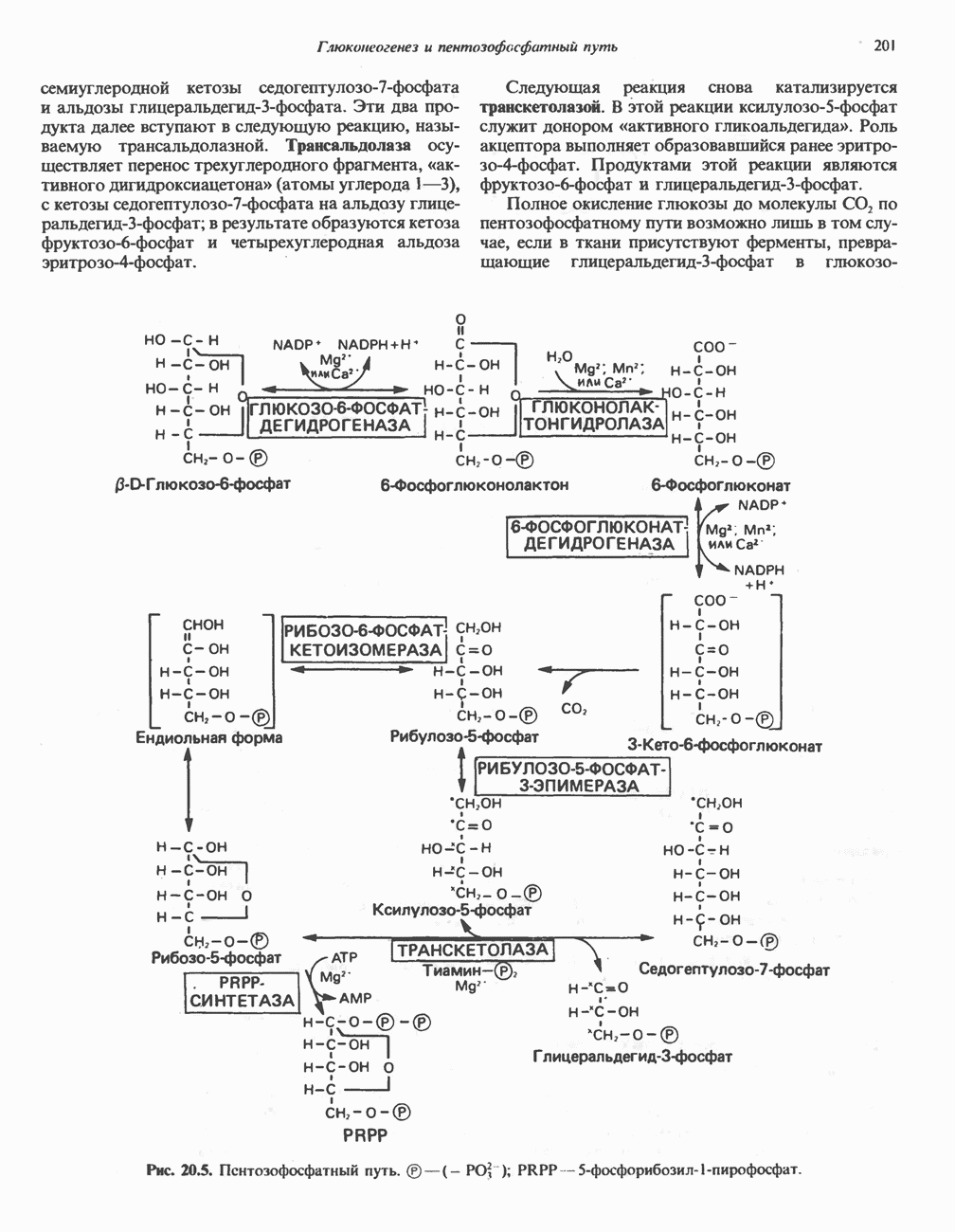
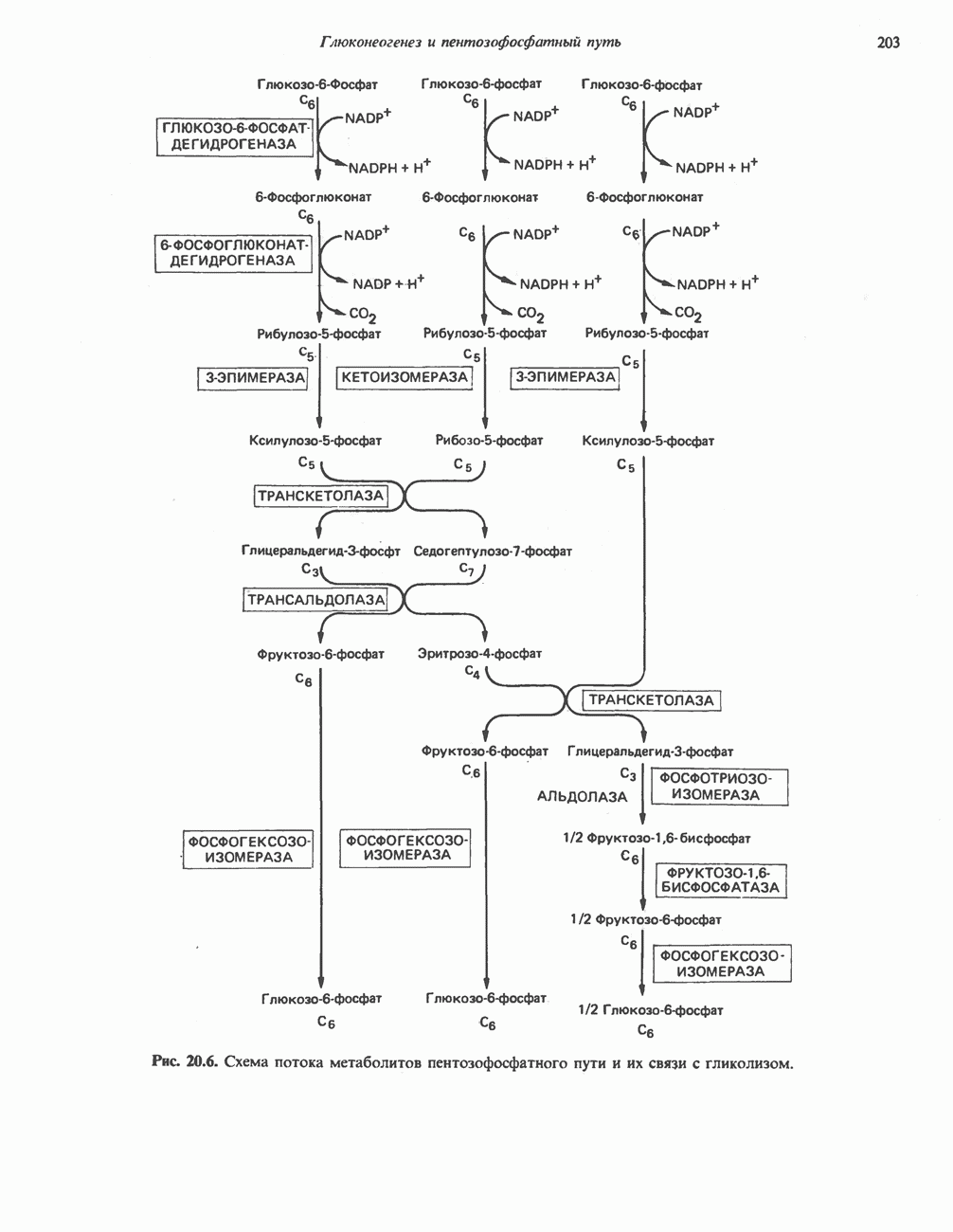
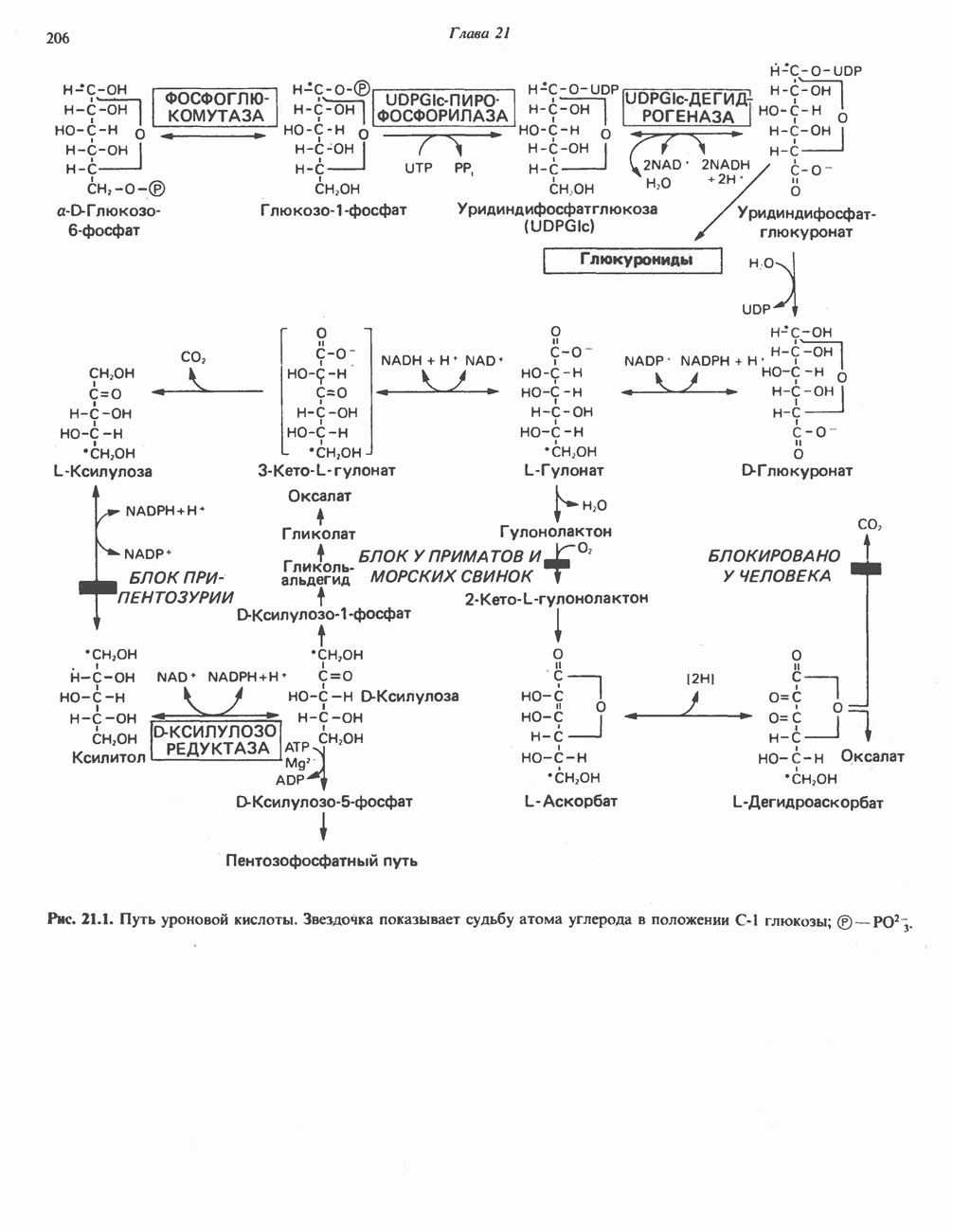
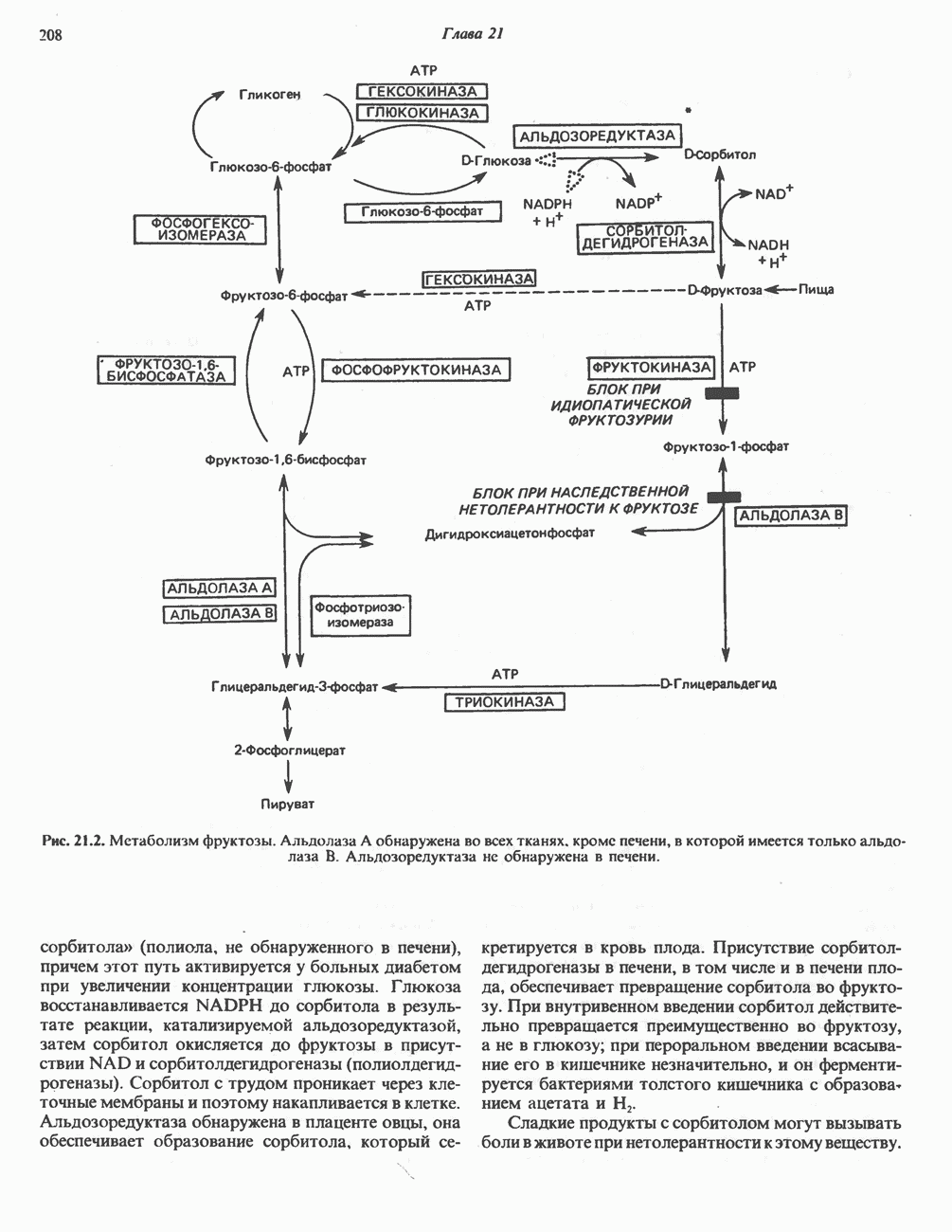
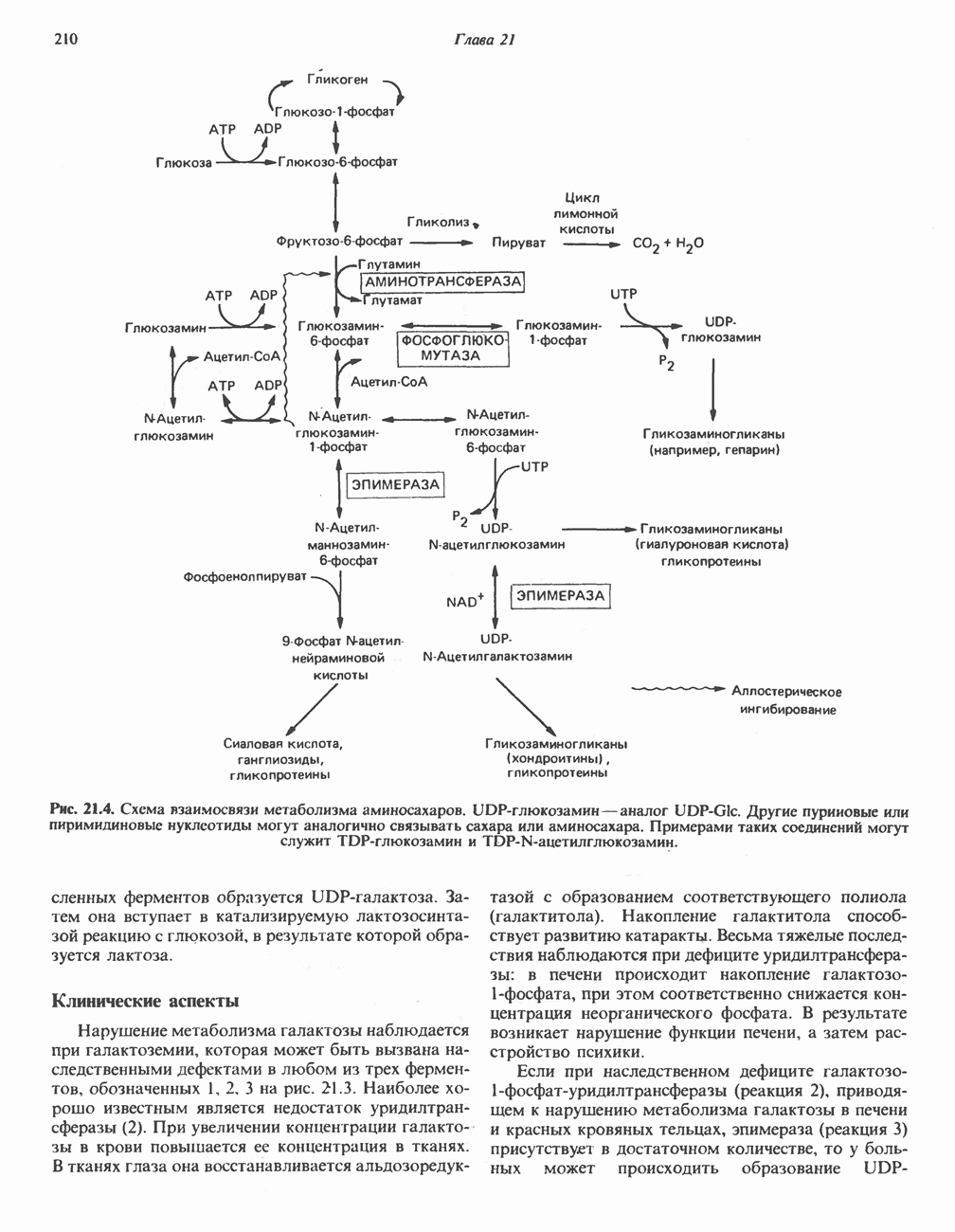
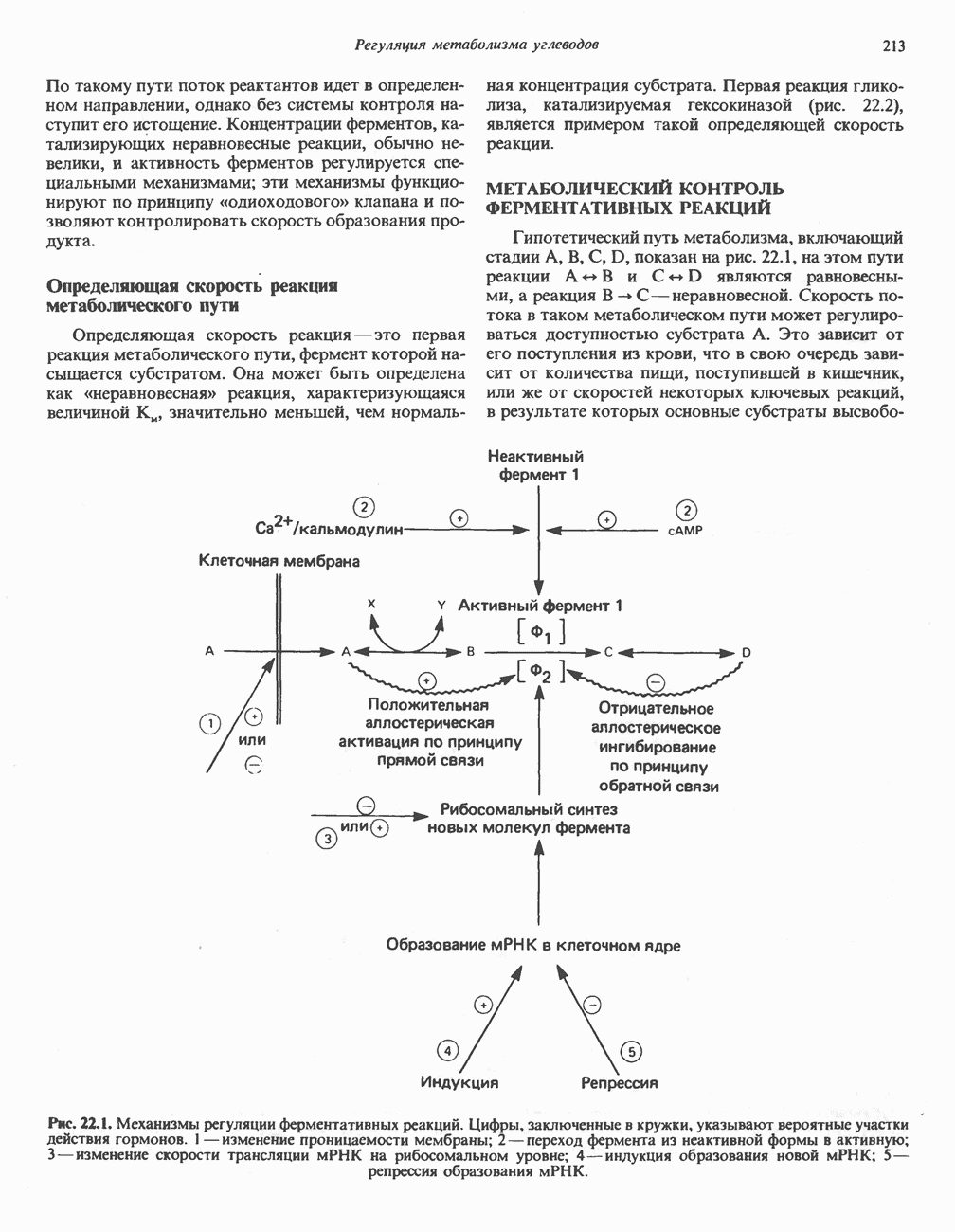
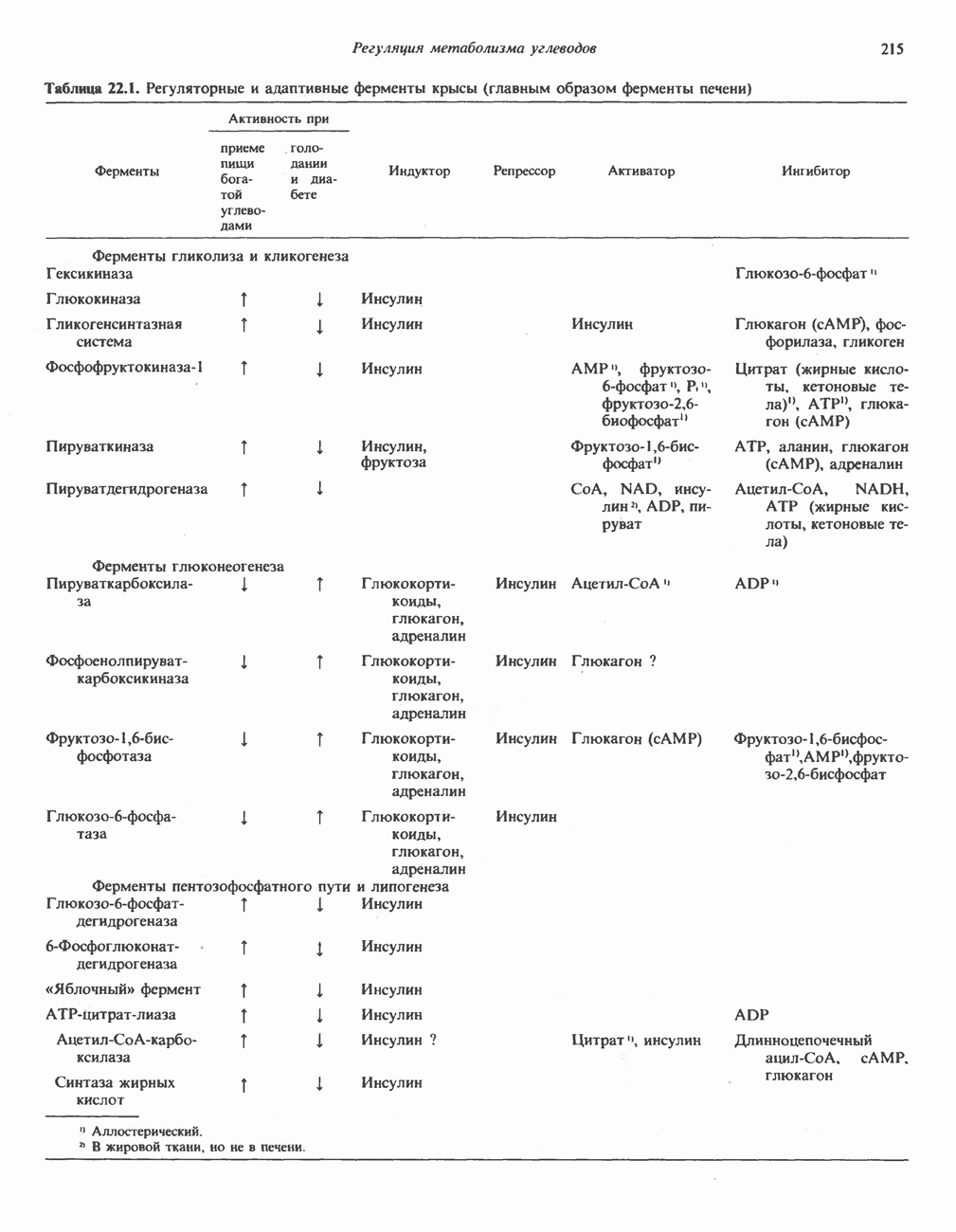
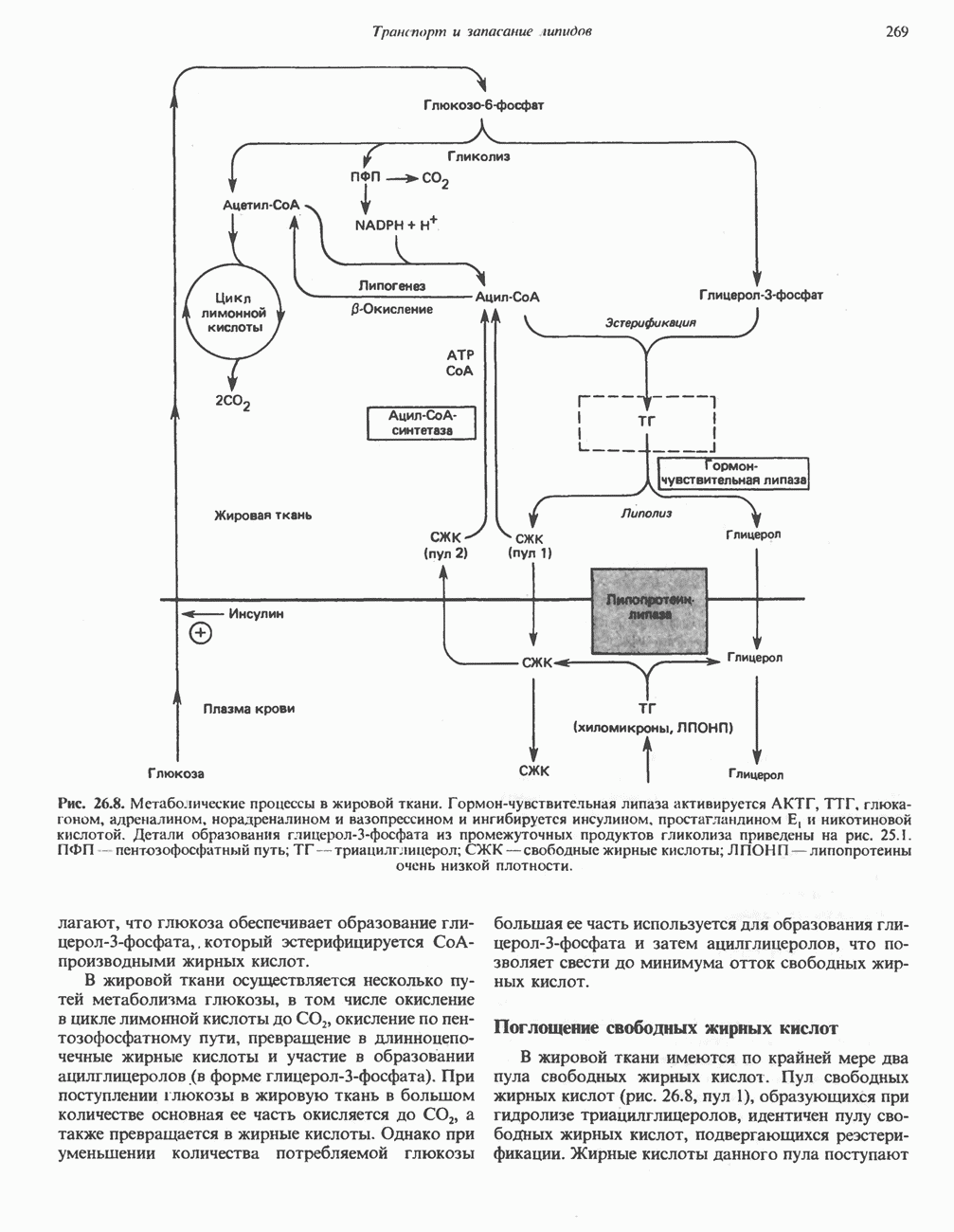
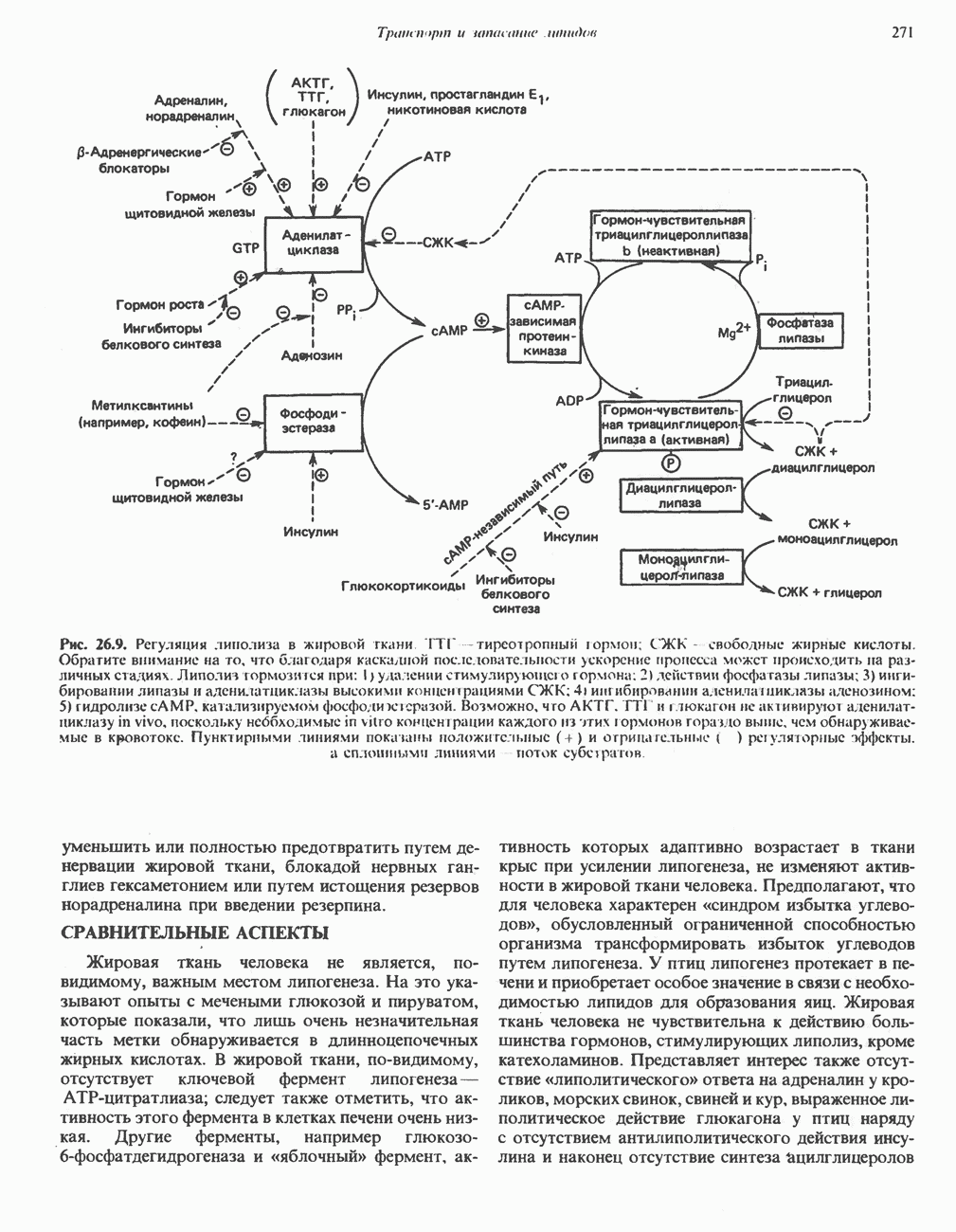
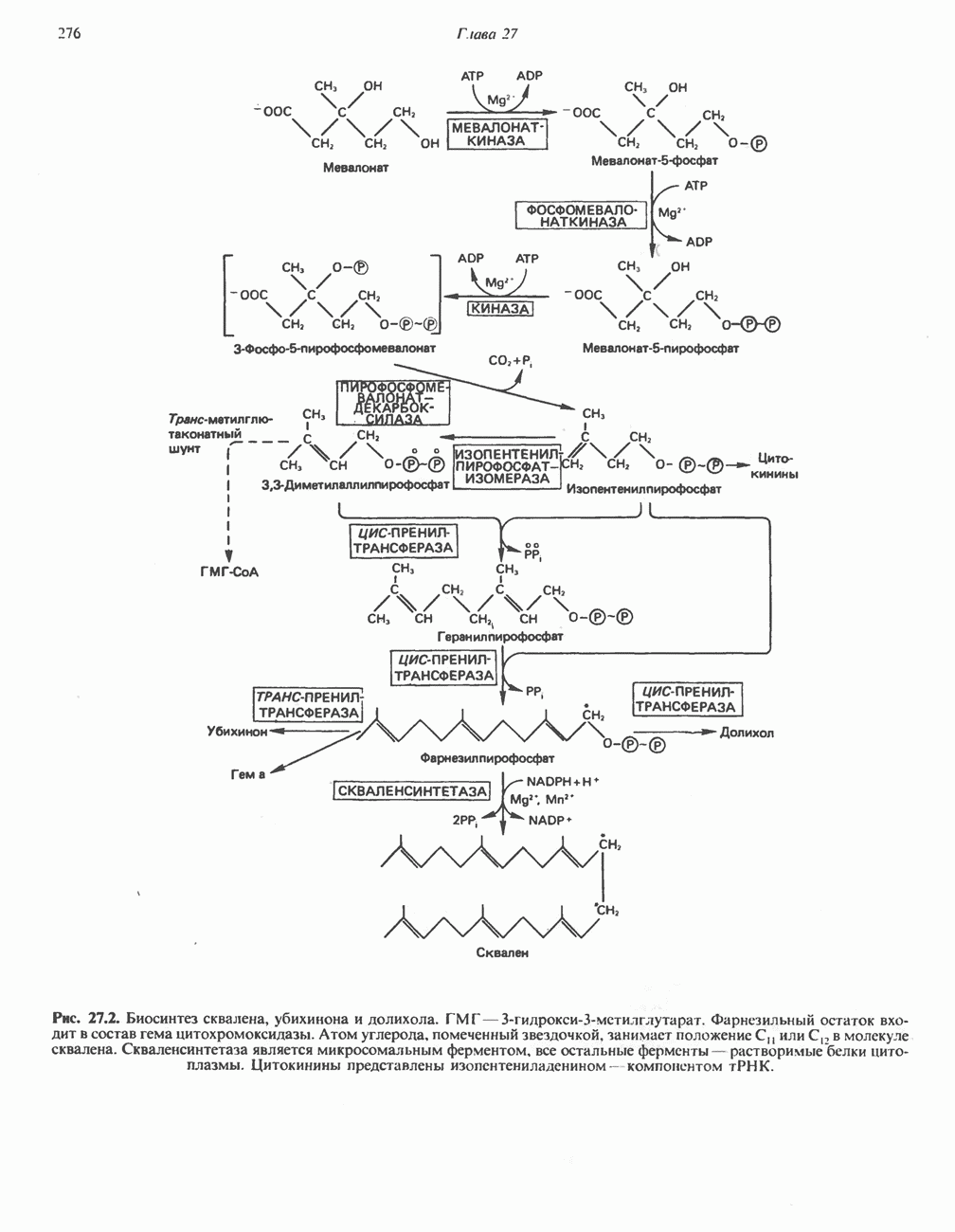
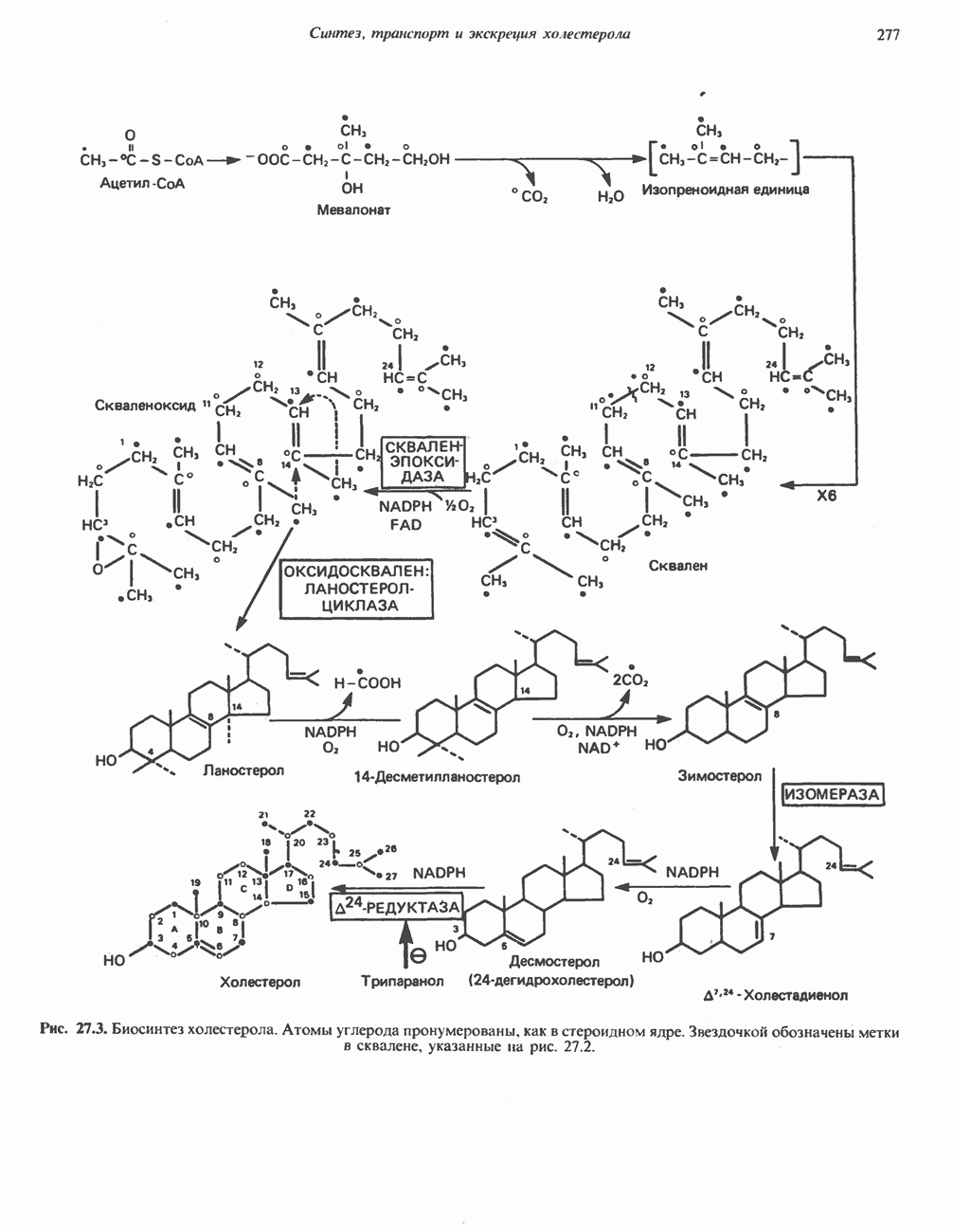
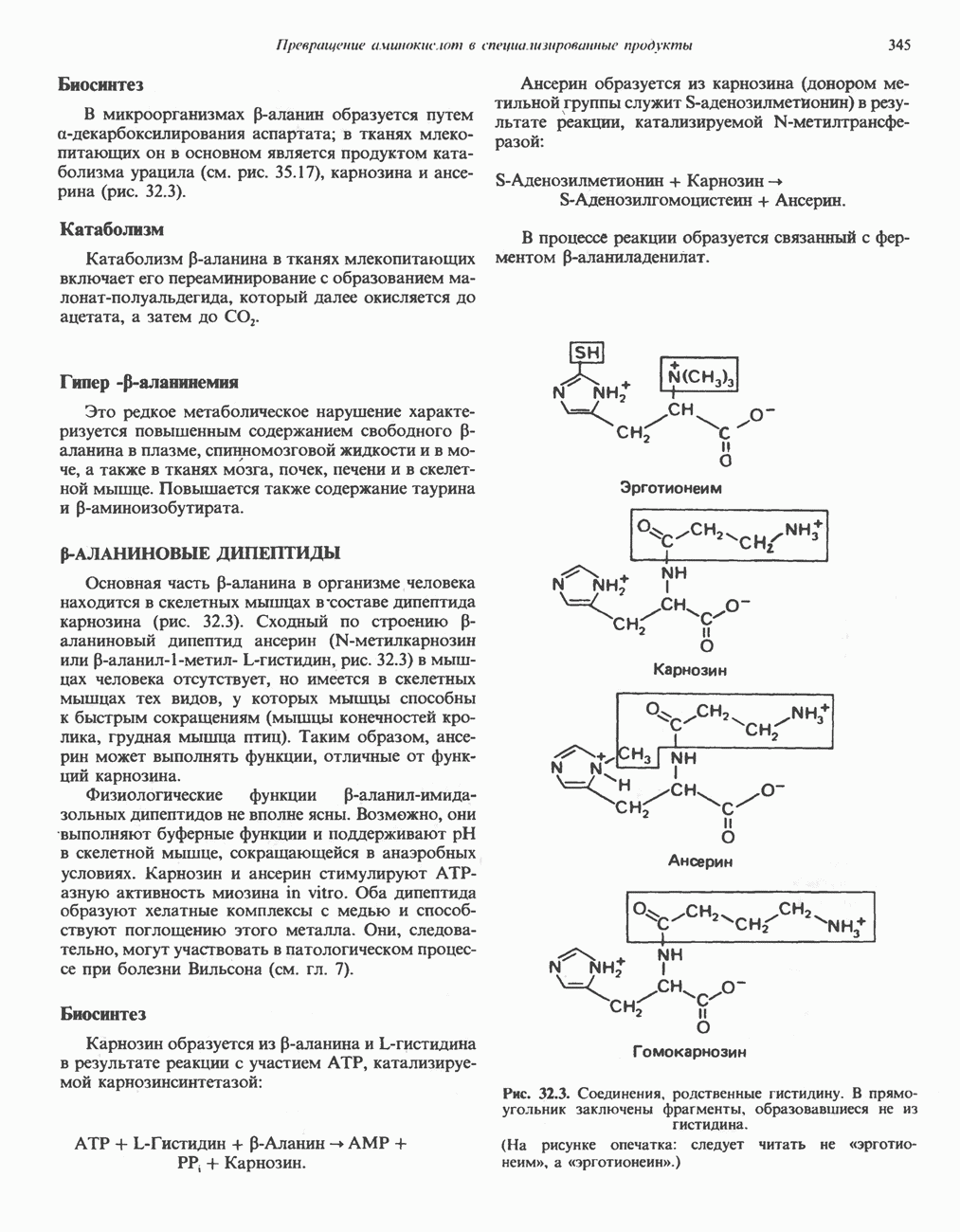
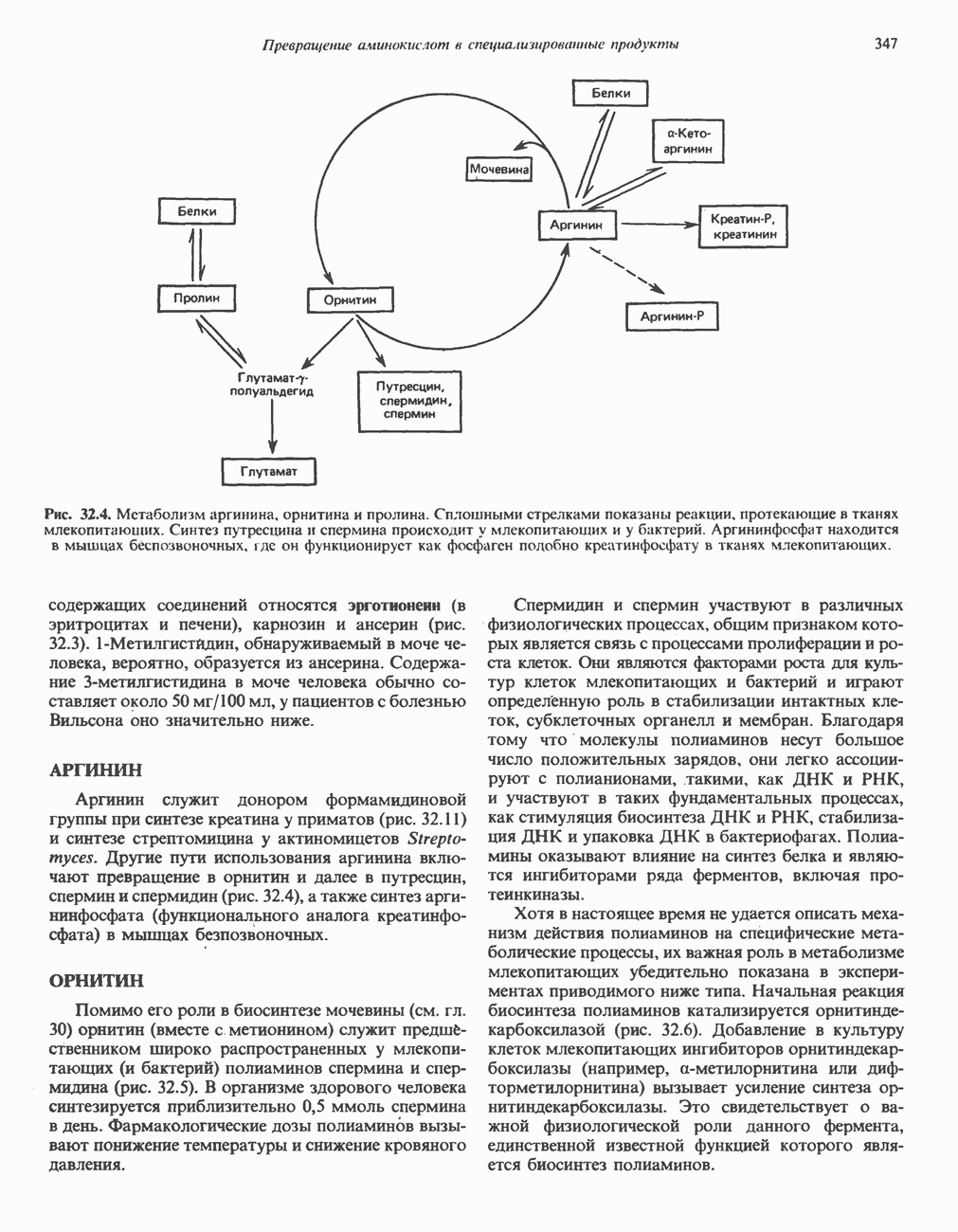
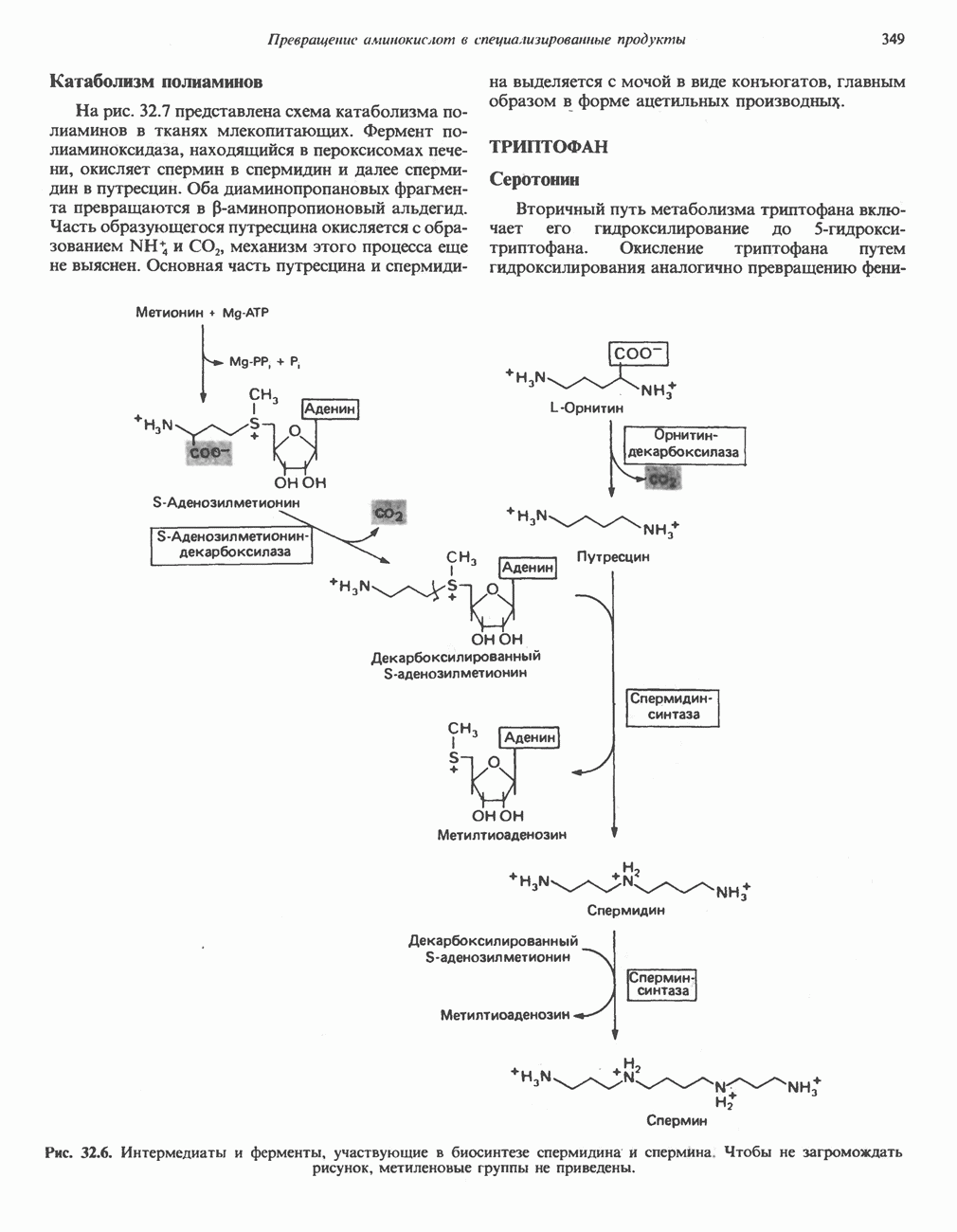
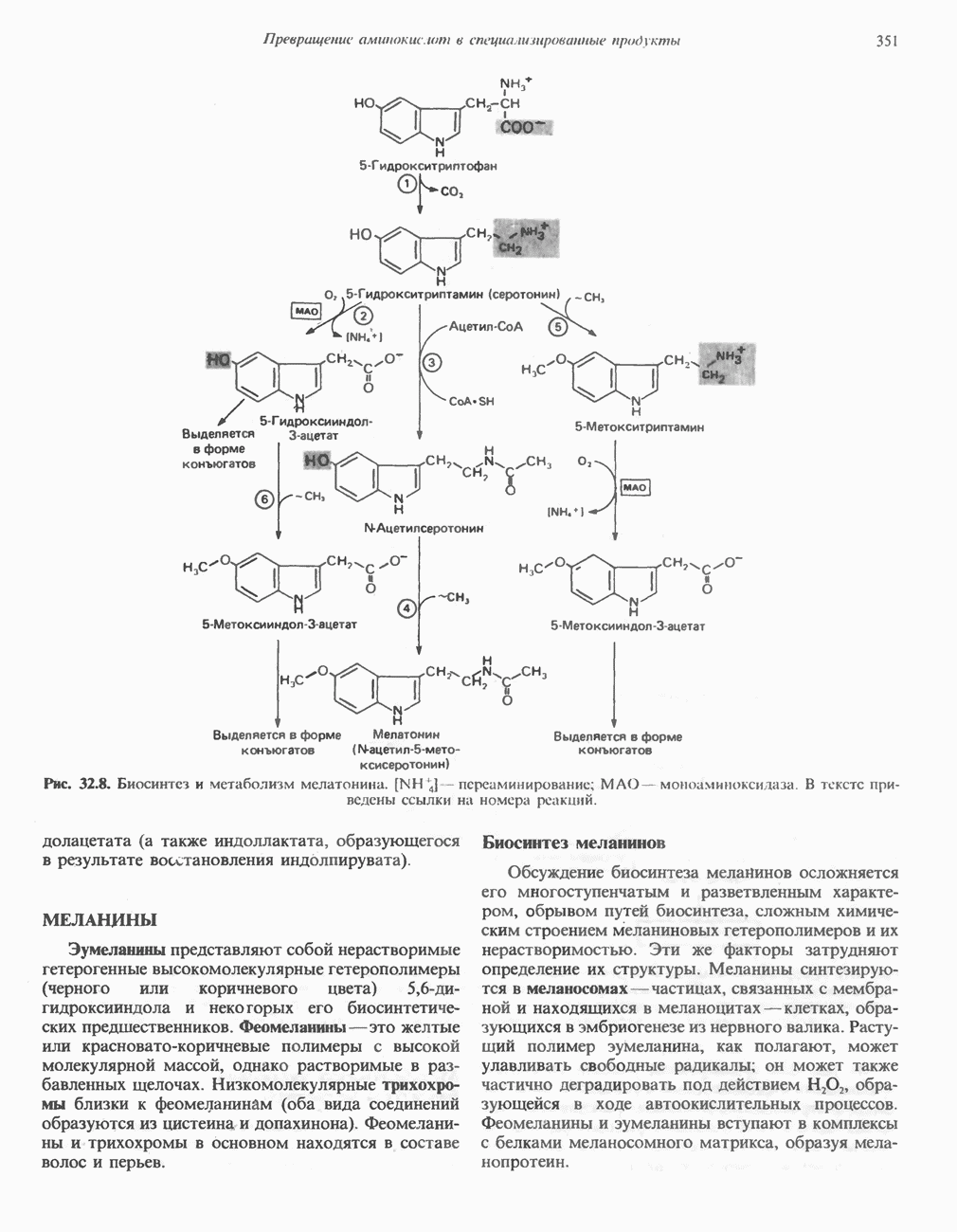
БИОСИНТЕЗ МОЧЕВИНЫ
Общая картина
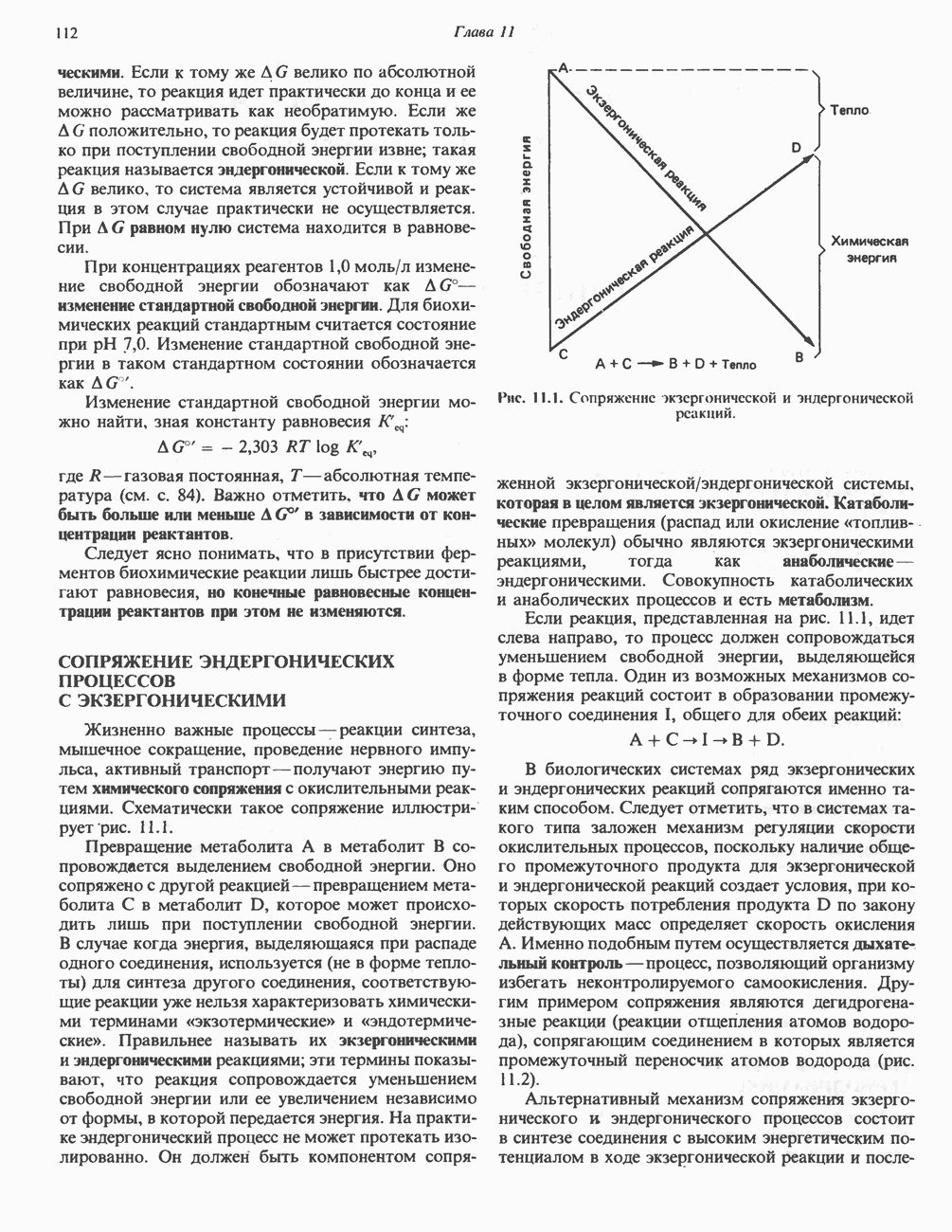
Умеренно активный человек, потребляющий в день около 300 г углеводов, 100 i жира и 100 г пищевого белка, должен за сутки выделять около 16,5 г азота. 95% азота удаляется через почки и остальные 5% - в составе фекалий. Главный путь экскреции азота у человека — в составе мочевины, которая синтезируется в печени, затем поступает в кровь и экскретируется почками. У людей с режимом питания, характерным для западных стран, на долю мочевины приходится 80—90% экскретируемого азота.
Реакции цикла мочевины
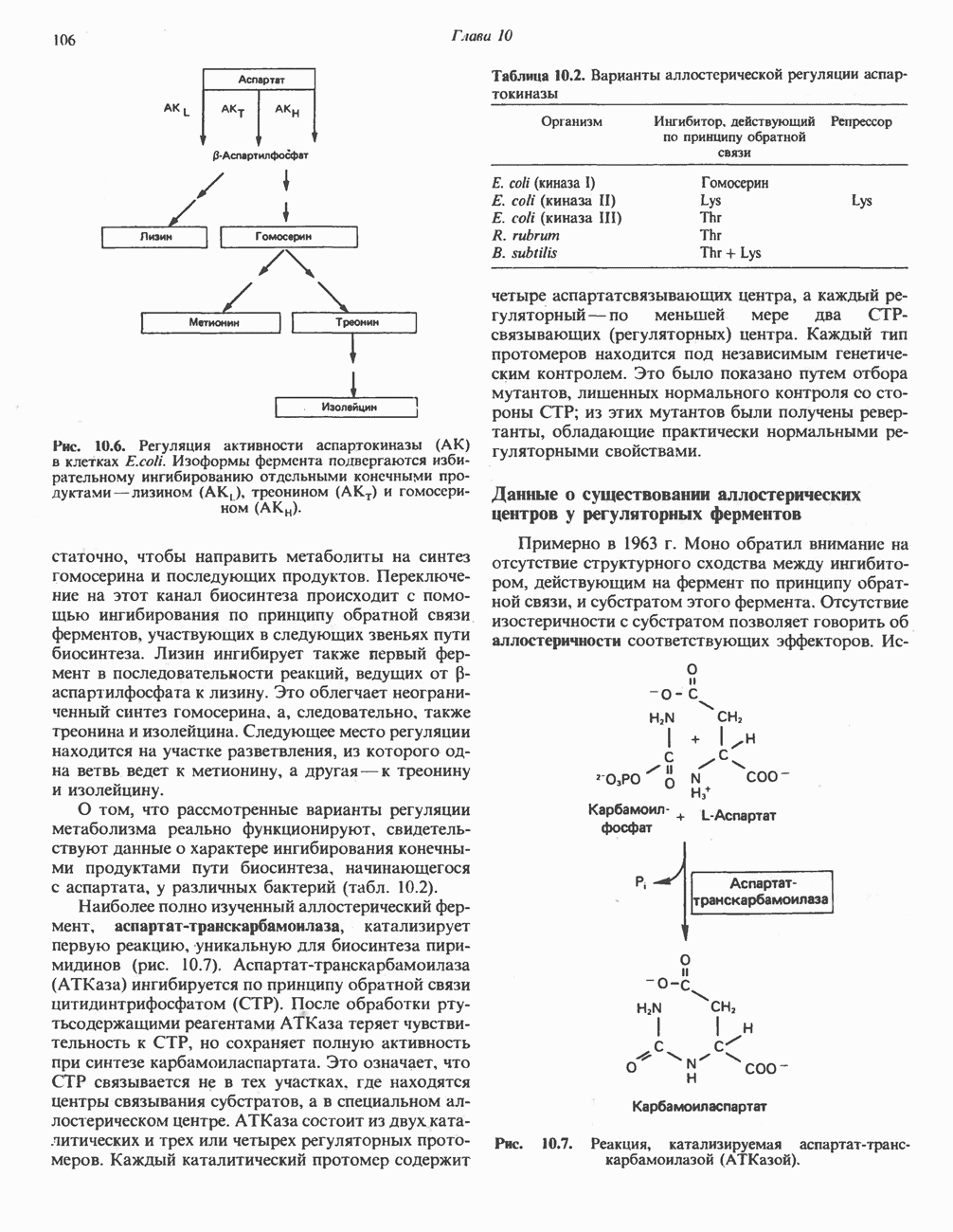
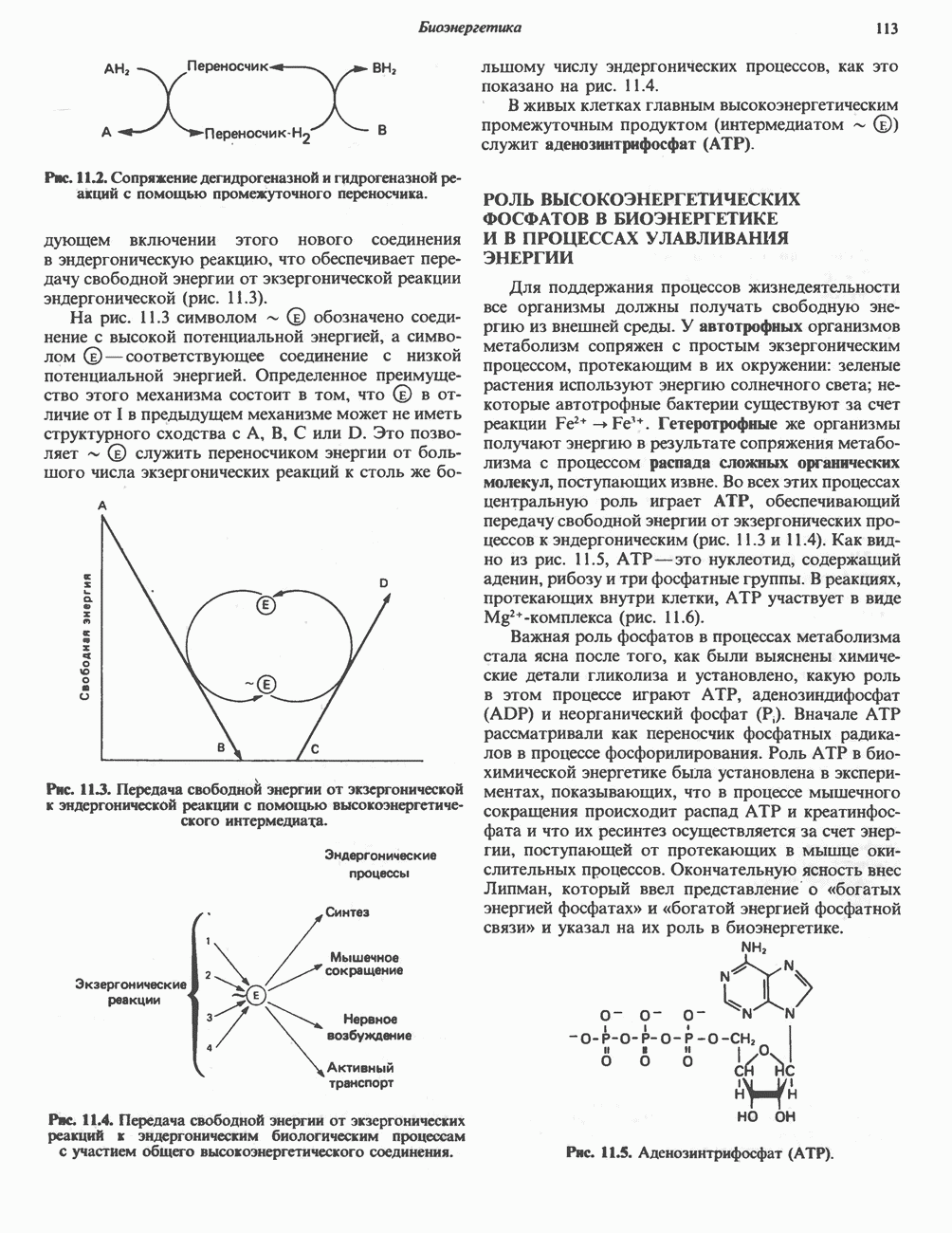
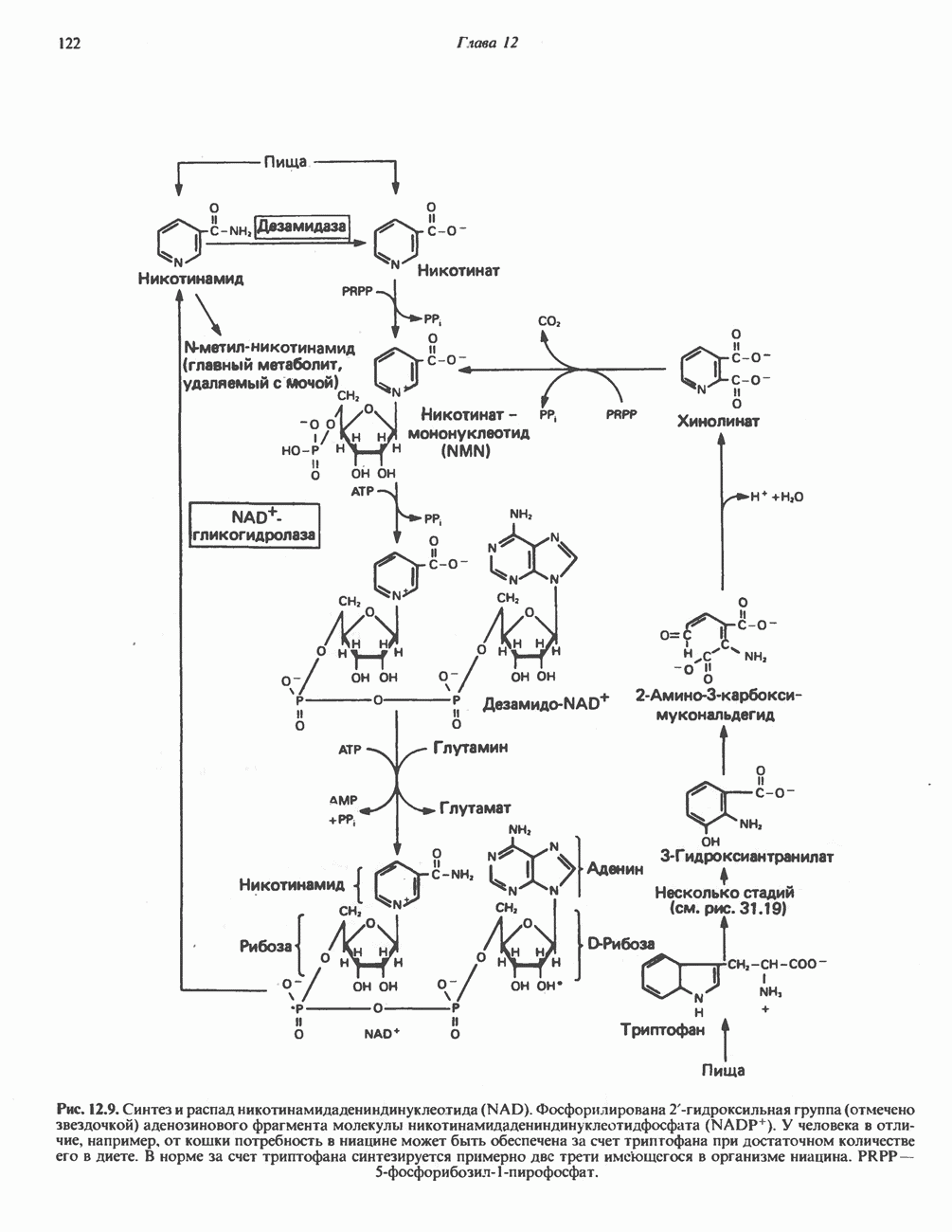
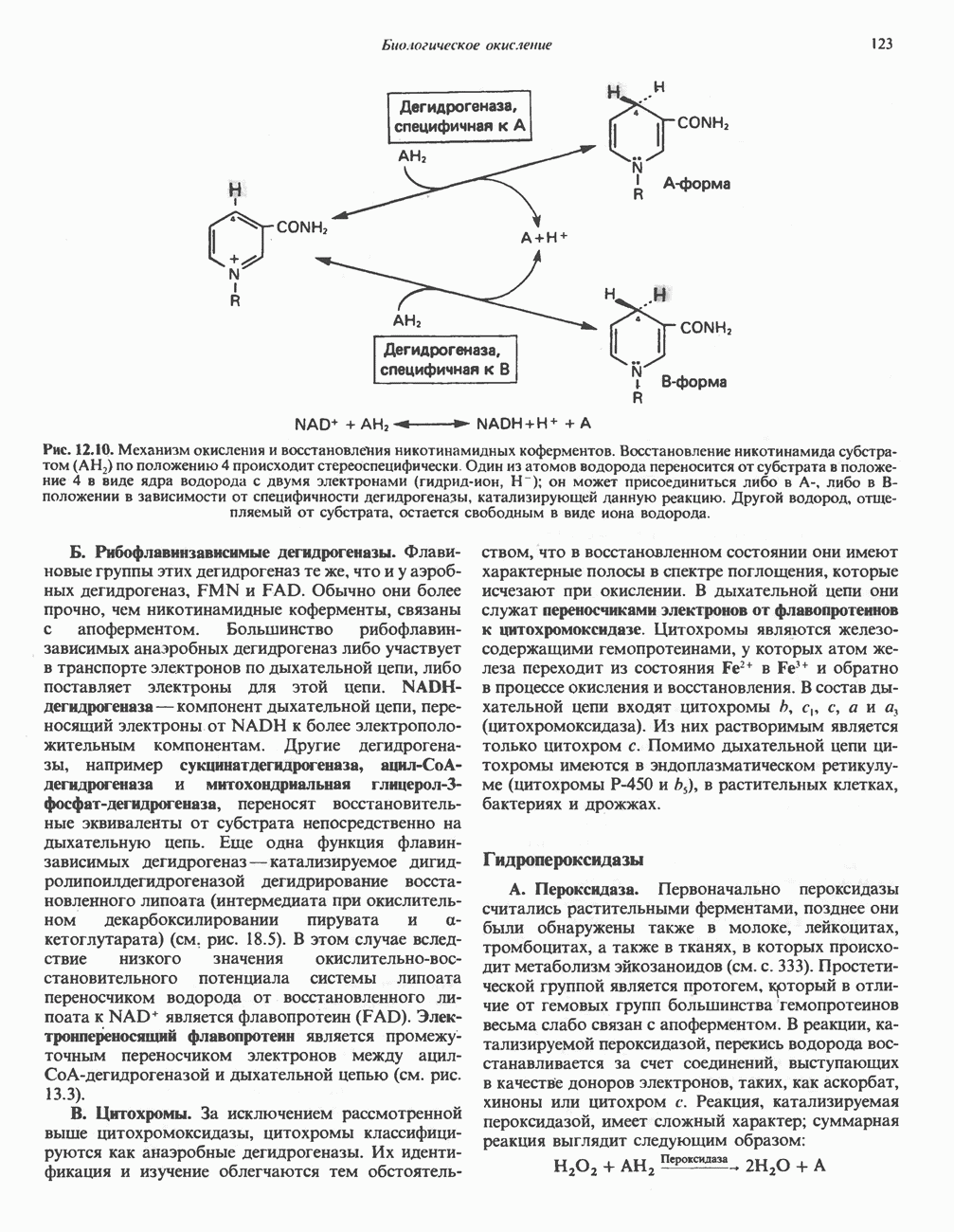
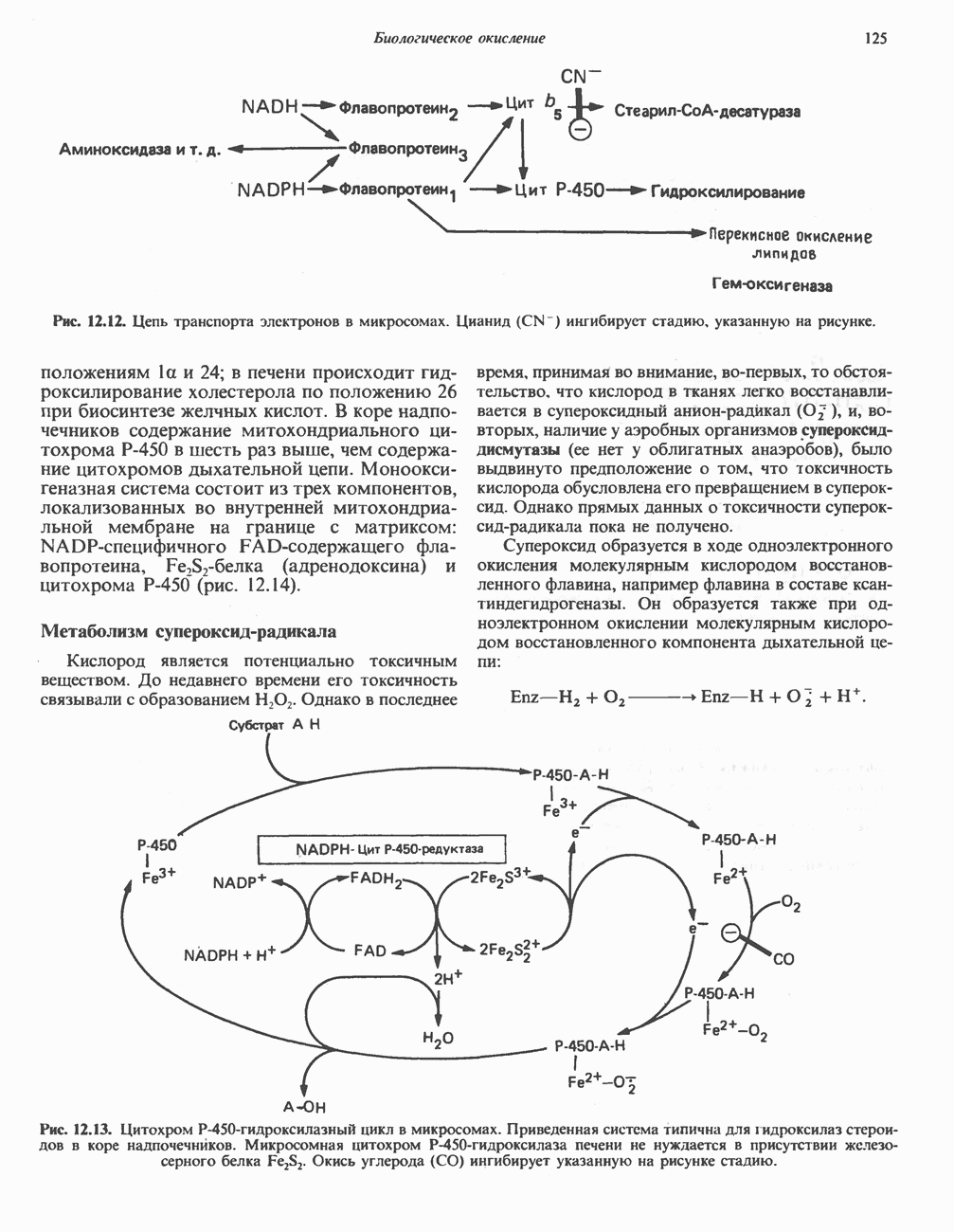
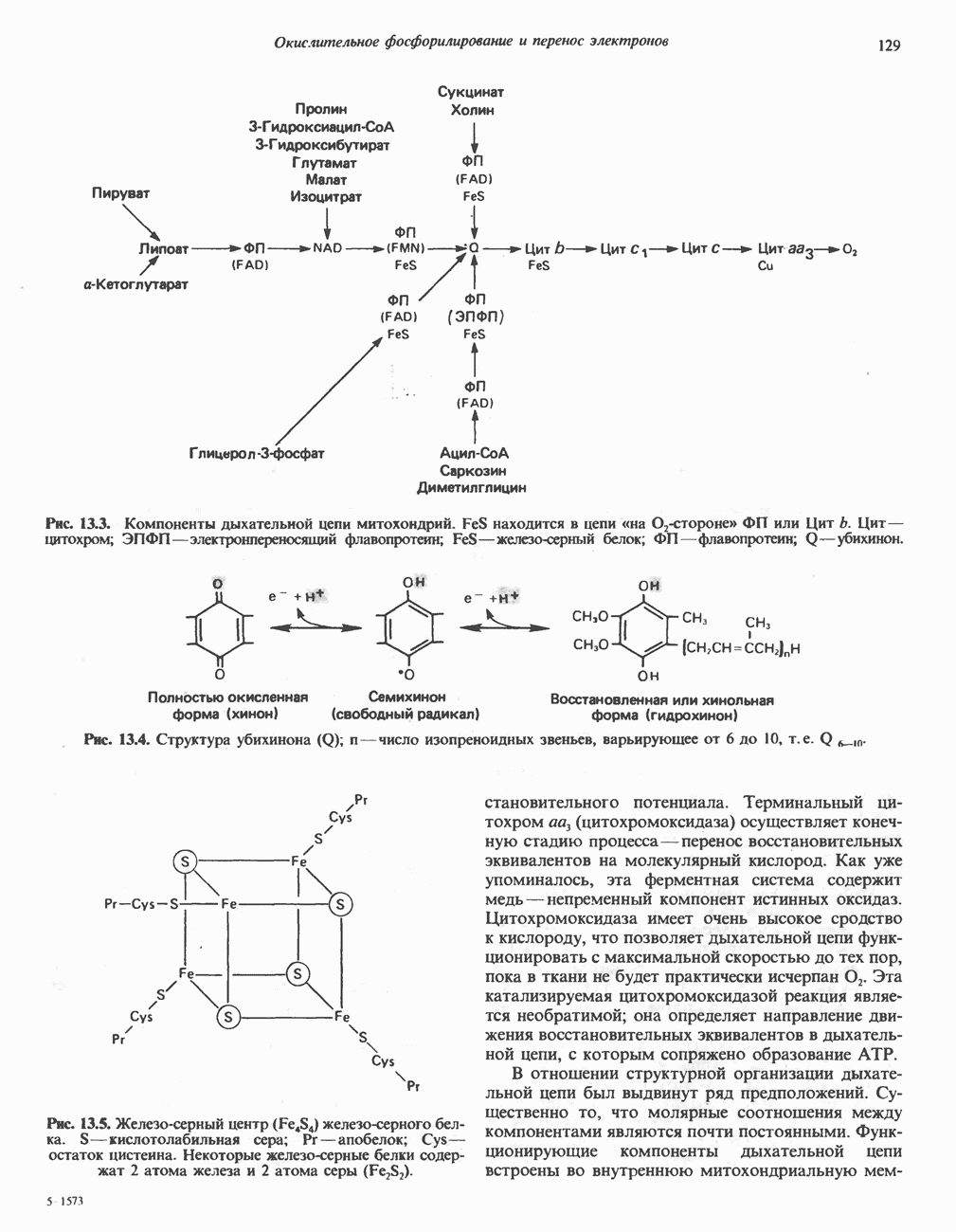
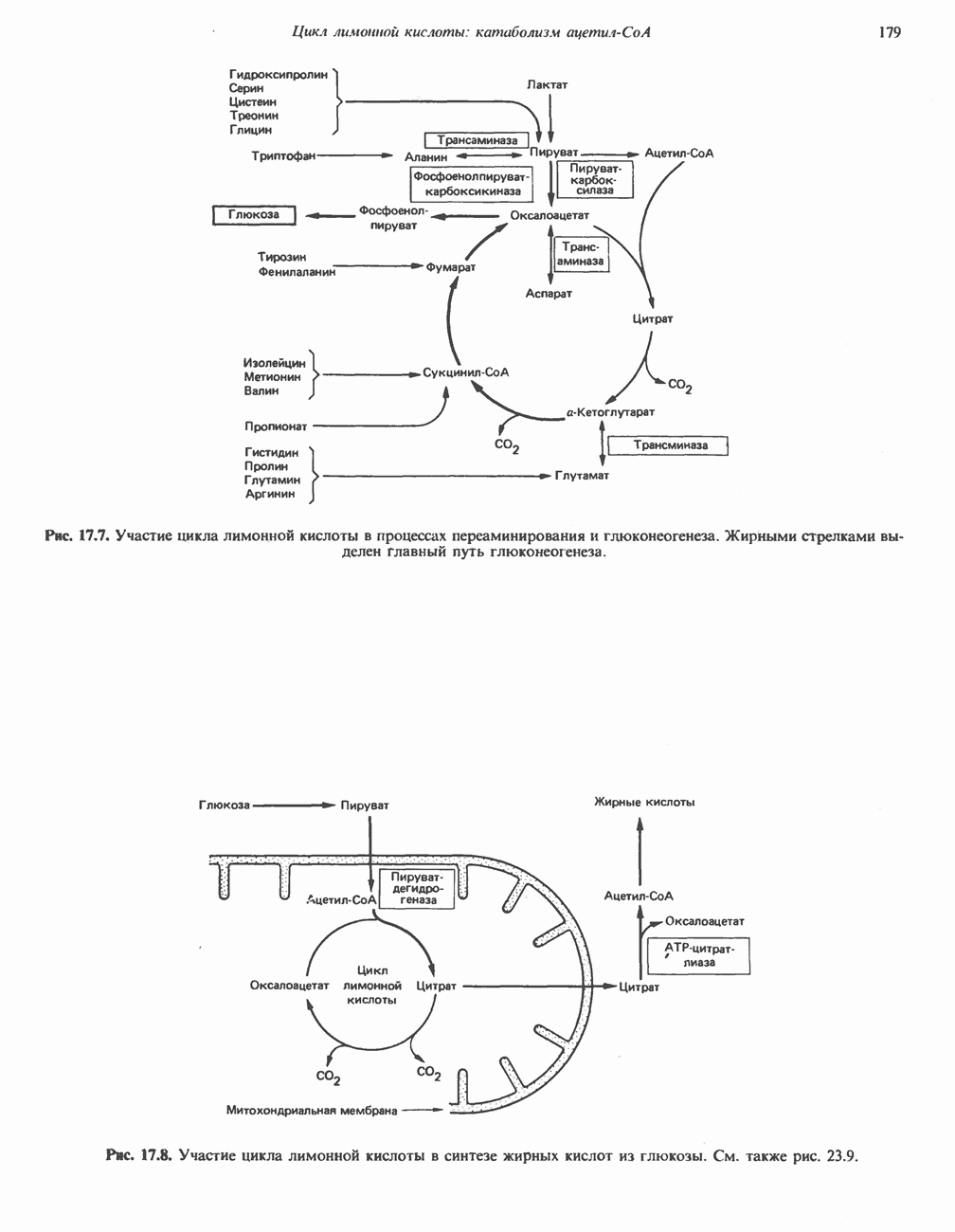
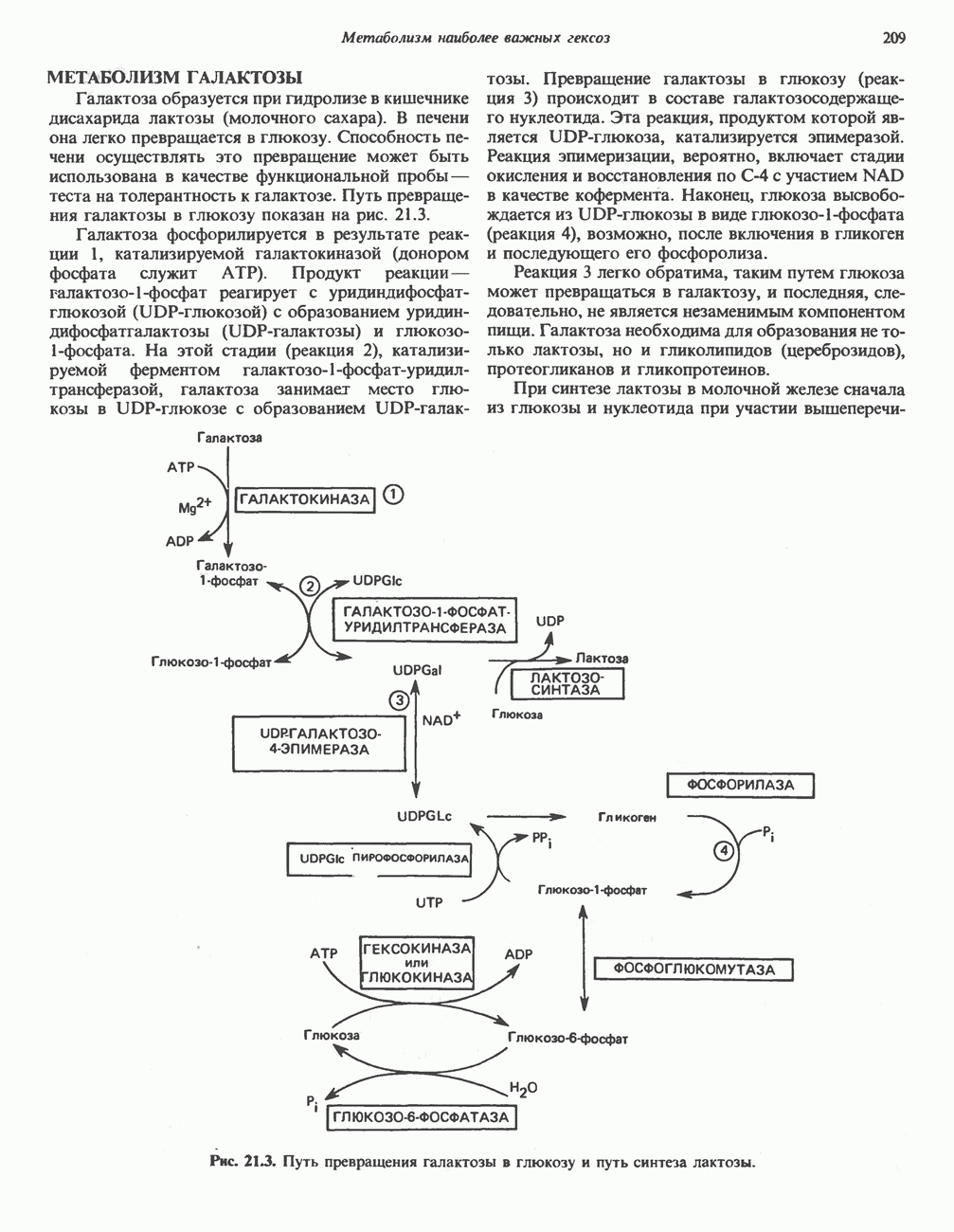
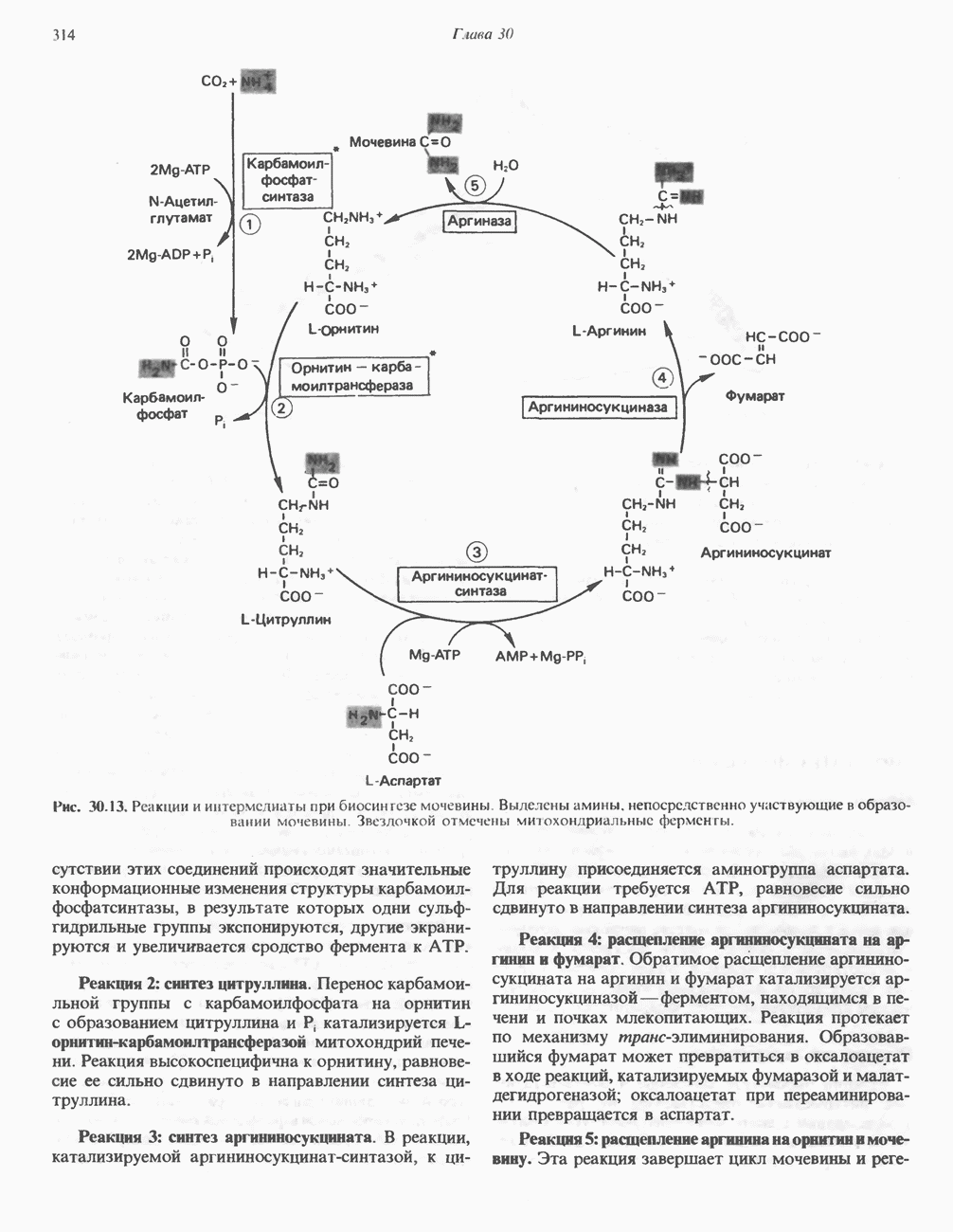
Реакции биосинтеза мочевины и соответствующие интермедиаты - представлены на рис. 30.13; в образовании 1 моля мочевины участвуют 1 моль ионов аммония. 1 моль двуокиси углерода (активируемой  и АТР) и 1 моль а-аминного азота аспартата. В ходе синтеза потребляются 3 моля АТР (2 из них превращаются в ADP и
и АТР) и 1 моль а-аминного азота аспартата. В ходе синтеза потребляются 3 моля АТР (2 из них превращаются в ADP и  а третий — в АМР и
а третий — в АМР и  ), в нем последовательно участвуют пять ферментов, катализирующих реакции 1—5 на рис. 30.13. Из 6 аминокислот, вовлекаемых в синтез мочевины, одна (
), в нем последовательно участвуют пять ферментов, катализирующих реакции 1—5 на рис. 30.13. Из 6 аминокислот, вовлекаемых в синтез мочевины, одна ( -ацетилглутамат) служит активатором одного из ферментов и в химических превращениях не участвует. Остальные пять—аспартат, аргинин, орнитин, цитруллин и аргининосукцинат — служат переносчиками атомов, которые в итоге образуют молекулу мочевины. Первые две из этих аминокислот входят в состав белков, тогда как три другие (орнитин, цитруллин и аргининосукцинат) в состав белков не входят. Главной метаболической ролью этих трех последних аминокислот у млекопитающих является участие в синтезе мочевины. Обратите внимание, что образование мочевины является частично циклическим процессом. Орнитин, участвующий в реакции 2, регенерируется в ходе реакции 5. Таким образом, ни потерь, ни накопления орнитина, ци-труллина, аргининосукцината и аргинина в ходе синтеза мочевины не происходит; потребляются только ион аммония,
-ацетилглутамат) служит активатором одного из ферментов и в химических превращениях не участвует. Остальные пять—аспартат, аргинин, орнитин, цитруллин и аргининосукцинат — служат переносчиками атомов, которые в итоге образуют молекулу мочевины. Первые две из этих аминокислот входят в состав белков, тогда как три другие (орнитин, цитруллин и аргининосукцинат) в состав белков не входят. Главной метаболической ролью этих трех последних аминокислот у млекопитающих является участие в синтезе мочевины. Обратите внимание, что образование мочевины является частично циклическим процессом. Орнитин, участвующий в реакции 2, регенерируется в ходе реакции 5. Таким образом, ни потерь, ни накопления орнитина, ци-труллина, аргининосукцината и аргинина в ходе синтеза мочевины не происходит; потребляются только ион аммония,  , АТР и аспартат.
, АТР и аспартат.
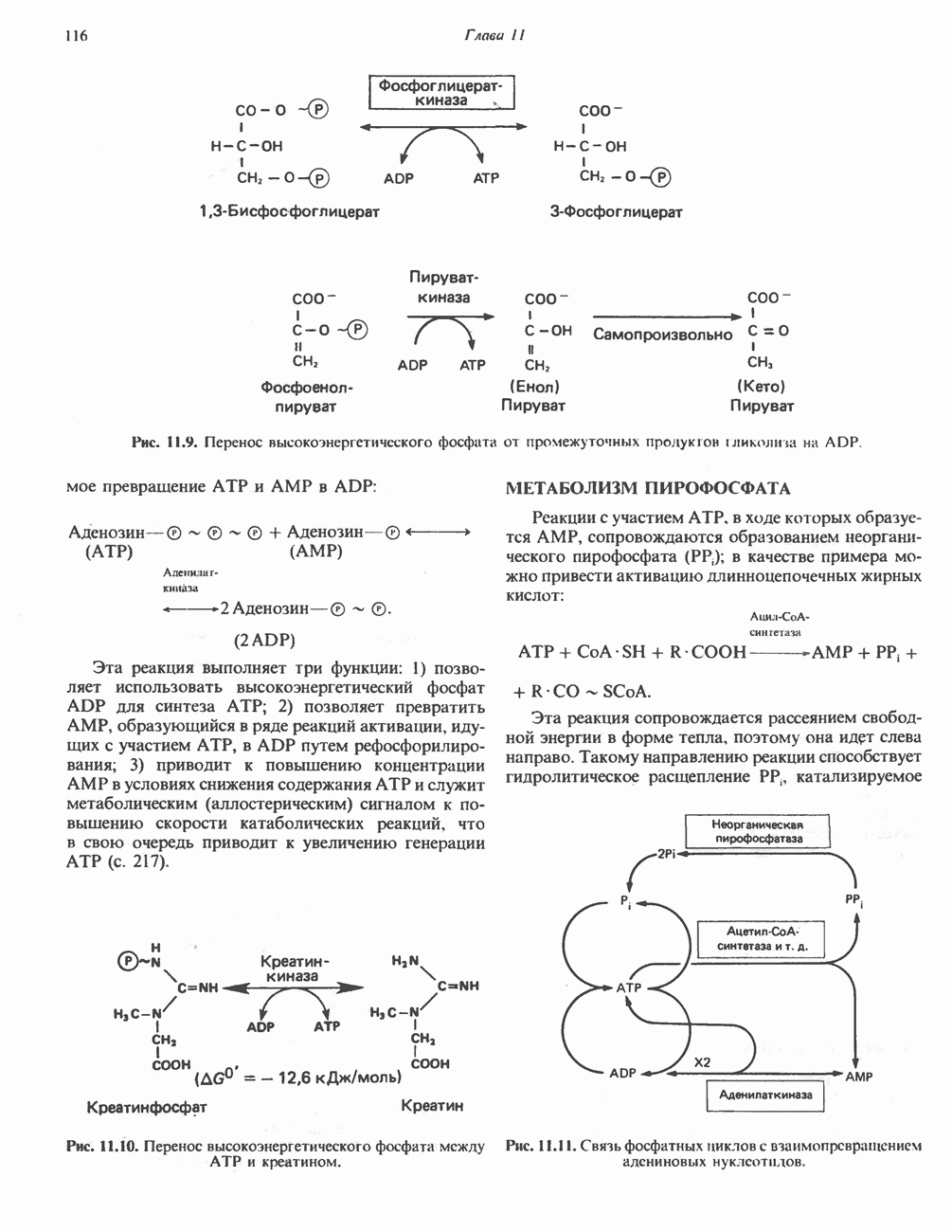
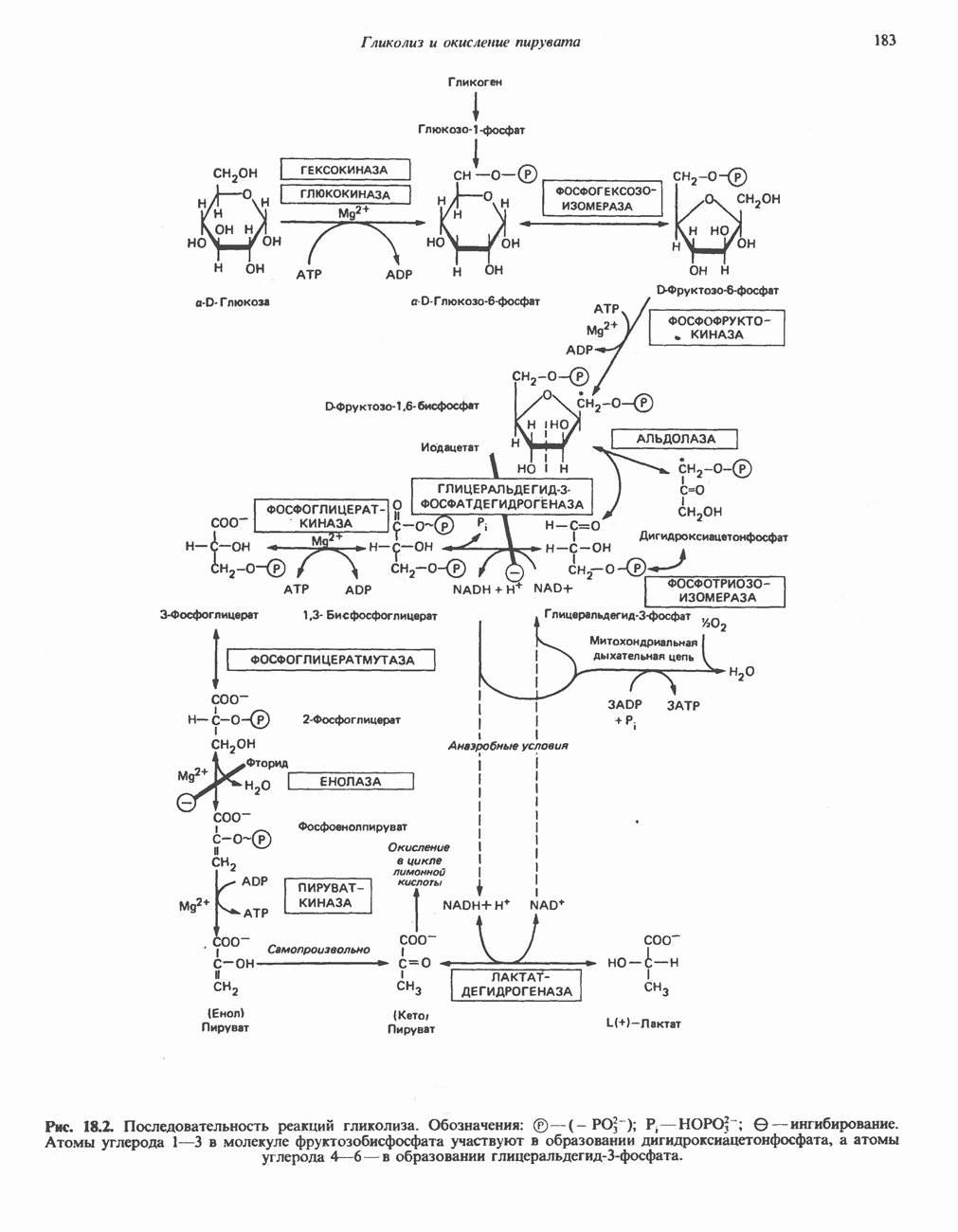
Реакция 1: синтез карбамонлфосфата. Конденсация иона аммония, двуокиси углерода и фосфата (поступаюше! о от АТР), которая приводит к образованию карбамонлфосфата, катализируется карбамоилфосфатсинтазой—ферментом, находящимся в митохондриях печени всех уреотелических организмов, включая человека. Осуществляемый в ходе этой реакции гидролиз двух молекул АТР обеспечивает энергией образование двух ковалентных связей: амидной связи и ангидридной связи при образовании карбамоил-фосфата из карбоновой и фосфорной кислот. Для данной реакции требуются ионы  , а также дикарбоновая кислота, предпочтительно N-ацетилглутамат. В присутствии
, а также дикарбоновая кислота, предпочтительно N-ацетилглутамат. В присутствии
(см. скан)
Рис. 30.13. Реакции и интермедиаты при биосинтезе мочевины. Вылетены амины, непосредственно участвующие в образовании мочевины. Звезлочкой отмечены митохондриальные ферменты.
этих соединений происходят значительные конформационные изменения структуры карбамоил-фосфатсинтазы, в результате которых одни сульфгидрильные группы экспонируются, другие экранируются и увеличивается сродство фермента к АТР.
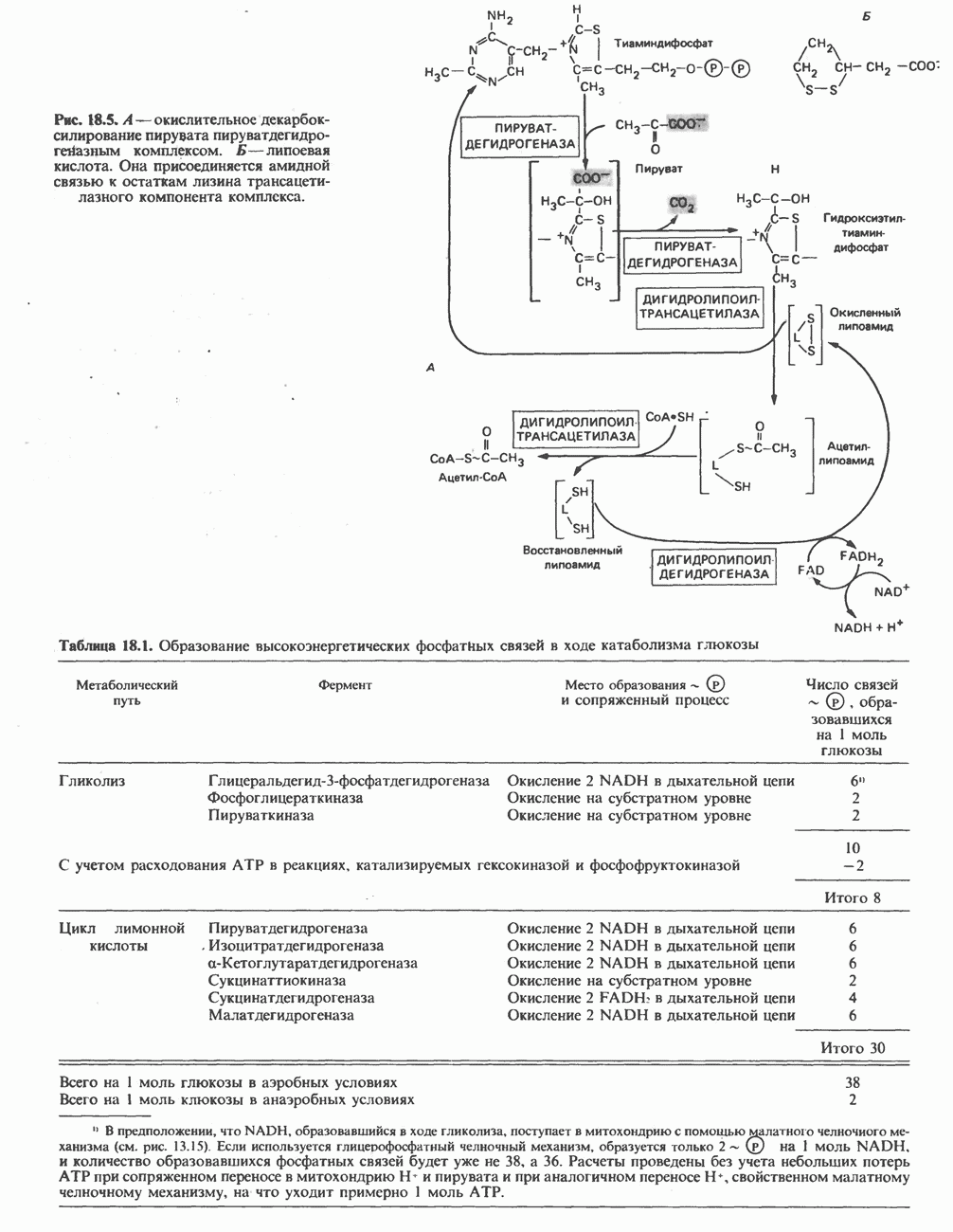
Реакция 2: синтез цитруллнна. Перенос карбамоильной группы с карбамонлфосфата на орнитин с образованием цитруллина и  катализируется L-орнитин-карбамоилтрансферазой митохондрий печени. Реакция высокоспецифична к орнитину, равновесие ее сильно сдвинуто в направлении синтеза цитруллина.
катализируется L-орнитин-карбамоилтрансферазой митохондрий печени. Реакция высокоспецифична к орнитину, равновесие ее сильно сдвинуто в направлении синтеза цитруллина.
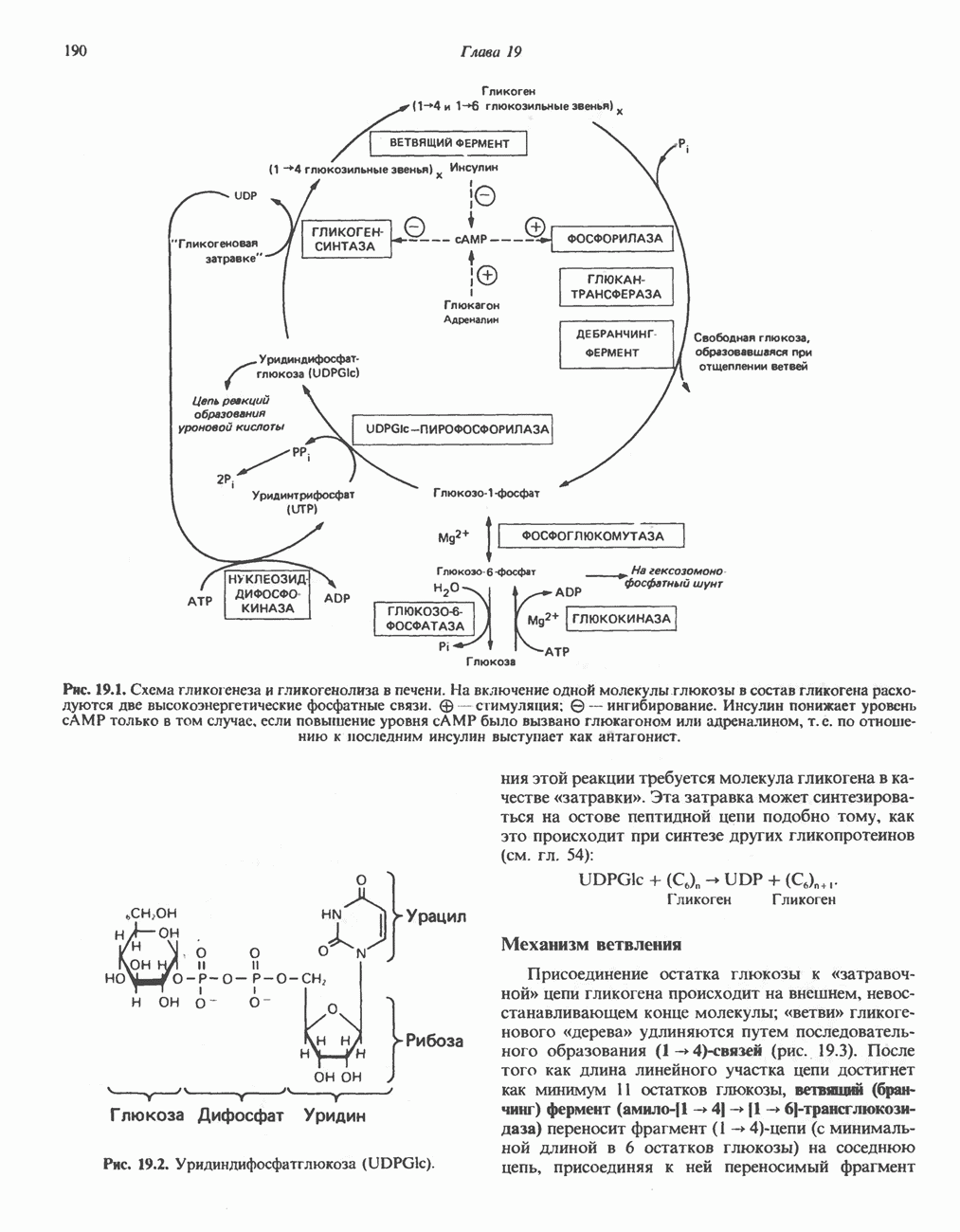
Реакция 3: синтез аргининосукцината. В реакции, катализируемой аргининосукцинат-синтазой, к цитруллину присоединяется аминогруппа аспартата. Для реакции требуется АТР, равновесие сильно сдвинуто в направлении синтеза аргининосукцината.
Реакция 4: расщепление аргининосукцината на аргинин и фумарат. Обратимое расщепление аргининосукцината на аргинин и фумарат катализируется аргининосукциназой—ферментом, находящимся в печени и почках млекопитающих. Реакция протекает по механизму элиминирования. Образовавшийся фумарат может превратиться в оксалоацетат в ходе реакций, катализируемых фумаразой и малат-дегидрогеназой; оксалоацетат при переаминировании превращается в аспартат.
Реакция 5: расщепление аргинина на орнитин и мочевину. Эта реакция завершает цикл мочевины и регенерирует











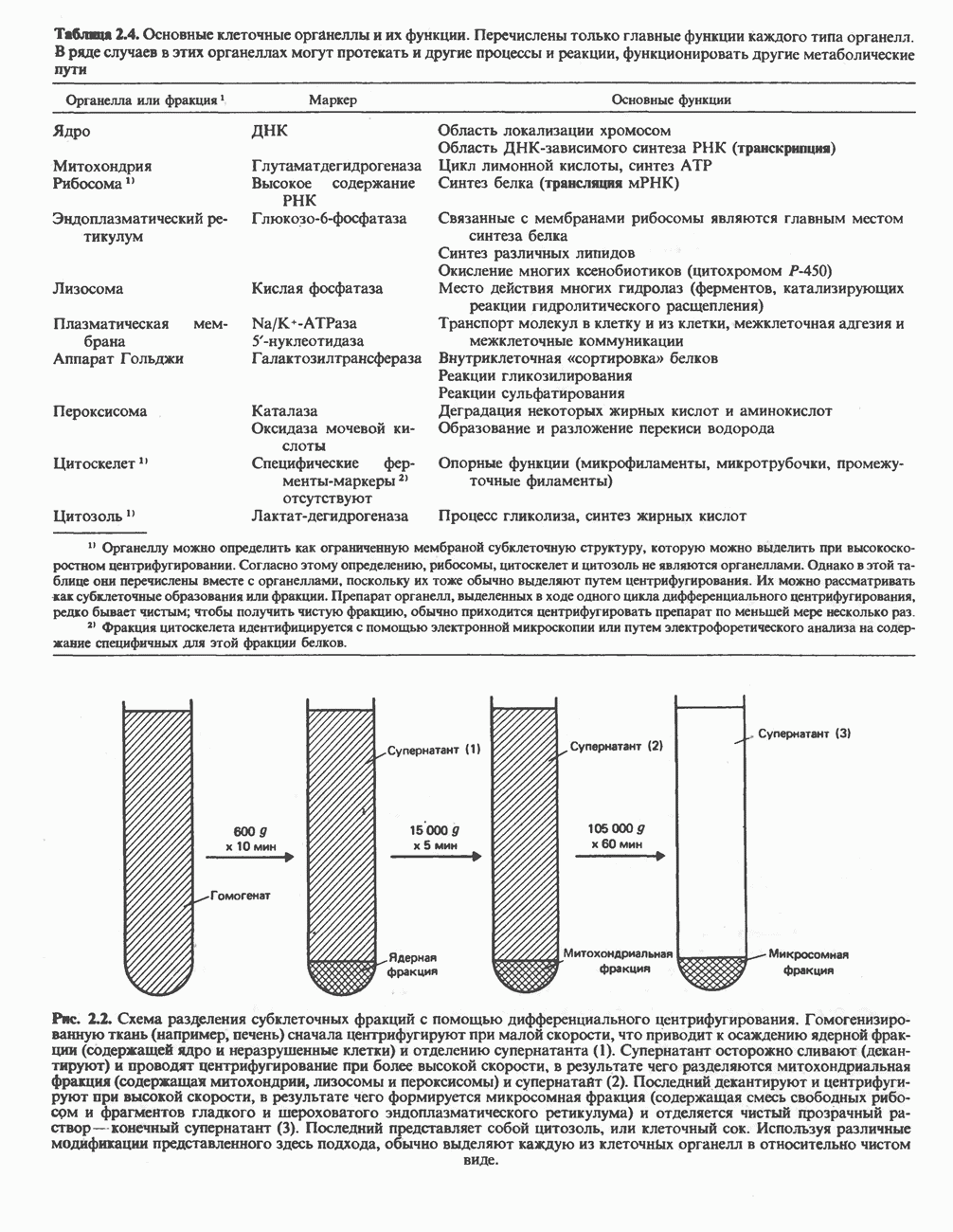







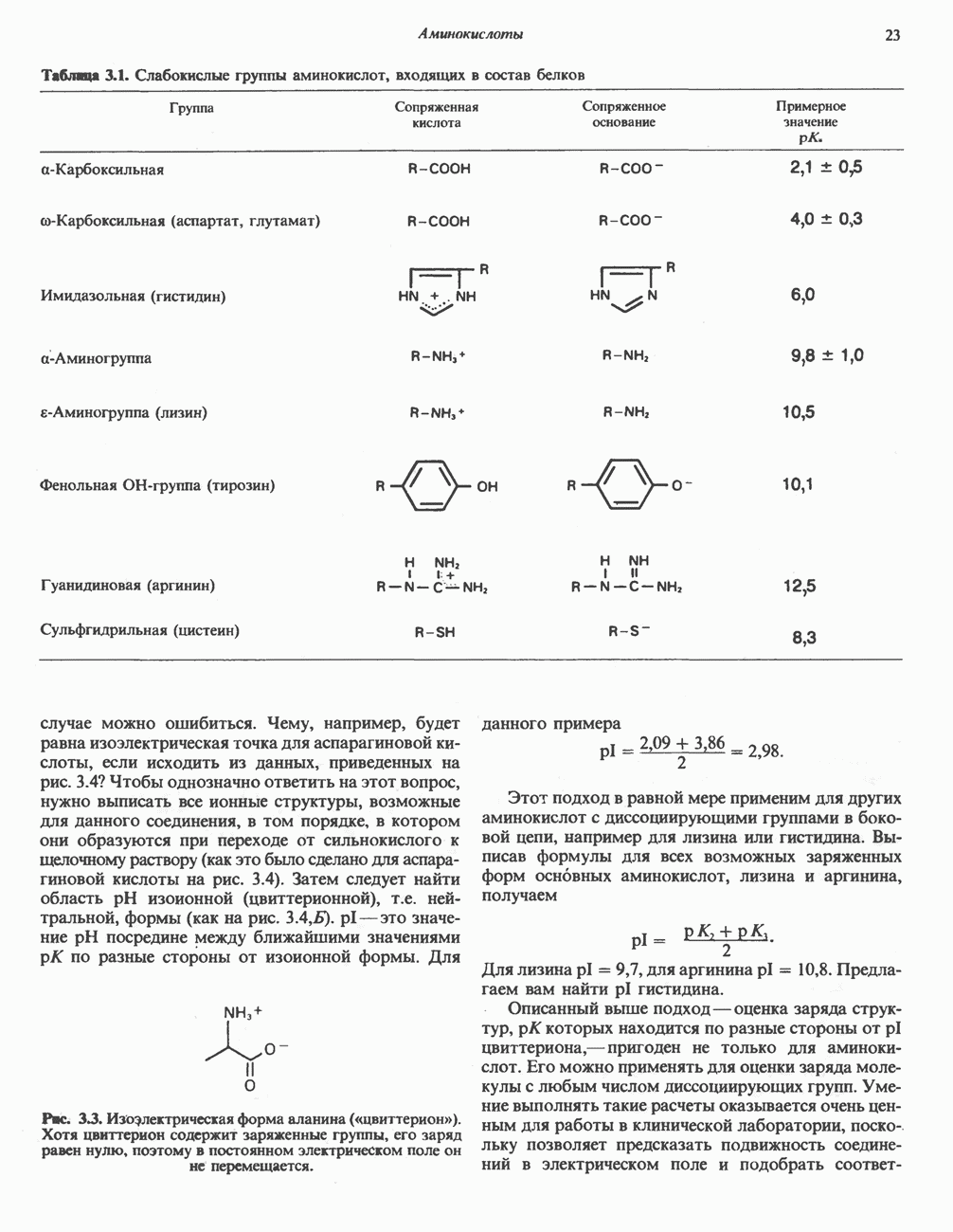





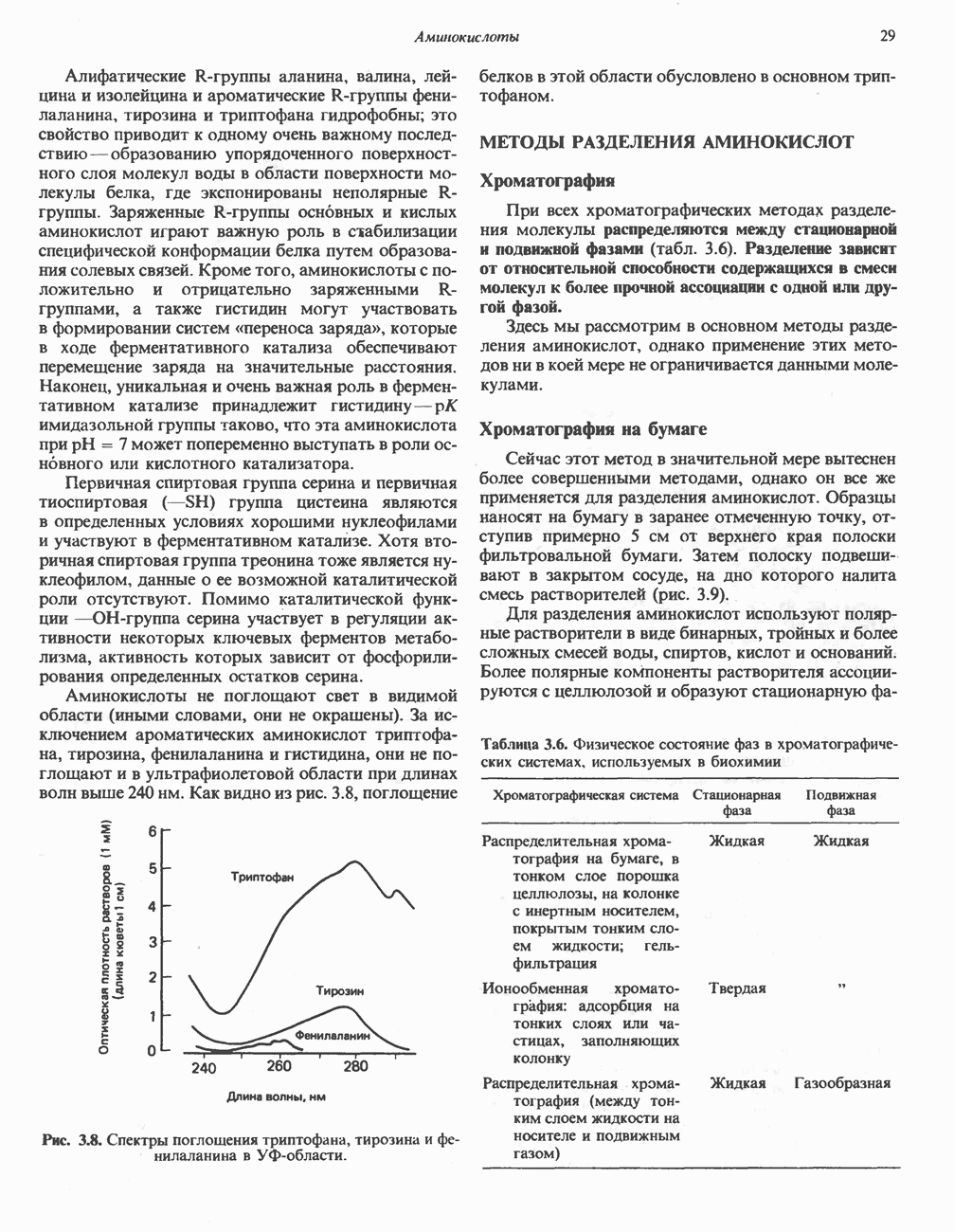
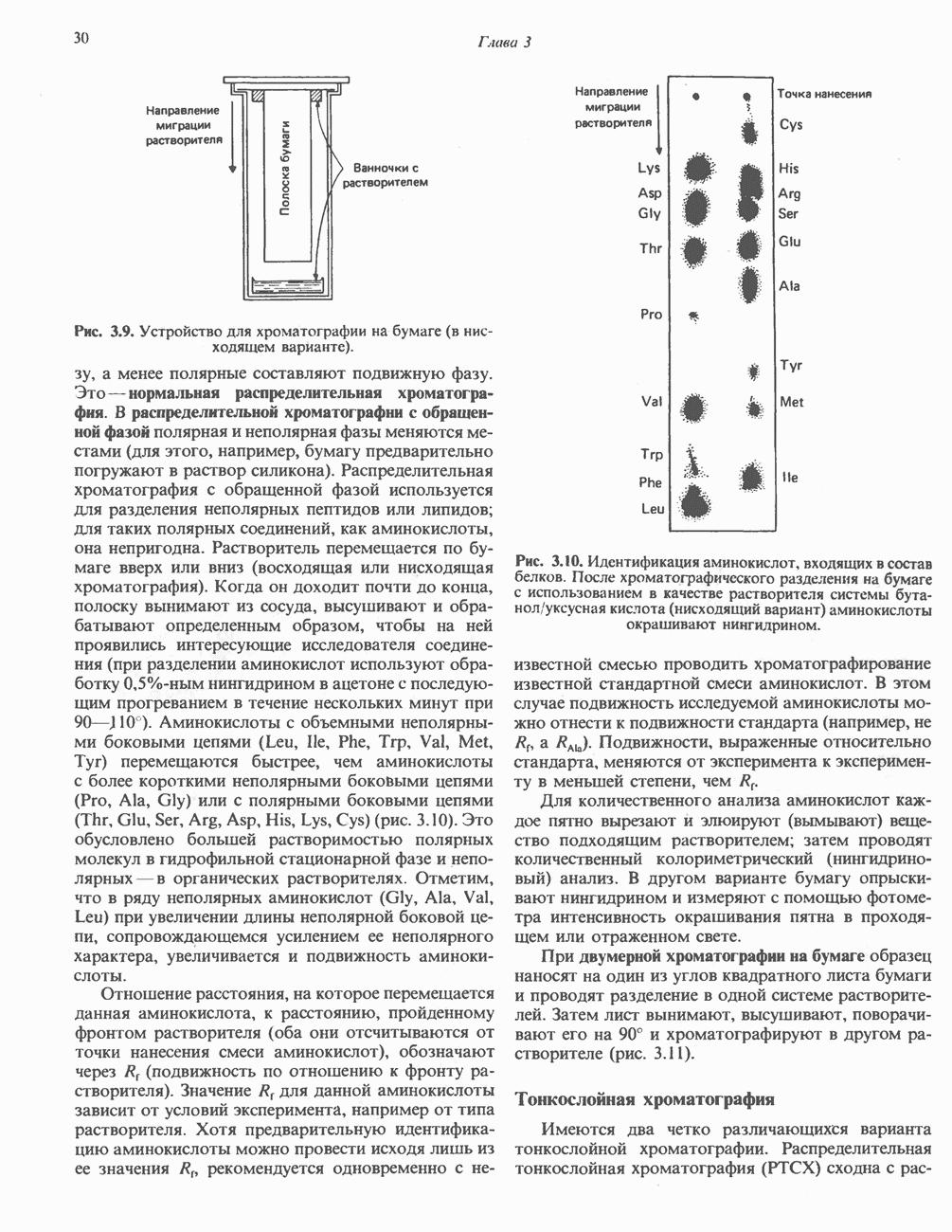

































































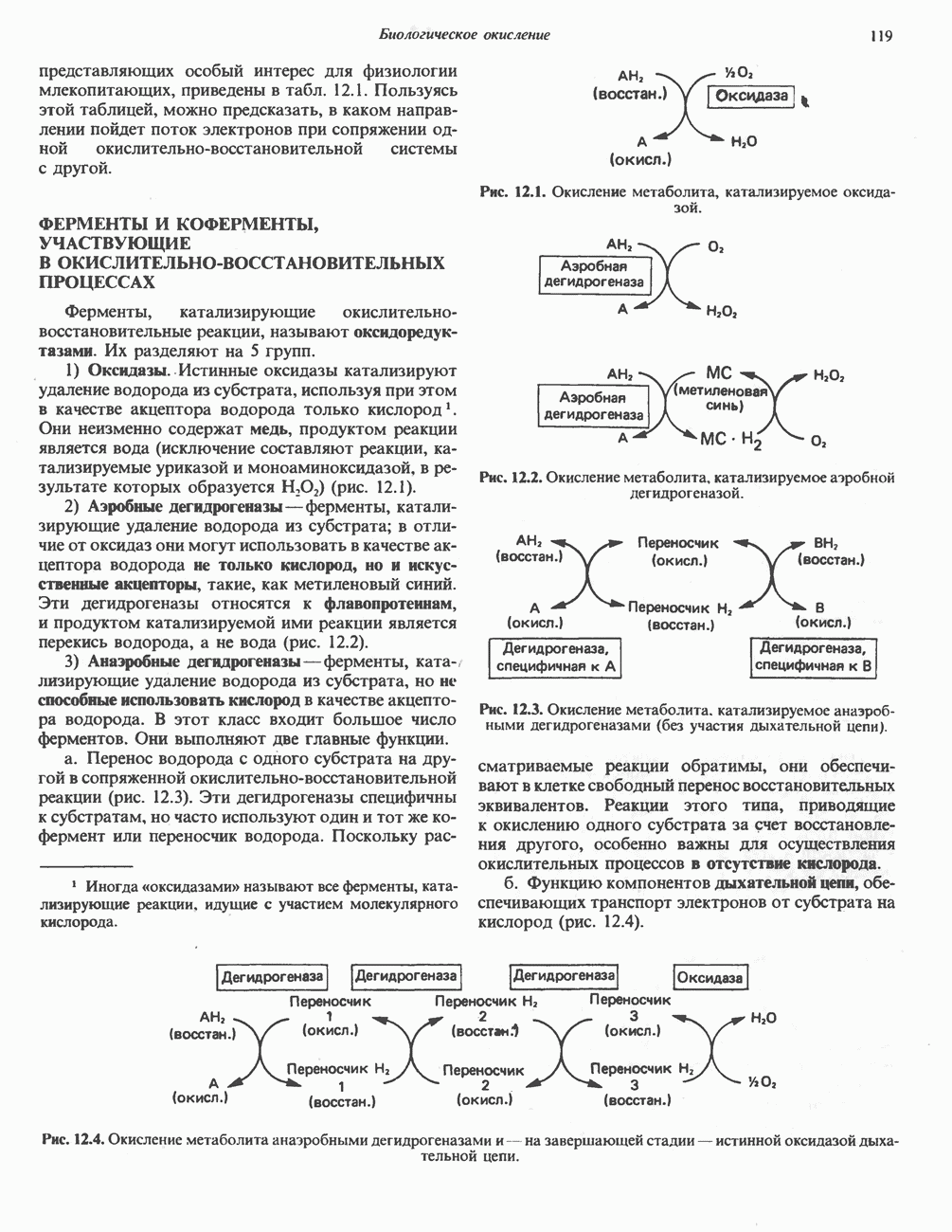














































































































































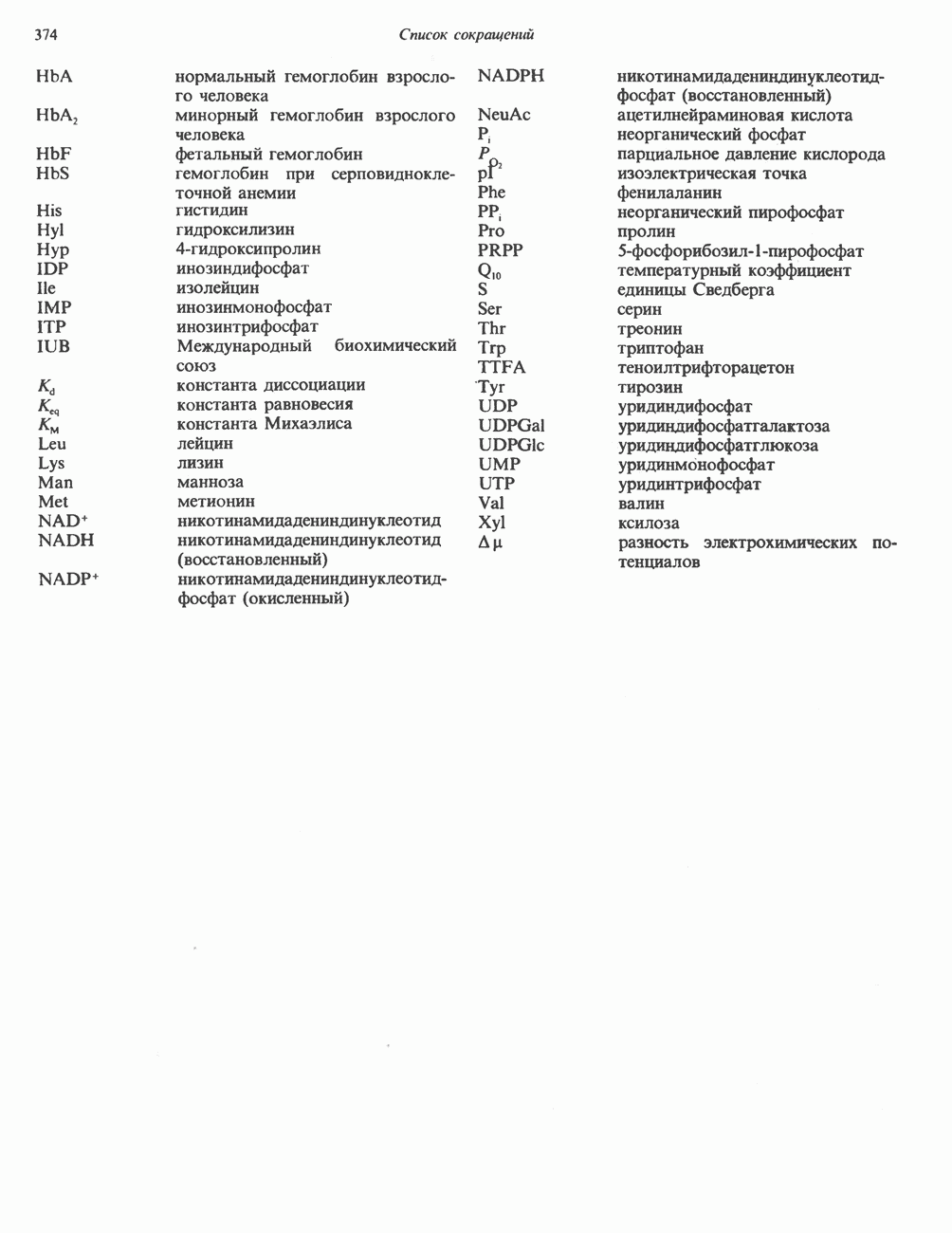
 или
или  Сильными ингибиторами фермента являются орнитин и лизин, конкурирующие с аргинином
Сильными ингибиторами фермента являются орнитин и лизин, конкурирующие с аргинином 
