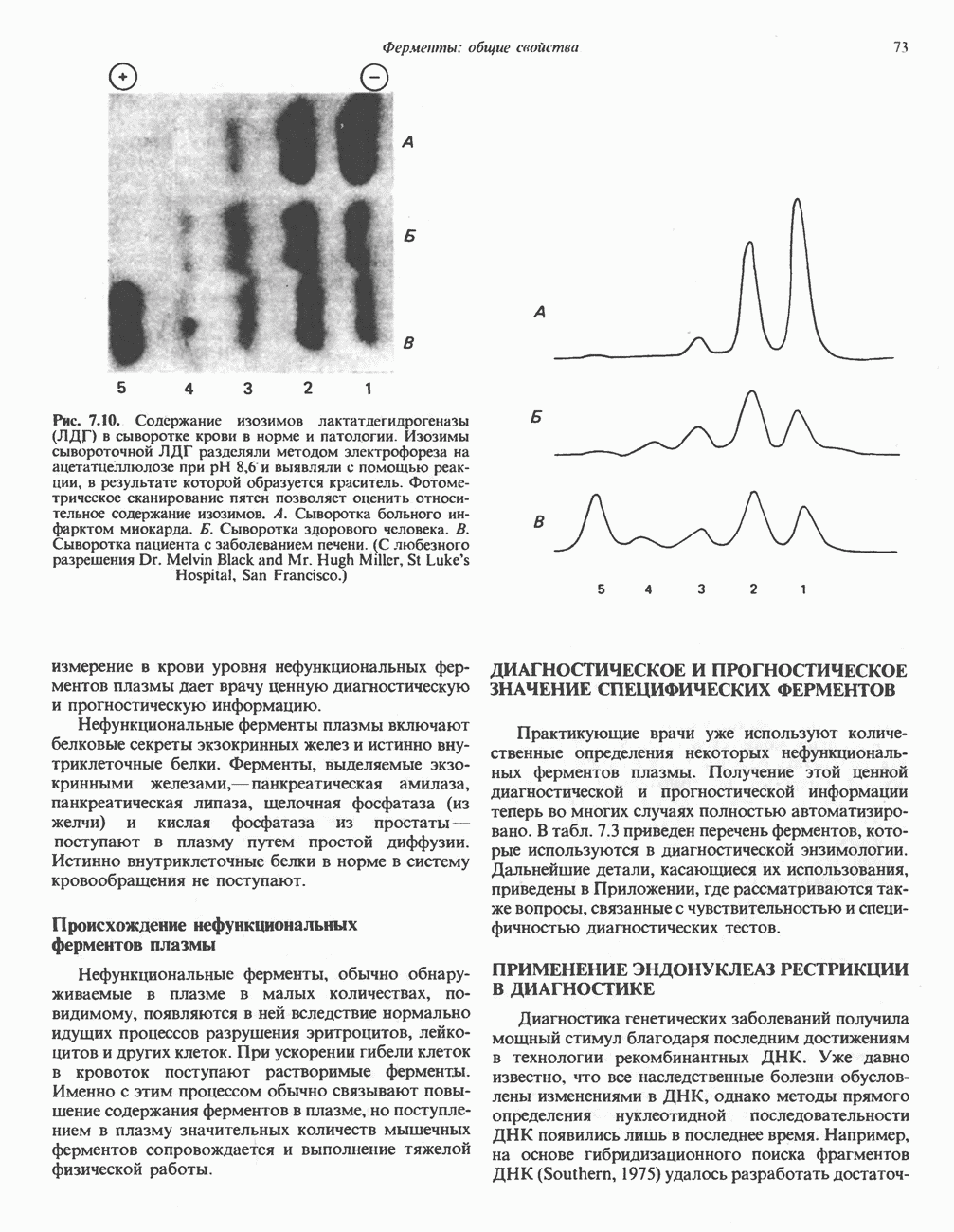
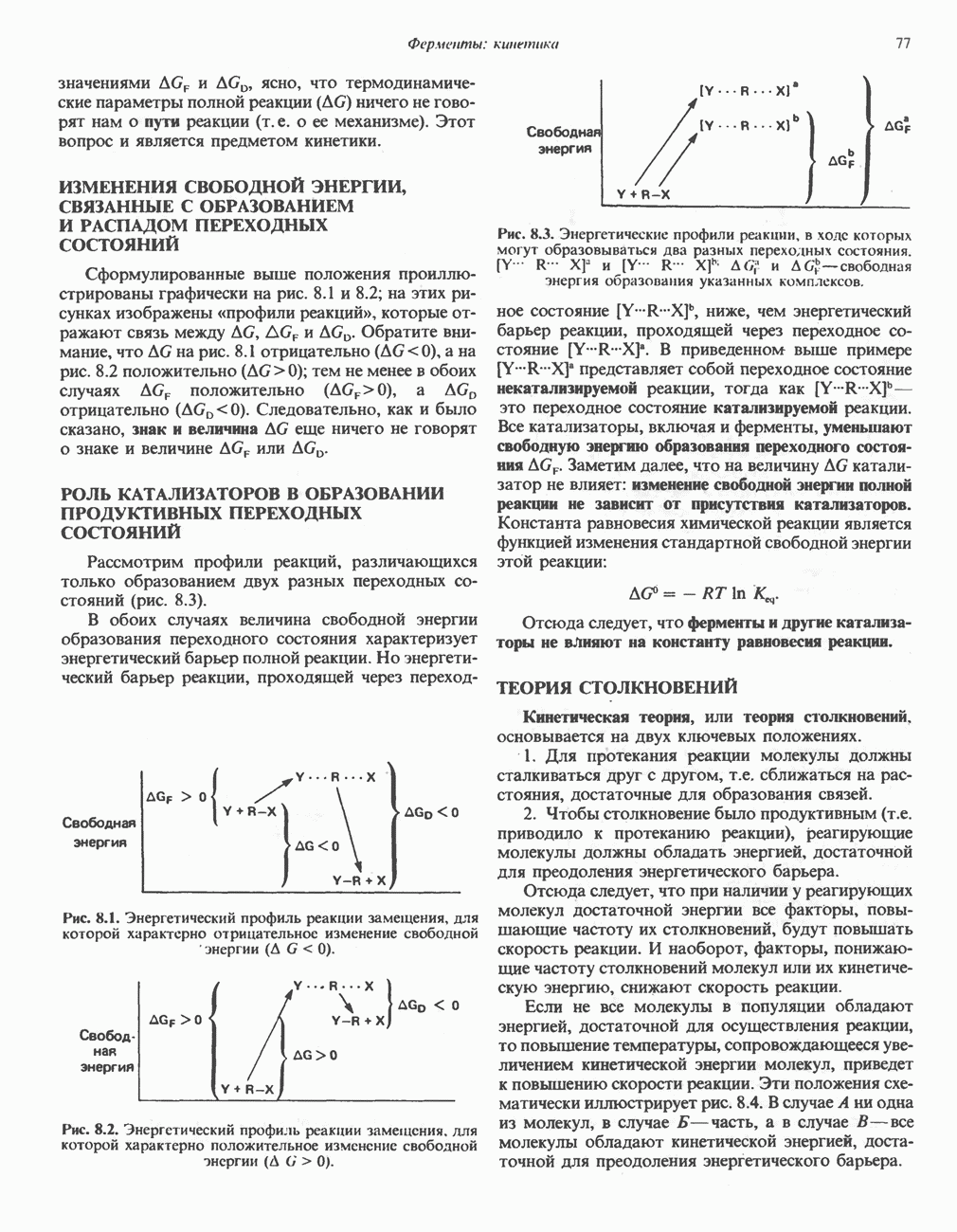
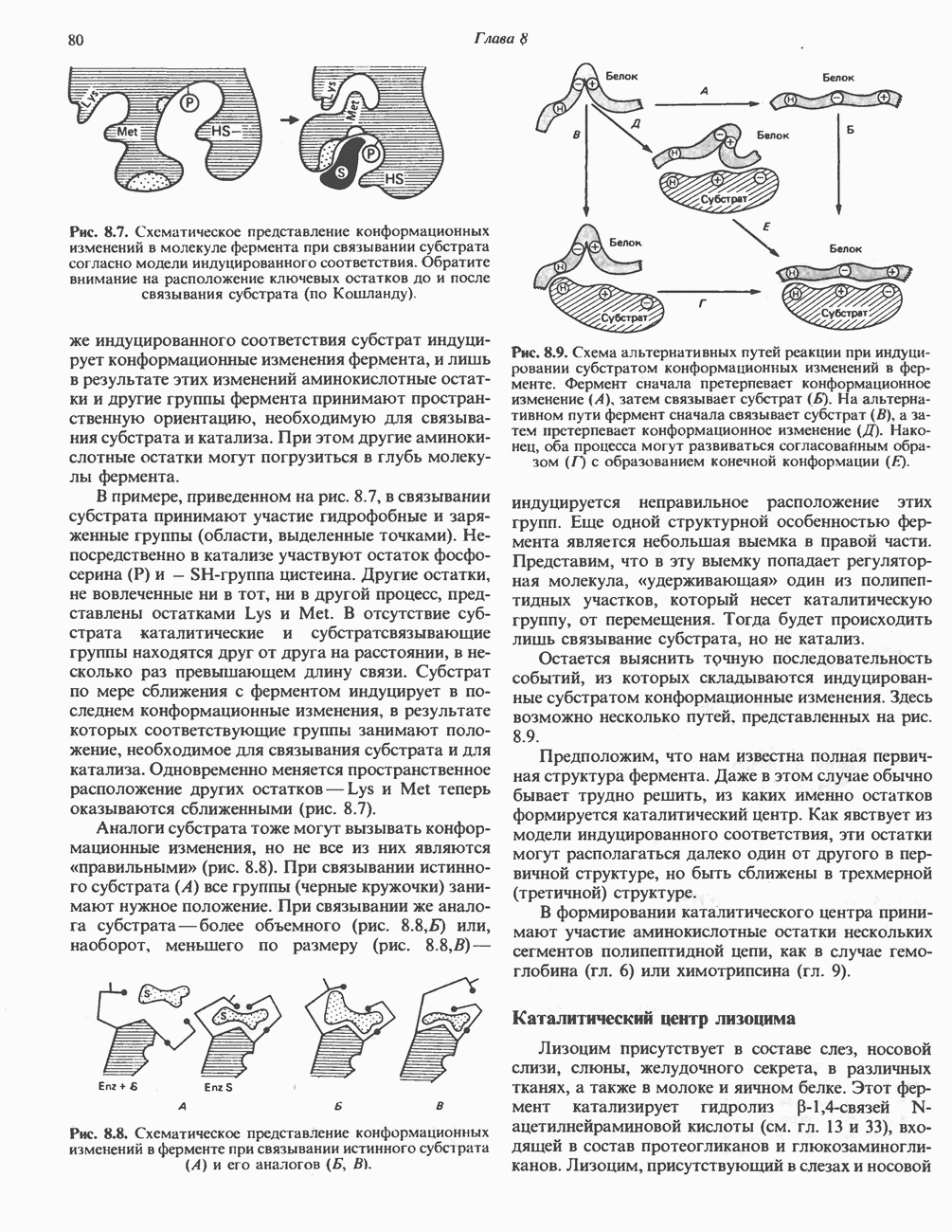
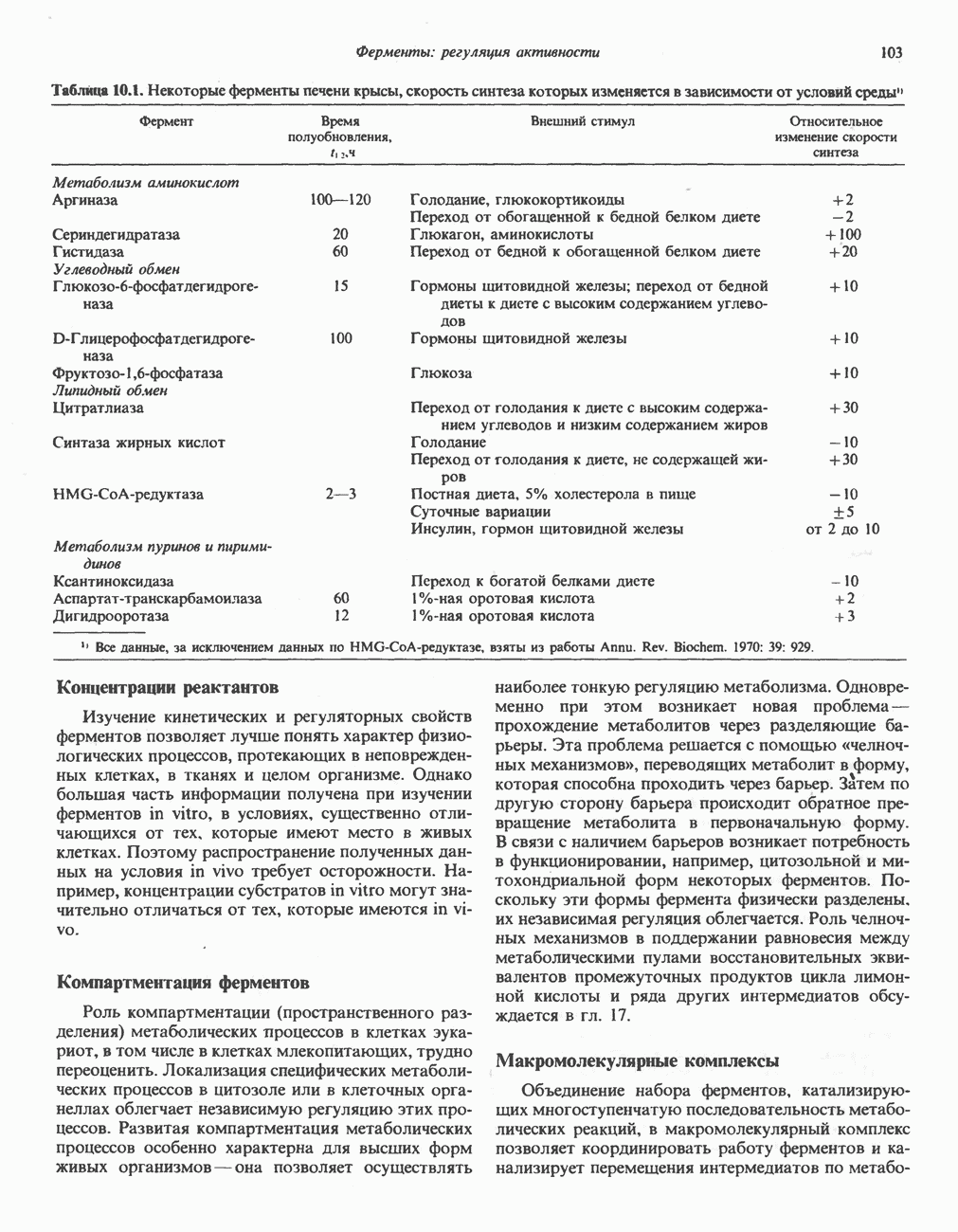
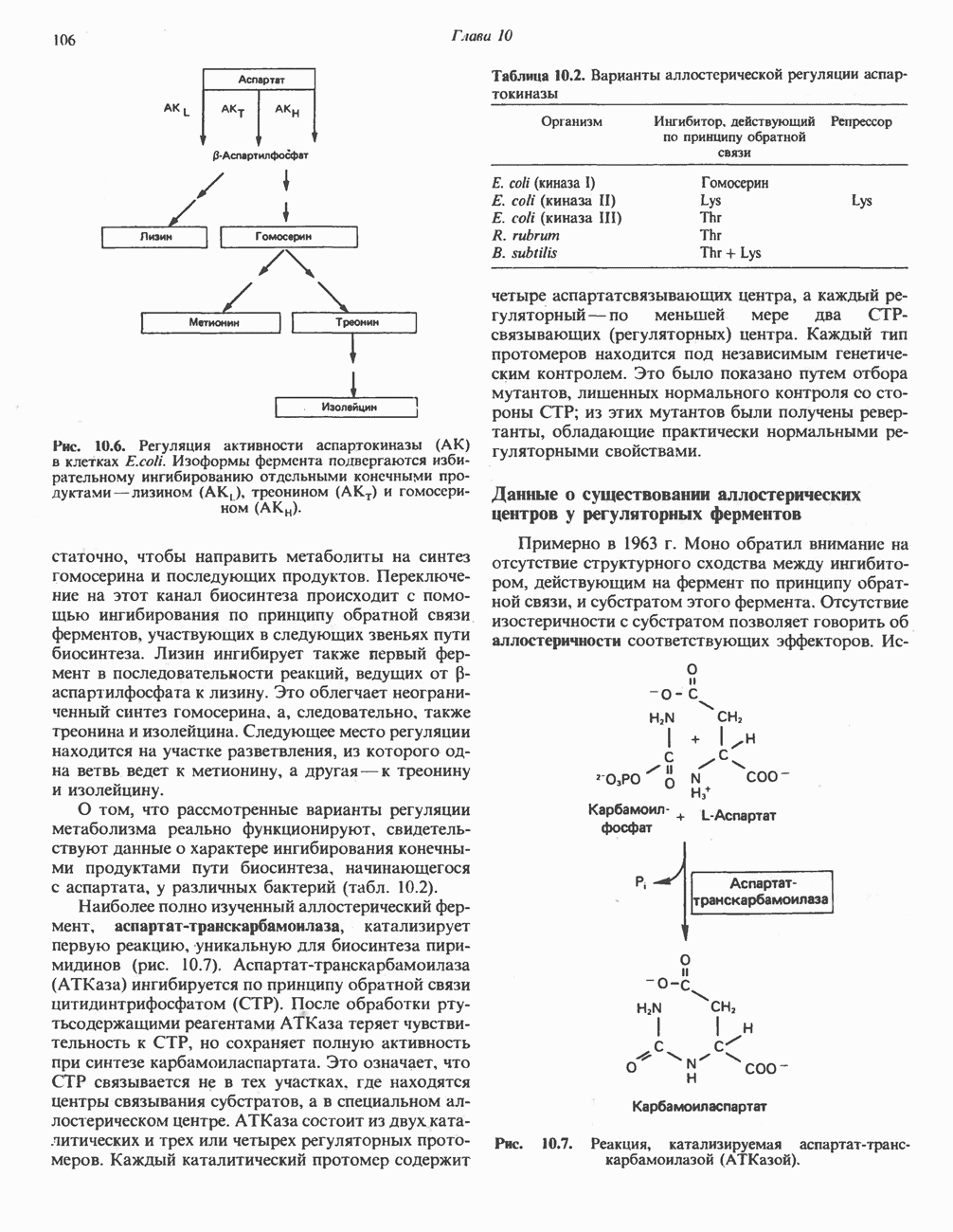
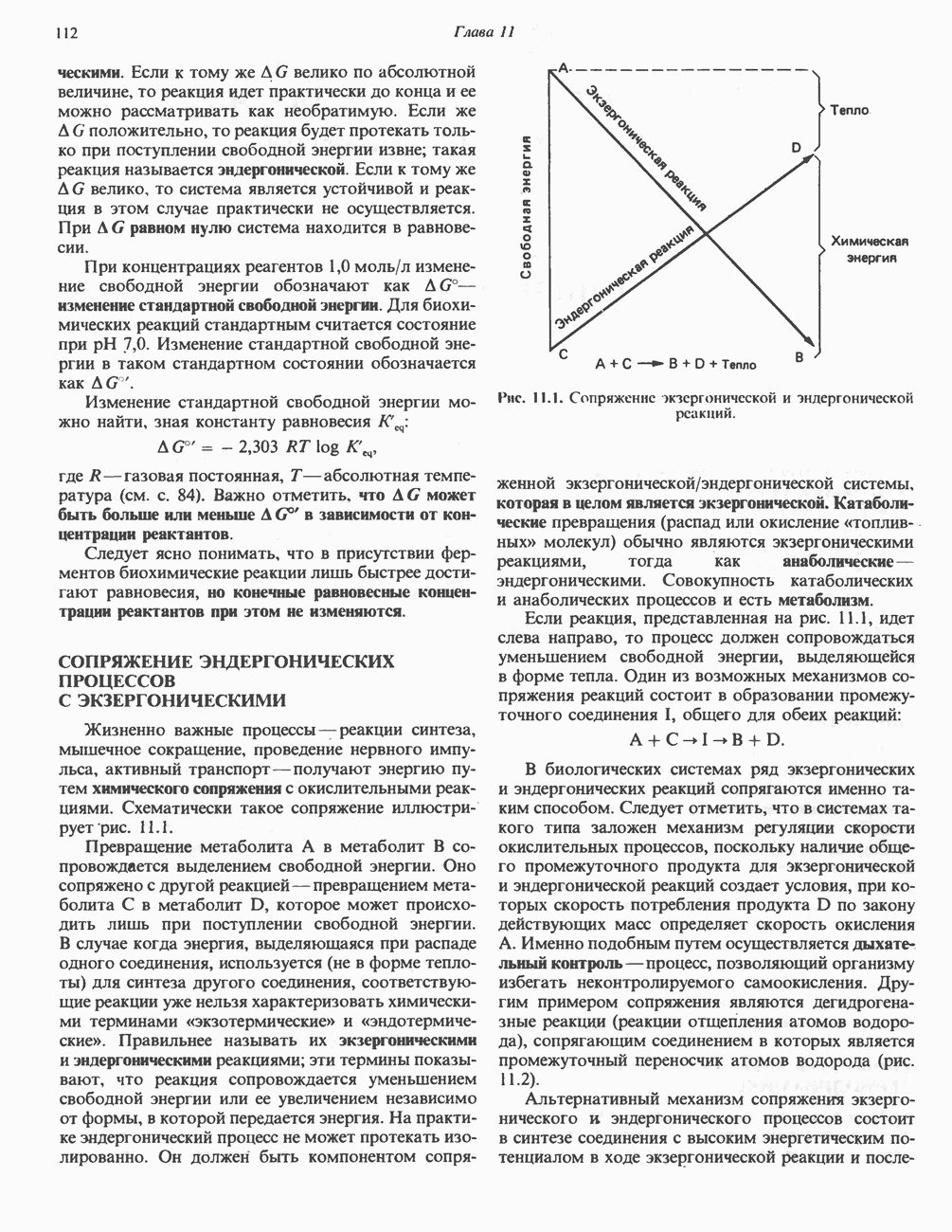
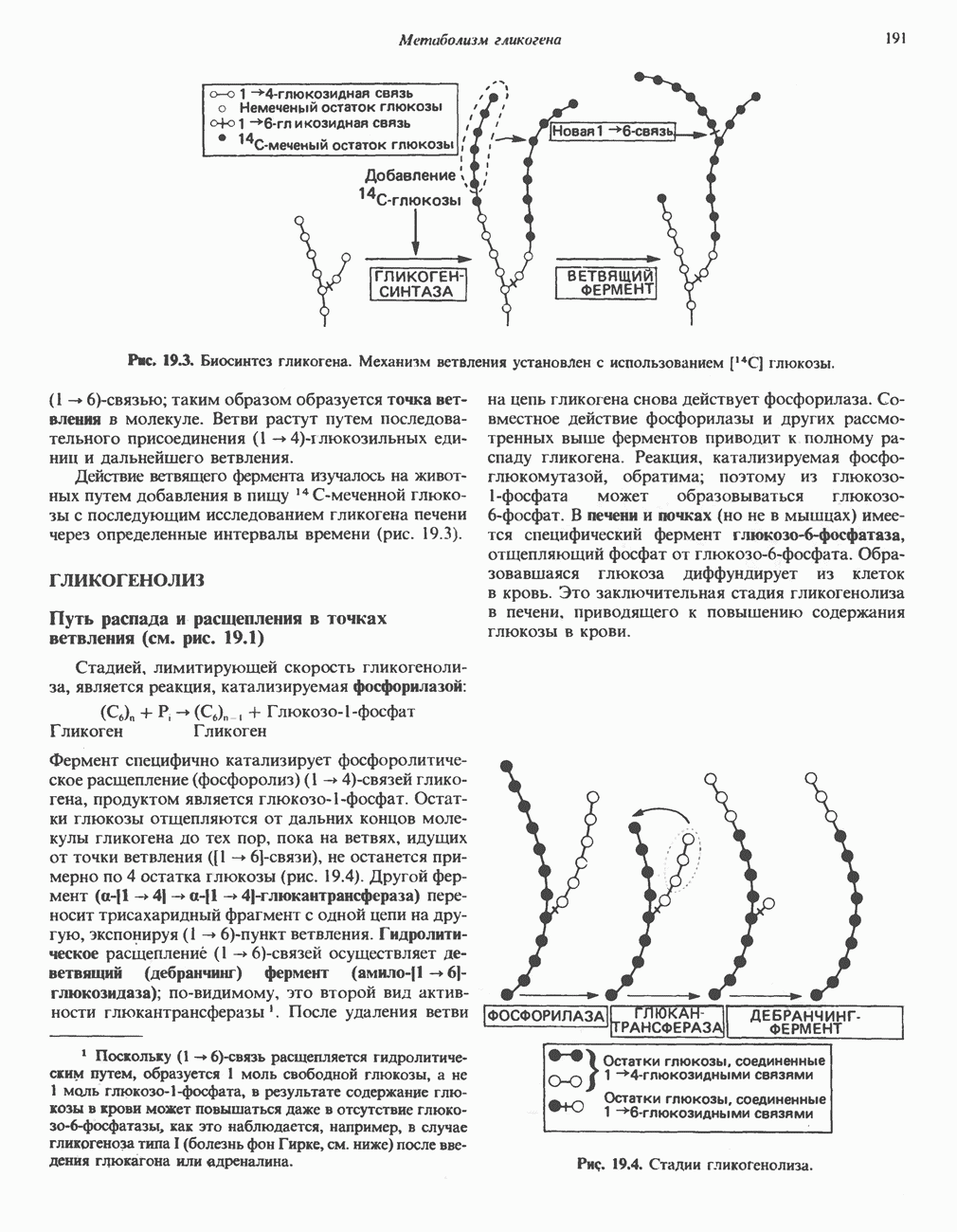
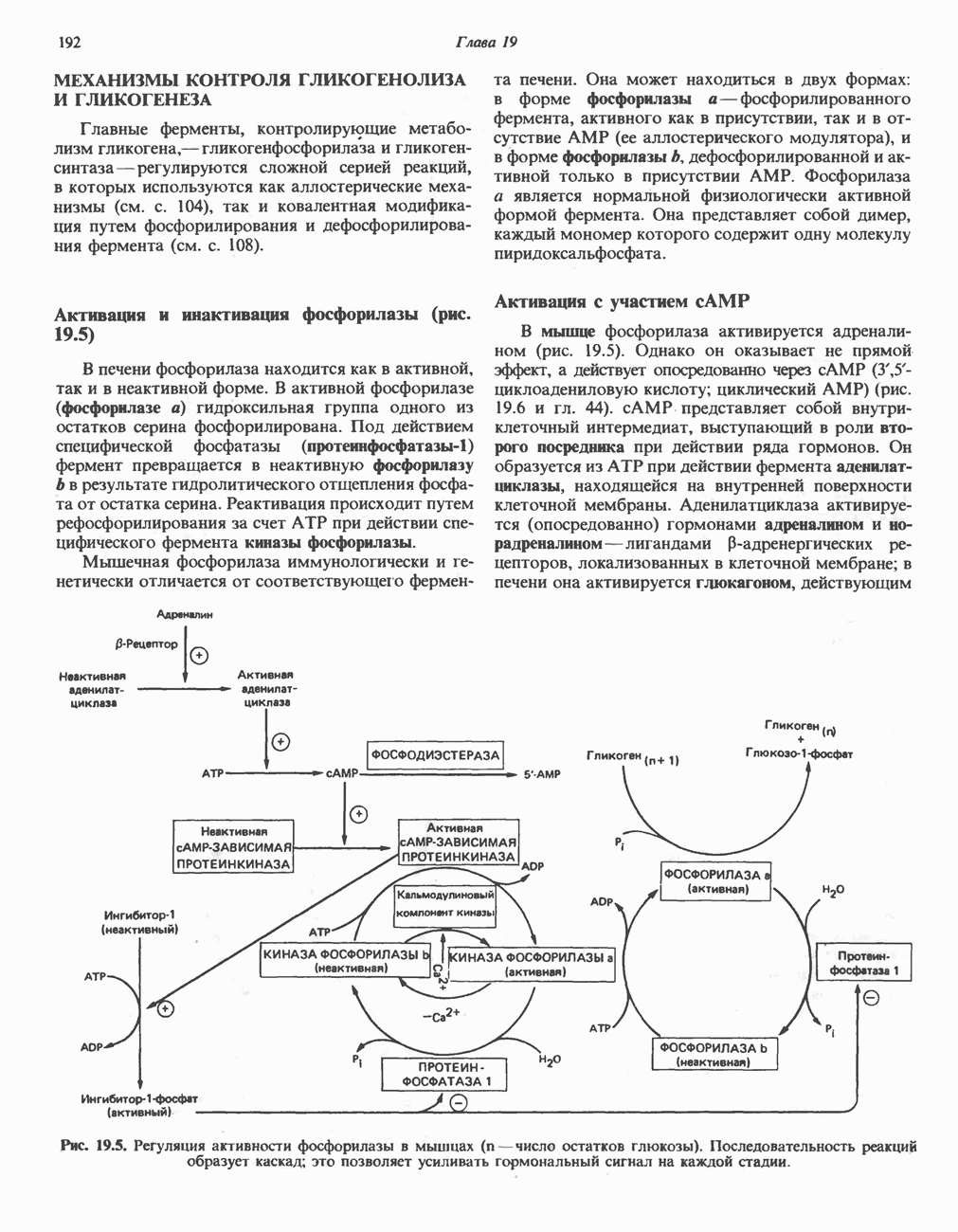
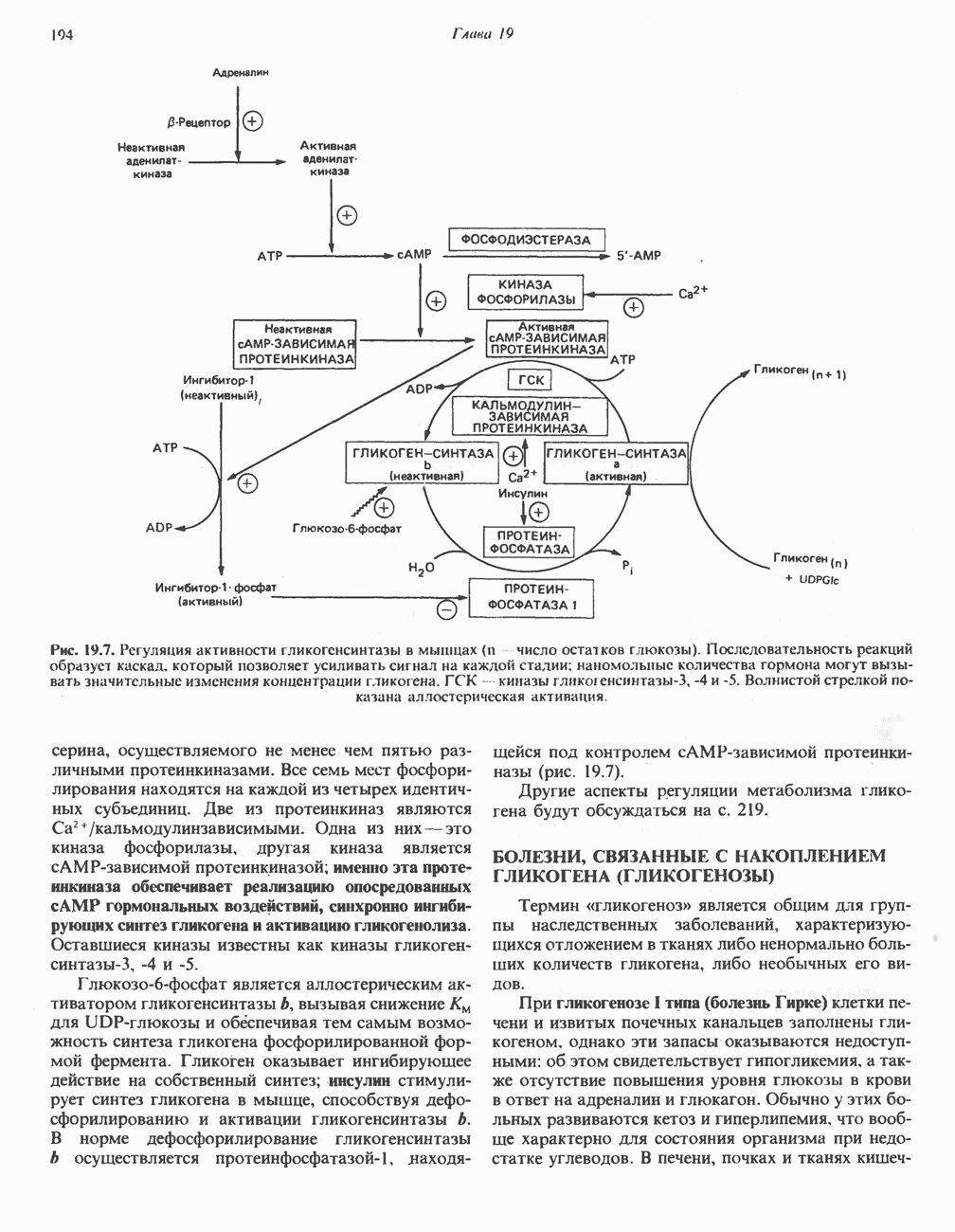
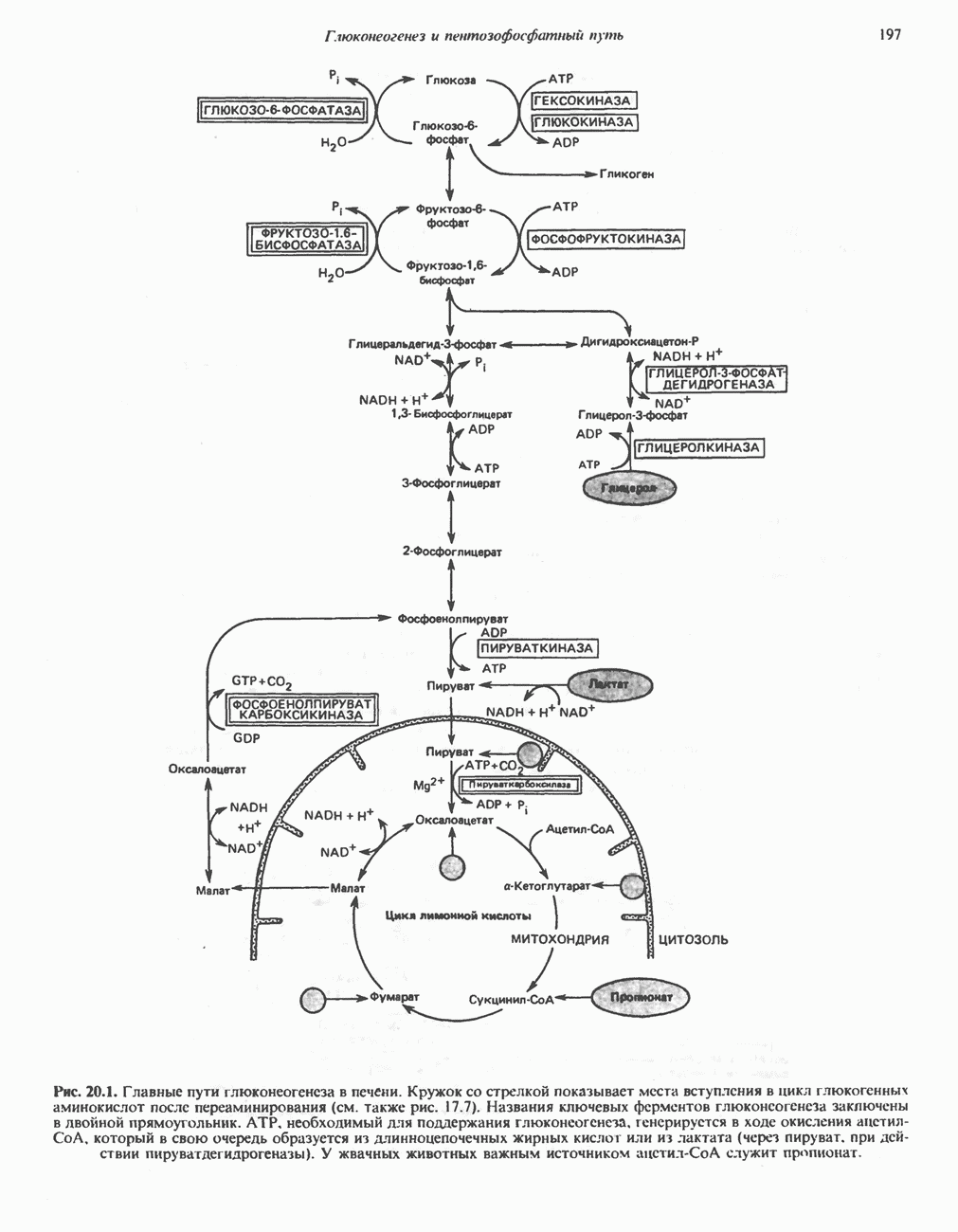
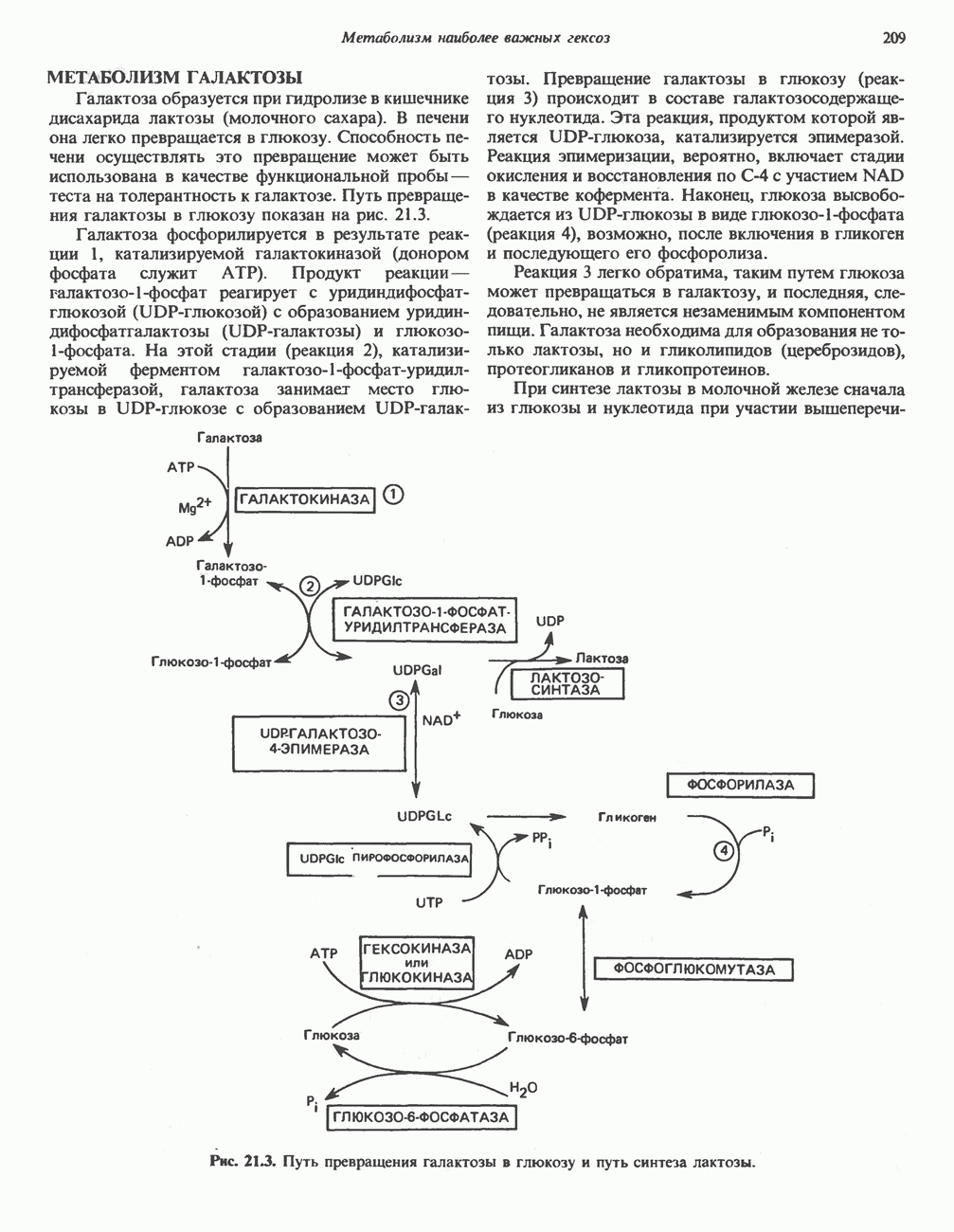
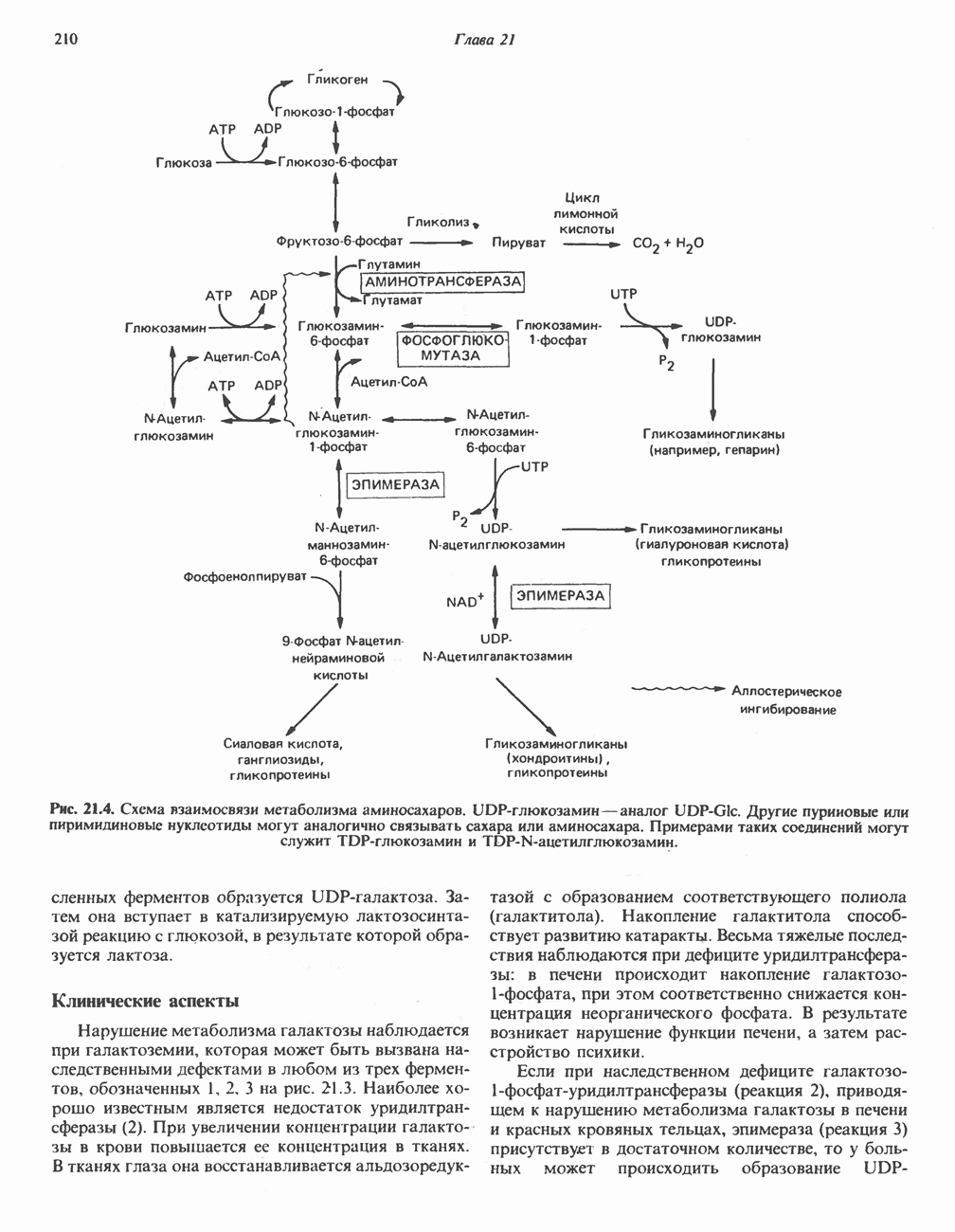
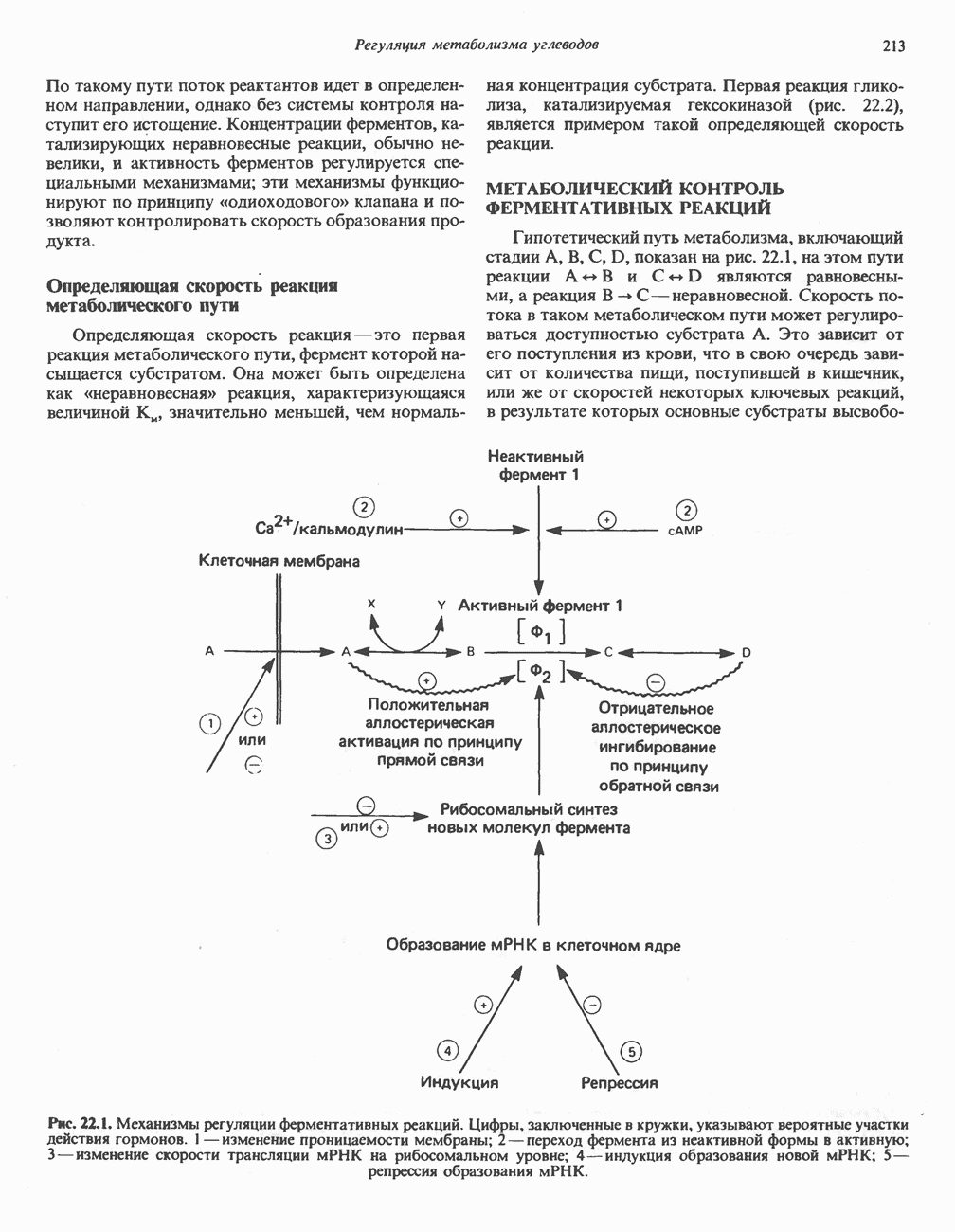
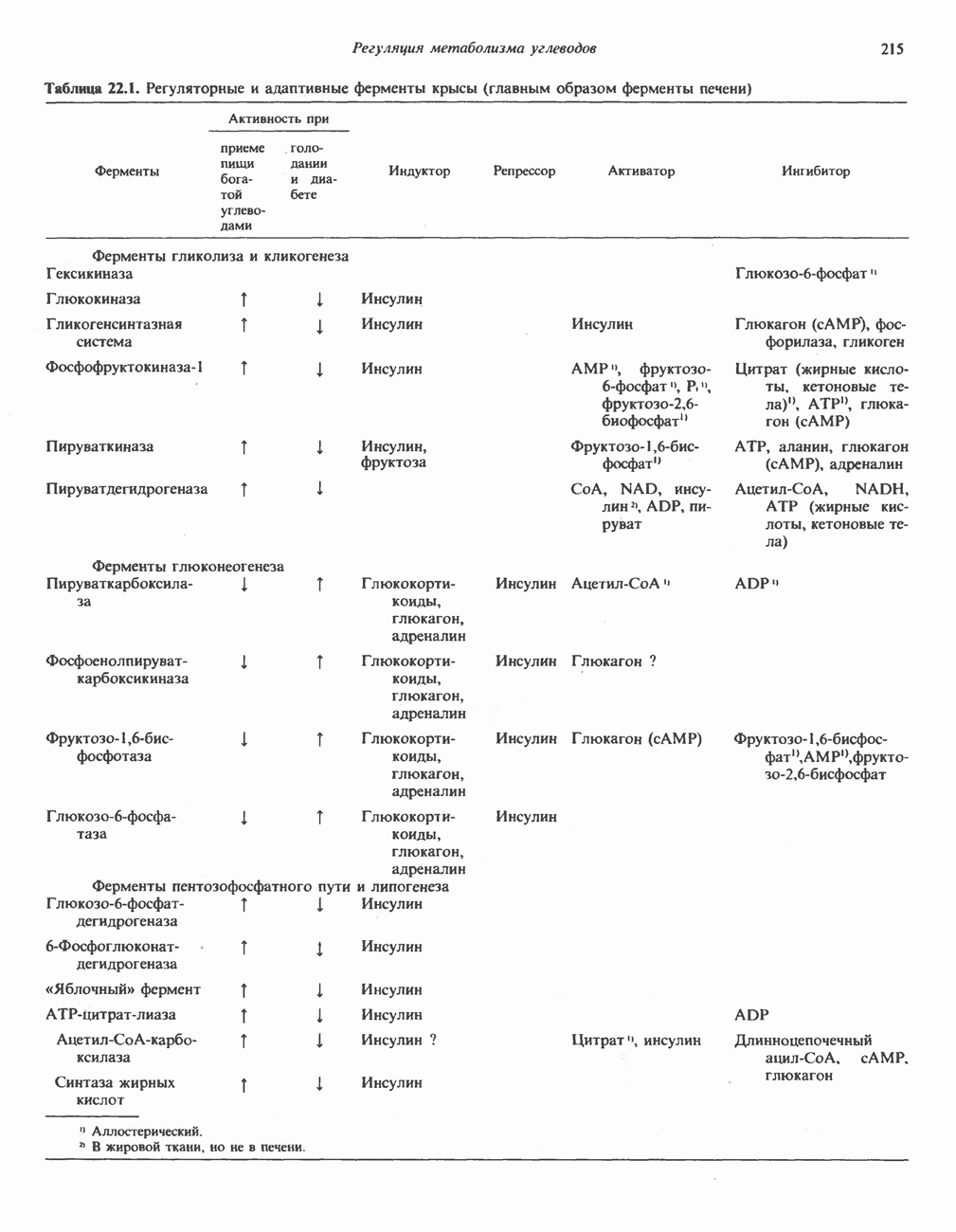
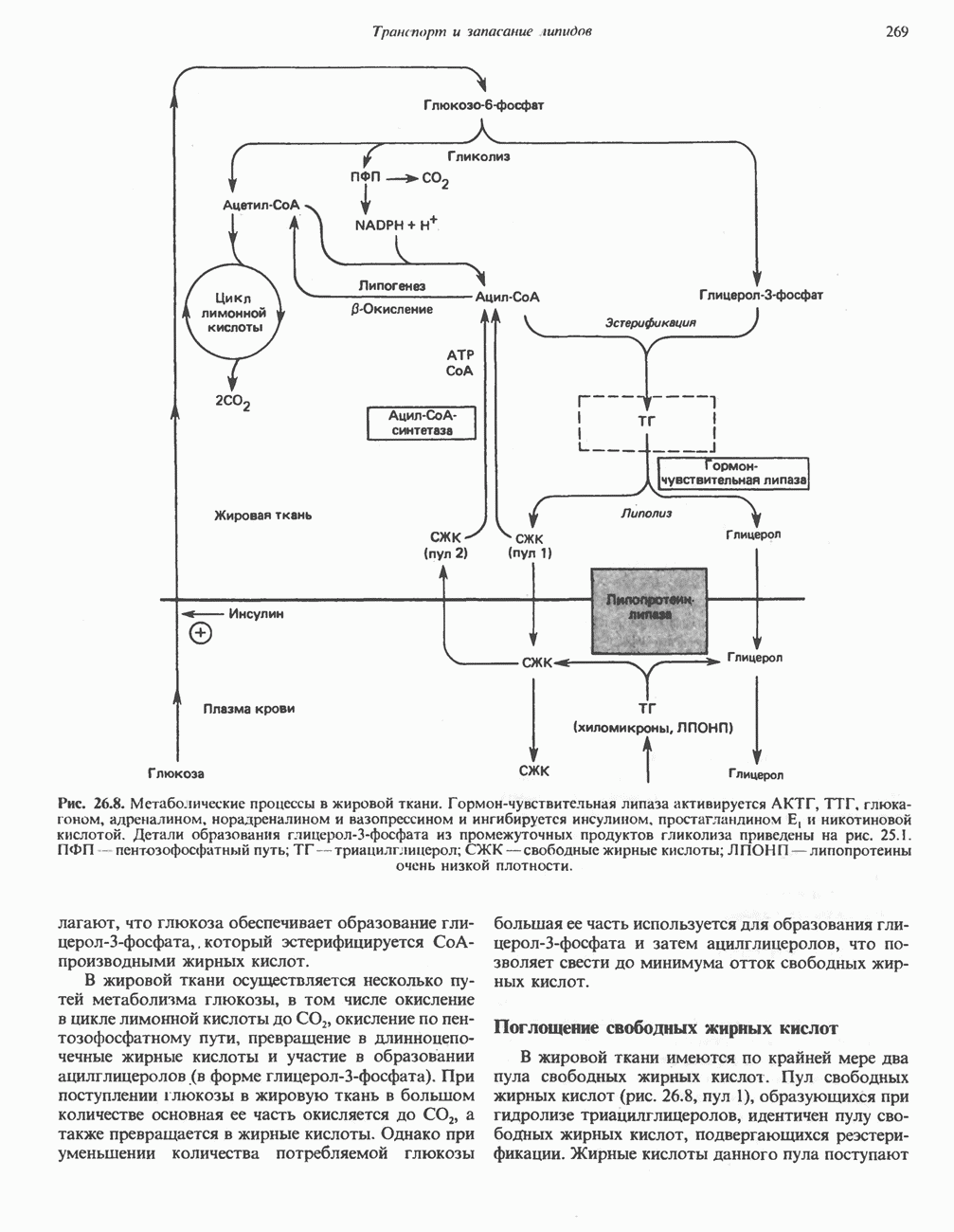
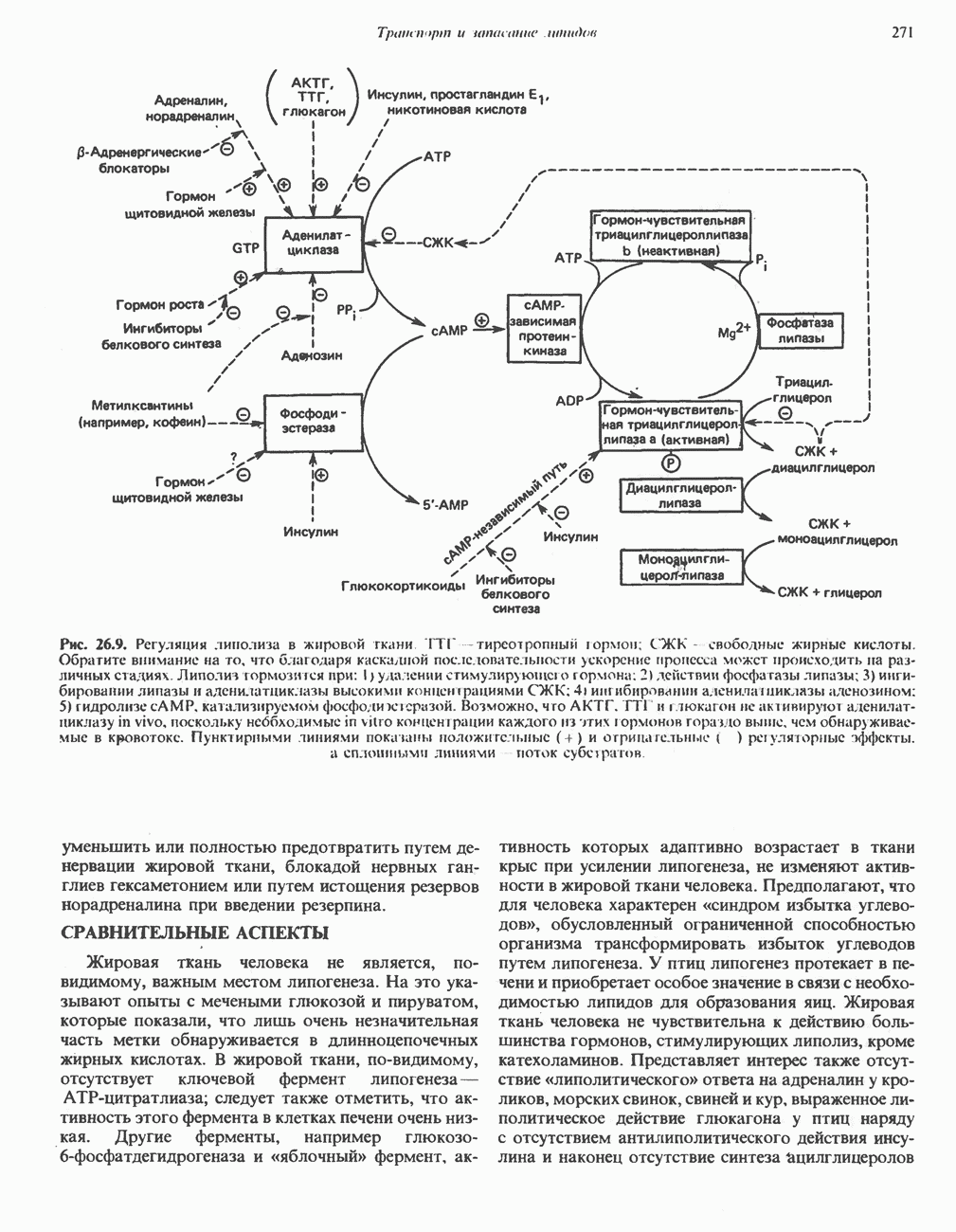
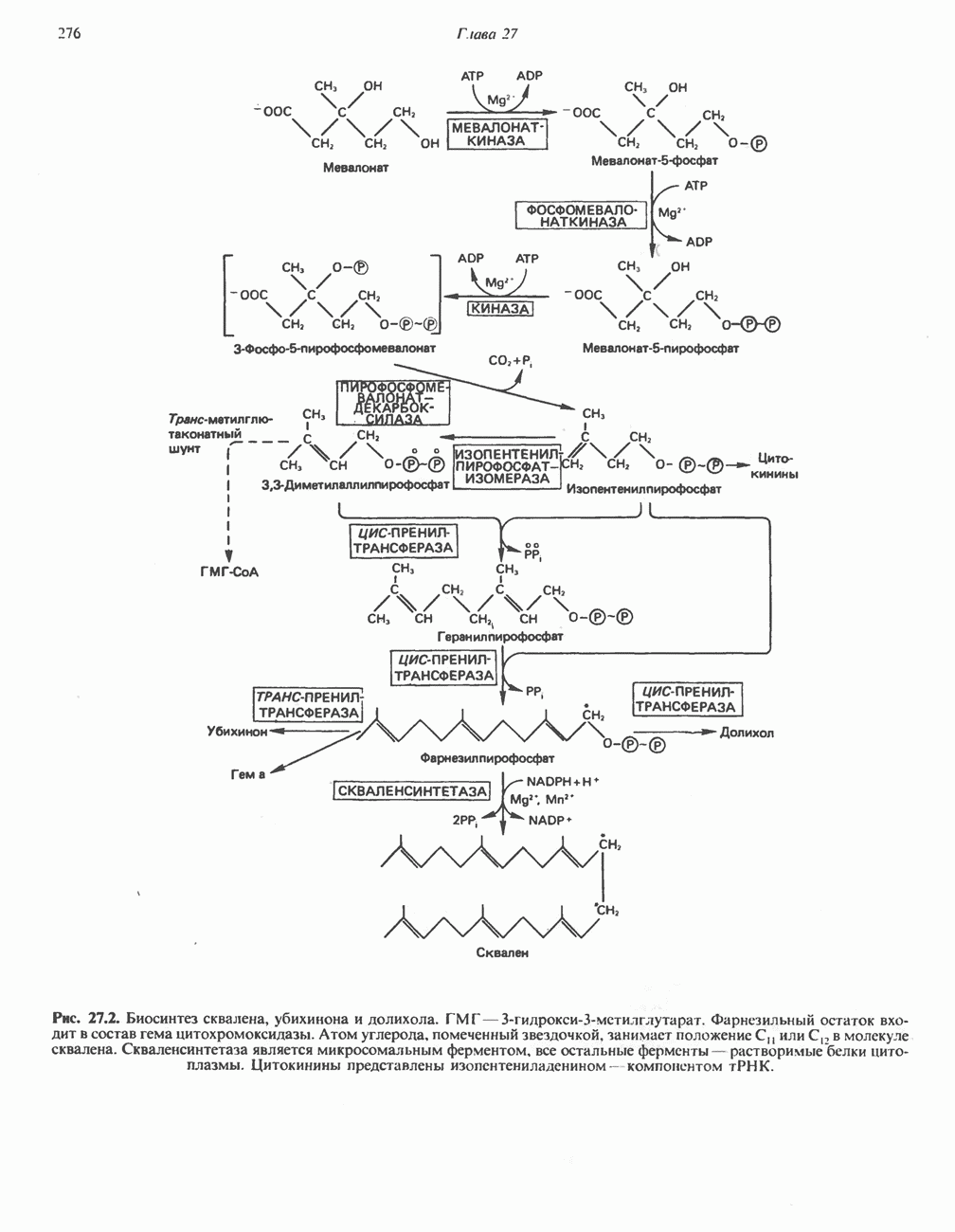
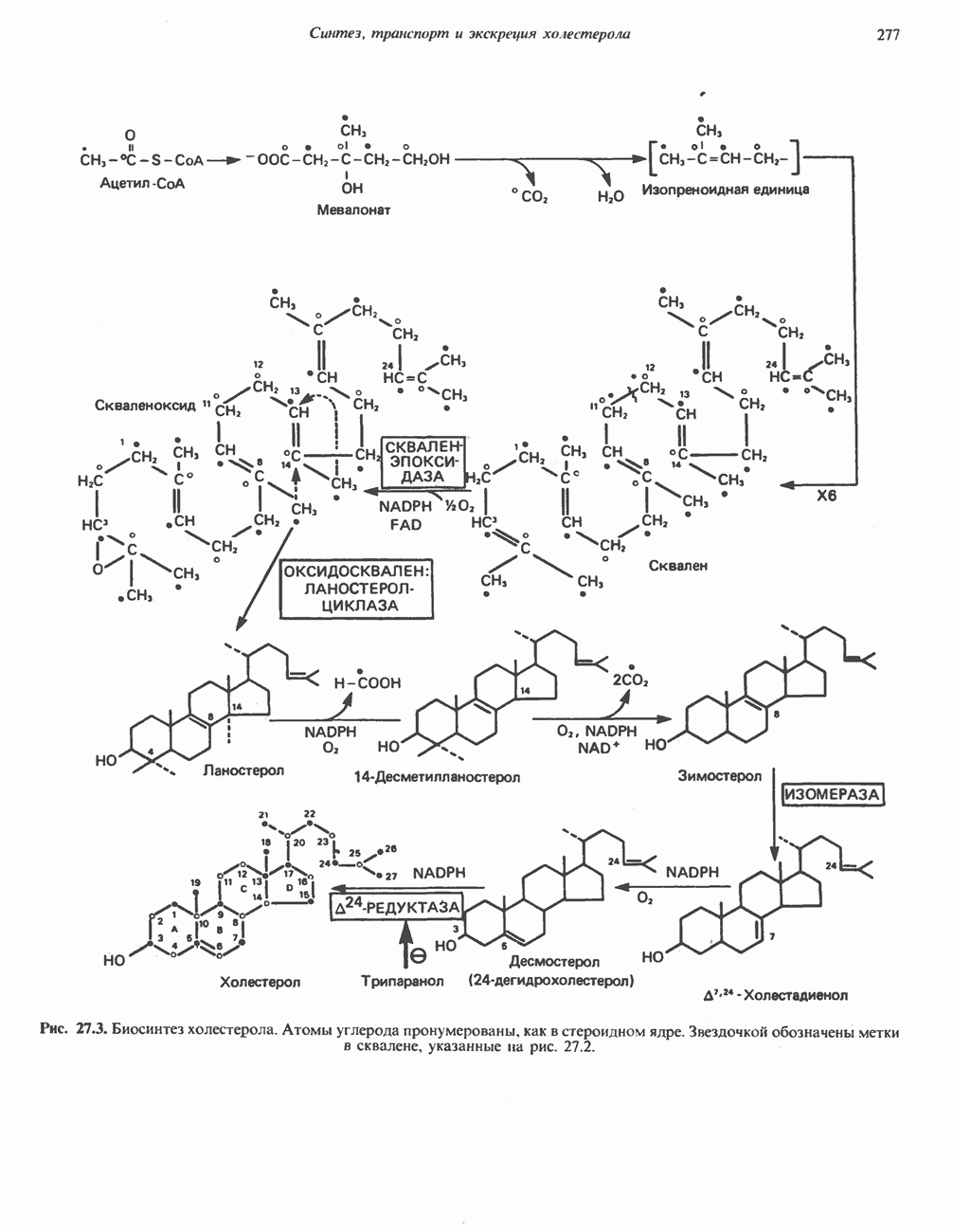
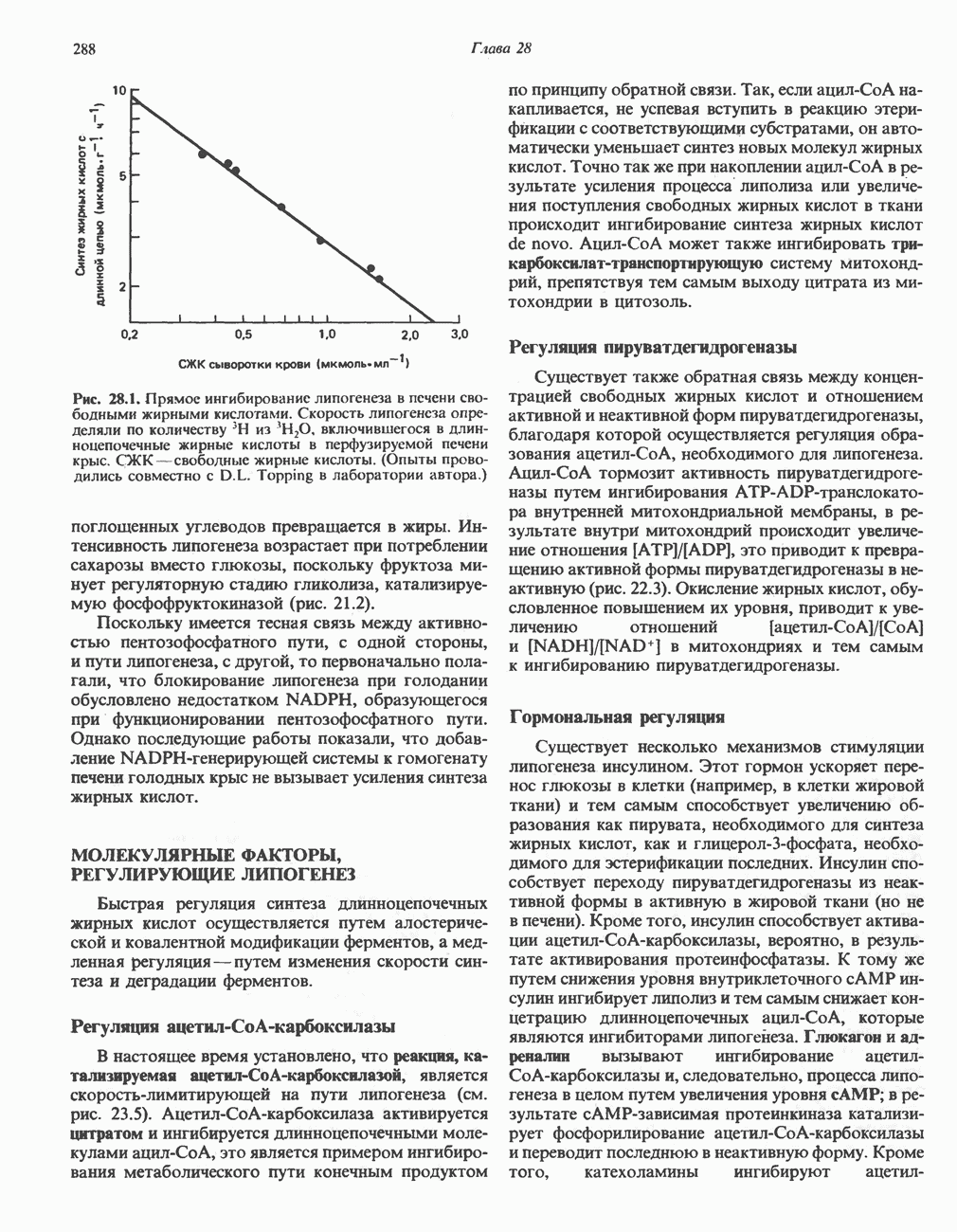
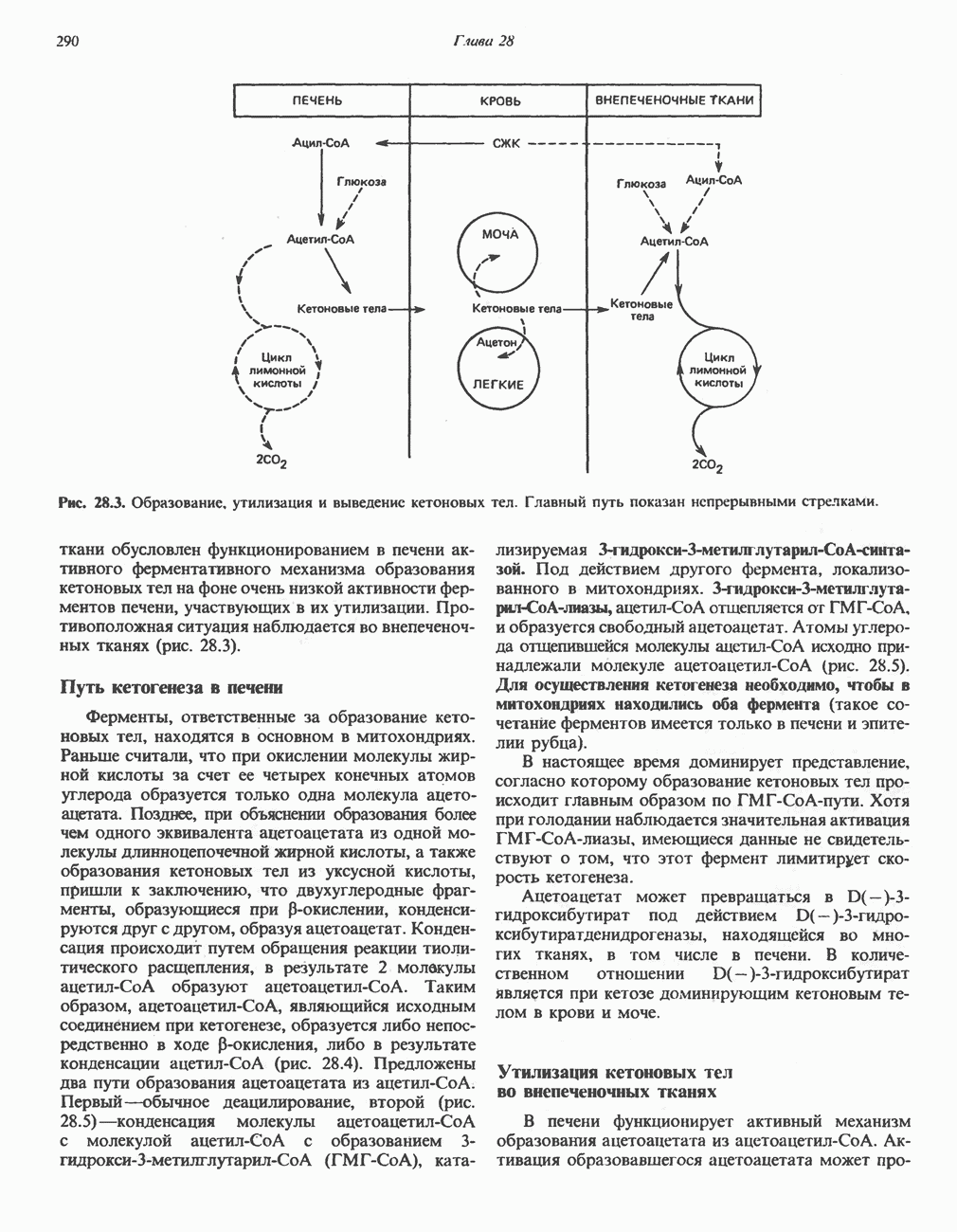
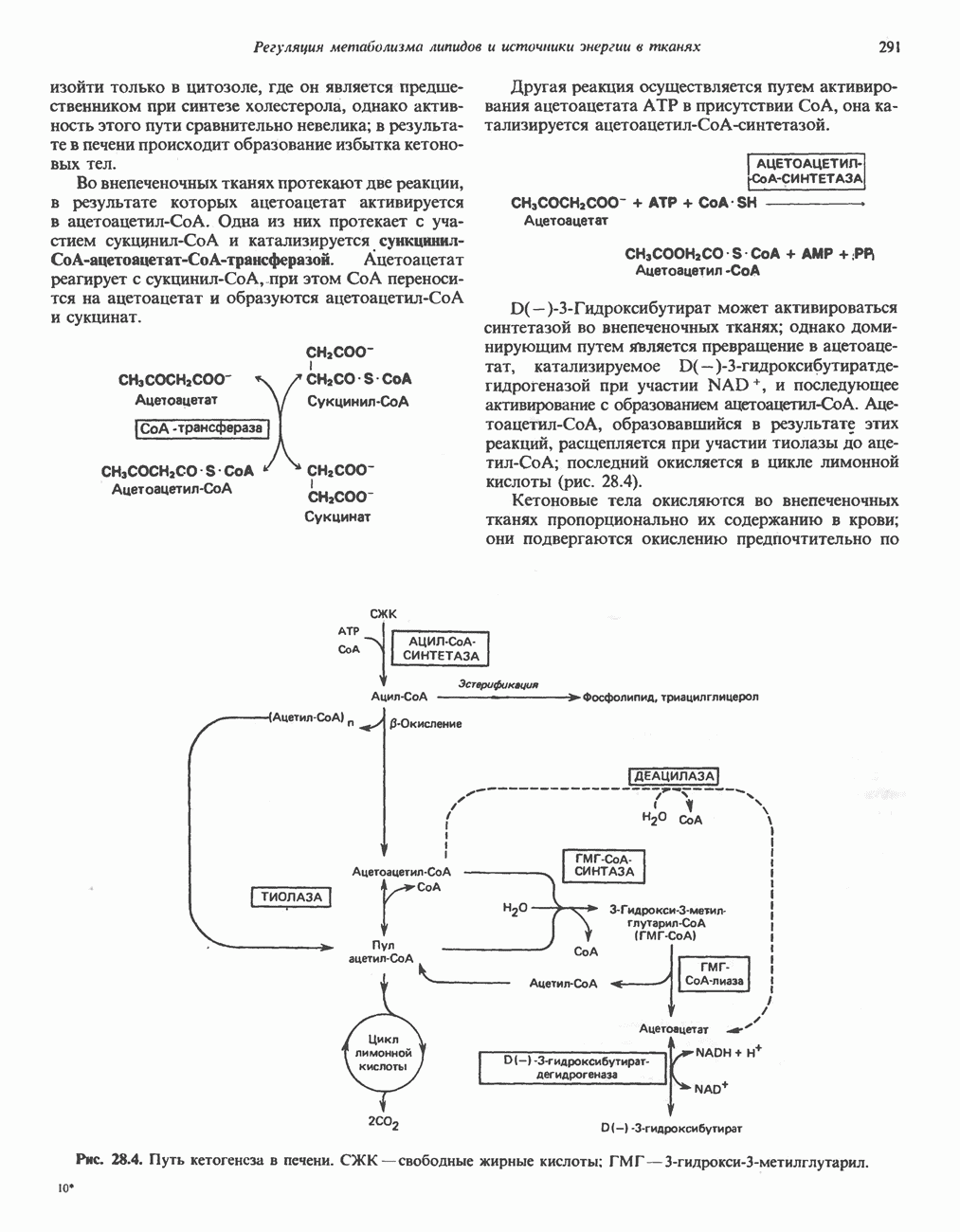
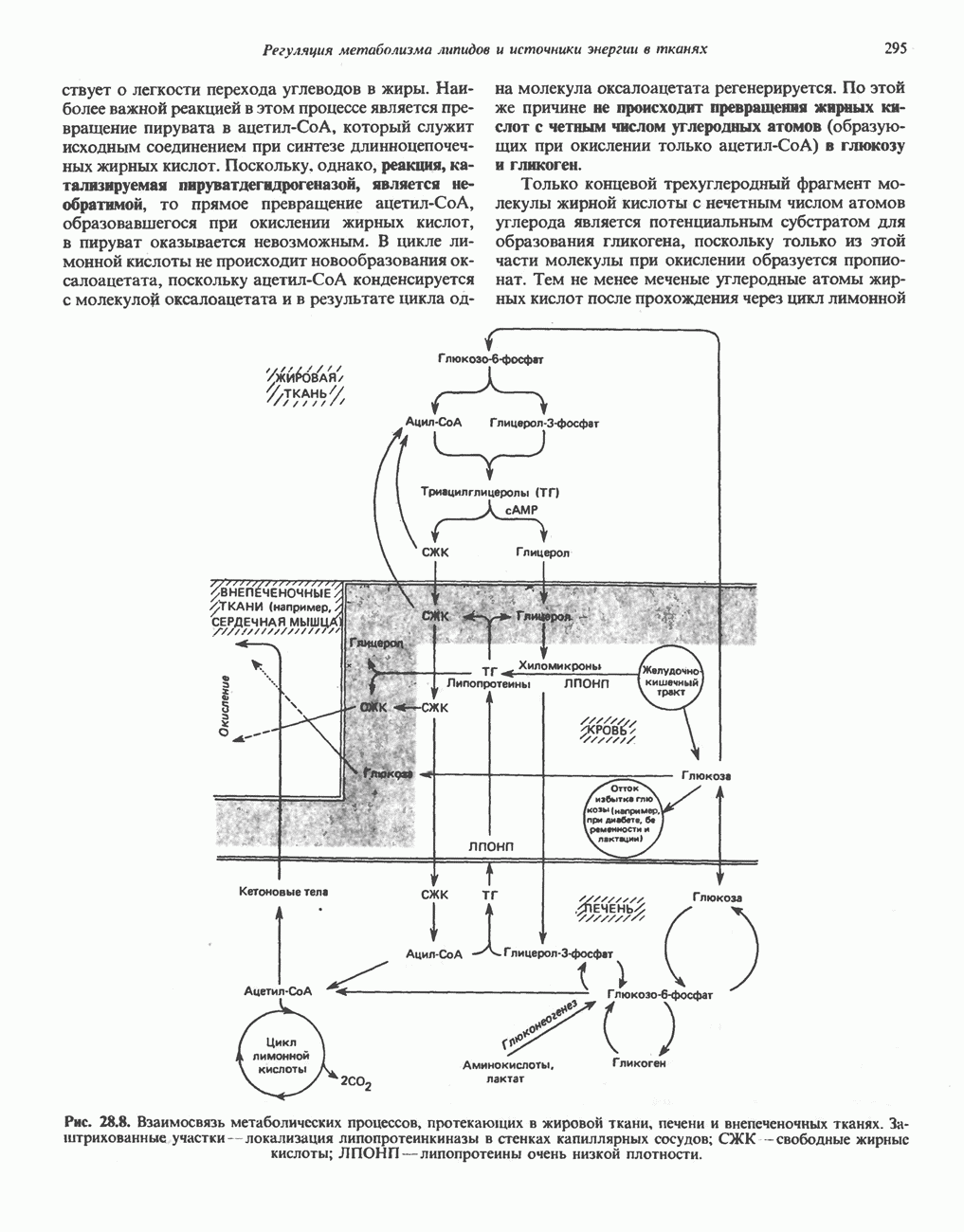
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
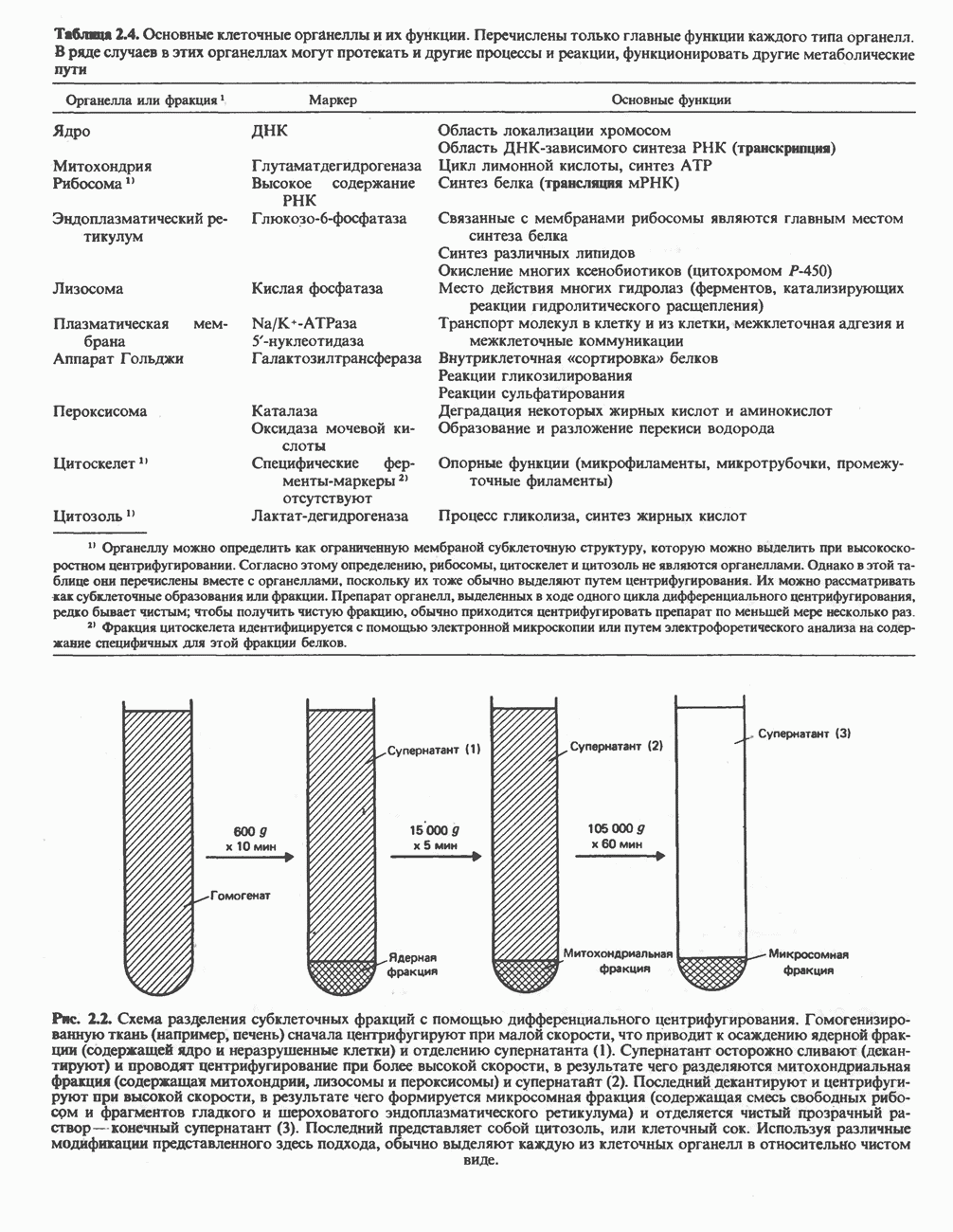
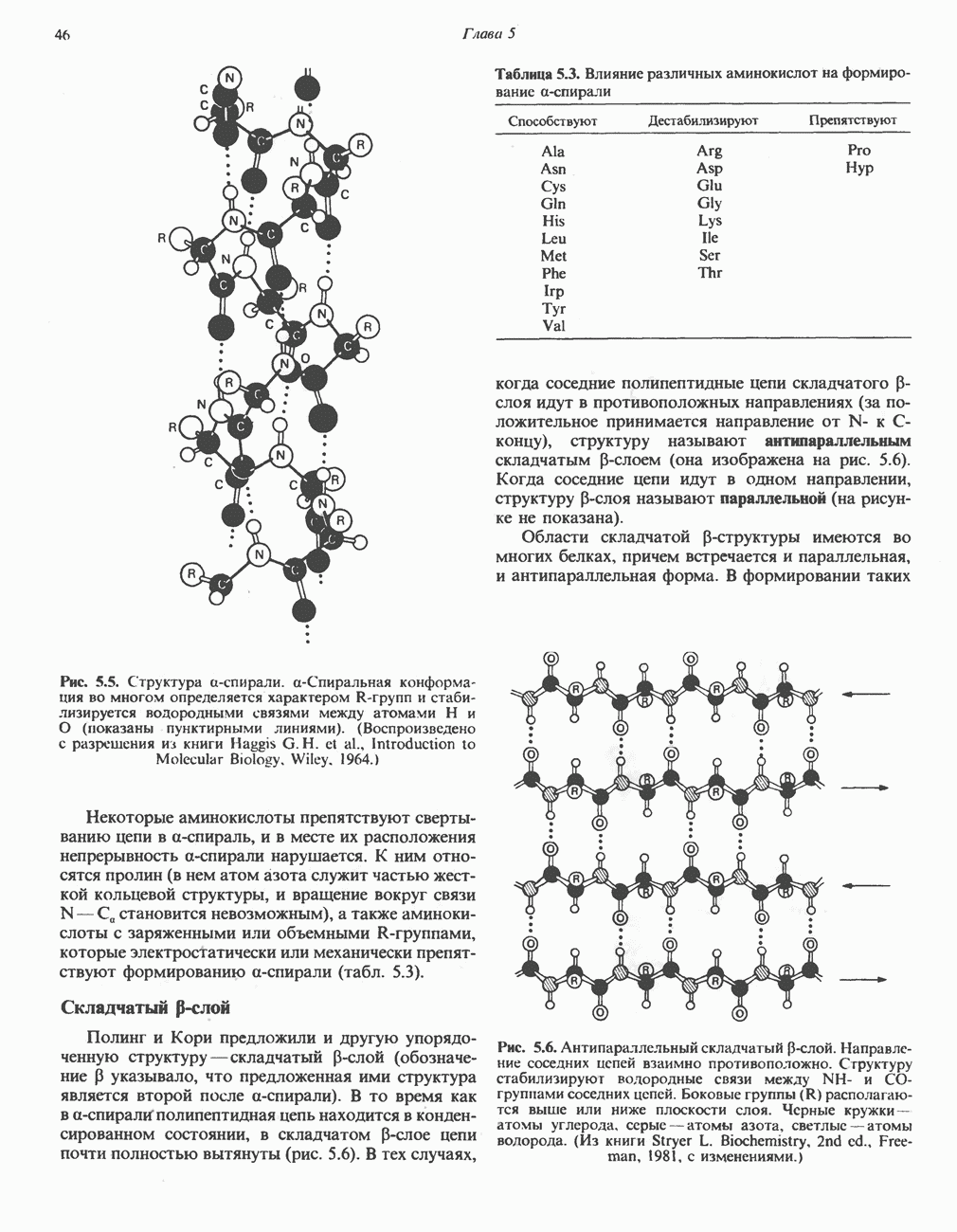
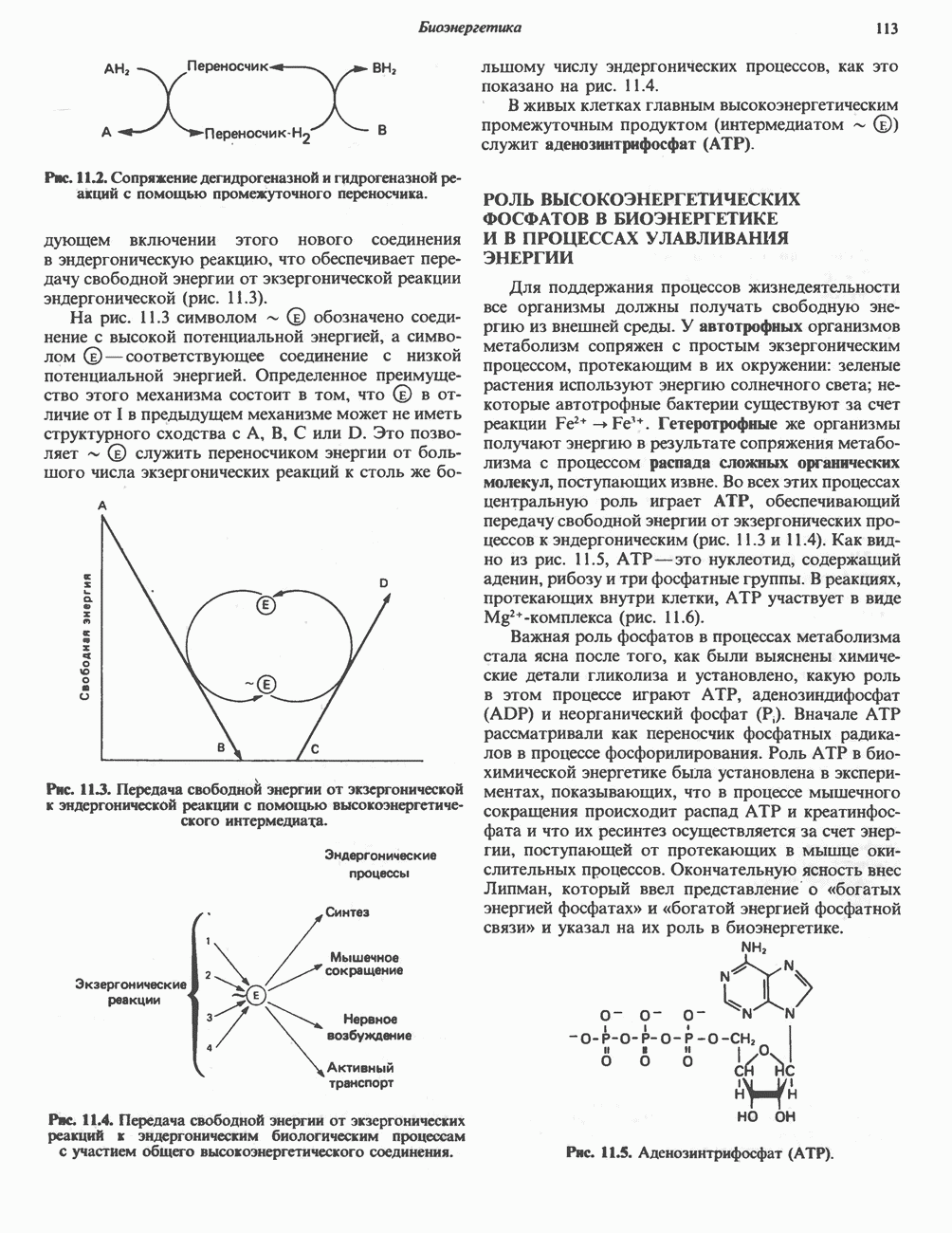
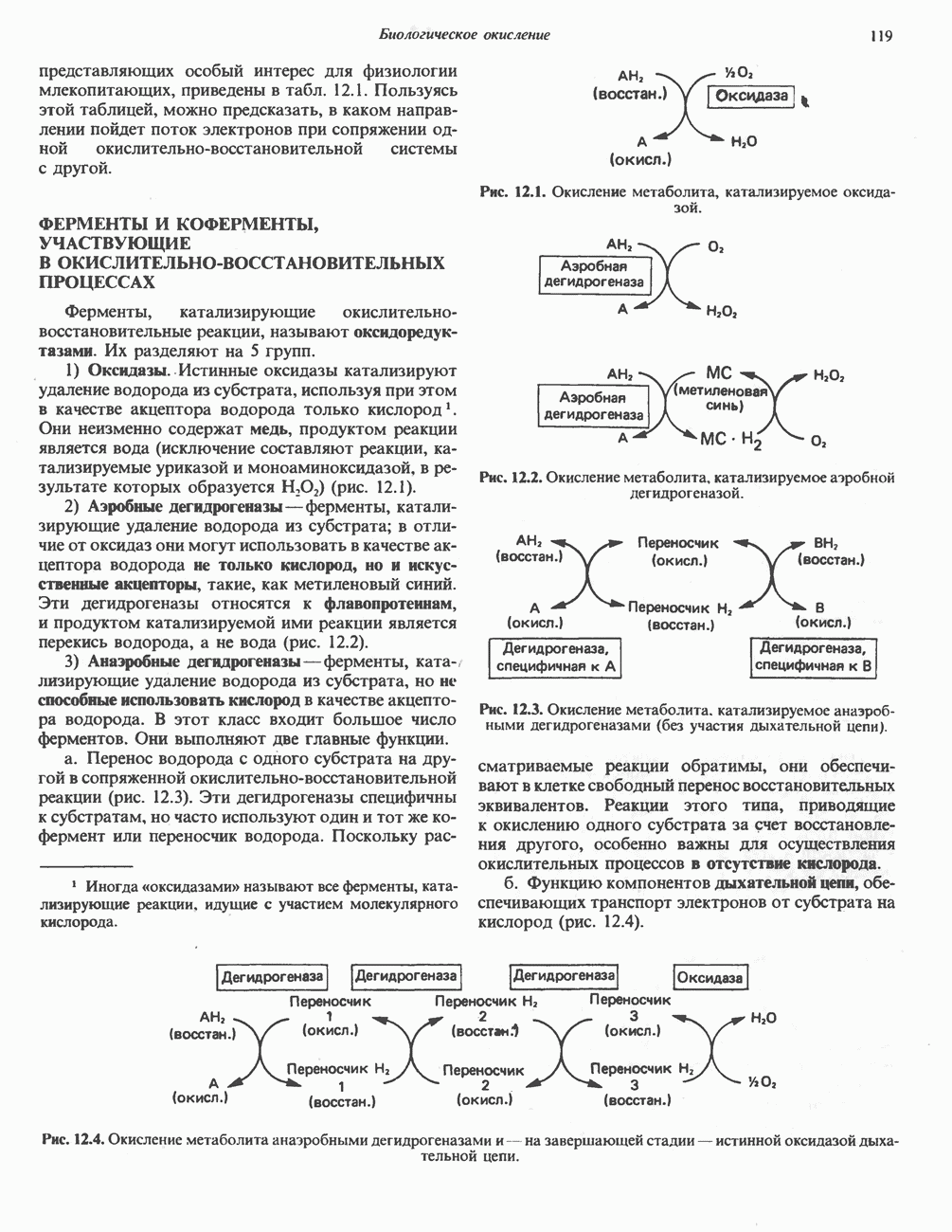
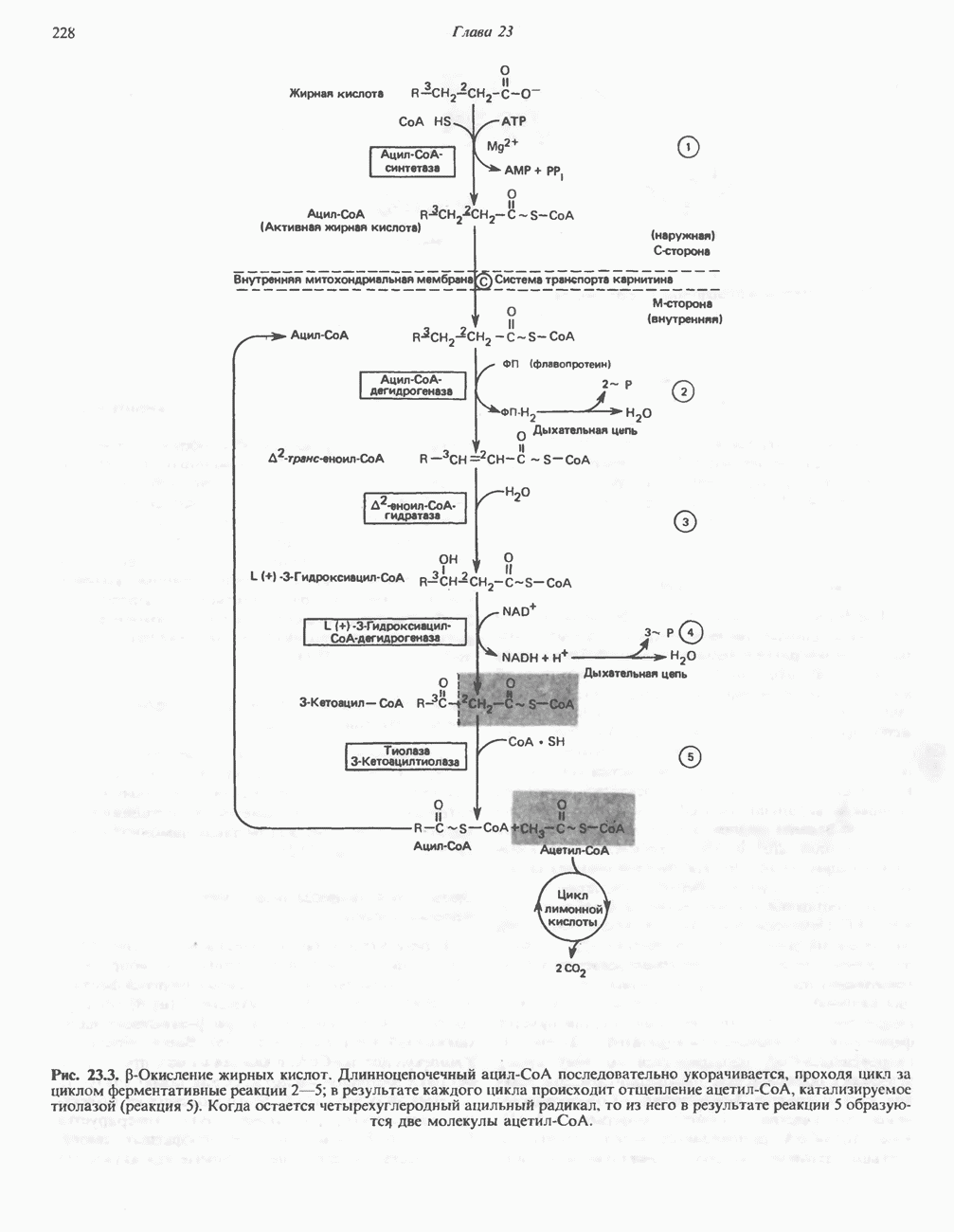
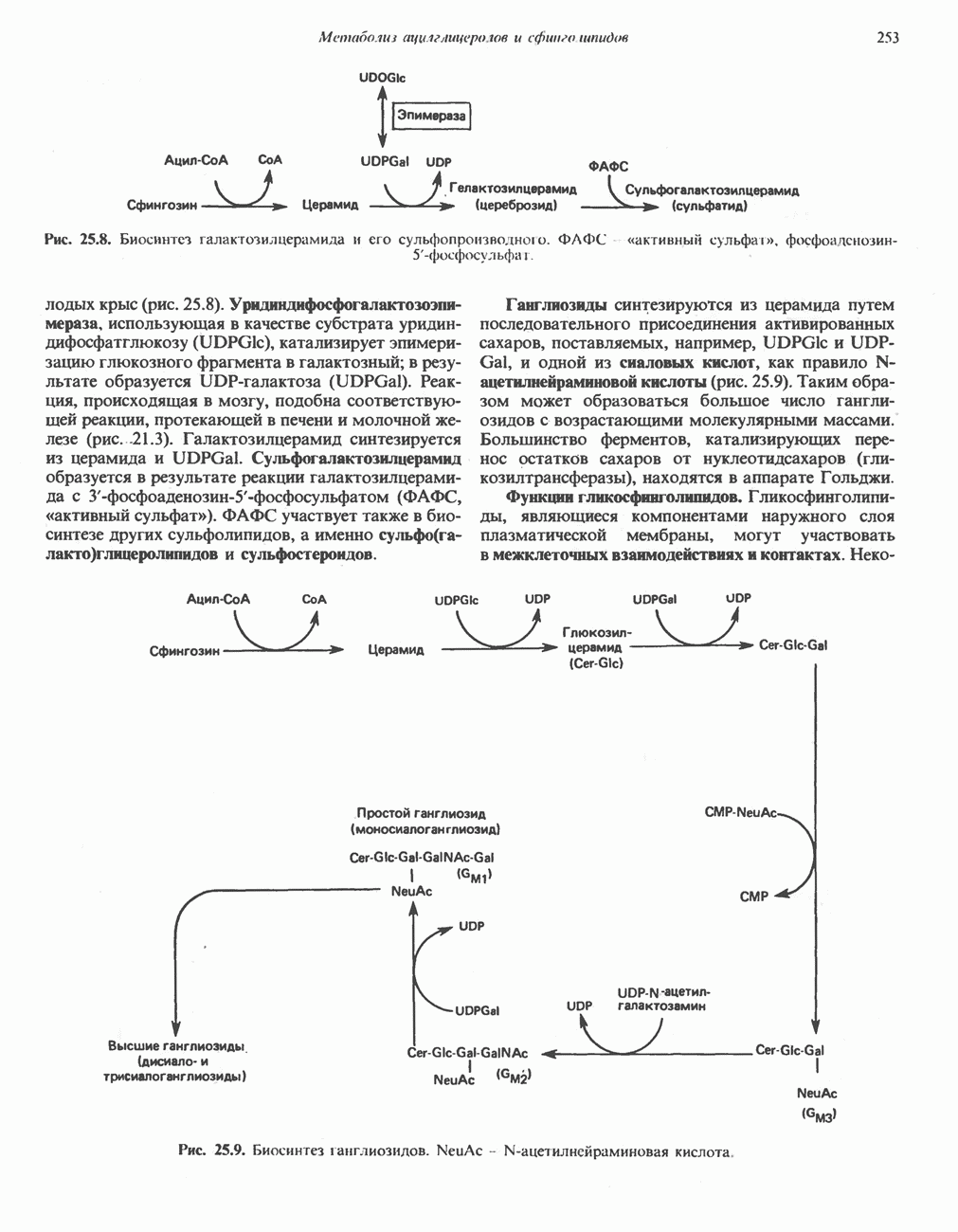
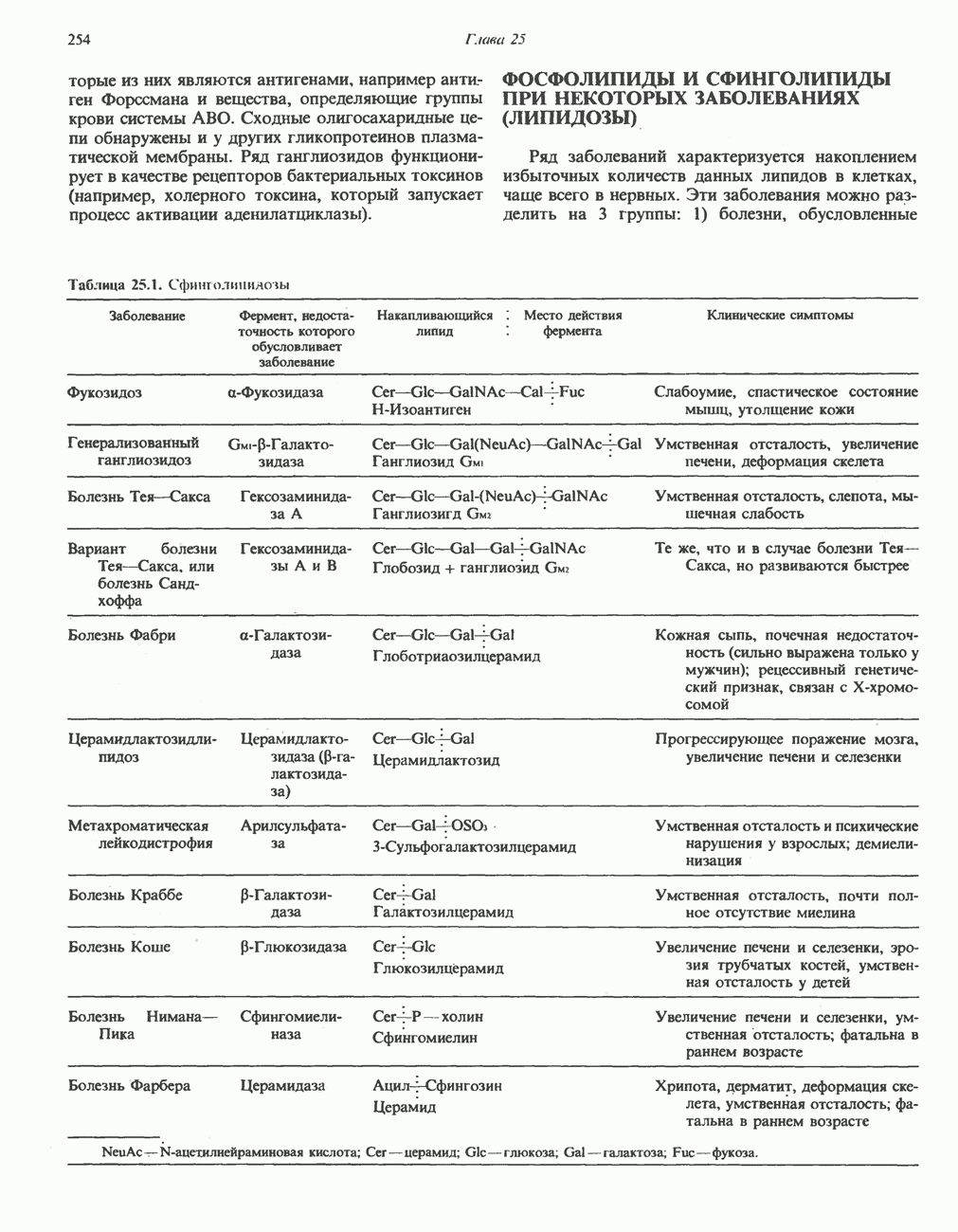
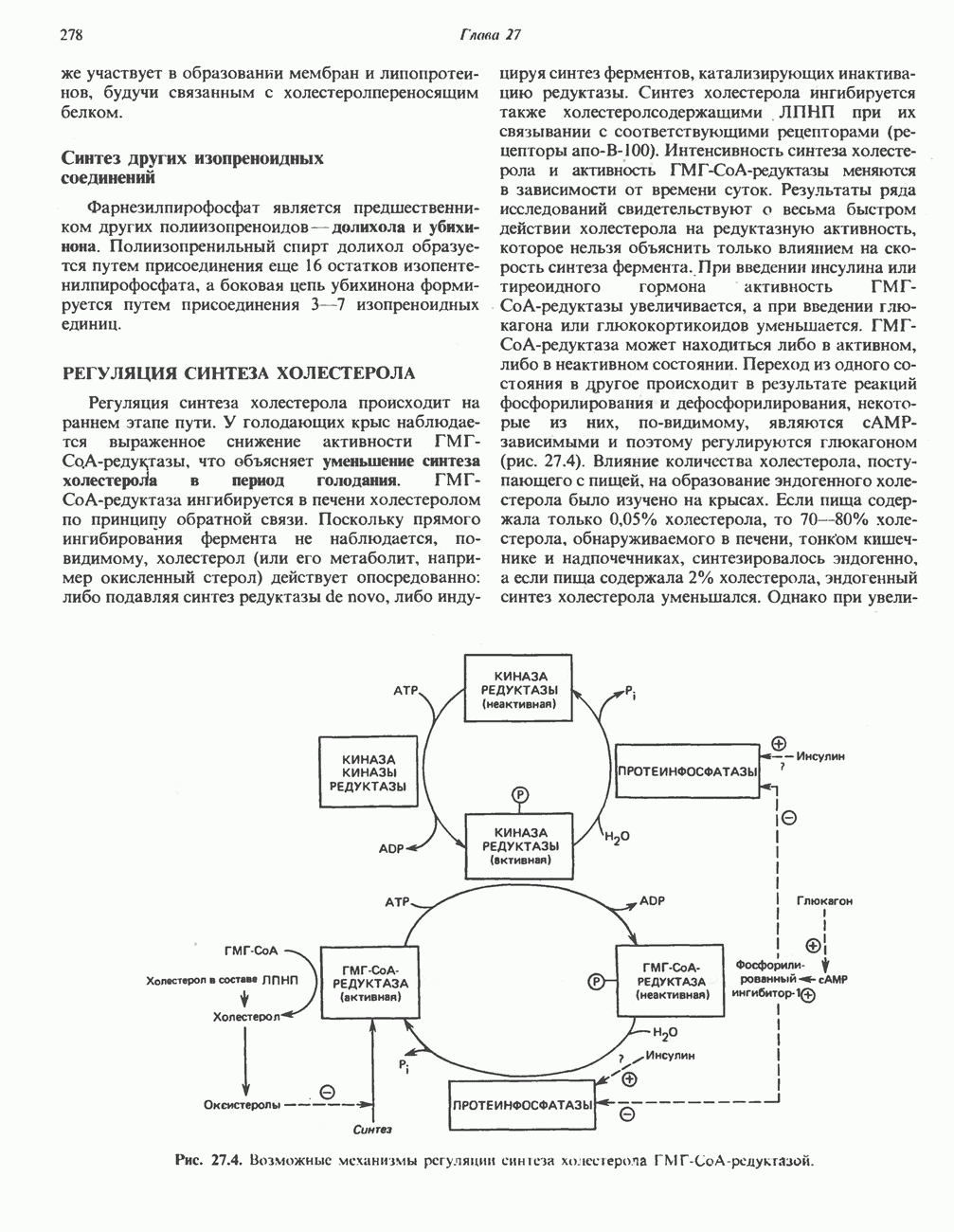
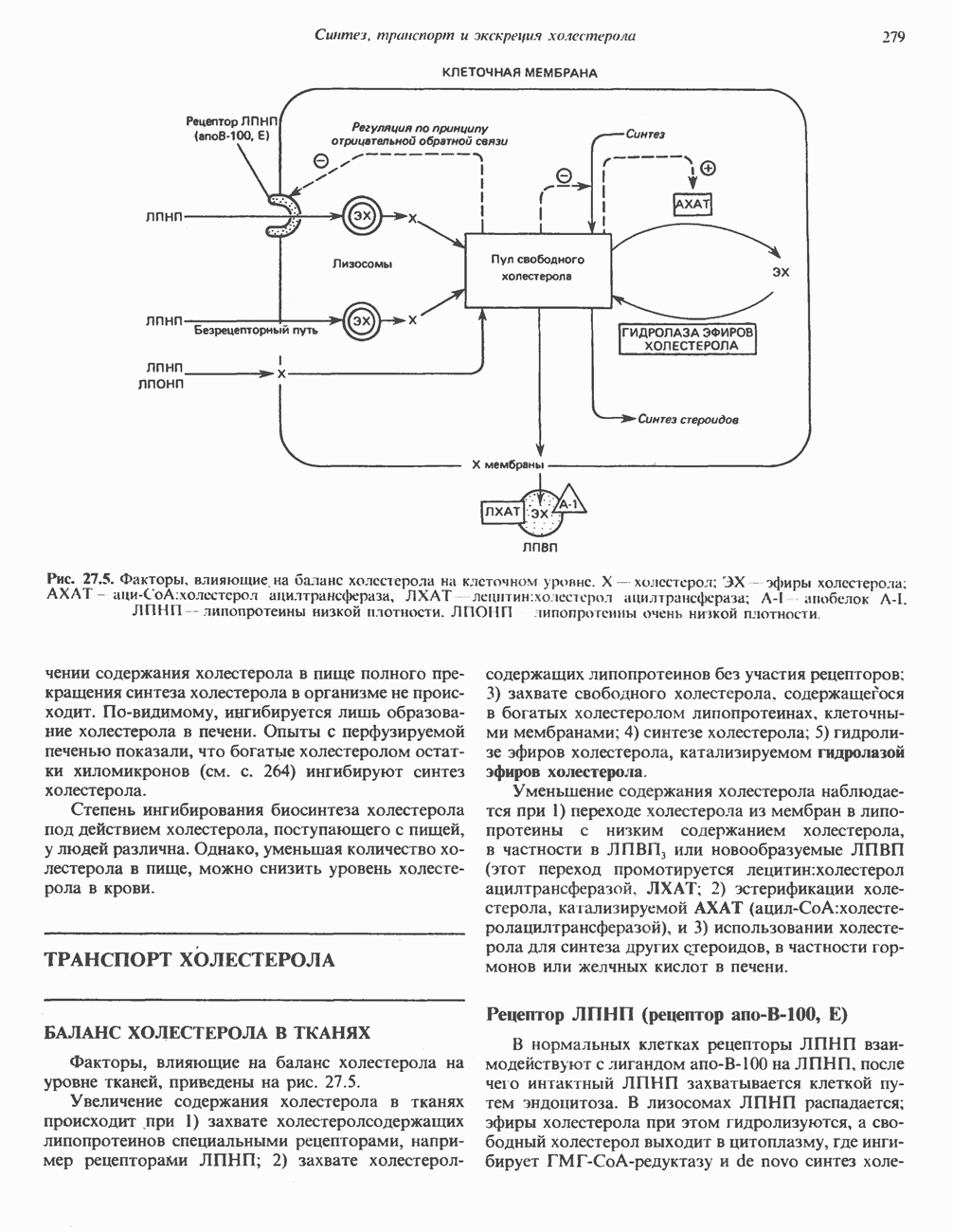
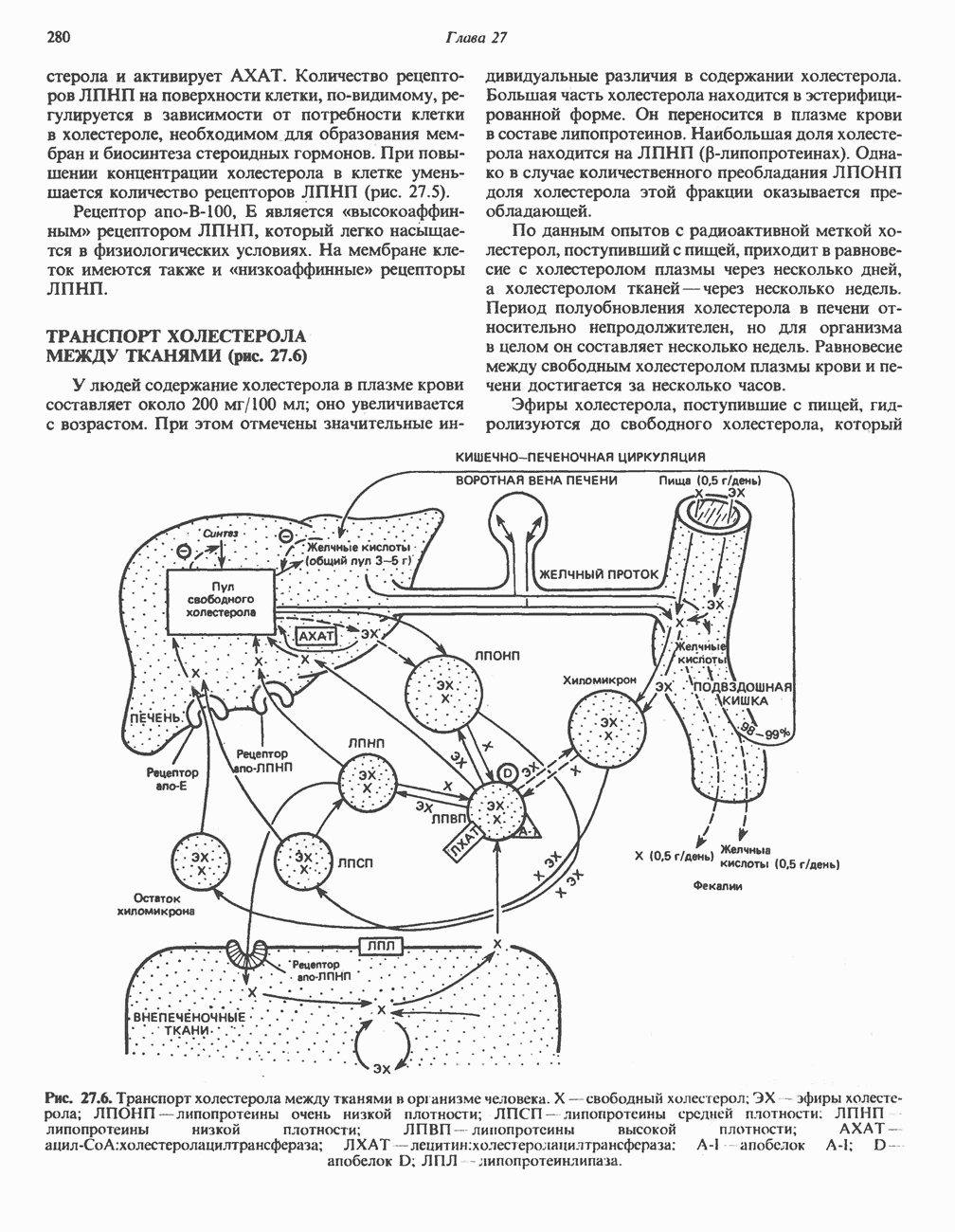
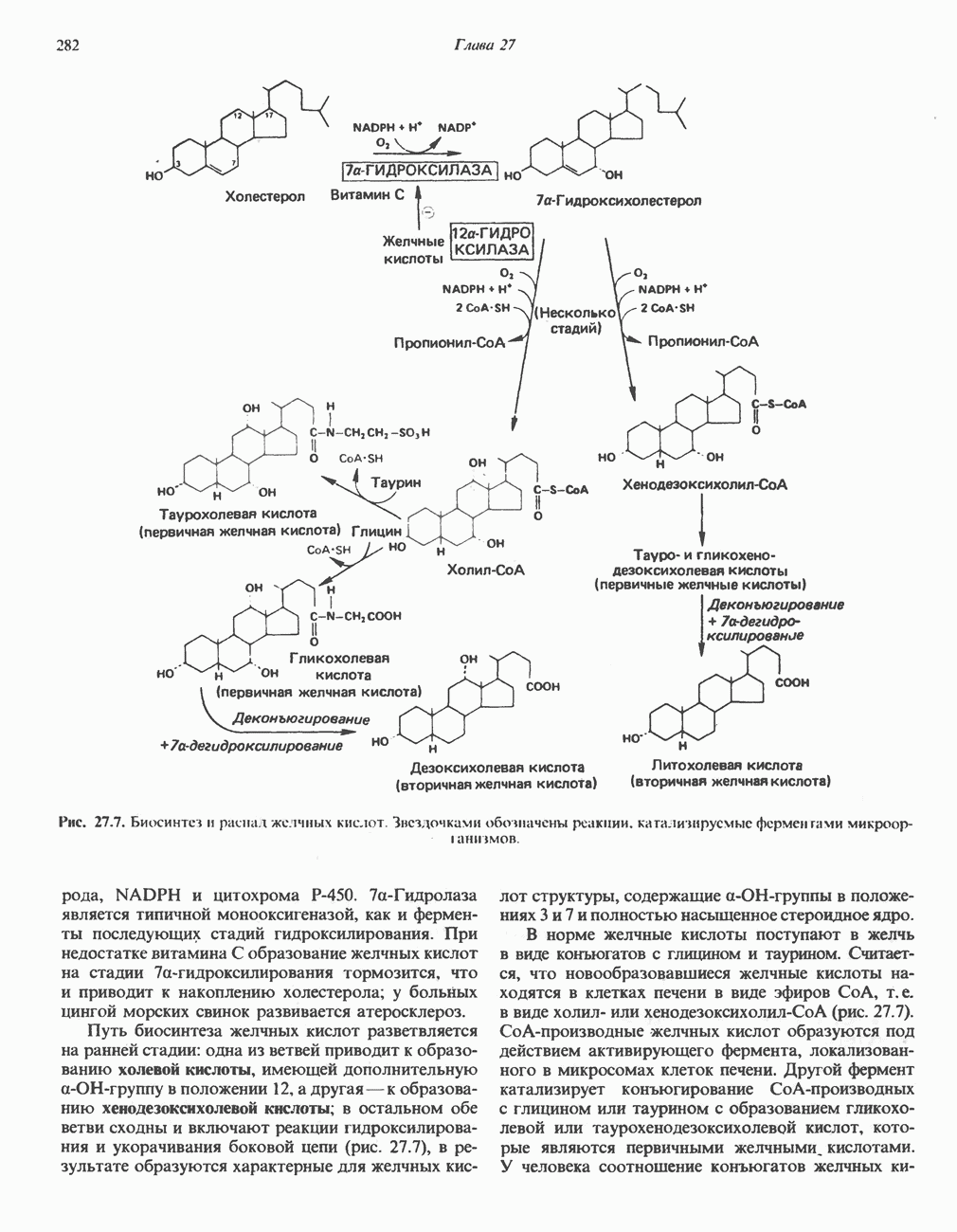
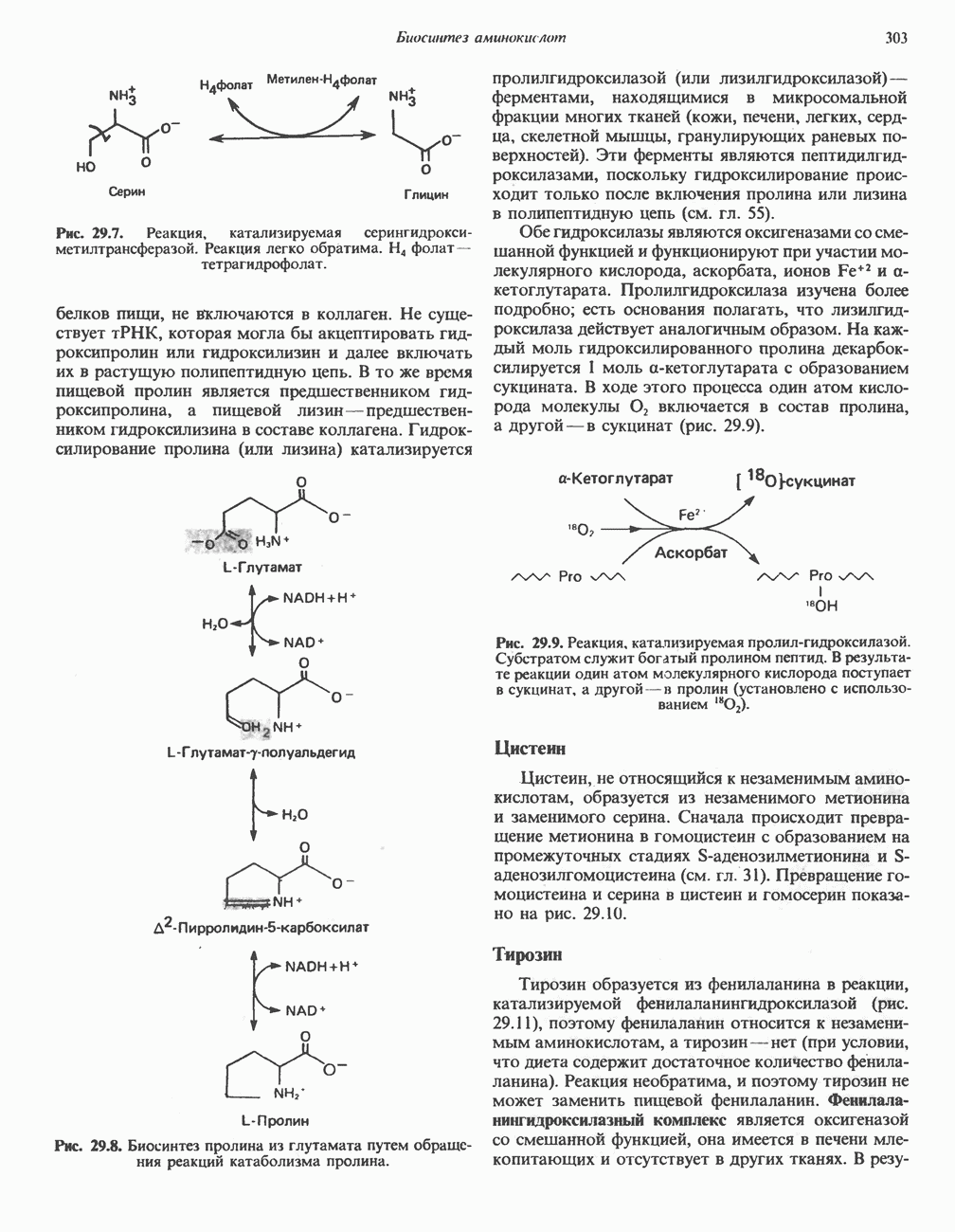
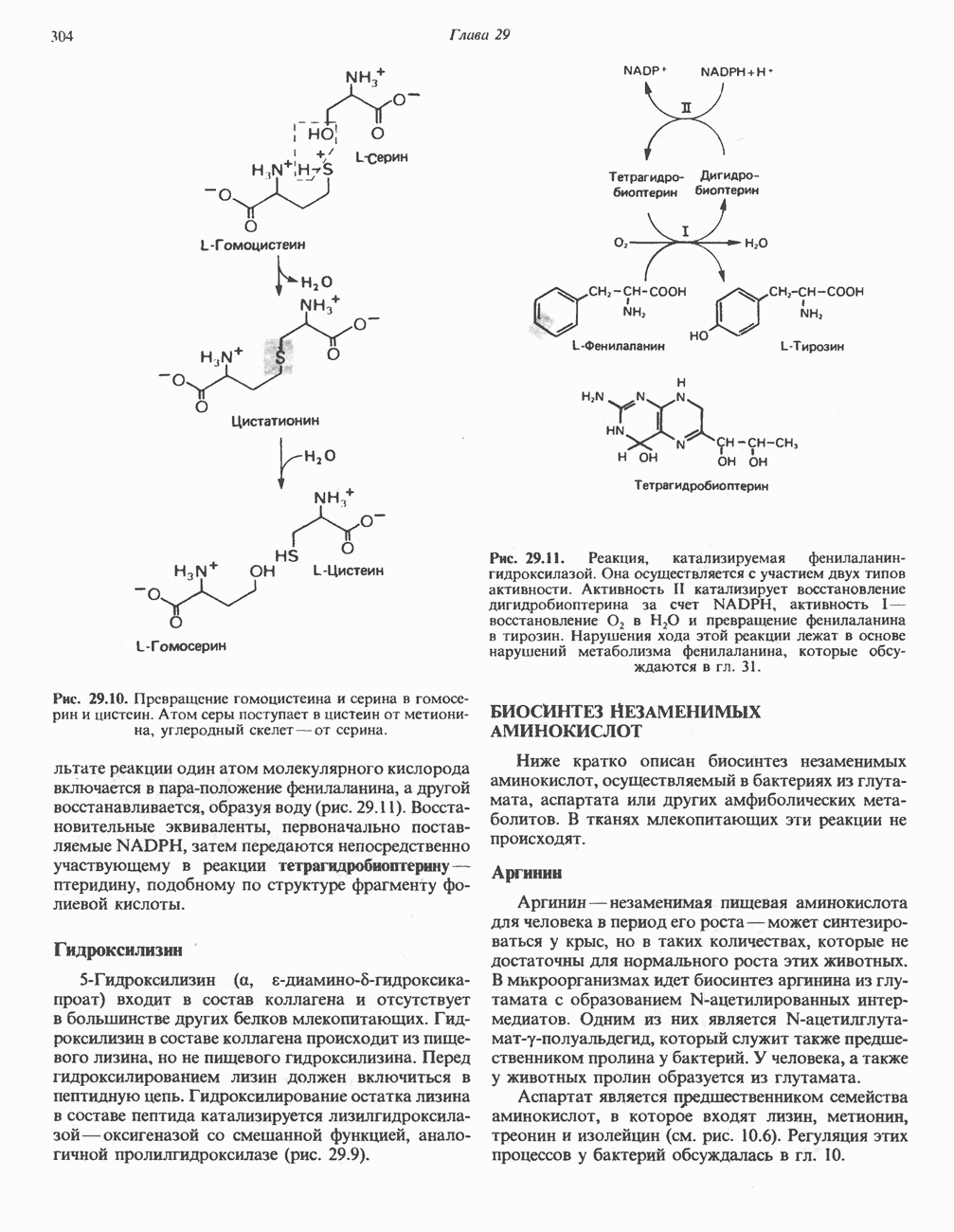
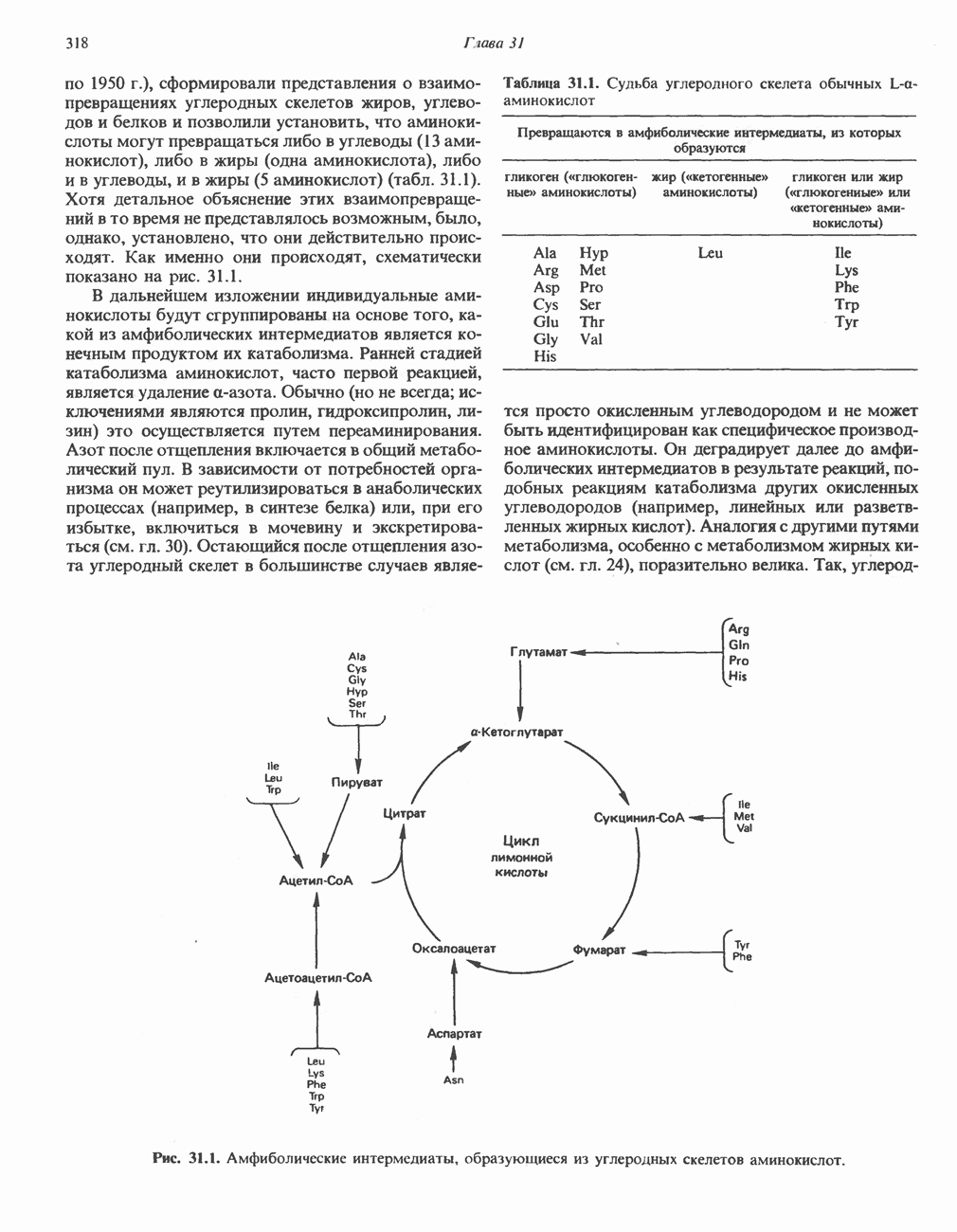
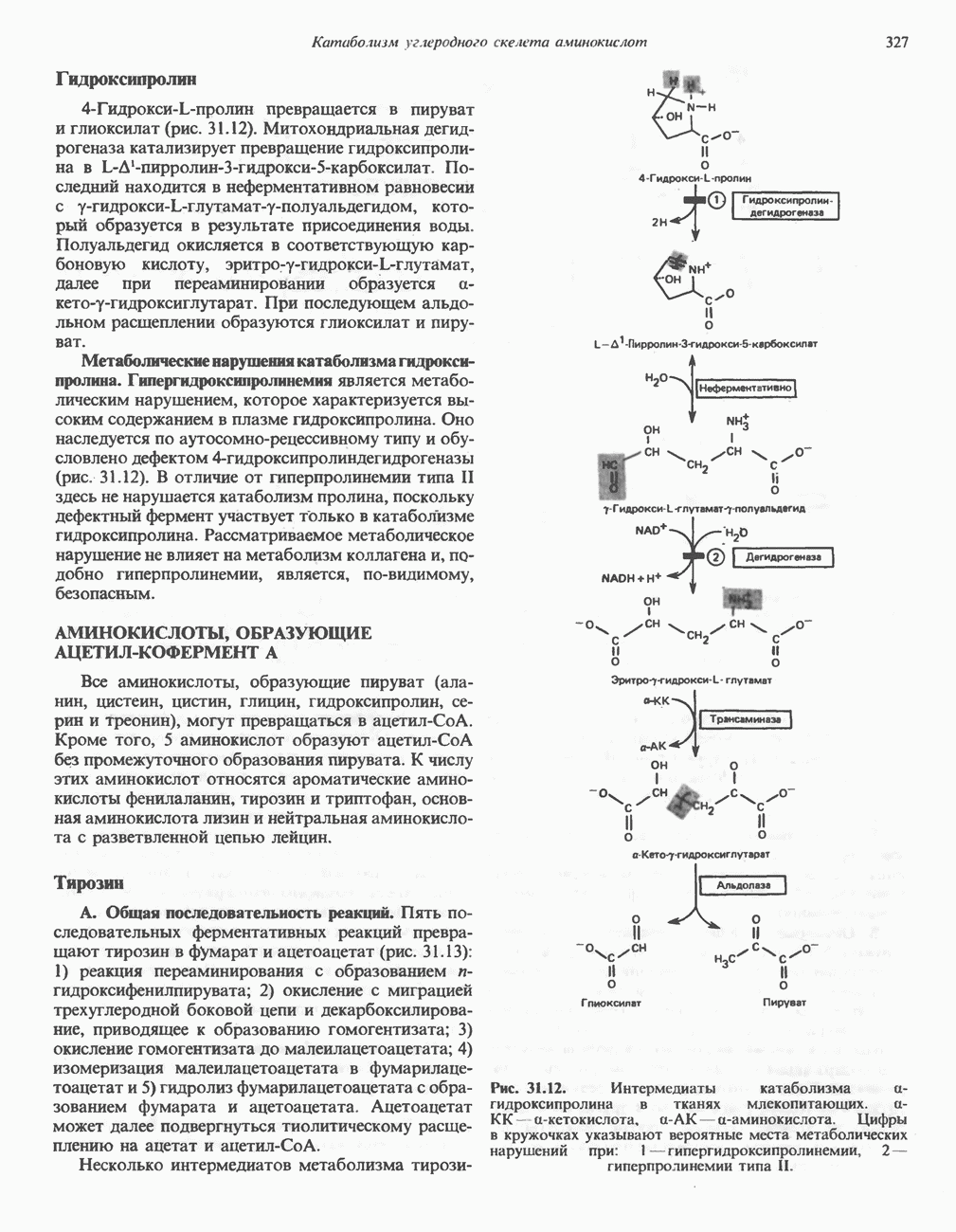
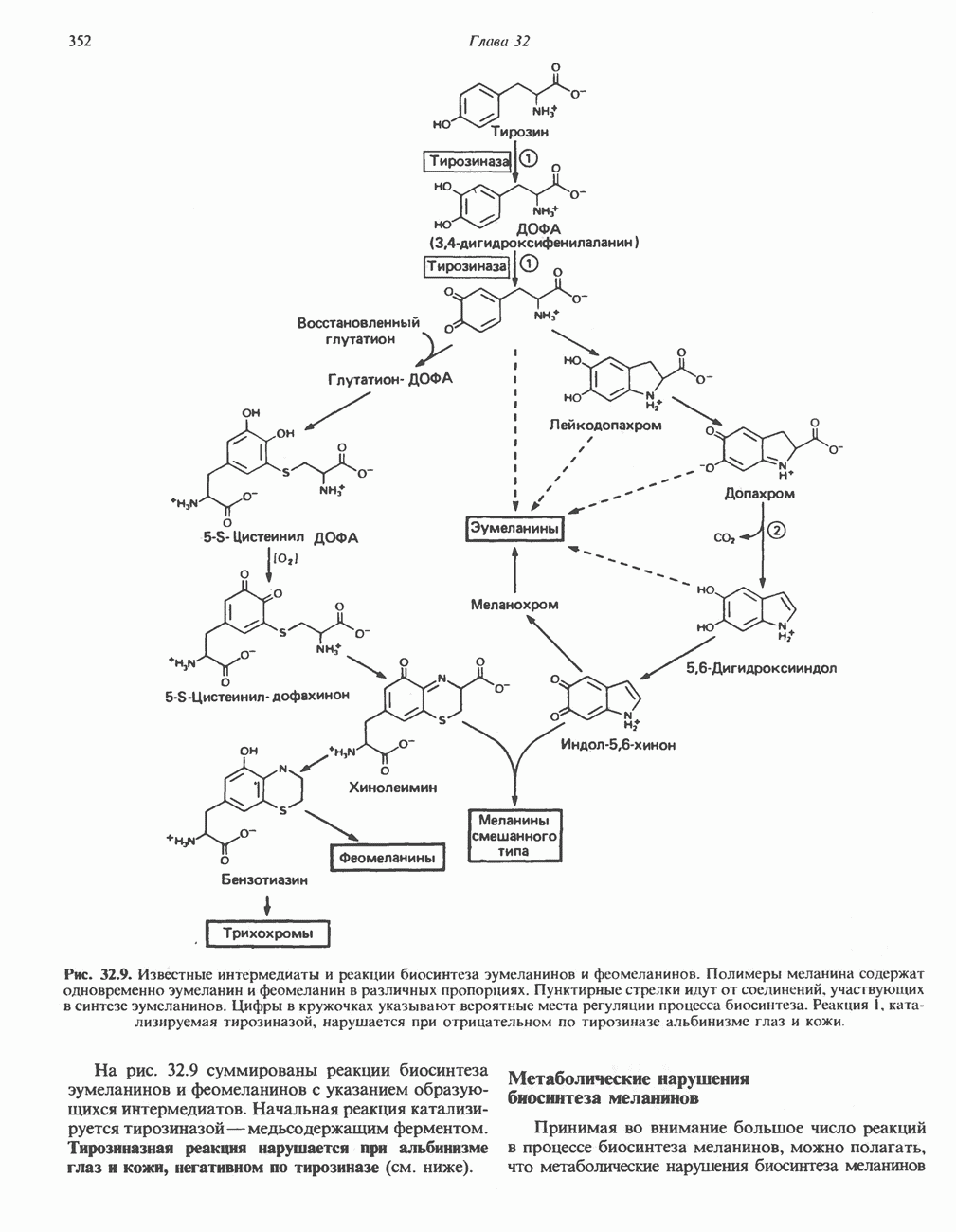
ФЕРМЕНТЫ И КОФЕРМЕНТЫ, УЧАСТВУЮЩИЕ В ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНЫХ ПРОЦЕССАХФерменты, катализирующие окислительновосстановительные реакции, называют оксидоредуктазами. Их разделяют на 5 групп. 1) Оксидазы. Истинные оксидазы катализируют удаление водорода из субстрата, используя при этом в качестве акцептора водорода только кислород. Они неизменно содержат медь, продуктом реакции является вода (исключение составляют реакции, катализируемые уриказой и моноаминоксидазой, в результате которых образуется 2) Аэробные дегидрогеназы—ферменты, катализирующие удаление водорода из субстрата; в отличие от оксидаз они могут использовать в качестве акцептора водорода не только кислород, но и искусственные акцепторы, такие, как метиленовый синий. Эти дегидрогеназы относятся к флавопротеинам, и продуктом катализируемой ими реакции является перекись водорода, а не вода (рис. 12.2). 3) Анаэробные дегидрогеназы—ферменты, ката-, лизирующие удаление водорода из субстрата, но не способные использовать кислород в качестве акцептора водорода. В этот класс входит большое число ферментов. Они выполняют две главные функции. а. Перенос водорода с одного субстрата на другой в сопряженной окислительно-восстановительной реакции (рис. 12.3). Эти дегидрогеназы специфичны к субстратам, но часто используют один и тот же кофермент или переносчик водорода.
Рис. 12.1. Окисление метаболита, катализируемое оксидазой.
Рис. 12.2. Окисление метаболита, катализируемое аэробной дегидрогеназой.
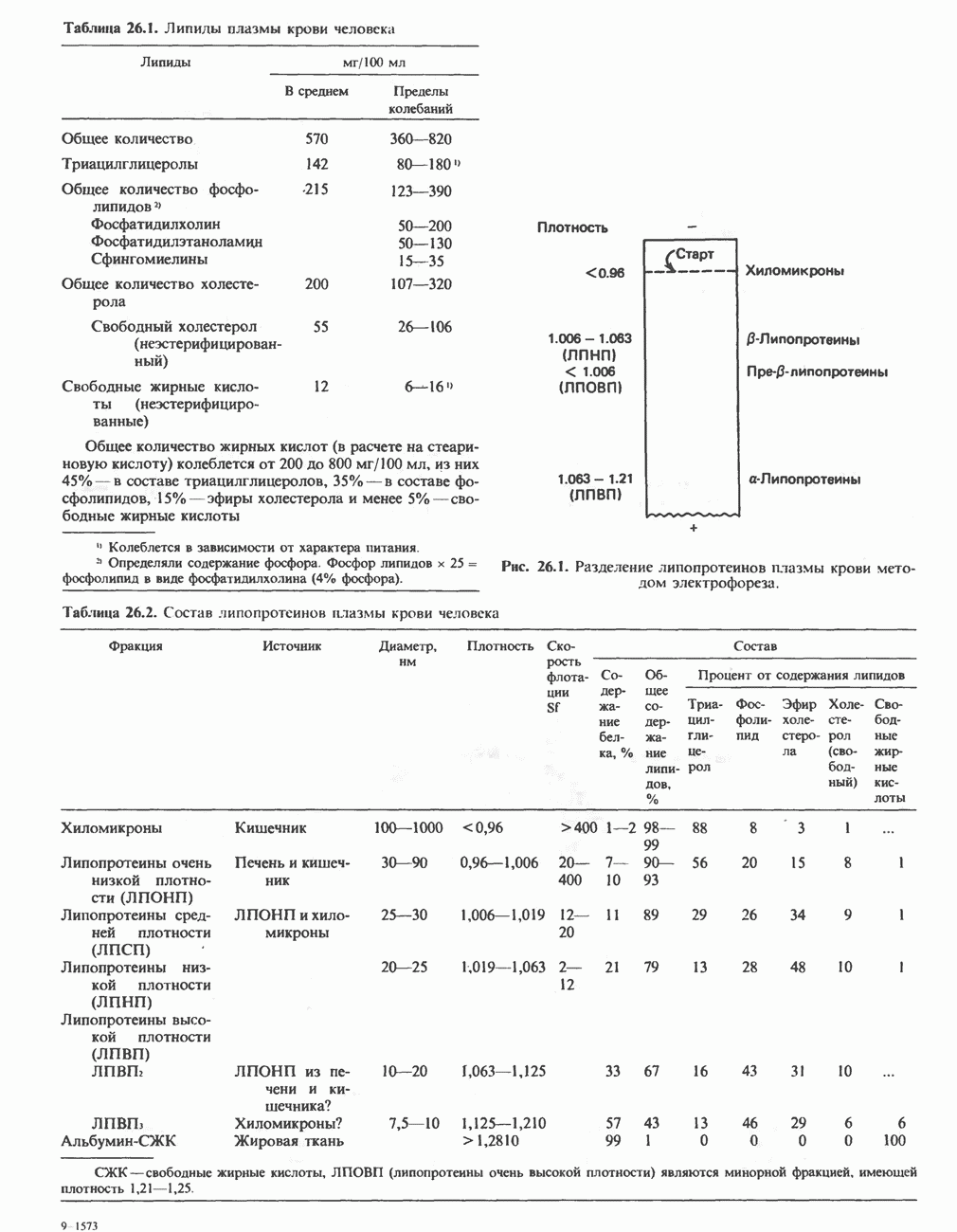
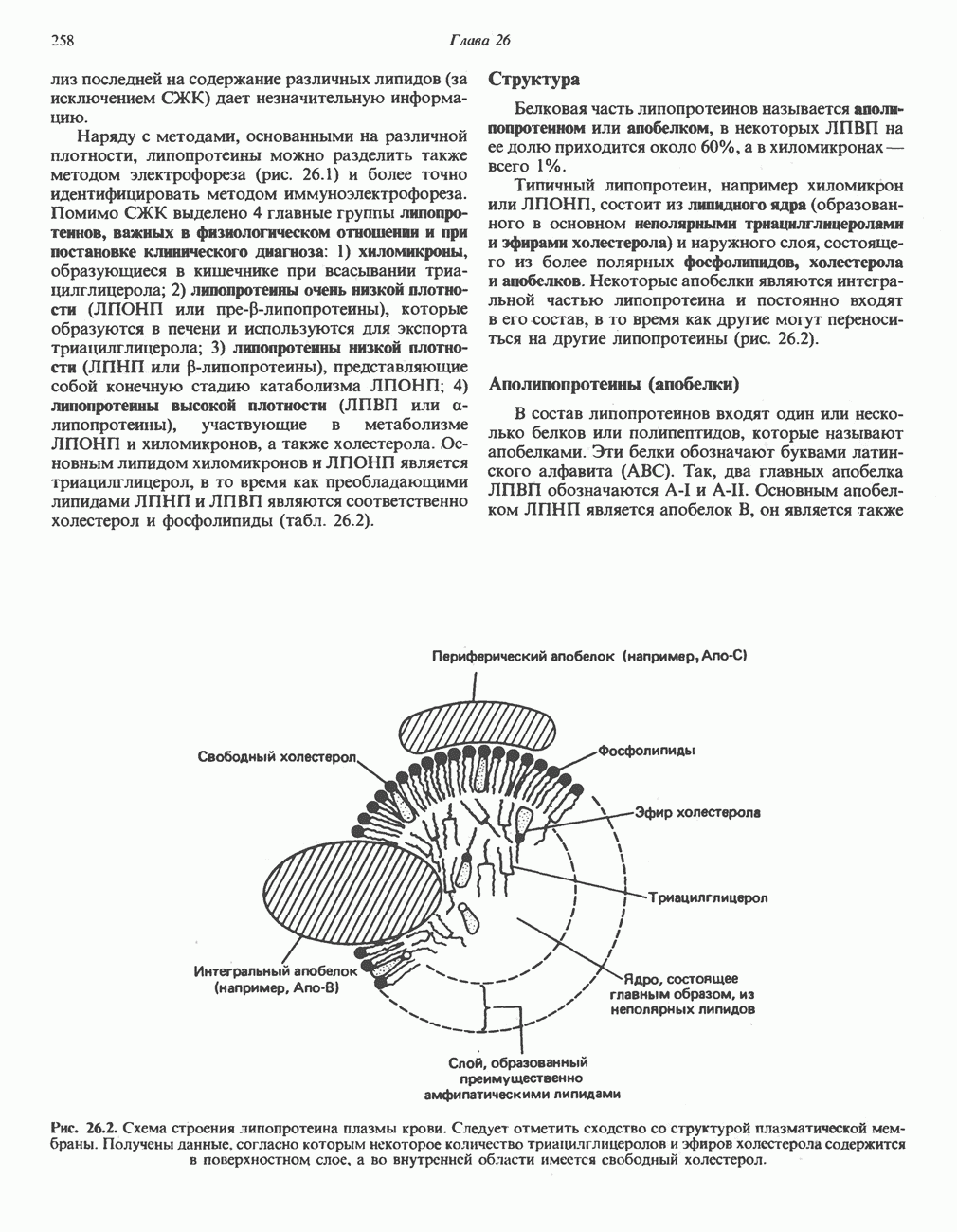
Рис. 12.3. Окисление метаболита, катализируемое анаэробными дегидрогеназами (без участия дыхательной цепи). Поскольку рассматриваемые реакции обратимы, они обеспечивают в клетке свободный перенос восстановительных эквивалентов. Реакции этого типа, приводящие к окислению одного субстрата за счет восстановления другого, особенно важны для осуществления окислительных процессов в отсутствие кислорода. б. Функцию компонентов дыхательной цепи, обеспечивающих транспорт электронов от субстрата на кислород (рис. 12.4).
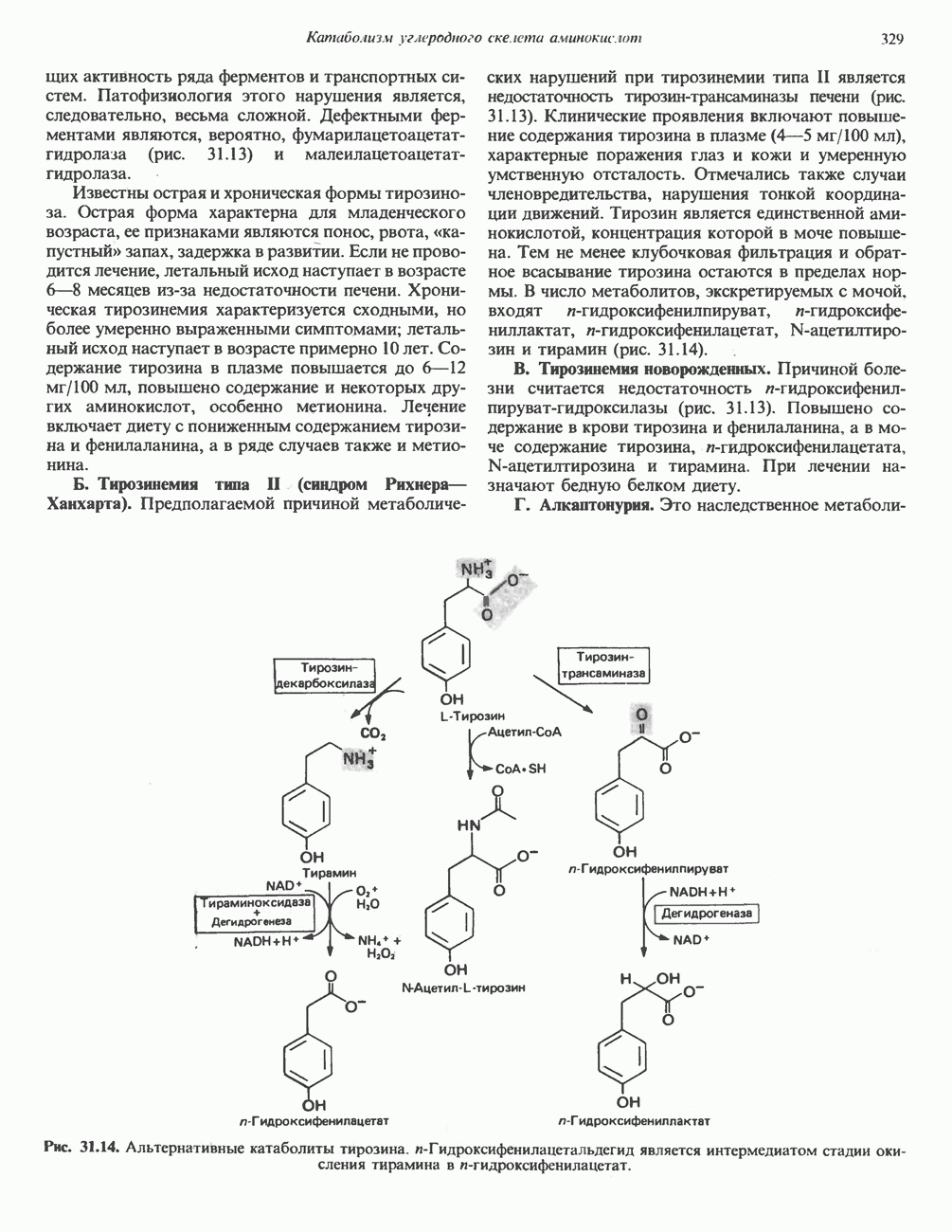
Рис. 12.4. Окисление метаболита анаэробными дегидрогеназами и — на завершающей стадии — истинной оксидазой дыхательной цепи. 4) Гидрокснпероксидазы — ферменты, использующие в качестве субстрата перекись водорода или органические перекиси. К этой категории относятся два типа ферментов: пероксидазы, находящиеся в составе молока, в растениях, лейкоцитах, тромбоцитах, эритроцитах и т. д., и каталаза, функционирующая в тканях животных и растений. 5) Окснгеназы — ферменты, катализирующие прямое введение кислорода в молекулу субстрата. ОксидазыЦитохромоксидаза—гемопротеин, широко распространенный в растительных и животных тканях. Она служит конечным компонентом цепи дыхательных переносчиков, локализованных в митохондриях, и катализирует реакцию, в результате которой электроны, высвобождающиеся из молекул субстрата при их окислении дегидрогеназами, переносятся на конечный акцептор — кислород. Данный фермент отравляется окисью углерода, цианидом и сероводородом. Иногда цитохромоксидазу называют цитохромом Фенолаза (тирозиназа, полифенолоксидаза, катехолоксидаза) — это медьсодержащий фермент с широкой специфичностью. Он катализирует превращение монофенола (в присутствии о-дифенола) в о-хинон. Медь обнаружена в ряде других ферментов, в частности в уриказе, катализирующей окисление мочевой кислоты в аллантоин, и в моноаминоксндазе, окисляющей адреналин и тирамин в митохондриях. Аэробные дегидрогеназыАэробные дегидрогеназы являются флавопротеи-нами; они содержат в качестве простетической группы флавинмононуклеотид (FMN) или флавинадрнин-дннуклеотнд (FAD). FMN и FAD образуются в организме человека Рибофлавин (витамин
Рис. 12.5. Рибофлавин. В тканях рибофлавин путем FMN и FAD обычно прочно — но не ковалентно—связаны с соответствующим апоферментом. Многие флавопротеиновые ферменты содержат один или несколько ионов металлов, выполняющих роль кофакторов; такие флавопротеиновые ферменты называют металлофлавопротеннамн. К ферментам группы аэробных дегидрогеназ относится также дегидрогеназа
Рис. 12.6. Рибофлавинфосфат (флавинмононуклеотид,
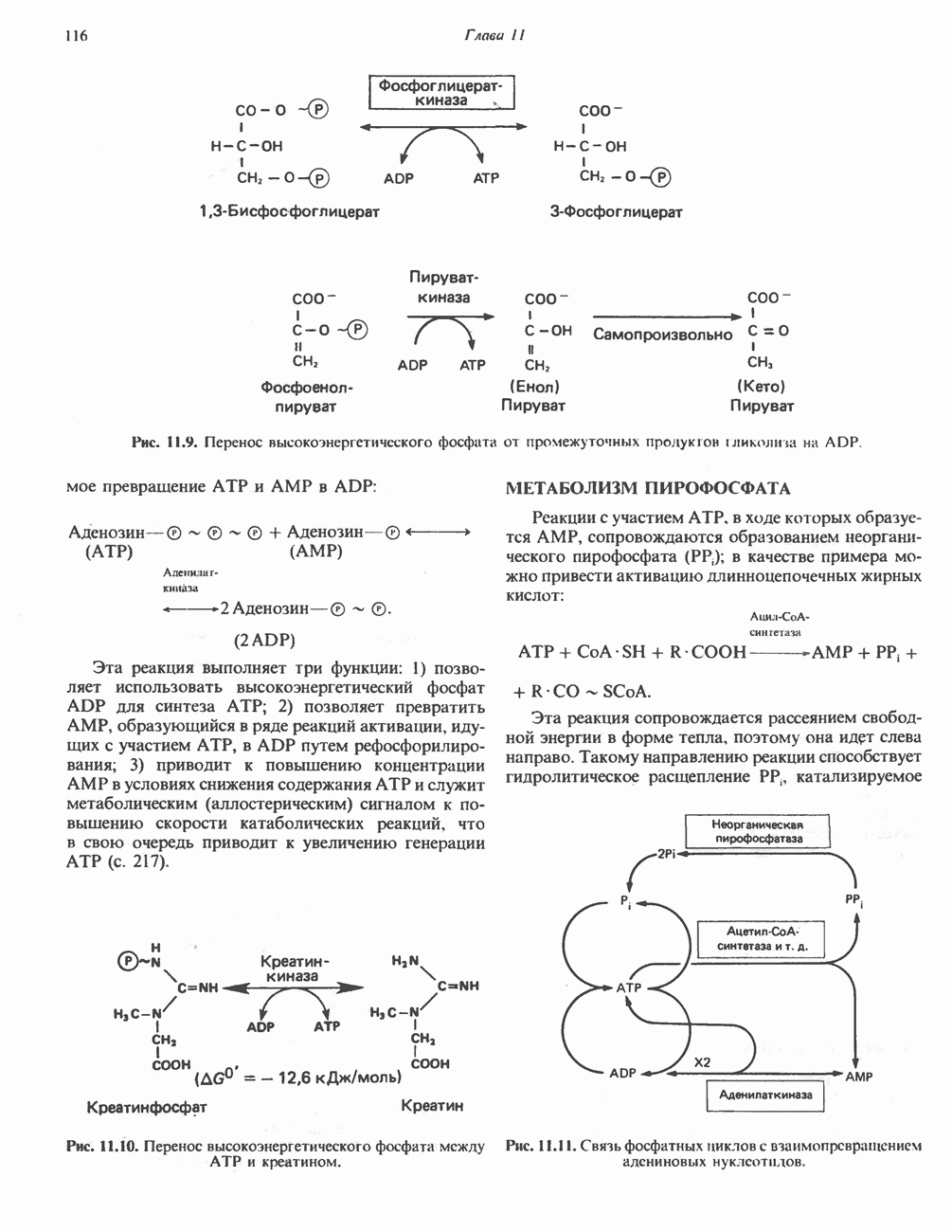
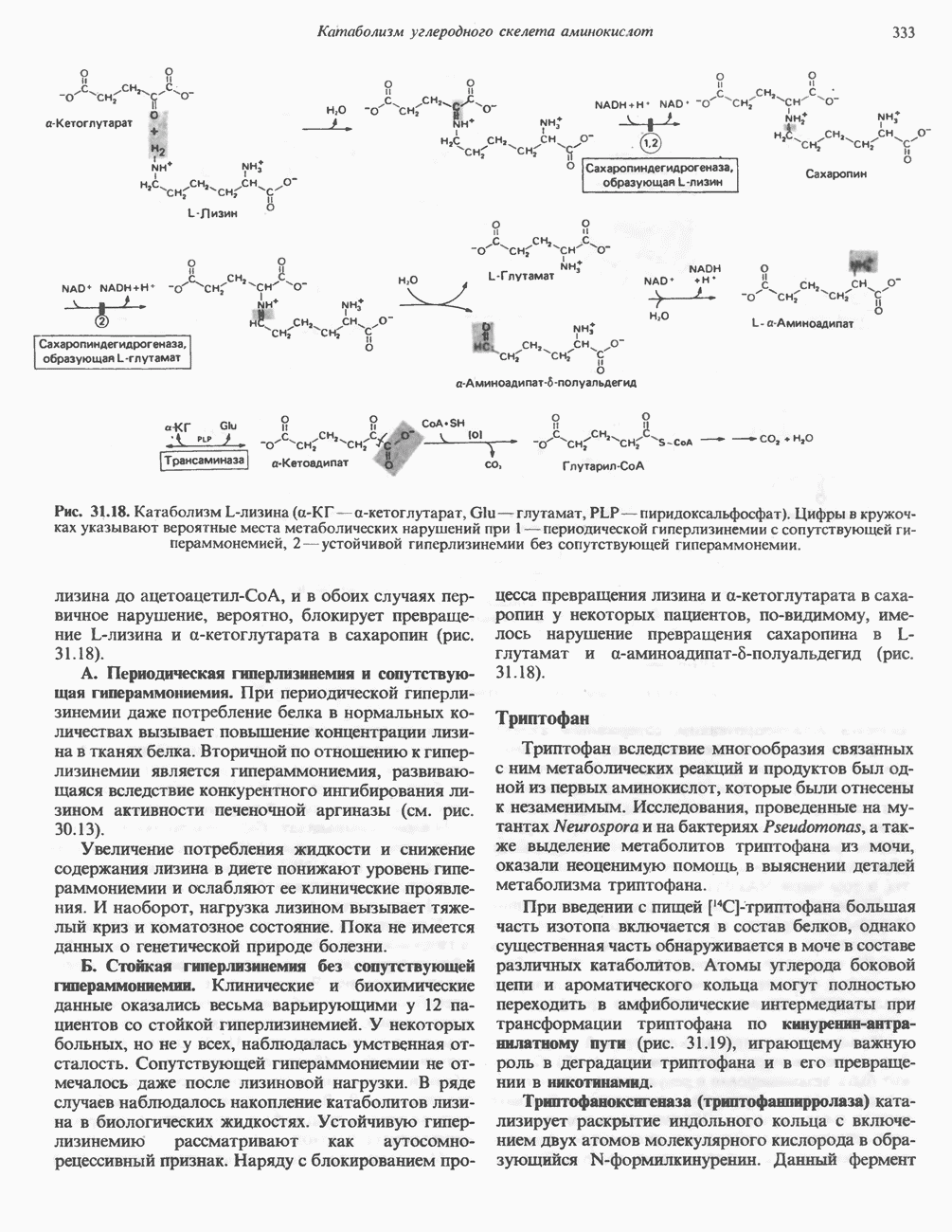
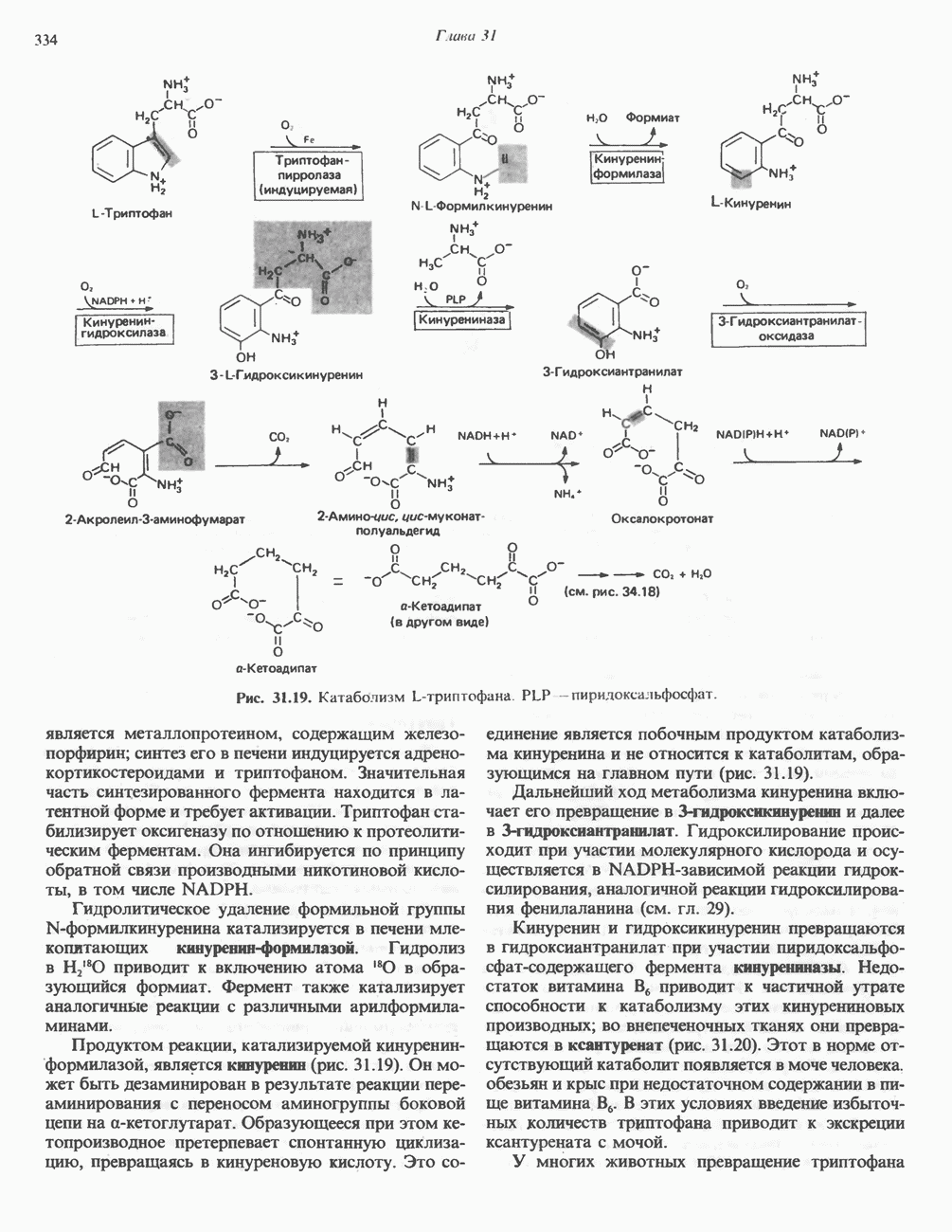
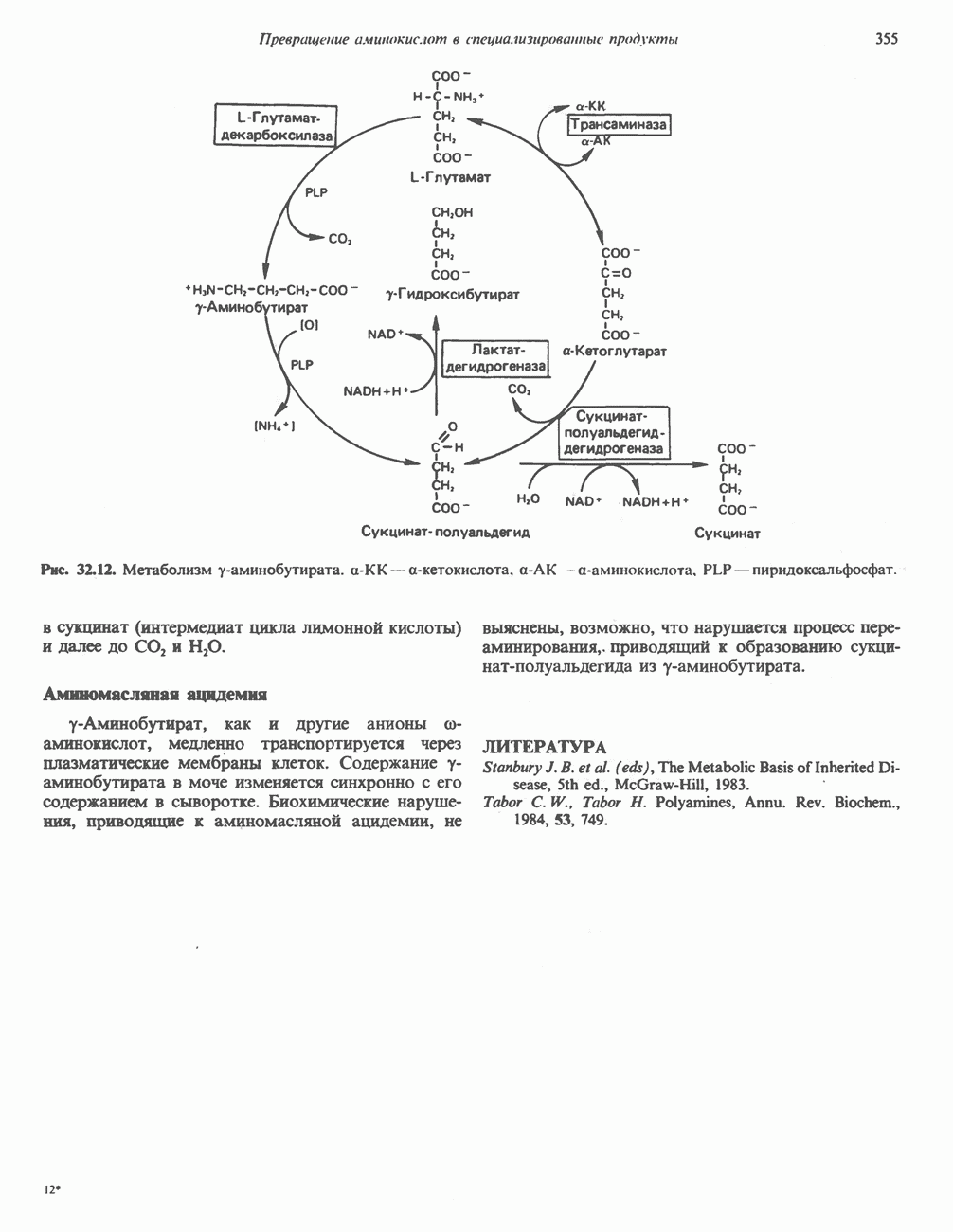
Рис. 12.7. Флавинадениндинуклеотид (FAD). Альдегиддегидрогеназа - FAD-содержащий фермент, находящийся в печени млекопитающих. Это — металлофлавопротеин, содержащий молибден и негемовое железо, окисляющий альдегиды и Глюкозооксидаза-FAD - специфичный фермент, получаемый из грибов. Она важна тем, что используется при определении глюкозы. Механизм окисления и восстановления, осуществляемый этими ферментами, весьма сложен. Судя по имеющимся данным, происходит двустадийное восстановление изоаллоксазинового кольца с промежуточным образованием семихинона (свободного радикала) (рис. 12.8). Анаэробные дегидрогеназы А. Дегидрогеназы, зависимые от никотинамидных коферментов. В эту категорию попадает большое число дегидрогеназ. Они специфичны либо к никотинамидадениндинуклеотиду В общем случае NAD-зависимые дегидрогеназы катализируют окислительно-восстановительные реакции окислительных путей метаболизма — гликолиза, цикла лимонной кислоты, дыхательной цепи митохондрий. NADP-завнснмые дегидрогеназы участвуют в процессах восстановительного синтеза, в частности во внемитохондриальном синтезе жирных кислот и стероидов; они также являются коферментами дегидрогеназ пентозофосфатного пути. Некоторые дегидрогеназы, функционирующие с никотинамидными коферментами, содержат ион цинка, в частности алкогольдегидрогеназа печени и глице-ральдегид-3-фосфат-дегидрогеназа скелетных мышц. Полагают, что ионы цинка не участвуют непосредственно в процессах окисления и восстановления.
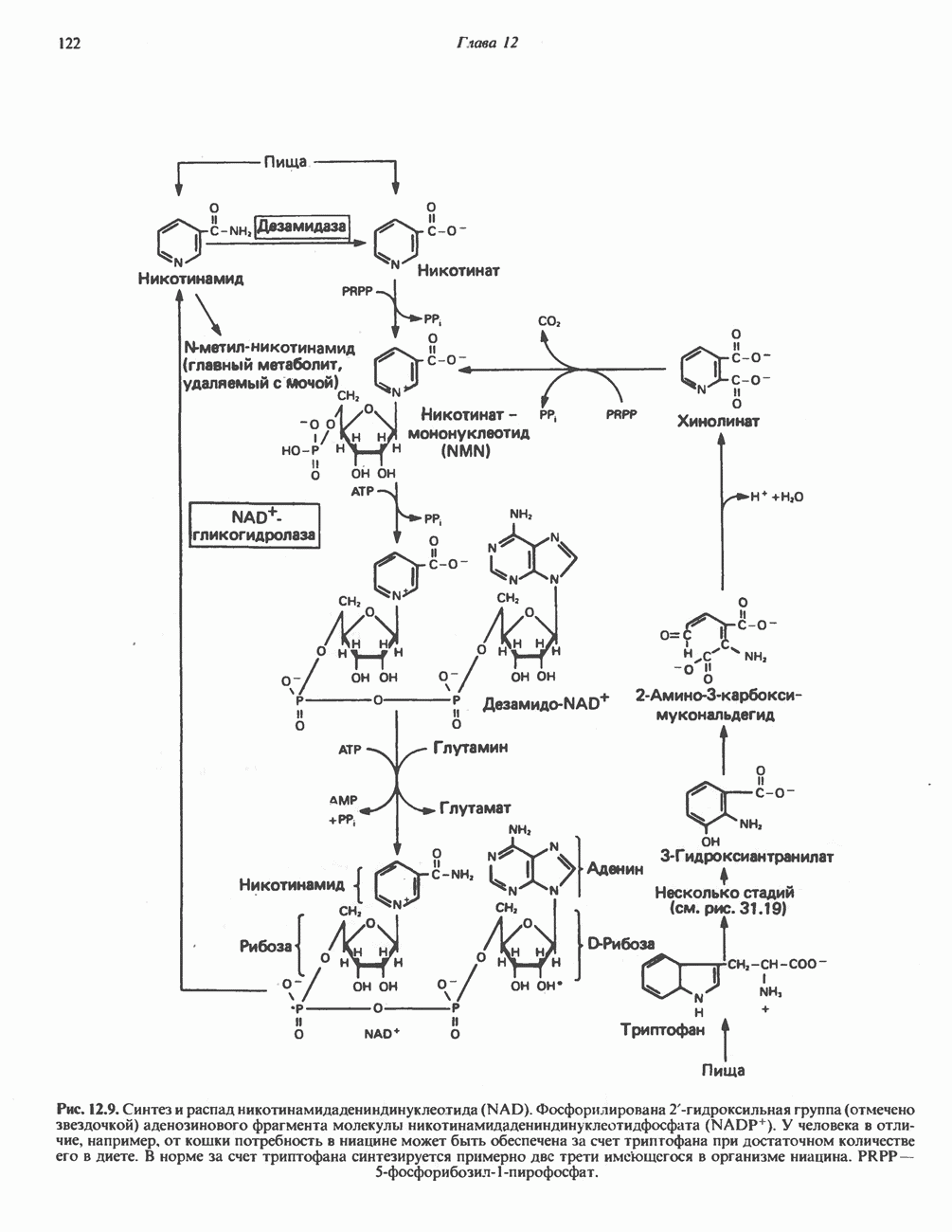
Рис. 12.8. Восстановление изоаллоксазинового кольца флавиновых нуклеотидов. (см. скан) Рис. 12.9. Синтез и распад никотинамидадениндинуклеотида (NAD). Фосфорилирована 2-гидроксильная группа (отмечено звездочкой) аденозинового фрагмента молекулы никотинамидадениндинуклеотидфосфата
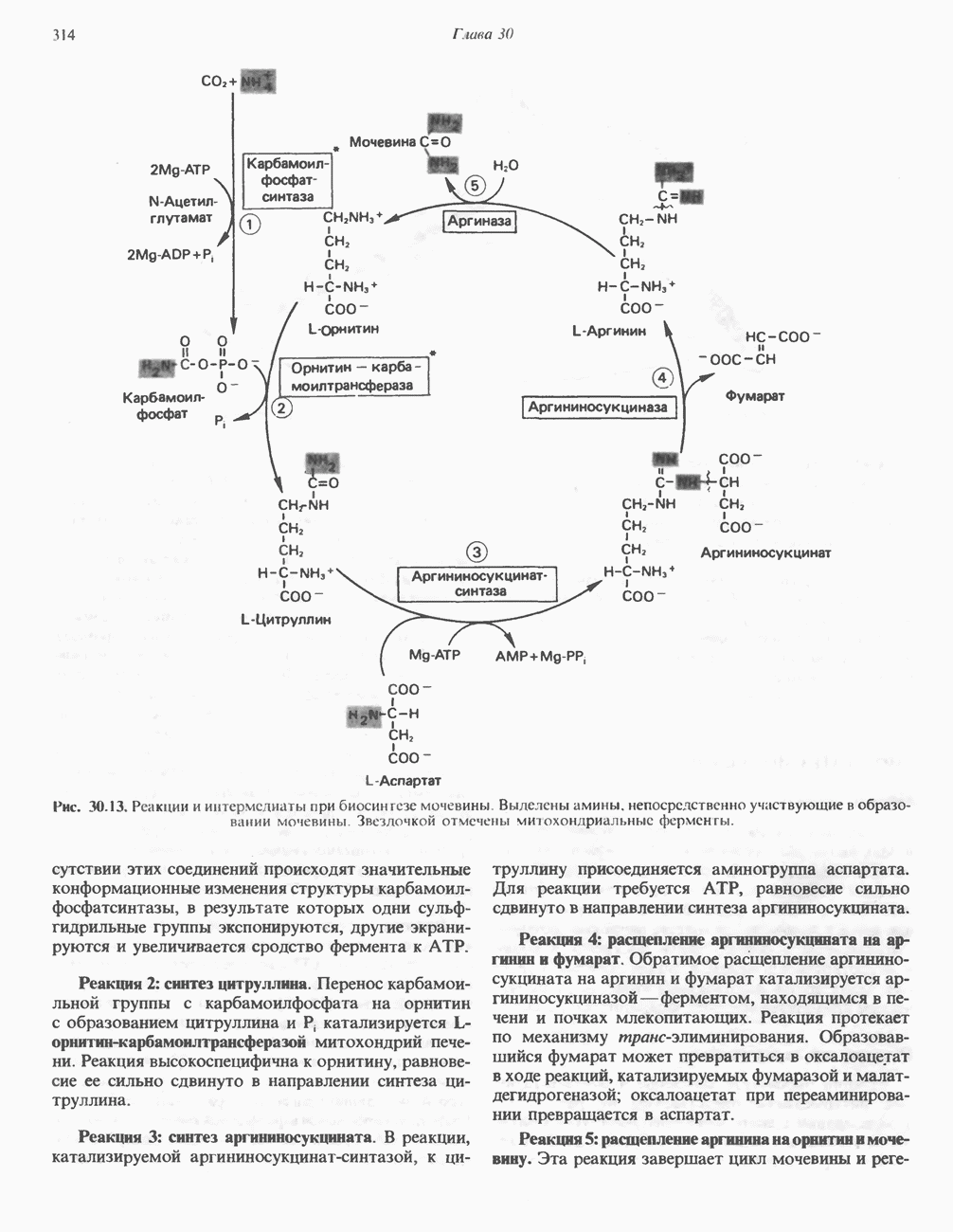
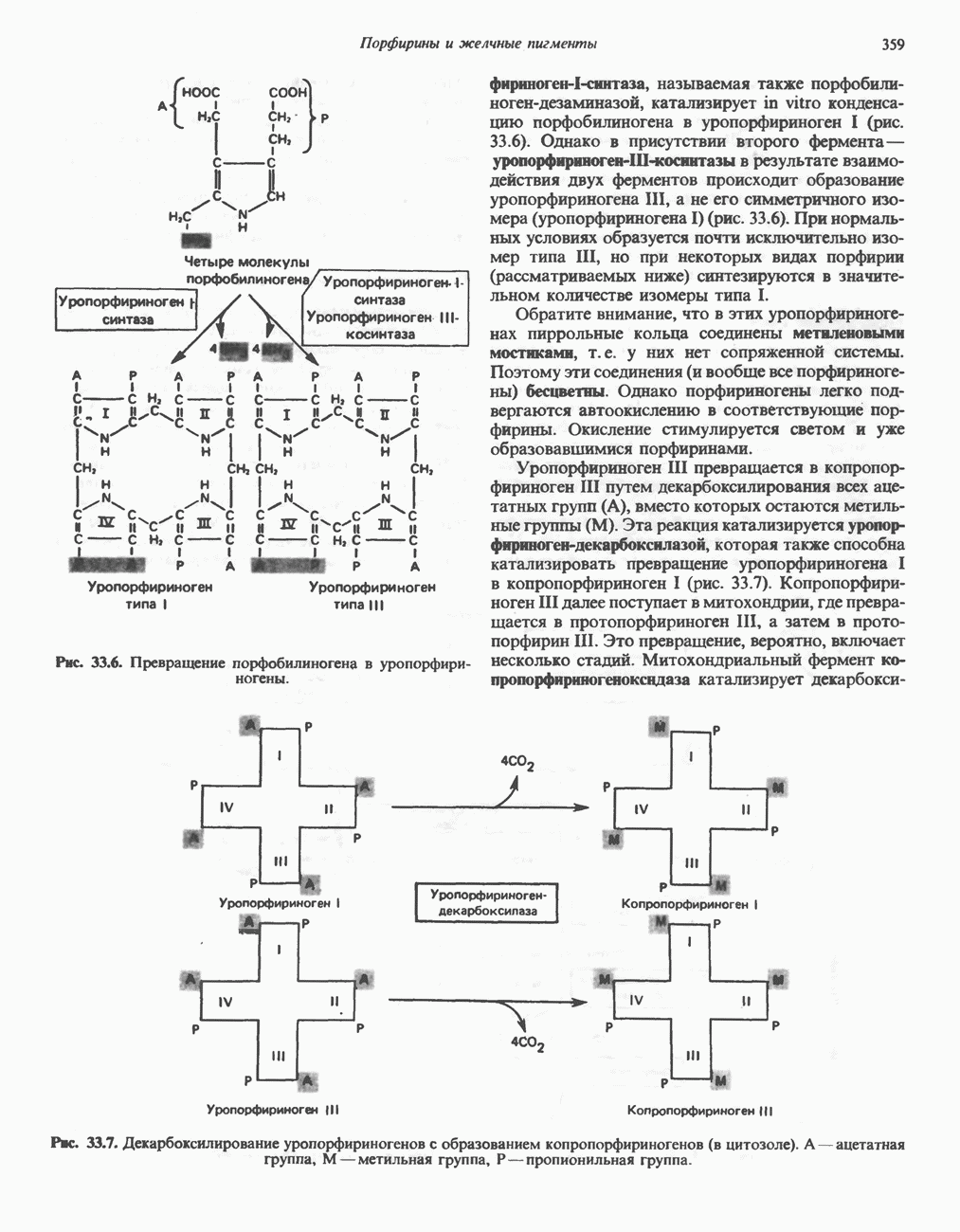
Рис. 12.10. Механизм окисления и восстановления никотинамидных коферментов. Восстановление никотинамида субстратом Б. Рибофлавннзавнсимые дегидрогеназы. Флави-новые группы этих дегидрогеназ те же, что и у аэробных дегидрогеназ, FMN и FAD. Обычно они более прочно, чем никотинамидные коферменты, связаны с апоферментом. Большинство рибофлавин-зависимых анаэробных дегидрогеназ либо участвует в транспорте электронов по дыхательной цепи, либо поставляет электроны для этой цепи. В. Цитохромы. За исключением рассмотренной выше цитохромоксидазы, цитохромы классифицируются как анаэробные дегидрогеназы. Их идентификация и изучение облегчаются тем обстоятельством, что в восстановленном состоянии они имеют характерные полосы в спектре поглощения, которые исчезают при окислении. В дыхательной цепи они служат переносчиками электронов от флавопротеинов к цитохромоксндазе. Цитохромы являются железосодержащими гемопротеинами, у которых атом железа переходит из состояния ГидропероксидазыА. Пероксидаза. Первоначально пероксидазы считались растительными ферментами, позднее они были обнаружены также в молоке, лейкоцитах, тромбоцитах, а также в тканях, в которых происходит метаболизм эйкозаноидов (см. с. 333). Простетической группой является протогем, который в отличие от гемовых групп большинства гемопротеинов весьма слабо связан с апоферментом. В реакции, катализируемой пероксидазой, перекись водорода восстанавливается за счет соединений, выступающих в качестве доноров электронов, таких, как аскорбат, хиноны или цитохром с. Реакция, катализируемая пероксидазой, имеет сложный характер; суммарная реакция выглядит следующим образом:
В эритроцитах глутатионпероксндаза, содержащая в качестве простетической группы селен, катализирует разложение Б. Каталаза. Это — гемопротеин, содержащий четыре гемовые группы. Наряду с пероксидазной активностью каталаза способна использовать одну молекулу
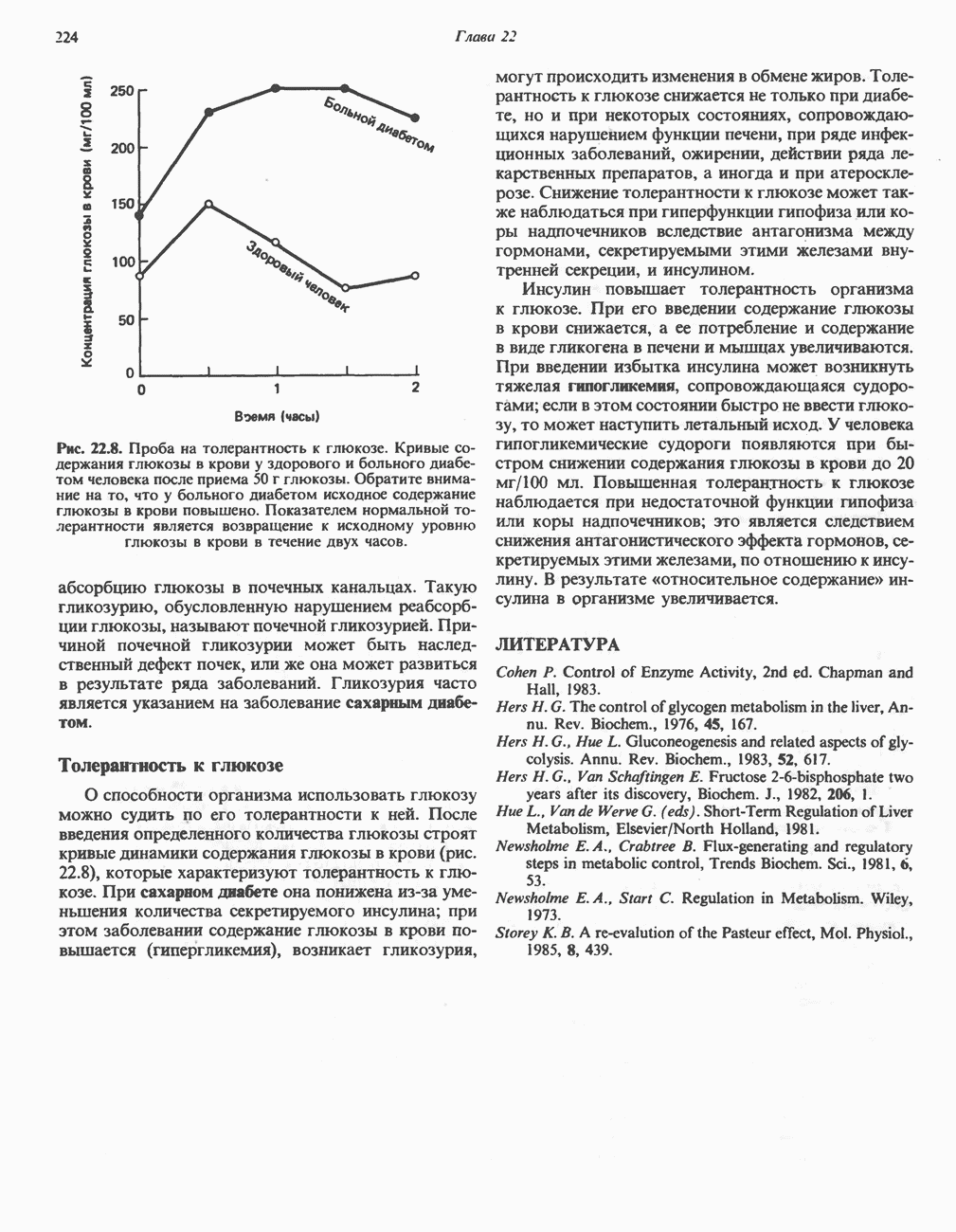
Каталаза имеется в крови, костном мозге, мембранах слизистых оболочек, почках и печени. Ее функцией считается разложение перекиси водорода, образующейся при действии аэробных дегидрогеназ. Во многих тканях, включая и печень, обнаружены микротельца, пероксисомы, которые богаты аэробными дегидрогеназами и каталазой. По-видимому, биологически выгодно группировать в одном месте как ферменты, приводящие к образованию ОксигеназыОксигеназы не относятся к ферментам, катализирующим реакции, снабжающие клетку энергией; они участвуют в синтезе и деградации многих типов метаболитов. Ферменты этой группы катализируют включение кислорода в молекулу субстрата. Оно происходит в две стадии: 1) кислород связывается с активным центром фермента; 2) происходит реакция, в результате которой связанный кислород восстанавливается или переносится на субстрат. Оксигеназы подразделяются на две подгруппы. А. Дноксигеназы (кнслород-трансферазы, истинные оксигеназы). Эти ферменты катализируют включение в молекулу субстрата обоих атомов молекулы кислорода:
Примером служат железосодержащие ферменты гомогентнзатдноксигеназа и Б. Монооксигеназы (оксидазы со смешанной функцией, гидроксилазы). Эти ферменты катализируют включение в субстрат только одного из атомов молекулы кислорода. Другой атом кислорода восстанавливается до воды; для этой цели необходим дополнительный донор электронов (косубстрат):
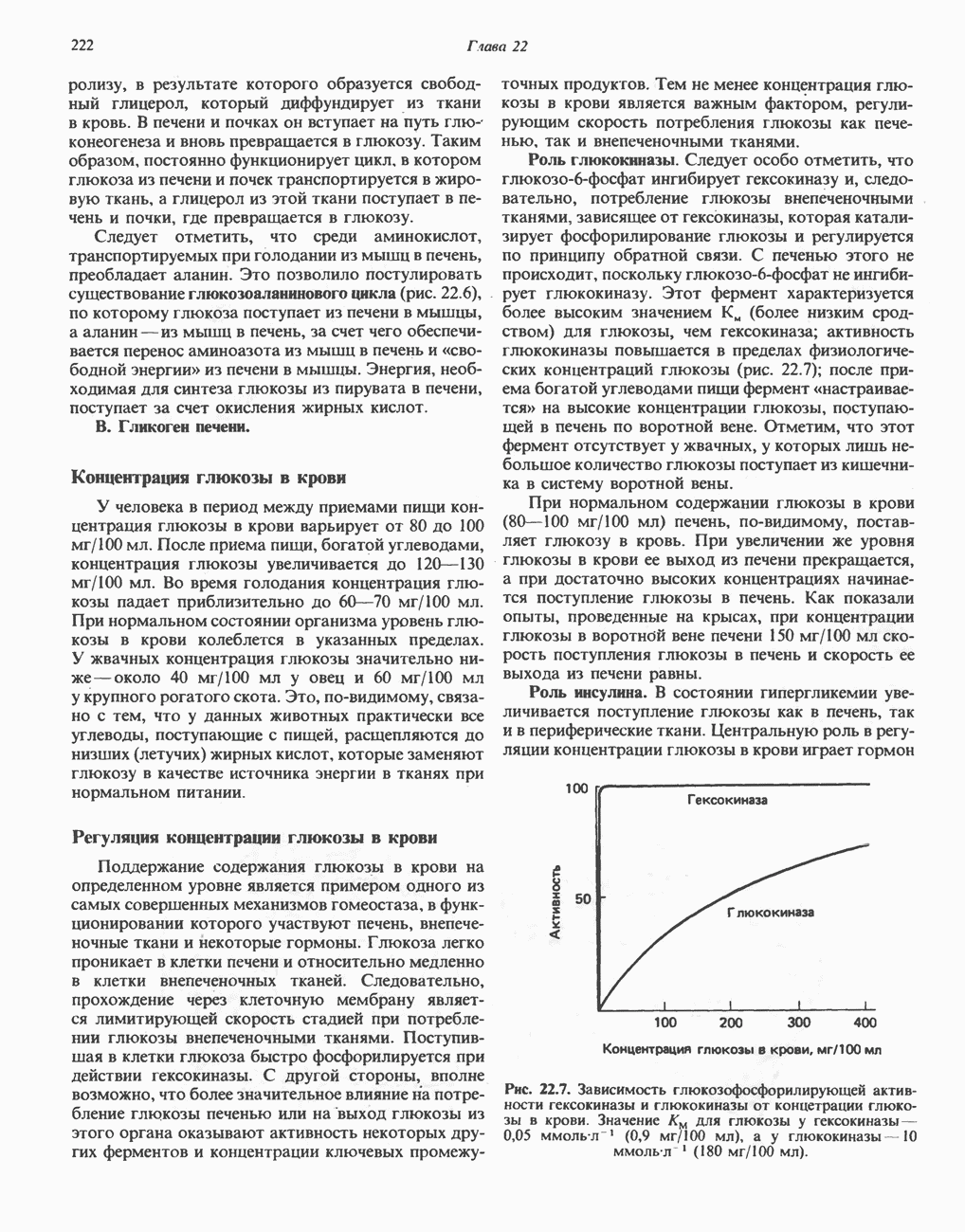
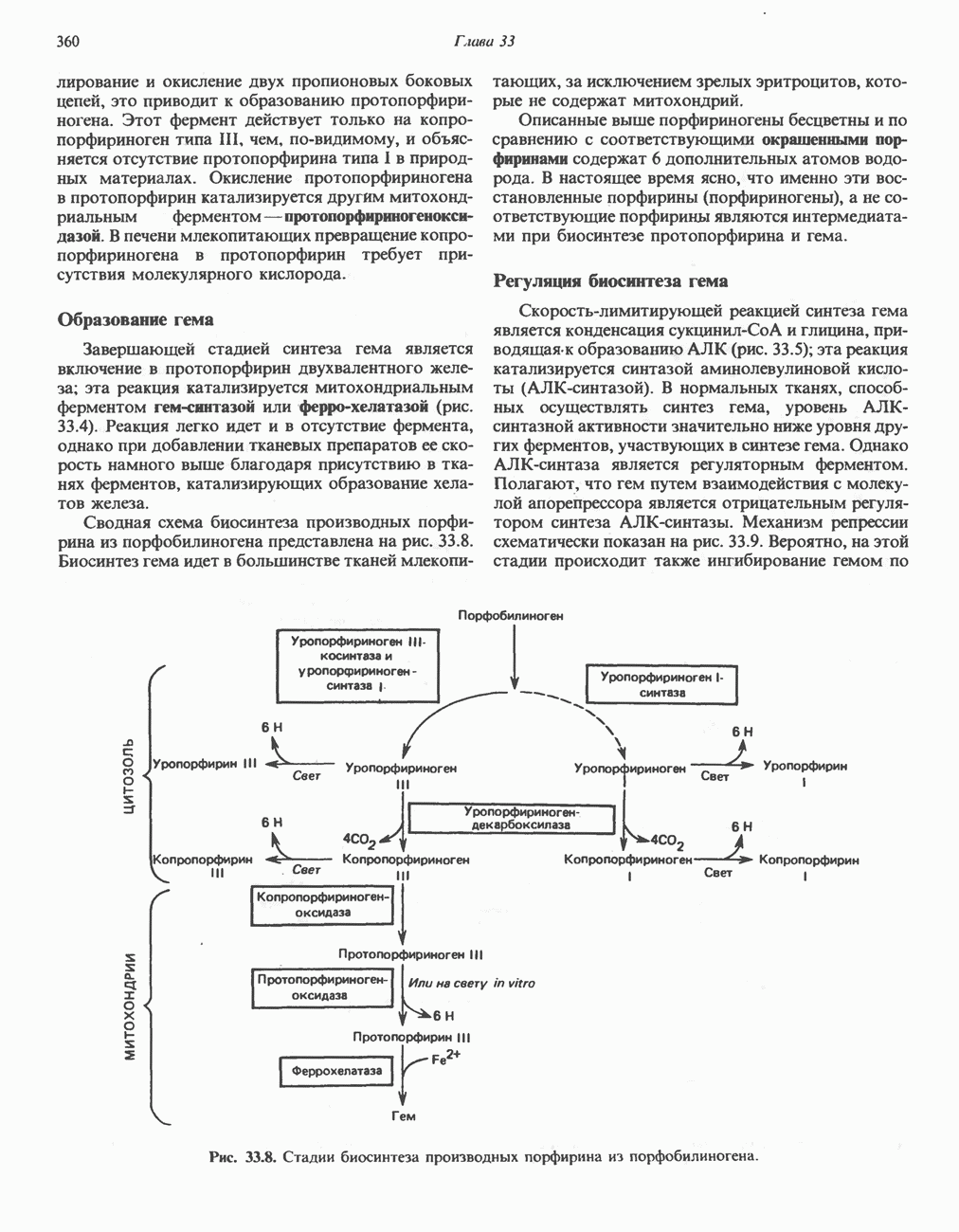
Микросомные цитохром P-450-содержащие монооксигеназные системыК этой группе относятся ферменты, участвующие в метаболизме многих лекарственных веществ путем их гидроксилирования. Они находятся в микросомах печени вместе с цитохромом Р-450 и цитохромом
(Лекарственное вещество). К лекарственным веществам, метаболизм которых идет при участии рассматриваемых систем, относятся бензпирен, аминопирин, анилин, морфин и бензофетамин. Многие лекарственные вещества, например фенобарбитал, способны индуцировать синтез микросомных ферментов и цитохрома Митохондриальные цитохром Р-450-содержащие моноокснгеназные системыЭти системы находятся в стероидогенных тканях — в коре надпочечников, в семенниках, яичниках и плаценте; они участвуют в биосинтезе стероидных гормонов из холестерола (гидроксилирование по
Рис. 12.12. Цепь транспорта электронов в микросомах. Цианид положениям Метаболизм супероксид-радикалаКислород является потенциально токсичным веществом. До недавнего времени его токсичность связывали с образованием Супероксид образуется в ходе одноэлектронного окисления молекулярным кислородом восстановленного флавина, например флавина в составе ксантиндегидрогеназы. Он образуется также при одноэлектронном окислении молекулярным кислородом восстановленного компонента дыхательной цепи:
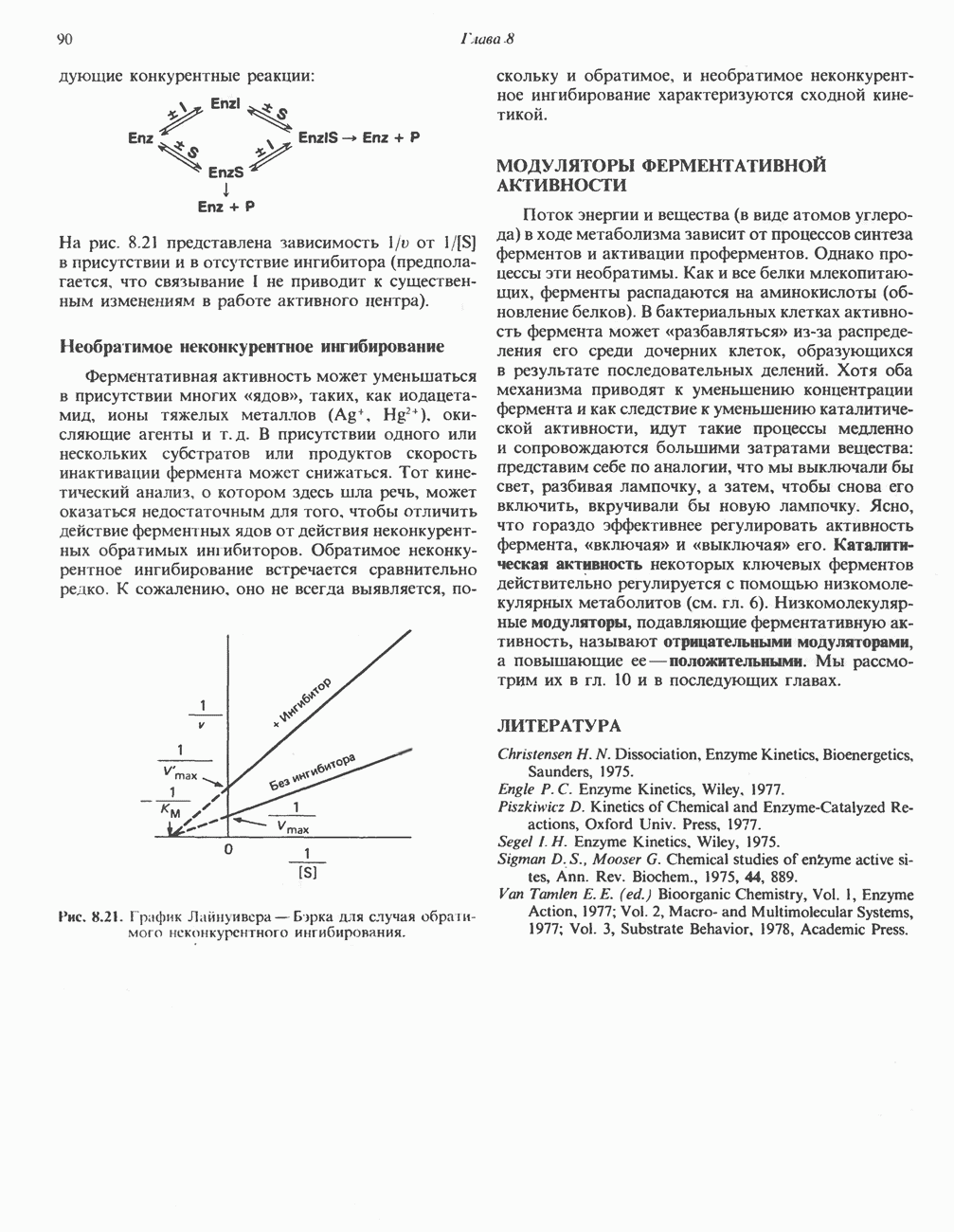
Рис. 12.13. Цитохром Р-450-гидроксилазный цикл в микросомах. Приведенная система типична для гидроксилаз стероидов в коре надпочечников. Микросомная цитохром Р-450-гидроксилаза печени не нуждается в присутствии железосерного белка
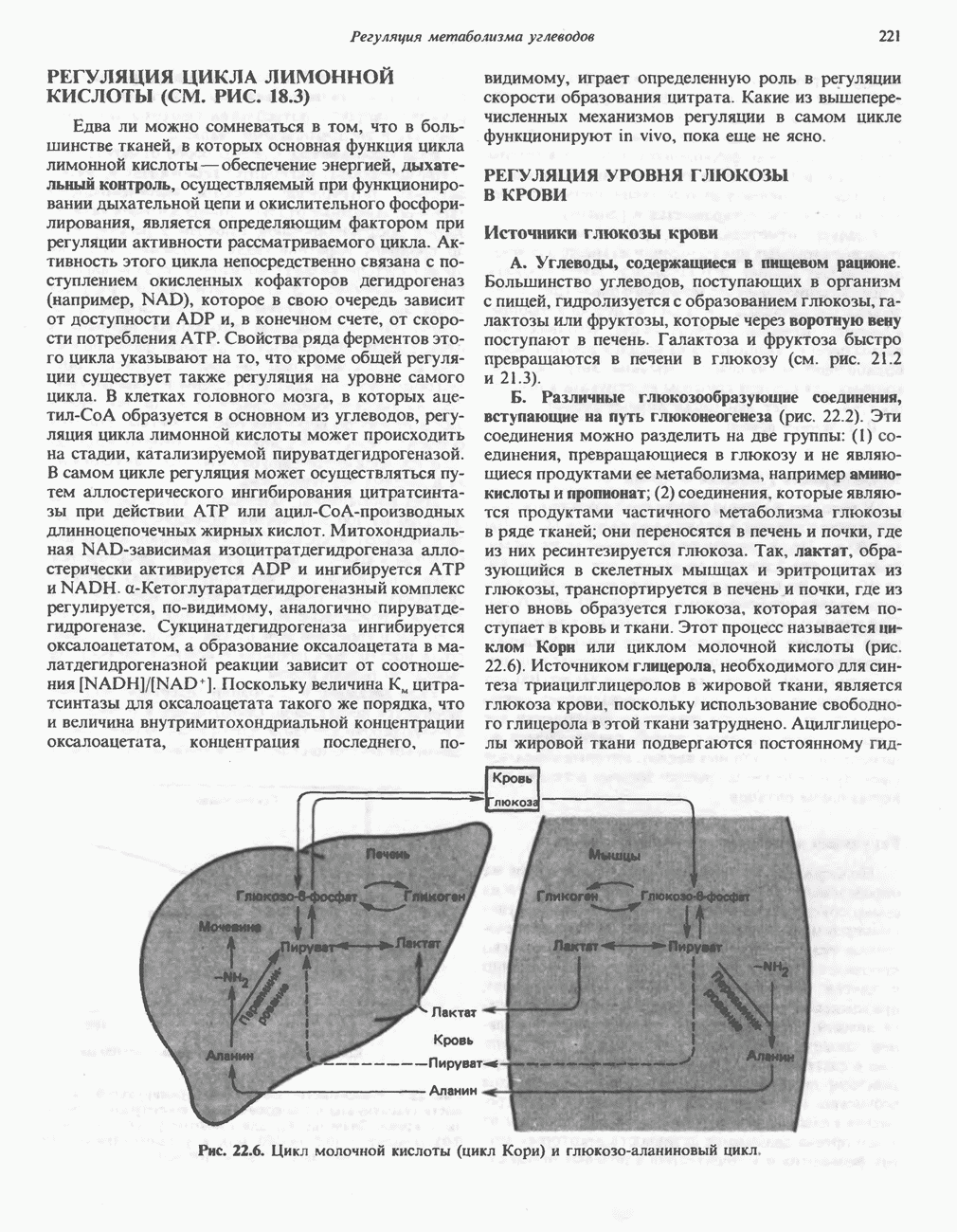
Рис. 12.14. Митохондриальная цитохром Р-450-монооксигеназная система. Супероксид может восстанавливать окисленный нитохром
Он удаляется также специфическим ферментом — супероксиддисмутазой:
В этой реакции супероксид выступает одновременно как окислитель и как восстановитель. Химическое действие супероксида в тканях усиливается в результате инициирования цепной реакции образования свободных радикалов. Было высказано предположение, что О связанный с цитохромом Р-450, является интермедиатом при активации кислорода в процессе реакций гидроксилирования (рис. 12.13). Функцией супероксиддисмутазы является, по-видимому, защита аэробных организмов от повреждающего действия супероксида. Фермент обнаруживается в нескольких внутриклеточных компартментах. Цитозольный фермент состоит из двух сходных субъединиц, содержащих по одному иону ЛИТЕРАТУРАBonnett R. Oxygen activation and tetrapyrroles, Essays Biochem., 1981, 17, 1. Ernster L. (ed.) Bioenergetics, Elsevier, 1984. Fleisher S., Packer L. (ed.) Biological oxidations, microsomal, cytochrome P-450, and other hemoprotein systems. In: Methods in Enzymology, Vol. 52, Biomembranes, part C, Academic Press, 1978. Friedovich I. Superoxide dismutases, Annu. Rev. Biochem., 1975, 44, 147. Salemme F. R. Structure and function of cytochromes c, Annu. Rev. Biochem., 1977, 46, 299. Schenk man J. B., Jansson /.. Rohie-Svh К. M. The many roles of cytochrome b5 in hepatic microsomes, Life Sci., 1976, 19, 611. Tolbert N. E. Metabolic pathways in peroxisomes and glyoxyso-mes. Annu. Rev. Biochem., 1981, 50, 133. Tyler D. D., Sutton С. M. Respiratory enzyme systems in mitochondrial membranes, Page 33. In: Membrane Structure and Function, Vol. 5, Bittar E. E. (ed.), Wiley, 1984. White R.E., Coon M.J. Oxygen activation by cytochrome P-450, Annu. Rev. Biochem., 1980, 49, 315.
|
 |
Оглавление
|


















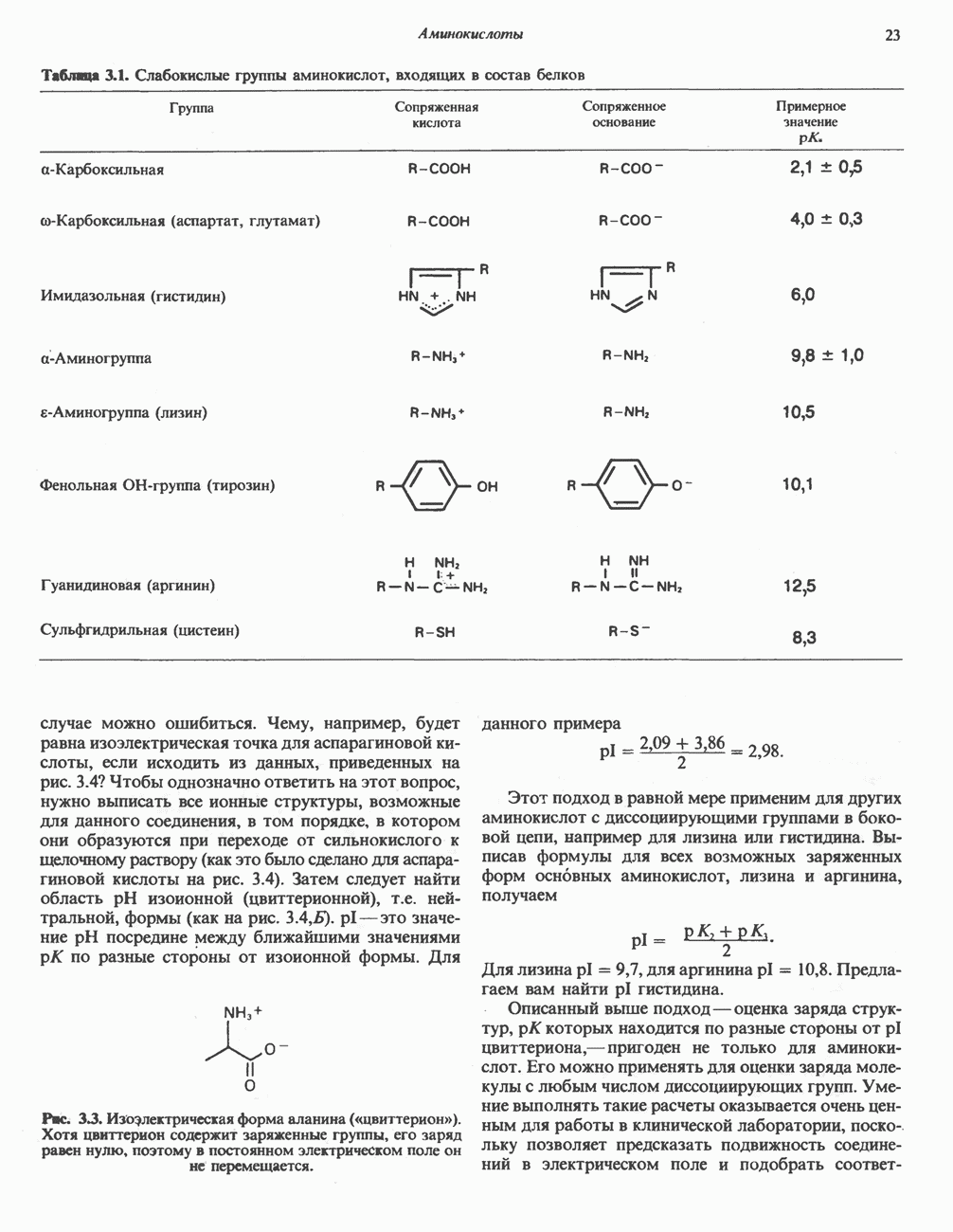





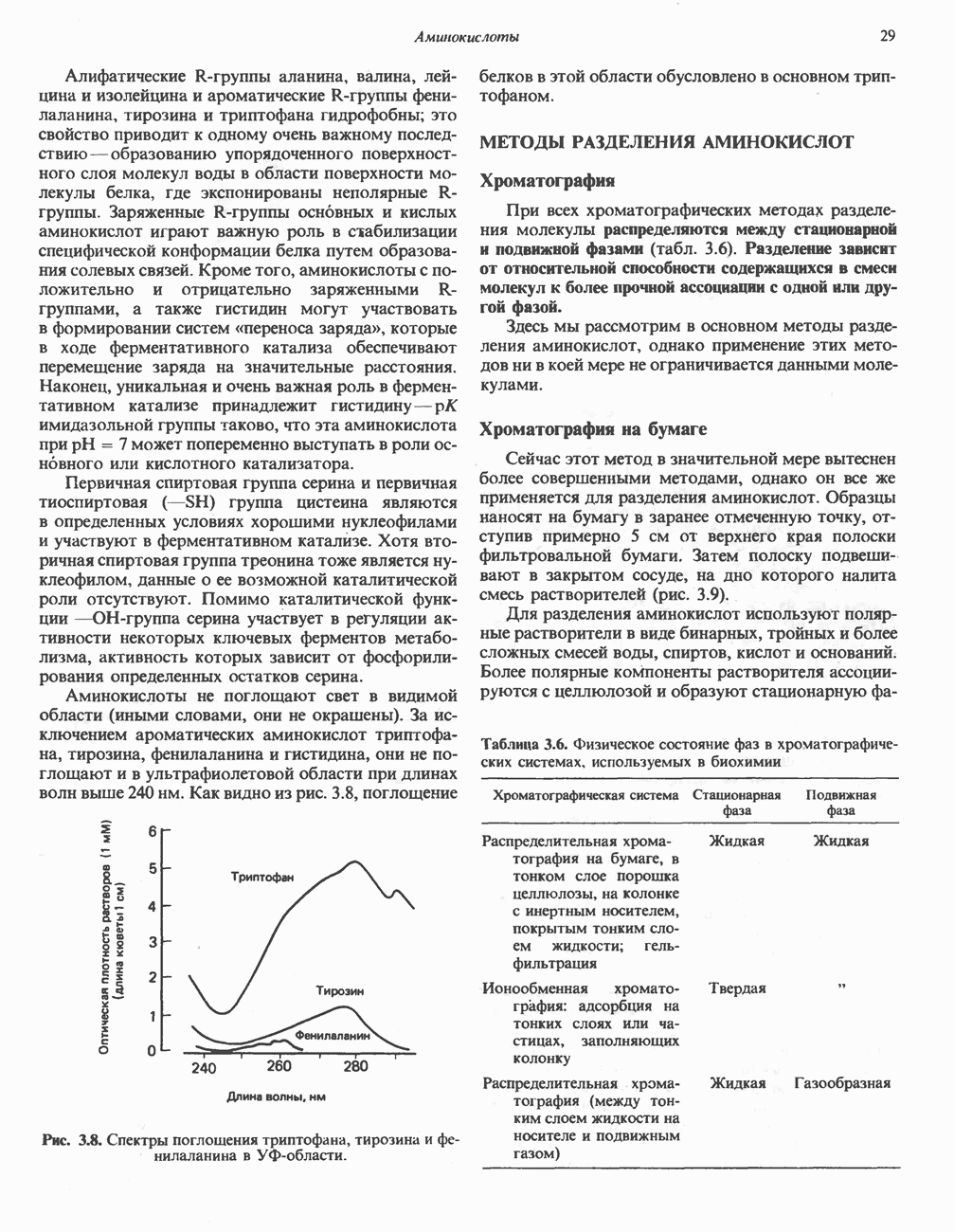
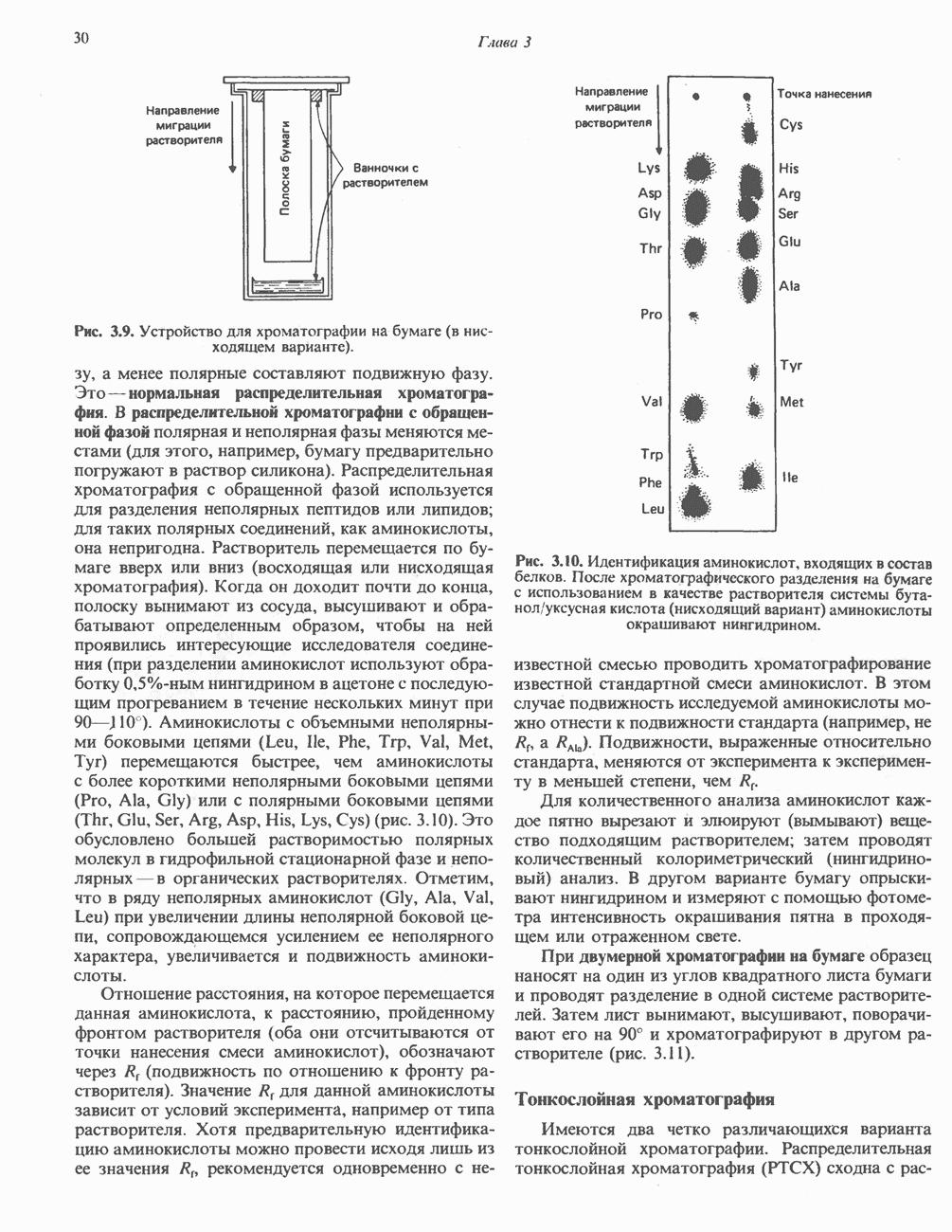












































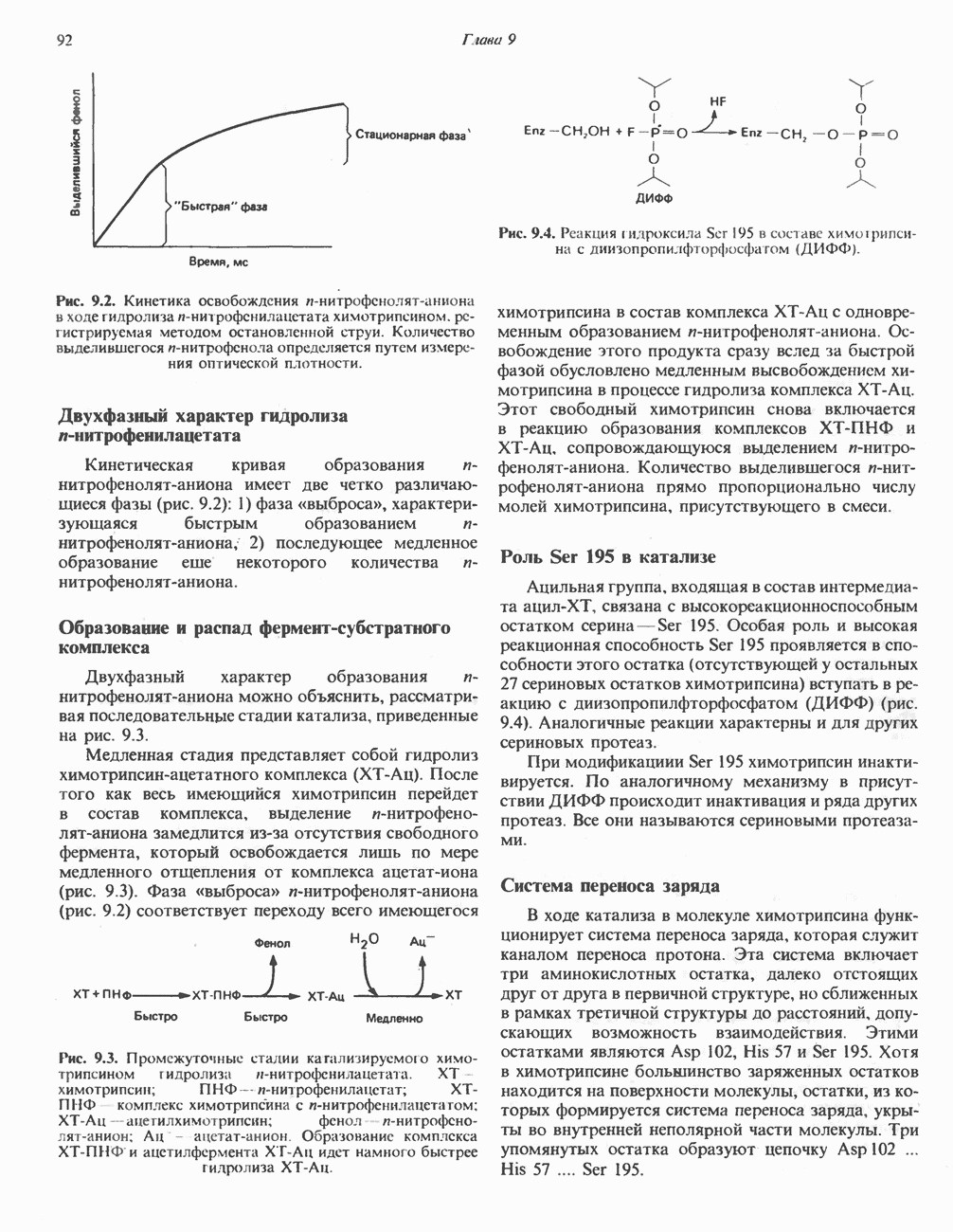

















































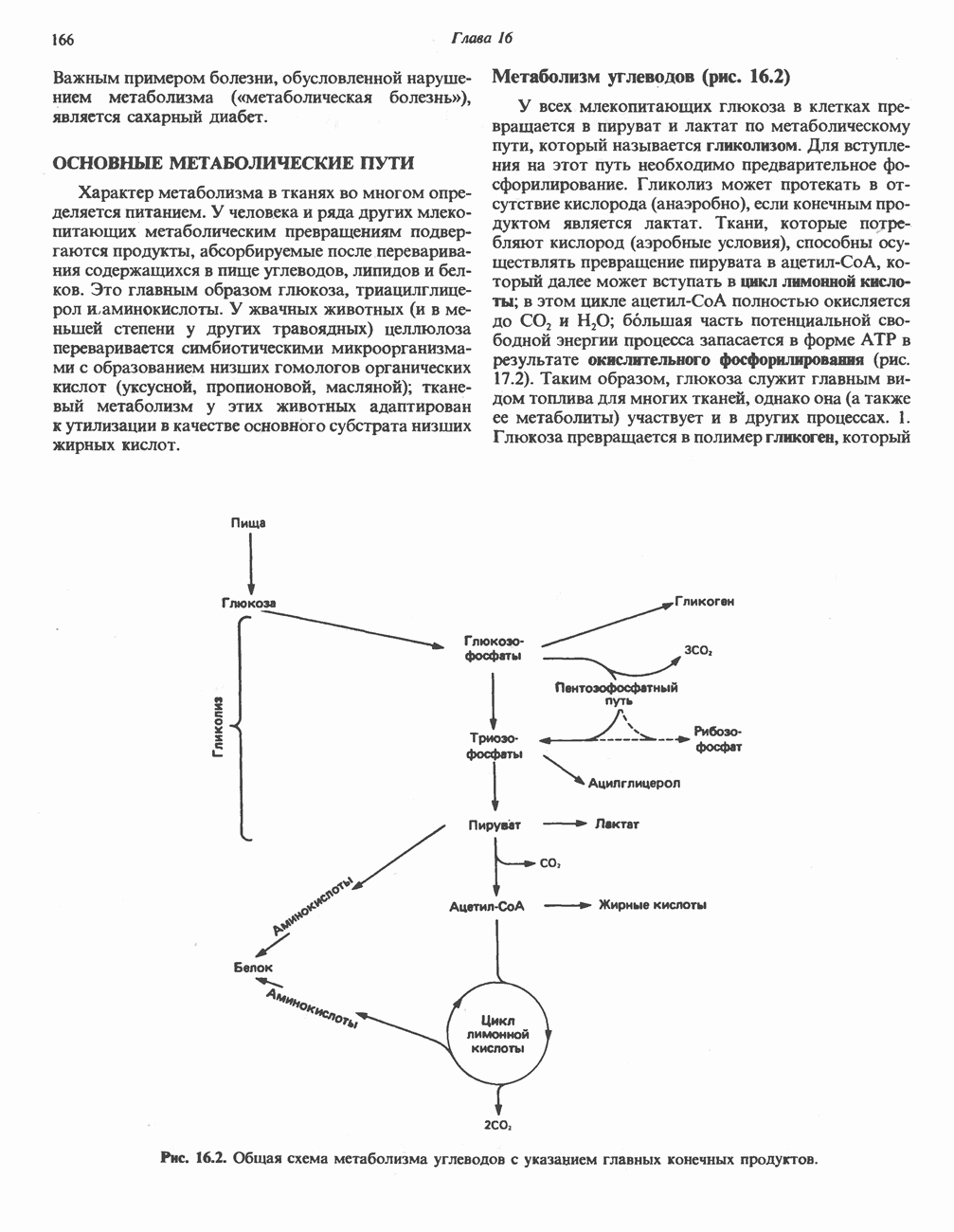


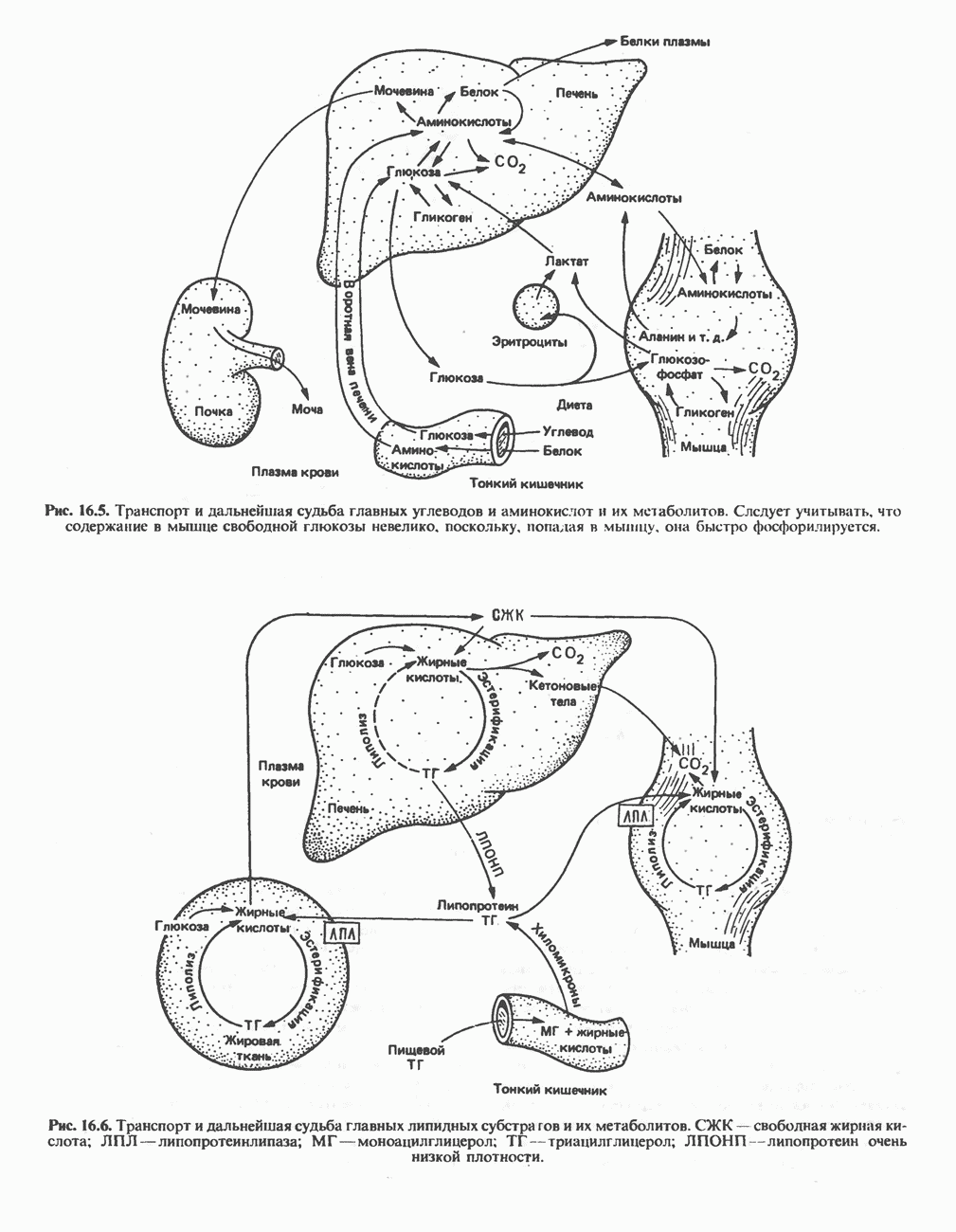































































































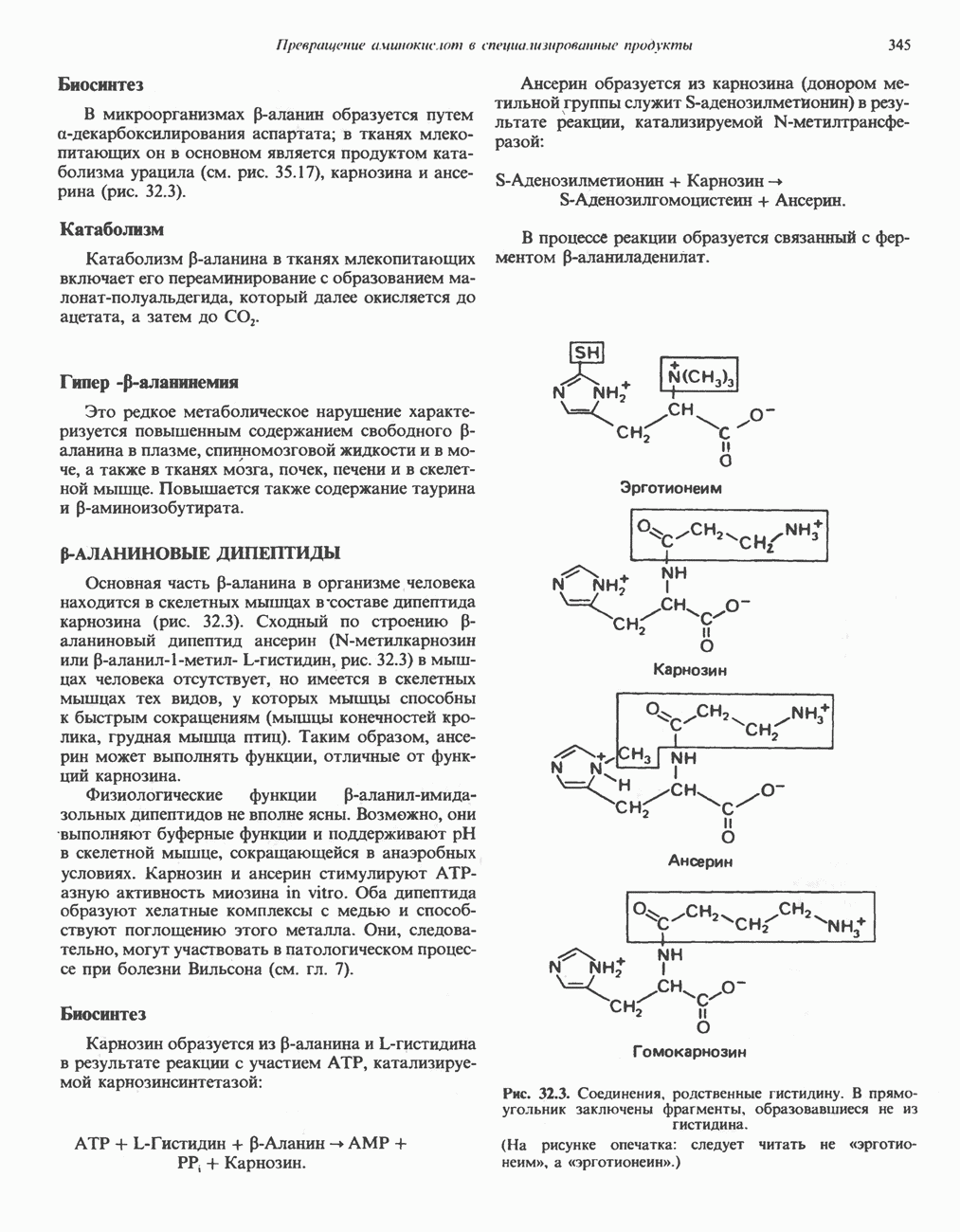

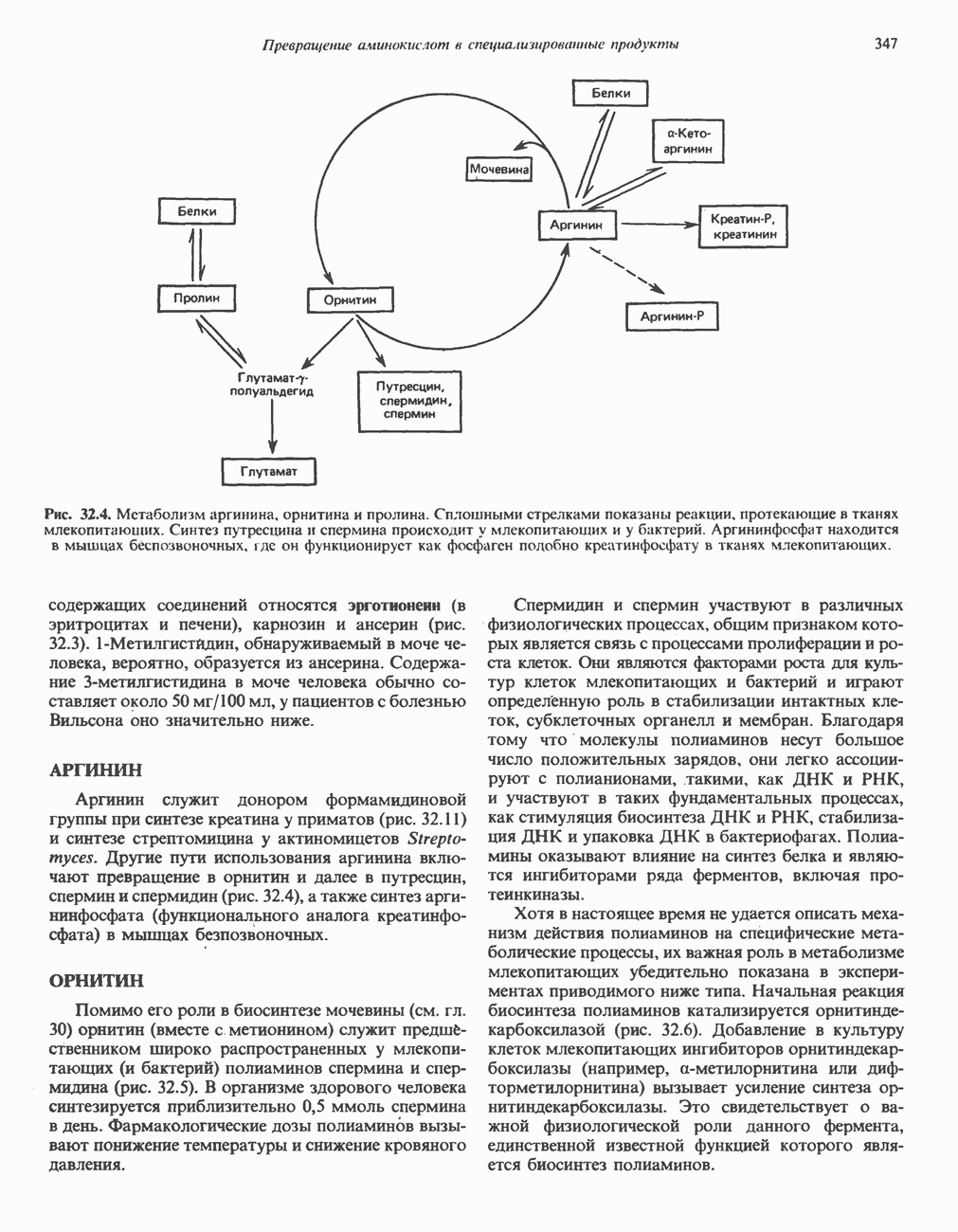

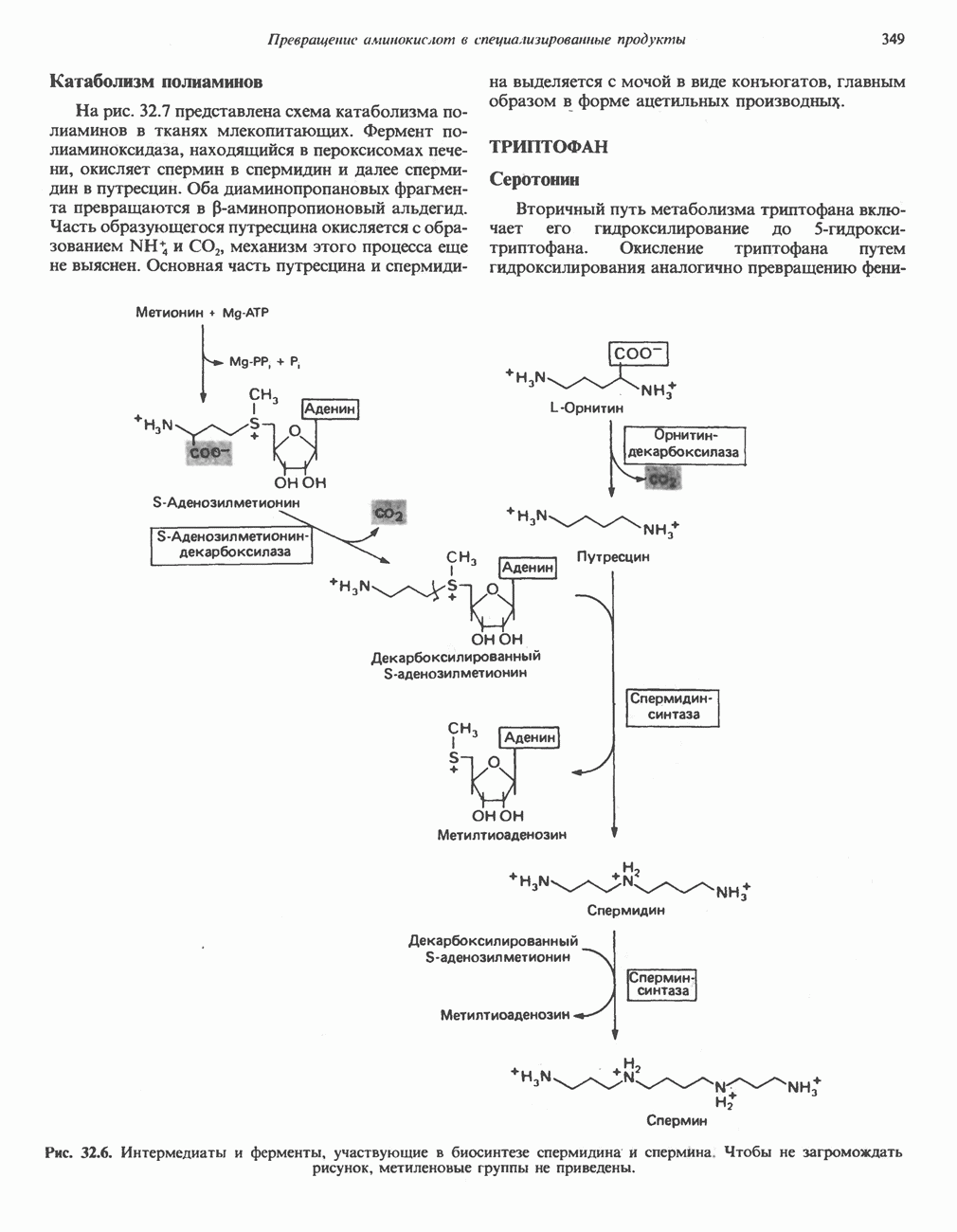

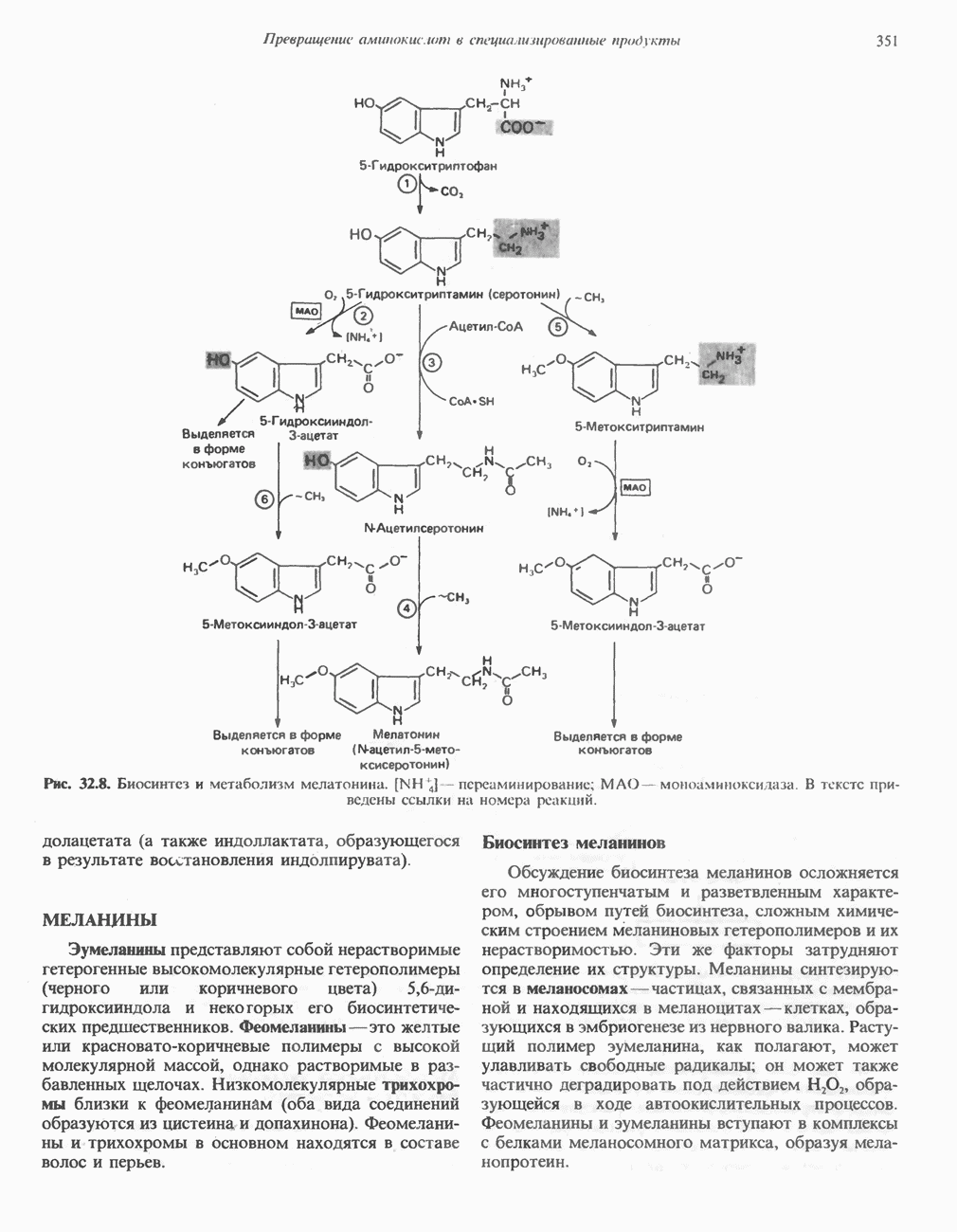














 ) (рис. 12.1).
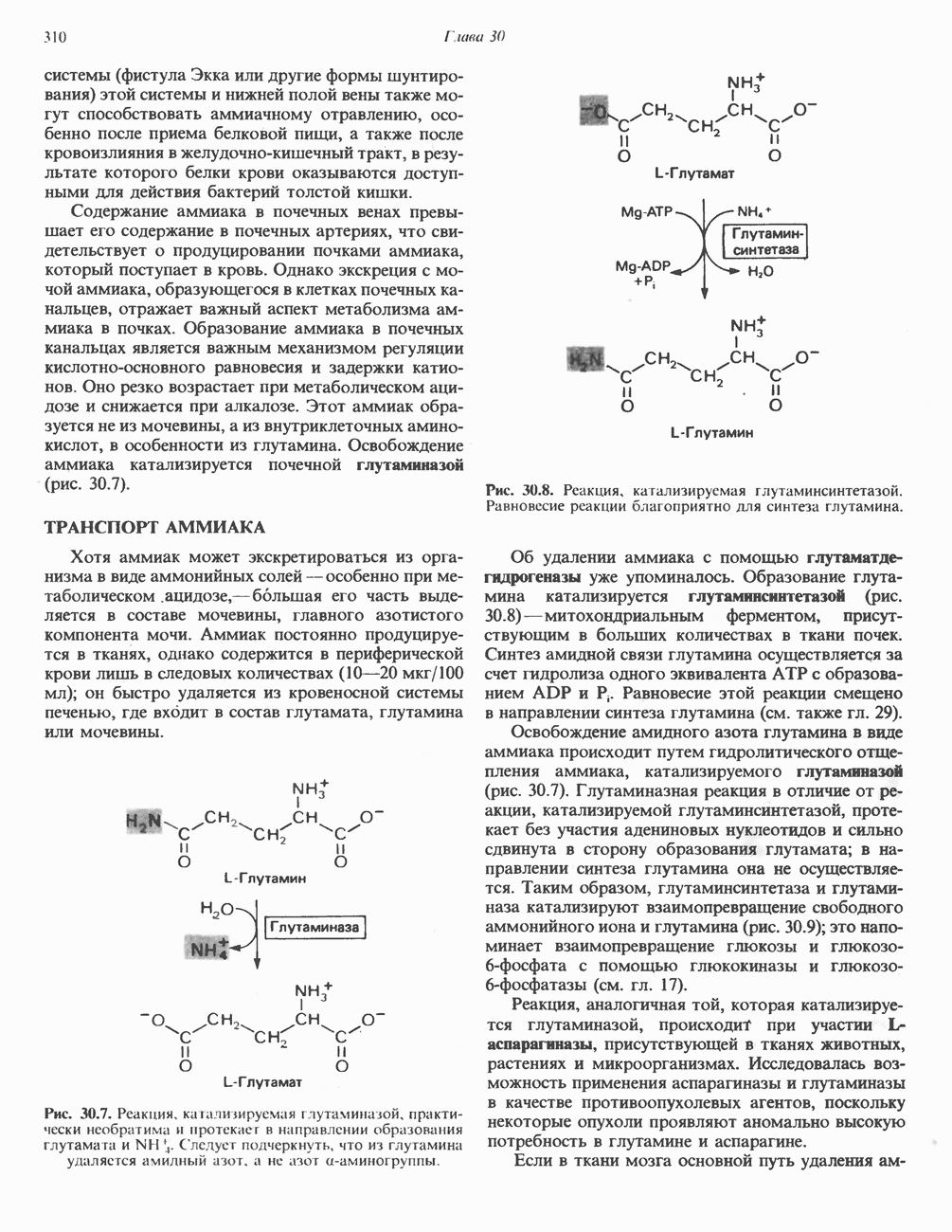
) (рис. 12.1).




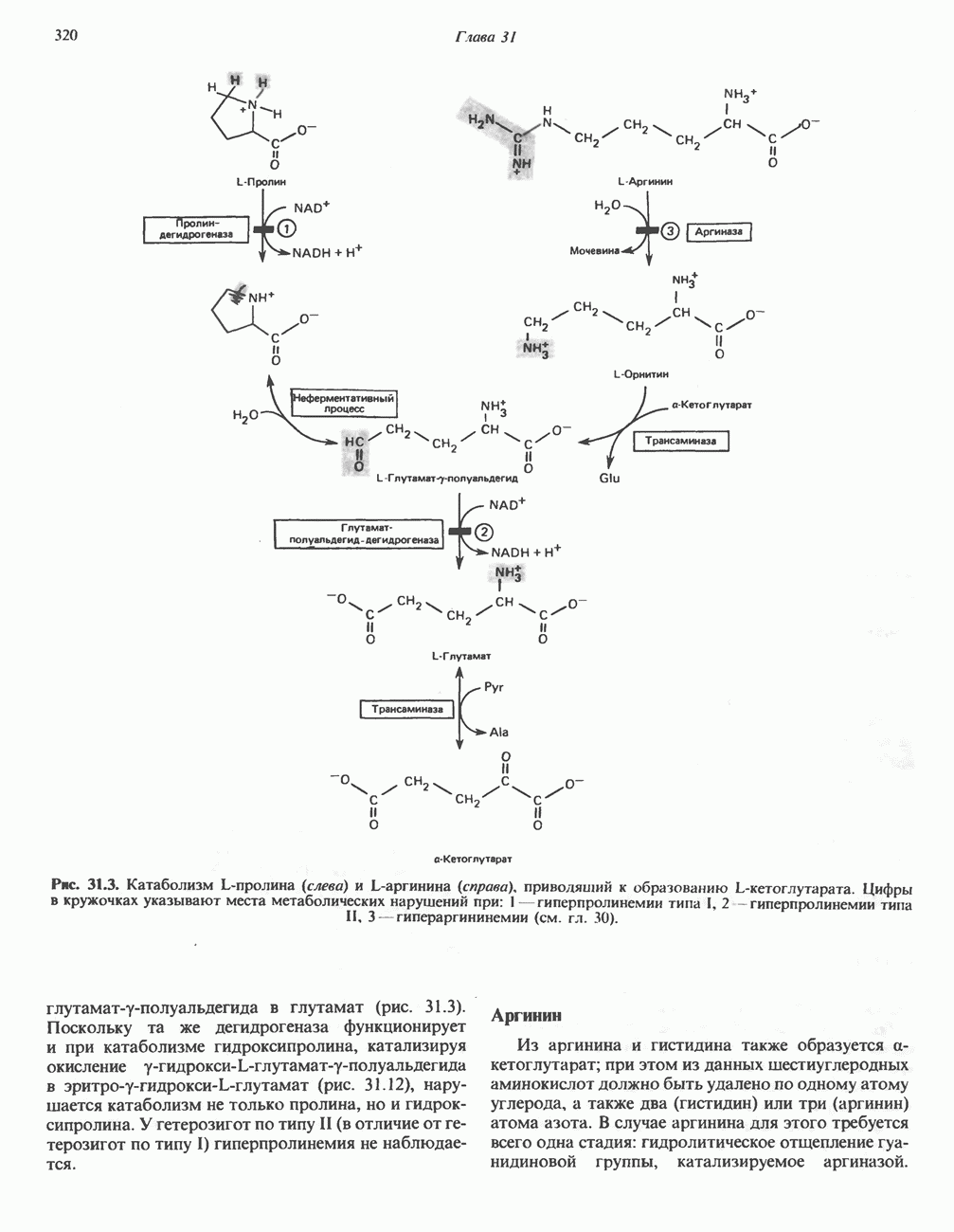
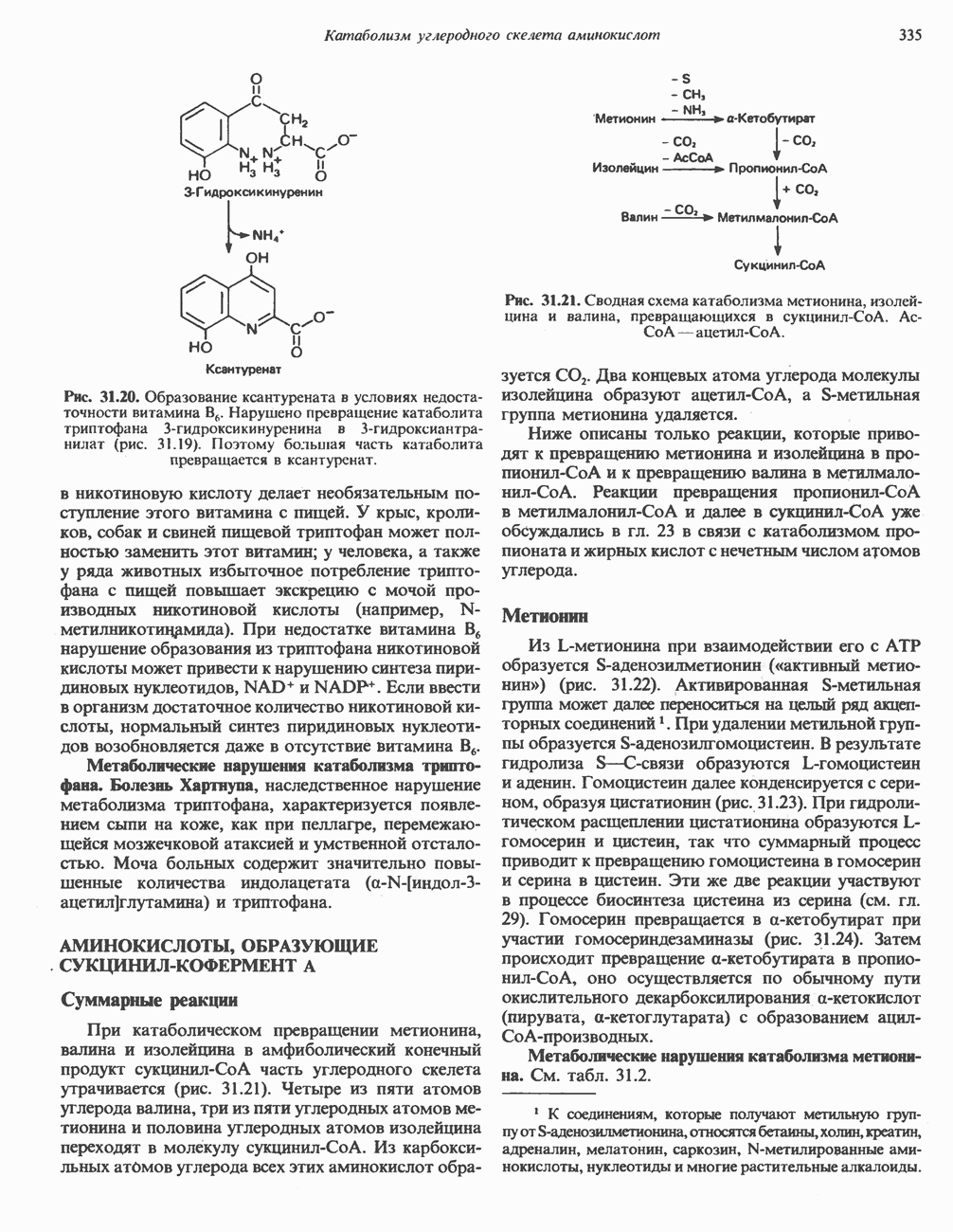
 Первоначально предполагали, что цитохром а и цитохром
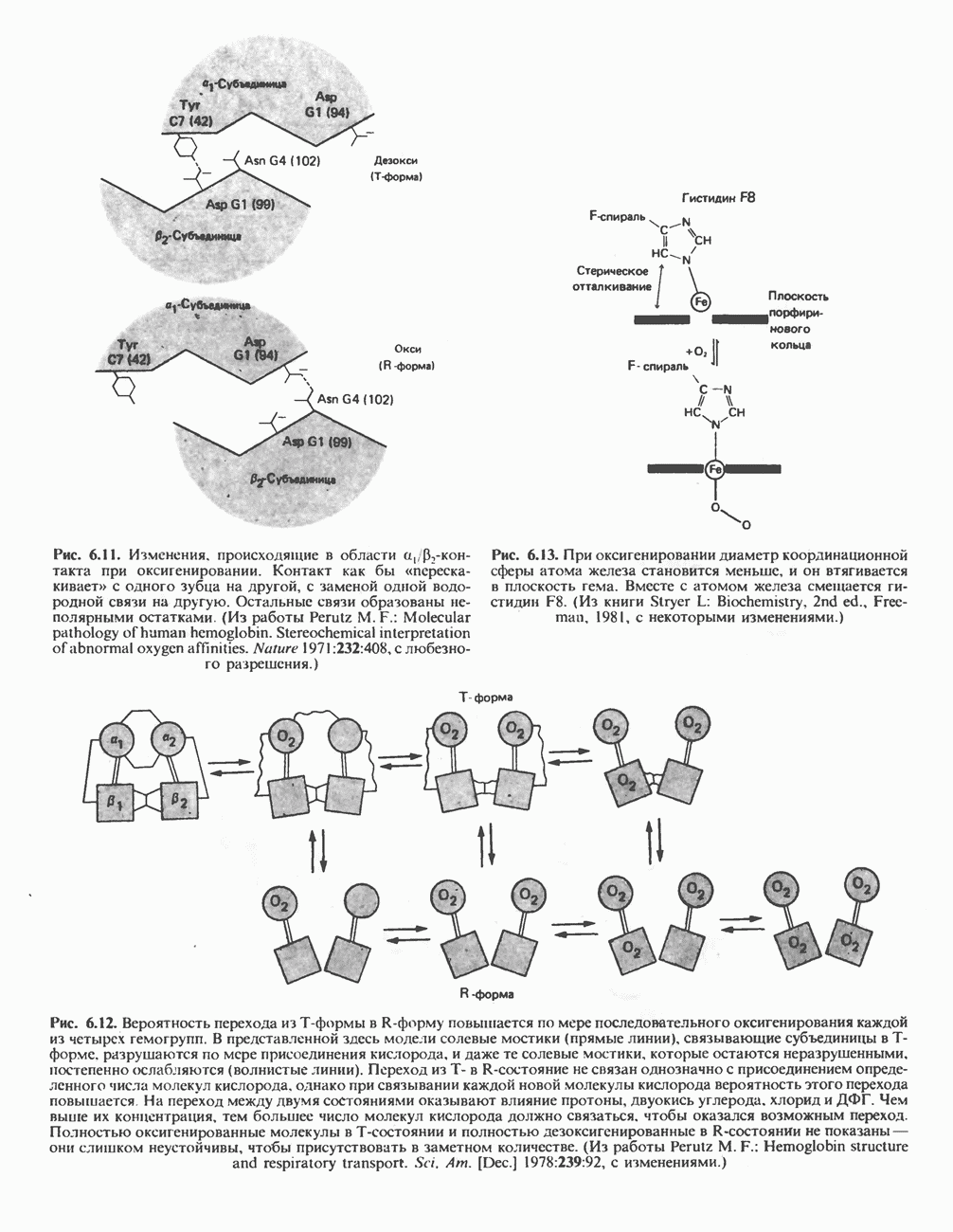
Первоначально предполагали, что цитохром а и цитохром  -это автономные гемопротеины, поскольку каждый из них характеризуется определенным спектром, кроме того, они проявляют разную чувствительность к действию окиси углерода и цианида. В дальнейшем же было показано, что эти два цитохрома входят в состав комплекса, который получил название цитохром
-это автономные гемопротеины, поскольку каждый из них характеризуется определенным спектром, кроме того, они проявляют разную чувствительность к действию окиси углерода и цианида. В дальнейшем же было показано, что эти два цитохрома входят в состав комплекса, который получил название цитохром  Он содержит две молекулы гема, в каждой из которых атом железа может переходить из состояния
Он содержит две молекулы гема, в каждой из которых атом железа может переходить из состояния  в состояние
в состояние  и обратно в ходе окисления и восстановления, а также два атома Си, каждый из которых взаимодействует с одним из гемов.
и обратно в ходе окисления и восстановления, а также два атома Си, каждый из которых взаимодействует с одним из гемов.
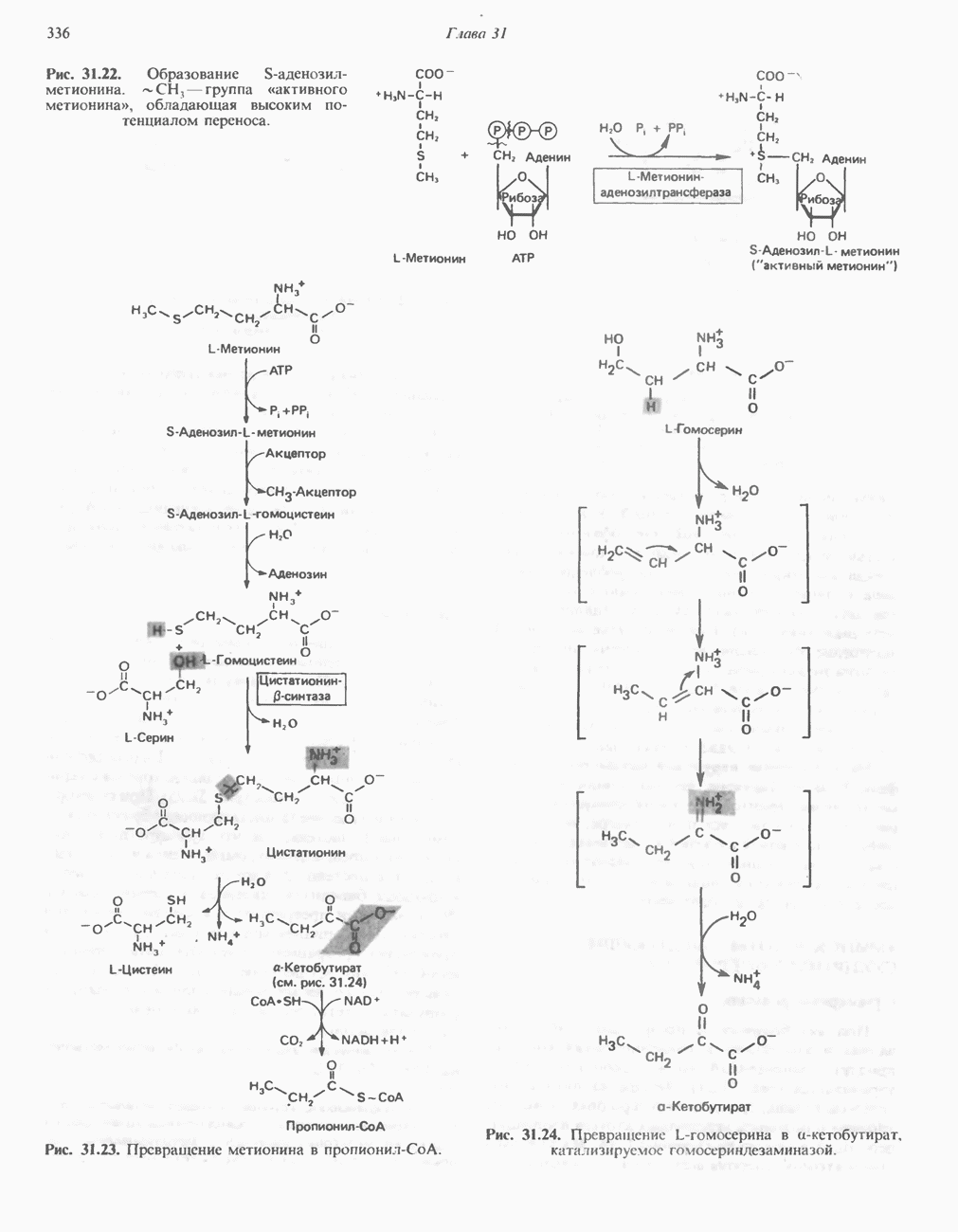
 витамина рибофлавина (рис. 12.5).
витамина рибофлавина (рис. 12.5).
 ) в тканях млекопитающих не синтезируется; он образуется в растениях и микроорганизмах и является, следовательно, незаменимым компонентом диеты. В состав молекулы рибофлавина входят сахарный спирт рибитол и гетероциклический флавин.
) в тканях млекопитающих не синтезируется; он образуется в растениях и микроорганизмах и является, следовательно, незаменимым компонентом диеты. В состав молекулы рибофлавина входят сахарный спирт рибитол и гетероциклический флавин.

 -зависимого фосфорилирования превращается в FMN (рис. 12.6); далее путем переноса АМР от другой молекулы АТР образуется FAD (рис. 12.7).
-зависимого фосфорилирования превращается в FMN (рис. 12.6); далее путем переноса АМР от другой молекулы АТР образуется FAD (рис. 12.7).
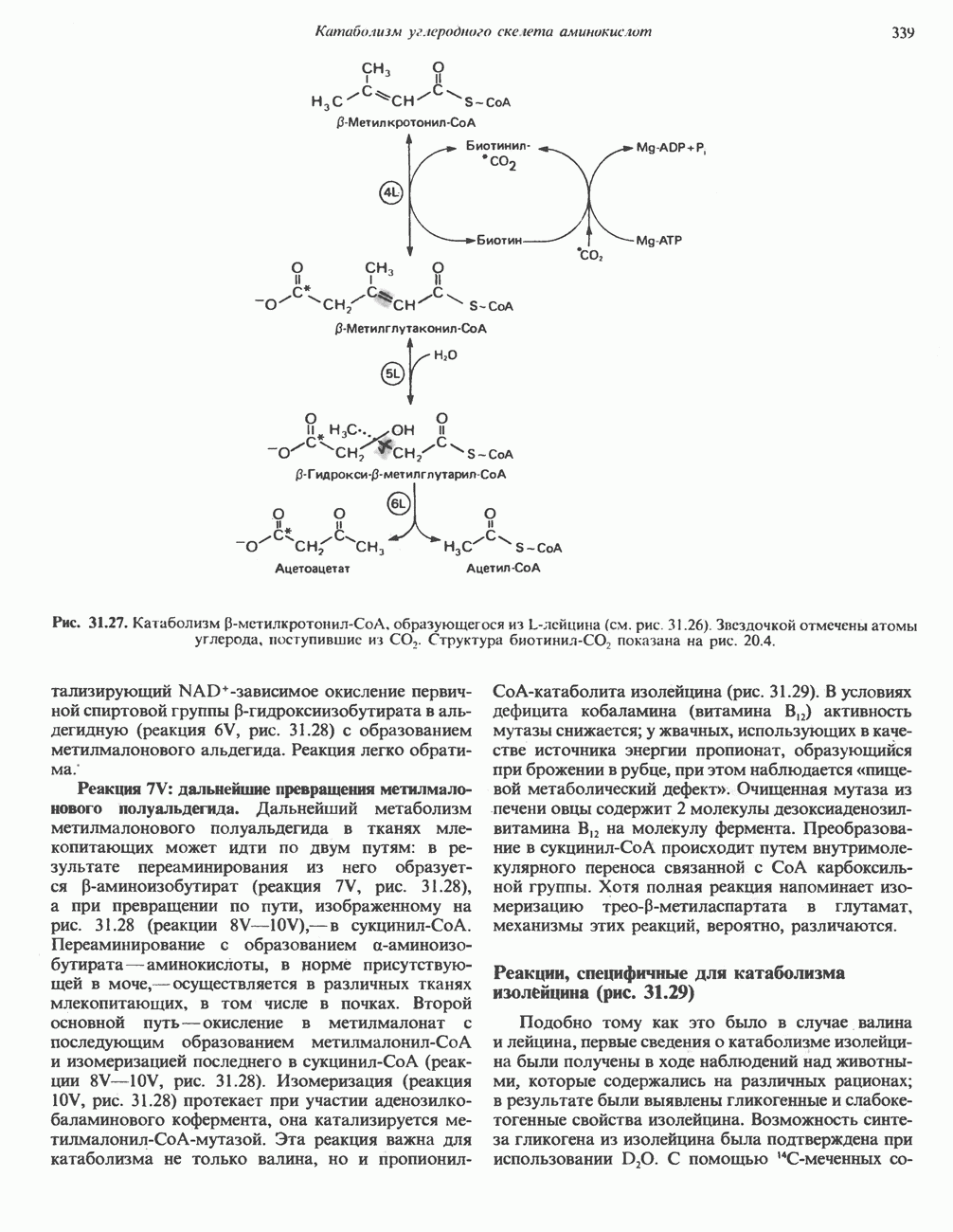
 -аминокислот (окси-даза
-аминокислот (окси-даза  -аминокислот) — находящийся в почках FMN - содержащий фермент с широкой специфичностью, катализирующий окислительное дезаминирование природных
-аминокислот) — находящийся в почках FMN - содержащий фермент с широкой специфичностью, катализирующий окислительное дезаминирование природных  -аминокислот. Широко распространена ксантиндегидрогеназа (ксантиноксидаза); она обнаружена в молоке, в тонком кишечнике, почках и печени. Ксантиндегидрогеназа содержит молибден; она играет важную роль в превращении пуриновых оснований в мочевую кислоту и особое значение имеет в печени и почках птиц, которые экскретируют мочевую кислоту как главный конечный азотсодержащий продукт метаболизма пуринов, а также катаболизма белков и аминокислот.
-аминокислот. Широко распространена ксантиндегидрогеназа (ксантиноксидаза); она обнаружена в молоке, в тонком кишечнике, почках и печени. Ксантиндегидрогеназа содержит молибден; она играет важную роль в превращении пуриновых оснований в мочевую кислоту и особое значение имеет в печени и почках птиц, которые экскретируют мочевую кислоту как главный конечный азотсодержащий продукт метаболизма пуринов, а также катаболизма белков и аминокислот.

 ).
).

 -гетероциклические субстраты.
-гетероциклические субстраты.
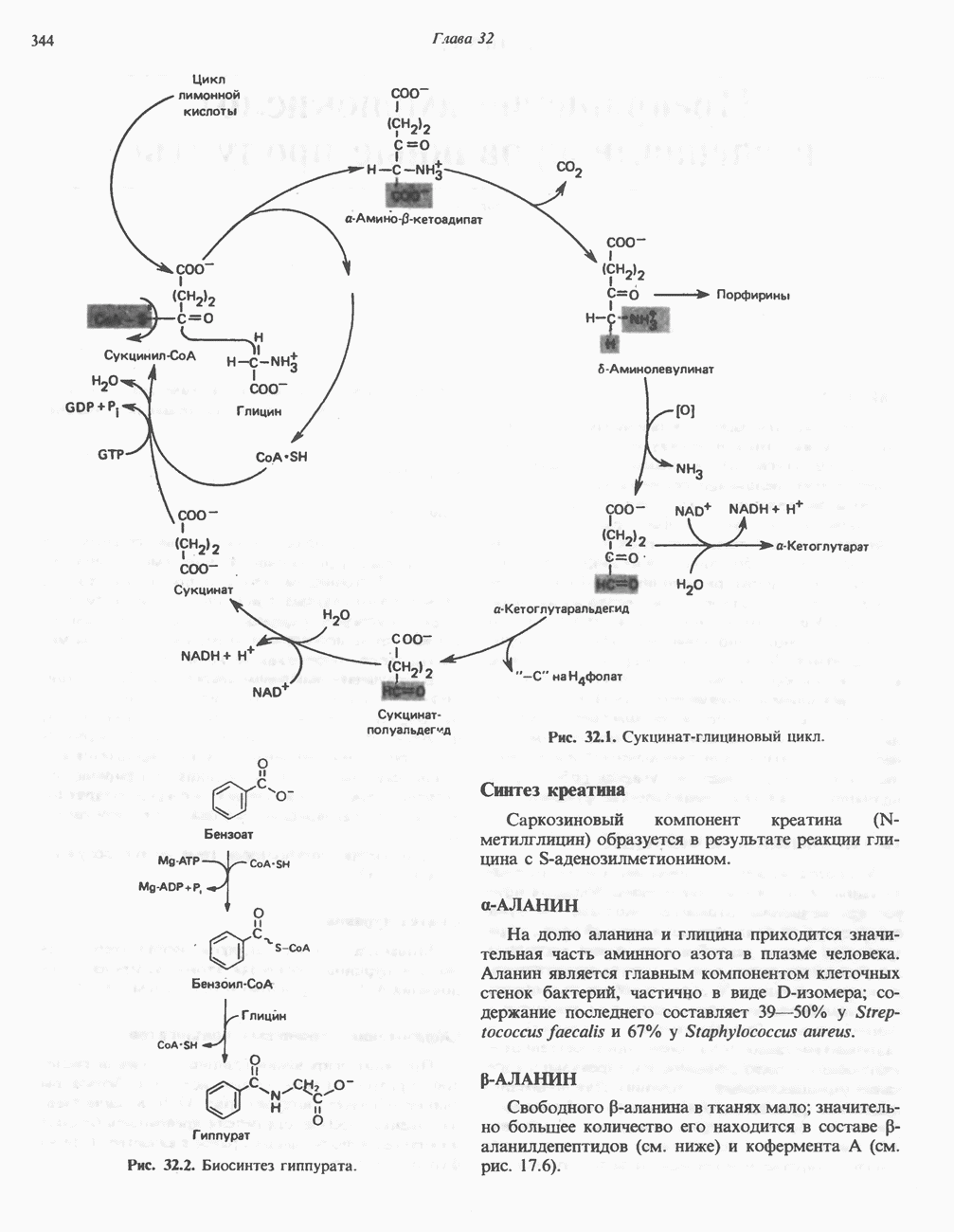
 либо к никотинамидадениндинуклеотидфосфату
либо к никотинамидадениндинуклеотидфосфату  выполняющим роль коферментов (рис. 12.9). Имеются, однако, дегидрогеназы, у которых коферментом может быть как
выполняющим роль коферментов (рис. 12.9). Имеются, однако, дегидрогеназы, у которых коферментом может быть как  так и
так и  образуются в организме человека из витамина ннацина. Ниацин включает никотиновую кислоту и ее амид (никотинамид) — каждое из этих соединений может, выполнять функции витамина в пищевом рационе. Для синтеза
образуются в организме человека из витамина ннацина. Ниацин включает никотиновую кислоту и ее амид (никотинамид) — каждое из этих соединений может, выполнять функции витамина в пищевом рационе. Для синтеза  или
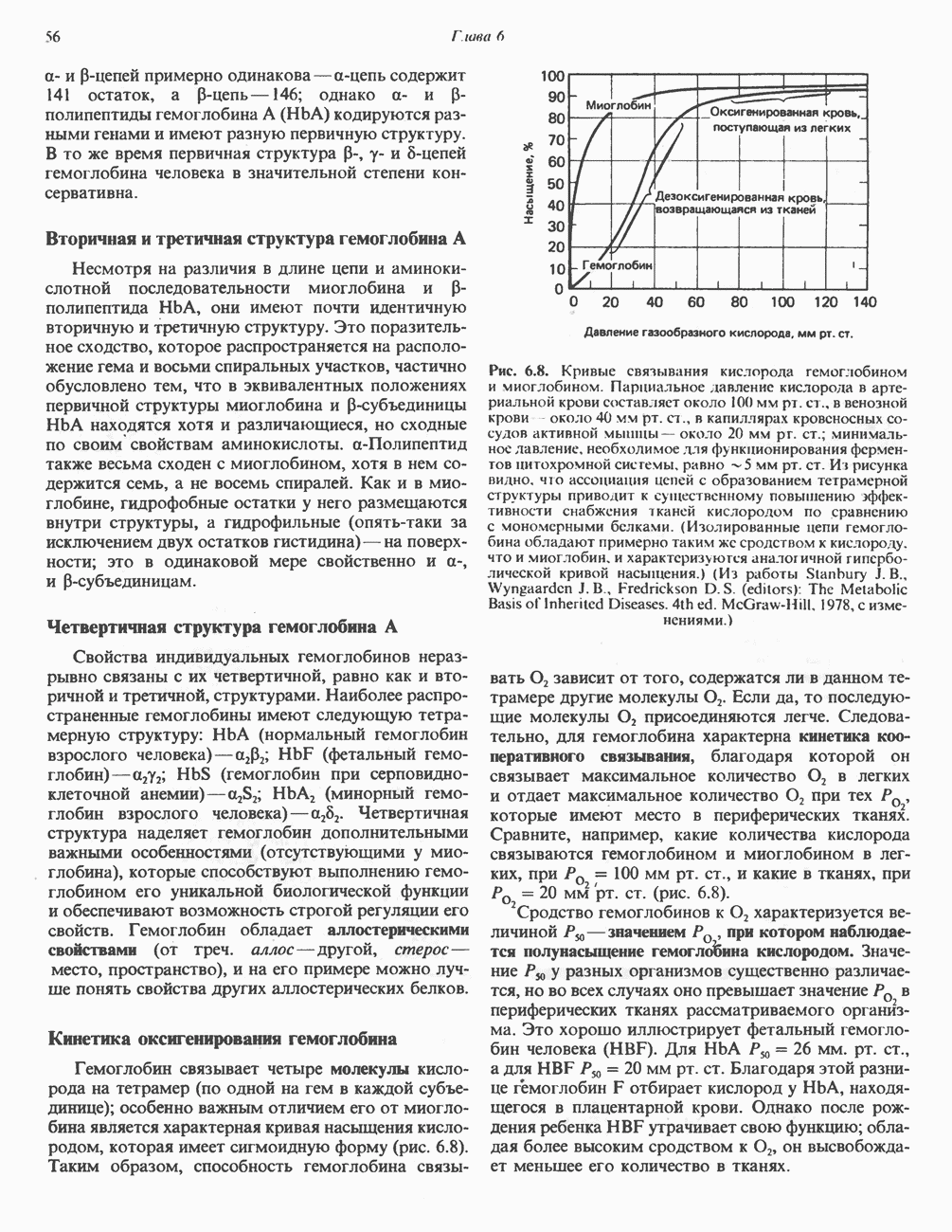
или  ферменты, находящиеся в цитозоле большинства клеток, используют только никотиновую кислоту, но не никотинамид. Никотинамидный фрагмент
ферменты, находящиеся в цитозоле большинства клеток, используют только никотиновую кислоту, но не никотинамид. Никотинамидный фрагмент  образуется из никотинатного фрагмента, когда последний находится в составе нуклеотида; амидная группа поступает из глутамина (рис. 12.9). Имеются данные о том, что в митохондриях для синтеза
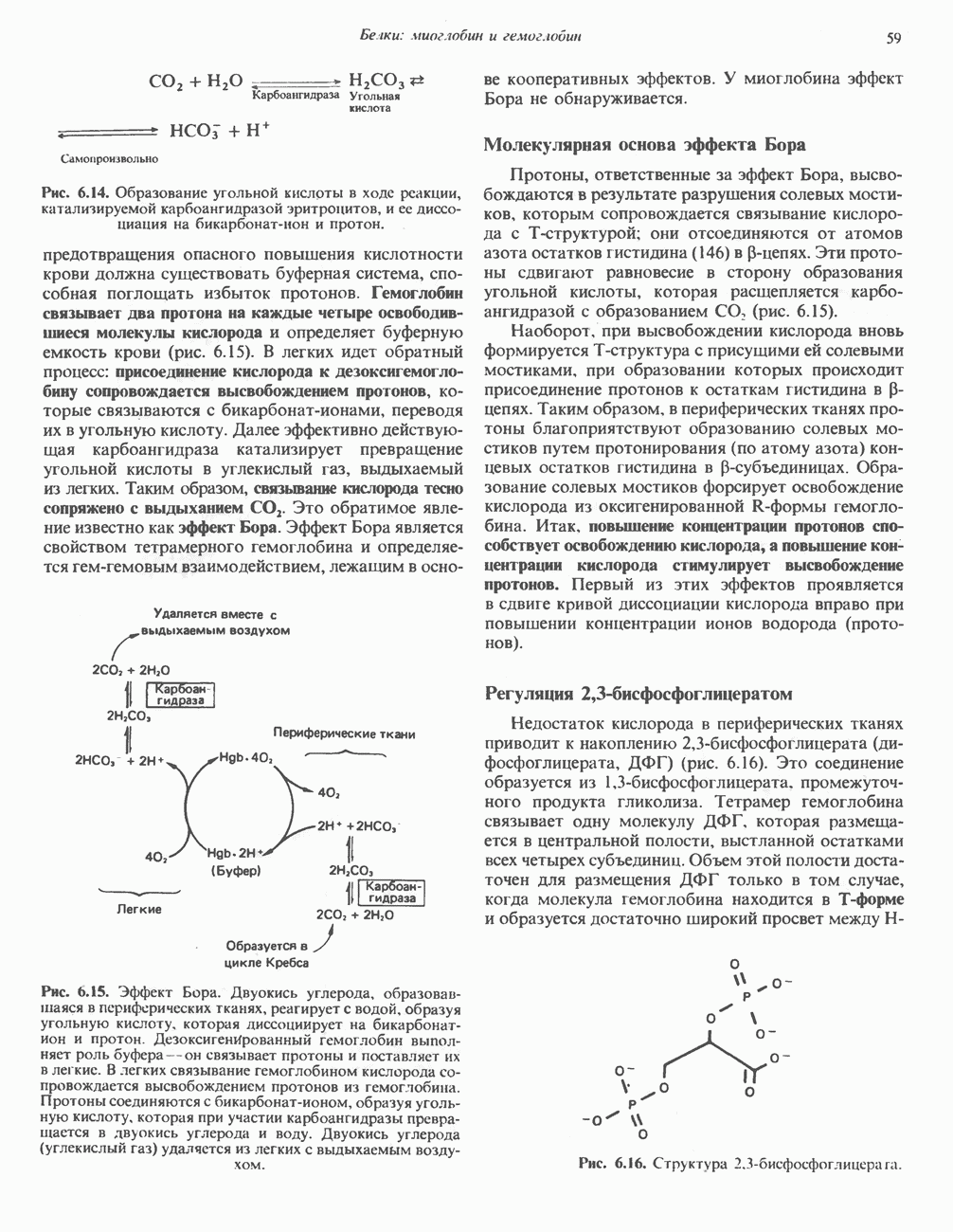
образуется из никотинатного фрагмента, когда последний находится в составе нуклеотида; амидная группа поступает из глутамина (рис. 12.9). Имеются данные о том, что в митохондриях для синтеза  используется никотинамид. Коферменты восстанавливаются специфическими субстратами дегидрогеназ и окисляются адекватным акцептором электронов (рис. 12.10). Коферменты могут свободно и обратимо диссоциировать из комплекса с соответствующими апоферментами.
используется никотинамид. Коферменты восстанавливаются специфическими субстратами дегидрогеназ и окисляются адекватным акцептором электронов (рис. 12.10). Коферменты могут свободно и обратимо диссоциировать из комплекса с соответствующими апоферментами.

 . У человека в отличие, например, от кошки потребность в ниацине может быть обеспечена за счет триптофана при достаточном количестве его в диете. В норме за счет триптофана синтезируется примерно две трети имеющегося в организме ниацина. PRPP-5-фосфорибозил-1-пирофосфат.
. У человека в отличие, например, от кошки потребность в ниацине может быть обеспечена за счет триптофана при достаточном количестве его в диете. В норме за счет триптофана синтезируется примерно две трети имеющегося в организме ниацина. PRPP-5-фосфорибозил-1-пирофосфат.

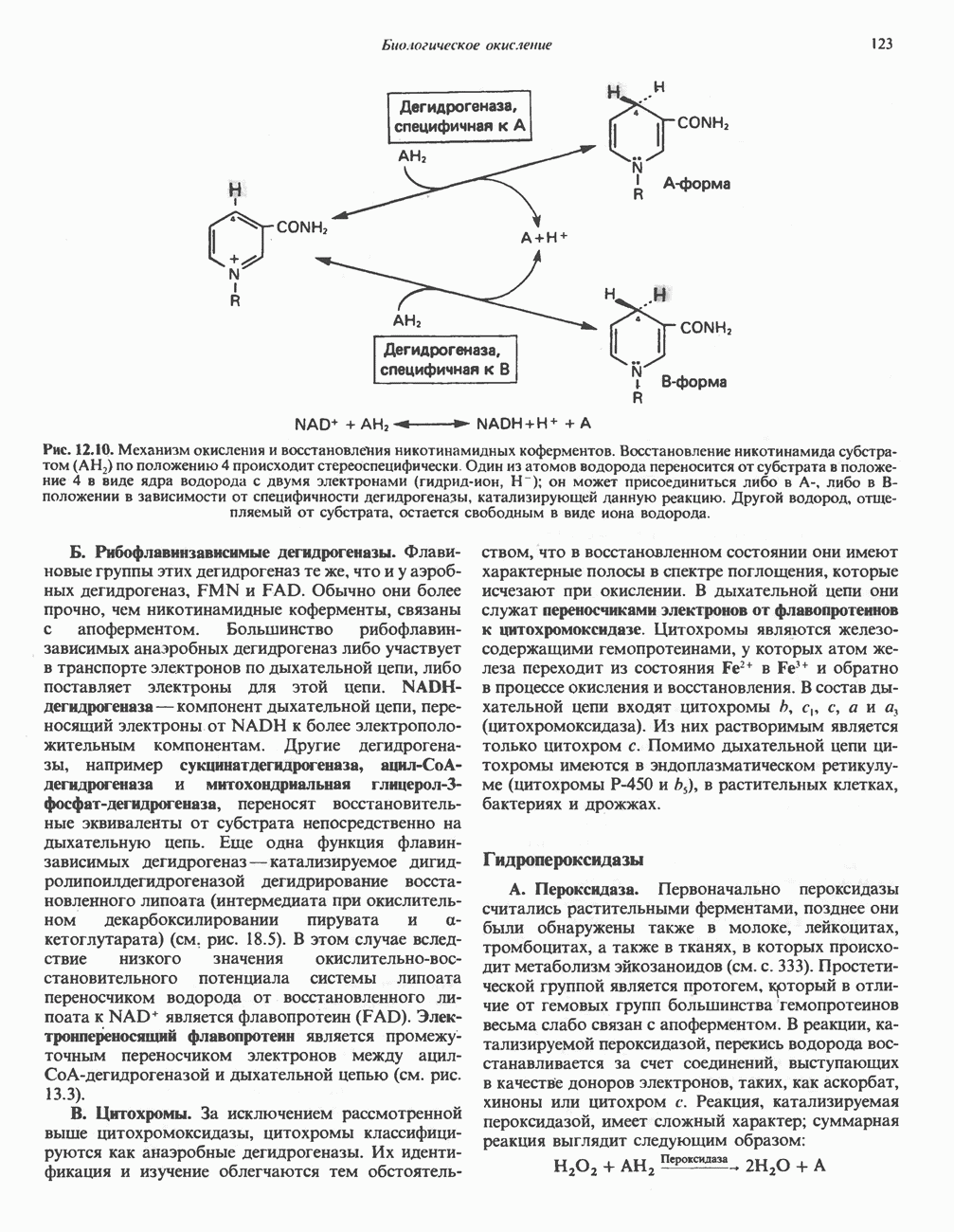
 по положению 4 происходит стереоспецифически. Один из атомов водорода переносится от субстрата в положение 4 в виде ядра водорода с двумя электронами (гидрид-ион,
по положению 4 происходит стереоспецифически. Один из атомов водорода переносится от субстрата в положение 4 в виде ядра водорода с двумя электронами (гидрид-ион,  ); он может присоединиться либо в
); он может присоединиться либо в  либо в В-положении в зависимости от специфичности дегидрогеназы, катализирующей данную реакцию. Другой водород, отщепляемый от субстрата, остается свободным в виде иона водорода.
либо в В-положении в зависимости от специфичности дегидрогеназы, катализирующей данную реакцию. Другой водород, отщепляемый от субстрата, остается свободным в виде иона водорода.
 -дегидрогеназа— компонент дыхательной цепи, переносящий электроны от NADH к более электроположительным компонентам. Другие дегидрогеназы, например сукцинатдегидрогеназа, ацил-СоА-дегидрогеназа и митохондриальная глицерол-3-фосфат-дегидрогеназа, переносят восстановительные эквиваленты от субстрата непосредственно на дыхательную цепь. Еще одна функция флавин-зависимых дегидрогеназ—катализируемое дигид-ролипоилдегидрогеназой дегидрирование восстановленного липоата (интермедиата при окислительном декарбоксилировании пирувата и а-кетоглутарата) (см. рис. 18.5). В этом случае вследствие низкого значения окислительно-восстановительного потенциала системы липоата переносчиком водорода от восстановленного липоата к
-дегидрогеназа— компонент дыхательной цепи, переносящий электроны от NADH к более электроположительным компонентам. Другие дегидрогеназы, например сукцинатдегидрогеназа, ацил-СоА-дегидрогеназа и митохондриальная глицерол-3-фосфат-дегидрогеназа, переносят восстановительные эквиваленты от субстрата непосредственно на дыхательную цепь. Еще одна функция флавин-зависимых дегидрогеназ—катализируемое дигид-ролипоилдегидрогеназой дегидрирование восстановленного липоата (интермедиата при окислительном декарбоксилировании пирувата и а-кетоглутарата) (см. рис. 18.5). В этом случае вследствие низкого значения окислительно-восстановительного потенциала системы липоата переносчиком водорода от восстановленного липоата к  является флавопротеин (FAD). Элек-тронпереносящнн флавопротеин является промежуточным переносчиком электронов между ацил-СоА-дегидрогеназой и дыхательной цепью (см. рис. 13.3).
является флавопротеин (FAD). Элек-тронпереносящнн флавопротеин является промежуточным переносчиком электронов между ацил-СоА-дегидрогеназой и дыхательной цепью (см. рис. 13.3).
 и обратно в процессе окисления и восстановления. В состав дыхательной цепи входят цитохромы
и обратно в процессе окисления и восстановления. В состав дыхательной цепи входят цитохромы  (цитохромоксидаза). Из них растворимым является только цитохром с. Помимо дыхательной цепи цитохромы имеются в эндоплазматическом ретикулуме
(цитохромоксидаза). Из них растворимым является только цитохром с. Помимо дыхательной цепи цитохромы имеются в эндоплазматическом ретикулуме  в растительных клетках, бактериях и дрожжах.
в растительных клетках, бактериях и дрожжах.

 и гидроперекисей липидов восстановленным глутатионом и таким образом защищает липиды мембран и гемоглобин от окисления перекисями.
и гидроперекисей липидов восстановленным глутатионом и таким образом защищает липиды мембран и гемоглобин от окисления перекисями.
 в качестве донора электронов, а другую — как окислитель, т.е. акцептор электронов. In vivo в большинстве случаев каталаза разлагает пероксид водорода
в качестве донора электронов, а другую — как окислитель, т.е. акцептор электронов. In vivo в большинстве случаев каталаза разлагает пероксид водорода

 так и ферменты, разлагающие ее (рис. 12.11). К ферментам, обеспечивающим образование
так и ферменты, разлагающие ее (рис. 12.11). К ферментам, обеспечивающим образование  помимо пероксисомных ферментов относятся также митохондриальные и микросомные системы транспорта электронов.
помимо пероксисомных ферментов относятся также митохондриальные и микросомные системы транспорта электронов.


 -гидрокснантранилдт-дно-кснгеназа из супернатантной фракции гомогената печени, а также гемсодержащие ферменты, в частности
-гидрокснантранилдт-дно-кснгеназа из супернатантной фракции гомогената печени, а также гемсодержащие ферменты, в частности  -триптофандиоксигеназа (триптофанпирролаза) из печени.
-триптофандиоксигеназа (триптофанпирролаза) из печени.

 Восстановителями этих цитохромов являются NADH и NADPH (рис. 12.12); цитохромы окисляются субстратами в результате серии ферментативных реакций, составляющих так называемый гидроксилазный цикл (рис. 12.13):
Восстановителями этих цитохромов являются NADH и NADPH (рис. 12.12); цитохромы окисляются субстратами в результате серии ферментативных реакций, составляющих так называемый гидроксилазный цикл (рис. 12.13):


 при отщеплении боковой цепи и по положениям
при отщеплении боковой цепи и по положениям  и
и  ). Ферменты почечной системы катализируют гидроксилирование
). Ферменты почечной системы катализируют гидроксилирование  -гидроксихолекальциферола по
-гидроксихолекальциферола по

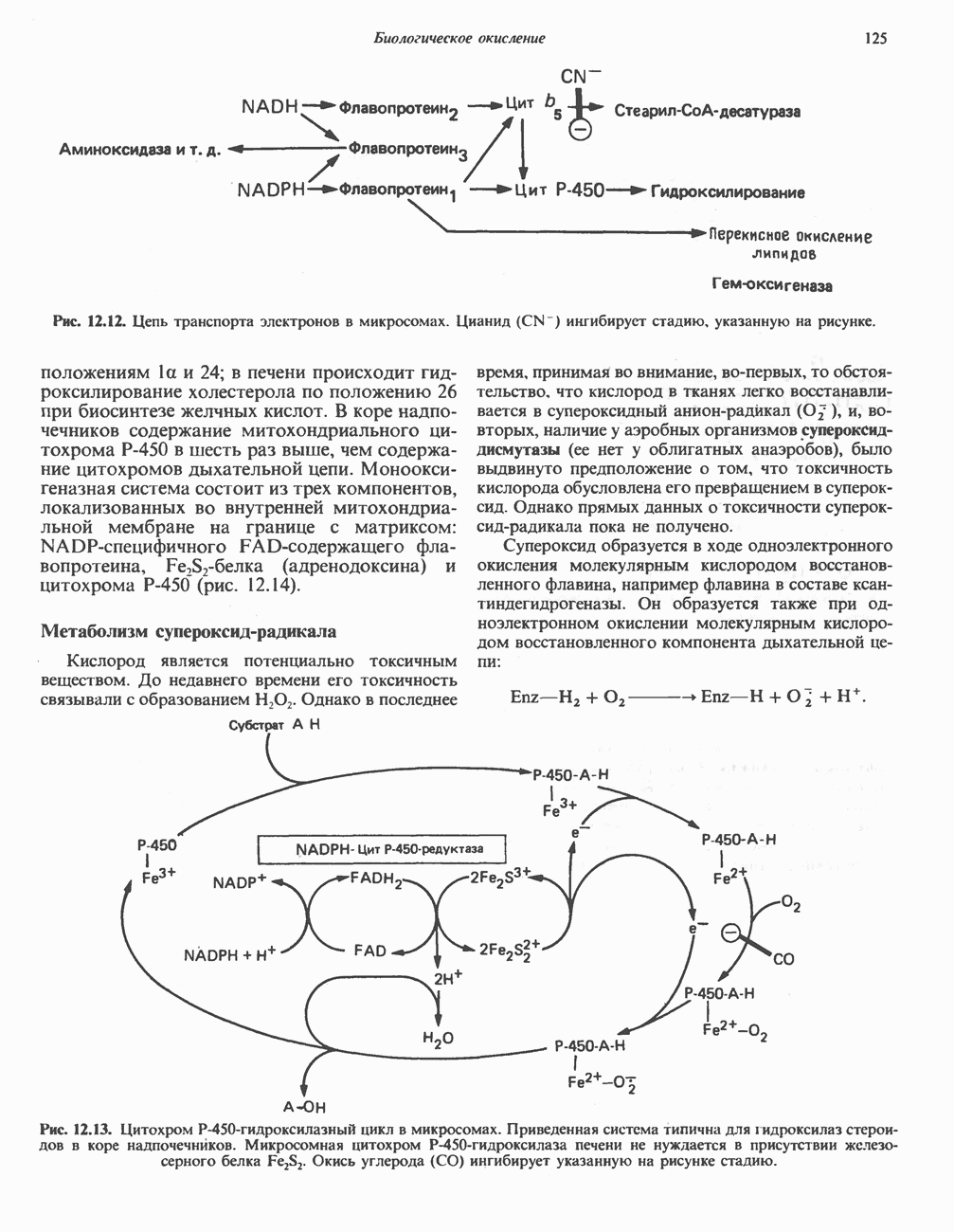
 ингибирует стадию, указанную на рисунке.
ингибирует стадию, указанную на рисунке.
 и
и  ; в печени происходит гидроксилирование холестерола по положению 26 при биосинтезе желчных кислот. В коре надпочечников содержание митохондриального цитохрома Р-450 в шесть раз выше, чем содержание цитохромов дыхательной цепи. Монооксигеназная система состоит из трех компонентов, локализованных во внутренней митохондриальной мембране на границе с матриксом: NADP-специфичного FАD - содержащего флавопротеина,
; в печени происходит гидроксилирование холестерола по положению 26 при биосинтезе желчных кислот. В коре надпочечников содержание митохондриального цитохрома Р-450 в шесть раз выше, чем содержание цитохромов дыхательной цепи. Монооксигеназная система состоит из трех компонентов, локализованных во внутренней митохондриальной мембране на границе с матриксом: NADP-специфичного FАD - содержащего флавопротеина,  -белка (адренодоксина) и цитохрома Р-450 (рис. 12.14).
-белка (адренодоксина) и цитохрома Р-450 (рис. 12.14).
 Однако в последнее время, принимая во внимание, во-первых, то обстоятельство, что кислород в тканях легко восстанавливается в супероксидный анион-радикал
Однако в последнее время, принимая во внимание, во-первых, то обстоятельство, что кислород в тканях легко восстанавливается в супероксидный анион-радикал  , и, во-вторых, наличие у аэробных организмов супероксид-дисмутазы (ее нет у облигатных анаэробов), было выдвинуто предположение о том, что токсичность кислорода обусловлена его превращением в супероксид. Однако прямых данных о токсичности супероксидрадикала пока не получено.
, и, во-вторых, наличие у аэробных организмов супероксид-дисмутазы (ее нет у облигатных анаэробов), было выдвинуто предположение о том, что токсичность кислорода обусловлена его превращением в супероксид. Однако прямых данных о токсичности супероксидрадикала пока не получено.


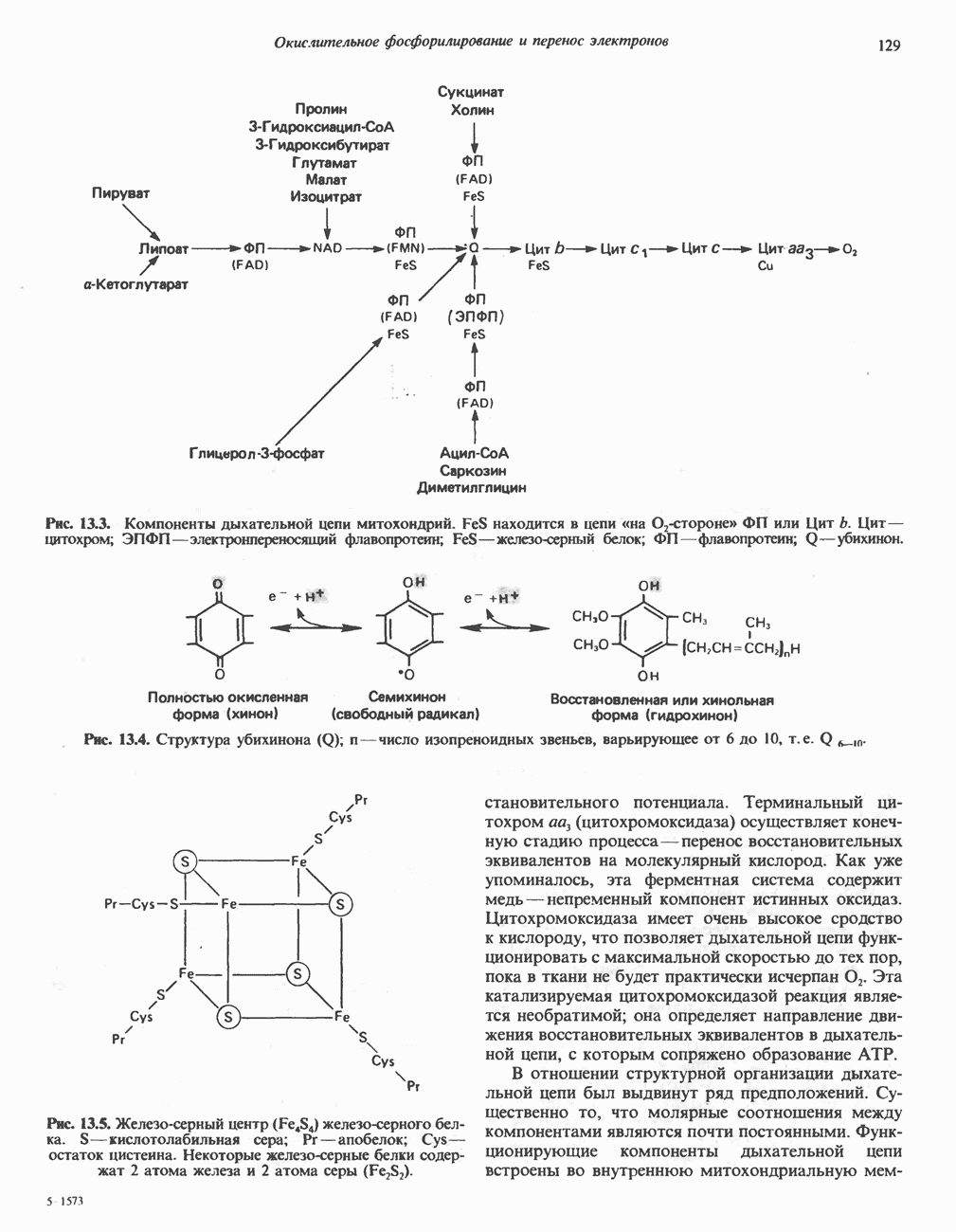
 Окись углерода (СО) ингибирует указанную на рисунке стадию.
Окись углерода (СО) ингибирует указанную на рисунке стадию.

 —железо-серный белок (адренодоксин). Поскольку NADP(H) не может проникать в митохондриальную мембрану, источниками восстановительных эквивалентов являются такие субстраты, как малат и изоцитрат, для которых внутри митохондрий имеются специфические NADP - зависимые дегидрогеназы.
—железо-серный белок (адренодоксин). Поскольку NADP(H) не может проникать в митохондриальную мембрану, источниками восстановительных эквивалентов являются такие субстраты, как малат и изоцитрат, для которых внутри митохондрий имеются специфические NADP - зависимые дегидрогеназы.
 :
:


 ; митохондриальный фермент, так же как и фермент, обнаруженный у бактерий, содержит ион Мп2+. Это обстоятельство служит еще одним свидетельством в пользу гипотезы о происхождении митохондрий из прокариот, вступивших в симбиоз с протоэукариотами. Дисмутаза присутствует во всех основных тканях аэробов. Пребывание животных в атмосфере 100%-ного кислорода вызывает адаптивное повышение содержания дисмутазы, особенно в легких; длительное пребывание в такой атмосфере приводит к повреждению легких и летальному исходу. Антиоксиданты, например а-токоферол (витамин Е), способны улавливать свободные радикалы, такие, как
; митохондриальный фермент, так же как и фермент, обнаруженный у бактерий, содержит ион Мп2+. Это обстоятельство служит еще одним свидетельством в пользу гипотезы о происхождении митохондрий из прокариот, вступивших в симбиоз с протоэукариотами. Дисмутаза присутствует во всех основных тканях аэробов. Пребывание животных в атмосфере 100%-ного кислорода вызывает адаптивное повышение содержания дисмутазы, особенно в легких; длительное пребывание в такой атмосфере приводит к повреждению легких и летальному исходу. Антиоксиданты, например а-токоферол (витамин Е), способны улавливать свободные радикалы, такие, как  , снижая тем самым токсичность кислорода.
, снижая тем самым токсичность кислорода.