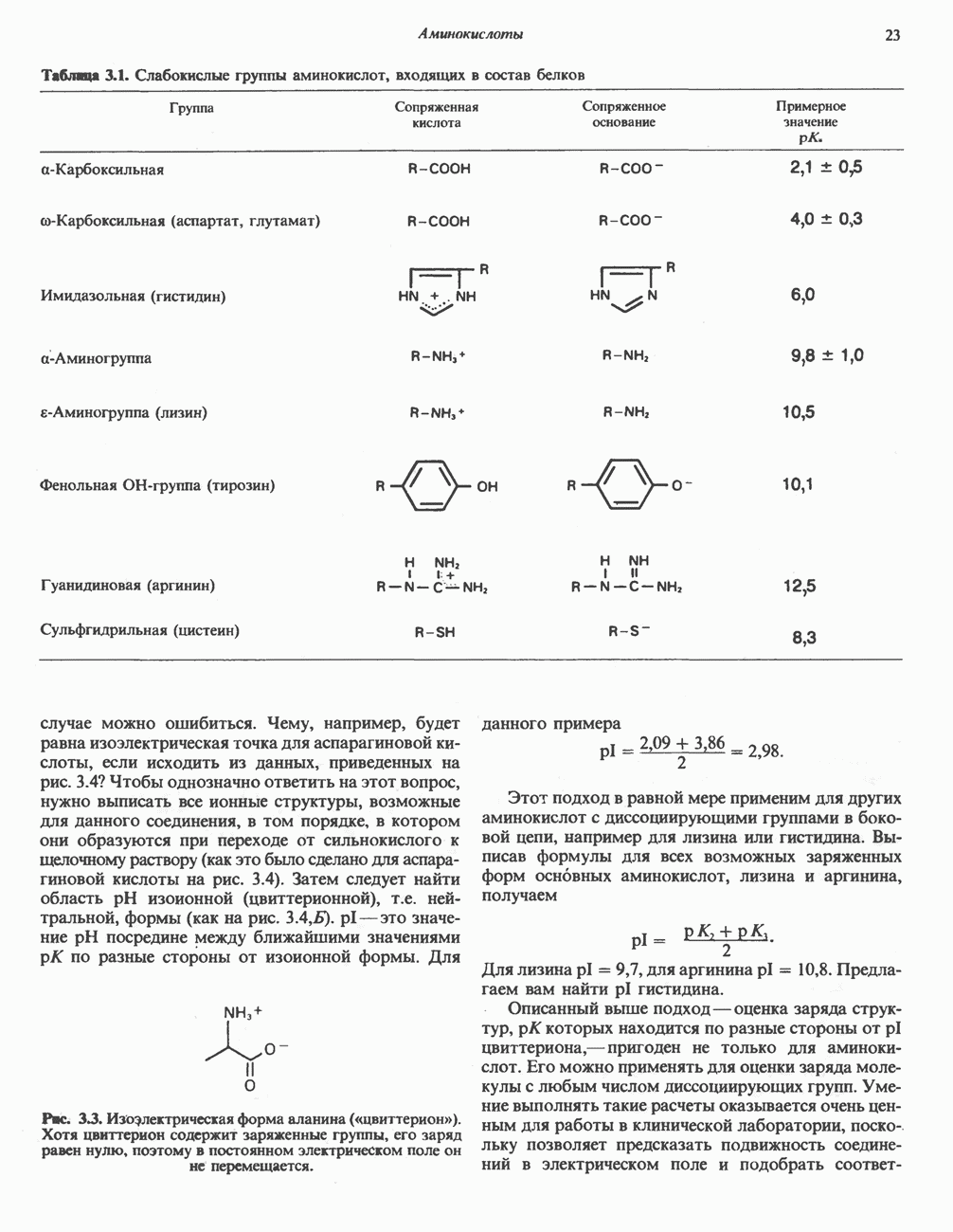
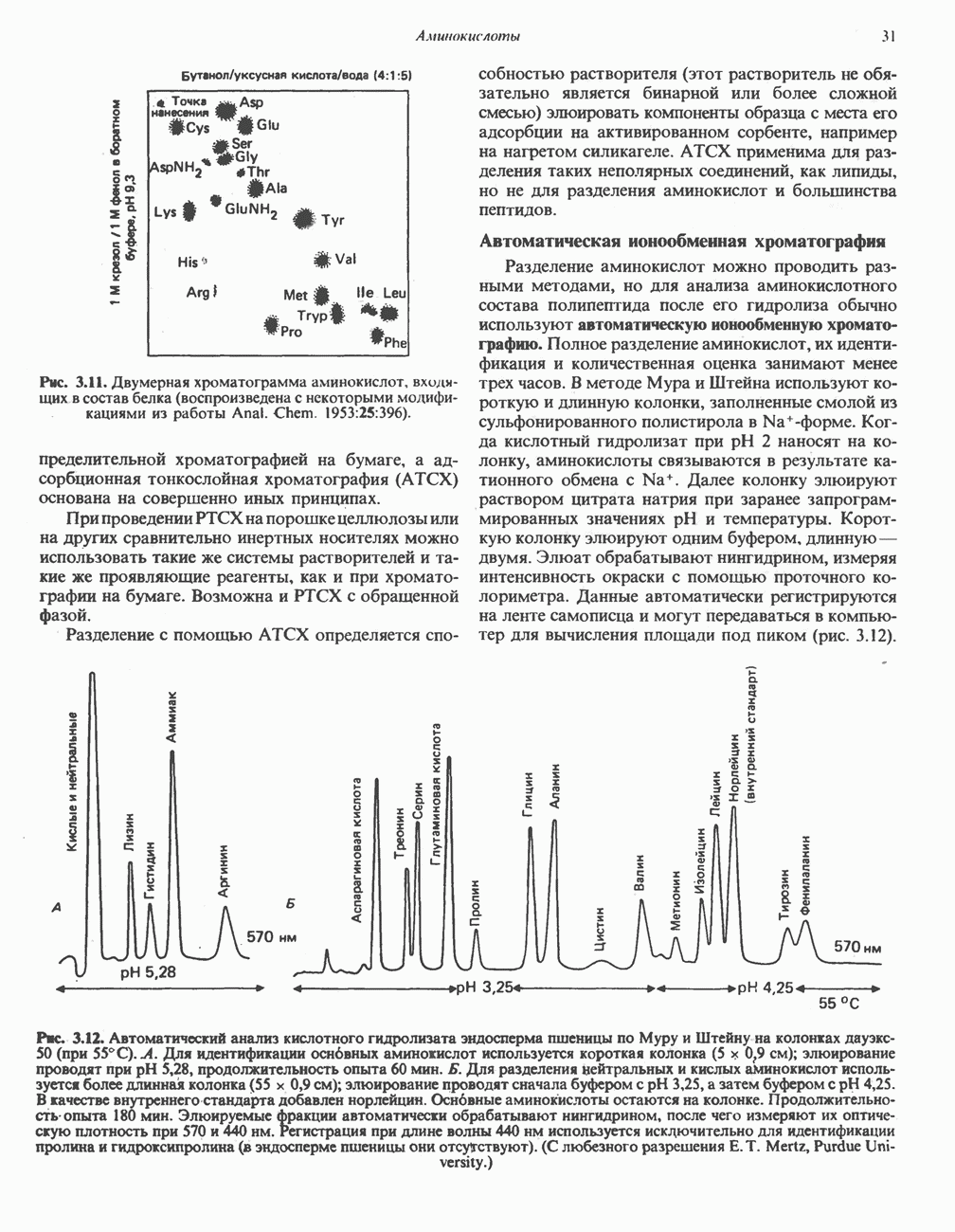
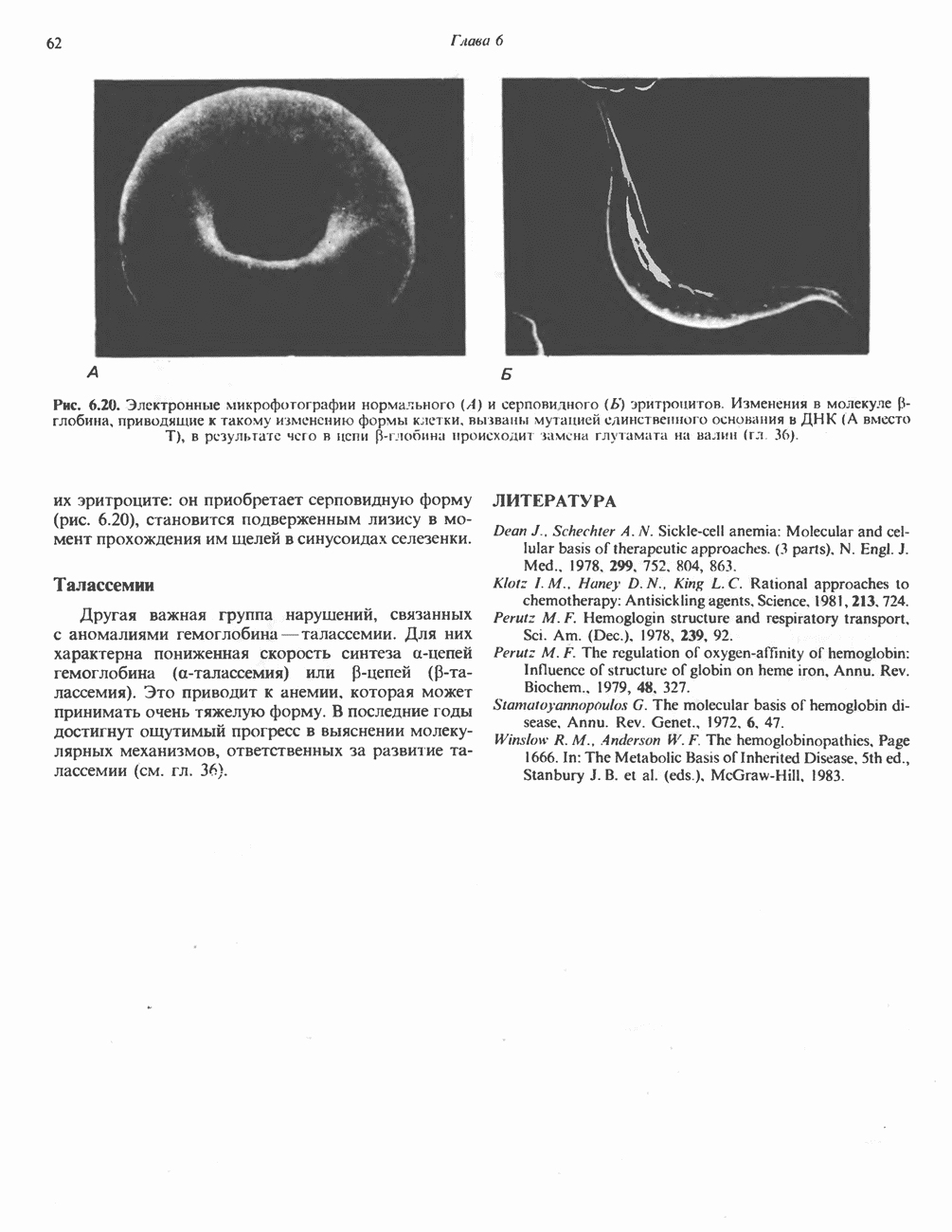
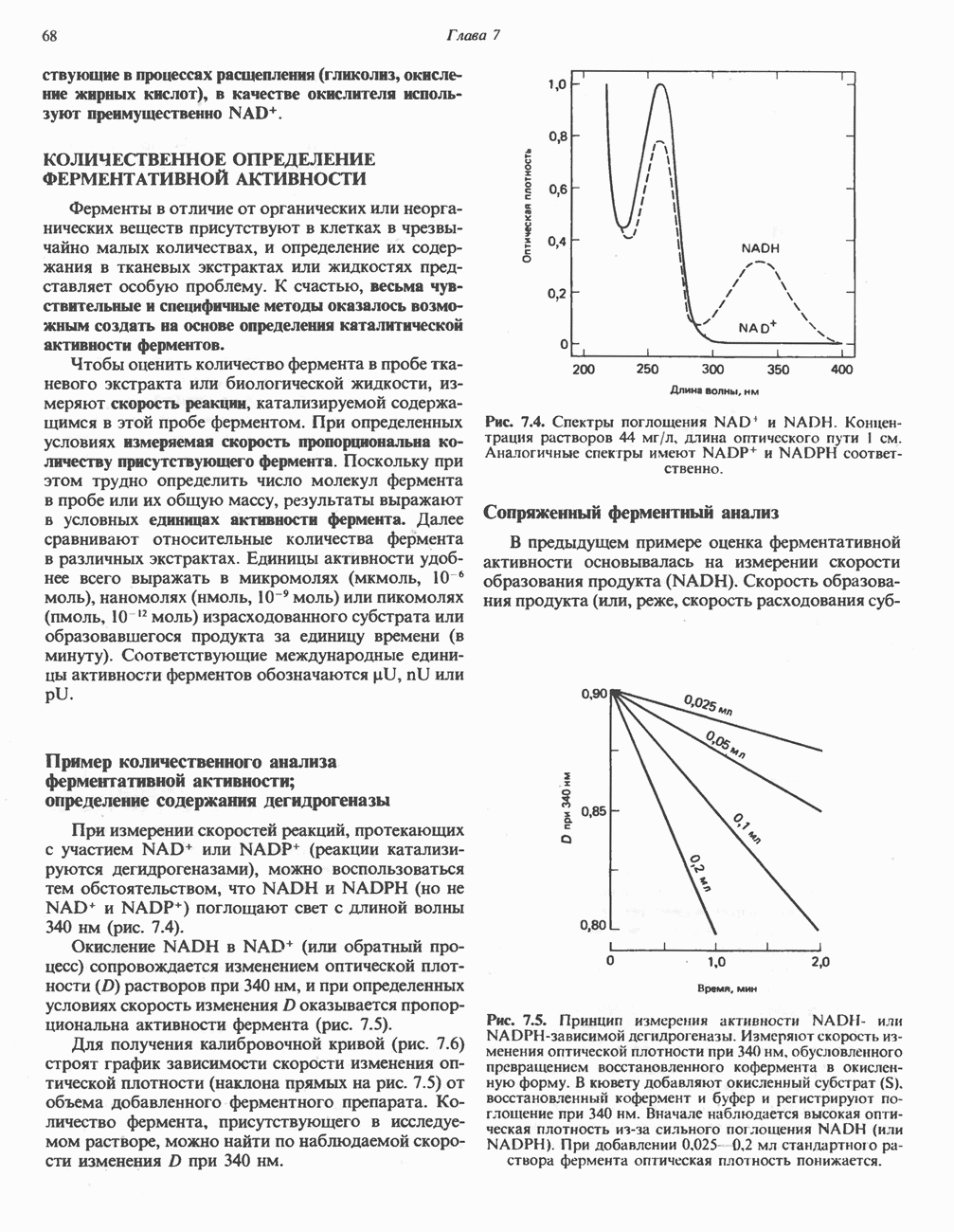
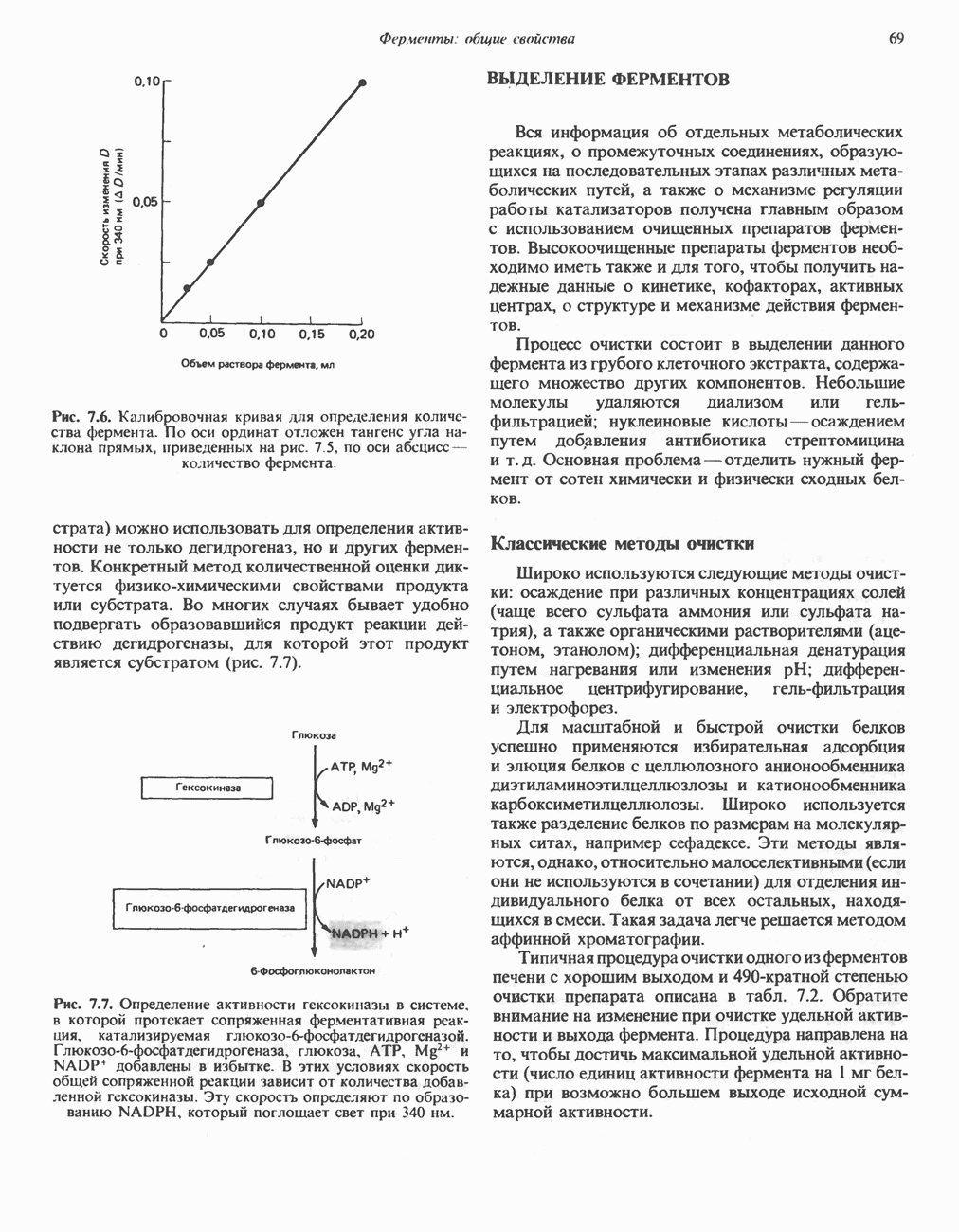
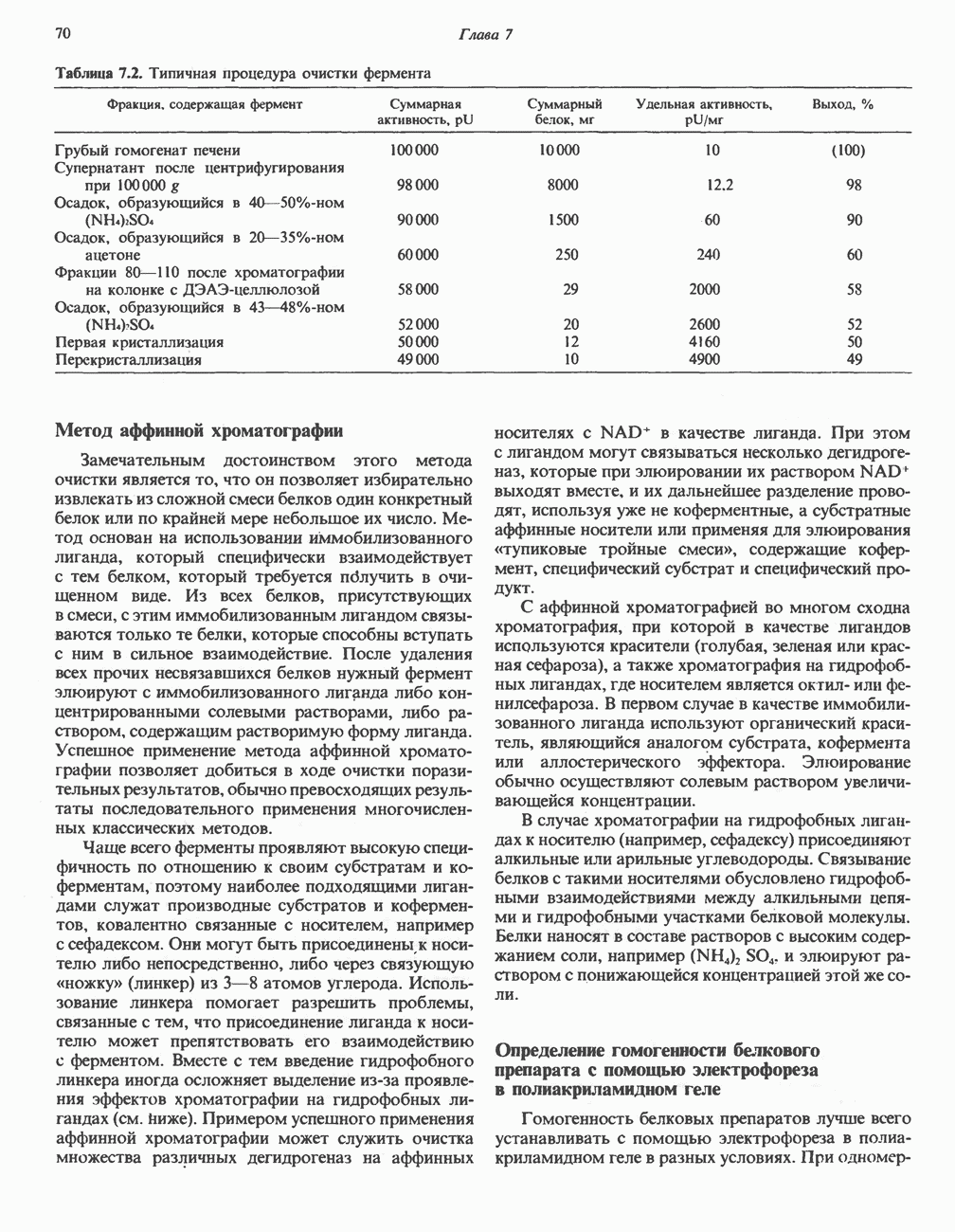
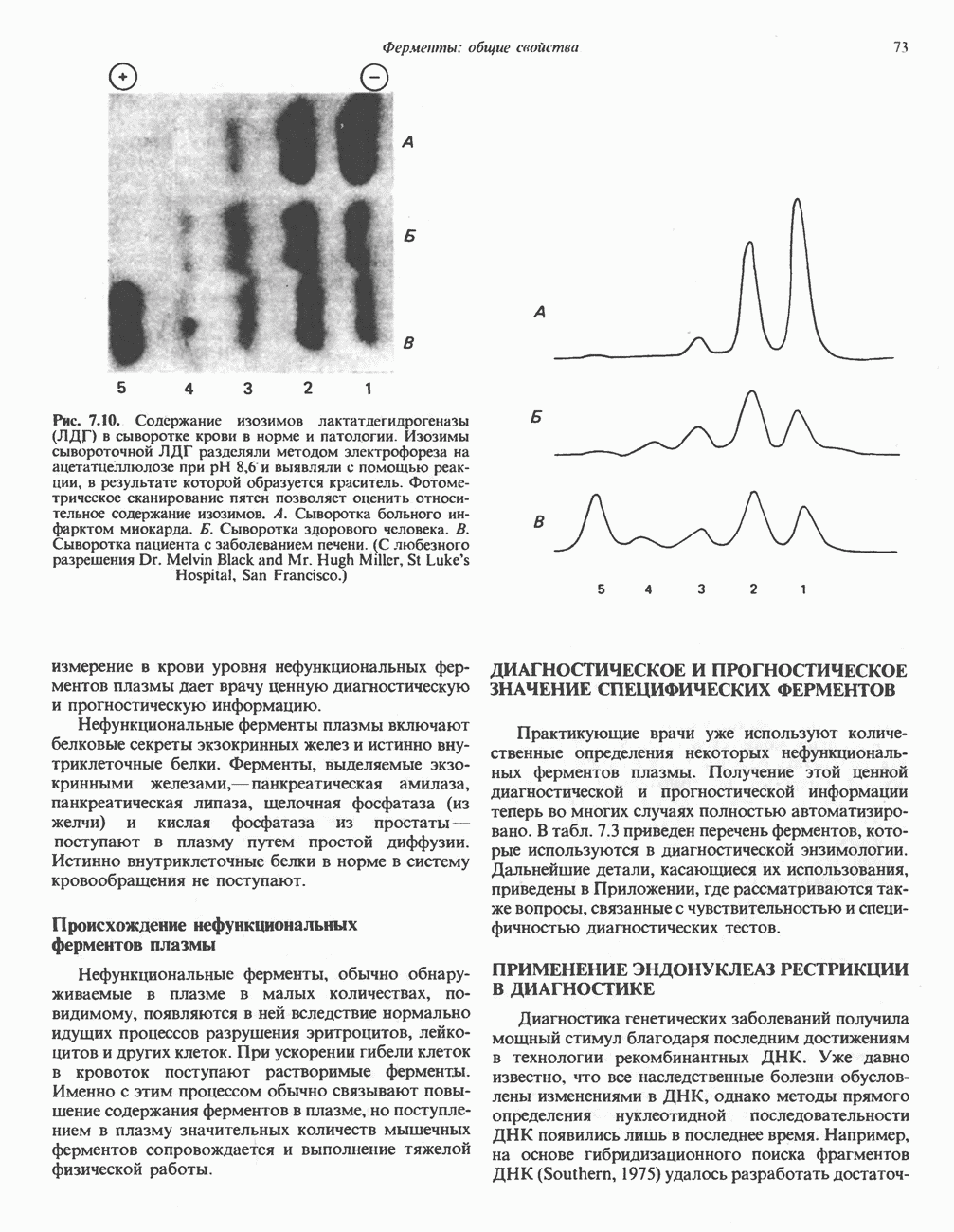
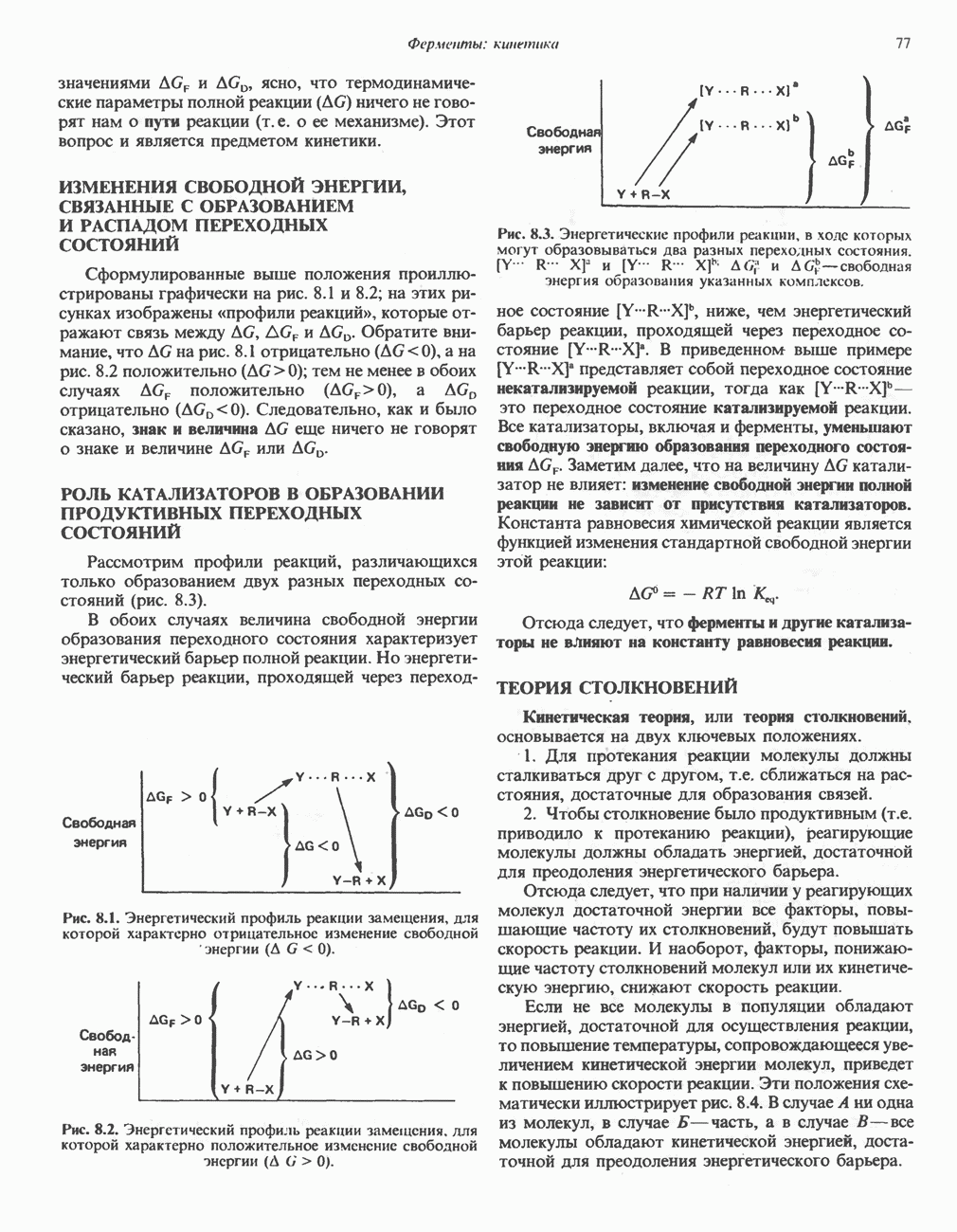
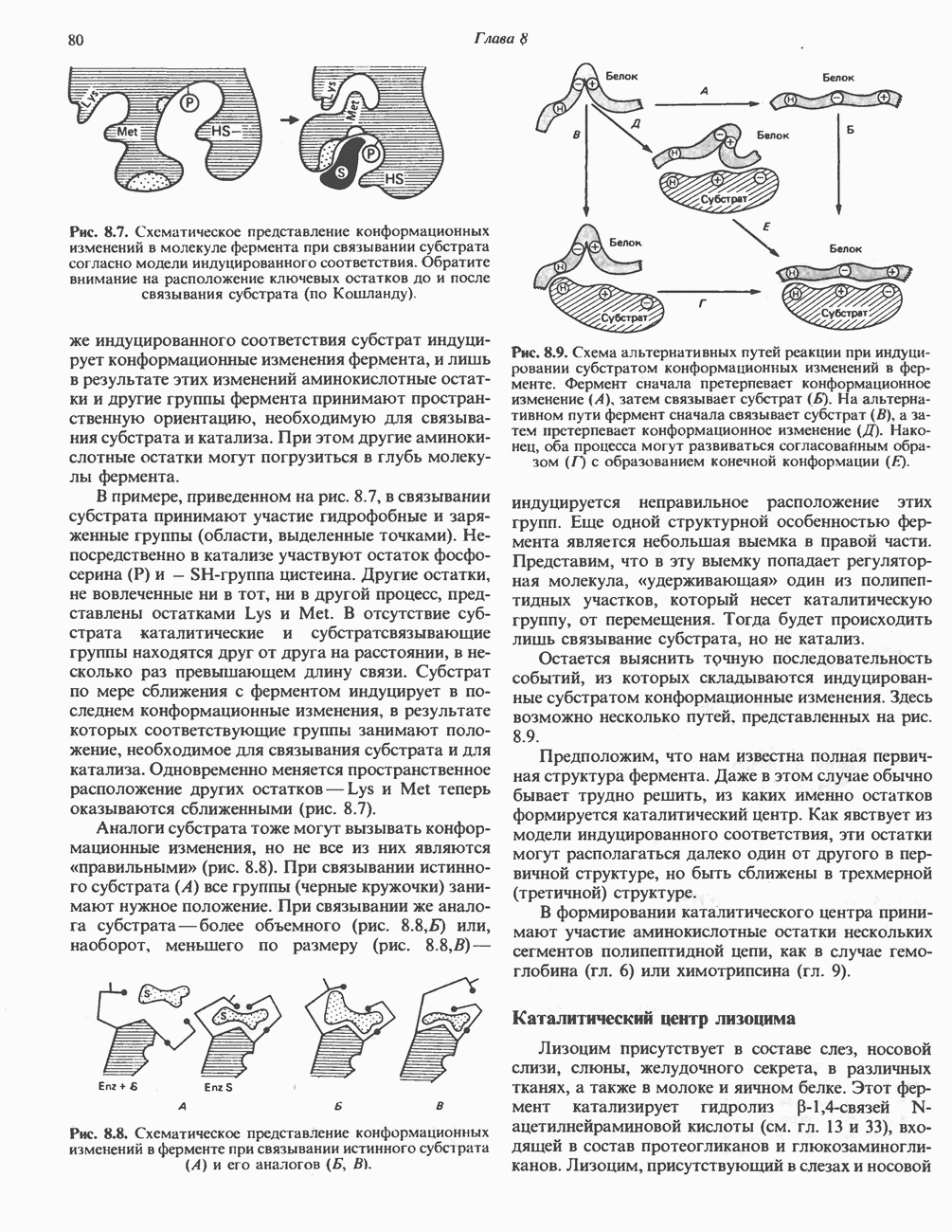
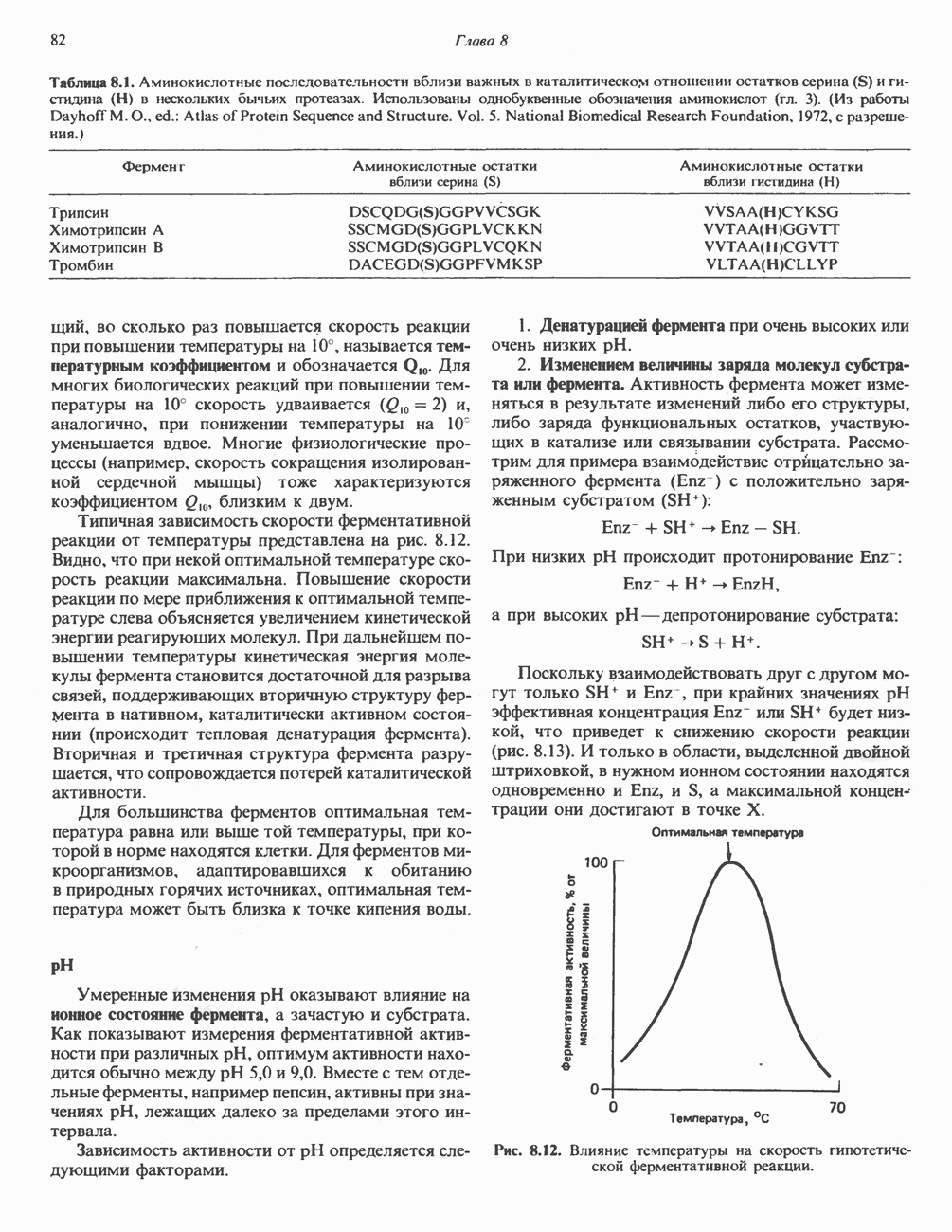
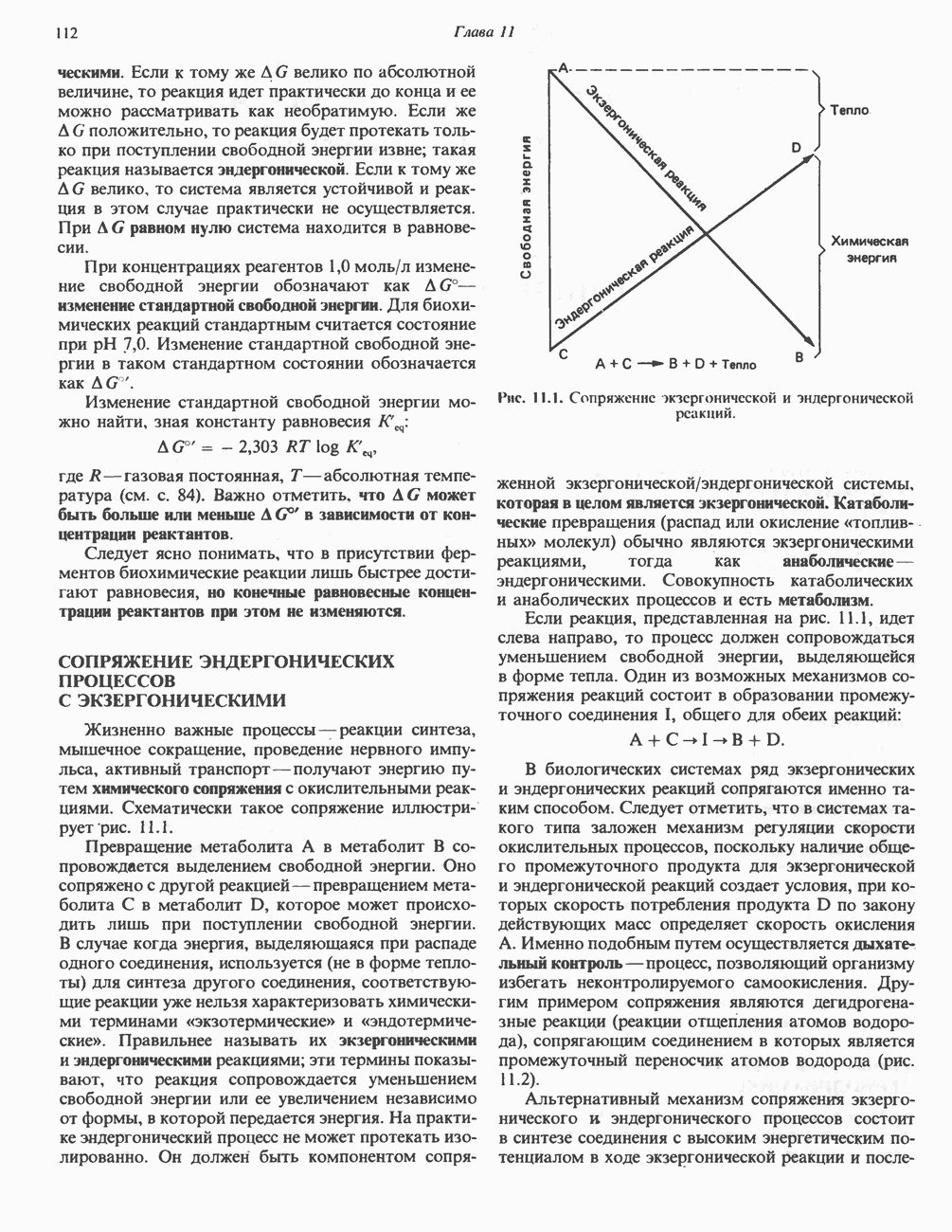
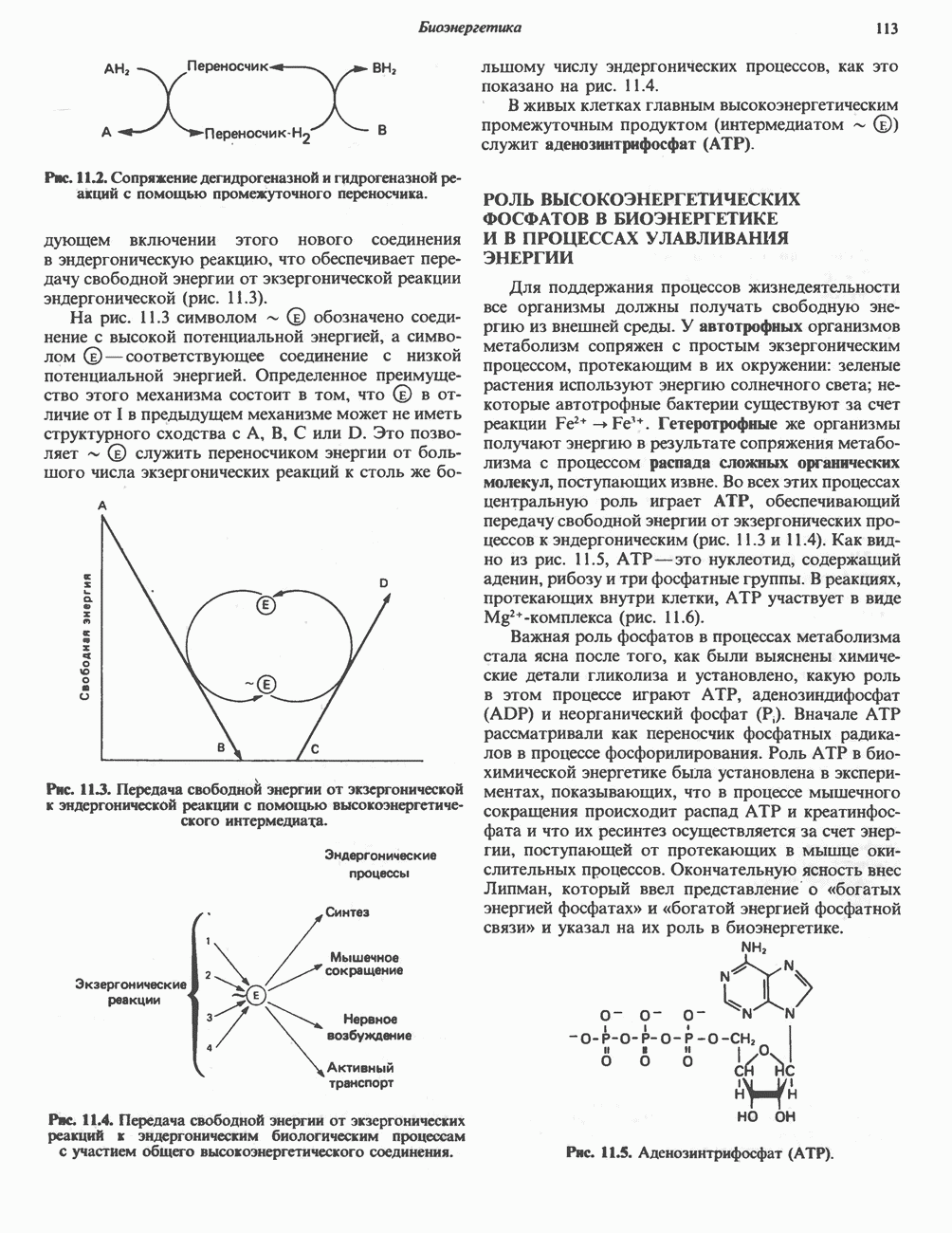
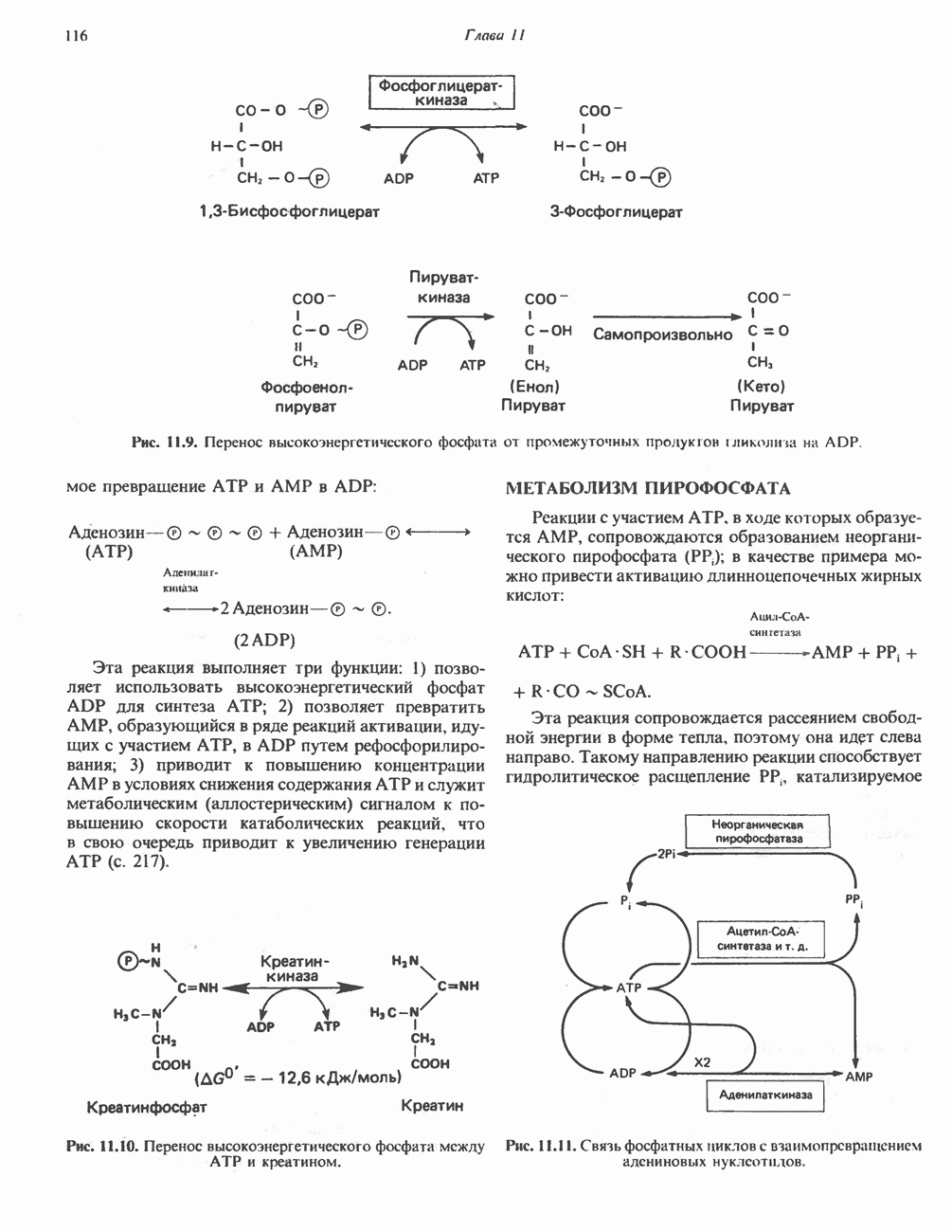
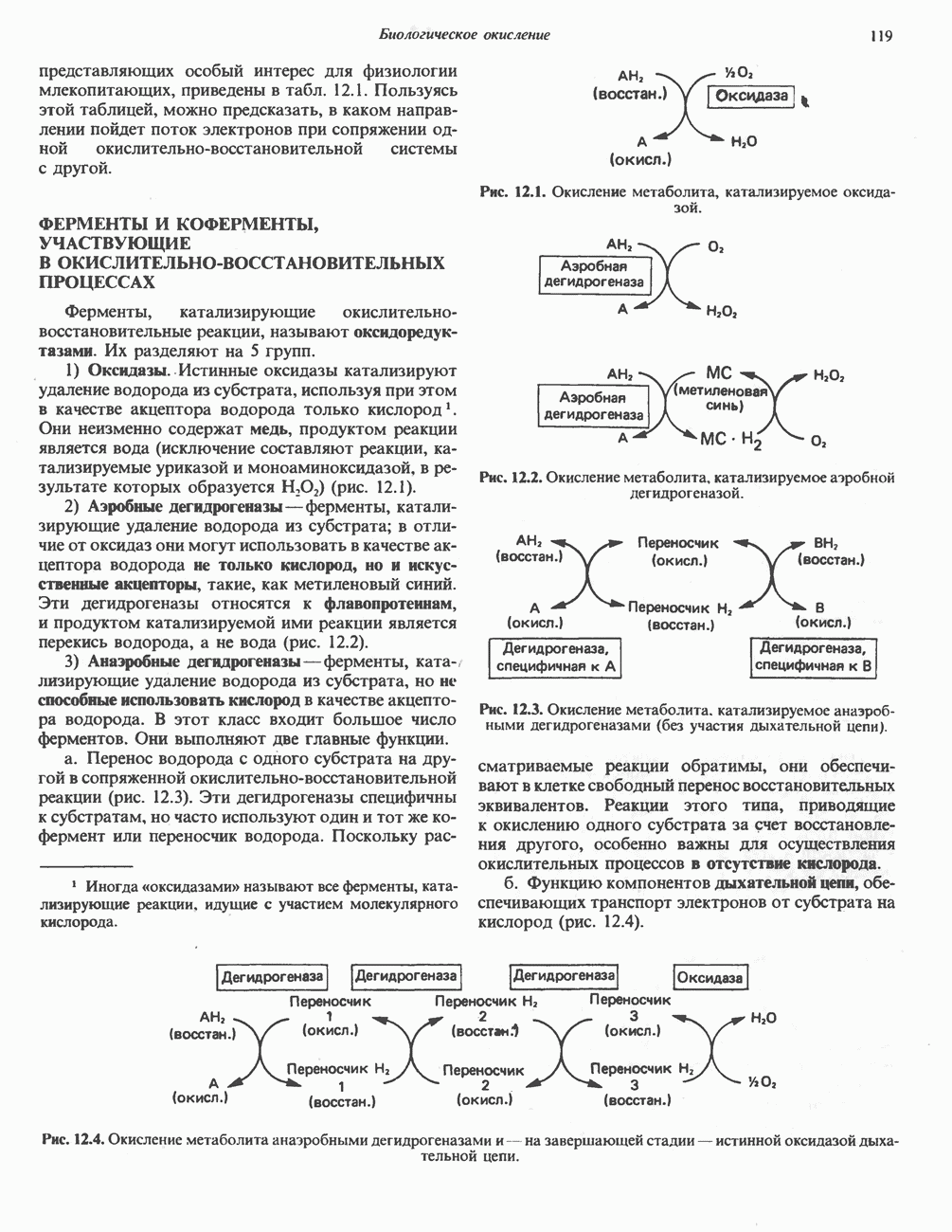
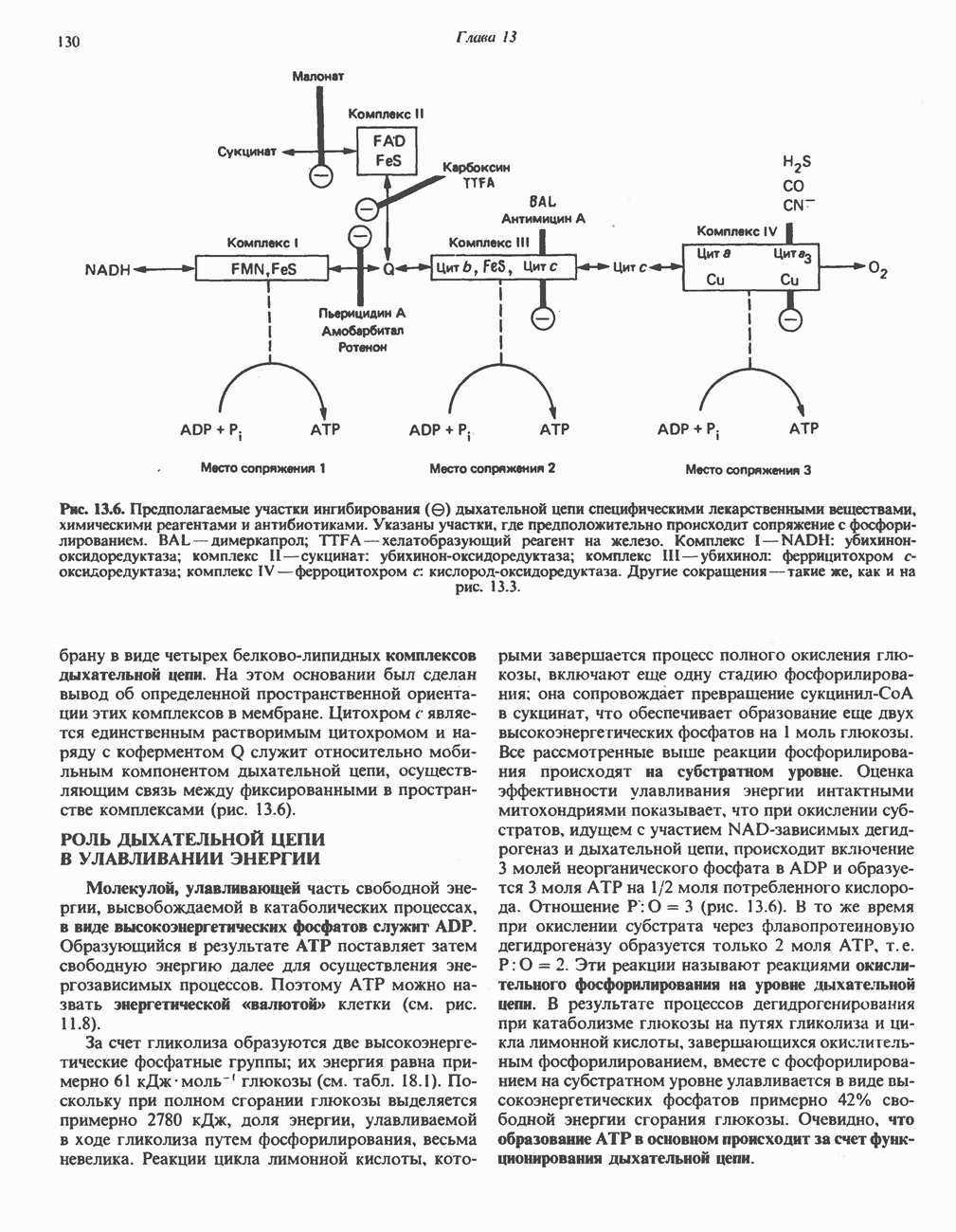
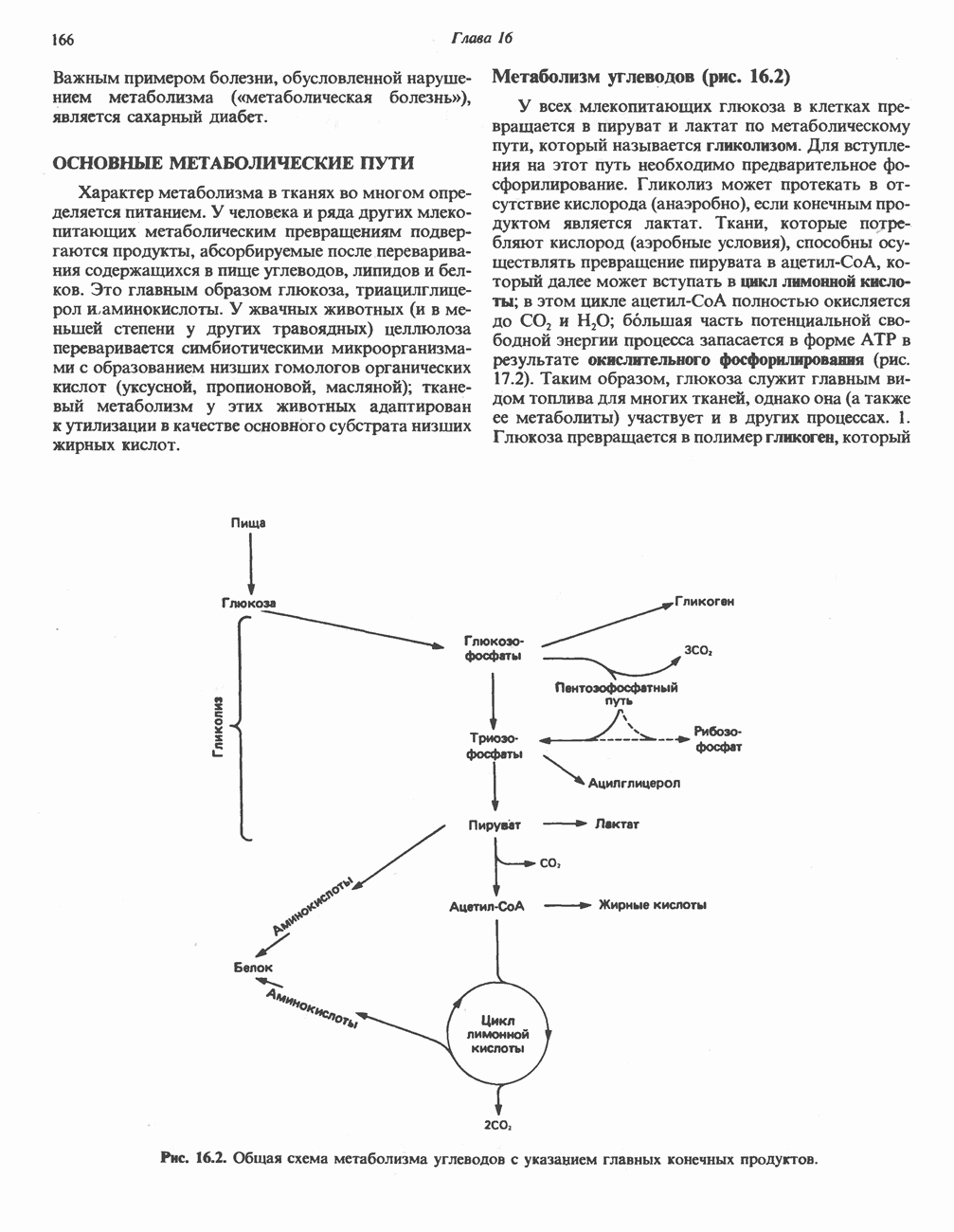
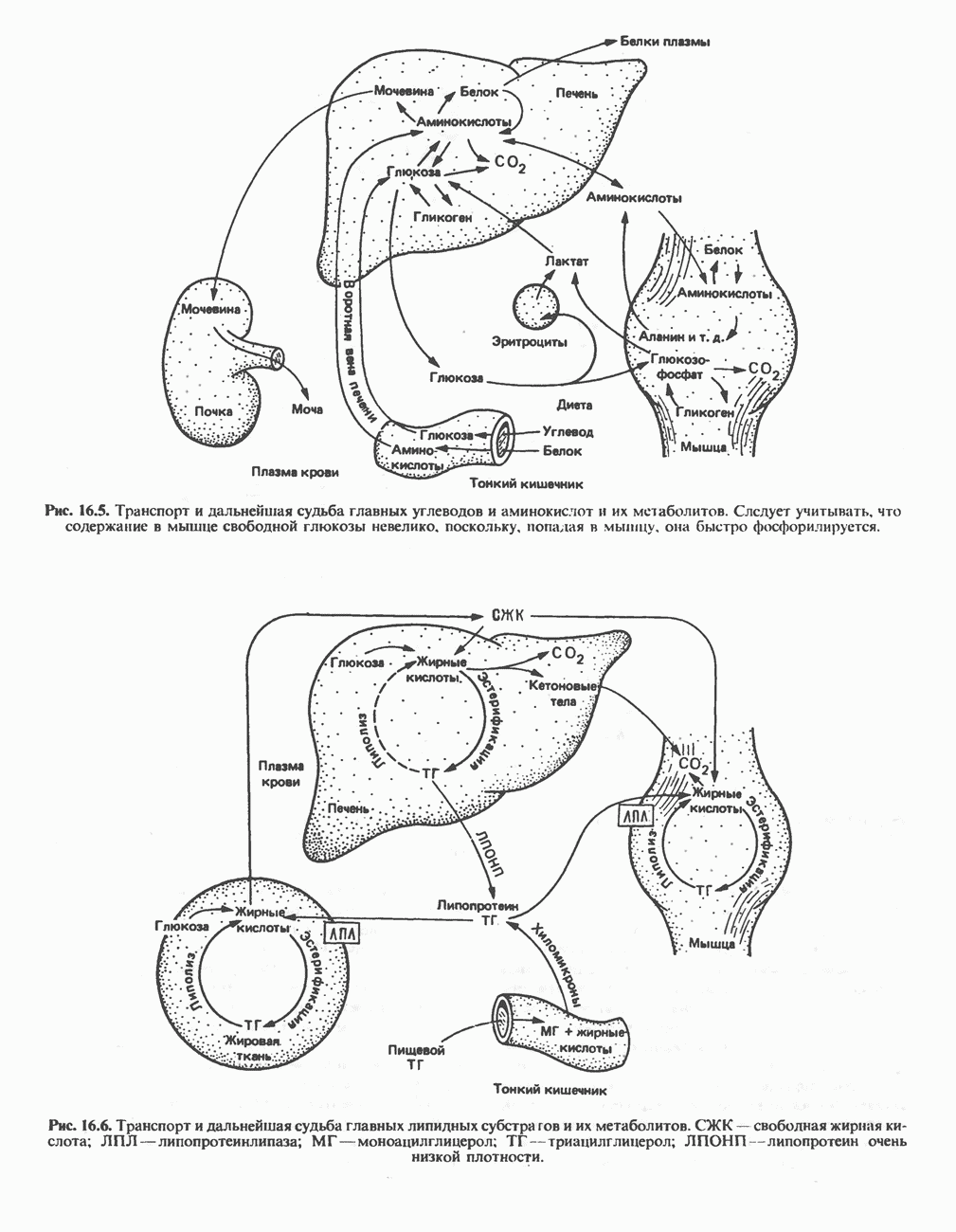
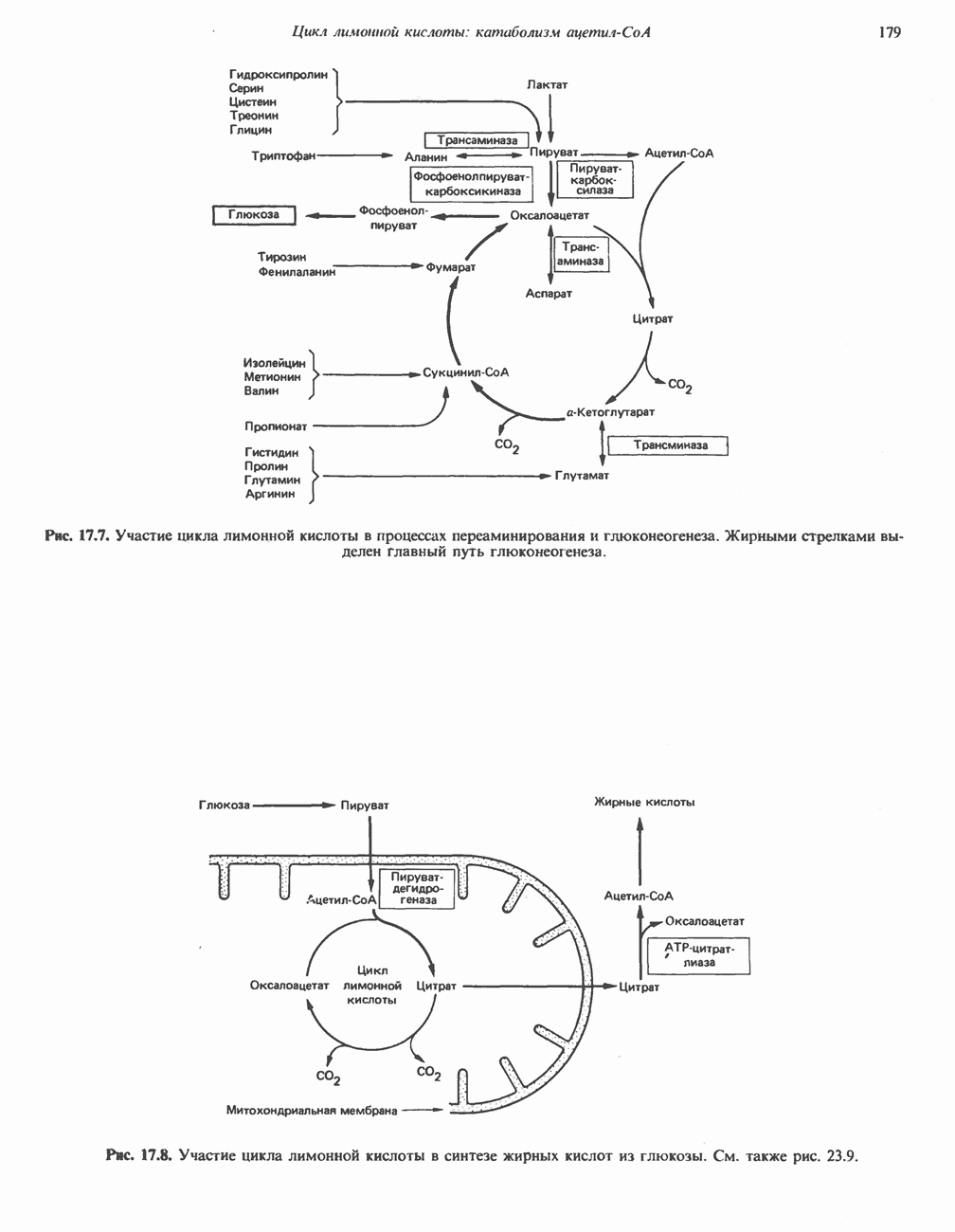
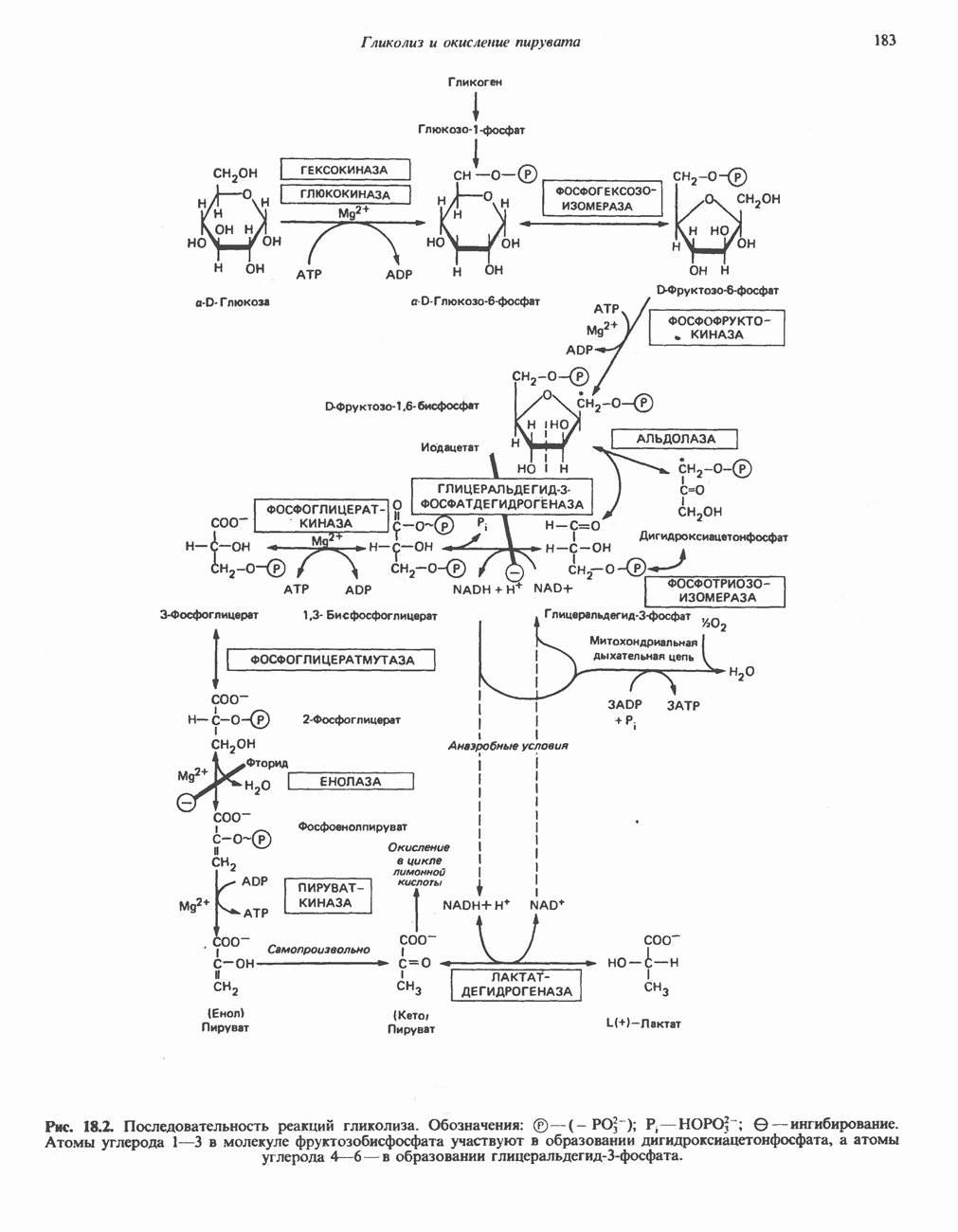
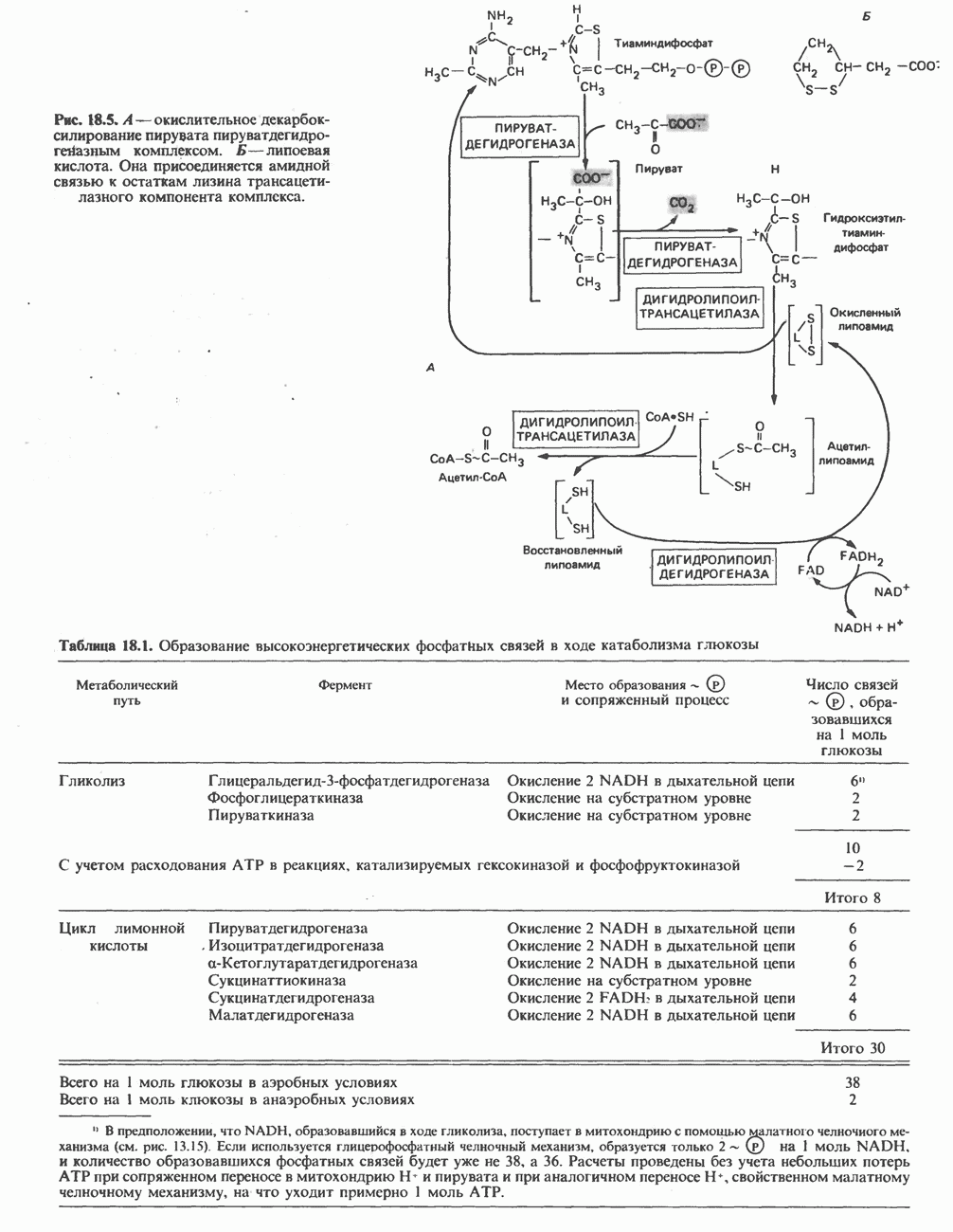
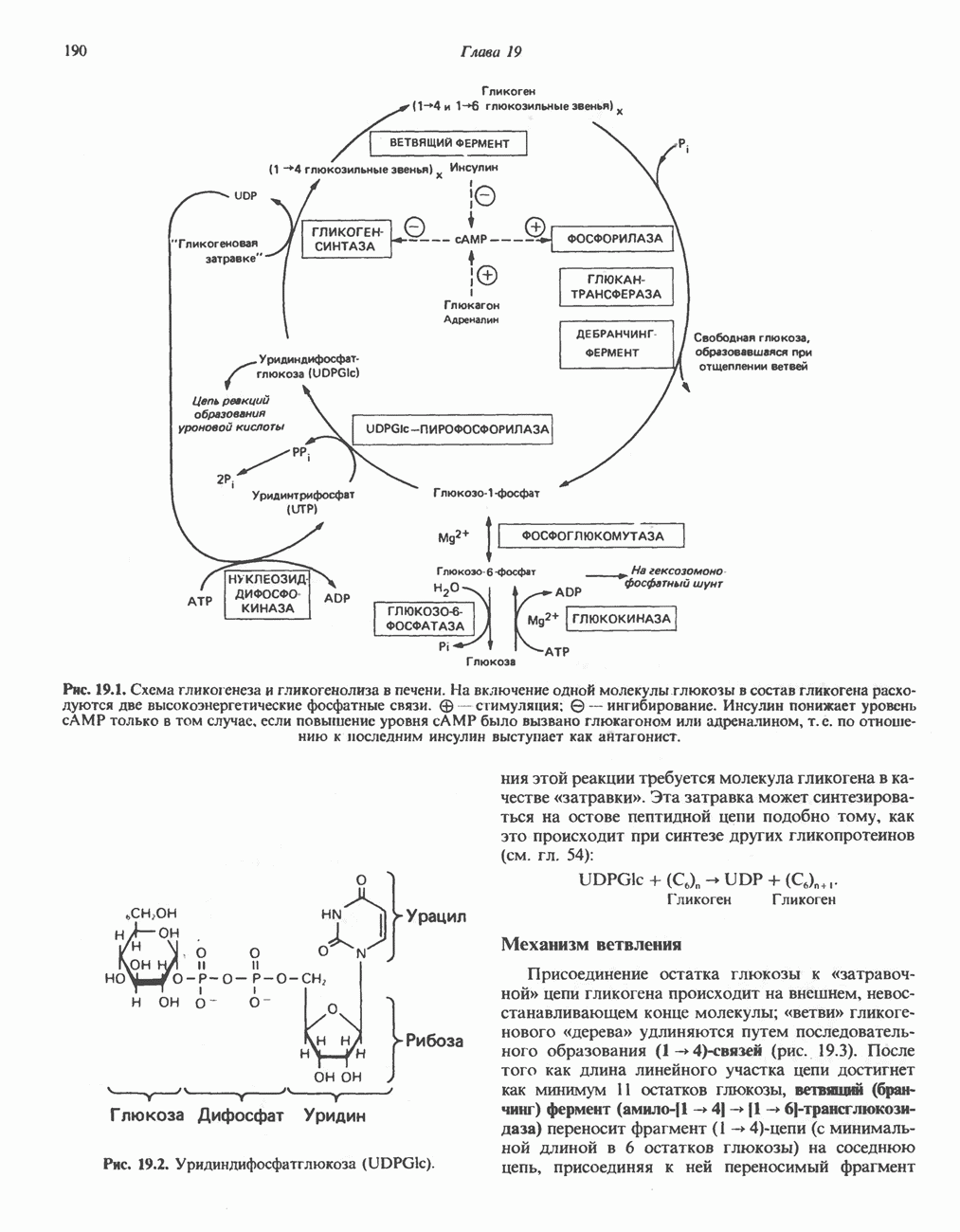
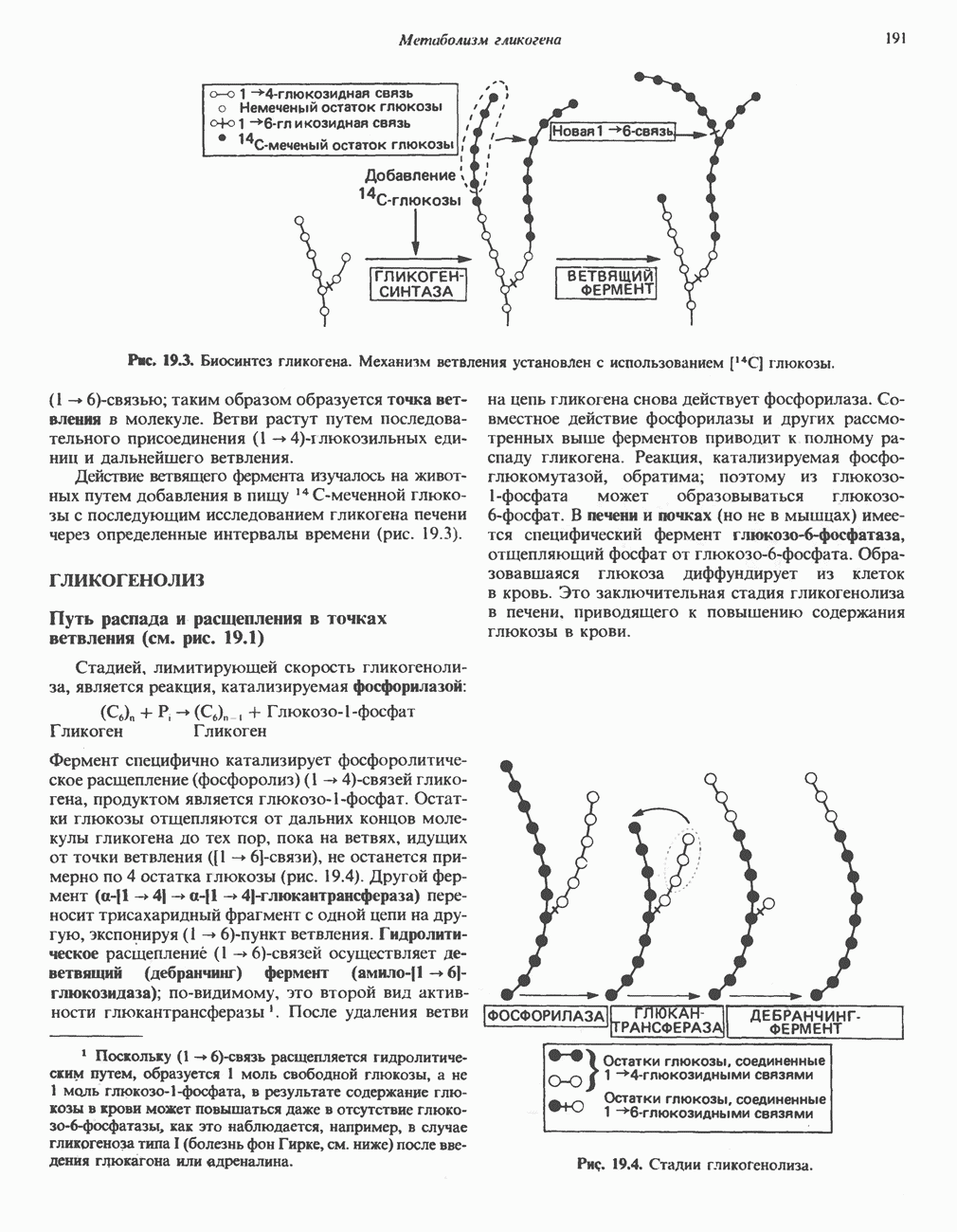
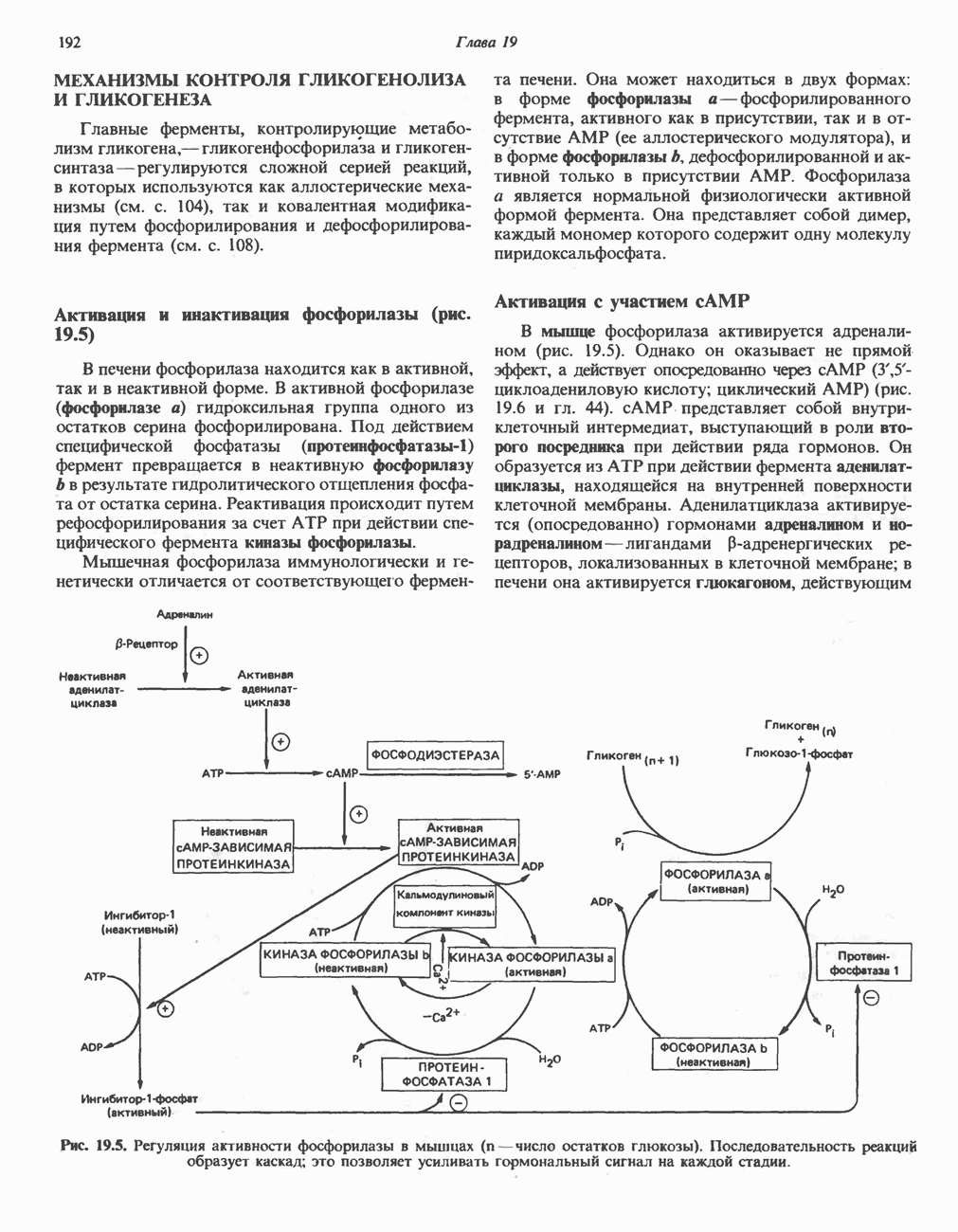
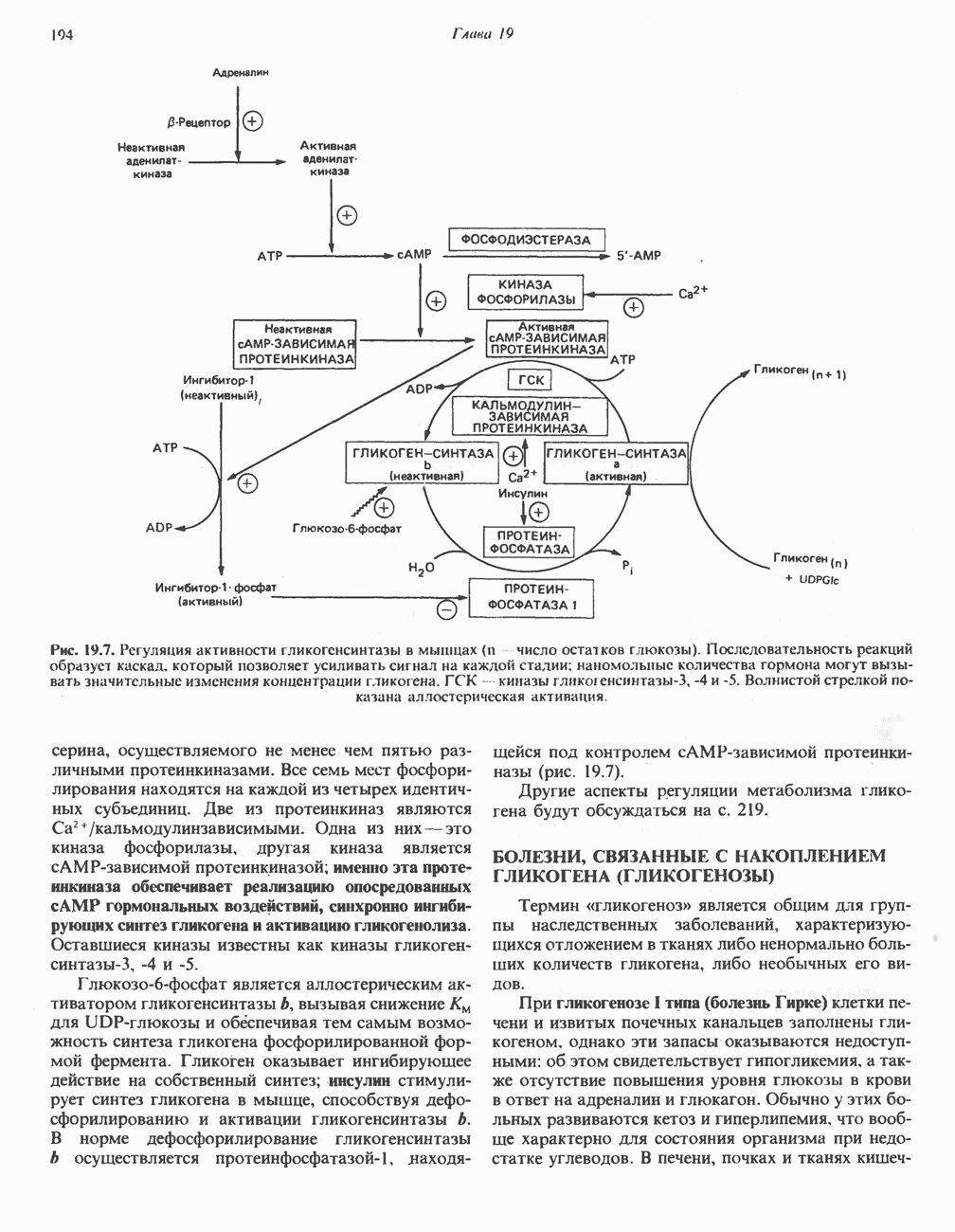
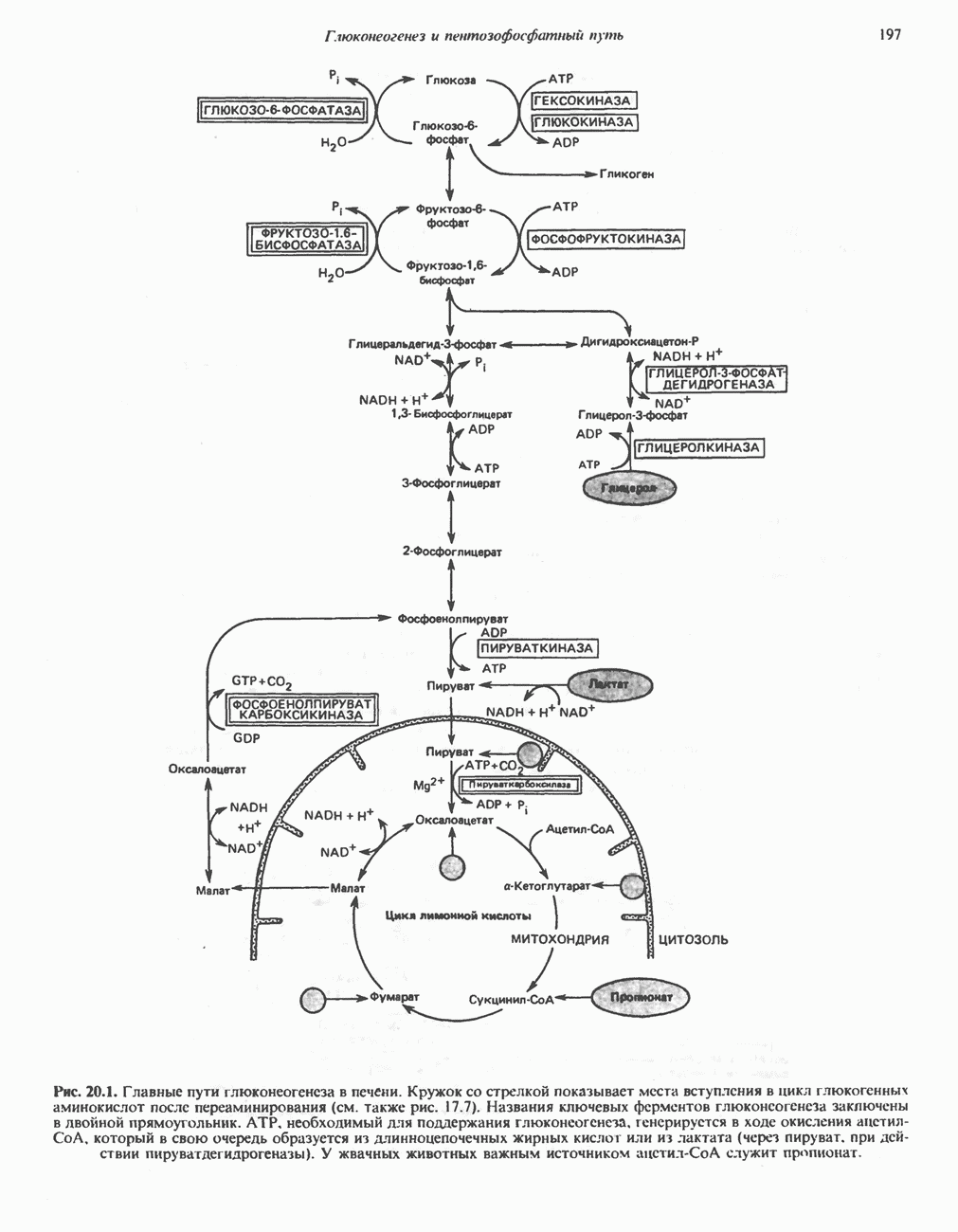
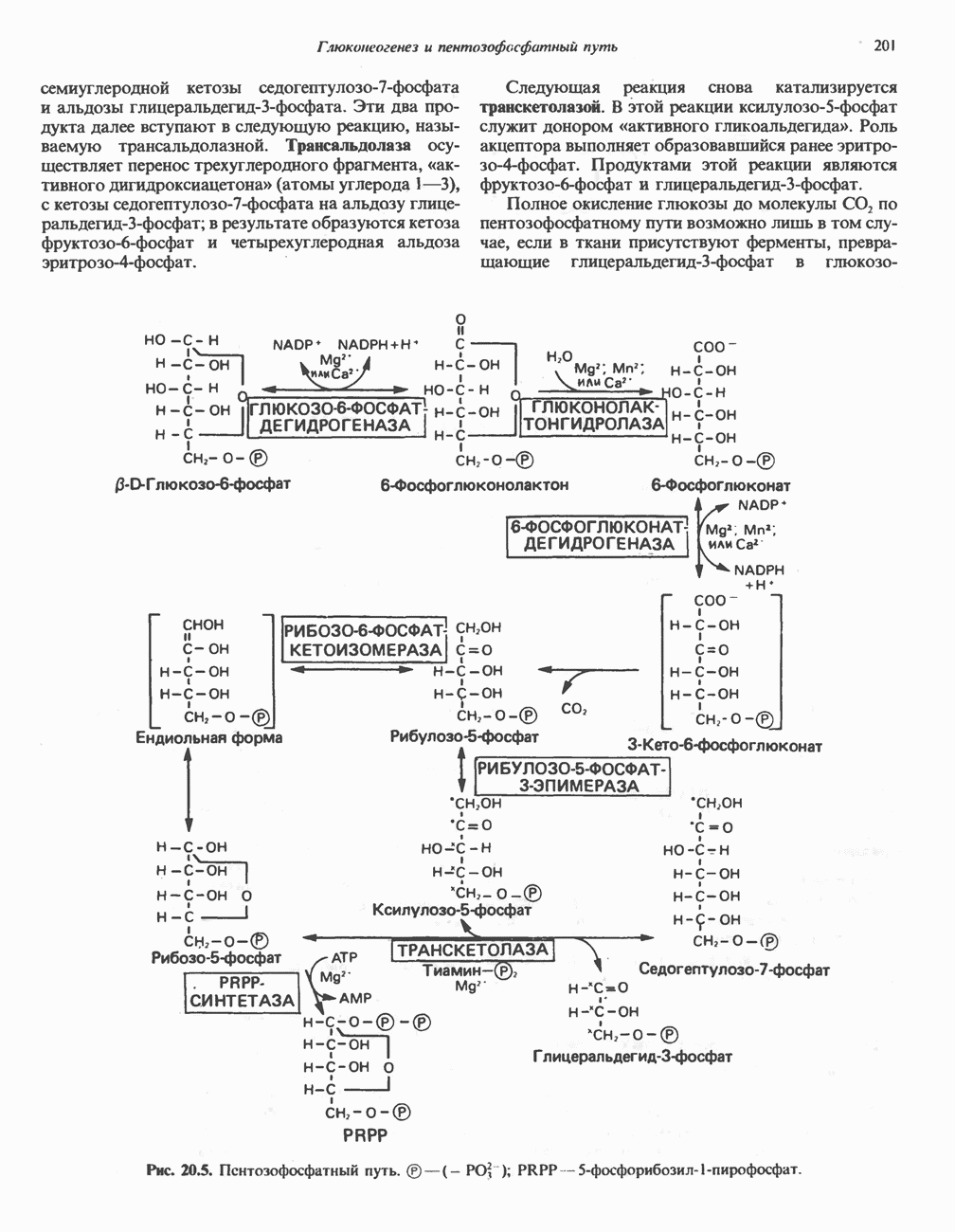
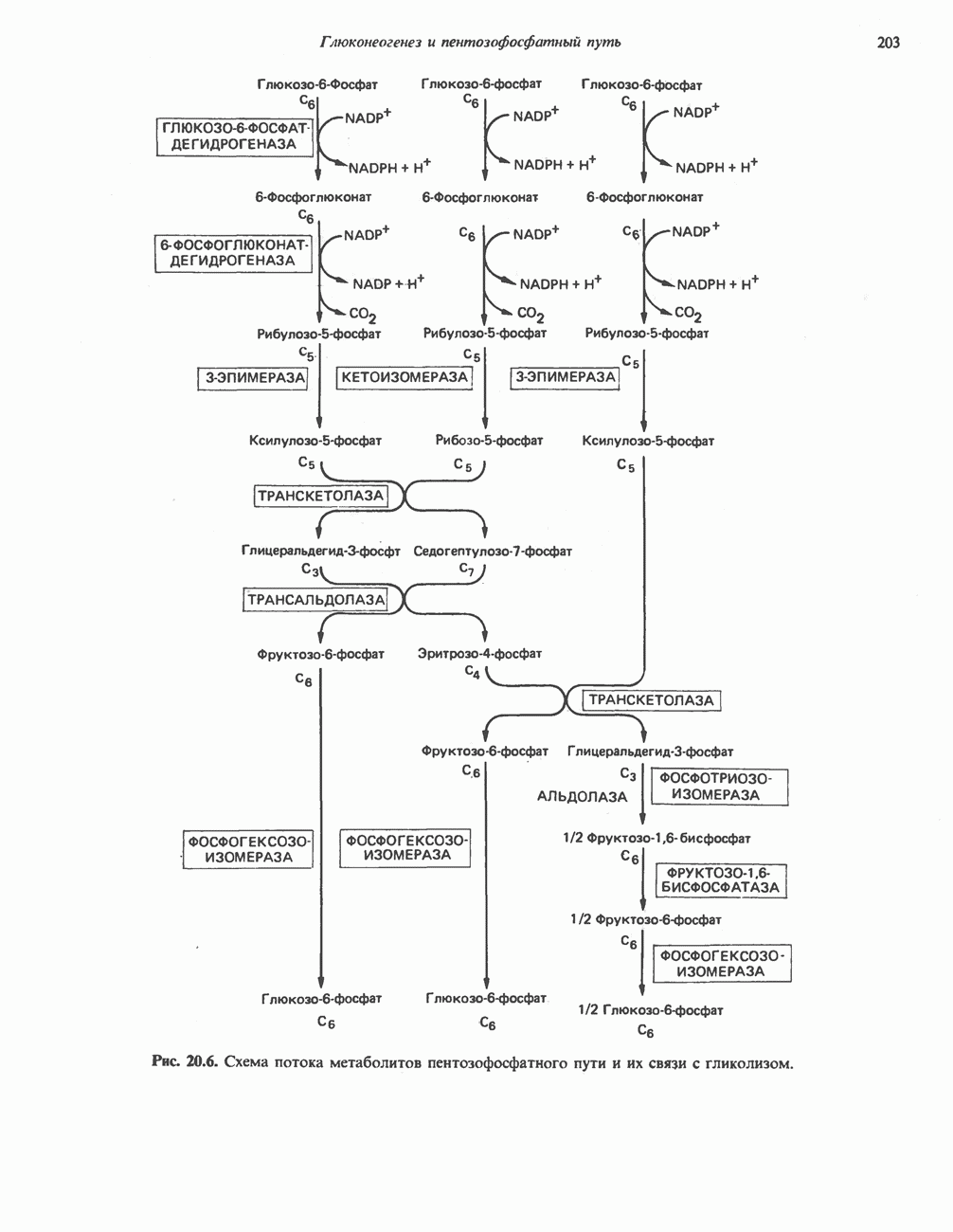
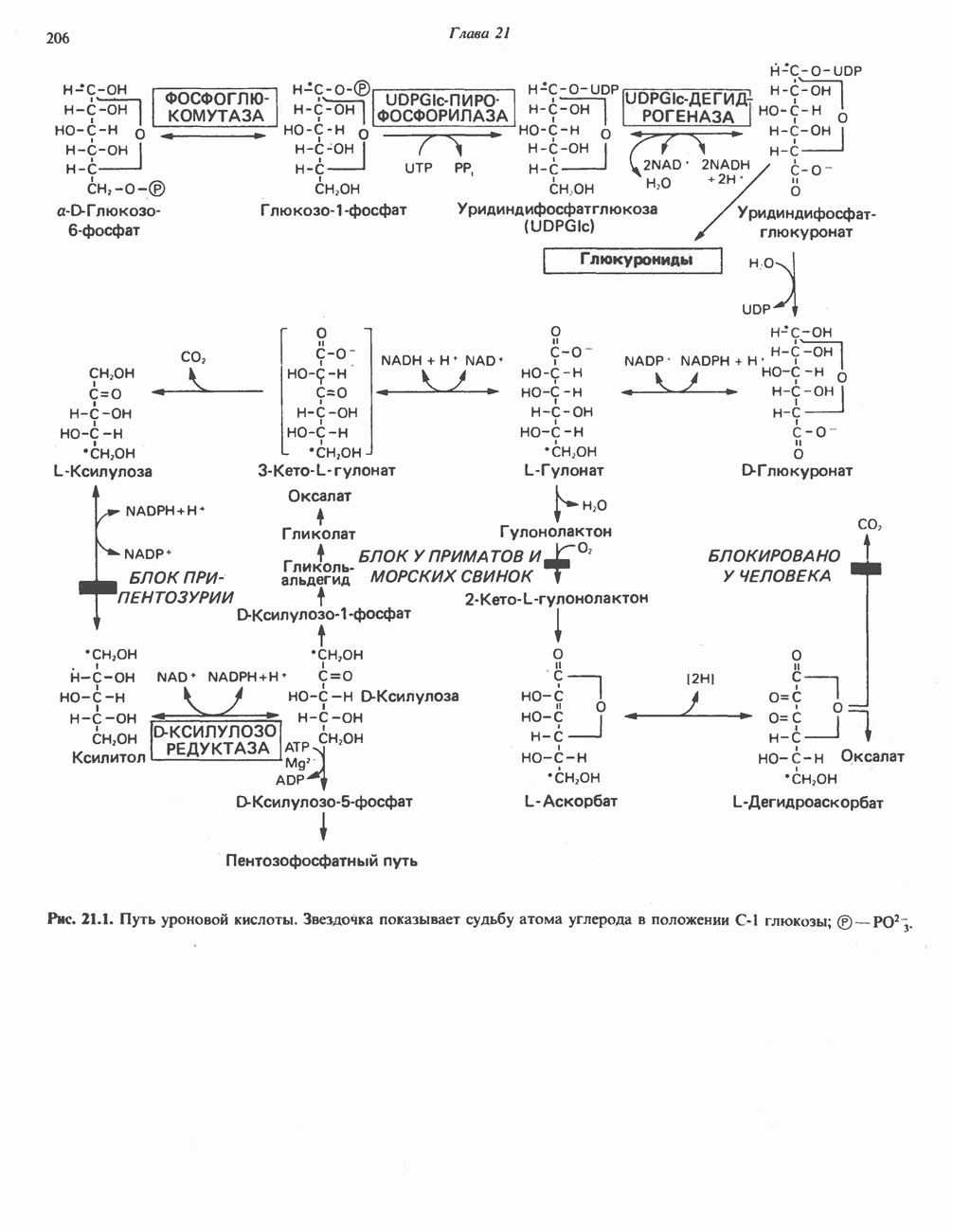
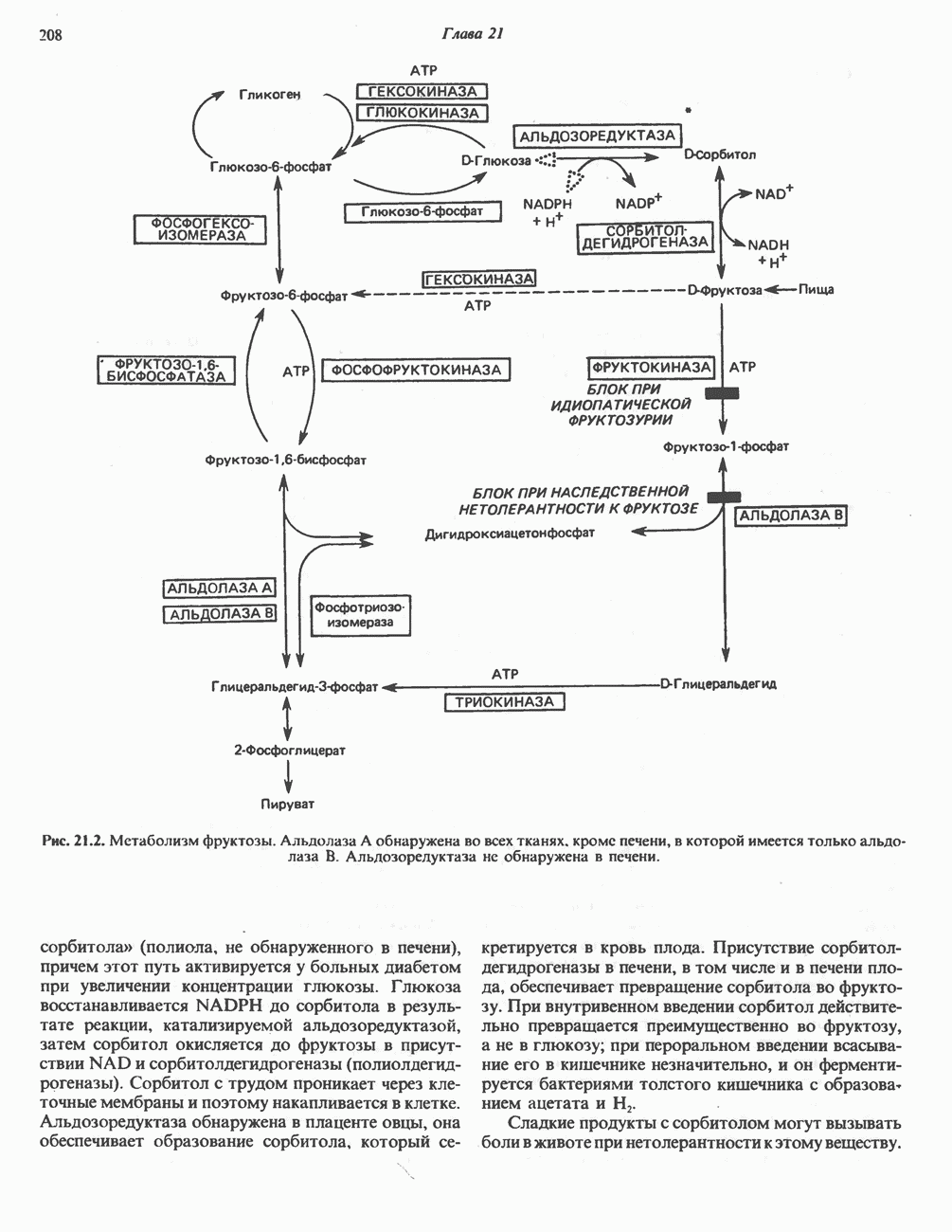
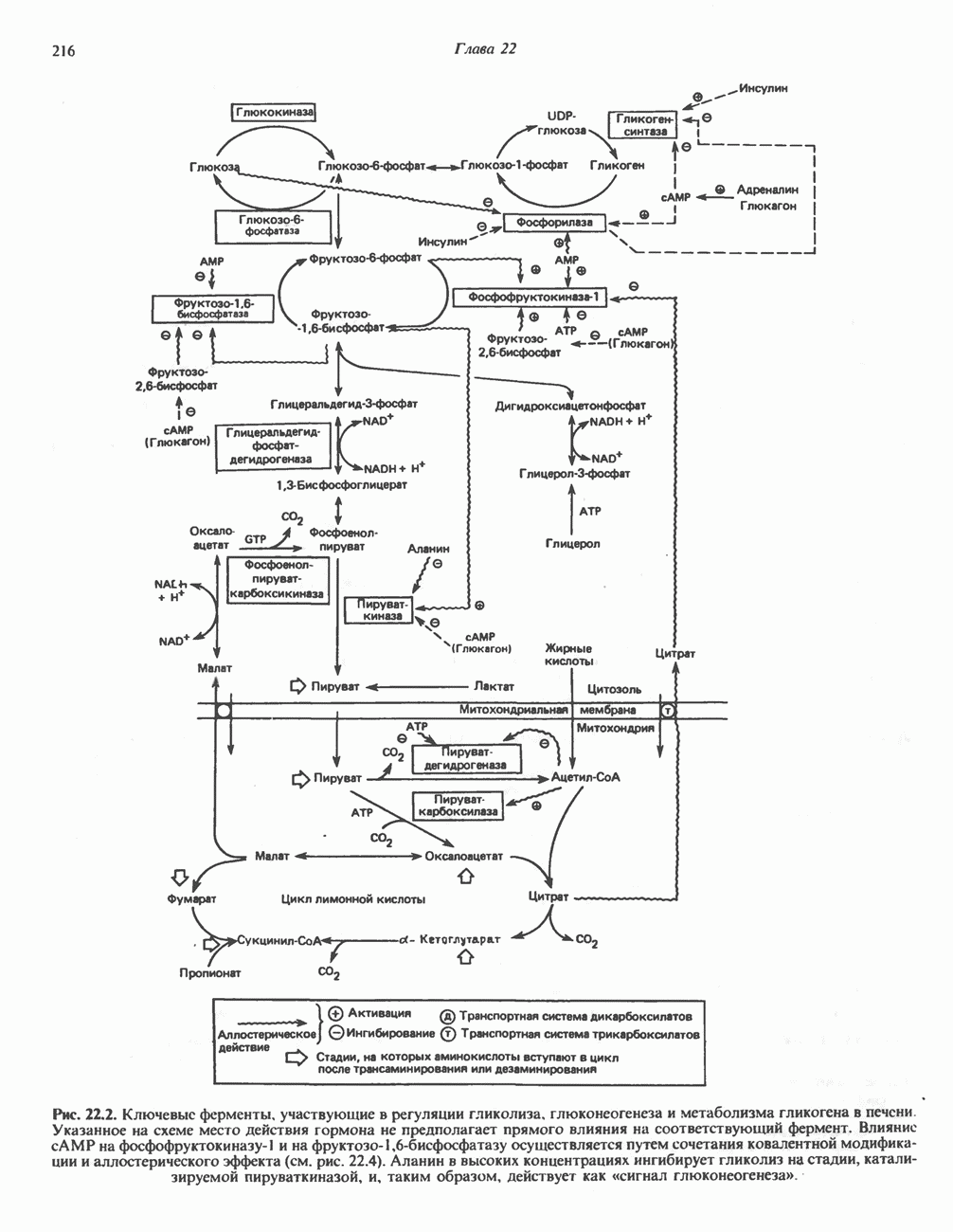
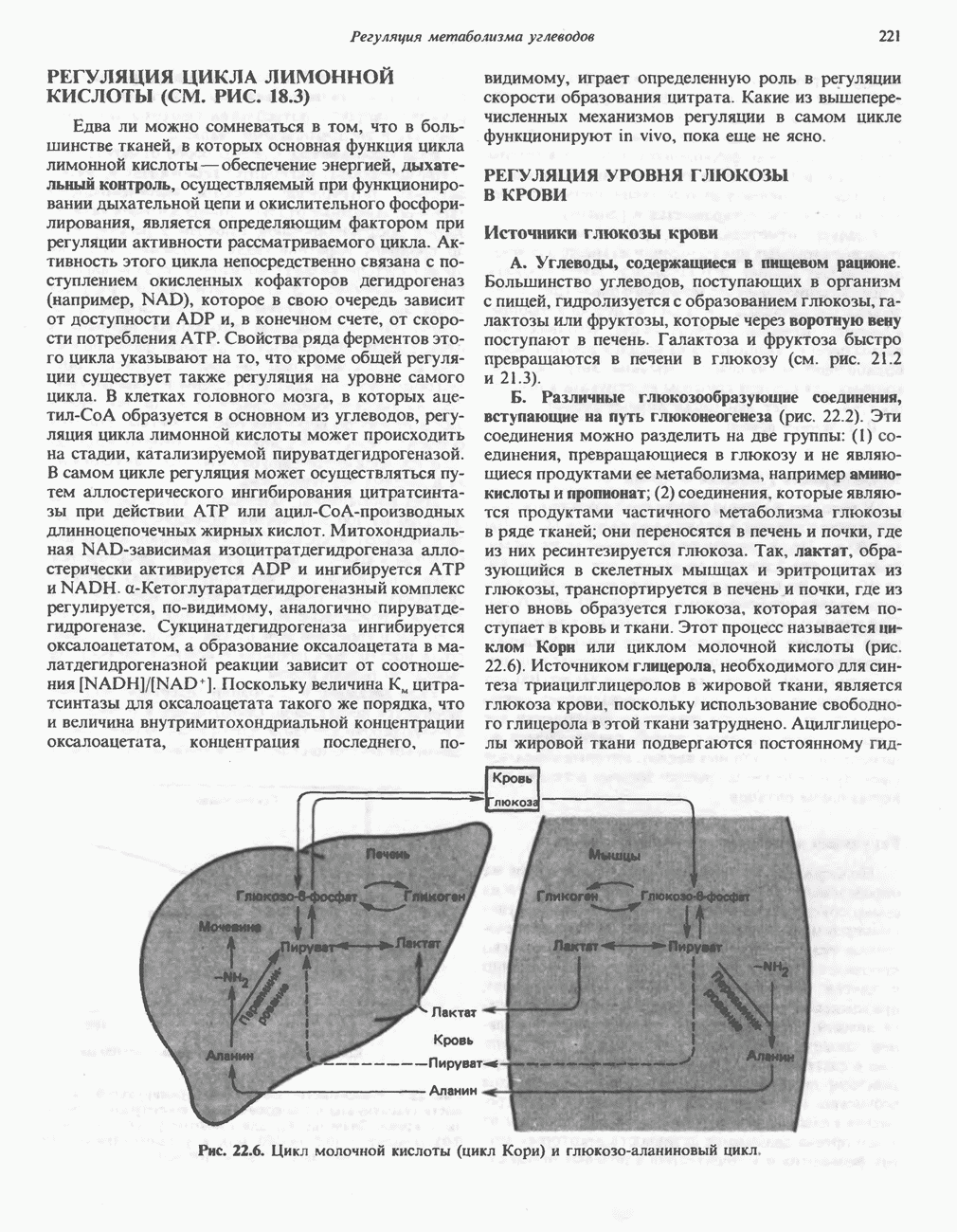
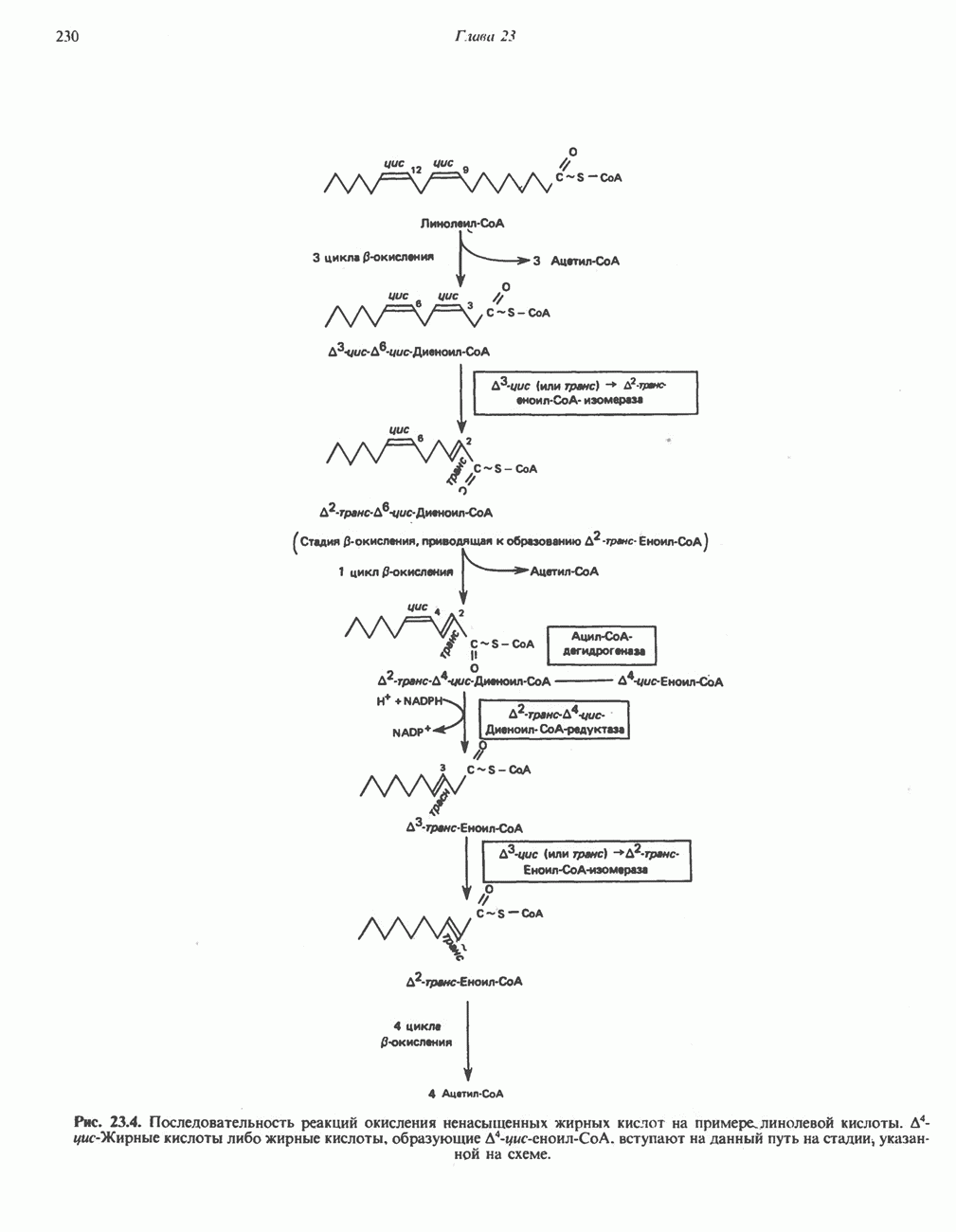
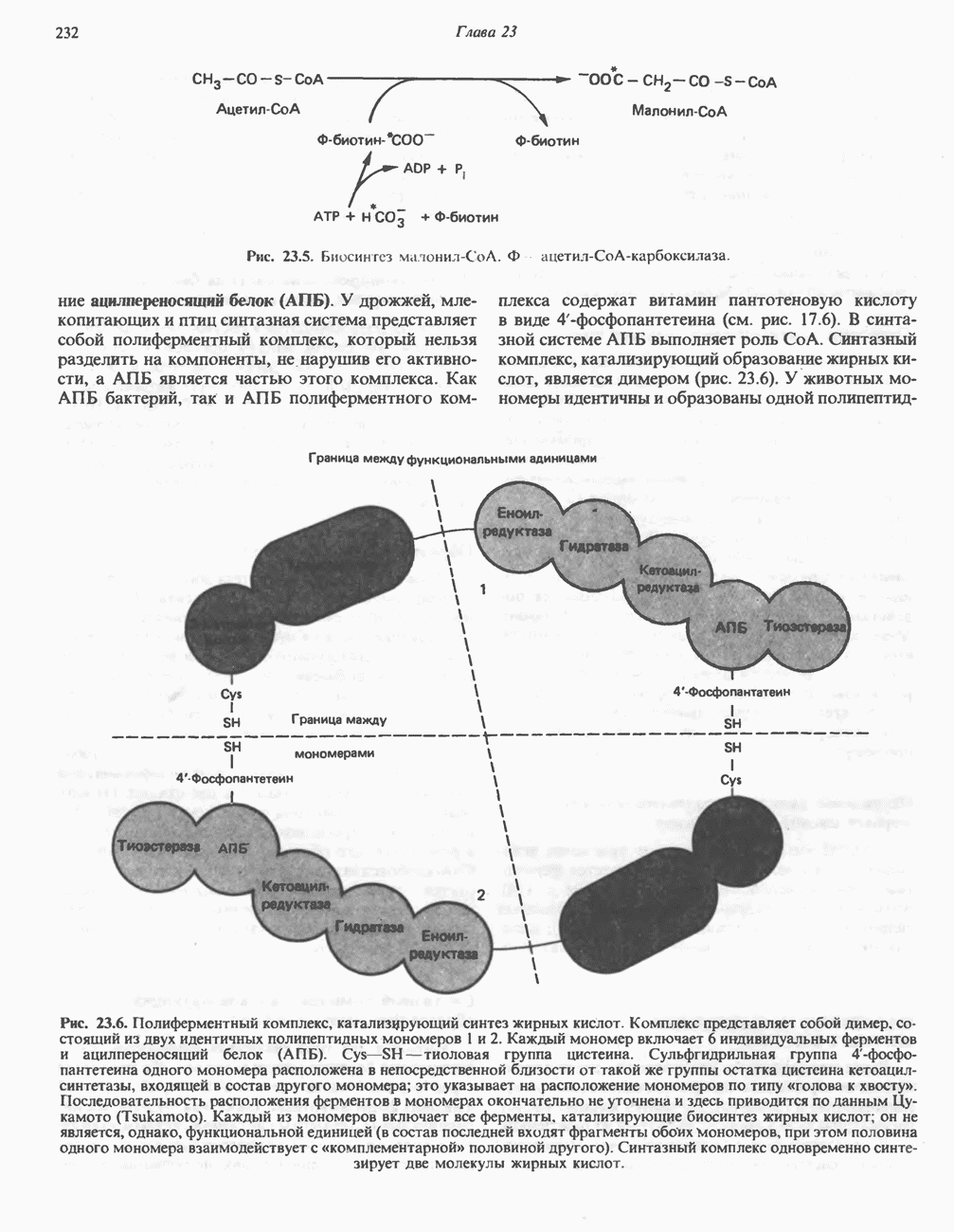
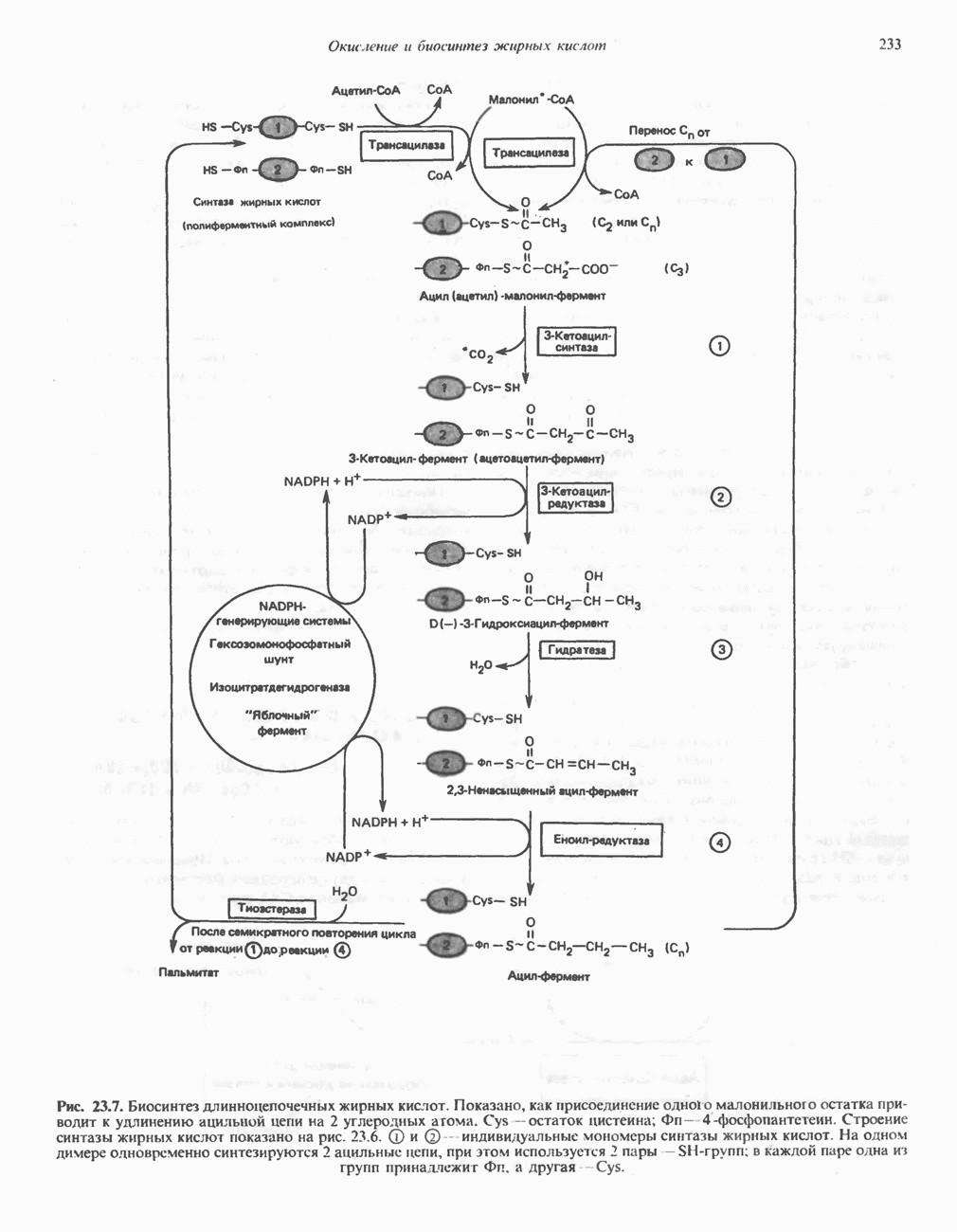
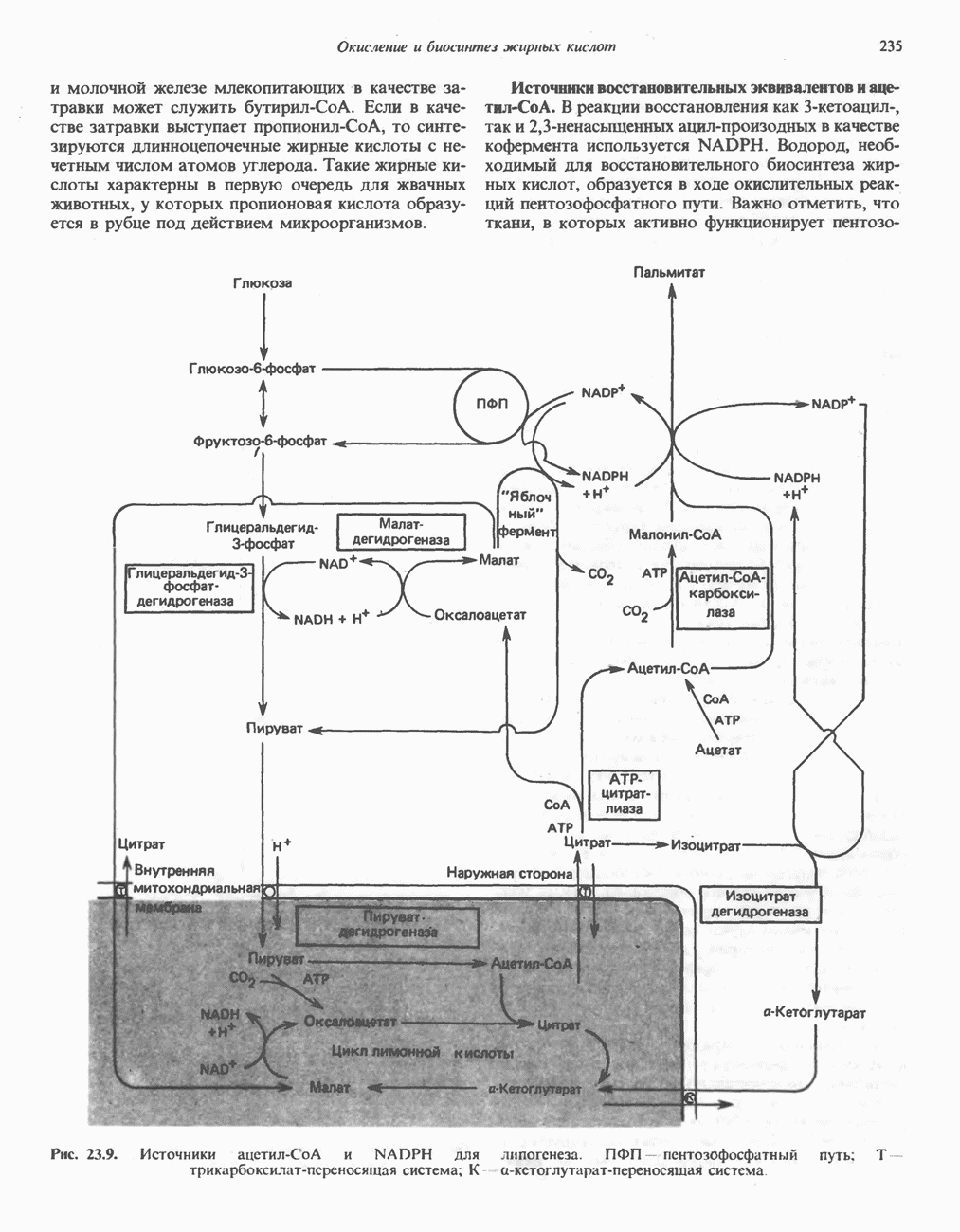
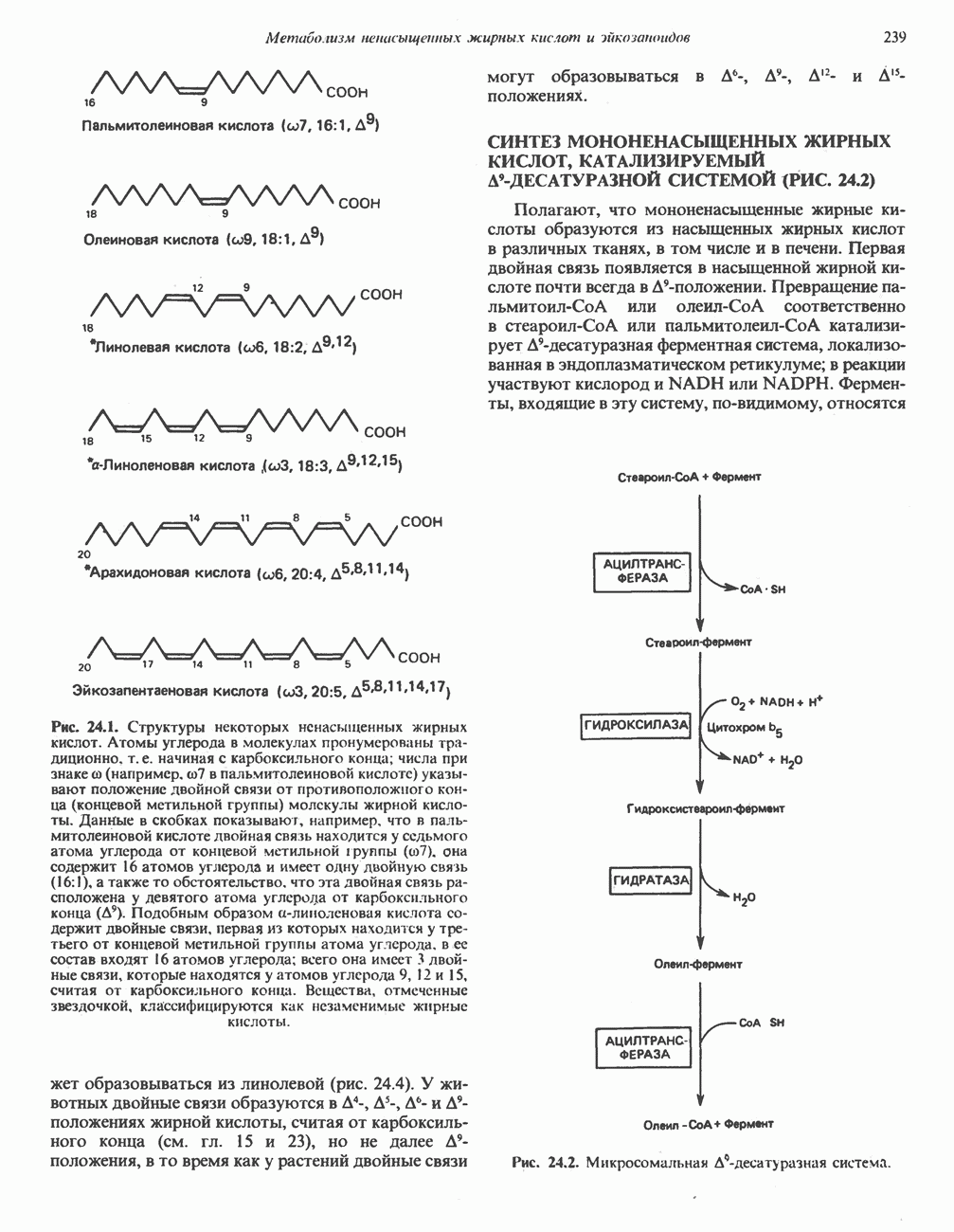
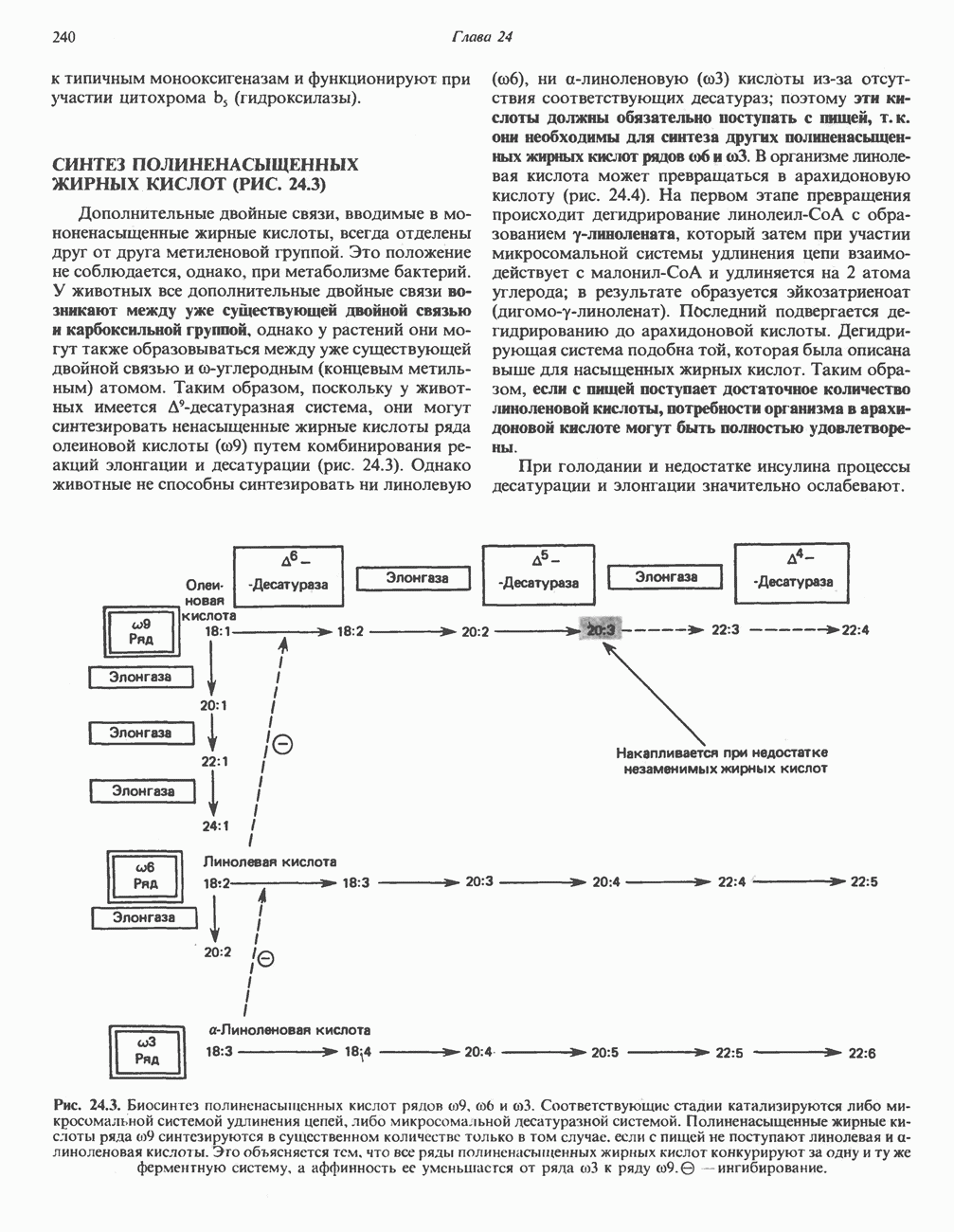
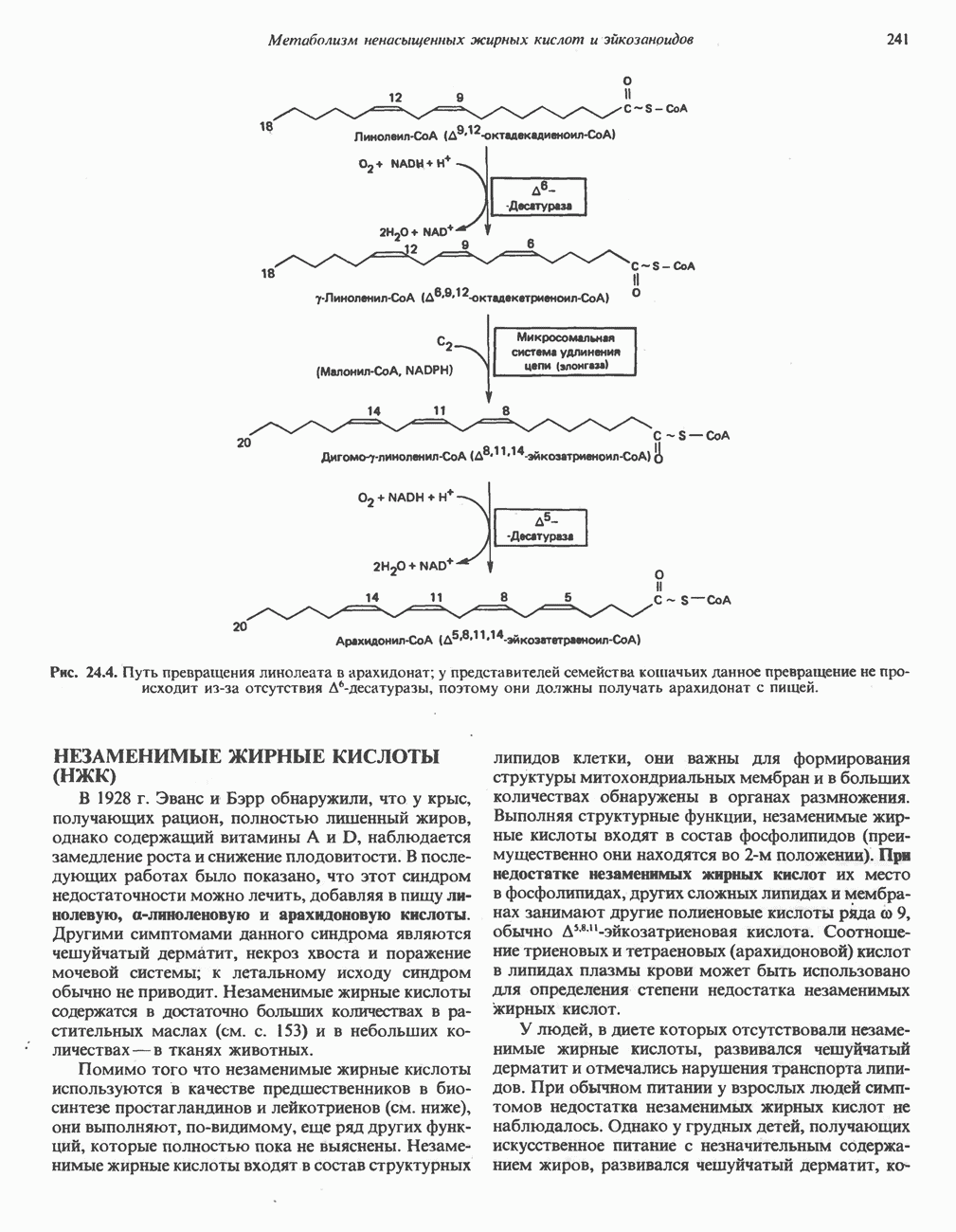
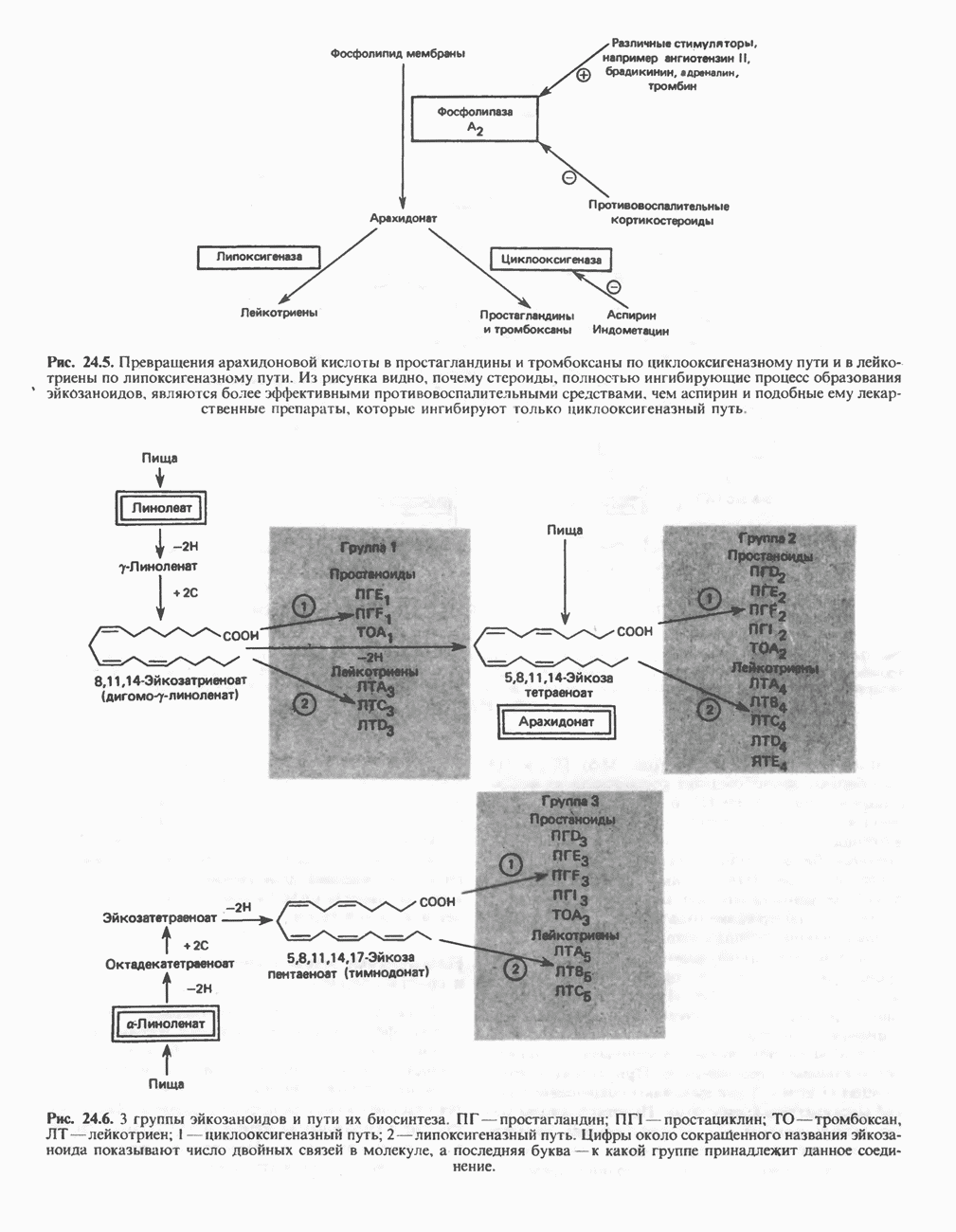
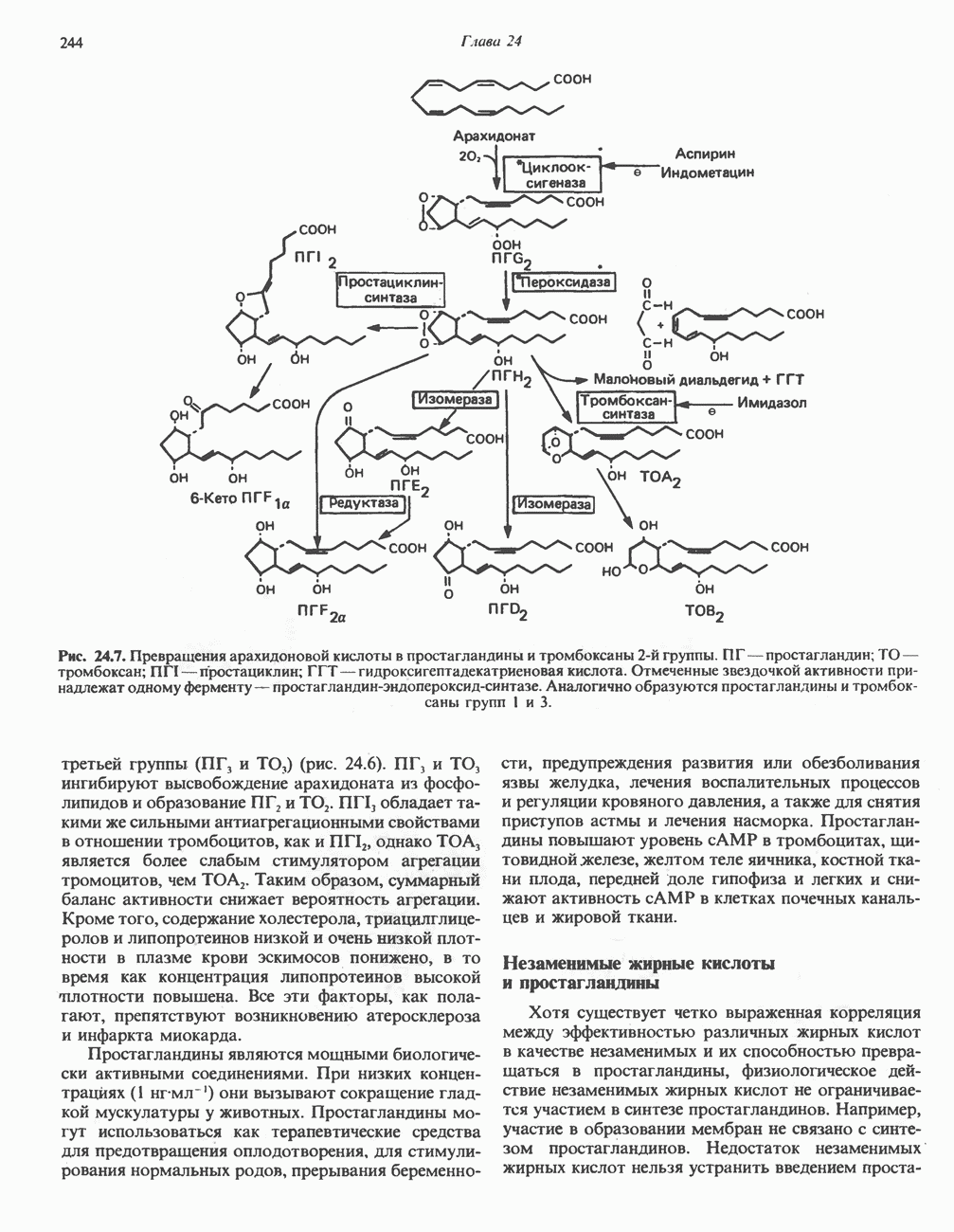
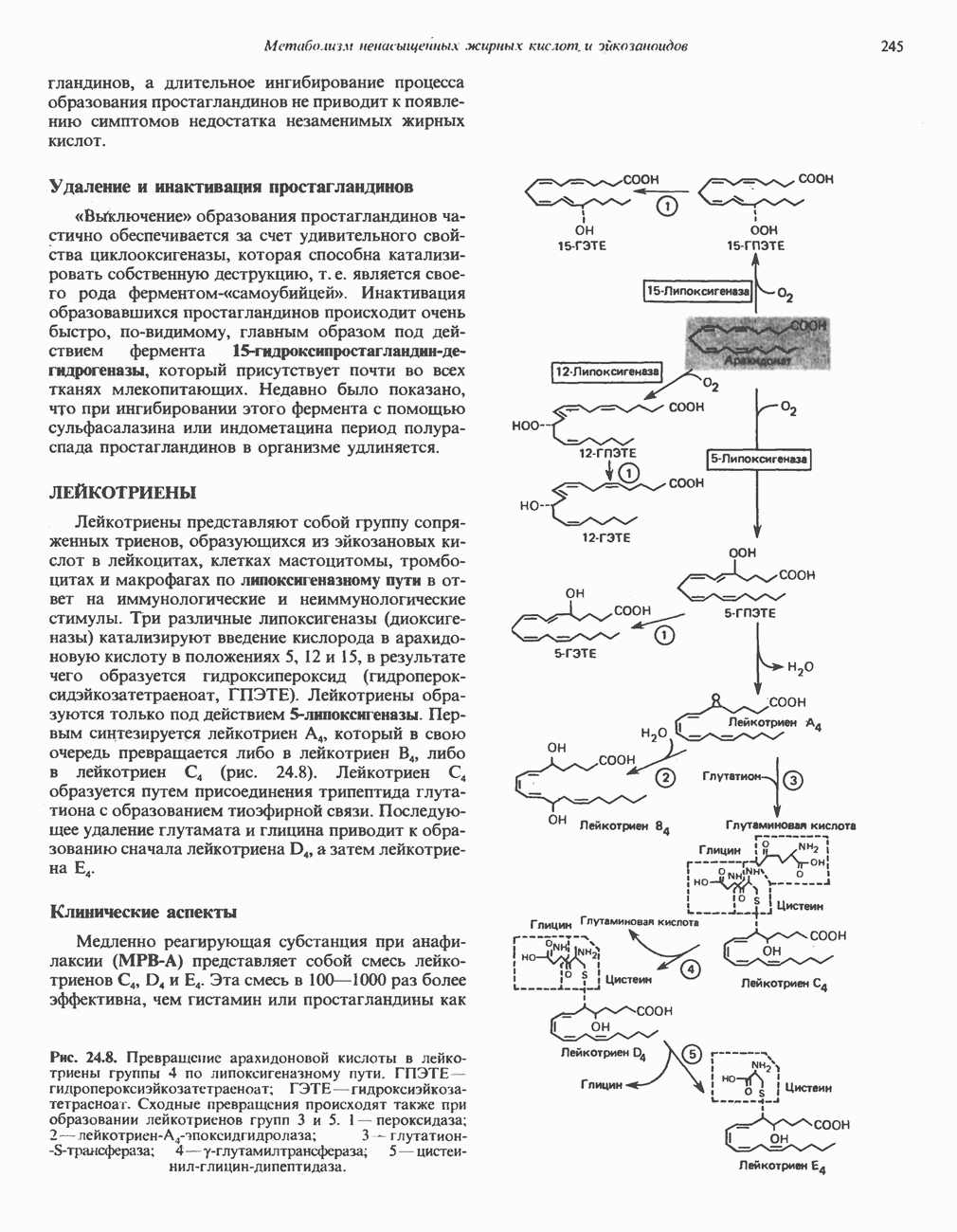
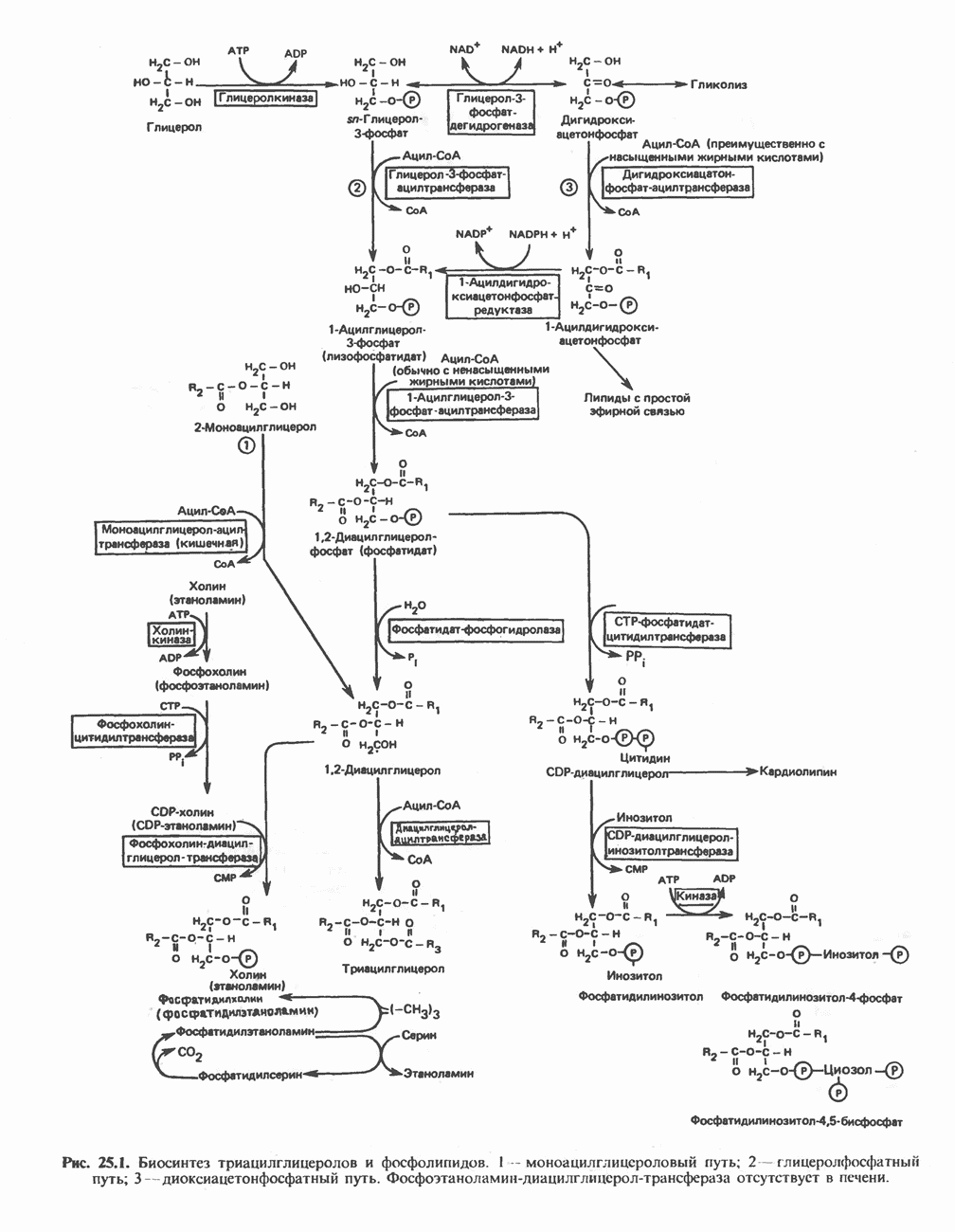
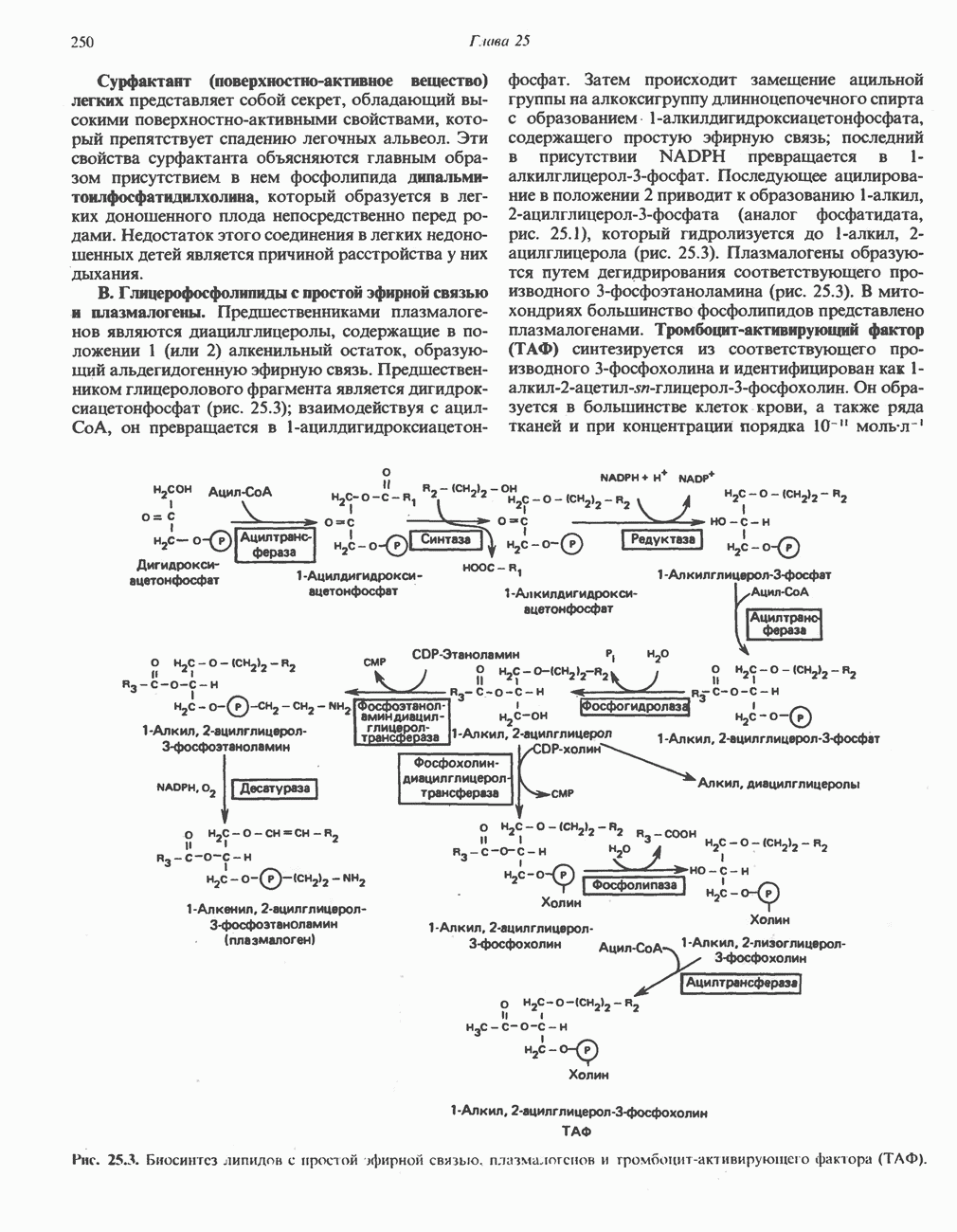
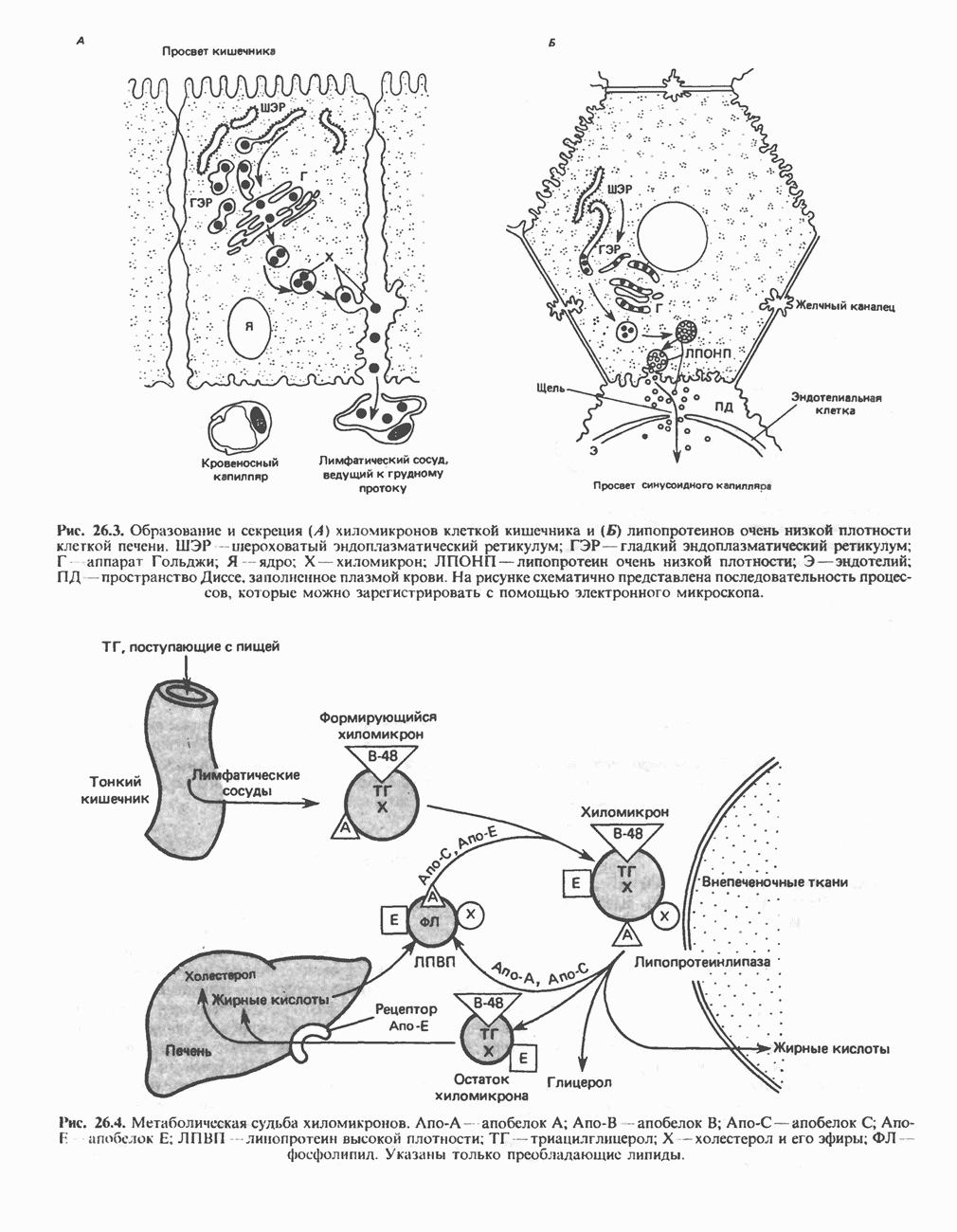
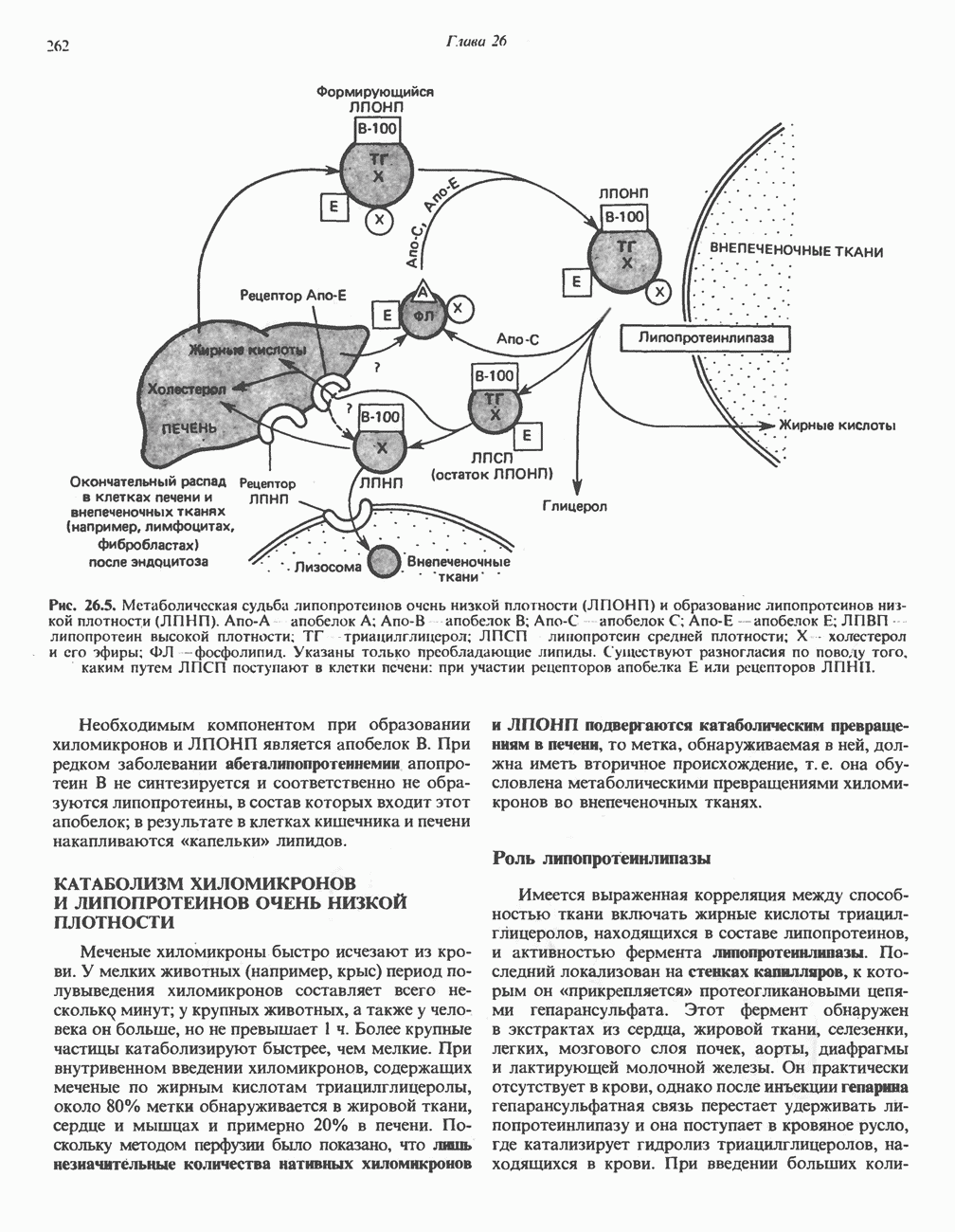
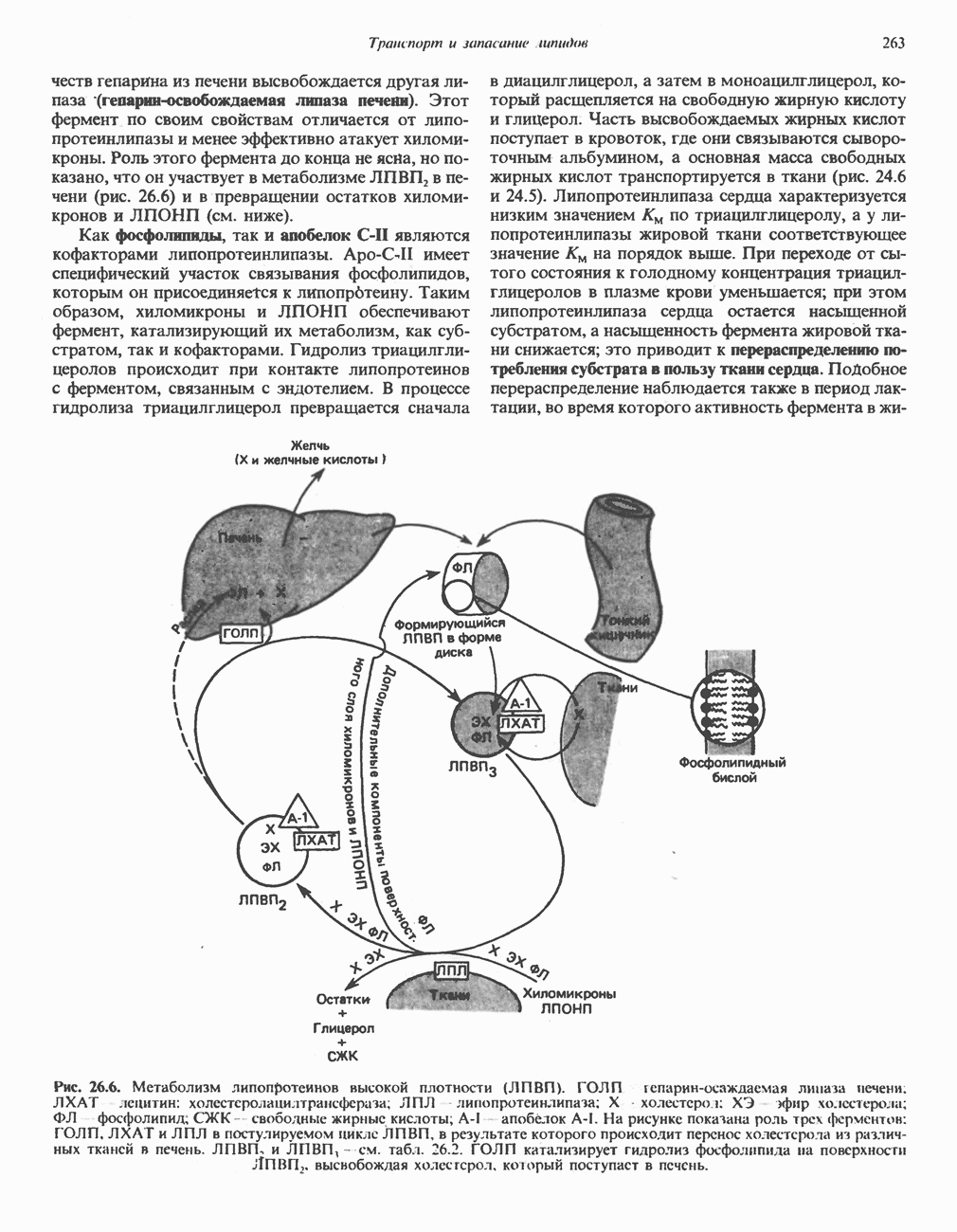
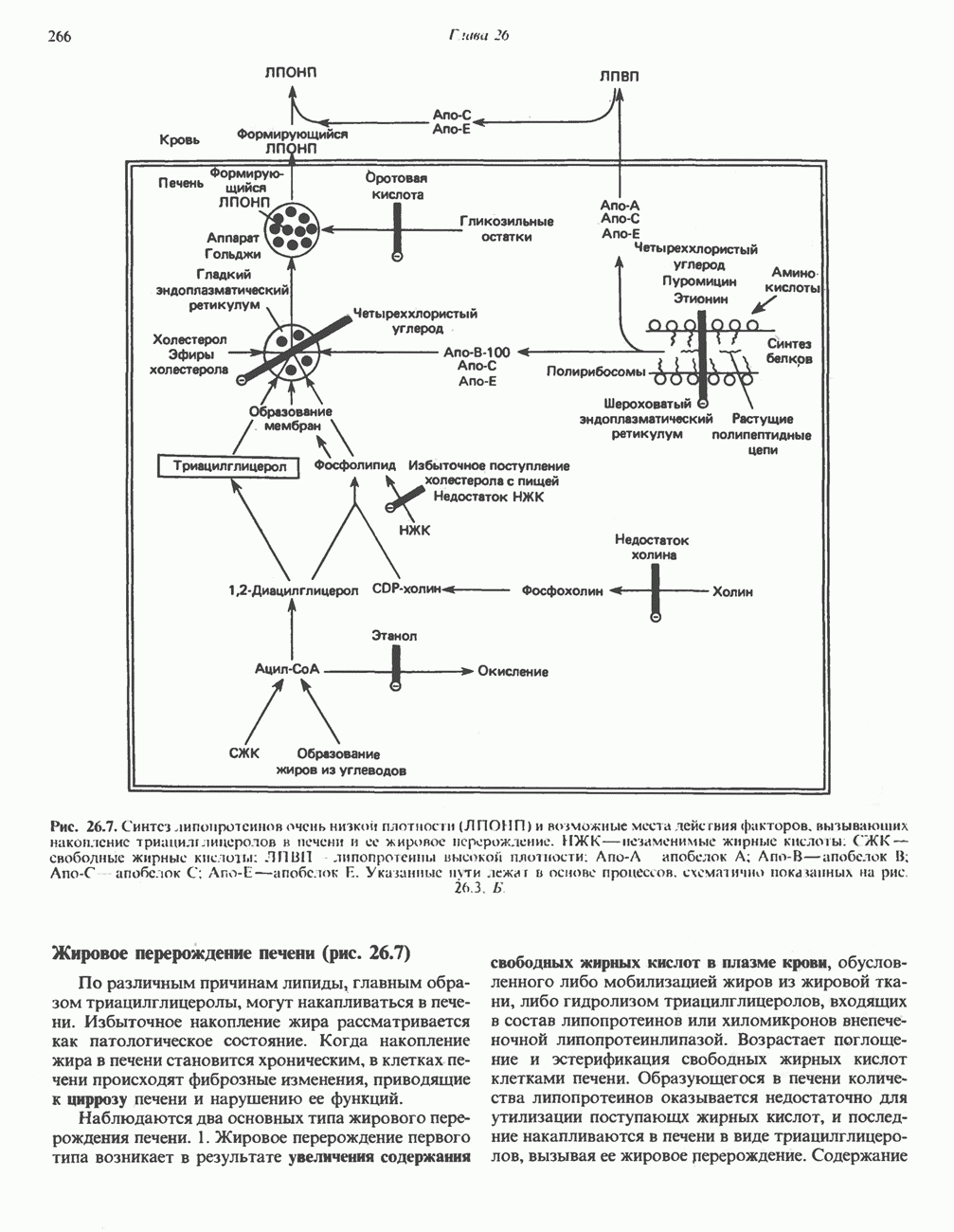
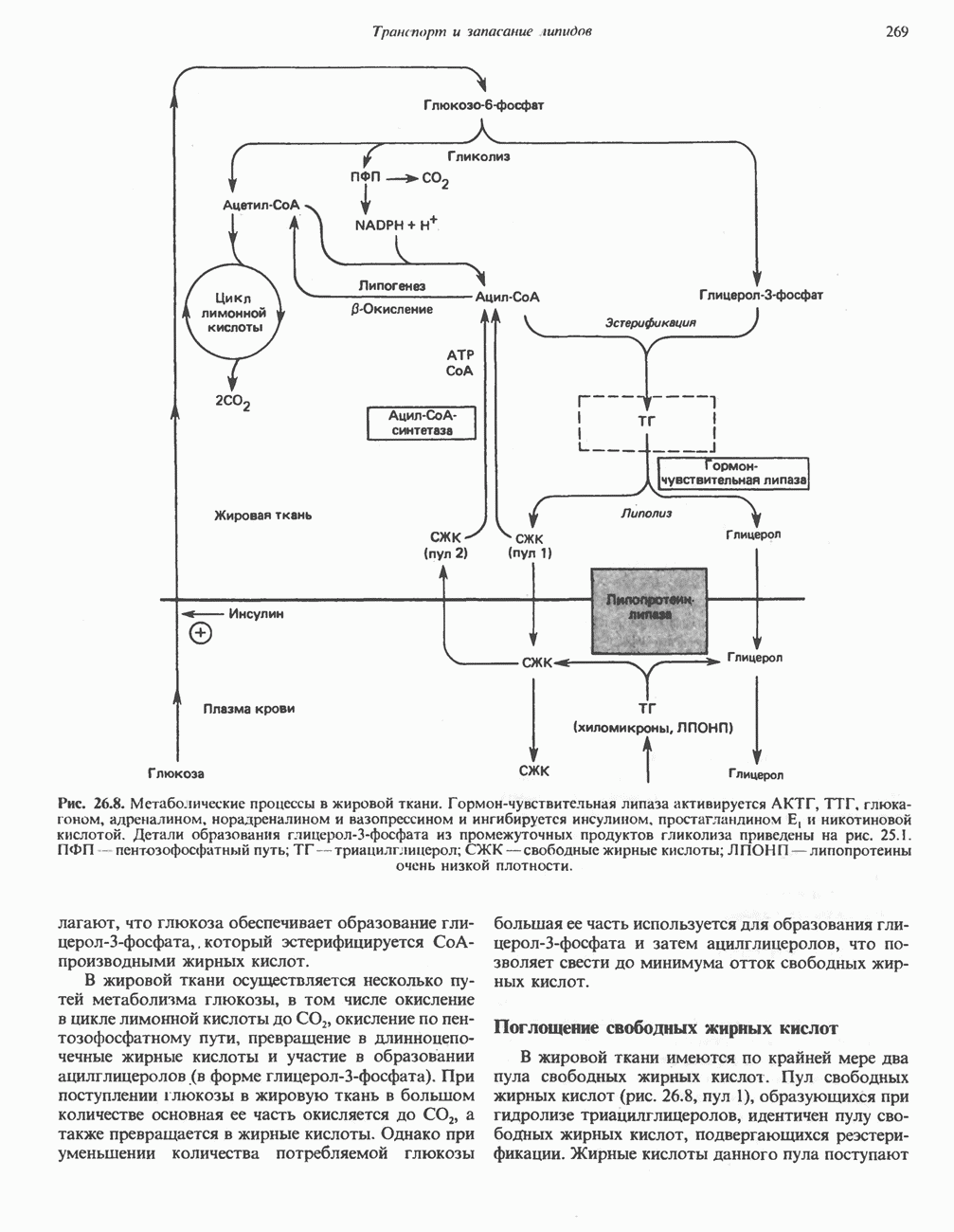
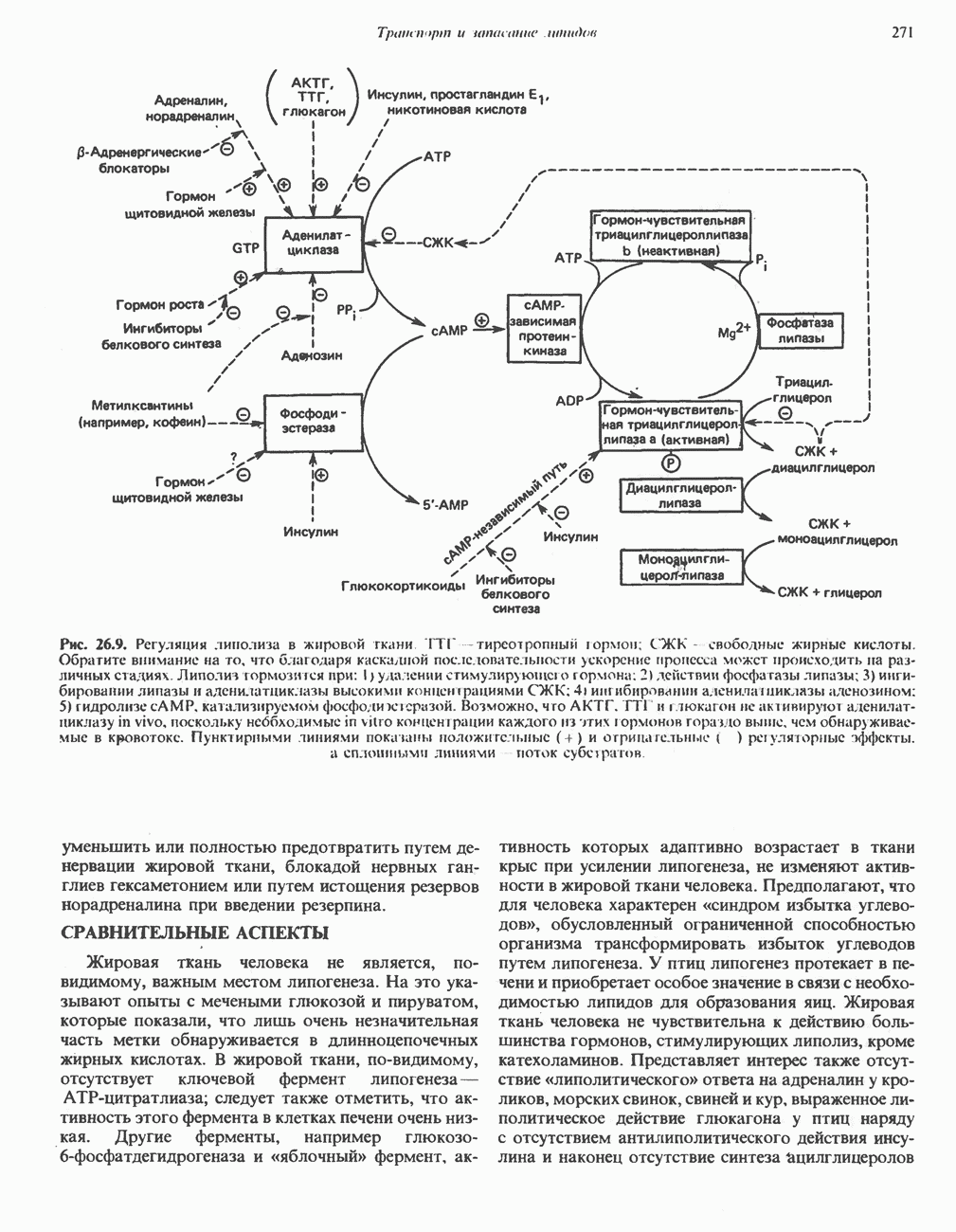
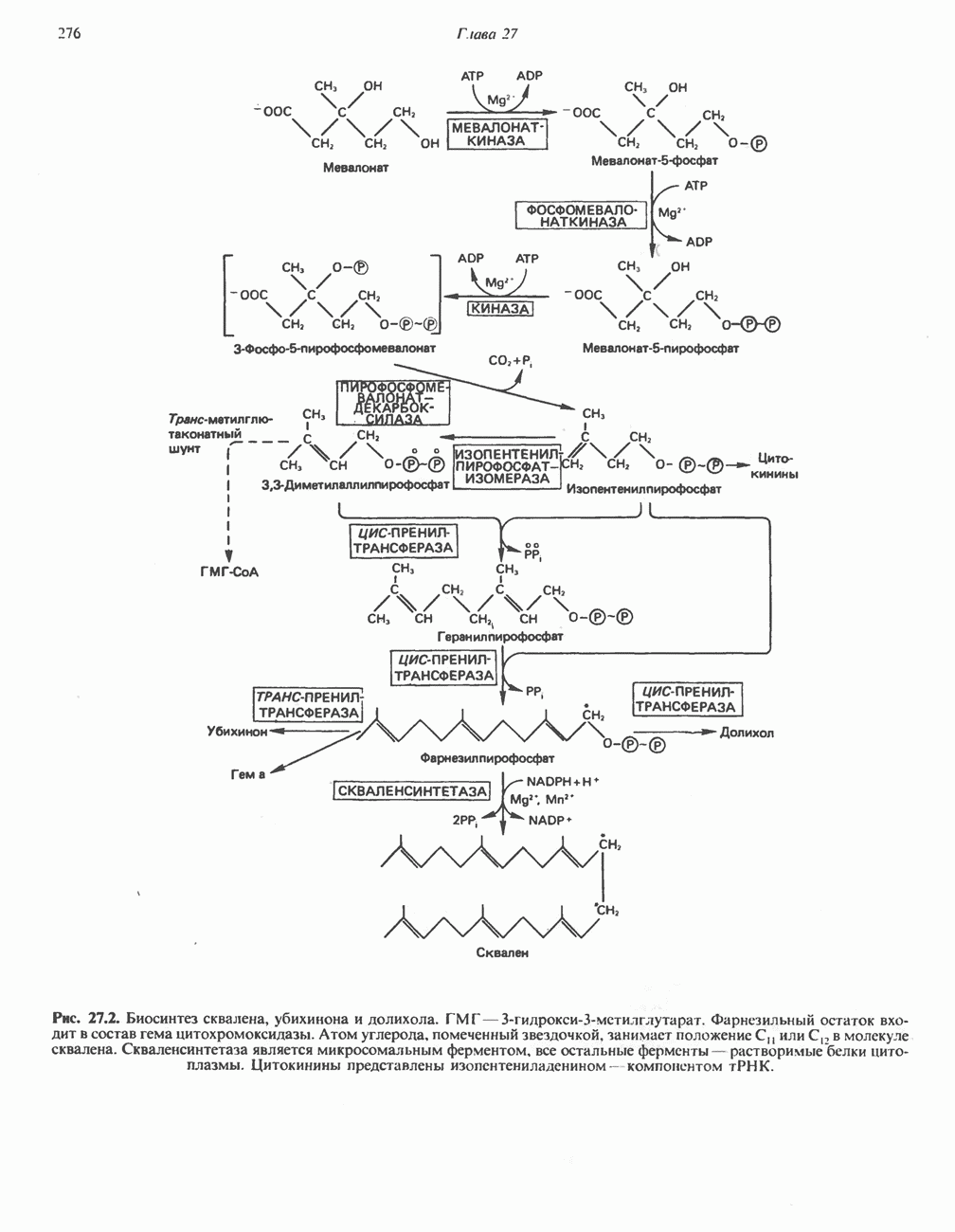
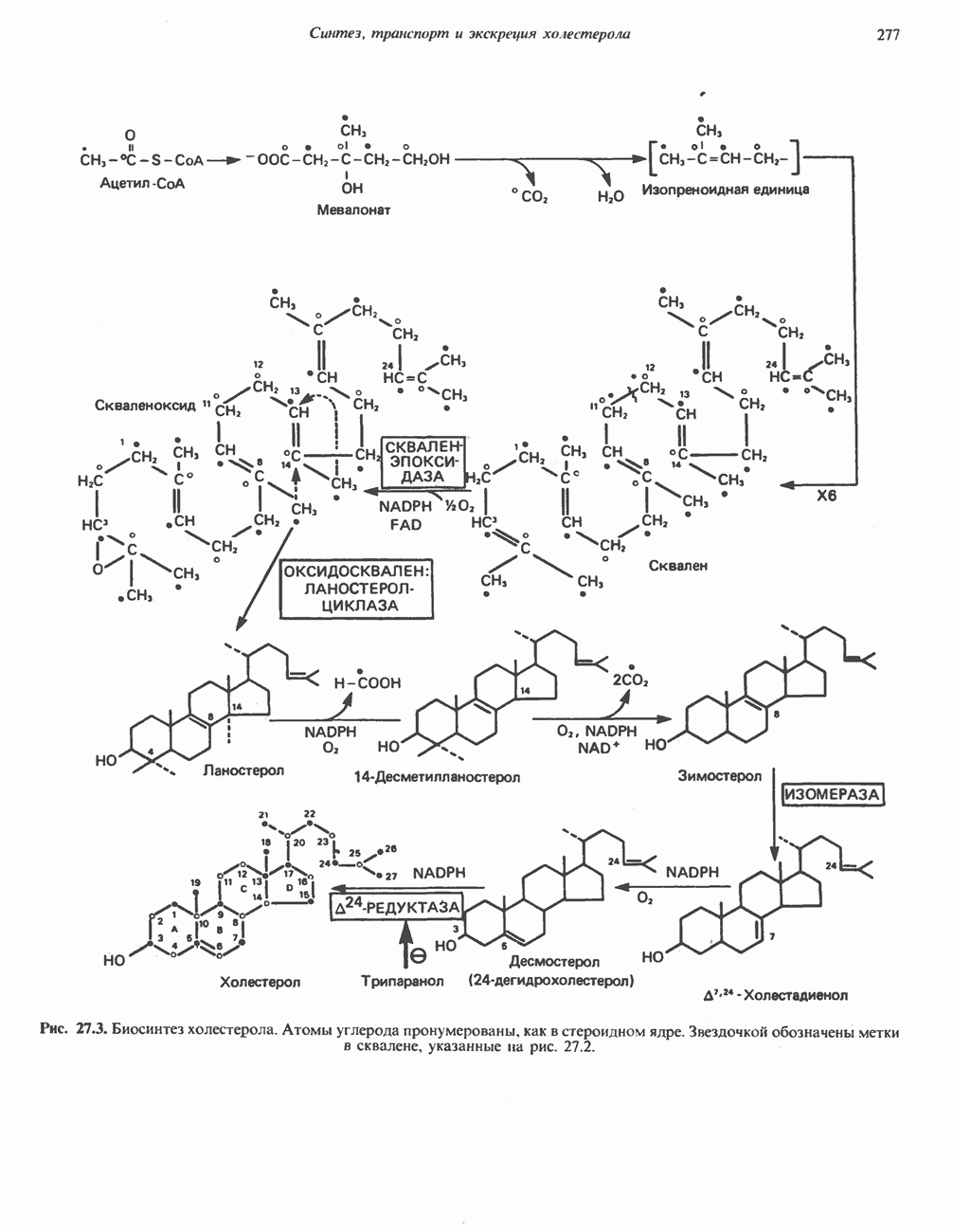
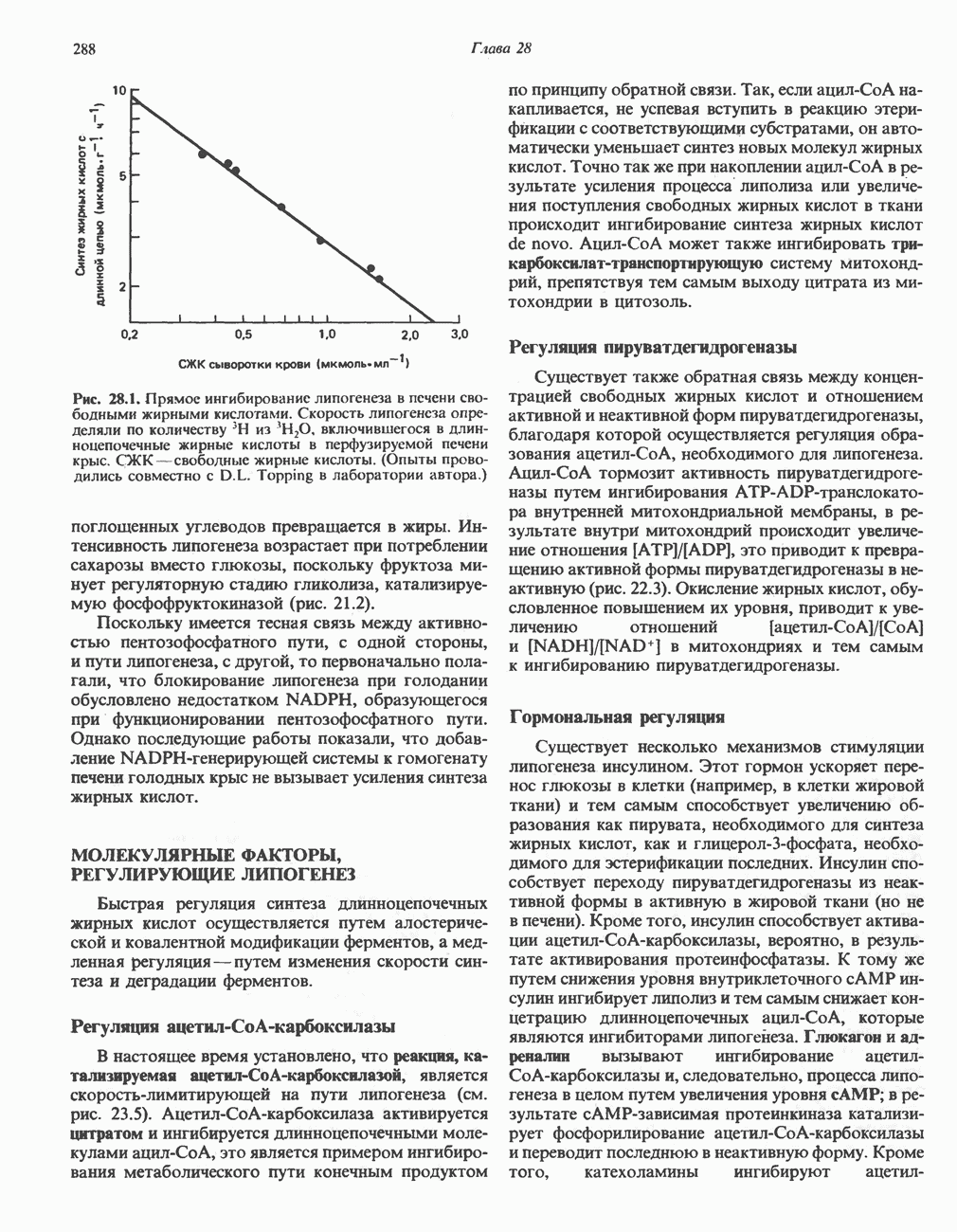
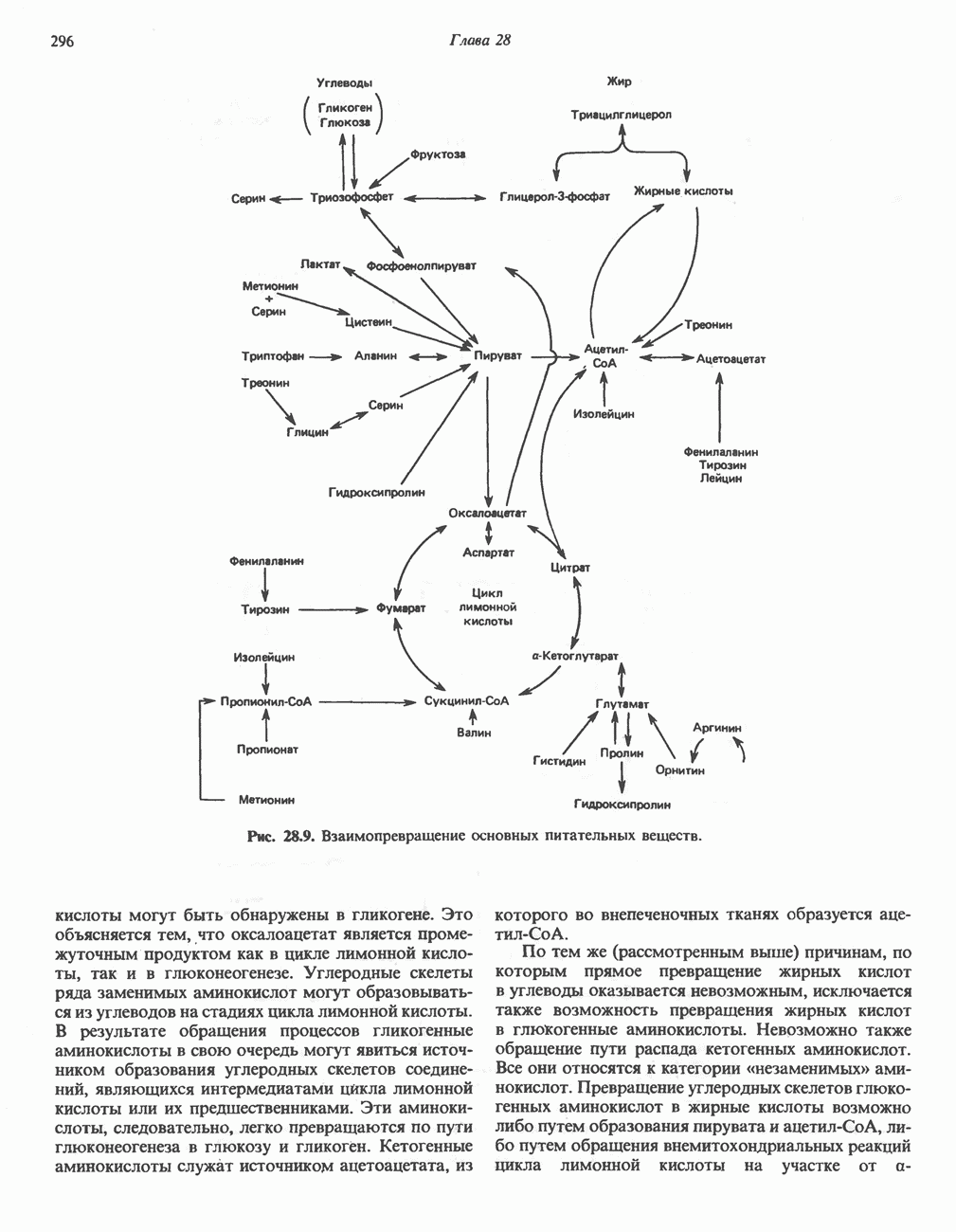
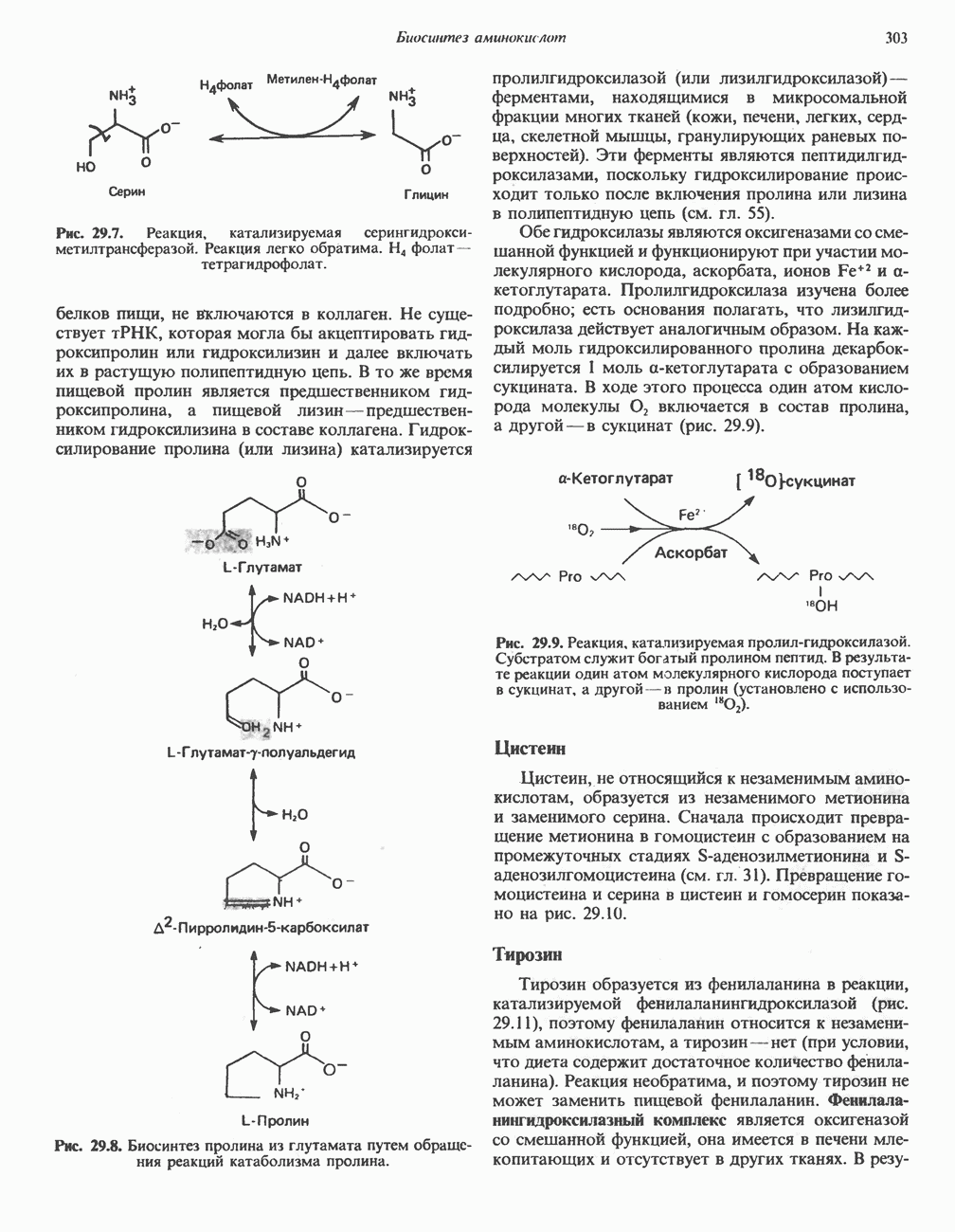
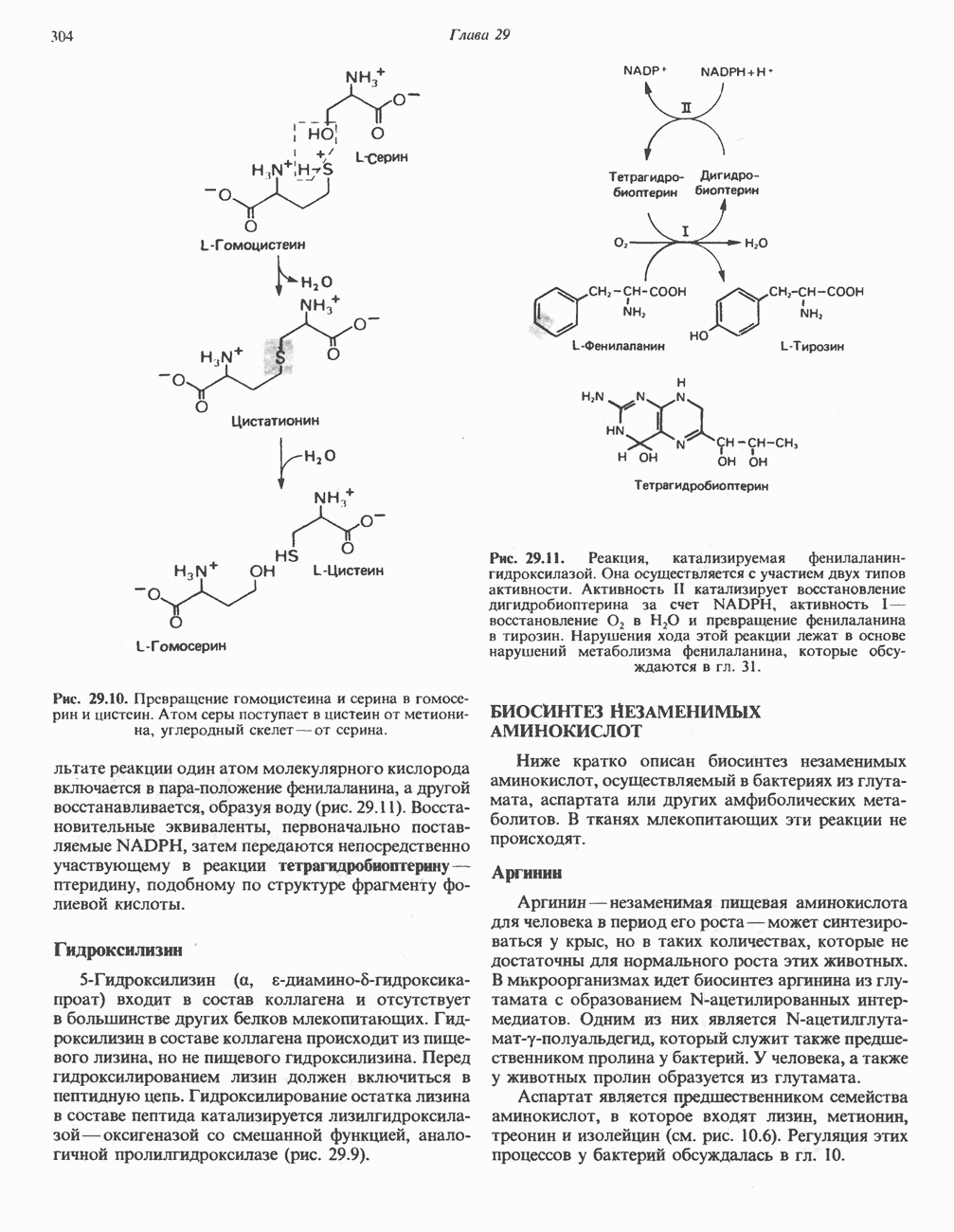
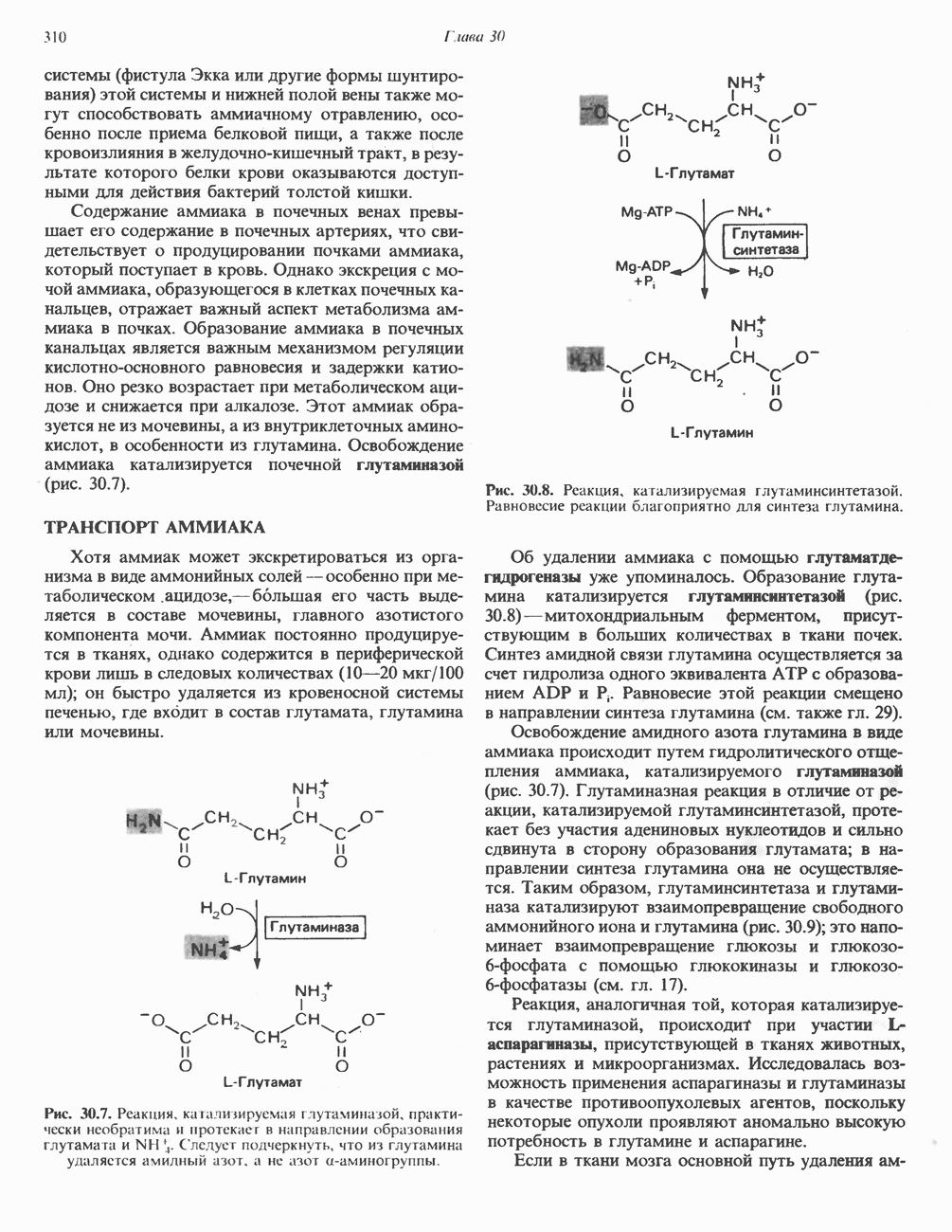
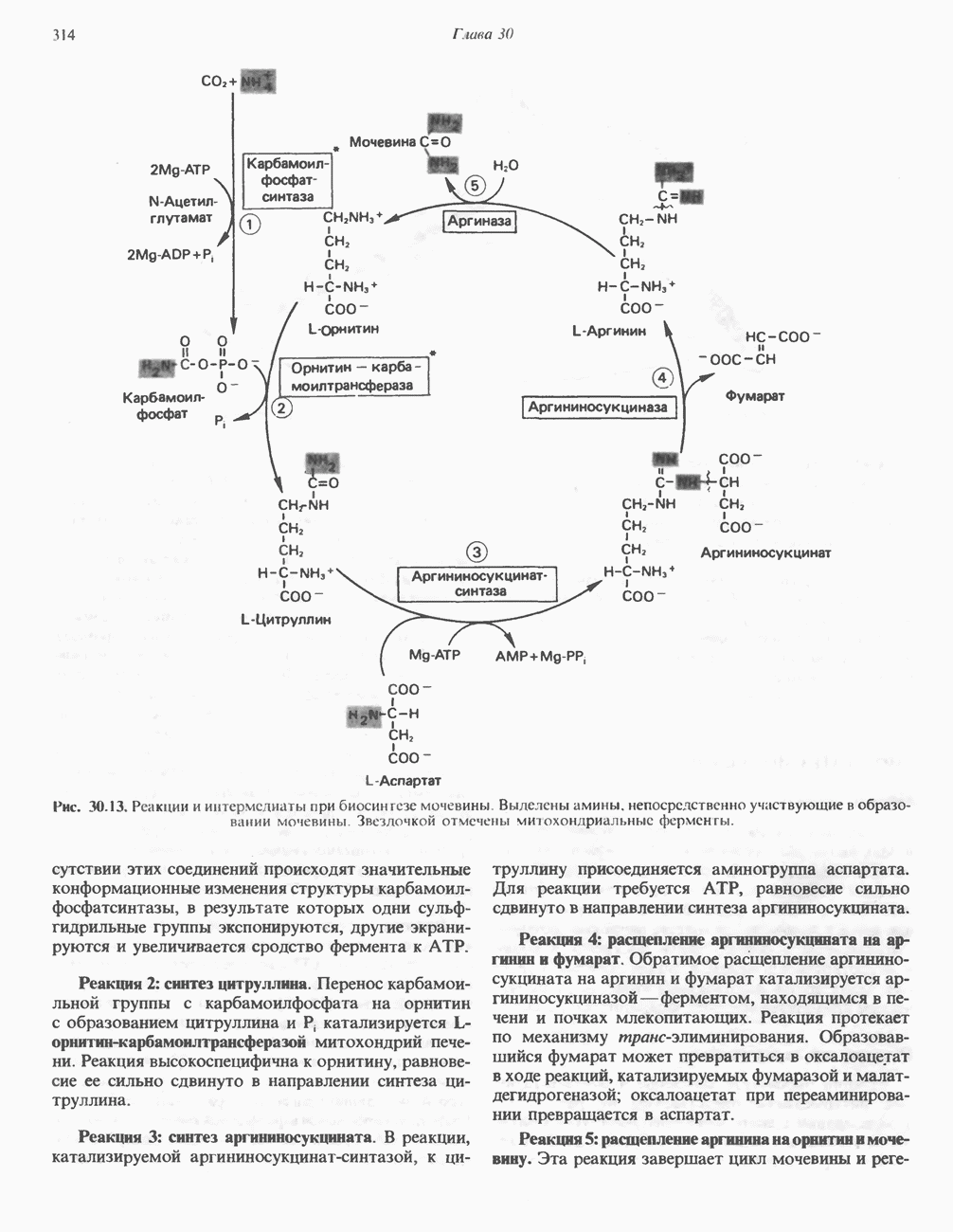
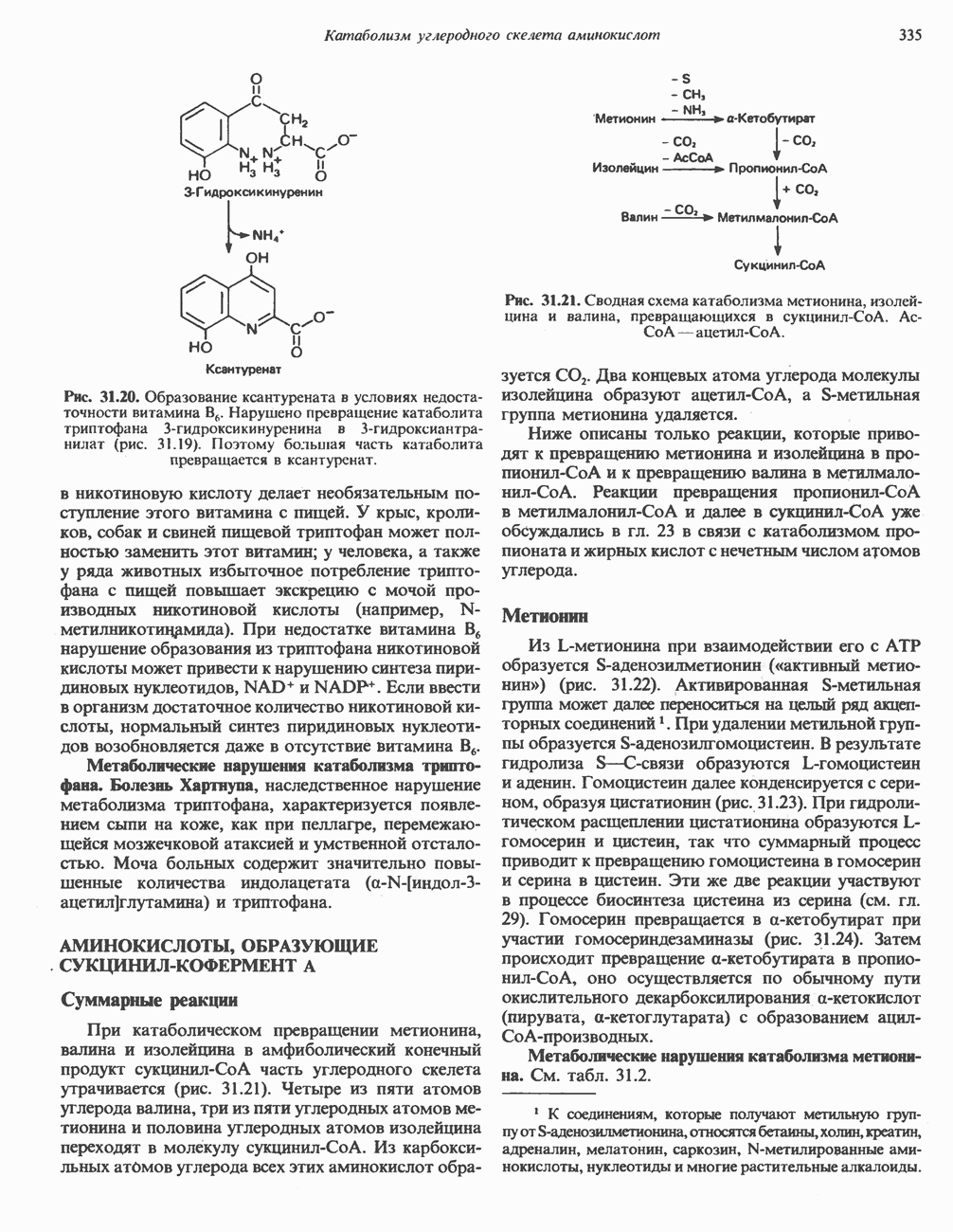
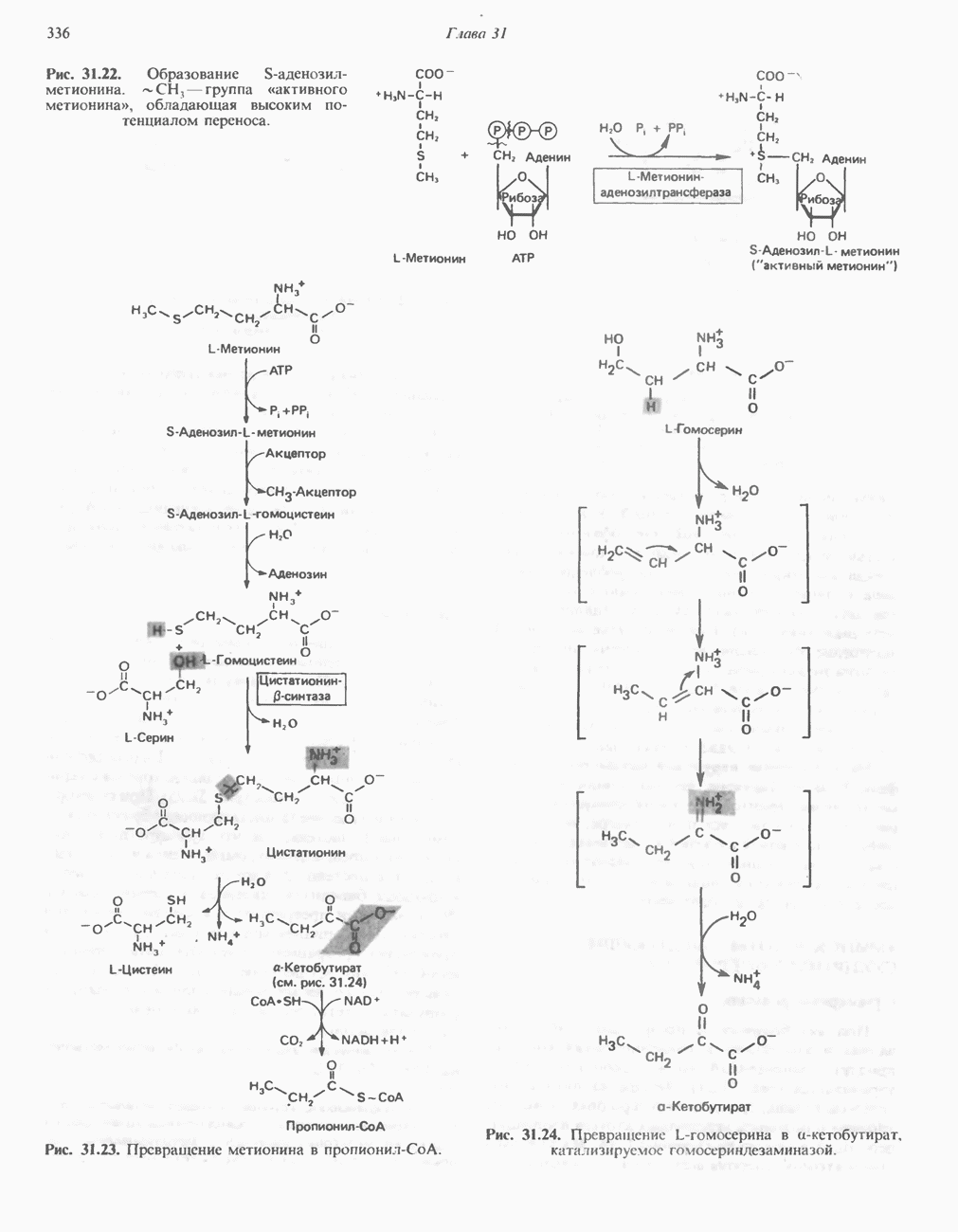
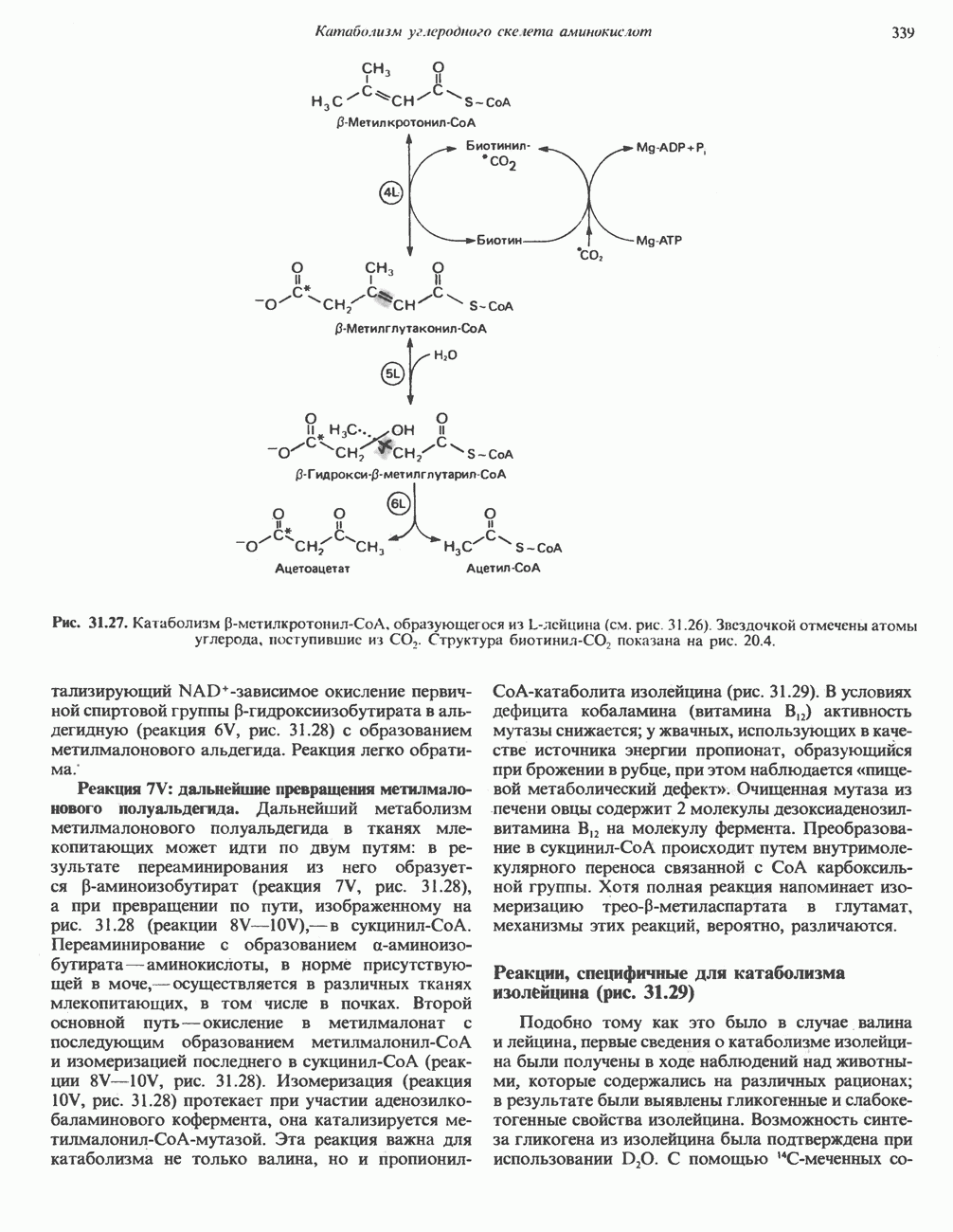
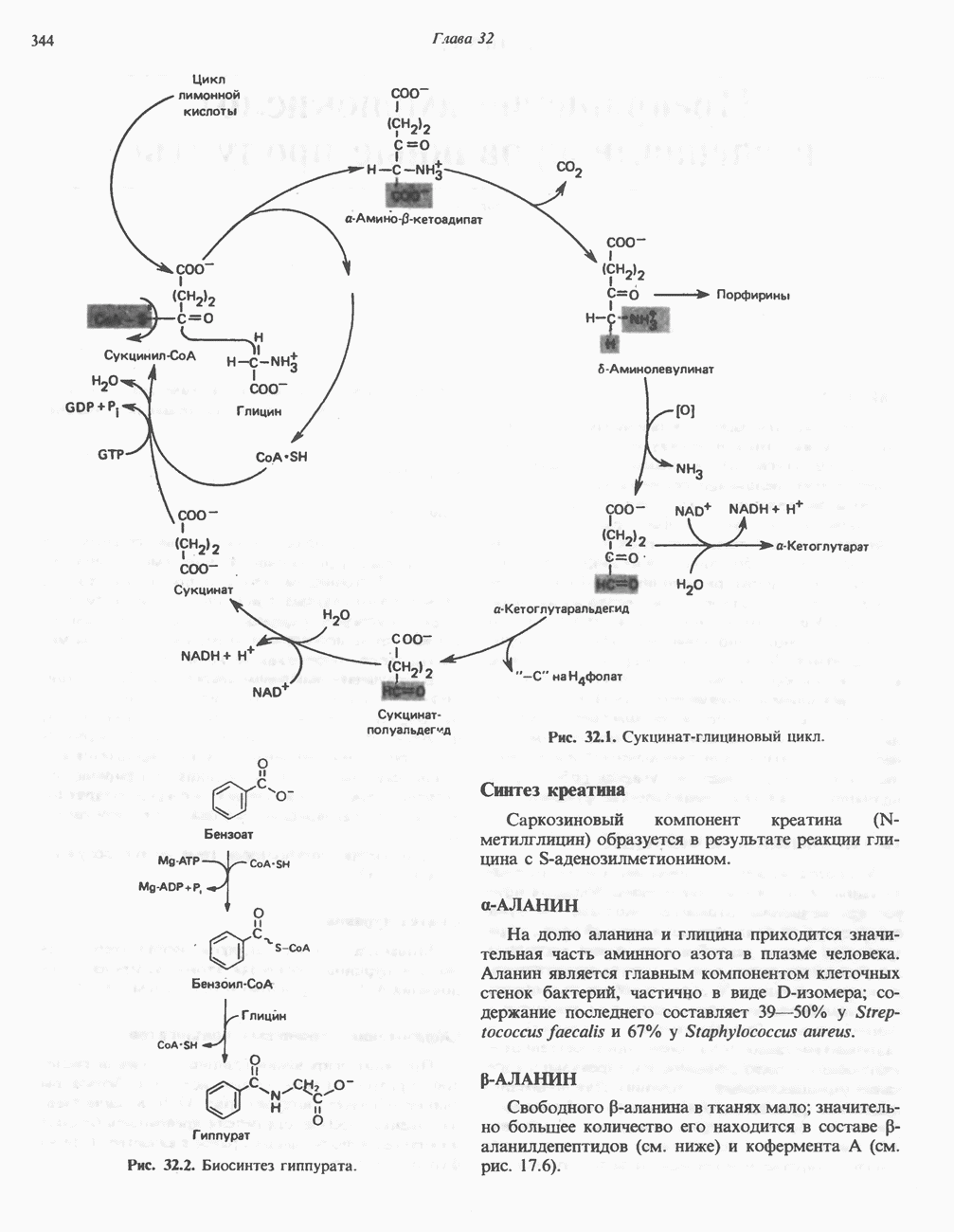
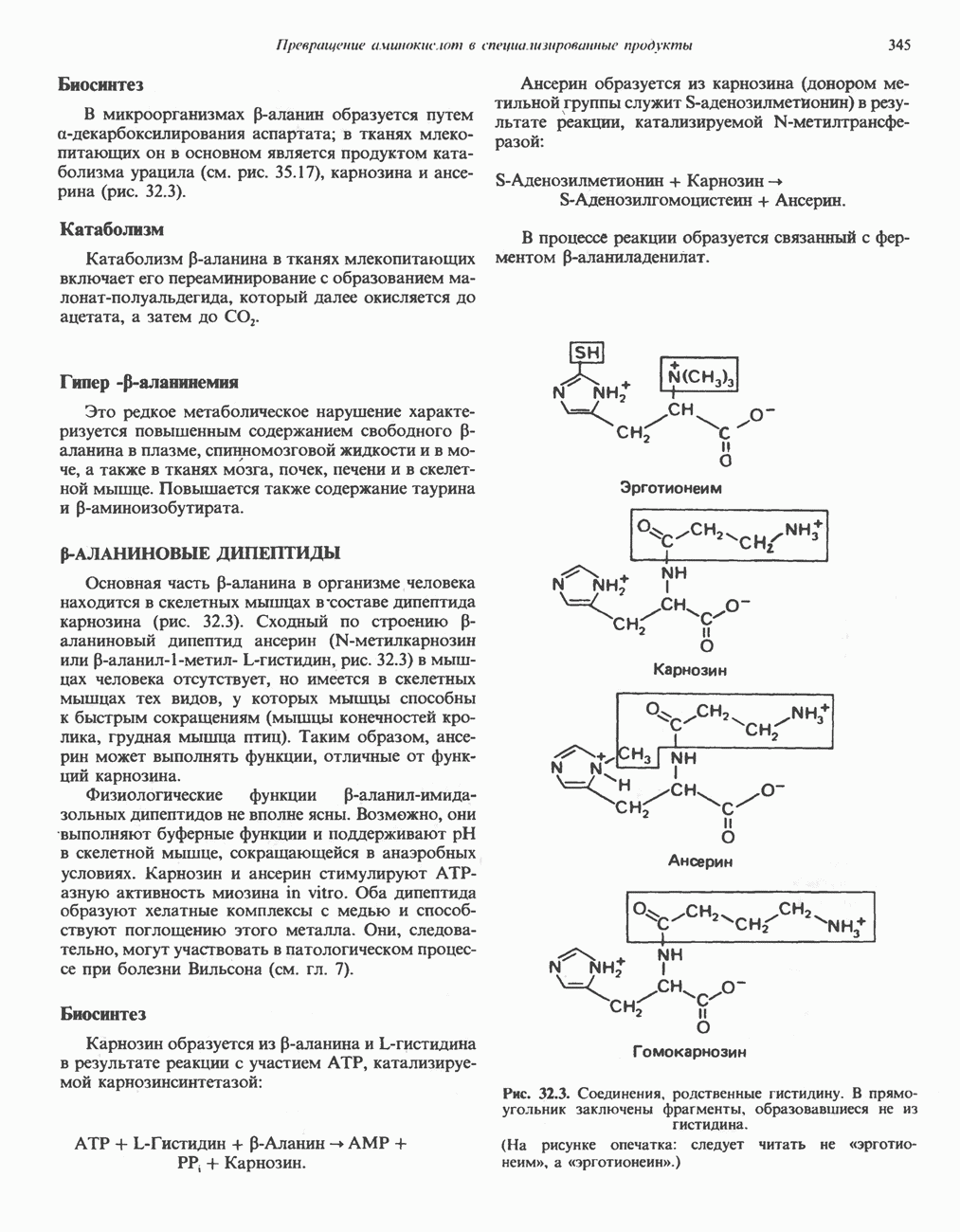
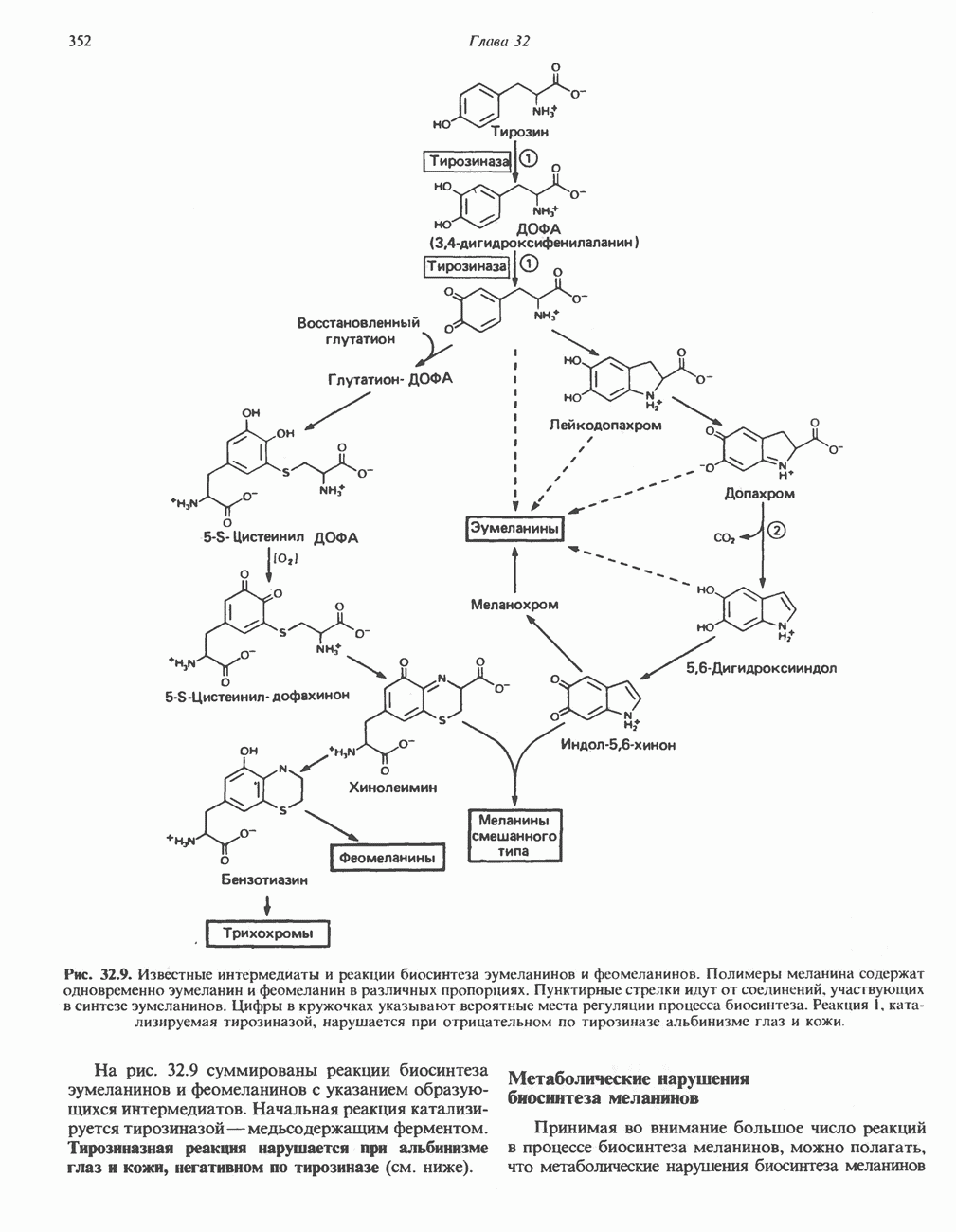
АМИНОКИСЛОТЫ, ОБРАЗУЮЩИЕ а-КЕТОГЛУТАРАТ
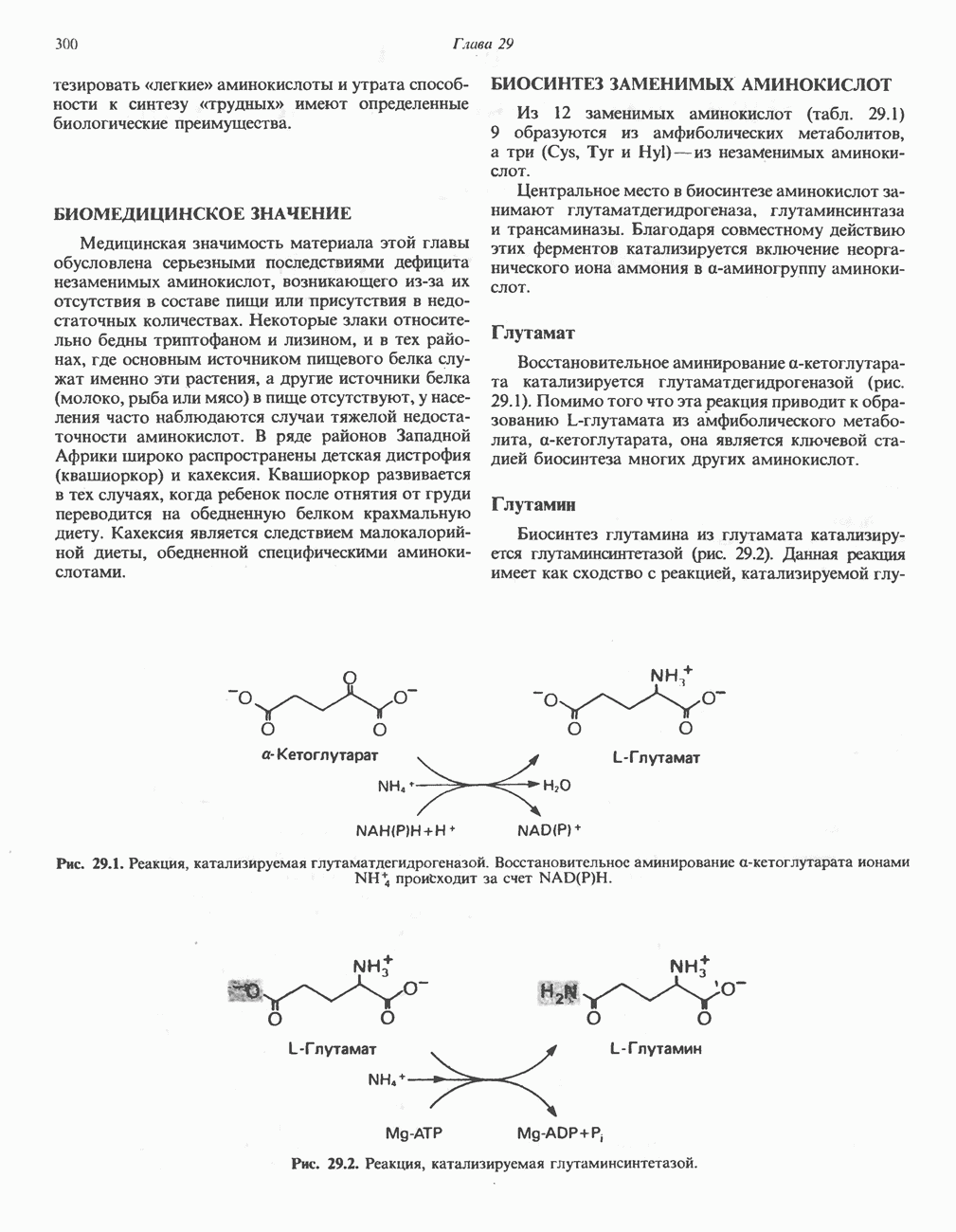
Глутамин и глутамат
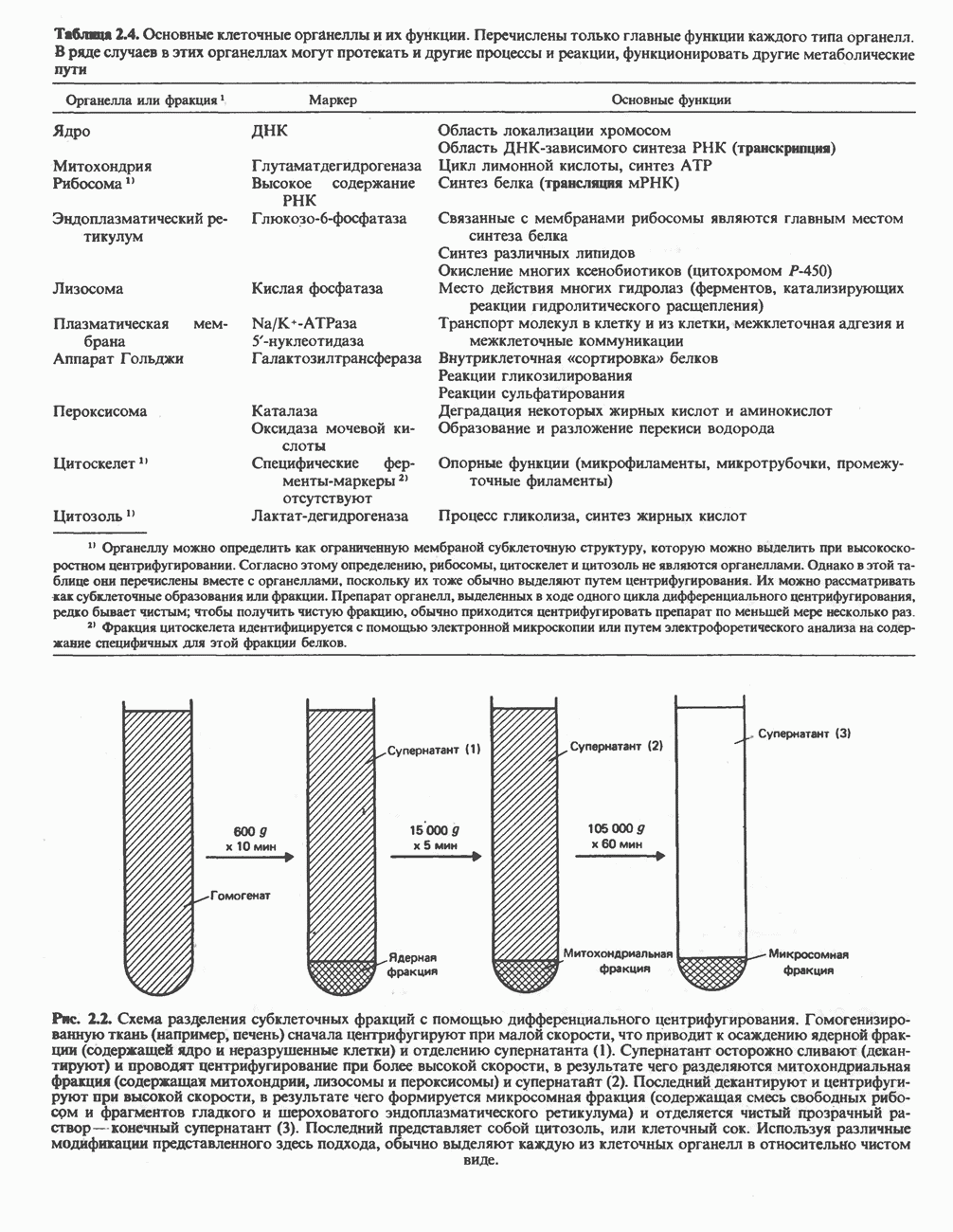
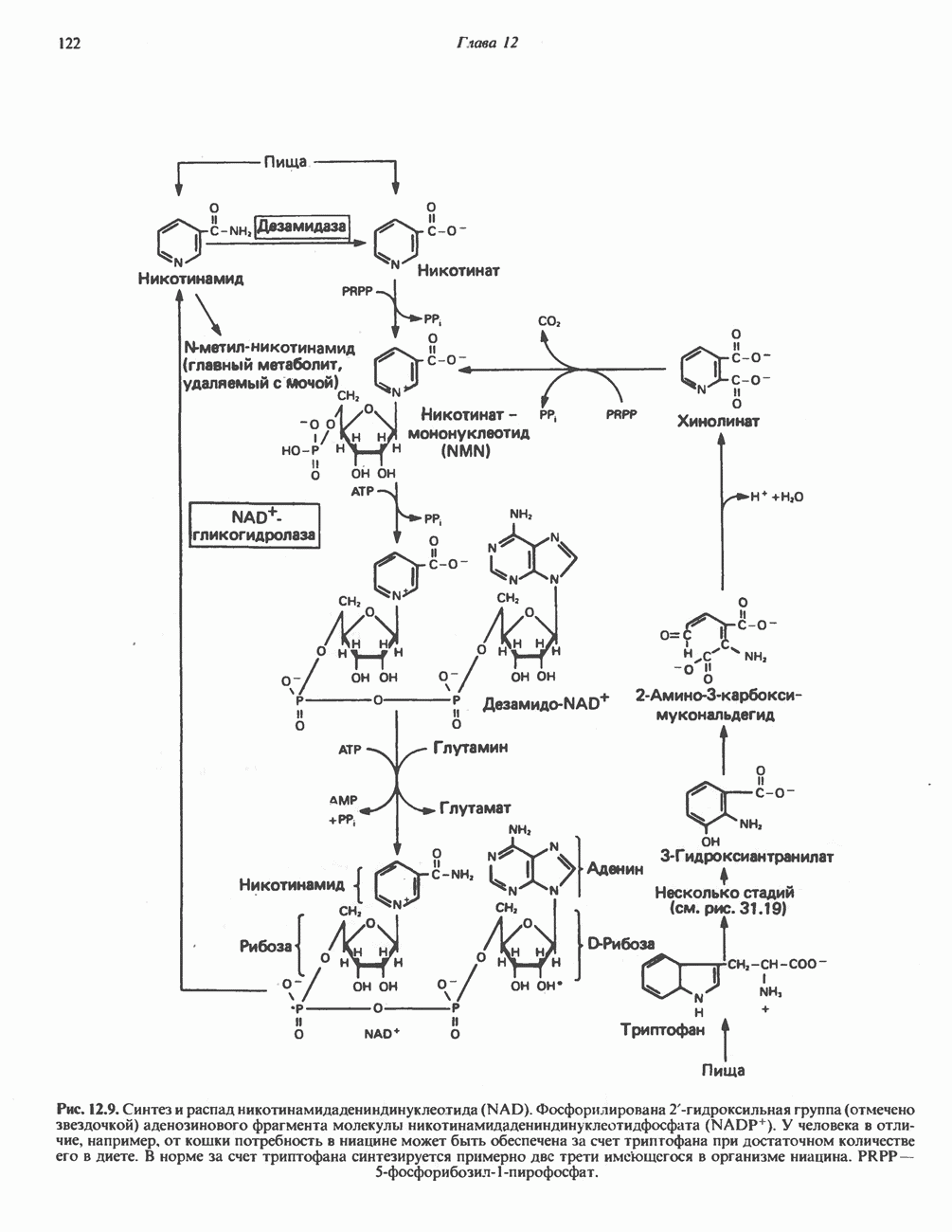
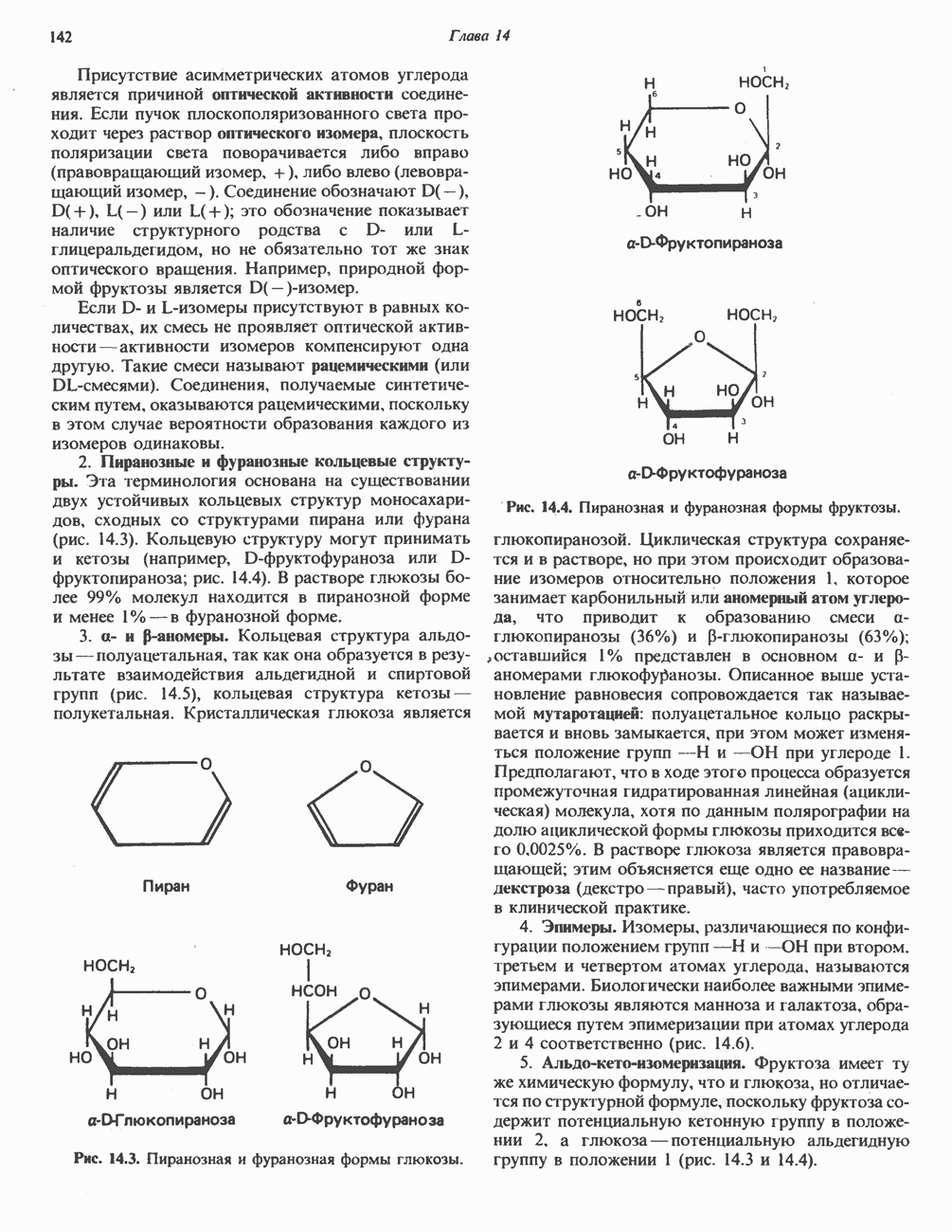
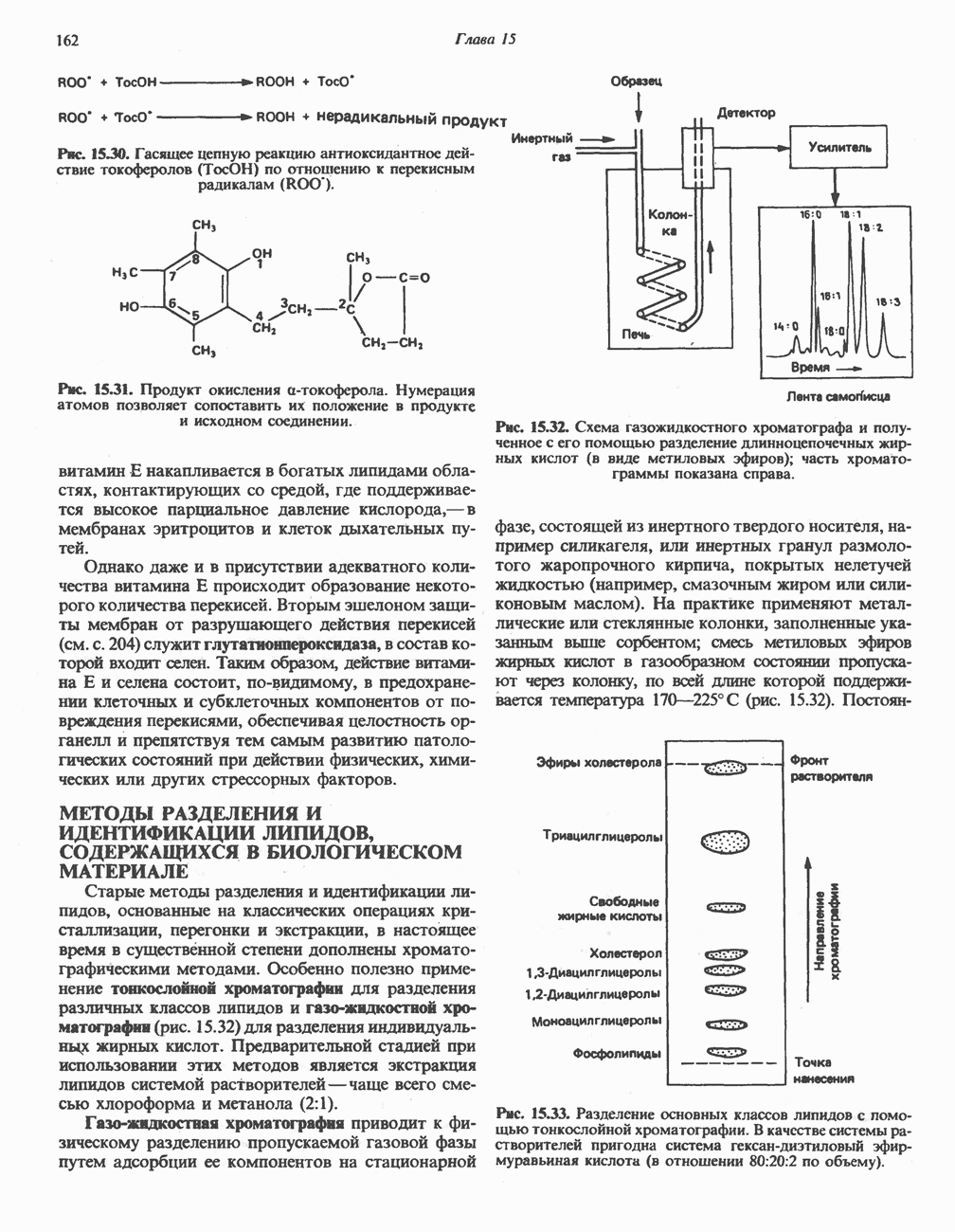
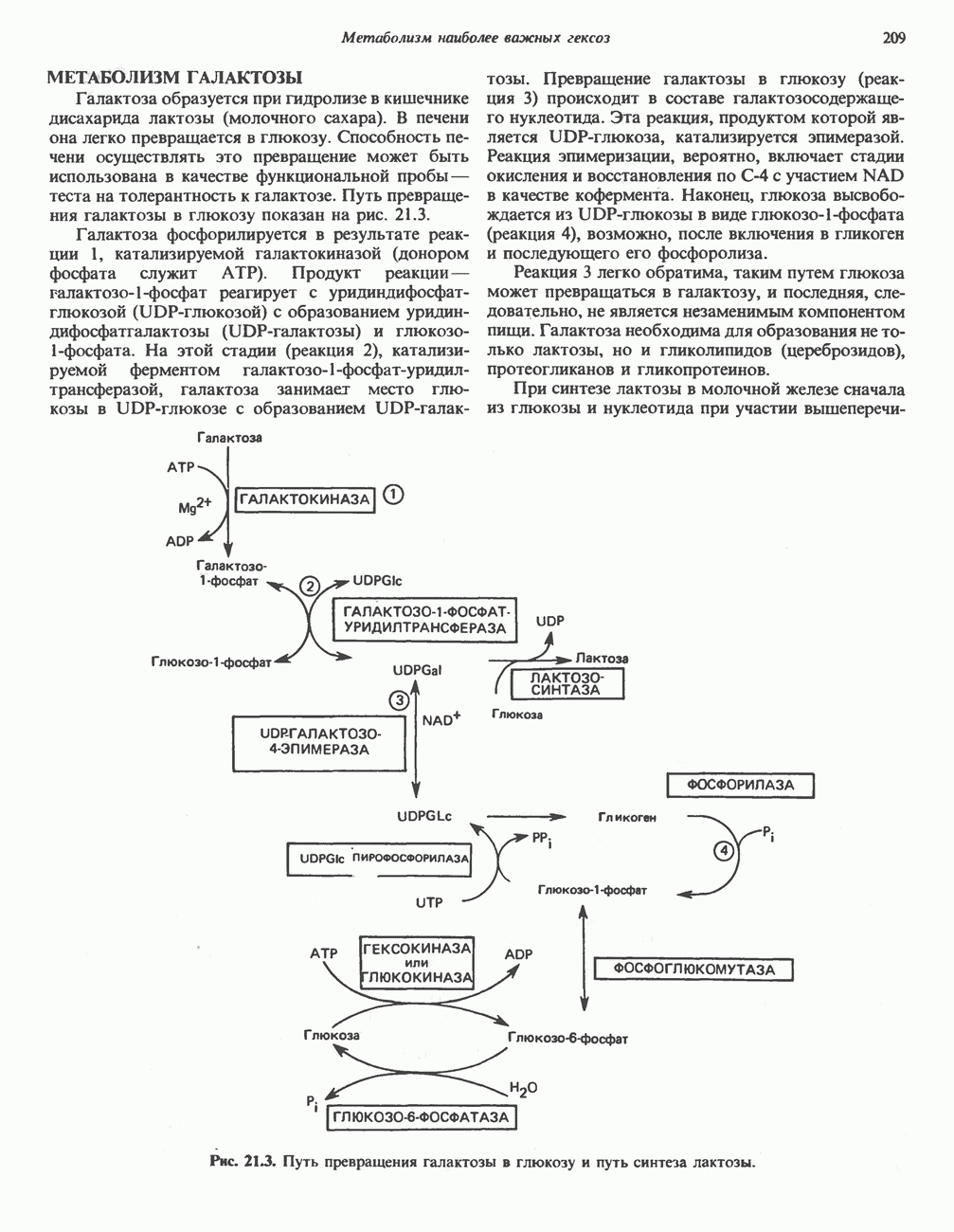
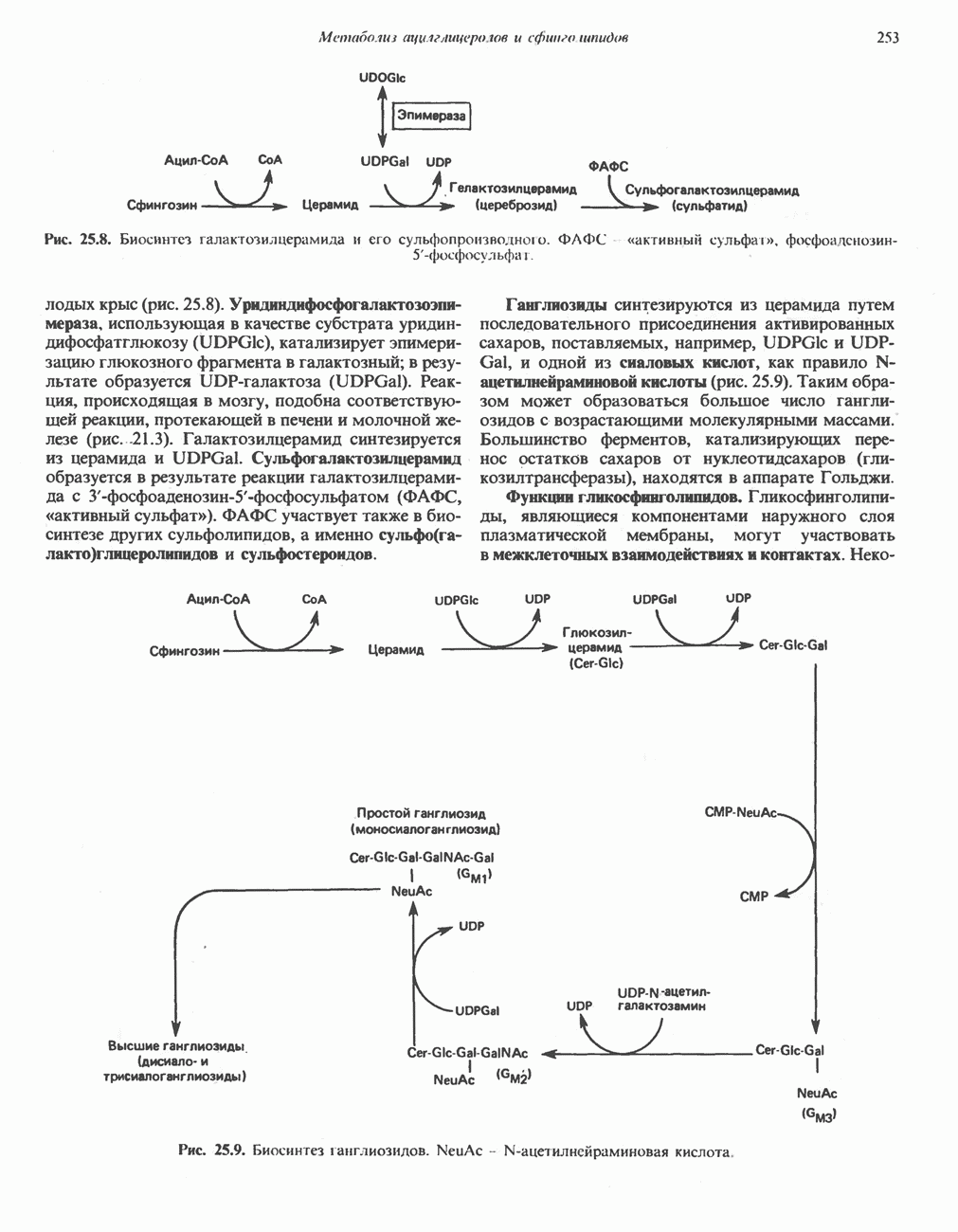
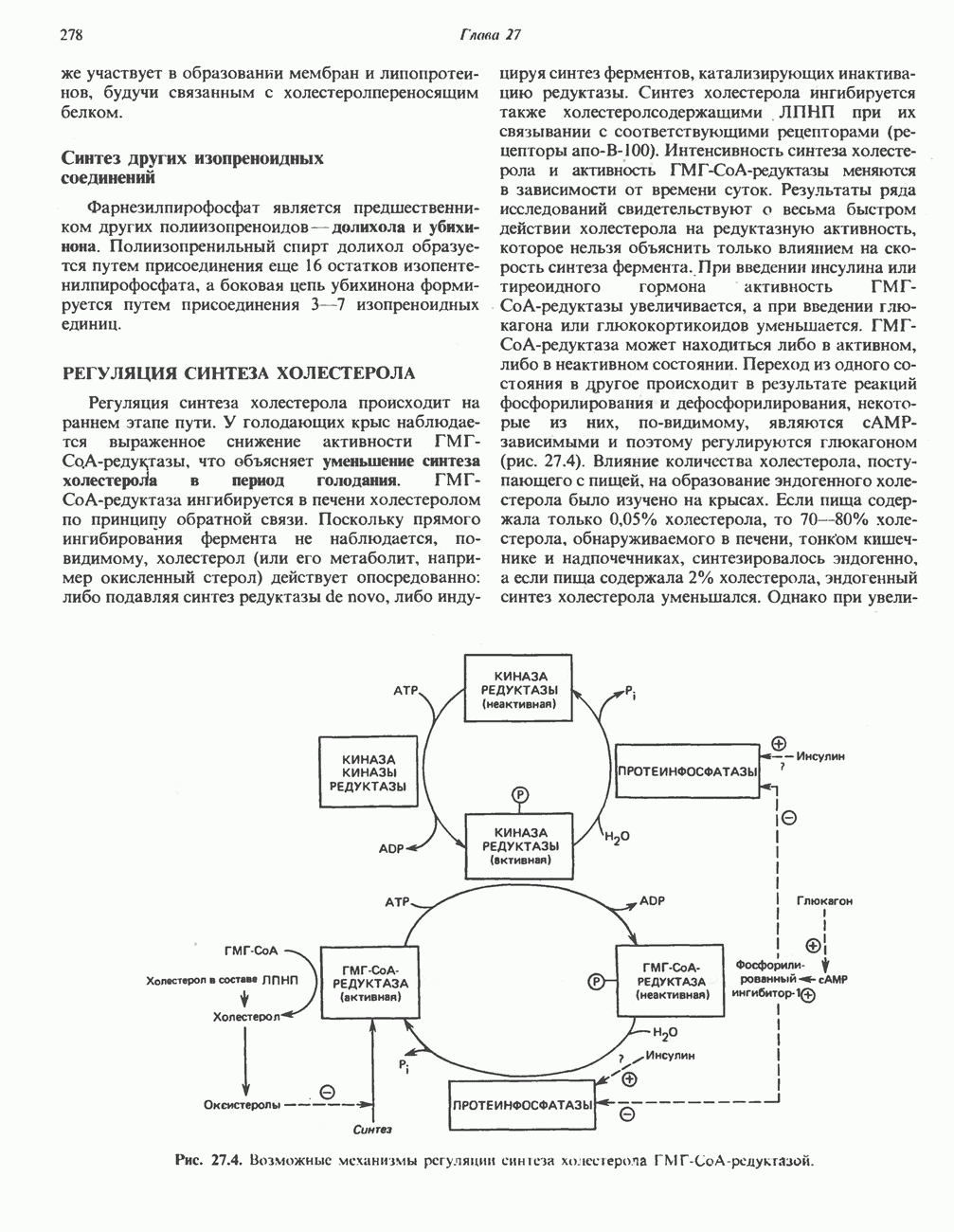
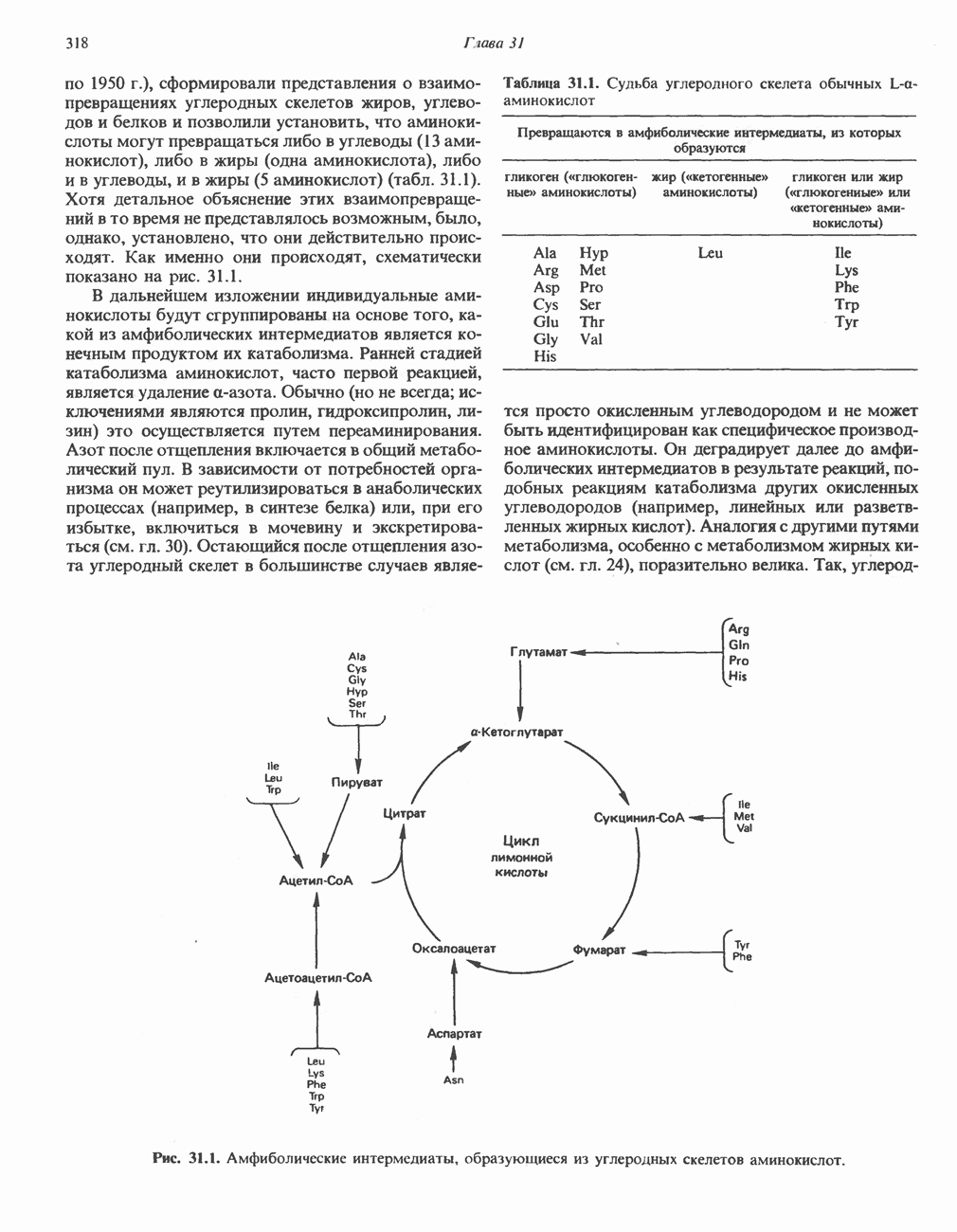
Катаболизм глутамина и глутамата протекает подобно катаболизму аспарагина и аспартата, но с рбразованием а-кетоглутарата—метиленового гомолога оксалоацетата (рис. 31.2, нижняя часть). В то время как глутамат и аспартат являются субстратами одной и той же трансаминазы, дезамидирование аспарагина и глутамина осуществляется различными ферментами. Ферменты, обладающие двойной специфичностью (глутаминазной и аспарагиназной),  некоторых бактепий.
некоторых бактепий.
Возможно, по тем же причинам, которые упоминались для аспарагина и аспартата, не зарегистрировано метаболических нарушений глутамин-глутаматного катаболического пути.
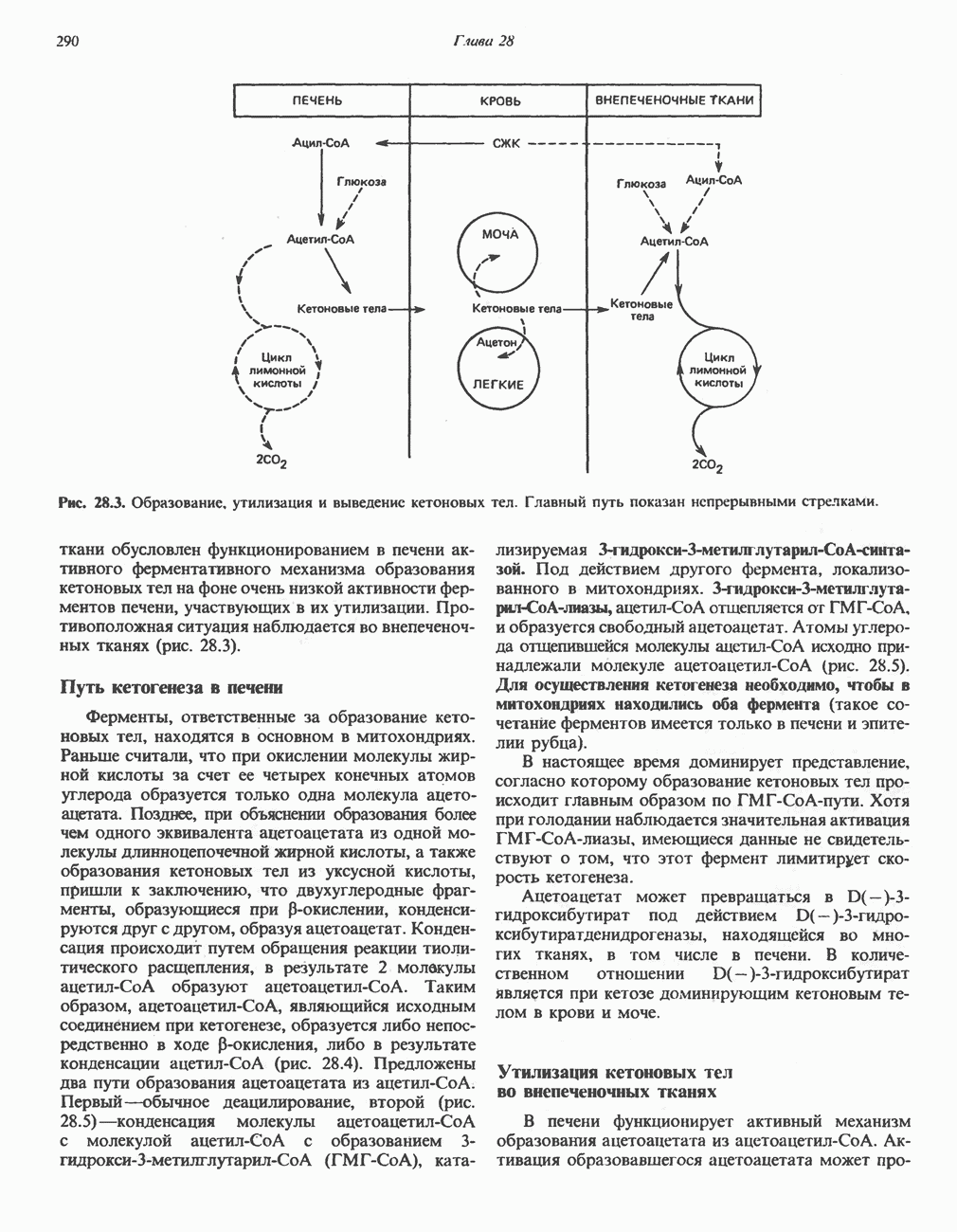
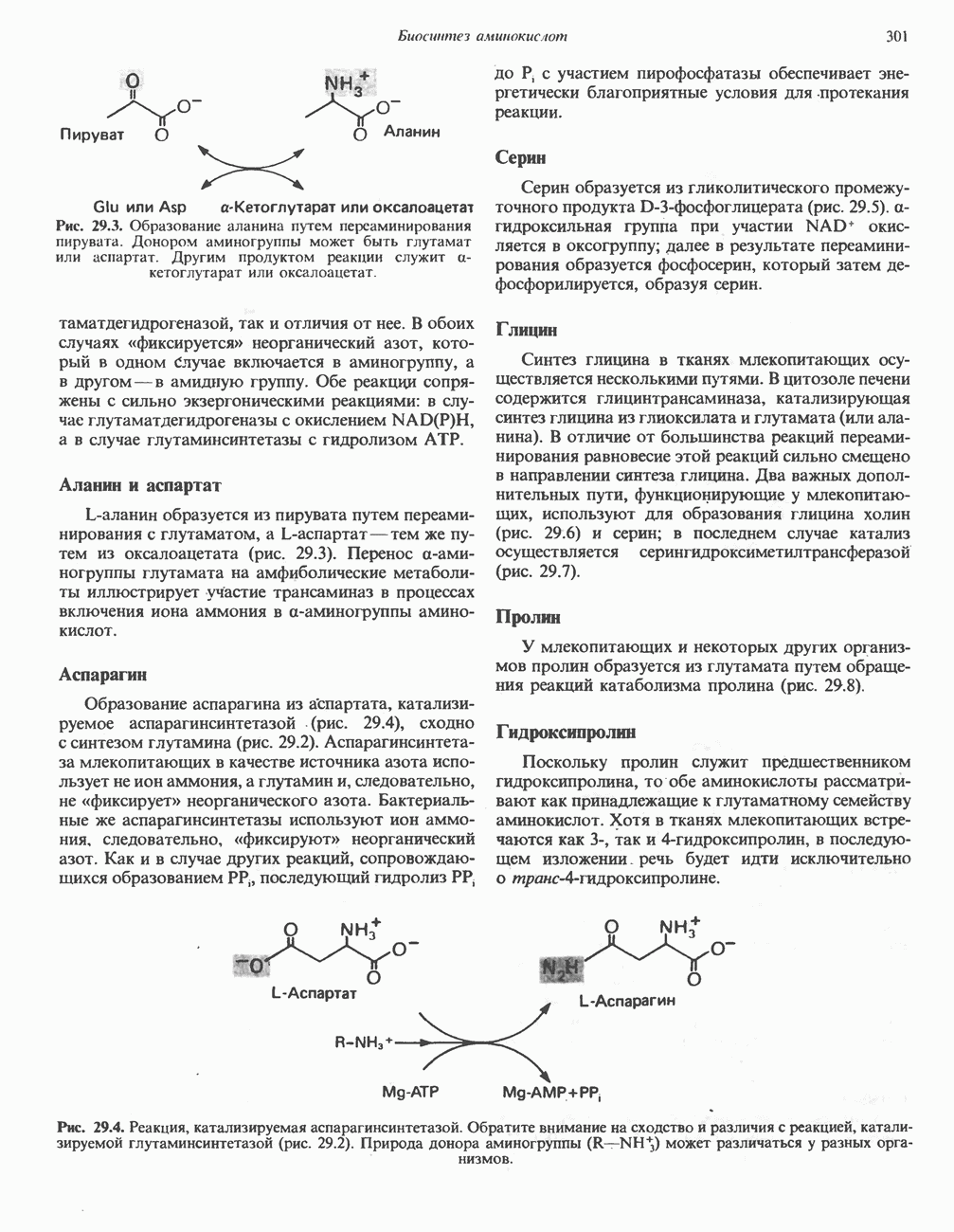
Пролин
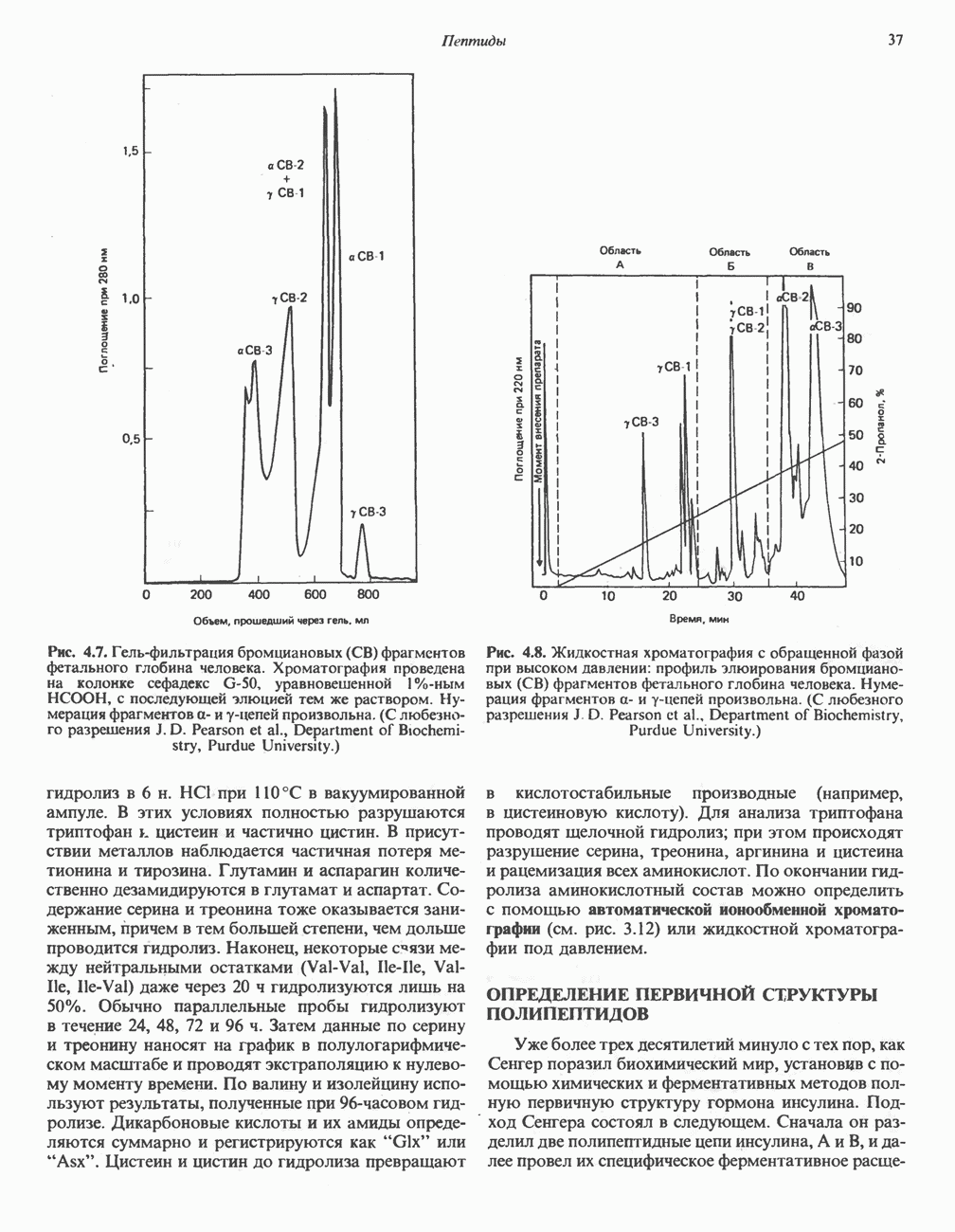
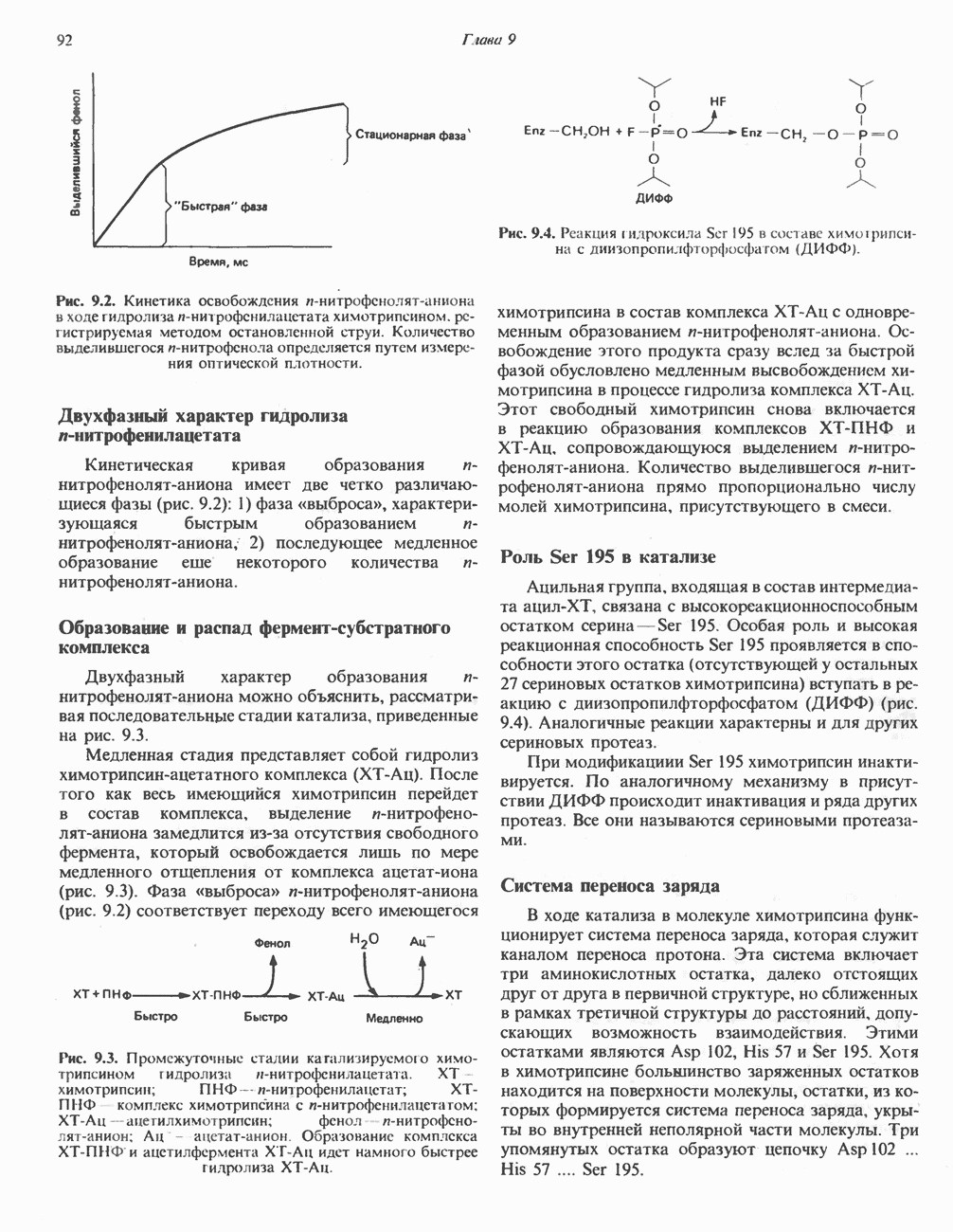
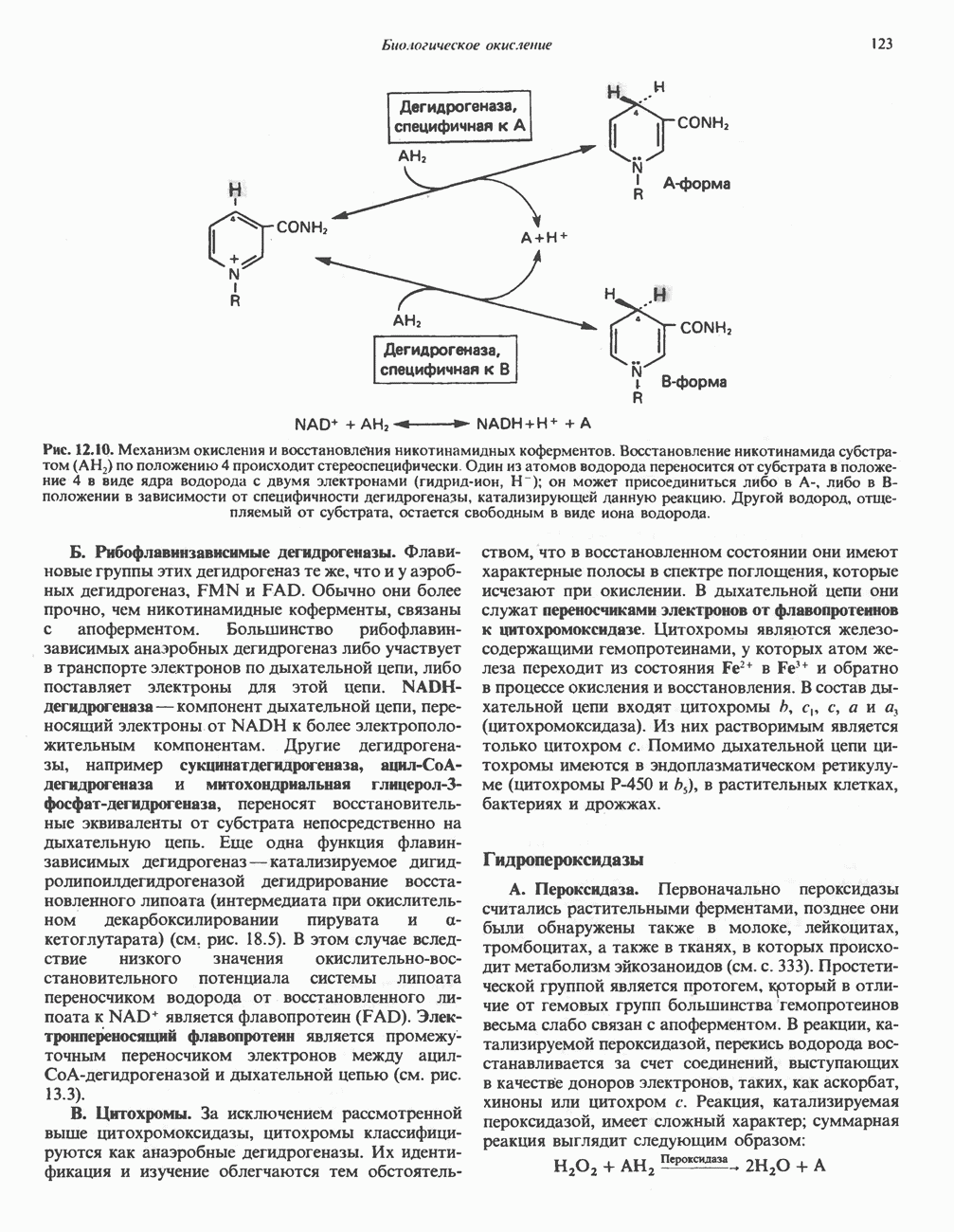
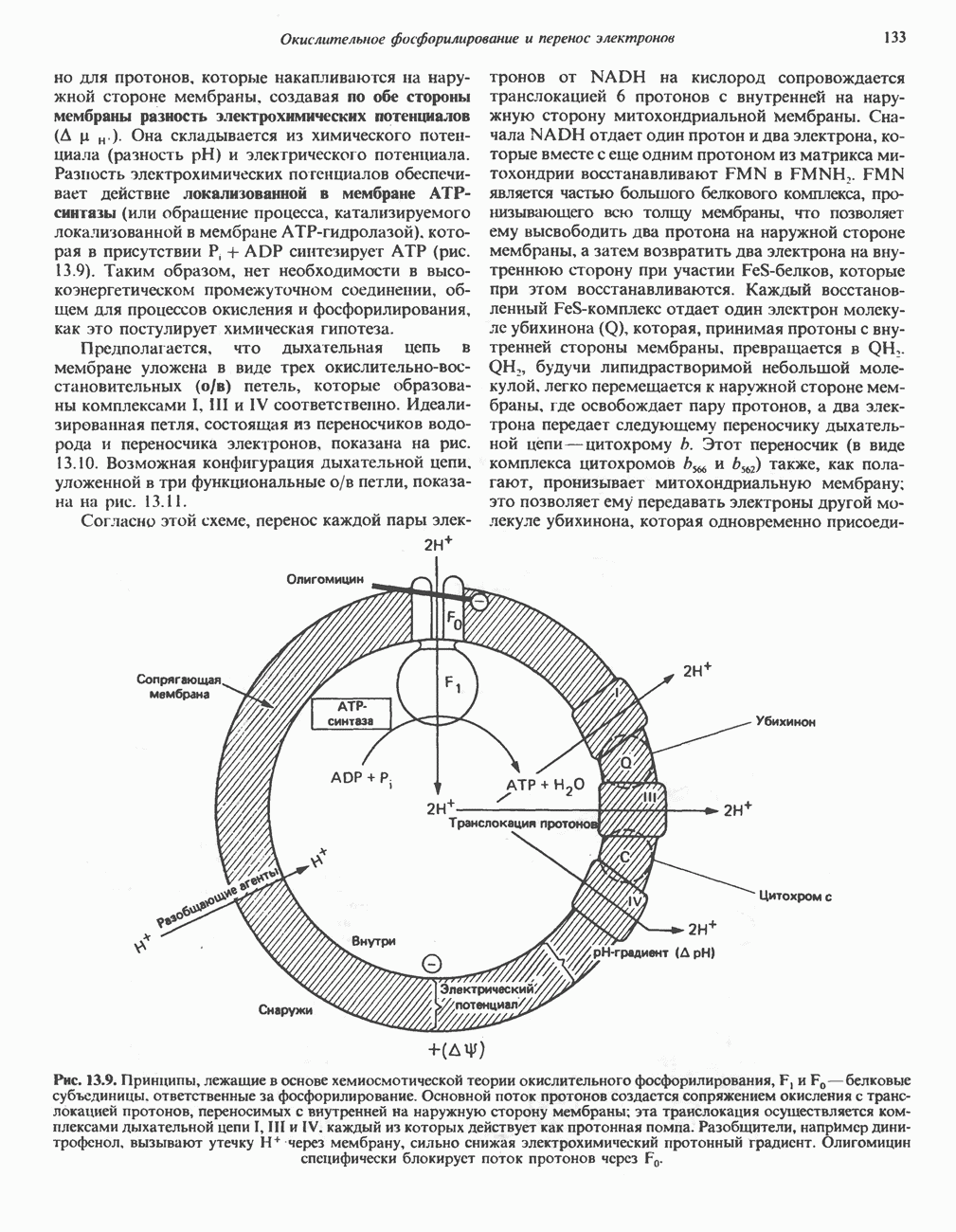
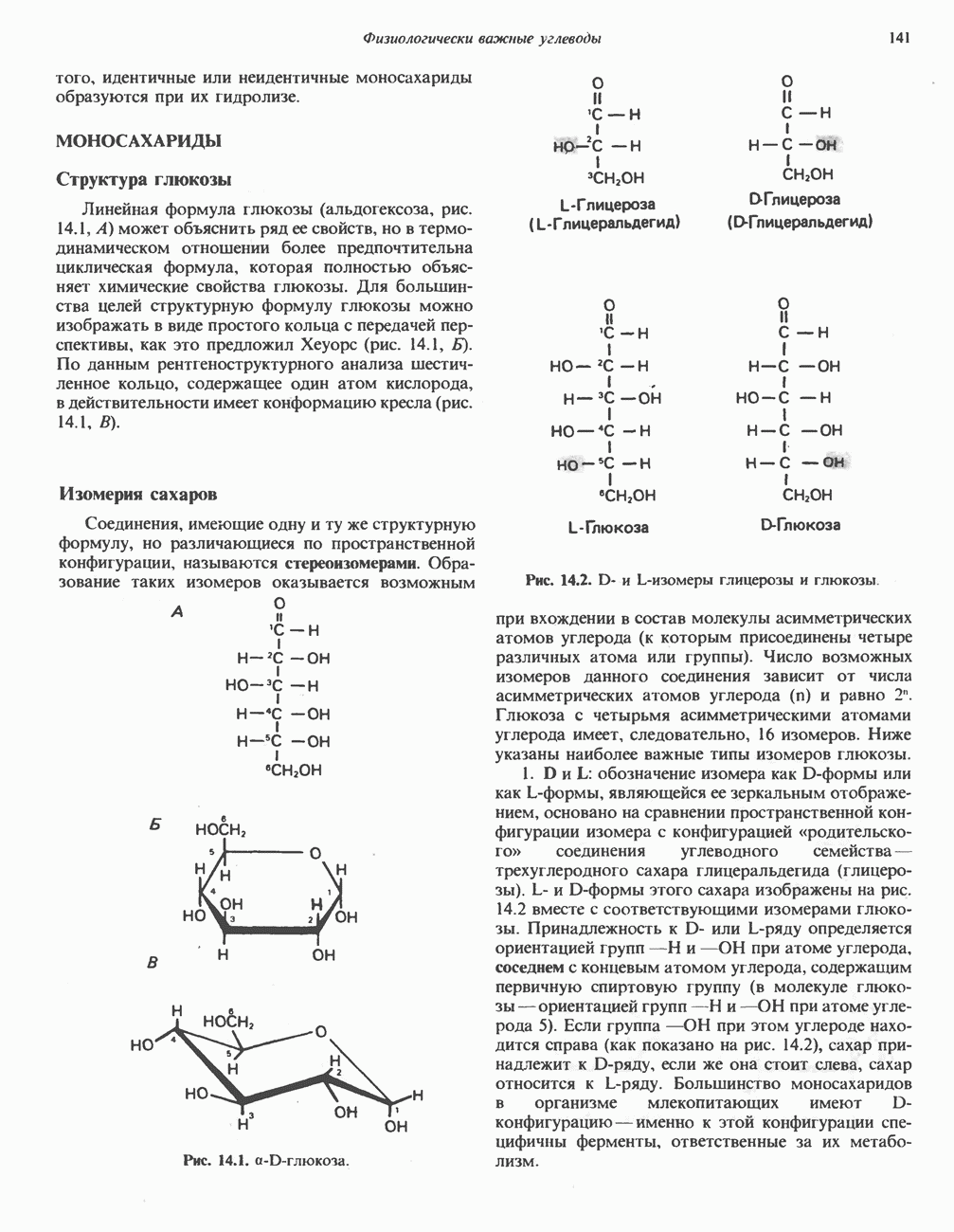
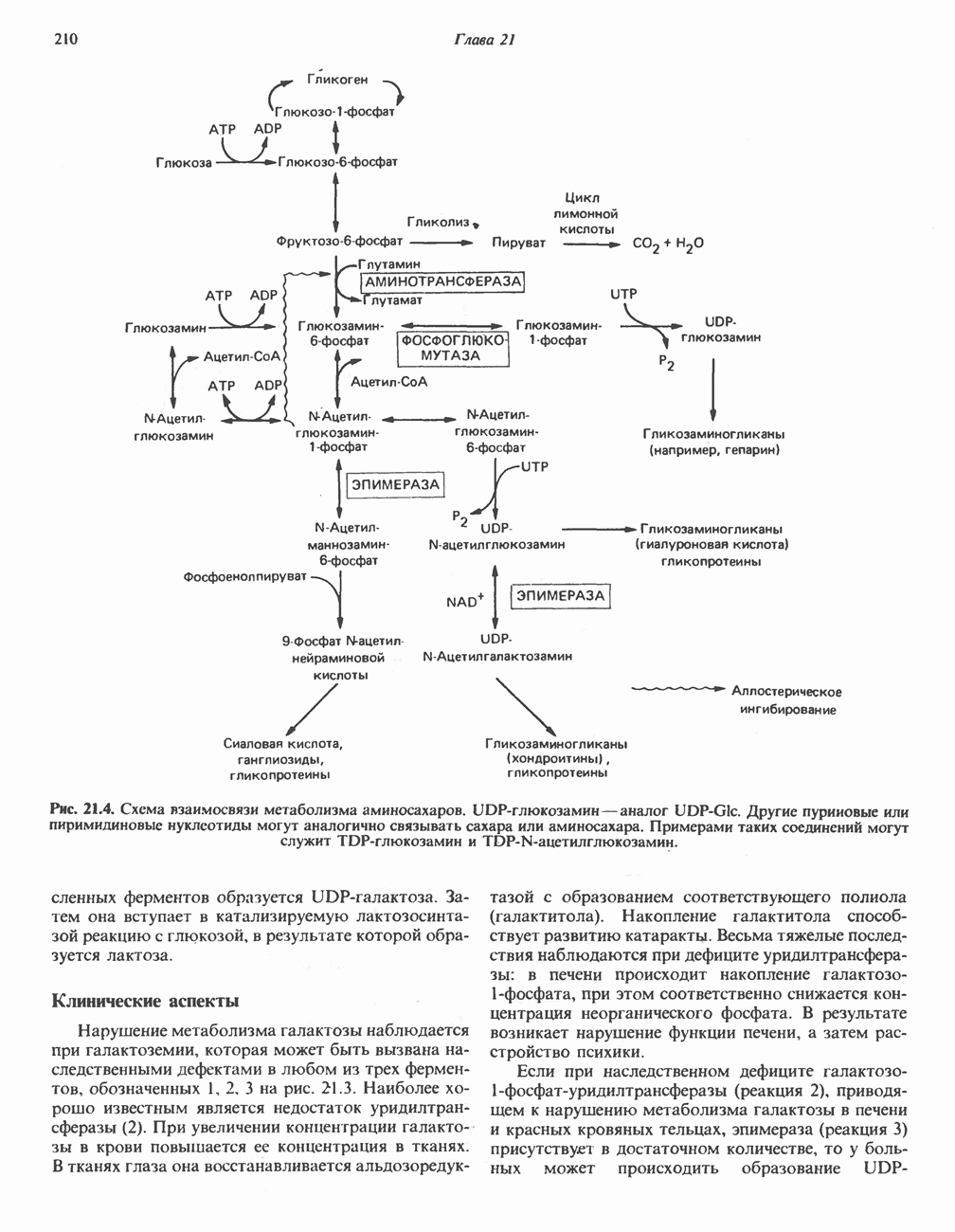
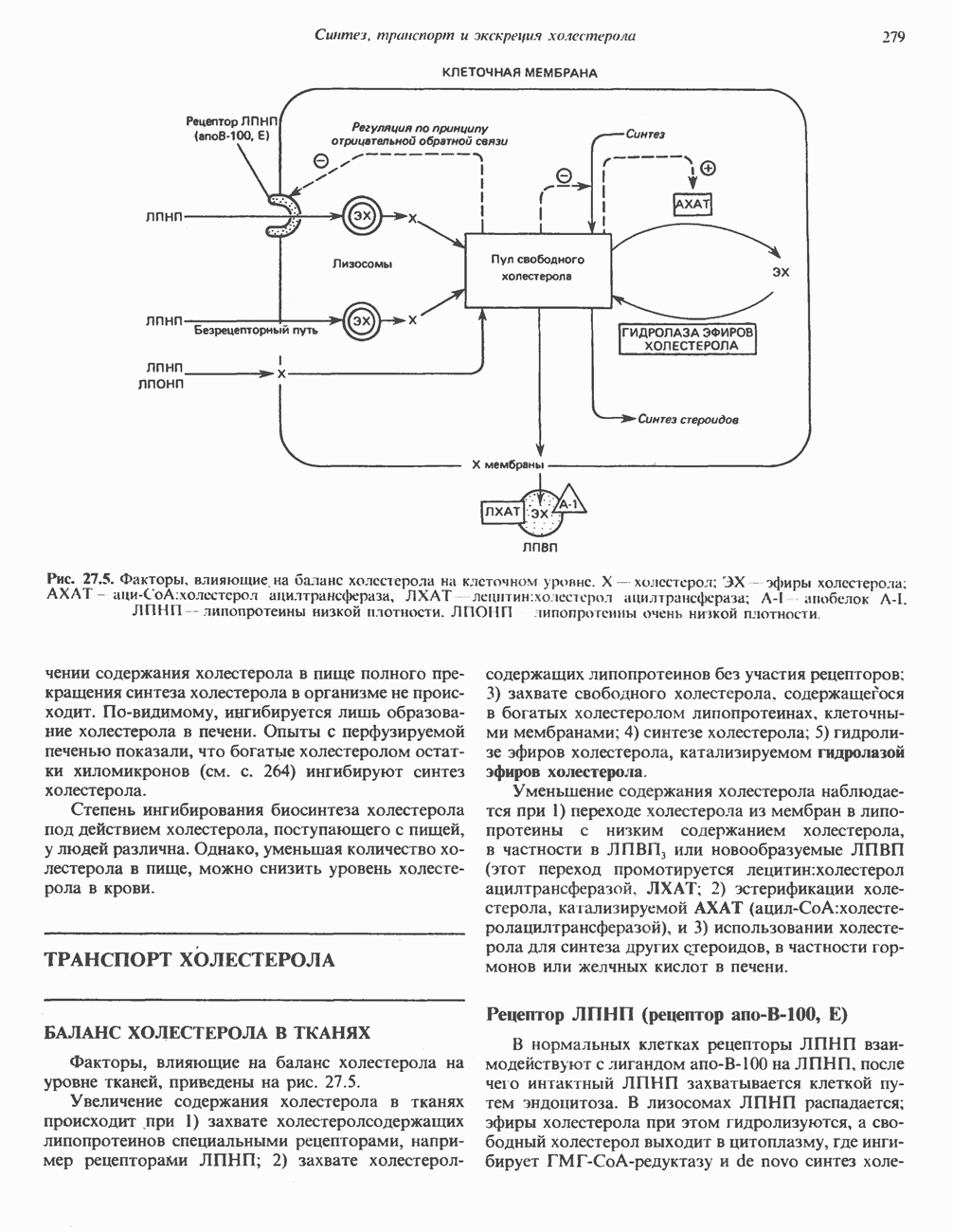
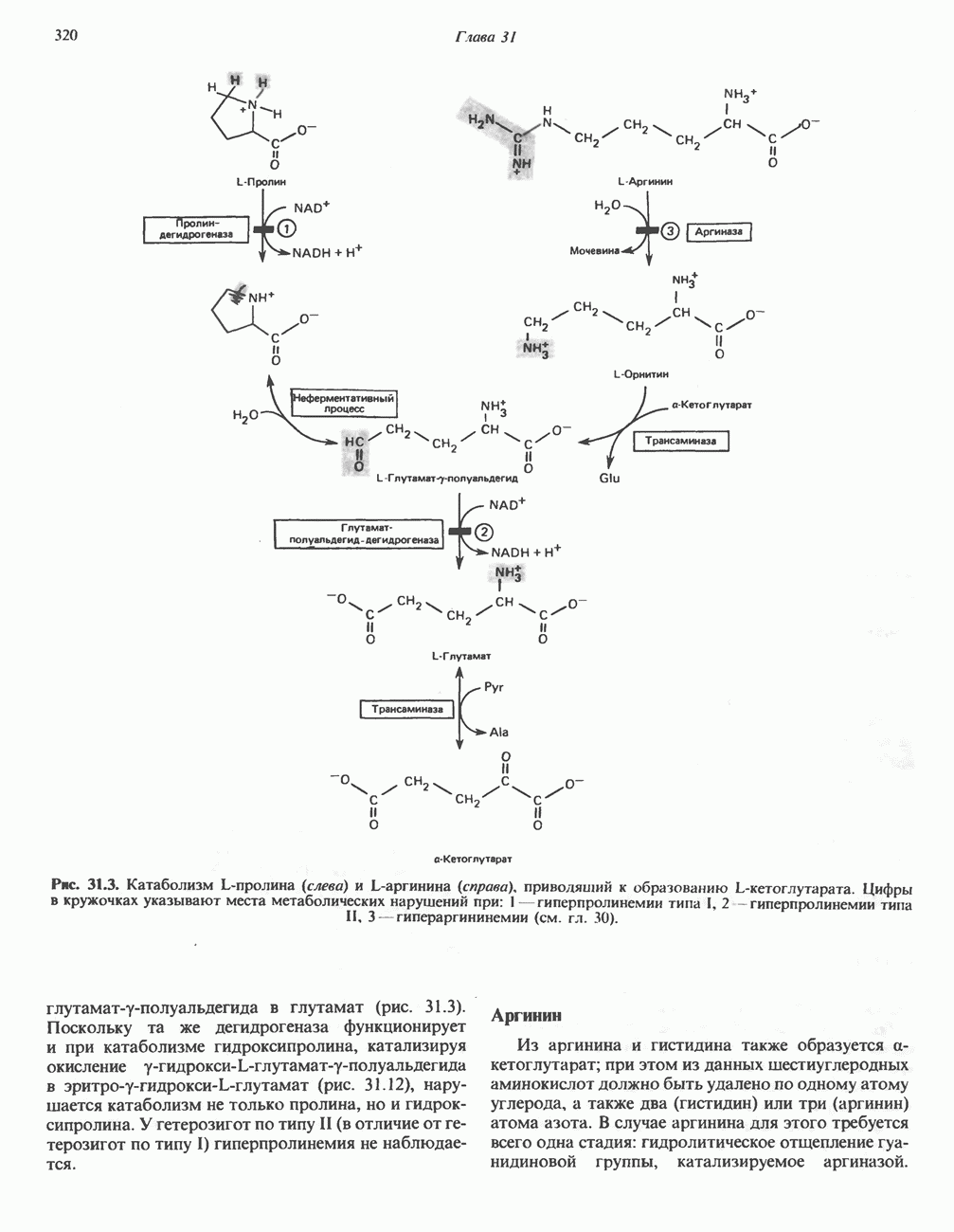
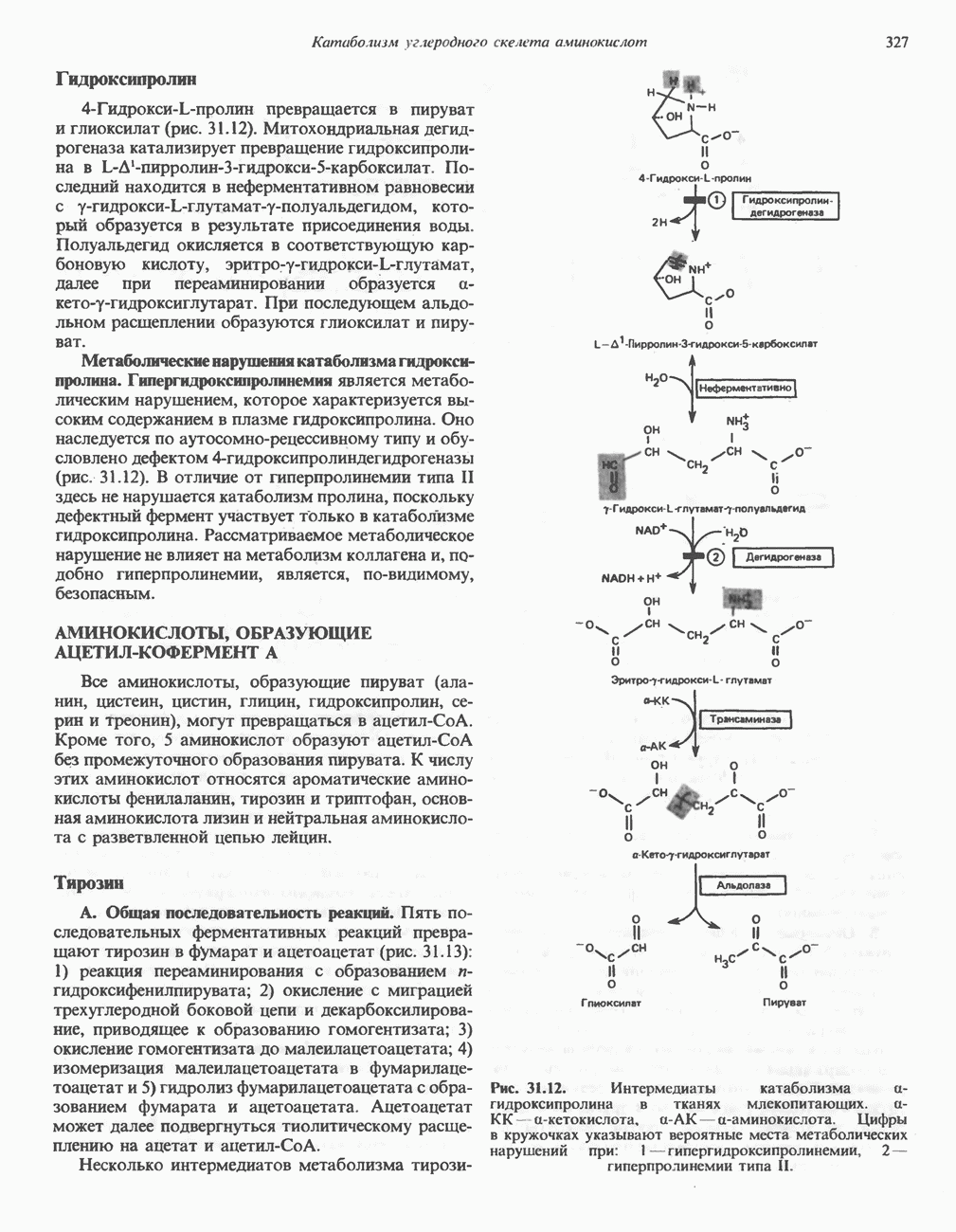
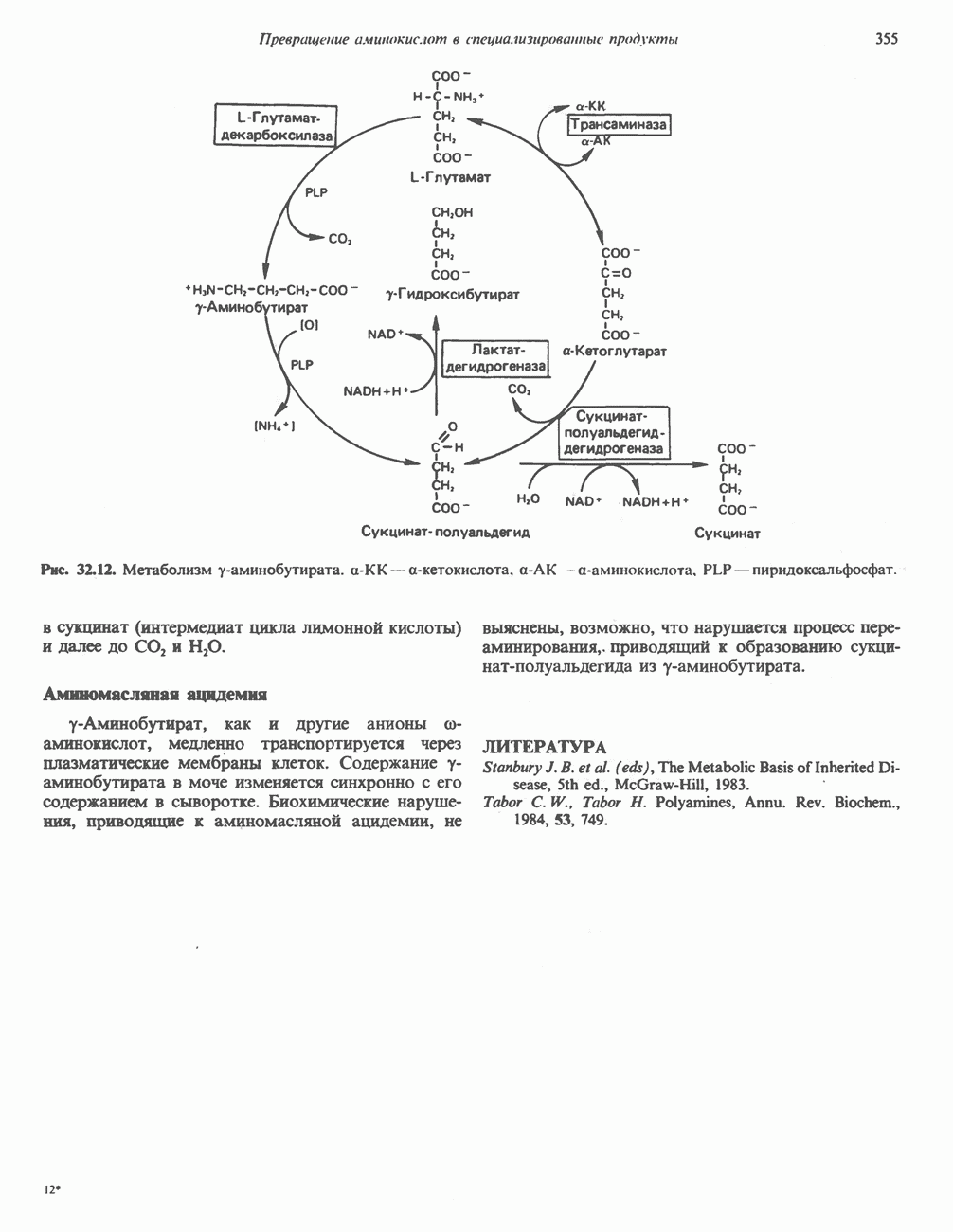
Все 5 углеродов L-пролина переходят в а-кетоглутарат (рис. 31.3, слева). Пролин окисляется в дегидропролин, который при присоединении воды превращается в глутамат-у-полуальдегид. Последний далее окисляется в глутамат, из которого при переаминировании образуется а-кетоглутарат.
Метаболические нарушения в катаболизме пролина. Описано два генетически различных типа гиперпролинемии. Оба типа—тип 1 и тип II — наследуются, вероятно, по аутосомно-рецессивному типу. Хотя в половине зарегистрированных случаев наблюдалась умственная отсталость, оба типа гиперпролинемии не считаются опасными для здоровья.
А. Гиперпролинемия типа I. Участком метаболического блока при гиперпролинемии типа I является пролиндегидрогеназа (рис. 31.3). В отличие от гиперпролинемии типа II в этом случае не наблюдается нарушений в катаболизме гидроксипротлна. На экспериментальной модели гиперпролинемии типа I, мышах линии  показано, что активность пролиндегидрогеназы печени составляет только 10% от нормы. У гетерозигот типа I гиперпролинемия выражена в легкой форме.
показано, что активность пролиндегидрогеназы печени составляет только 10% от нормы. У гетерозигот типа I гиперпролинемия выражена в легкой форме.
Б. Гиперпролинемия типа II. Степень гиперпролинемии в этом случае выше, чем при гиперпролинемии типа I. Моча содержит А-пирролин-3-гидрокси-5-карбоксилат. При гиперпролинемии типа II участком метаболического блока является дегидрогеназа, катализирующая окисление
(см. скан)
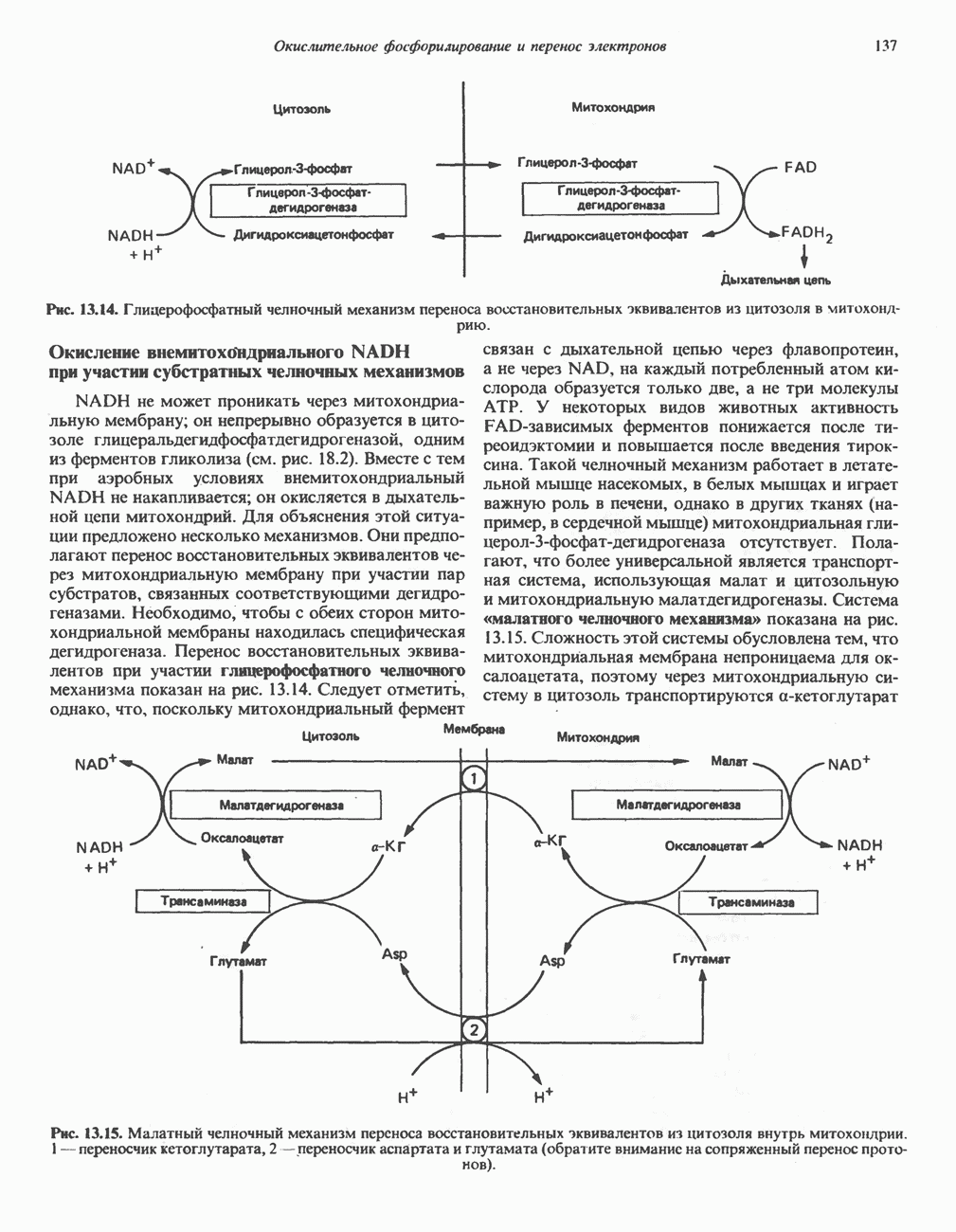
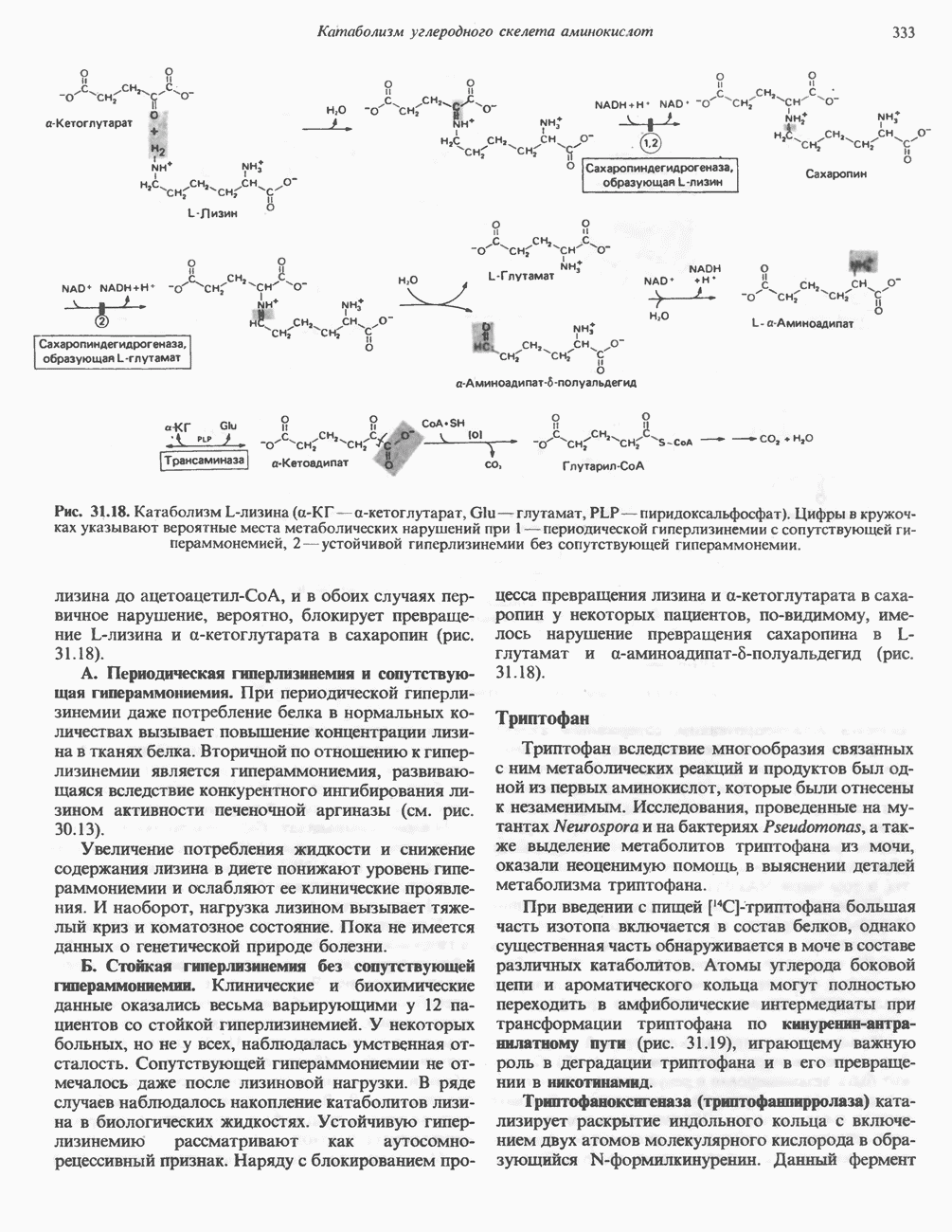
Рис. 31.3. Катаболизм L-пролина (слева) и L-аргинина (справа), приводяший к образованию L-кетоглутарата. Цифры в кружочках указывают места метаболических нарушений при: 1 - гиперпролинемии типа I, 2 - гиперпролинемии типа II, 3 - гипераргининемии (см. гл. 30).
глутамат-у-полуальдегида в глутамат (рис. 31.3). Поскольку та же дегидрогеназа функционирует и при катаболизме гидроксипролина, катализируя окисление у-гидрокси-Ь-глутамат-у-полуальдегида в эритро-у-гидрокси-Ь-глутамат (рис. 31.12), нарушается катаболизм не только пролина, но и гидроксипролина. У гетерозигот по типу II (в отличие от гетерозигот по типу I) гиперпролинемия не наблюдается.
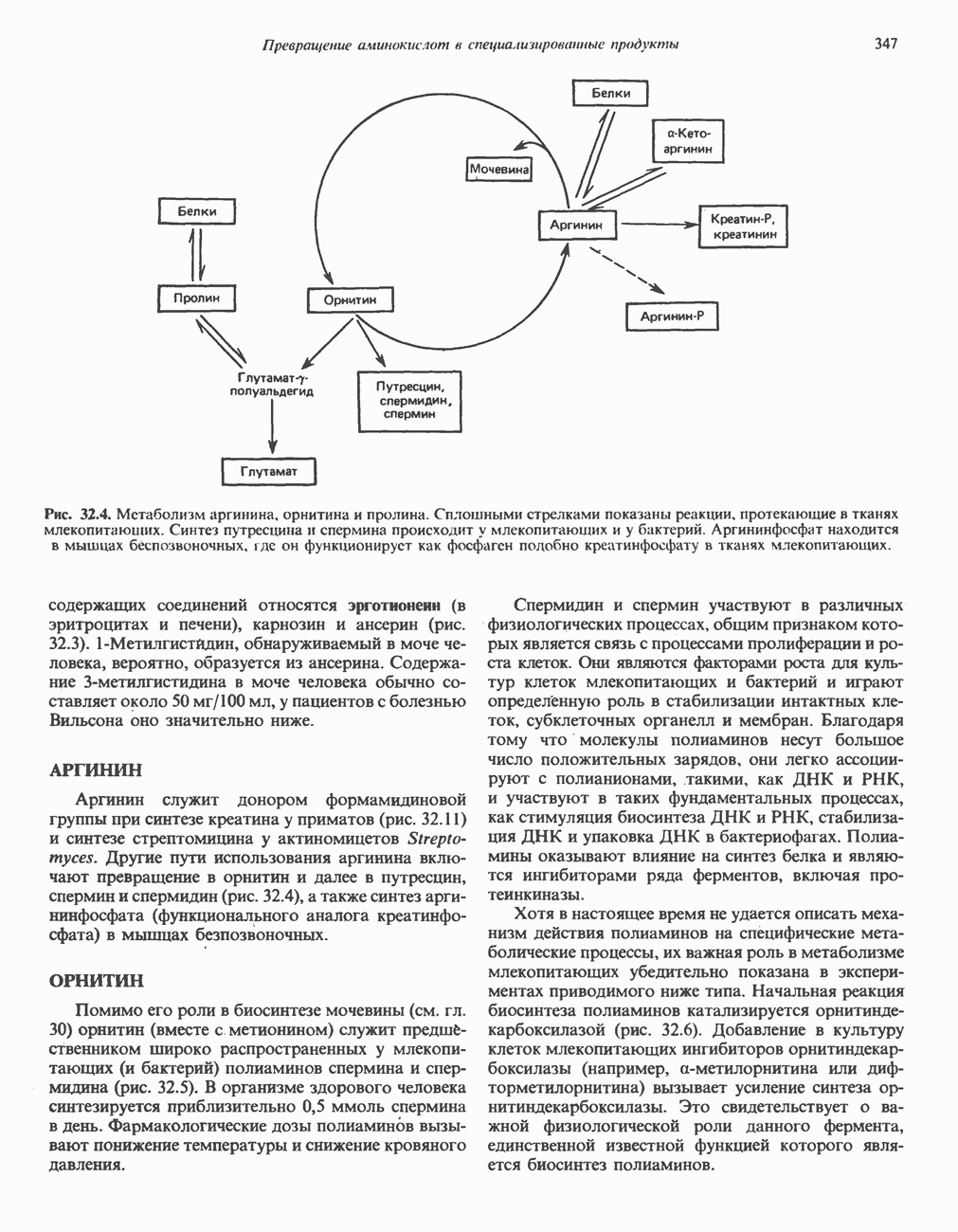
Аргинин
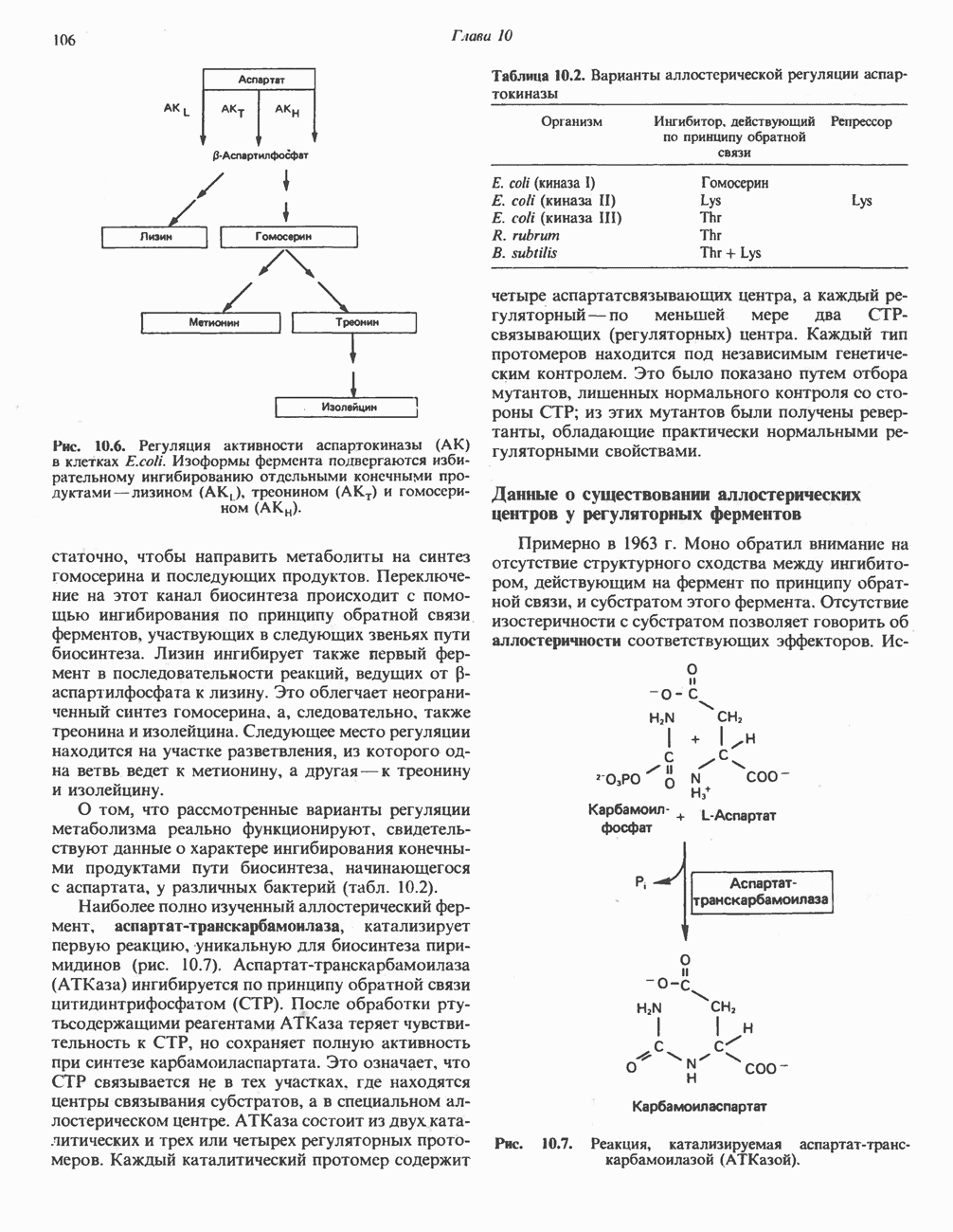
Из аргинина и гистидина также образуется а-кетоглутарат; при этом из данных шестиуглеродных аминокислот должно быть удалено по одному атому углерода, а также два (гистидин) или три (аргинин) атома азота. В случае аргинина для этого требуется всего одна стадия: гидролитическое отщепление гуанидиновой группы, катализируемое аргиназой.
Образующийся продукт — орнитин далее вступает в реакцию переаминировання по 6-аминогруппе, в результате образуется глутамат-у-полуальдегид, который затем превращается в а-кетоглутарат, как это описано выше для пролина (рис. 31.3).
Гипераргннннемия — метаболическое нарушение катаболизма аргинина, обусловленное недостаточностью фермента аргиназы печени, рассматривалось в гл. 30 в связи с метаболическими нарушениями цикла мочевины.
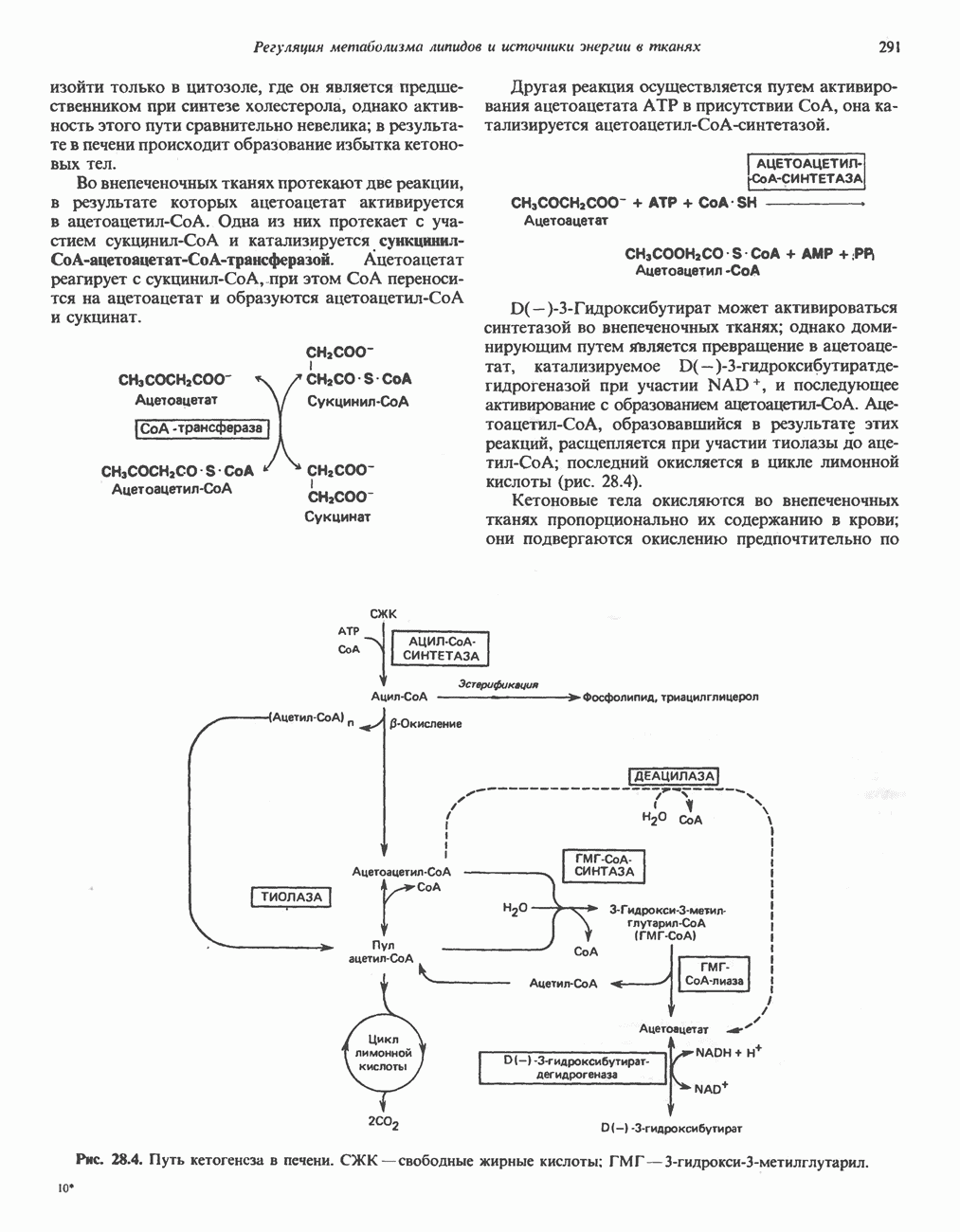
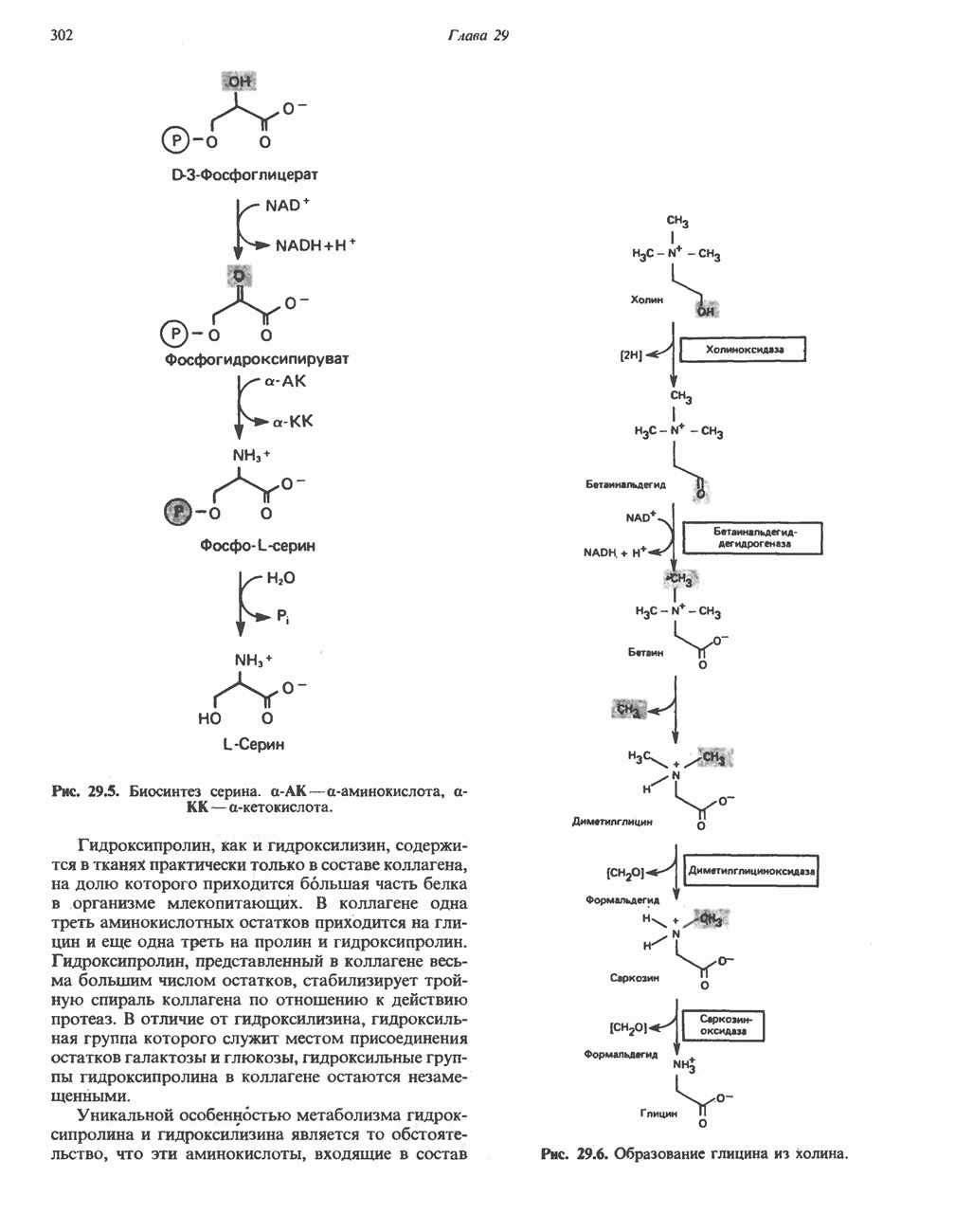
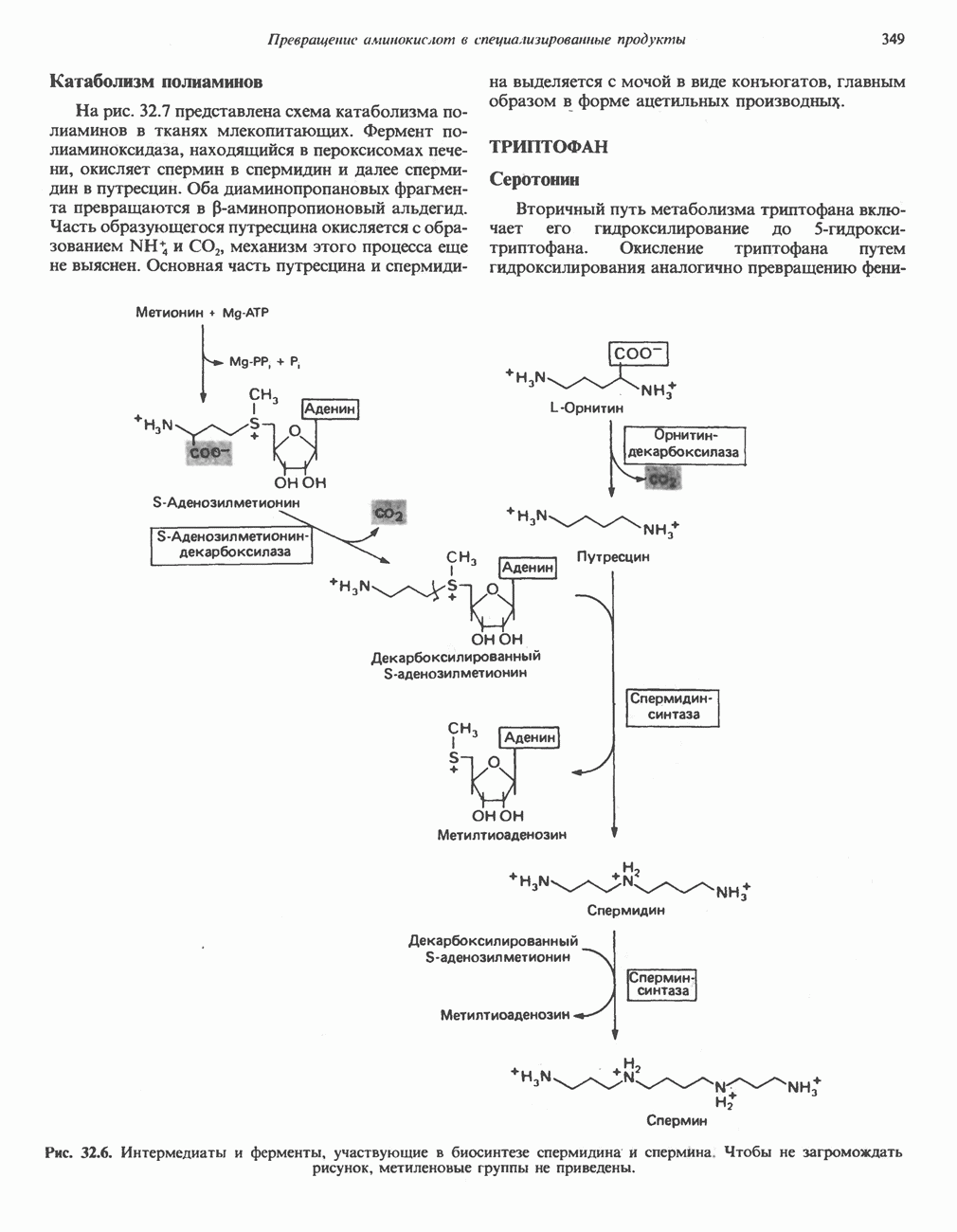
Гистидин
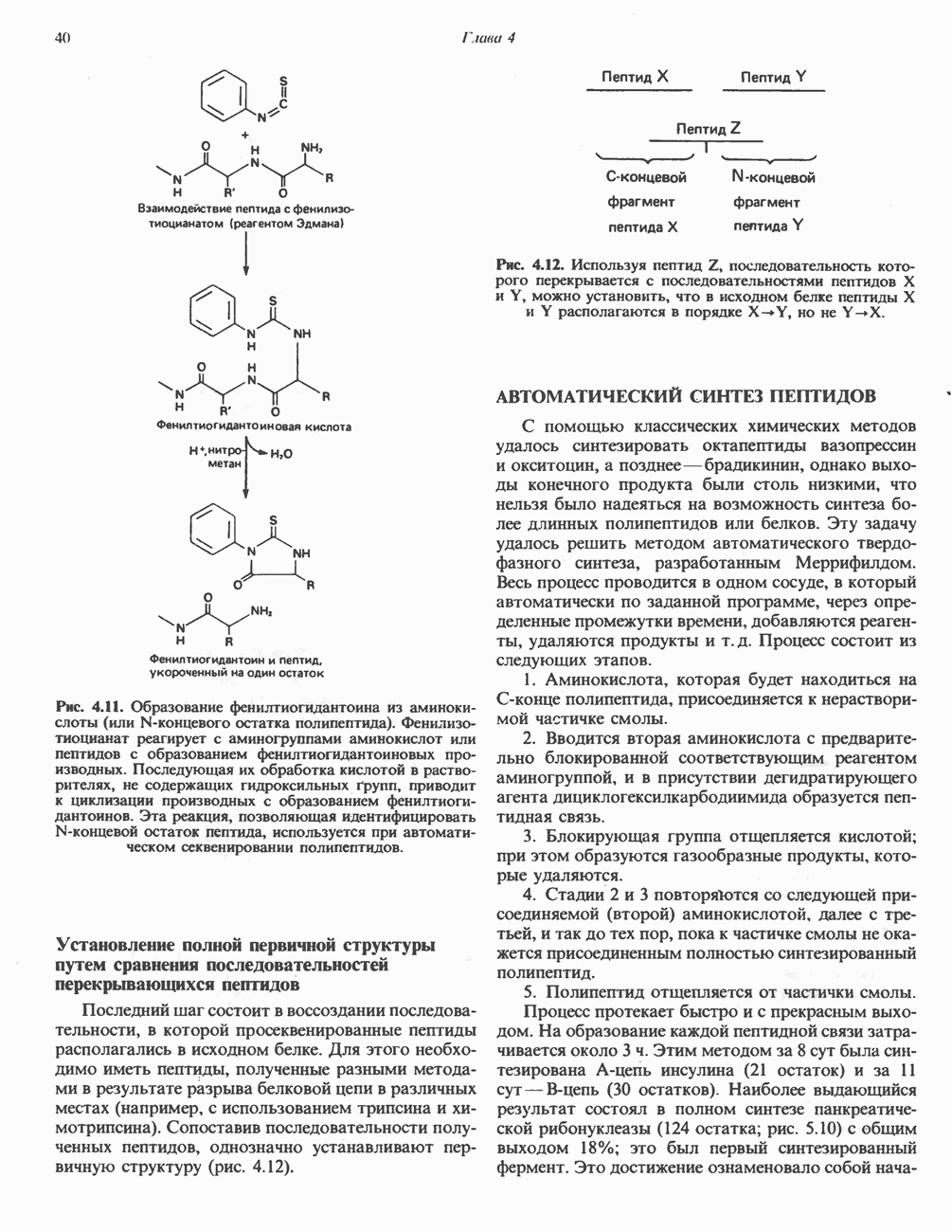
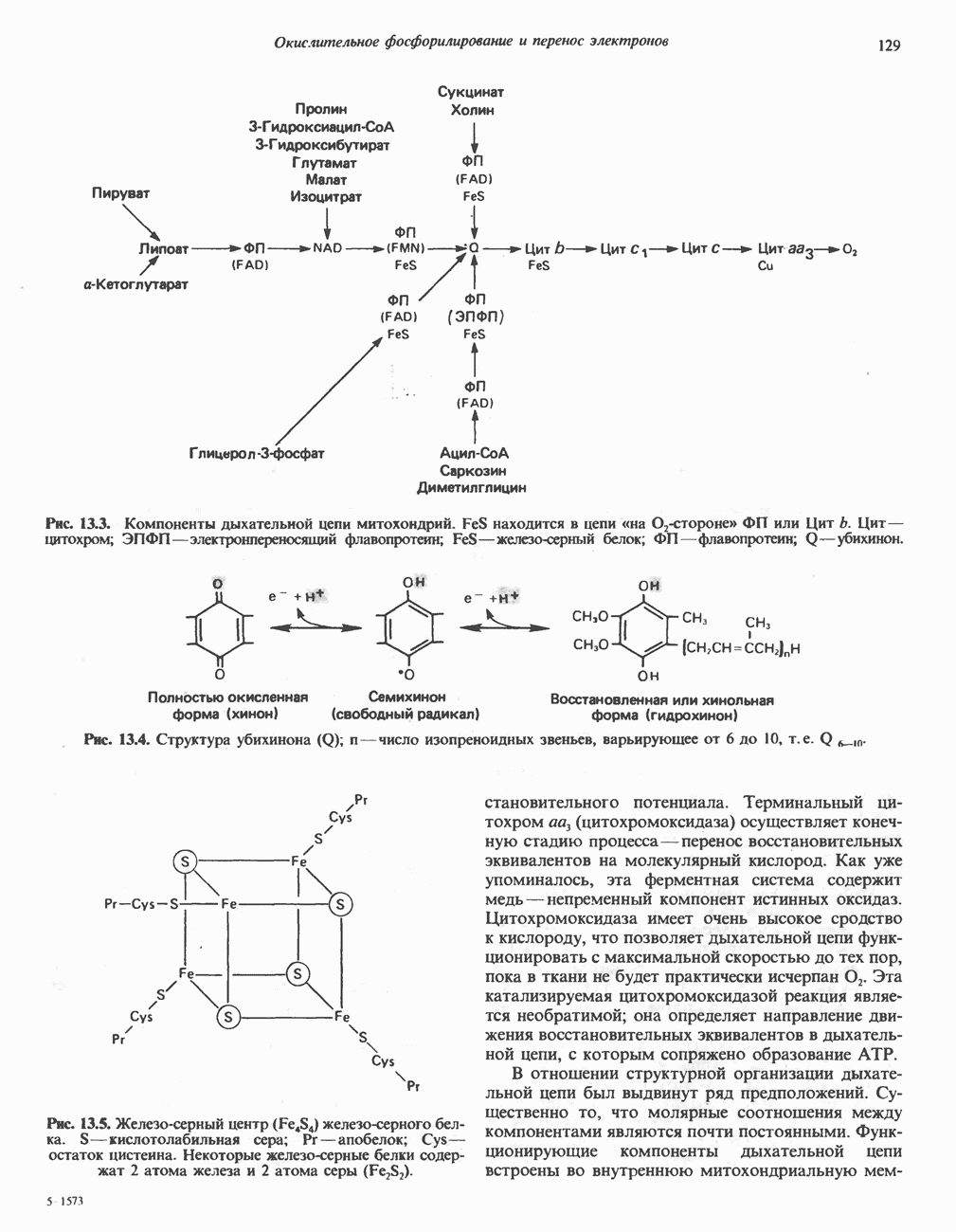
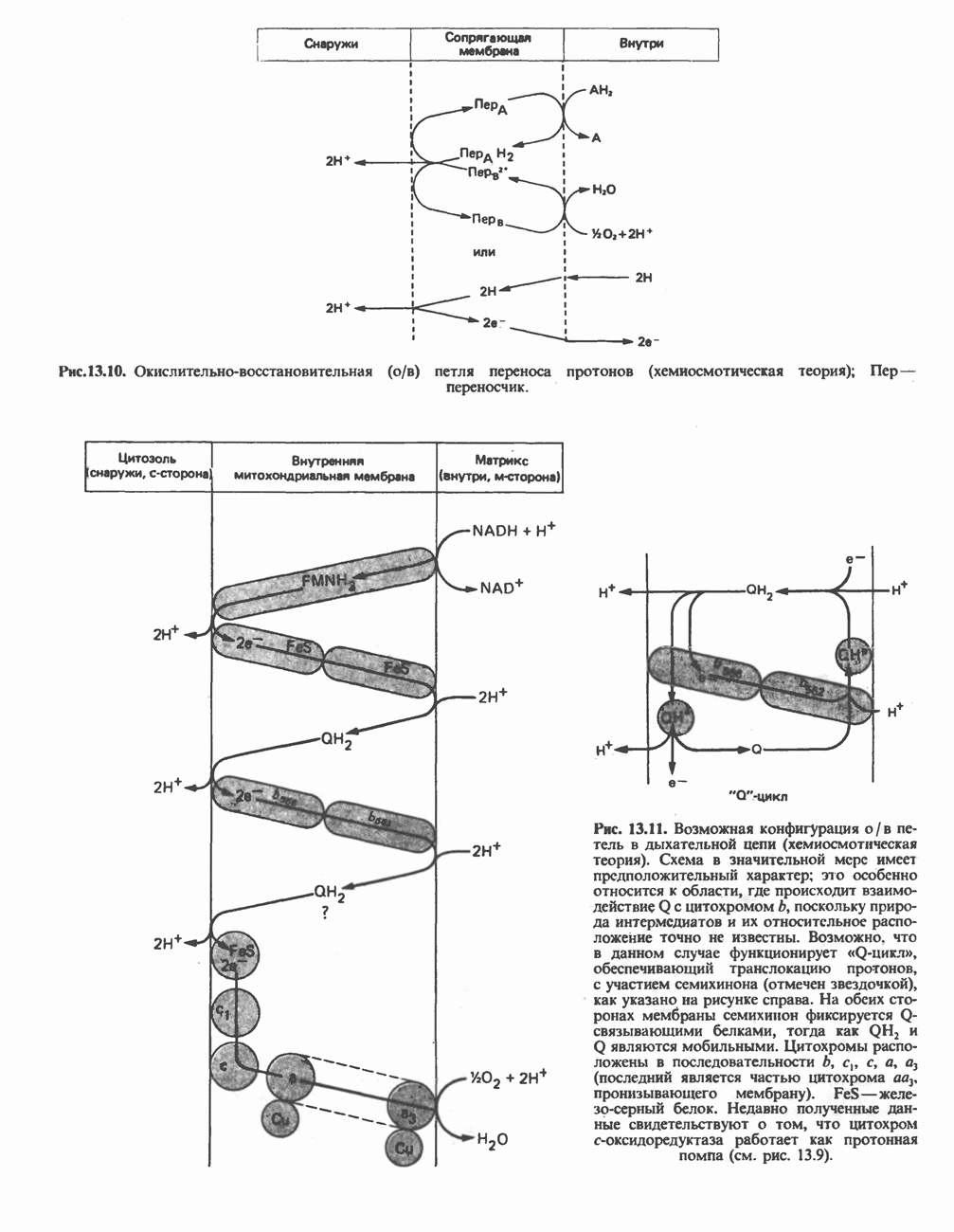
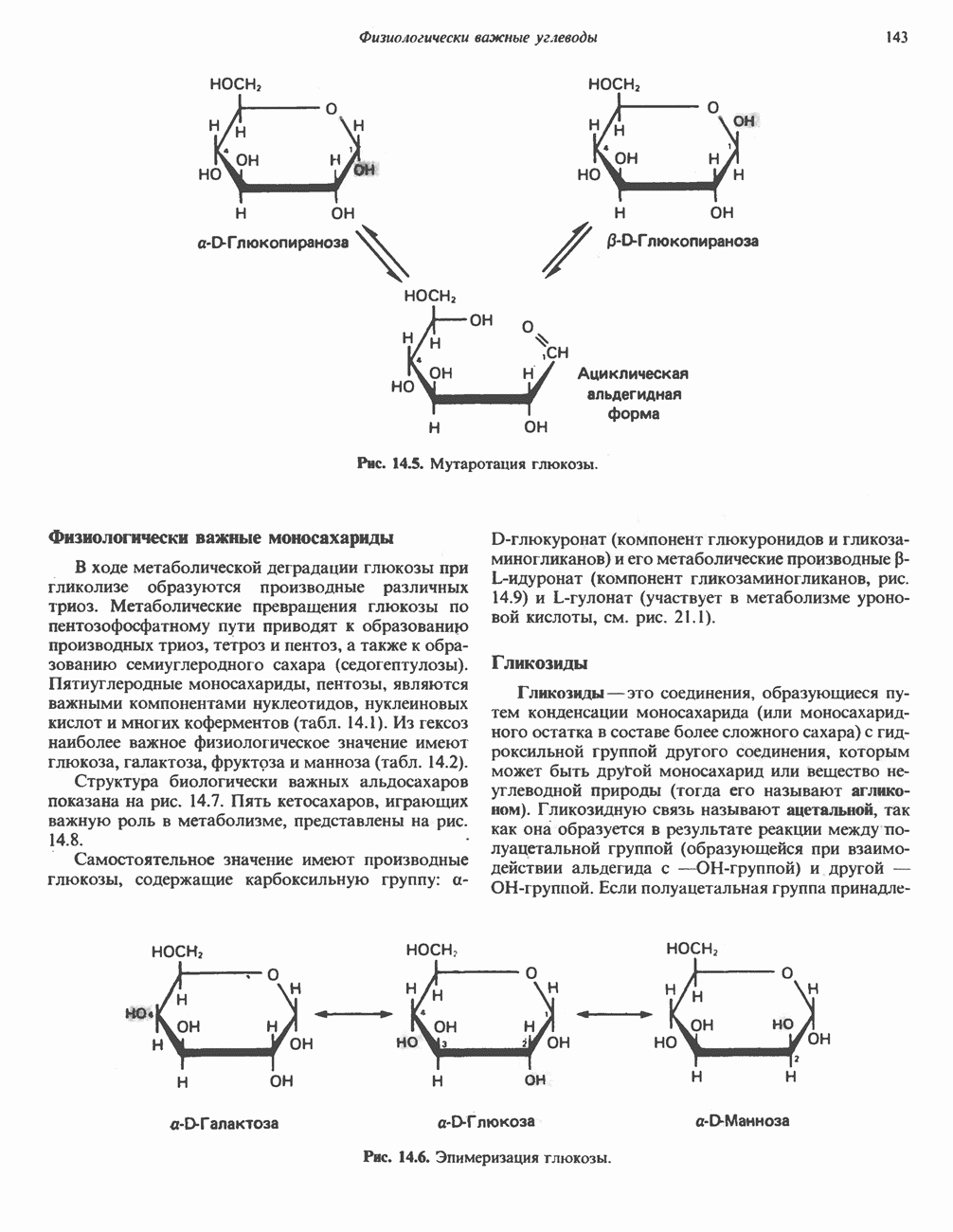
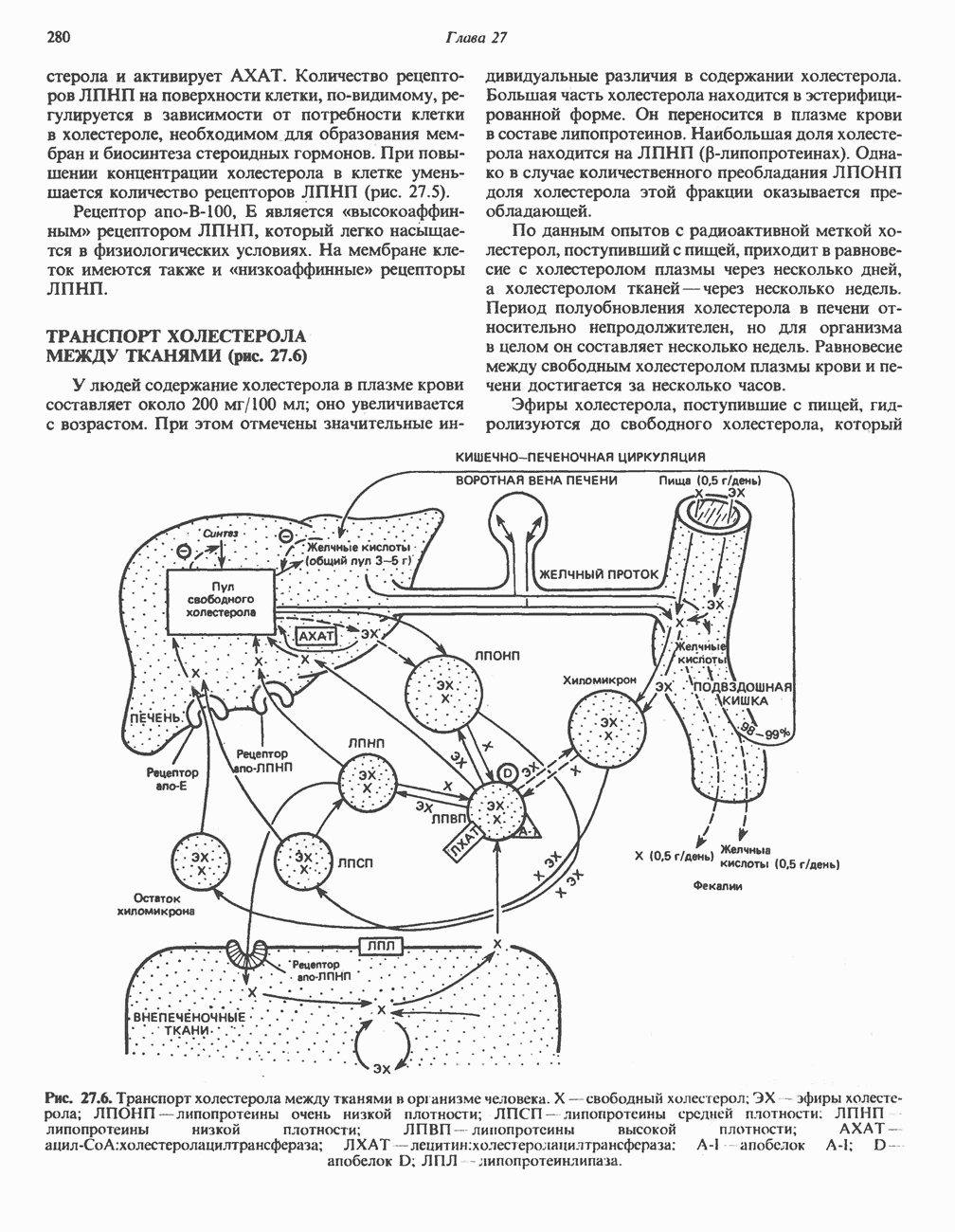
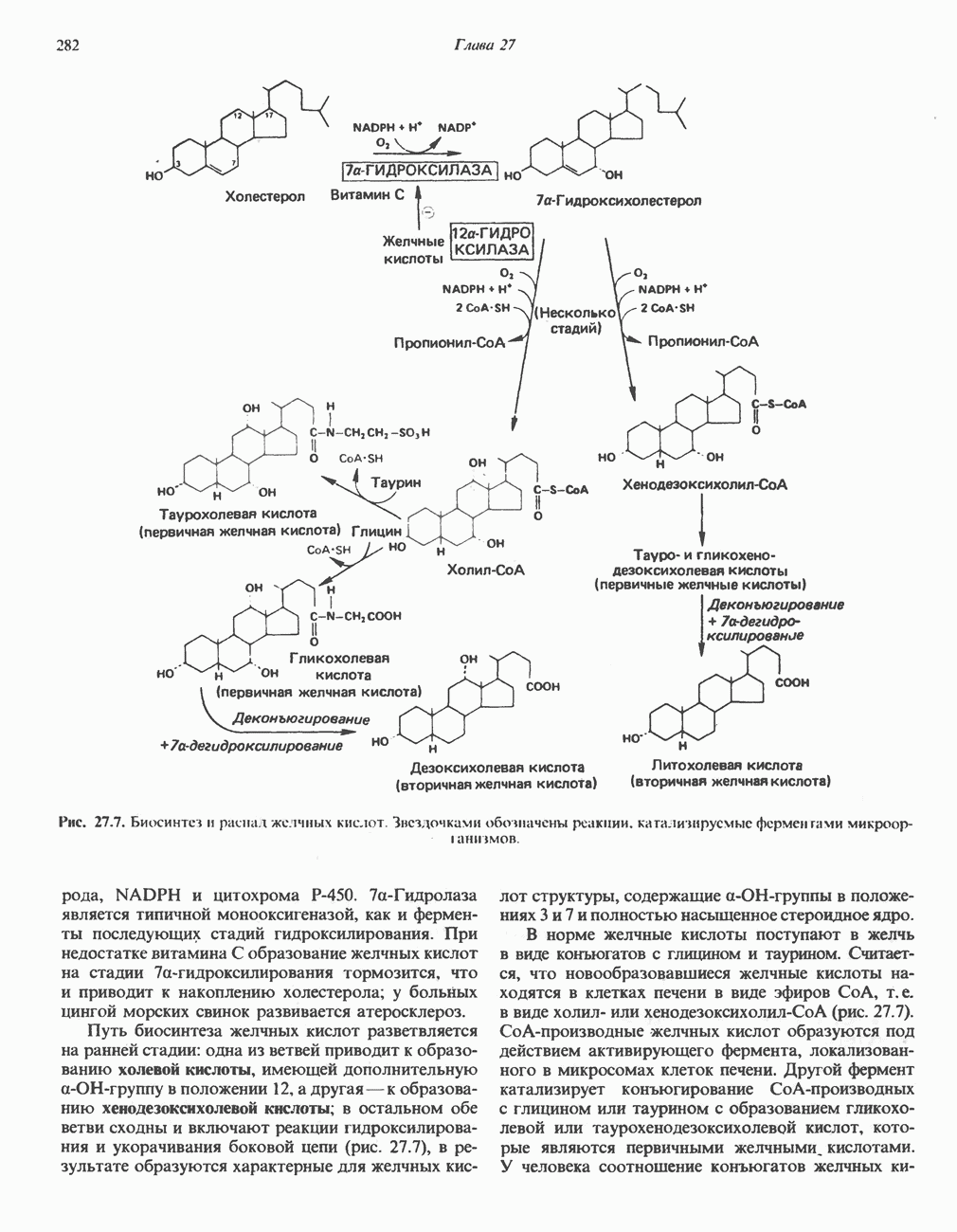
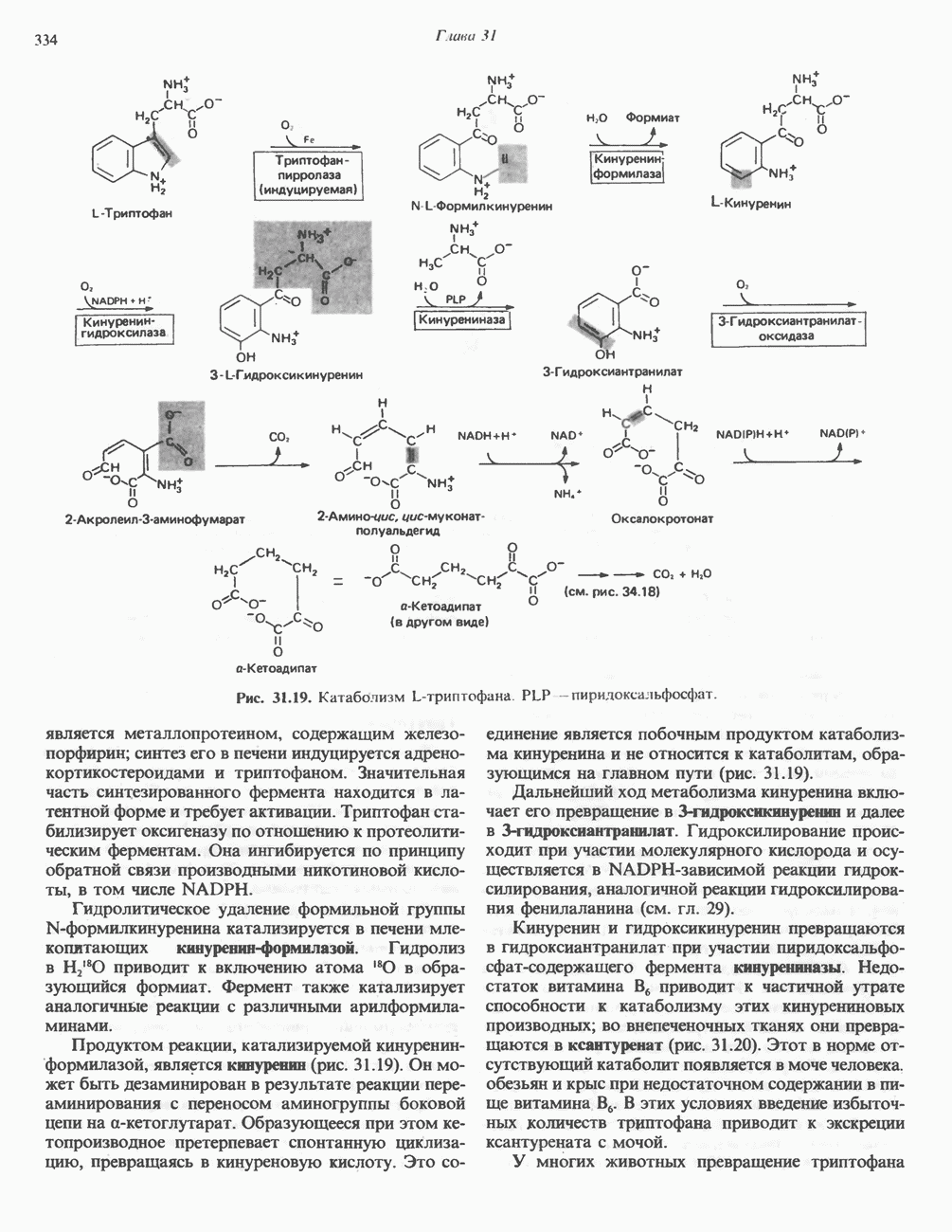
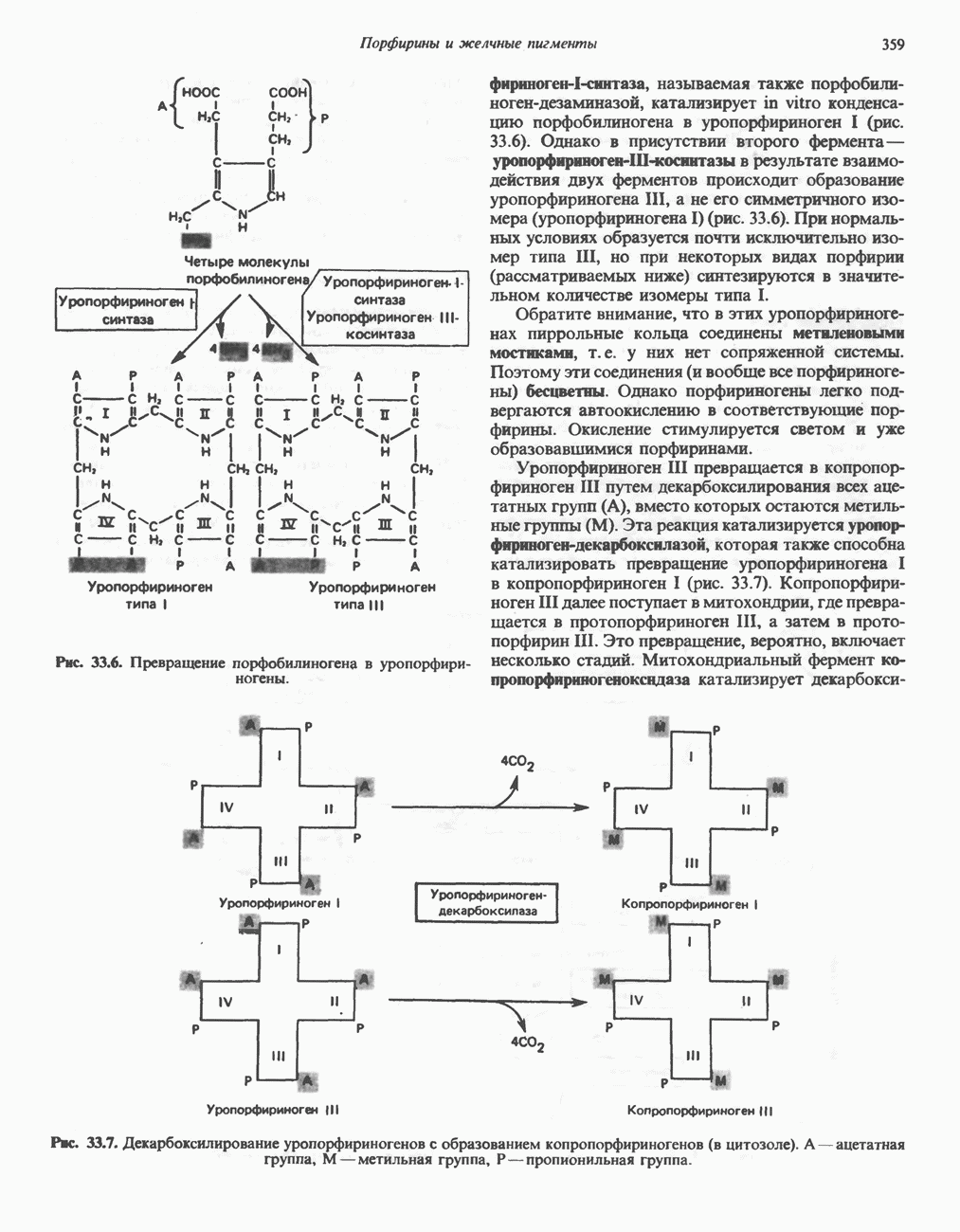
Для удаления из гистидина «лишних» атомов углерода и азота требуется 4 реакции (рис. 31.4). При дезаминировании гистидина образуется уроканат. Превращение уроканата в 4-имидазолон-5-пропионат, катализируемое уроканазой, включает присоединение воды и внутримолекулярный окислительно-восстановительный процесс. Превращение 4-имидазолон-5-пропионата может идти по ряду путей; на пути образования а-кетоглутарата сначала происходит гидролиз с образованием N-формиминоглутамата, далее формиминогруппа переносится на а-углерод гетрагидрофолата, при этом образуются глутамат и ГФ-формимино-тетрагидрофолат. У пациентов, страдающих от недостатка фолиевой кислоты, последняя реакция частично или полностью блокируется и N-формиминоглутамат экскретнруется с мочой. На этом основан тест на недостаточность фолиевой кислоты; после нагрузки гистидином в моче обнаруживается N-формиминоглутамат.
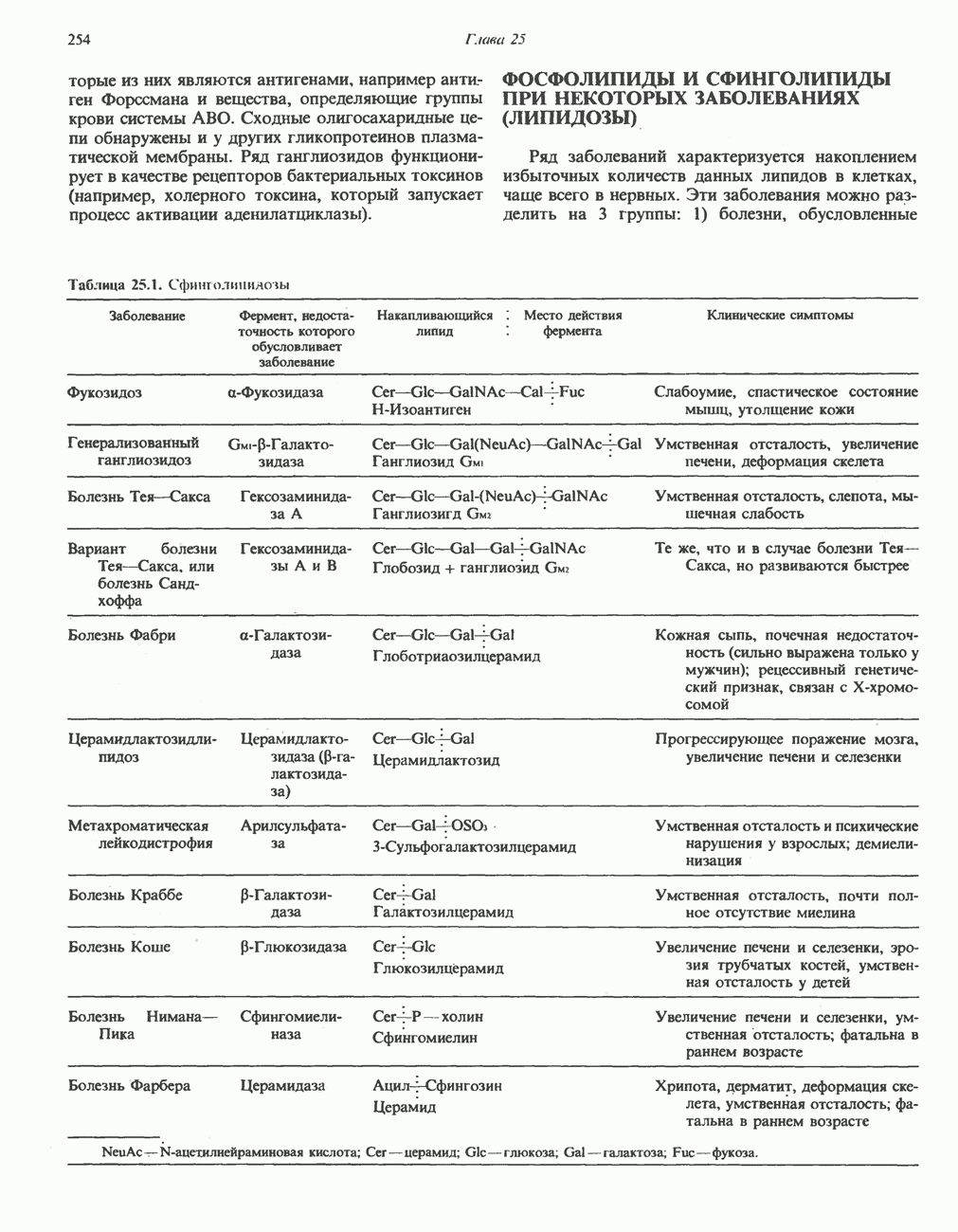
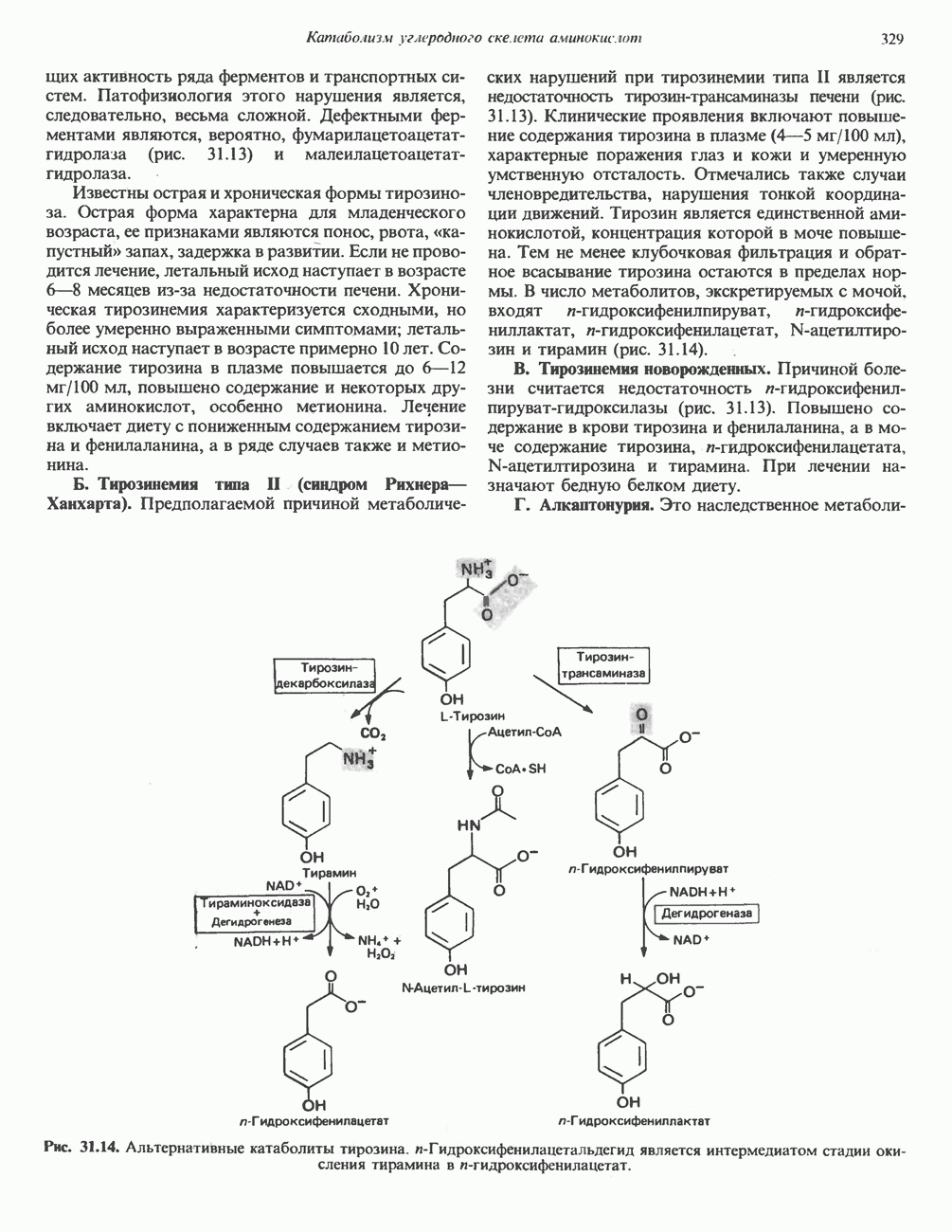
Метаболические нарушения катаболизма гистидина. Гистидинемия—метаболическое нарушение катаболизма гистидина, обусловленное дефицитом фермента гистидазы (рис. 31.4), наследуется по аутосомно-рецессивному пути. Свыше половины больных гистидинемией характеризуются умственной отсталостью и дефектами речи.
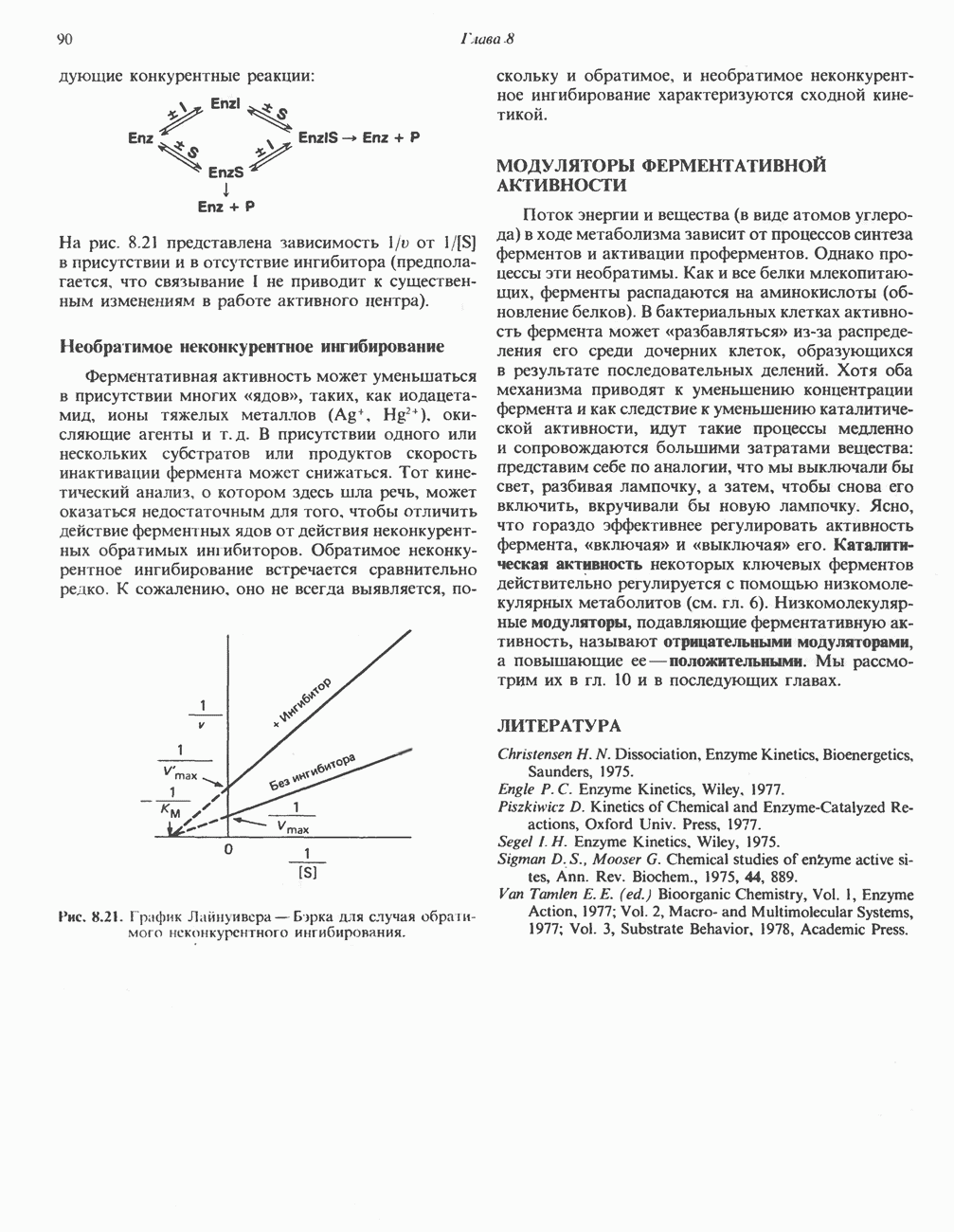
Наряду с повышением уровня гистидина в крови и моче наблюдается также возрастание экскреции имидазолпирувата (при проведении цветной реакции с феррихлоридом следует иметь в виду, что окраску дает также фенилпируват, поэтому иногда ошибочно ставят диагноз «фенилкетонурия»). Метаболической причиной гистидинемии является недостаточная активность гистидазы в печени, вследствие этого замедляется превращение гистидина в уроканат (рис. 31.4). В этом случае создаются благоприятные возможности для реализации альтернативного метаболического
(см. скан)
Рис. 31.4, Катаболическое превращение  -гистидина с образованием а-кетоглутарата.
-гистидина с образованием а-кетоглутарата.  -тетрагидрофолат. Реакция, катализируемая гистидазой. является вероятным местом метаболическою нарушения при гистидинемии.
-тетрагидрофолат. Реакция, катализируемая гистидазой. является вероятным местом метаболическою нарушения при гистидинемии.
пути гистидин вступает в реакцию переаминирования и превращается в имидазолпируват; избыток последнего экскретнруется с мочой. В моче пациентов с гистидинемией обнаружены также продукты превращения имидазолпируватаимида-золацетат и имидазоллактат.
В норме содержание гистидина в моче достаточно велико, и его легко детектировать. Заметное повышение содержания  истидина в моче может служить характерным тестом на нормальную беременность, в то же время содержание аминокислоты не повышается, когда беременность сопровождается возрастанием кровяного давления. Повышенное выделение гистидина при беременности не следует рассматривать как результат нарушения метаболизма этой аминокислоты. Наблюдаемые явления можно объяснить изменением функции почек при нормальной беременности а также при беременности, сопровождающейся гипертонией. Следует отметить что в период беременности повышается экскреция не только гистидина, но и ряда других аминокислот.
истидина в моче может служить характерным тестом на нормальную беременность, в то же время содержание аминокислоты не повышается, когда беременность сопровождается возрастанием кровяного давления. Повышенное выделение гистидина при беременности не следует рассматривать как результат нарушения метаболизма этой аминокислоты. Наблюдаемые явления можно объяснить изменением функции почек при нормальной беременности а также при беременности, сопровождающейся гипертонией. Следует отметить что в период беременности повышается экскреция не только гистидина, но и ряда других аминокислот.