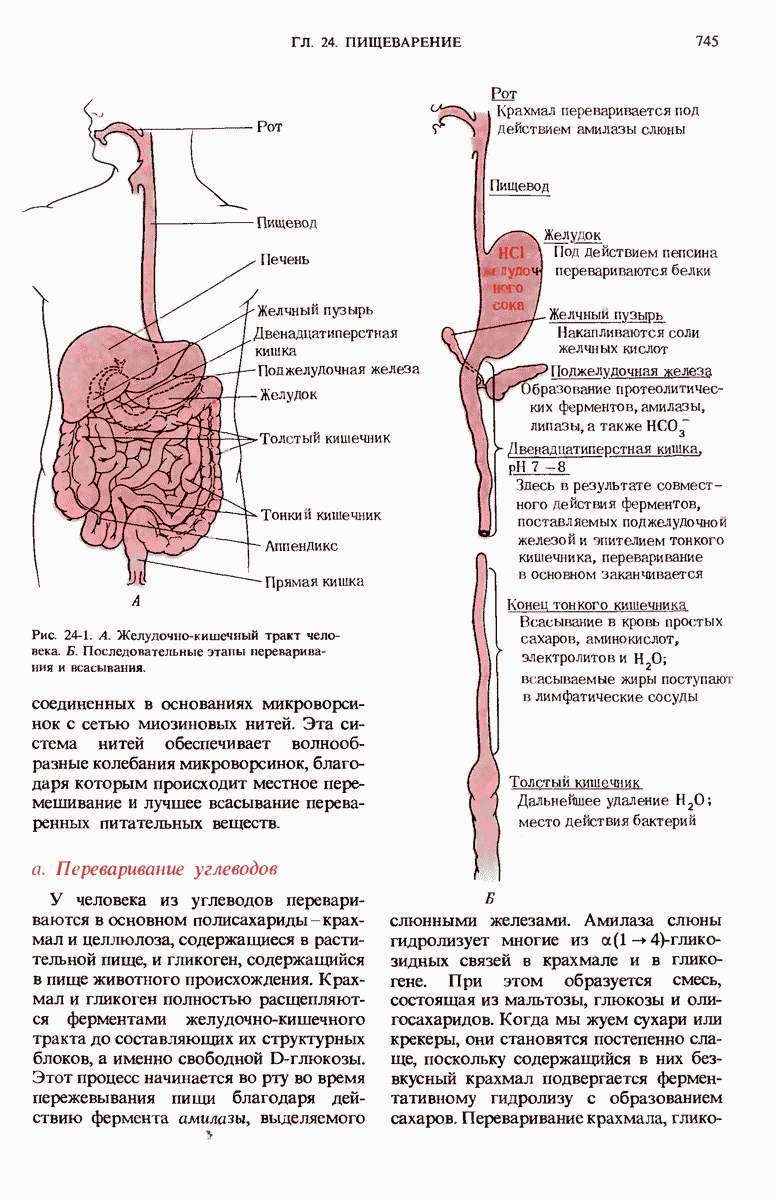
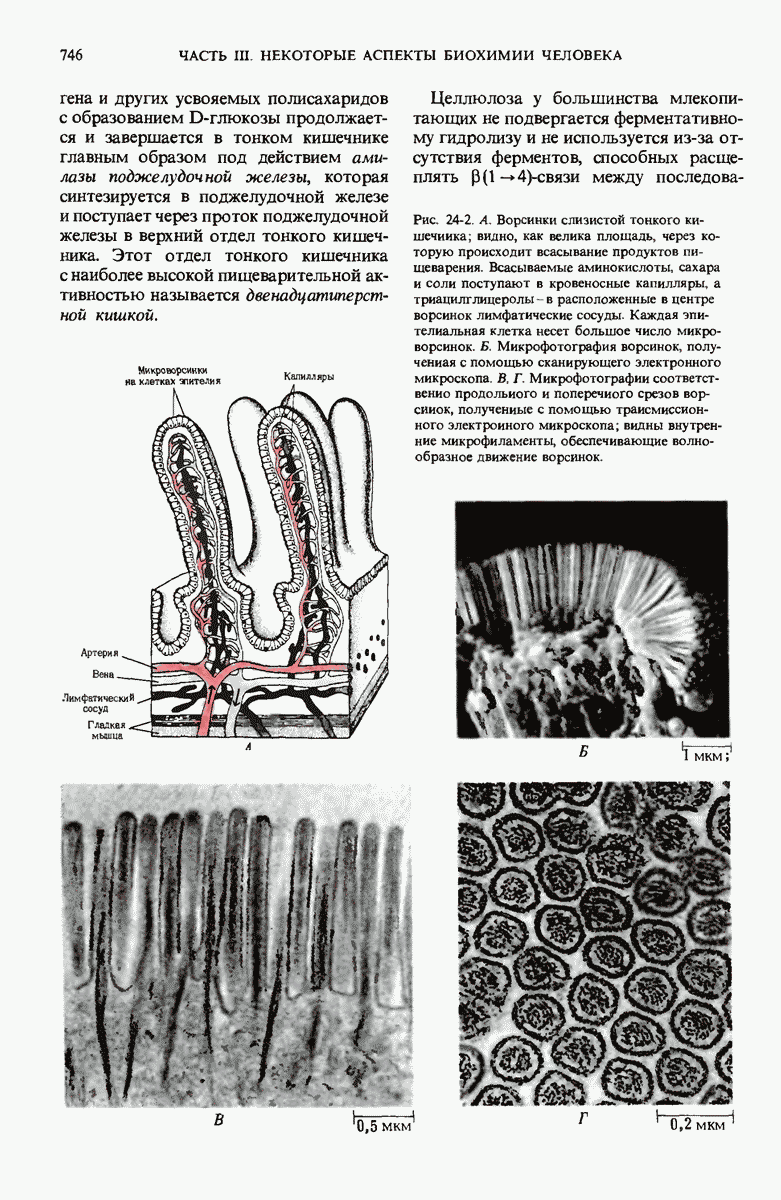
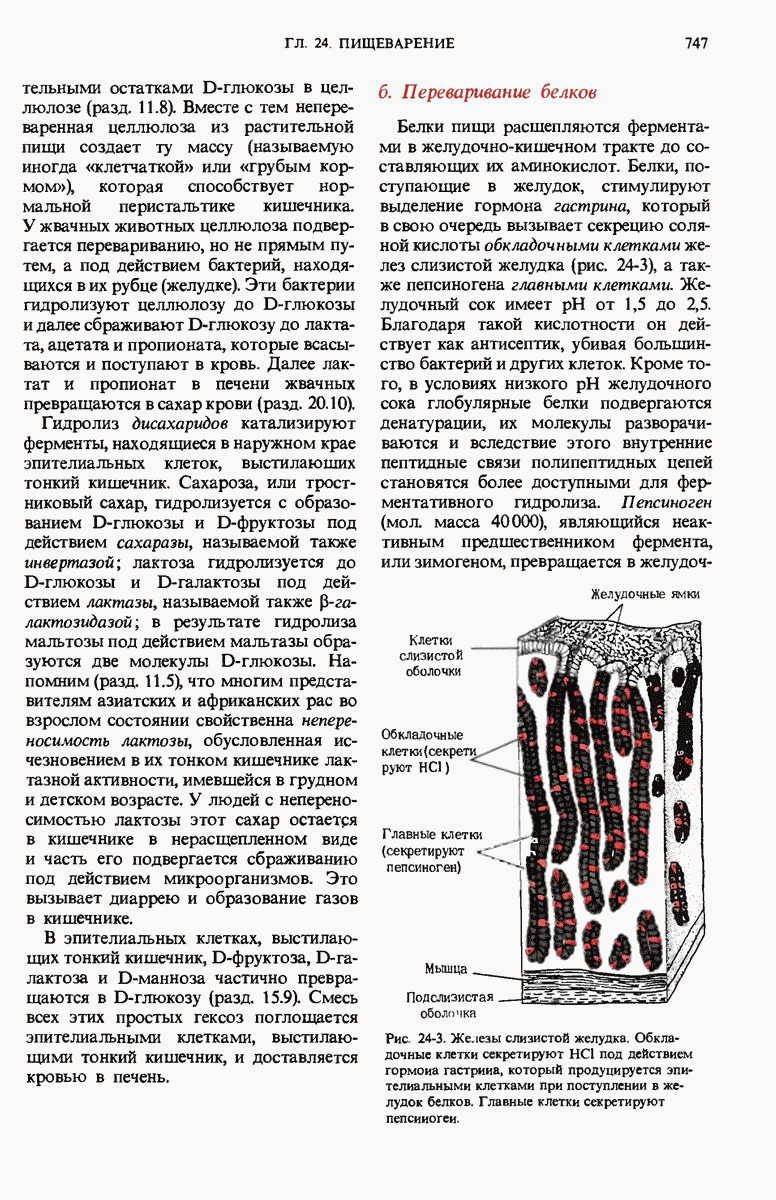
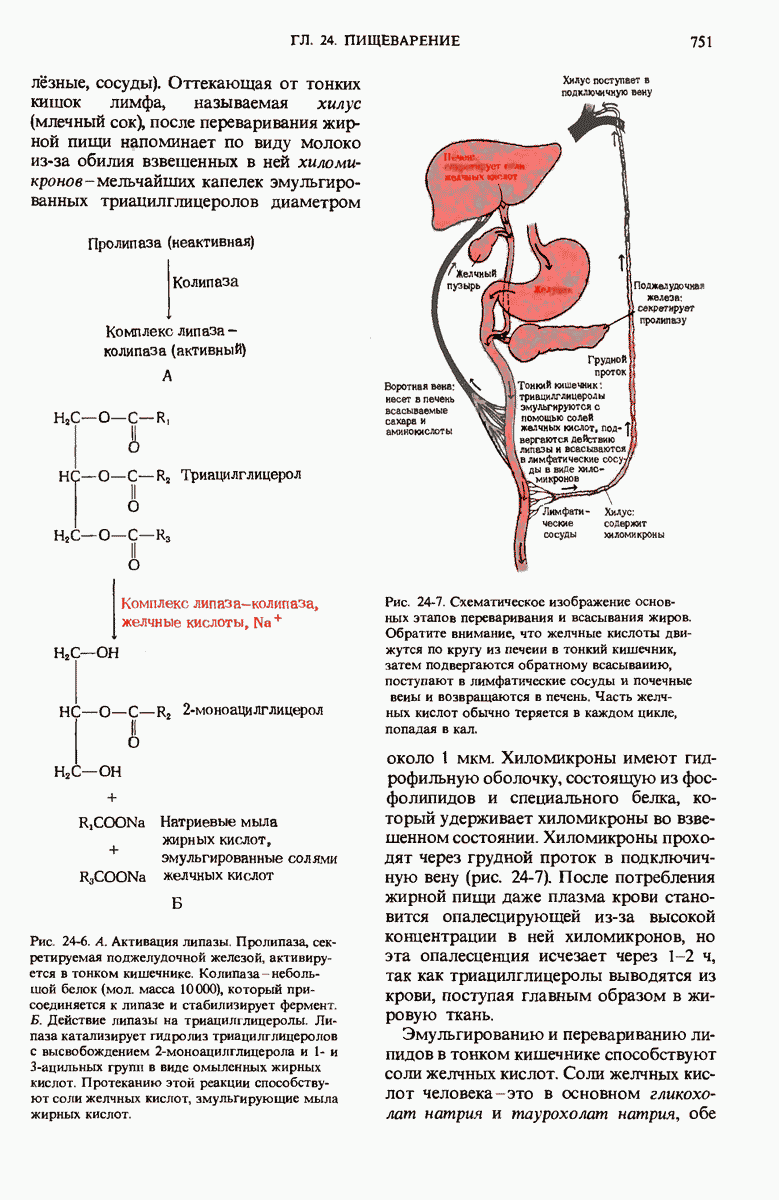
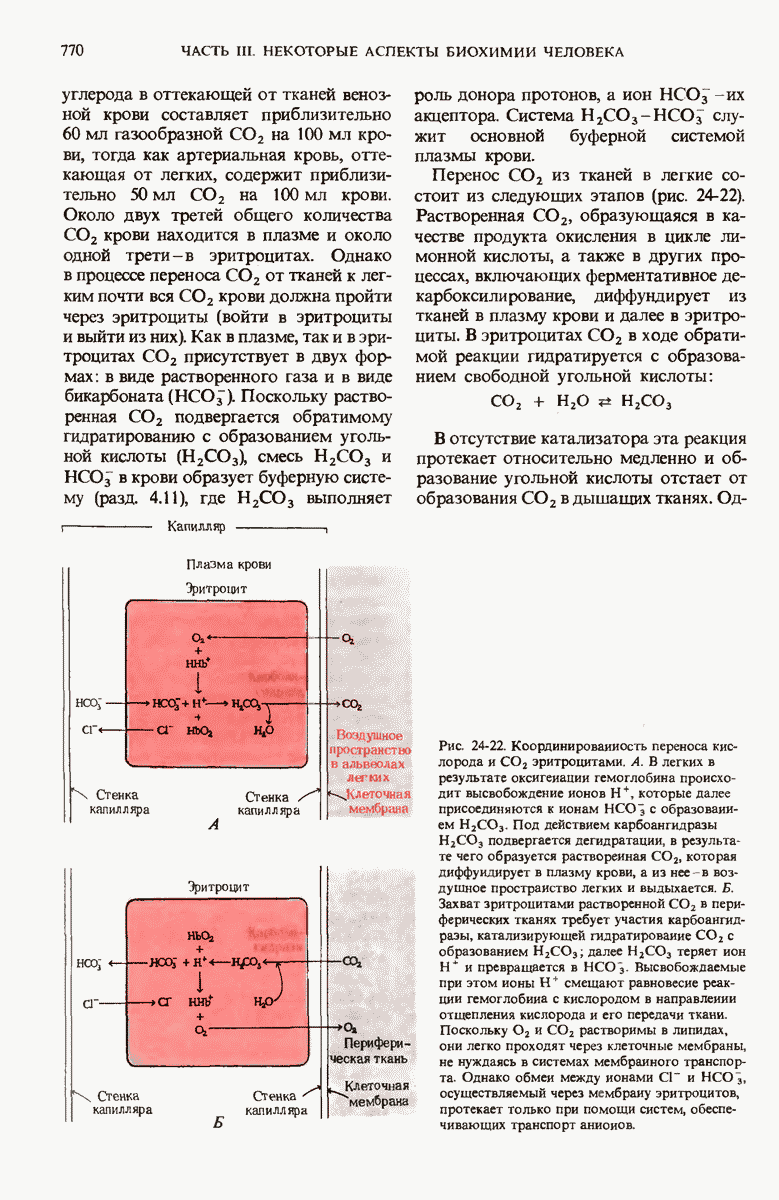
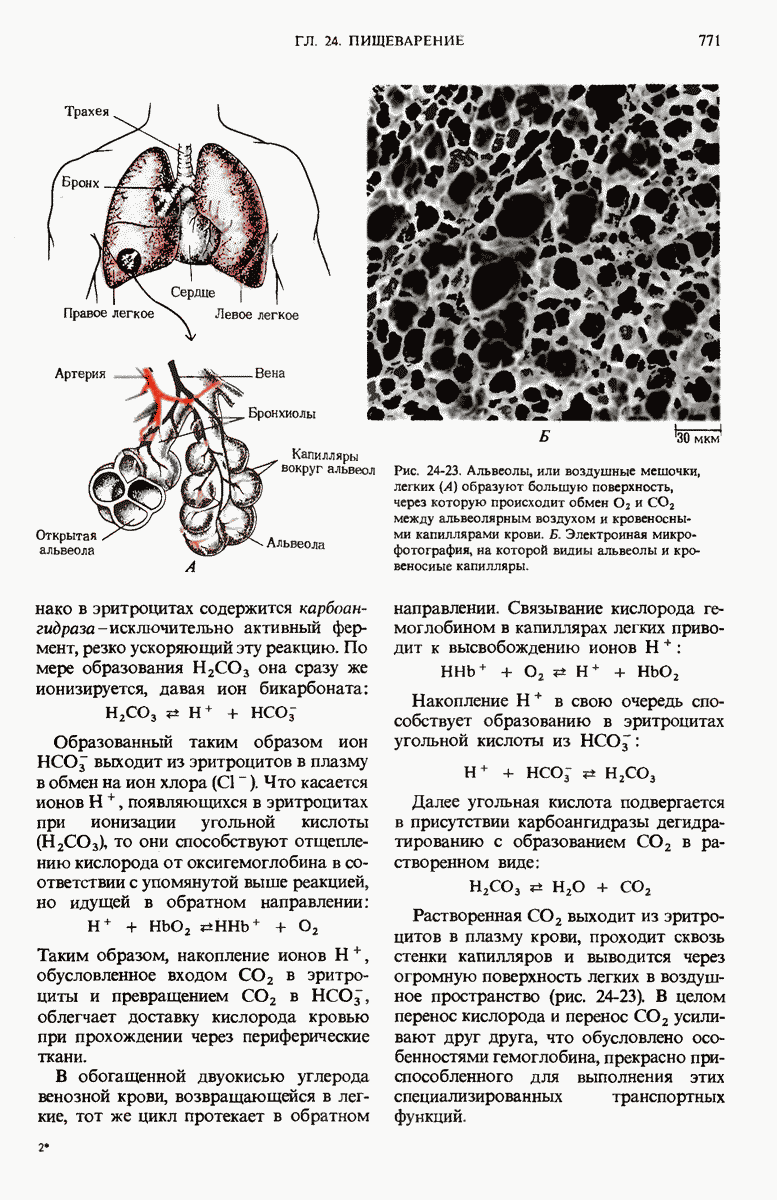
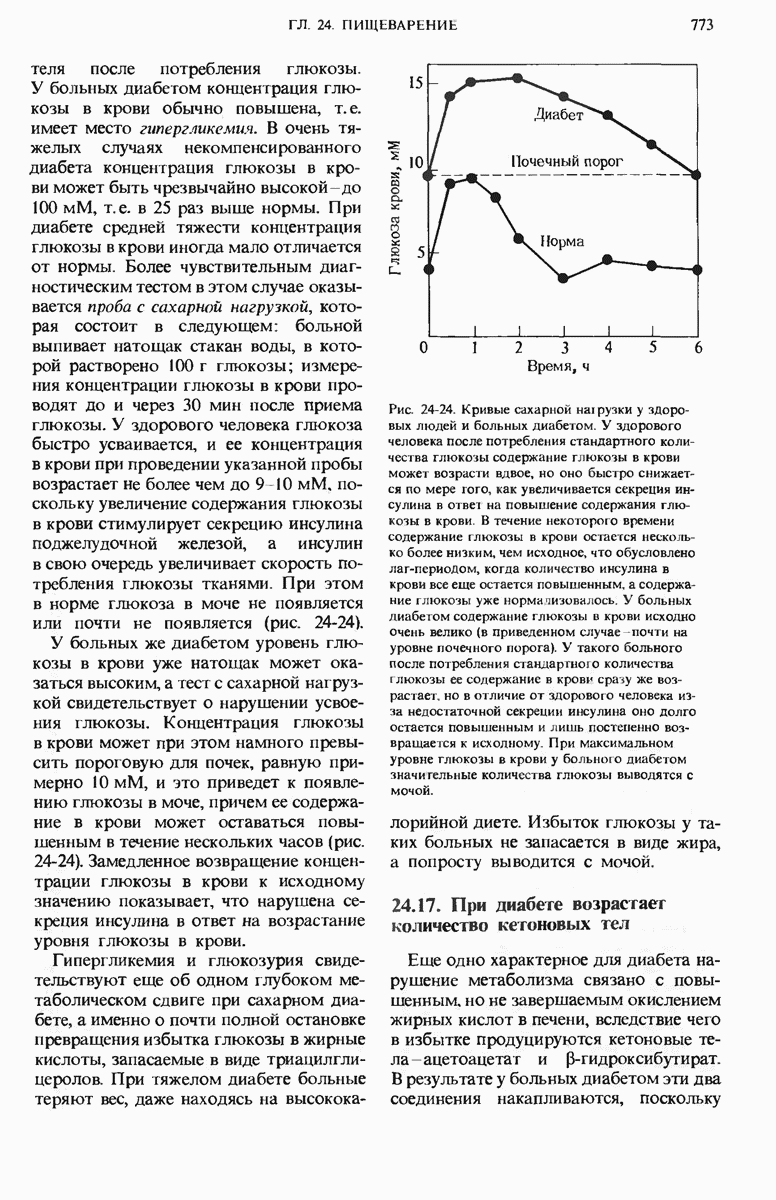
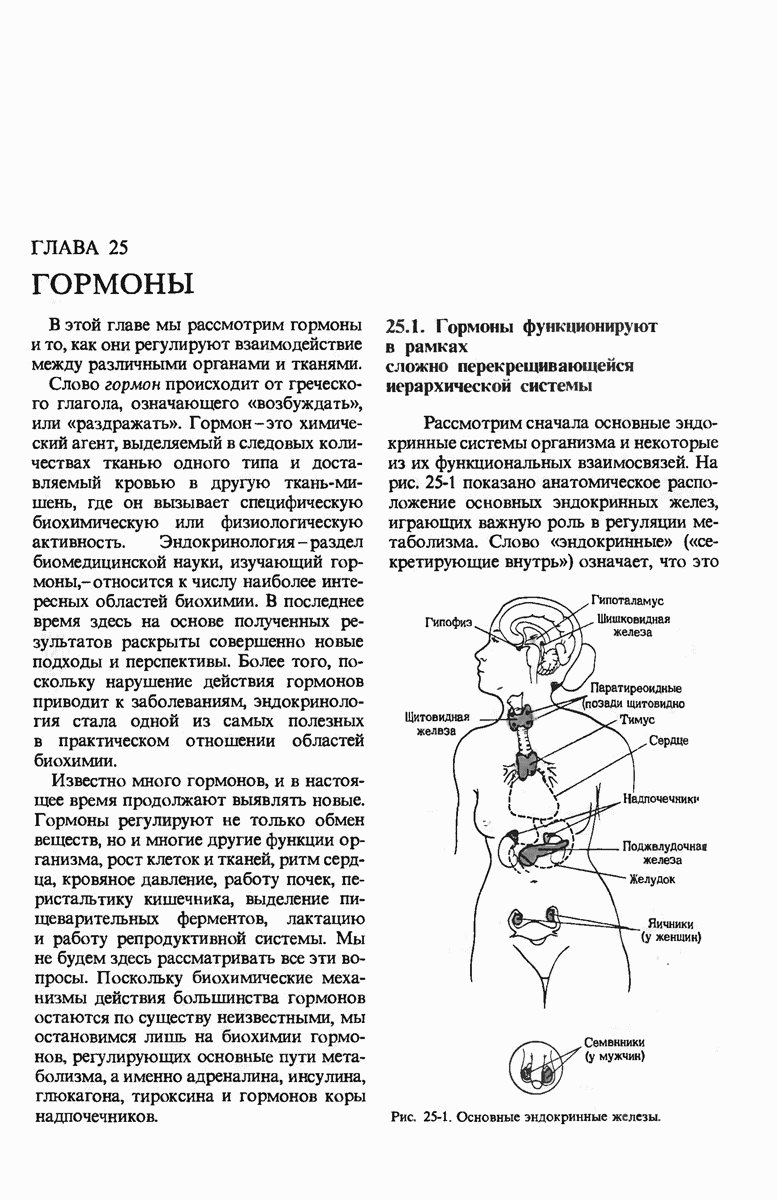
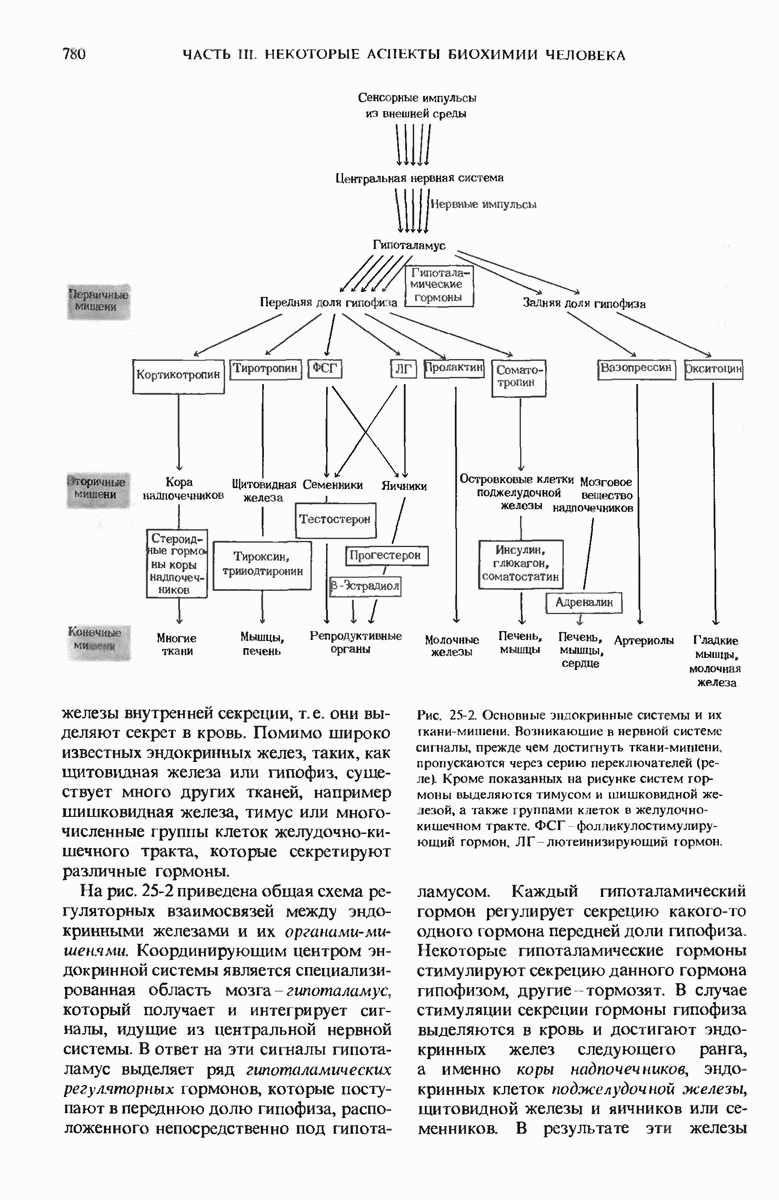
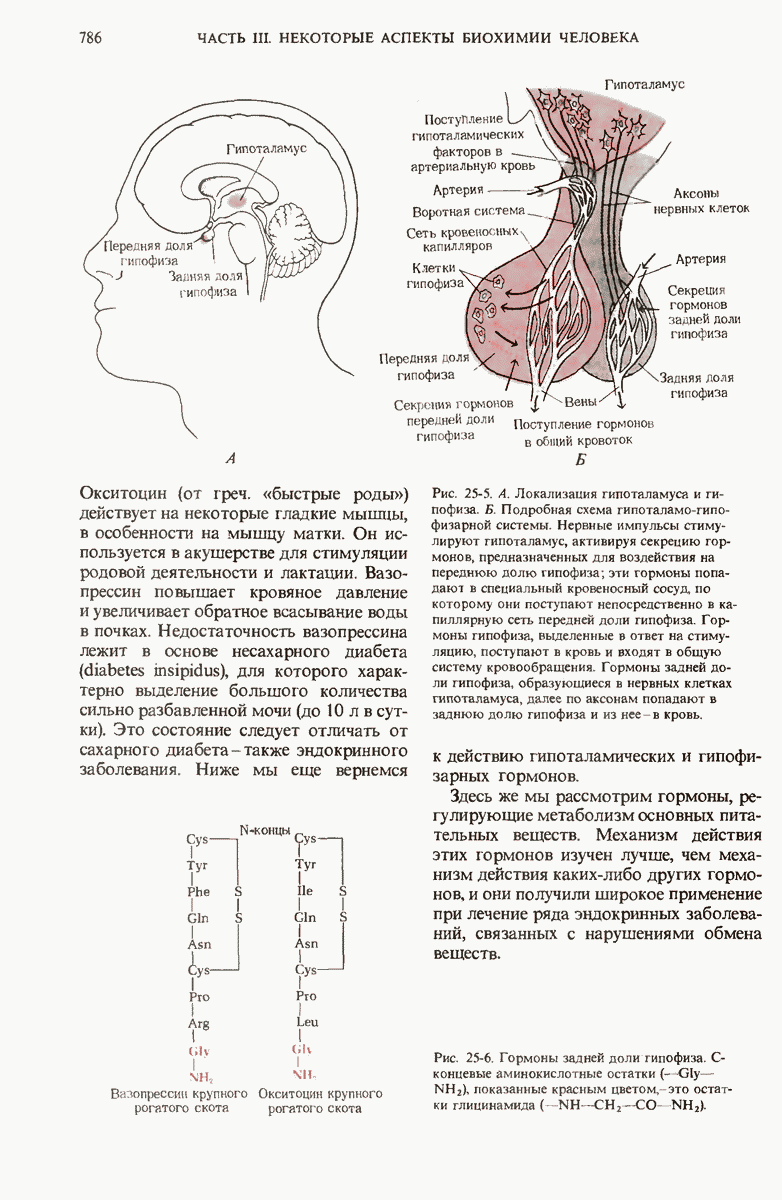
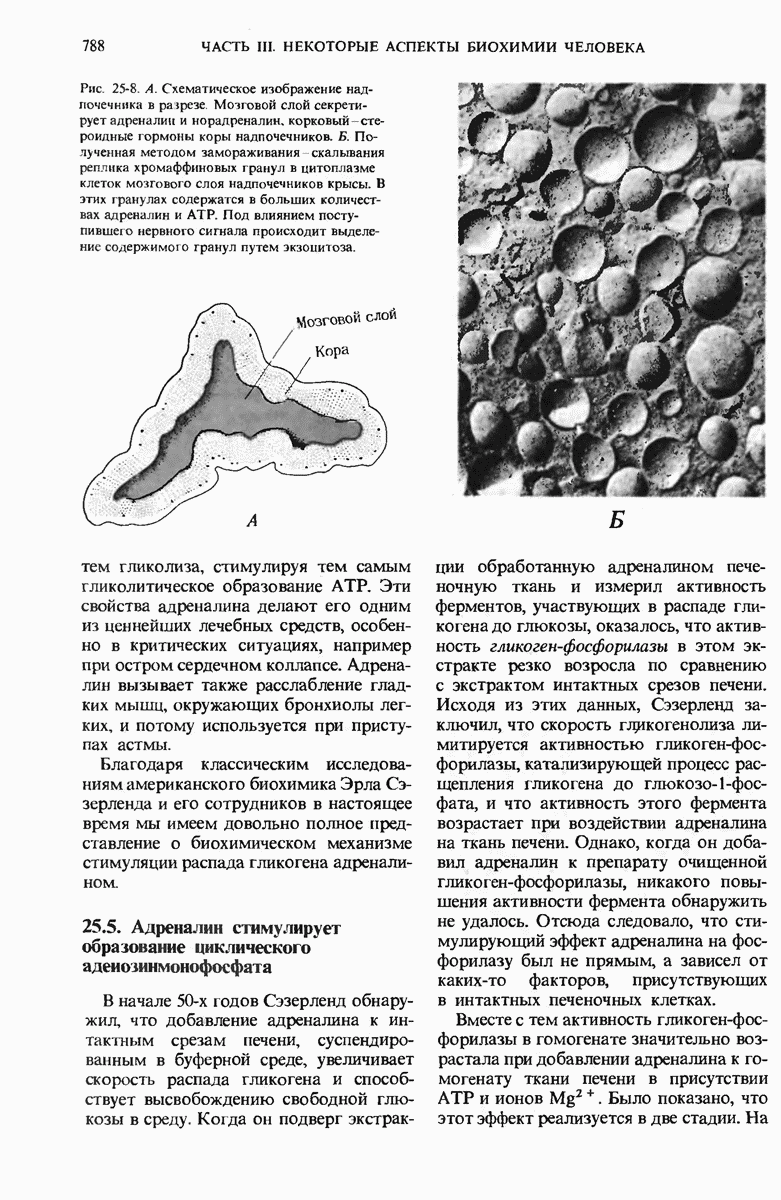
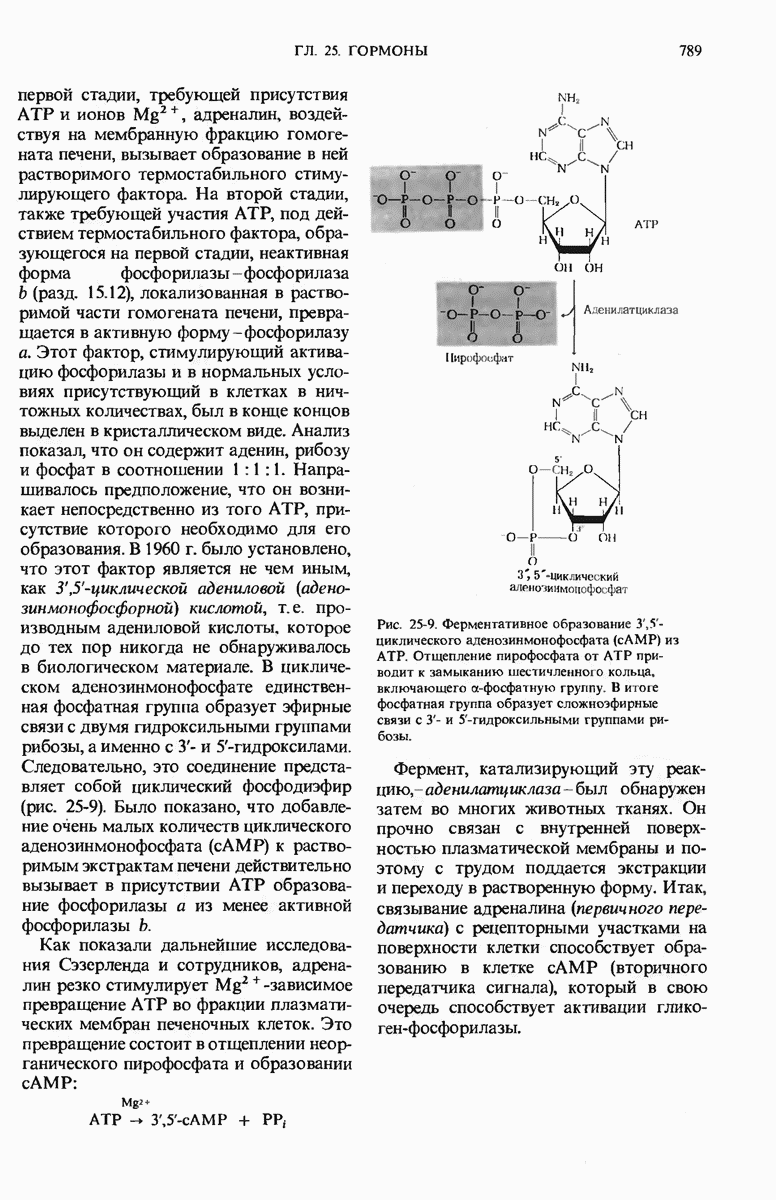
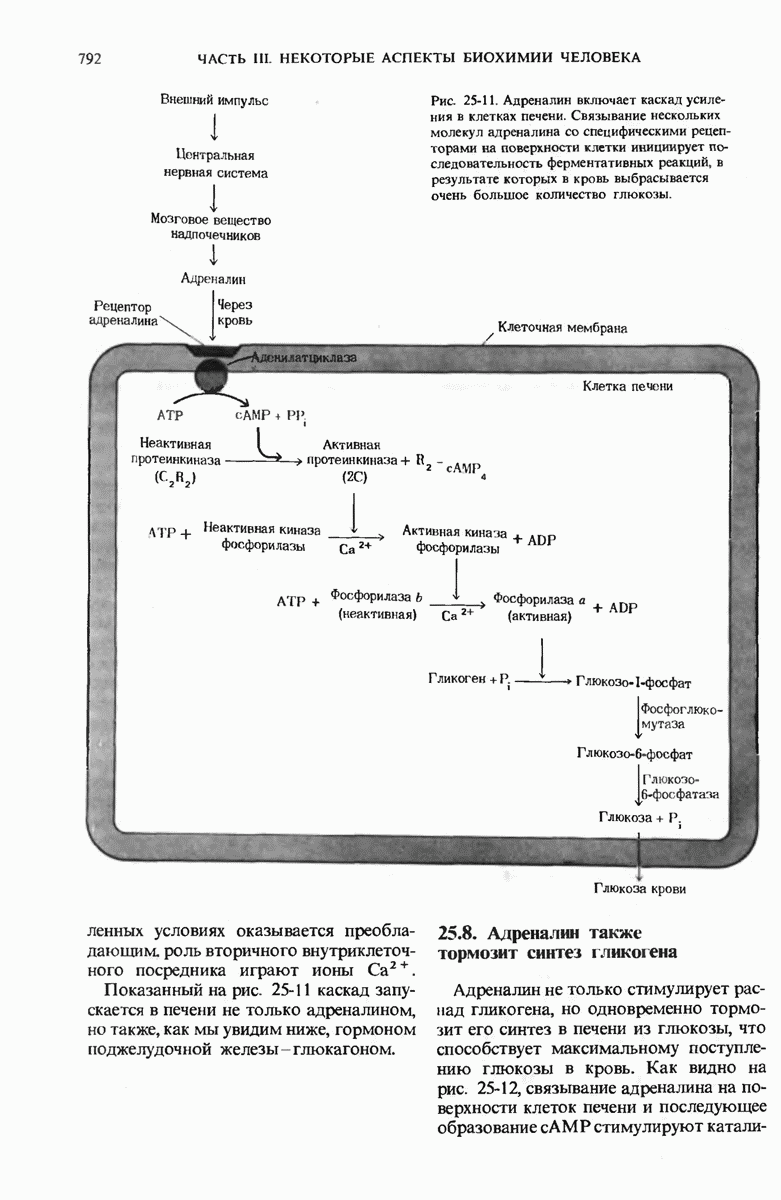
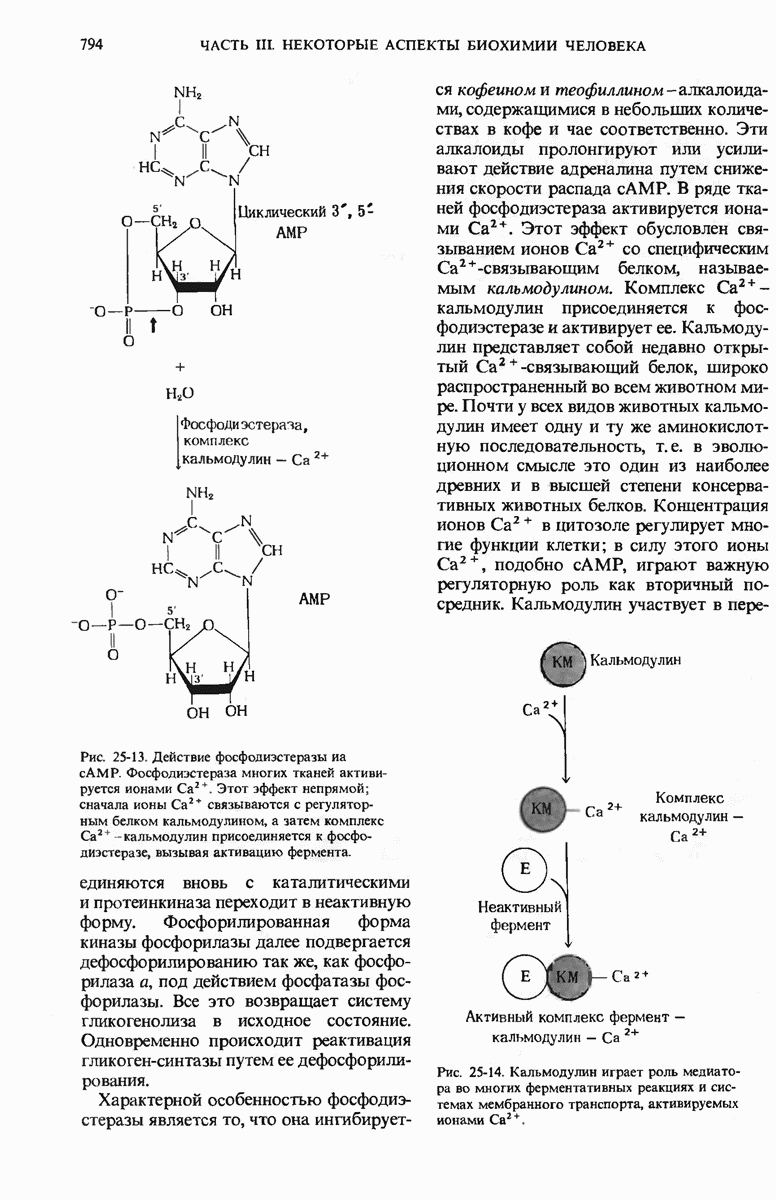
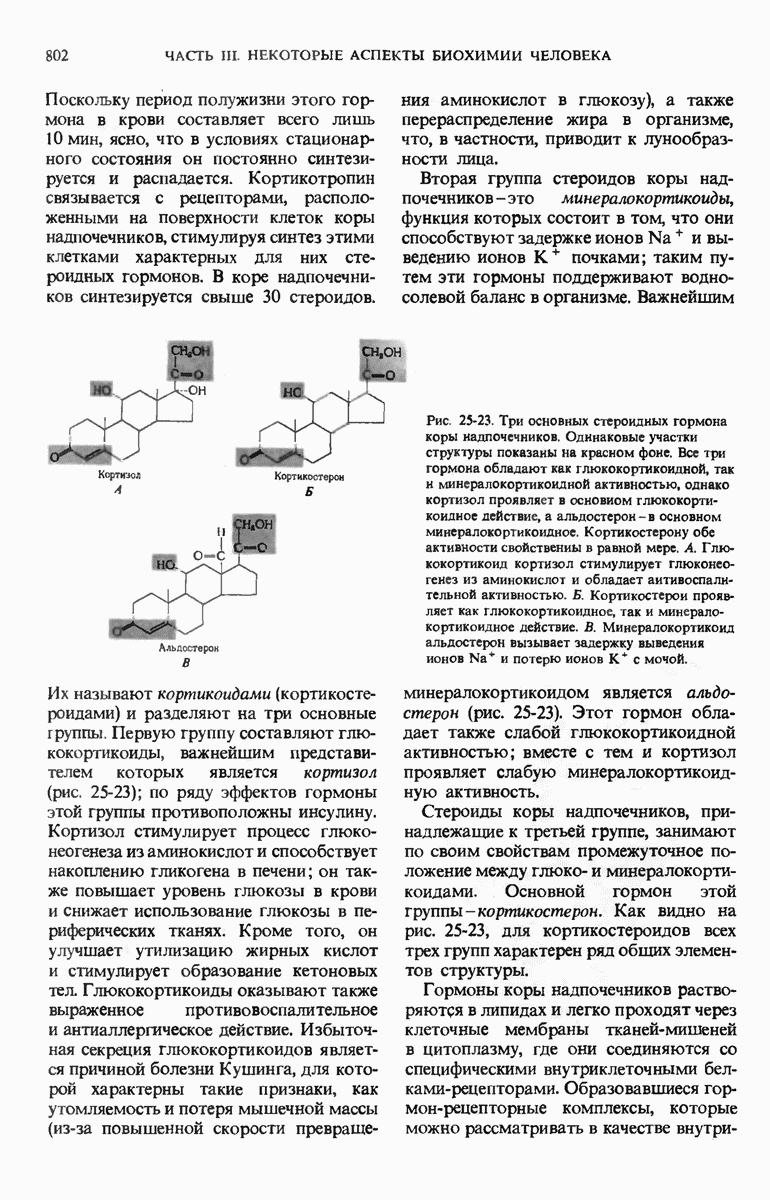
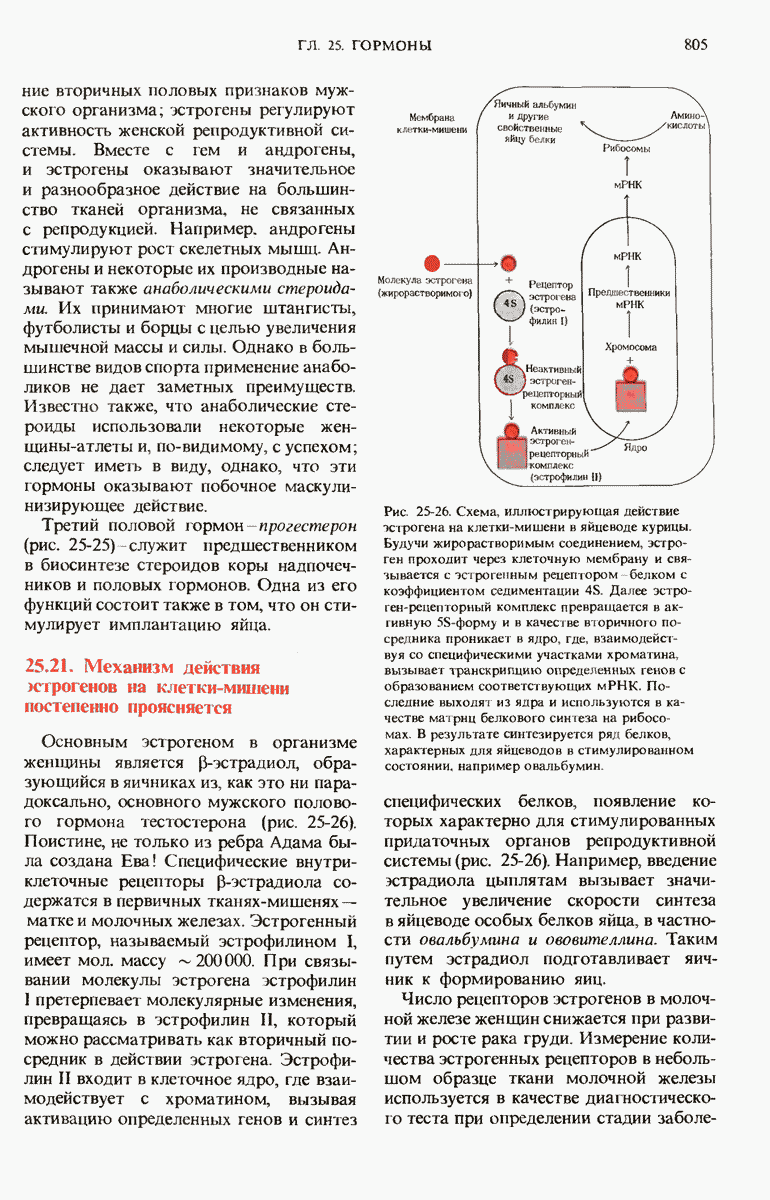
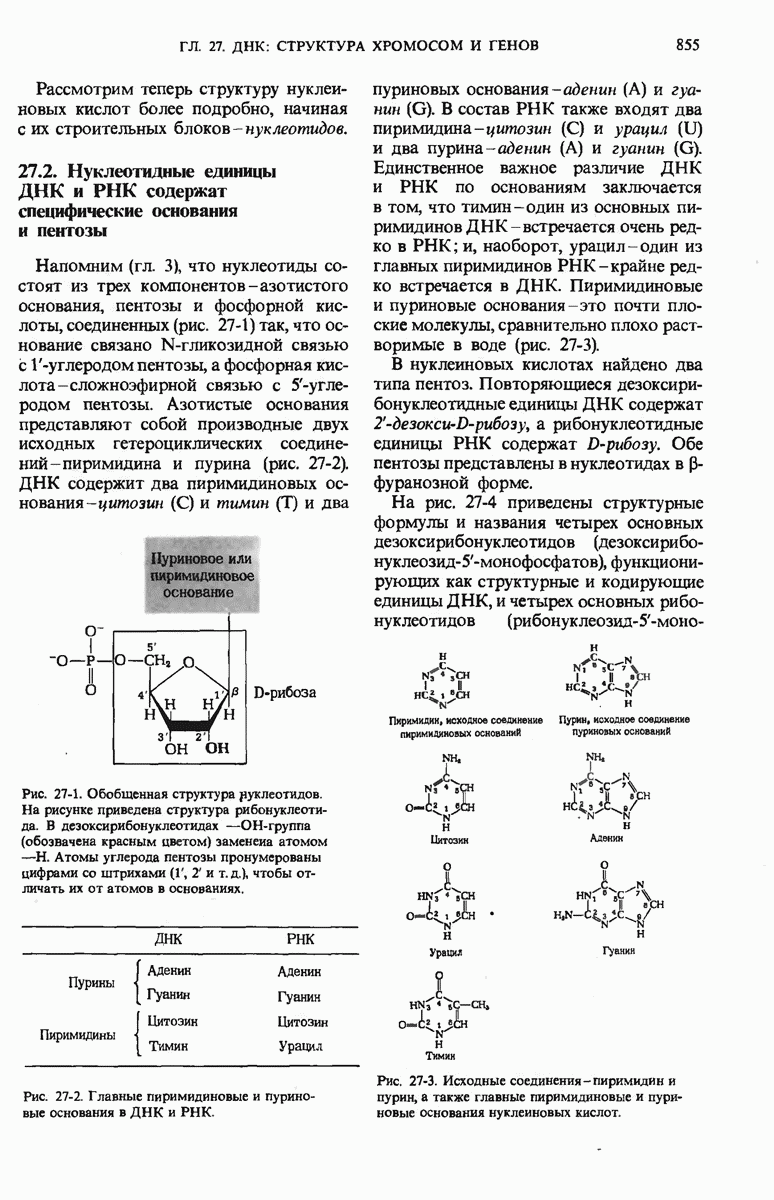
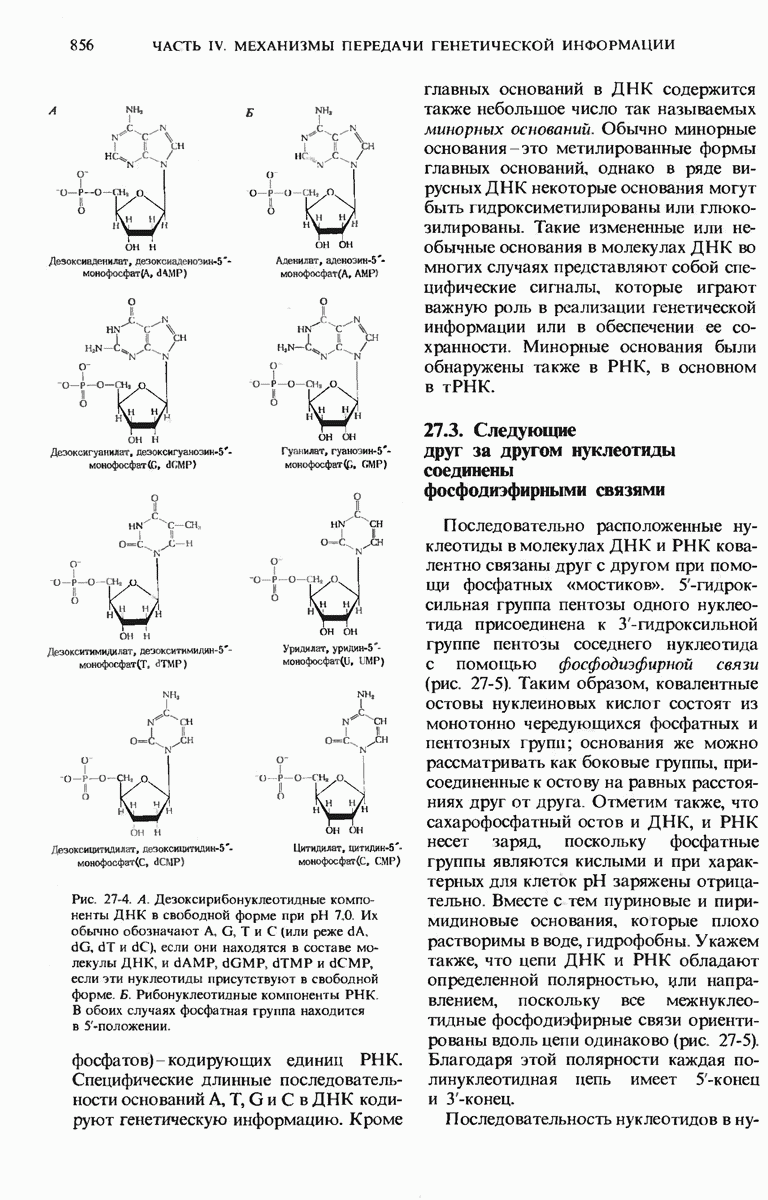
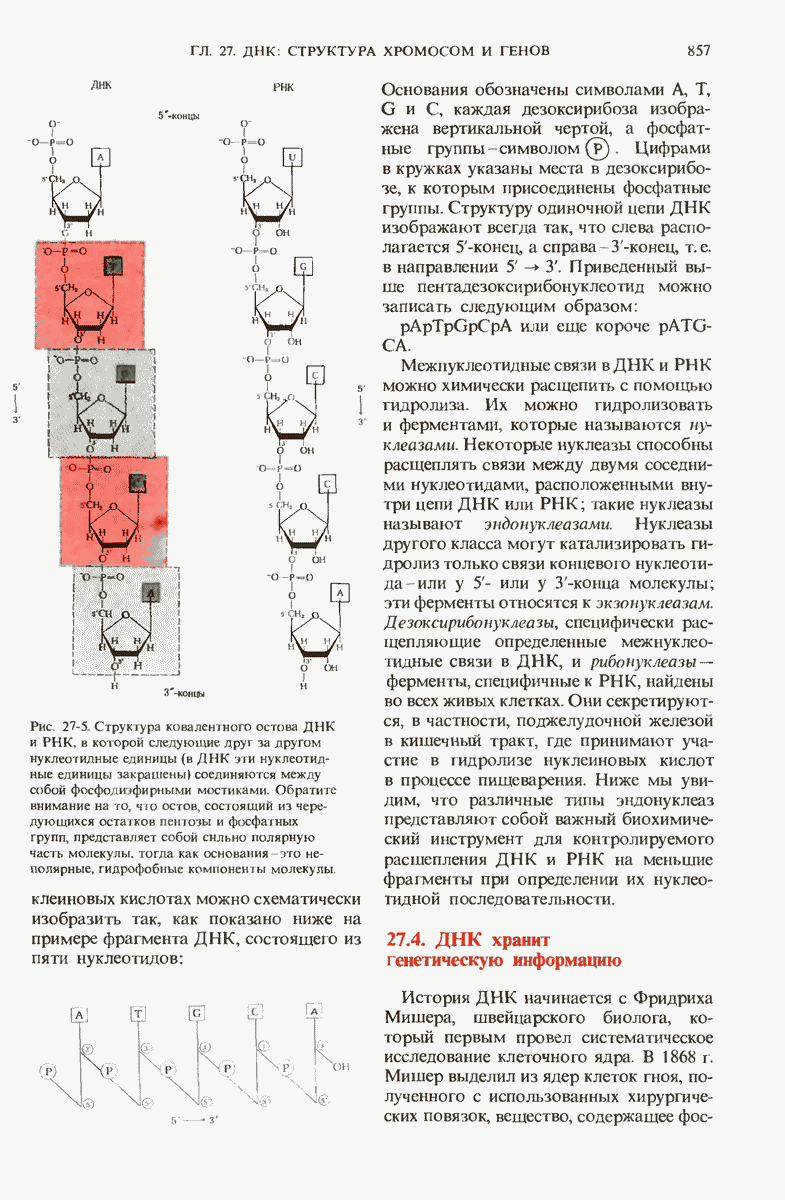
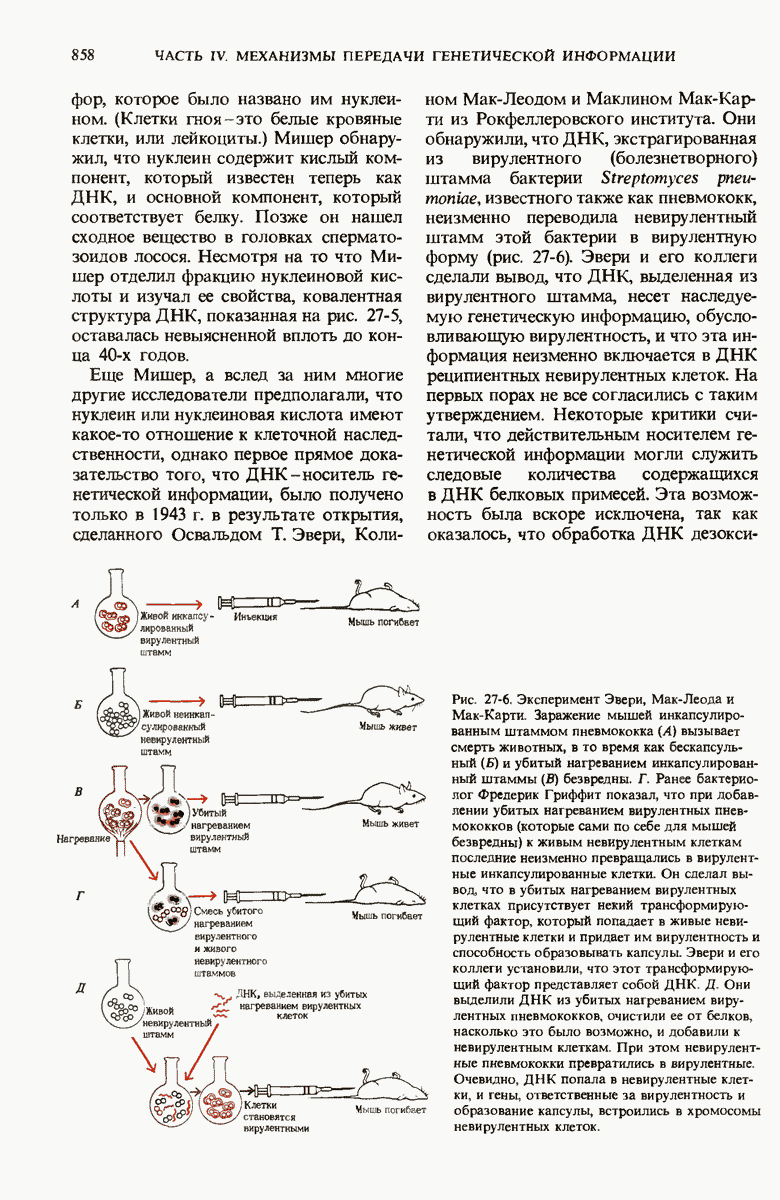
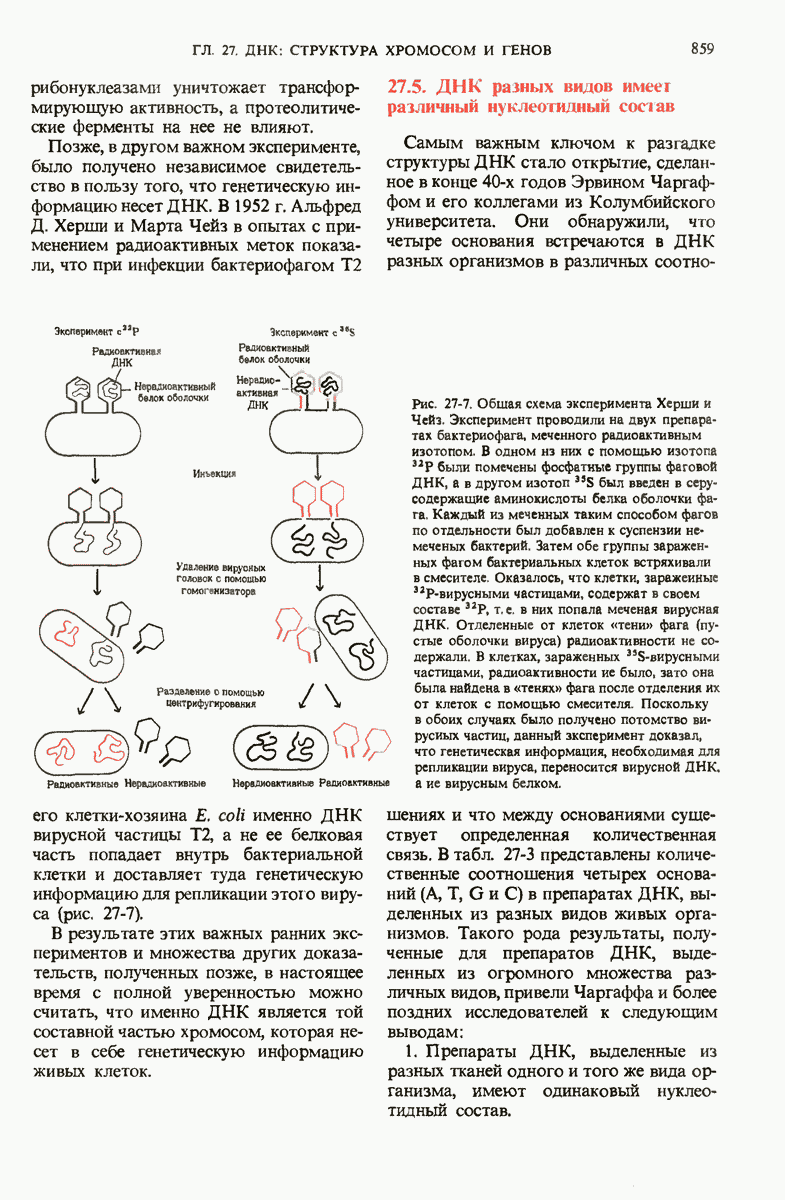
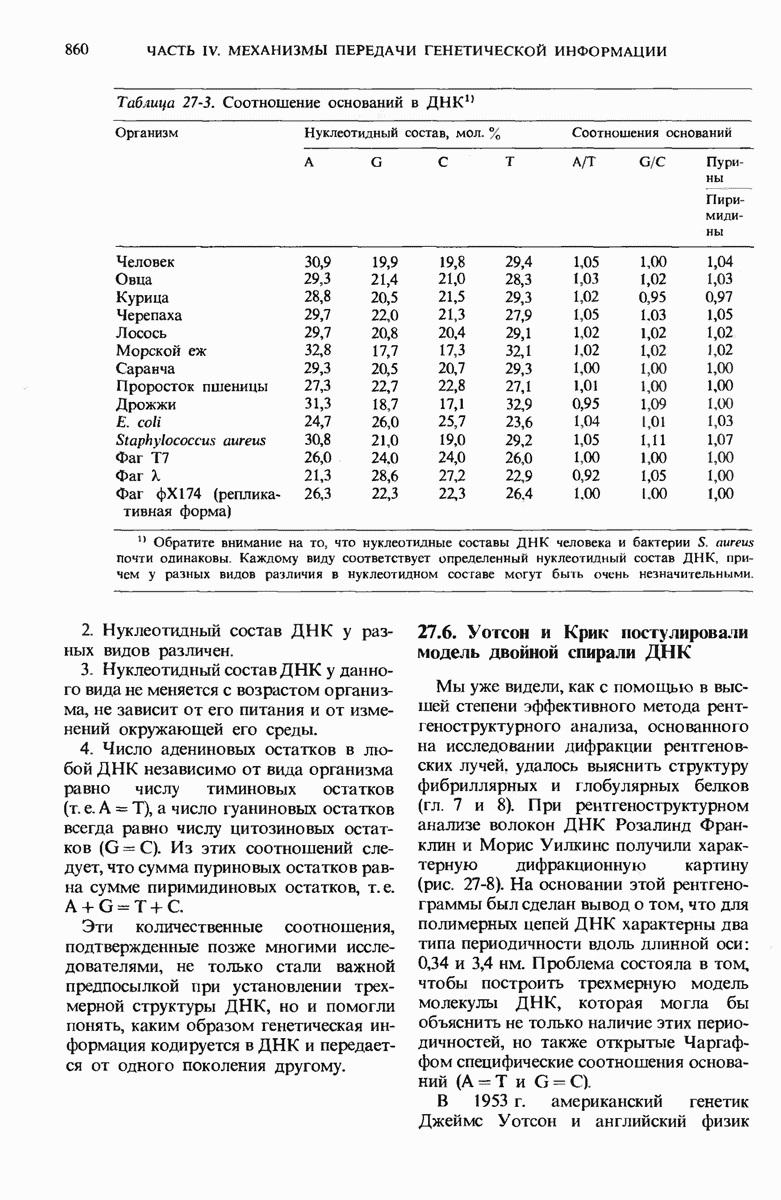
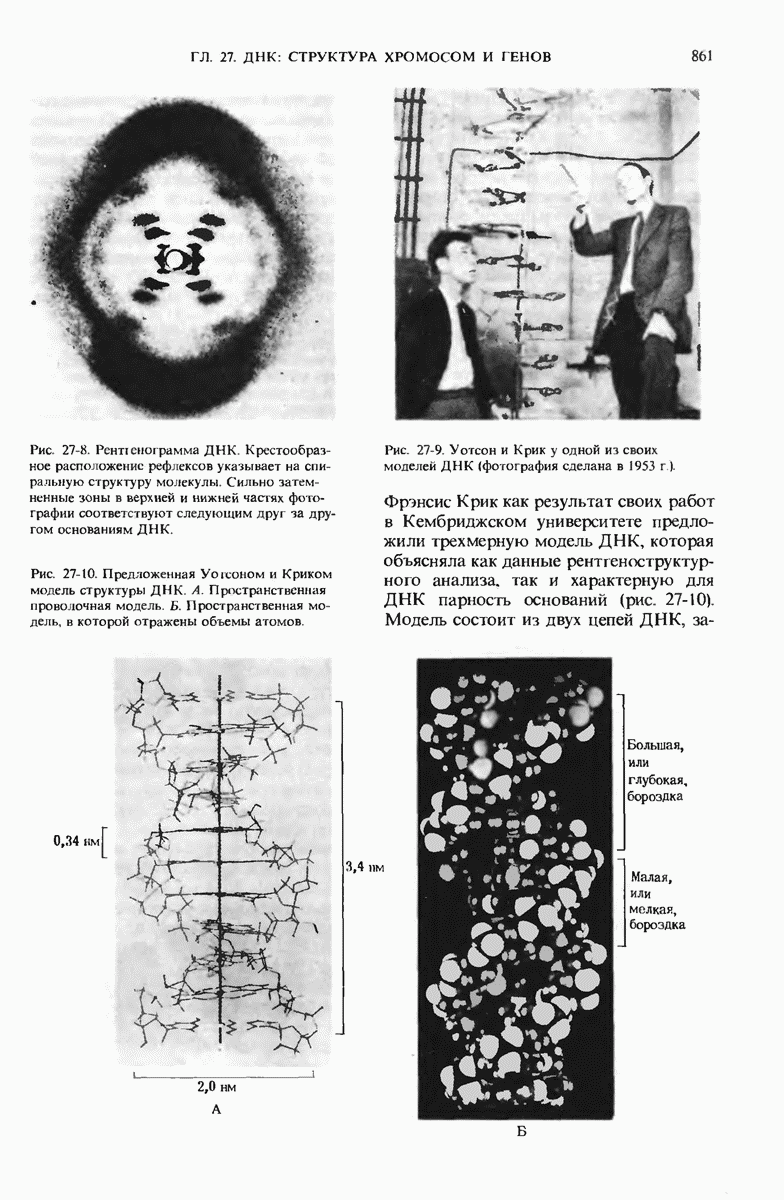
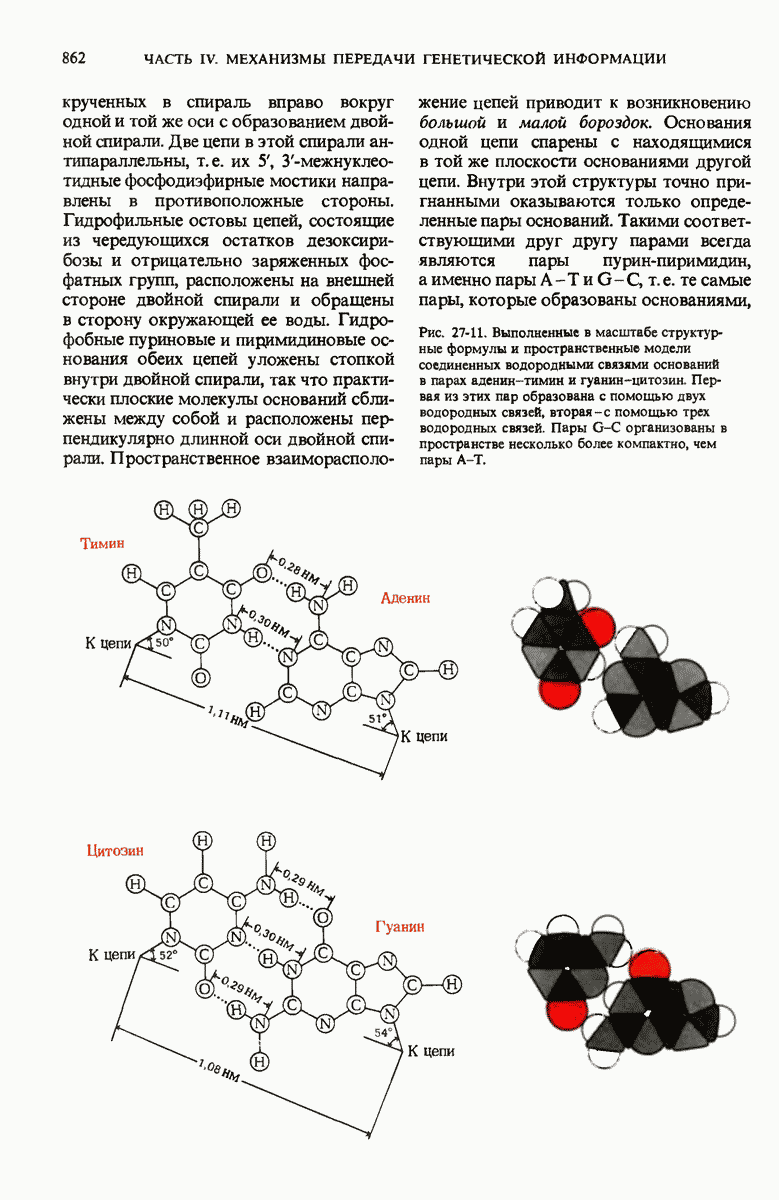
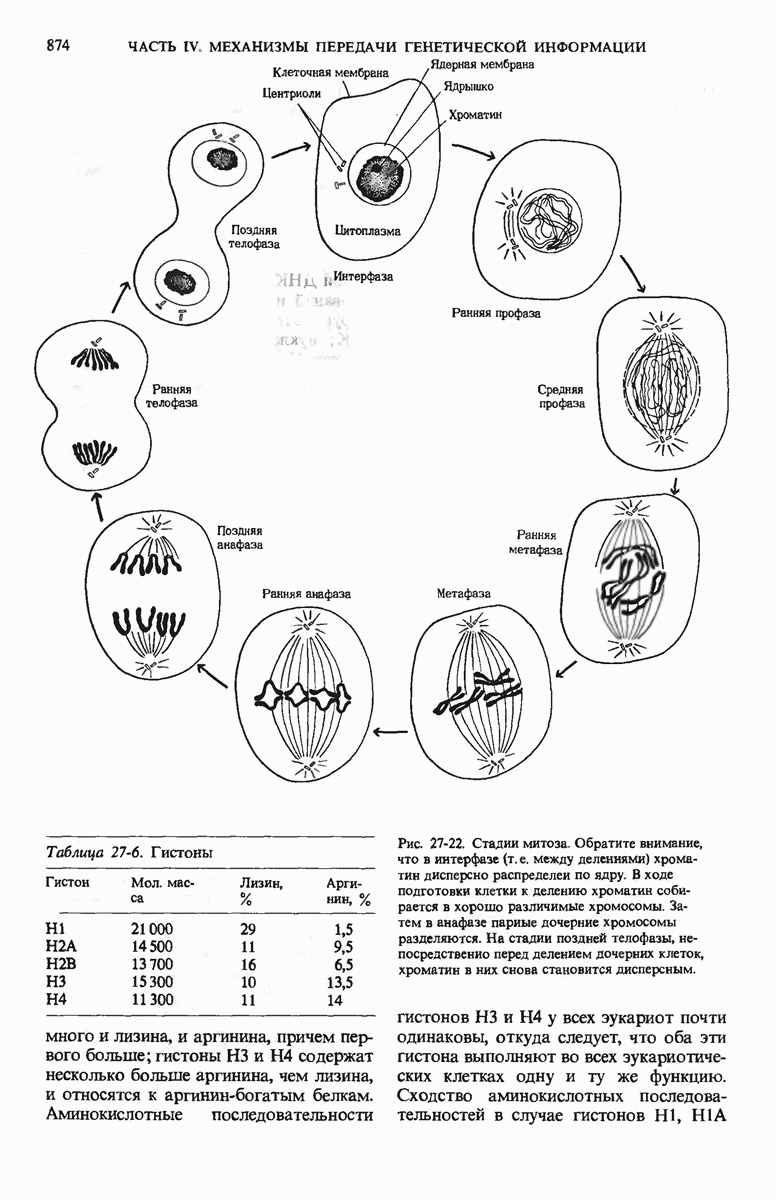
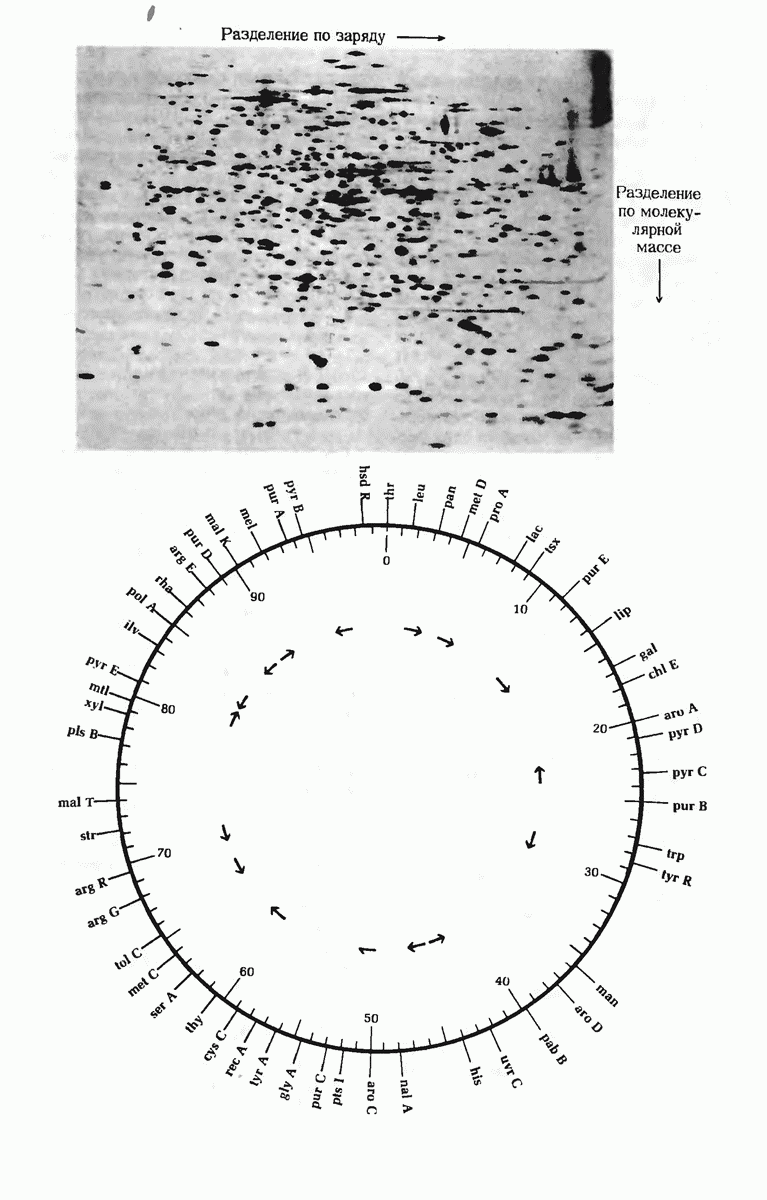
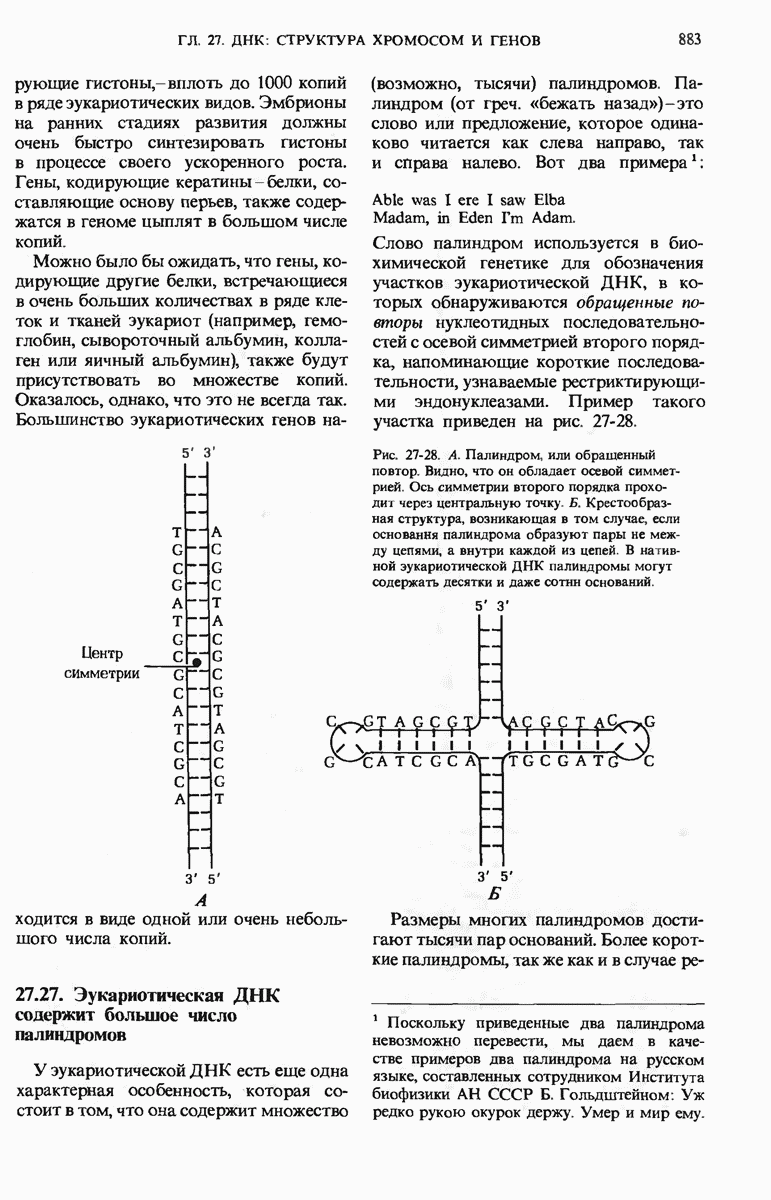
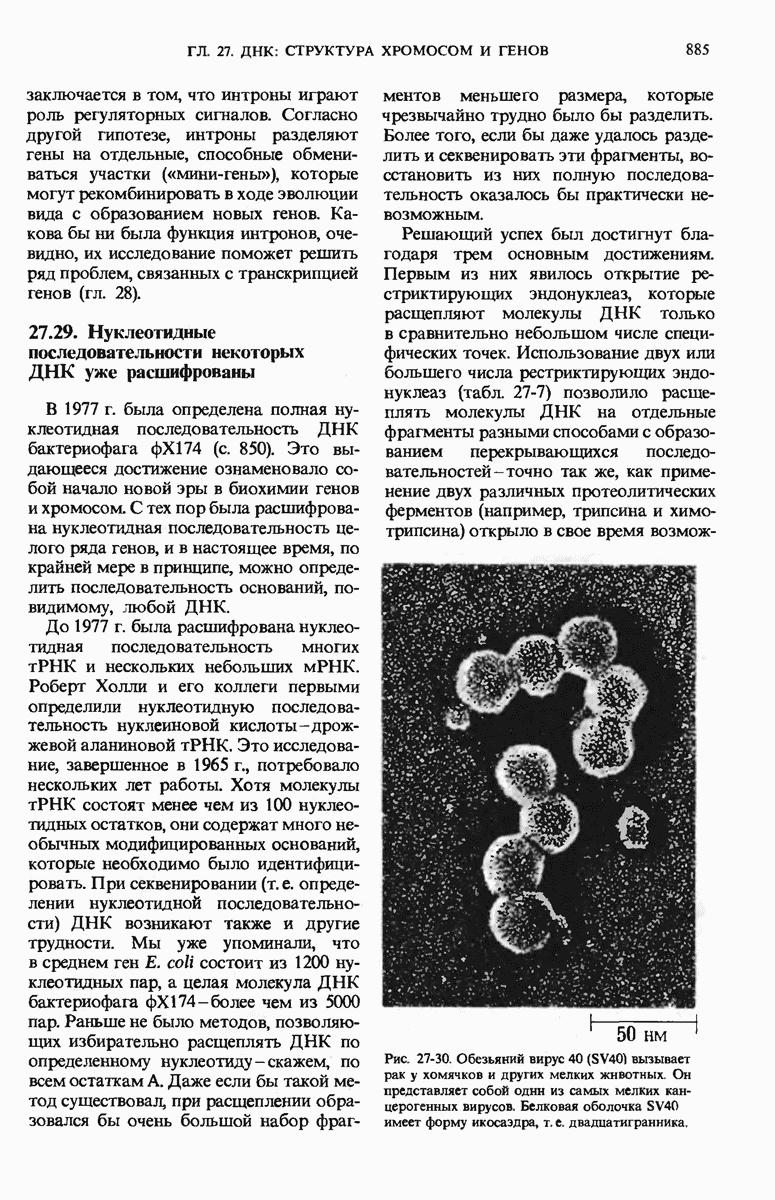
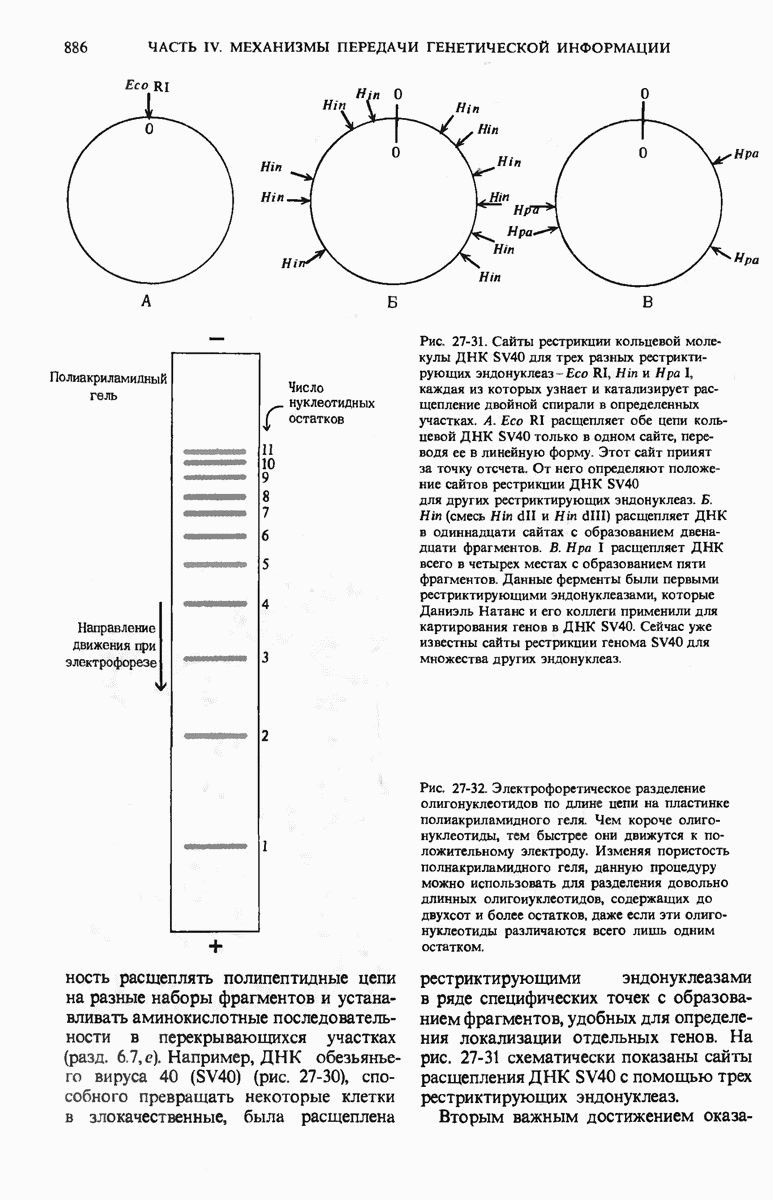
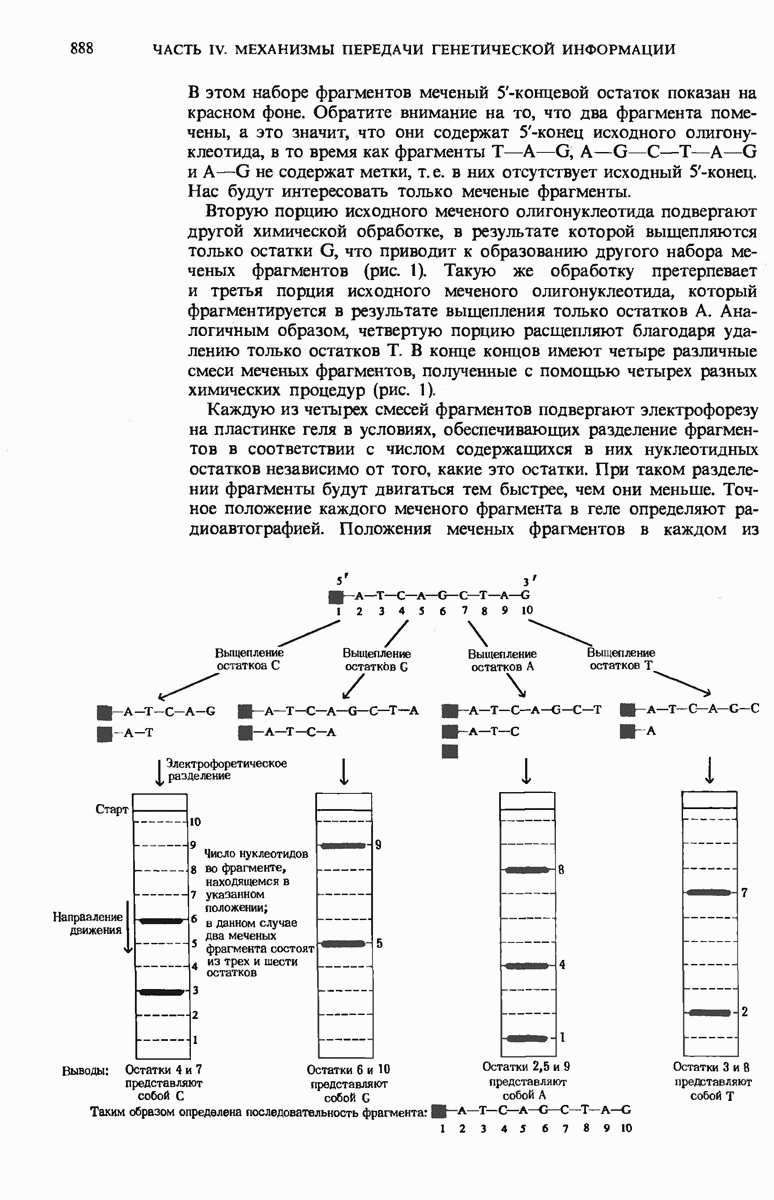
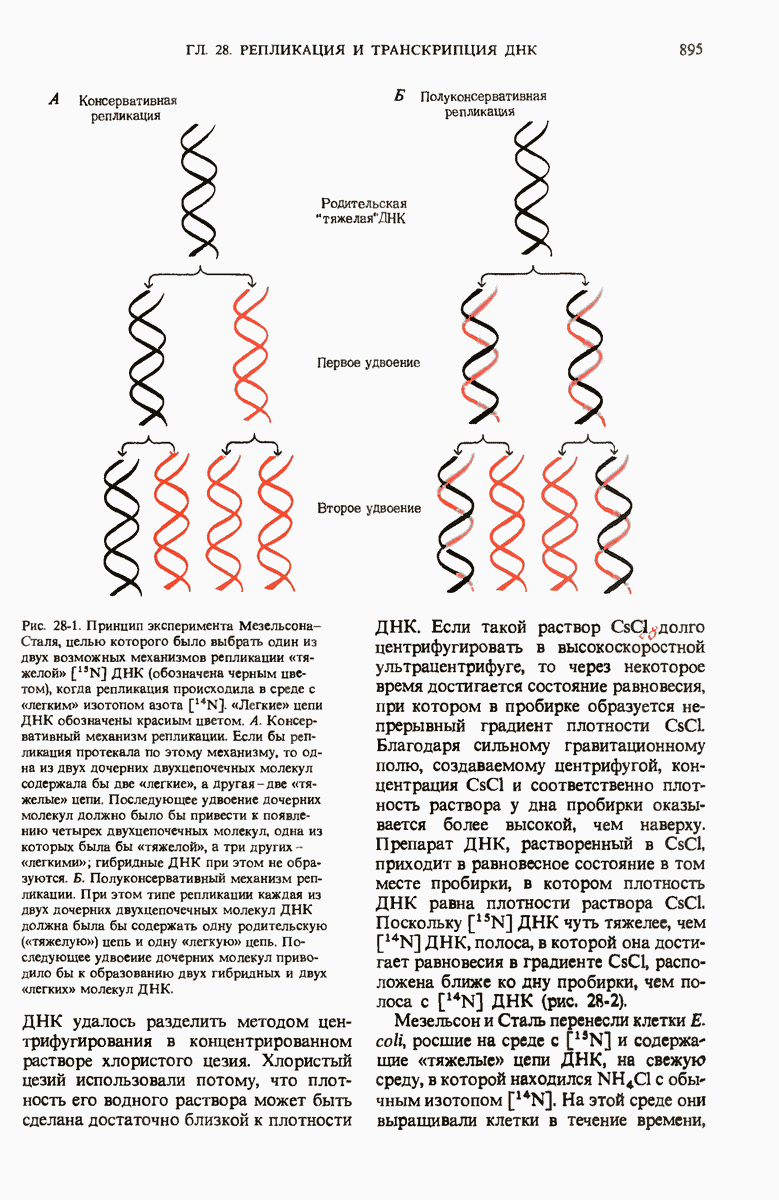
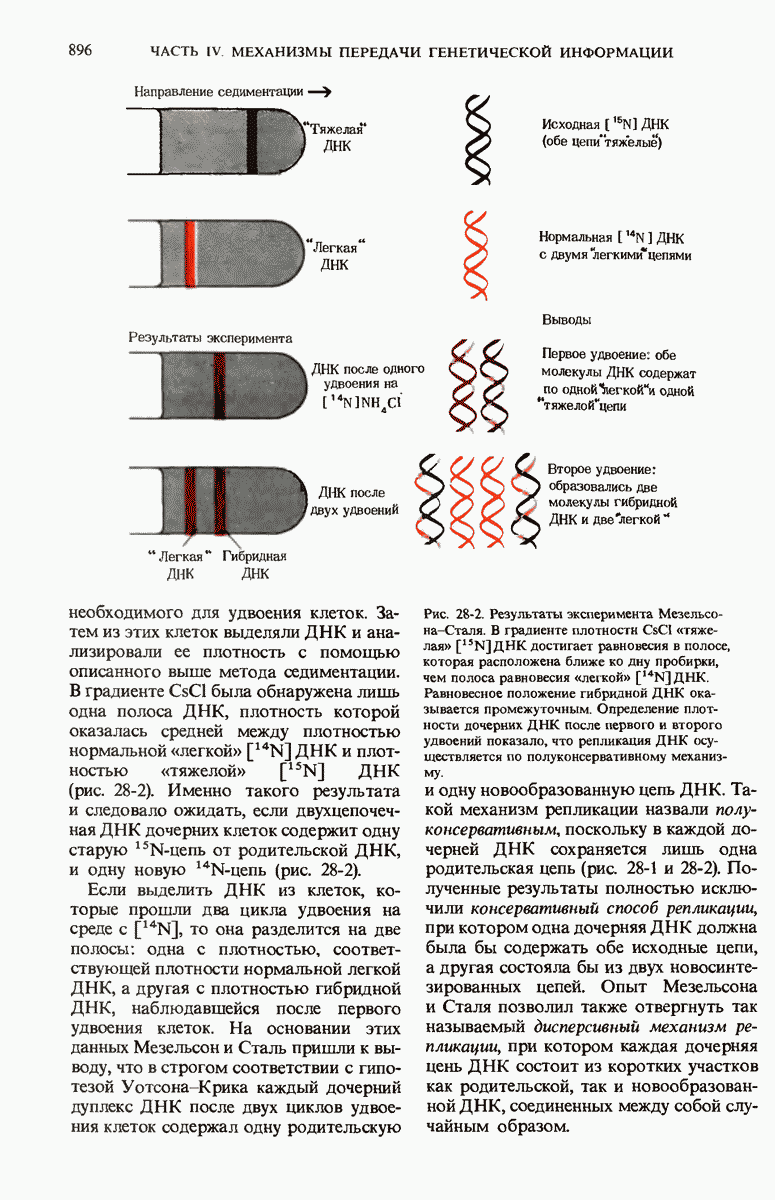
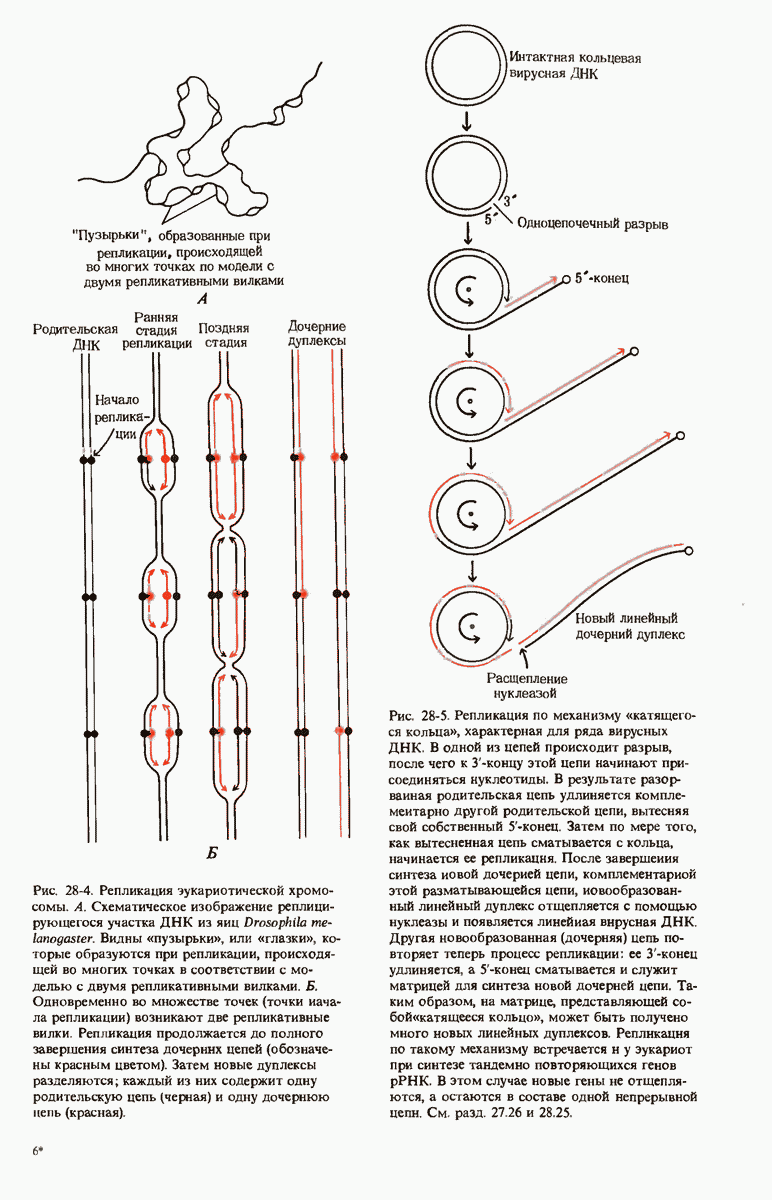
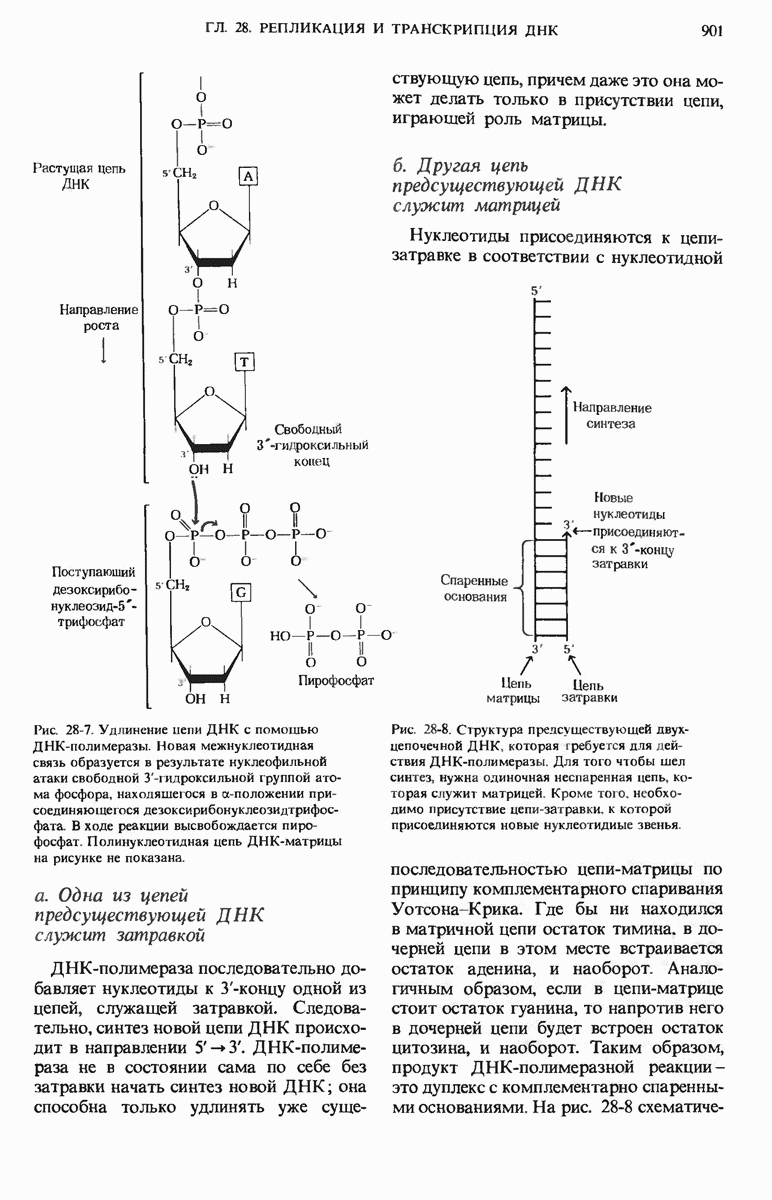
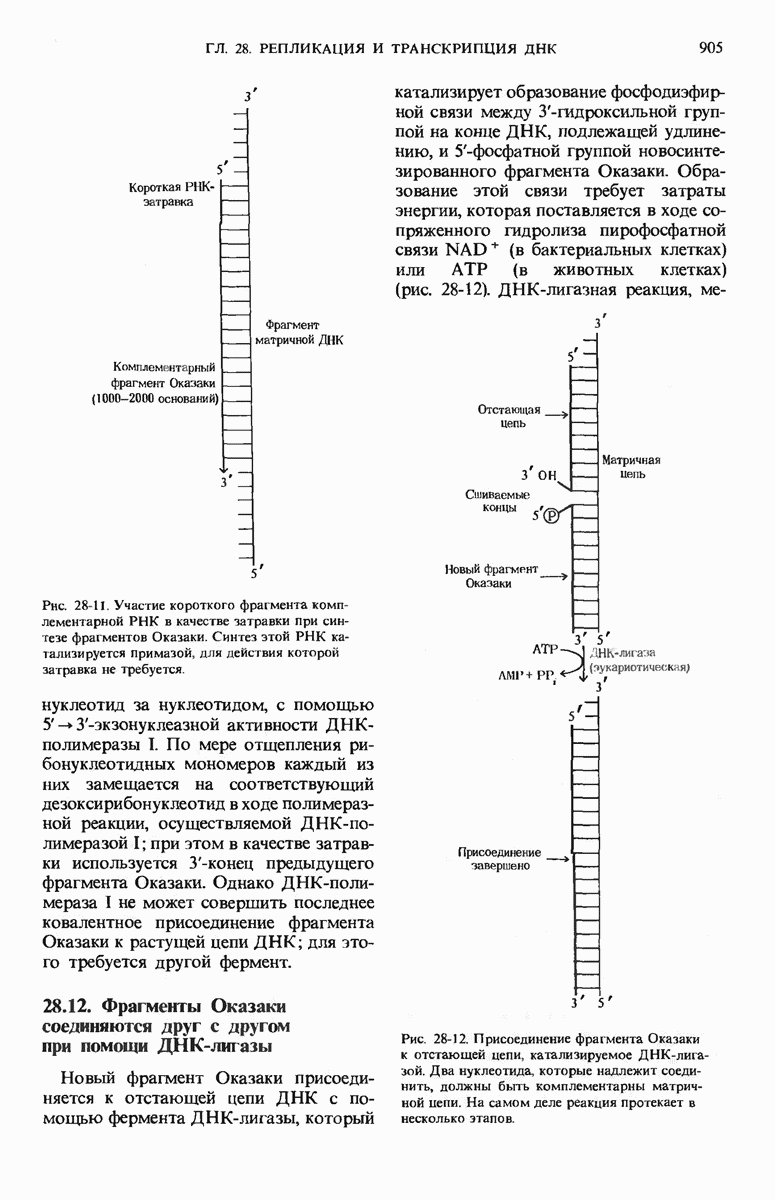
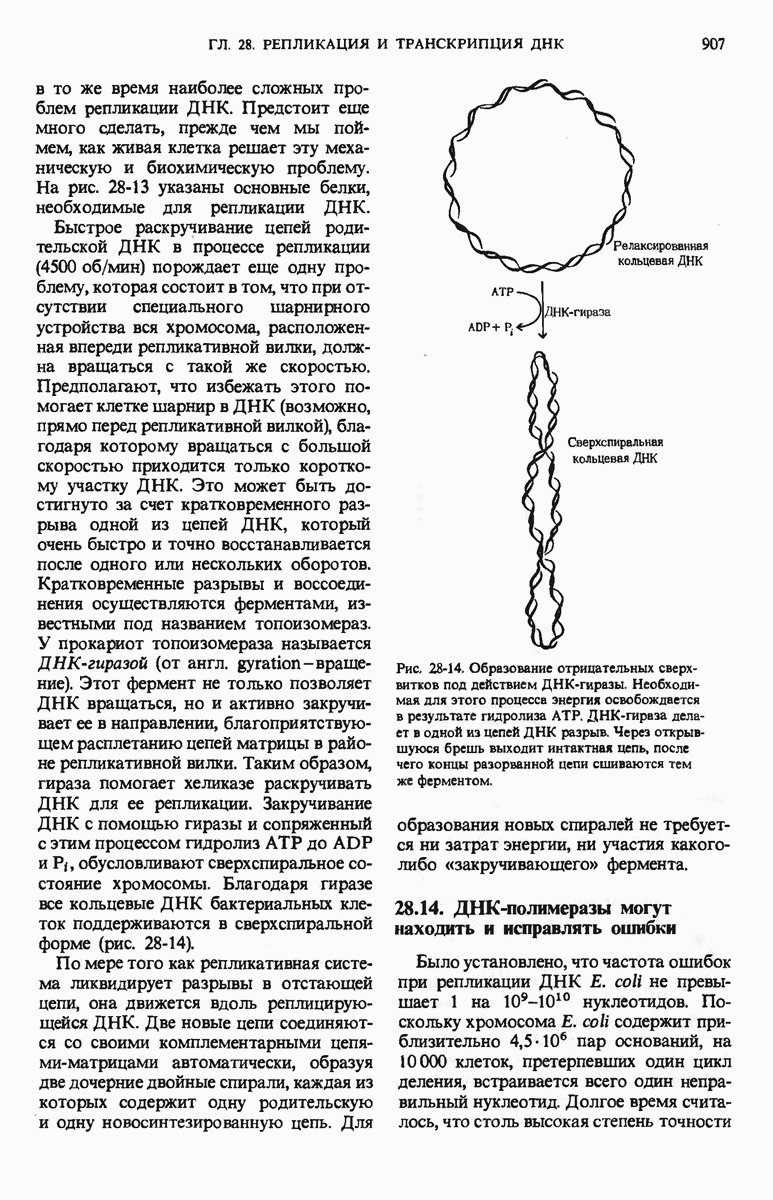
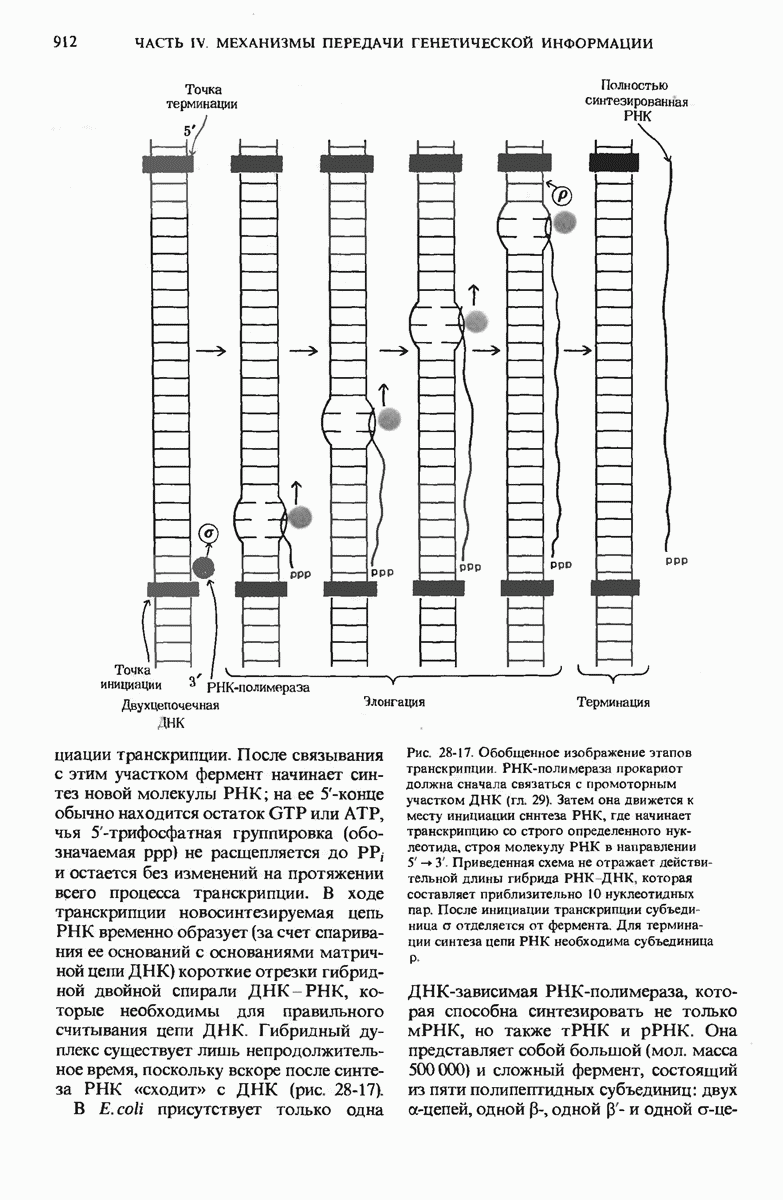
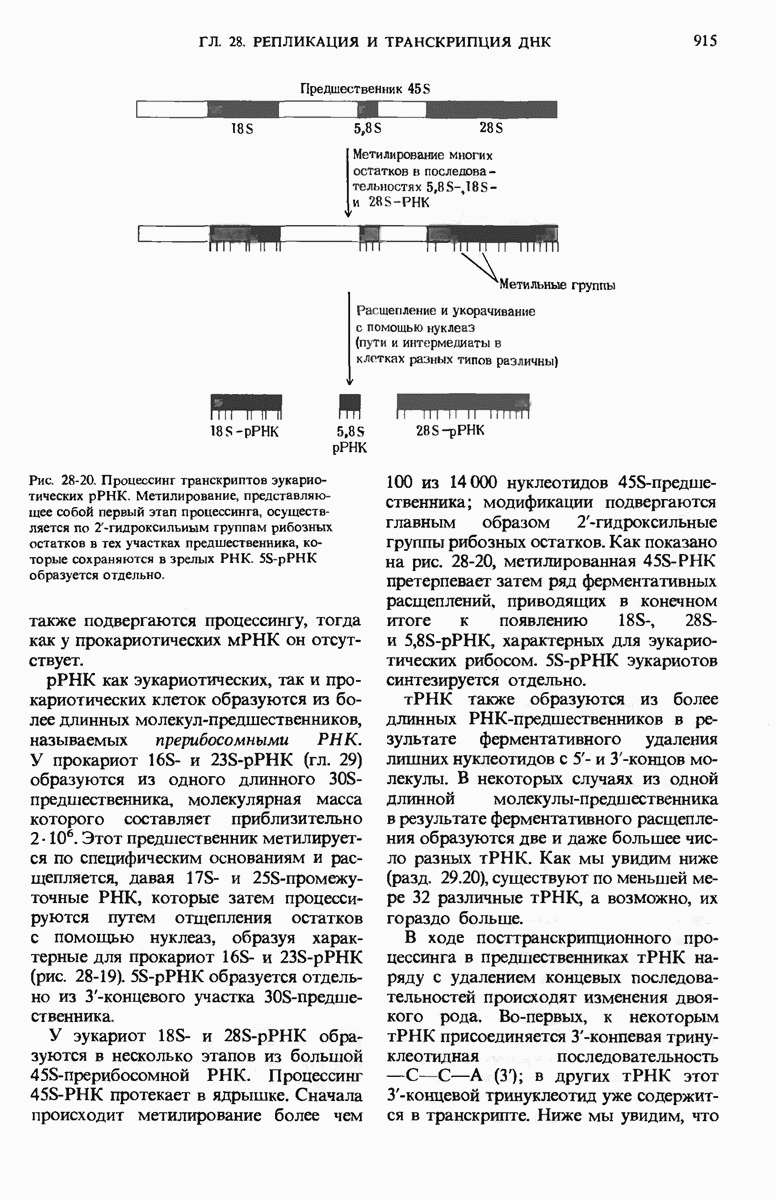
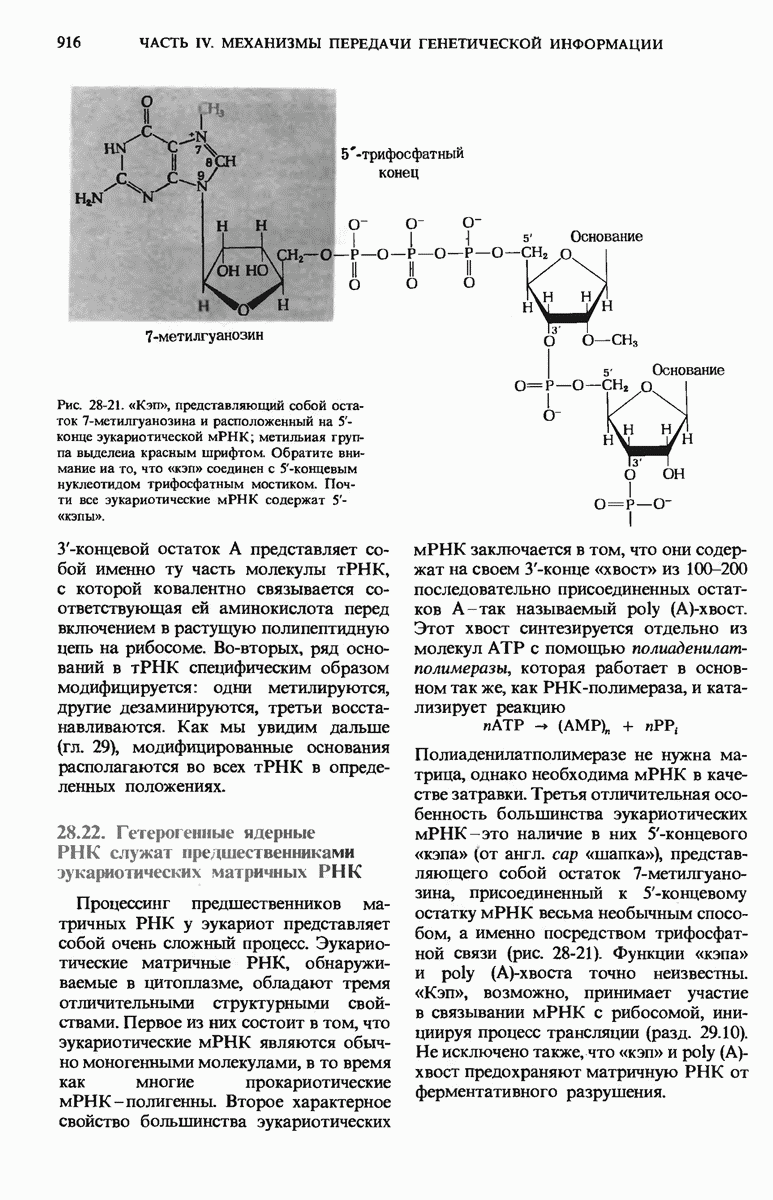
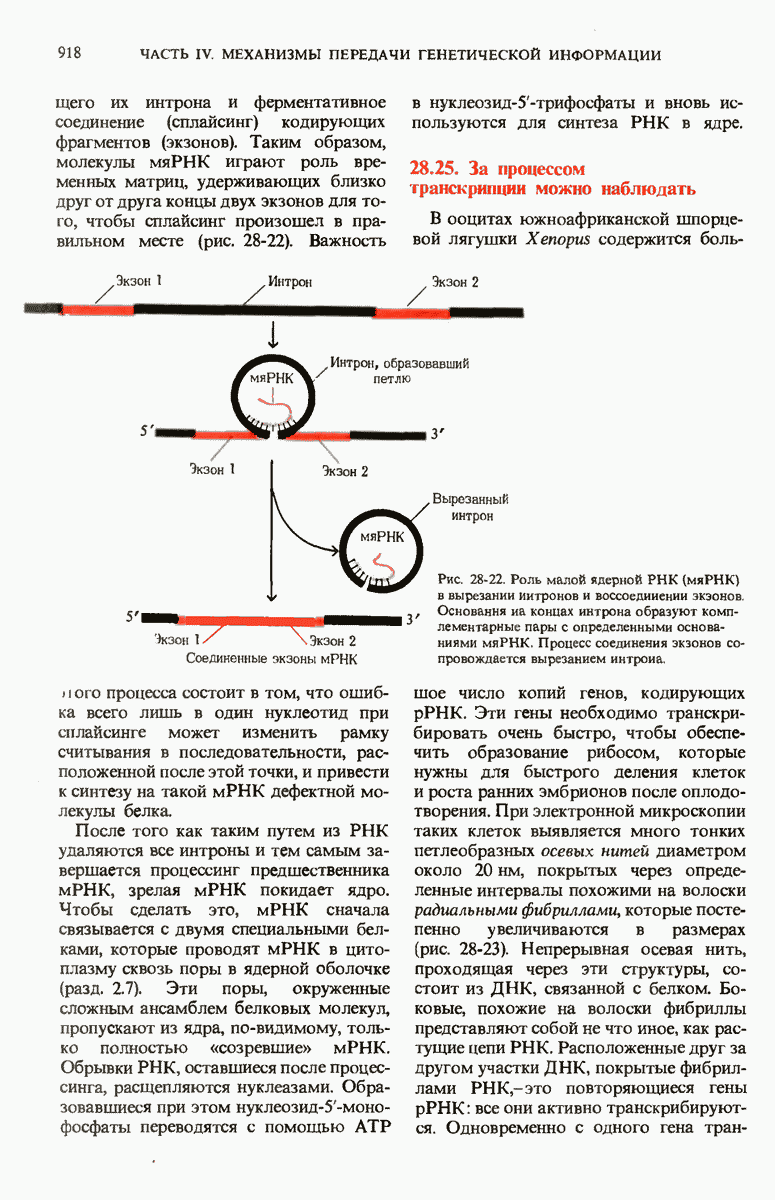
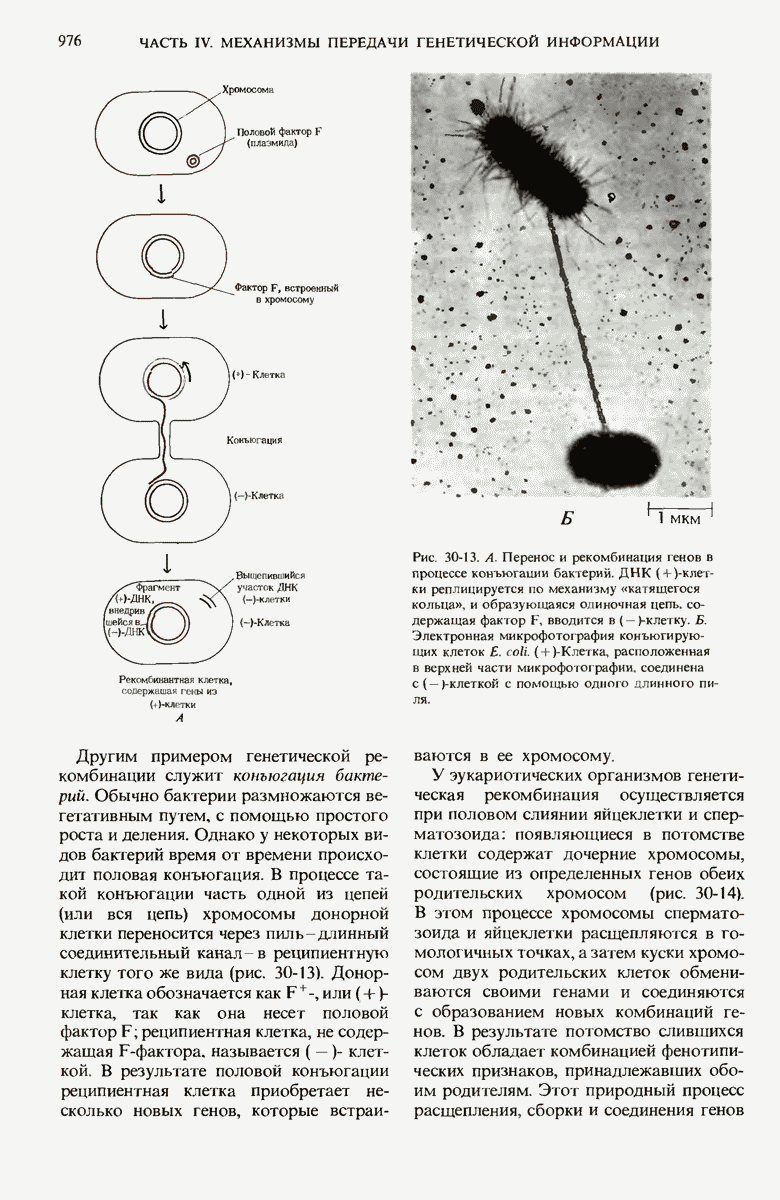
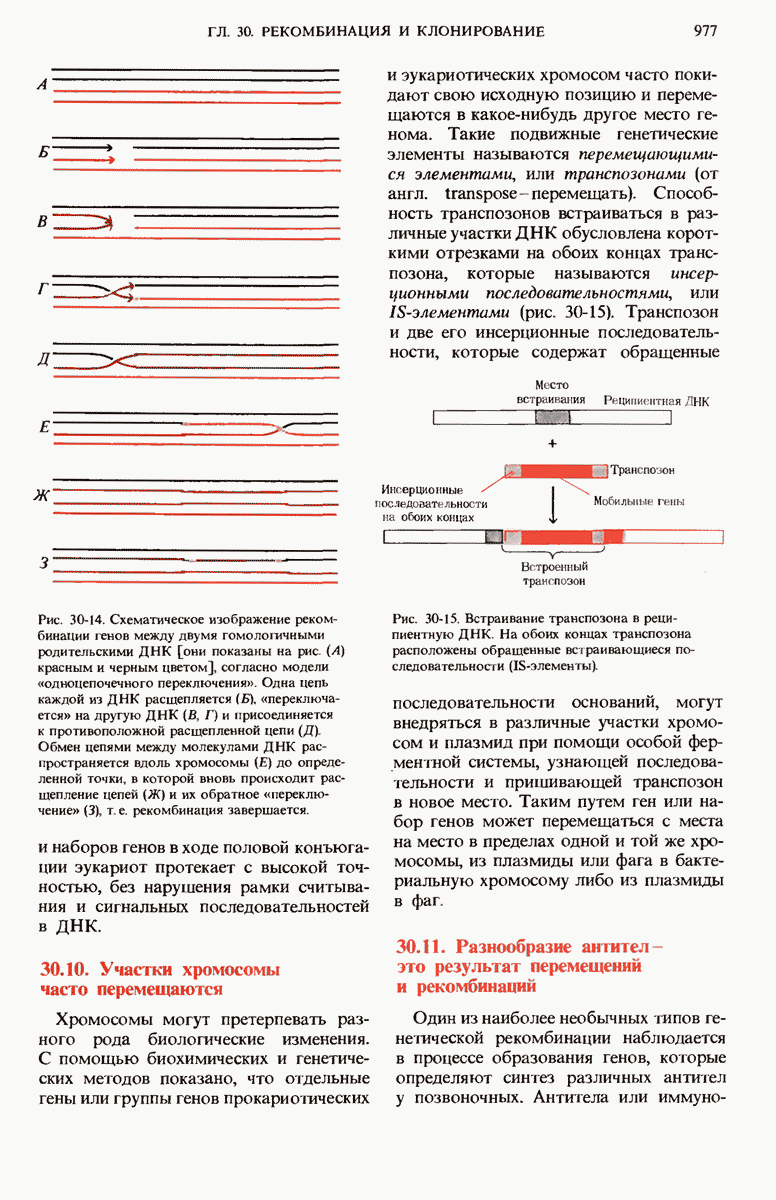
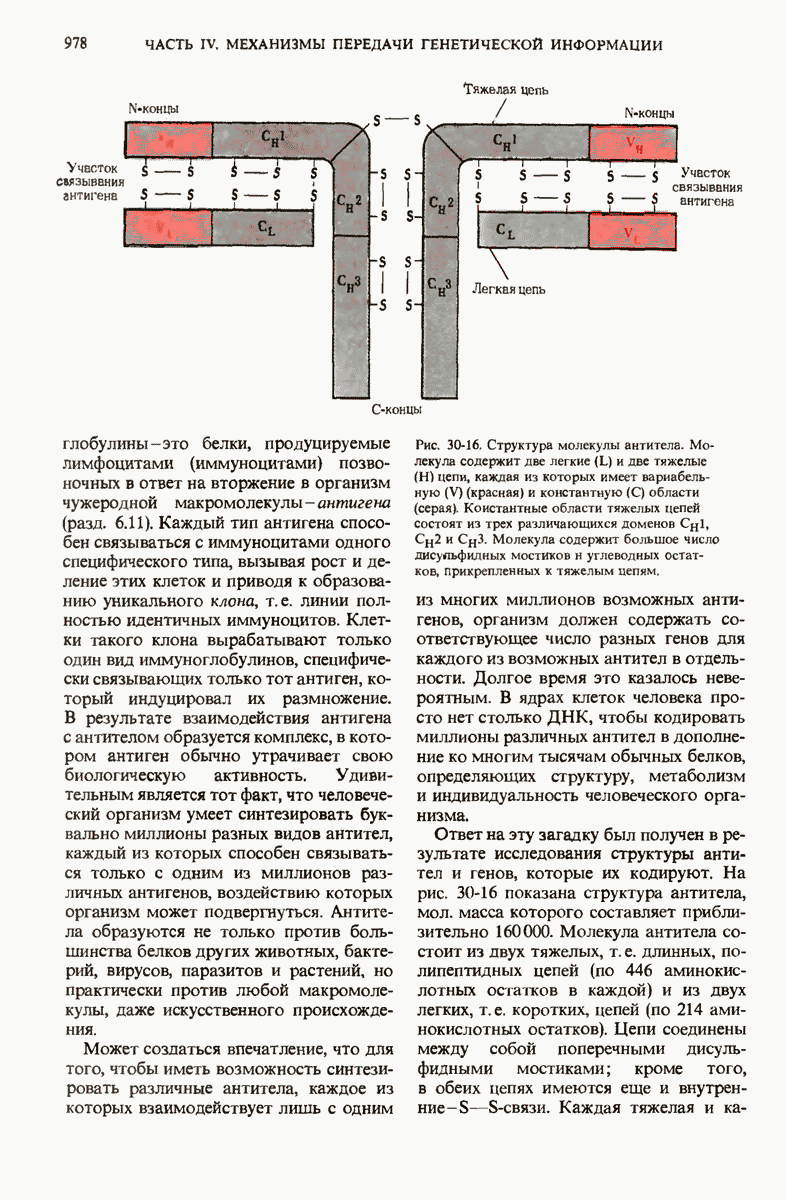
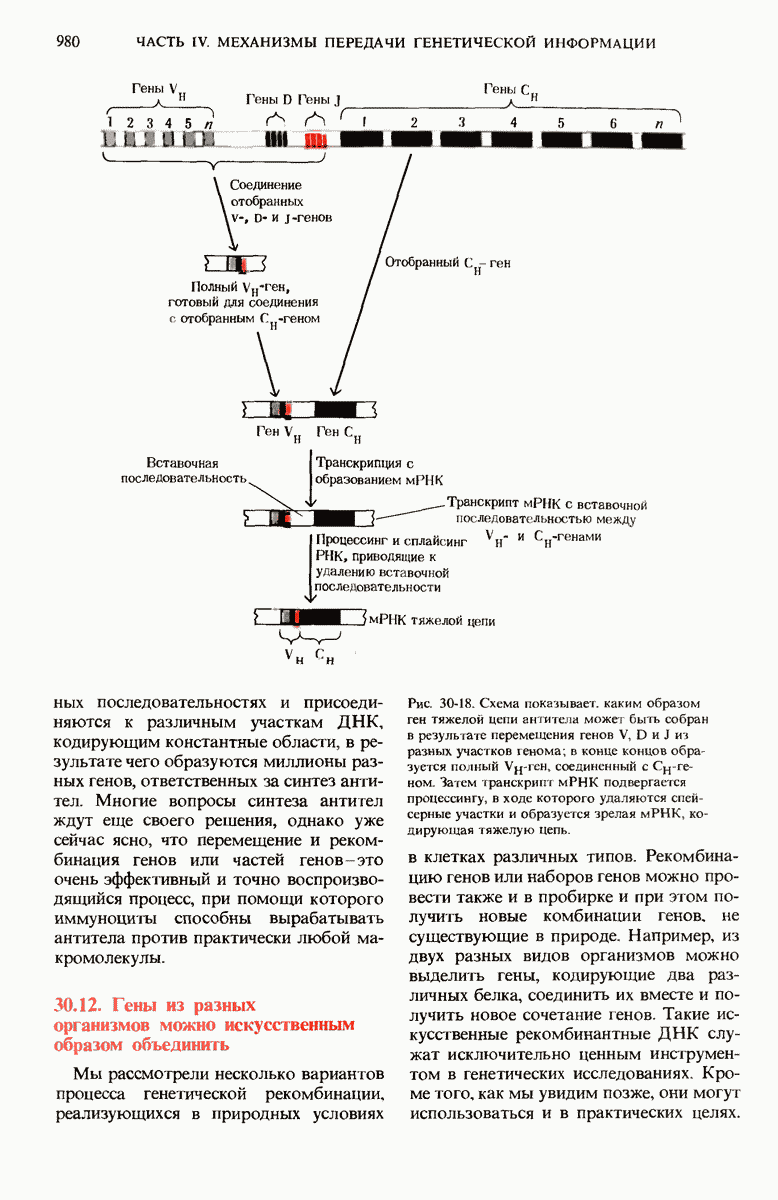
ГЛАВА 29. СИНТЕЗ БЕЛКА И ЕГО РЕГУЛЯЦИЯ
Механизм биосинтеза белков со всем многообразием их биологической активности и видовой специфичности был одной из крупнейших проблем в истории биохимии. В течение многих лет невозможно было ответить даже на очень простые вопросы относительно белкового синтеза. Например, образуются ли белки сразу как одно целое или же создаются путем сборки из множества коротких предварительно синтезированных пептидов? Или такой вопрос: может быть, все белки клетки образуются из одного длинного полипептида-предшественника в результате специфических изменений его боковых (R) групп? До начала 1950-х годов не было достоверно установлено даже то, что белки - это индивидуальные химические соединения с определенной молекулярной массой, определенным аминокислотным составом и определенной последовательностью аминокислотных остатков.
Сегодня мы уже многое знаем о процессе белкового синтеза, однако не исключено, что это лишь малая часть того, что нам еще предстоит узнать. По всей вероятности, синтез белка представляет собой самый сложный из биосинтетических процессов: он требует очень большого числа ферментов и других специфических макромолекул. В эукариотических клетках в белковом синтезе принимают участие свыше 70 различных рибосомных белков, не менее 20 ферментов, необходимых для активации аминокислот-предшественников, более десятка вспомогательных ферментов и других особых белковых факторов инициации, элонгации и терминации синтеза полипептидов, а также, возможно, не менее 100 дополнительных ферментов, участвующих в процессинге (созревании) белков. Если добавить к этому списку еще свыше 70 видов транспортных и рибосомных РНК, то станет ясно, что синтез полипептидов требует совместного действия почти трехсот различных макромолекул. К тому же многие из этих макромолекул организованы в сложные трехмерные структуры рибосом, в которых по мере синтеза полипептидной цепи происходит перемещение (транслокация) мРНК.
Несмотря на большую сложность аппарата биосинтеза белковых молекул, их синтез протекает с чрезвычайно высокой скоростью. К примеру, на образование полипептидной цепи, состоящей из ста аминокислотных остатков, рибосоме Е. coli достаточно пяти секунд. Более того, как мы увидим ниже, синтез тысяч различных белков в каждой клетке строго упорядочен, так что при данных условиях метаболизма синтезируется лишь необходимое число молекул каждого белка.
29.1. Открытия раннего периода заложили основу исследований биосинтеза белка
Основа наших сегодняшних представлений о биосинтезе белка была заложена в результате трех главных открытий 50-х годов. В начале 50-х годов Пол Замечник и его коллеги из Главного Массачусетского Госпиталя задались вопросом: в каком месте клетки синтезируются белки? В поисках ответа они вводили крысам радиоактивные аминокислоты.
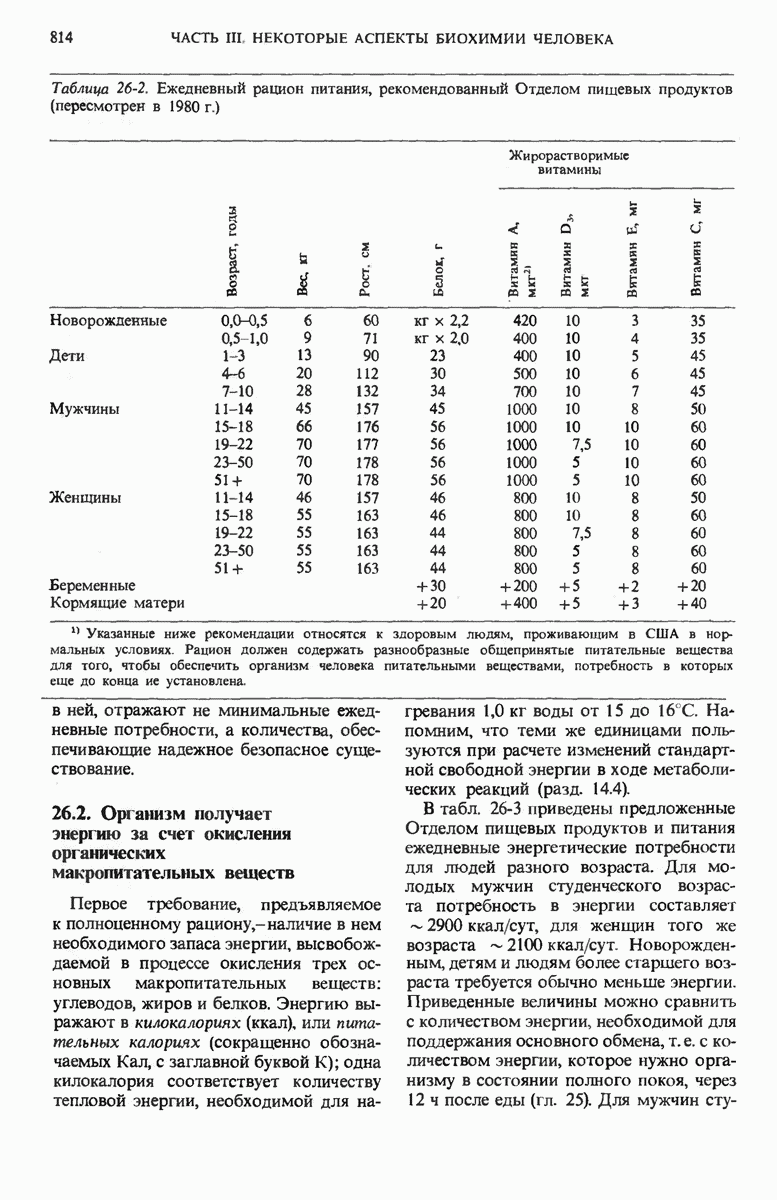
Через разные промежутки времени после инъекции они извлекали печень, гомогенизировали ее, фракционировали путем центрифугирования (разд. 13.16) и проверяли затем полученные субклеточные фракции на наличие в них радиоактивного белка. Если после введения меченых аминокислот проходили часы или дни, то меченые белки обнаруживали во всех внутриклеточных фракциях. Если же печень извлекали и фракционировали всего через несколько минут после инъекции меченых аминокислот, то новообразованный меченый белок обнаруживали лишь во фракции, содержащей небольшие рибонуклеопротеиновые частицы.

Рис. 29-1. Участок клетки поджелудочной железы, на котором видны рибосомы, прикрепленные к внешней поверхности эндоплазматического ретнкулума.
Исходя из этих данных, был сделан вывод о том, что местом синтеза белка из аминокислот являются рибонуклеопротеиновые частицы, которые ранее уже были обнаружены в животных тканях с помощью электронного микроскопа (разд. 2.17); позже они получили название рибосом (рис. 29-1).
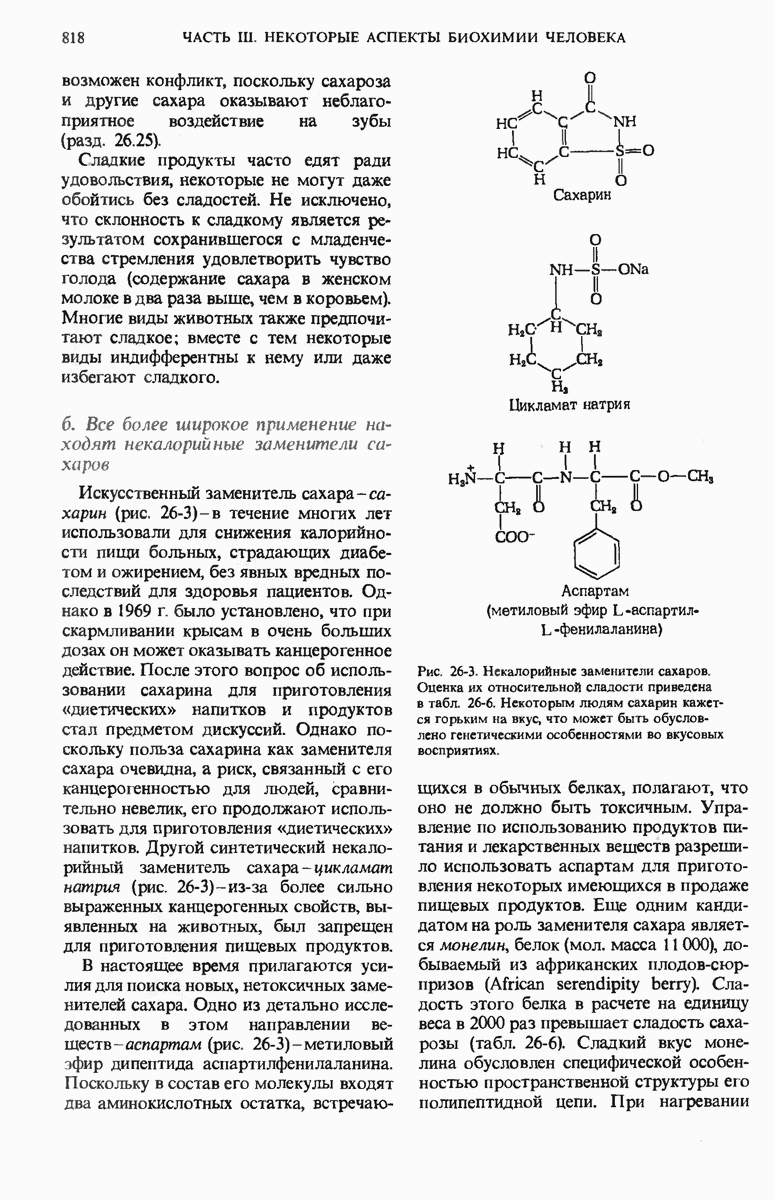
Второе открытие было сделано Мэлоном Хоглендом и Полом Замечником, которые обнаружили, что инкубация аминокислот с АТР и цитозольной фракцией клеток печени приводит к активации аминокислот. В ходе этого ферментативного процесса аминокислоты присоединялись к термостабильной растворимой РНК особого типа, названной позднее транспортной РНК (тРНК).
Третье из упомянутых выше главных открытий принадлежит Френсису Крику, который задумался над вопросом о том, каким образом генетическая информация, закодированная в нуклеиновых кислотах с помощью четырехбуквенного языка, переводится на двадцатибуквенный язык белков. Крик пришел к выводу, что тРНК должны выполнять в данном процессе роль адаптера. При этом одна часть молекулы тРНК может связываться со специфической аминокислотой, а какая-то другая ее часть - узнавать в мРНК короткую нуклеотидную последовательность, которая кодирует эту аминокислоту (рис. 29-2).
Именно эти три открытия привели вскоре к выяснению основных этапов биосинтеза белка и в конечном счете к установлению генетического кода для аминокислот.

Рис. 29-2. Гипотеза Крика об адапторной функции тРНК. Сегодня мы знаем, что аминокислота соединена с тРНК ковалентно, а специфический триплет нуклеотидов в другом участке молекулы тРНК «узнает» кодирующий триплет мРНК посредством образования водородных связей между комплементарными основаниями.