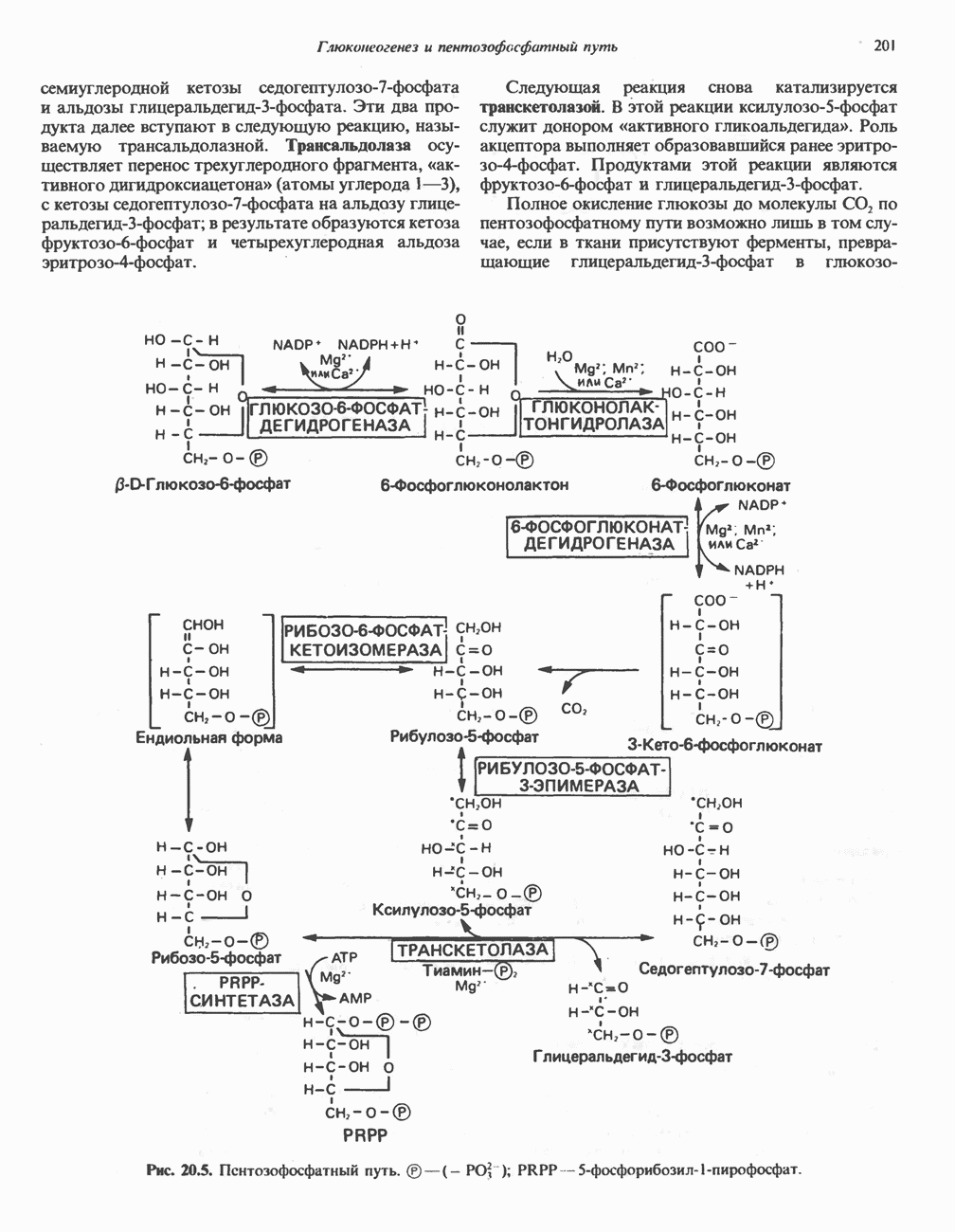
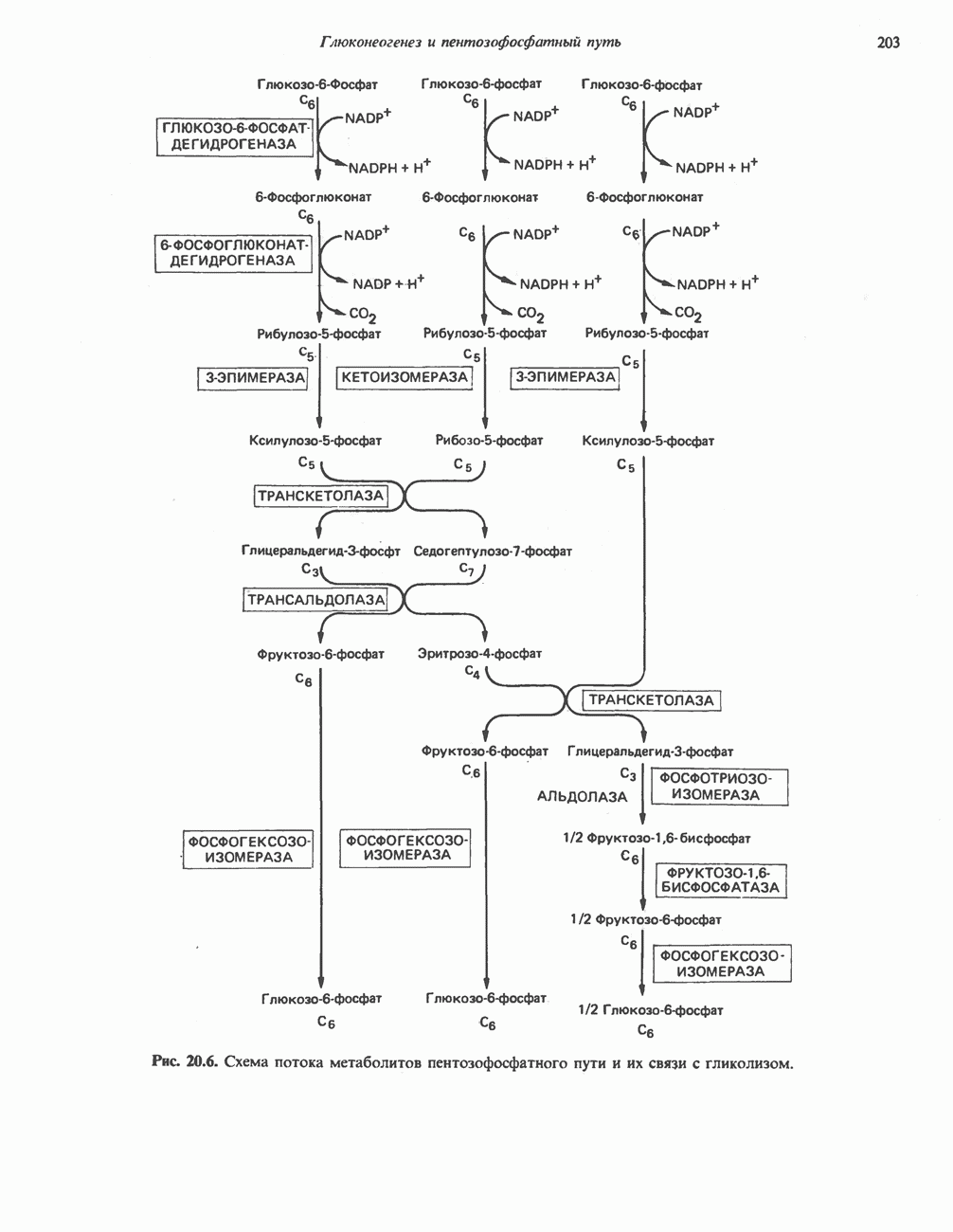
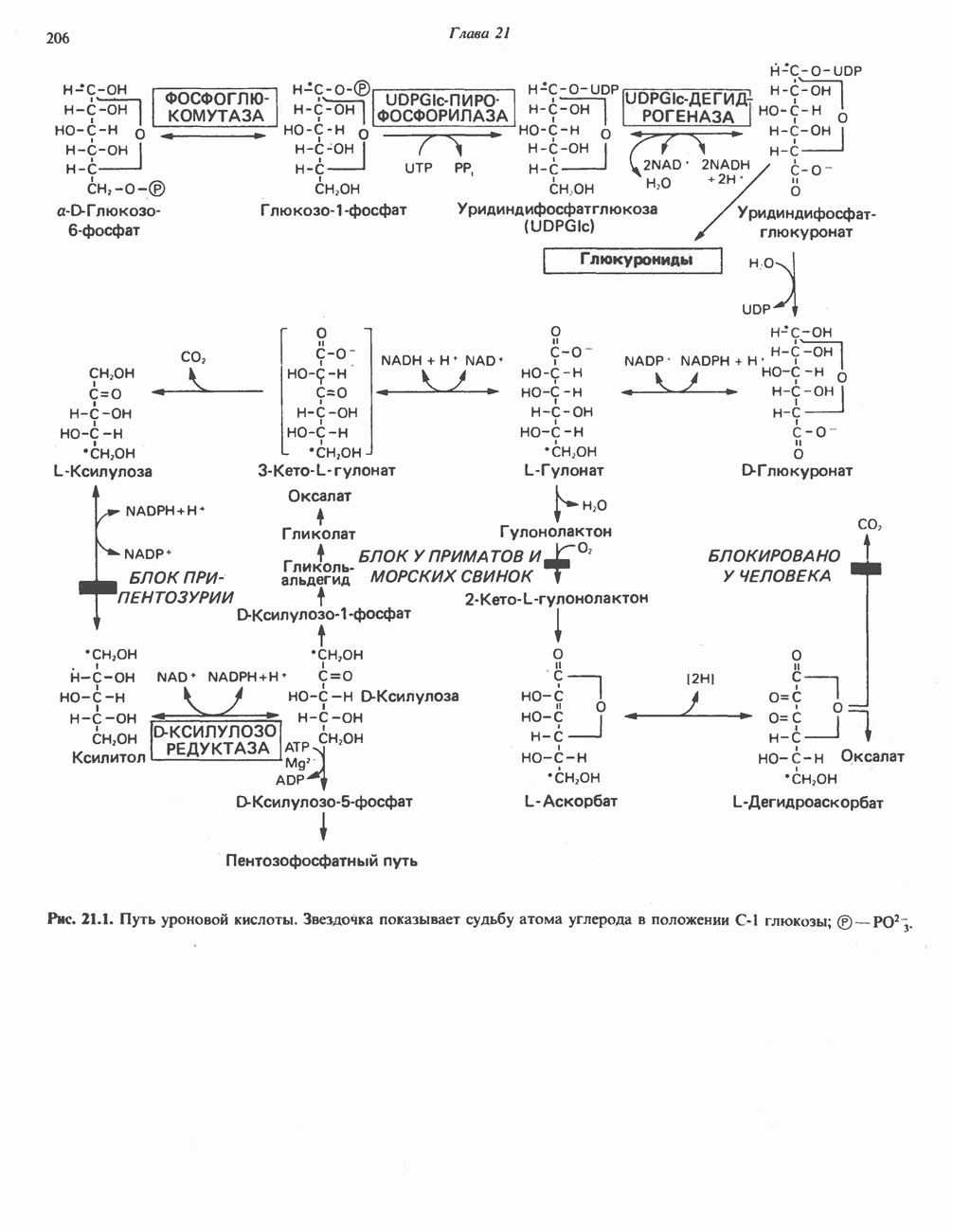
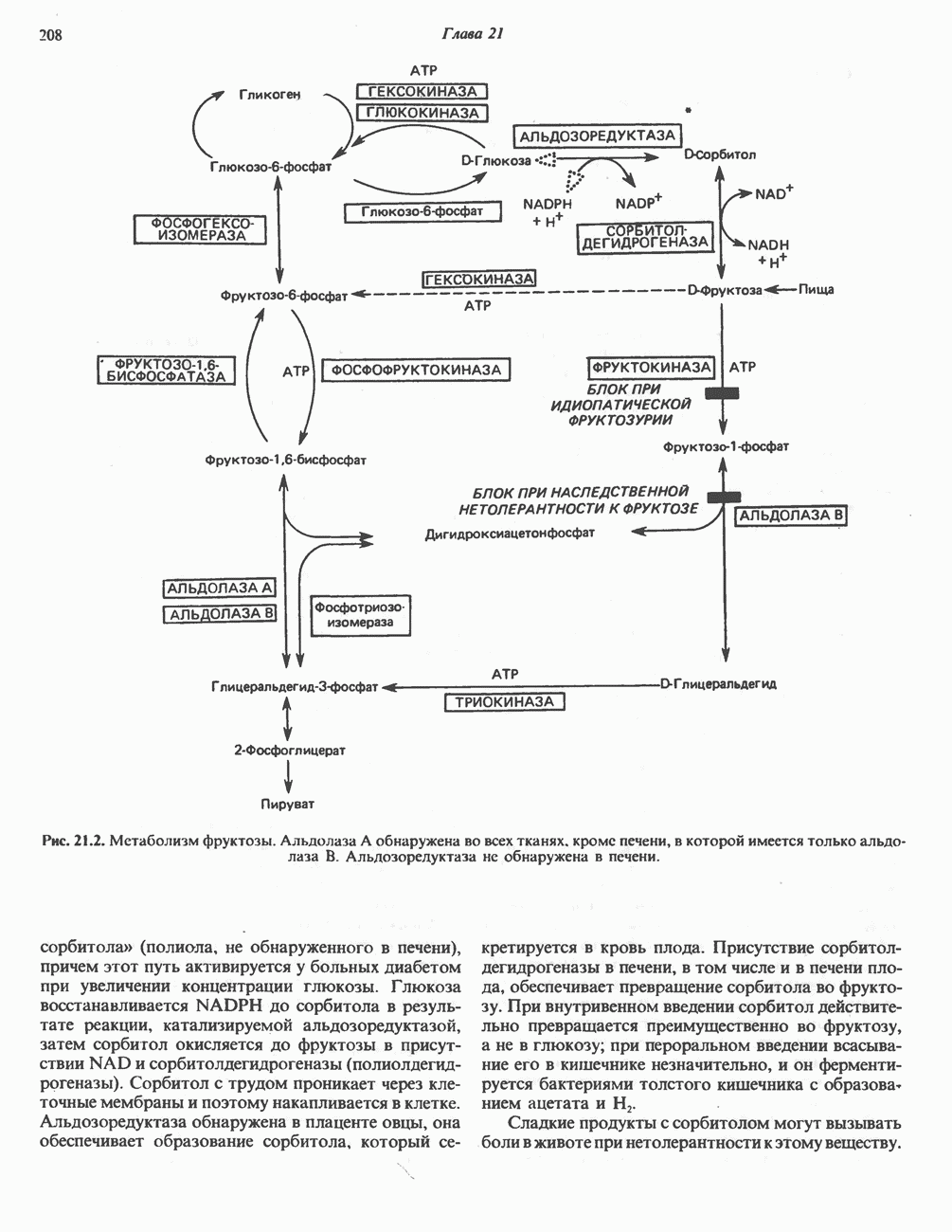
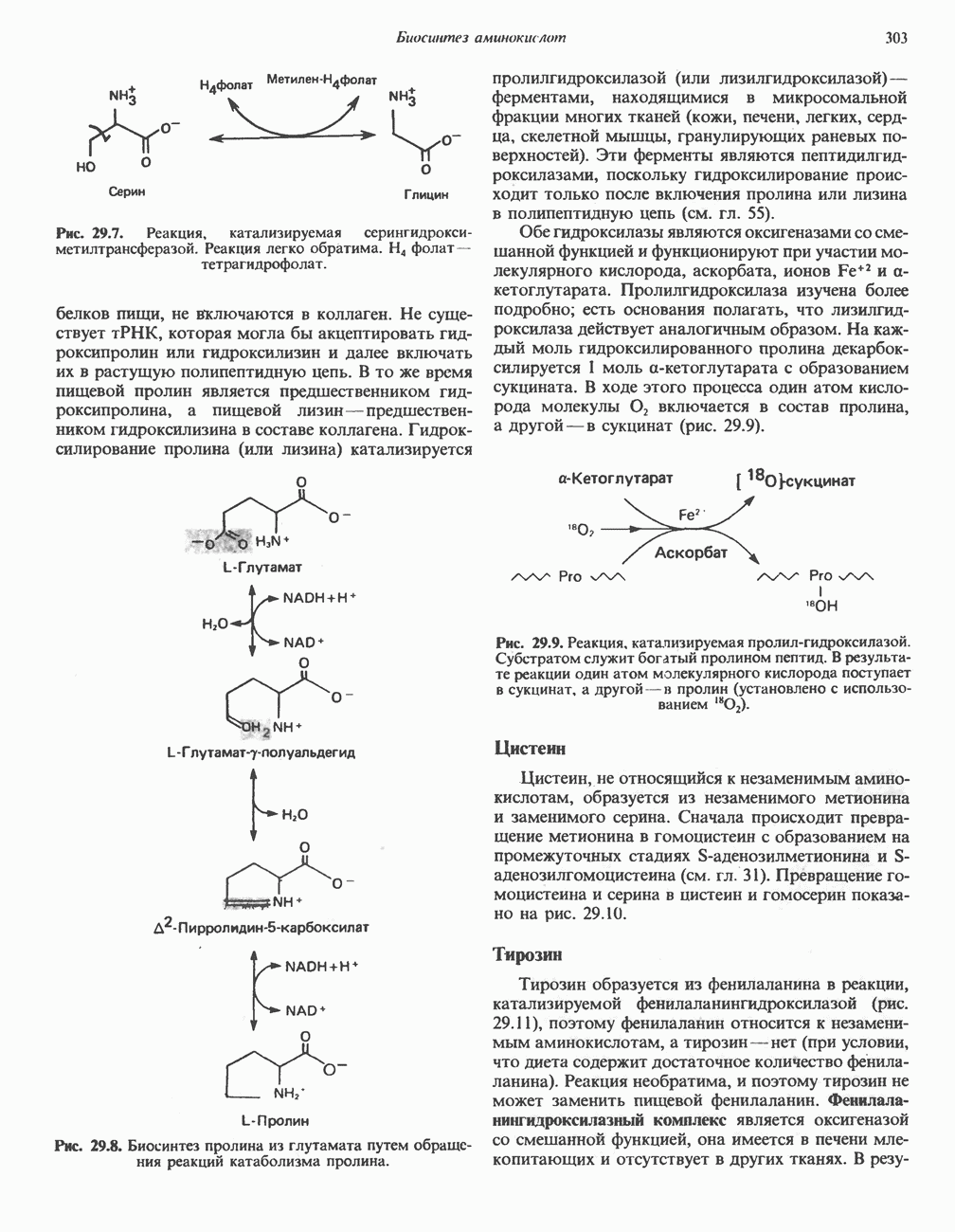
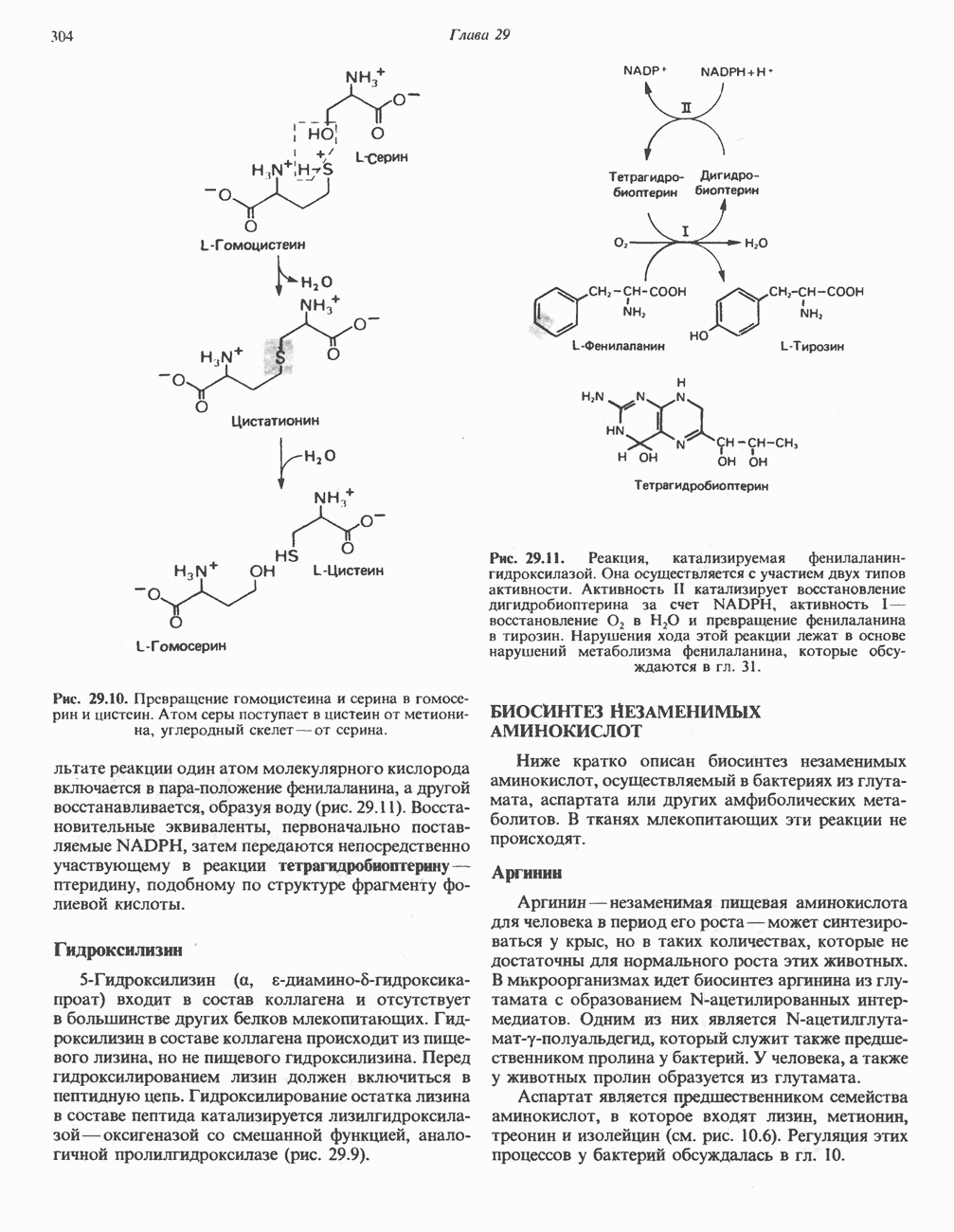
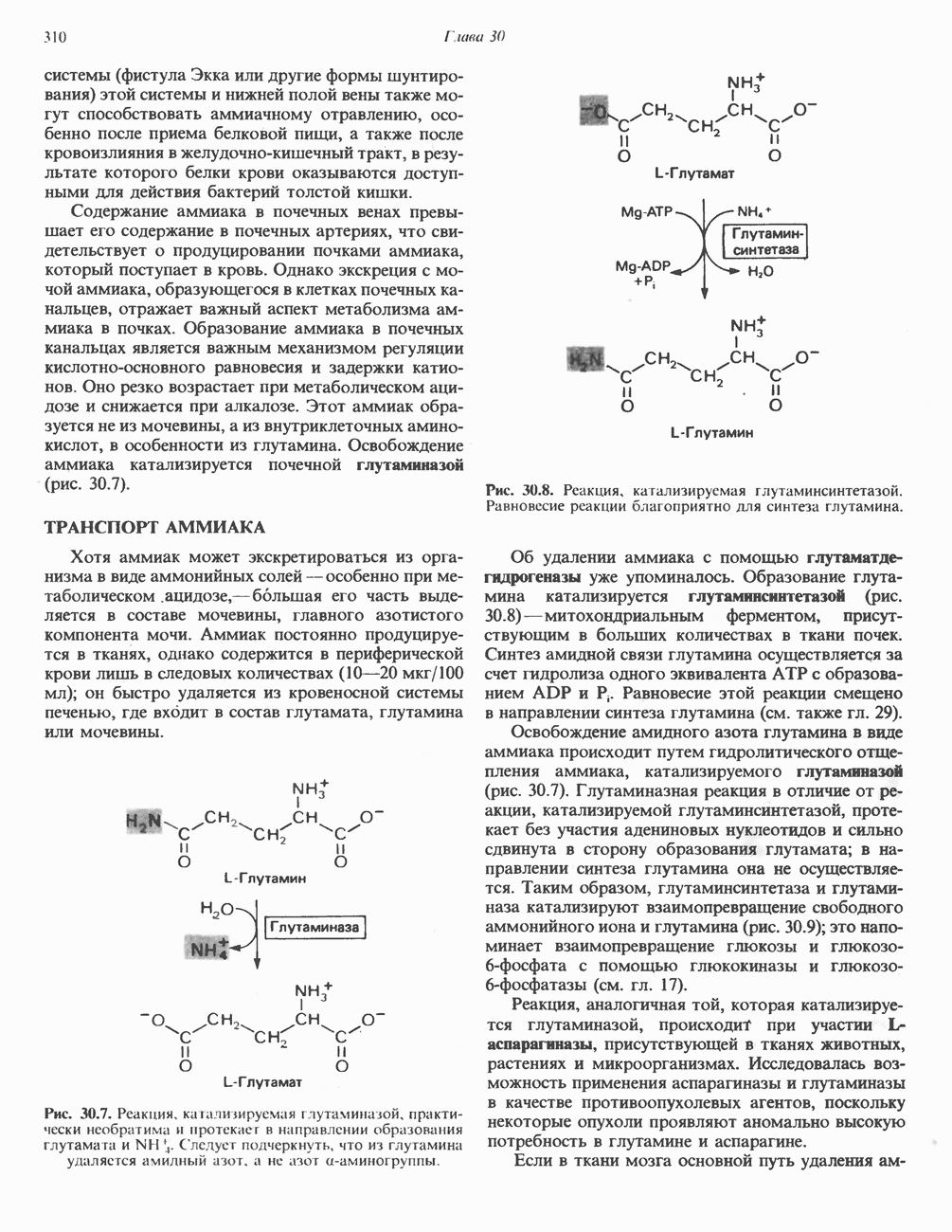
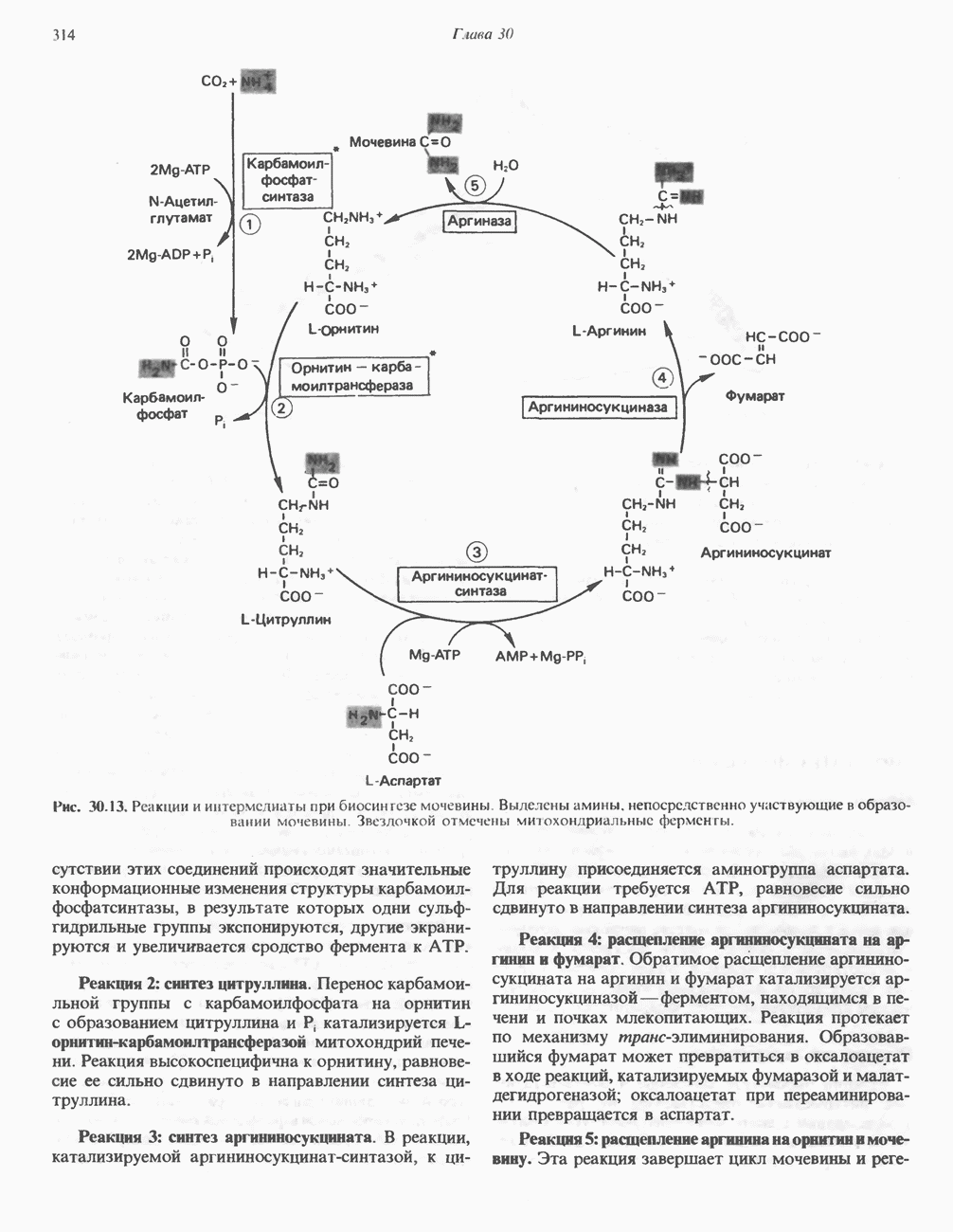
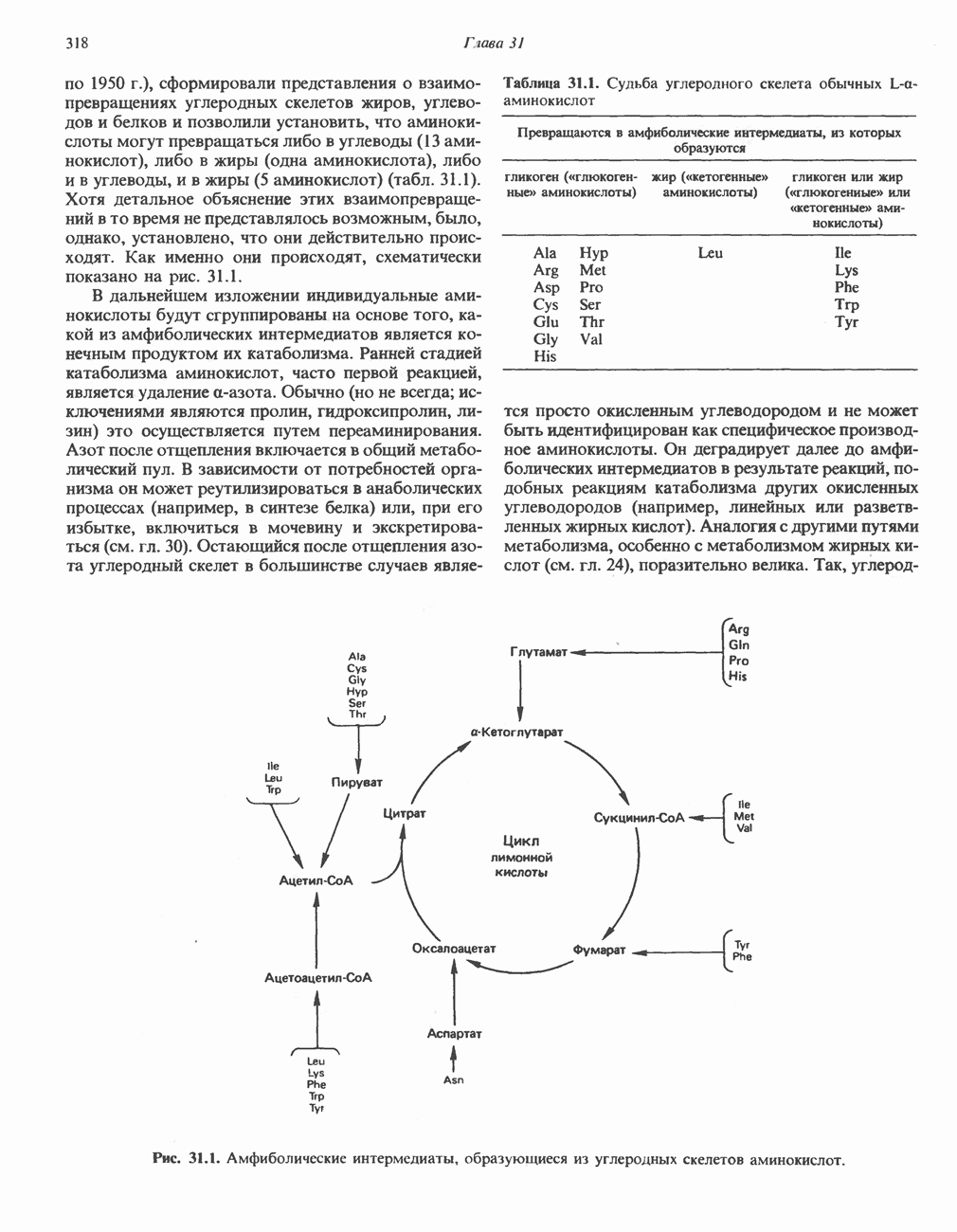
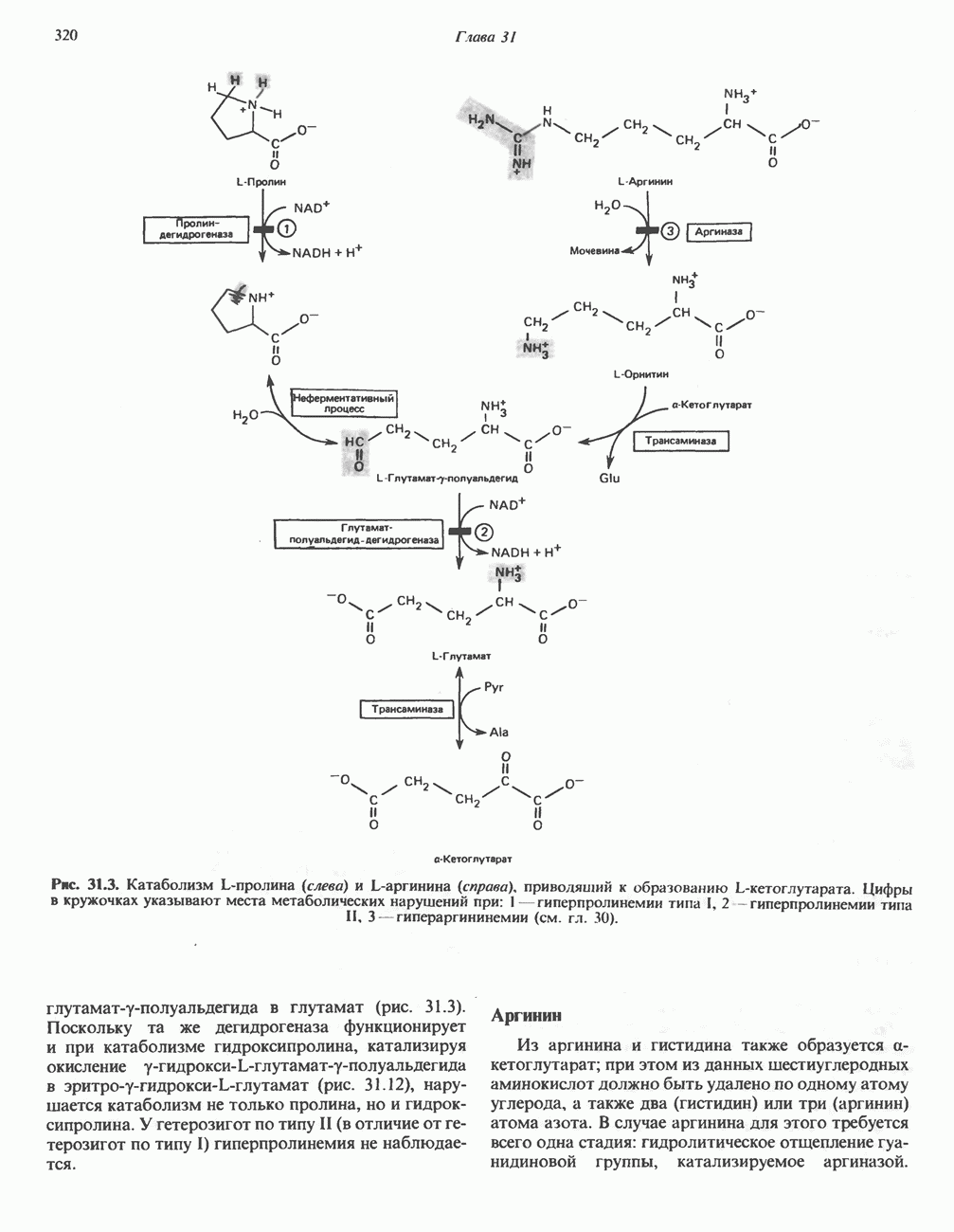
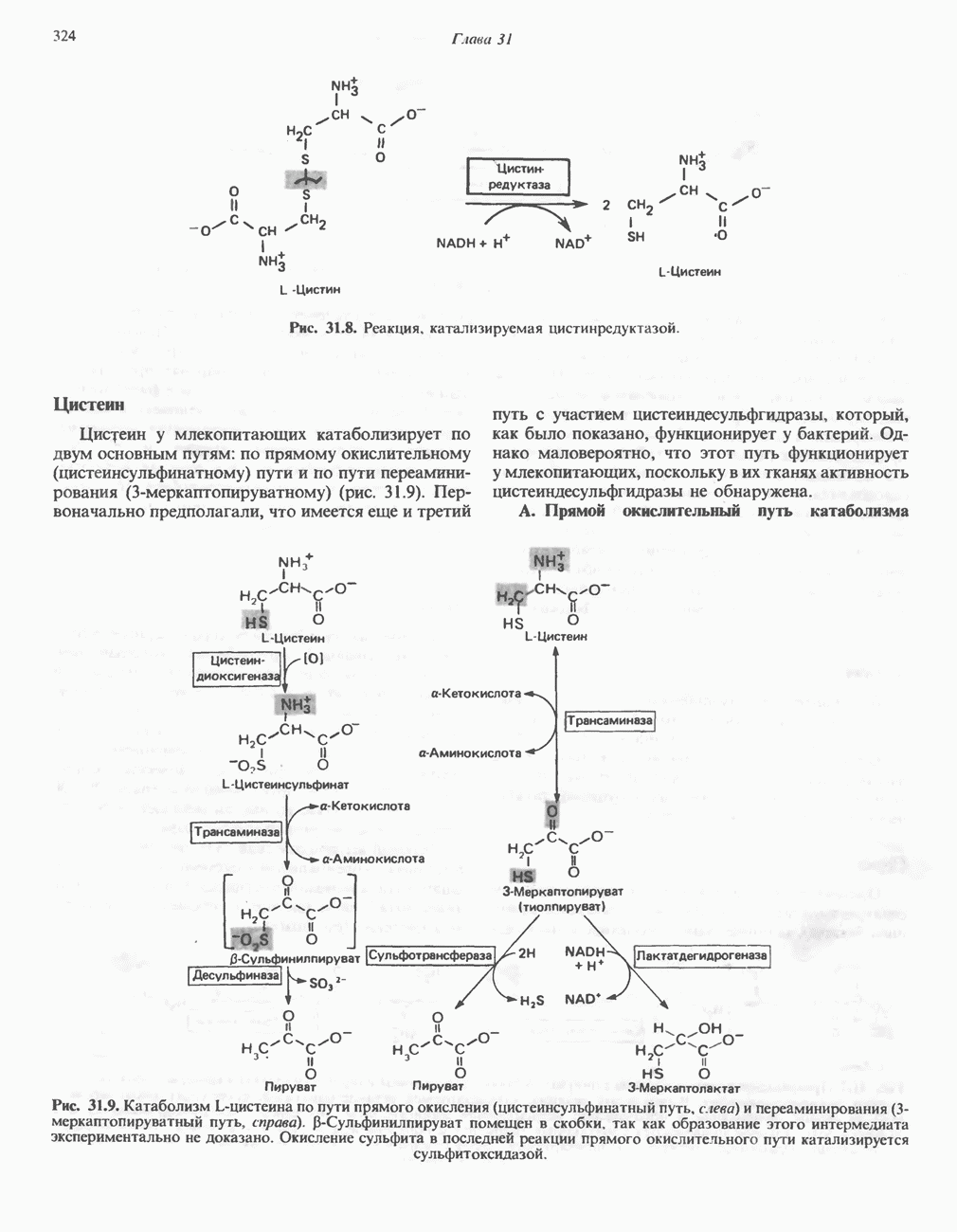
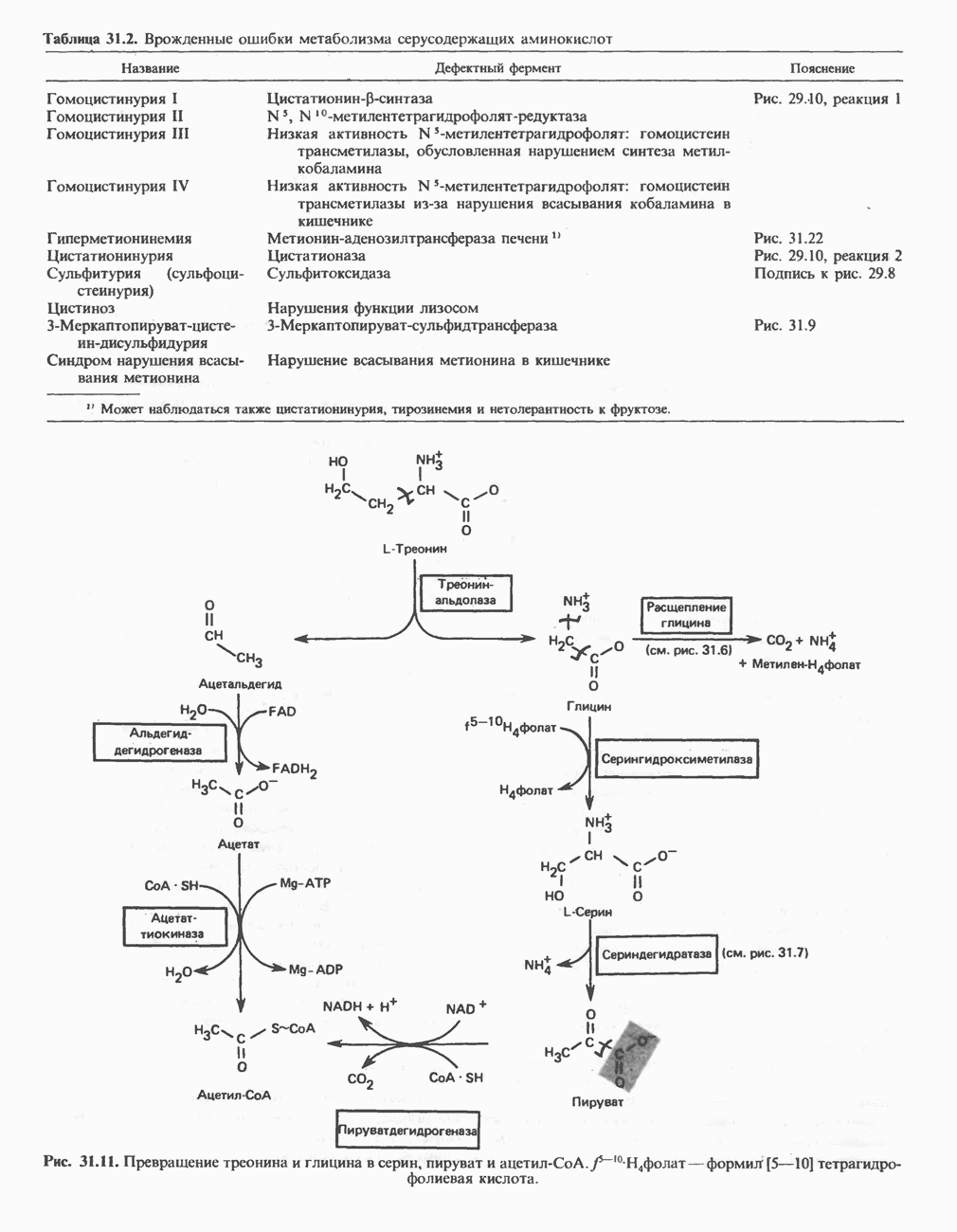
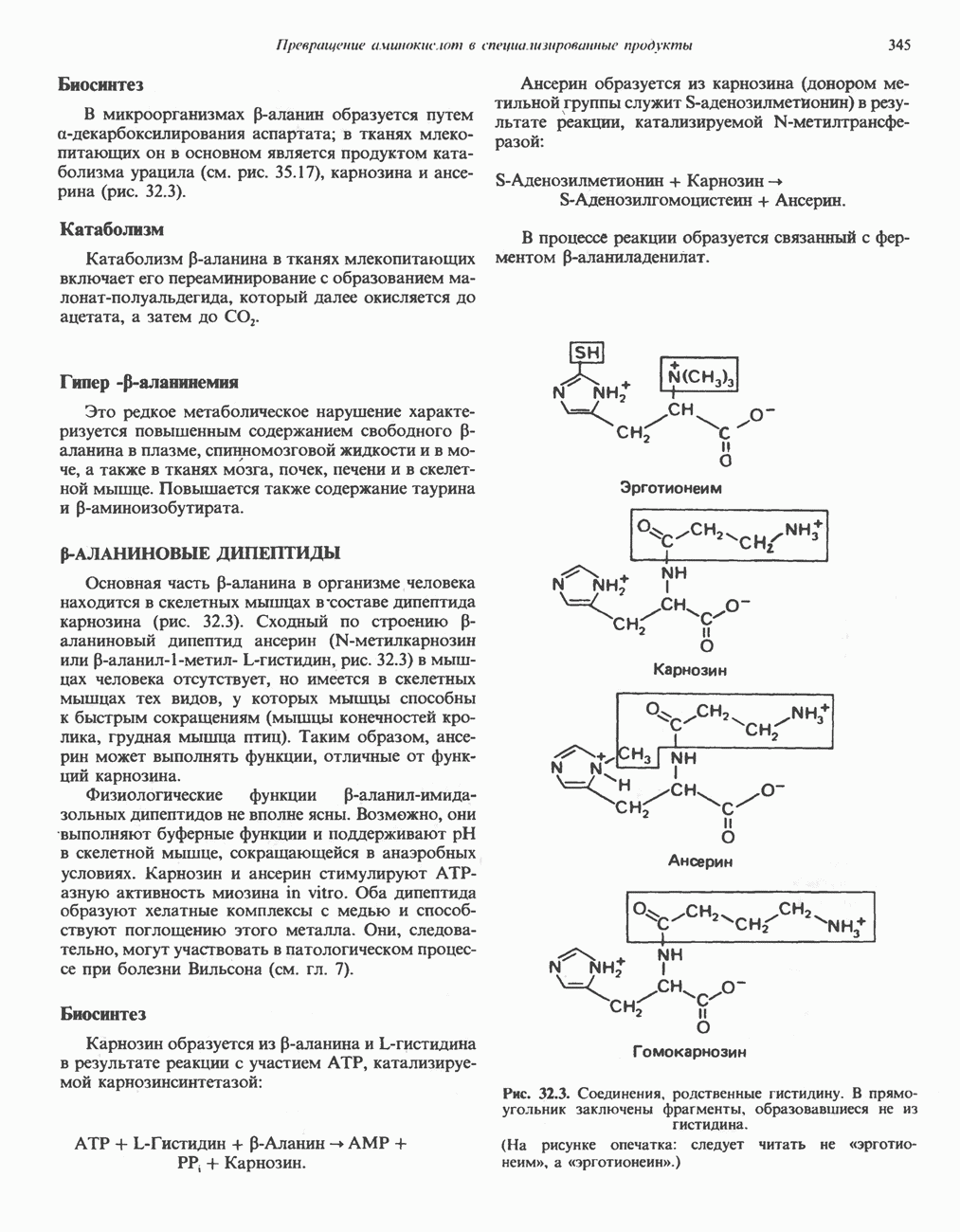
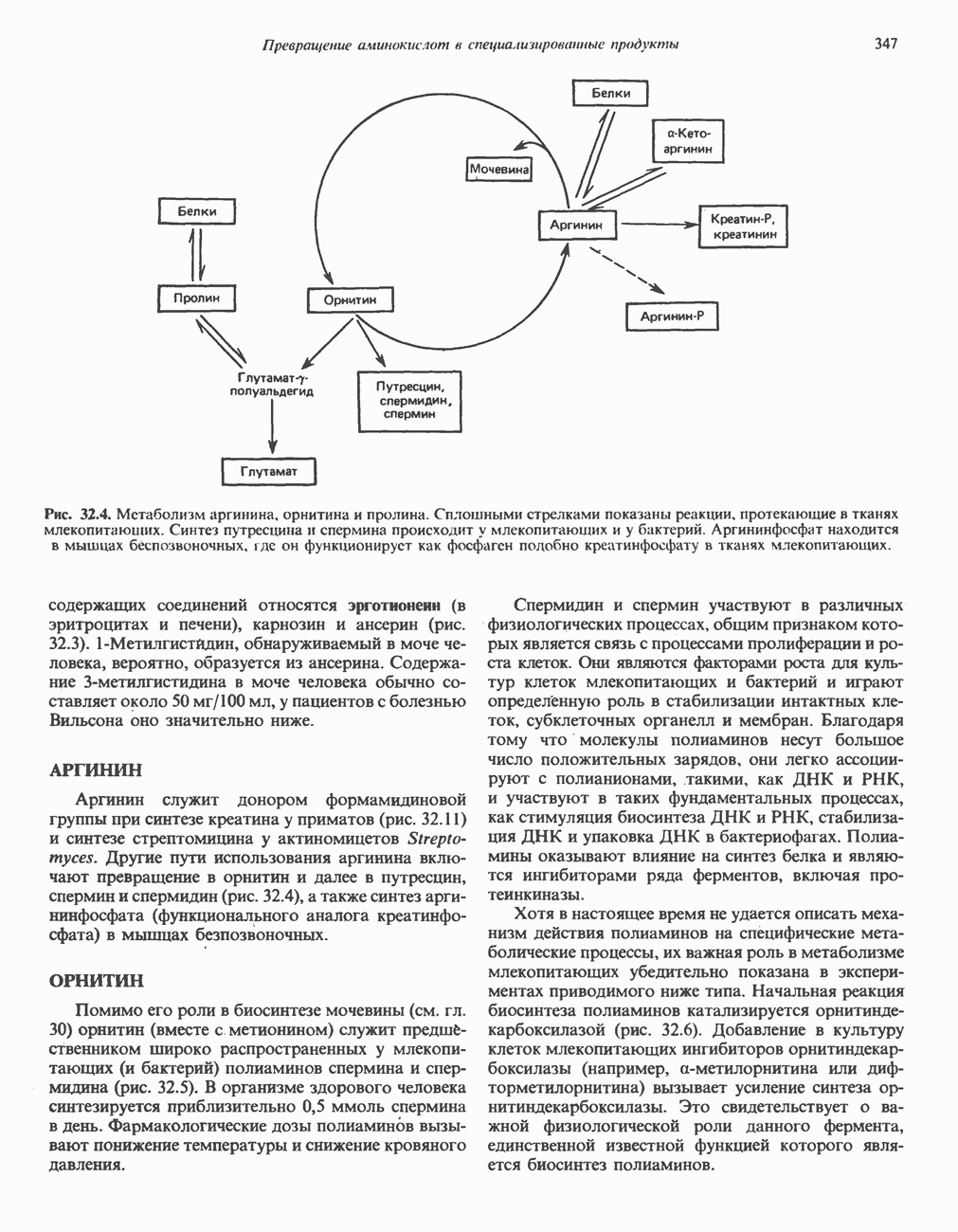
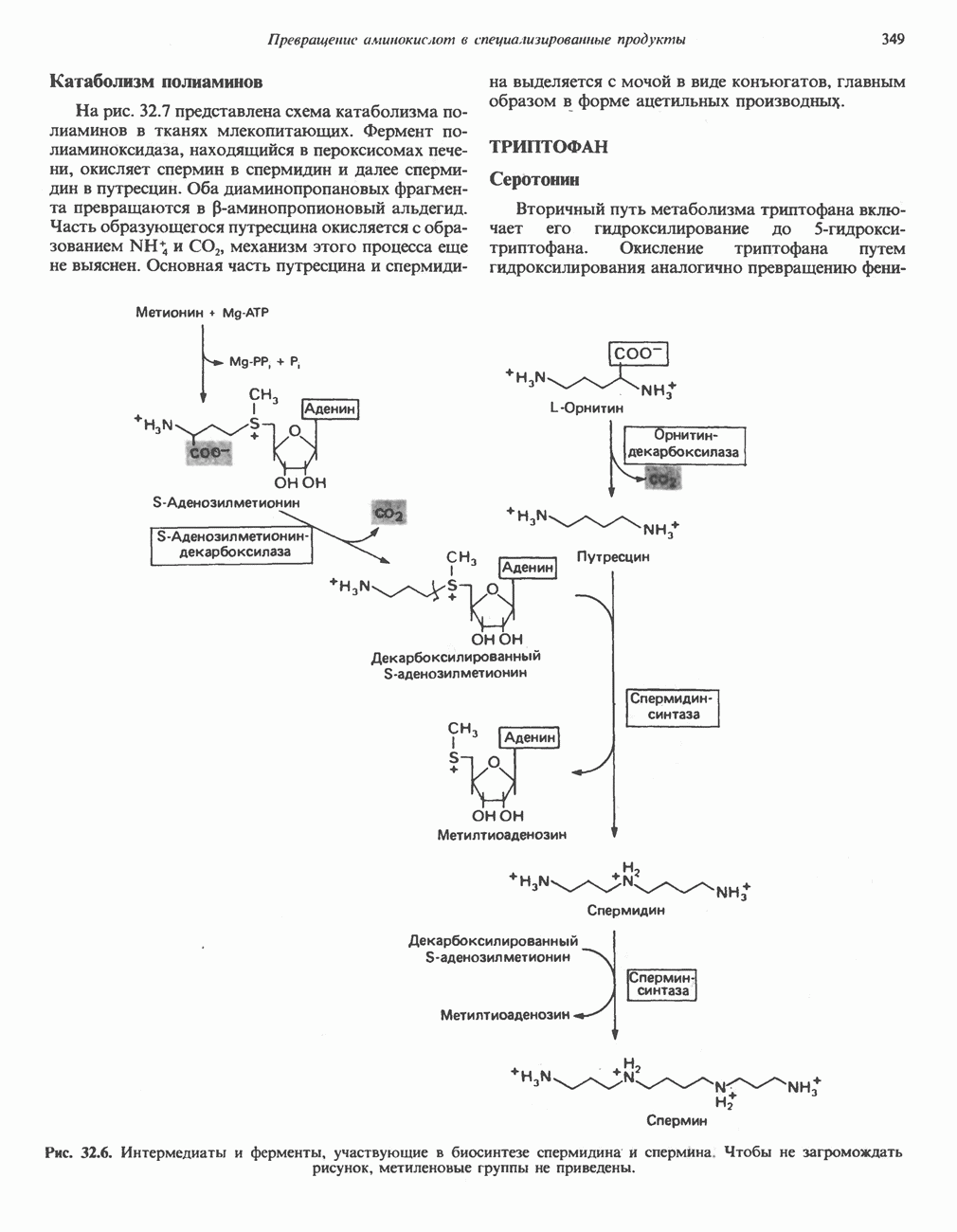
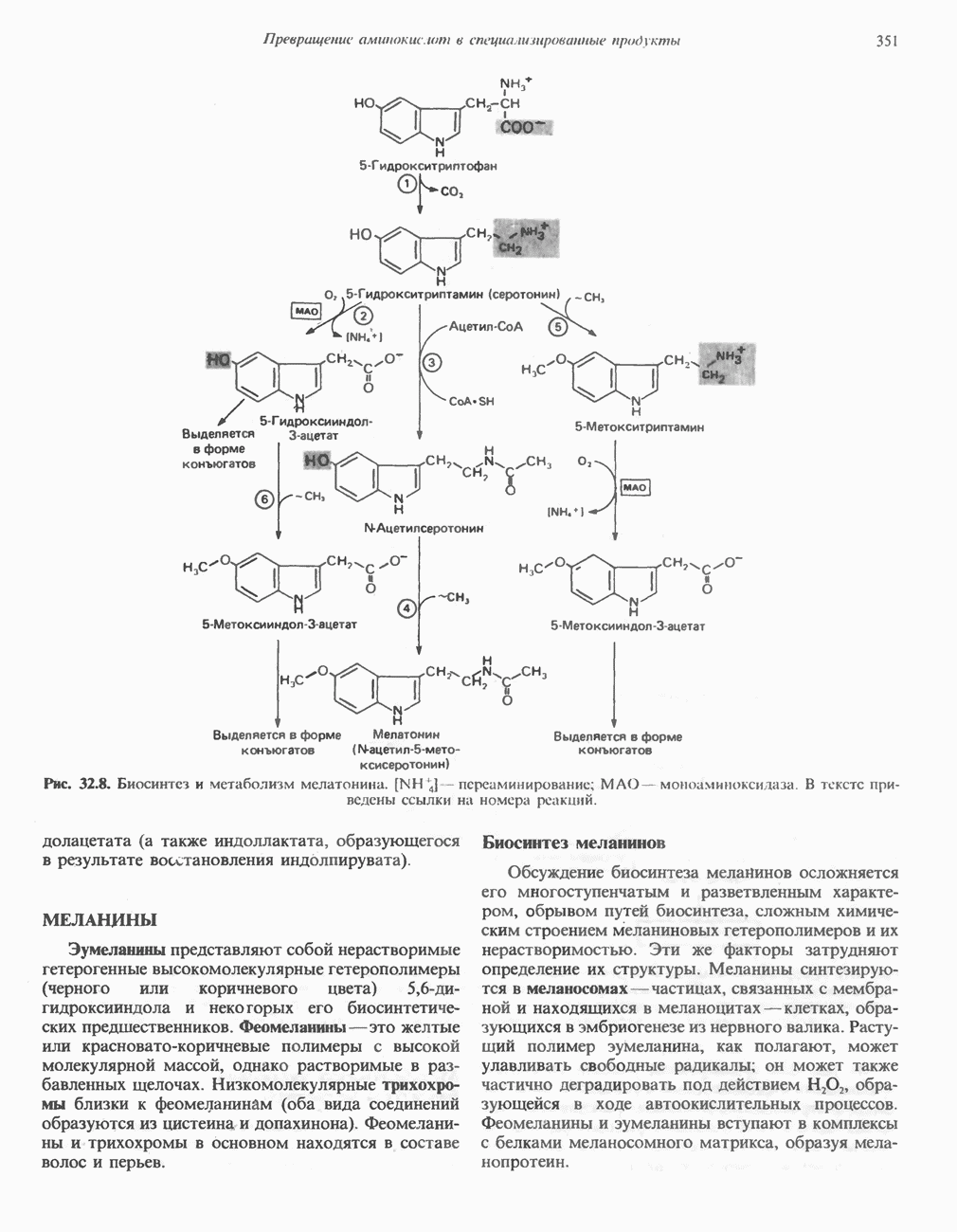
Пред.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 145 146 147 148 149 150 151 152 153 154 155 156 157 158 159 160 161 162 163 164 165 166 167 168 169 170 171 172 173 174 175 176 177 178 179 180 181 182 183 184 185 186 187 188 189 190 191 192 193 194 195 196 197 198 199 200 201 202 203 204 205 206 207 208 209 210 211 212 213 214 215 216 217 218 219 220 221 222 223 224 225 226 227 228 229 230 231 232 233 234 235 236 237 238 239 240 241 242 243 244 245 246 247 248 249 250 251 252 253 254 255 256 257 258 259 260 261 262 263 264 265 266 267 268 269 270 271 272 273 274 275 276 277 278 279 280 281 282 283 284 285 286 287 288 289 290 291 292 293 294 295 296 297 298 299 300 301 302 303 304 305 306 307 308 309 310 311 312 313 314 315 316 317 318 319 320 321 322 323 324 325 326 327 328 329 330 331 332 333 334 335 336 337 338 339 340 341 342 343 344 345 346 347 348 349 350 351 352 353 354 355 356 357 358 359 360 361 362 363 364 365 366 367 368 369 370 След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
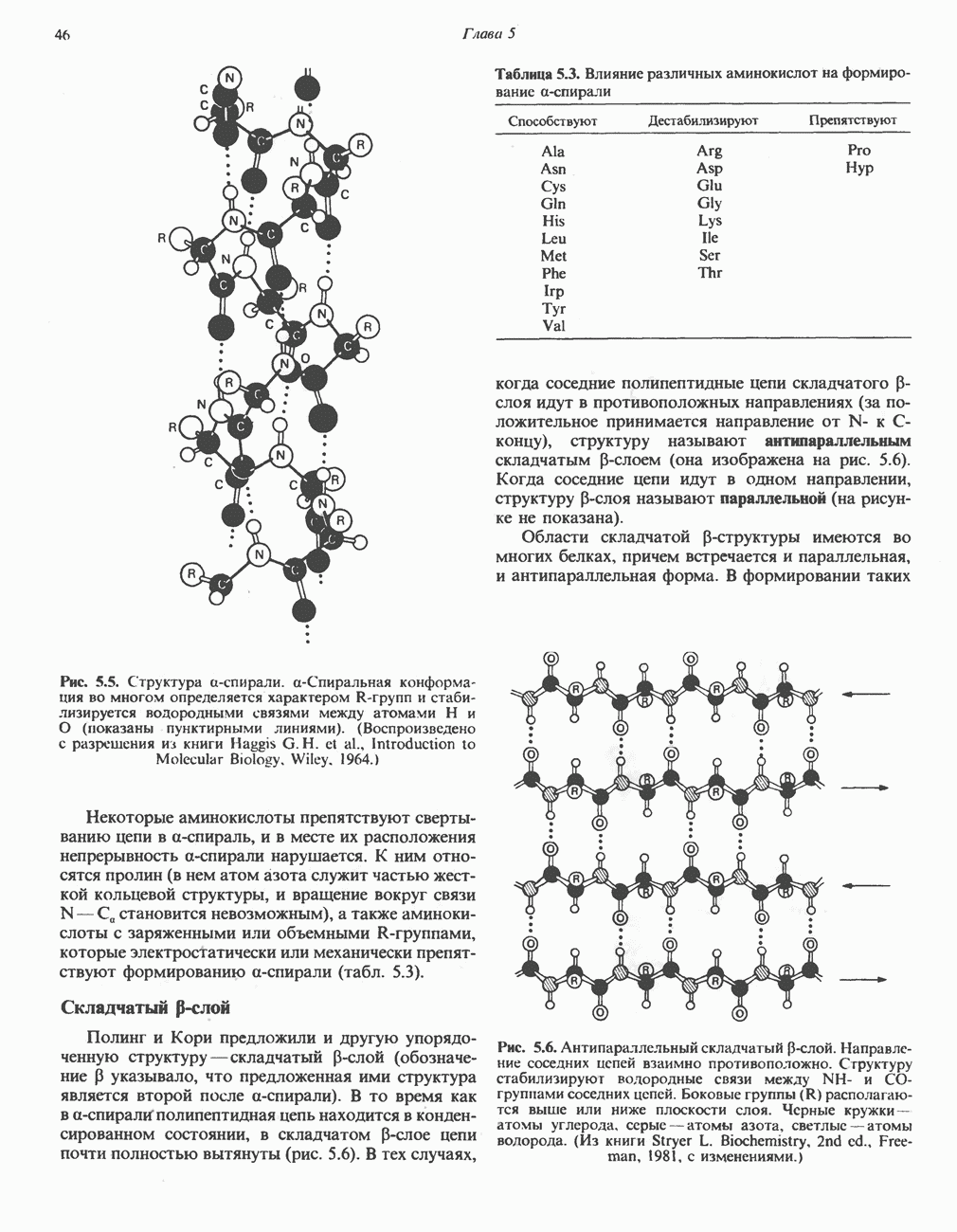
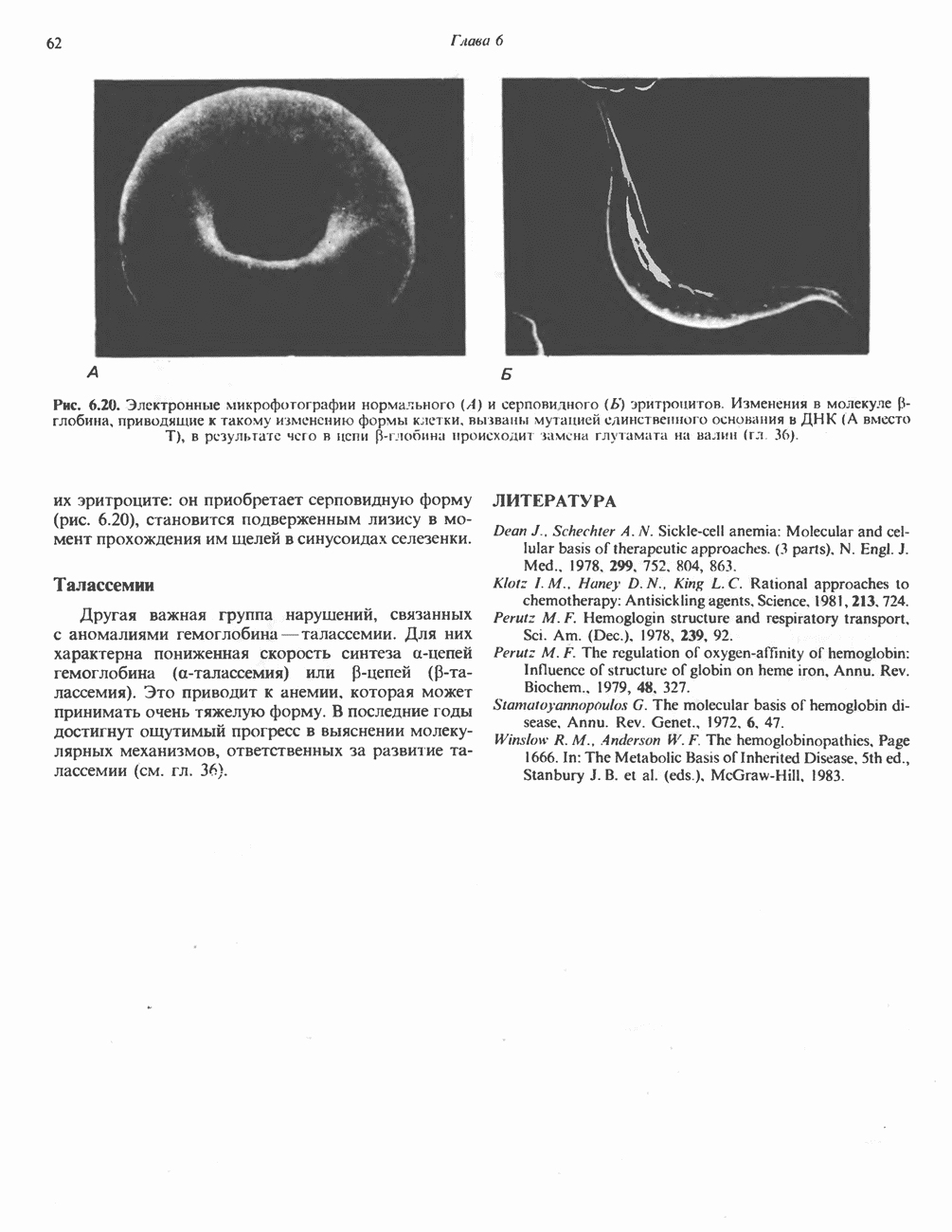
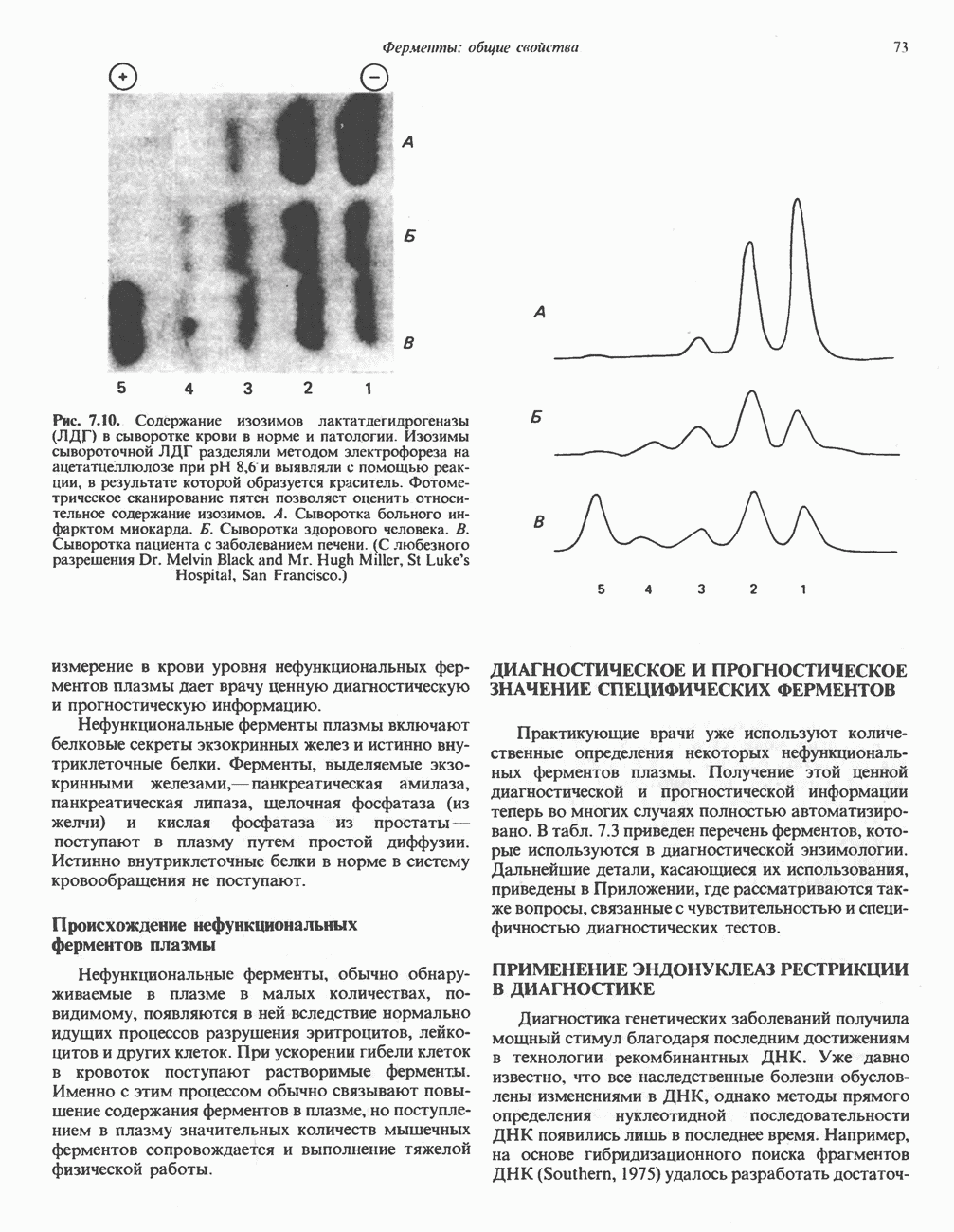
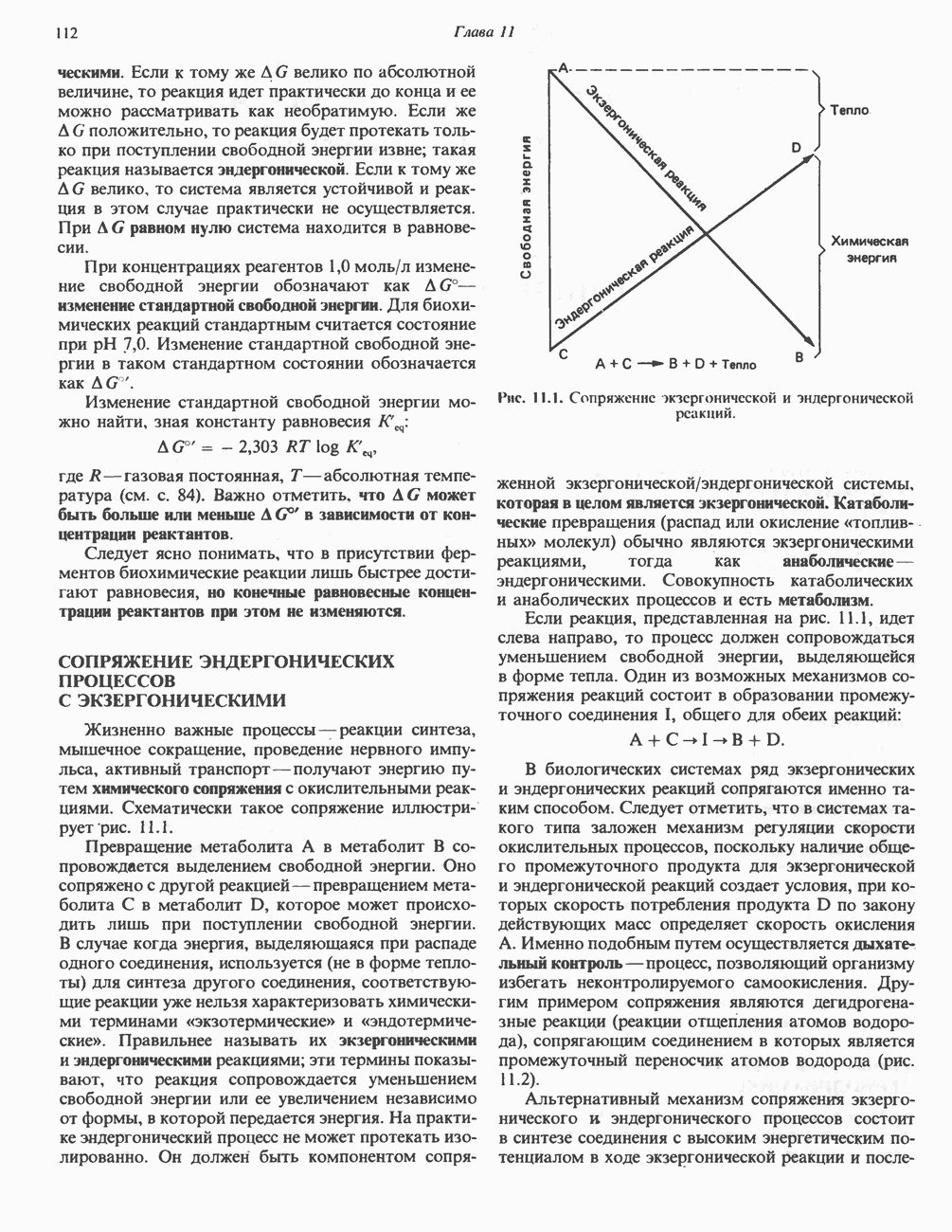
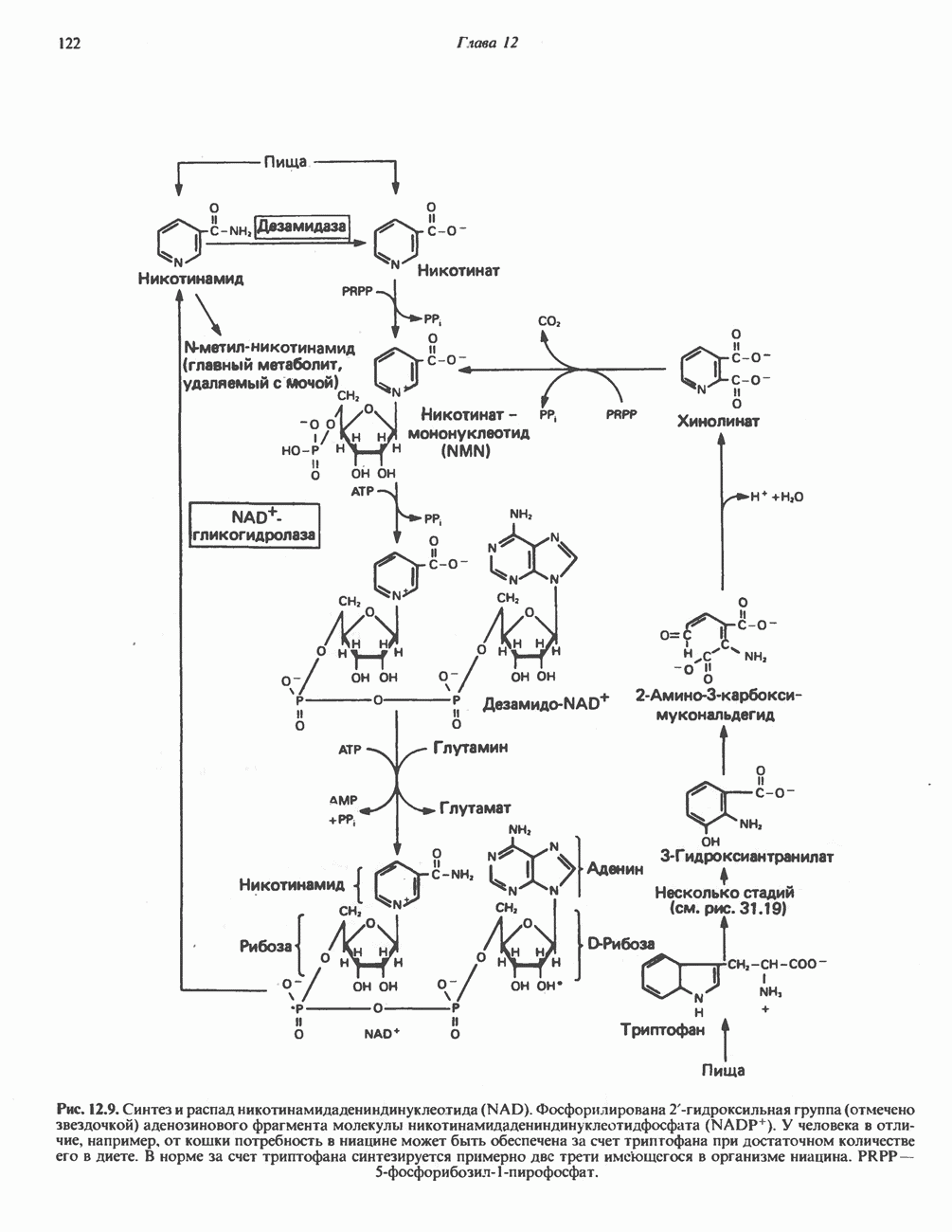
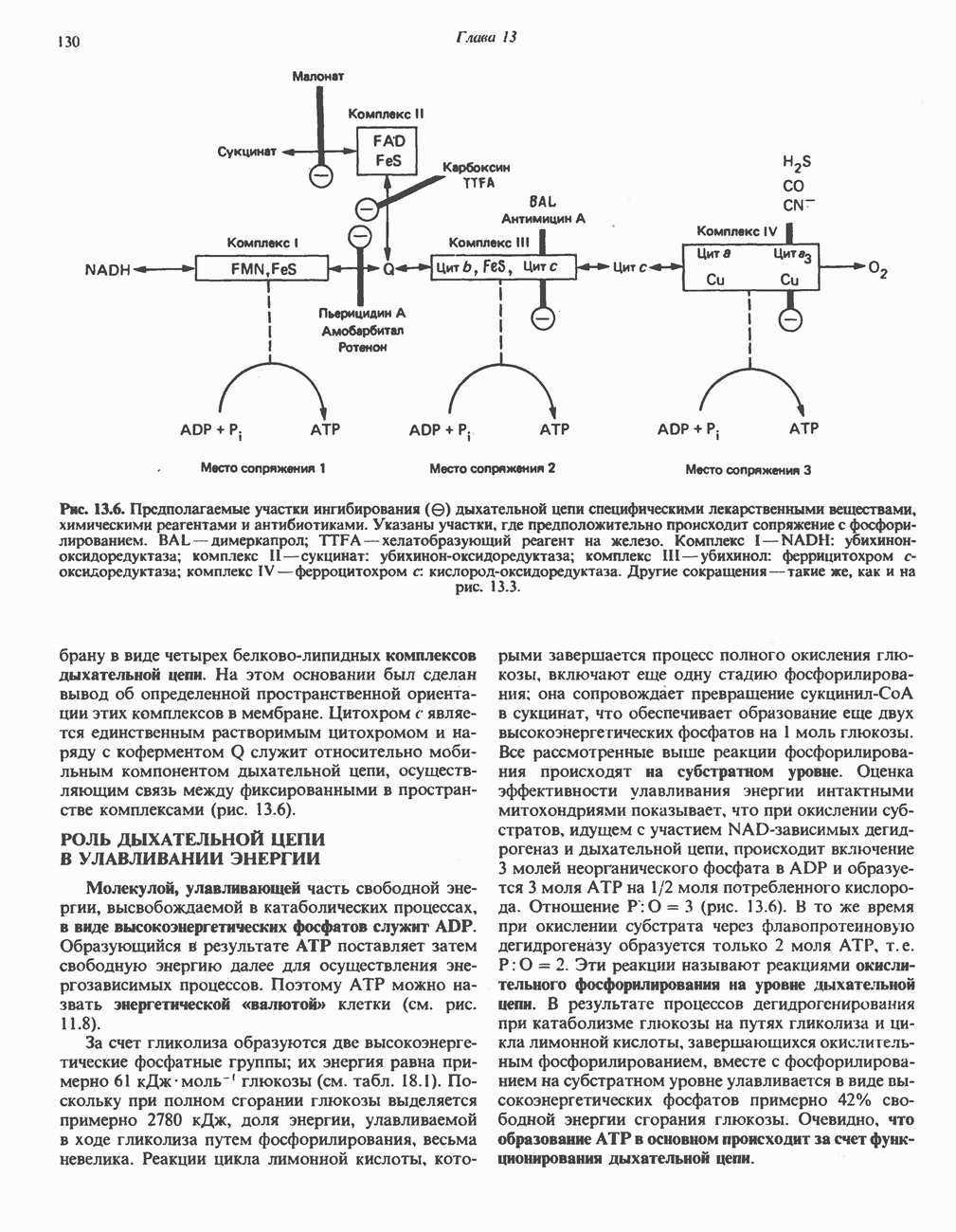
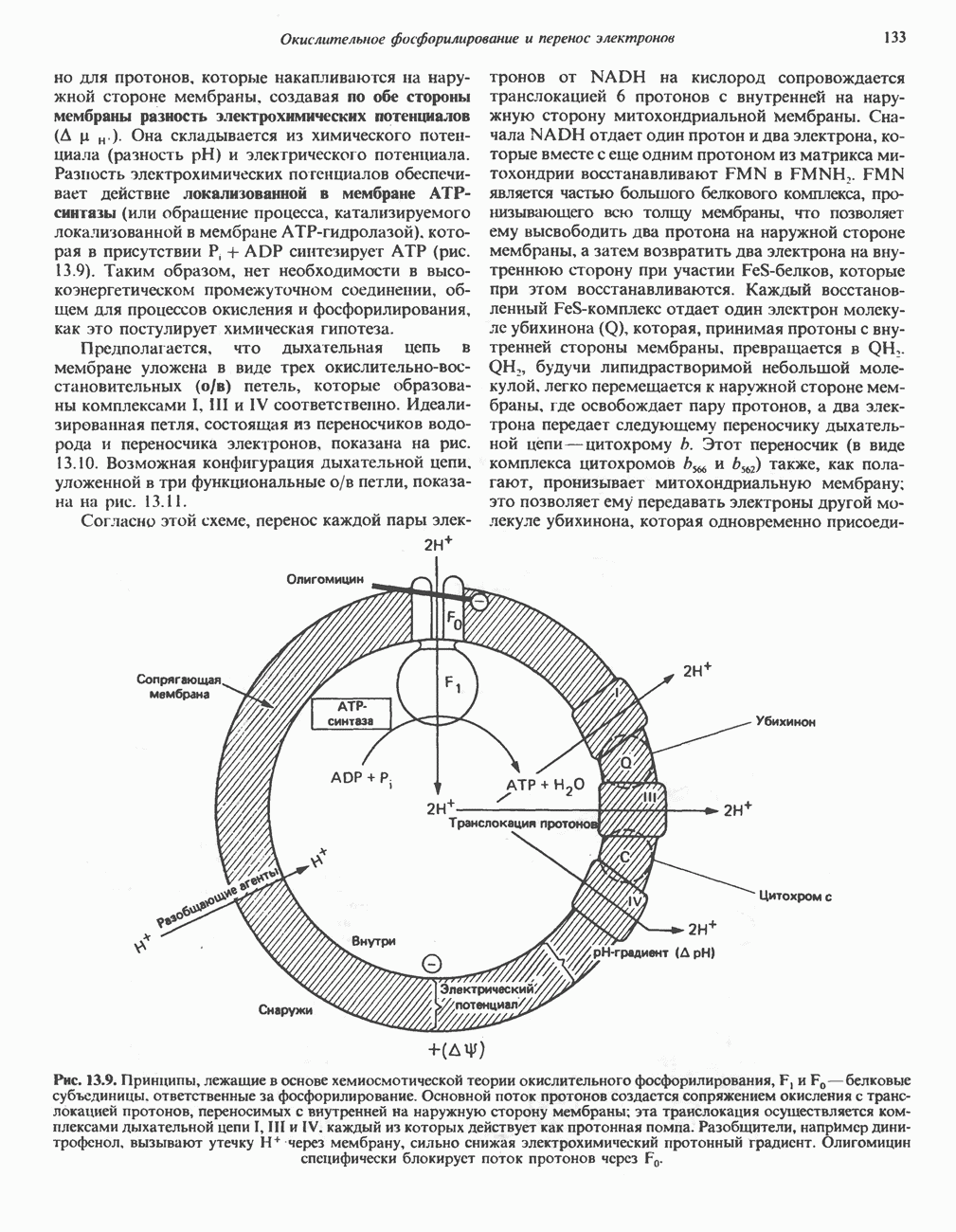
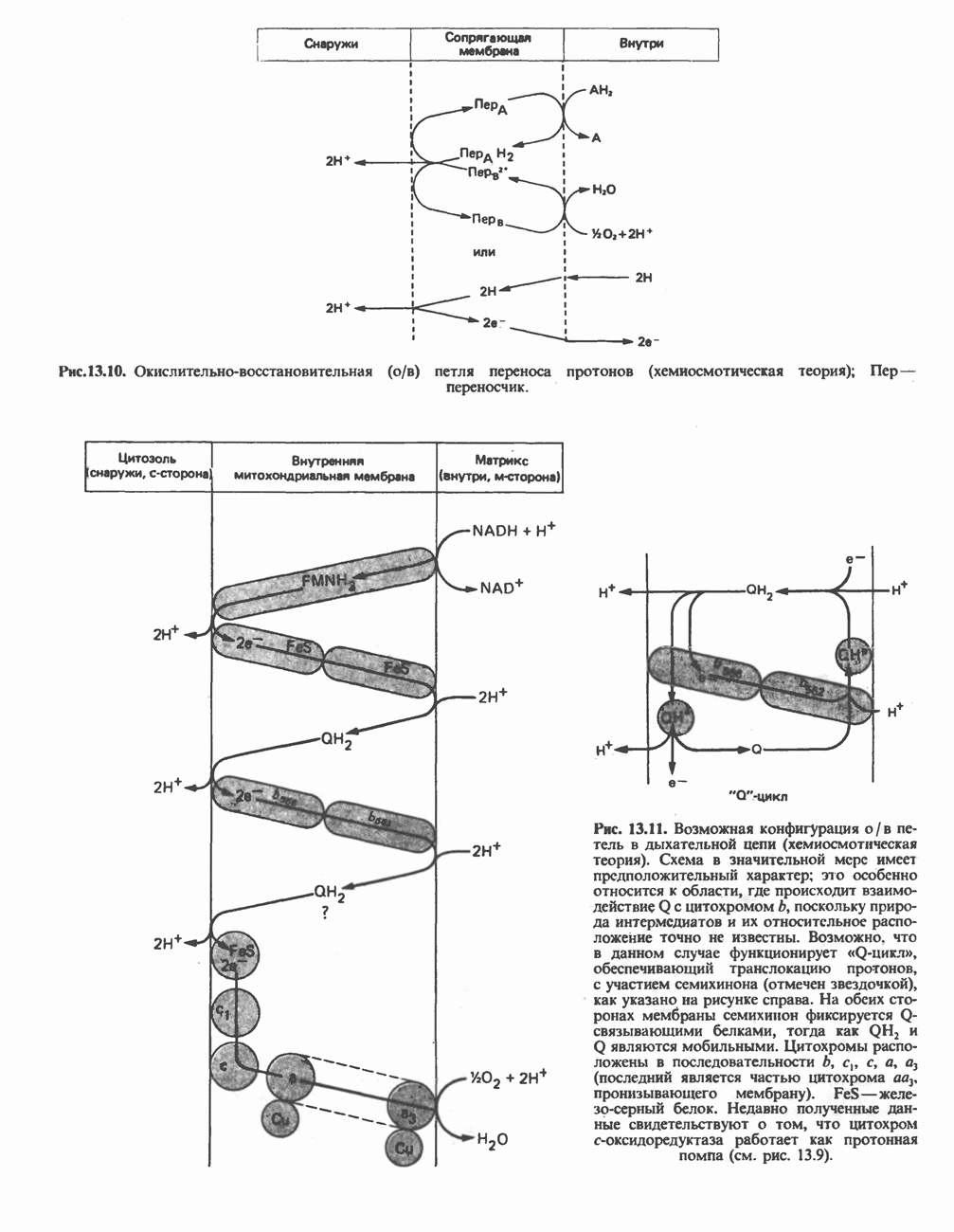
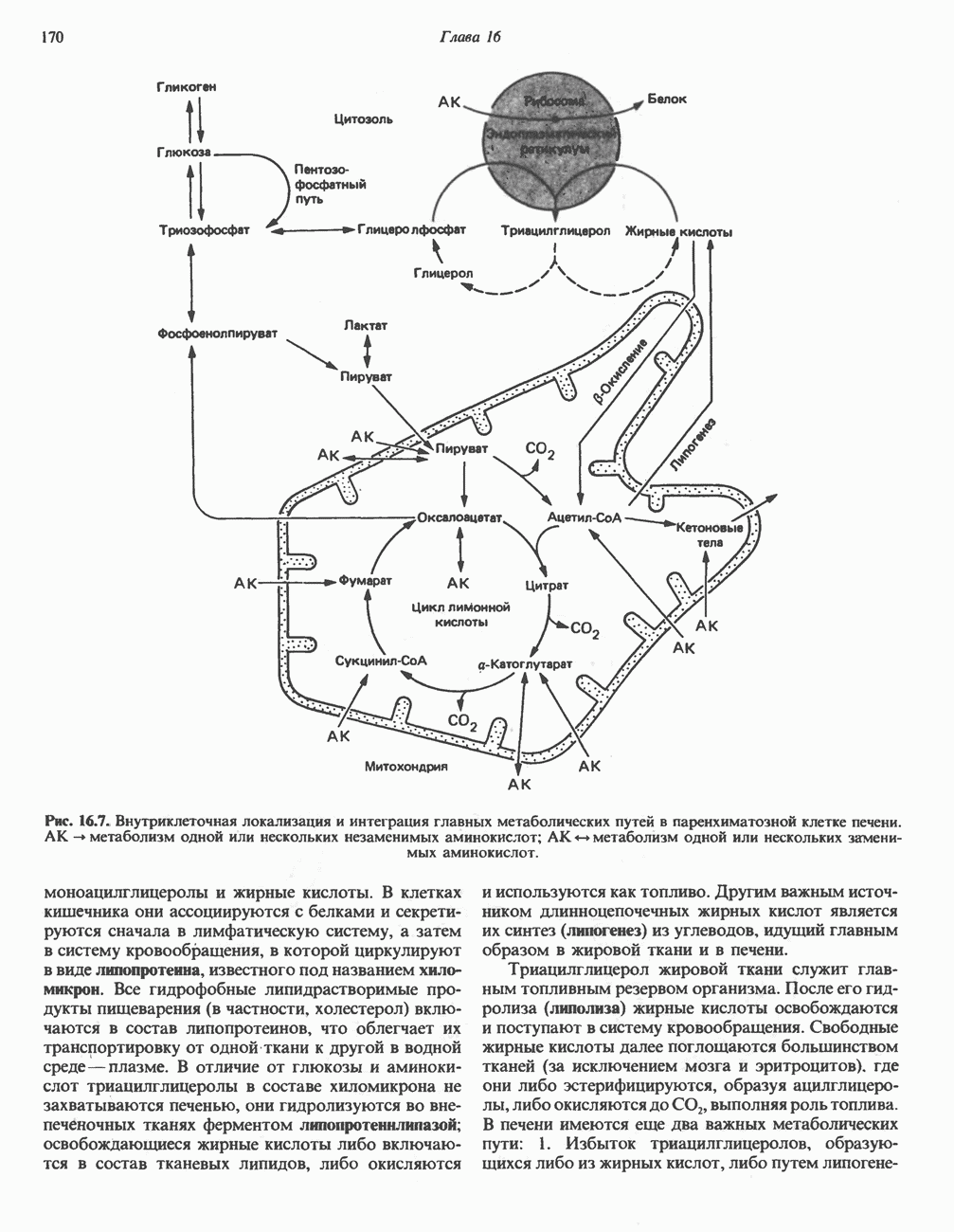
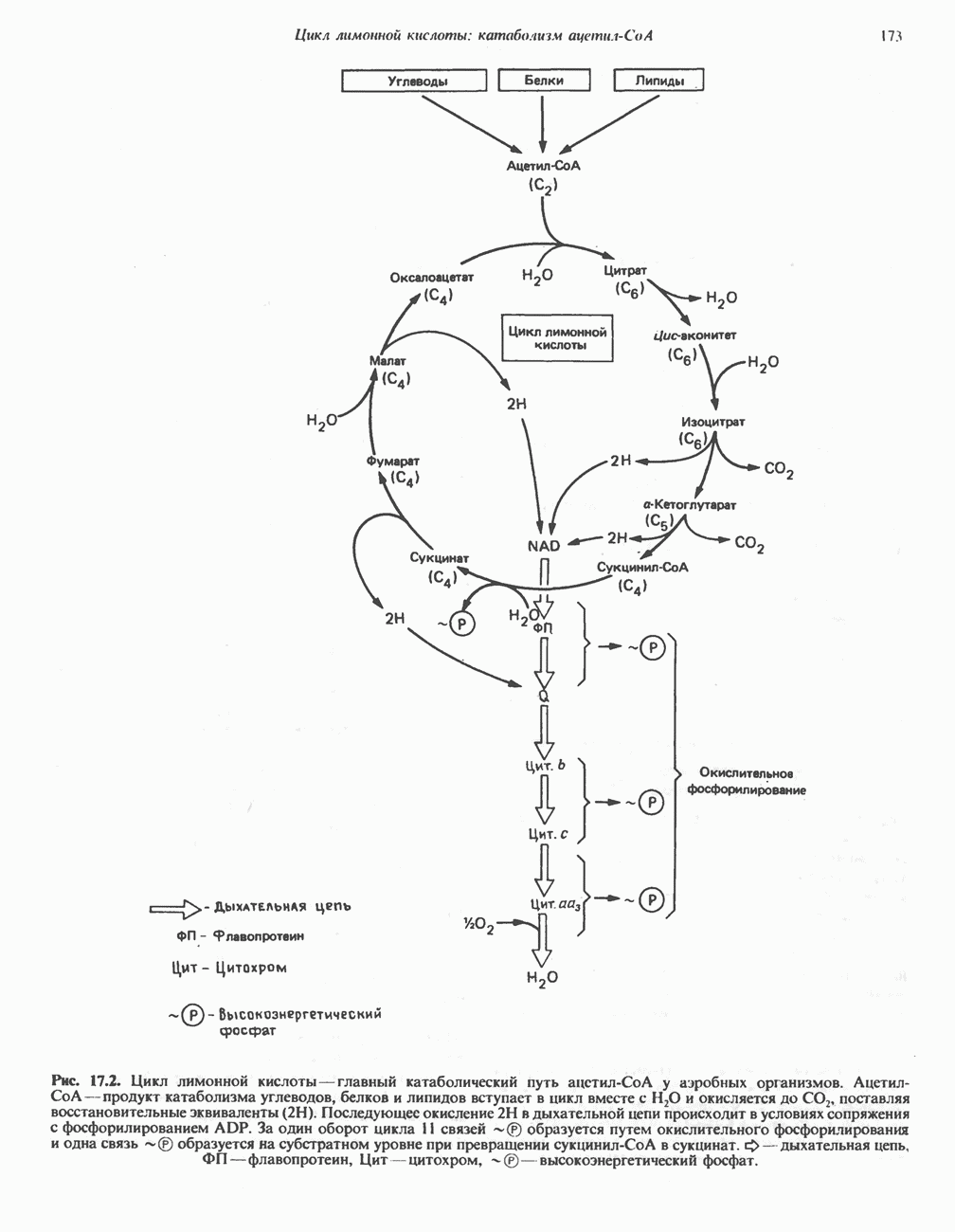
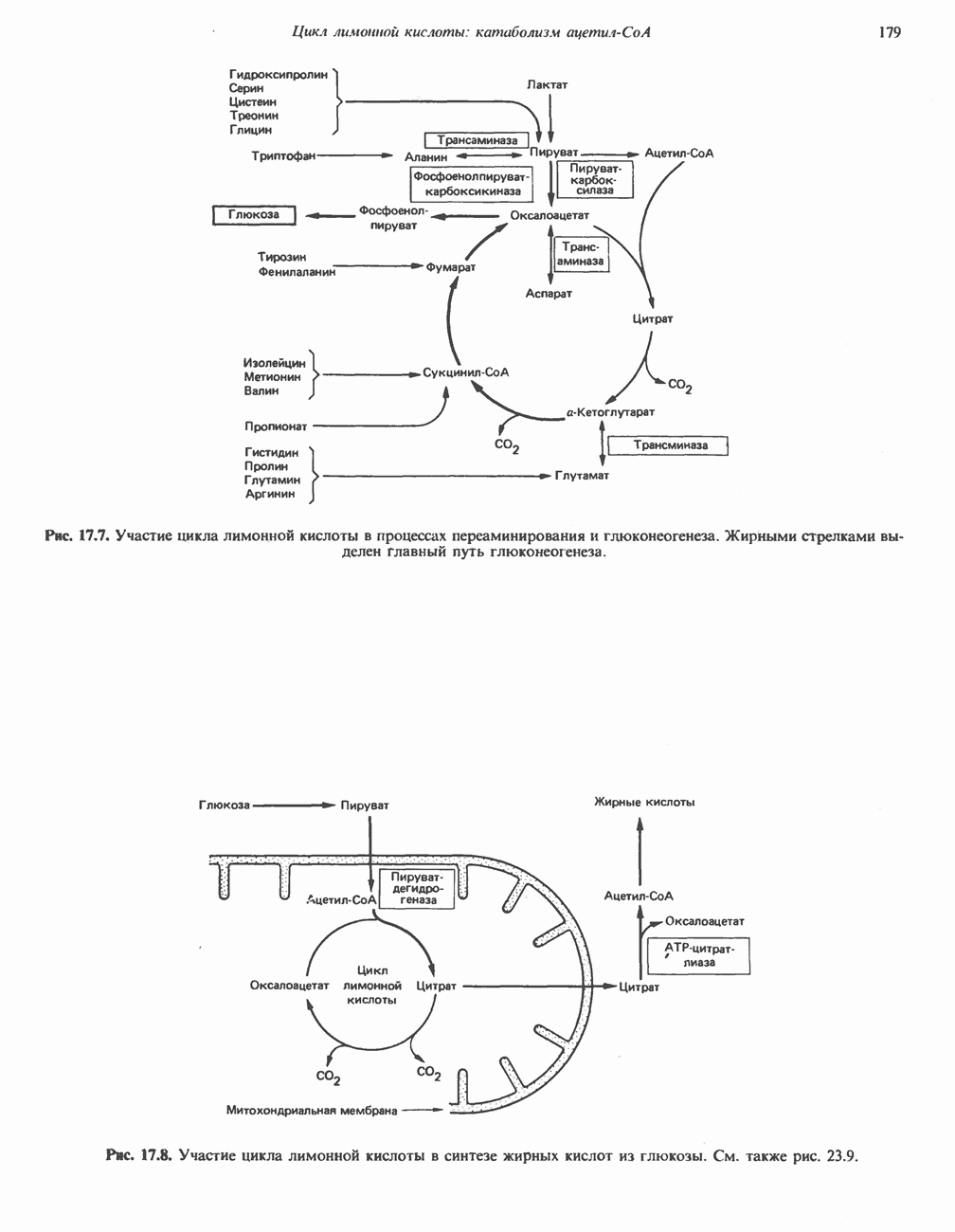
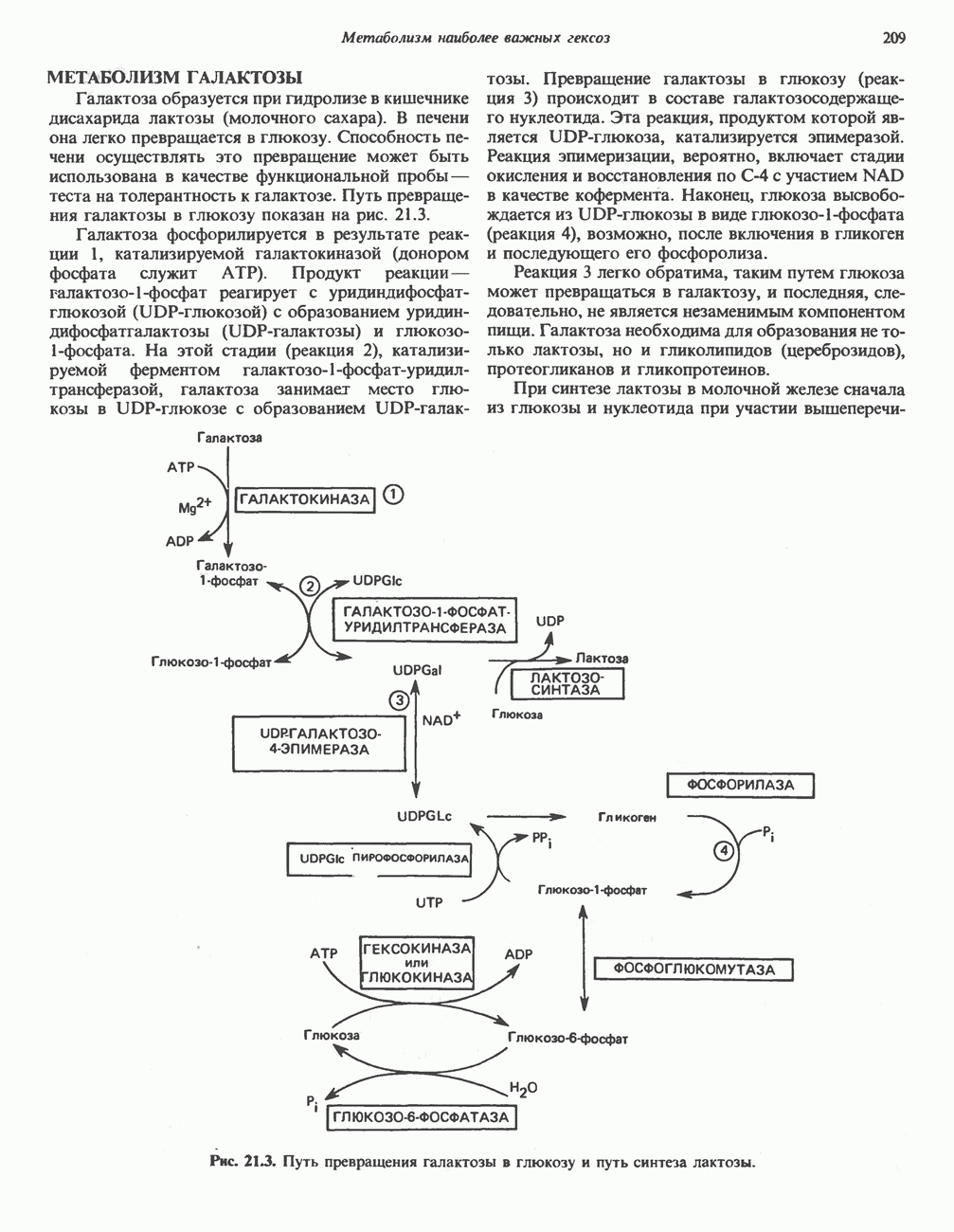
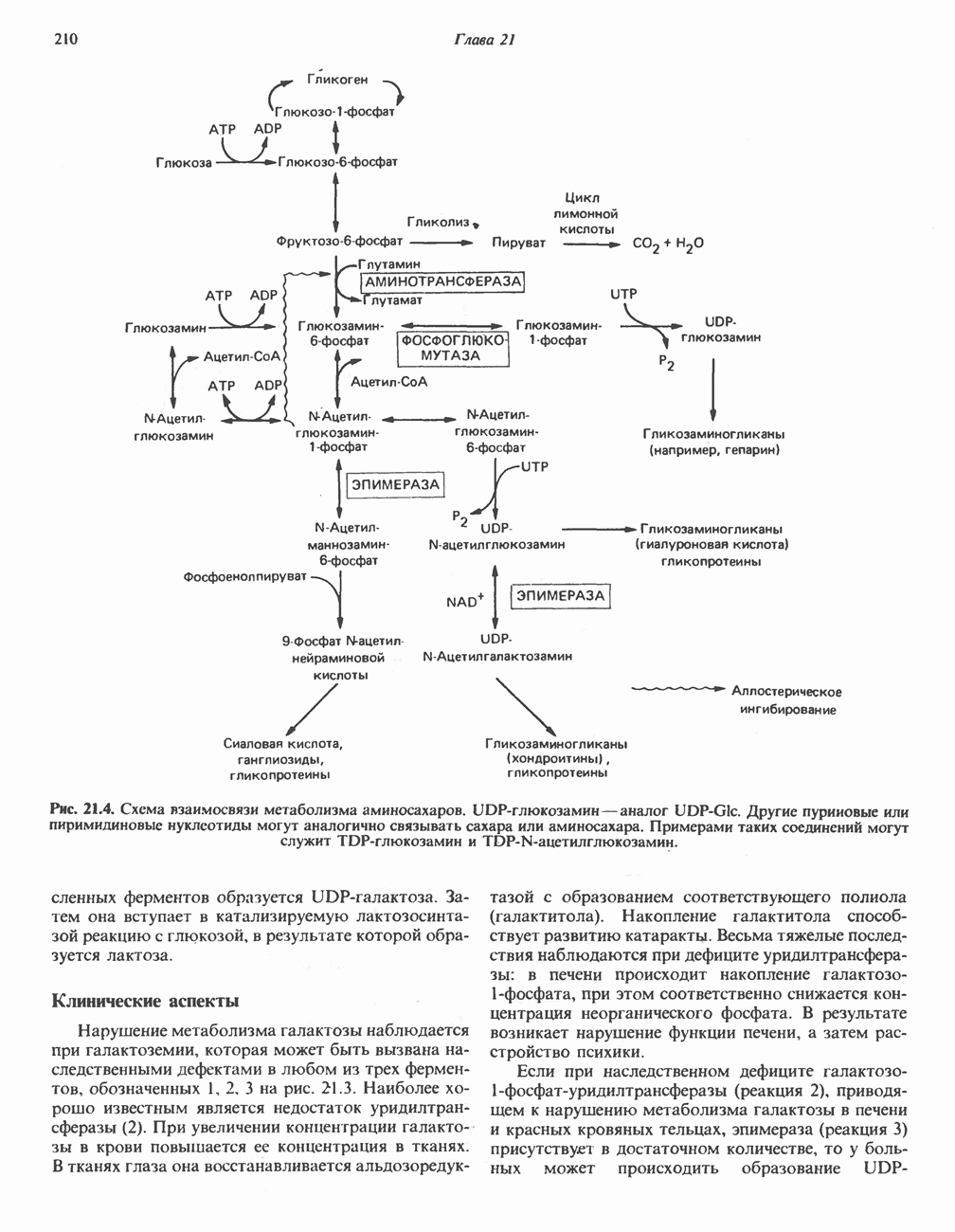
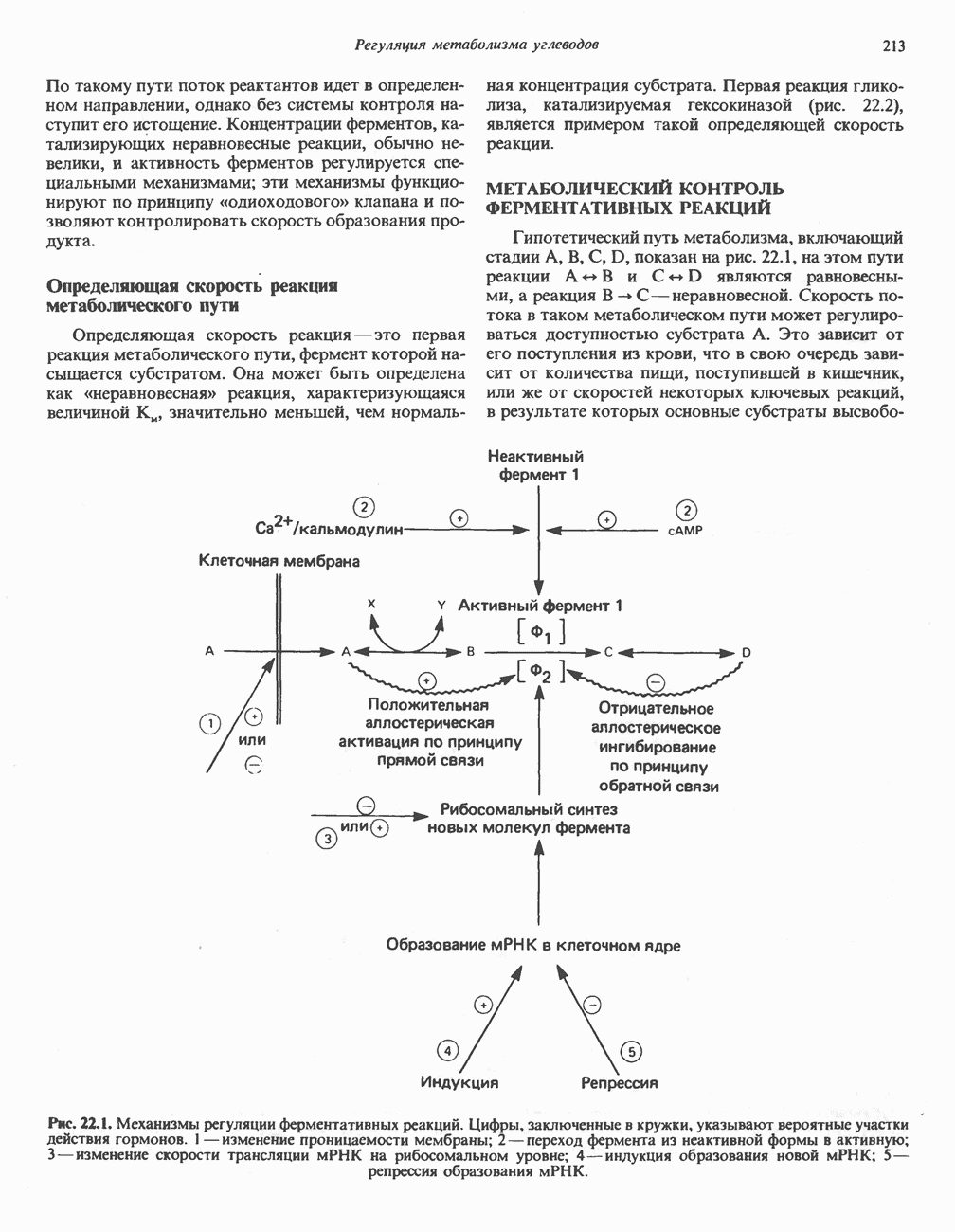
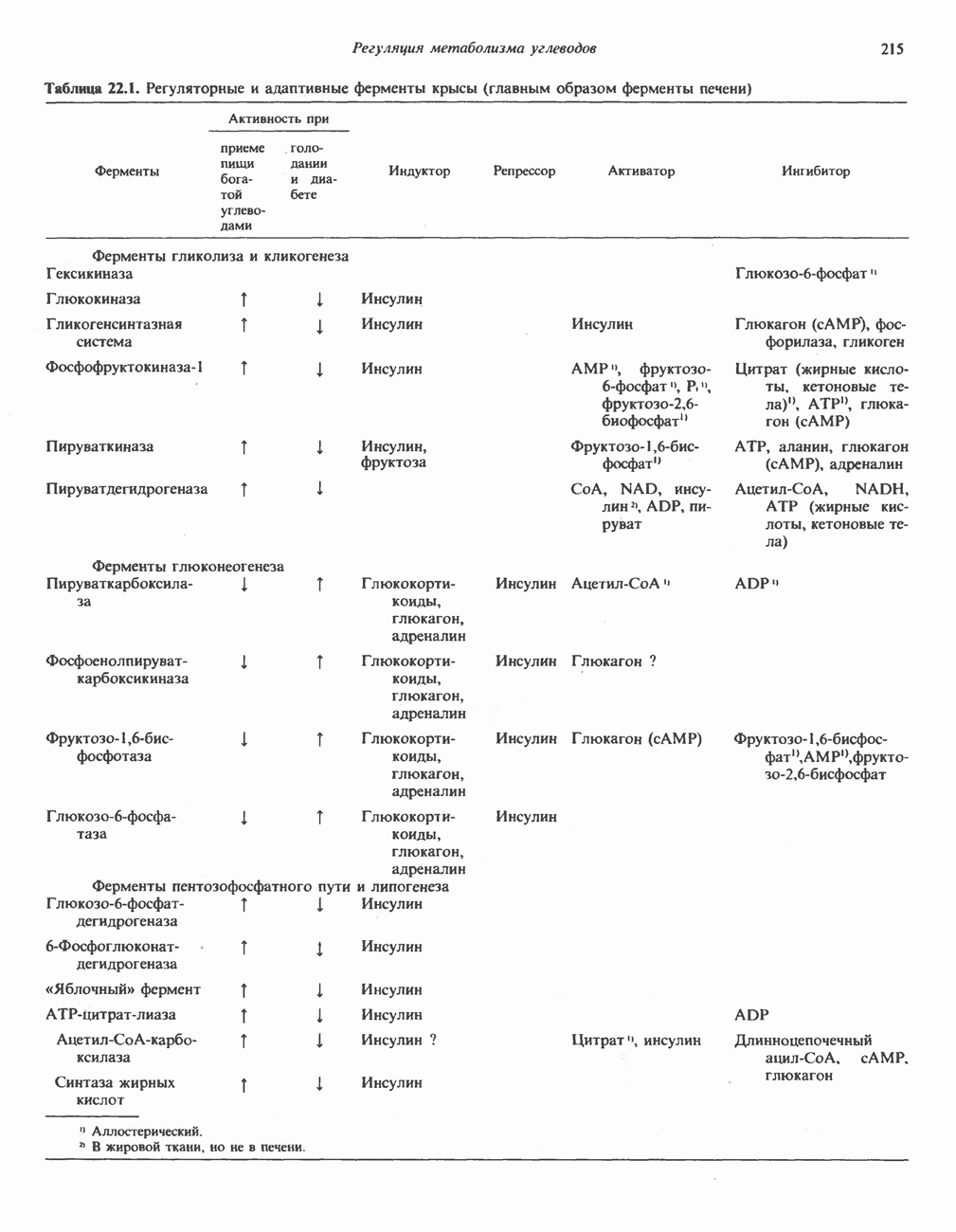
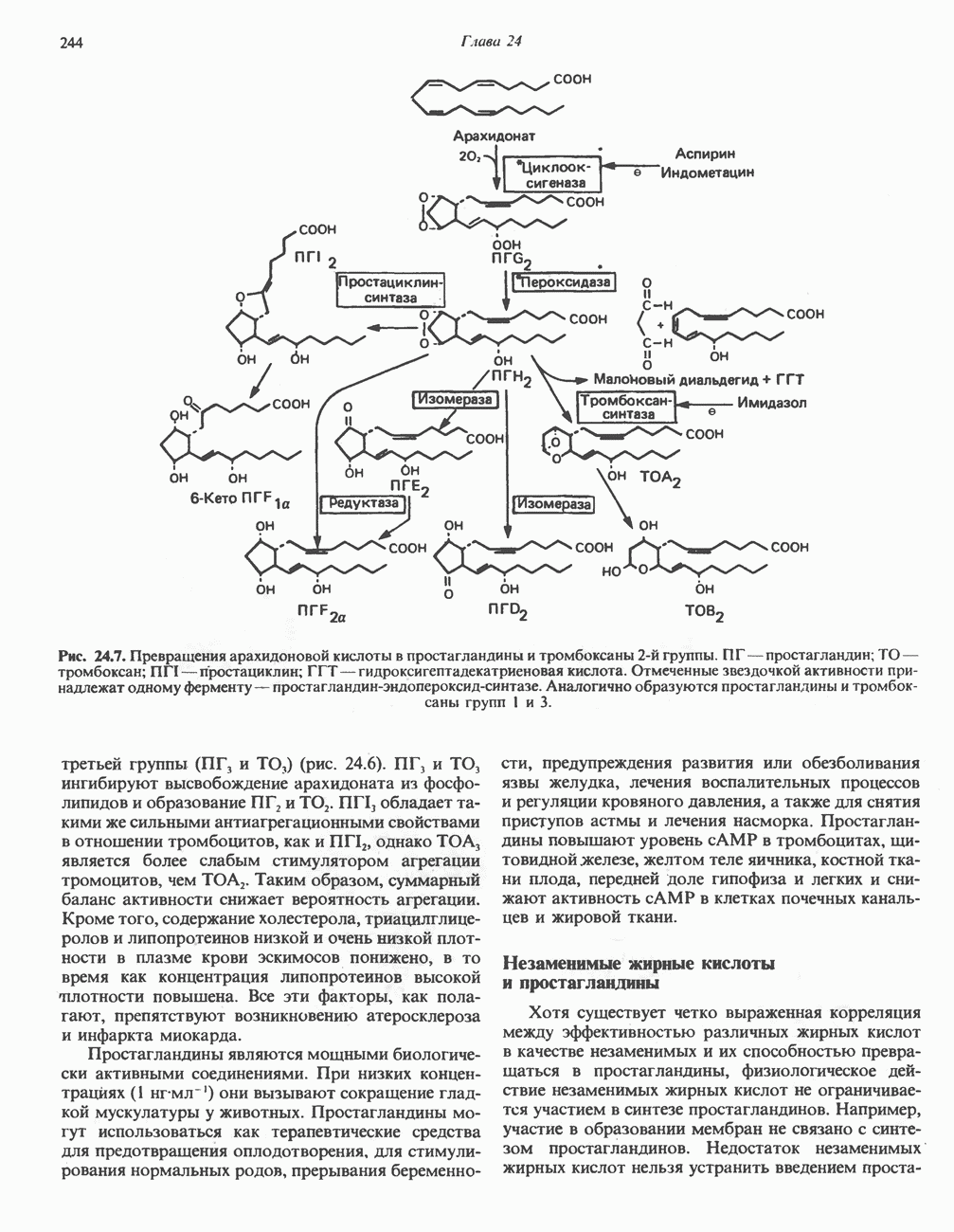
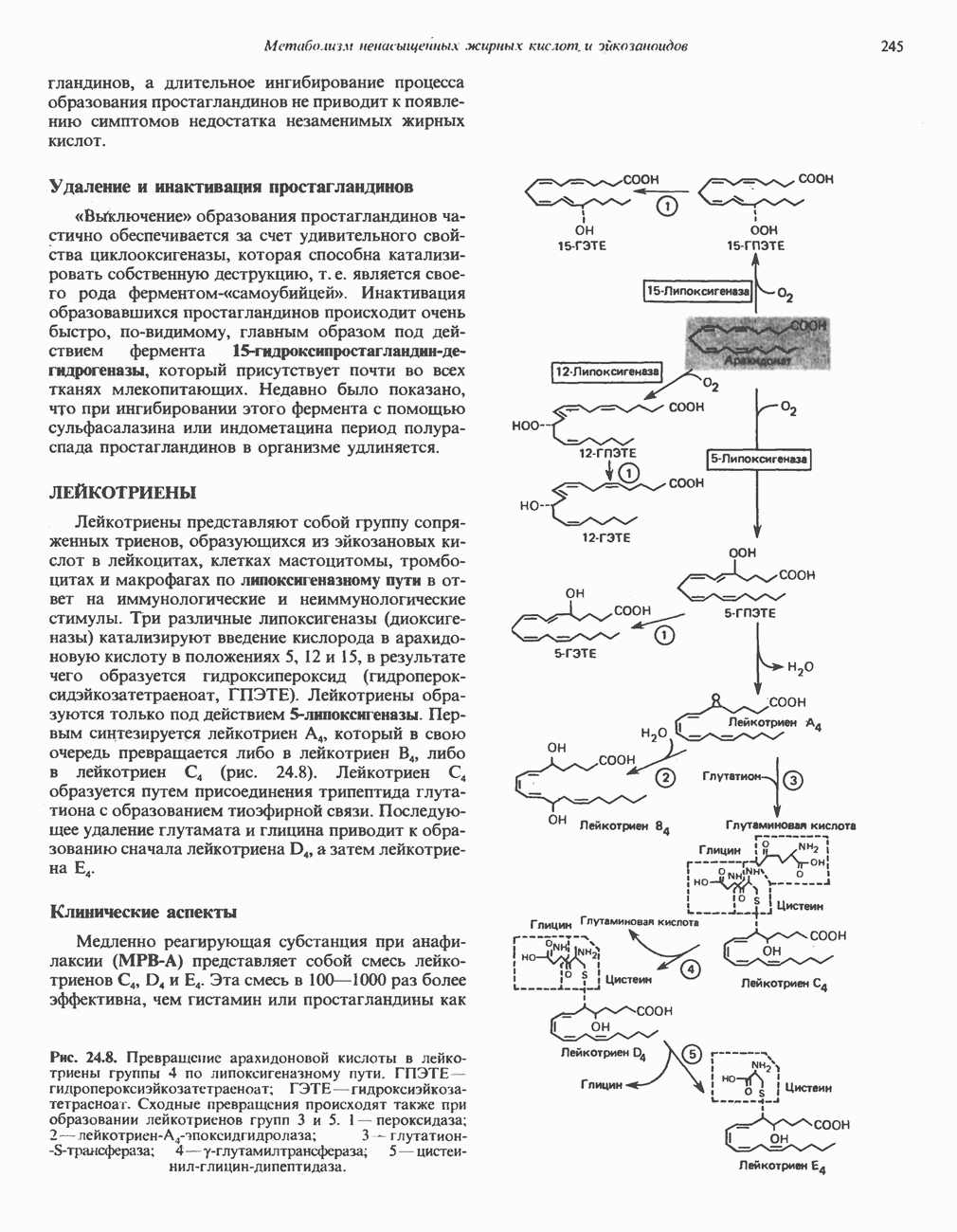
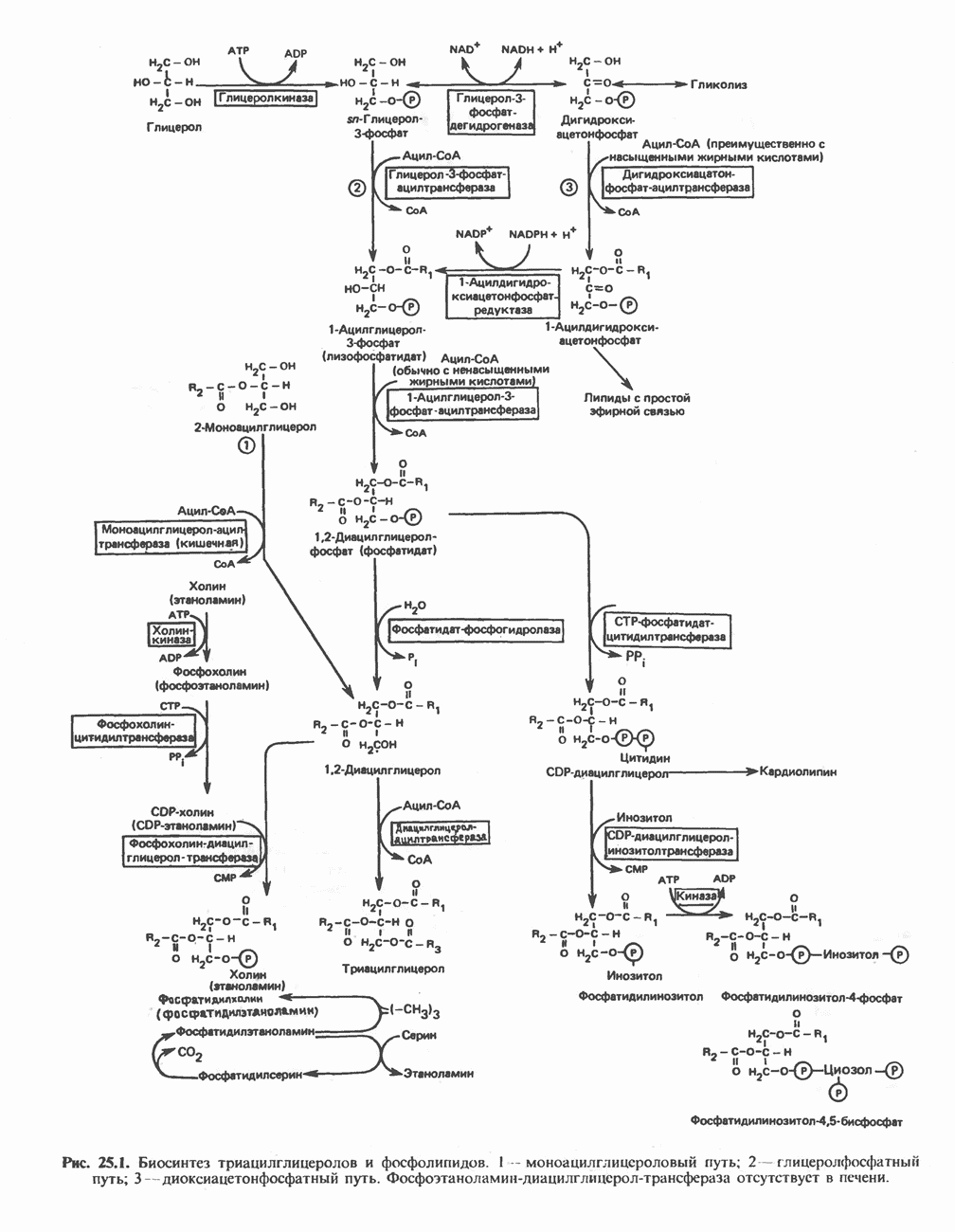
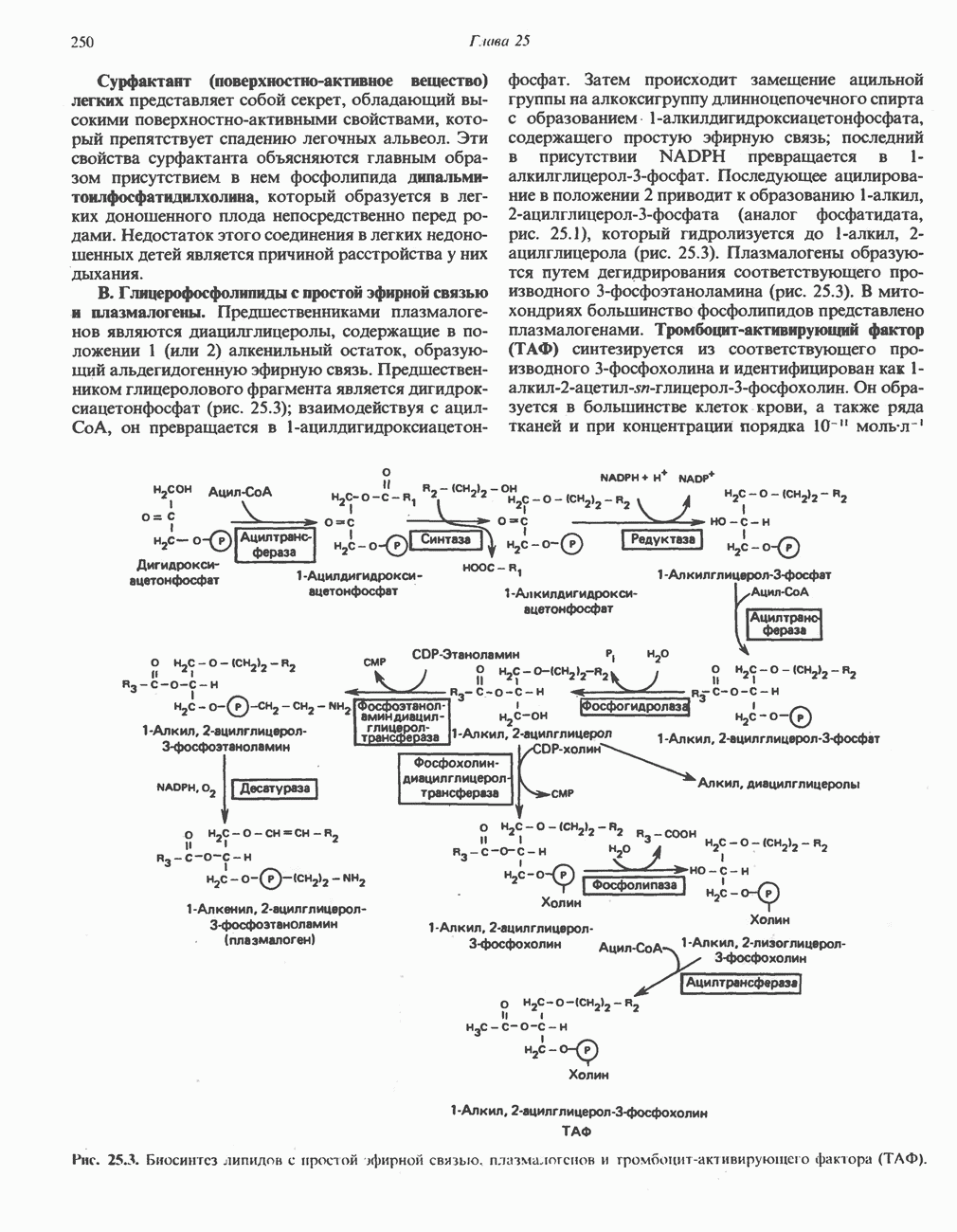
БИОСИНТЕЗ НАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТРанее предполагали, что процессы расщепления являются обращением процессов синтеза (например, гликогенолиз и гликогенез), а синтез жирных кислот рассматривали как процесс, обратный их окислению. В настоящее время установлено, что митохондриальная система биосинтеза жирных кислот, включающая несколько модифицированную последовательность реакции Внемитохондриальная система биосинтеза de novo жирных кислот (липогенез)Эта система находится в растворимой (цитозольной) фракции клеток многих органов, в частности печени, почек, мозга, легких, молочной железы, а также в жировой ткани. Биосинтез жирных кислот протекает с участием NADPH, АТР, Образование малонил-СоАПервой реакцией биосинтеза жирных кислот, катализируемой ацети Синтазный комплекс, катализирующий образование жирных кислотИмеются два типа синтазных комплексов, катализирующих биосинтез жирных кислот; оба находятся в растворимой части клетки. У бактерий, растений и низших форм животных, таких, как эвглена, все индивидуальные ферменты синтазной системы находятся в виде автономных полипептидов; ацильные радикалы связаны с одним из них, получившим название
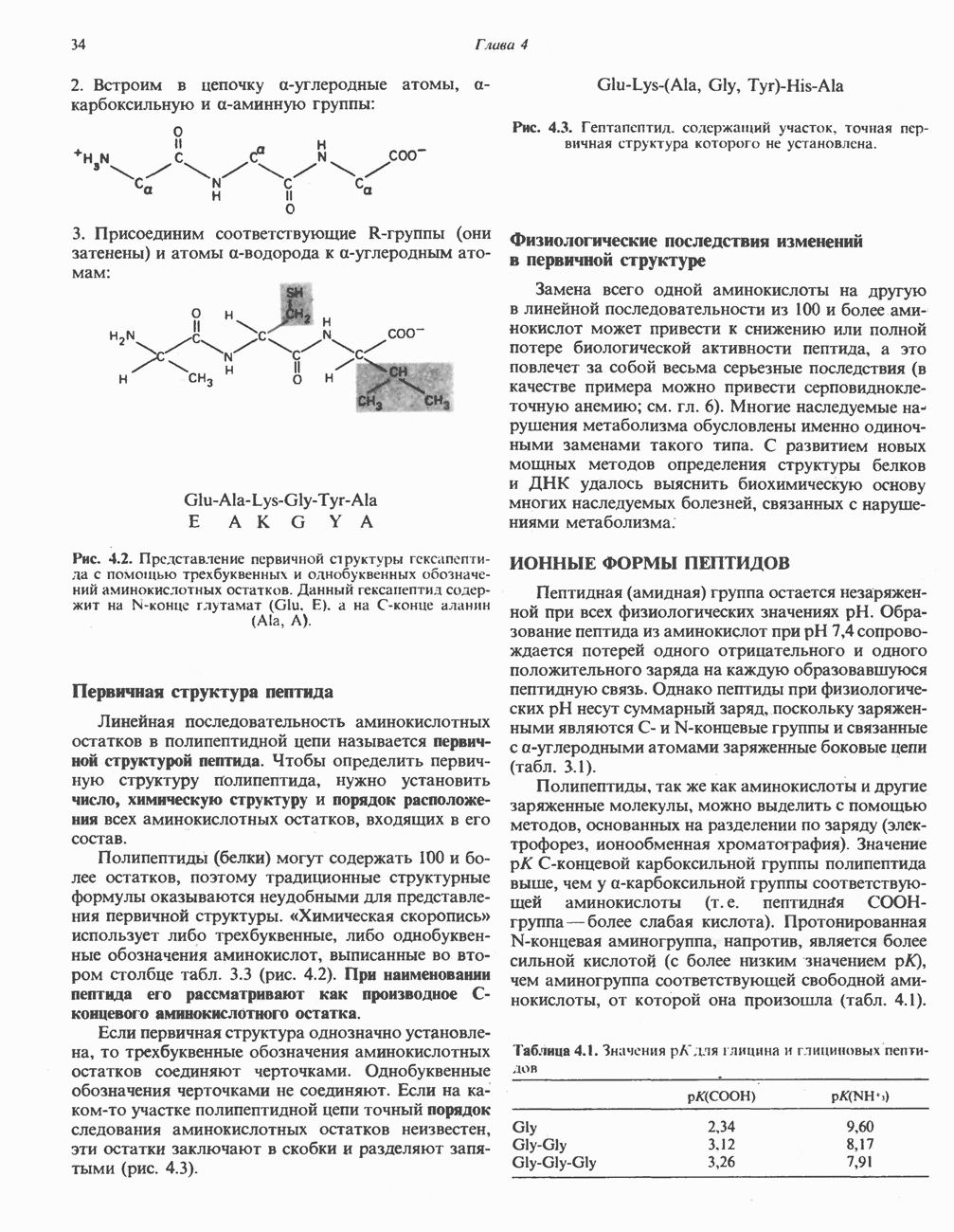
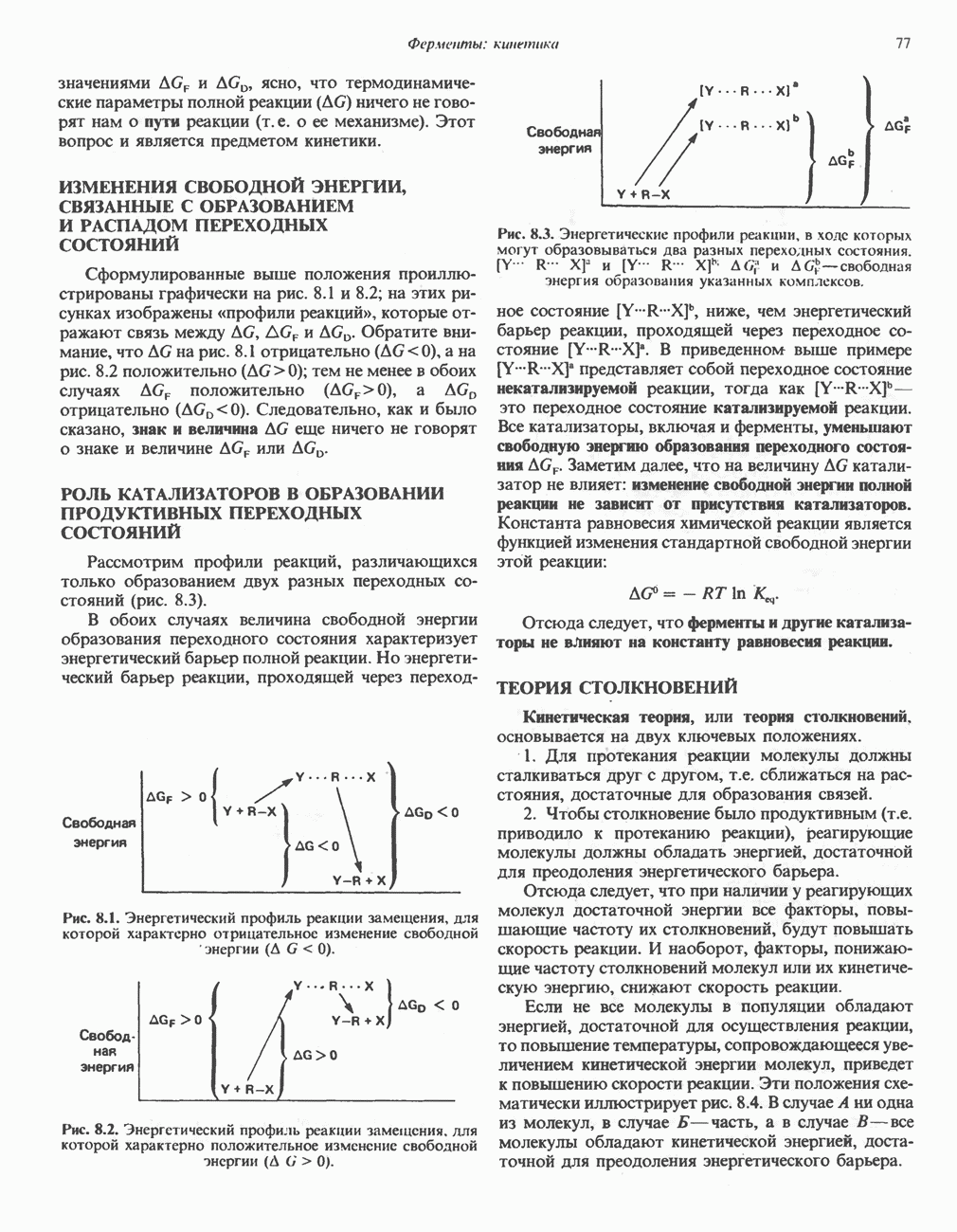
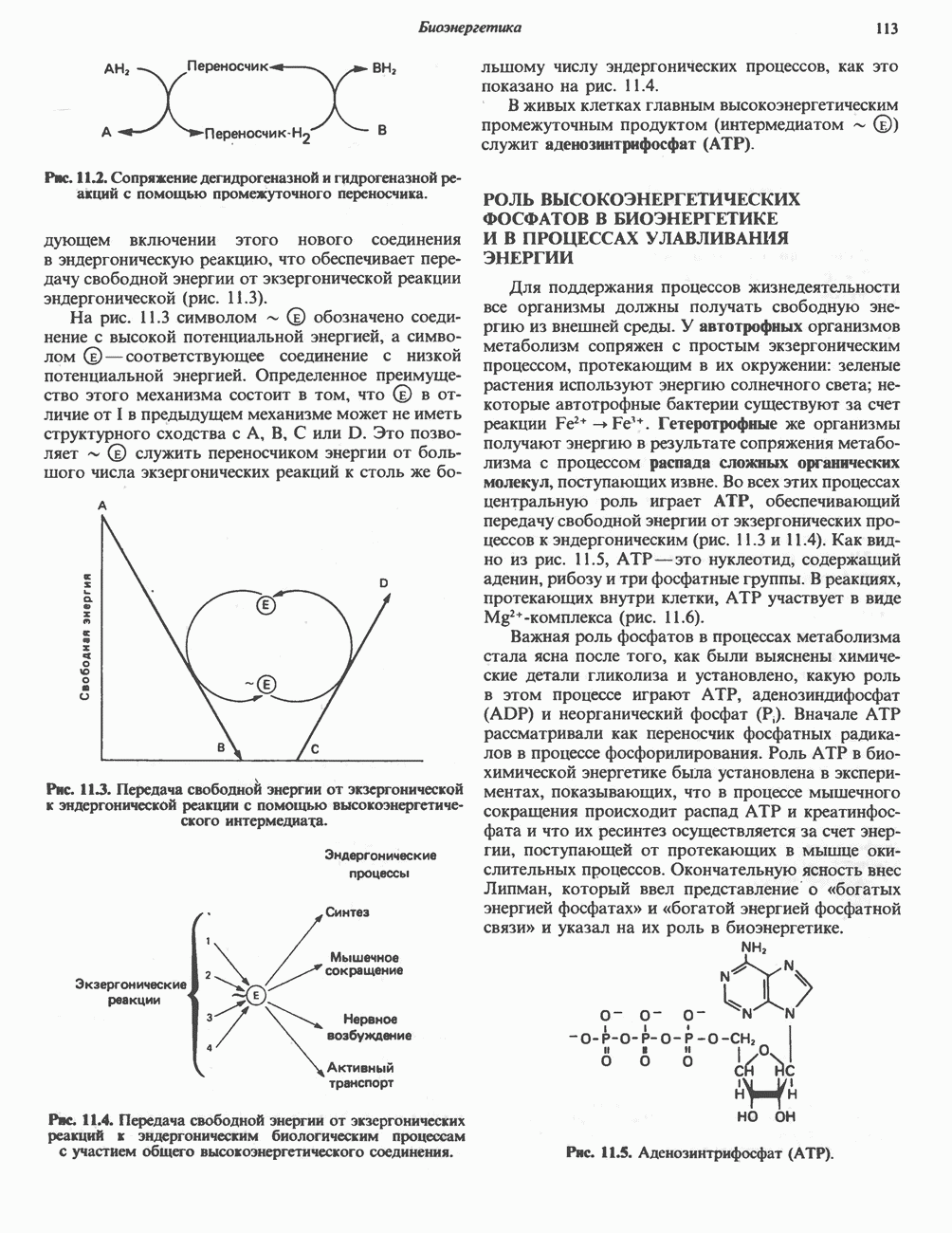
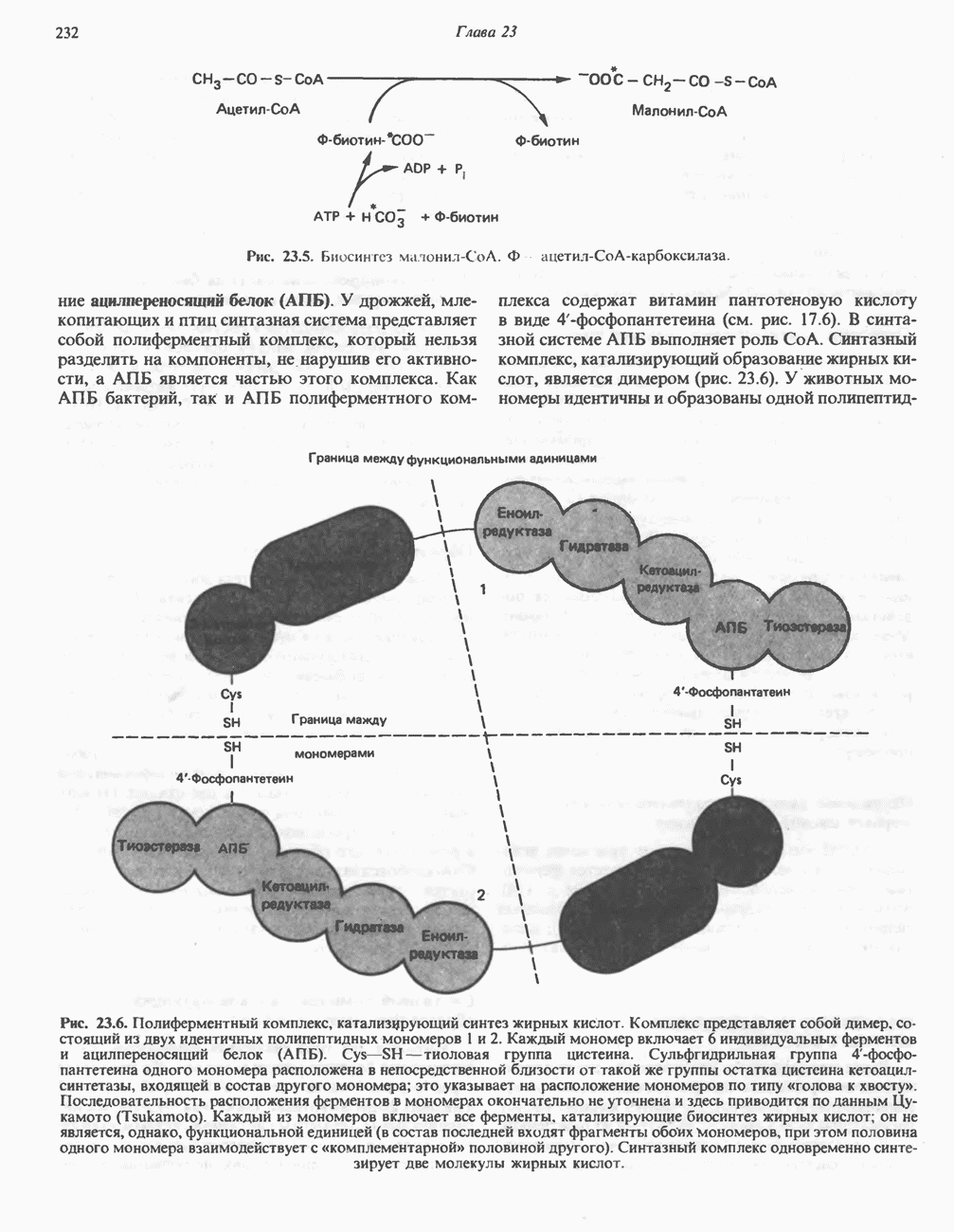
Рис. 23.5. Биосинтез малонил-СоА. Фацетил-СоА-карбоксилаза. ацилпереносящин белок (АПБ). У дрожжей, млекопитающих и птиц синтазная система представляет собой полиферментный комплекс, который нельзя разделить на компоненты, не нарушив его активности, а АПБ является частью этого комплекса. Как АПБ бактерий, так и АПБ полиферментного комплекса содержат витамин пантотеновую кислоту в виде 4-фосфопантетеина (см. рис. 17.6). В синтазной системе АПБ выполняет роль СоА. Синтазный комплекс, катализирующий образование жирных кислот, является димером (рис. 23.6). У животных мономеры идентичны и образованы одной полипептидной
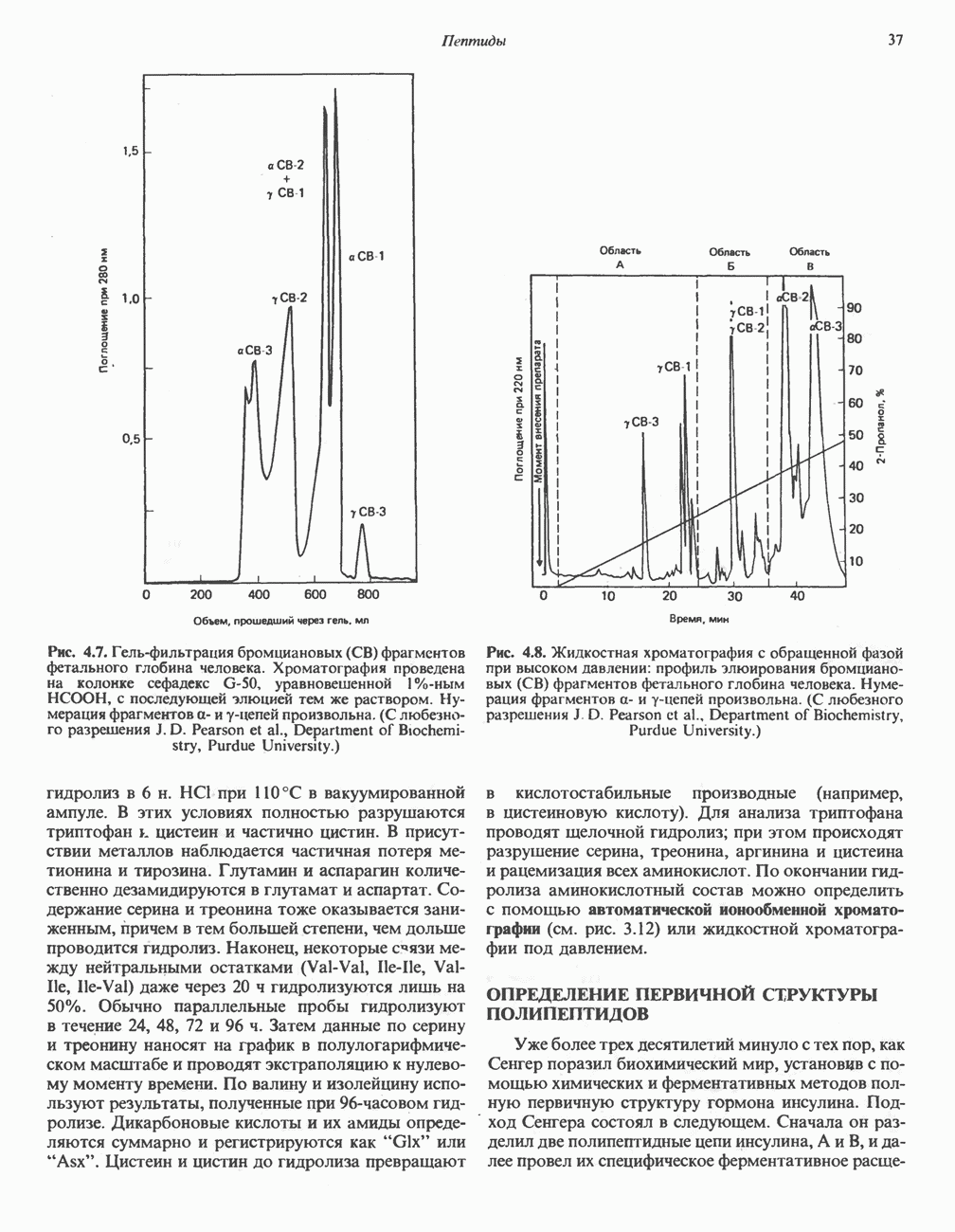
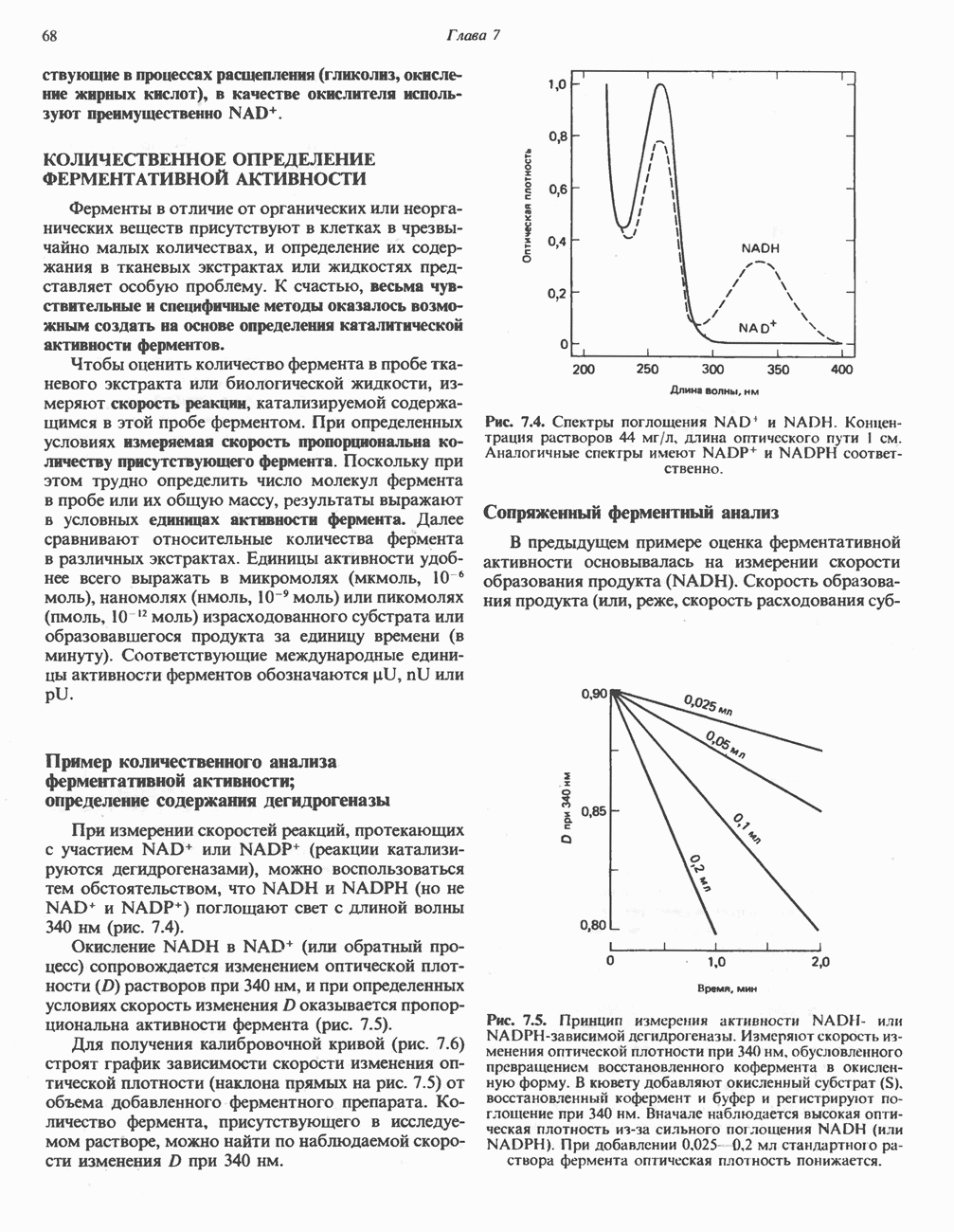
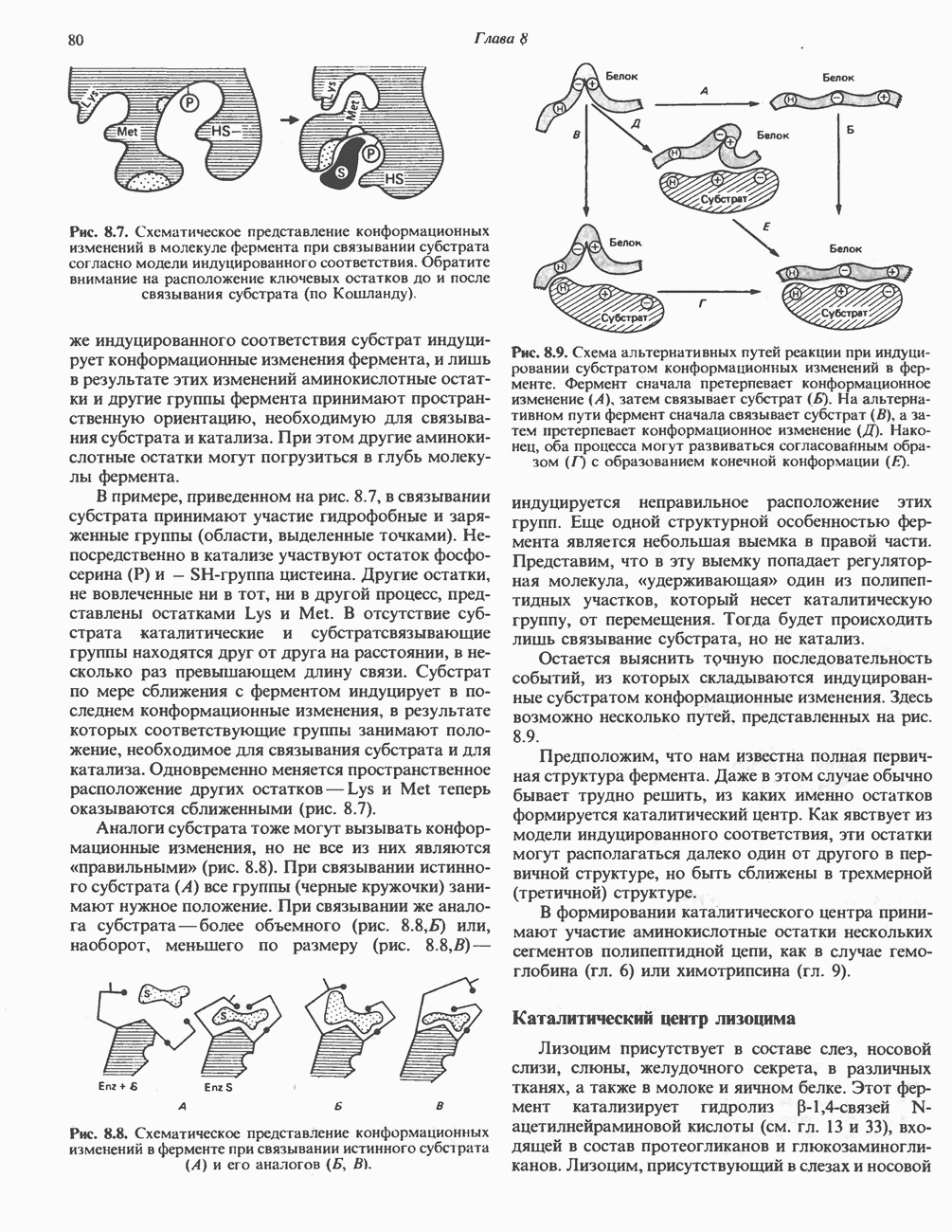
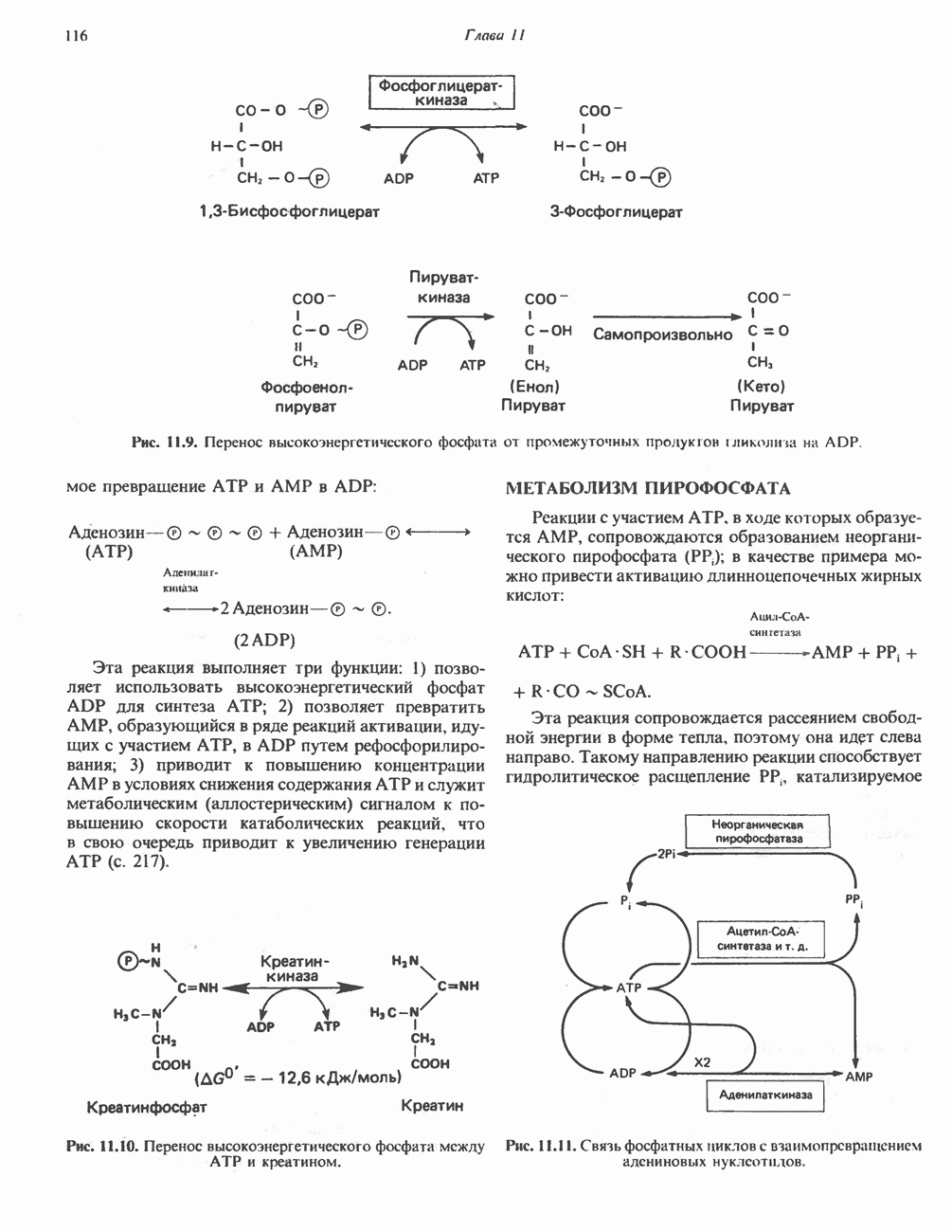
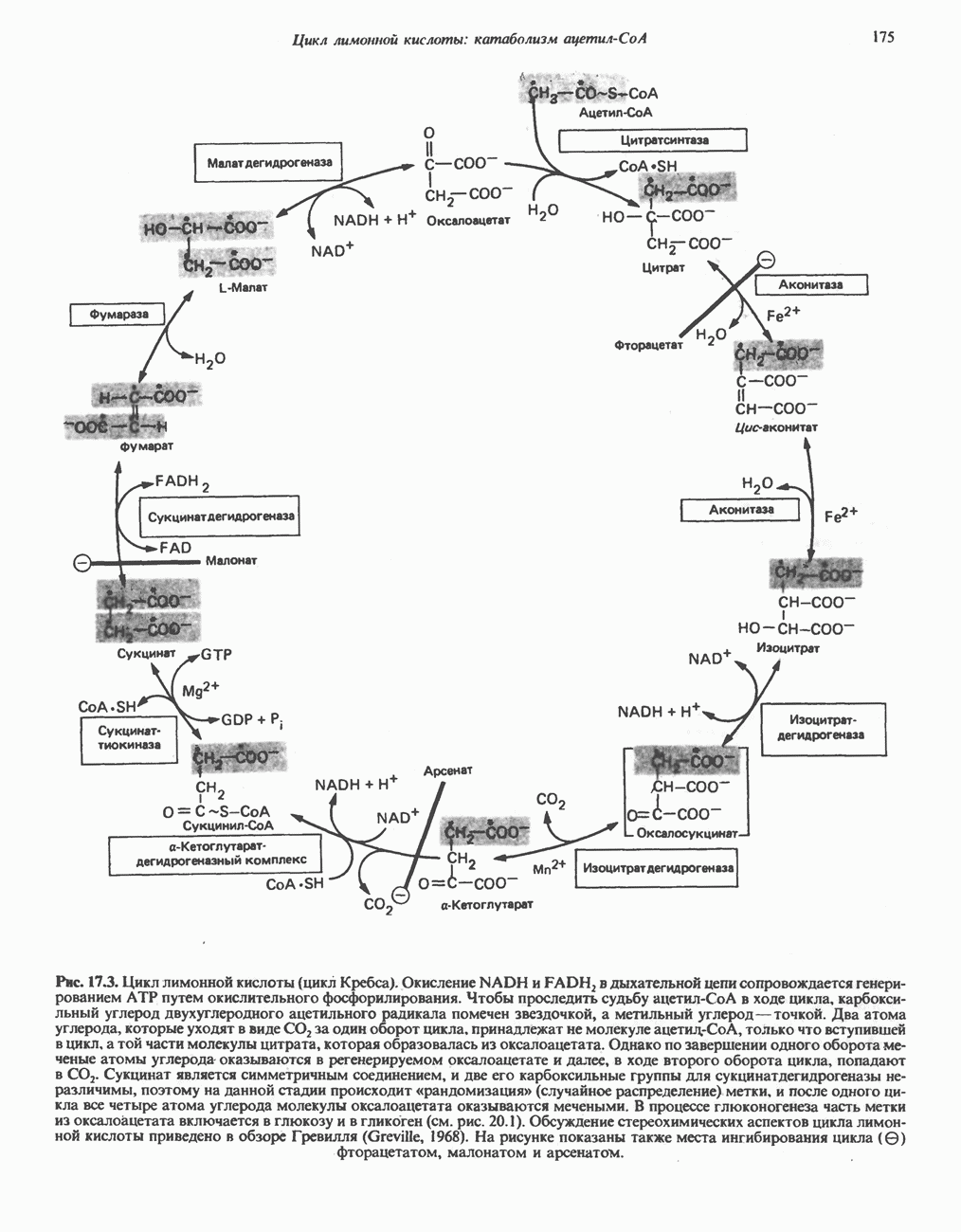
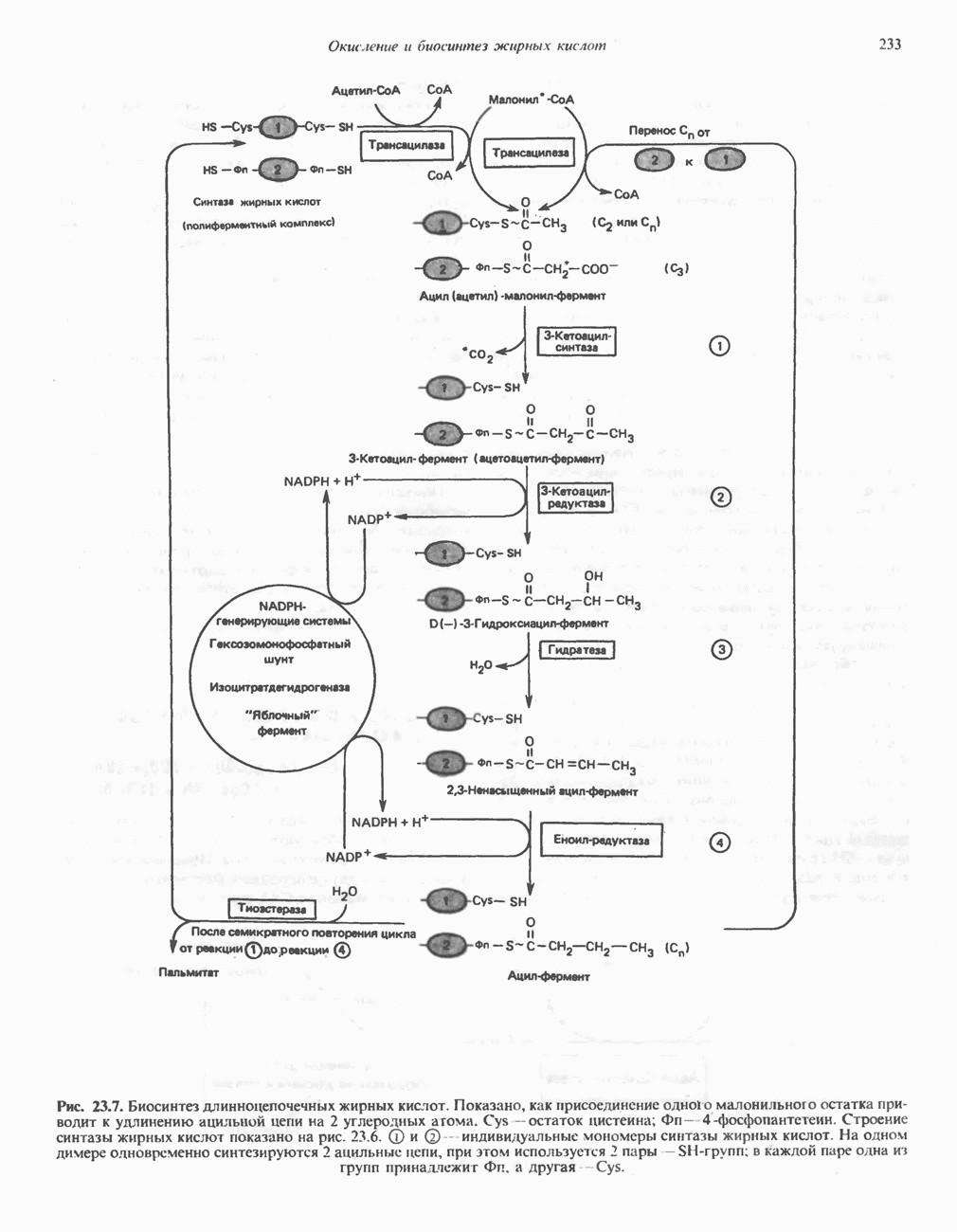
Рис. 23.6. Полиферментный комплекс, катализирующий синтез жирных кислот. Комплекс представляет собой димер, состоящий из двух идентичных полипептидных мономеров 1 и 2. Каждый мономер включает 6 индивидуальных ферментов и ацилпереносящий белок (АПБ). Cys—SH—тиоловая группа цистеина. Сульфгидрильная группа 4-фосфопантетеина одного мономера расположена в непосредственной близости от такой же группы остатка цистсина кетоацил-синтетазы, входящей в состав другого мономера; это указывает на расположение мономеров по типу «голова к хвосту». Последовательность расположения ферментов в мономерах окончательно не уточнена и здесь приводится по данным Цукамото (Tsukamoto). Каждый из мономеров включает все ферменты, катализирующие биосинтез жирных кислот; он не является, однако, функциональной единицей (в состав последней входят фрагменты обоих мономеров, при этом половина одного мономера взаимодействует с «комплементарной» половиной другого). Синтазный комплекс одновременно синтезирует две молекулы жирных кислот. (см. скан) Рис. 23.7. Биосинтез длинноцепочечных жирных кислот. Показано, как присоединение одною малонильного остатка приводит к удлинению ацилыюй цепи на 2 углеродных агома. Cys - остаток цистеина; Фп - 4-фосфопантетеин. Строение синтазы жирных кислот показано на рис. 23.6. цепью, включающей 6 ферментов, катализирующих биосинтез жирных кислот, и АПБ с реакционноспособной На первом этапе процесса инициирующая молекула В молочной железе имеется особая тиоэстераза, специфичная к ацильным остаткам По-видимому, в одном димерном синтазном комплексе имеются 2 активных центра, функционирующие независимо друг от друга, в результате одновременно образуются 2 молекулы пальмитиновой кислоты. Объединение всех ферментов рассматриваемого метаболического пути в единый полиферментный комплекс обеспечивает его высокую эффективность и устраняет конкуренцию других процессов, в результате достигается эффект компартментации данного пути в клетке без участия дополнительных барьеров проницаемости. Ниже приводится суммарная реакция биосинтеза пальмитиновой кислоты из ацетил-СоА и малонил-СоА:
Из молекулы
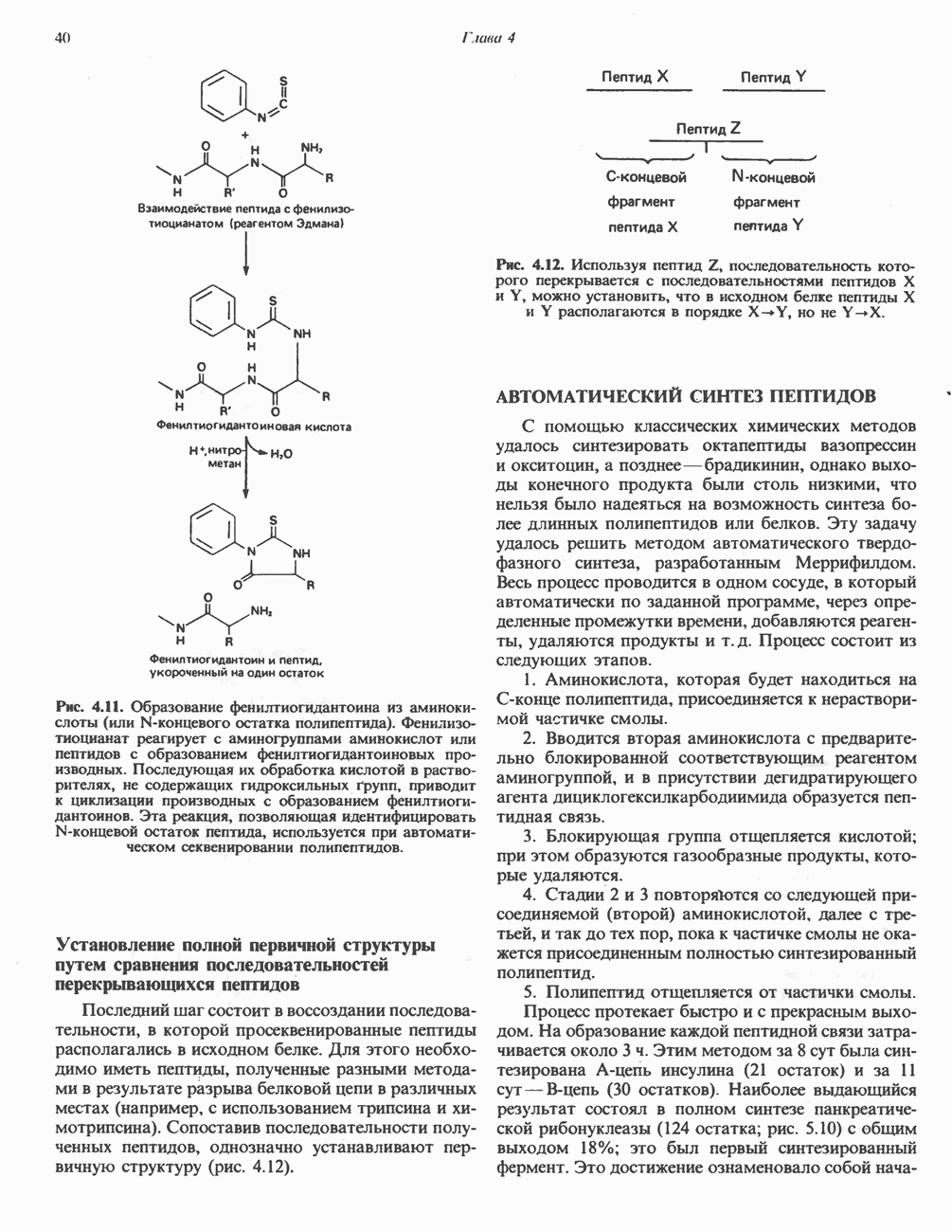
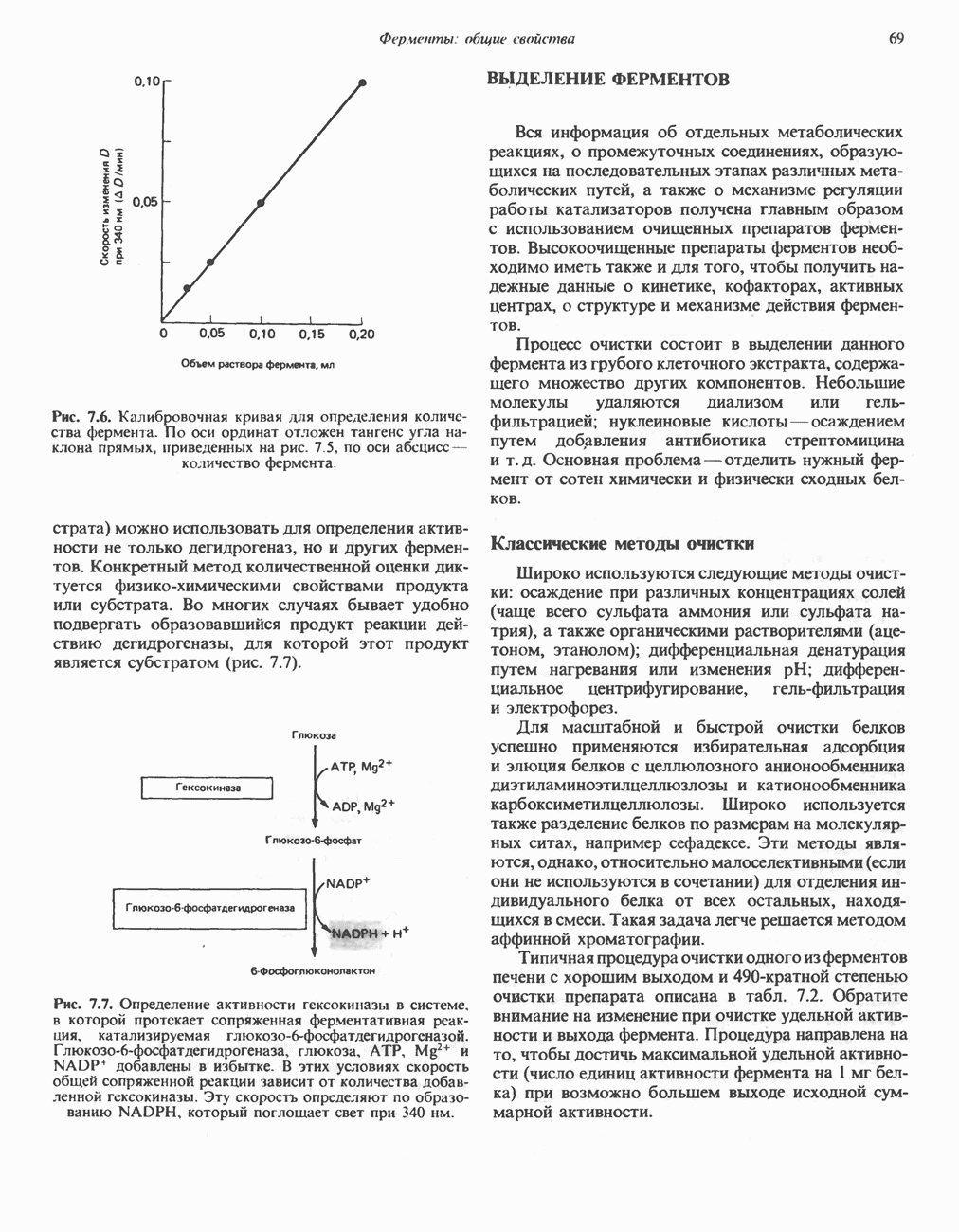
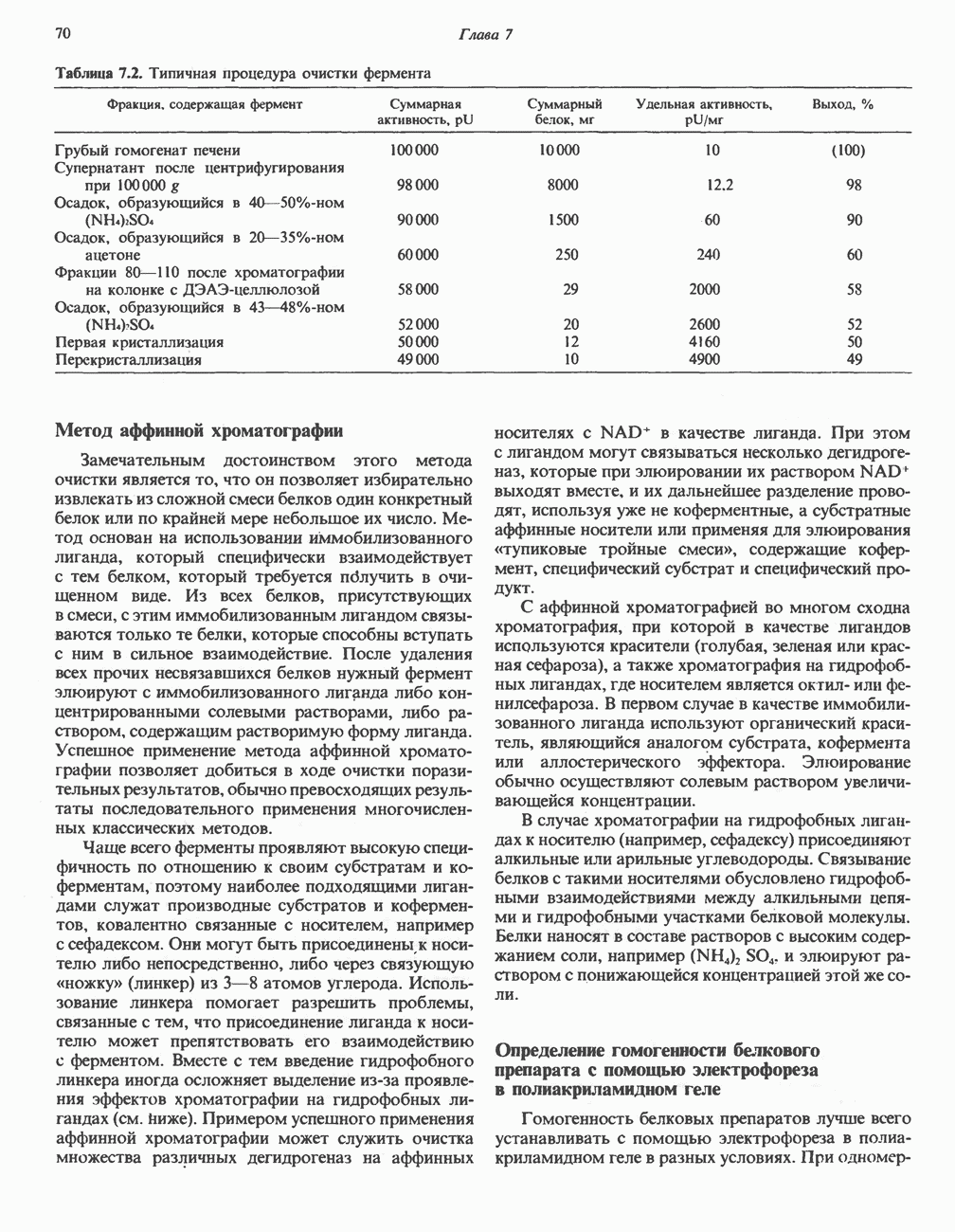
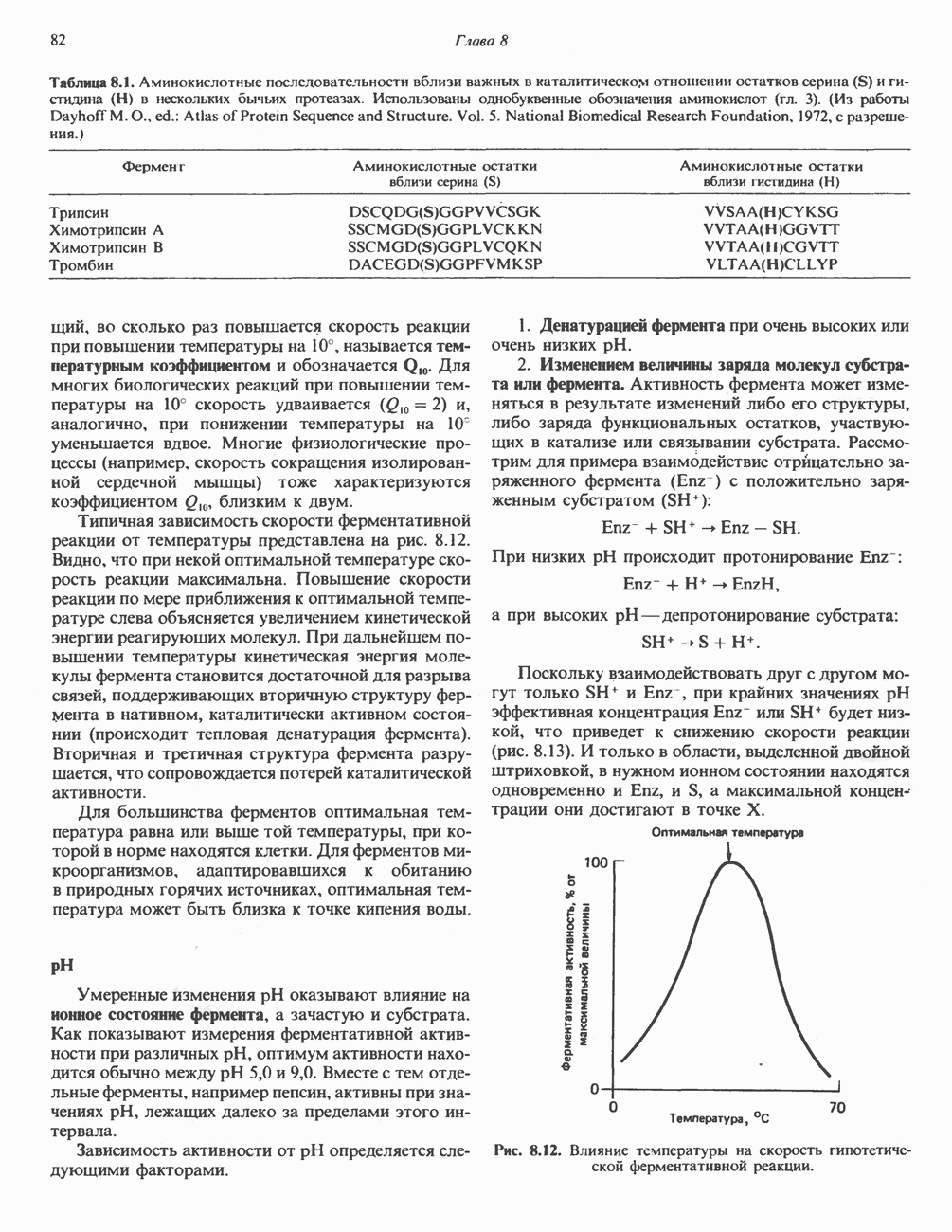
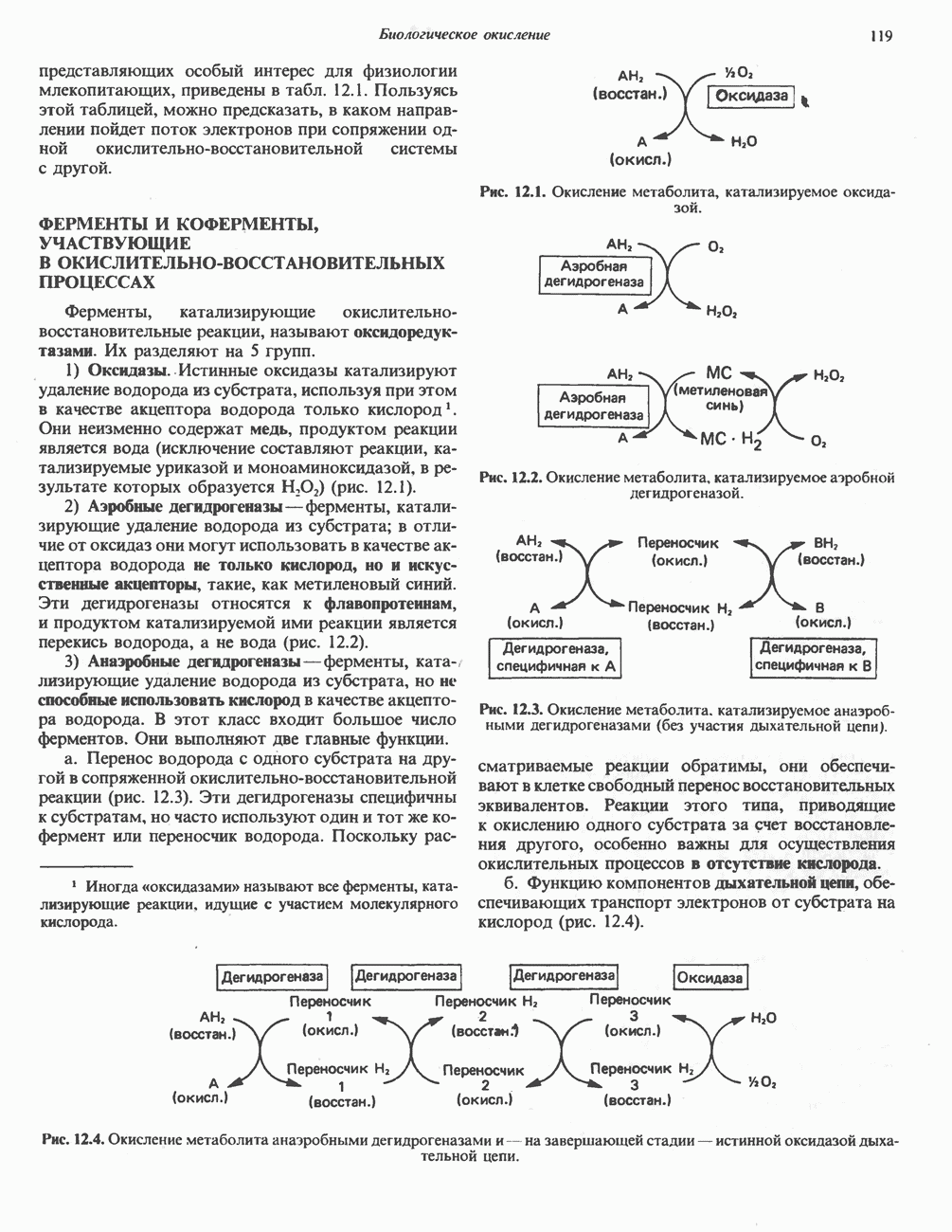
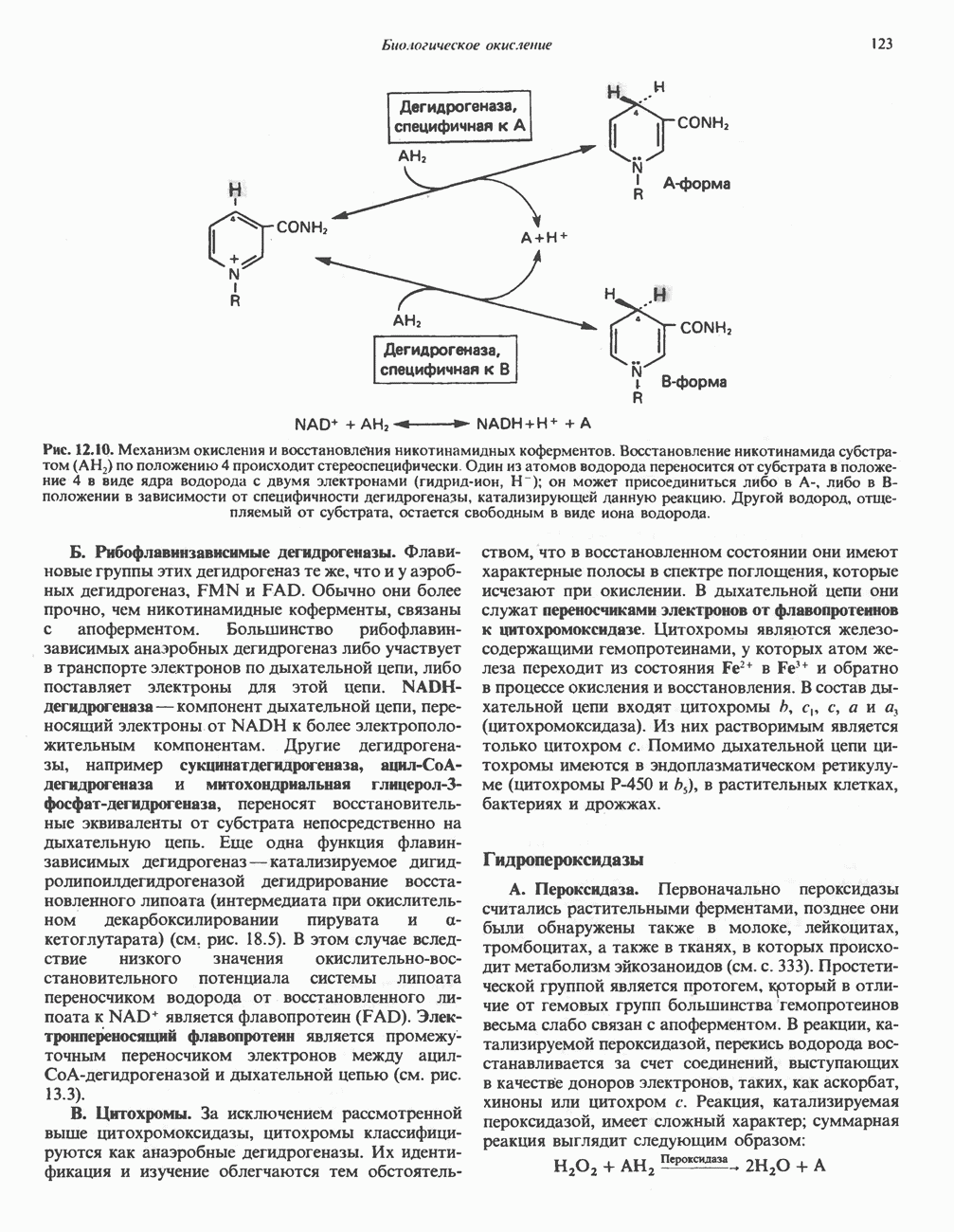
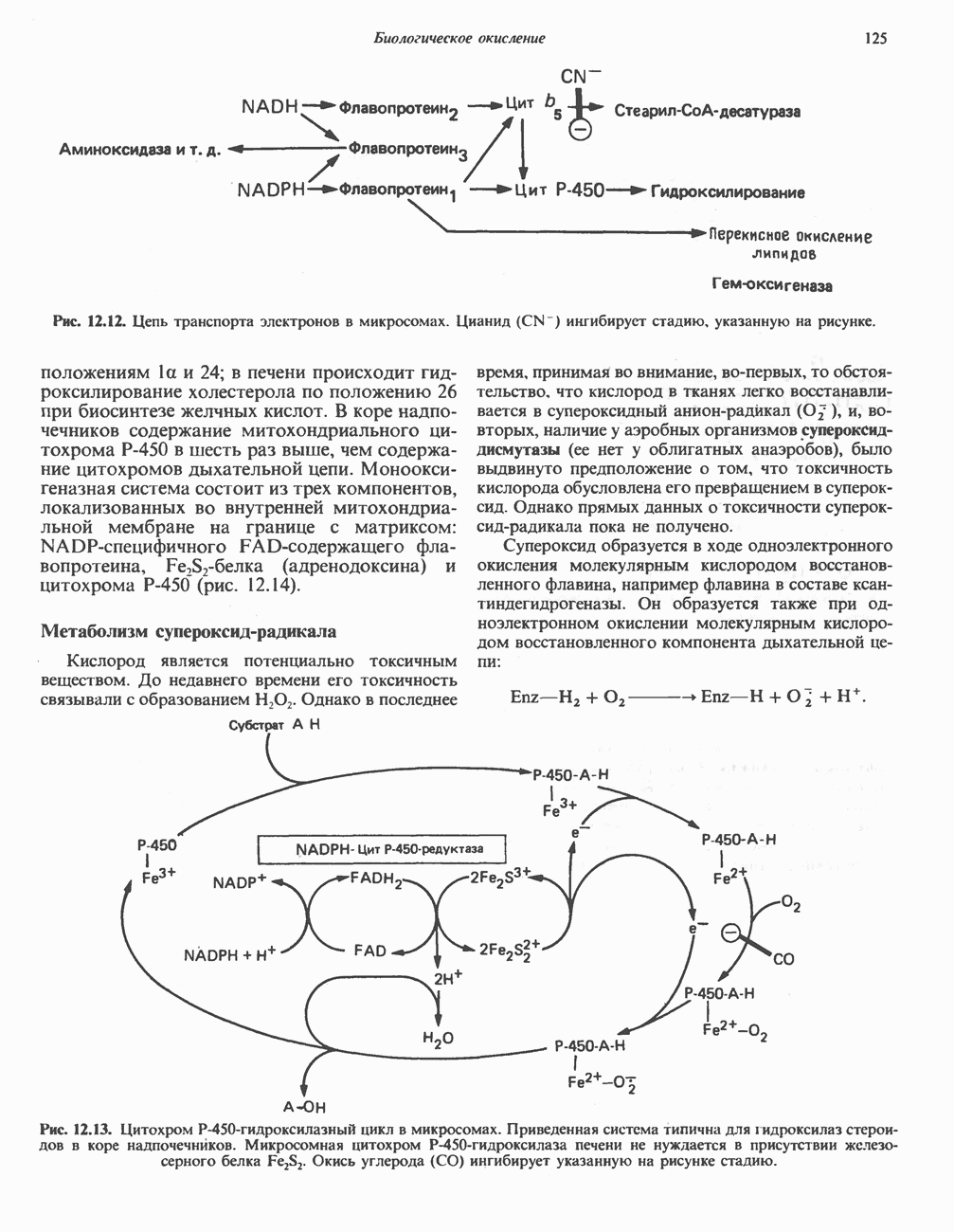
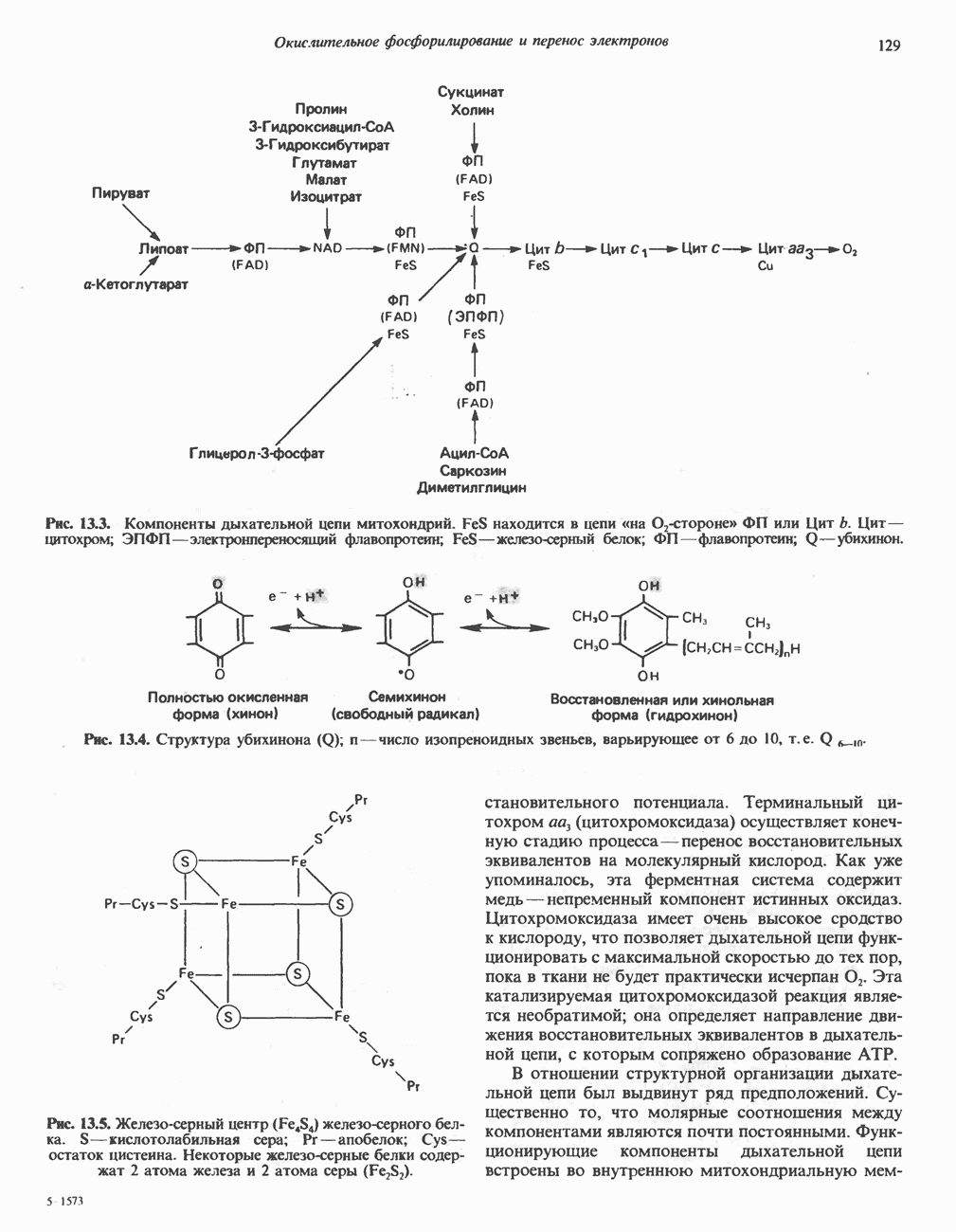
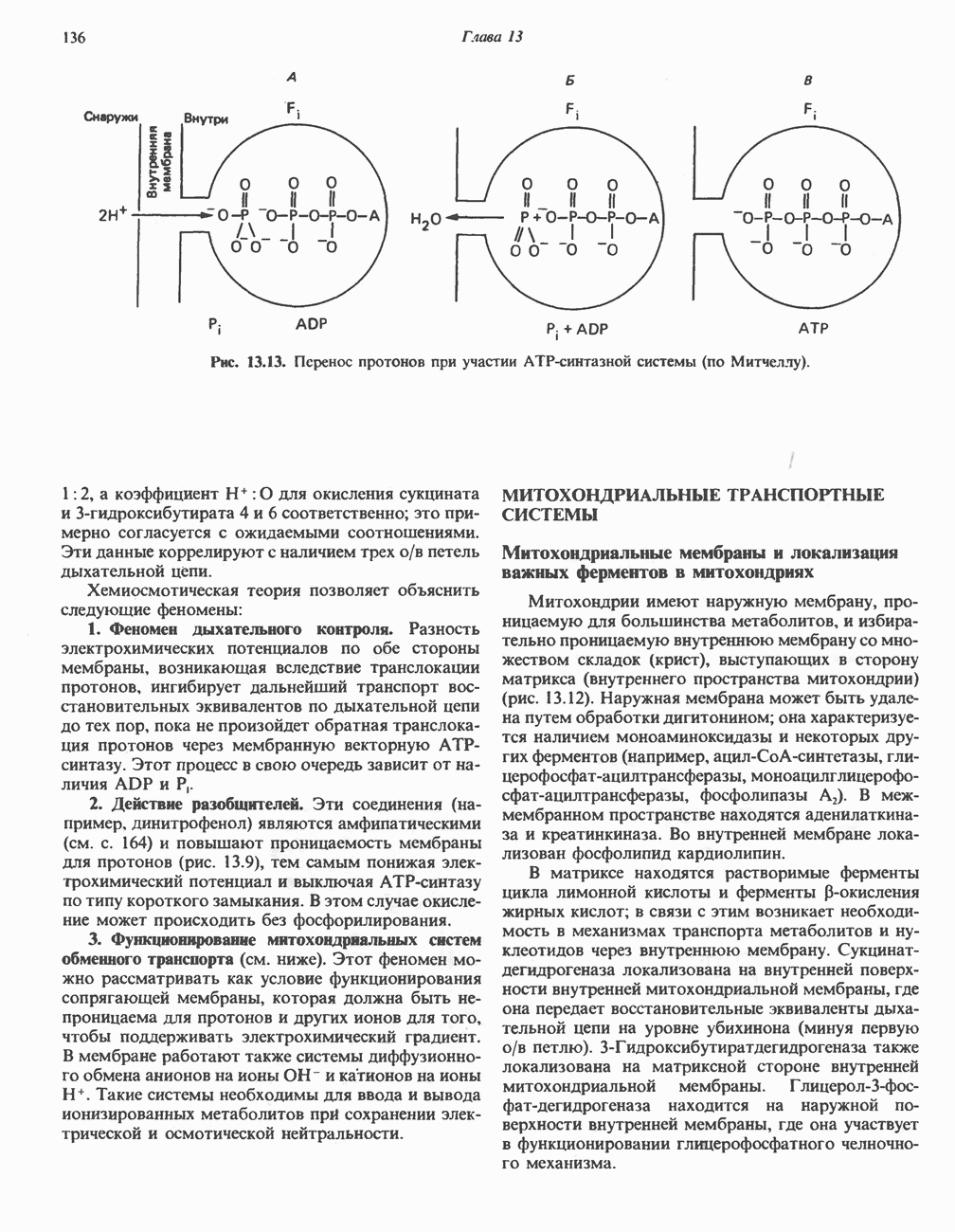
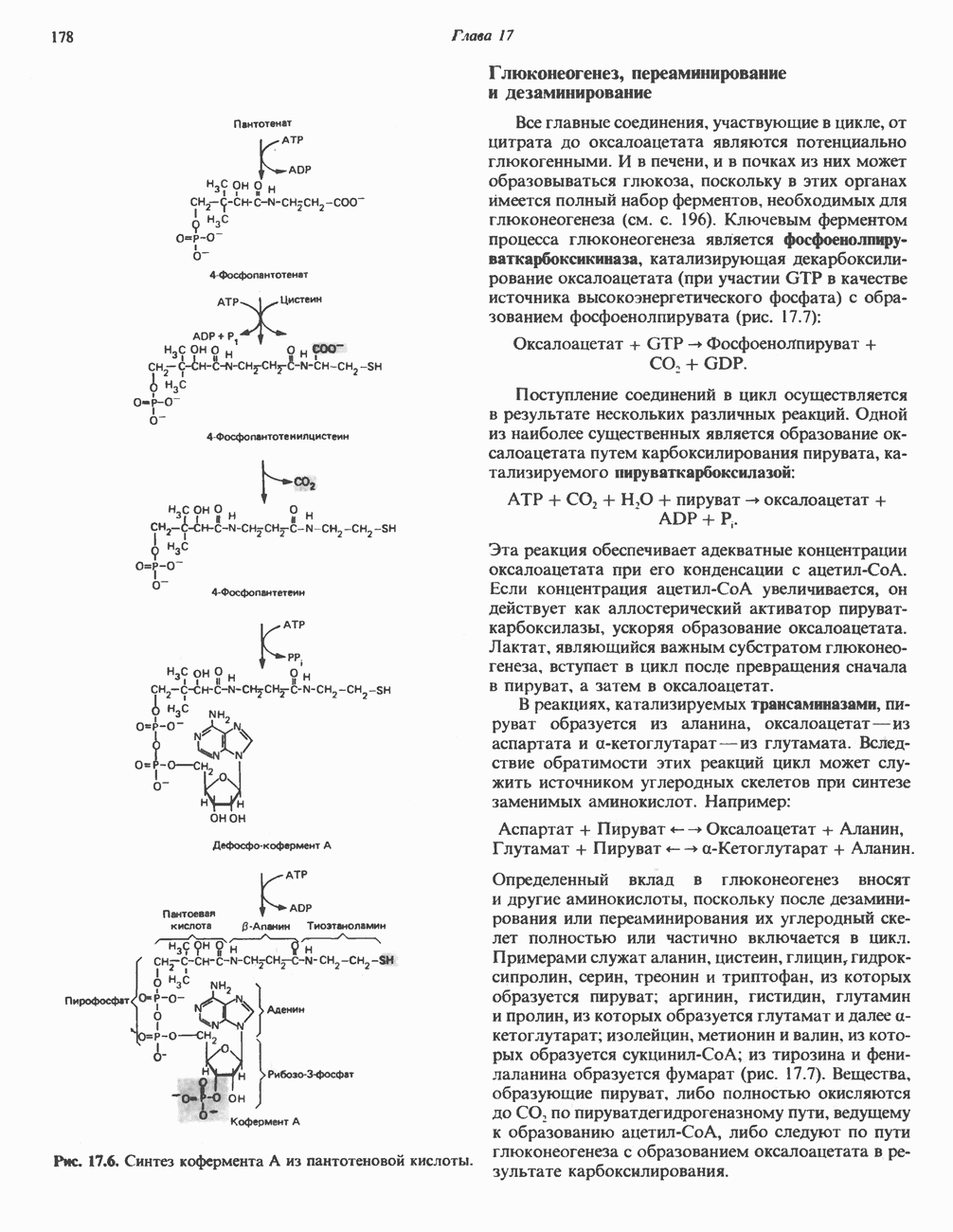
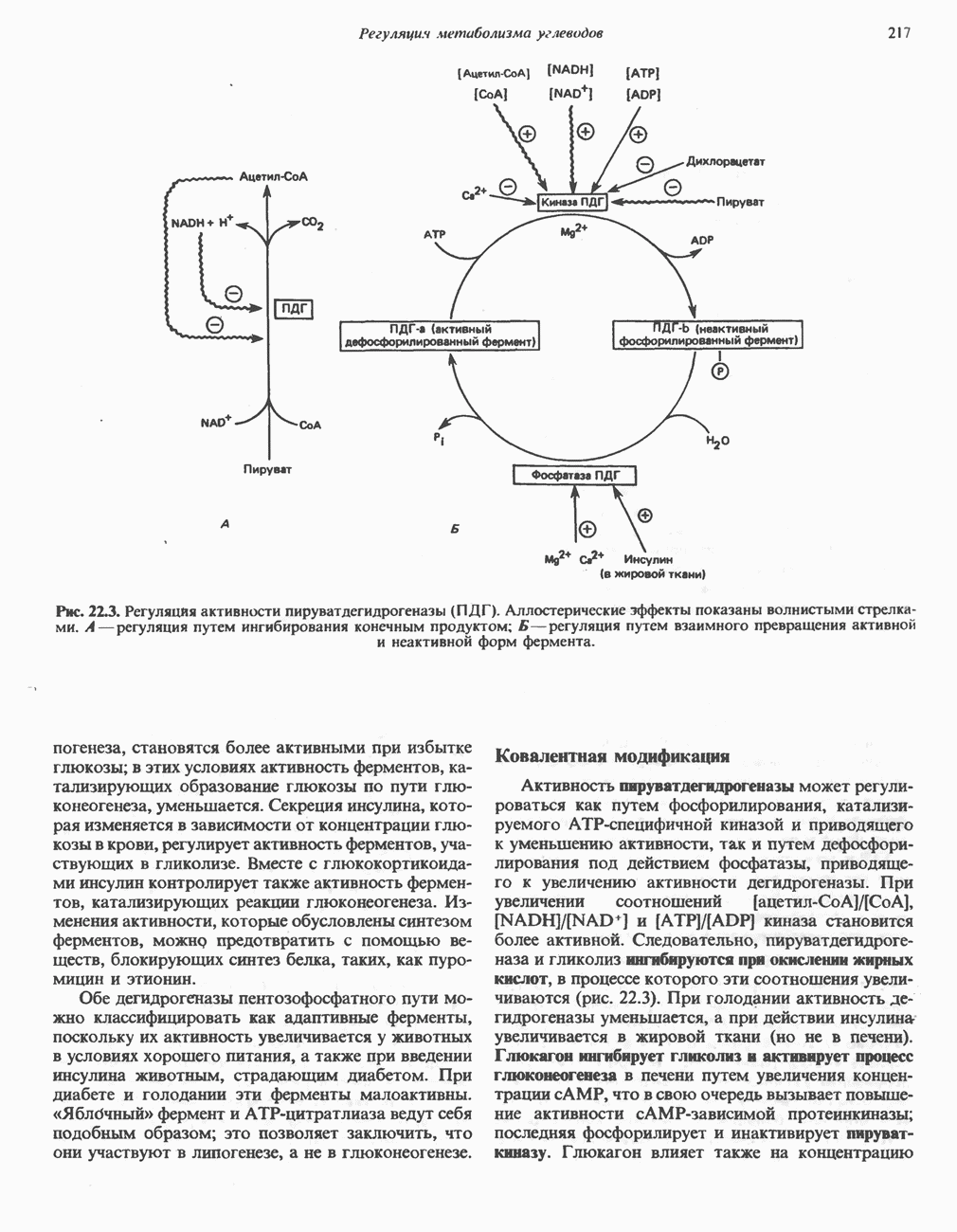
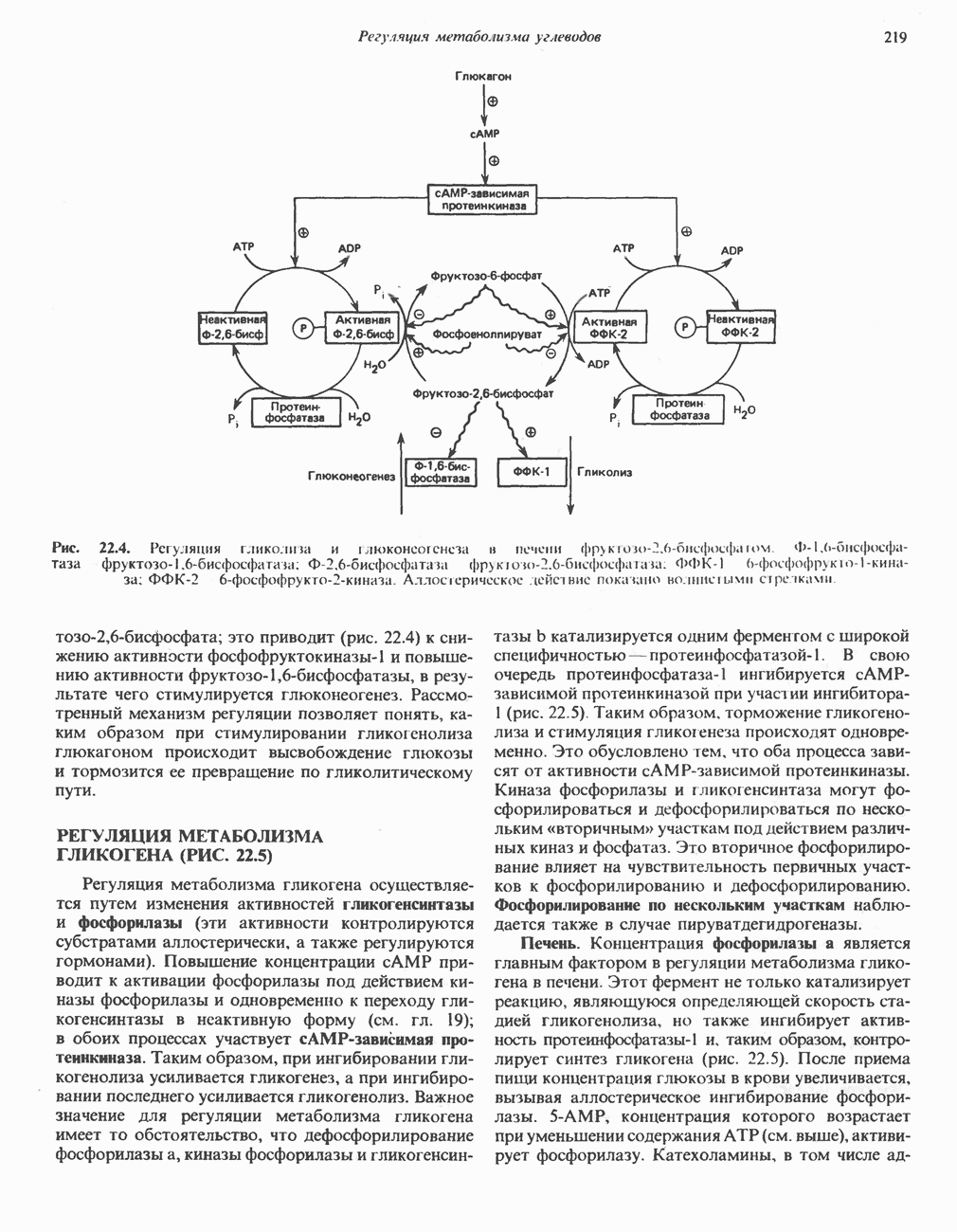
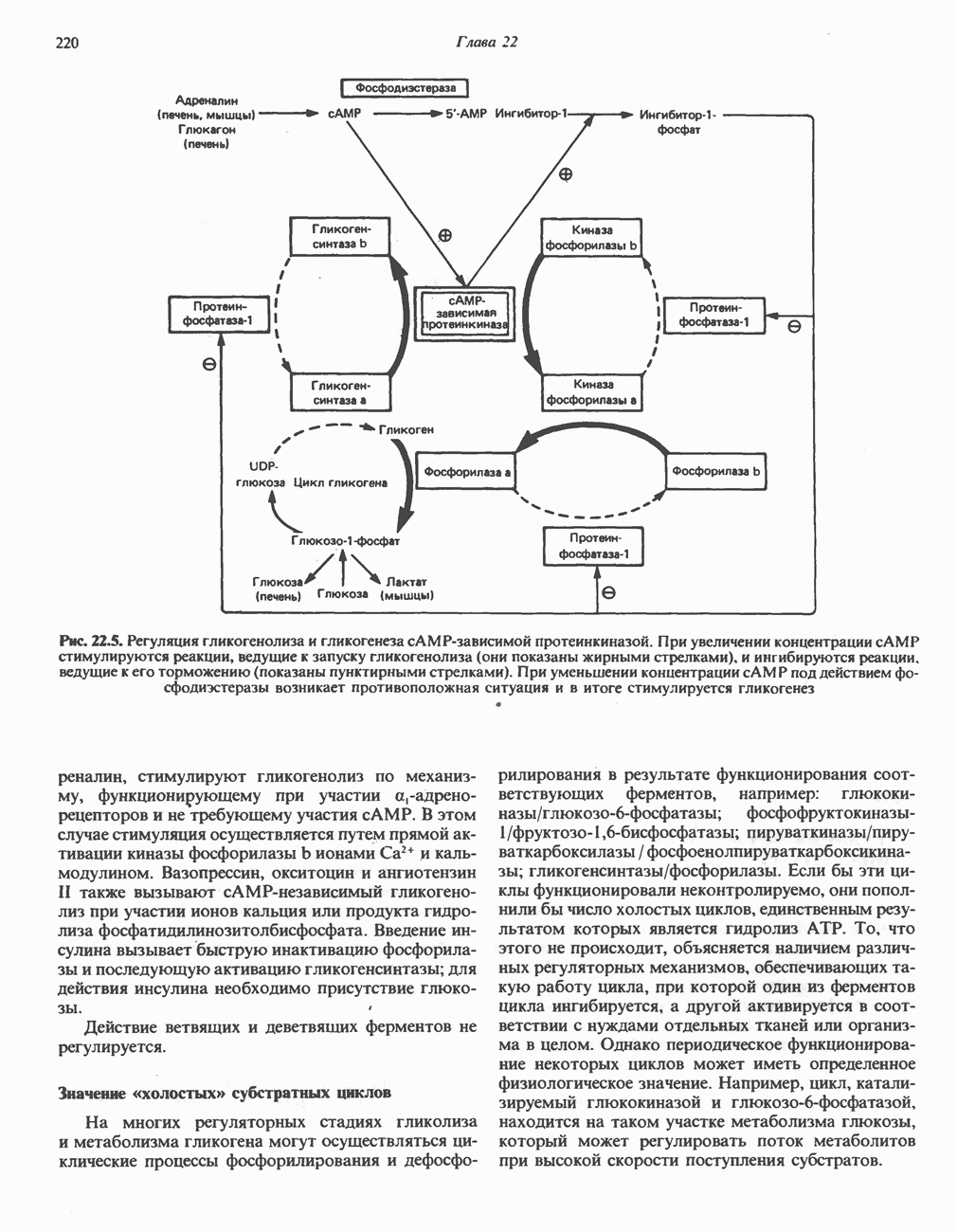
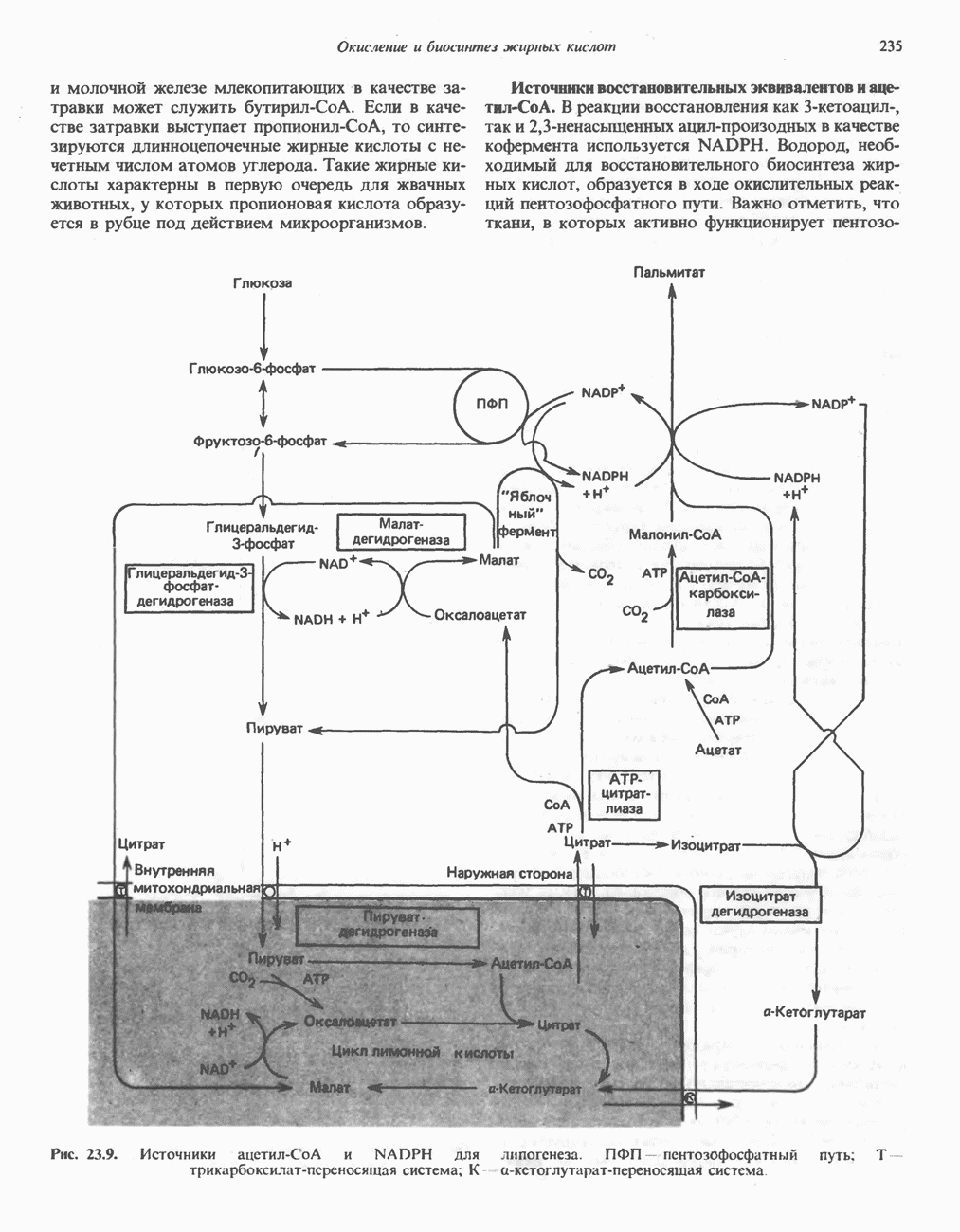
Рис. 23.8. Судьба пальмитата. и молочной железе млекопитающих в качестве затравки может служить бутирил-СоА. Если в качестве затравки выступает пропионил-СоА, то синтезируются длинноцепочечные жирные кислоты с нечетным числом атомов углерода. Такие жирные кислоты характерны в первую очередь для жвачных животных, у которых пропионовая кислота образуется в рубце под действием микроорганизмов. Источники восстановительных эквивалентов и ацетил-СоА. В реакции восстановления как 3-кетоацил-, так и 2,3-ненасыщенных ацилпроизодных в качестве кофермента используется NADPH. Водород, необходимый для восстановительного биосинтеза жирных кислот, образуется в ходе окислительных реакций пентозофосфатного пути. Важно отметить, что ткани, в которых активно функционирует пентозо- (см. скан) Рис. 23.9. Источники ацетил-СоА и NADPH для липогснеза. ПФП — пентозофосфатный путь: Т трикарбоксилат-псреносящая система; К а-кетоглутарат-переносяшая система фосфатный путь, способны эффективно осуществлять липогенез (например, печень, жировая ткань и молочная железа в период лактации). Кроме того, оба метаболических пути протекают в клетке вне митохондрий, поэтому переходу NADPH/NADP от одного метаболического пути к другому не препятствуют мембраны или другие барьеры. Другими источниками NADPH являются реакция превращения малата в пируват, катализируемая «яблочным» ферментом ( Ацетил-СоА, являющийся строительным блоком для синтеза жирных кислот, образуется в митохондриях из углеводов в результате окисления пирувата. Однако ацетил-СоА не может свободно проникать во внемитохондриальный компартмент — главное место биосинтеза жирных кислот. Активности внеми-тохондриальной АТР-цитрат-лиазы и «яблочного» фермента при хорошем питании увеличиваются У жвачных содержание АТР-цитратлиазы и «яблочного» фермента в тканях, осуществляющих липогенез, незначительно. Это связано, по-видимому, с тем, что у этих животных основным источником ацетил-СоА является ацетат, образующийся в рубце. Поскольку ацетат активируется до ацетил-СоА внемитохондриально, ему не нужно проникать в митохондрии и превращаться в цитрат, прежде чем включиться в путь биосинтеза длинноцепочечных жирных кислот. У жвачных животных из-за низкой активности «яблочного» фермента особое значение приобретает образование NADPH, катализируемое
Рис. 23.10. Микросомальная система удлинения цепи жирной кислоты (элонгазная система). внемитохондриальной изоцитратдегидрогеназой. Микросомальная система удлинения цепей жирных кислот (элонгаза)Микросомы, по-видимому, являются основным местом, где происходит удлинение, длинноцепочечных жирных кислот. Ацил-СоА-производные жирных кислот превращаются в соединения, содержащие на 2 атома углерода больше; малонил-СоА является донором ацетильной группы, a NADPH— восстановителем. Промежуточными соединениями рассматриваемого пути являются тиоэфиры СоА. Затравочными молекулами могут служить насыщенные (С10 и выше) и ненасыщенные жирные кислоты. При голодании процесс удлинения цепей жирных кислот затормаживается. При образовании миелиновых оболочек нервных клеток в мозгу резко усиливается процесс удлинения стеарил-СоА, в результате образуются ЛИТЕРАТУРАBoyer P. D. (ed.). The Enzymes, 3rd ed.. Vol. 16 of Lipid Enzymology, Academic Press, 1983. - Debeer L. J., Mannaerts G. P. The mitochondrial and peroxisomal pathways of fatty acid oxidation in rat liver, Diabete Metab. (Paris), 1983, 9, 134. Goodridge A.G. Fatty acid synthesis in eukaryotes, Page 143. In: Biochemistry of Lipids and Membranes, Vance D. E., Vance J. E. (eds.), Benjamin/Cummings, 1985. Gurr M.I., James A.I. Lipid Biochemistry: An Introduction, 3rd ed., Wiley, 1980. Pande S. V., Parvin R. Page 143. In: Carnitine Biosynthesis, Metabolism, and Functions, Frenkel R. A., McGarry J. D. (eds.), Academic Press, 1980. Schulz H. Oxidation of fatty acids, Page 116. In: Biochemistry of Lipids and Membranes, Vance D. E., Vance J. E. (eds.), Benjamin/Cummings, 1985. Singh N.. Wak.il S.J., Stoops J.K. On the question of half- or fullsite reactivity of animal fatty acid synthetase, J. Biol. Chem., 1984, 259, 3605. Tsukamoto Y. et al. The architecture of the animal fatty acid synthetase complex, J. Biol. Chem., 1983, 258, 15312. Various authors. Disorders characterized by evidence of abnormal lipid metabolism. In: The Metabolic Basis of Inherited Disease, 5th ed., Stanbury J. B. et al. (eds.), McGraw-Hill, 1983.
|
 |
Оглавление
|























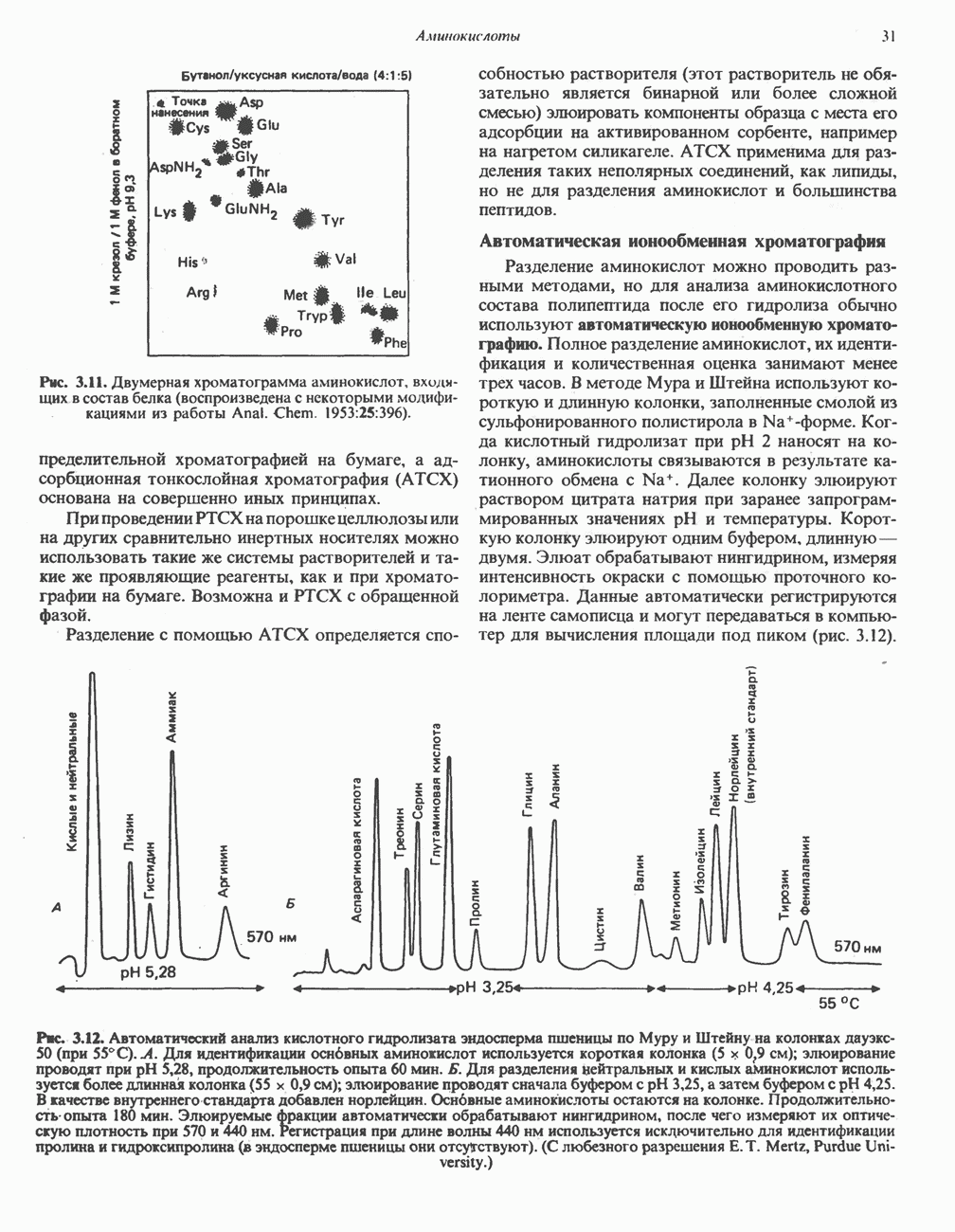












































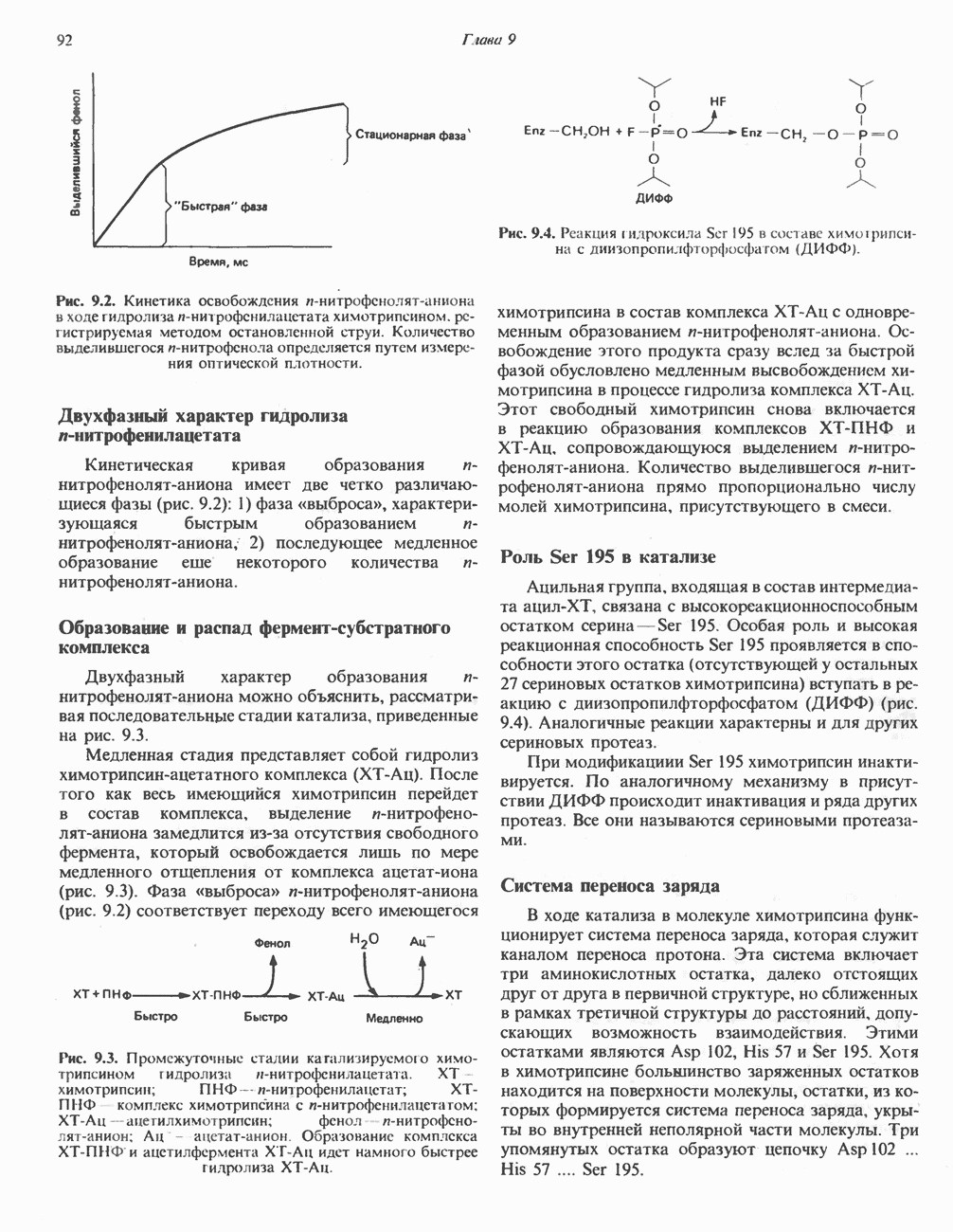



















































































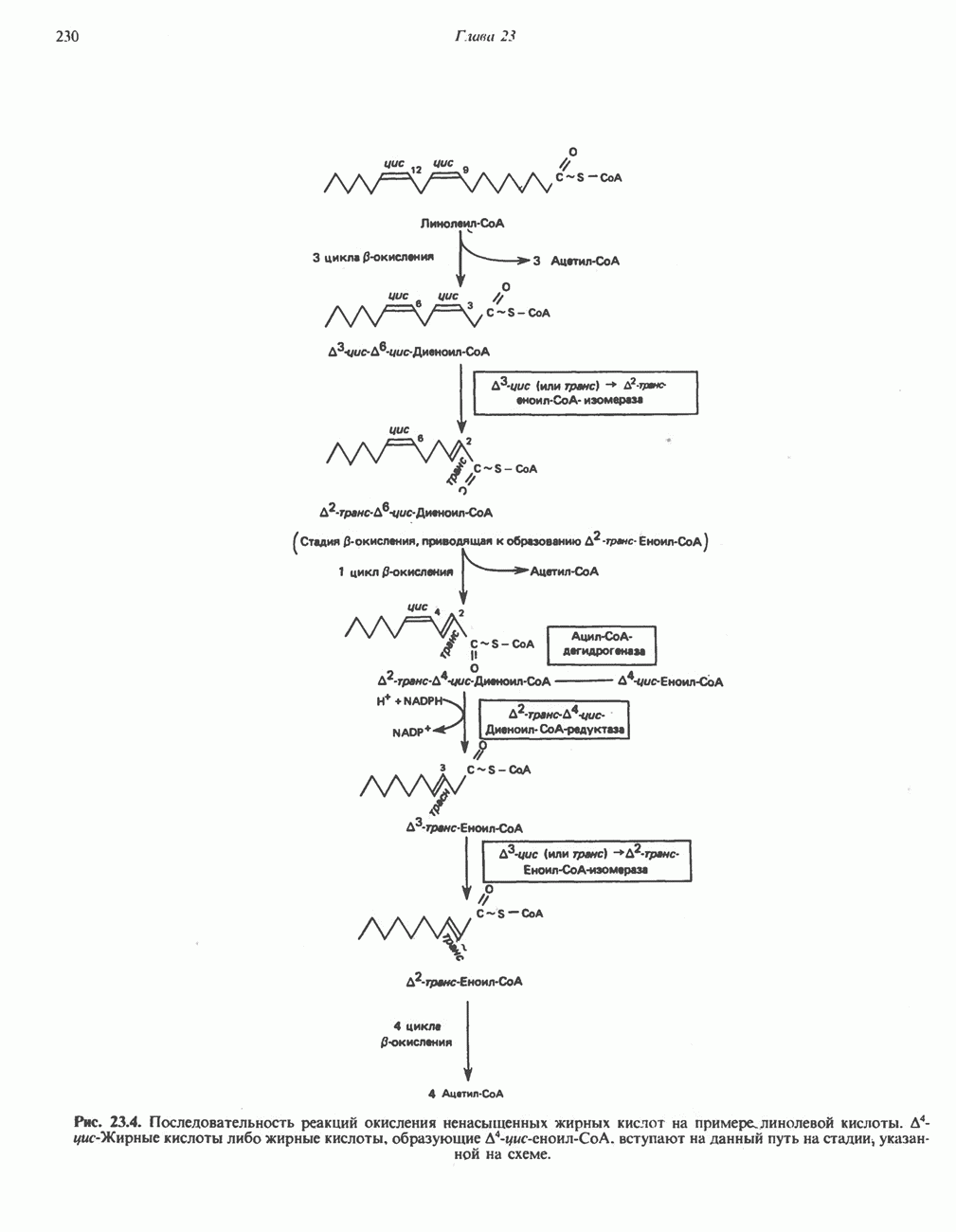
















































































 -окисления, осуществляет только удлинение уже существующих в организме среднецепочечных жирных кислот, в то время как полный биосинтез пальмитиновой
-окисления, осуществляет только удлинение уже существующих в организме среднецепочечных жирных кислот, в то время как полный биосинтез пальмитиновой  активно протекает вне митохондрий по совершенно другому пути. Активная система, обеспечивающая удлинение цепей
активно протекает вне митохондрий по совершенно другому пути. Активная система, обеспечивающая удлинение цепей  качестве источника
качестве источника  ); субстратом является
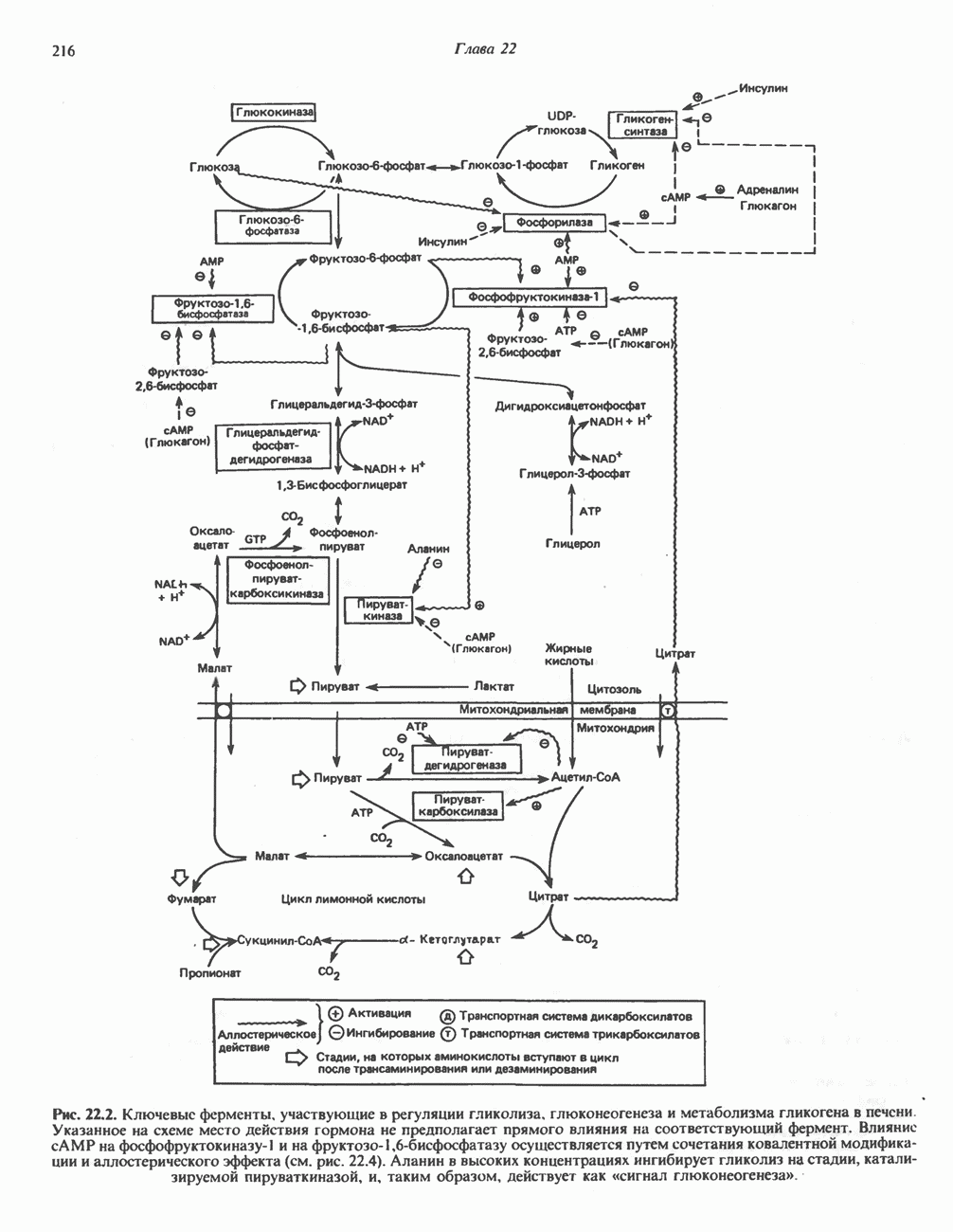
); субстратом является  конечным продуктом — пальмитиновая
конечным продуктом — пальмитиновая  -окисления значительно различаются.
-окисления значительно различаются.
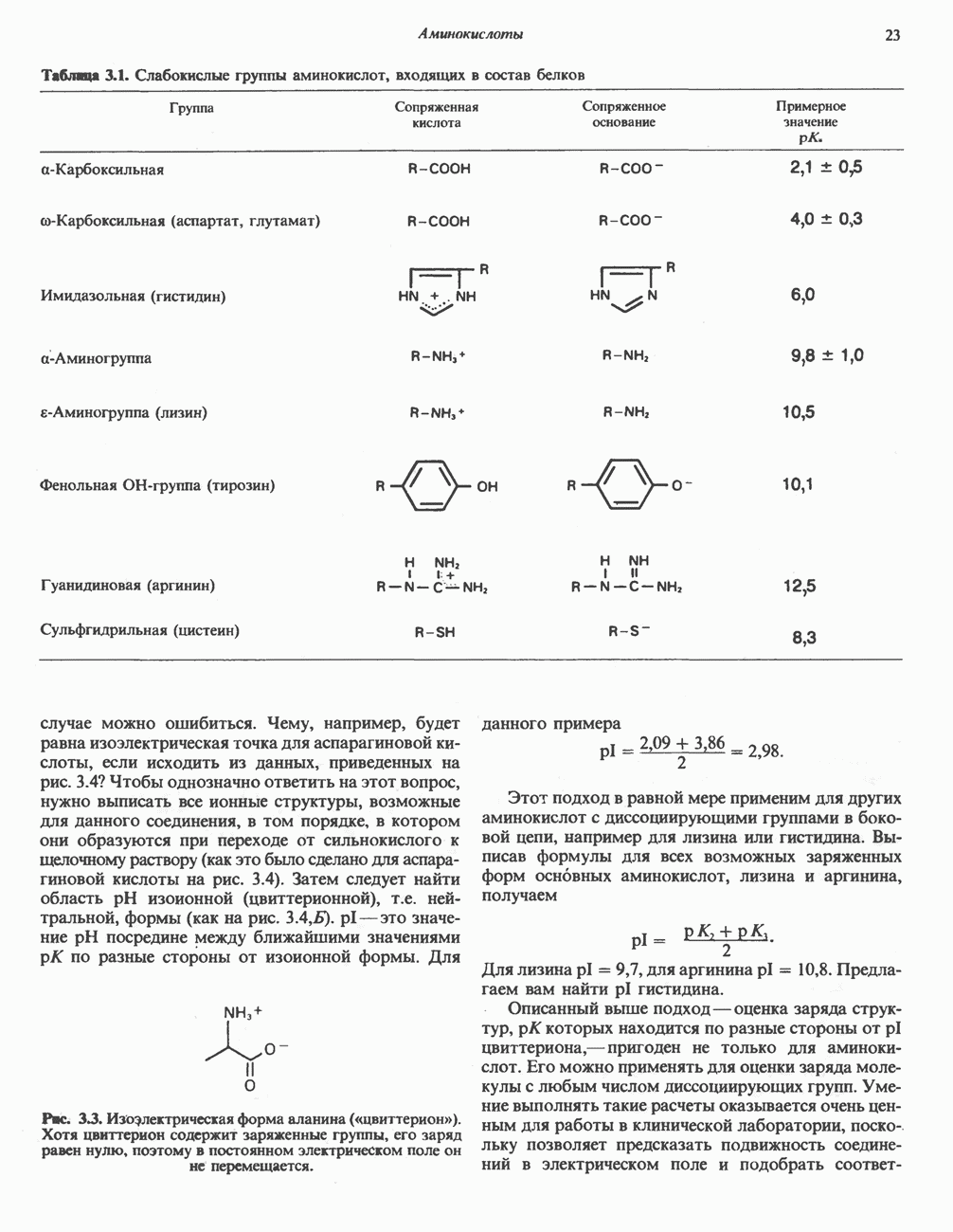
 арбоксилазой и осуществляемой за счет энергии АТР, является карбоксилирование
арбоксилазой и осуществляемой за счет энергии АТР, является карбоксилирование  источником
источником  является бикарбонат. Для функционирования фермента необходим
является бикарбонат. Для функционирования фермента необходим  активируется цитратом и ингибируется длинноцепочечными
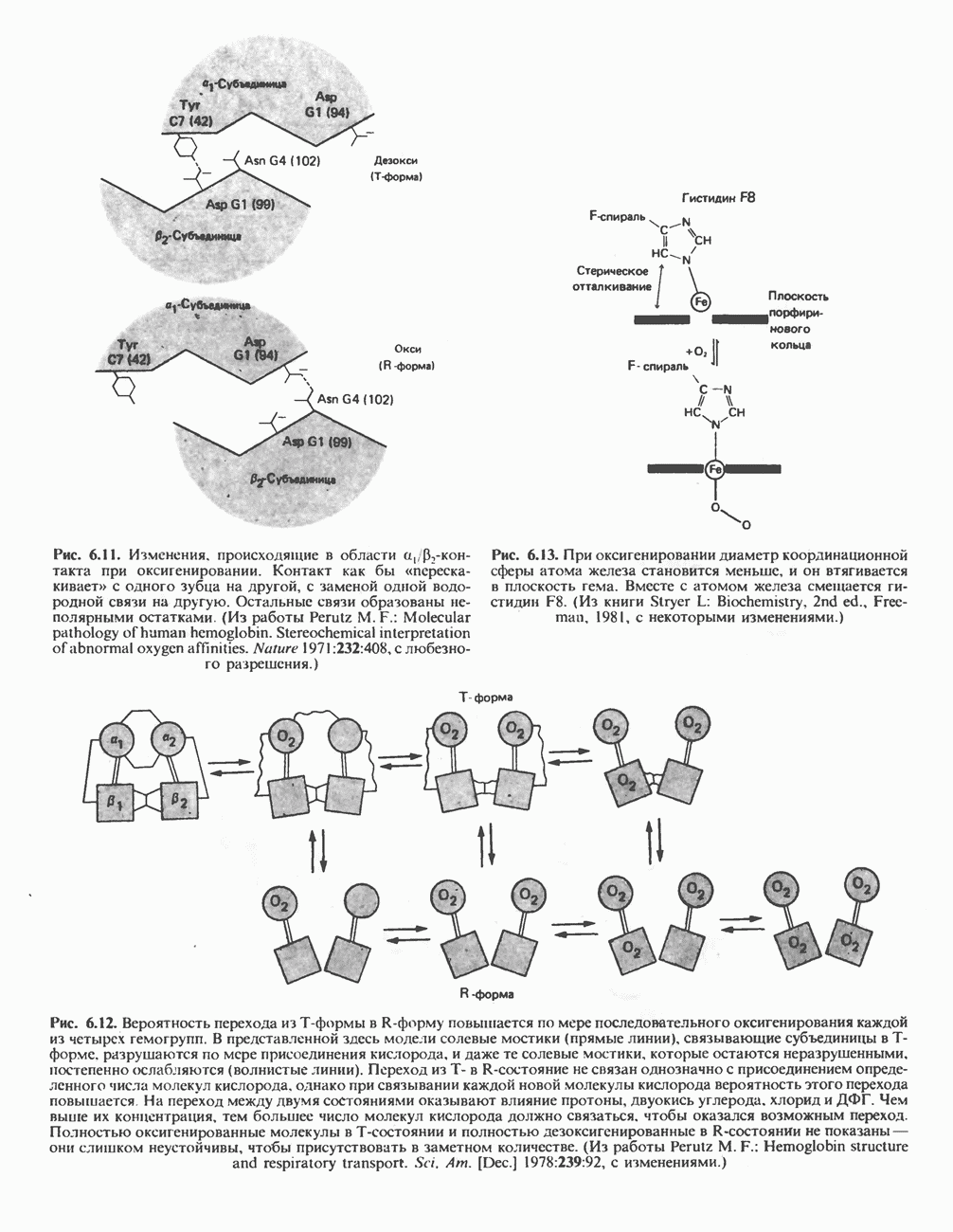
активируется цитратом и ингибируется длинноцепочечными  Активированная форма фермента легко полимеризуется с образованием нитей, состоящих из 10—20 протомеров.
Активированная форма фермента легко полимеризуется с образованием нитей, состоящих из 10—20 протомеров.


 - индивидуальные мономеры синтазы жирных кислот. На одном димере одновременно синтезируются 2 ацильные цепи, при этом используется 2 пары -
- индивидуальные мономеры синтазы жирных кислот. На одном димере одновременно синтезируются 2 ацильные цепи, при этом используется 2 пары -  -групп; в каждой паре одна из групп принадлежит Фп, а другая - Cys.
-групп; в каждой паре одна из групп принадлежит Фп, а другая - Cys.
 -группой, принадлежащей
-группой, принадлежащей  -фосфопантетеину. В непосредственной близости от этой группы расположена другая сульфгидрильная группа, принадлежащая остатку цистеина, входящего в состав
-фосфопантетеину. В непосредственной близости от этой группы расположена другая сульфгидрильная группа, принадлежащая остатку цистеина, входящего в состав  -кетоацил-синтазы (конденсирующего фермента), которая входит в состав другого мономера (рис. 23.6). Поскольку для проявления синтазной активности необходимо участие обеих сульфгидрильных групп, синтазный комплекс активен только в виде димера.
-кетоацил-синтазы (конденсирующего фермента), которая входит в состав другого мономера (рис. 23.6). Поскольку для проявления синтазной активности необходимо участие обеих сульфгидрильных групп, синтазный комплекс активен только в виде димера.
 при участии трансацилазы взаимодействует с
при участии трансацилазы взаимодействует с  -группой цистеина
-группой цистеина  под действием того же фермента (трансацилазы) взаимодействует с соседней
под действием того же фермента (трансацилазы) взаимодействует с соседней  -группой, принадлежащей
-группой, принадлежащей  -фосфопантетеину, локализованному в АПБ другого мономера. В результате этой реакции образуется ацетил (ацил) малонил-фермент. З-Кетоацилсннтаза катализирует взаимодействие ацетильной группы фермента с метиленовой группой малонила и высвобождение
-фосфопантетеину, локализованному в АПБ другого мономера. В результате этой реакции образуется ацетил (ацил) малонил-фермент. З-Кетоацилсннтаза катализирует взаимодействие ацетильной группы фермента с метиленовой группой малонила и высвобождение  в результате образуется
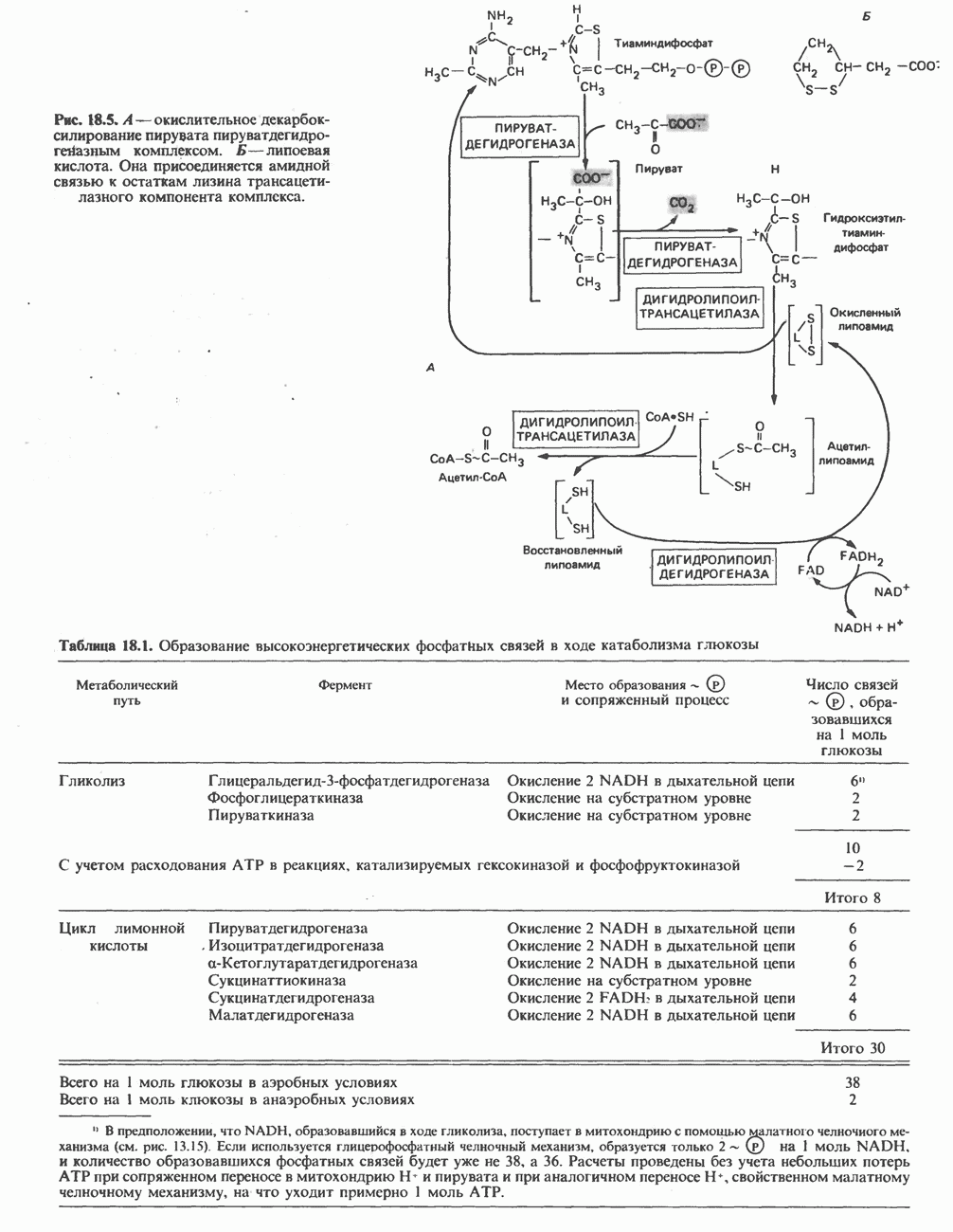
в результате образуется  -кетоацил-фермент (ацетоацетил-фермент); при этом освобождается сульфгидрильная группа цистеина, ранее занятая ацетильной группой. Декарбоксилирование позволяет реакции пройти до конца и является движущей силой биосинтеза. 3-Кетоацильная группа восстанавливается, затем дегидратируется и вновь восстанавливается, в результате образуется соответствующий насыщенный ацил-8-фермент. Эти реакции сходны с соответствующими реакциями Р-окисления; отличие заключается, в частности, в том, что при биосинтезе образуется D(-)-изомер 3-гидроксикислоты, а не
-кетоацил-фермент (ацетоацетил-фермент); при этом освобождается сульфгидрильная группа цистеина, ранее занятая ацетильной группой. Декарбоксилирование позволяет реакции пройти до конца и является движущей силой биосинтеза. 3-Кетоацильная группа восстанавливается, затем дегидратируется и вновь восстанавливается, в результате образуется соответствующий насыщенный ацил-8-фермент. Эти реакции сходны с соответствующими реакциями Р-окисления; отличие заключается, в частности, в том, что при биосинтезе образуется D(-)-изомер 3-гидроксикислоты, а не  кроме того, NADPH, а не NADH является донором водорода в реакциях восстановления. Далее новая
кроме того, NADPH, а не NADH является донором водорода в реакциях восстановления. Далее новая  взаимодействует с —
взаимодействует с — -группой фосфопантетеина, при этом насыщенный ацильный остаток перемещается на свободную —
-группой фосфопантетеина, при этом насыщенный ацильный остаток перемещается на свободную — -группу цистеина. Цикл реакций повторяется еще 6 раз, и каждый новый остаток малоната встраивается в углеродную цепь, до тех пор пока не образуется насыщенный 16-углеродный ацилрадикал (пальмитоил). Последний высвобождается из полиферментного комплекса под действием шестого фермента, входящего в состав комплекса, - тиоэстеразы (деацилазы). Свободная пальмитиновая
-группу цистеина. Цикл реакций повторяется еще 6 раз, и каждый новый остаток малоната встраивается в углеродную цепь, до тех пор пока не образуется насыщенный 16-углеродный ацилрадикал (пальмитоил). Последний высвобождается из полиферментного комплекса под действием шестого фермента, входящего в состав комплекса, - тиоэстеразы (деацилазы). Свободная пальмитиновая  Затем активированный пальмитат обычно подвергается эстерификации с образованием ацилглицеролов (рис. 23.8).
Затем активированный пальмитат обычно подвергается эстерификации с образованием ацилглицеролов (рис. 23.8).
 или
или  -жирных кислот, входящих в состав липидов молока. В молочной железе жвачных животных этот
-жирных кислот, входящих в состав липидов молока. В молочной железе жвачных животных этот 
 выступающей в качестве затравки, образуются 15-й и 16-й углеродные атомы пальмитиновой
выступающей в качестве затравки, образуются 15-й и 16-й углеродные атомы пальмитиновой  В печени
В печени

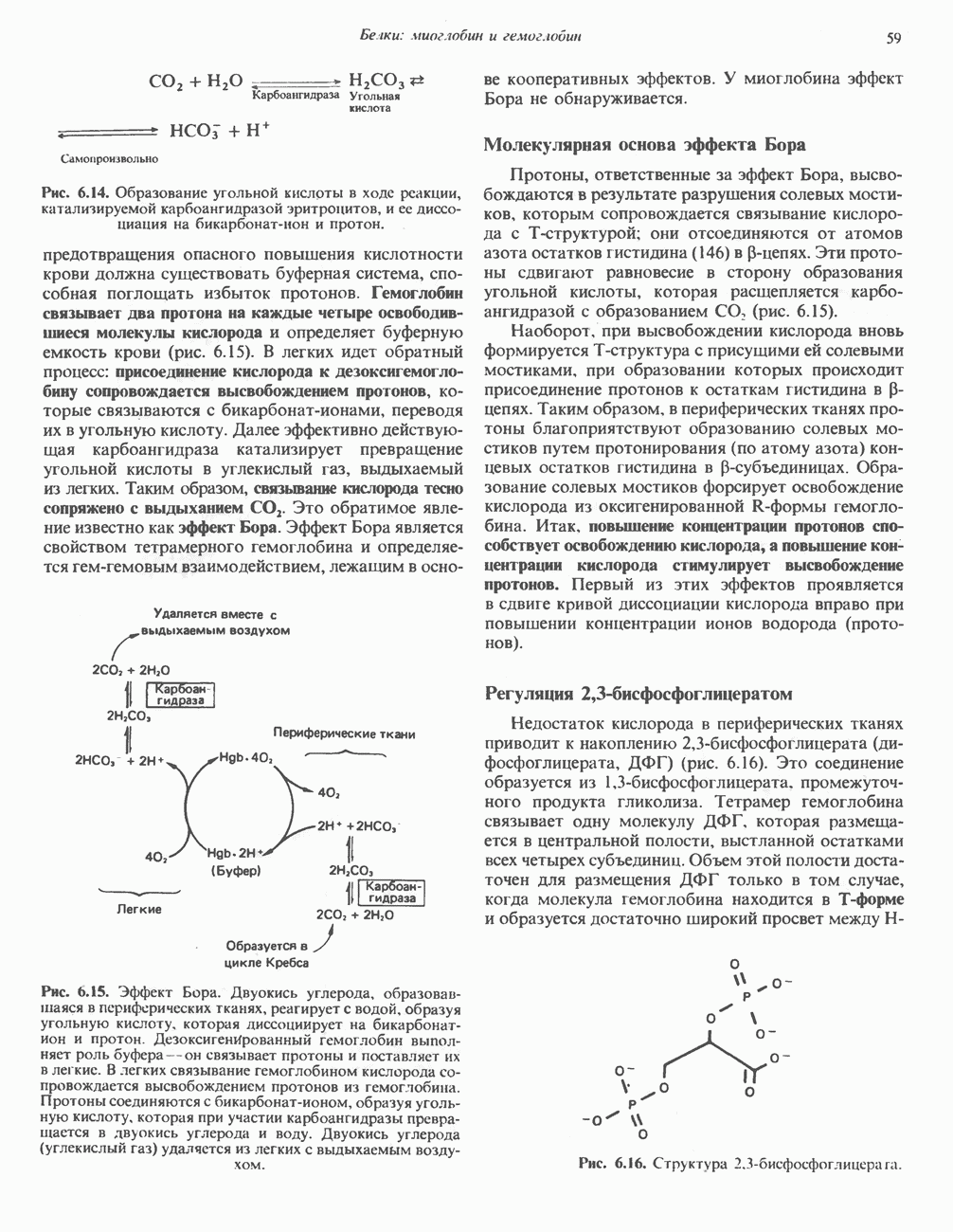
 -малатдегидрогеназой) (рис. 23.9), а также внемитохондриальная реакция, катализируемая нзоцитратдегидрогеназой (вероятно, роль ее незначительна).
-малатдегидрогеназой) (рис. 23.9), а также внемитохондриальная реакция, катализируемая нзоцитратдегидрогеназой (вероятно, роль ее незначительна).
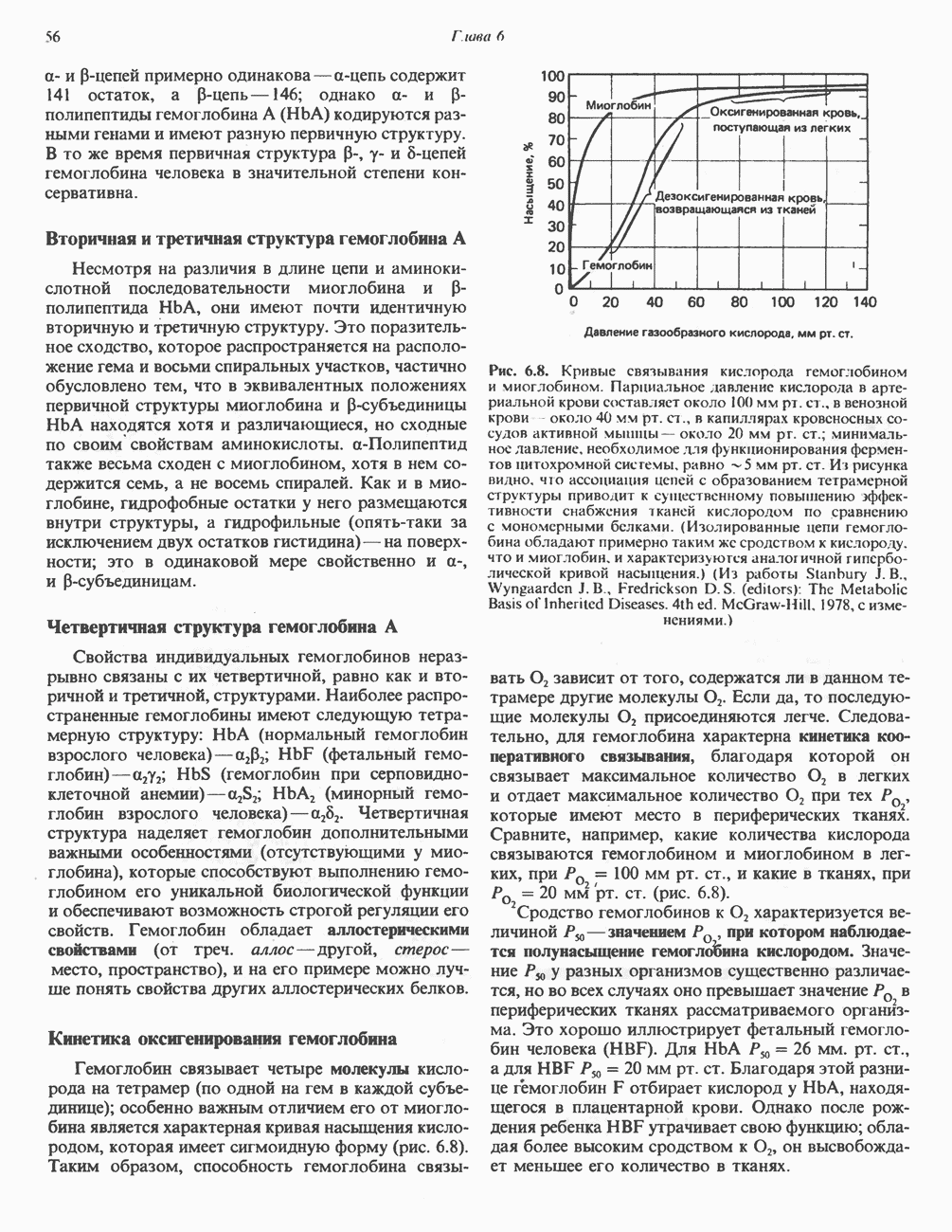
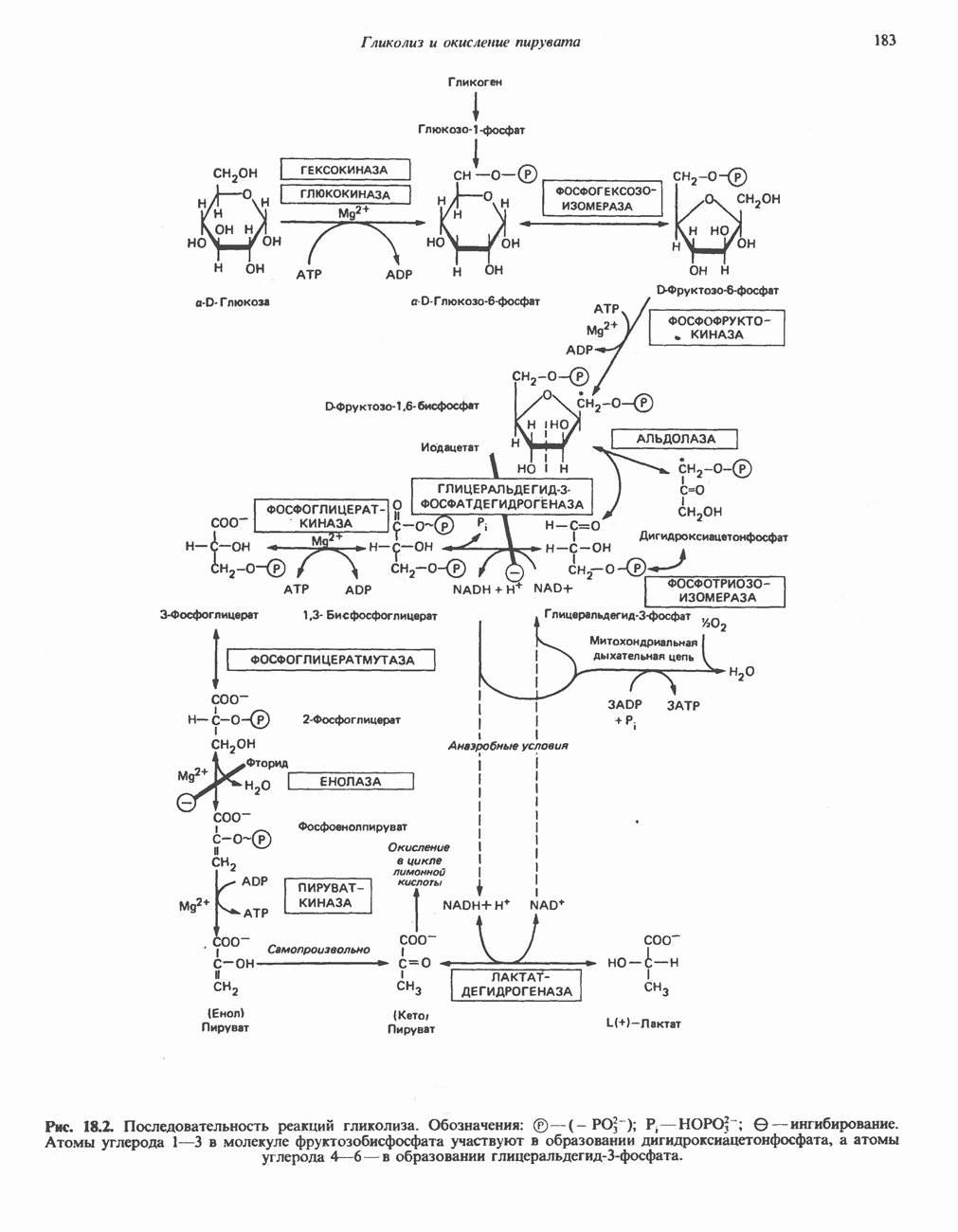
 -раллельно активностям ферментов, участвующих в биосинтезе жирных кислот. В настоящее время полагают, что путь использования пирувата в процессе липогенеза проходит через стадию образования цитрата. Этот метаболический путь включает гликолиз, затем окислительное декарбоксилирование пирувата до ацетил-СоА в митохондриях и последующую реакцию
-раллельно активностям ферментов, участвующих в биосинтезе жирных кислот. В настоящее время полагают, что путь использования пирувата в процессе липогенеза проходит через стадию образования цитрата. Этот метаболический путь включает гликолиз, затем окислительное декарбоксилирование пирувата до ацетил-СоА в митохондриях и последующую реакцию 
 -жирные
-жирные