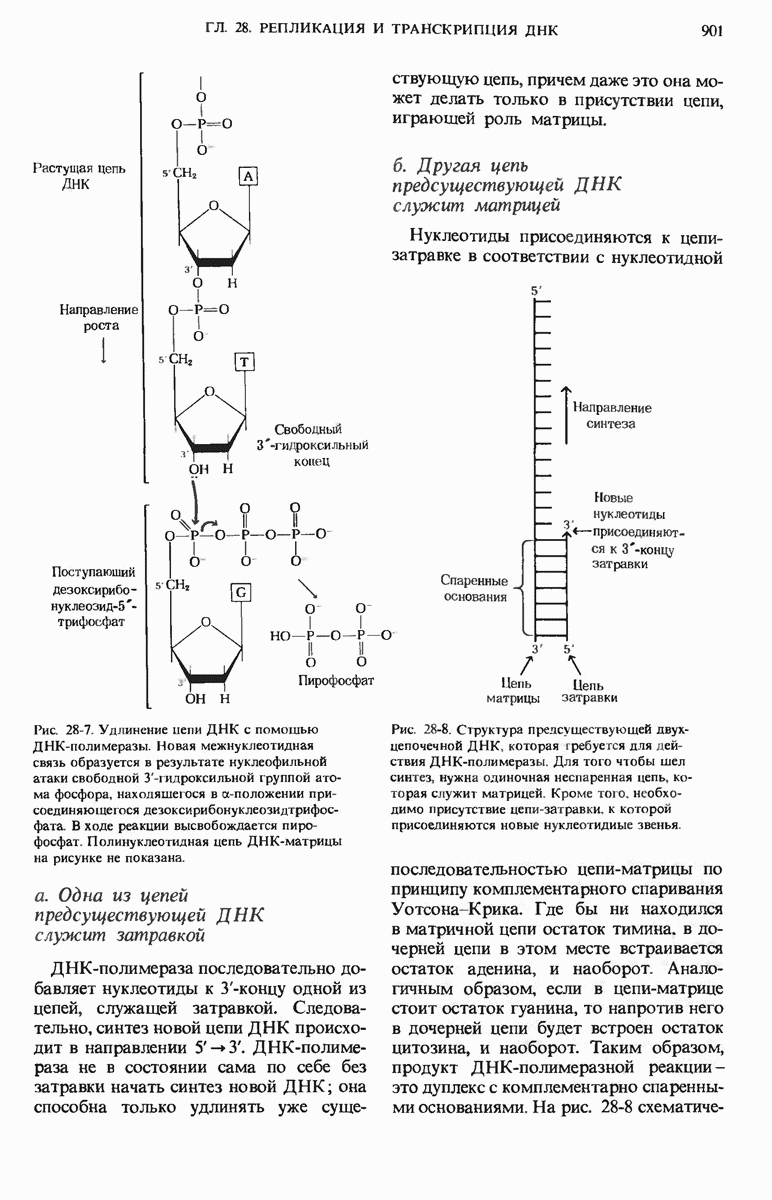
ДНК некоторых вирусов реплицируются в одном направлении по механизму «катящегося кольца», вариант которого представлен на рис. 28-5. Вначале одна из двух цепей кольцевой родительской ДНК расщепляется ферментом. Затем к З-концу расщепленной цепи присоединяется несколько новых нуклеотидов. Рост новой цепи на кольцевой матрице осуществляется за счет постепенного вытеснения 5-концевой части расщепленной цепи из катящейся кольцевой матрицы. По мере роста новой цепи вытесненный 5-хвост становится линейной матрицей для синтеза новой комплементарной цепи. Этот синтез на линейной матрице продолжается до тех пор, пока не образуется дочерняя цепь ДНК, комплементарная одному обороту кольцевой матрицы. Двухцепочечный хвост отщепляется затем с помощью фермента, и на 5-конце опять может начинаться процесс репликации. Таким путем с кольцевой матрицы может сходить множество комплементарных копий кольцевой ДНК. Механизм «катящегося кольца» используется в ооцитах в процессе синтеза генов рРНК; он позволяет получать большое число копий этих генов, расположенных в тандемной последовательности, что в свою очередь дает возможность синтезировать одновременно много рРНК. Этот механизм необходим ооцитам для того, чтобы производить много рибосом для быстрого синтеза клеточных белков в процессе ускоренного роста эмбриона на ранних стадиях развития.

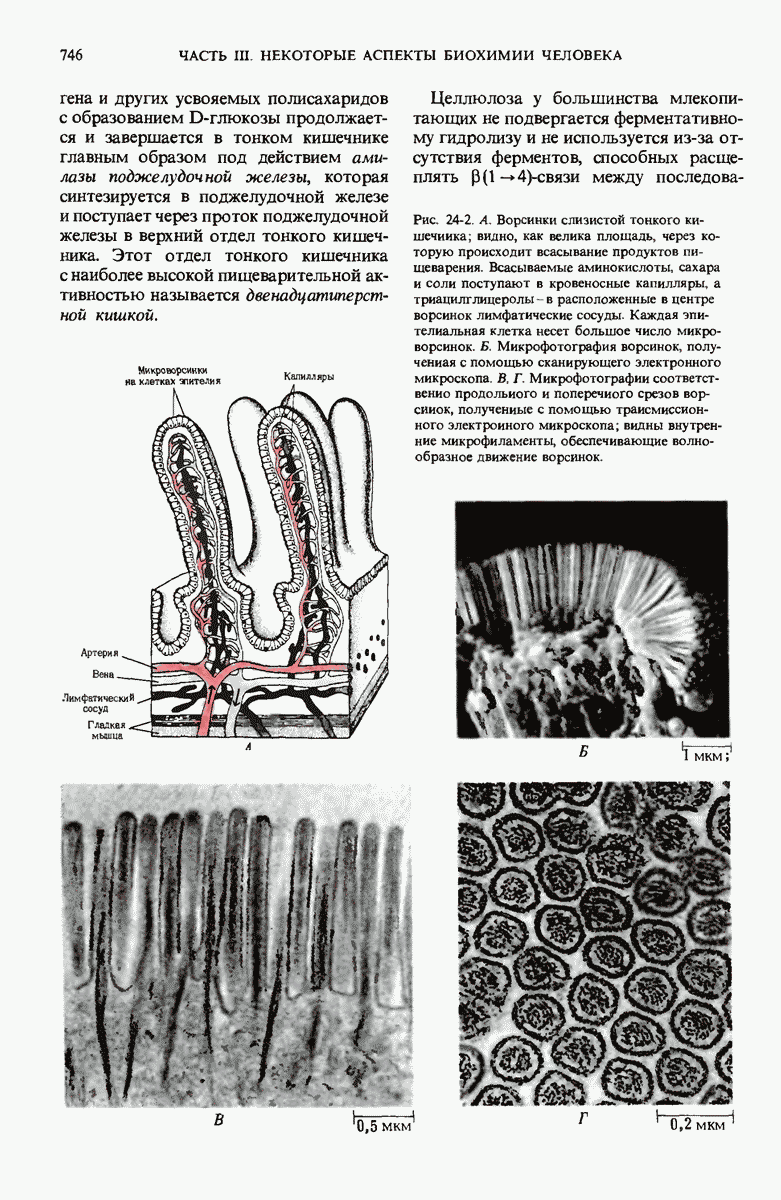
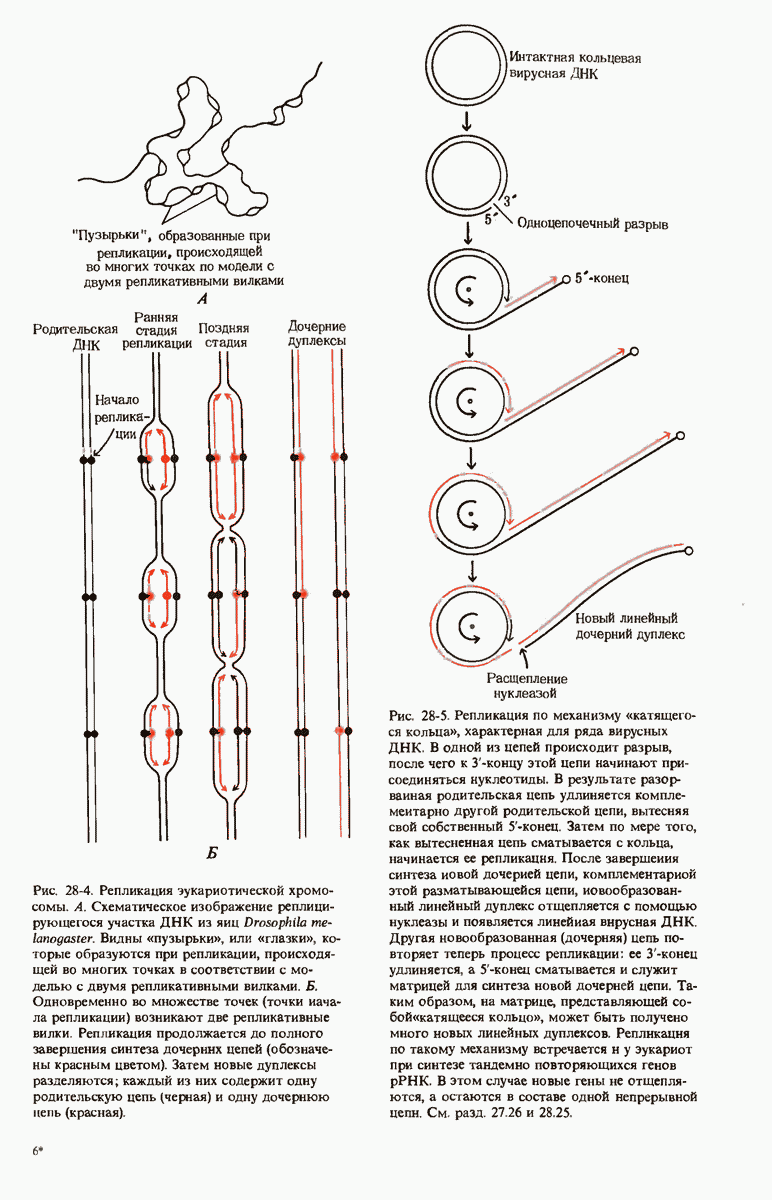
Рис. 28-4. Репликация эукариотической хромосомы. А. Схематическое изображение реплицирующегося участка ДНК из яиц Drosophila melanogaster. Видны «пузырьки», или «глазки», ко торые образуются при репликации, происходящей во многих точках в соответствии с моделью с двумя репликативными вилками. Б. Одновременно во множестве точек (точки начала репликации) возникают две репликативные вилки. Репликация продолжается до полного завершения синтеза дочерних цепей (обозначены красным цветом). Затем новые дуплексы разделяются; каждый из них содержит одну родительскую цепь (черная) и одну дочернюю цепь (красная).

Рис. 28-5. Репликация по механизму «катящегося кольца», характерная для ряда вирусных ДНК. В одной из цепей происходит разрыв, после чего к З-концу этой цепи начинают присоединяться нуклеотиды. В результате разорванная родительская цепь удлиняется комплементарно другой родительской цепи, вытесняя свой собственный 5-конец. Затем по мере того, как вытесненная цепь сматывается с кольца, начинается ее репликация. После завершения синтеза новой дочерней цепи, комплементарной этой разматывающейся цепи, новообразованный линейный дуплекс отщепляется с помощью нуклеазы и появляется линейная вирусная ДНК. Другая новообразованная (дочерняя) цепь повторяет теперь процесс репликации: ее 3-конец удлиняется, а 5-конец сматывается и служит матрицей для синтеза новой дочерней цепи. Таким образом, на матрице, представляющей собой «катящееся кольцо», может быть получено много новых линейных дуплексов. Репликация по такому механизму встречается и у эукариот при синтезе тандемно повторяющихся генов рРНК. В этом случае новые гены не отщепляются, а остаются в составе одной непрерывной цепн. См. разд. 27.26 и 28.25.
28.5. Бактериальные экстракты содержат ДНК-полимеразу
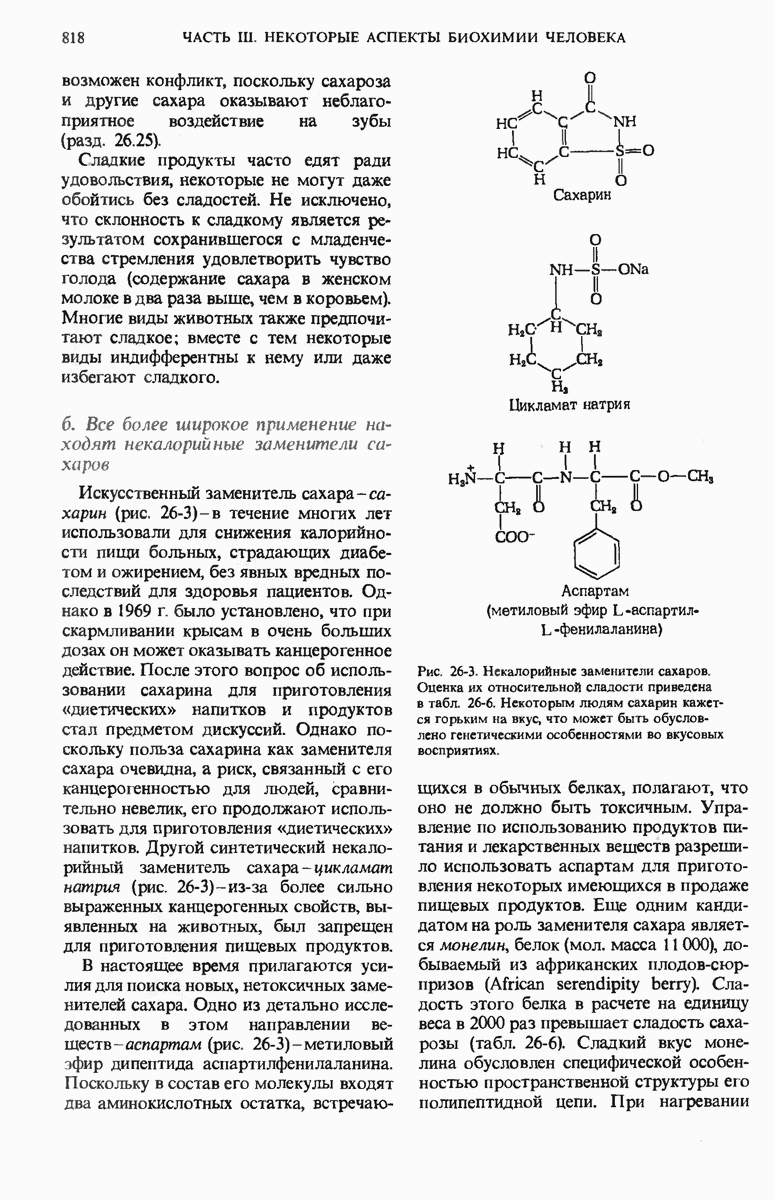
Теперь, когда мы получили общее представление о репликации ДНК, рассмотрим вопрос об участии ферментов в этом процессе. Впервые механизмы действия ферментов при репликации стали доступны прямому биохимическому исследованию благодаря важным работам Артура Корнберга и его коллег, начатым в 1956 г. Они инкубировали экстракты из клеток Е. coli со смесью dATP, dTTP, dGTP и dCTP,  -фосфатная группа которых была помечена изотопом
-фосфатная группа которых была помечена изотопом  (рис. 28-6).
(рис. 28-6).

Рис. 28-6. Дезоксирибонуклеозид-5-трифосфат, меченный  в
в  -положении.
-положении.
Было обнаружено, что при этом синтезируется очень небольшое количество новой ДНК, содержащей в своих фосфатных группах изотоп  Фермент, катализирующий эту реакцию и названный ДНК-полимеразой , был в конце концов очищен и его свойства были подробно изучены. Оказалось, что он катализирует последовательное присоединение дезоксирибонуклеотидных остатков к концу цепи ДНК с одновременным высвобождением неорганического пирофосфата, содержащего
Фермент, катализирующий эту реакцию и названный ДНК-полимеразой , был в конце концов очищен и его свойства были подробно изучены. Оказалось, что он катализирует последовательное присоединение дезоксирибонуклеотидных остатков к концу цепи ДНК с одновременным высвобождением неорганического пирофосфата, содержащего  -фосфатные группы каждого встраивающегося дезоксирибонуклеозид-5-трифосфата. Уравнение реакции в простейшей форме имеет вид
-фосфатные группы каждого встраивающегося дезоксирибонуклеозид-5-трифосфата. Уравнение реакции в простейшей форме имеет вид

где dNMP и dNTP означают соответственно дезоксирибонуклеозид-5-монофосфат и дезоксирибонуклеозид-5-трифосфат. Если хотя бы один из четырех предшественников отсутствует, то новая ДНК не образуется, т.е. синтез новой ДНК идет только в присутствии всех четырех предшественников. 5-трифосфаты всех четырех дезоксирибонуклеозидов не могут быть заменены соответствующими 5-дифосфатами или 5-монофосфа-тами; фермент не работает также с рибонуклеозид-5-трифосфатами. Для работы ДНК-полимеразе необходимы ионы  а в ее активном центре содержится прочно связанный с ферментом ион
а в ее активном центре содержится прочно связанный с ферментом ион 
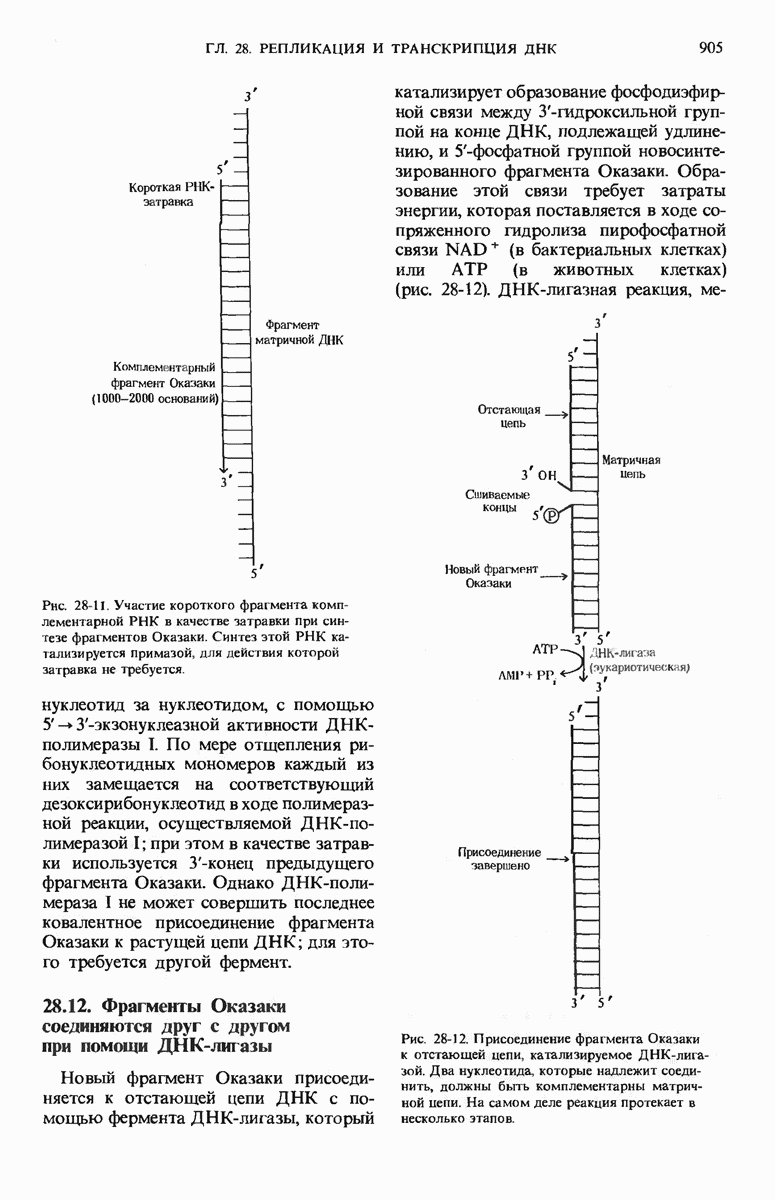
ДНК-полимераза катализирует ковалентное связывание новых дезоксирибонуклеотидов, которое происходит благодаря присоединению их  -фосфатных групп к свободному З-гидроксильному концу предсуществующей цепи ДНК; следовательно, синтез цепи ДНК происходит в направлении
-фосфатных групп к свободному З-гидроксильному концу предсуществующей цепи ДНК; следовательно, синтез цепи ДНК происходит в направлении  (рис. 28-7). Энергия, затрачиваемая на образование каждой новой фосфодиэфирной связи в остове ДНК, обеспечивается расщеплением пирофосфатной связи между
(рис. 28-7). Энергия, затрачиваемая на образование каждой новой фосфодиэфирной связи в остове ДНК, обеспечивается расщеплением пирофосфатной связи между  и
и  -фосфатными группами предшественников - дезоксирибонуклеозид-5-трифосфатов. Образующийся при этом пирофосфат разрушается затем до фосфата, который может сдвигать реакцию в сторону ее завершения. В процессе работы было сделано очень важное наблюдение: было отмечено, что ДНК-полимеразная реакция протекает только в том случае, если в системе уже находится некоторое количество предсуществующей двухцепочечной ДНК.
-фосфатными группами предшественников - дезоксирибонуклеозид-5-трифосфатов. Образующийся при этом пирофосфат разрушается затем до фосфата, который может сдвигать реакцию в сторону ее завершения. В процессе работы было сделано очень важное наблюдение: было отмечено, что ДНК-полимеразная реакция протекает только в том случае, если в системе уже находится некоторое количество предсуществующей двухцепочечной ДНК.