Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
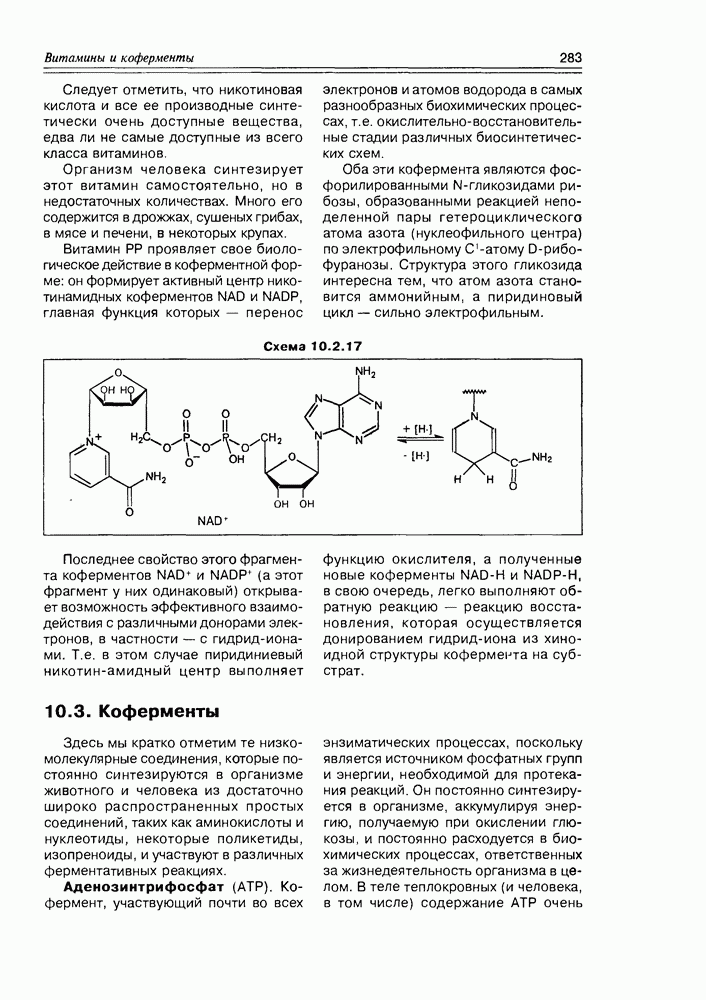
4.3. Биосинтез аминокислотСинтез молекул а-аминокислот требует обязательного включения следующих этапов: а) процесса формирования углеродного скелета; б) фиксации азота с доведением его до состояния аминогруппы; в) введения специфических функциональных групп. Нетрудно увидеть, анализируя материал предыдущего параграфа, что углеродные цепочки размера Азотофиксация столь же важный процесс в природе, что и фиксация углекислого газа, так как осуществляется переход неорганического инертного атмосферного азота в органические азотистые соединения — аминокислоты и белки, в первую очередь, а на последующих этапах — в нуклеиновые основания, алкалоиды и др. Растениями атмосферный азот не фиксируется и не усваивается. Эти функции выполняют прокариоты — бактерии рода Rhizobium, живущие симбиотрофно в корневых клетках растения-хозяина (бобовые растения, облепиха, ольха), свободно живущие бактерии типа Klebsiella pneumoniae и фотосинтезирующие сине-зеленые водоросли (цианобактерии). Фиксация азота сводится к поглощению его бактериями, где он связывается в виде комплекса ферментом нитрогеназой, осуществляющим постадийное восстановление азота до аммиака. Нитрогеназа представляет собой сложный фермент, в состав которого входят несколько белковых молекул, связанных с достаточно большим количеством атомов сульфидной серы, атомами железа в виде кластеров — это мощная восстановительная система — и двумя атомами молибдена, выполняющими функцию комплексообразователя по отношению к азоту. Схема 4.3.1
Второй путь усвоения азота осуществляется растениями, которые поглощают его в виде нитратов, т.е. в связанном виде — из почвы, где они образуются различными способами: либо это результат окисления аммиака, выделяющегося при разложении органических веществ; либо это соли азотной кислоты (селитры), содержащиеся в некоторых минералах; либо это окислы азота, образующиеся в результате атмосферных процессов. Восстановление нитратов до аммиака протекает в два этапа: сначала под действием нитрат-редуктазы образуются нитриты, которые на втором этапе нитрит- редуктазой восстанавливаются до аммиака (схема 4.3.1). Следующий этап усвоения азота — это фиксация аммиака — реакции, которые приводят к образованию аминокислот. В основе этих процессов лежат реакции нуклеофильного присоединения аммиака по карбонильной группе а-кетоглутаровой кислоты с последующей дегидратацией а-аминоспирта и восстановления имина до глутаминовой кислоты. Первые две стадии этой схемы (до образования имина) протекают легко и не нуждаются в стереоспецифичном катализе. Восстановление иминокислоты до аминокислоты строго стереоспецифично и контролируется соответствующим ферментом. Второй путь ассимиляции аммиака основан на реакции нуклеофильного замещения в у-карбоксильной группе глутаминовой кислоты, ведущей к образованию амида этой кислоты — аминокислоте глутамину. Настоящая реакция является основной по объему азота “перевозимого” в аминокислоты, хотя и представляет собой продолжение первого, вышеописанного пути усвоения азота по схеме “кетоглутаровая кислота плюс аммиак” (схема 4.3.2). Высокая производительность этой реакции ассимиляции аммиака обязана ее циклическому характеру, так как глутамин далее участвует в биосинтезе других аминокислот, выполняя роль донора аминогруппы, при этом сам снова преобразуется в глутаминовую кислоту. Схемы биосинтеза основных протеиногенных аминокислот достаточно многоступенчаты, но их можно свести к двум основным этапам — это формирование соответствующих окси- и оксокислот и аминирование последних до аминокислот В качестве исходных при синтезе аминокислот, как правило, выступают Кислоты с большим количеством атомов углерода образуются из вышеуказанных присоединением к ним активированных Схема 4.3.2
Схема 4.3.3
в условиях ферментативного катализа. Пути биосинтеза конкретных аминокислот различаются деталями схемы и природой исходной окси- или оксокислоты. По этому последнему фактору аминокислоты подразделяются на аминокислоты, происходящие из пировиноградной кислоты — лейцин, изолейцин, валин, лизин, аланин; аминокислоты, происходящие из щавелевоуксусной кислоты — аспарагиновая кислота, аспарагин, треонин, метионин; аминокислоты, происходящие из 2-оксоглу-таровой кислоты —аргинин, пролин, глутаминовая кислота, глутамин; аминокислоты, происходящие из продуктов цикла Кальвина — глицин, серин, цистеин. Забегая немного вперед, включим сюда шикимовую кислоту (ключевое соединение в биоситезе феноло-кислот), из которой образуются фенилаланин, тирозин, триптофан. Можно сказать, особняком стоит и биосинтез гистидина, который выпадает из общего комплекса реакций, ведущих к аминокислотам. Установлено, что предшественниками гистидина являются
|
 |
Оглавление
|













































































































































































































































































































































































 с карбоксильной группой образуются в ходе фотосинтеза (пировиноградная и щавелевоукесусная кислоты). Кроме того, из пировиноградной кислоты реакцией окислительного декарбоксилирования получается уксусная кислота, столь необходимая для биосинтеза а-аминокислот и широко используемая Природой в других биосинтетических схемах.
с карбоксильной группой образуются в ходе фотосинтеза (пировиноградная и щавелевоукесусная кислоты). Кроме того, из пировиноградной кислоты реакцией окислительного декарбоксилирования получается уксусная кислота, столь необходимая для биосинтеза а-аминокислот и широко используемая Природой в других биосинтетических схемах.

 -окси- и оксокислоты, образующиеся в фотосинтетическом цикле Кальвина (схема 4.3.3).
-окси- и оксокислоты, образующиеся в фотосинтетическом цикле Кальвина (схема 4.3.3).
 -фрагментов, образующихся из пировиноградной кислоты
-фрагментов, образующихся из пировиноградной кислоты


 и 5-фосфорибозил-1-пирофосфат, а сложная схема этого биосинтеза включает стадии деградации пуринового гетероцикла, формирования нового имидазольного фрагмента, а потом уже введения аминокислотного остатка.
и 5-фосфорибозил-1-пирофосфат, а сложная схема этого биосинтеза включает стадии деградации пуринового гетероцикла, формирования нового имидазольного фрагмента, а потом уже введения аминокислотного остатка.