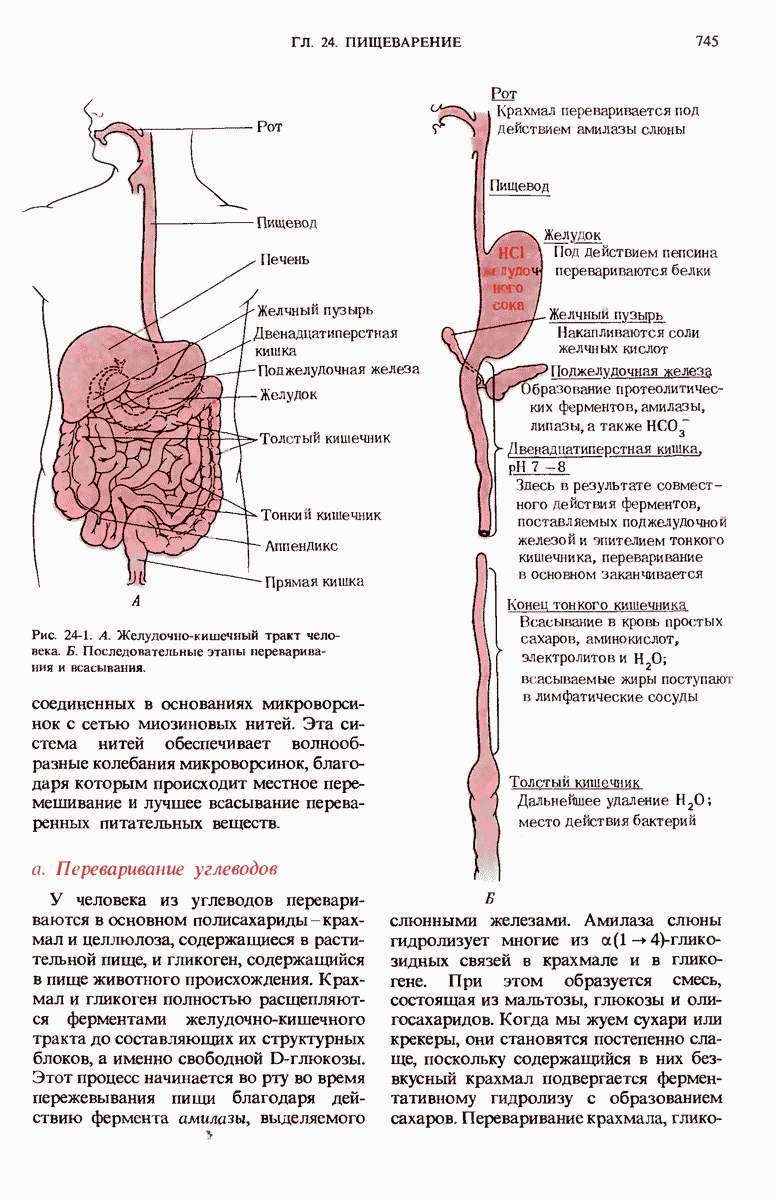
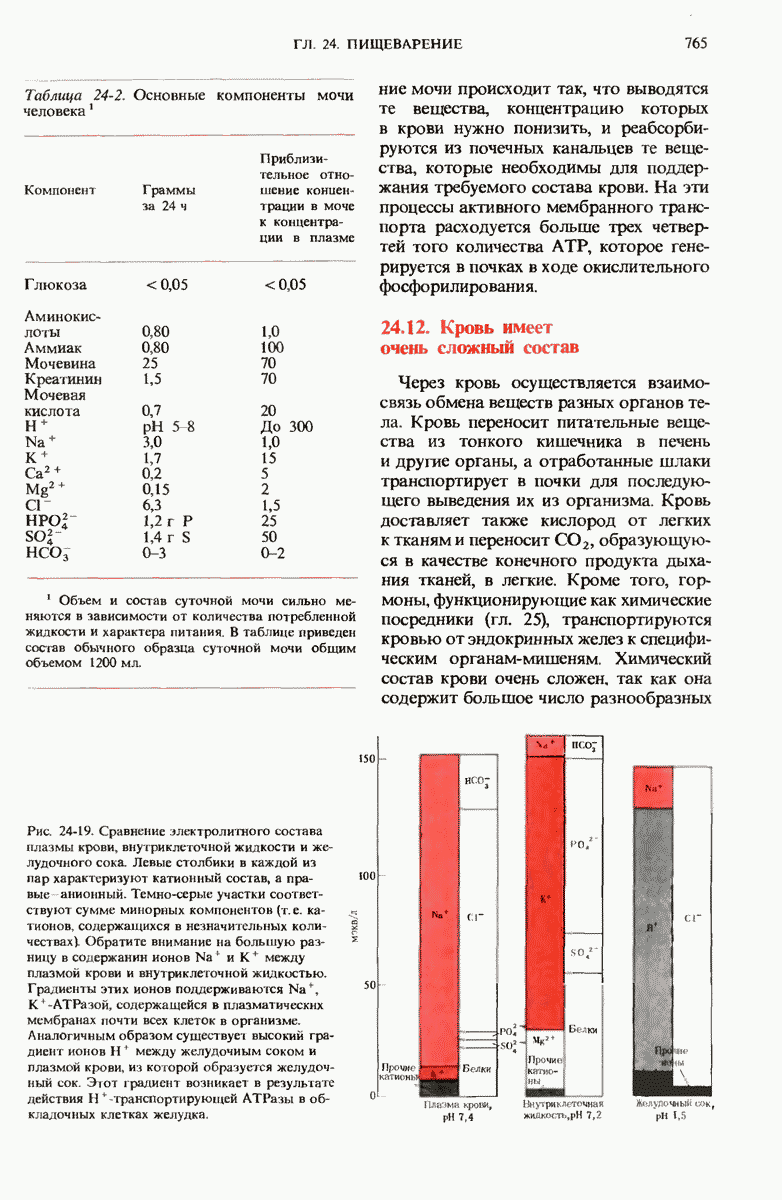
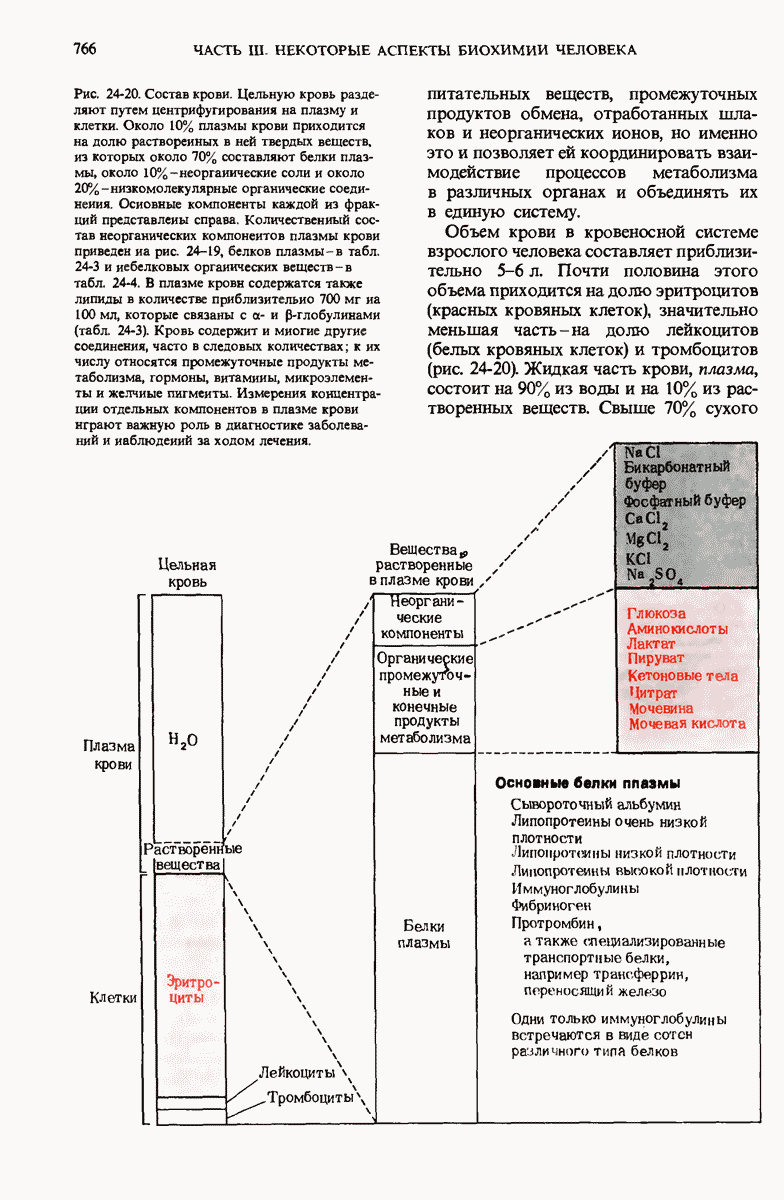
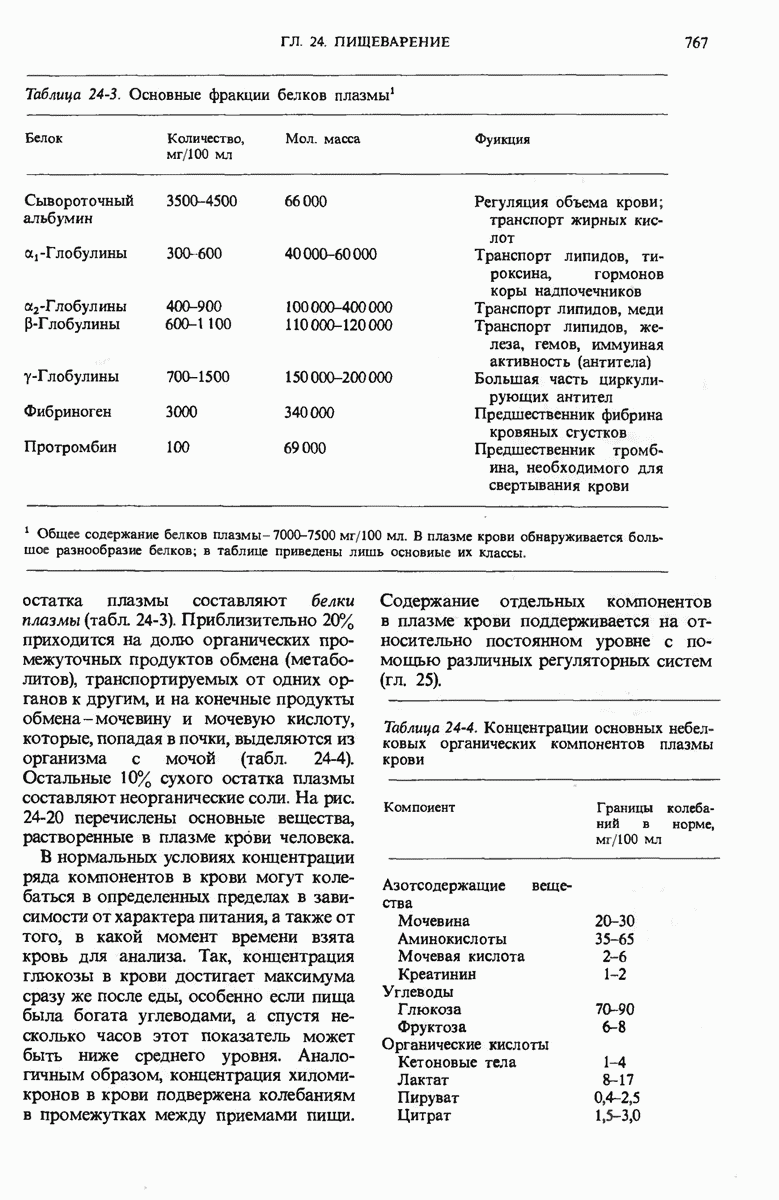
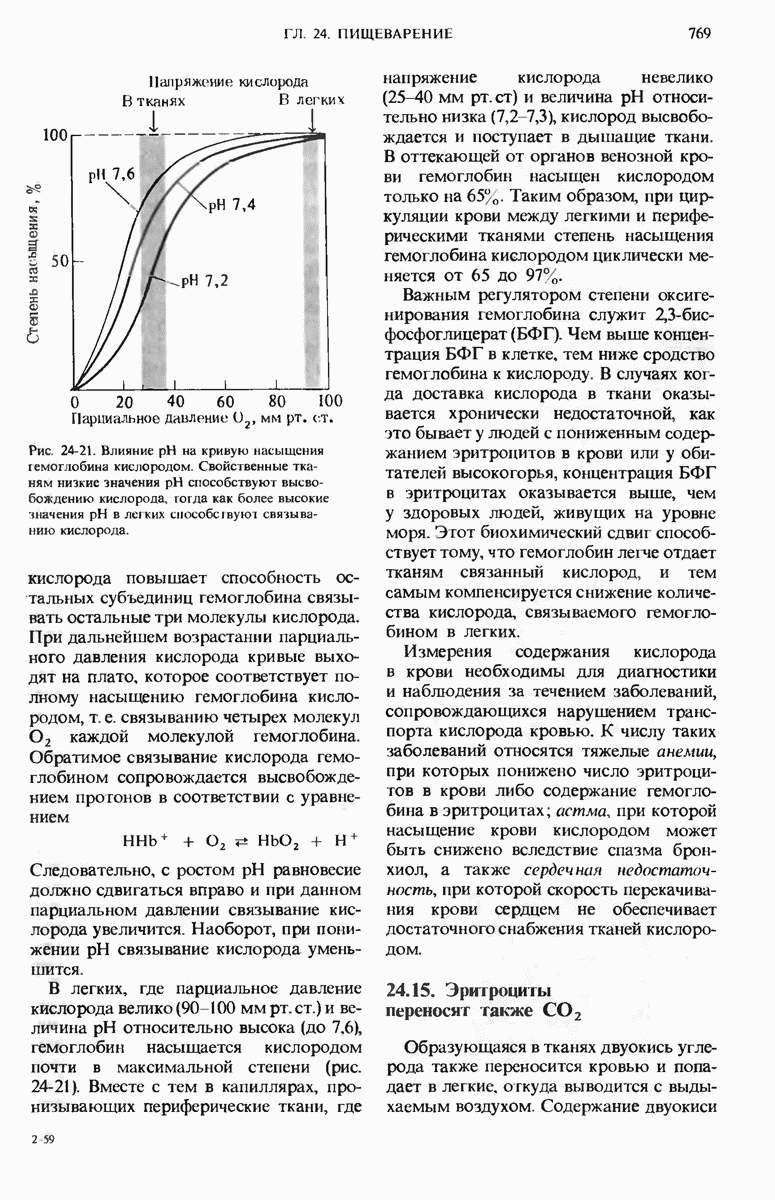
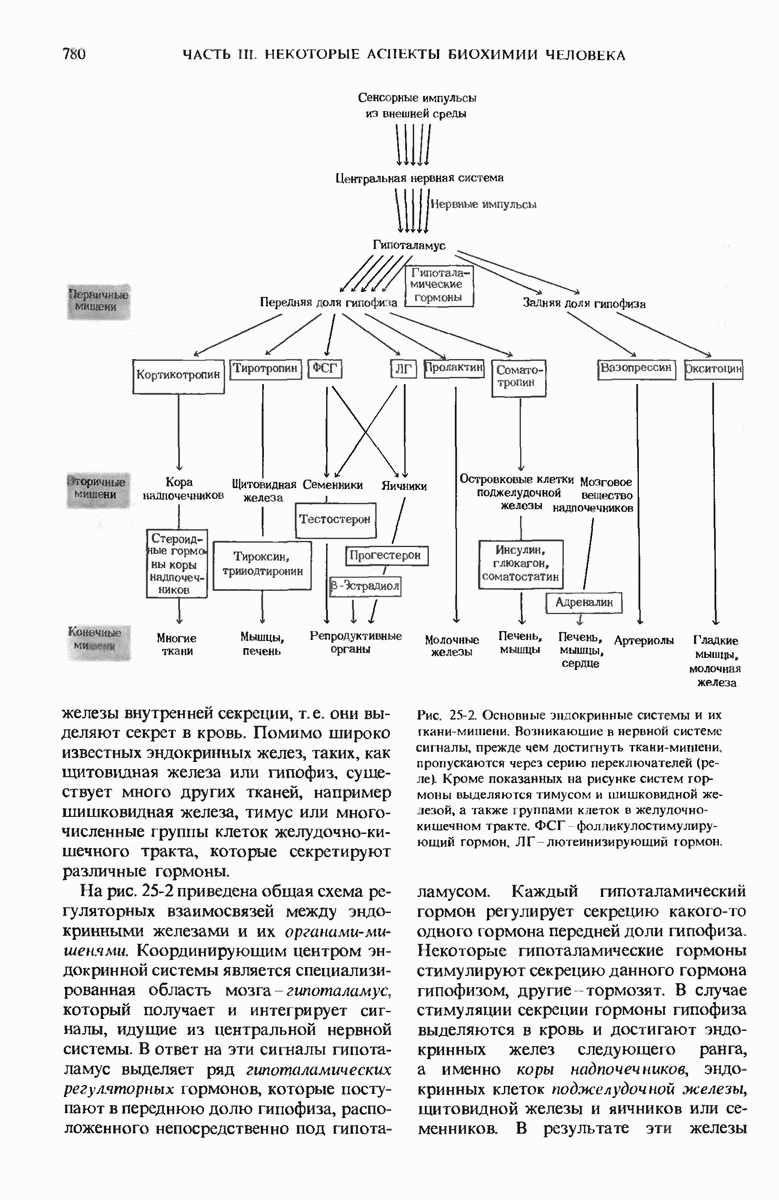
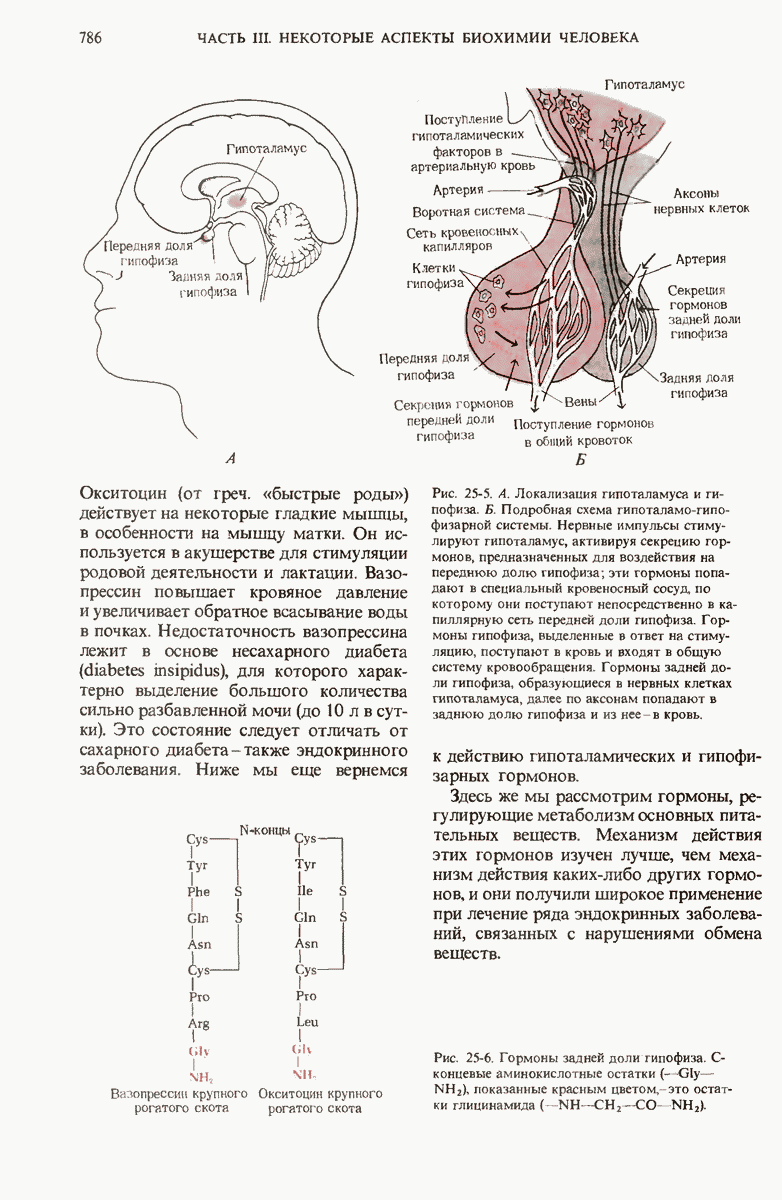
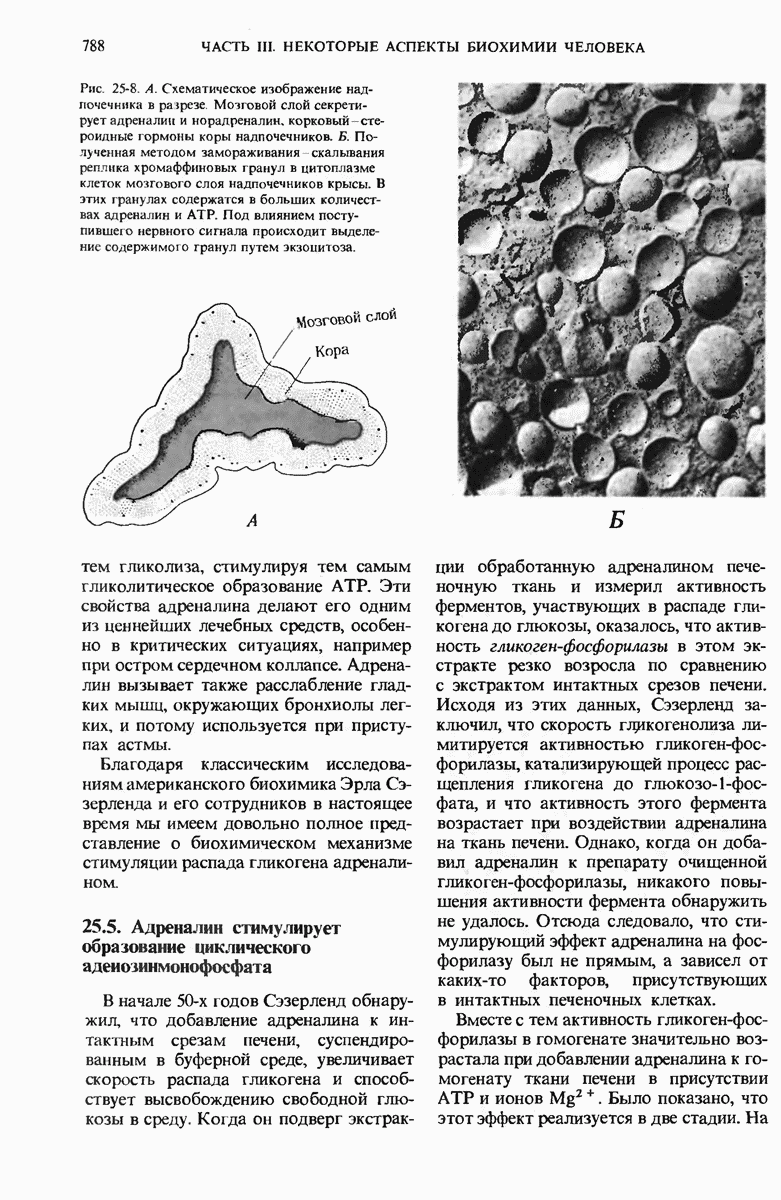
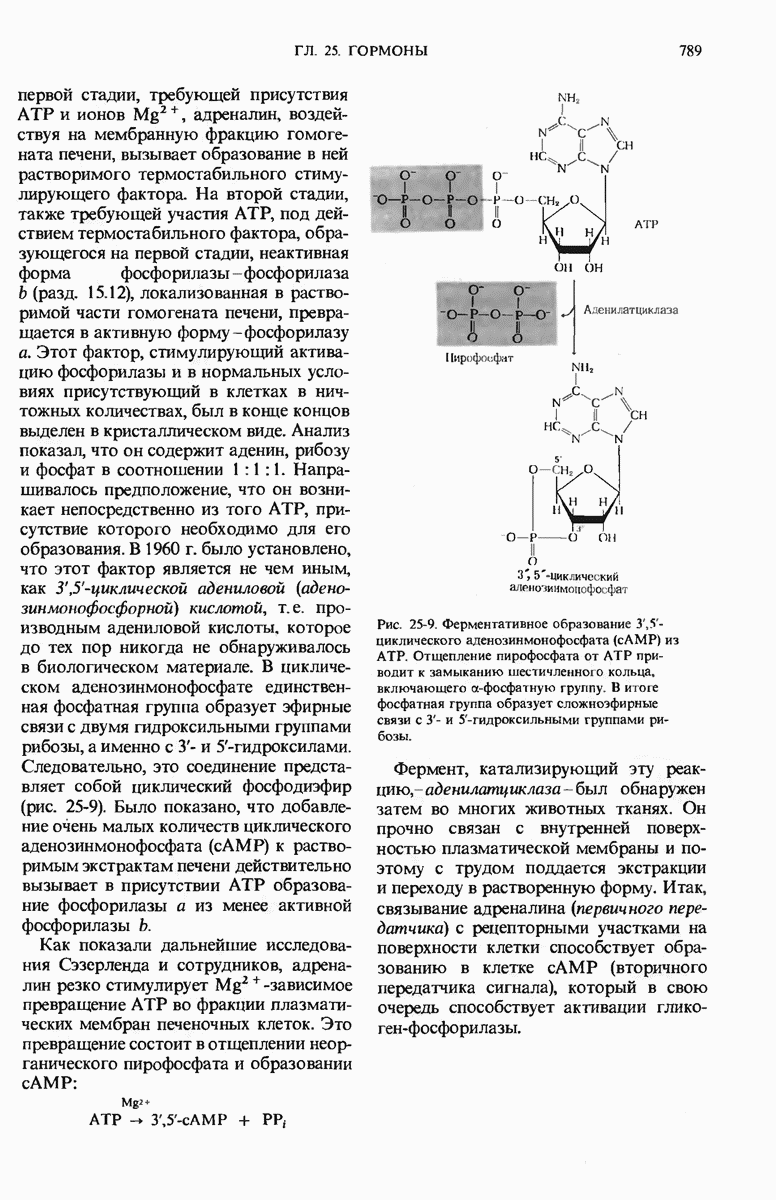
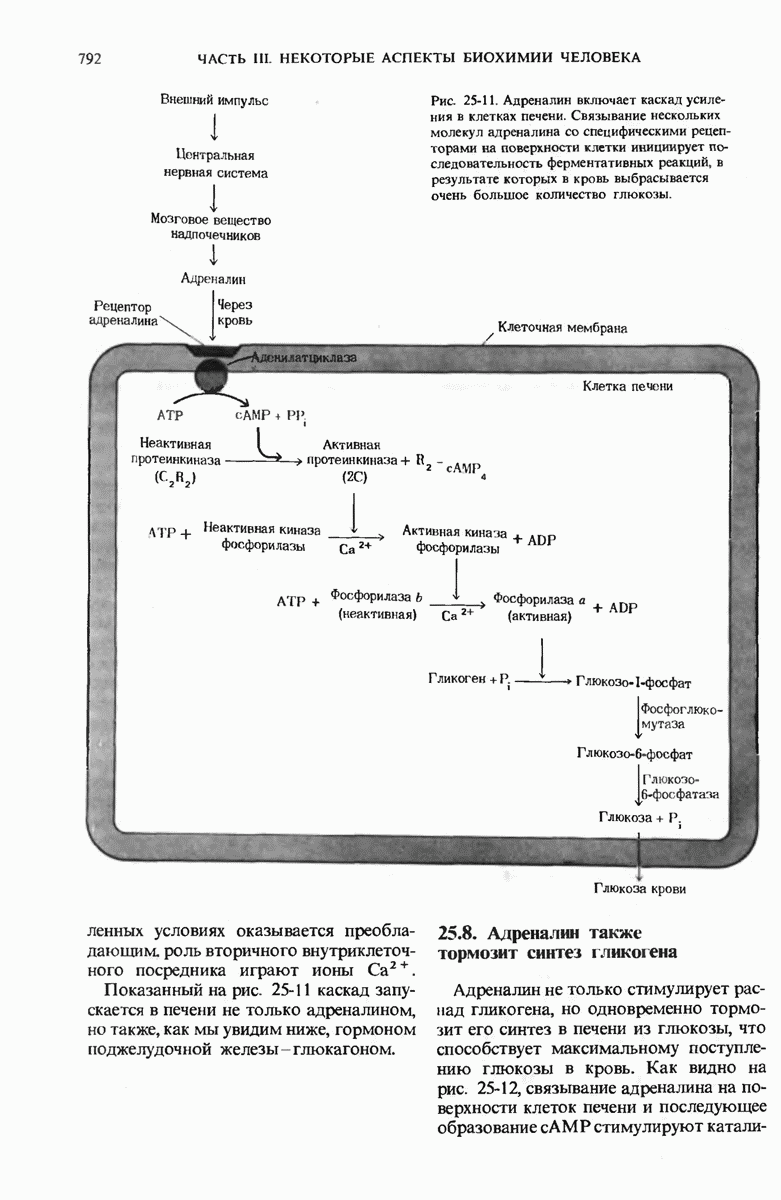
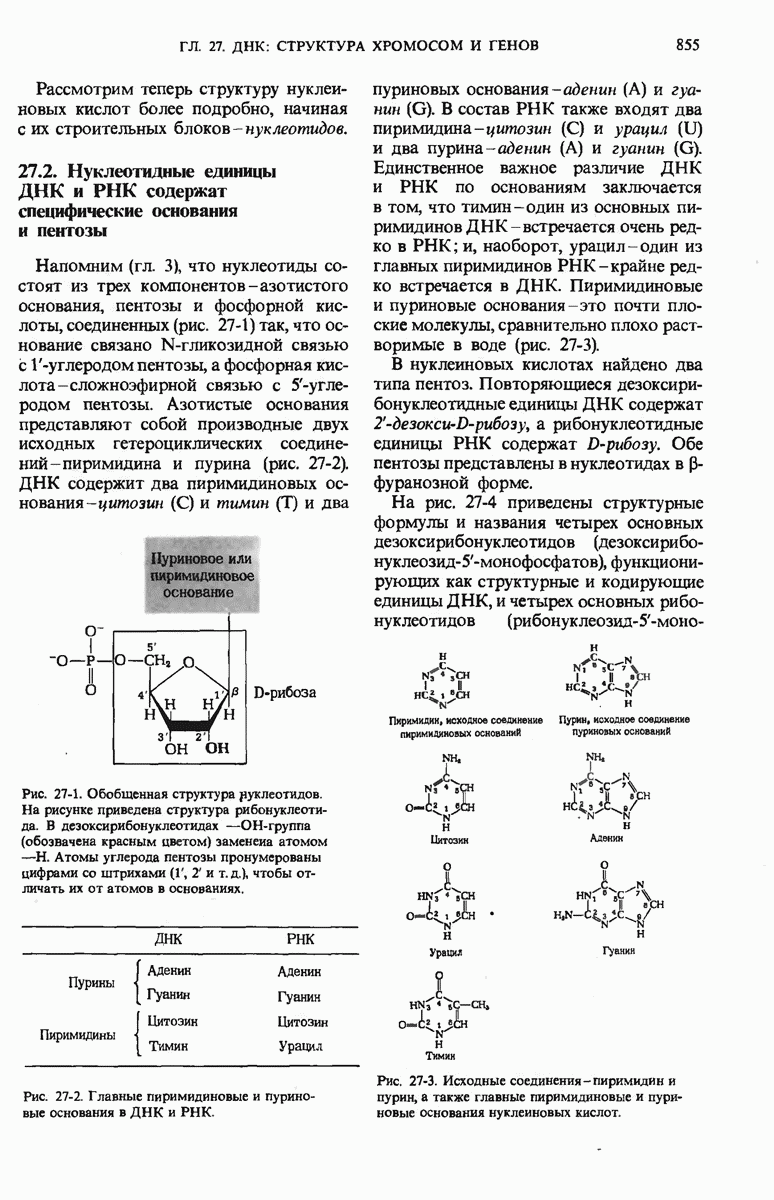
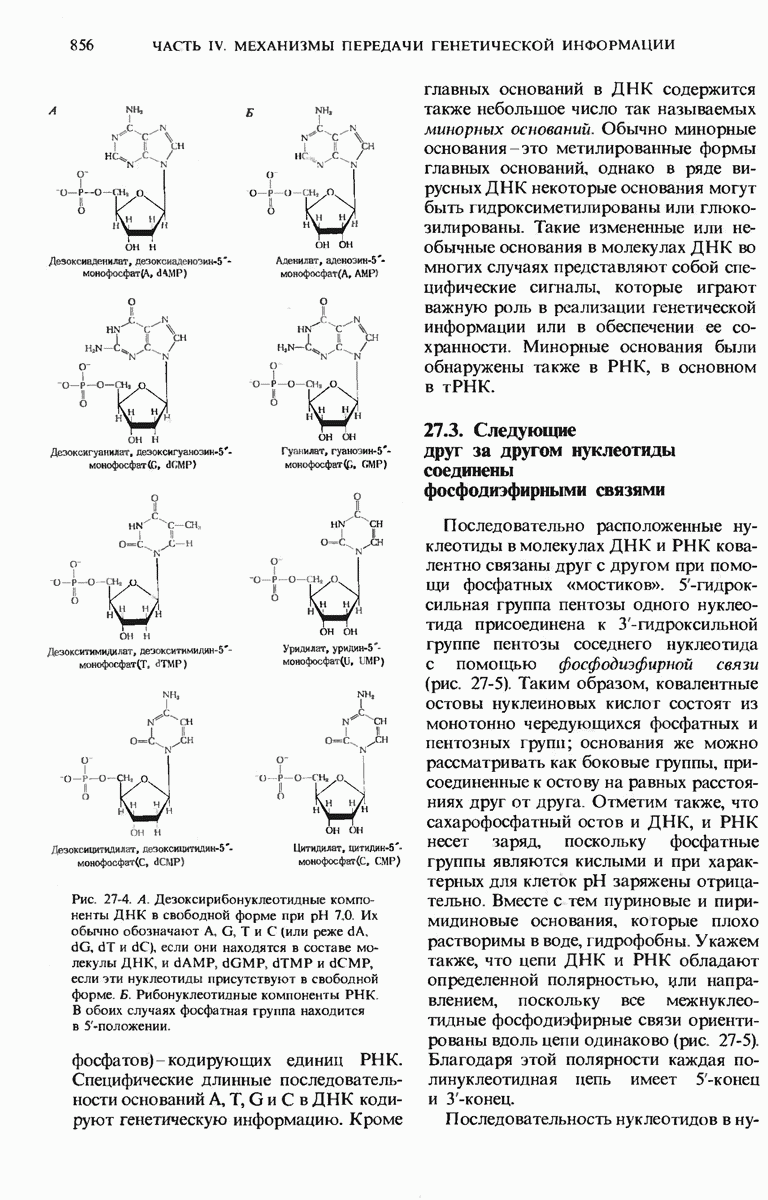
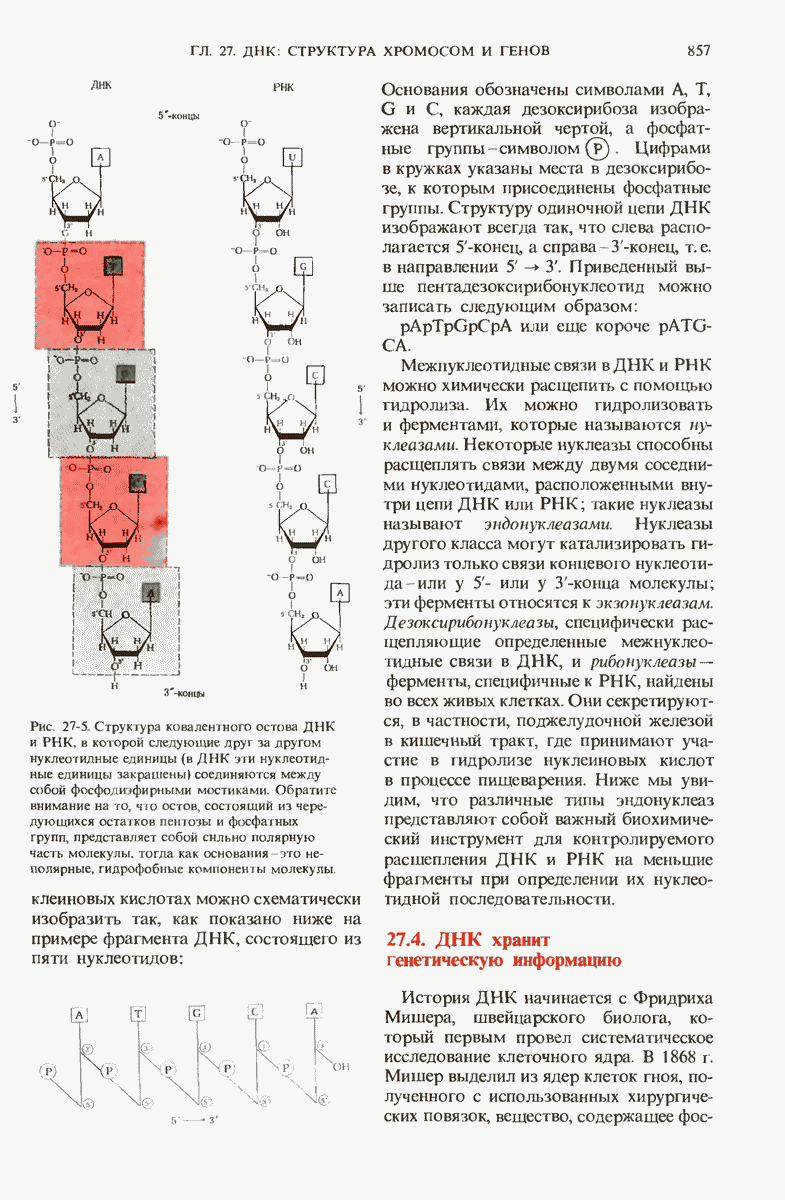
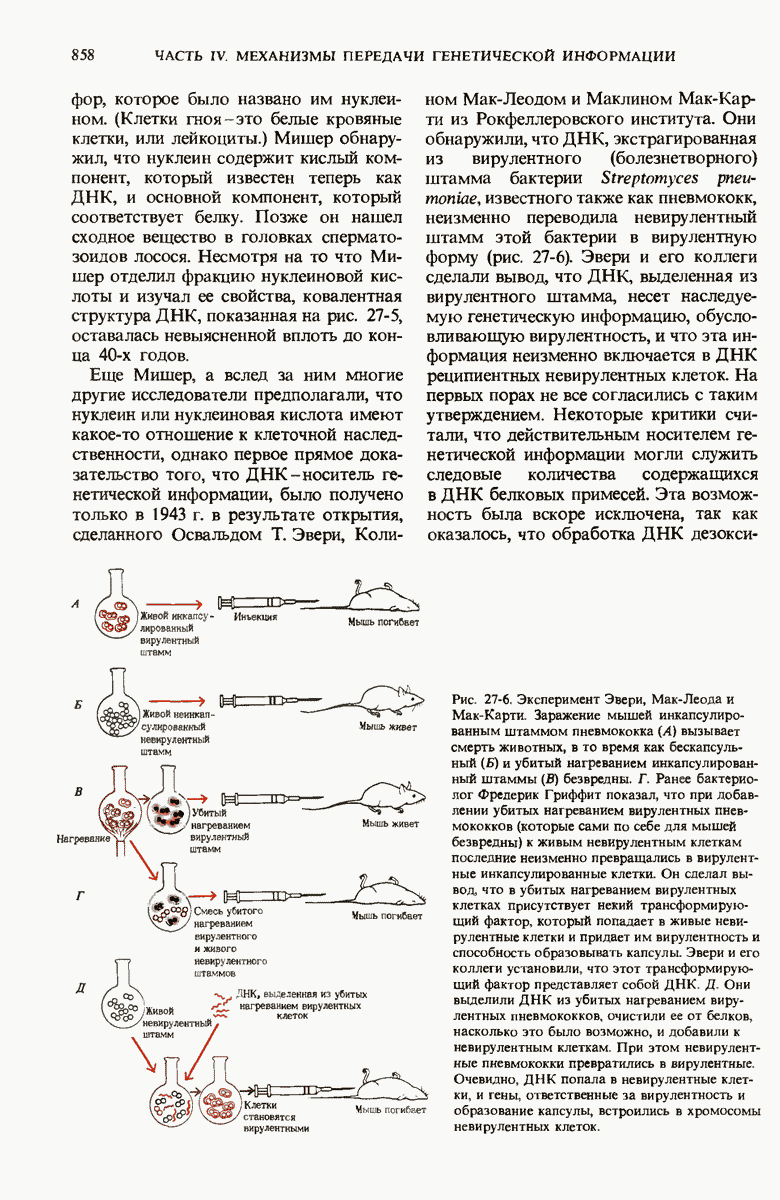
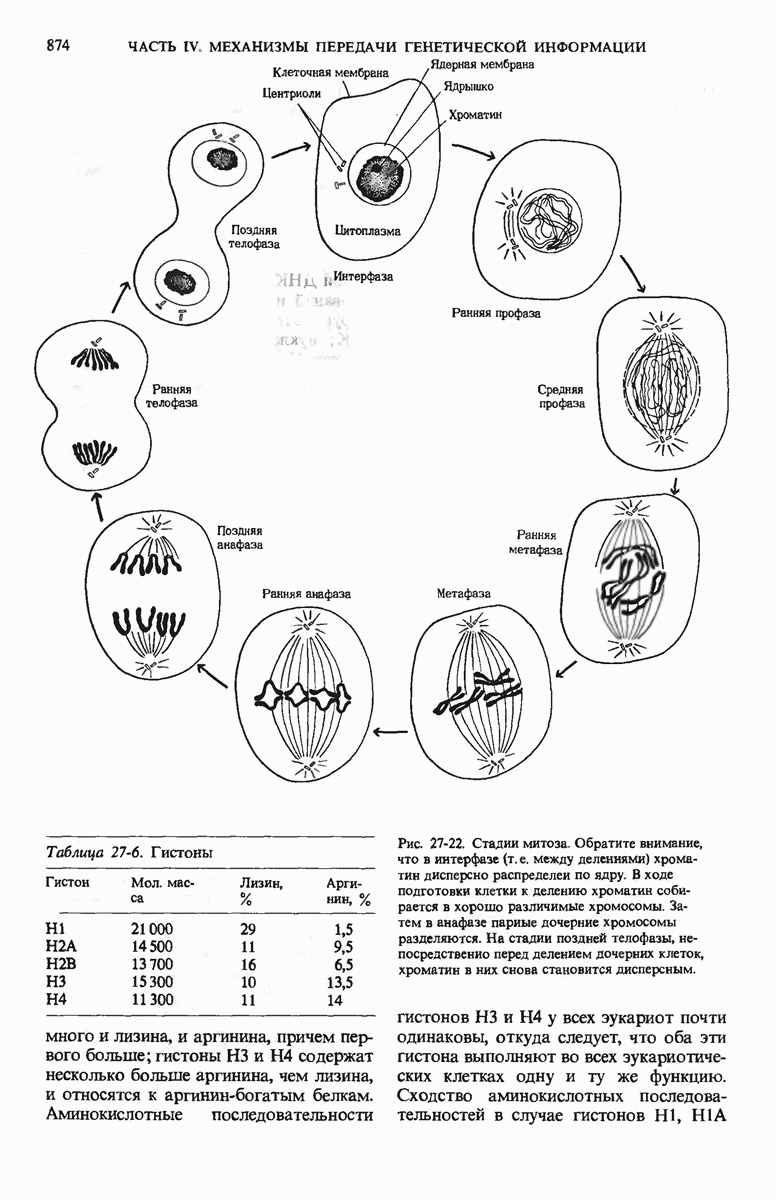
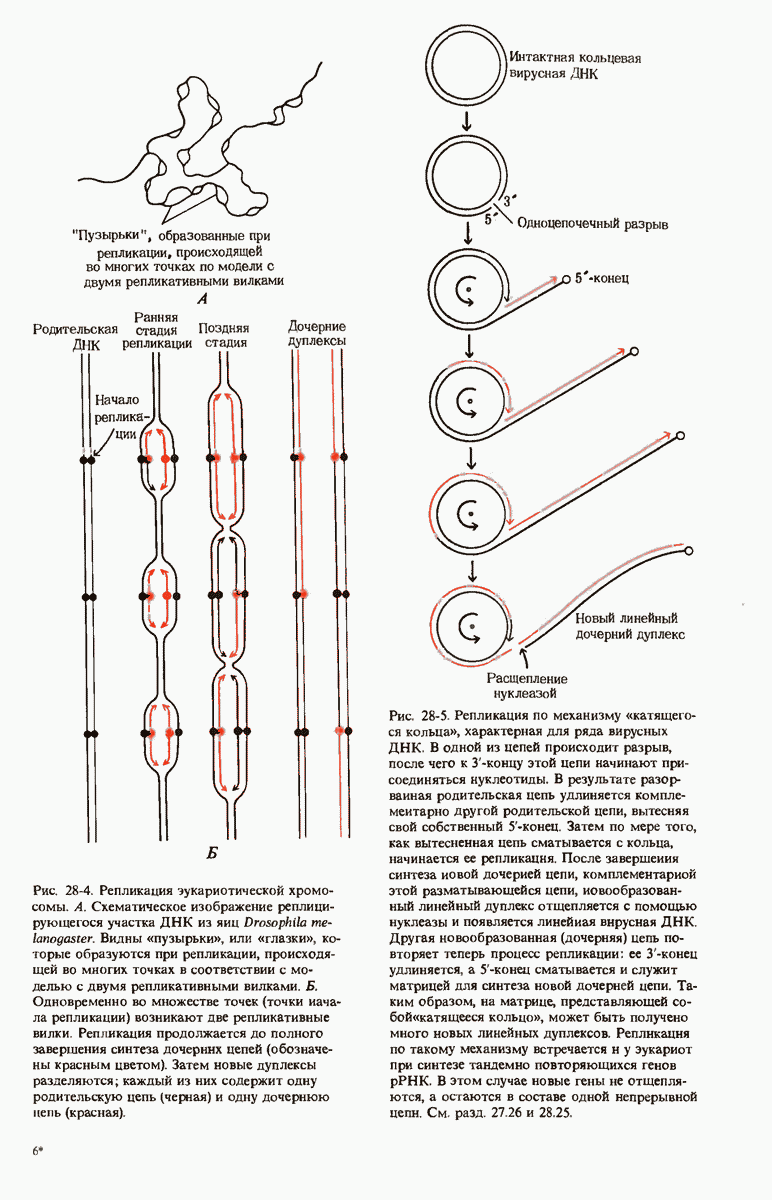
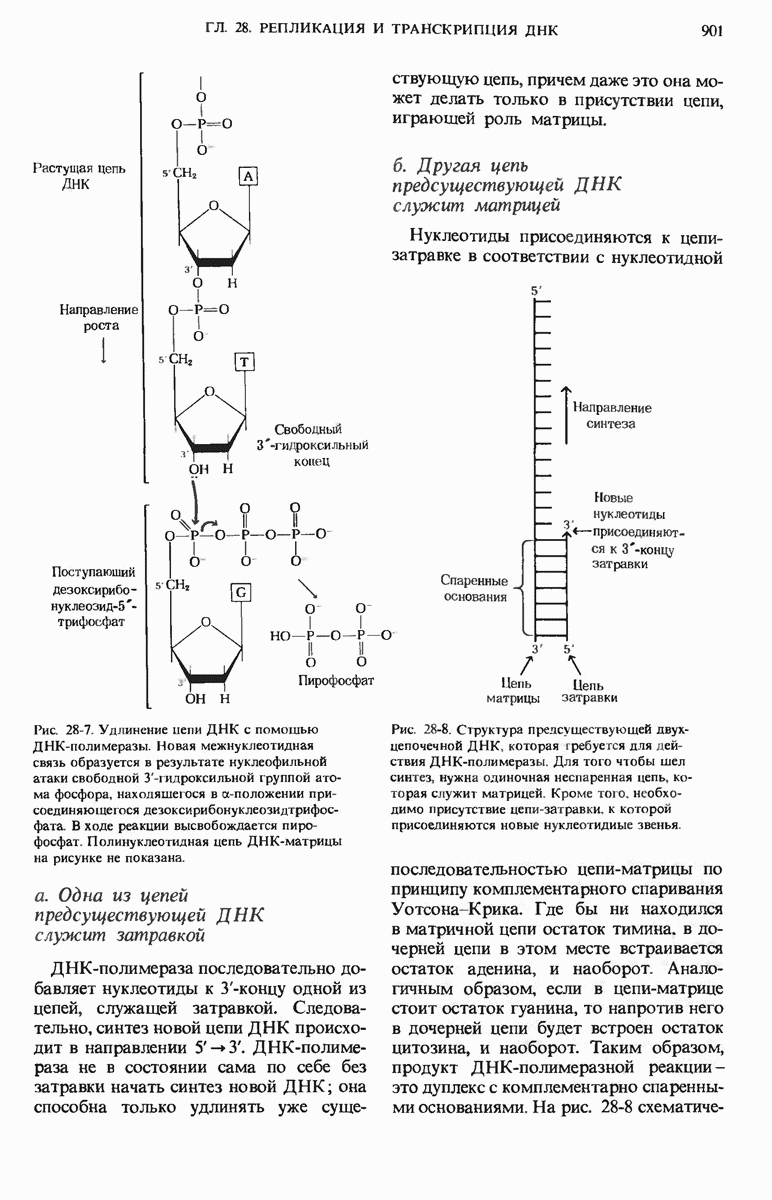
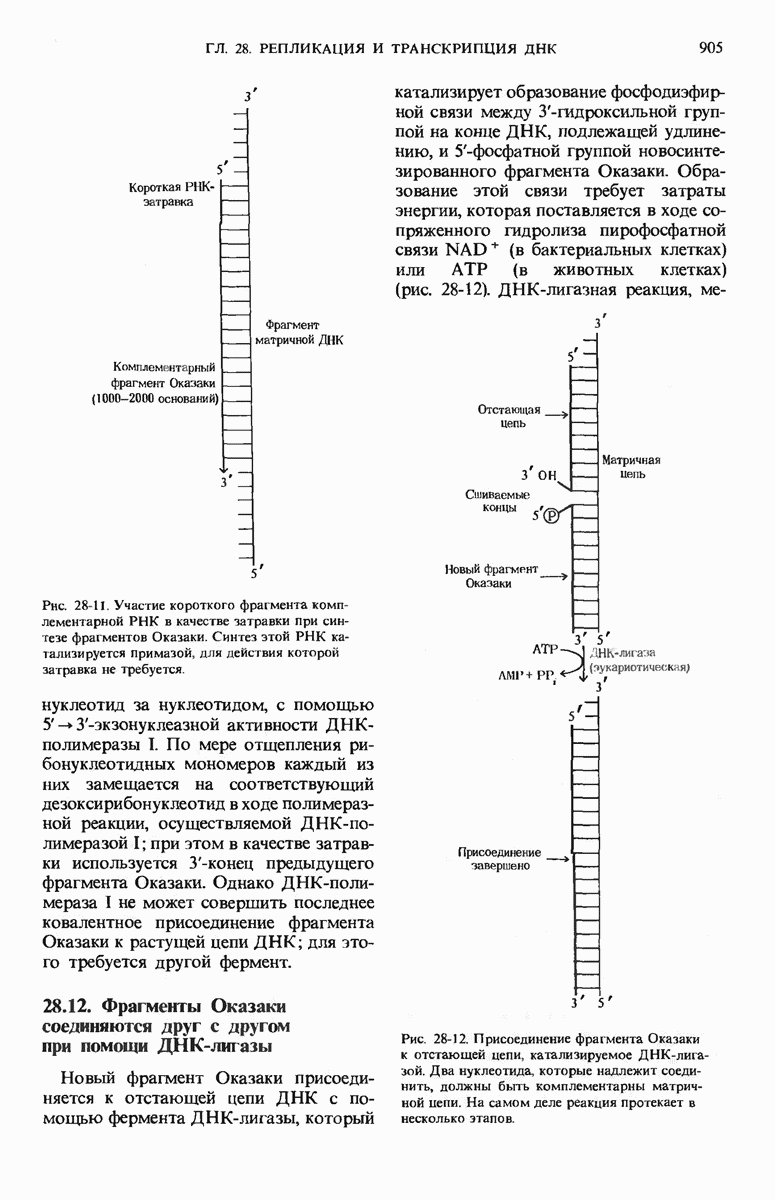
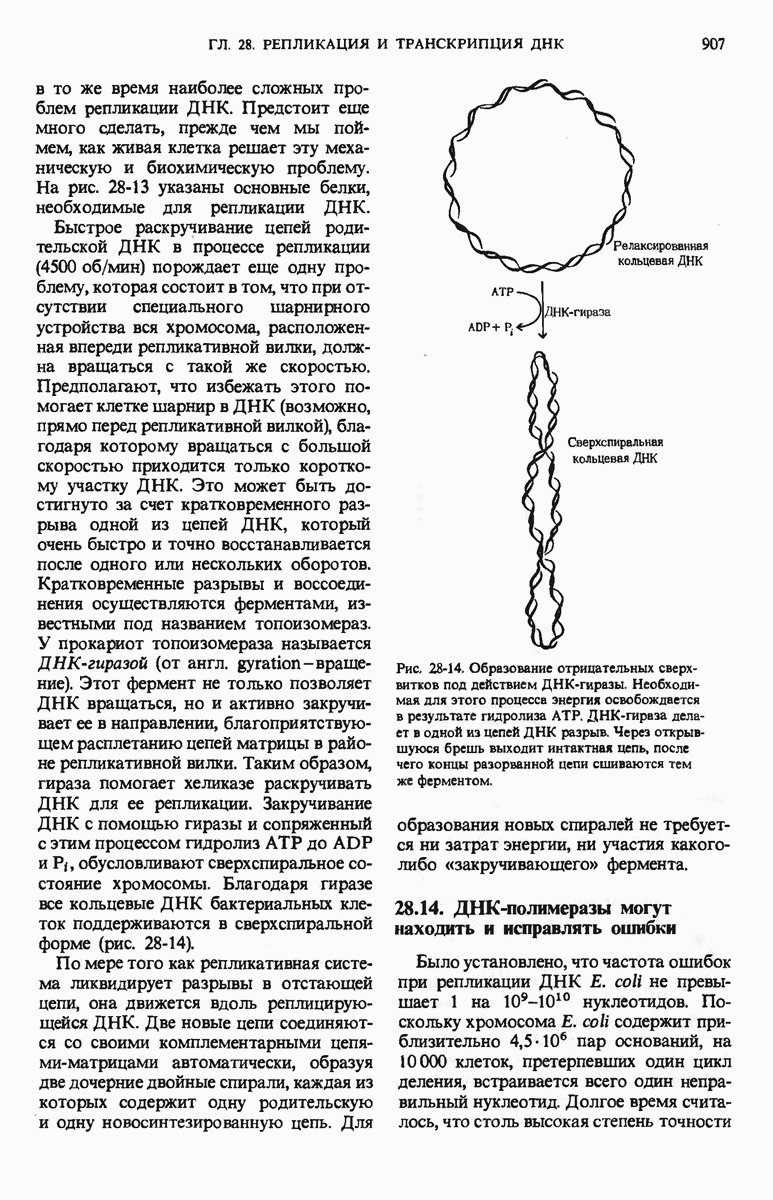
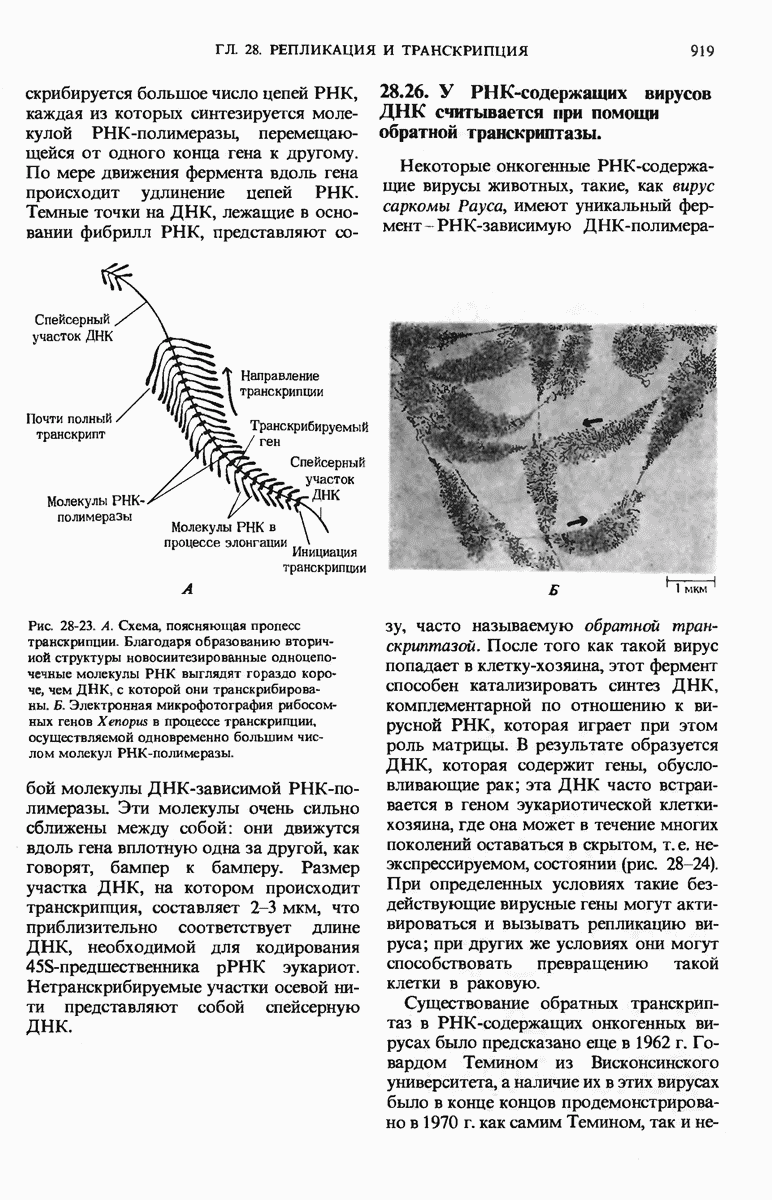
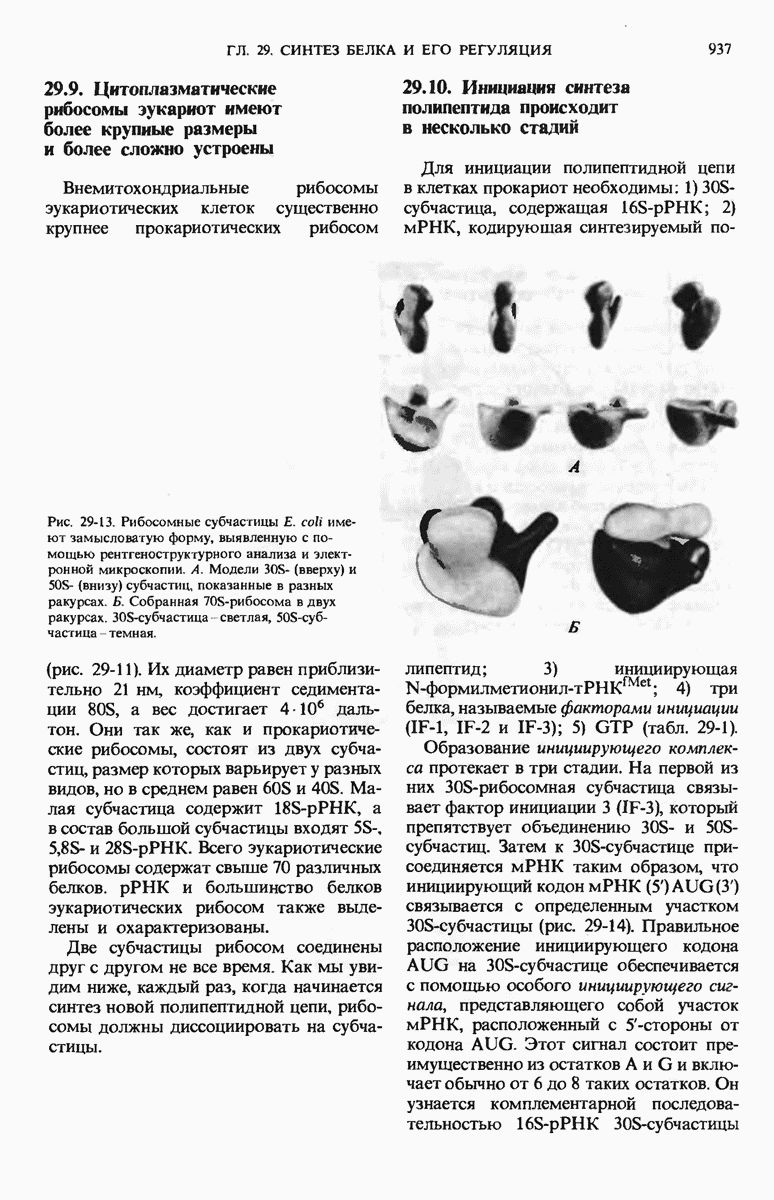
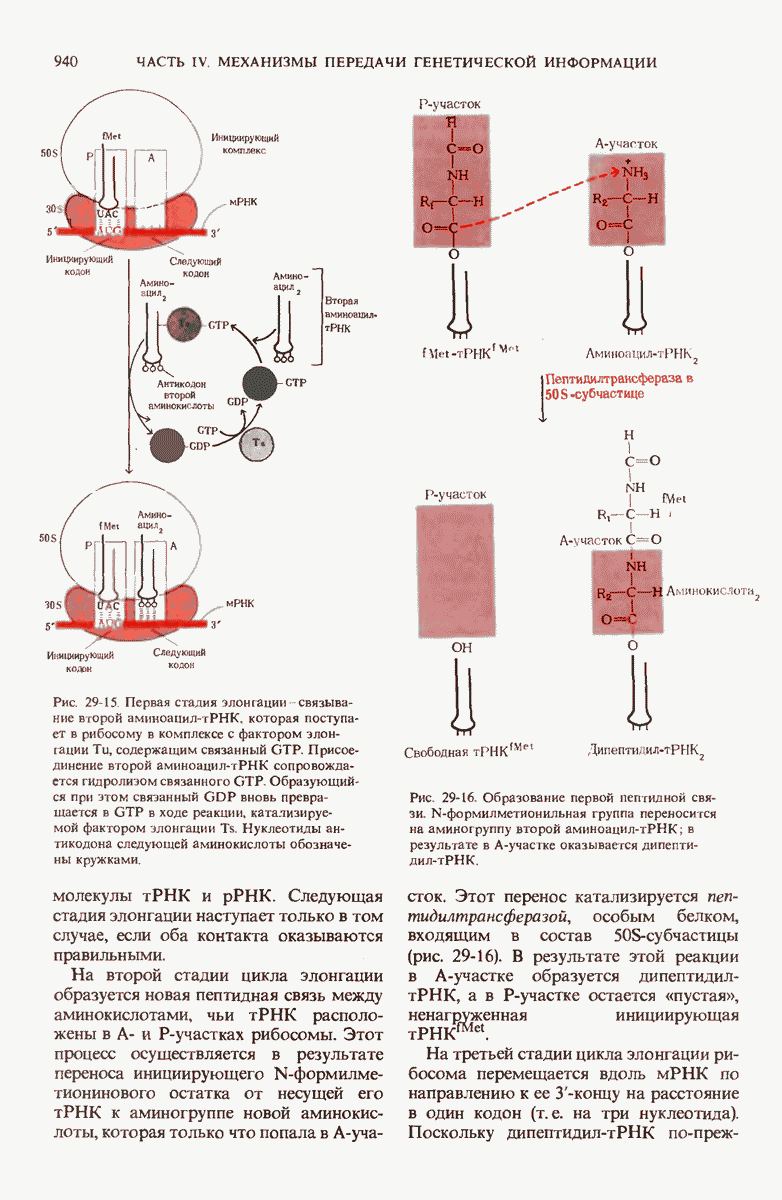
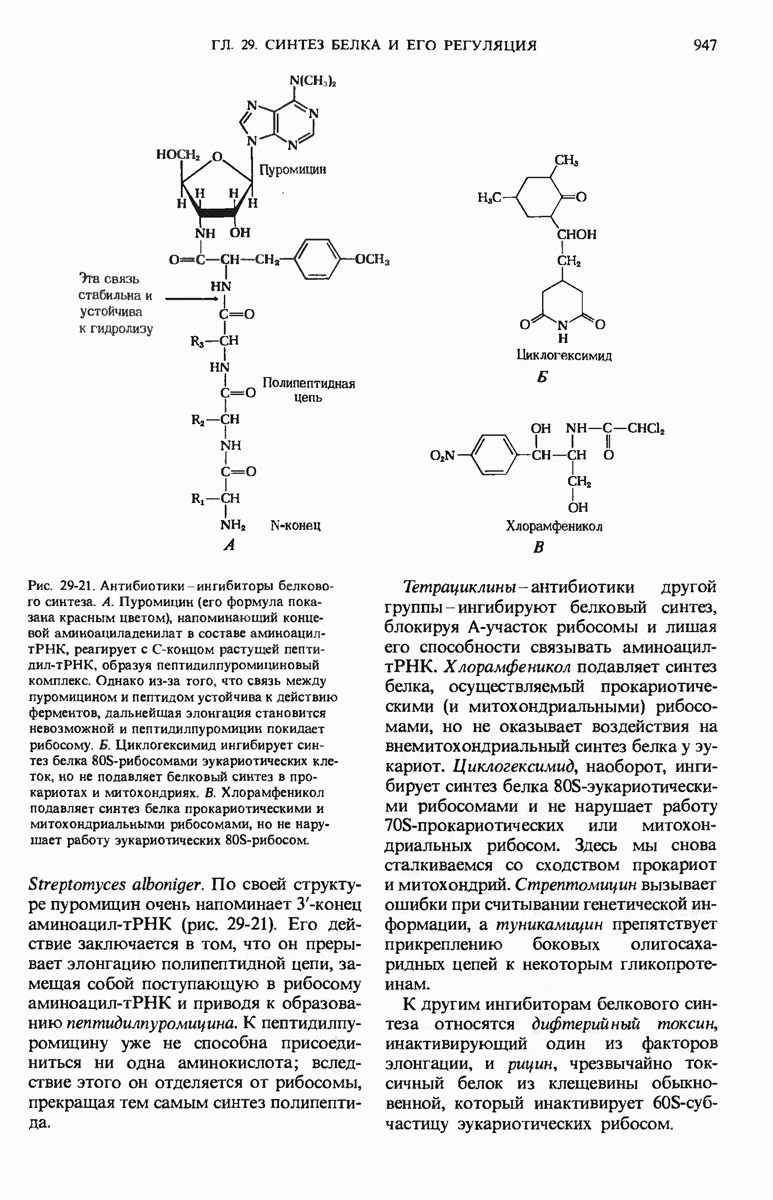
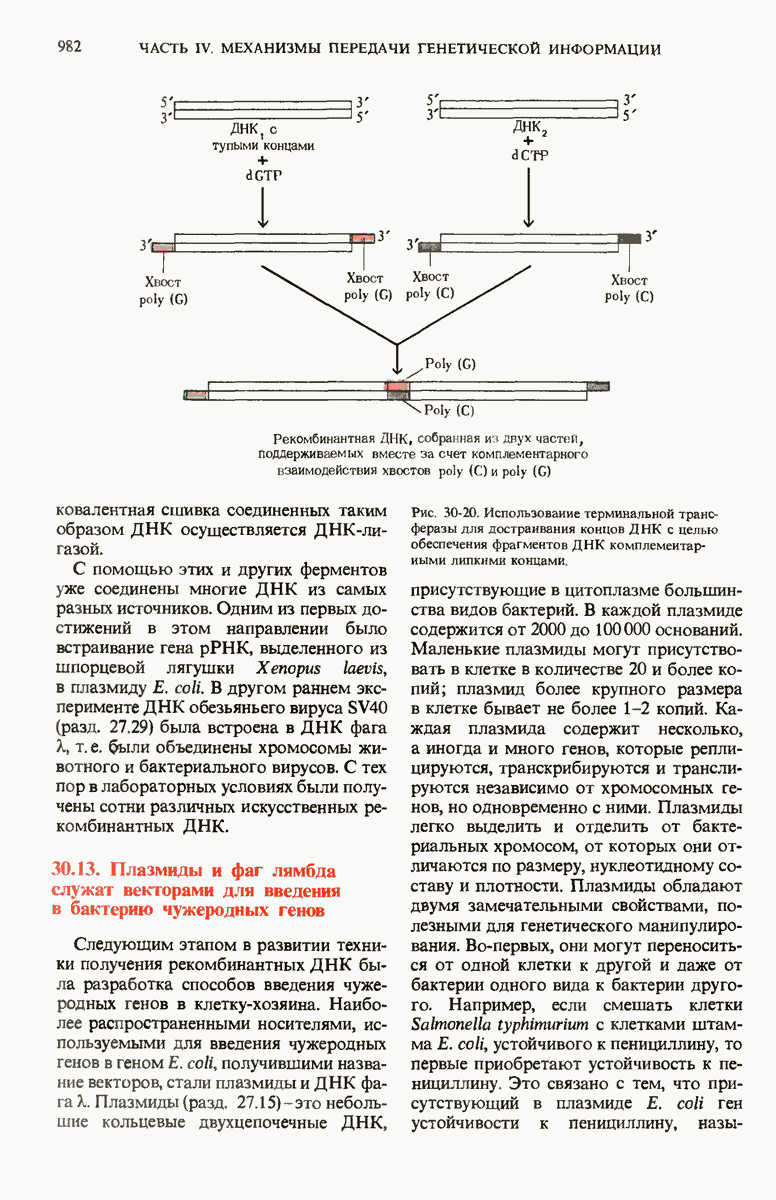
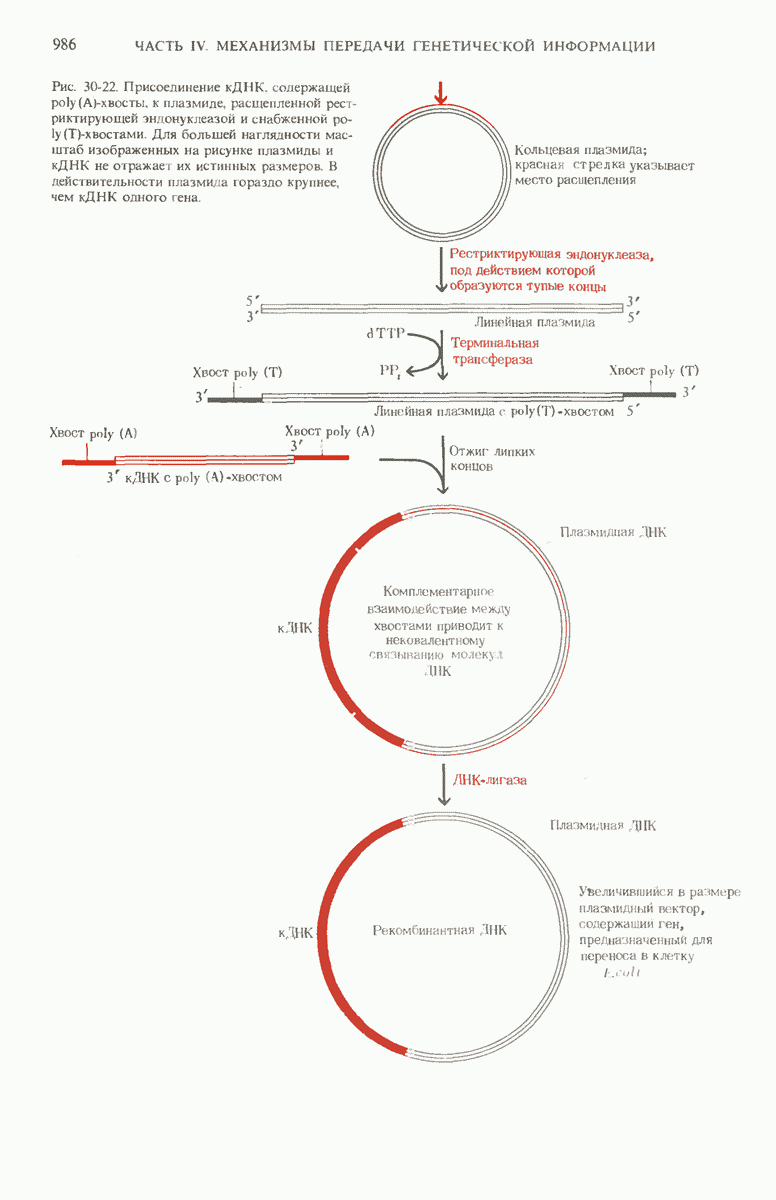
29.27. В оперонах имеется еще промоторный участок
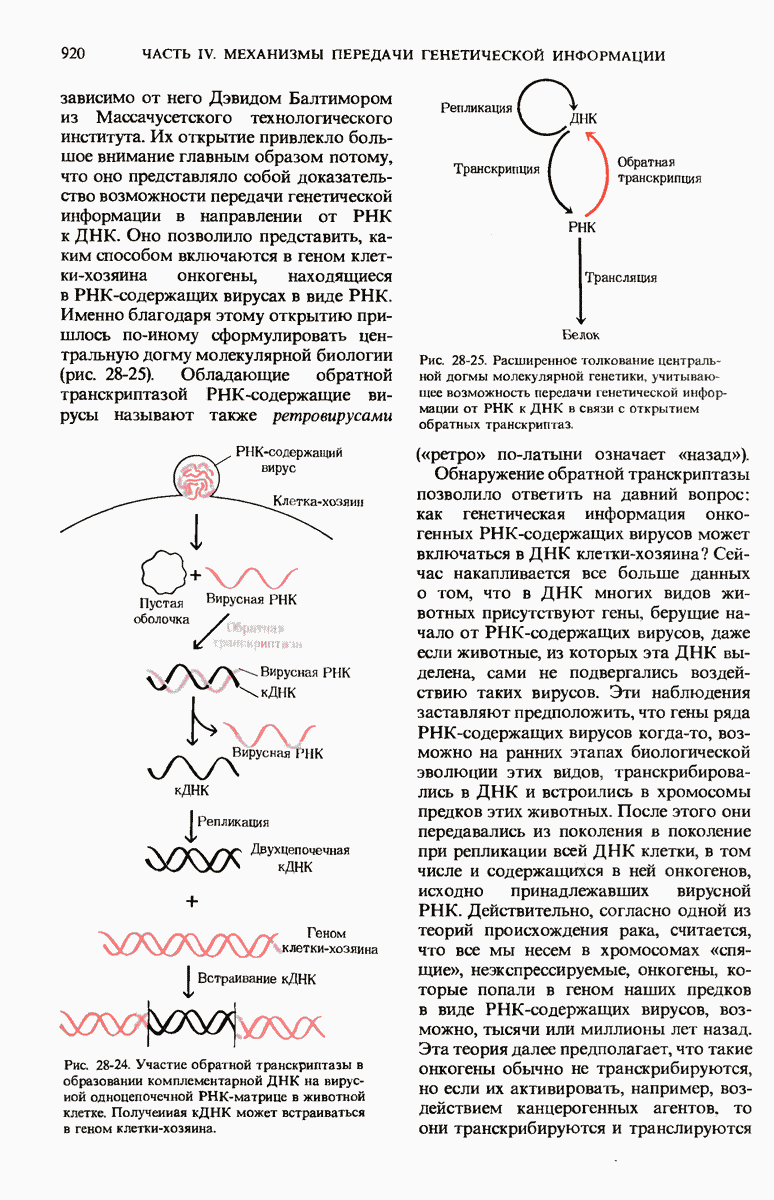
Мы уже видели, что если в среде присутствует лактоза и нет глюкозы, то индуктор, соединяясь с репрессором, «снимает» его с оператора и тем самым дает возможность транскрибироваться  -генам и соответственно синтезироваться
-генам и соответственно синтезироваться  -белкам. Предположим теперь, что в среде находятся и лактоза, и глюкоза. В этих условиях Е. coli использует только глюкозу, пренебрегая лактозой. Более того, клетки перестают синтезировать
-белкам. Предположим теперь, что в среде находятся и лактоза, и глюкоза. В этих условиях Е. coli использует только глюкозу, пренебрегая лактозой. Более того, клетки перестают синтезировать  -белки. Репрессия синтеза
-белки. Репрессия синтеза  -белков глюкозой называется катаболитной репрессией. Клетки Е. coli способны чувствовать, доступна ли глюкоза, с помощью другого регуляторного механизма, который совместно с
-белков глюкозой называется катаболитной репрессией. Клетки Е. coli способны чувствовать, доступна ли глюкоза, с помощью другого регуляторного механизма, который совместно с  -репрессором и оператором контролирует синтез
-репрессором и оператором контролирует синтез  -ферментов.
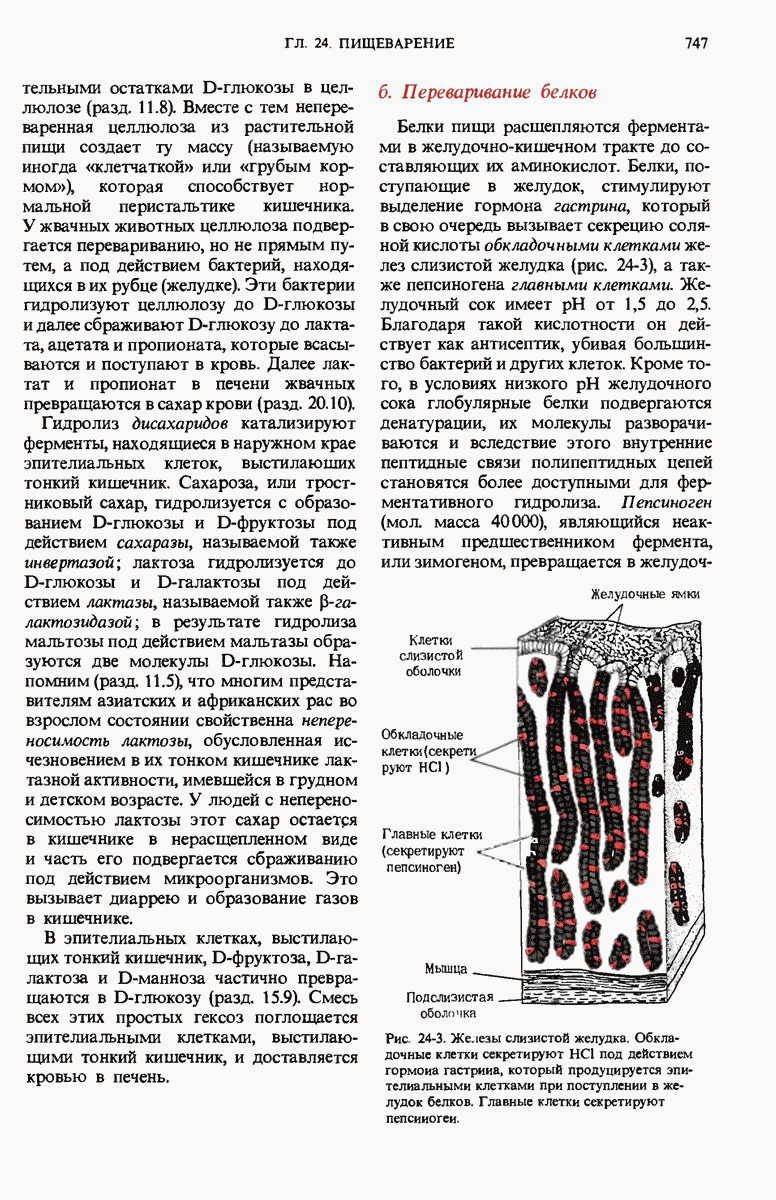
-ферментов.

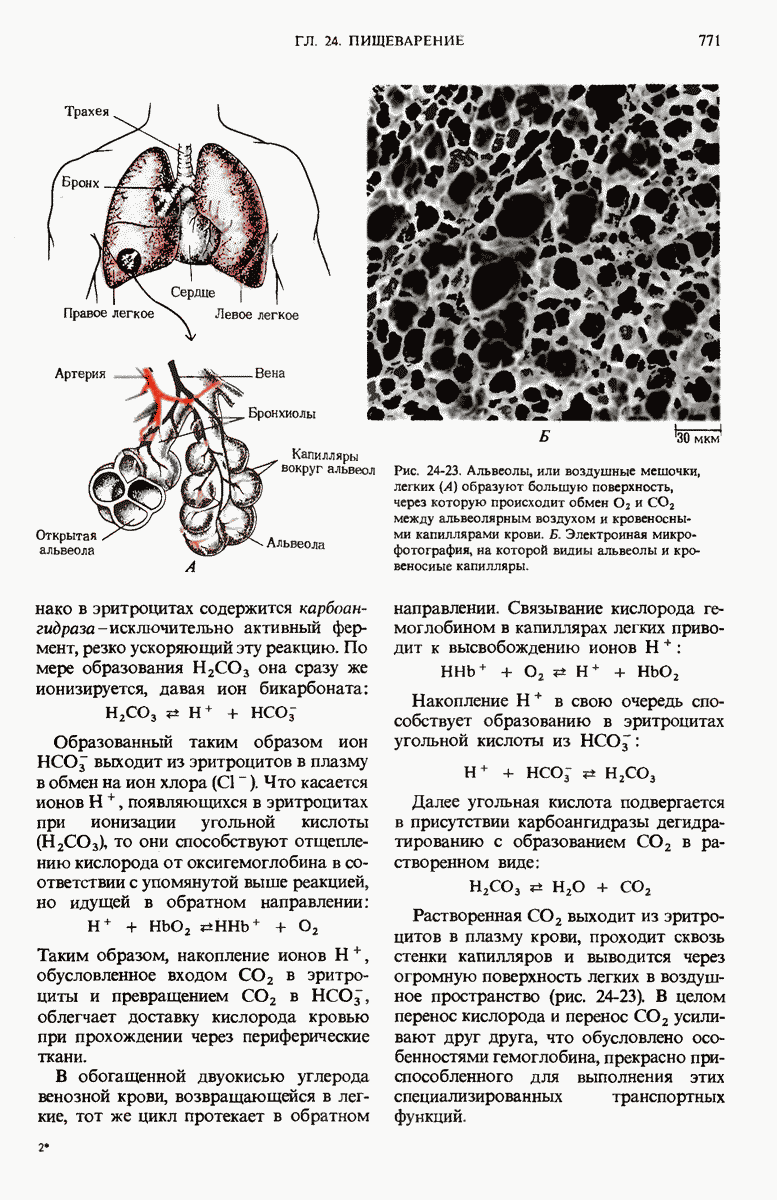
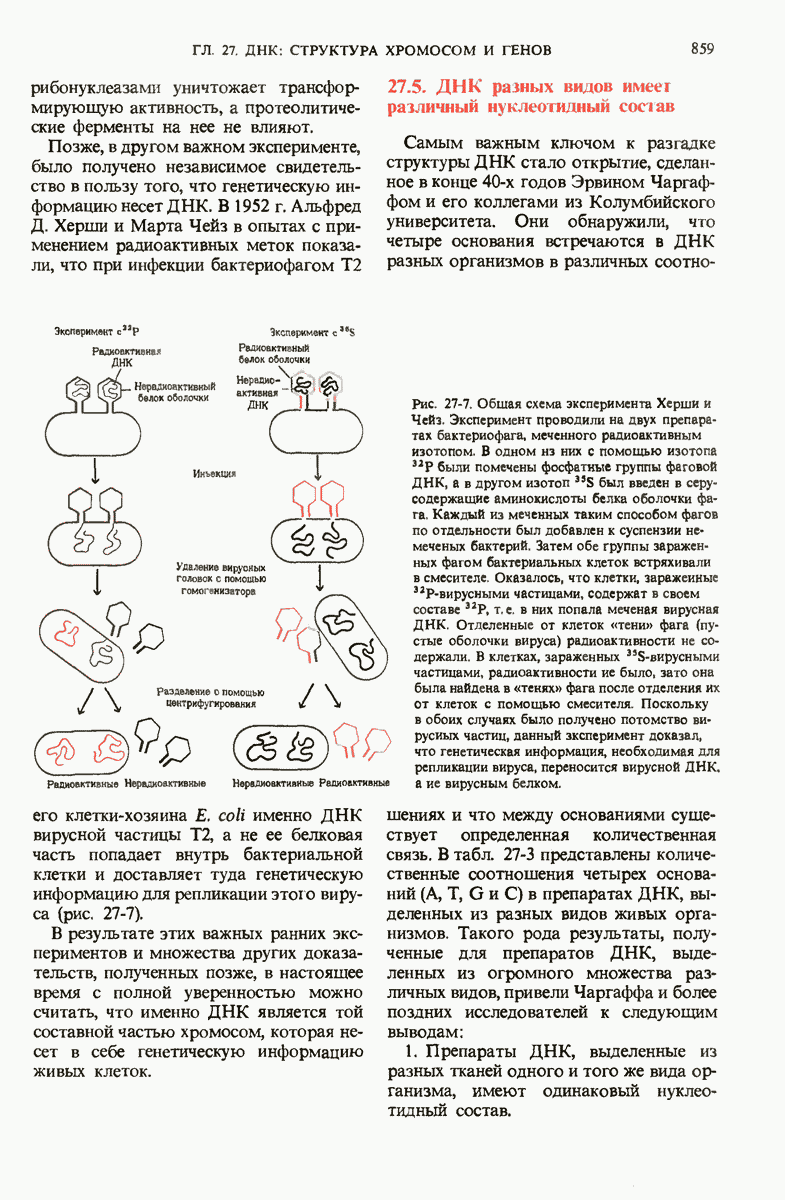
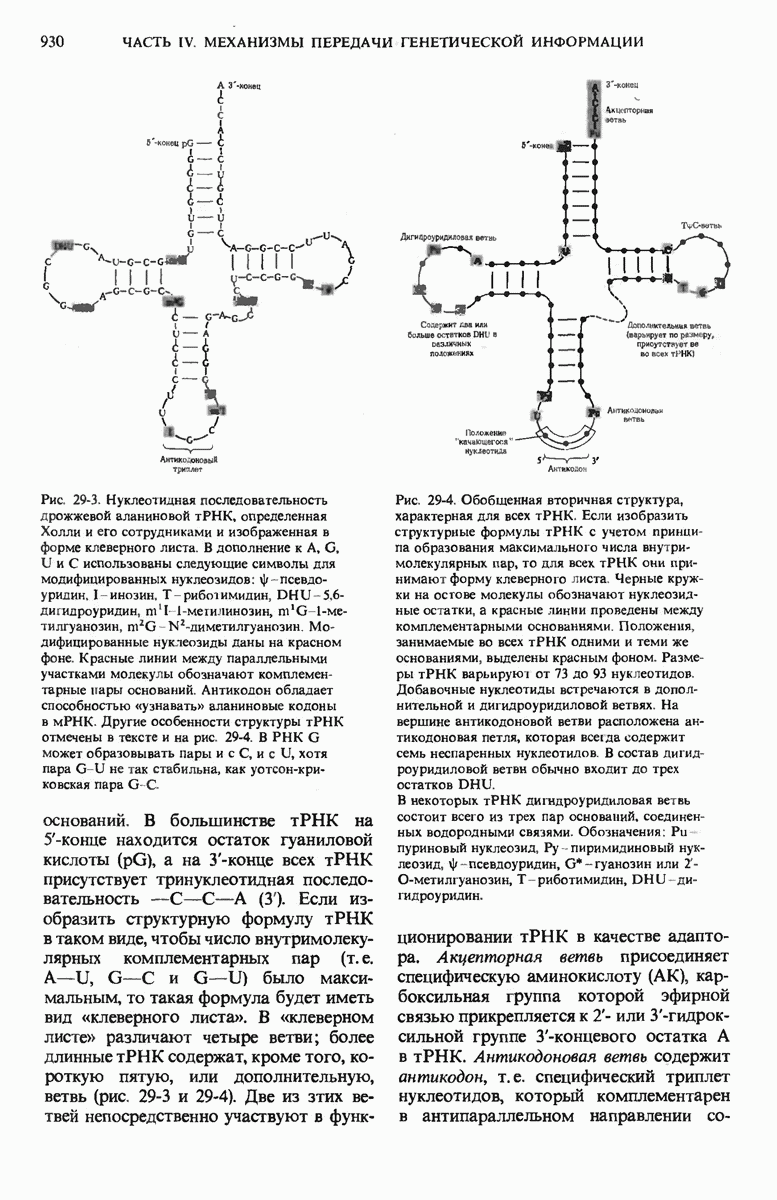
Рис. 29-26. Электронная микрофотография участка ДНК Е. coli, на которой видна молекула  -репрессора (указана стрелкой), присоединенная к
-репрессора (указана стрелкой), присоединенная к  -оператору.
-оператору.
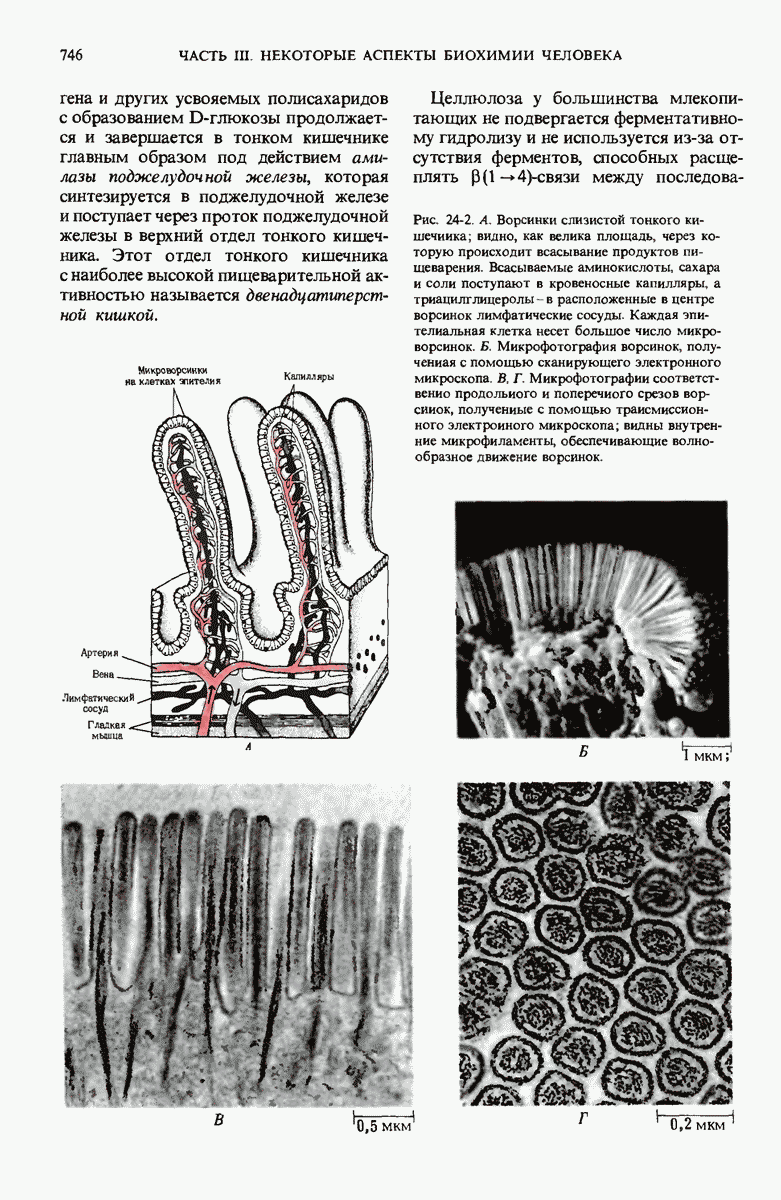
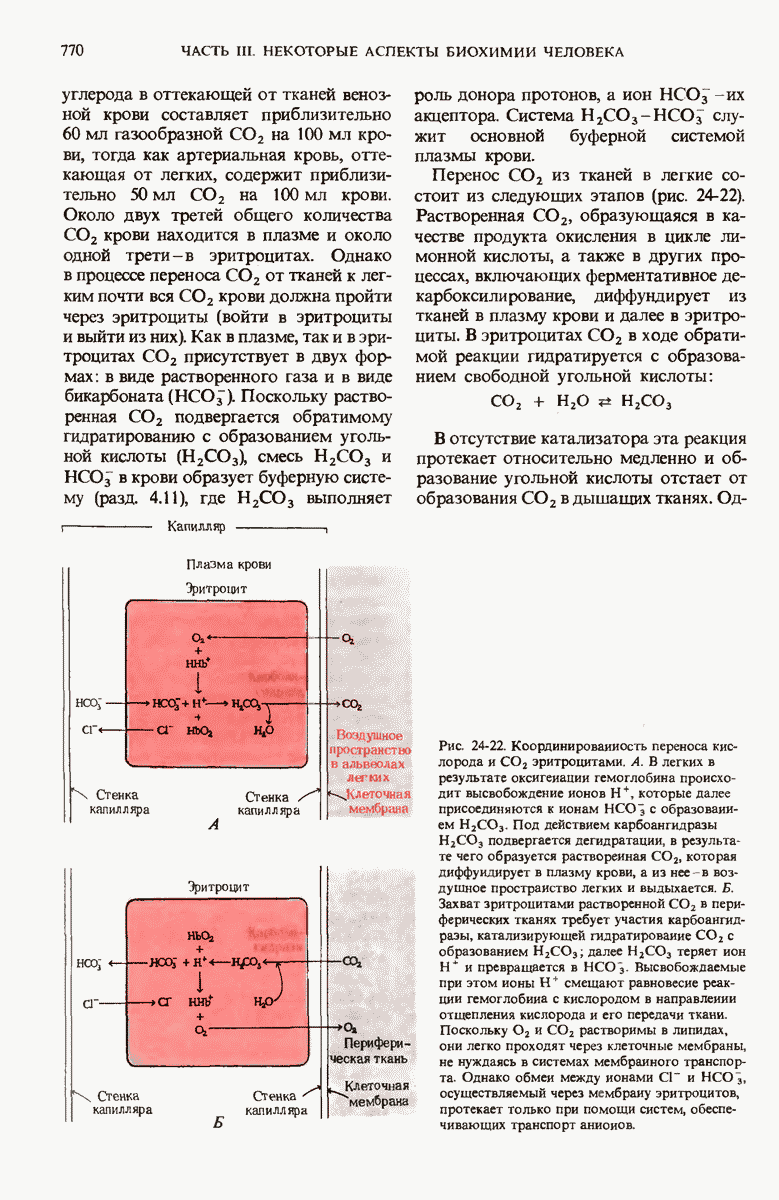
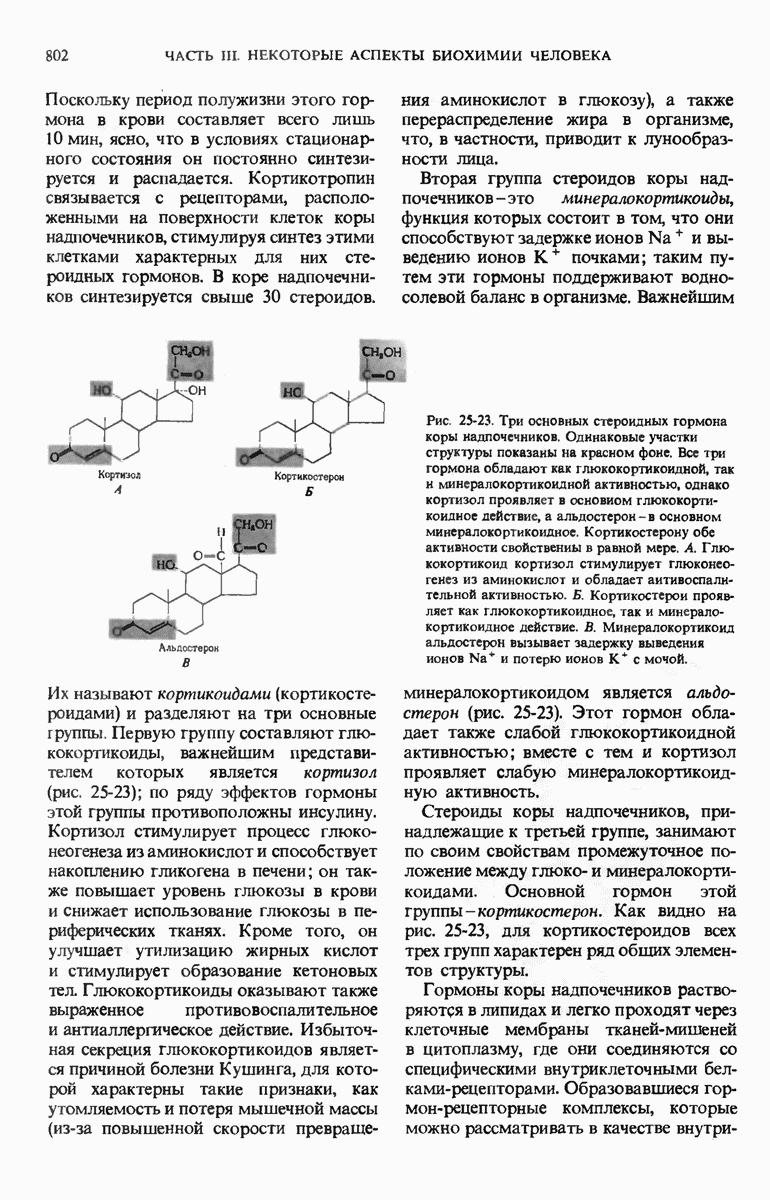
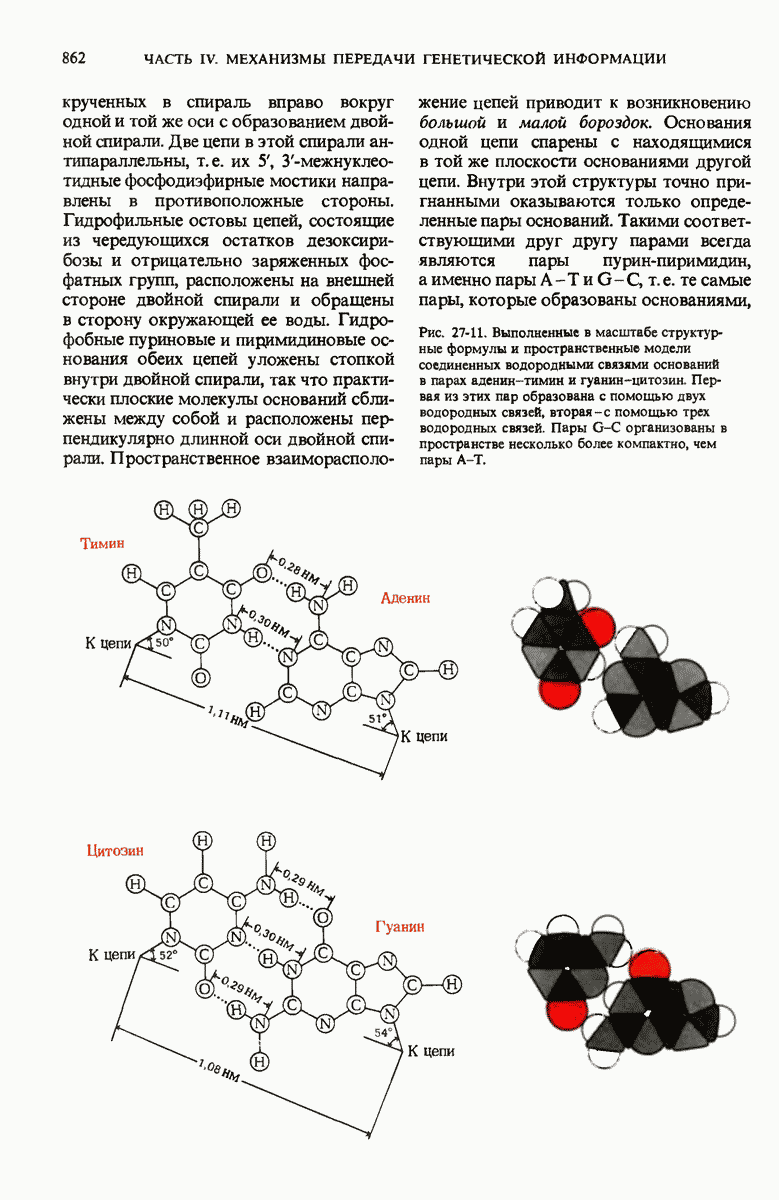
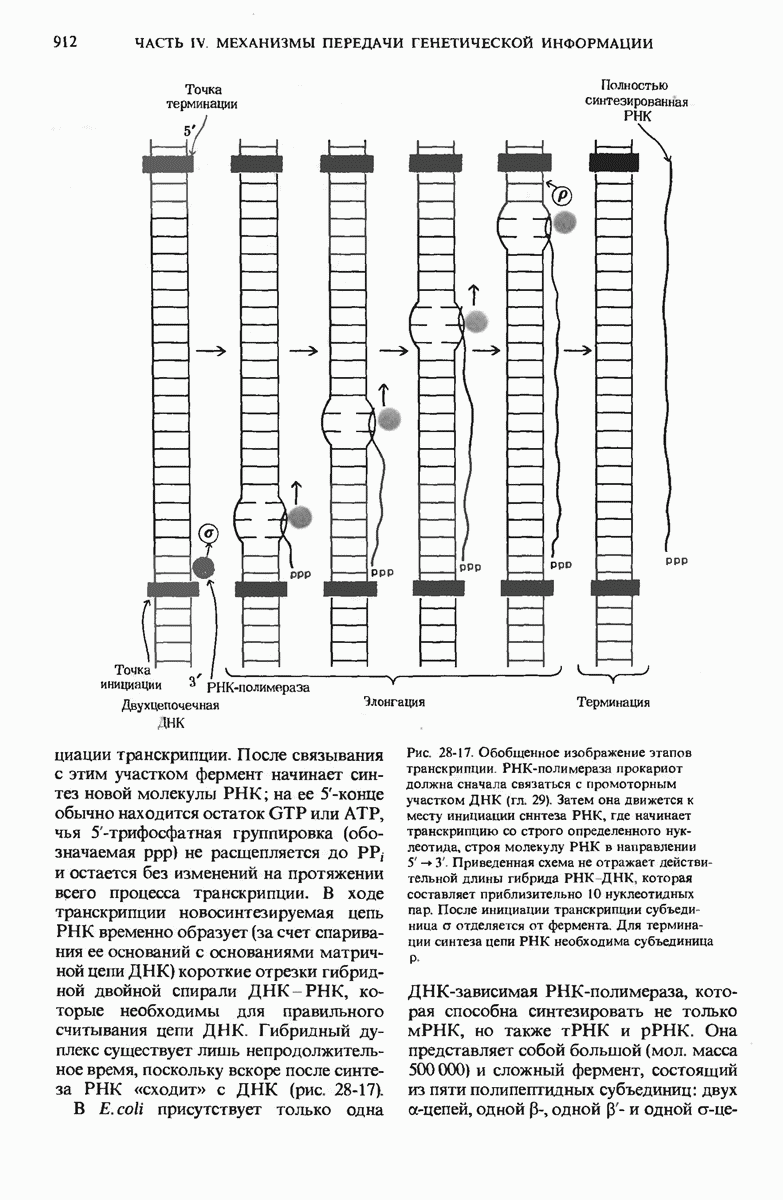
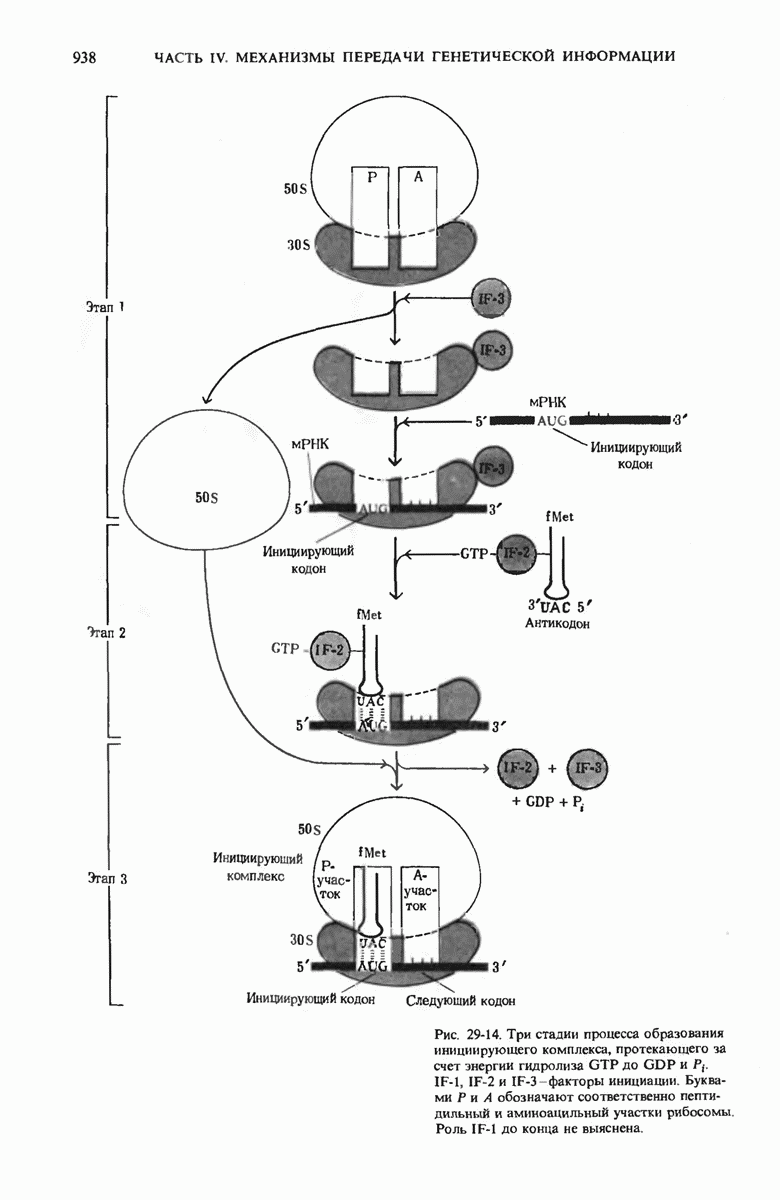
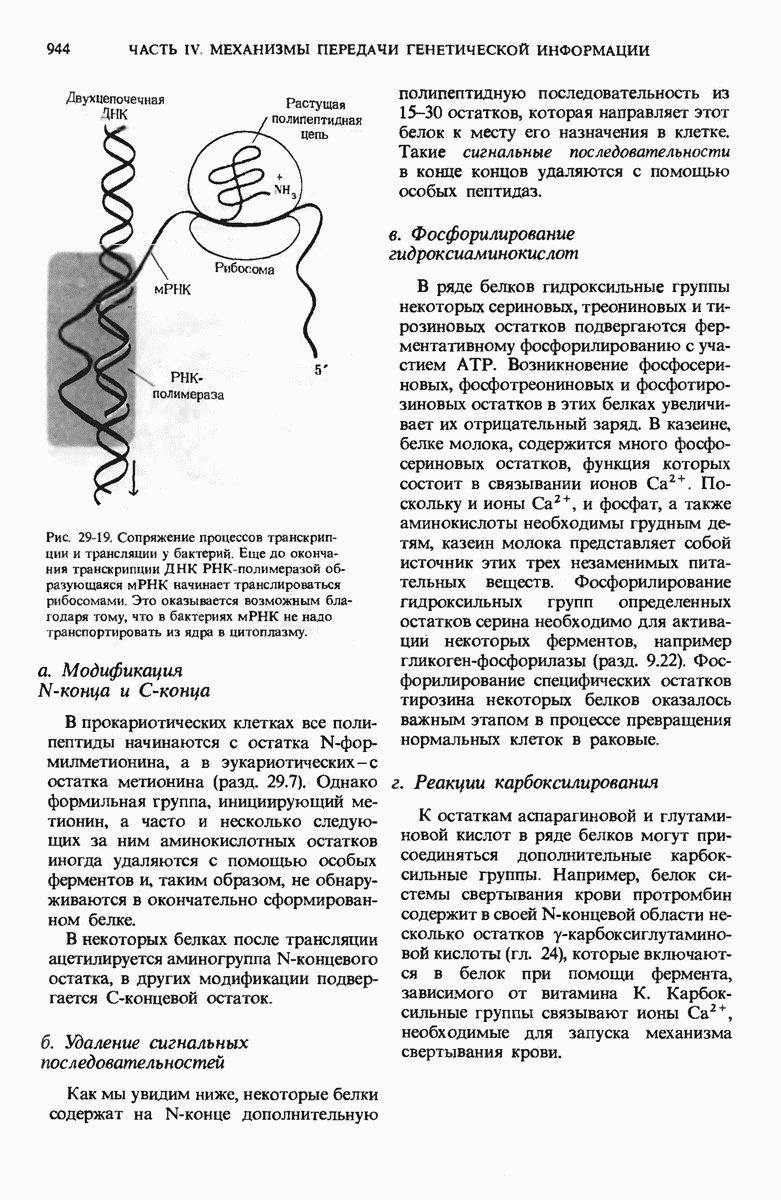
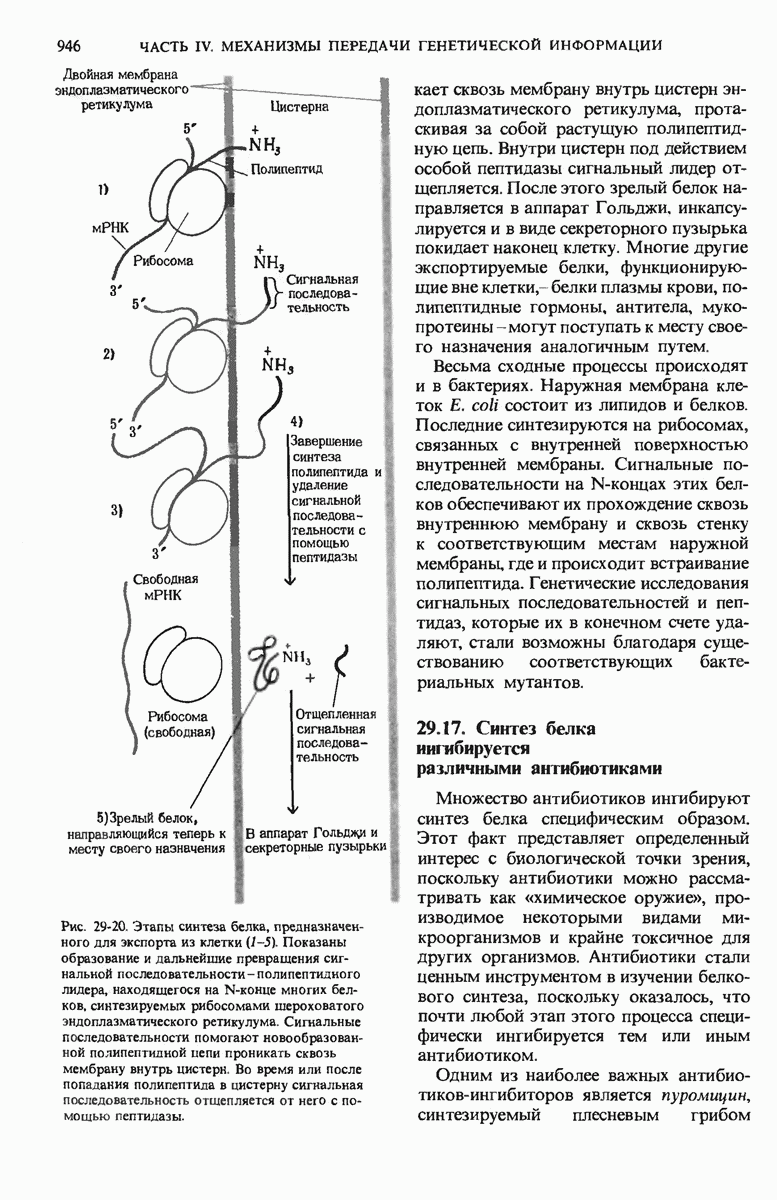
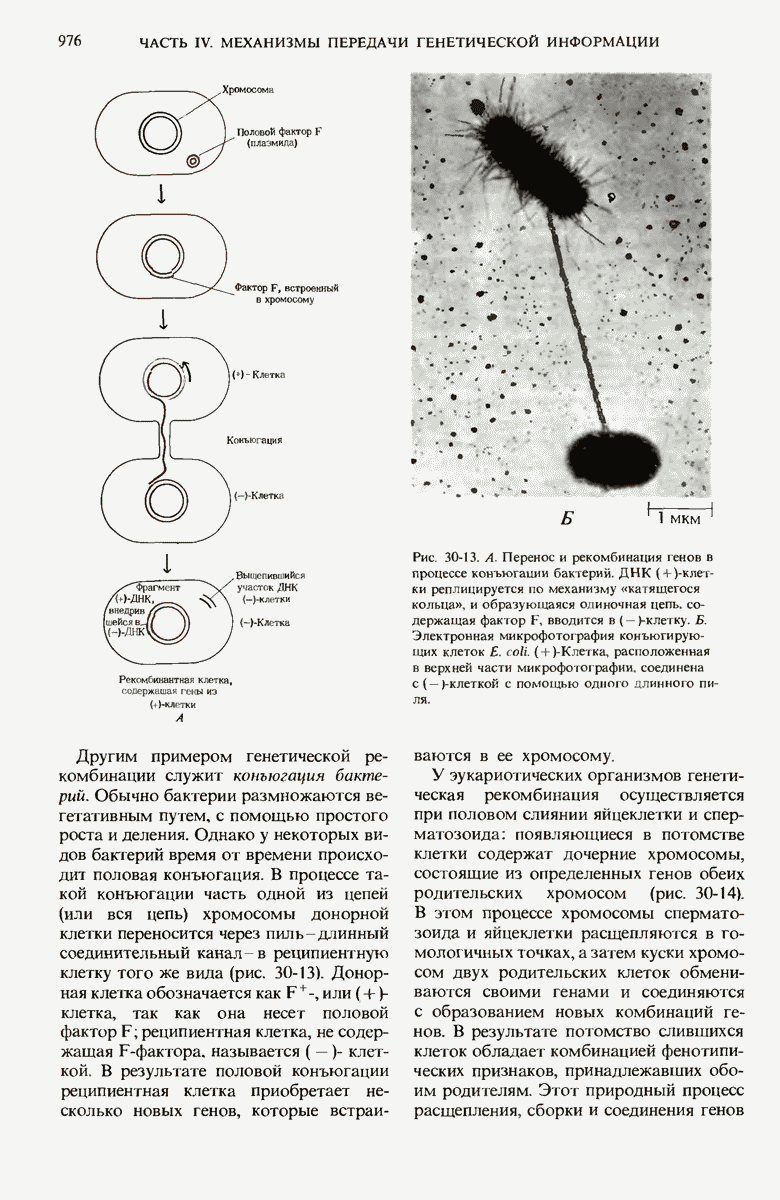
В дополнение к i-гену и оператору (о-участку) в ДНК существует еще один особый регуляторный участок -промотор, или  -участок, расположенный между геном i и оператором (рис. 29-27). Промотор в свою очередь состоит из двух функционально различных частей. Рядом с оператором находится «вход для РНК-полимеразы», т.е. участок, в котором происходит первоначальное связывание РНК-полимеразы. Вторая часть промотора представляет собой специфический участок связывания другого регуляторного белка - белка, активирующего катаболитный ген (БАК или САР от англ. catabolite activator protein). Эта часть промоторного участка держит под своим контролем другую его часть, ответственную за связывание РНК-полимеразы.
-участок, расположенный между геном i и оператором (рис. 29-27). Промотор в свою очередь состоит из двух функционально различных частей. Рядом с оператором находится «вход для РНК-полимеразы», т.е. участок, в котором происходит первоначальное связывание РНК-полимеразы. Вторая часть промотора представляет собой специфический участок связывания другого регуляторного белка - белка, активирующего катаболитный ген (БАК или САР от англ. catabolite activator protein). Эта часть промоторного участка держит под своим контролем другую его часть, ответственную за связывание РНК-полимеразы.

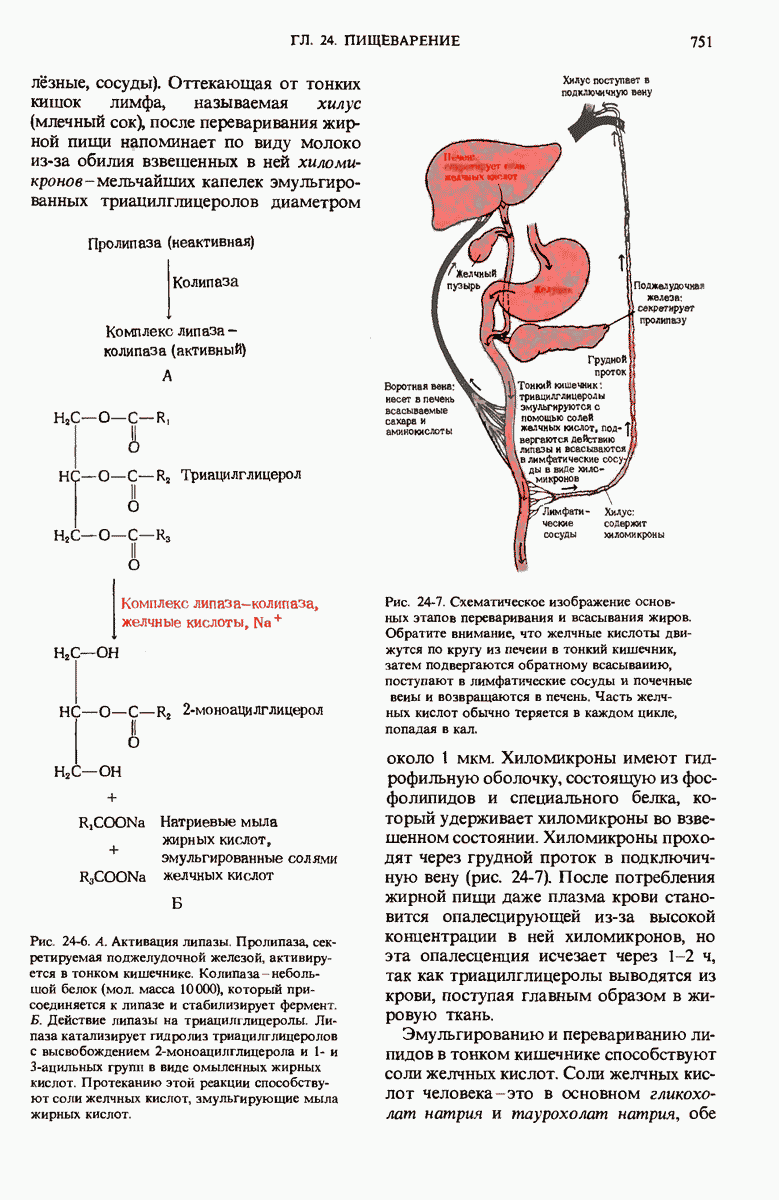
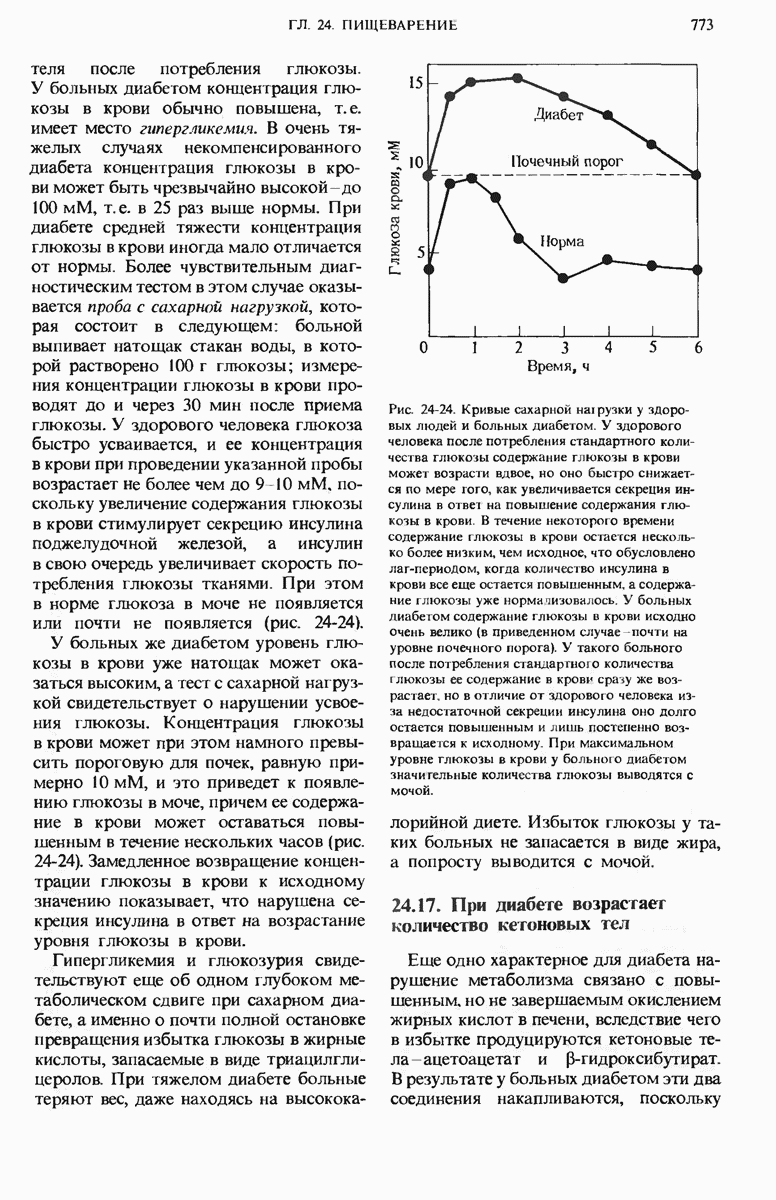
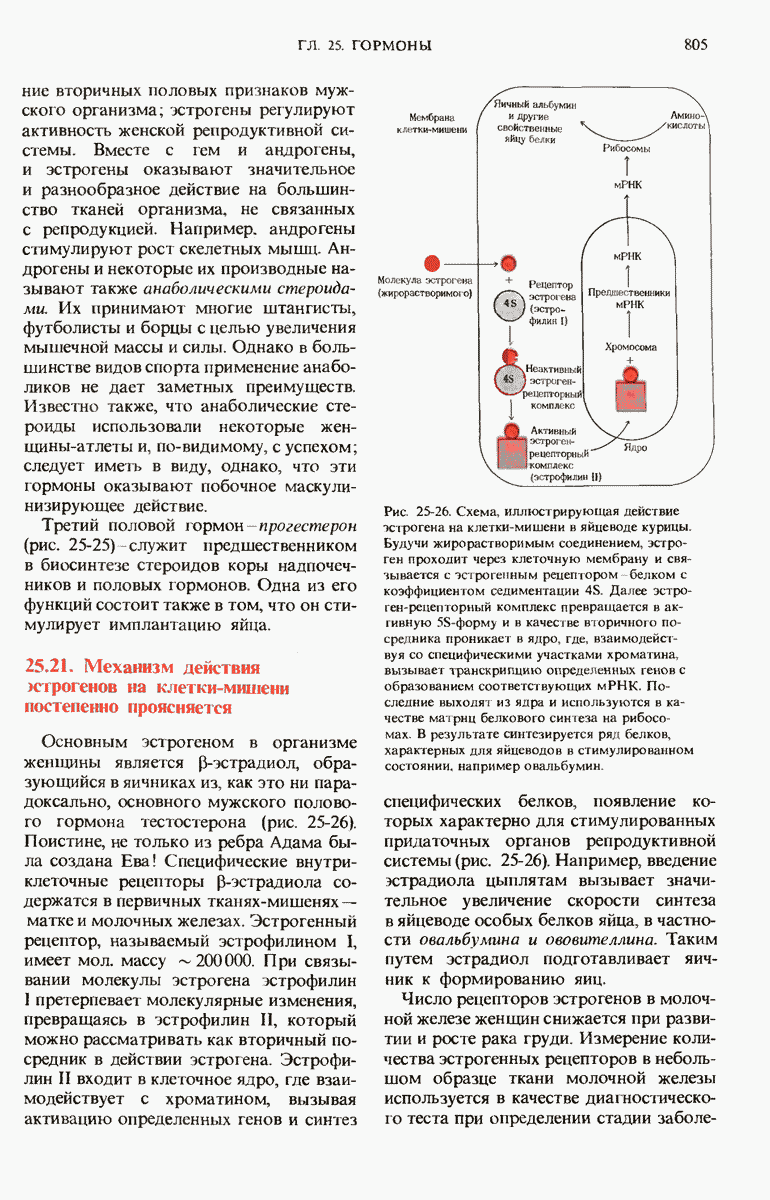
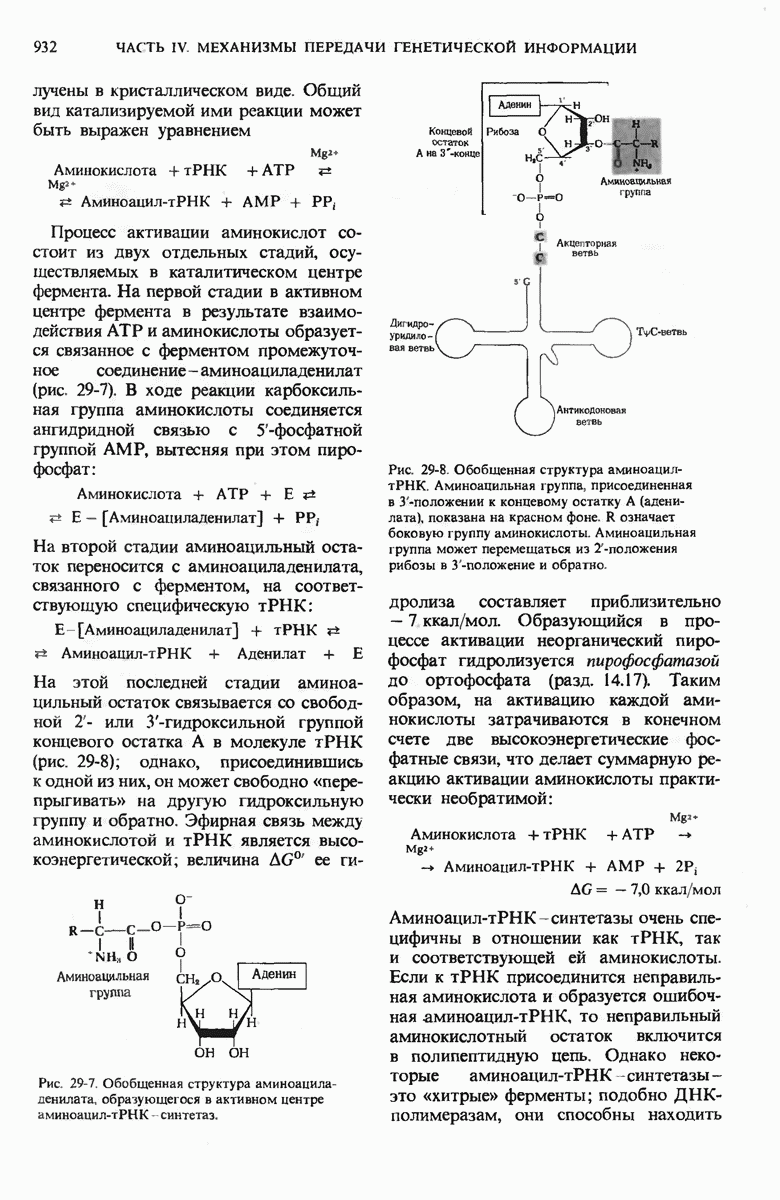
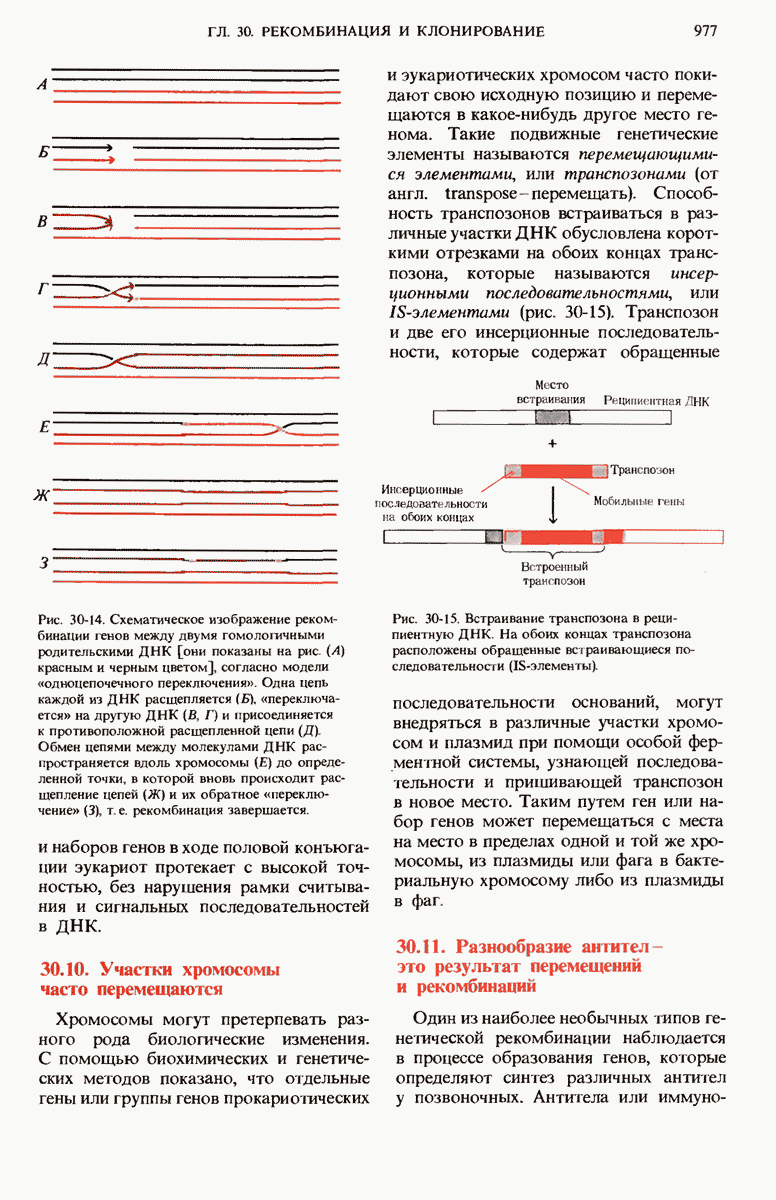
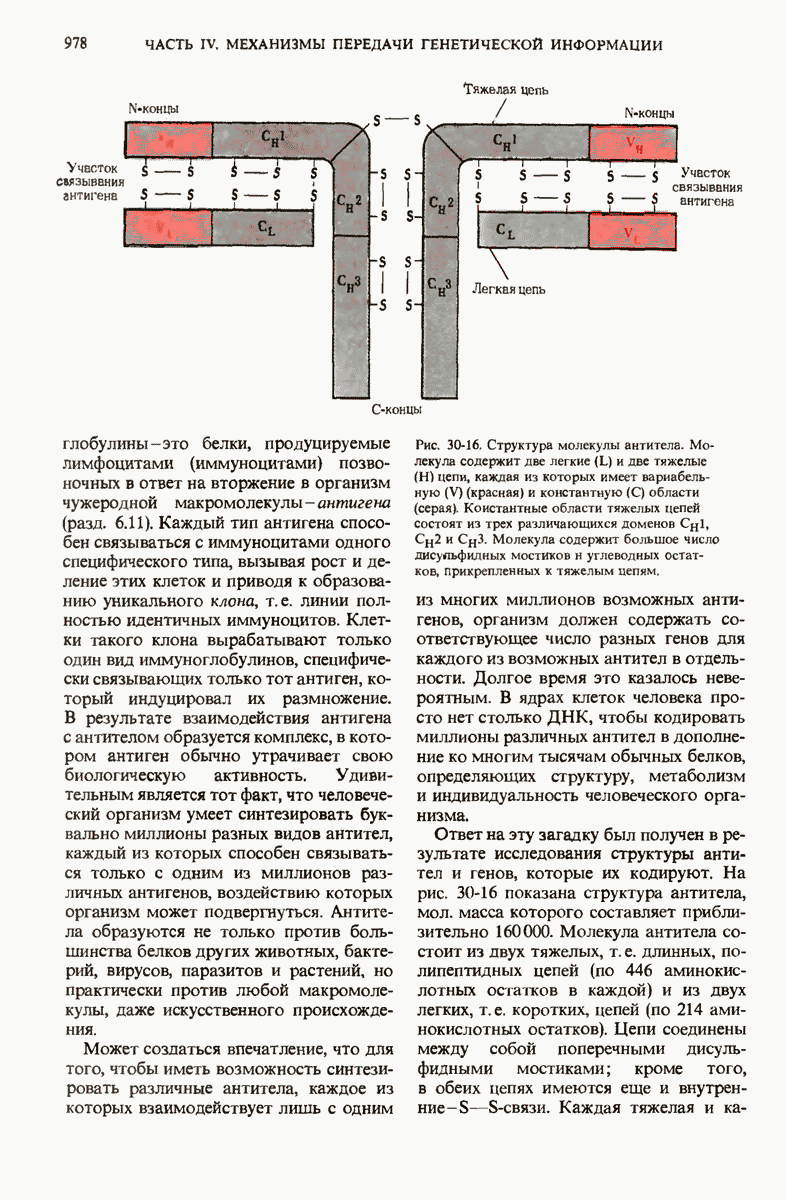
Рис. 29-27. А. Регуляторные участки lac-onepoна. САР-участок промотора способен связывать САР лишь в том случае, если он находится в комплексе с сАМР. РНК-полимераза может попасть в участок первоначального связывания только при условии, если САР-участок занят. Репрессор взаимодействует с операторол лишь в отсутствие индуктора. Б. Три структурных гена z, у и  lac-оперона транскрибируются при условии, что в среде нет глюкозы, а присутствует лактоза. В этом случае оператор свободен от репрессора и комплекс САР-сАМР соединяется с промотором, позволяя РНК-полимеразе попасть в участок первоначального связывания, «спуститься» к инициирующему кодону и начать транскрибировать три структурных гена. В. Если глюкозы в среде много, то сАМР не образуется и САР поэтому не в состоянии связаться с промотором. В этих условиях РНК-полимераза не может получить доступ к промотору и lac-гены не транскрибируются.
lac-оперона транскрибируются при условии, что в среде нет глюкозы, а присутствует лактоза. В этом случае оператор свободен от репрессора и комплекс САР-сАМР соединяется с промотором, позволяя РНК-полимеразе попасть в участок первоначального связывания, «спуститься» к инициирующему кодону и начать транскрибировать три структурных гена. В. Если глюкозы в среде много, то сАМР не образуется и САР поэтому не в состоянии связаться с промотором. В этих условиях РНК-полимераза не может получить доступ к промотору и lac-гены не транскрибируются.
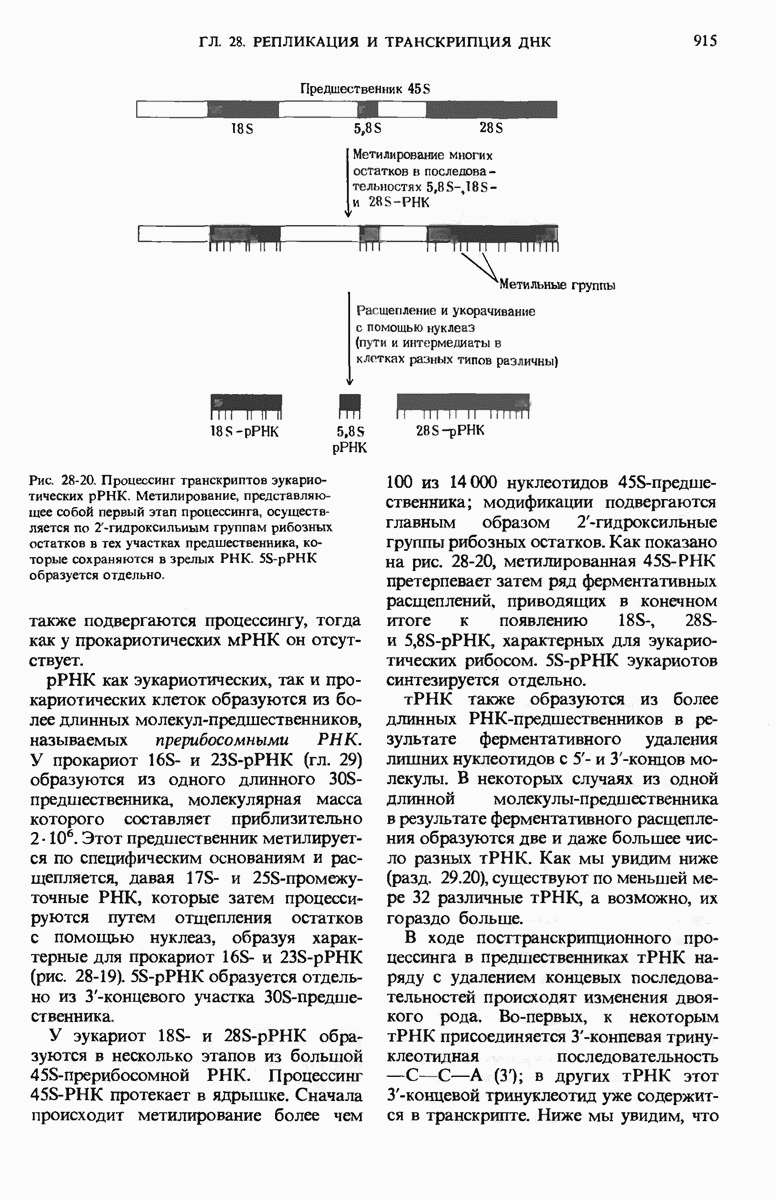
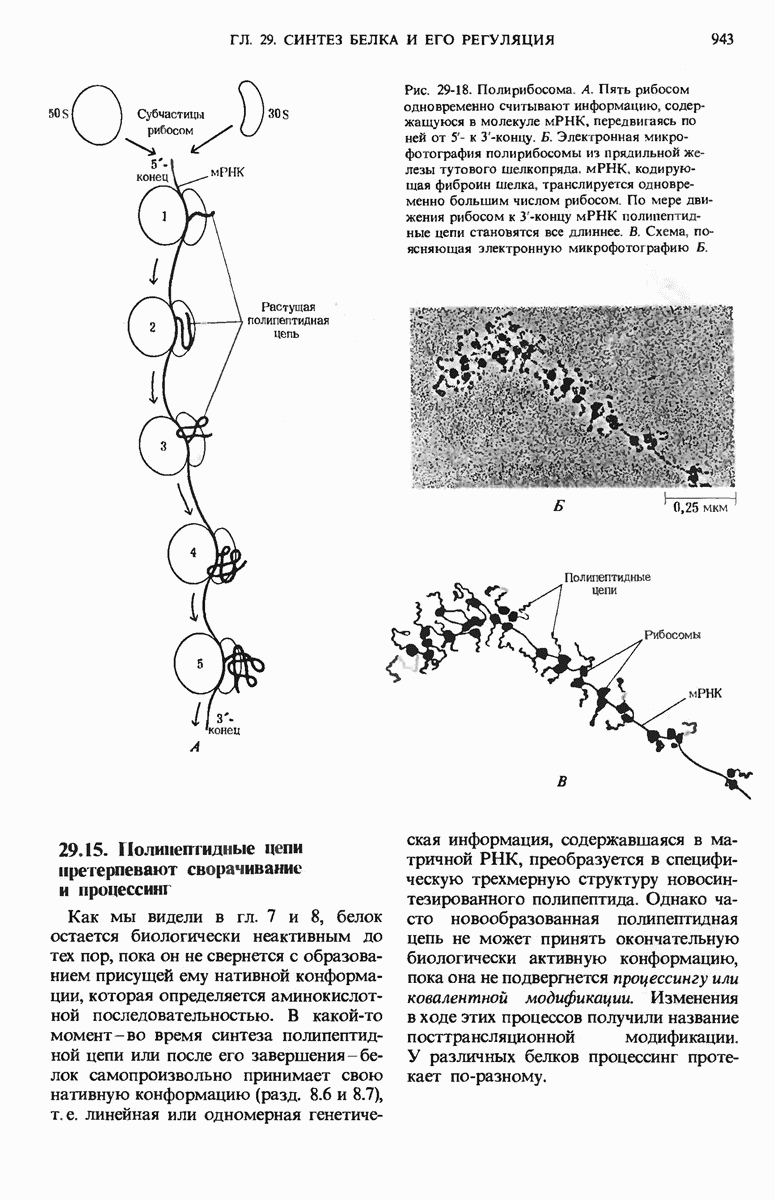
Если в среде отсутствует глюкоза, то в клетке формируется комплекс между САР и циклическим АМР (сАМР); этот комплекс соединяется с САР-участком ДНК и дает возможность РНК-полимеразе попасть в участок первоначального связывания. Если среда содержит лактозу, то о-участок открыт, поскольку оператор не может взаимодействовать с индуктор-репрессорным комплексом.
В этих условиях РНК-полимераза перемещается от места своего первоначального связывания через область оператора и начинает транскрибировать три lac-гена. Если же в среде присутствует глюкоза, то концентрация сАМР сильно снижается и комплекс САР-сАМР не может образоваться. Короче говоря, САР-участок обеспечивает доступность участка первоначального связывания РНК-полимеразы только будучи соединенным с комплексом САР-сАМР. Поэтому, когда такой комплекс не образуется, РНК-полимераза не может связаться с промоторным участком и lac-гены не транскрибируются. Таким образом, транскрипция lac-генов возможна только при отсутствии глюкозы; следовательно, lac-оперон находится как под позитивным ( -участок), так и под негативным (о-участок) контролем.
-участок), так и под негативным (о-участок) контролем.
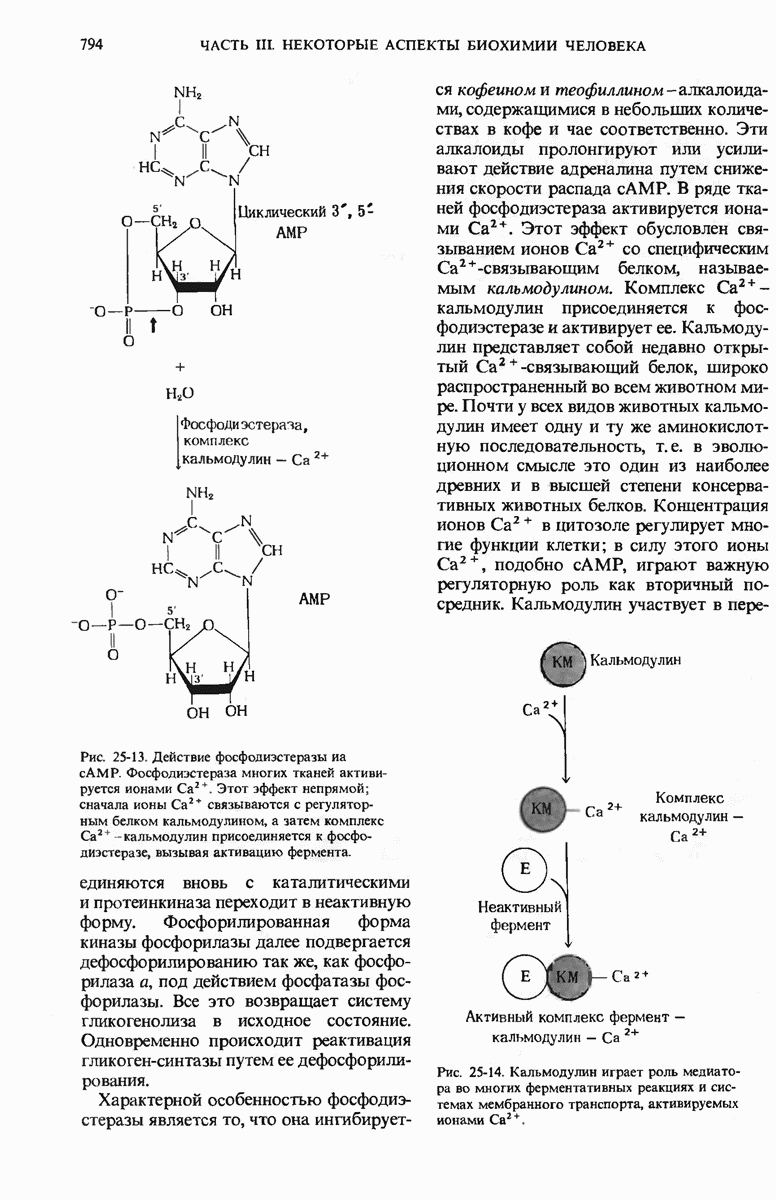
Возникает естественный вопрос, как САР улавливает присутствие глюкозы? В молекуле САР есть два центра - один, как указано выше, для связывания с САР-участком промотора и второй для связывания сАМР. Вспомним, что сАМР - это внутриклеточный посредник при действии ряда гормонов в клетках позвоночных (гл. 25). У Е. coli сАМР также служит посредником, но для другой цели: он сигнализирует о том, есть ли в ростовой среде глюкоза. Клетки Е. coli содержат фермент аденилатциклазу (разд. 25.5), который катализирует образование сАМР из АТР; в этих клетках имеется также фосфодиэстераза, которая гидролизует сАМР и тем самым инактивирует его. Если концентрация глюкозы высока и соответствует потребностям клетки, концентрация сАМР в клетке очень мала (рис. 29-27). Однако, когда концентрация глюкозы в клетке падает, содержание в ней сАМР возрастает благодаря увеличению активности аденилатциклазы и уменьшению активности фосфодиэстеразы (разд. 25.9). Образующийся сАМР взаимодействует с САР, а комплекс САР-сАМР в свою очередь связывается с САР-участком промотора. Только в этом случае РНК-полимераза может присоединиться к месту инициации и начать синтезировать на lac-генах мРНК (рис. 29-27). Поэтому сАМР в бактериях получил название «сигнал голода».
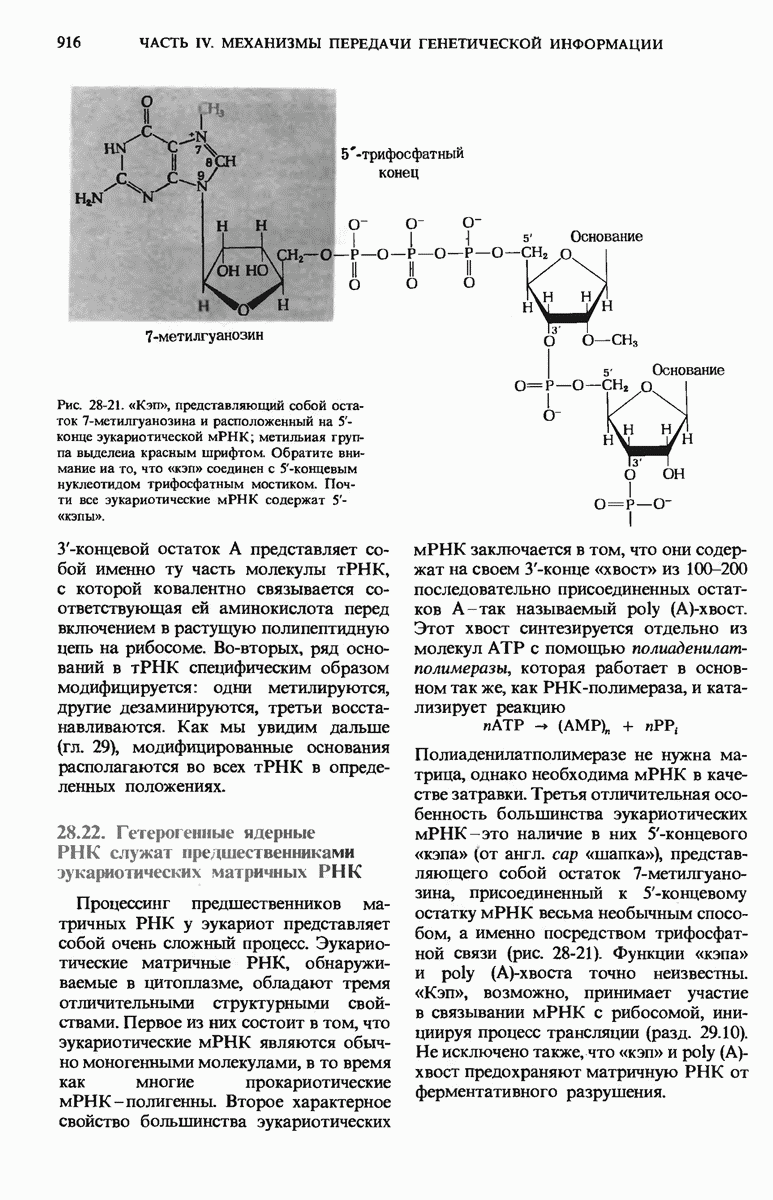
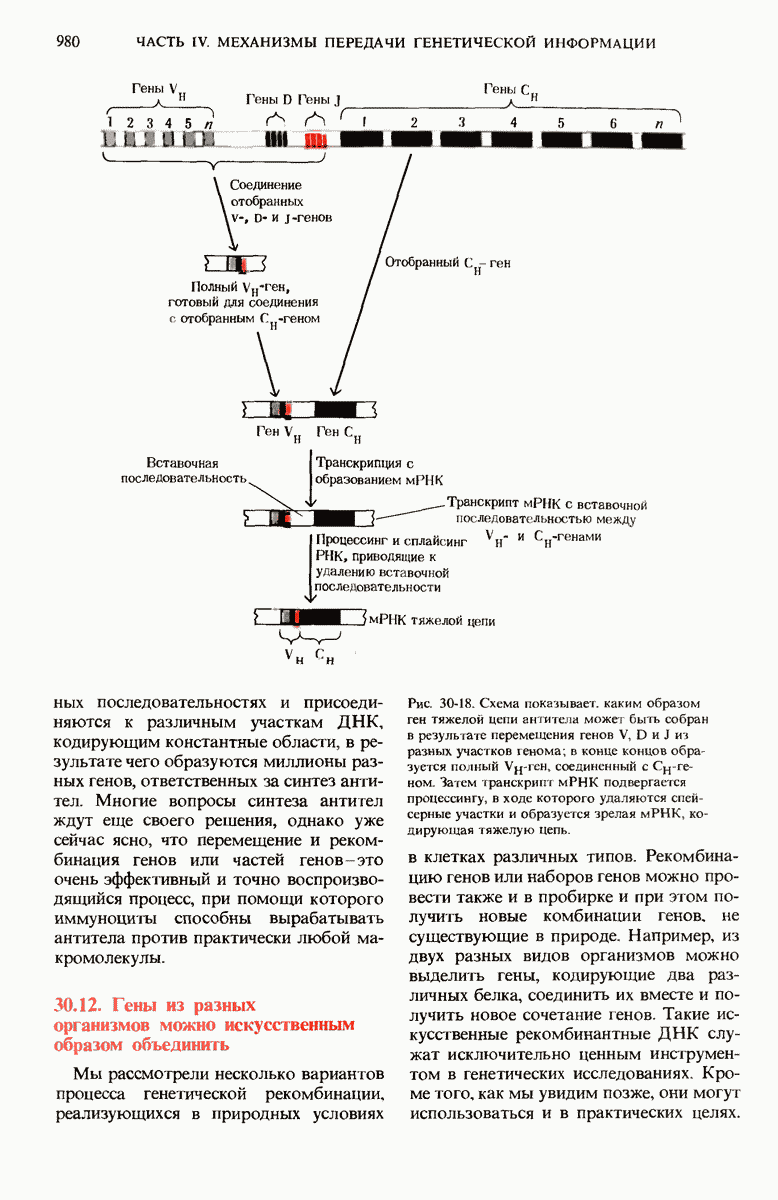
Большая часть нуклеотидной последовательности lac-оперона Е. coli уже определена. Известна полная нуклеотидная последовательность оператора и промотора.
Весь промотор состоит, как оказалось, из 85 нуклеотидных пар, причем на долю САР-участка приходится около 38, а на долю участка первоначального связывания РНК-полимеразы - около 40 пар оснований (рис. 29-28).
Кроме ряда оперонов с их регуляторными генами бактерии обладают и другими механизмами регуляции белкового синтеза. Некоторые из них позволяют осуществлять регуляцию не по принципу «все или ничего», а за счет постепенной аттенуации, т. е. снижения скорости синтеза белка. Механизмы, чувствительные к концентрации аммиака или других источников азота, дают возможность бактериям приспособить свое белковое хозяйство к скудным условиям существования. Из сказанного ясно, что бактерии обладают тончайшими механизмами регуляции синтеза своих ферментов, позволяющими им оптимизировать свой метаболизм в соответствии с принципом максимальной экономии.