Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
5.4. Биосинтез жирных кислот и их производныхСтартовой реакцией биосинтеза жирных кислот считается (и это вполне надежно доказано) образование так называемого “активного” ацетата из пировиноградной кислоты и кофермента А. Суммарное уравнение реакции включает нуклеофильную атаку тиольной группой углерода карбонильной функции и декарбоксилирование пируватного фрагмента с сопутствующими окислительно-восстановительными процессами (схема 5.4.1). Тиольная группа является активным центром кофермента Учитывая тот факт, что все ферменты этого процесса связаны в единую тесную систему, можно представить и сами реакции в виде единой системы, которая отражает как химическую природу основных этапов, так и определенную степень их согласованности. Молекула Схема 5.4.1
Электрофильным центром является углеродный атом карбонильной группы, при атаке которого нуклеофилом (карбанионом, например) реализуется замещение коферментной функции Но, как оказалось, Природа очень хорошо "изучила" классическую органическую химию, из которой мы знаем, что карбанионный центр наиболее легко генерируется в производных малоновой кислоты — даже слабые основания отщепляют протон от метиленовой группы малонового эфира. Поэтому второй путь образования ацетоацетил-SCoA выглядит так: ацетил-ScoA переводится в малонил-SCoA; активированный основанием Процесс нуклеофильного замещения Полное восстановление всех кетонных групп поликетидов, которое обычно происходит в процессе сборки цепи, и гидролиз концевой ацетатной группы приводит к насыщенным жирным кислотам. Кстати, из этой схемы вполне очевидным становится правило “четного количества углеродов” в основных нормальных жирных кислотах. Схема 5.4.2 (см. скан) Схема 5.4.3 (см. скан) Пальмитиновая кислота является первым самостоятельным продуктом биосинтеза жирных кислот. В последующем она может удлиняться до насыщенных кислот ион железа Схема 5.4.4
Следующий важный, распространенный и разнообразный метаболический процесс в ряду жирных кислот — это их оксигенирование. Оксигенированию, как правило, подвергаются ненасыщенные (особенно метилен-разделенные полиеновые) жирные кислоты, образуя гидроксипроизводные (например, лейкотриены), эпоксикислоты и фурановые кислоты, а также карбоциклические оксилипины (например, простогландины). Реакции оксигенирования катализируются большой группой ферментов — липоксигеназами. Очевидно, что это близкие по структуре ферменты, так как они все активируют молекулу кислорода и различаются лишь своей селективностью по структуре жирной кислоты и месту кислородной атаки на ее углеродной цепи. Окисление жирных кислот, имеющих метилен разделенные олефиновые связи, может протекать по двум механизмам: либо это первоначальное отщепление радикала водорода от метиленового звена с образованием сопряженного пентадиенил радикала, оксигенирование которого проходит по одному из концевых углеродов этой сопряженной системы, либо это синхронное присоединение Экспериментально показано, что образование лактоновых метаболитов жирных кислот, нередко встречающихся в различных организмах, также контролируется ферментом с участием Схема 5.4.5
Схема 5.4.6
замещающего S-CoA фрагмент. Эта циклизация может быть осуществлена на различных стадиях формирования и функционализации углеродного скелета молекулы. Вышеописанное образование лактона фактически может быть определено как внутримолекулярная реакция перезтерификации с участием тиозфиражирной кислоты. Такая реакция может протекать и межмолекулярно (и это интересно), если в качестве спиртовой компоненты выступает глицерин, а вернее, его производное или предшественник, а жирная кислота, как и в предыдущем случае, активирована по карбоксильной группе коферментом А. Теперь вполне очевидно, что продуктами такой перезтерификации будут жиры и жироподобные соединения — липиды. В различных организмах биосинтез липидов осуществляется различными, но достаточно близкими путями. Как уже упомянуто, жирная кислота Схема 5.4.7
синтезируется самостоятельно и в коферментно связанной форме взаимодействует с глицерином Но вариант участия собственно глицерина редок, так как процесс с участием трех свободных гидроксилов не обеспечит структурной селективности, если необходимо ввести различные кислотные остатки. Обычно реакция начинается от дигидроксиацетона или от 2,3-дигидроксипропанола — оба они образуются при катаболизме углеводов. Здесь мы опишем схему биосинтеза липидов (как наиболее селективную), стартуя от дигидроксиацетона (схема 5.4.7). На первом этапе дигидроксиацетон фосфорилируется по одной спиртовой функции, образуя соответствующий фосфат. Оставшаяся свободная гидроксильная группа теперь может быть достаточно однозначно ацилирована коферментно связанной жирной кислотой На следующей стадии карбональная группа с помощью NAD-H восстанавливается до спиртовой, которая в свою очередь может быть этерифицирована уже другой молекулой коферментно активированной жирной кислоты Таким образом, образуется фосфатидная кислота, которая далее может идти на формирование фосфолипидов либо при гидролизе высвободить еще одну спиртовую функцию для окончательного ацилирования, образующего триглицериды — жиры.
|
 |
Оглавление
|

























































































































































































































































































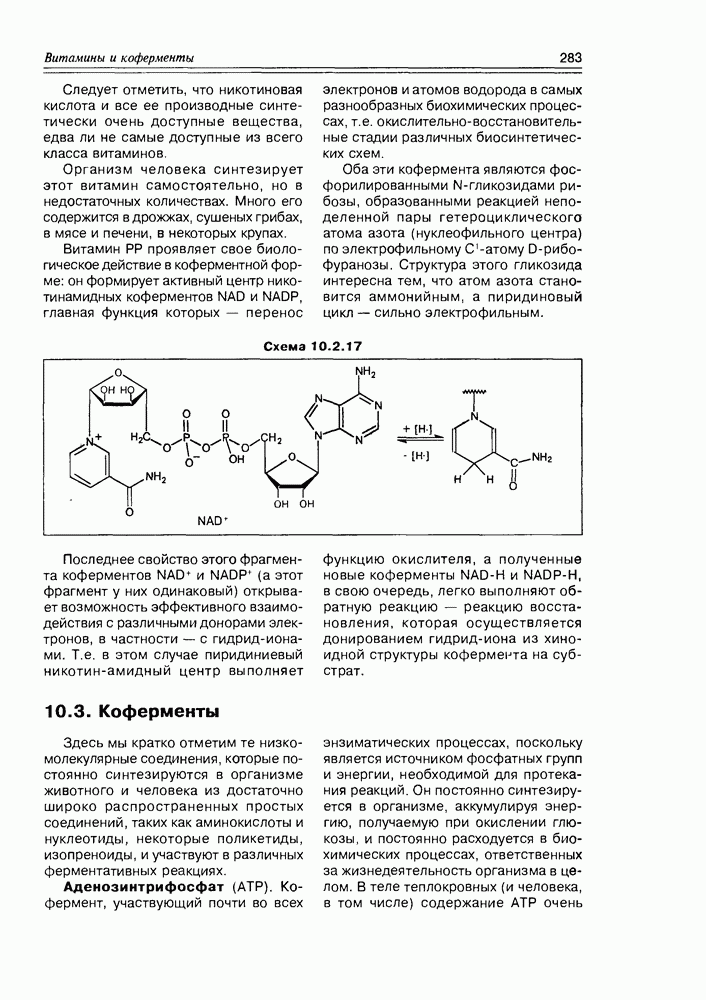




















































































 , в качестве окислителя выступает кофермент
, в качестве окислителя выступает кофермент  в общем, весь процесс катализируется мультиферментным комплексом, представленным высокоинтегрированной системой из трех видов ферментов.
в общем, весь процесс катализируется мультиферментным комплексом, представленным высокоинтегрированной системой из трех видов ферментов.
 замечательна тем, что она имеет два реакционных центра — электрофильный и нуклеофильный, которые обеспечивают процесс сшивки ацетильных фрагментов.
замечательна тем, что она имеет два реакционных центра — электрофильный и нуклеофильный, которые обеспечивают процесс сшивки ацетильных фрагментов.

 . Нуклеофильную активность в молекуле “активного ацетила" в условиях основного катализа проявляет метильная группа: основание отщепляет от метила ацетильного фрагмента закисленный водород, формируя таким способом карбанионный центр. Суммируя эти возможности ацетил-S-CoA, нетрудно увидеть, что одна молекула активного ацетила выступает в роли нуклеофила и может замещать коферментный фрагмент другой такой же молекулы, образуя уже четырехуглеродный фрагмент - ацетоацетил-S-CoA.
. Нуклеофильную активность в молекуле “активного ацетила" в условиях основного катализа проявляет метильная группа: основание отщепляет от метила ацетильного фрагмента закисленный водород, формируя таким способом карбанионный центр. Суммируя эти возможности ацетил-S-CoA, нетрудно увидеть, что одна молекула активного ацетила выступает в роли нуклеофила и может замещать коферментный фрагмент другой такой же молекулы, образуя уже четырехуглеродный фрагмент - ацетоацетил-S-CoA.
 атакует вторую, немодифицированную, молекулу ацетил-SCoA, образуя малоно-ацетил-SCoA; последний декарбоксилируется до ацето-ацетил-SCoA (схема 5.4.2). Второй путь несколько более длинный, но зато он более энергетически выгодный, так как каждая стадия требует небольшой энергии активации, т.е. общий энергетический барьер процесса преодолевается по ступенькам, что более удобно, а иногда и единственно возможно для живых систем.
атакует вторую, немодифицированную, молекулу ацетил-SCoA, образуя малоно-ацетил-SCoA; последний декарбоксилируется до ацето-ацетил-SCoA (схема 5.4.2). Второй путь несколько более длинный, но зато он более энергетически выгодный, так как каждая стадия требует небольшой энергии активации, т.е. общий энергетический барьер процесса преодолевается по ступенькам, что более удобно, а иногда и единственно возможно для живых систем.
 фрагмента уже в молекуле ацето-ацетил-S-CoA может совершиться вновь по одной из приведенных выше схем по точно тому же механизму, образуя уже шестиуглеродный фрагмент все с той же коферментной группой на конце молекулы. Таким образом, процесс наращивания углеродной цепи может продолжаться и далее, образуя соединения общей формулы
фрагмента уже в молекуле ацето-ацетил-S-CoA может совершиться вновь по одной из приведенных выше схем по точно тому же механизму, образуя уже шестиуглеродный фрагмент все с той же коферментной группой на конце молекулы. Таким образом, процесс наращивания углеродной цепи может продолжаться и далее, образуя соединения общей формулы  называемые поликетидами. А описанный выше биосинтетический путь называется поликетидным.
называемые поликетидами. А описанный выше биосинтетический путь называется поликетидным.
 и т.д. по тому же механизму “ацильной сборки”, либо включаться в последующие биосинтетические превращения. Из последующих (после сборки алкильной цепи) реакций насыщенных жирных кислот необходимо выделить образование ненасыщенных жирных кислот — олеиновой кислоты из стеариновой, как первоочередной. Эта реакция катализируется ферментом дэдесатуразой в присутствии молекулярного кислорода и NAD-H. Этот фермент селективен по месту образования двойной связи
и т.д. по тому же механизму “ацильной сборки”, либо включаться в последующие биосинтетические превращения. Из последующих (после сборки алкильной цепи) реакций насыщенных жирных кислот необходимо выделить образование ненасыщенных жирных кислот — олеиновой кислоты из стеариновой, как первоочередной. Эта реакция катализируется ферментом дэдесатуразой в присутствии молекулярного кислорода и NAD-H. Этот фермент селективен по месту образования двойной связи  и ее конфигурации
и ее конфигурации  Эта селективность указывает на механизм процесса, как синхронного цис-элиминирования с предварительной фиксацией карбоксильной группы кислоты на определенной функции фермента. Известно, что фермент десатураза в своей простетической группе содержит
Эта селективность указывает на механизм процесса, как синхронного цис-элиминирования с предварительной фиксацией карбоксильной группы кислоты на определенной функции фермента. Известно, что фермент десатураза в своей простетической группе содержит
 участвующий в переносе электрона, поскольку процесс дегидрирования, в общем случае — это процесс окисления. Установлено также, что кислород вступает в реакцию на уровне десатуразы, являясь акцептором водородных атомов. Так что процесс цис-дегидрирования стеариновой кислоты до олеиновой может быть представлен схемой с элементом переходного состояния, включающего атом железа
участвующий в переносе электрона, поскольку процесс дегидрирования, в общем случае — это процесс окисления. Установлено также, что кислород вступает в реакцию на уровне десатуразы, являясь акцептором водородных атомов. Так что процесс цис-дегидрирования стеариновой кислоты до олеиновой может быть представлен схемой с элементом переходного состояния, включающего атом железа  молекула кислорода
молекула кислорода  фрагмент алкильной цепи в гош-конформации (схема 5.4.4).
фрагмент алкильной цепи в гош-конформации (схема 5.4.4).

 по гиперсопряженной аллильной системе. И тот и другой механизмы контролируются ферментами гипоксигеназами, которые обеспечивлюг активацию молекулы кислорода (обычно они содержат ион железа) и конфигурационную селективность гидроксилирования. В любом случае, мы получаем гидро-перекисную кислоту с вторичной аллильно-перекисной функцией и сопряженным диеновым фрагментом Е, Z-конфигурации. Гидроперекиси непредельных жирных кислот далее претерпевают несложные in vivo трансформации либо в соответствующие гидроксикислоты, либо в фурановые жирные кислоты (схема 5.4.5).
по гиперсопряженной аллильной системе. И тот и другой механизмы контролируются ферментами гипоксигеназами, которые обеспечивлюг активацию молекулы кислорода (обычно они содержат ион железа) и конфигурационную селективность гидроксилирования. В любом случае, мы получаем гидро-перекисную кислоту с вторичной аллильно-перекисной функцией и сопряженным диеновым фрагментом Е, Z-конфигурации. Гидроперекиси непредельных жирных кислот далее претерпевают несложные in vivo трансформации либо в соответствующие гидроксикислоты, либо в фурановые жирные кислоты (схема 5.4.5).
 . Гидроксикислоты в коферментно связанной форме используют свою спиртовую функцию в качестве нуклеофила, для этого она может активироваться любым основанием из этого же кофермента (фосфат-анион или азотистый гетероцикл — аденин),
. Гидроксикислоты в коферментно связанной форме используют свою спиртовую функцию в качестве нуклеофила, для этого она может активироваться любым основанием из этого же кофермента (фосфат-анион или азотистый гетероцикл — аденин),


