Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
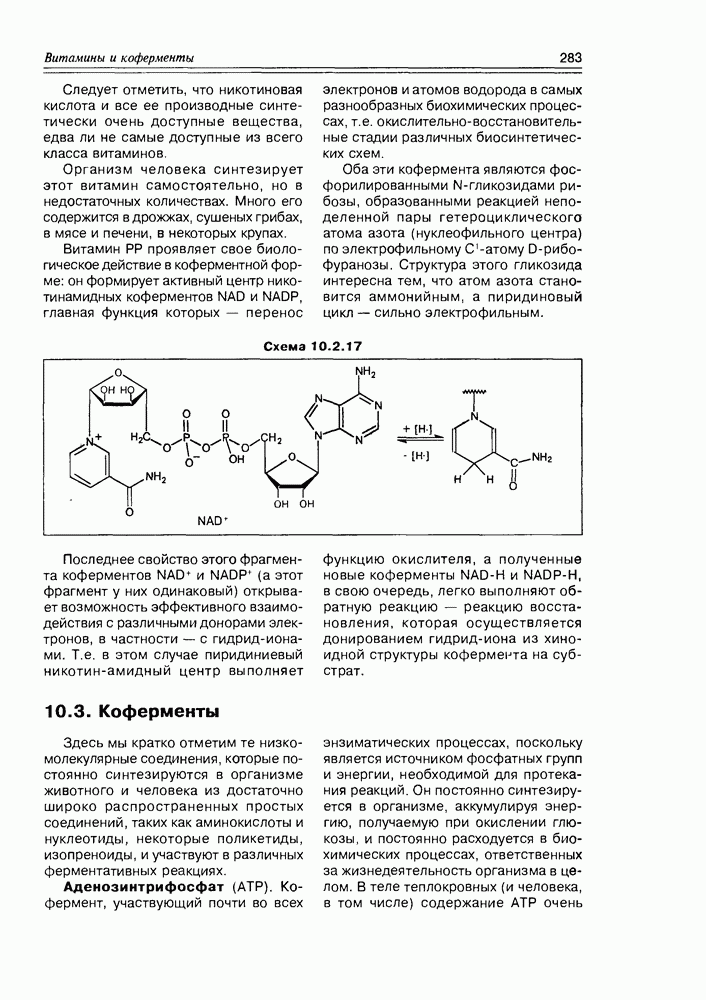
4.8. БелкиБелки или протеины — наиболее важные производные а-аминокислот, а с позиции их биологических свойств и особого положения в живой природе, белки представляют собой отдельный класс природных соединений, причем очень важный класс. Но учитывая их биологическую значимость и высокую молекулярную массу (от История исследований белков, по сравнению с другими классами природных соединений, наиболее богата событиями и открытиями, поскольку эти вещества вездесущи в живой природе, очень многообразны и наиболее сложны по структуре. Кроме того, их сложность и большие молекулярные размеры сочетаются с низкой устойчивостью и трудностью индивидуального выделения. Но к настоящему времени многие барьеры на этом пути преодолены. Достаточно быстро и надежно хроматографически определяется аминокислотный состав белков и последовательность их соединения между собой; рентгеноструктурный анализ позволяет установить пространственную структуру тех белковых молекул, которые удается получить в виде кристаллов; различными вариантами метода ЯМР успешно исследуется поведение белков в растворах, в процессах комплексообразования, т.е. в ситуации, близкой к той, которая имеет место в живой клетке. В настоящее время принято различать четыре структурных уровня в архитектуре белковых молекул: первичная, вторичная, третичная и четвертичная структуры белков. Первичная структура белков. Под первичной структурой белковой молекулы подразумевается знание аминокислотного состава ее и последовательность распределения аминокислотных остатков в полимерной белковой цепочке. Определение первичной структуры до недавнего времени было очень сложной задачей, стратегия решения которой состоит из следующих нескольких этапов: после гидролиза белка на мономерные аминокислоты, необходимо определить мольное соотношение их; определить молекулярную массу белка одним из физических методов; расщепить белок на фрагменты, в которых можно определить последовательность аминокислот каким-либо методом, например с помощью ступенчатой деградации по Эдману и ряд других химических и физико-химических операций для выявления различных структурных деталей белка (наличие простетических групп, дисульфидных мостиков и т.д.). Первый белок, структура которого была полностью расшифрована — это инсулин. На эту работу Сэндшэр с сотрудниками затратил несколько лет (Нобелевская премия за 1958 г.). С помощью современной техники ту же работу можно осуществить за несколько недель. Из всех химических реакций, используемых для определения различных структурных моментов белковой проблемы, следует отметить те, которые прошли испытание временем и используются в современной методологии. Метод Эдмана последовательной деградации пептидов и белков основан на реакции фенилизотиоцианата с концевой аминогруппой в щелочной среде, которая приводит сначала к образованию тиомочевинной метки концевой аминокислоты, последняя при действии Схема 4.8.1
со следующей концевой аминокислотой: образующиеся производные фенилтиогидантоина идентифицируются хроматографически, поскольку все они различаются по структуре радикалов R соответствующих аминокислот (схема 4.8.1). Так как метод Эдмана позволяет проанализировать полипептидные цепочки, состоящие примерно из 50 остатков, что явно недостаточно для анализа всей белковой молекулы, последнюю предварительно расщепляют селективными реакциями на фрагменты. К таковым относится реакция белка с бромцианом, который расщепляет пептидную связь по остаткам метионина (схема 4.8.2). Схема 4.8.2
Пептидные связи, образованные триптофаном, легко разрываются N-бромсукцинимидом (схема 4.8.3). Наряду с химическими способами расщепления, плодотворно используют и ферментативные методы. Пептиды, выполняющие эту роль, подразделяют на те, которые отщепляют концевые или близкие к ним аминокислотные остатки (экзопептидазы), и те, которые Схема 4.8.3
Схема 4.8.4 (см. скан) расщепляют пептидные связи, удаленные от концов белковой молекулы (эндопептидазы). Для фрагментации последние, конечно же, более интересны, а наиболее перспективны среди них те, которые обладают специфичностью. Так, трипсин гидролизует пептидные связи, образованные аргинином или лизином с любой другой аминокислотой, кроме пролина. Специфичность реакций трипсина можно повысить, используя химическую модификацию аминокислотных остатков (как бы защищая одну из пептидных связей от ферментативного воздействия). Если в обычном варианте этот фермент расщепляет связи с участием аргинина и лизина, то после обработки белка малеиновым ангидридом, который взаимодействует с Так как белковые молекулы часто связаны между собой дисульфидными мостиками (или же такие мостики присутствуют внутримолекулярно), то важным химическим приемом, предворяющим все остальные, является расщепление этих связей. Для достижения цели используются различные реагенты, среди которых одним из наиболее популярных можно считать 2-меркаптоэтанол (схема 4.8.5). Схема 4.8.5
Безусловно, это далеко не полный перечень приемов, используемых для установления первичной структуры белковой молекулы, но, в общем, стратегия решения этой задачи такова; и после удачного ее применения мы можем указать строение полипептида или белка так, как это уже сделано выше — на схемах 4.4.1-4.4.4. Вторичная структура белков. Это первый этап пространственной организации полипептидных цепочек, контролируемый водородными связями пептидных групп, как внутримолекулярными, так и межмолекулярными. Основными видами вторичной структуры являются: а-спираль, характерная как для всей молекулы белка (кератин волос, миозин и тропомиозин мышц), так и только для отдельных участков белкового полимера (инсулин). Она стабилизирована внутримолекулярными водородными связями а-Спираль была теоретически предсказана Полингом и Кори за несколько лет до ее экспериментального обнаружения. Ими же открыта и Третичная структура белков. Это второй этап пространственной организации полипептидных биополимеров, контролируемый взаимодействиями боковых функциональных групп. Вид этих взаимодействий может быть различным: это разнообразные водородные связи, электростатическое притяжение ионов, ковалентные связи. Важнейшее место в формировании третичной Схема 4.8.6
структуры белка играют дисульфидные связи, образованные взаимодействием двух цистеиновых остатков. Эта связь выгодно отличается от слабых водородных своей прочностью в обычных условиях и в тоже время выигрышна, по сравнению с другими ковалентными связями, своей лабильностью, т.е. она может разрушаться и снова восстанавливаться в достаточно мягких условиях (схема 4.8.7). Форма молекулы белка с ярко выраженной третичной структурой может быть определена как форма “ клубка”, т.е. структуры неопределенной, но жестко фиксированной геометрической формы в отличие от клубка ниток, структура которого носит случайный характер и не воспроизводится при повторной процедуре его образования. Что это значит? Это значит, что белковый клубок, каждая петля которого Схема 4.8.7
фиксирована дисульфидным мостиком, после разрушающего действия какого-либо восстановителя существенно меняет свою структуру и от клубка переходит к неопределенной форме, соответствующей денатурированному белку. Но при проведении обратной реакции, реакции окисления, белок снова приобретает форму клубка, причем такого клубка, каким он был вначале. Если исходный белок имел при этом какую-либо биологическую активность (ферментативную, например), то он ее полностью теряет в процессе разрушения дисульфидных мостиков и полностью восстанавливает эту активность Схема 4.8.8
после образования дисульфидных мостиков, т.е. после воссоздания структуры клубка (схема 4.8.8). Из вышерассмотренного анализа третичной структуры белковой молекулы можно вывести следующее определение третичной структуры: это структура белка, обусловленная взаимодействием цистеиновых аминокислотных остатков, либо это клубок, фиксированный дисульфидными мостиками. Хотя в общем случае не исключено, что отдельные элементы (т.е. петли) клубка могут быть образованы взаимодействием и других аминокислот: водородными связями с участием ОН-групп серина и треонина, ионными связями аммонийно-карбоксилатного типа Четвертичная структура белка. Это, наверное, самая неопределенная структура. Единственное, что можно сказать о ней — то, что это комплекс из нескольких полипептидных цепочек, связанных между собой самыми различными связями: как слабыми водородными и ионными, так и прочными ковалентными, включая дисульфидные, сложноэфирные и амидные. Типичным случаем четвертичной структурной организации белка является молекула гемоглобина, состоящая из четырех полипептидных цепочек, связанных между собой водородными, гидрофобными и ионными связями. Особую роль выполняют ионные связи между аспарагиновой кислотой с одной стороны, лизином и аргинином с другой стороны — они образуются только в дезоксигемоглобине и разрываются при оксигенировании атома железа. В свою очередь, гемы связаны с белковыми Схема 4.8.9
Схема 4.8.10
цепями координационной связью атома железа и имидазольного фрагмента аминокислотного остатка гистидина (схема 4.8.9). Другим характерным примером белков с четвертичной структурной организацией являются иммуноглобулины, четыре полипептидных фрагмента которых связаны между собой дисульфидными мостиками (схема 4.8.10). В структуре этого класса белков обращает на себя внимание большое количество симметрично расположенных внутримолекулярных дисульфидных связей, которые в определенных условиях могут переходить в межмолекулярные дисульфидные мостики, тем самым меняя пространственную структуру белка в целом, и причем меняя ее существенно. В заключении о структурной организации белков следует отметить, что третичная и четвертичная структуры характерны для белков высокой биологической активности, ферментов в частности. Эти структуры позволяют молекулам формировать активные центры высокой эффективности и селективности. Определение третичной и четвертичной структур белковых молекул осуществляется физическими методами, в первую очередь, рентгеноструктурным анализом. Спектроскопические методы Химические свойства белков. Практически не имеет смысла говорить о химических свойствах каждого конкретного белка, поскольку любая белковая молекула имеет почти врсь арсенал функциональных групп, пожалуй, за исключением олефиновых и карбонильных, и в связи с этим, все белки должны обладать фиксированным набором химических реакций. В небольшом количестве случаев молекулы белков имеют какой-либо преобладающий состав. Так, в полипептидах коллагена треть аминокислотного состава приходится на глицин; в полипептидной цепи нейротоксина-1) (яд кобры) около 20% аминокислотного состава приходится на долю основных аминокислот (His, Lys, Arg), тогда как количество кислых аминокислот (Asp, Glu) - менее 10%. Поэтому имеет смысл говорить о химических свойствах не молекулы белка в целом, а отдельных ее участков. Во-первых, мы можем выделять гидрофильные и гидрофобные (липо-фильные) участки молекул, в молекулах белков могут быть участки цепи с доминирующими кислотными свойствами (локализация Asp и Glu) или доминирующими основными свойствами (локализация Arg, Lys и His) и т.д. Кроме того, определенные свойства участков цепи могут быть обусловлены взаимодействием соседних аминокислотных фрагментов; соседство аспаргиновой Схема 4.8.11
кислоты и лизина приводит к солеобразованию, а следовательно, к повышенной гидрофильности фрагмента цепи; соседство двух цистеиновых остатков — к образованию локального дисульфидного фрагмента, обладающего мягкими окислительными свойствами (схема 4.8.11). Следует отметить, что взаимодействие двух соседних групп в определенной степени связано с геометрией соответствующей пептидной связи, которая может иметь как транс-, так и цис-конфигурацию — достаточно жесткие структуры. Их жесткость обеспечена частичной двоесвязанностью связи -HN-CO- за счет Схема 4.8.12
Из последнего положения вытекает укореняющийся в последнее время, достаточно общий взгляд на главенствующее значение первичной структуры белка: последовательность и природа аминокислот в полипептидной цепи определяет ее пространственную структуру. Интересный пример специфического участия боковых функциональных групп аминокислот в реализации биологических функций белковых молекул в живом организме предоставляют нам медузы, Aequorea victoria, обладающие биолюминисцентной системой из двух ассоциированных белковых молекул, одна их которых обладает хромофорными свойствами (зеленая флюоресценция с Из схемы видно, что если этот процесс является чисто химическим (осуществляется под действием тепловой энергии на первой стадии и кислорода воздуха на второй), то скорее всего, он необратим. В случае его энзим-контролируемости, явление биофлюоресценции должно быть обратимым.
|
 |
Оглавление
|













































































































































































































































































































































































 до
до  kD), можно считать белковые соединения все-таки прерогативой биохимических и биофизических исследований. Поэтому здесь мы коснемся только их химической части: рассмотрим белки как производные а-аминокислот, т.е. как полипептиды высокой степени поликонденсации.
kD), можно считать белковые соединения все-таки прерогативой биохимических и биофизических исследований. Поэтому здесь мы коснемся только их химической части: рассмотрим белки как производные а-аминокислот, т.е. как полипептиды высокой степени поликонденсации.
 отщепляется, в конечном счете, в виде замещенного фенилтиогидантоина, после чего полипептидная цепь белка укорачивается на один аминокислотный остаток. Далее этот процесс повторяется вновь
отщепляется, в конечном счете, в виде замещенного фенилтиогидантоина, после чего полипептидная цепь белка укорачивается на один аминокислотный остаток. Далее этот процесс повторяется вновь



 -аминогруппой лизина, фермент расщепляет пептидные фрагменты только аргинина (схема 4.8.4).
-аминогруппой лизина, фермент расщепляет пептидные фрагменты только аргинина (схема 4.8.4).


 -складчатая структура белка, которая стабилизируется межмолекулярными водородными связями того же типа и характерна для таких белков, как фиброин шелка и спираль коллагена, которая представляет собой жгут из трех цепочек полипептида, стабилизированный межмолекулярными водородными связями (схема 4.8.6).
-складчатая структура белка, которая стабилизируется межмолекулярными водородными связями того же типа и характерна для таких белков, как фиброин шелка и спираль коллагена, которая представляет собой жгут из трех цепочек полипептида, стабилизированный межмолекулярными водородными связями (схема 4.8.6).



 остатков лизина (аргинина) и аспарагиновой (глутаминовой) кислоты. Но такие петли будут нестойкими и легко разрушаться при действии растворителя, изменении pH среды и т.д.
остатков лизина (аргинина) и аспарагиновой (глутаминовой) кислоты. Но такие петли будут нестойкими и легко разрушаться при действии растворителя, изменении pH среды и т.д.


 также приносят определенную информацию о пространственном строении белков, но эта информация носит дополнительный характер.
также приносят определенную информацию о пространственном строении белков, но эта информация носит дополнительный характер.

 взаимодействия Как правило, в полипептидных цепочках реализуется трансформа, которая не препятствует сближению функций боковых радикалов таких аминокислот, как Asp и Lys, Glu и Arg. Но тиольные группы цистеиновых остатков могут образовать дисульфидную связь только при цисформе пептидного звена, также стерически затрудненным оказывается в трансформе и солеобразование остатков аспарагиновой кислоты и гистидина. Поэтому такие трансстерически затрудненные реакции боковых функциональных групп в случае реализации взаимодействия будут менять геометрию пептидного звена с цис- на транс- и, таким образом, существенно влиять на вторичную структуру белковой молекулы.
взаимодействия Как правило, в полипептидных цепочках реализуется трансформа, которая не препятствует сближению функций боковых радикалов таких аминокислот, как Asp и Lys, Glu и Arg. Но тиольные группы цистеиновых остатков могут образовать дисульфидную связь только при цисформе пептидного звена, также стерически затрудненным оказывается в трансформе и солеобразование остатков аспарагиновой кислоты и гистидина. Поэтому такие трансстерически затрудненные реакции боковых функциональных групп в случае реализации взаимодействия будут менять геометрию пептидного звена с цис- на транс- и, таким образом, существенно влиять на вторичную структуру белковой молекулы.

 нм
нм  Хромофорная группа этого протеина создается в результате двух последовательных реакций (дегидратации и окисления с участием двух пептидных фрагментов цепи и фенольной функции тирозина (схема 4.8.12).
Хромофорная группа этого протеина создается в результате двух последовательных реакций (дегидратации и окисления с участием двух пептидных фрагментов цепи и фенольной функции тирозина (схема 4.8.12).