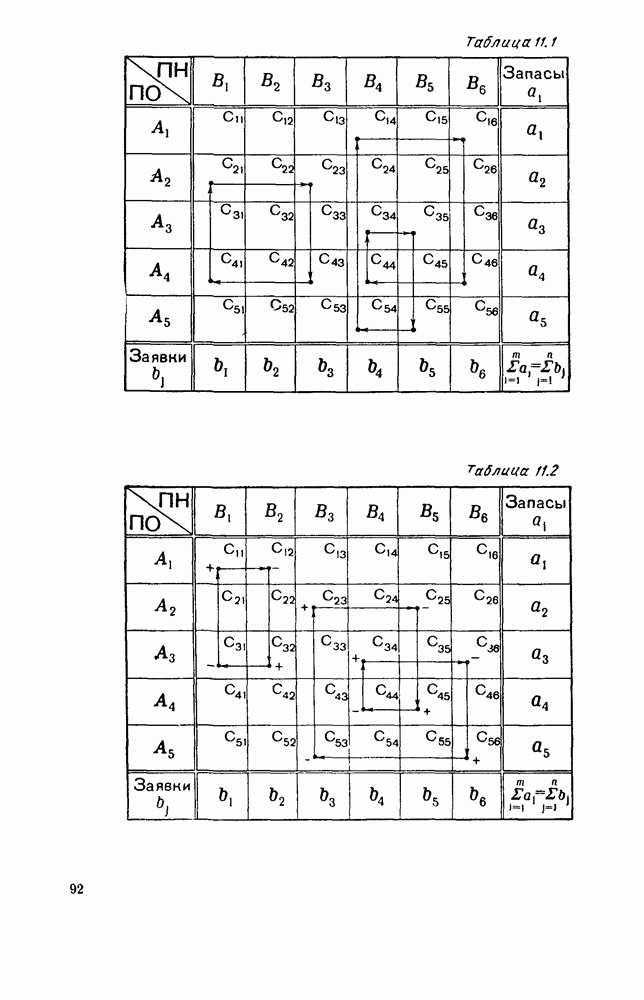
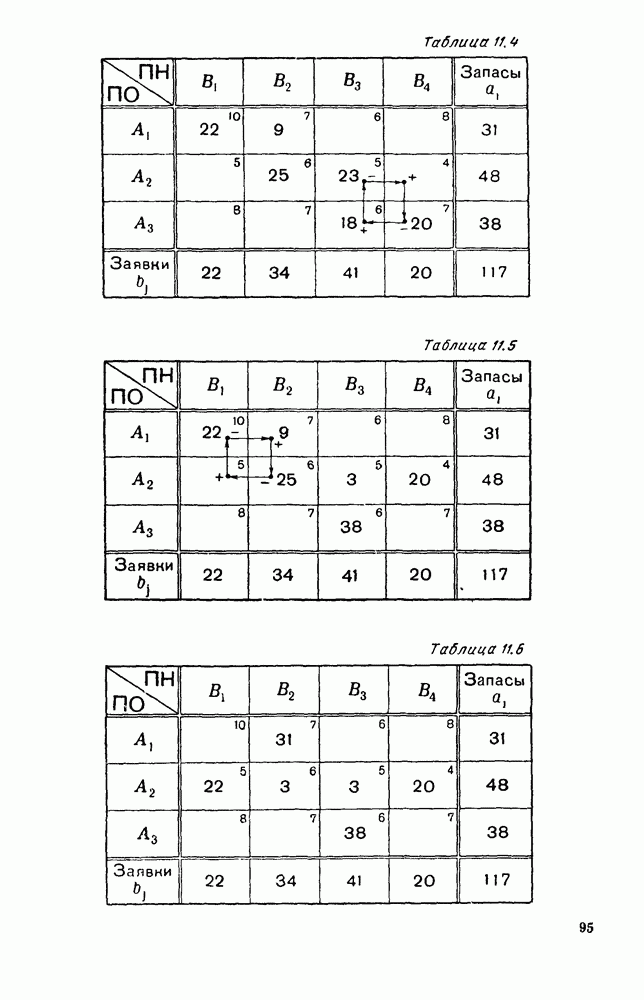
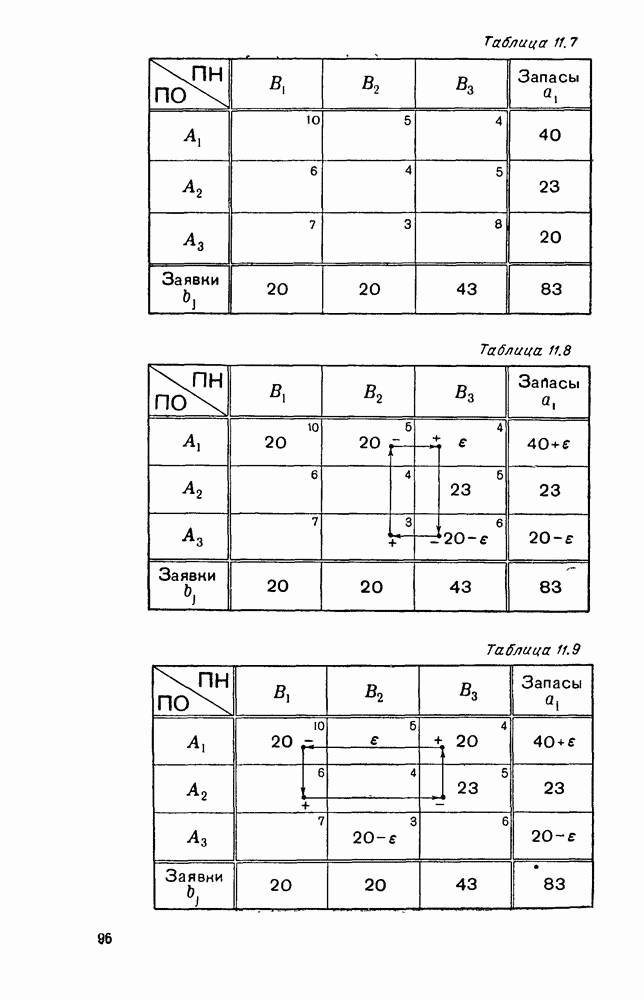
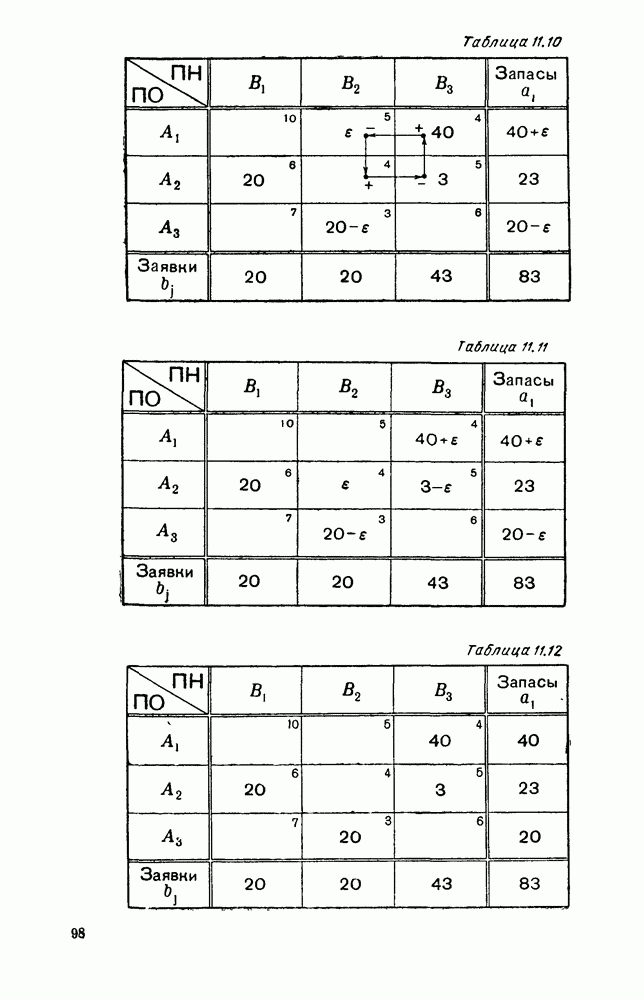
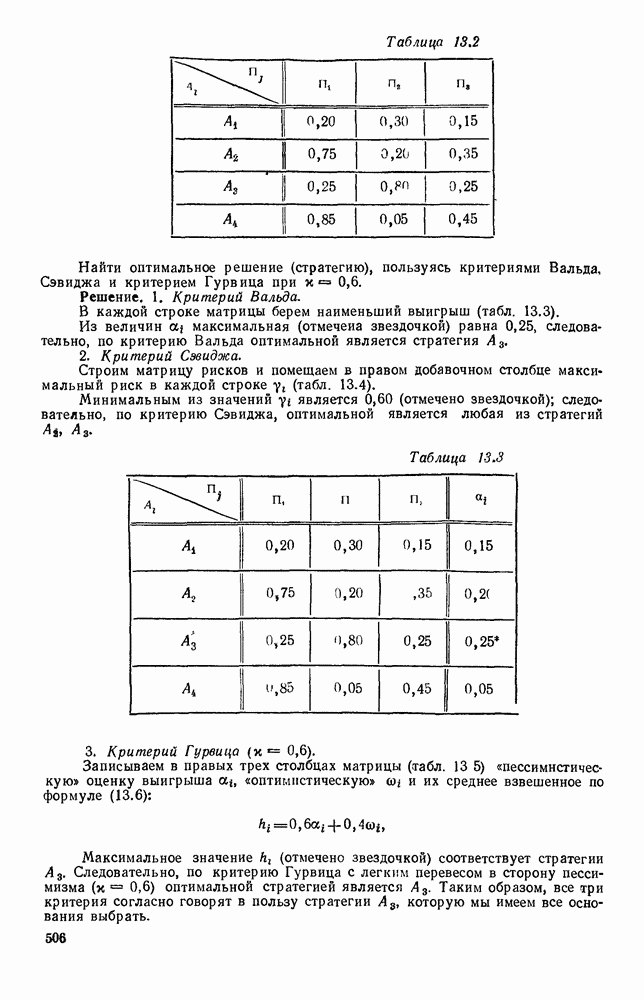
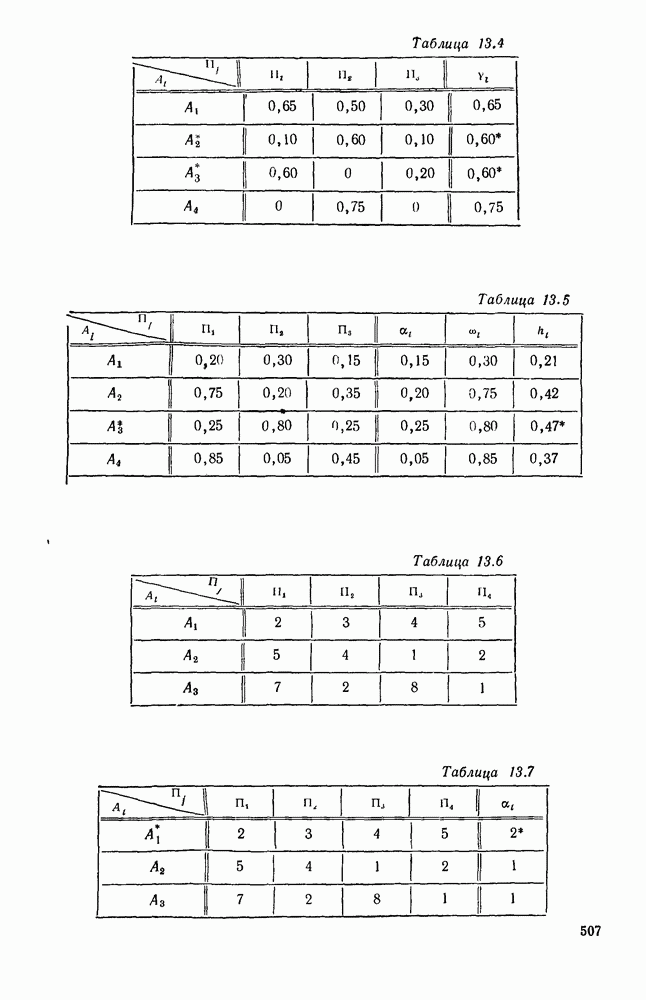
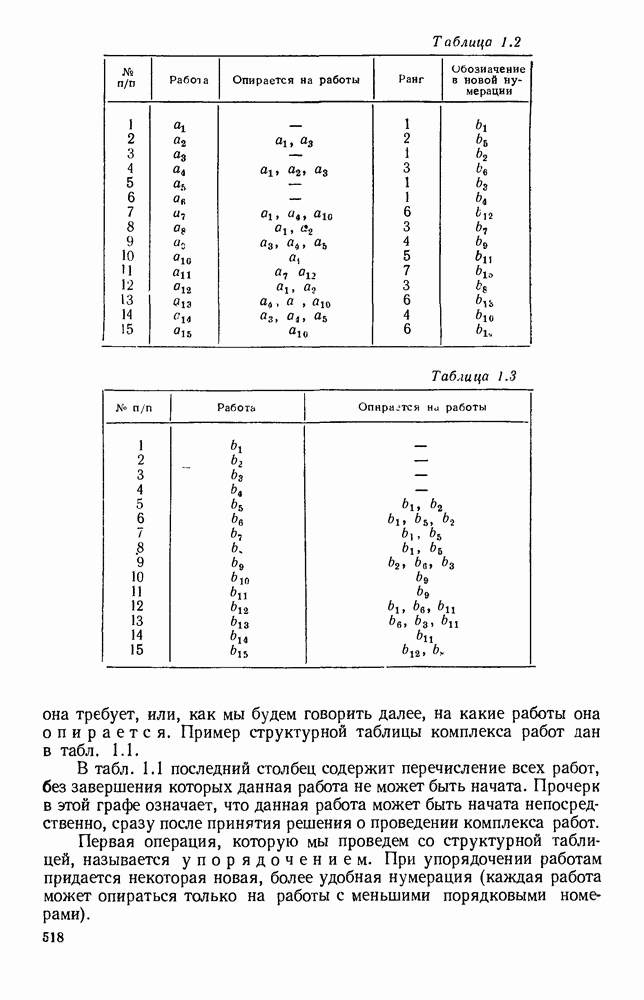
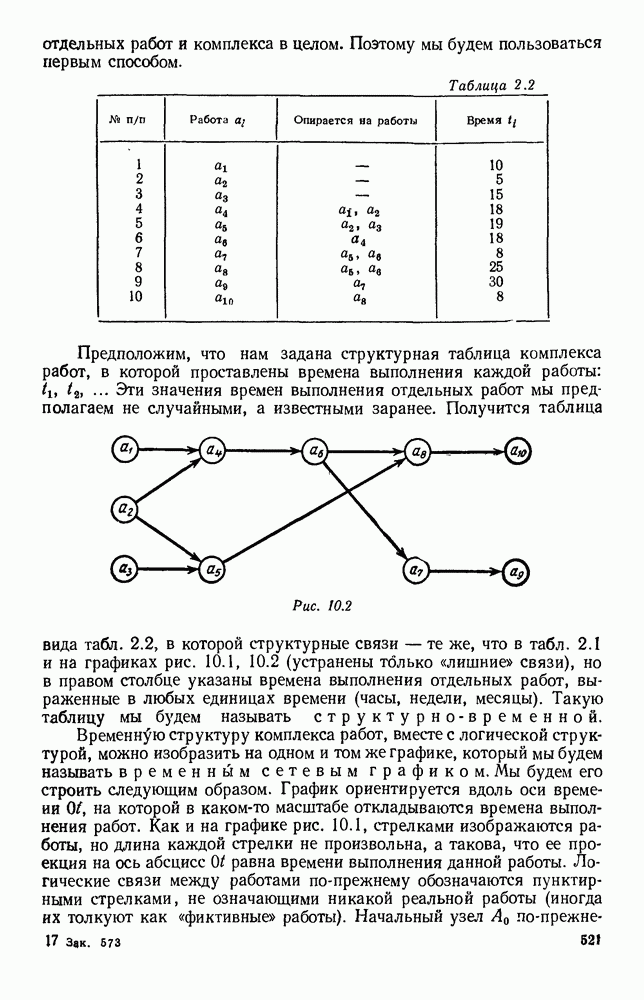
Пред.
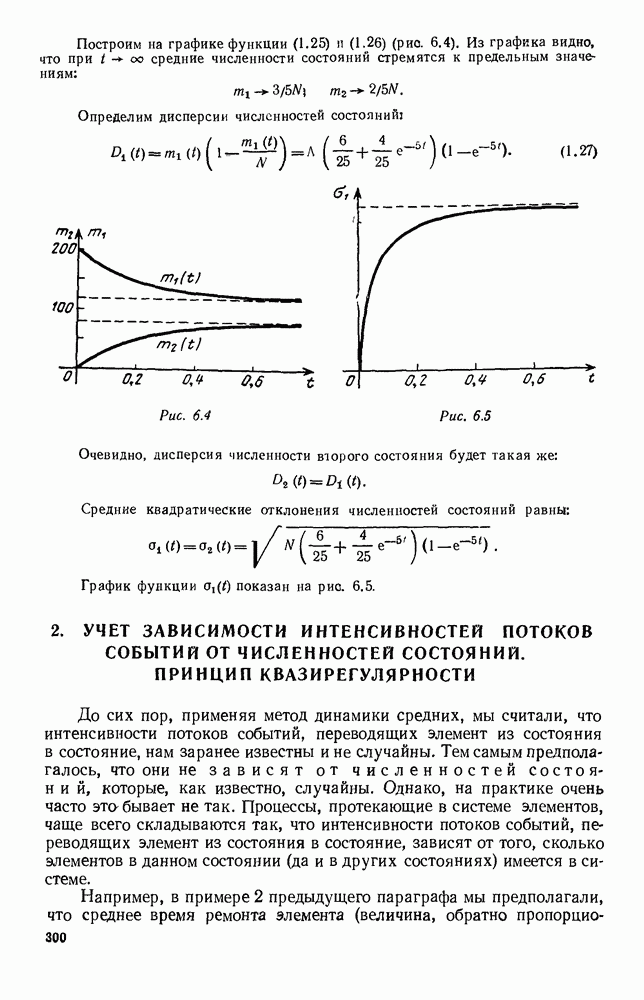
След.
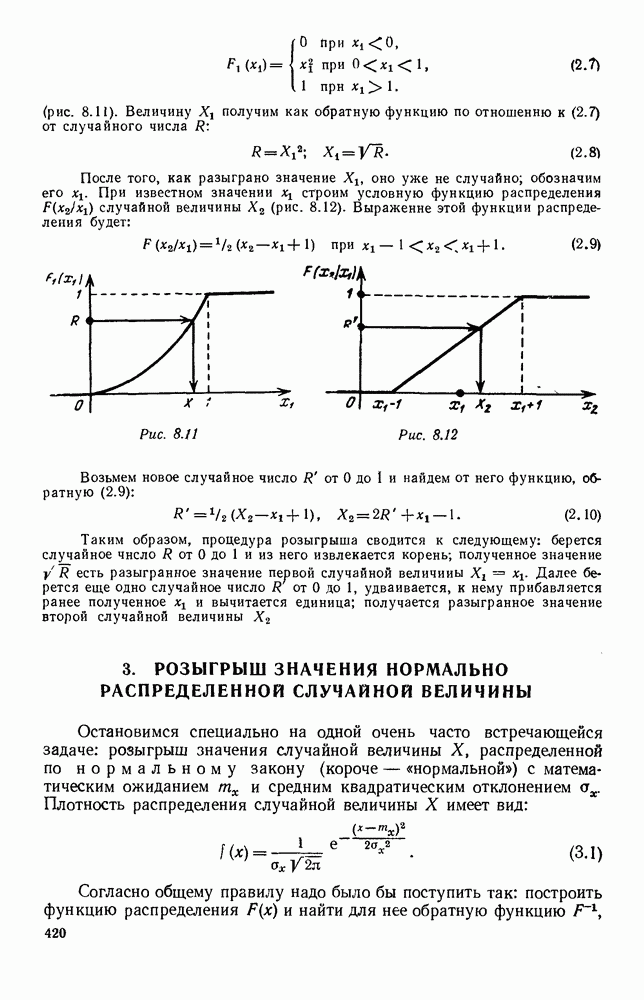
Макеты страниц
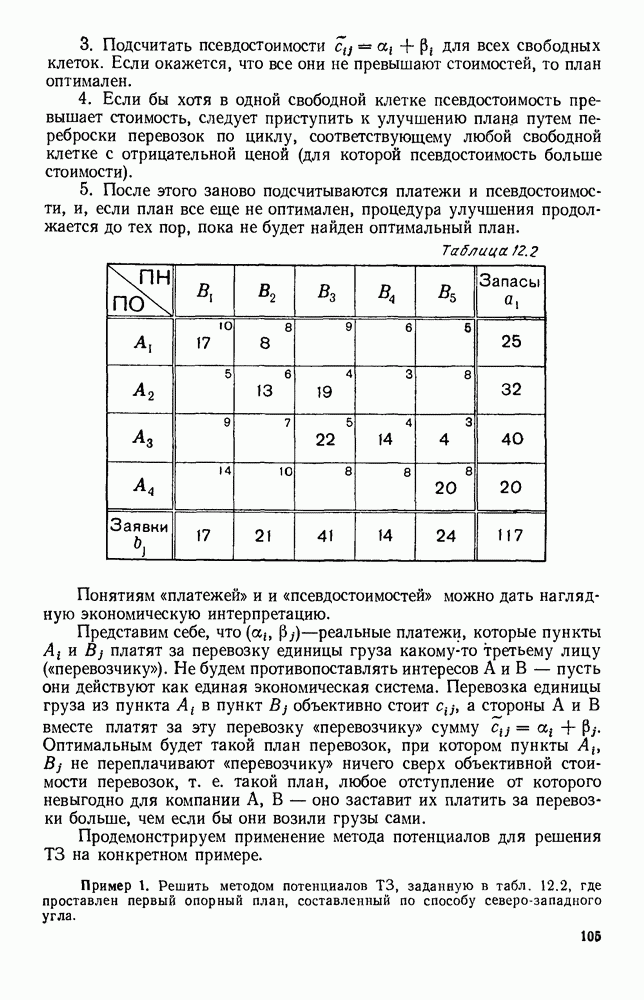
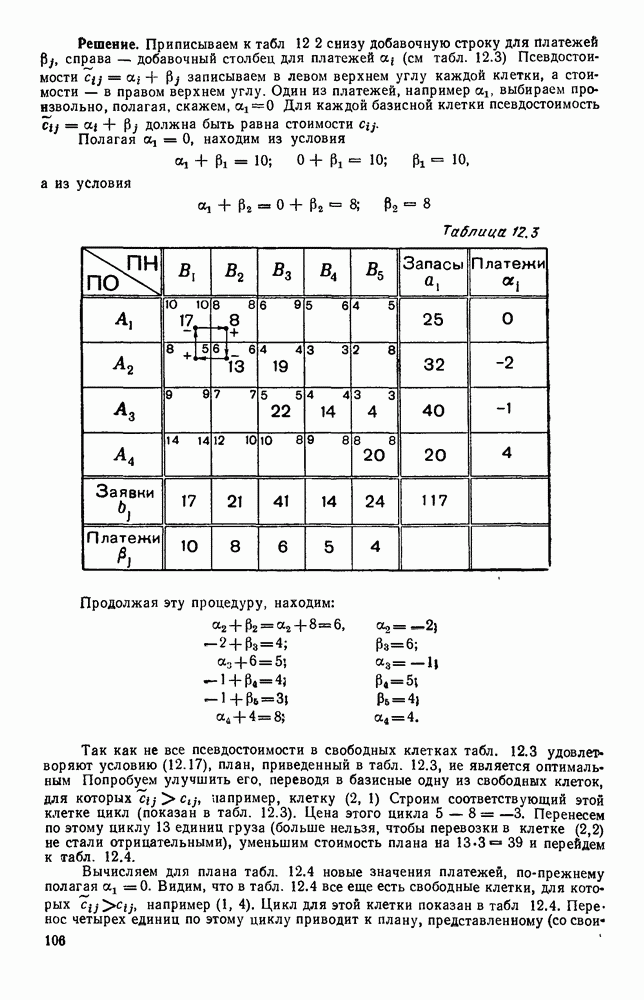
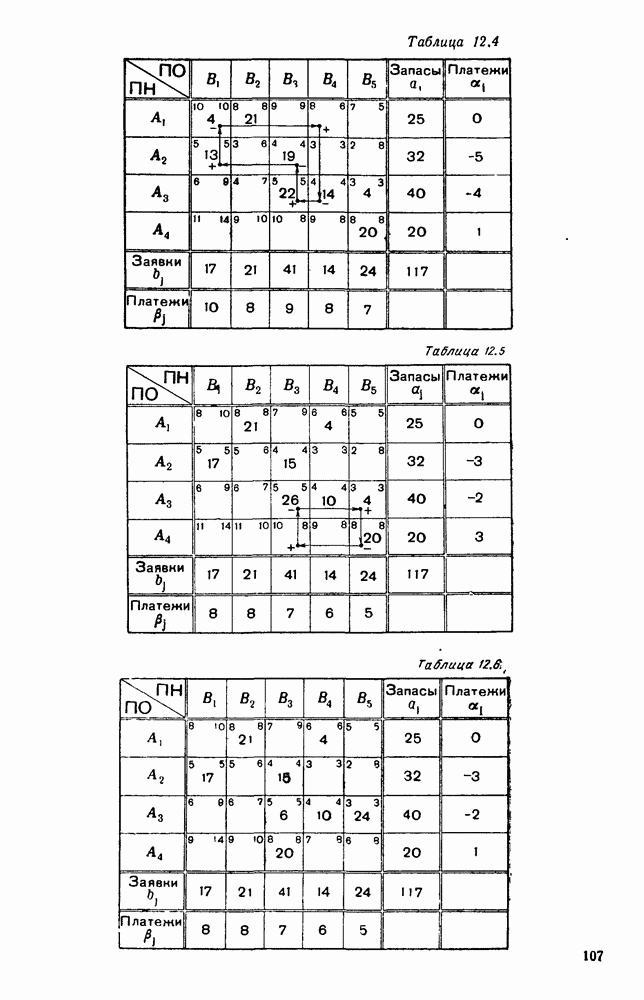
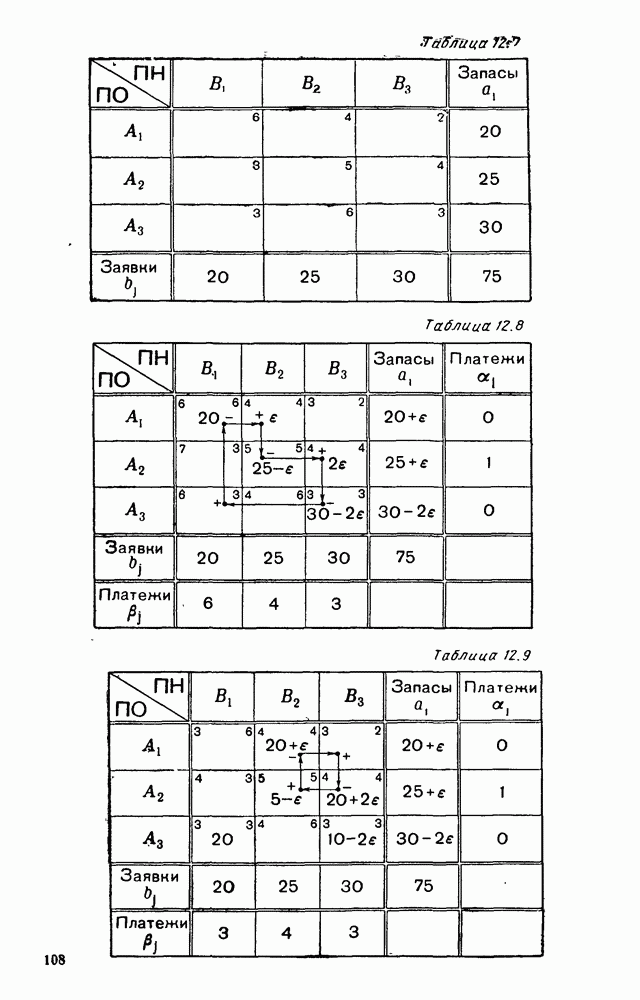
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
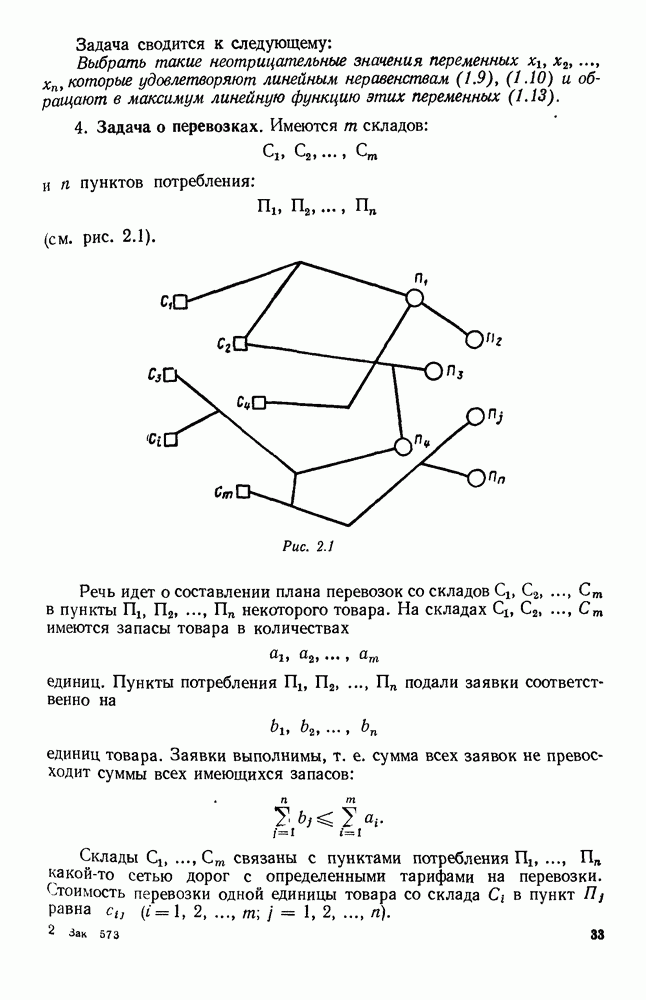
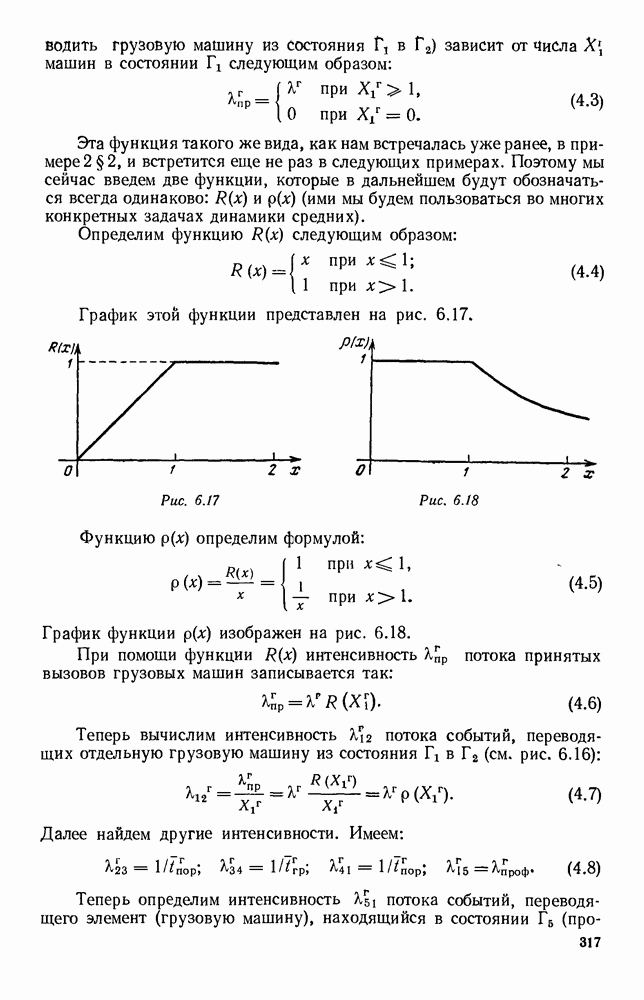
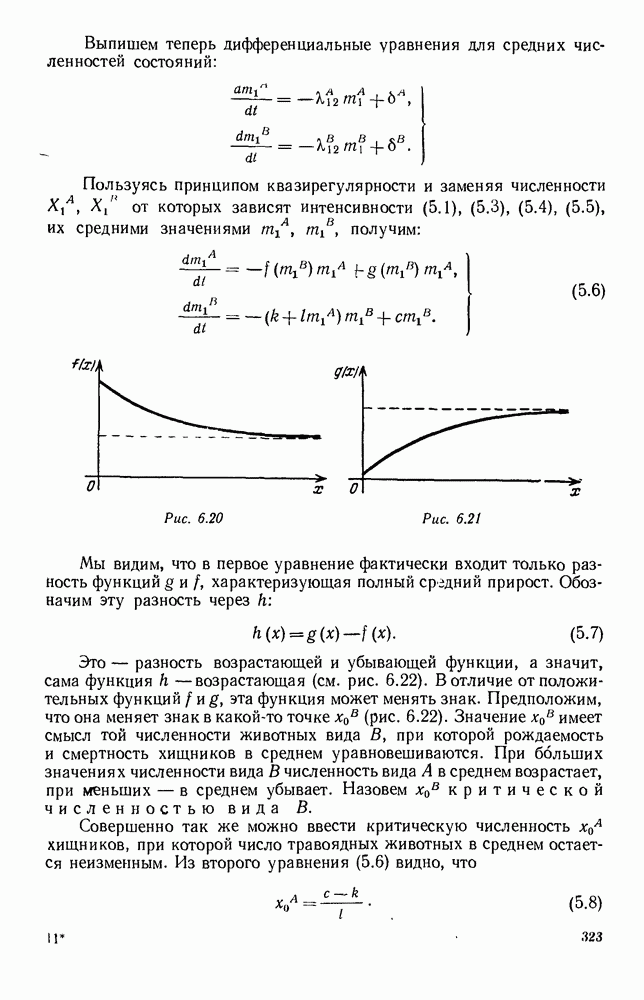
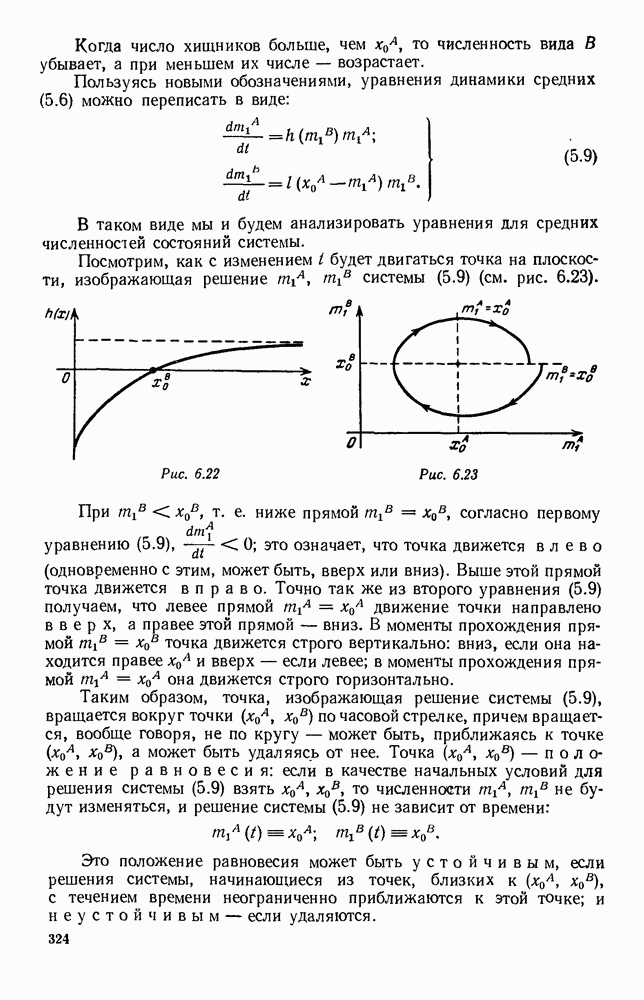
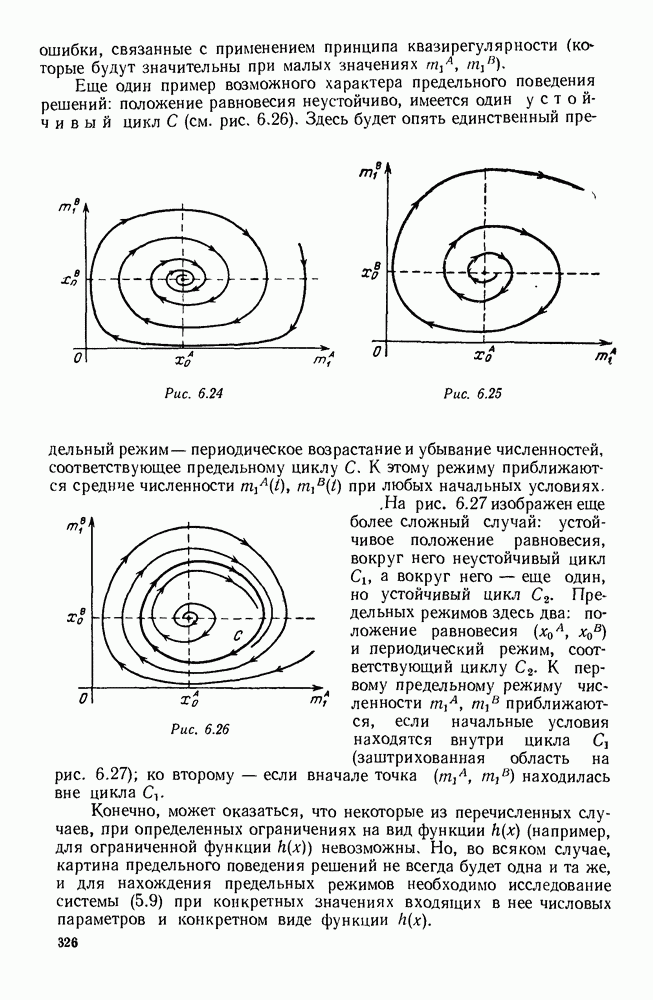
ZADANIA.TO
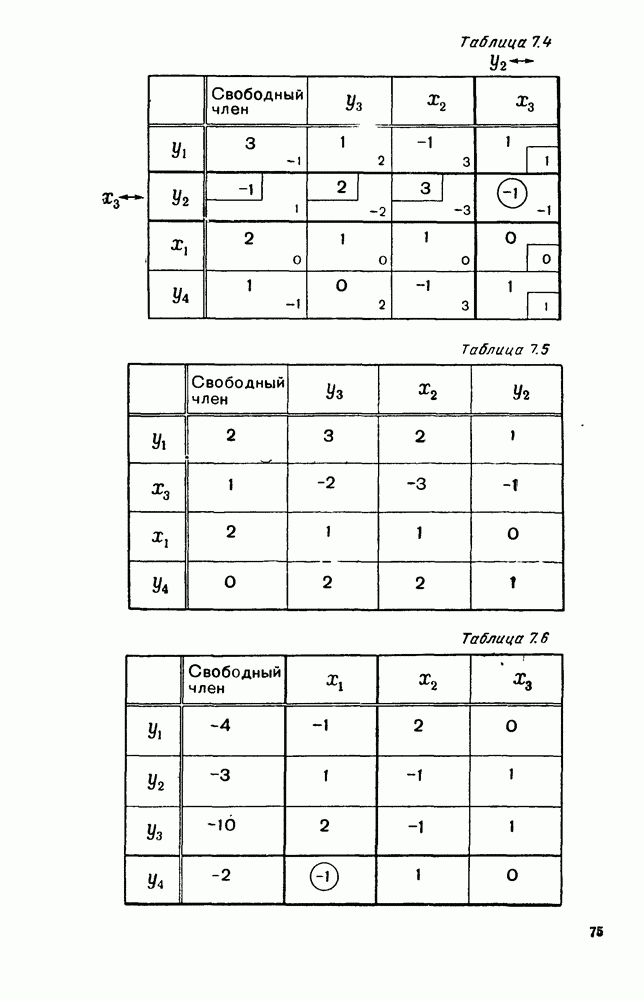
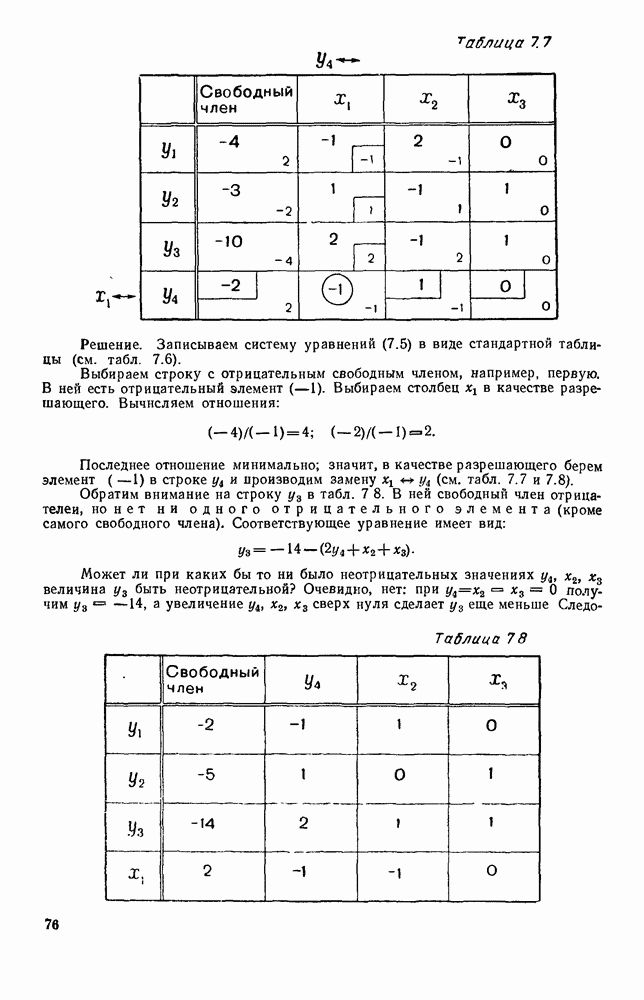
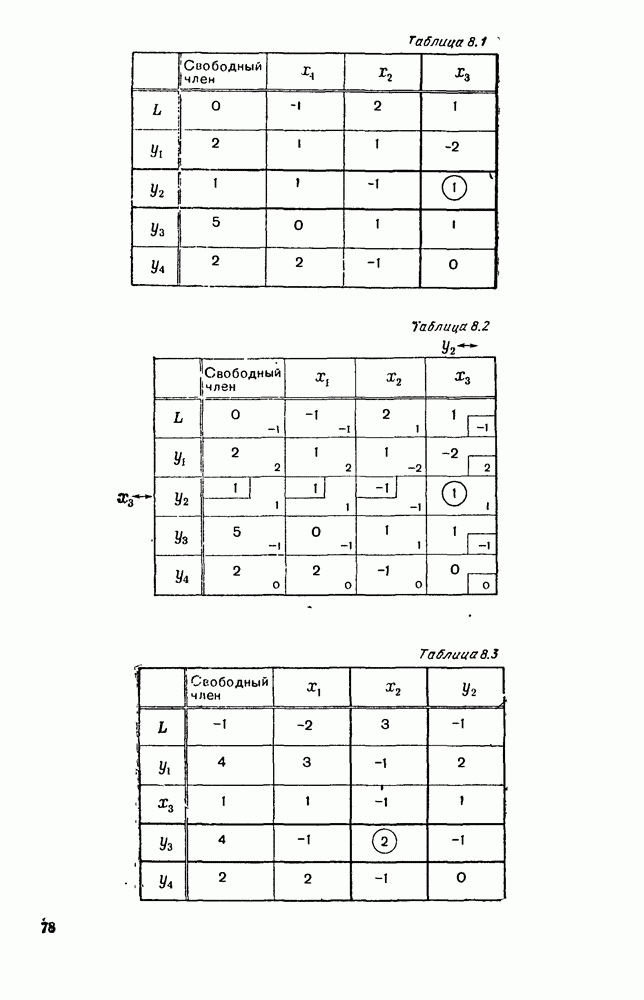
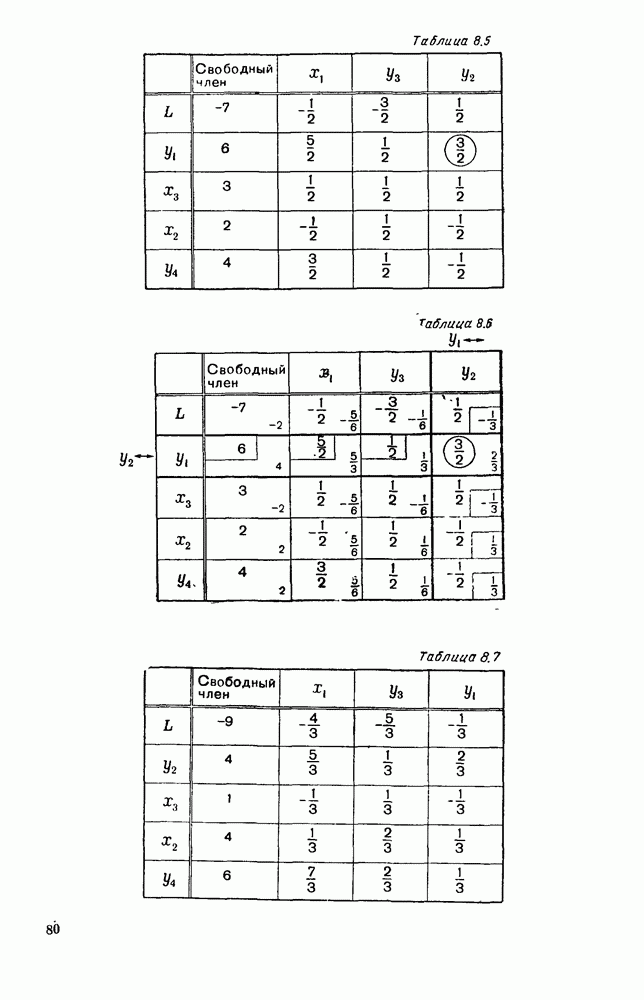
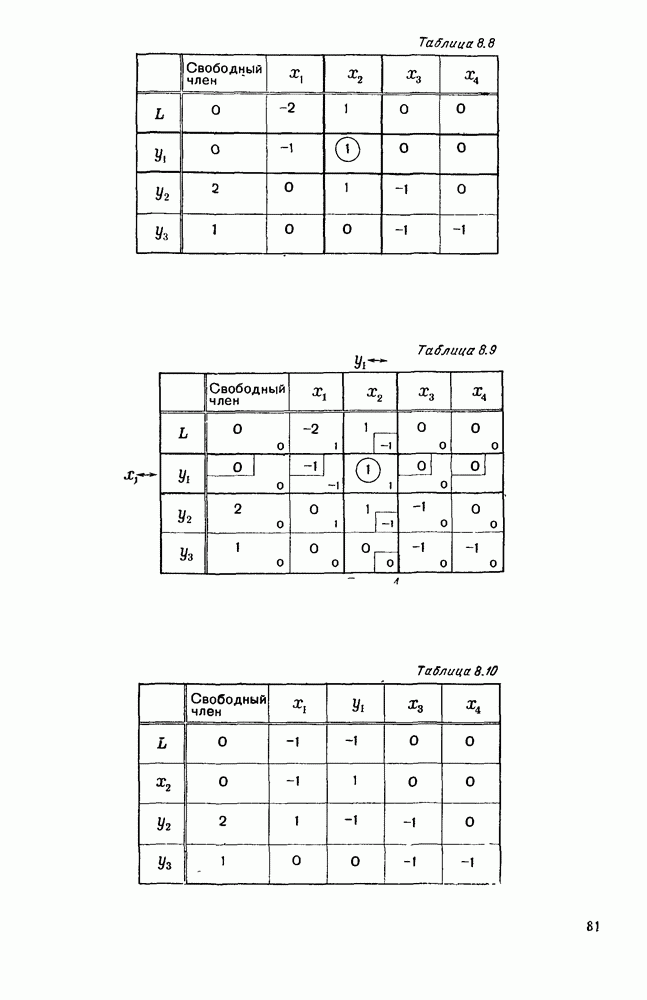
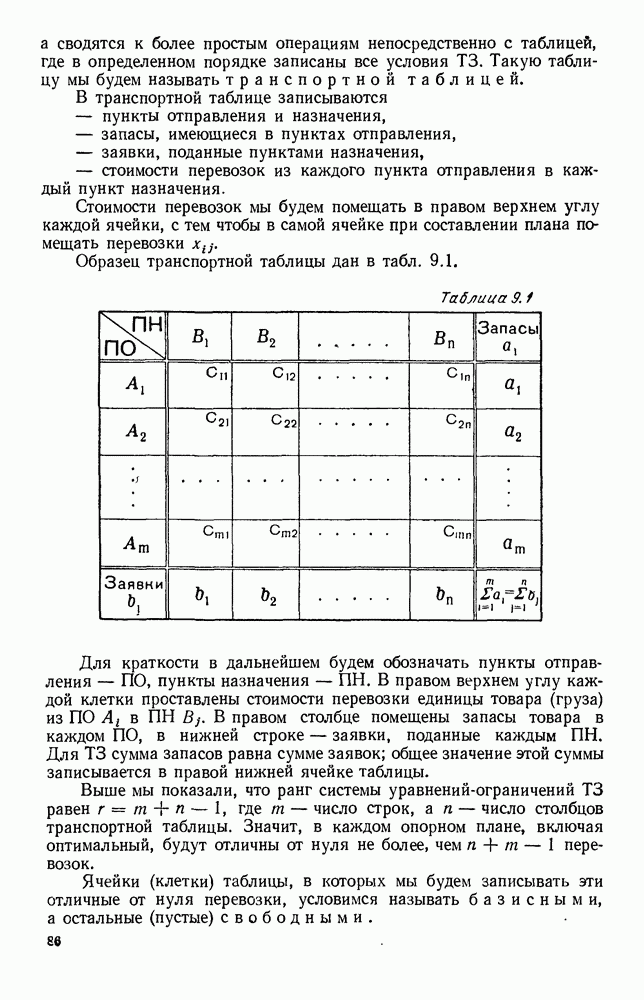
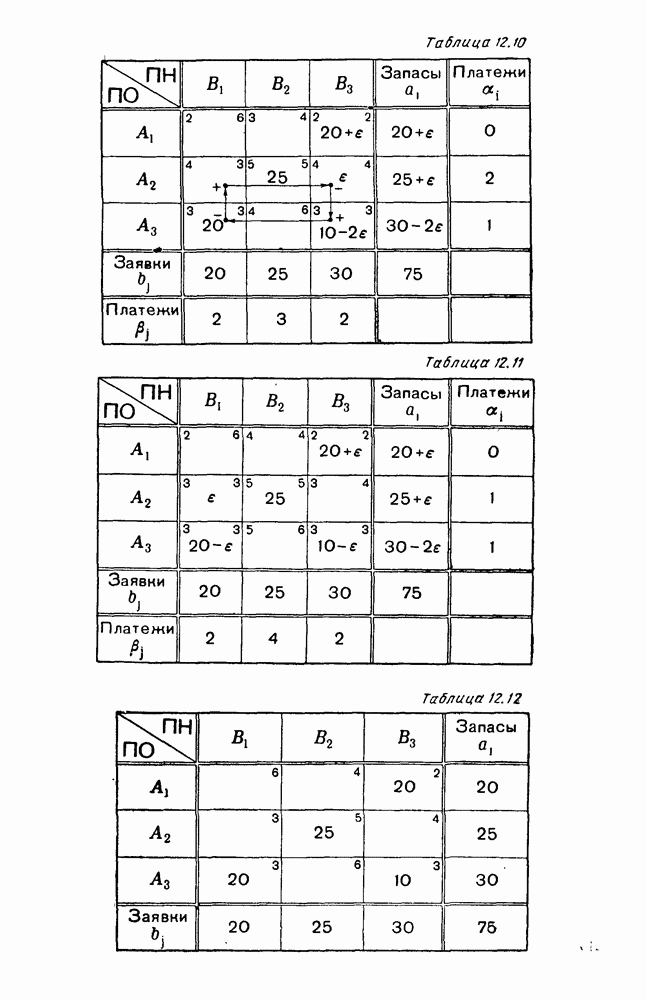
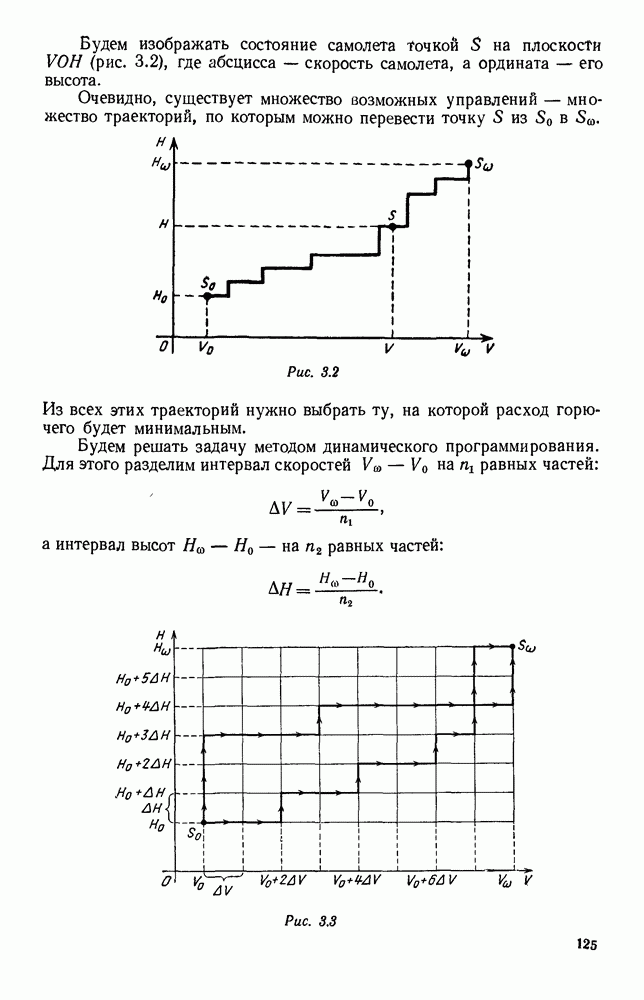
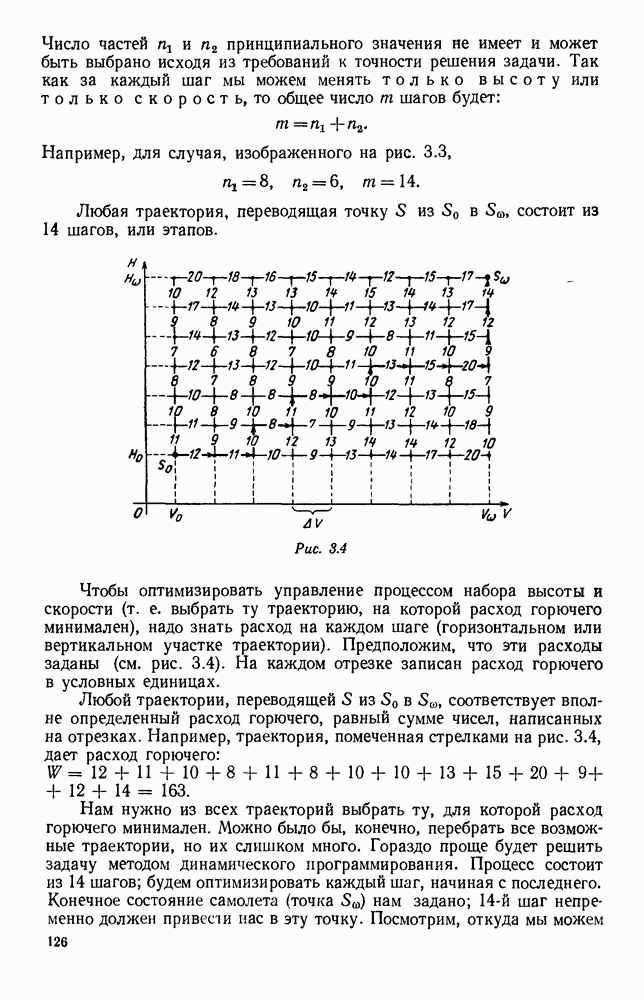
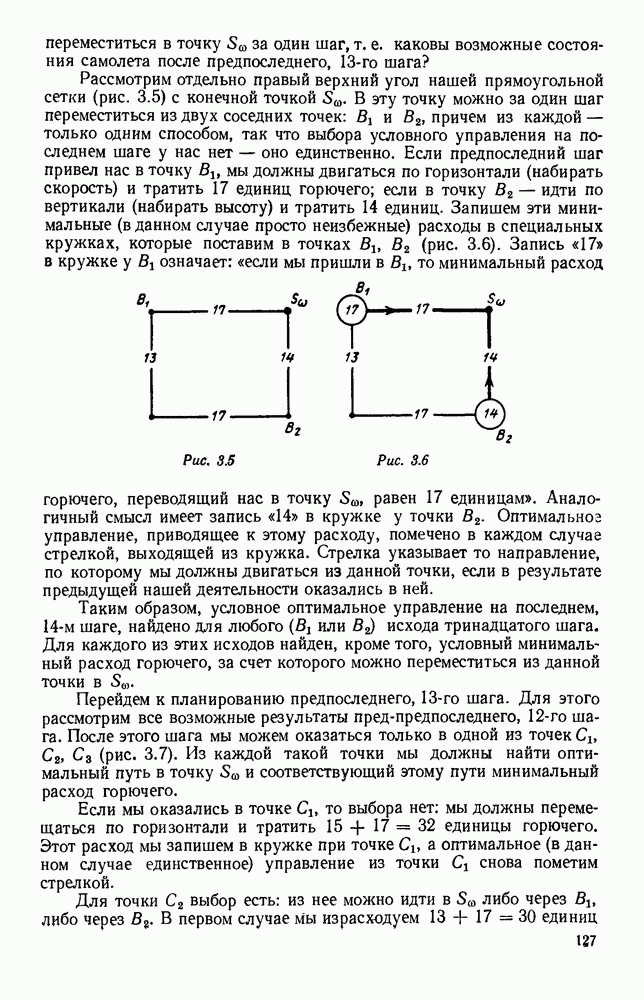
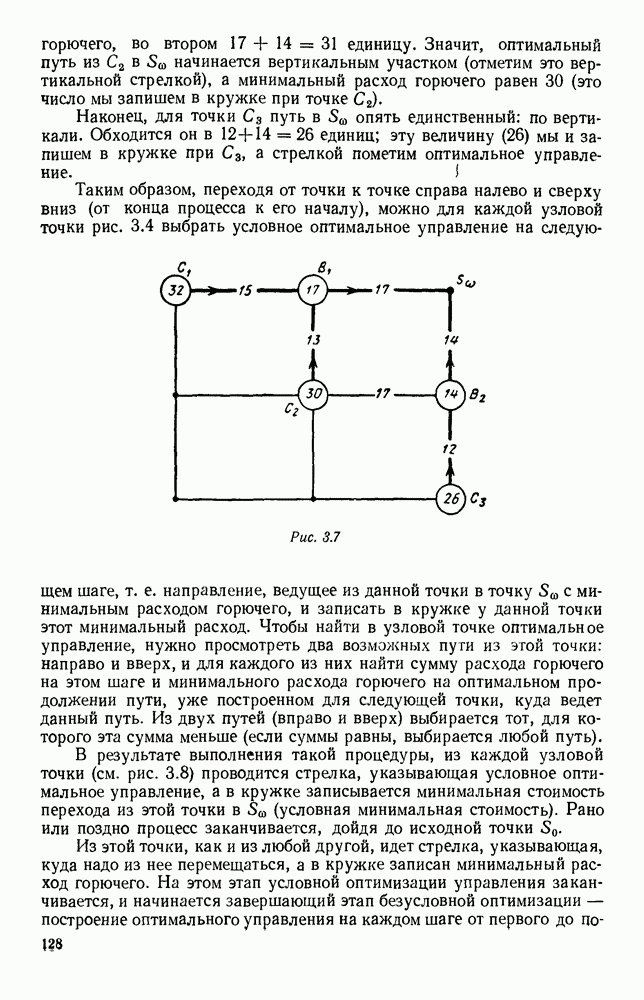
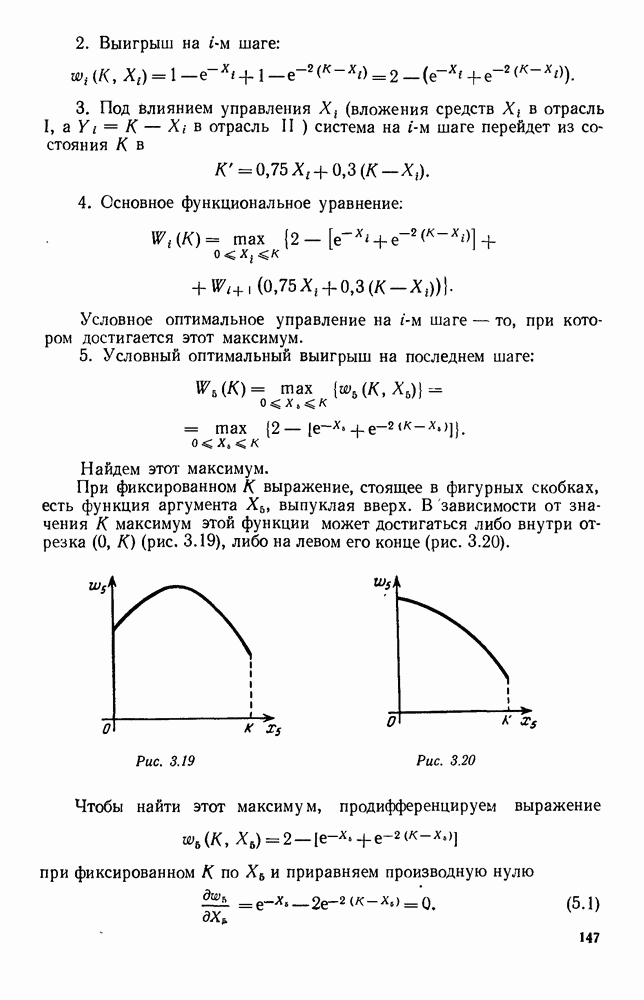
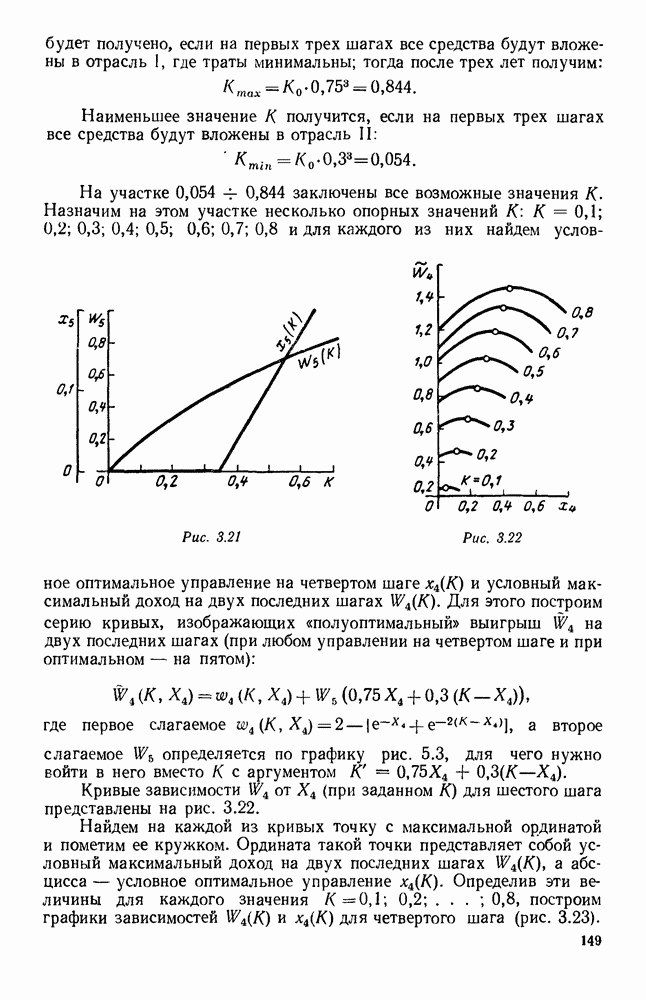
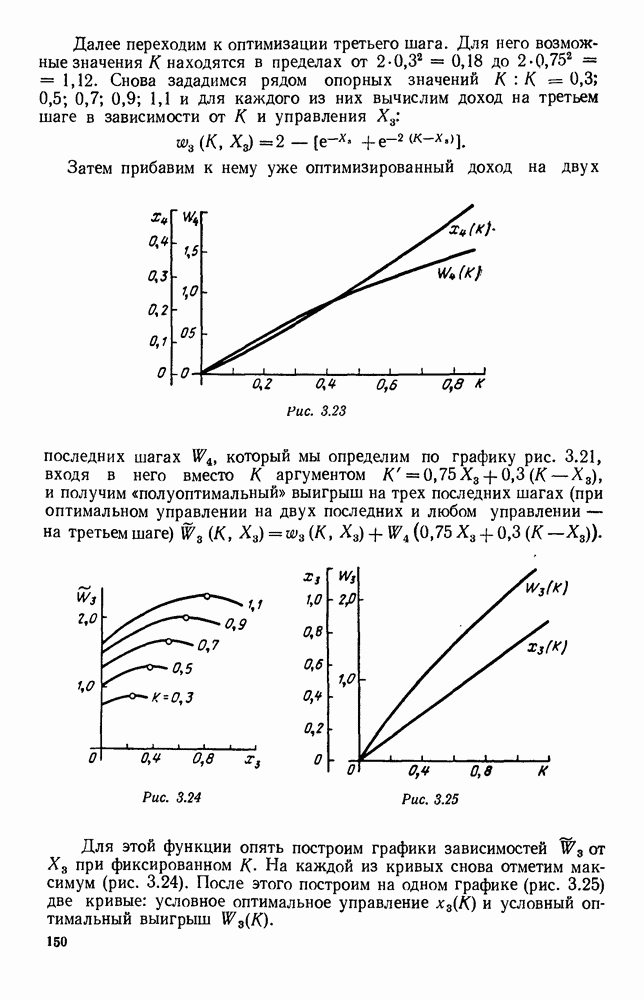
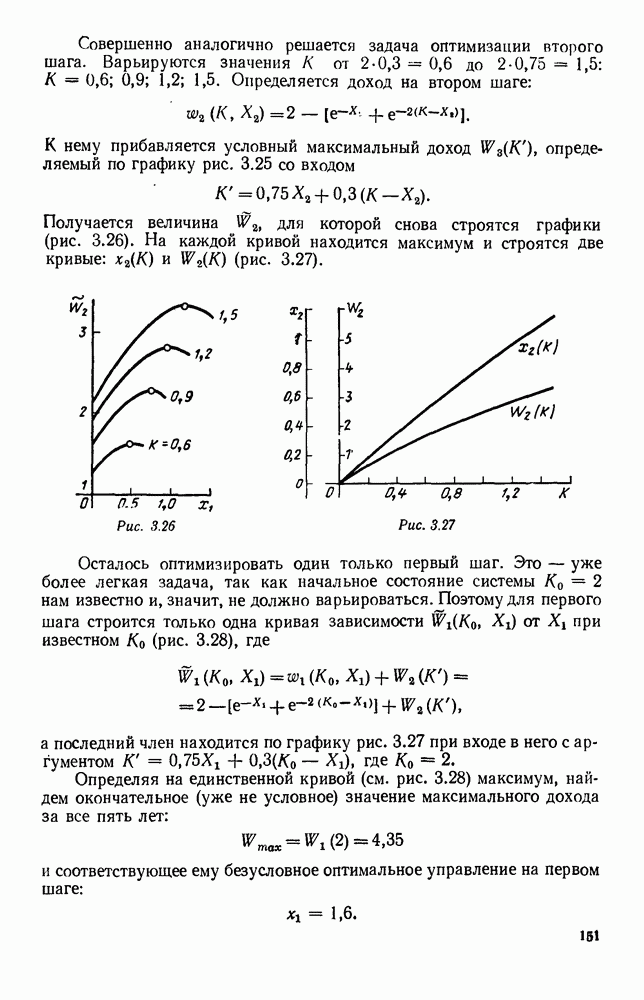
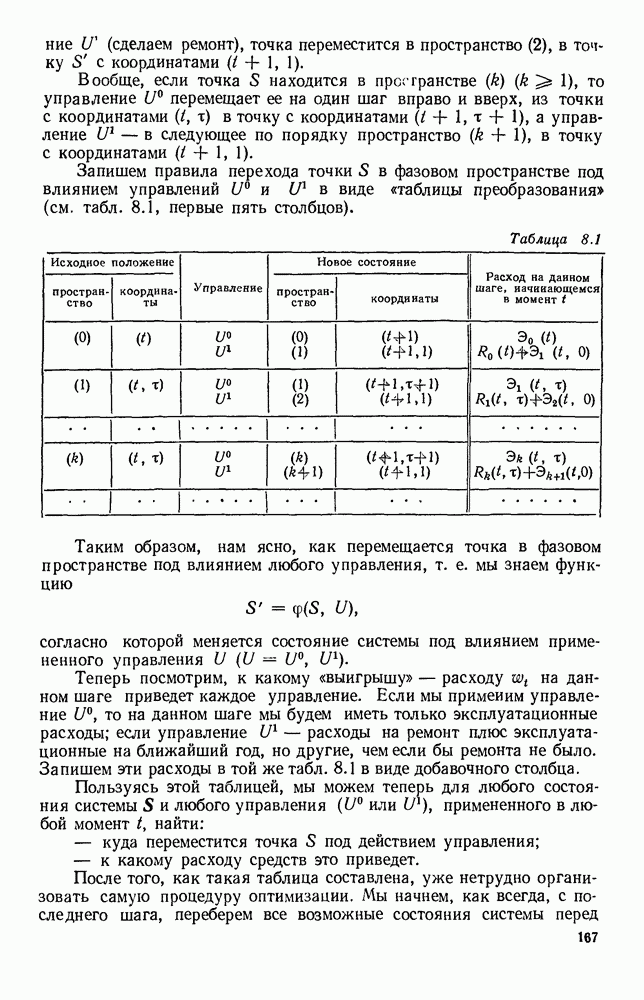
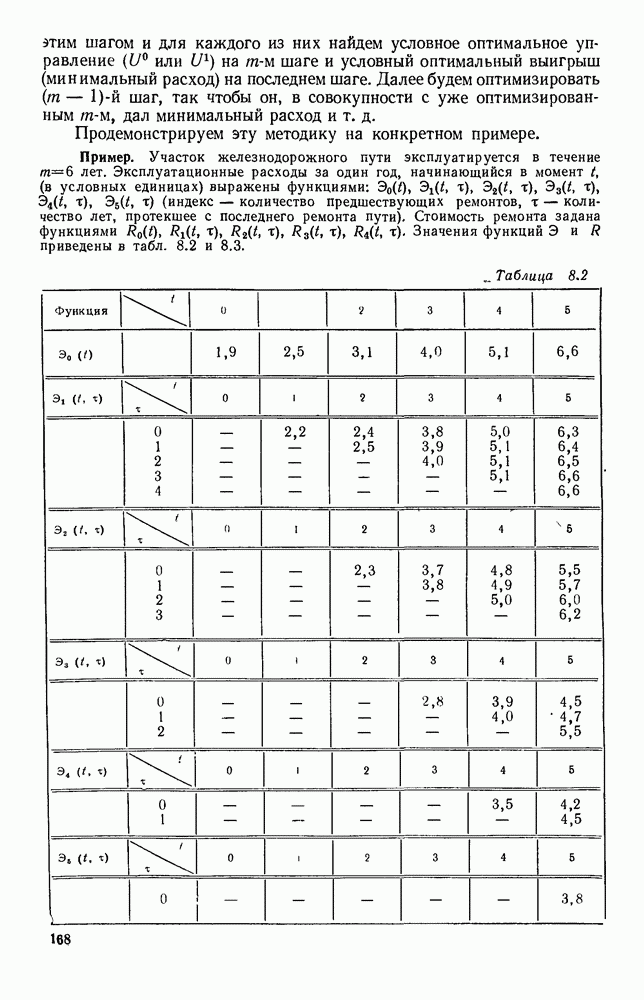
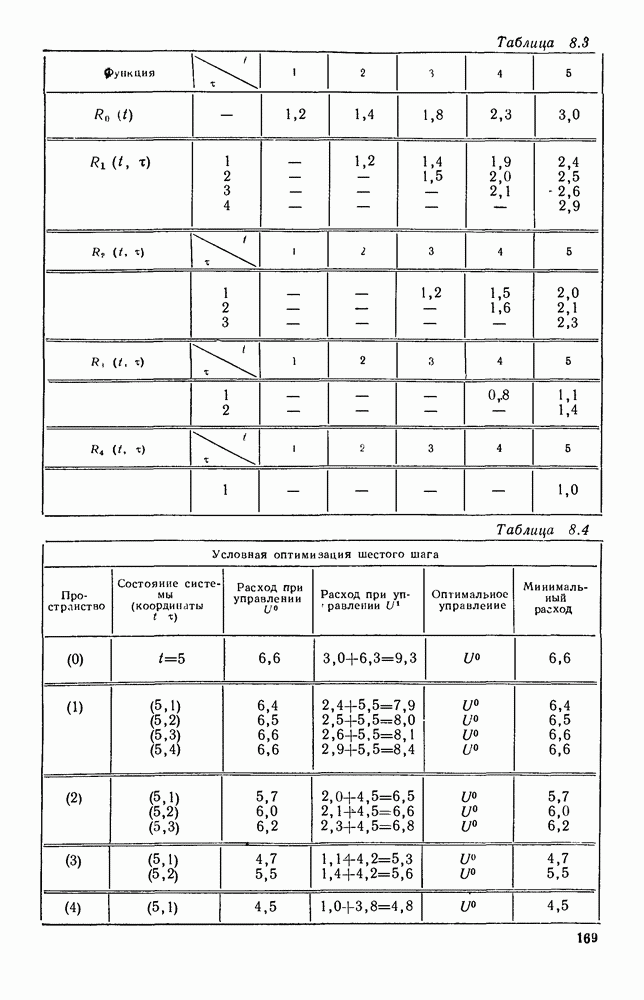
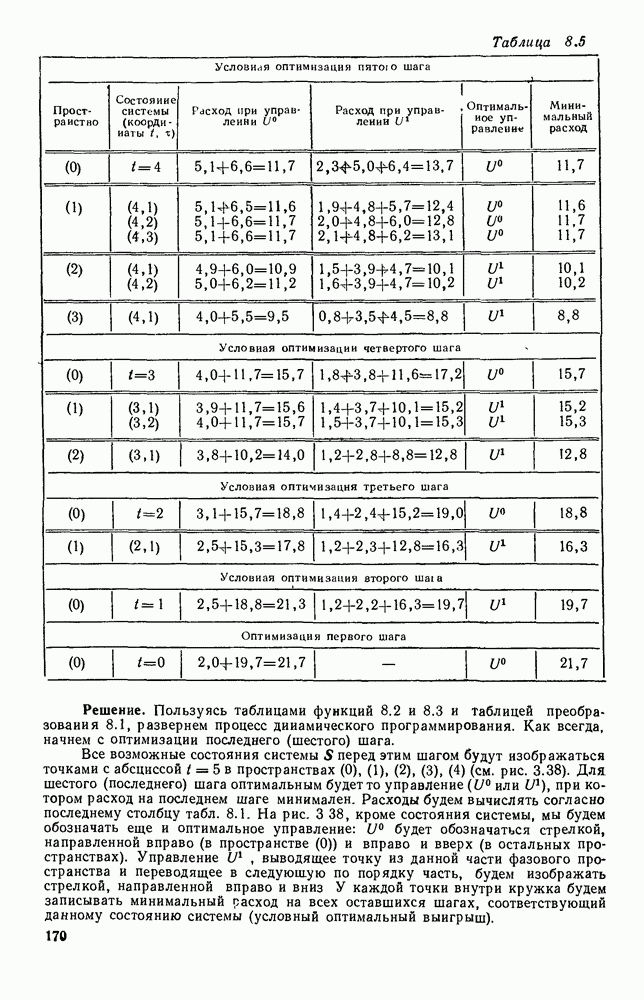
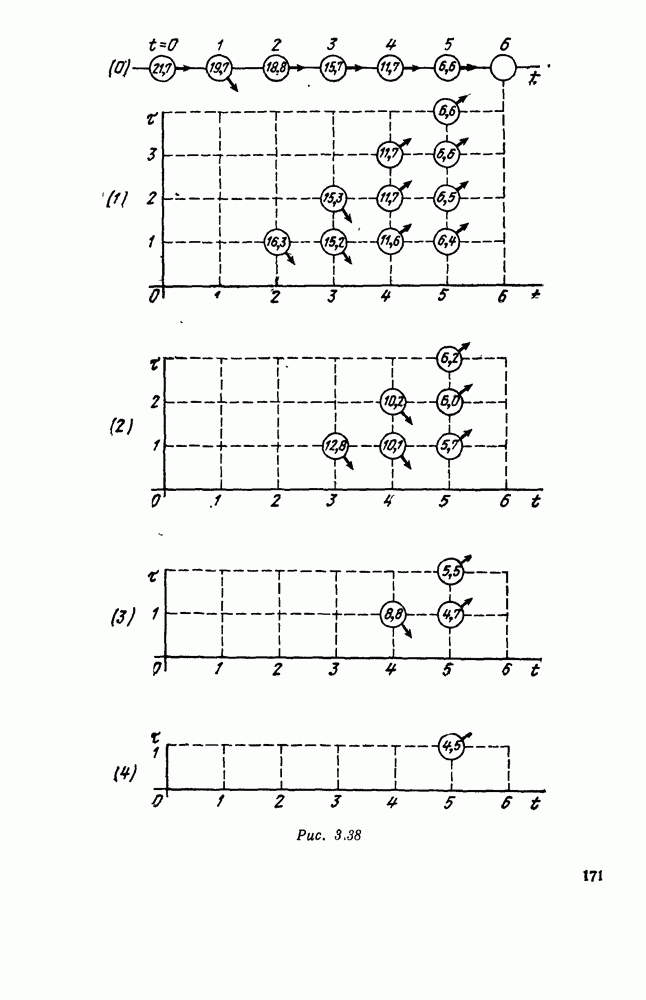
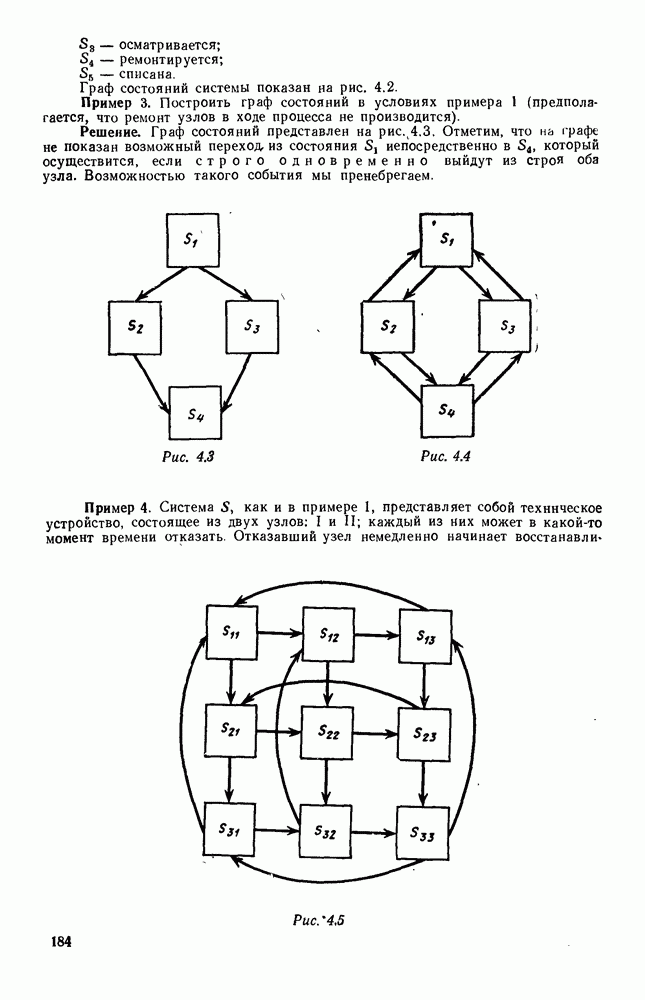
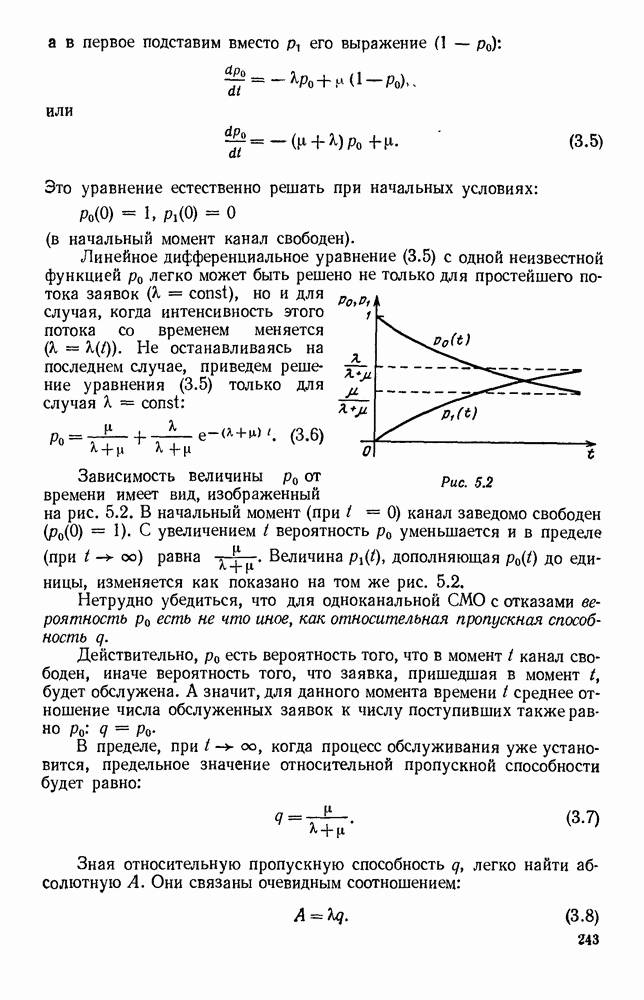
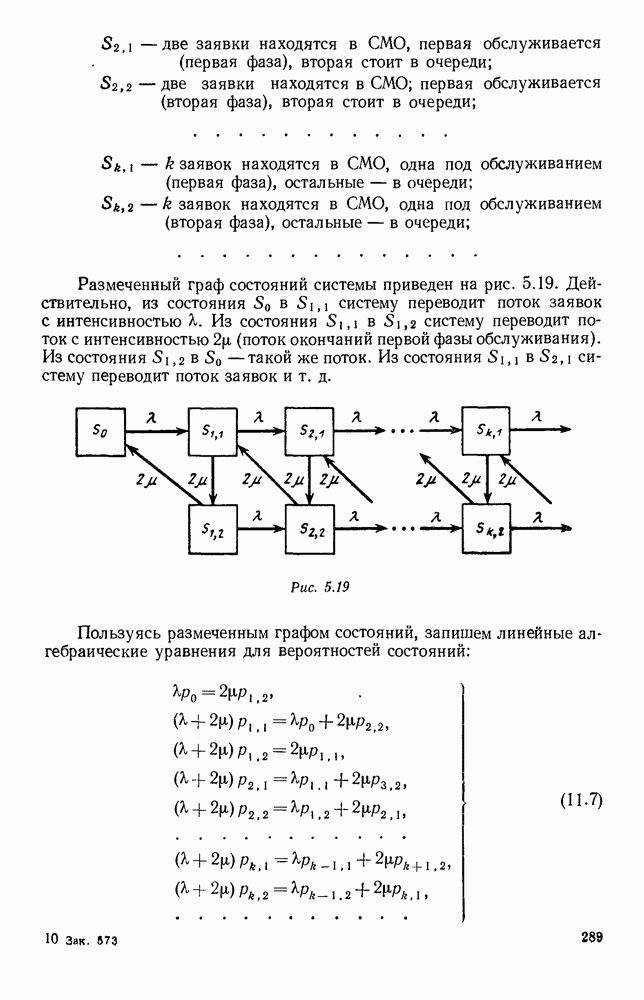
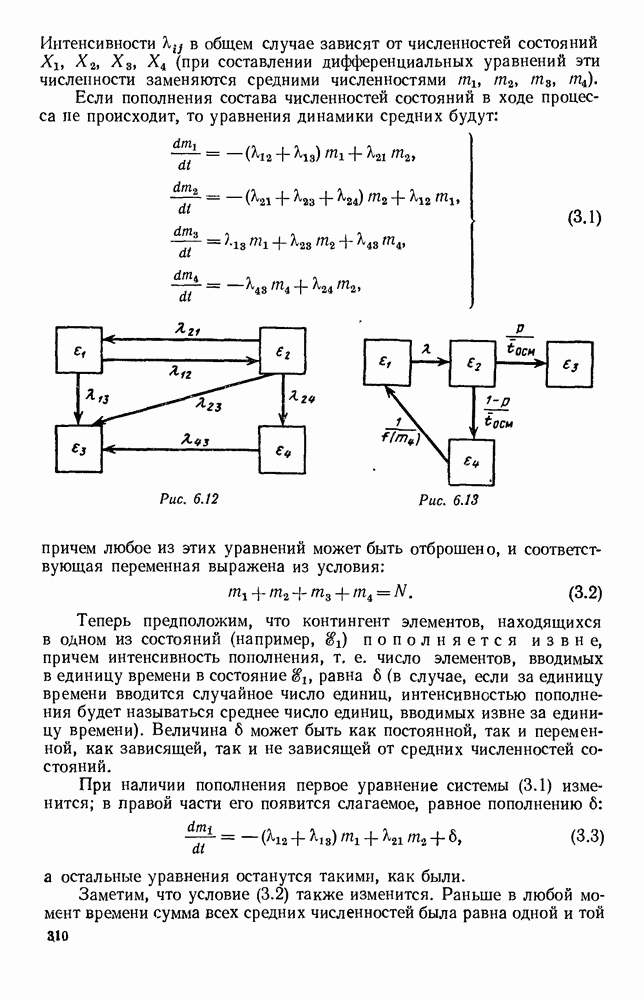
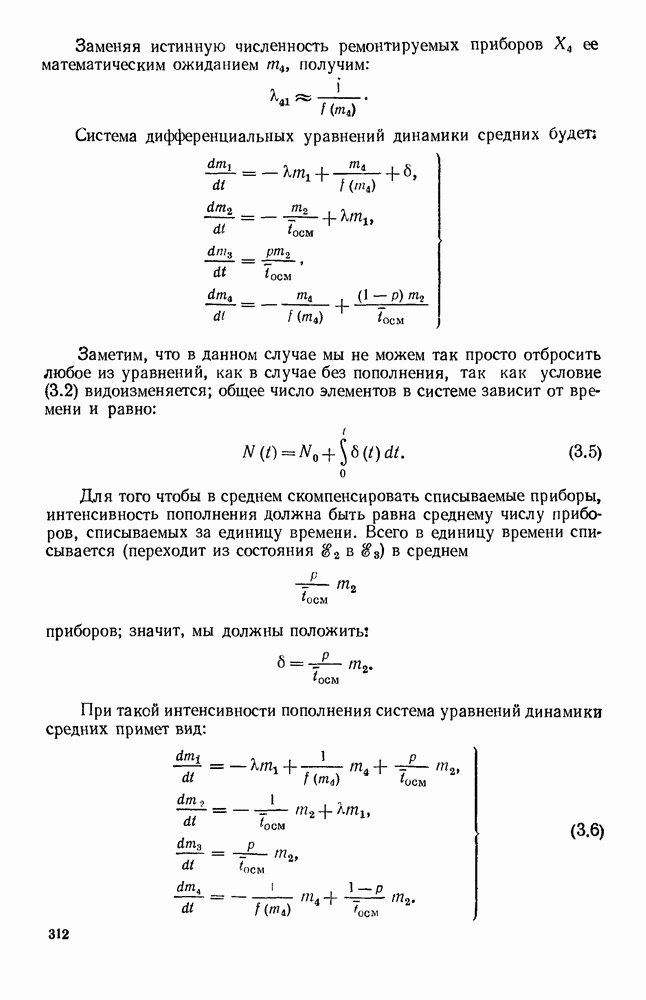
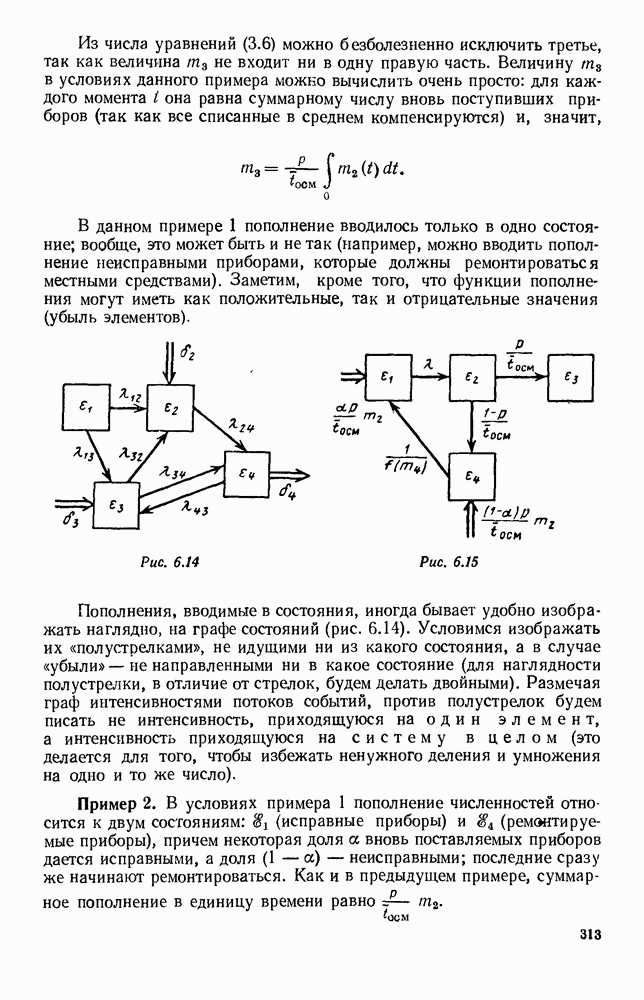
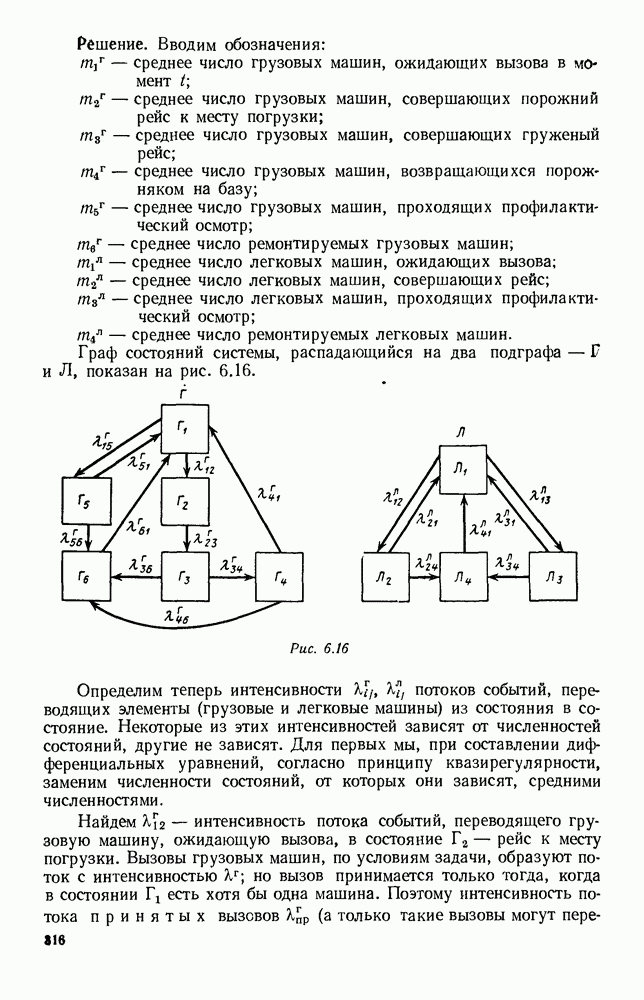
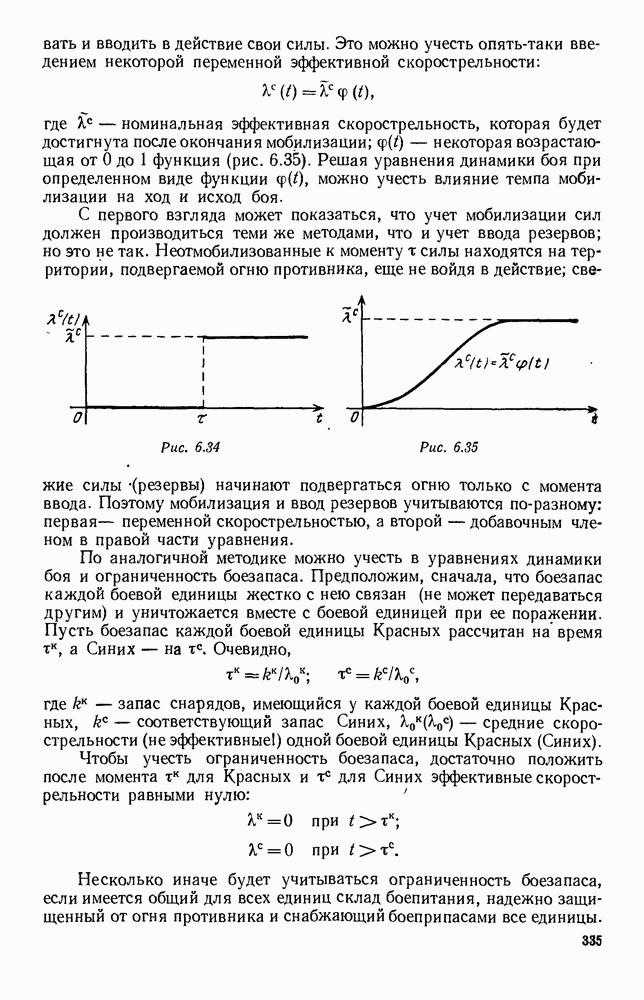
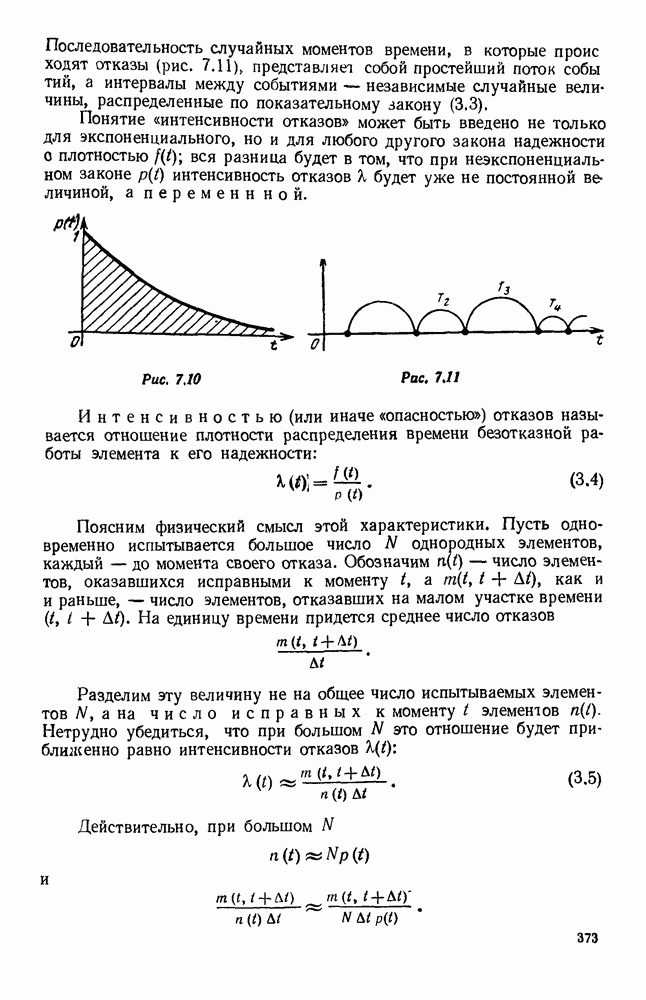
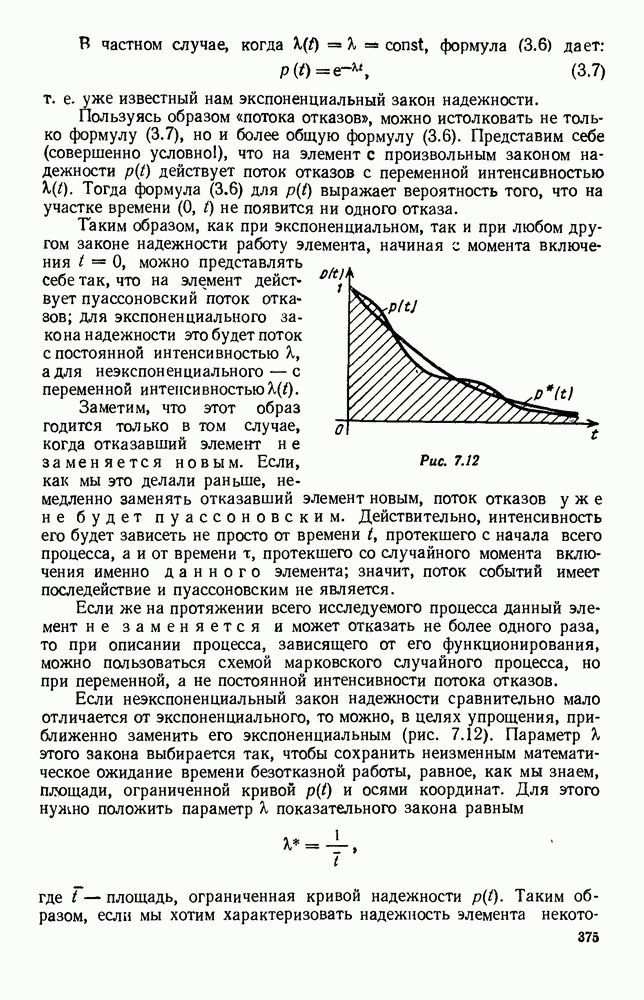
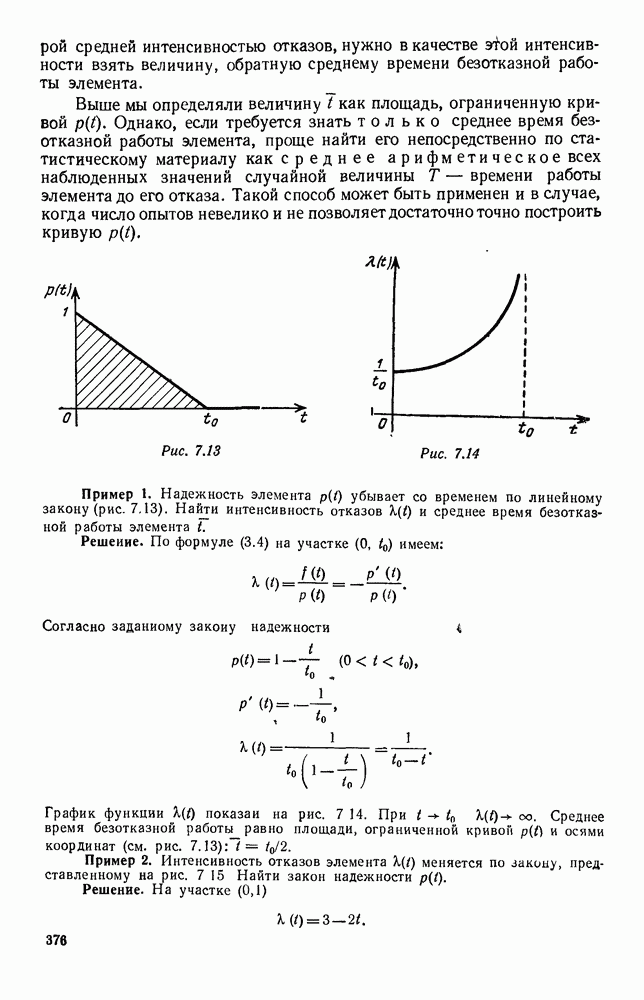
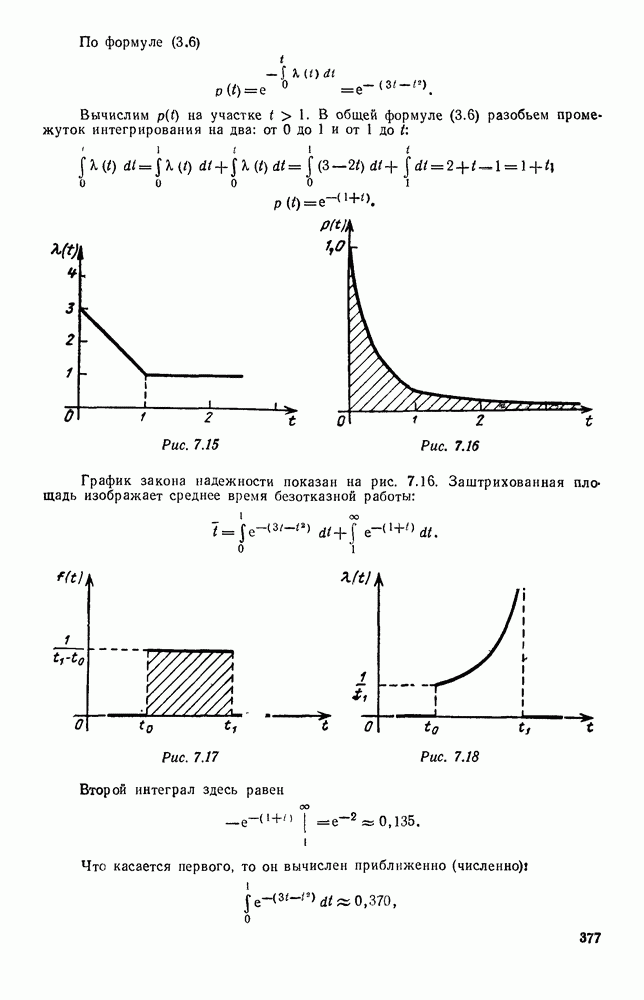
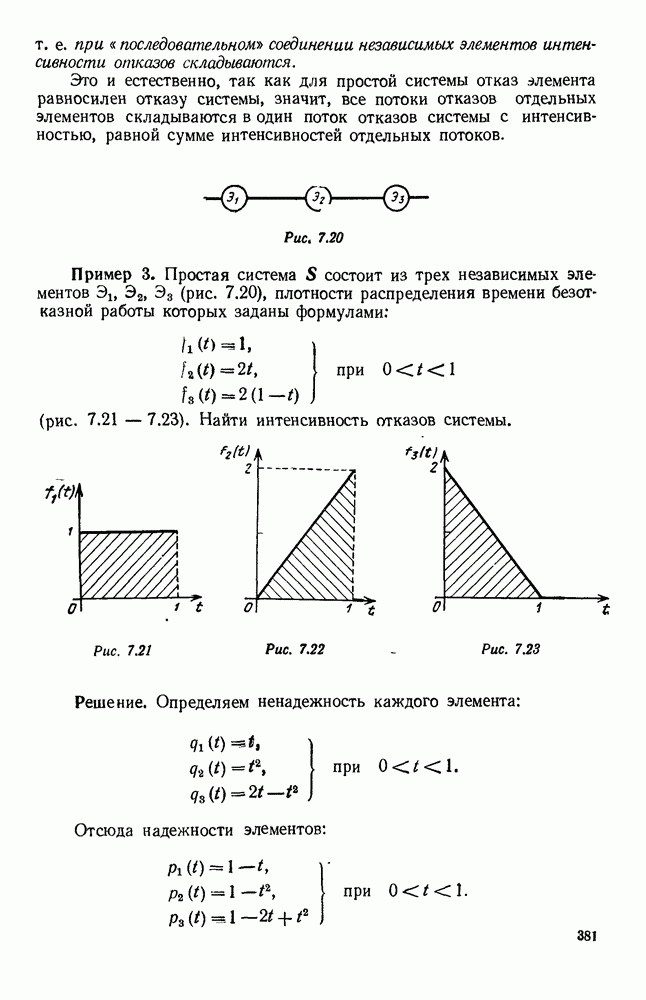
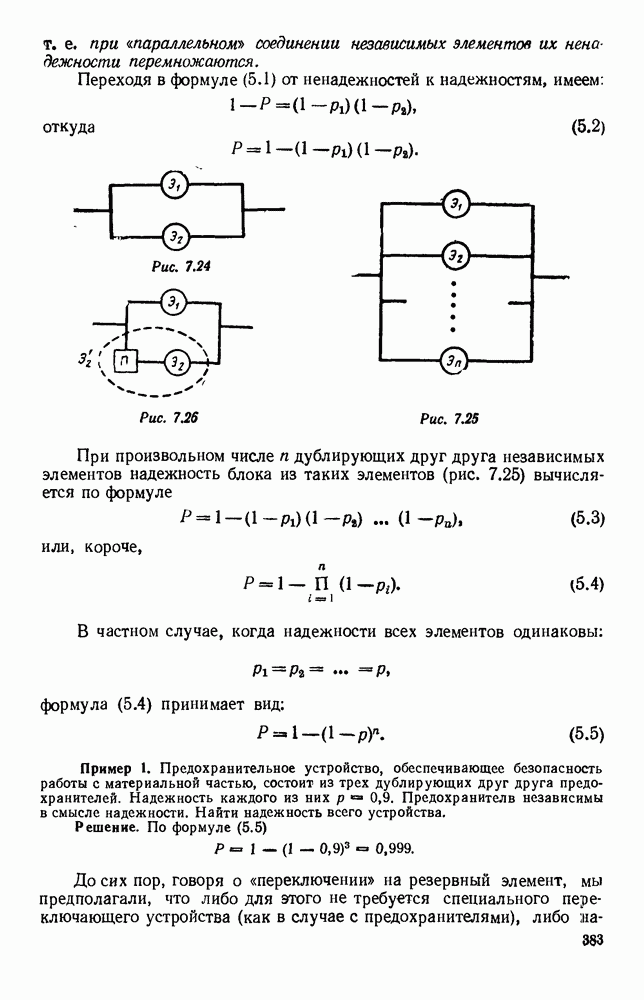
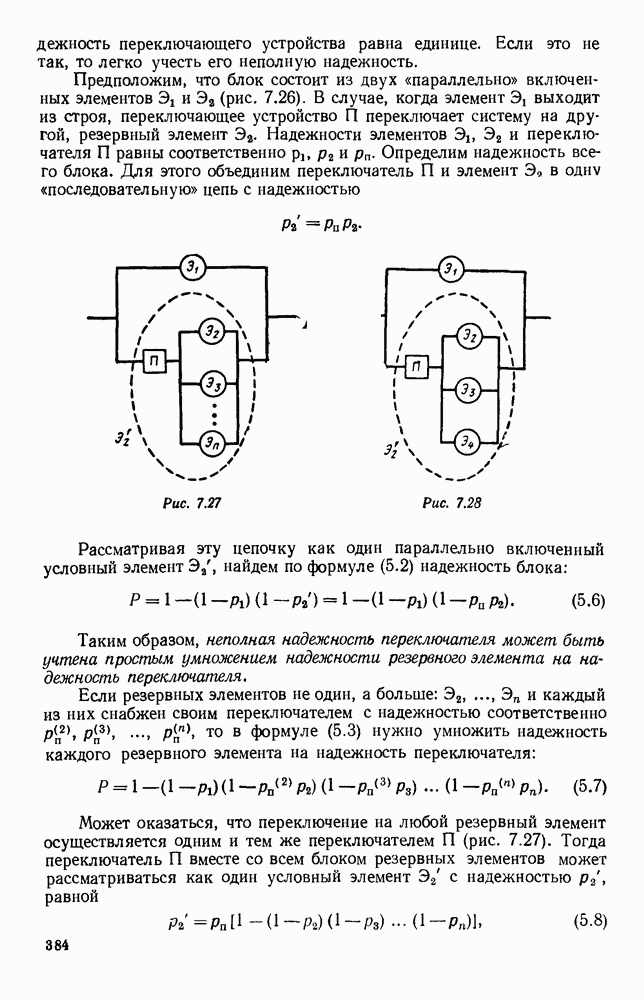
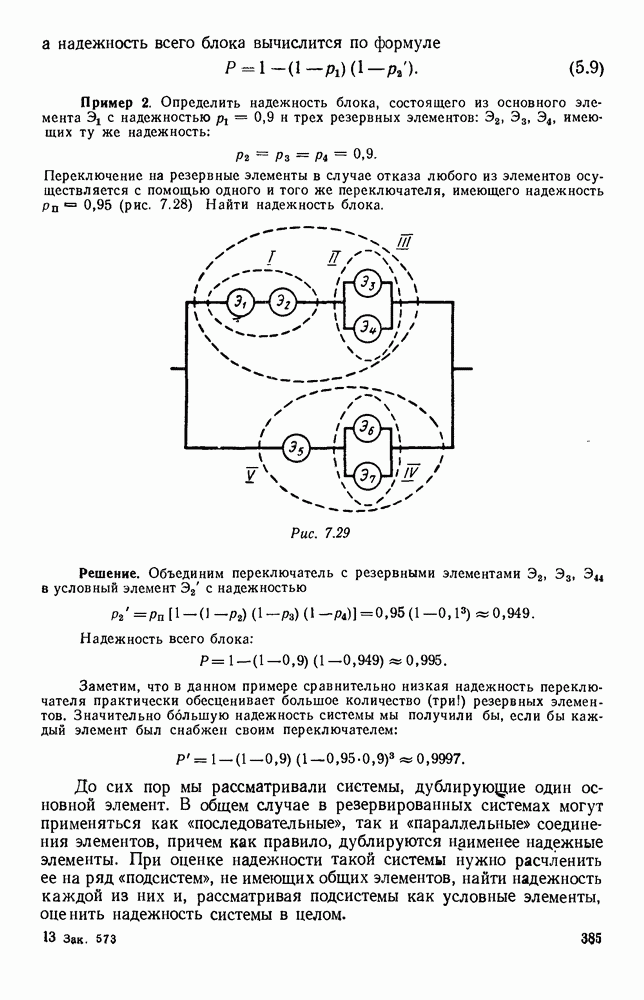
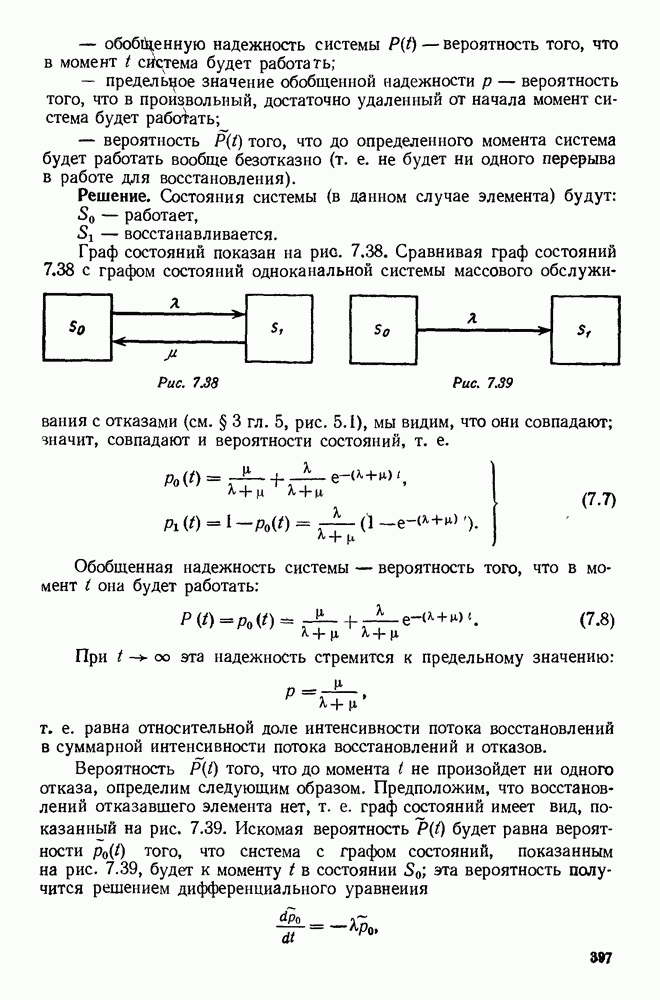
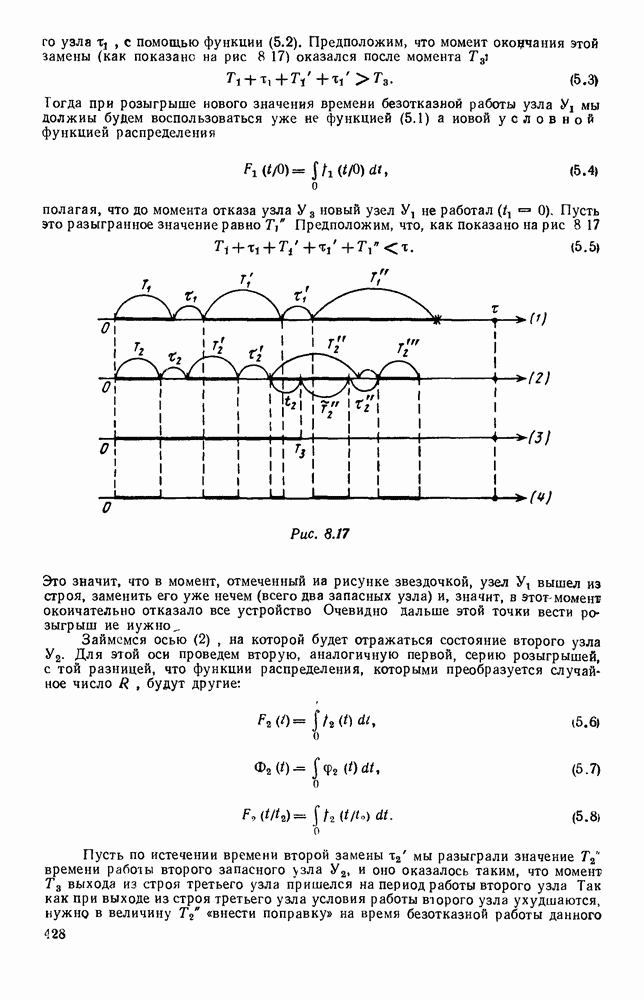
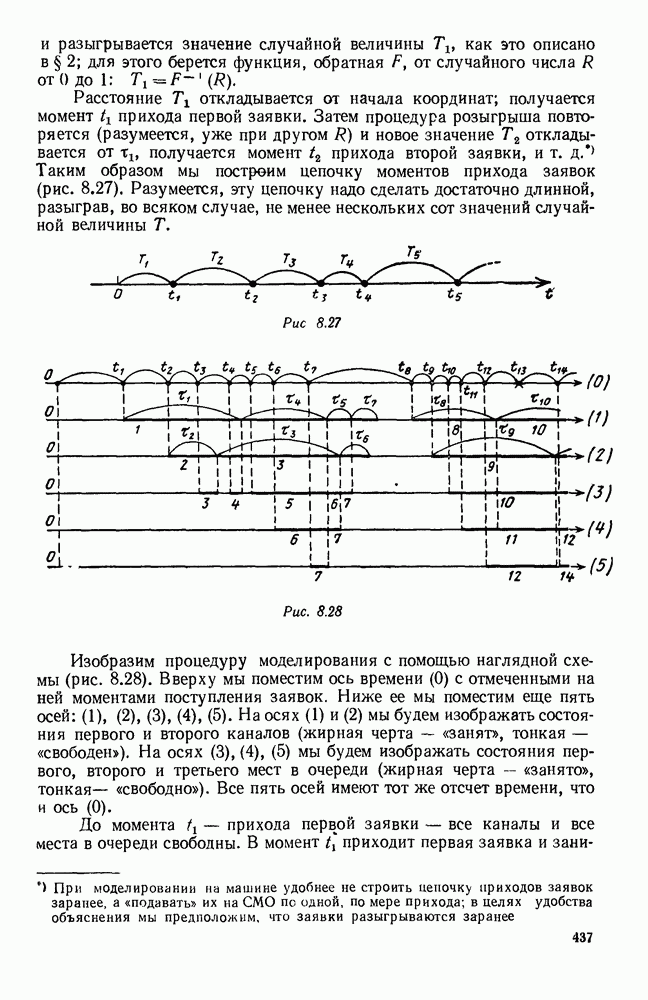
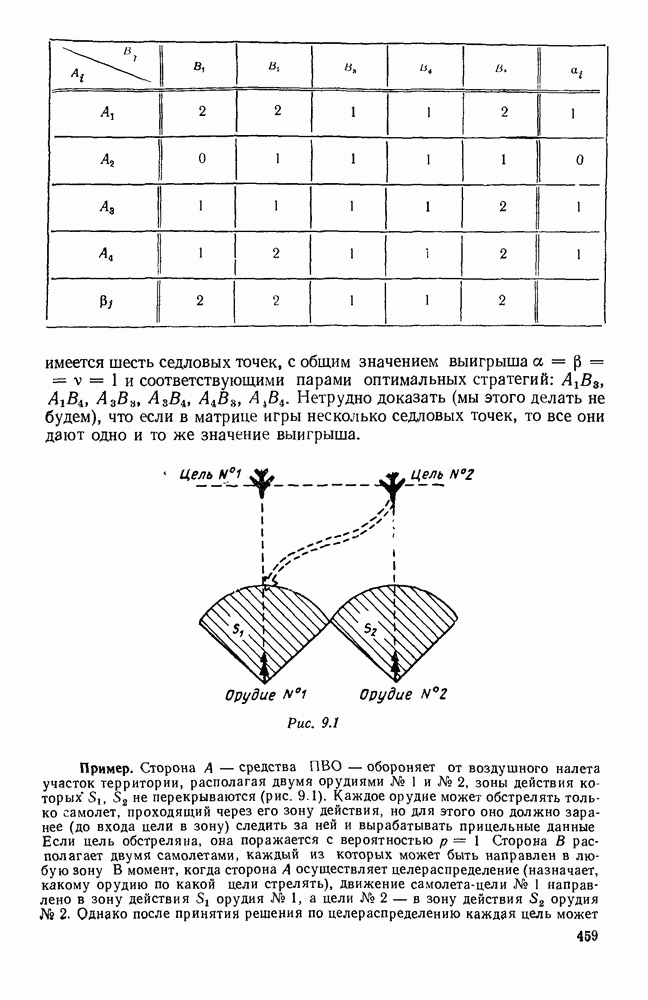
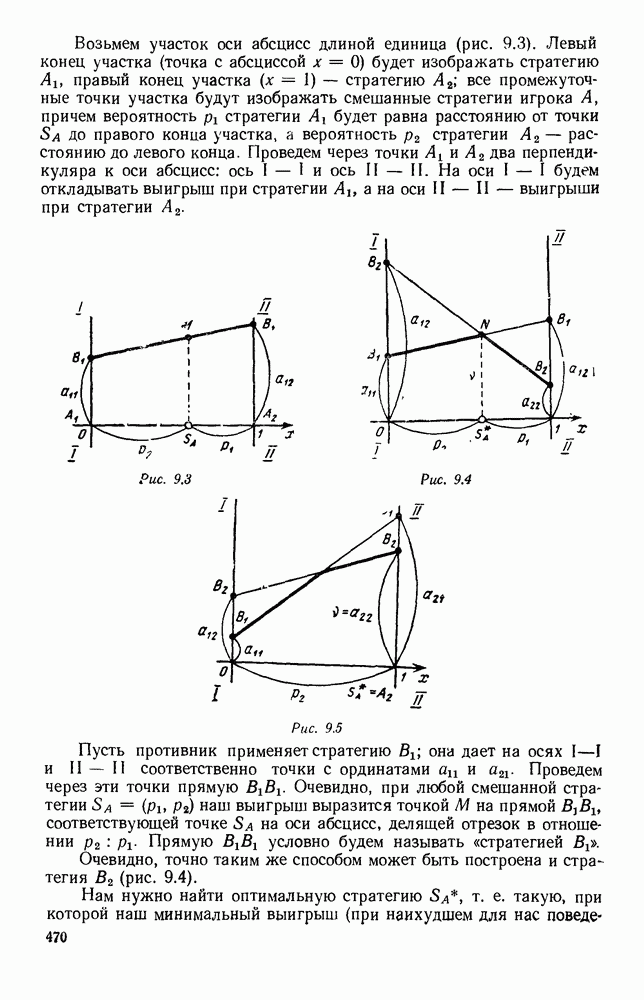
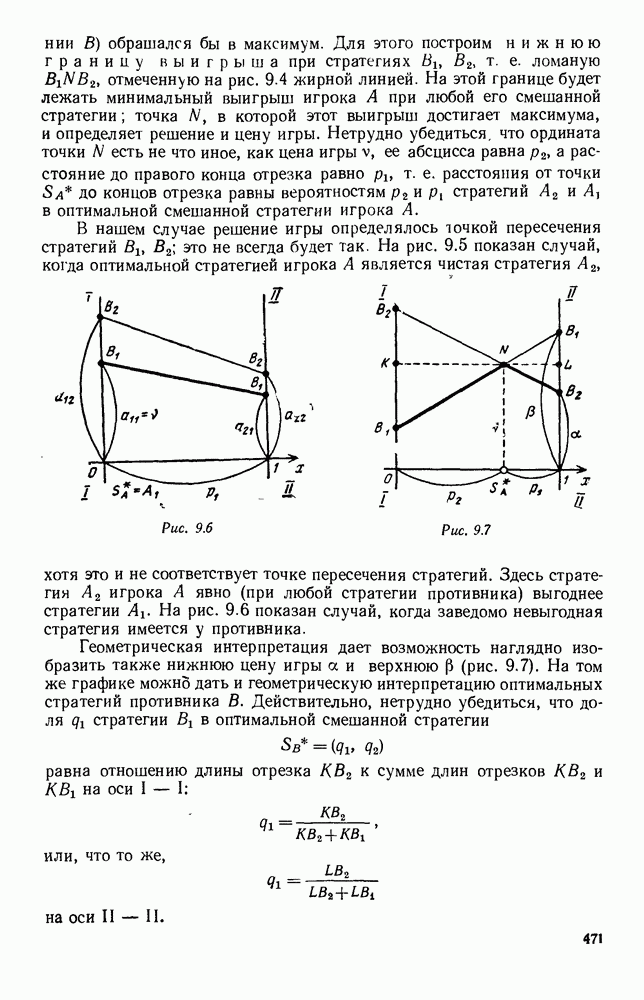
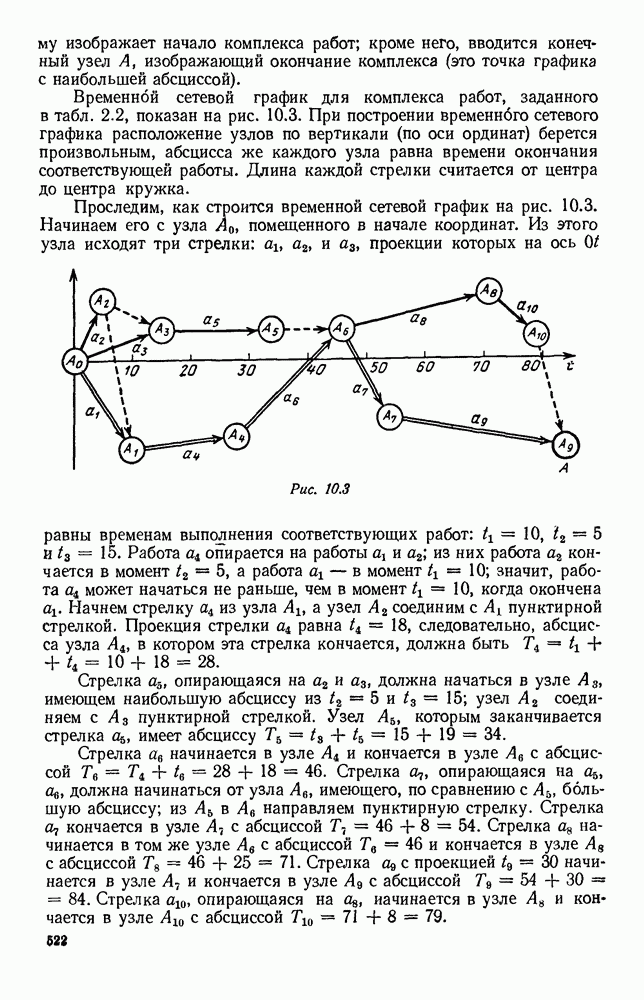
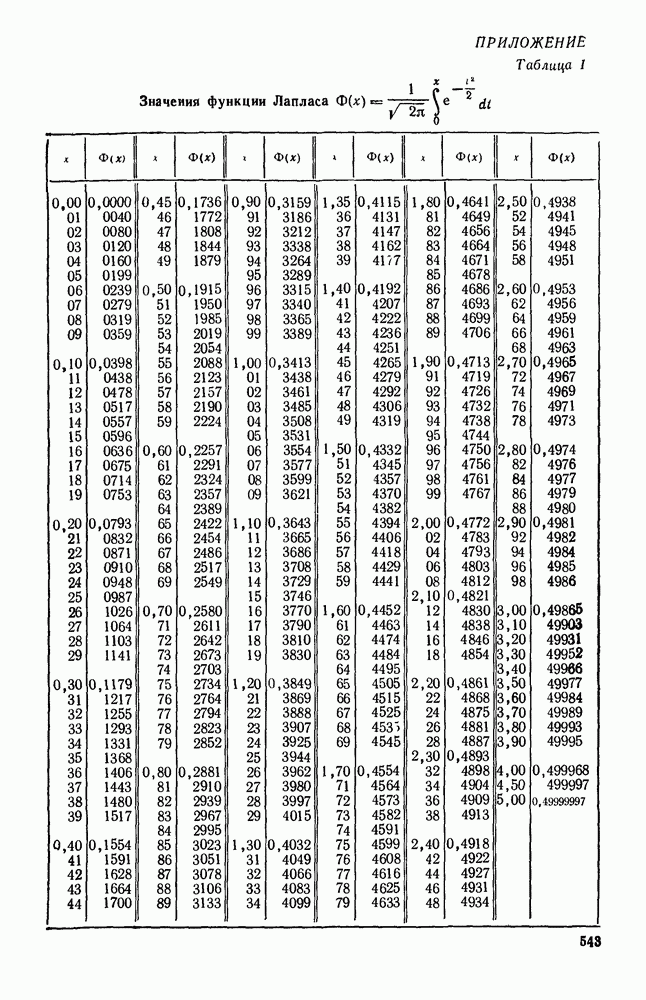
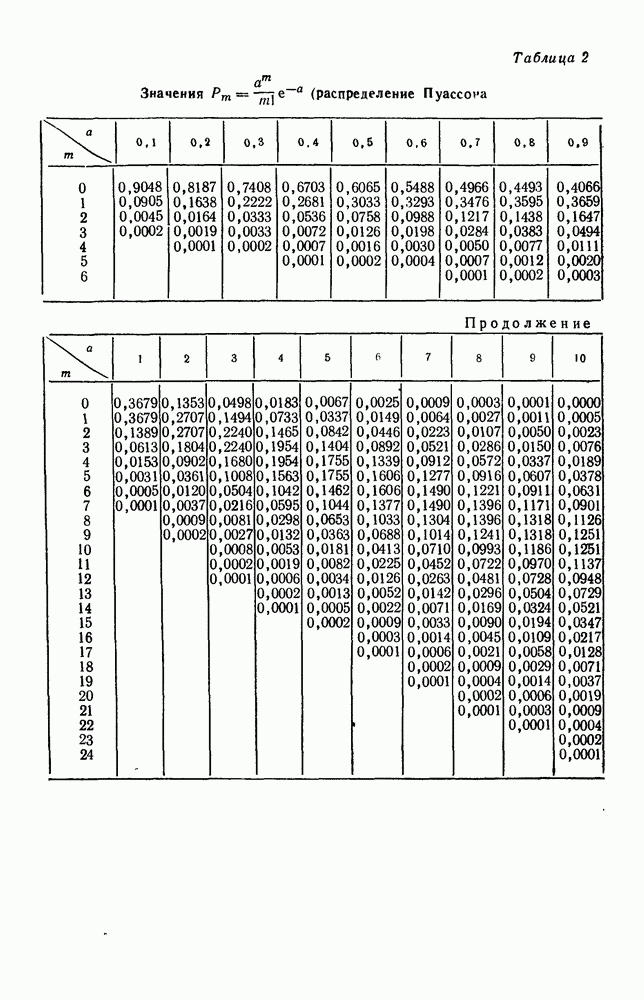
5. ПРЕДЕЛЬНОЕ ПОВЕДЕНИЕ СРЕДНИХ ЧИСЛЕННОСТЕЙ СОСТОЯНИЙВ предыдущих параграфах мы рассмотрели методику описания процесса, протекающего в сложной (многоэлементной) системе S с помощью уравнений динамики средних, в которых неизвестными функциями являются средние численности состояний: Естественно, возникает вопрос: к каким предельным значениям стремятся (если стремятся) эти средние численности при В случае, когда мы рассматривали уравнения для вероятностей состояний, вопрос о предельном режиме решался достаточно просто. Если из любого состояния система могла перейти в любое другое, а число состояний было конечно, то существовал предельный установившийся режим, не зависящий от начальных условий. Чтобы найти вероятности состояний в этом режиме, достаточно было положить левые части дифференциальных уравнений равными нулю и решить полученную систему линейных алгебраических уравнений. Для метода динамики средних дело обстоит несколько сложнее. Напомним, что уравнения динамики средних в общем случае нелинейны; так же нелинейны и алгебраические уравнения, получаемые из них, если левые части положить равными нулю. Может оказаться, что решение этой системы уравнений не единственно, тогда нужно рассмотреть совокупность решений и отбросить те из них, которые не отвечают физическим условиям задачи. Если даже решение единственно, нужно все же исследовать поведение решения системы дифференциальных уравнений при
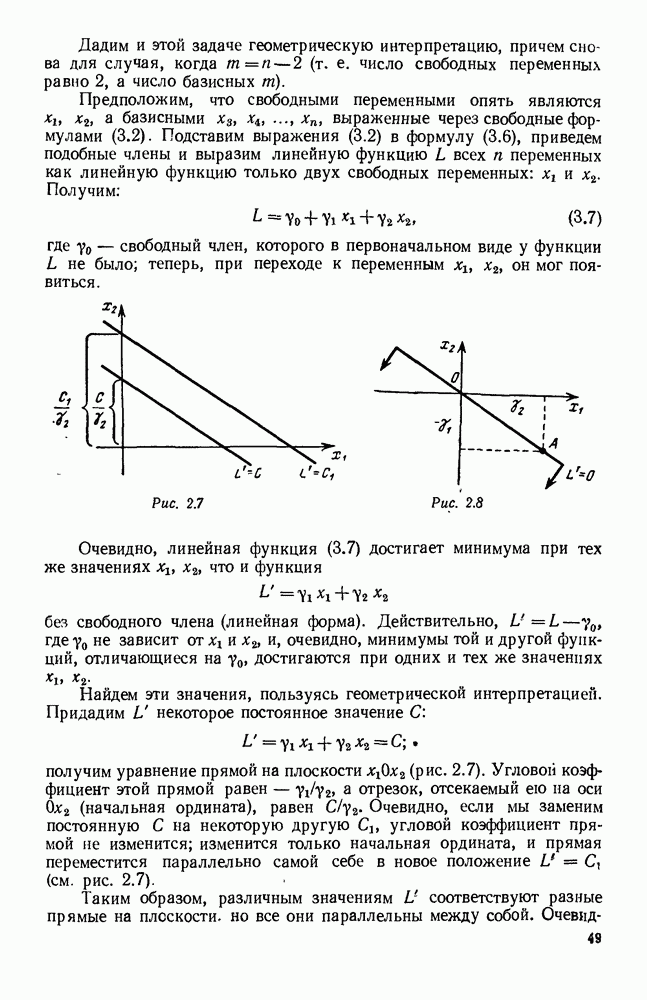
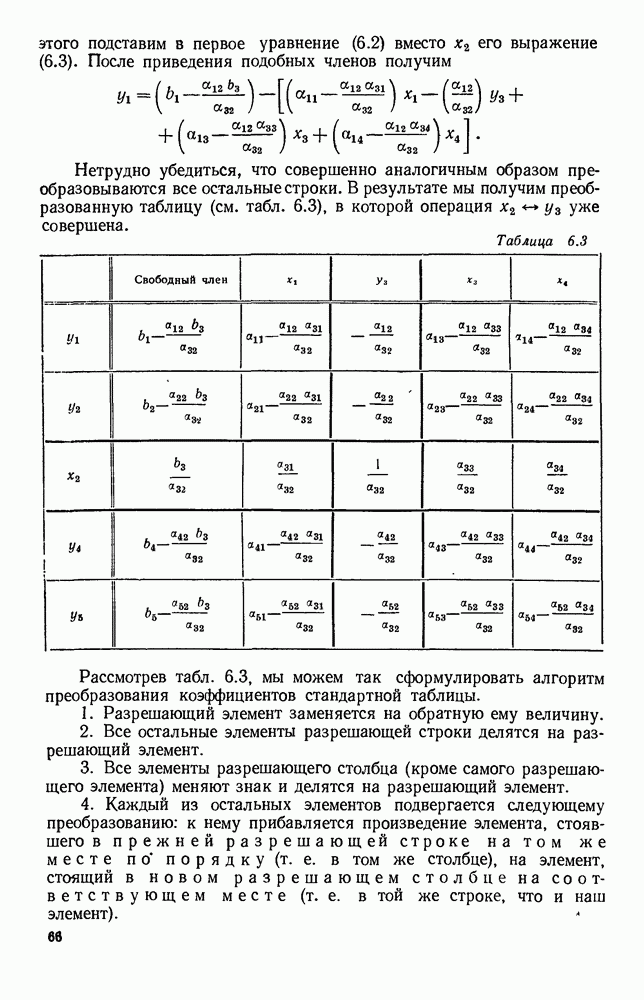
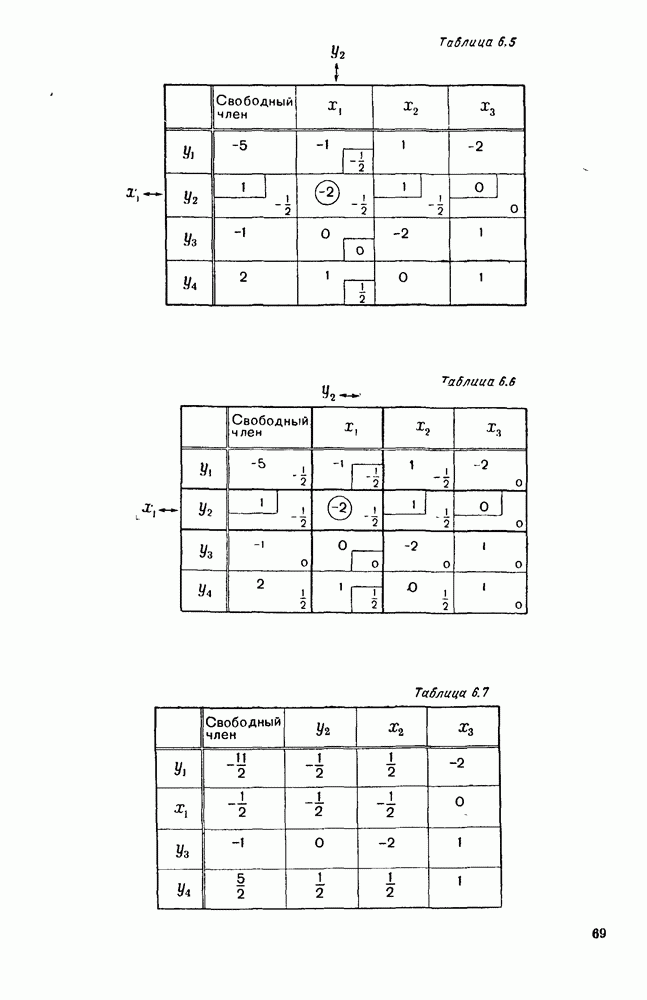
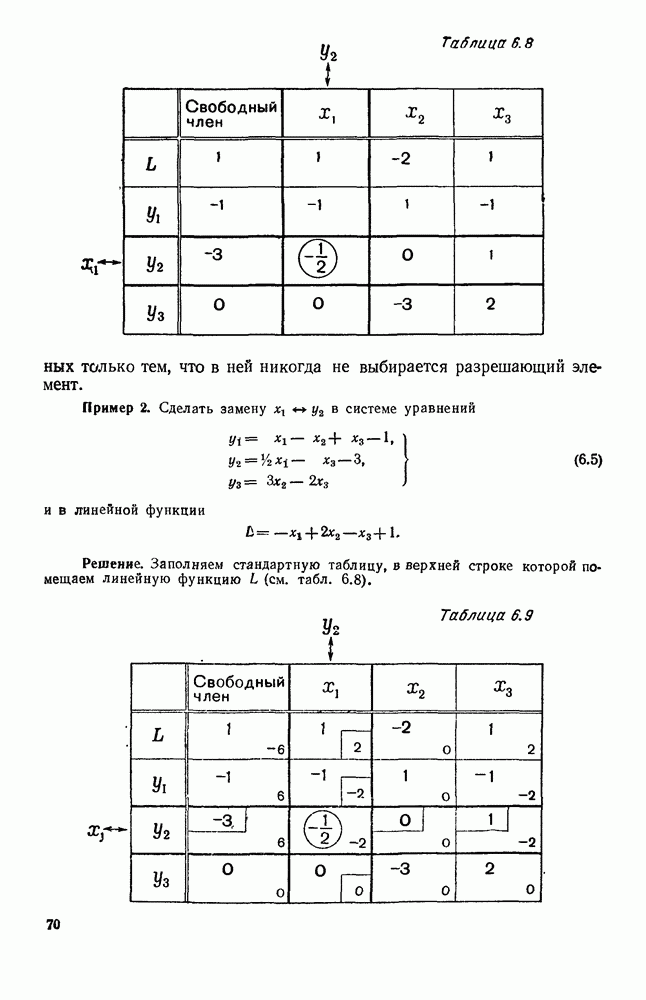
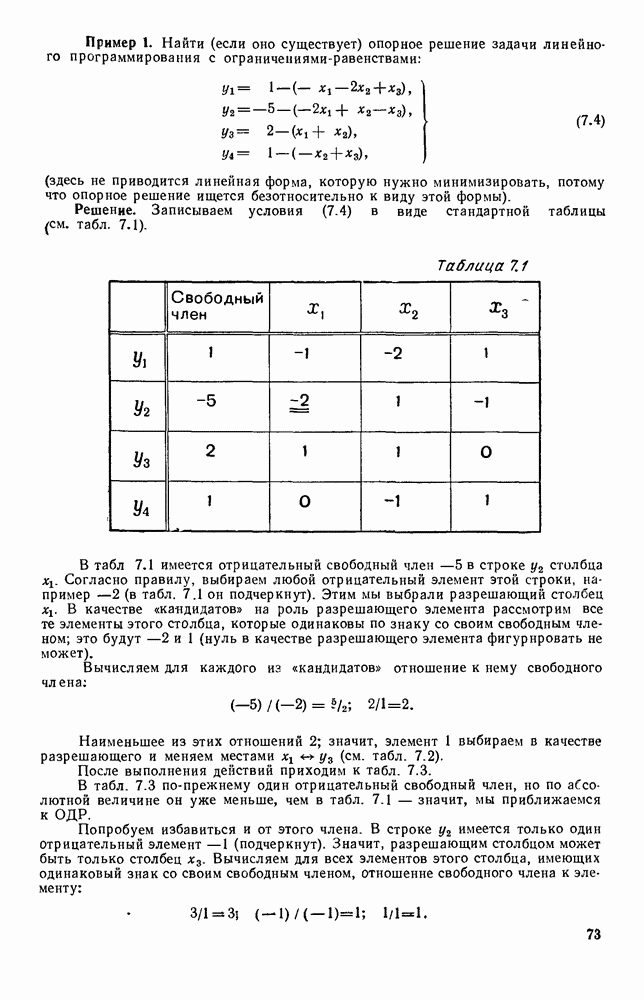
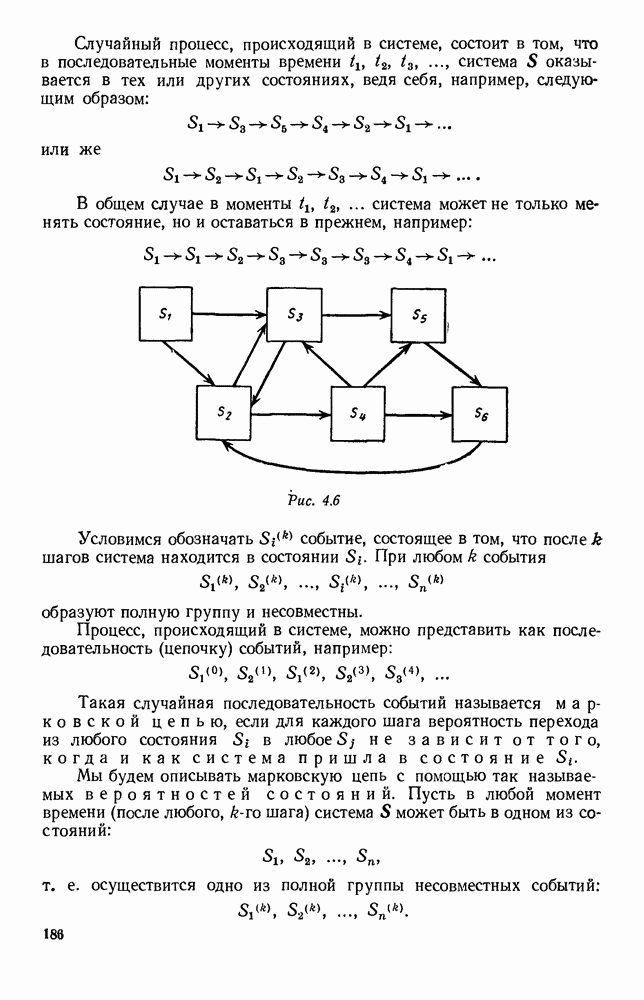
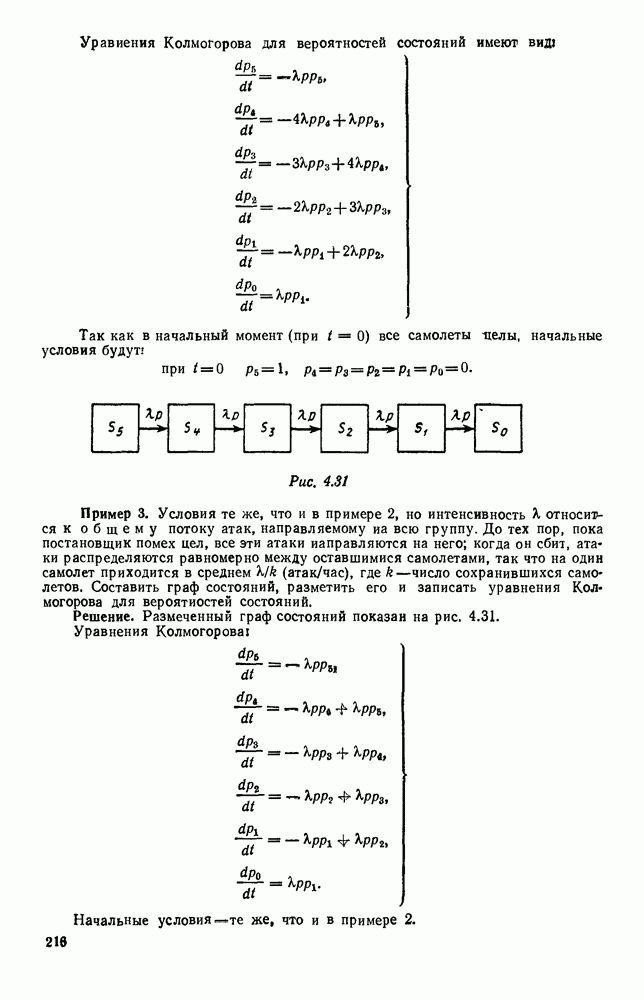
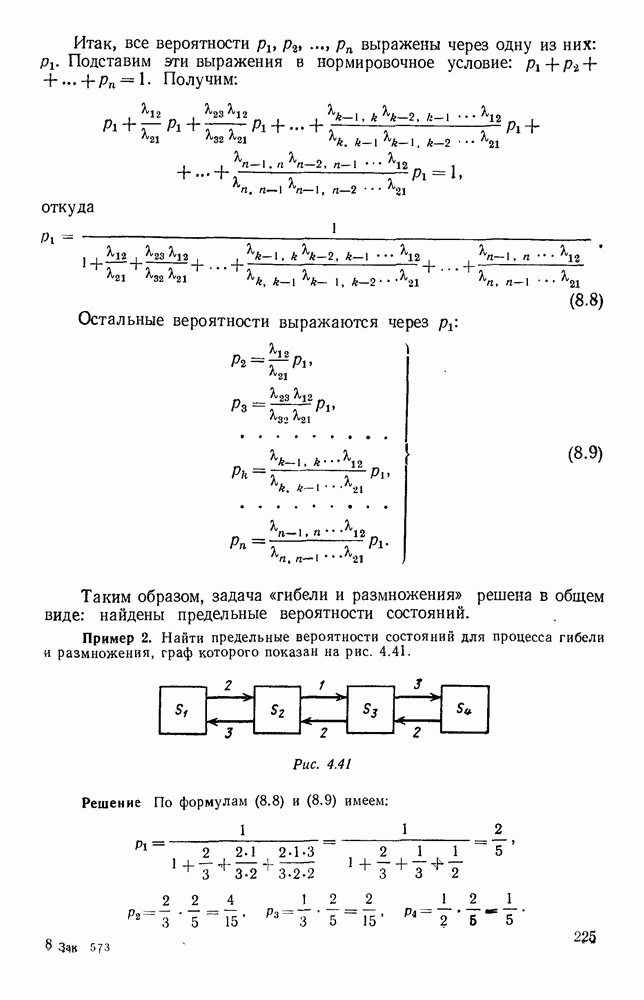
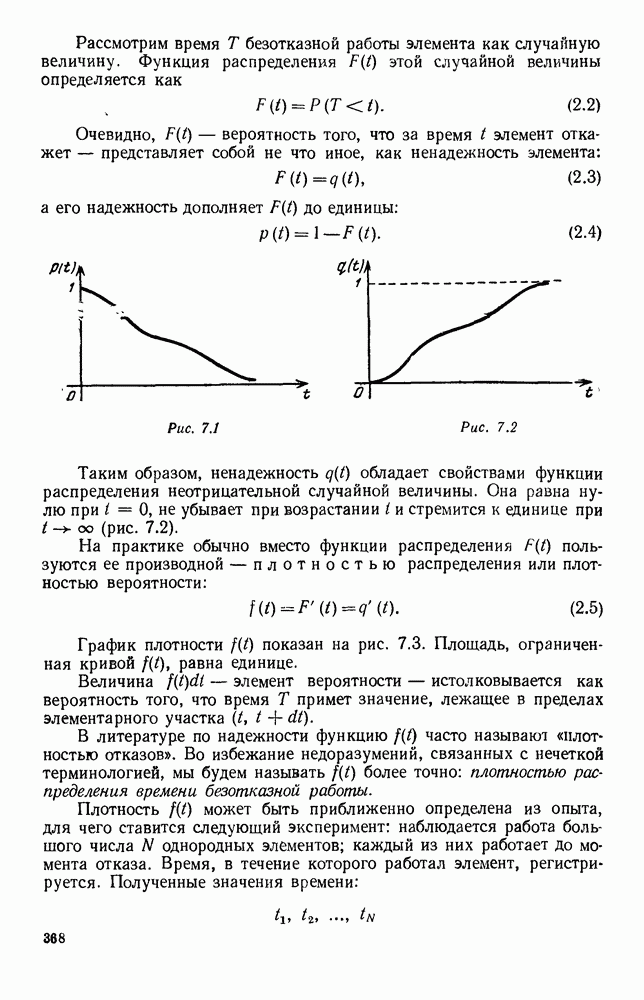
Рис. 6.19 Продемонстрируем особенности исследования предельного поведения средних численностей состояний на примере задачи, взятой, ради разнообразия, из области биологии. Пусть в некоторой местности обитают животные двух видов А и В, причем животные первого вида (А) — хищные, и они питаются животными второго вида (В), которые довольствуются растительной пищей. Будем характеризовать состояние каждого животного предельно грубо, учитывая только, живо ли оно еще или погибло. Построение графа состояний элементов системы не вызывает никаких трудностей: этот граф разобьется на два подграфа, соответствующих видам А я В (рис. 6.19). Здесь стрелки, ведущие из Обозначим численности элементов (животных) в состояниях Для того, чтобы составить дифференциальные уравнения, нужно задаться видом зависимости интенсивностей потоков Начнем с травоядных животных вида В. Предположим, что запасы пищи, доступной им, не зависят от численности животных ни вида А, ни вида В, и что пищи хватает на всех. Тогда естественно предположить, что средняя рождаемость в единицу времени (в пересчете на одну живую особь вида В) остается постоянной. Обозначим эту постоянную с; тогда прирост численности состояния
Что касается смертности
где I — константа. В пересчете на одну живую особь вида А число таких смертей («поеданий») в единицу времени будет равно
Теперь займемся животными вида А. Мы предположили, что их единственную пищу составляет вид В; поэтому естественно, что и рождаемость и смертность вида А будут зависеть от числа поедаемых в единицу времени животных, приходящихся на одного хищника. Число поедаемых животных задается формулой (5.2), и на одного хищника их приходится в среднем
Очевидно, функция Аналогично, рождаемость будет какой-то другой функцией от числа животных вида
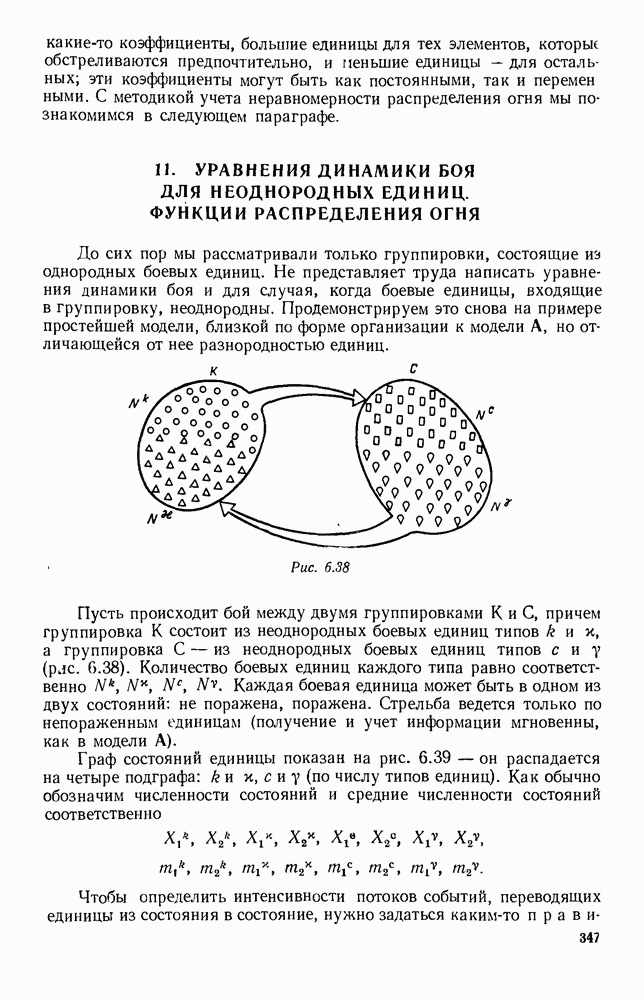
Функция естественно, возрастающая функция от Хрис. 6.21). Выпишем теперь дифференциальные уравнения для средних численностей состояний:
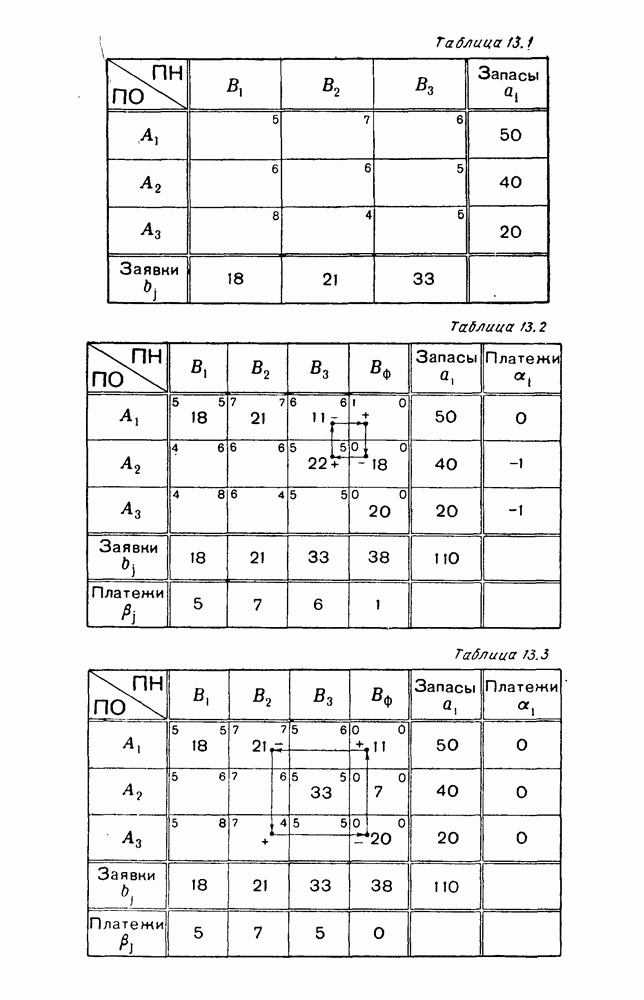
Пользуясь принципом квазирегулярности и заменяя численности
Рис. 6.20
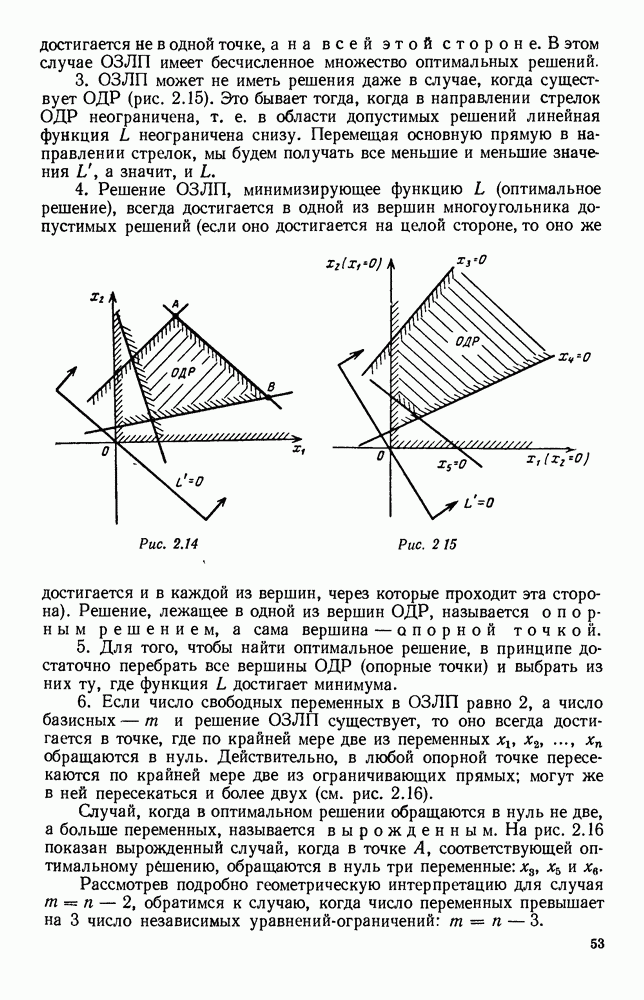
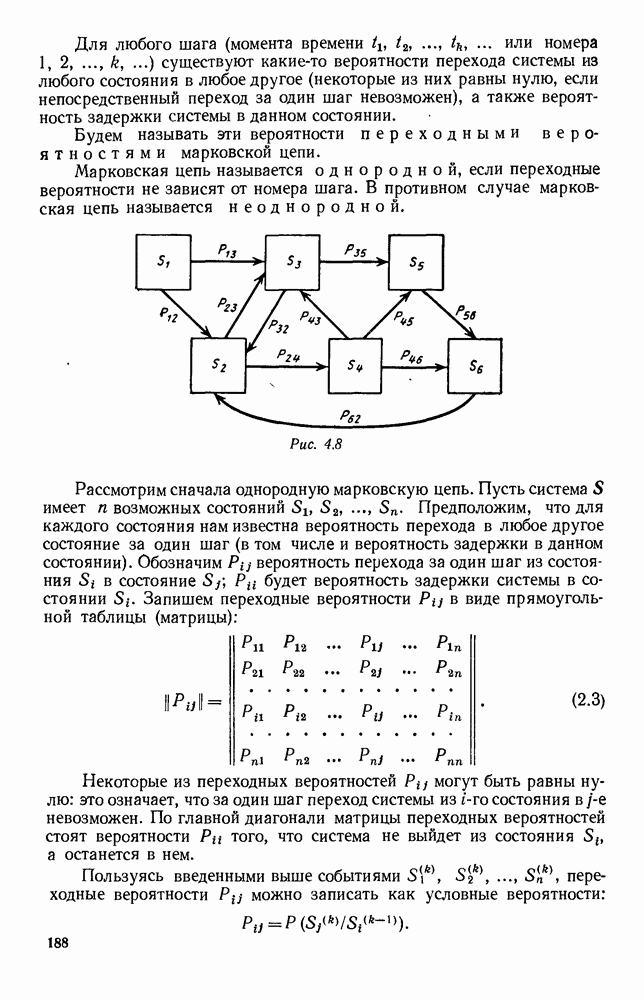
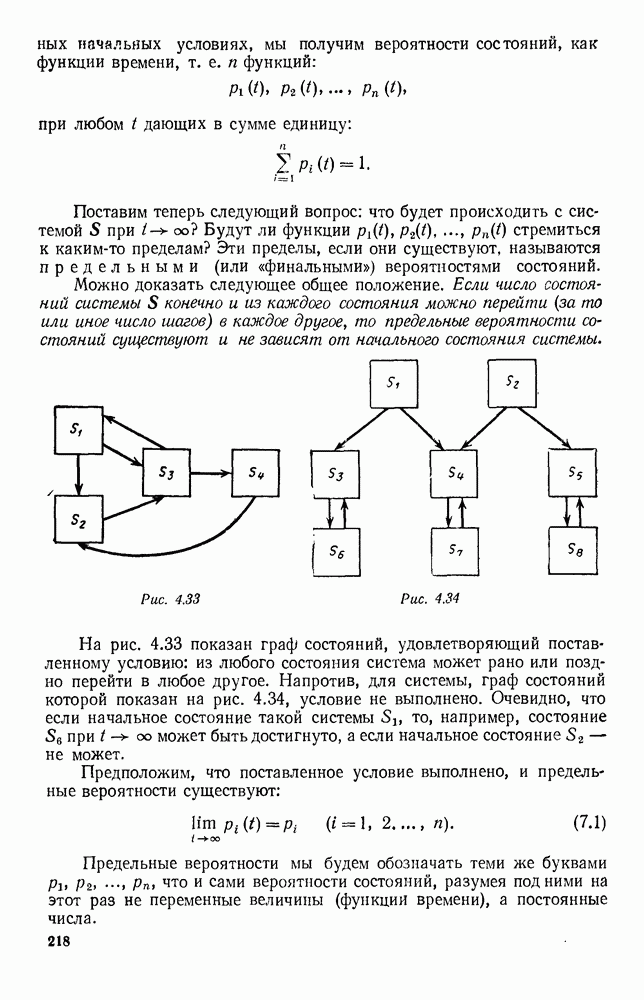
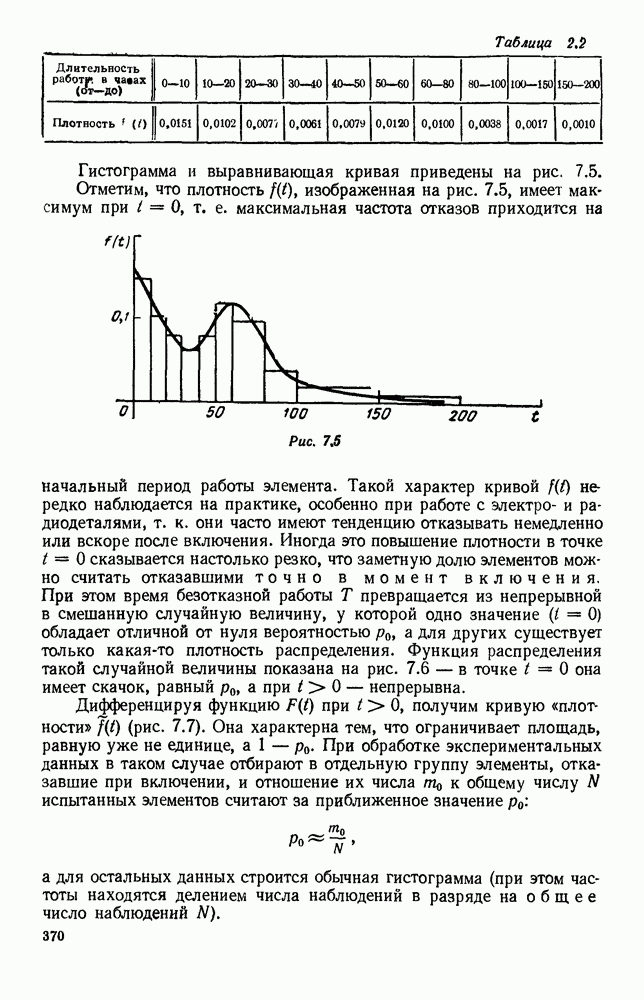
Рис. 6.21 Мы видим, что в первое уравнение фактически входит только разность функций g и
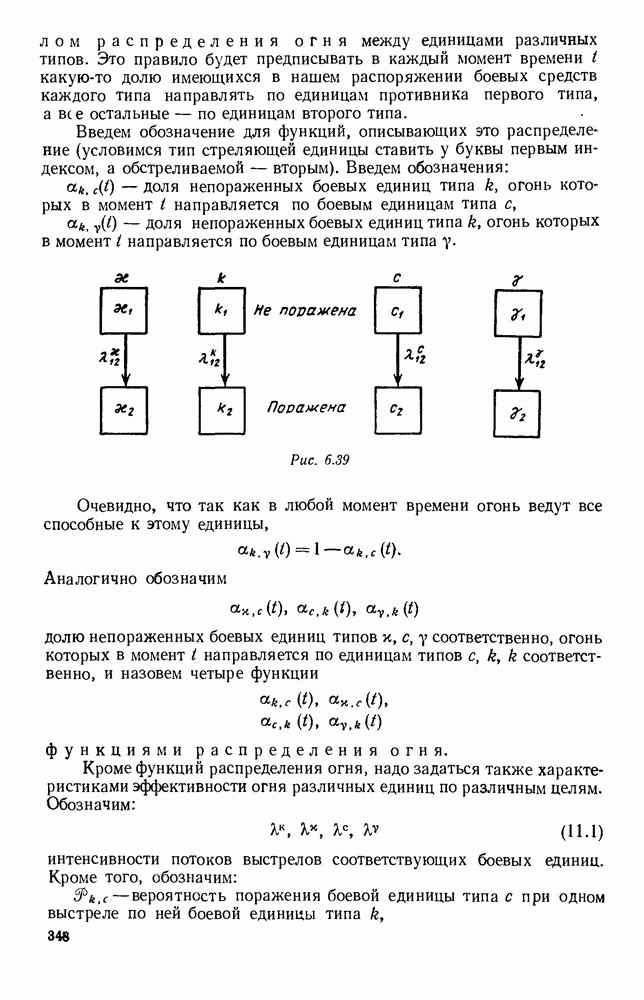
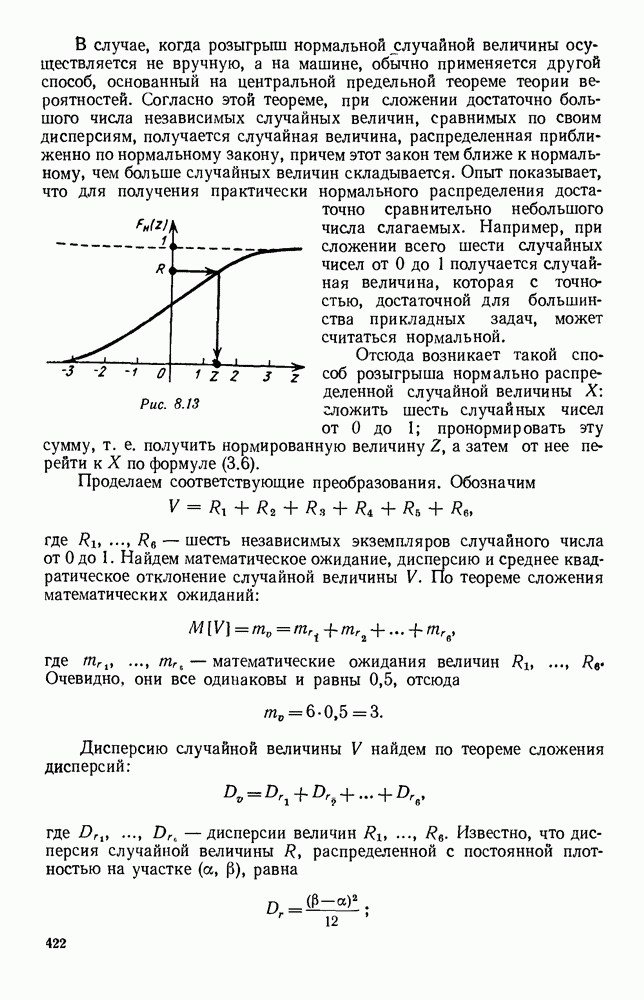
Это — разность возрастающей и убывающей функции, а значит, сама функция h — возрастающая (см. рис. 6.22). В отличие от положительных функций Совершенно так же можно ввести критическую численность
Когда число хищников больше, чем Пользуясь новыми обозначениями, уравнения динамики средних (5.6) можно переписать в виде:
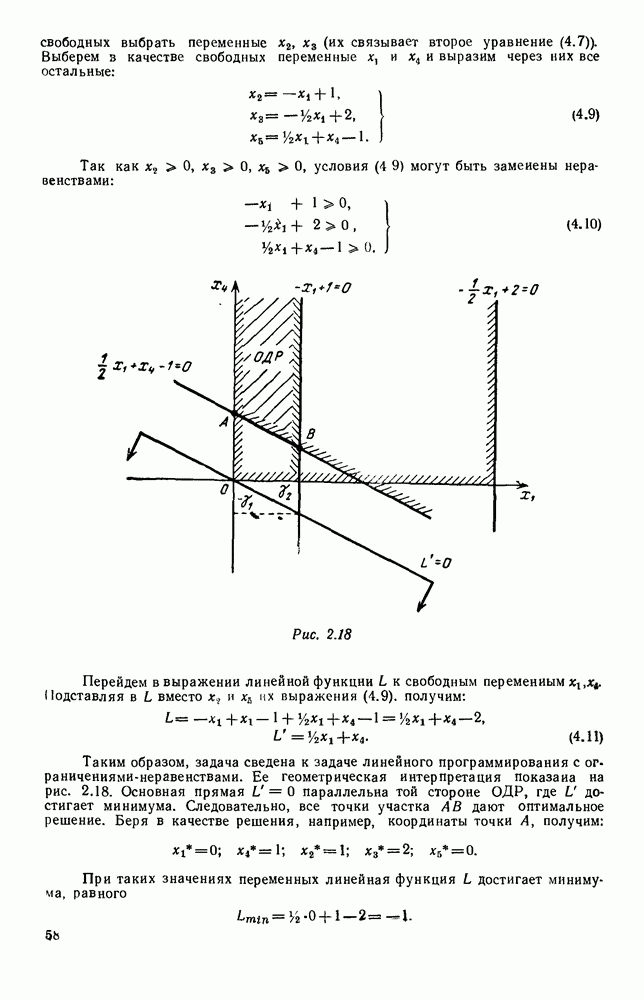
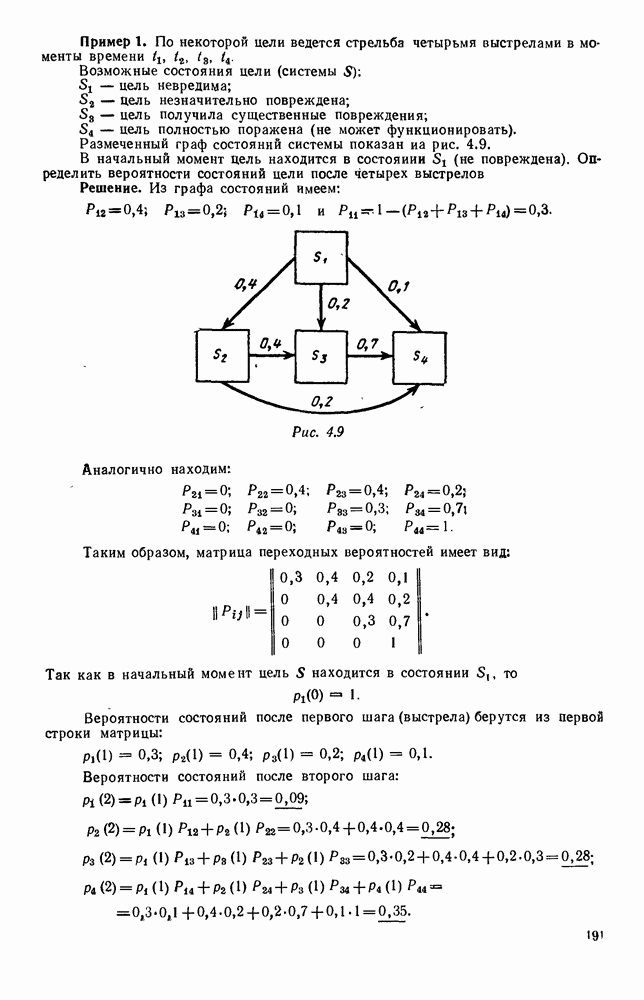
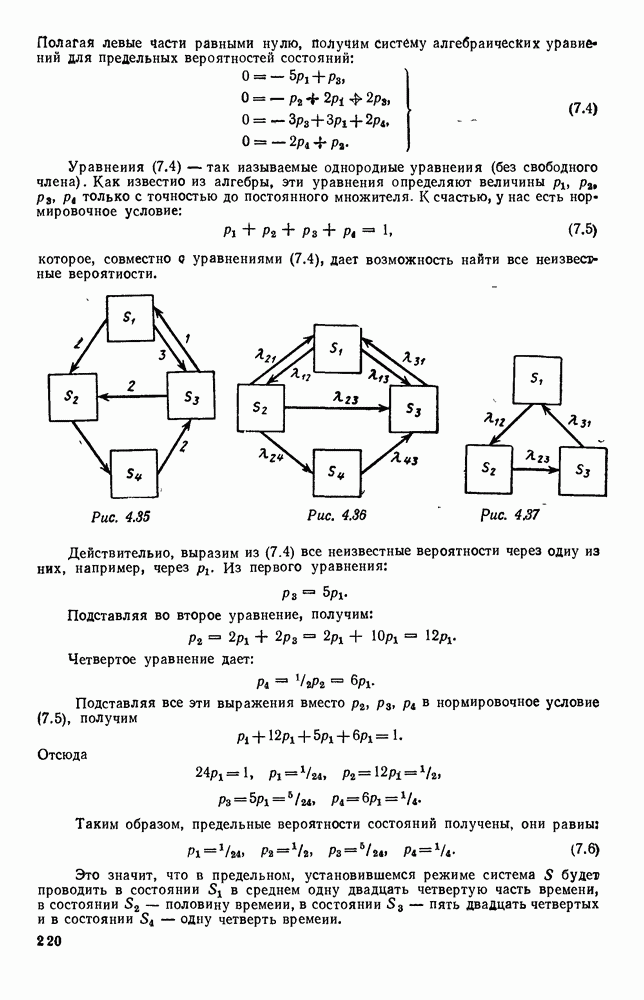
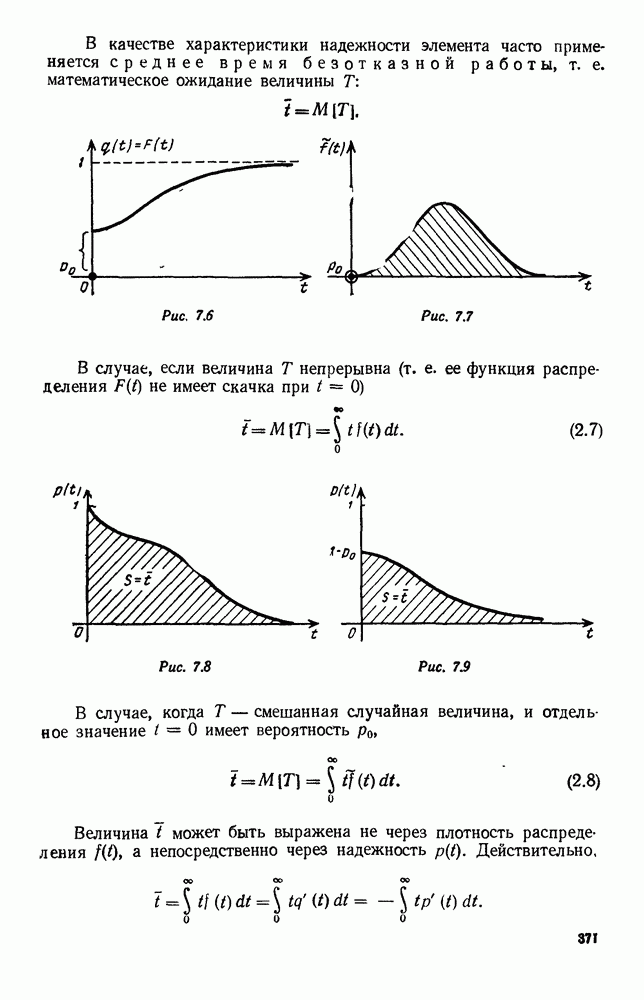
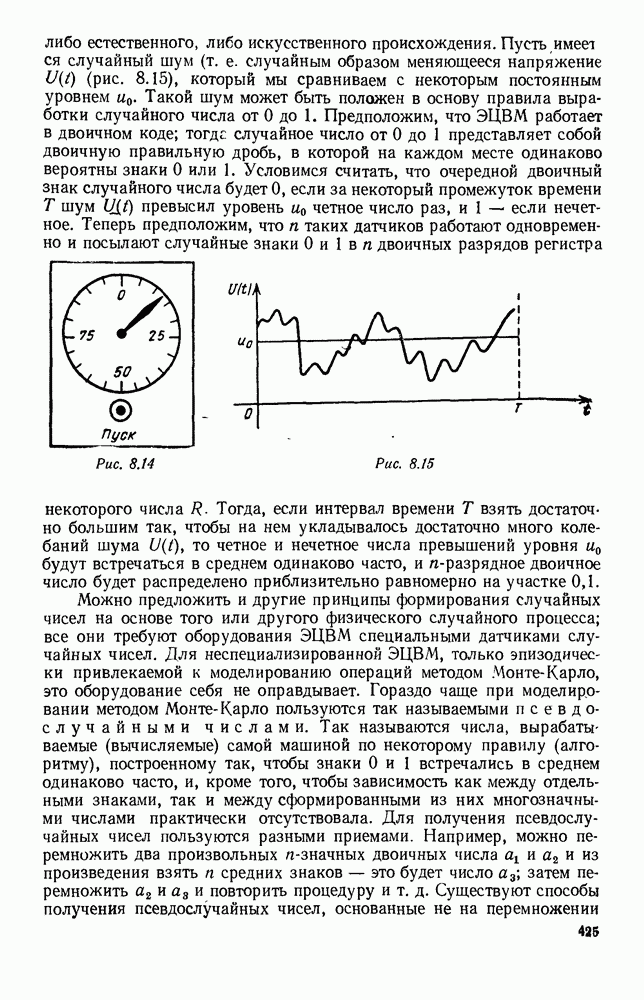
В таком виде мы и будем анализировать уравнения для средних численностей состояний системы. Посмотрим, как с изменением t будет двигаться точка на плоскости, изображающая решение
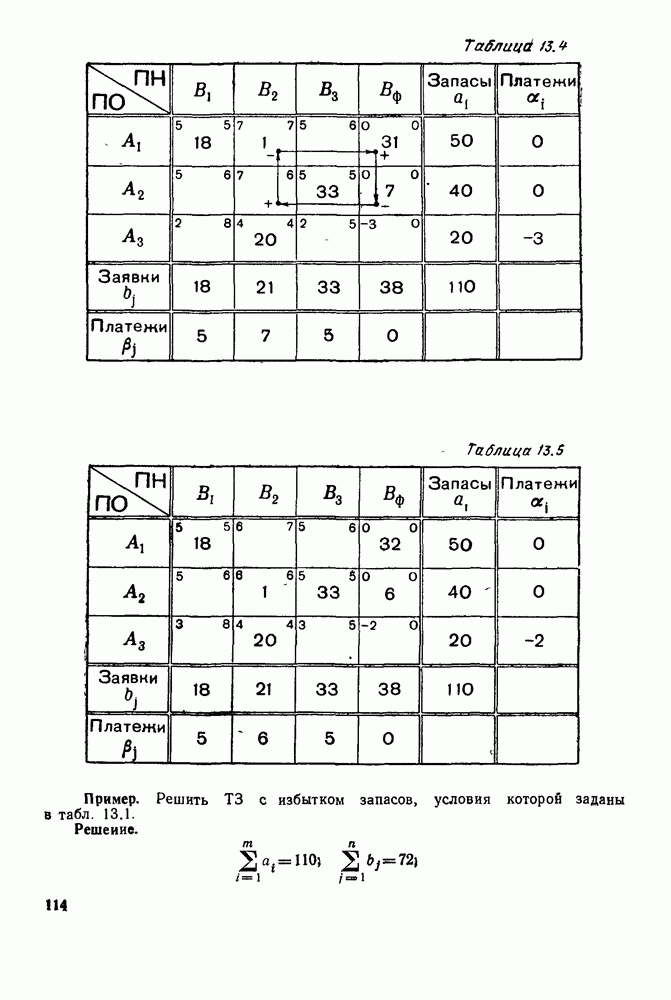
Рис. 6.22
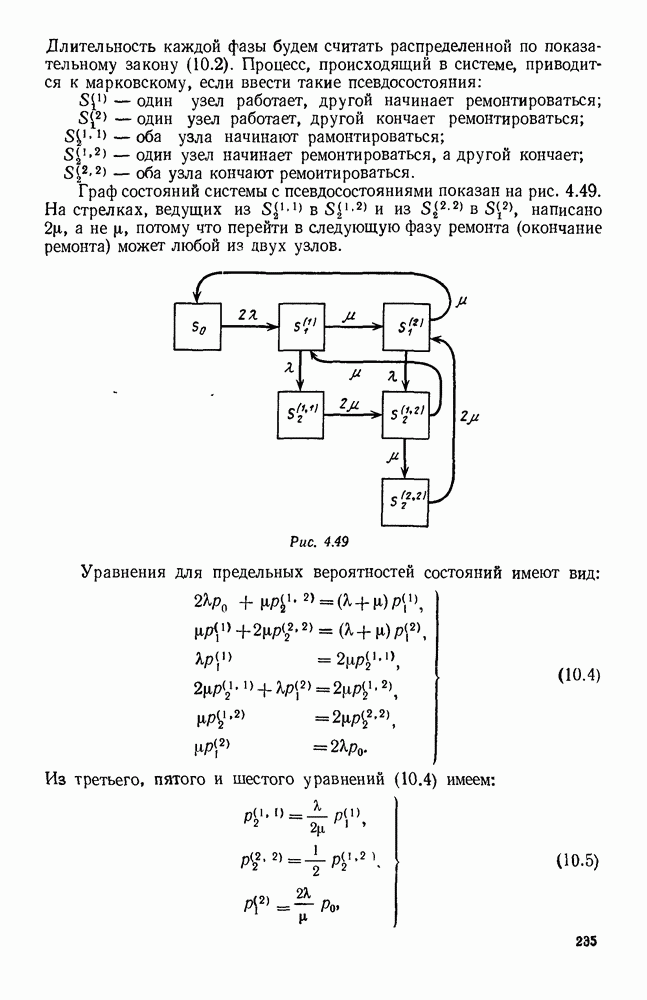
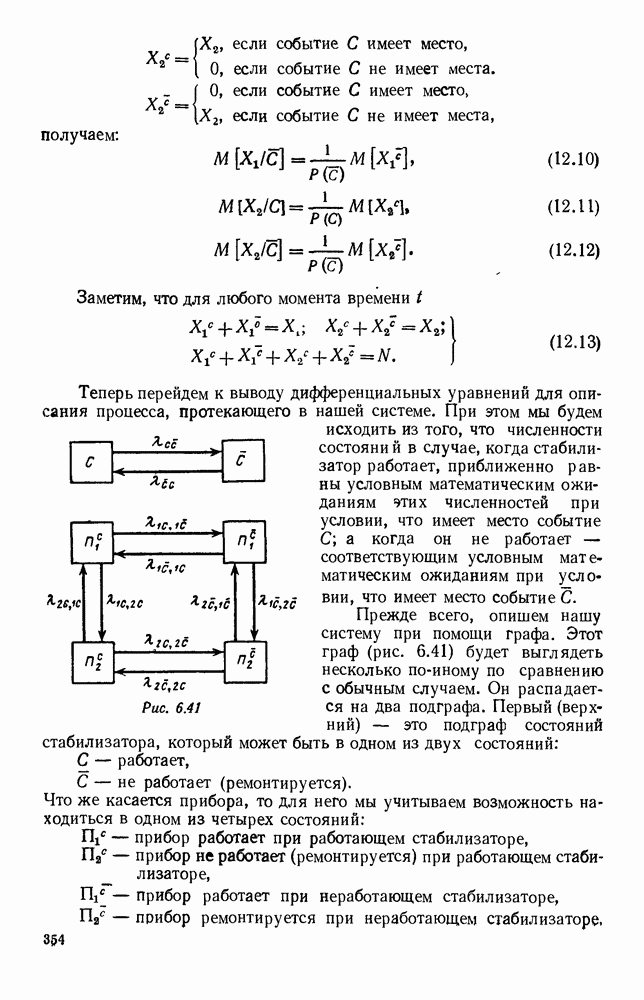
Рис. 6.23 При Таким образом, точка, изображающая решение системы (5.9), вращается вокруг точки
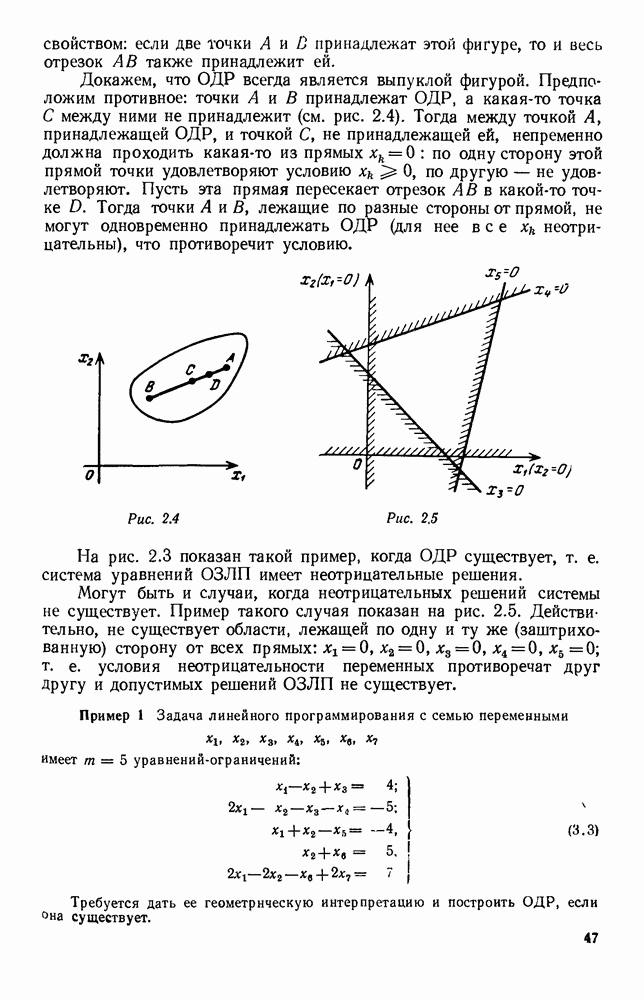
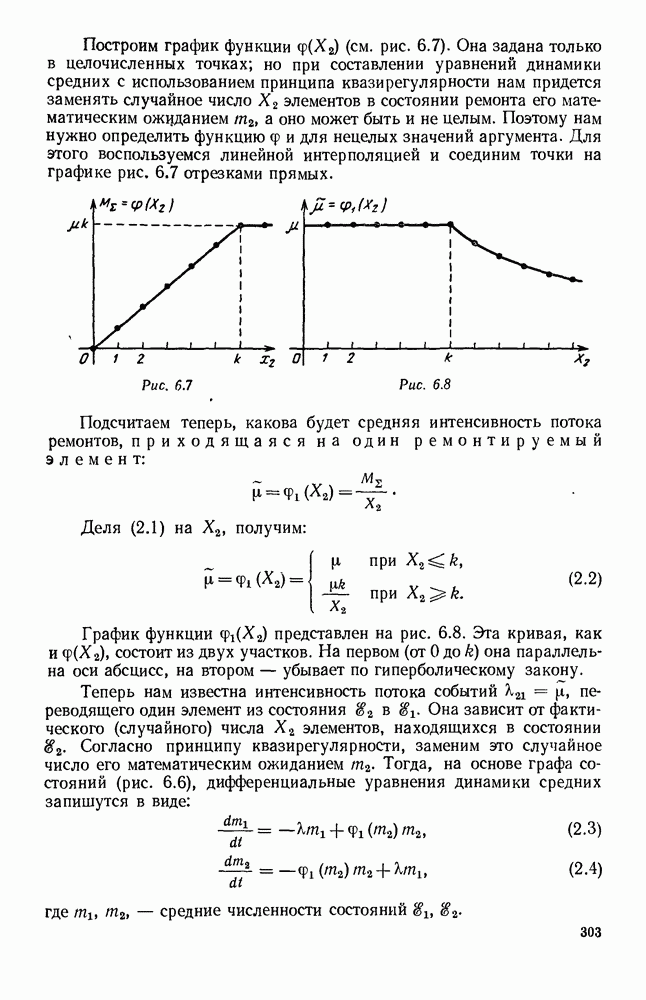
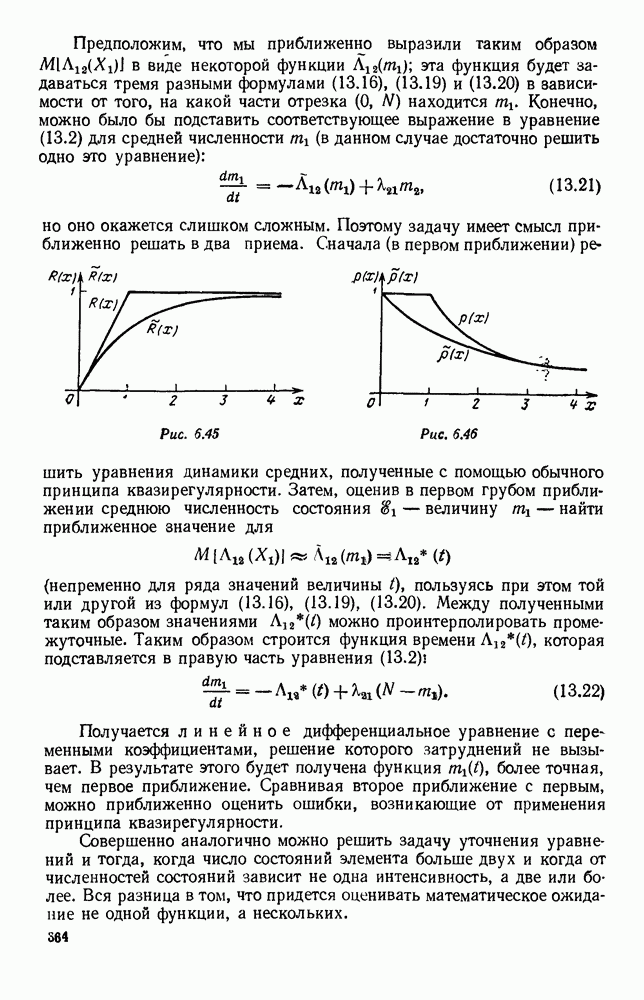
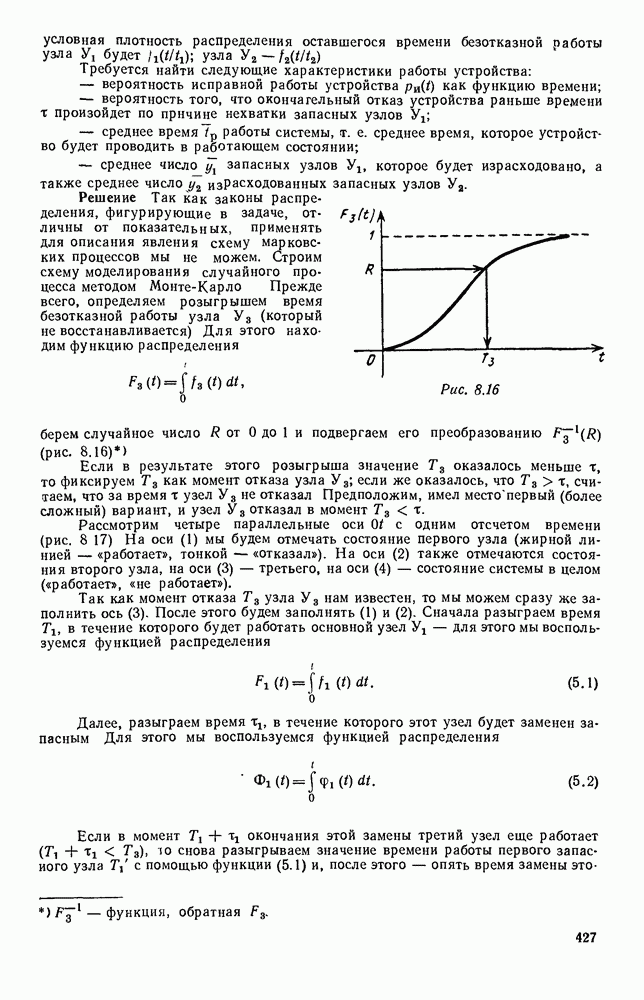
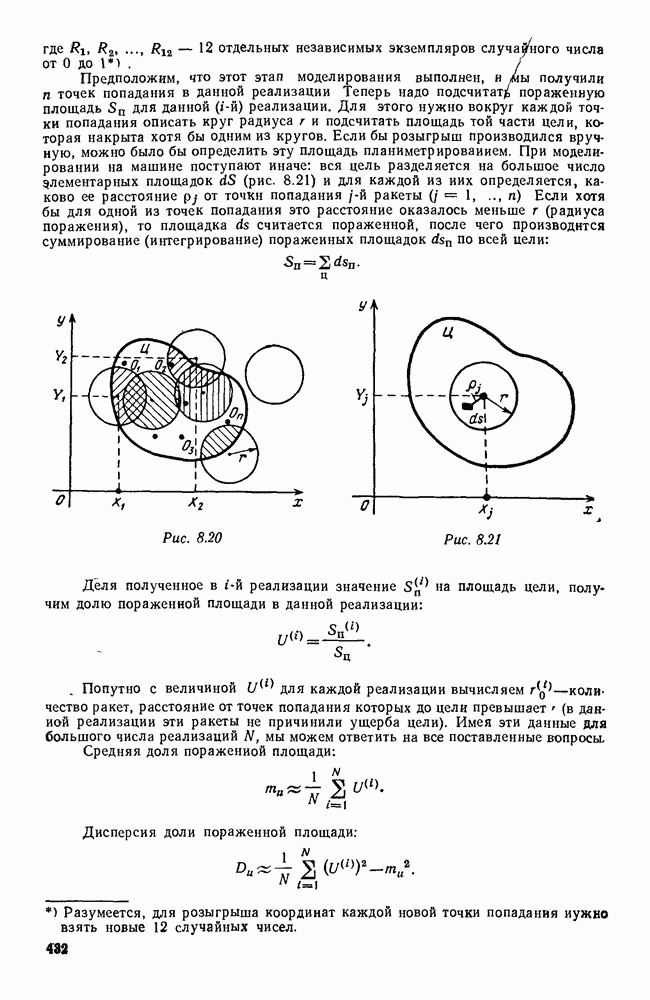
Это положение равновесия может быть устойчивым, если решения системы, начинающиеся из точек, близких к На рис. 6.23 изображен случай, когда решение, начинающееся на прямой — — после одного «оборота» точка окажется дальше от — после одного «оборота» точка в точности вернется к своему начальному положению. В последнем случае все последующие «обороты» точки Вернемся к случаю, изображенному на рис. 6.23. Следующий оборот точки Такие же рассуждения можно применить и в случае, когда после одного оборота точка Все эти характеры поведения решений системы (5.9) могут по-разному комбинироваться друг с другом, образуя иногда довольно сложную картину. Например, изобразим на рис. 6.24 случай, когда положение равновесия устойчиво, и никаких циклов (периодических решений) нет. Здесь будет единственный предельный режим Еще один пример возможного характера предельного поведения решений: положение равновесия неустойчиво, имеется один устойчивый цикл С (см. рис. 6.26).
Рис. 6.24
Рис. 6.25
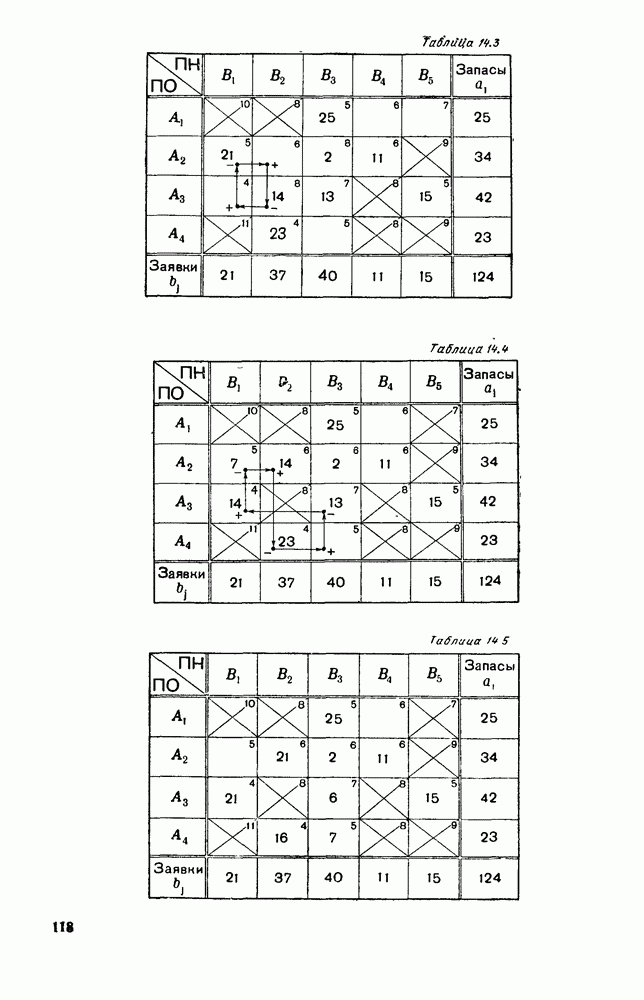
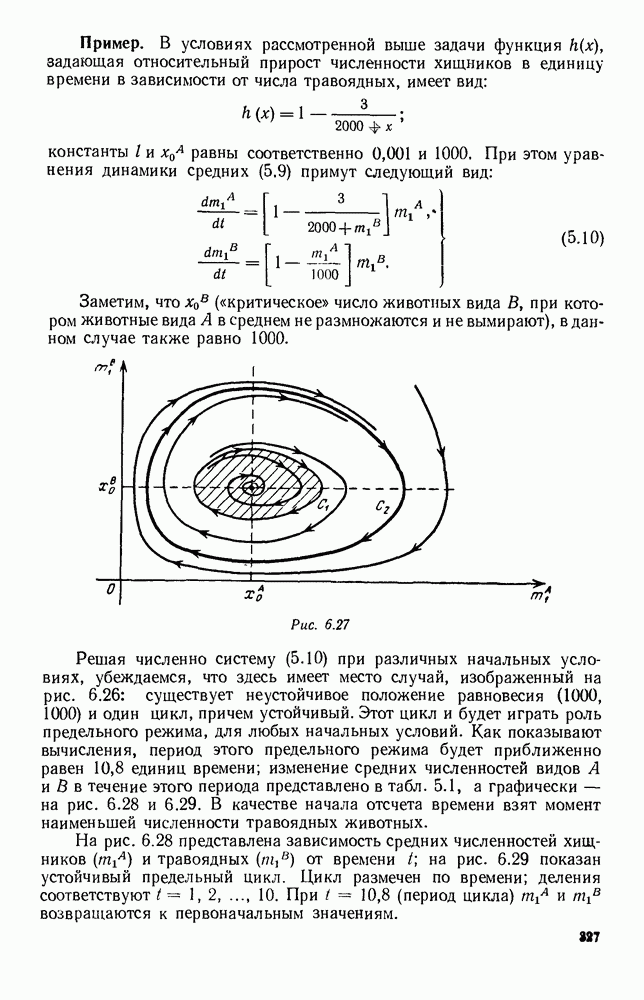
Рис. 6.26 Здесь будет опять единственный предельный режим — периодическое возрастание и убывание численностей, соответствующее предельному циклу С. К этому режиму приближаются средние численности На рис. 6.27 изображен еще более сложный случай: устойчивое положение равновесия, вокруг него неустойчивый цикл Конечно, может оказаться, что некоторые из перечисленных случаев, при определенных ограничениях на вид функции Пример. В условиях рассмотренной выше задачи функция
константы I и
Заметим, что
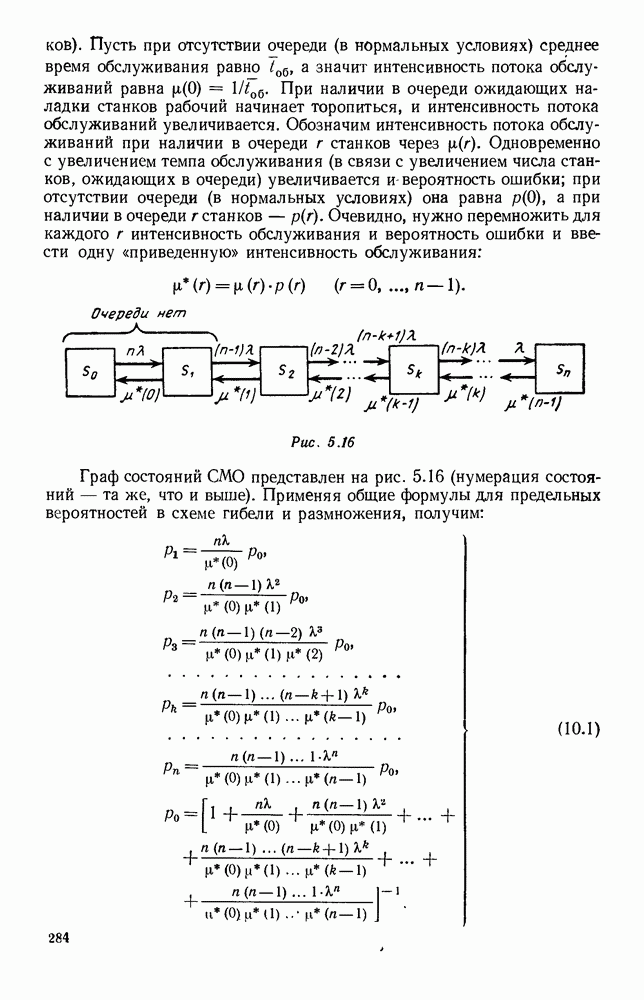
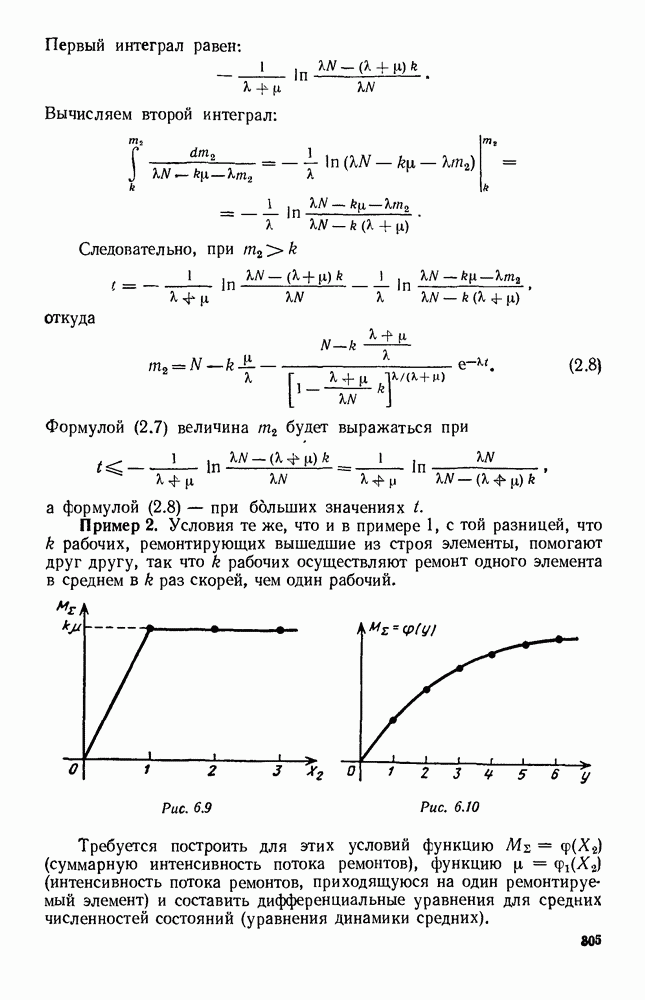
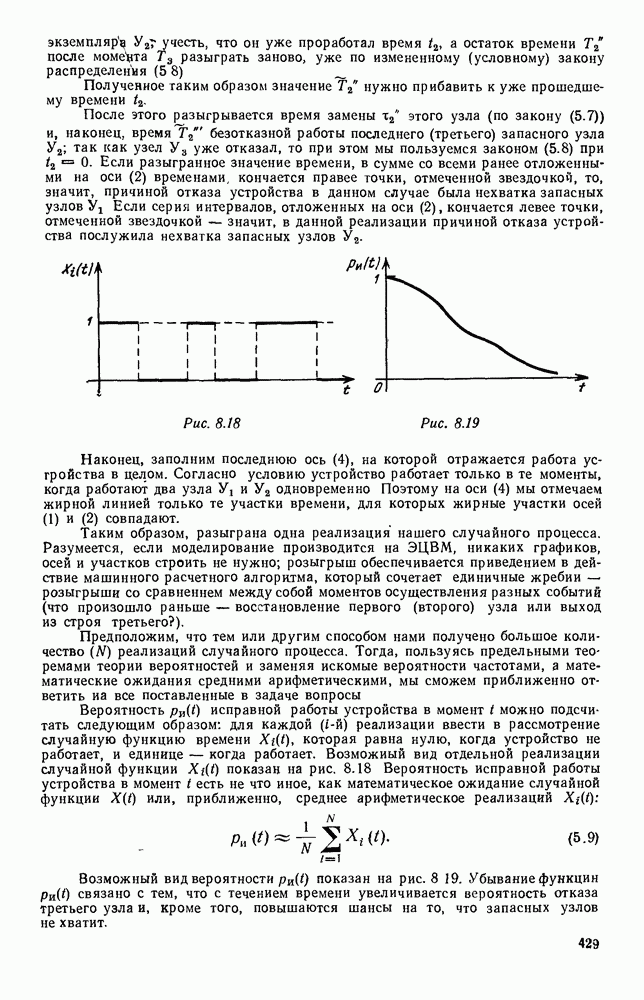
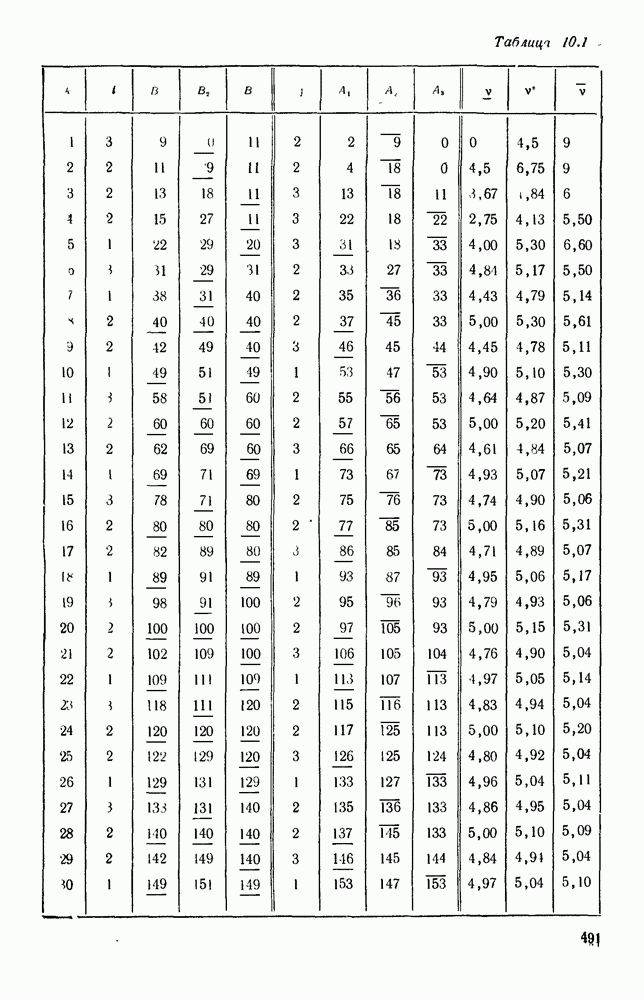
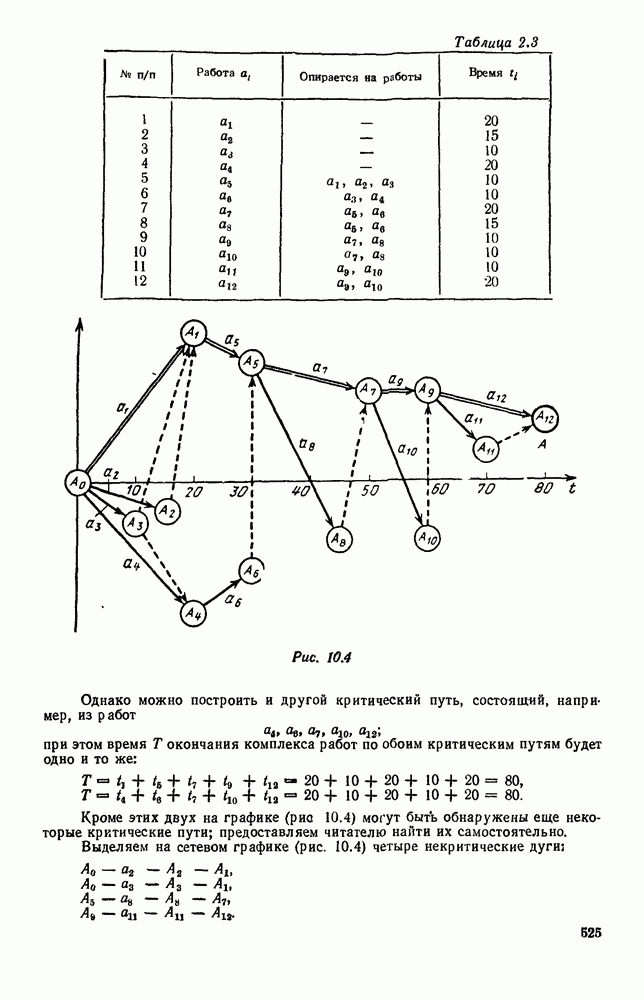
Рис. 6.27 Решая численно систему (5.10) при различных начальных условиях, убеждаемся, что здесь имеет место случай, изображенный на рис. 6.26: существует неустойчивое положение равновесия (1000, 1000) и один цикл, причем устойчивый. Этот цикл и будет играть роль предельного режима, для любых начальных условий. Как показывают вычисления, период этого предельного режима будет приближенно равен 10,8 единиц времени; изменение средних численностей видов А и В в течение этого периода представлено в табл. 5.1, а графически — на рис. 6.28 и 6.29. В качестве начала отсчета времени взят момент наименьшей численности травоядных животных. На рис. 6.28 представлена зависимость средних численностей хищников Таблица 5.1
Анализируя данные, приведенные в табл. 5.1, мы видим, что вначале, когда число травоядных мало (в среднем 450), хищники голодают, их число уменьшается, и поэтому число травоядных начинает расти.
Рис. 6.28
Рис. 6.29 К тому моменту, когда число травоядных достигает 1000 (это происходит немного больше, чем через 3 единицы времени от начала цикла), среднее число хищников достигает своего минимума (около 620). После того, как травоядных становится более 1000, число хищников опять начинает расти и снова достигает 1000 немного позже t = 6. Число травоядных в это время достигает максимума (2170 или немного больше), а потом снова начинает убывать, потому что число хищников больше критического 1000. Затем (при t между 8 и 9) число хищников достигает максимума, равного приблизительно 1550, а число травоядных в это время равно критическому 1000. На последнем участке цикла (до момента t = 10,8) убывает и число травоядных (потому что число хищников превосходит 1000), и число хищников (потому что травоядных меньше 1000). В конце периода, при t = 10,8, число хищников опять достигает критического значения 1000, а число травоядных возвращается к своему наименьшему значению 450. Любопытно, что в природе действительно встречаются такие периодические изменения численностей связанных друг с другом видов.
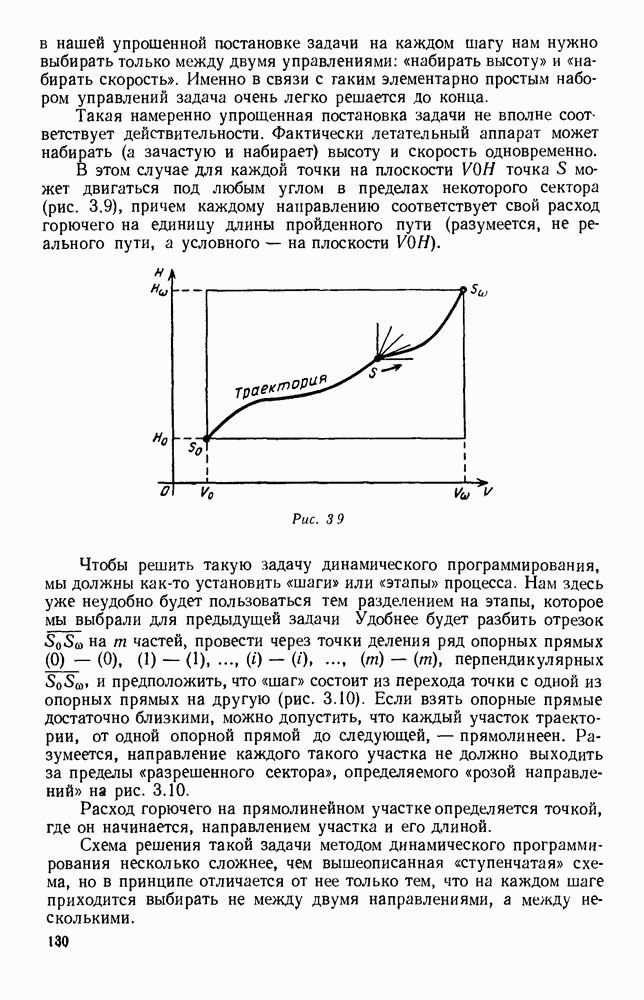
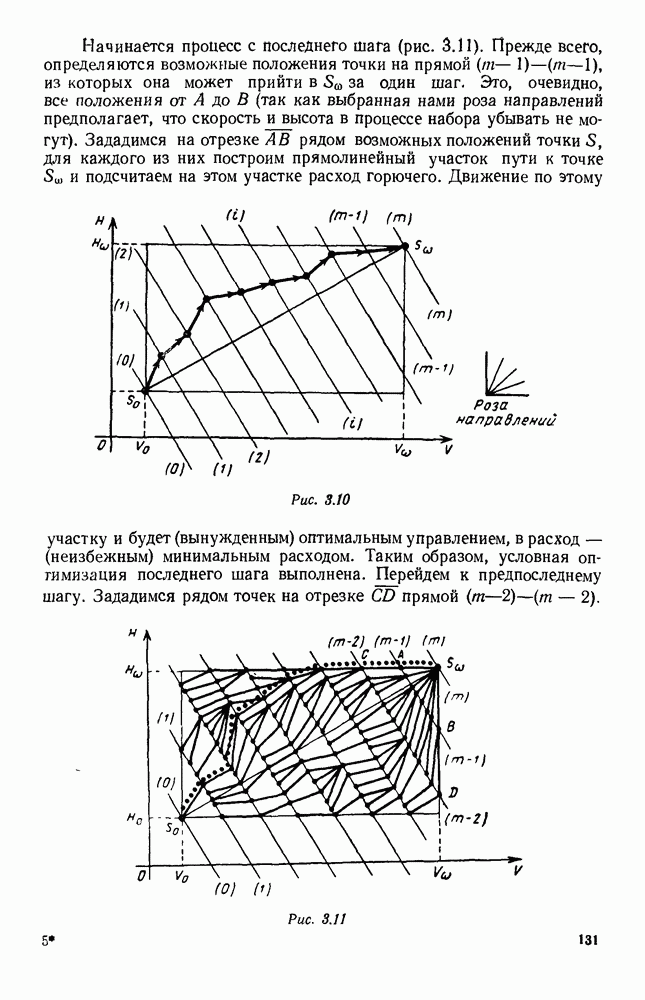
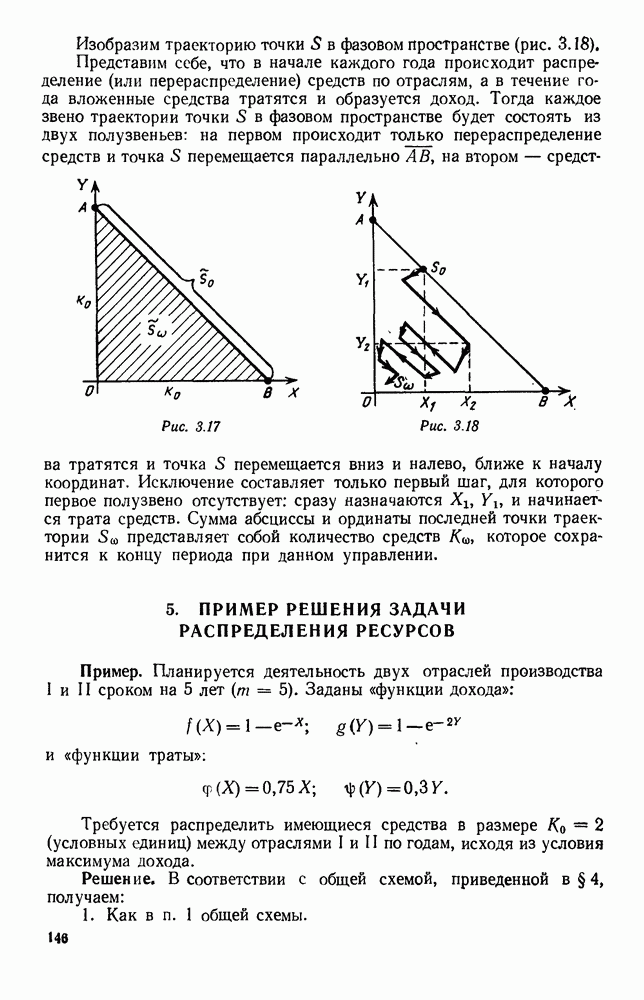
|
 |
Оглавление
|








































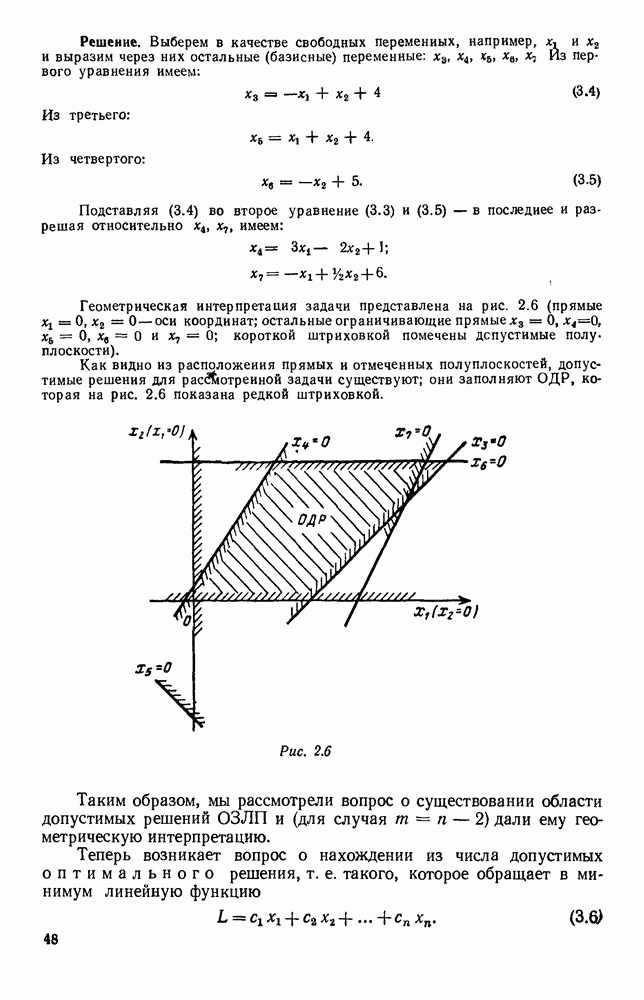
















































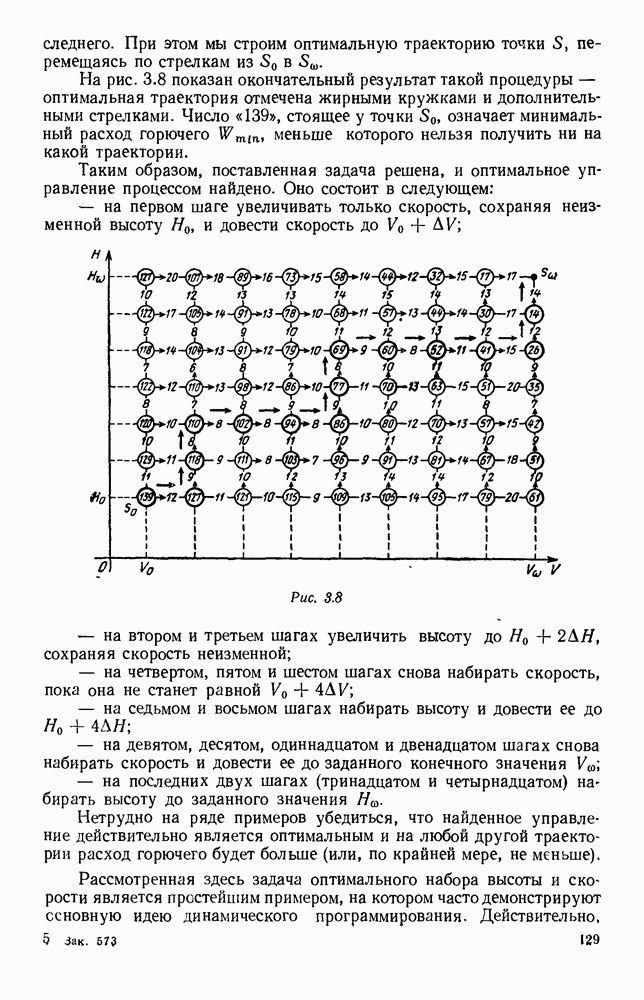
















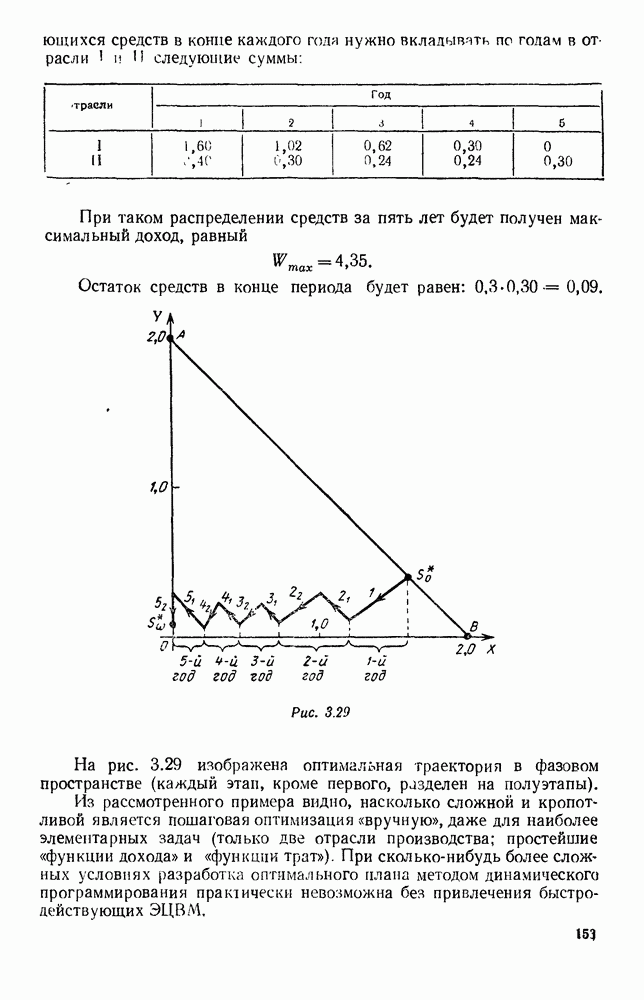



























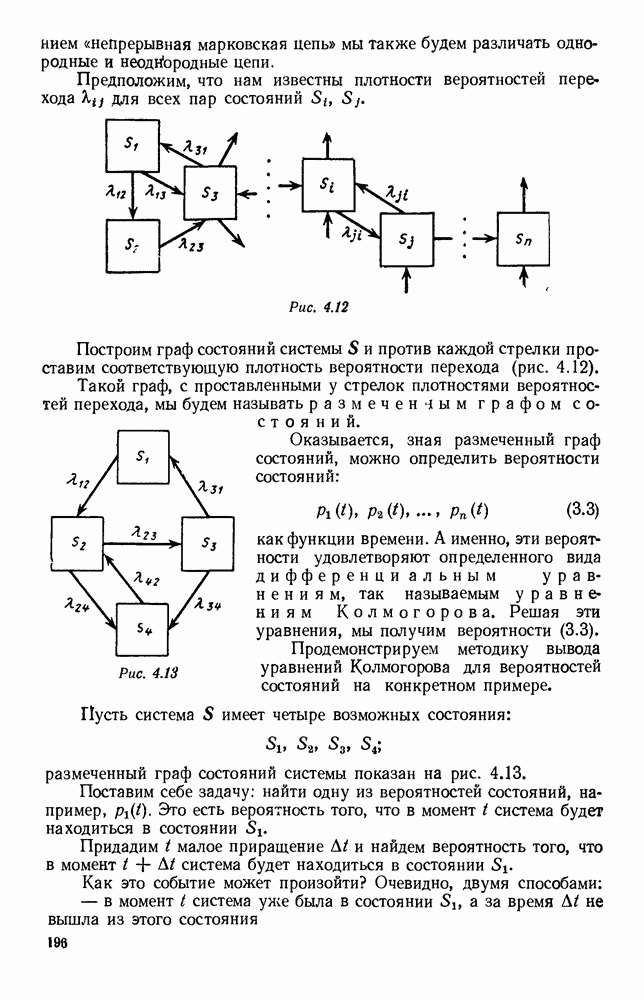








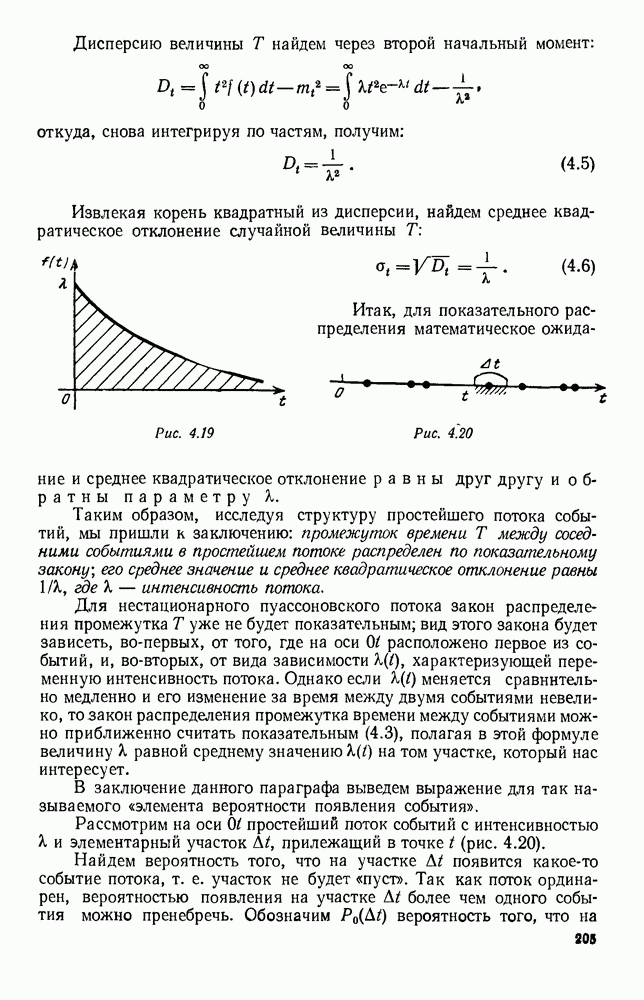





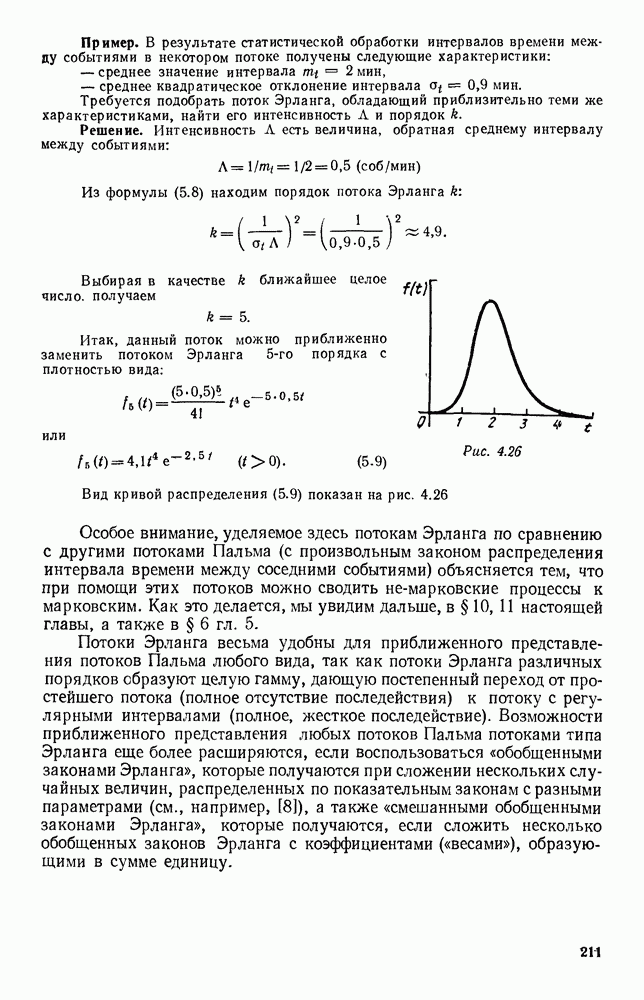

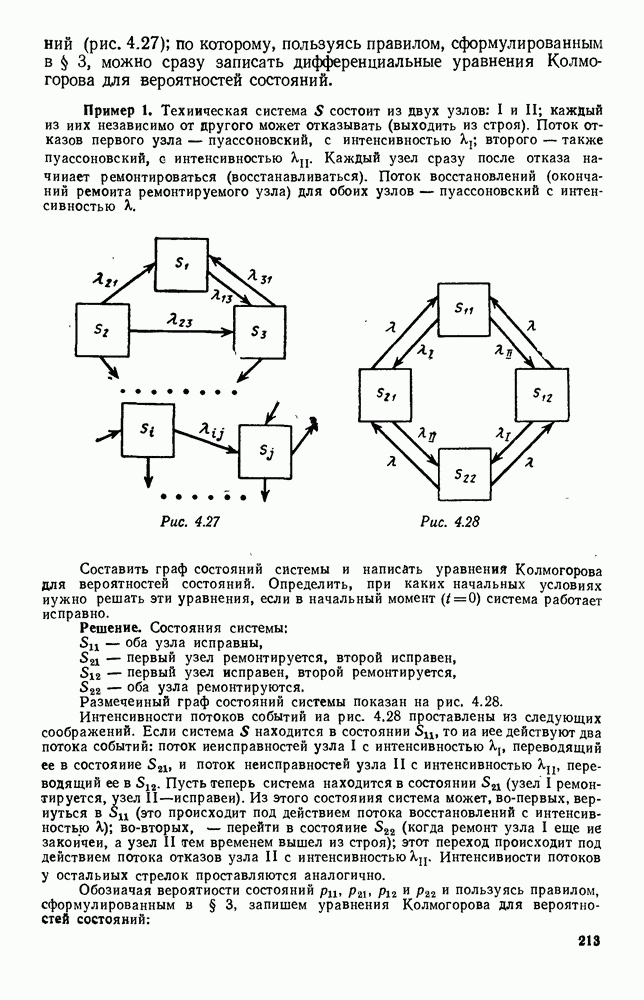
































































































































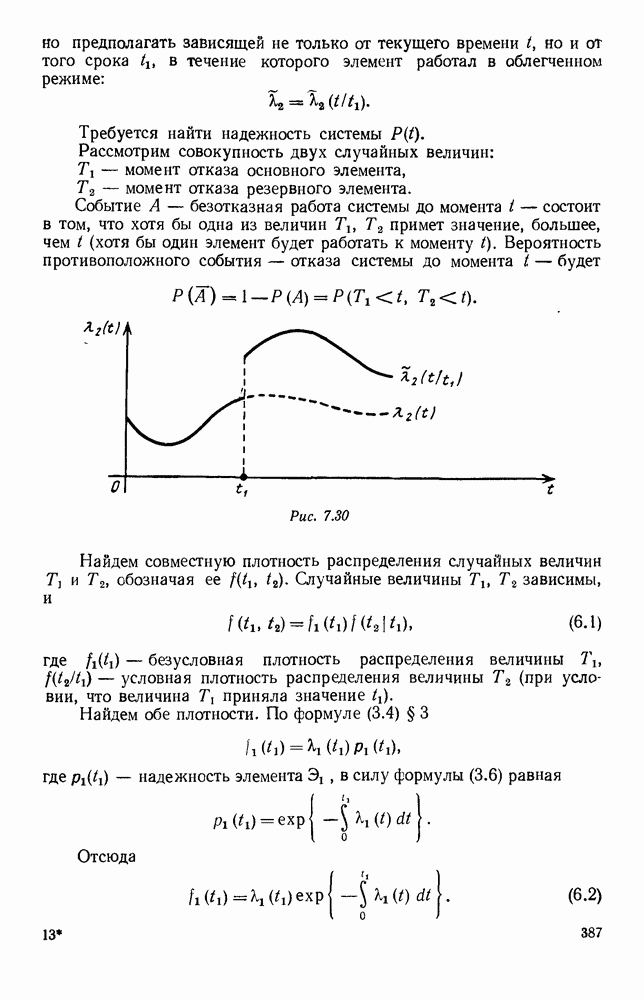
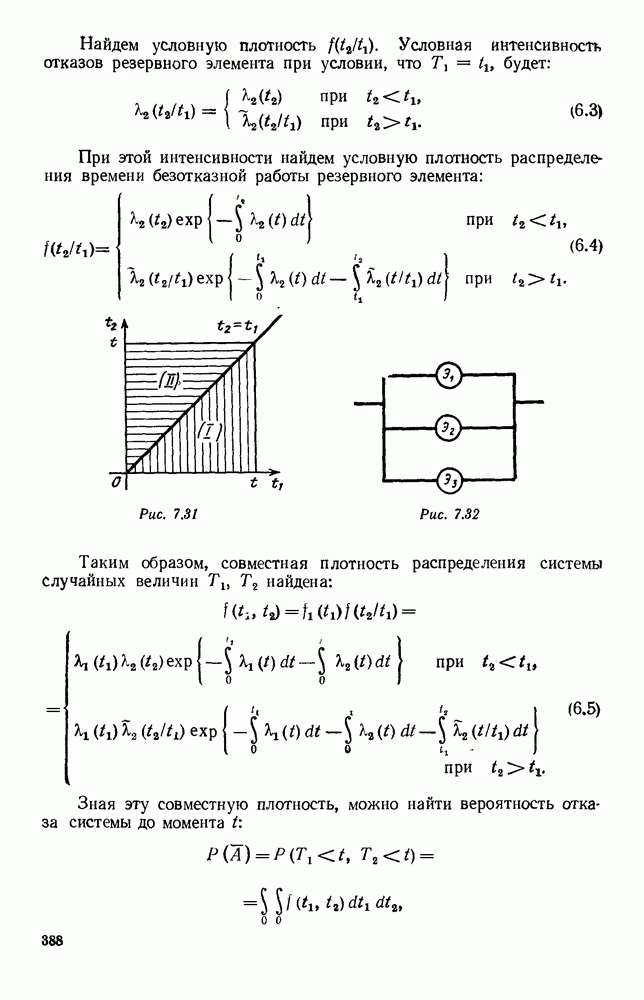



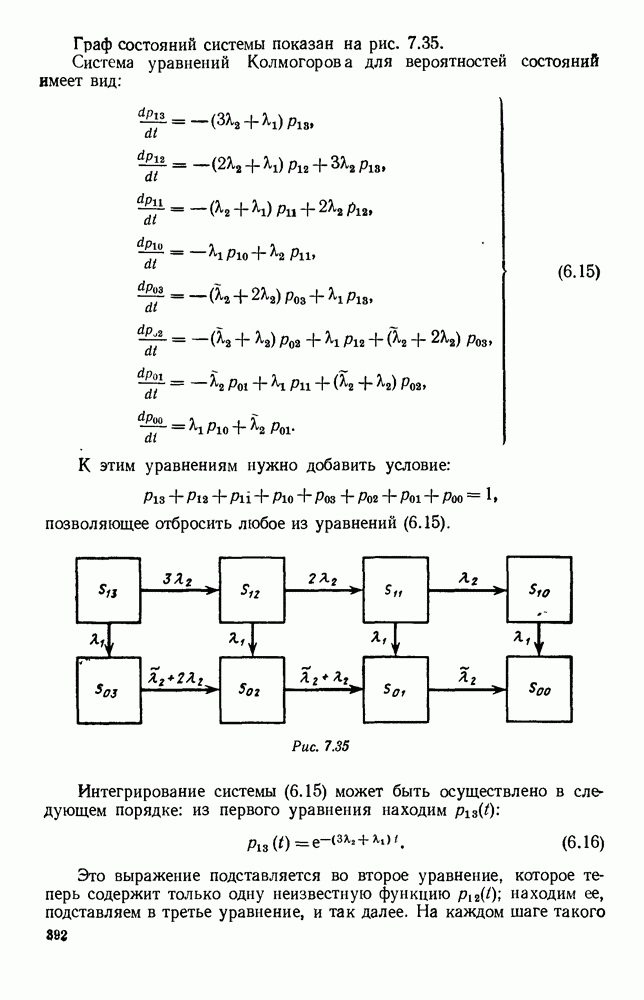


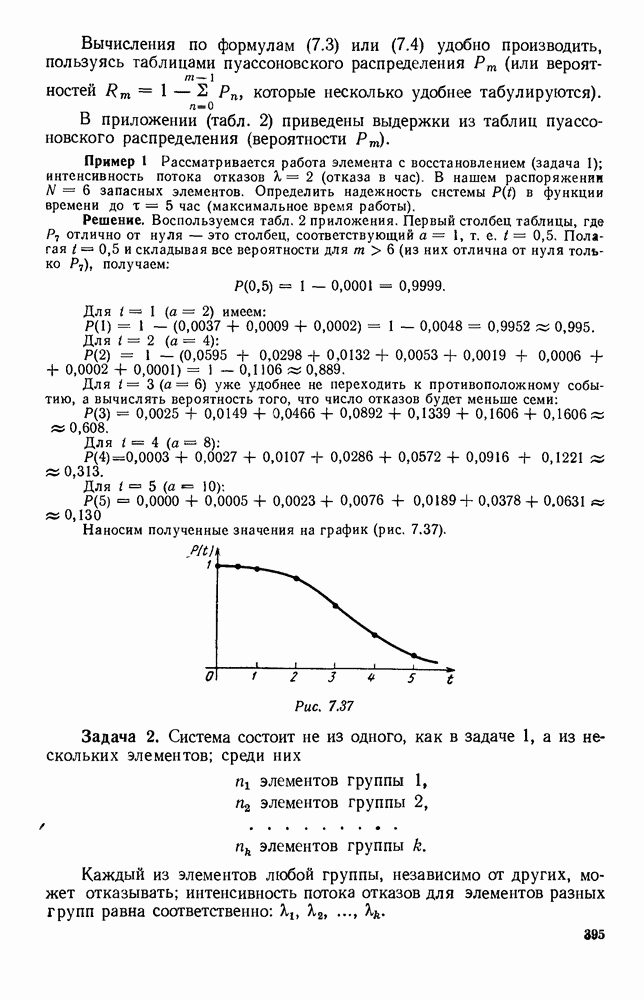





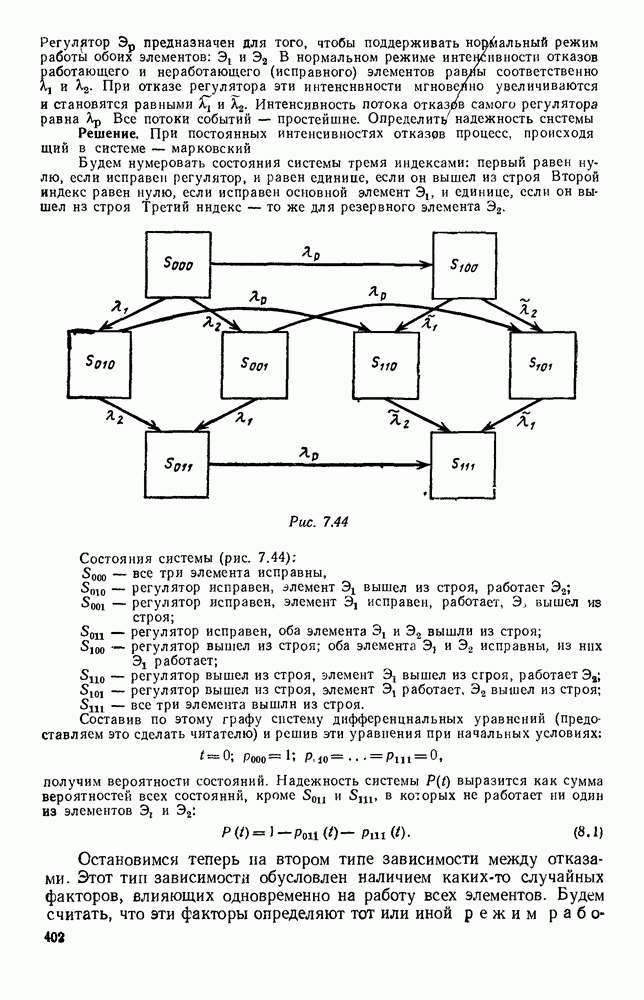























































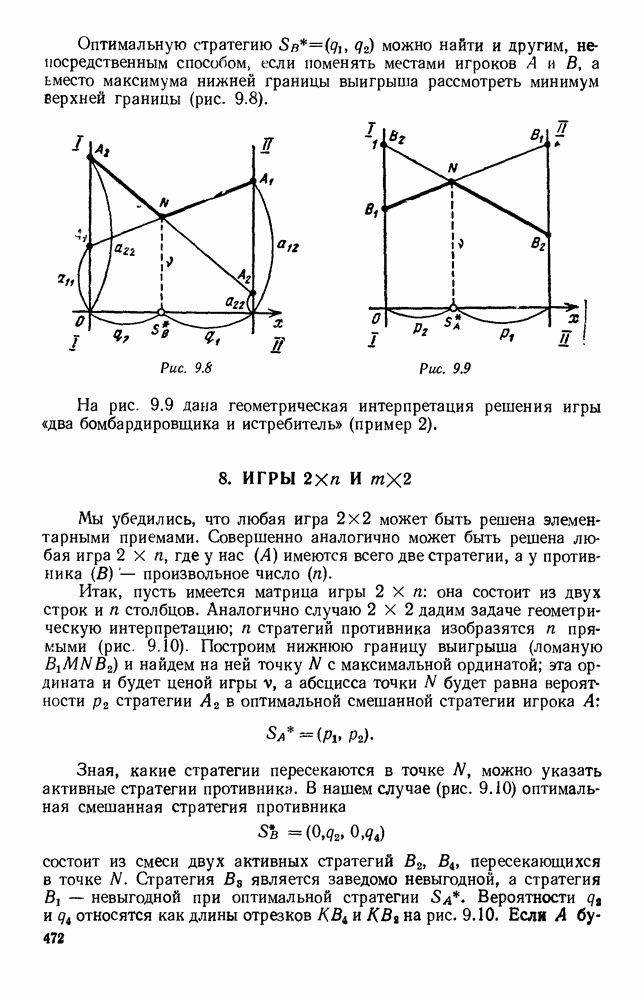
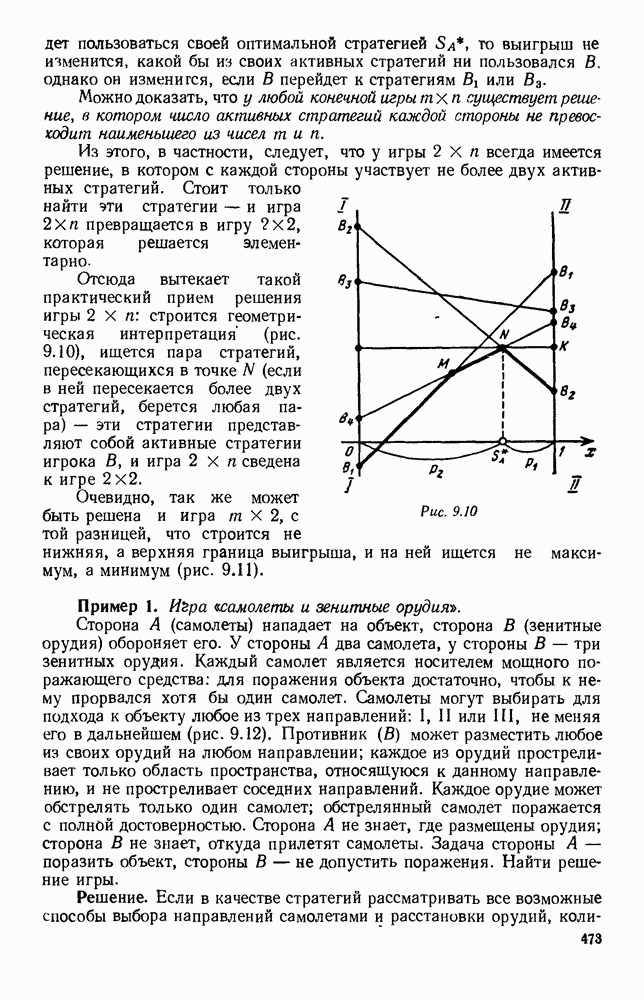


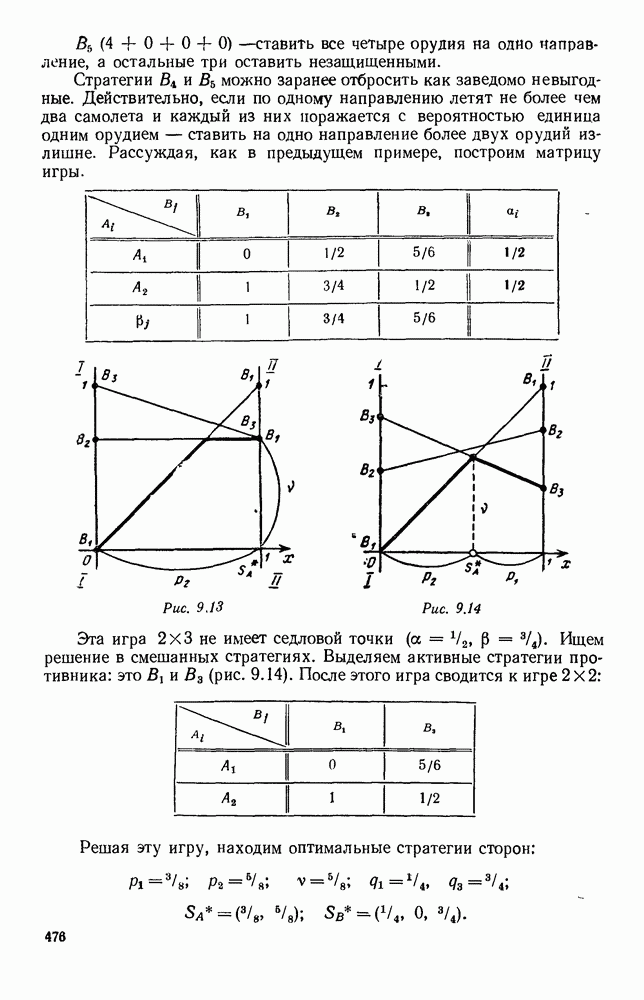







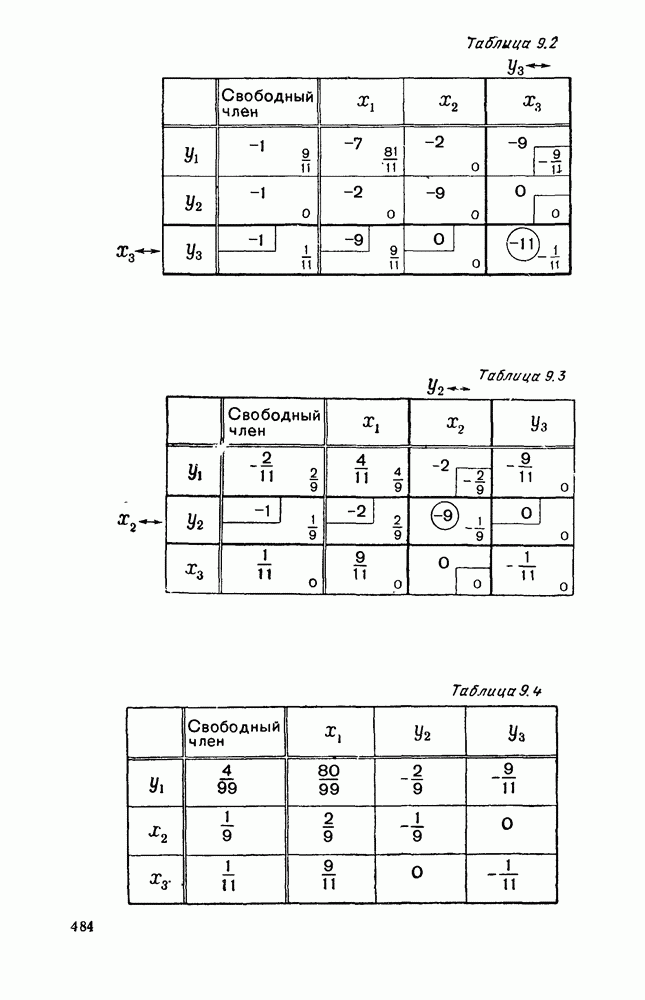
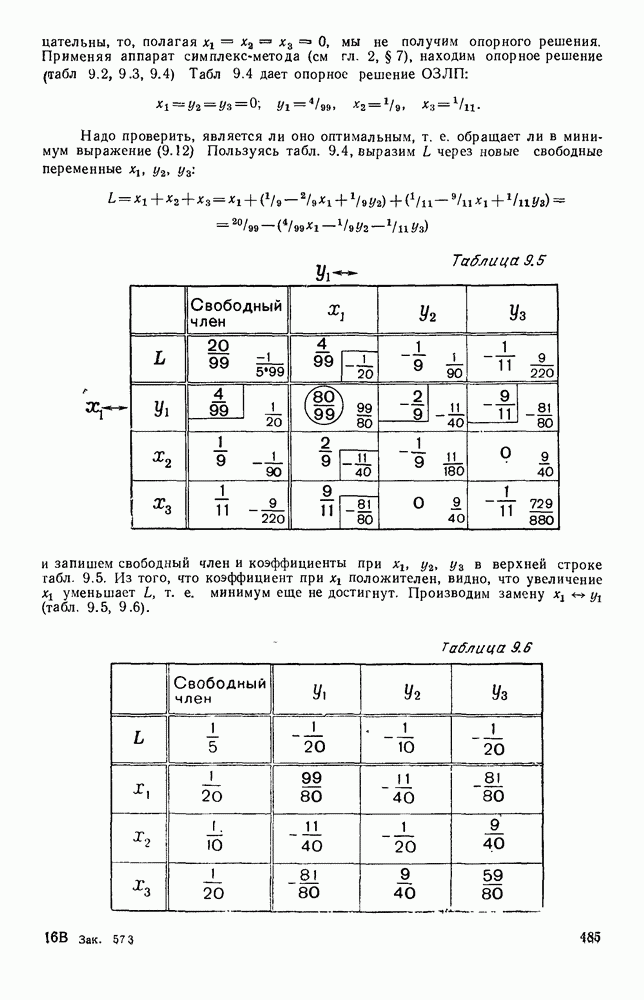

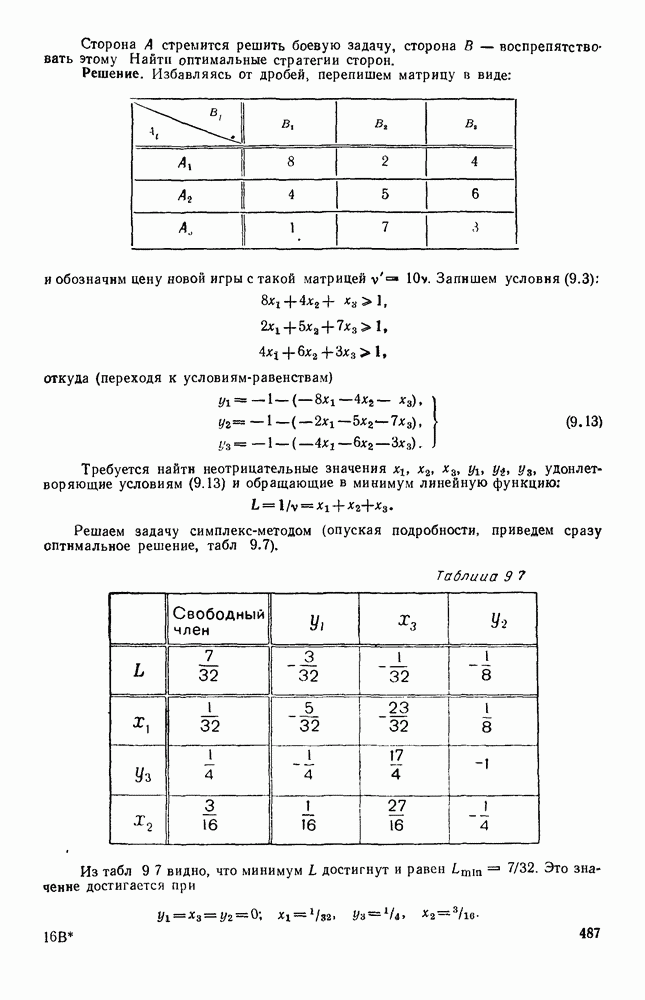
















































 Существует ли, и если существует, то каков установившийся режим?
Существует ли, и если существует, то каков установившийся режим?


 и из
и из  учитывают смертность животных, причем для вида В—смертность двух родов: и те случаи, когда особь погибает естественной смертью, и те, когда ее поедает животное вида А. Двойные полустрелки, ведущие в состояния
учитывают смертность животных, причем для вида В—смертность двух родов: и те случаи, когда особь погибает естественной смертью, и те, когда ее поедает животное вида А. Двойные полустрелки, ведущие в состояния  соответствуют пополнению численностей благодаря рождаемости.
соответствуют пополнению численностей благодаря рождаемости.
 соответственно через
соответственно через  а их математические ожидания через
а их математические ожидания через  . Мы хотим составить дифференциальные уравнения для
. Мы хотим составить дифференциальные уравнения для  (средние численности
(средние численности  предыдущих поколений, как окончательно выбывшие из «активных» элементов системы, не будут входить в эти уравнения, и мы можем ими совсем не интересоваться).
предыдущих поколений, как окончательно выбывшие из «активных» элементов системы, не будут входить в эти уравнения, и мы можем ими совсем не интересоваться).
 от
от  .
.
 за счет рождения новых животных выразится формулой:
за счет рождения новых животных выразится формулой:

 то мы уже сказали, что она складывается из двух слагаемых. Первое — это естественная смертность; мы будем считать ее постоянной (по тем же причинам, по которым считали постоянной рождаемость). Обозначим эту постоянную (среднее число естественных смертей в пересчете на одну живую особь вида В) через k; величина k может быть истолкована как доля животных вида В, погибающих в единицу времени естественной смертью. Второе слагаемое в составе
то мы уже сказали, что она складывается из двух слагаемых. Первое — это естественная смертность; мы будем считать ее постоянной (по тем же причинам, по которым считали постоянной рождаемость). Обозначим эту постоянную (среднее число естественных смертей в пересчете на одну живую особь вида В) через k; величина k может быть истолкована как доля животных вида В, погибающих в единицу времени естественной смертью. Второе слагаемое в составе  — это доля животных вида В, поедаемых в единицу времени хищниками вида А. Естественно предположить, что число встреч (в единицу времени) животных видов А и В, кончающихся тем, что А поедает В, прямо пропорционально числу
— это доля животных вида В, поедаемых в единицу времени хищниками вида А. Естественно предположить, что число встреч (в единицу времени) животных видов А и В, кончающихся тем, что А поедает В, прямо пропорционально числу  животных вида А (живых) и числу
животных вида А (живых) и числу  имеющихся животных вида В, т. е. что это число выражается формулой
имеющихся животных вида В, т. е. что это число выражается формулой

 так что интенсивность потока гибелей, приходящаяся на одну особь вида В в состоянии
так что интенсивность потока гибелей, приходящаяся на одну особь вида В в состоянии  выразится так:
выразится так:

 Таким образом, смертность хищников
Таким образом, смертность хищников  будет какой-то функцией от
будет какой-то функцией от  или, так как I — константа, какой-то функцией от Х:
или, так как I — константа, какой-то функцией от Х:

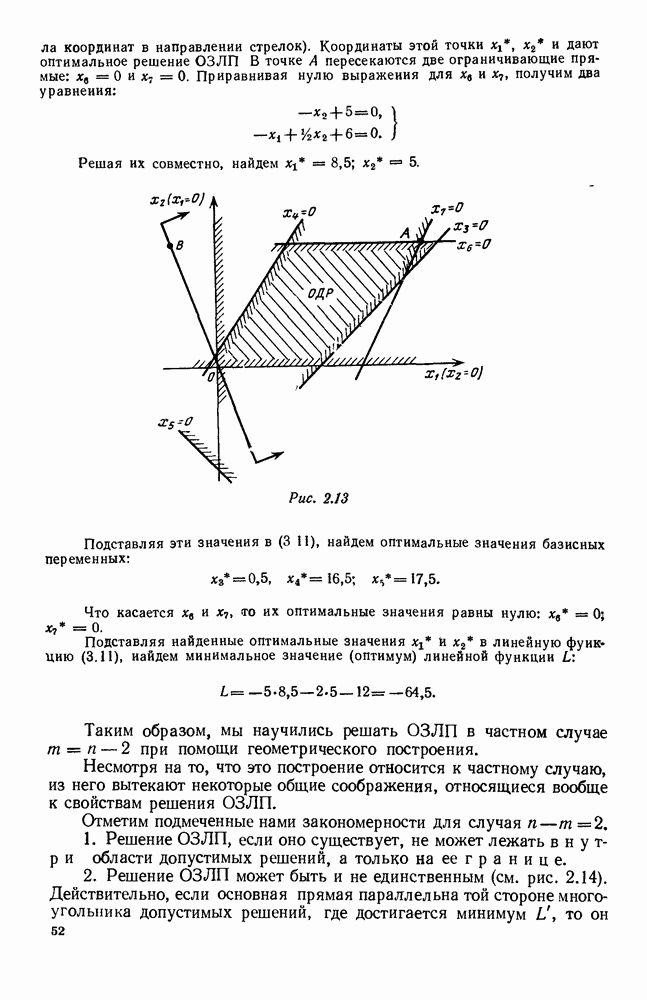
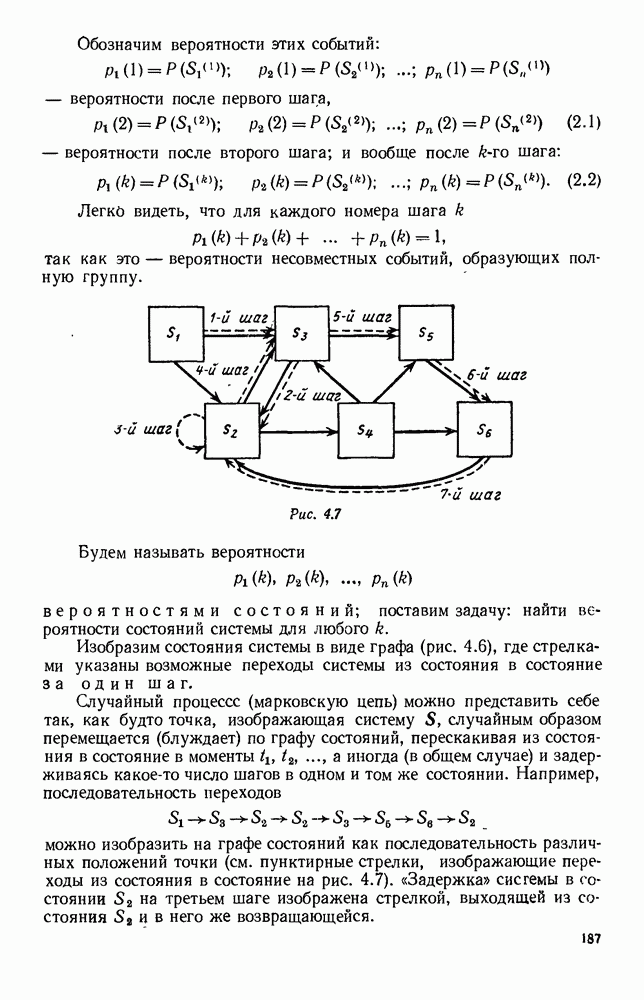
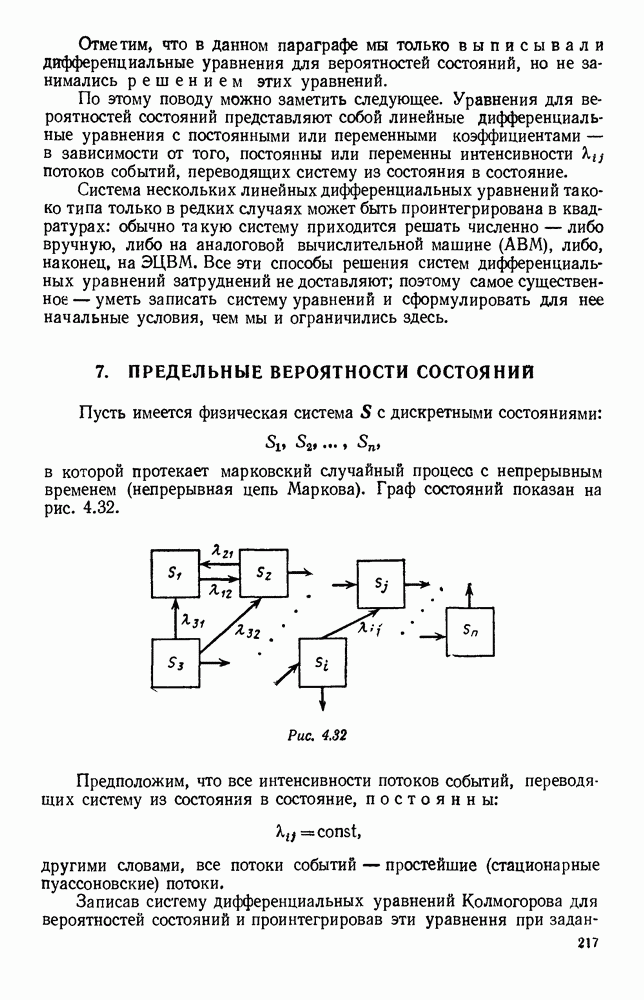
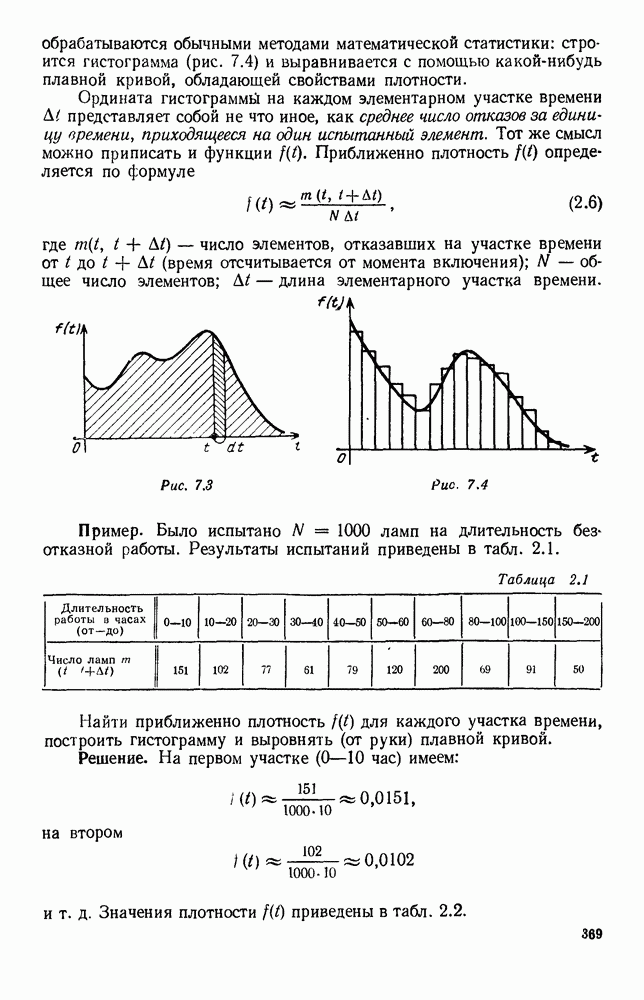
 будет убывающей функцией от
будет убывающей функцией от  (т. е. от количества пищи). Действительно, животные будут тем меньше умирать, чем больше у них будет пищи. Возможный вид функции
(т. е. от количества пищи). Действительно, животные будут тем меньше умирать, чем больше у них будет пищи. Возможный вид функции  представлен на рис. 6.20.
представлен на рис. 6.20.
 и средний прирост популяции хищников за счет рождаемости (в единицу времени) запишется так:
и средний прирост популяции хищников за счет рождаемости (в единицу времени) запишется так:


 от которых зависят интенсивности (5.1), (5.3), (5.4), (5.5), их средними значениями
от которых зависят интенсивности (5.1), (5.3), (5.4), (5.5), их средними значениями  получим:
получим:



 характеризующая полный средний прирост. Обозначим эту разность через
характеризующая полный средний прирост. Обозначим эту разность через 

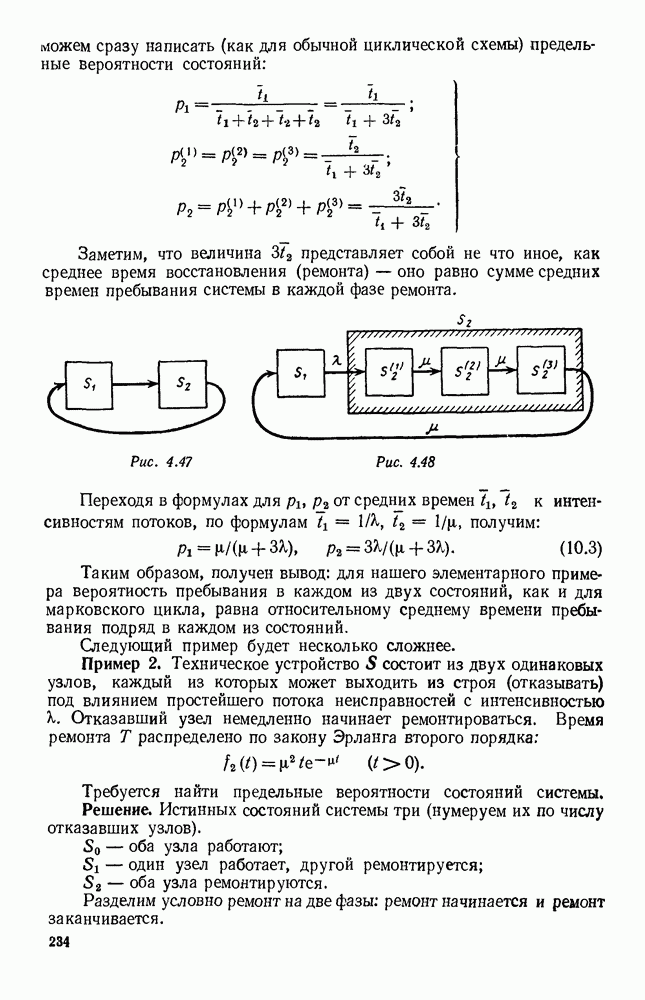
 и g, эта функция может менять знак. Предположим, что она меняет знак в какой-то точке
и g, эта функция может менять знак. Предположим, что она меняет знак в какой-то точке  (рис. 6.22). Значение
(рис. 6.22). Значение  имеет смысл той численности животных вида В, при которой рождаемость и смертность хищников в среднем уравновешиваются. При больших значениях численности вида В численность вида А в среднем возрастает, при меньших — в среднем убывает. Назовем
имеет смысл той численности животных вида В, при которой рождаемость и смертность хищников в среднем уравновешиваются. При больших значениях численности вида В численность вида А в среднем возрастает, при меньших — в среднем убывает. Назовем  критической численностью вида В.
критической численностью вида В.
 хищников, при которой число травоядных животных в среднем остается неизменным. Из второго уравнения (5.6) видно, что
хищников, при которой число травоядных животных в среднем остается неизменным. Из второго уравнения (5.6) видно, что

 то численность вида В убывает, а при меньшем их числе — возрастает.
то численность вида В убывает, а при меньшем их числе — возрастает.

 системы (5.9) (см. рис. 6.23).
системы (5.9) (см. рис. 6.23).


 т. е. ниже прямой
т. е. ниже прямой  согласно первому
согласно первому  уравнению (5.9),
уравнению (5.9),  это означает, что точка движется влево (одновременно с этим, может быть, вверх или вниз). Выше этой прямой точка движется вправо. Точно так же из второго уравнения (5.9) получаем, что левее прямой
это означает, что точка движется влево (одновременно с этим, может быть, вверх или вниз). Выше этой прямой точка движется вправо. Точно так же из второго уравнения (5.9) получаем, что левее прямой  движение точки направлено вверх, а правее этой прямой — вниз. В моменты прохождения прямой
движение точки направлено вверх, а правее этой прямой — вниз. В моменты прохождения прямой  точка движется строго вертикально: вниз, если она находится правее
точка движется строго вертикально: вниз, если она находится правее  и вверх — если левее; в моменты прохождения прямой
и вверх — если левее; в моменты прохождения прямой  она движется строго горизонтально.
она движется строго горизонтально.
 по часовой стрелке, причем вращается, вообще говоря, не по кругу — может быть, приближаясь к точке
по часовой стрелке, причем вращается, вообще говоря, не по кругу — может быть, приближаясь к точке  а может быть удаляясь от нее. Точка
а может быть удаляясь от нее. Точка  — положение равновесия: если в качестве начальных условий для решения системы (5.9) взять
— положение равновесия: если в качестве начальных условий для решения системы (5.9) взять  то численности
то численности  будут изменяться, и решение системы (5.9) не зависит от времени:
будут изменяться, и решение системы (5.9) не зависит от времени:

 с течением времени неограниченно приближаются к этой точке; и неустойчивым — если удаляются.
с течением времени неограниченно приближаются к этой точке; и неустойчивым — если удаляются.
 через один «оборот» оказывается ближе к точке
через один «оборот» оказывается ближе к точке  чем в начале. Очевидно, возможны еще два случая:
чем в начале. Очевидно, возможны еще два случая:
 чем в начале;
чем в начале;
 вокруг положения равновесия будут совершаться по одному и тому же замкнутому пути. Такой замкнутый путь в теории дифференциальных уравнений называется циклом; ему соответствует периодическое решение
вокруг положения равновесия будут совершаться по одному и тому же замкнутому пути. Такой замкнутый путь в теории дифференциальных уравнений называется циклом; ему соответствует периодическое решение  ): число хищников возрастает, поэтому число травоядных начинает убывать, достигает
): число хищников возрастает, поэтому число травоядных начинает убывать, достигает  ; тогда начинает убывать число хищников, и, когда оно минует критическое значение
; тогда начинает убывать число хищников, и, когда оно минует критическое значение  число травоядных начинает возрастать; наконец, численности тех и других животных достигают своих прежних значений и процесс начинается заново. Так же, как и положение равновесия, цикл может быть устойчивым (когда решения, начинающиеся вблизи него, неограниченно приближаются к циклу) и неустойчивым (когда они удаляются).
число травоядных начинает возрастать; наконец, численности тех и других животных достигают своих прежних значений и процесс начинается заново. Так же, как и положение равновесия, цикл может быть устойчивым (когда решения, начинающиеся вблизи него, неограниченно приближаются к циклу) и неустойчивым (когда они удаляются).
 вокруг положения равновесия еще больше приблизит ее к точке
вокруг положения равновесия еще больше приблизит ее к точке  и с каждым следующим оборотом точка
и с каждым следующим оборотом точка  будет все больше приближаться к точке
будет все больше приближаться к точке  Здесь опять-таки может быть два случая: либо точка будет неограниченно приближаться к положению равновесия — тогда, очевидно, это положение равновесия будет устойчивым; либо будет с каждым оборотом приближаться, но не неограниченно. В этом случае точка
Здесь опять-таки может быть два случая: либо точка будет неограниченно приближаться к положению равновесия — тогда, очевидно, это положение равновесия будет устойчивым; либо будет с каждым оборотом приближаться, но не неограниченно. В этом случае точка  будет неограниченно приближаться снаружи к какому-то циклу, который, конечно, будет устойчивым.
будет неограниченно приближаться снаружи к какому-то циклу, который, конечно, будет устойчивым.
 удалится от положения равновесия: либо с течением времени она будет неограниченно удаляться от точки
удалится от положения равновесия: либо с течением времени она будет неограниченно удаляться от точки  а значит, и от начала координат; либо она будет приближаться к какому-то устойчивому циклу (на этот раз изнутри).
а значит, и от начала координат; либо она будет приближаться к какому-то устойчивому циклу (на этот раз изнутри).
 и к нему будут стремиться все решения, каковы бы ни были начальные условия. На рис. 6.25 изображен другой случай, когда положение равновесия неустойчиво, а циклов, как и в предыдущем случае, нет. В этом случае никакого предельного режима не существует: численность каждого вида то убывает почти до нуля, то сильно возрастает, причем с каждым «оборотом» эти колебания становятся все больше, возрастая неограниченно. Разумеется, в точности такой характер предельного поведения численностей видов практически не может встретиться. Здесь начнут сказываться не учтенные при формализации задачи факторы: например, ограниченность запасов пищи у травоядных или ошибки, связанные с применением принципа квазирегулярности (которые будут значительны при малых значениях
и к нему будут стремиться все решения, каковы бы ни были начальные условия. На рис. 6.25 изображен другой случай, когда положение равновесия неустойчиво, а циклов, как и в предыдущем случае, нет. В этом случае никакого предельного режима не существует: численность каждого вида то убывает почти до нуля, то сильно возрастает, причем с каждым «оборотом» эти колебания становятся все больше, возрастая неограниченно. Разумеется, в точности такой характер предельного поведения численностей видов практически не может встретиться. Здесь начнут сказываться не учтенные при формализации задачи факторы: например, ограниченность запасов пищи у травоядных или ошибки, связанные с применением принципа квазирегулярности (которые будут значительны при малых значениях  ).
).



 при любых начальных условиях.
при любых начальных условиях.
 а вокруг него — еще один, но устойчивый цикл
а вокруг него — еще один, но устойчивый цикл  Предельных режимов здесь два: положение равновесия
Предельных режимов здесь два: положение равновесия  и периодический режим, соответствующий циклу
и периодический режим, соответствующий циклу  . К первому предельному режиму численности
. К первому предельному режиму численности  приближаются, если начальные условия находятся внутри цикла
приближаются, если начальные условия находятся внутри цикла  (заштрихованная область на рис. 6.27); ко второму — если вначале точка
(заштрихованная область на рис. 6.27); ко второму — если вначале точка  находилась вне цикла
находилась вне цикла 
 (например, для ограниченной функции
(например, для ограниченной функции  ) невозможны. Но, во всяком случае, картина предельного поведения решений не всегда будет одна и та же, и для нахождения предельных режимов необходимо исследование системы (5.9) при конкретных значениях входящих в нее числовых параметров и конкретном виде функции
) невозможны. Но, во всяком случае, картина предельного поведения решений не всегда будет одна и та же, и для нахождения предельных режимов необходимо исследование системы (5.9) при конкретных значениях входящих в нее числовых параметров и конкретном виде функции 
 задающая относительный прирост численности хищников в единицу времени в зависимости от числа травоядных, имеет вид:
задающая относительный прирост численности хищников в единицу времени в зависимости от числа травоядных, имеет вид:

 равны соответственно 0,001 и 1000. При этом уравнения динамики средних (5.9) примут следующий вид:
равны соответственно 0,001 и 1000. При этом уравнения динамики средних (5.9) примут следующий вид:

 («критическое» число животных вида В, при котором животные вида А в среднем не размножаются и не вымирают), в данном случае также равно 1000.
(«критическое» число животных вида В, при котором животные вида А в среднем не размножаются и не вымирают), в данном случае также равно 1000.

 и травоядных
и травоядных  от времени
от времени  на рис. 6.29 показан устойчивый предельный цикл. Цикл размечен по времени; деления соответствуют
на рис. 6.29 показан устойчивый предельный цикл. Цикл размечен по времени; деления соответствуют  При
При  (период цикла)
(период цикла)  возвращаются к первоначальным значениям.
возвращаются к первоначальным значениям.


