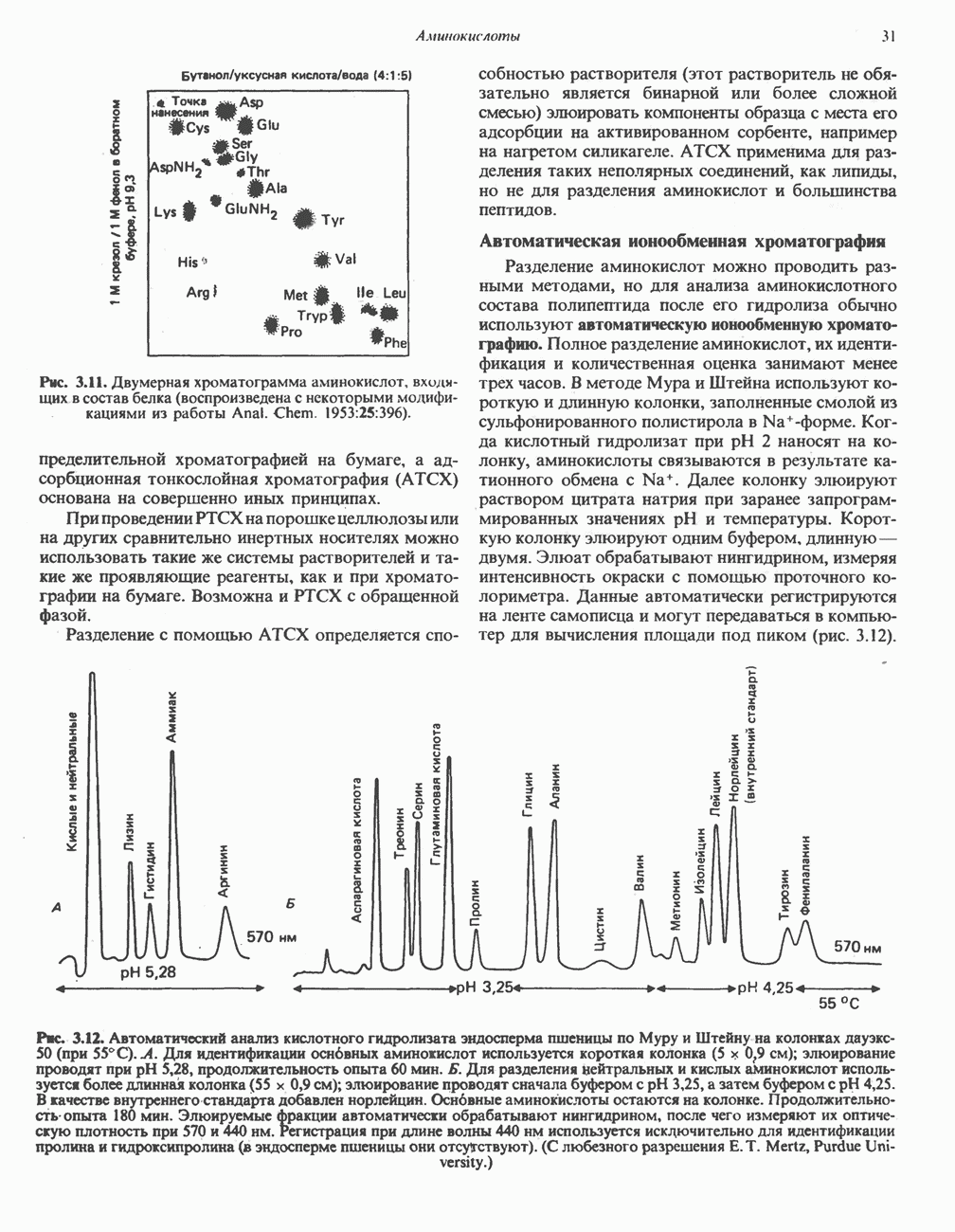
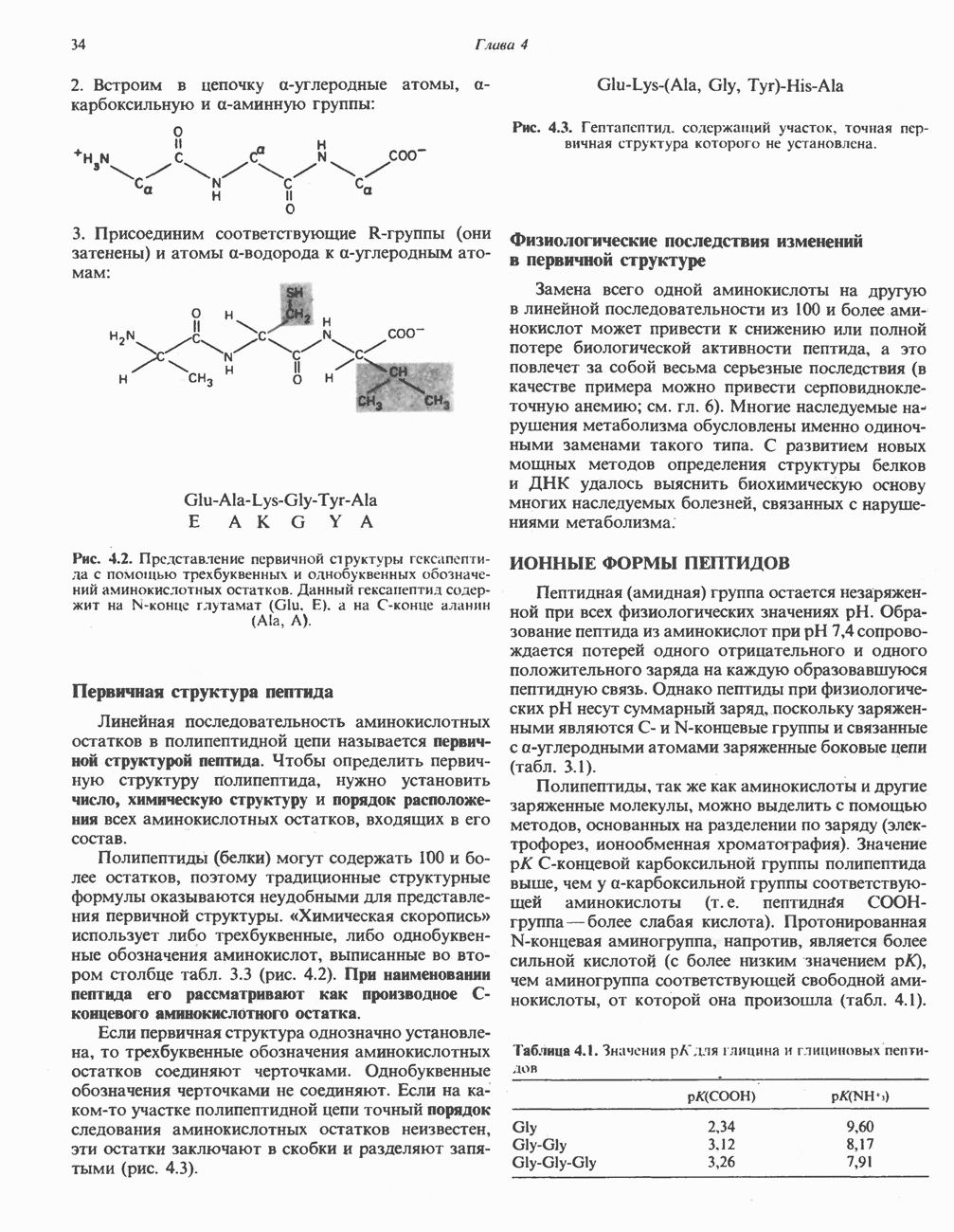
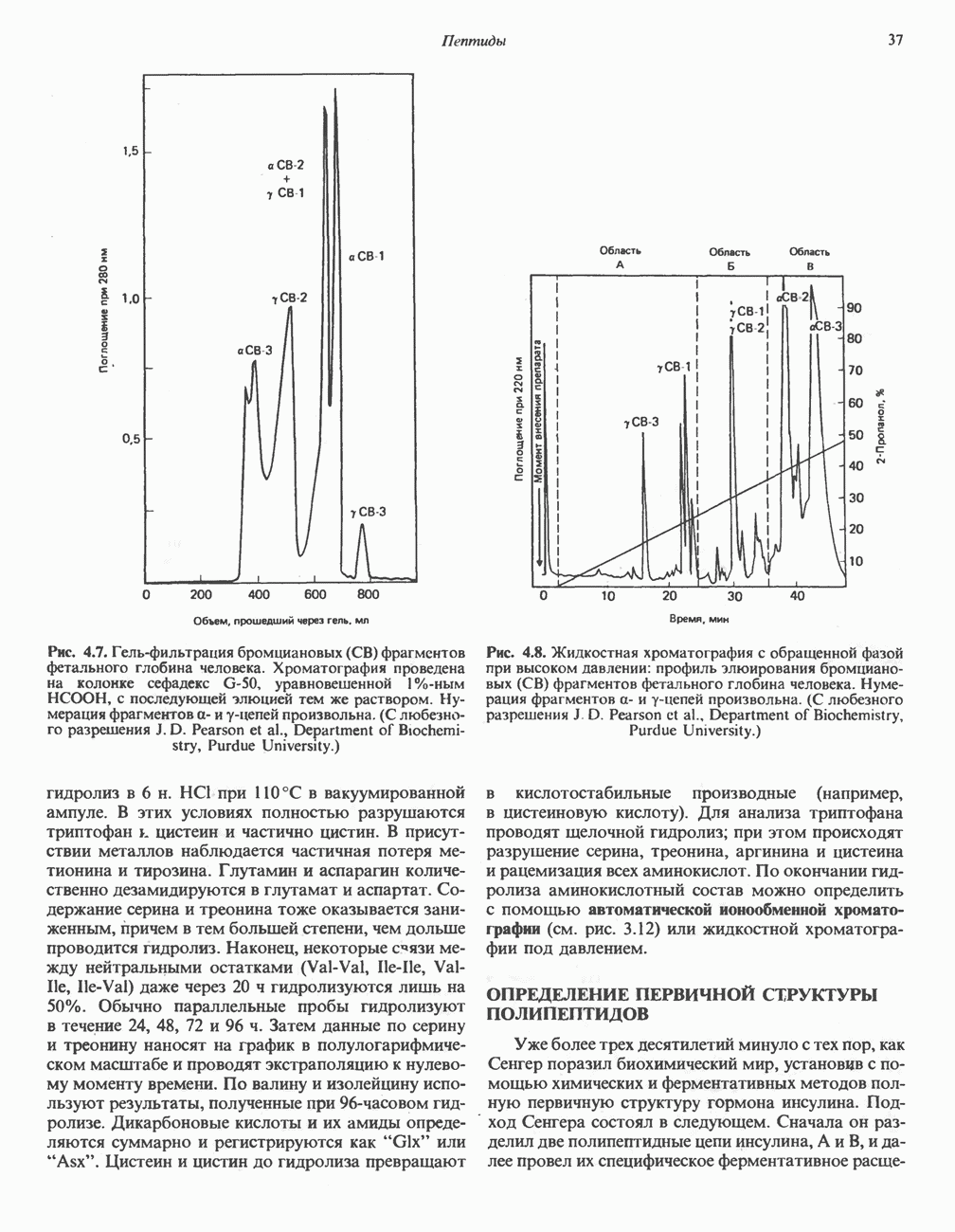
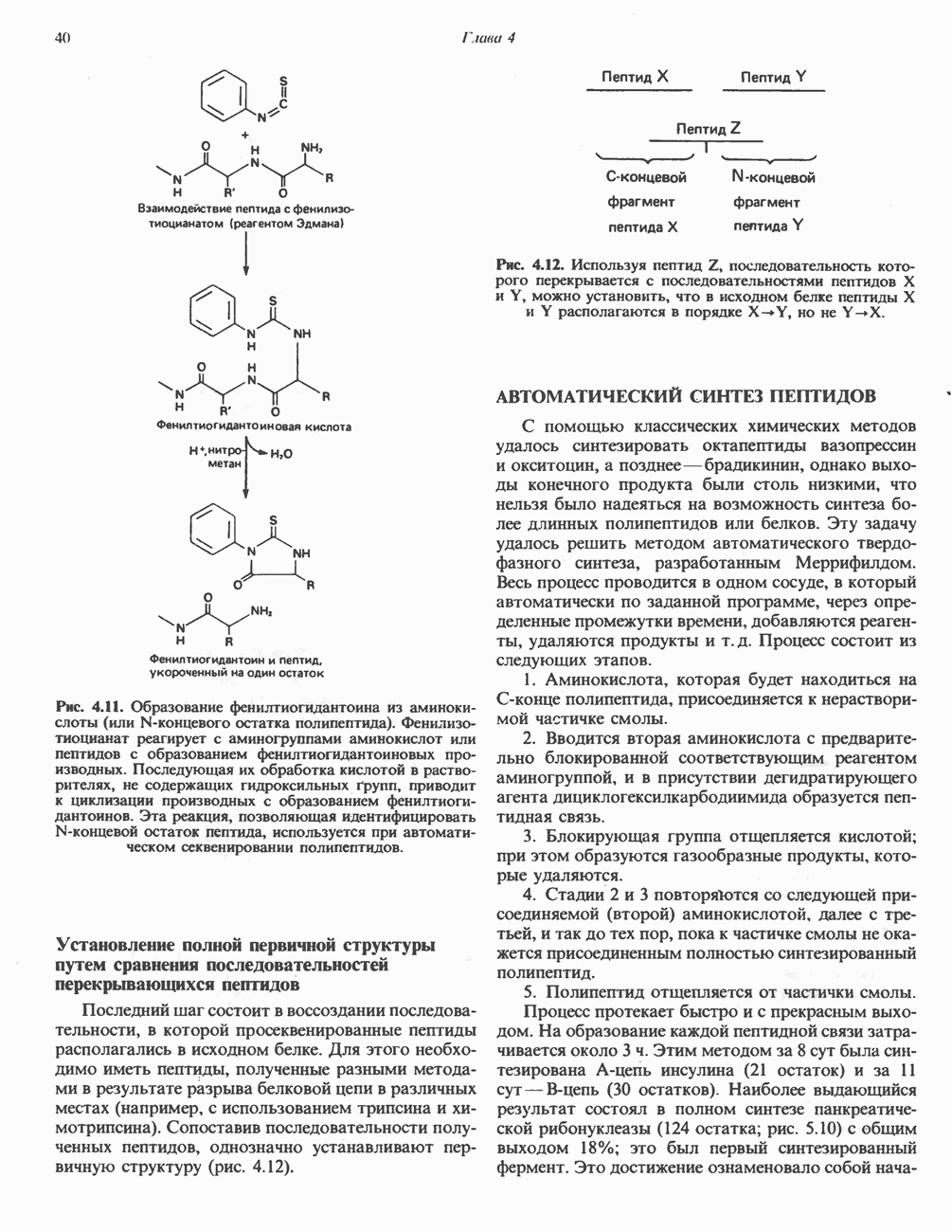
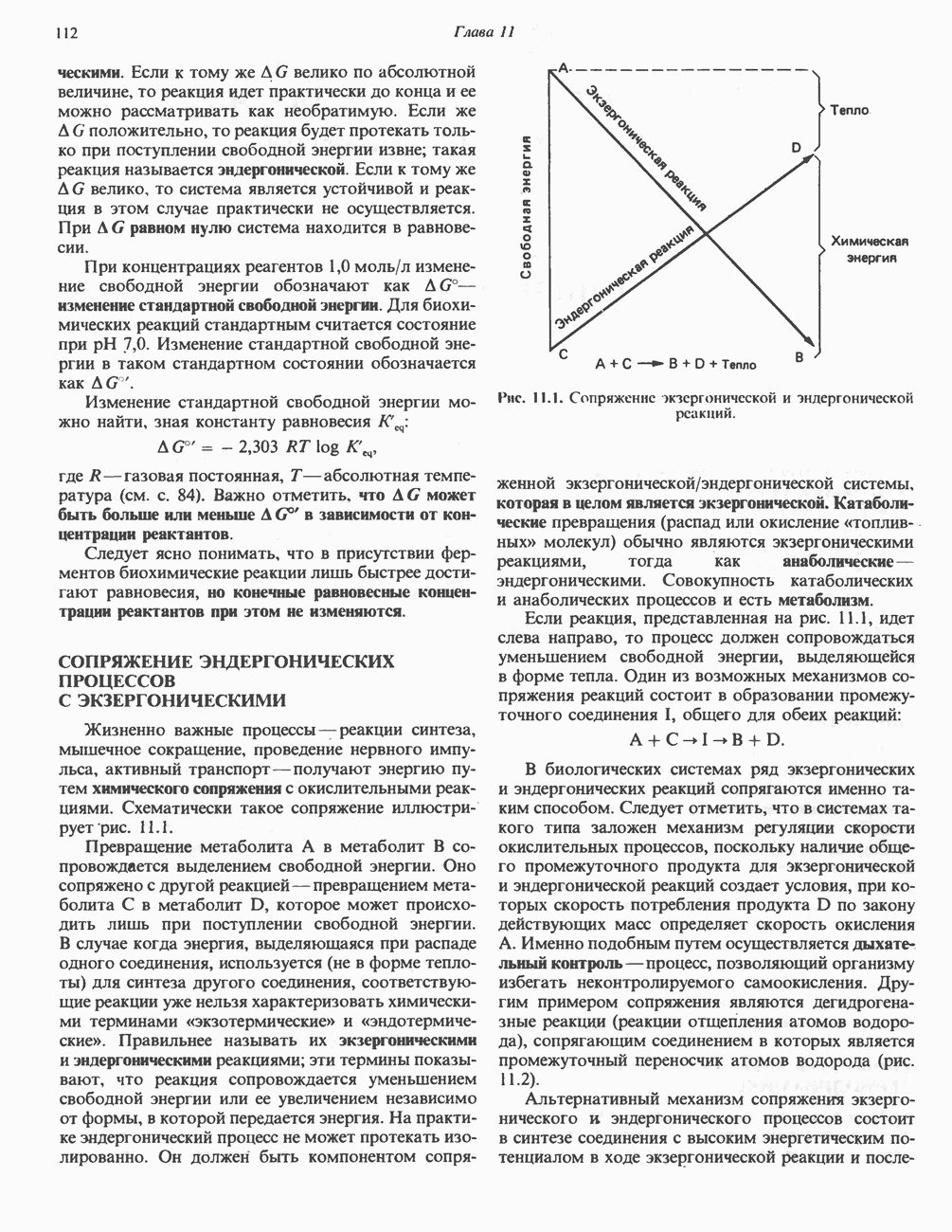
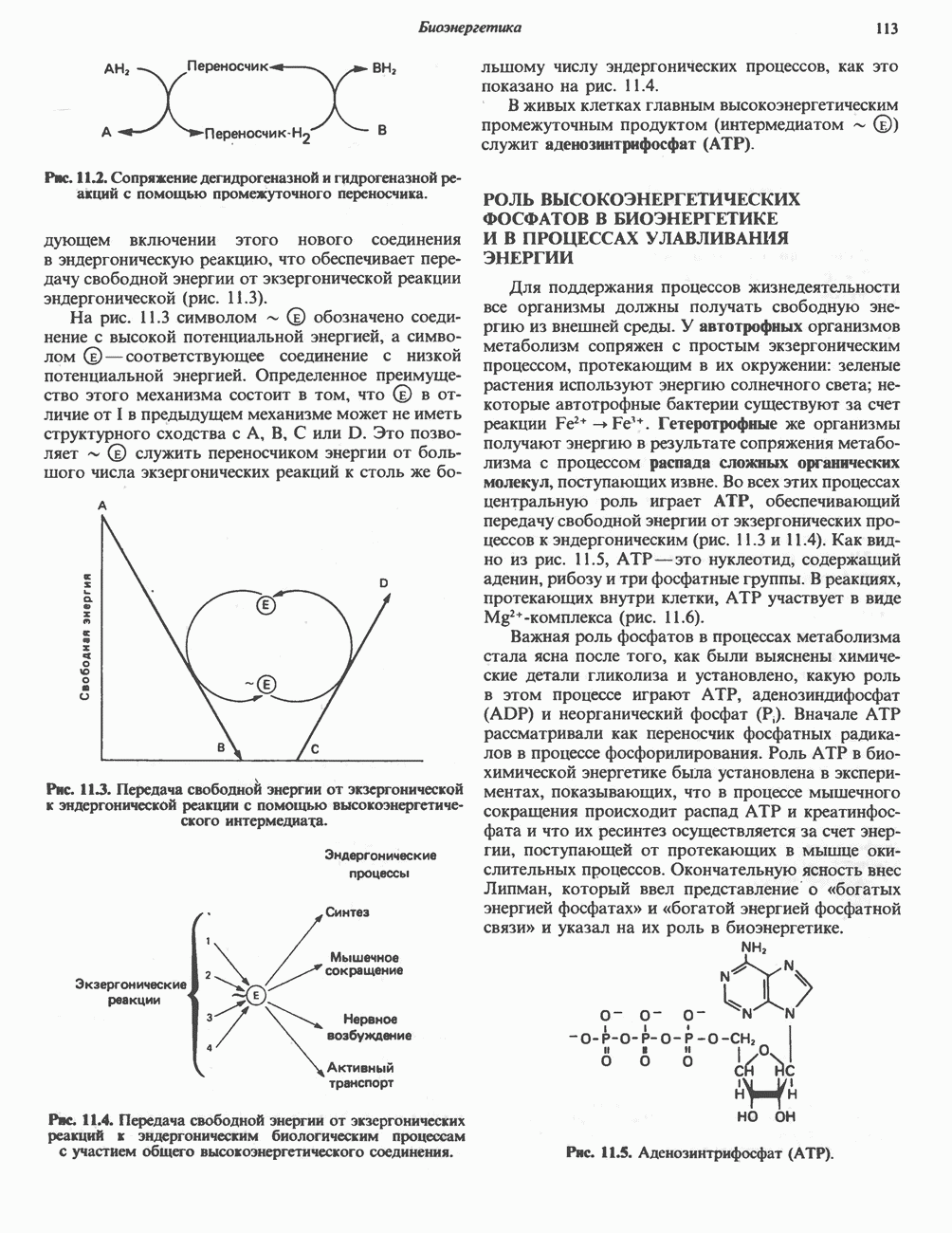
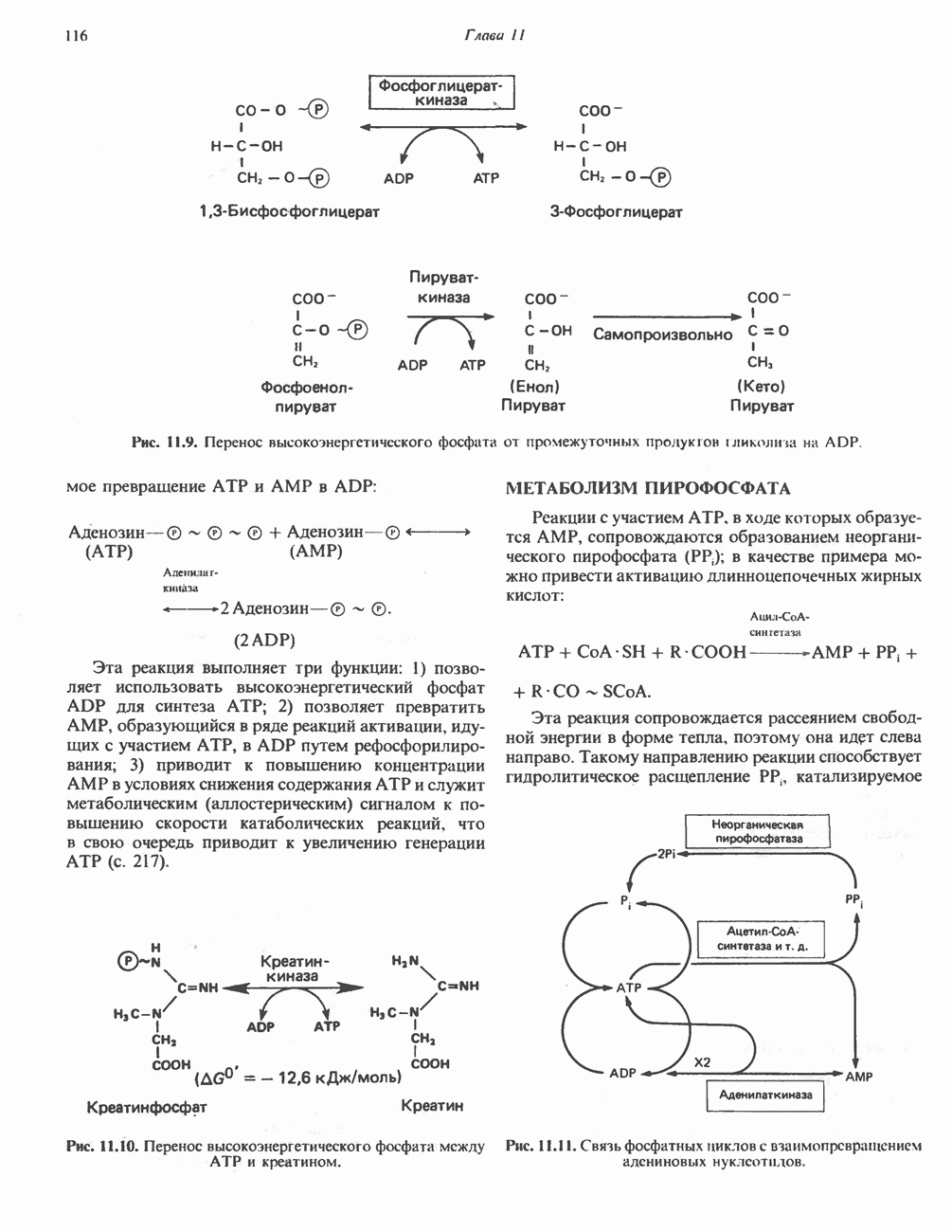
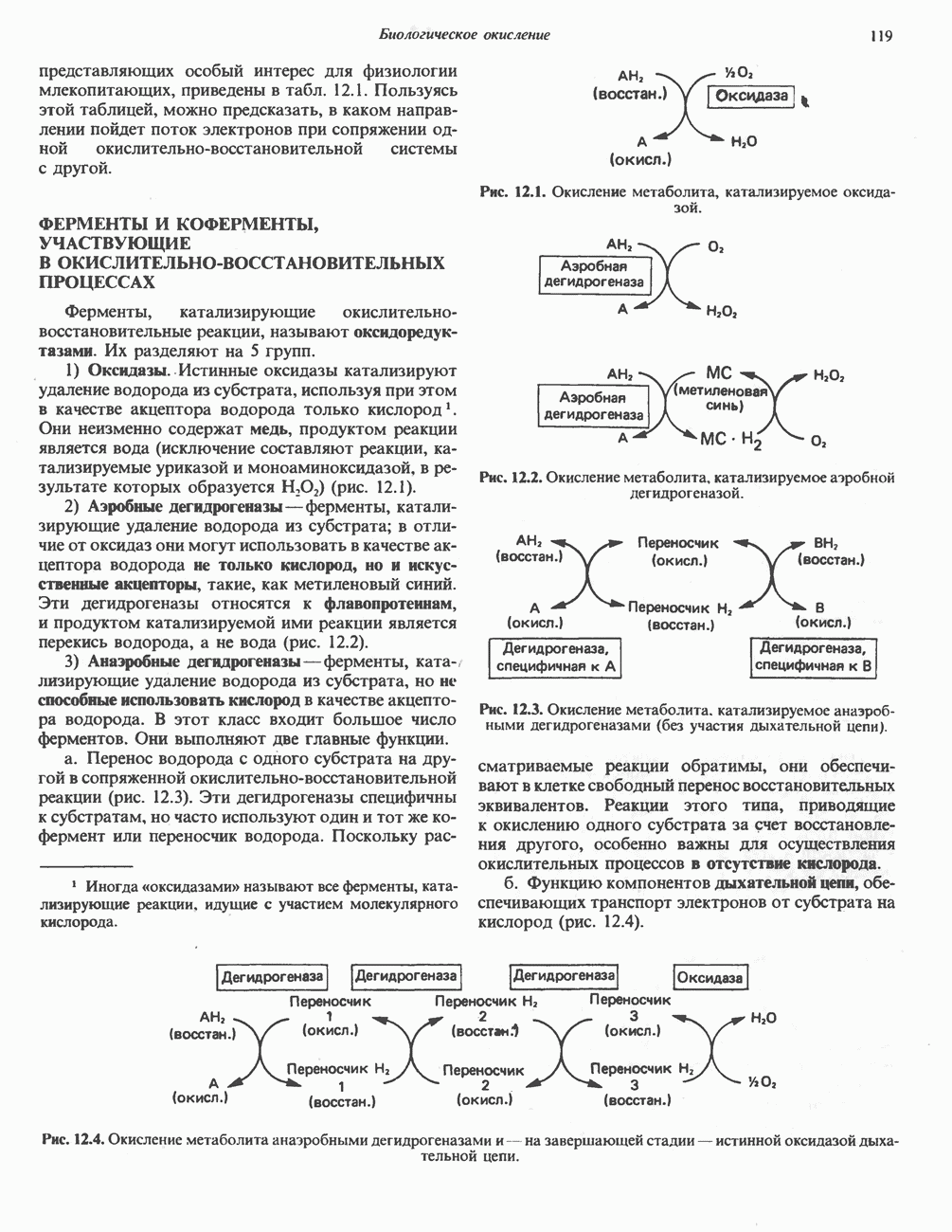
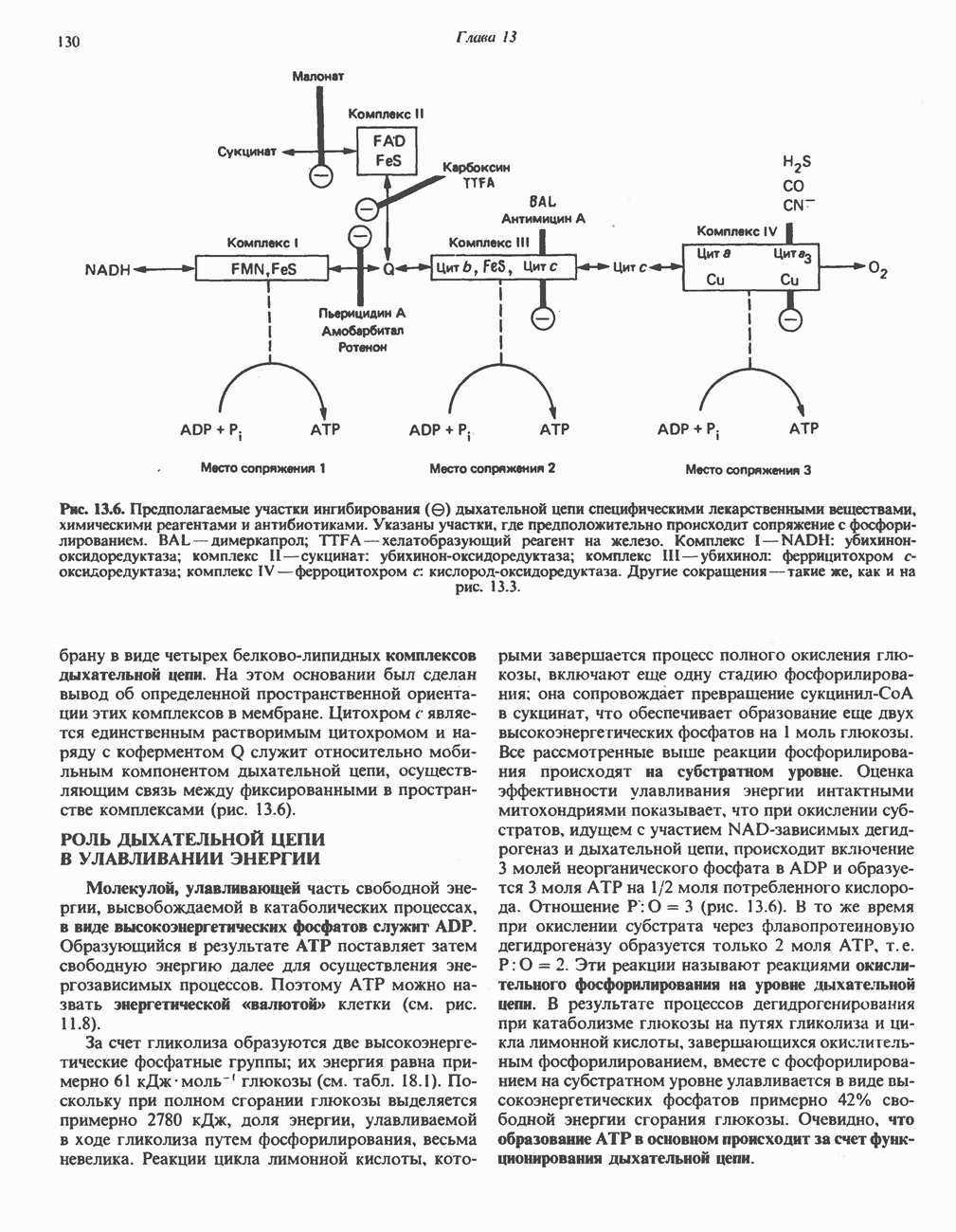
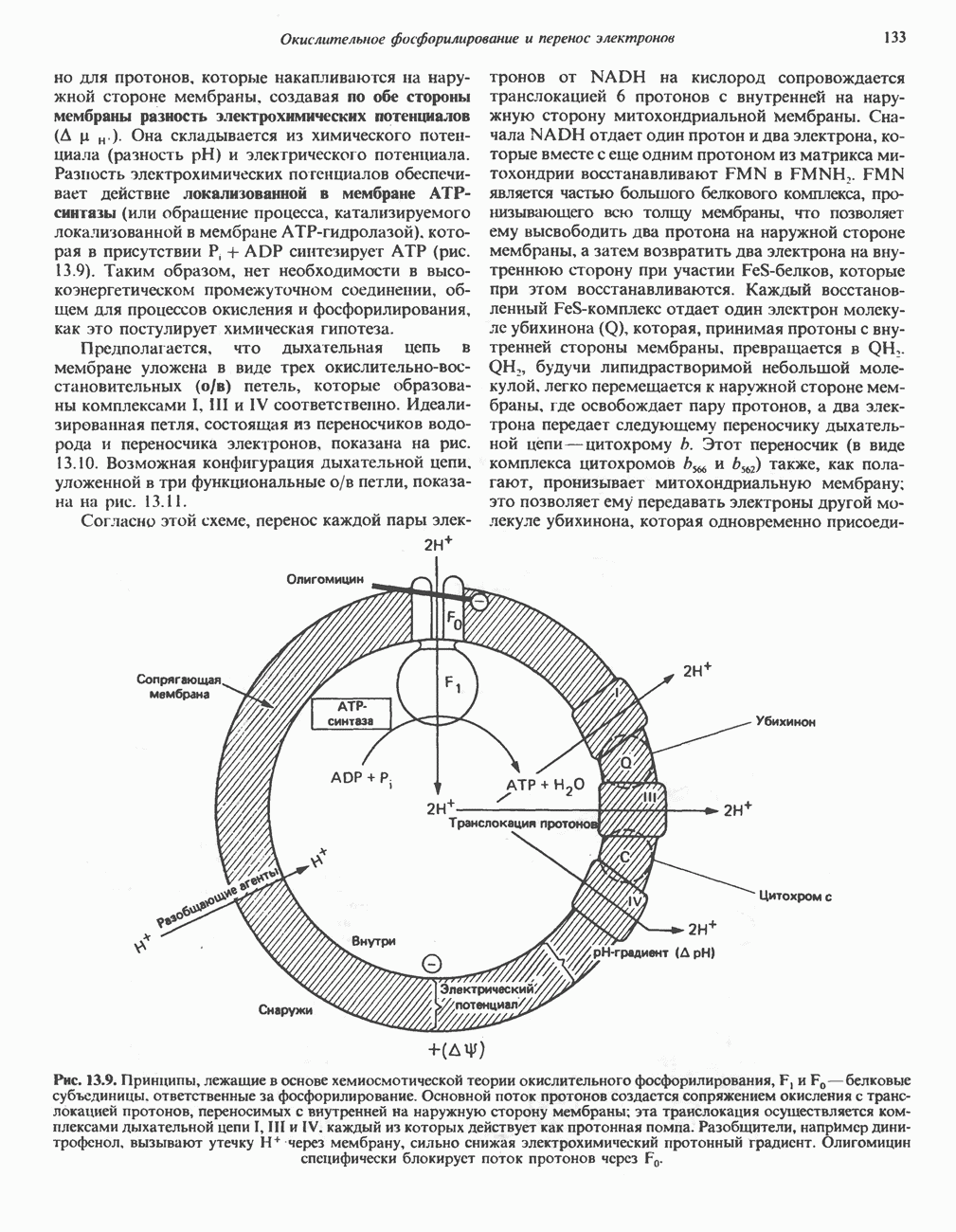
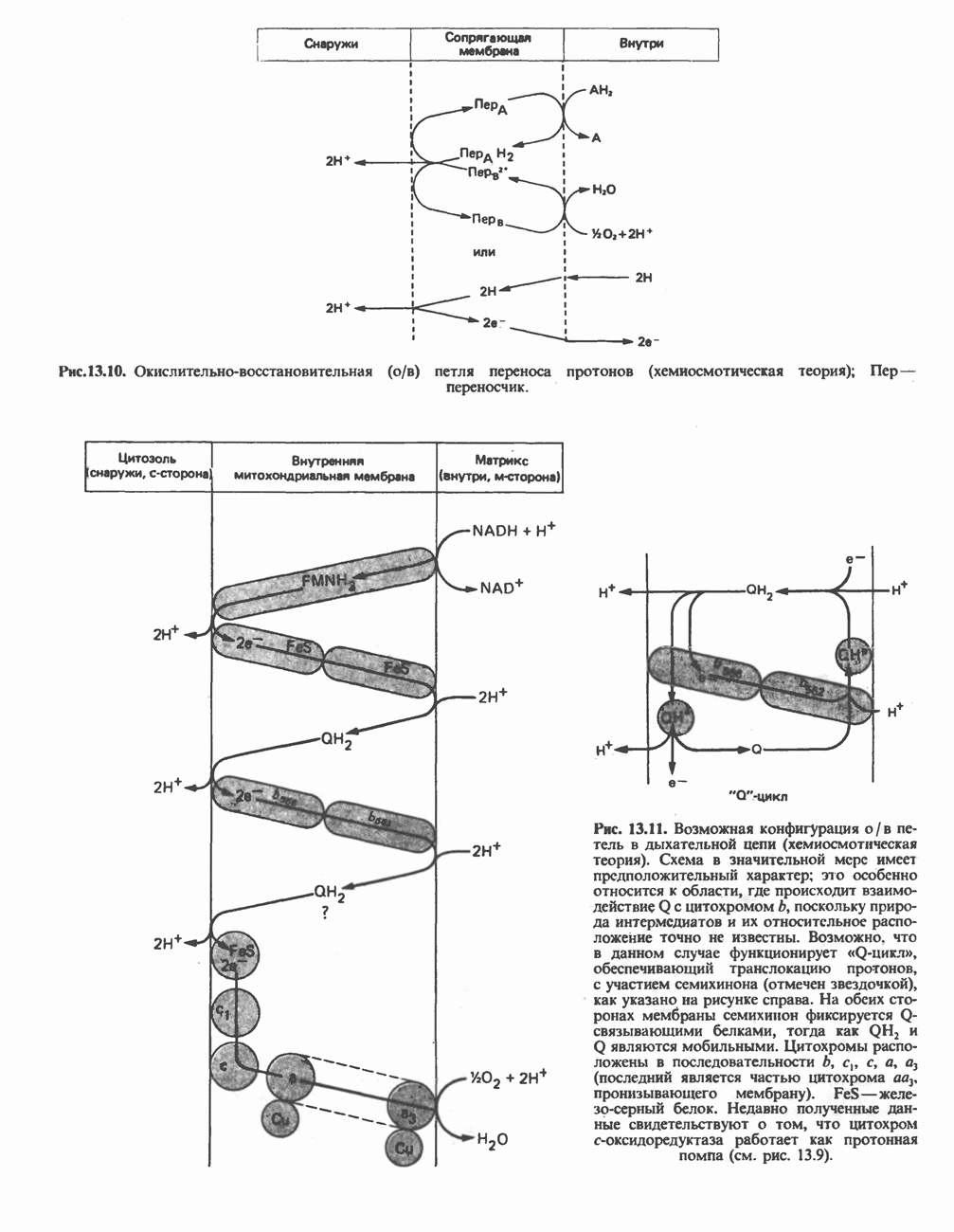
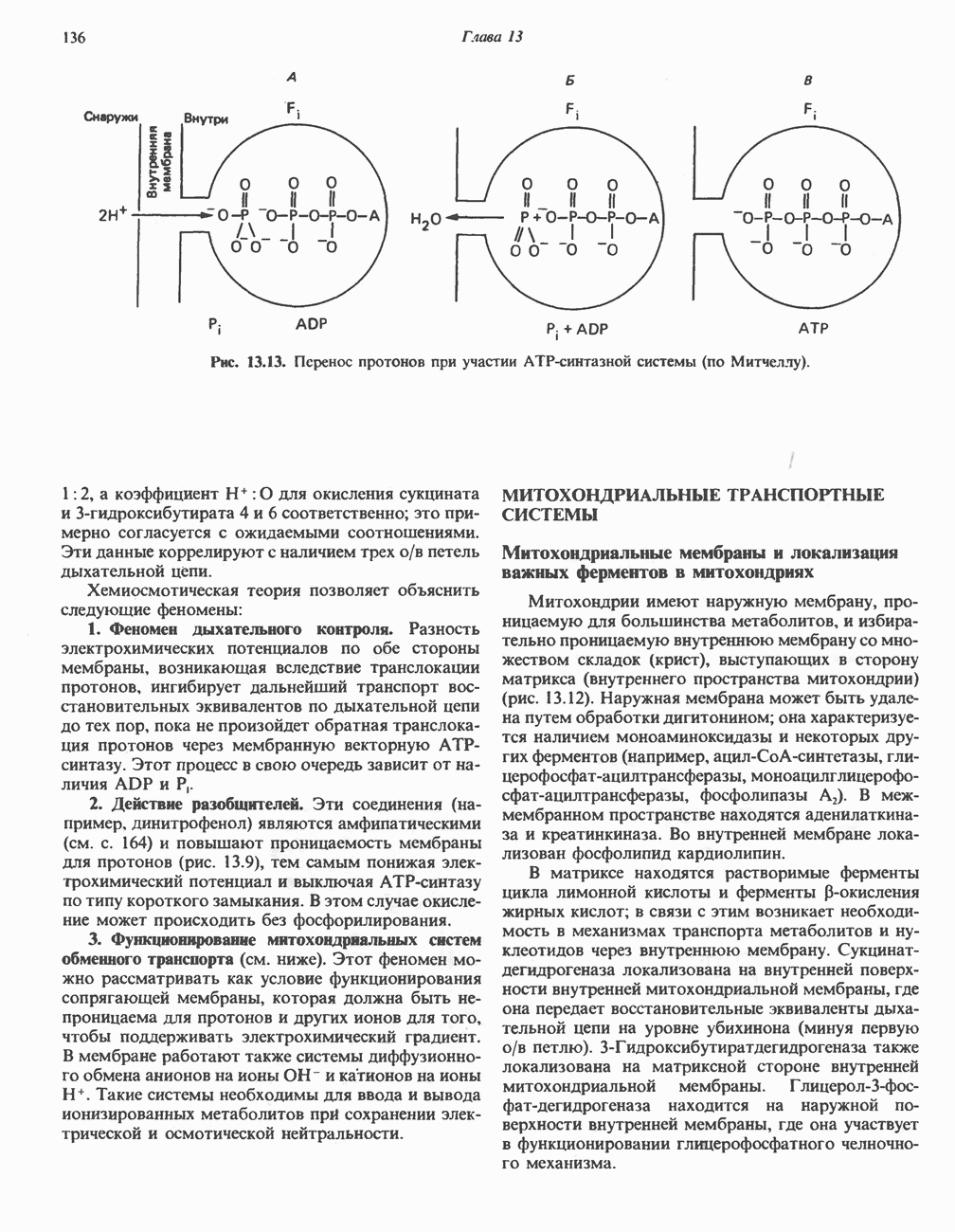
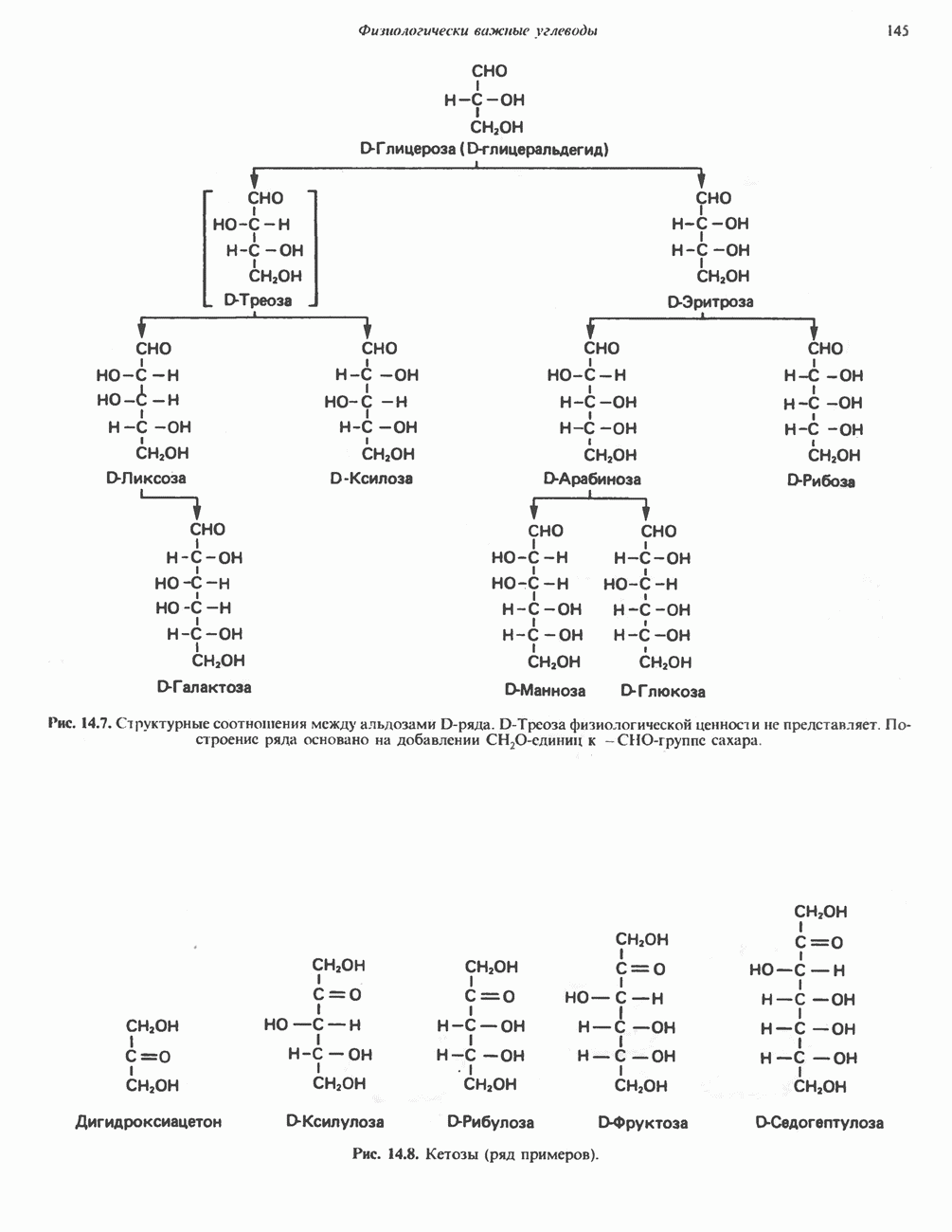
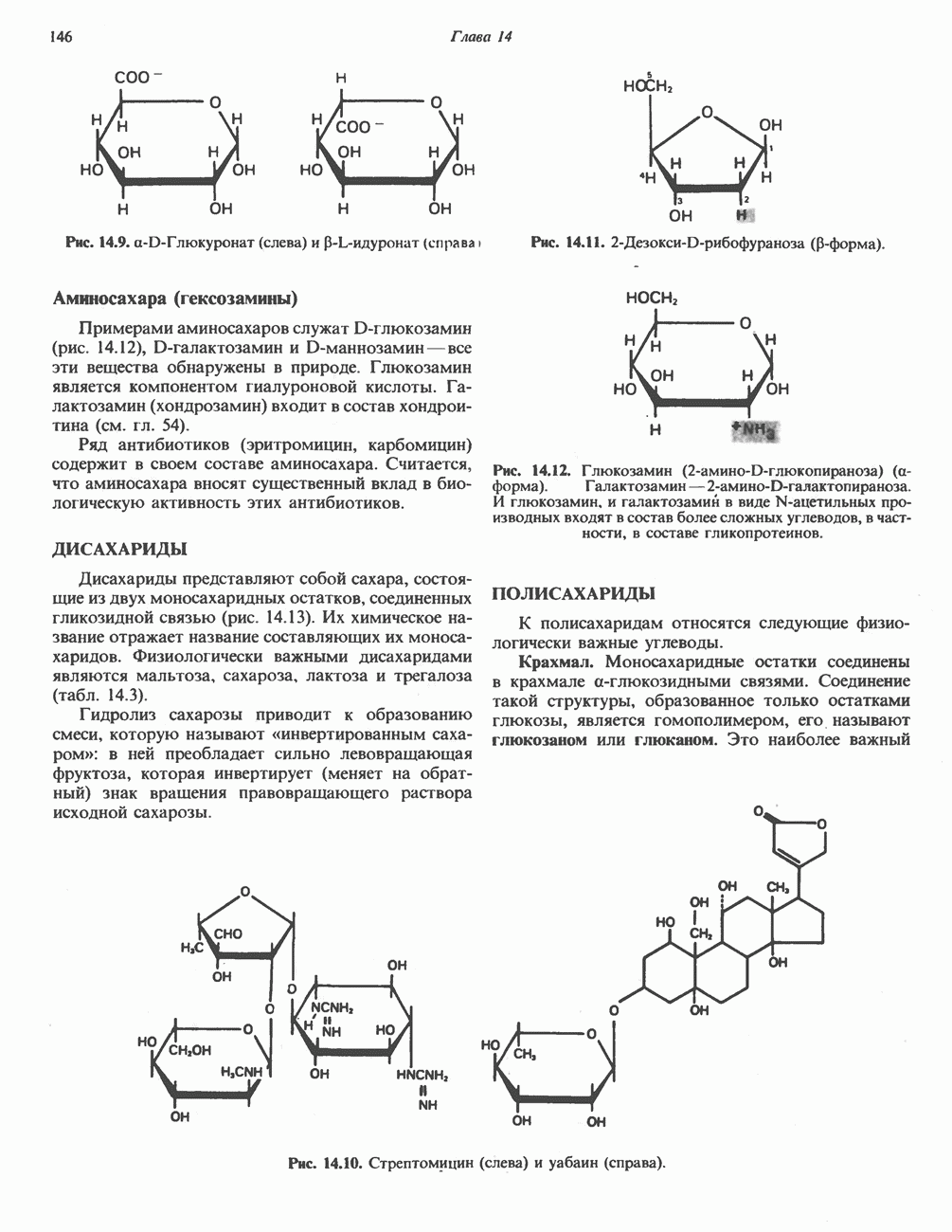
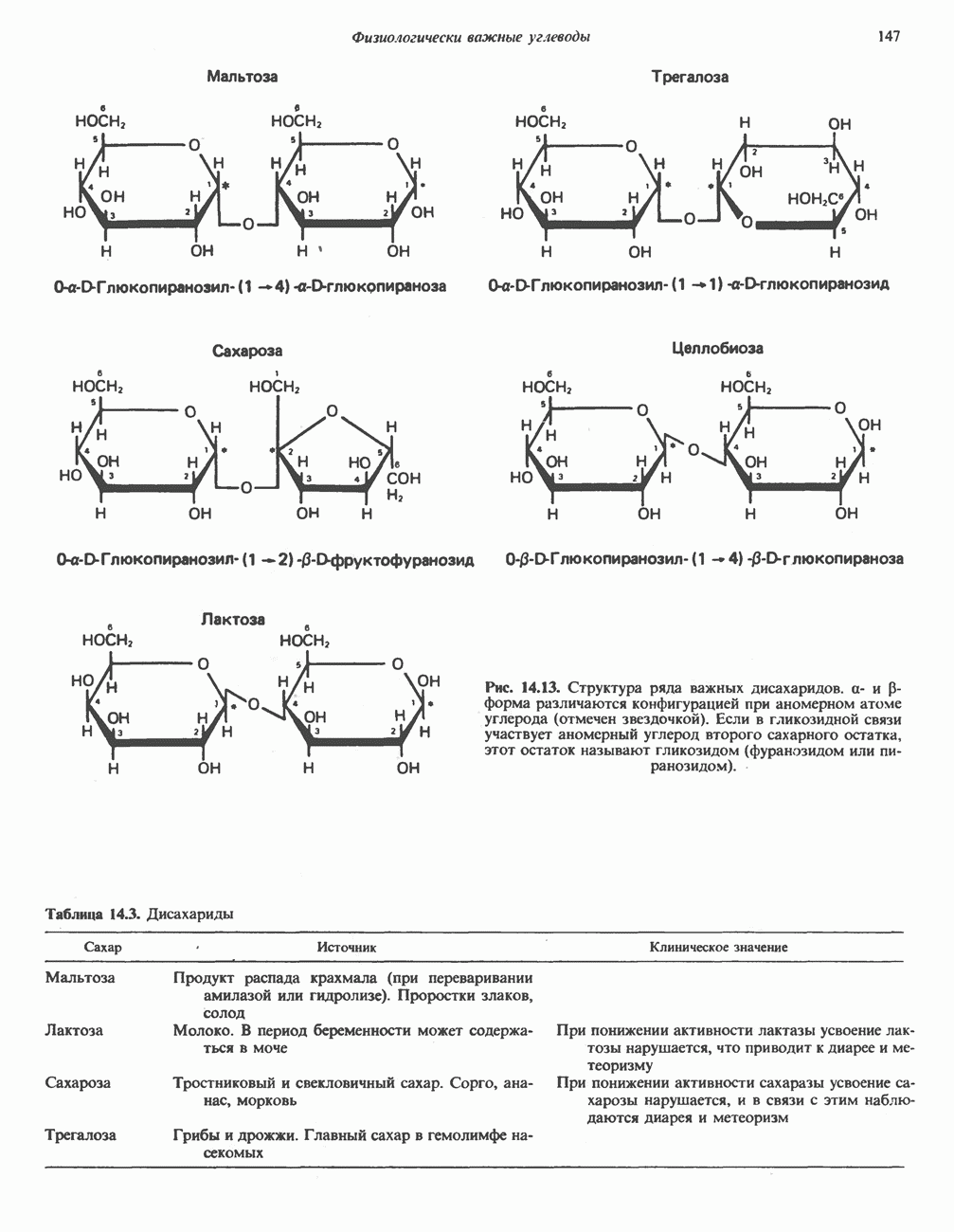
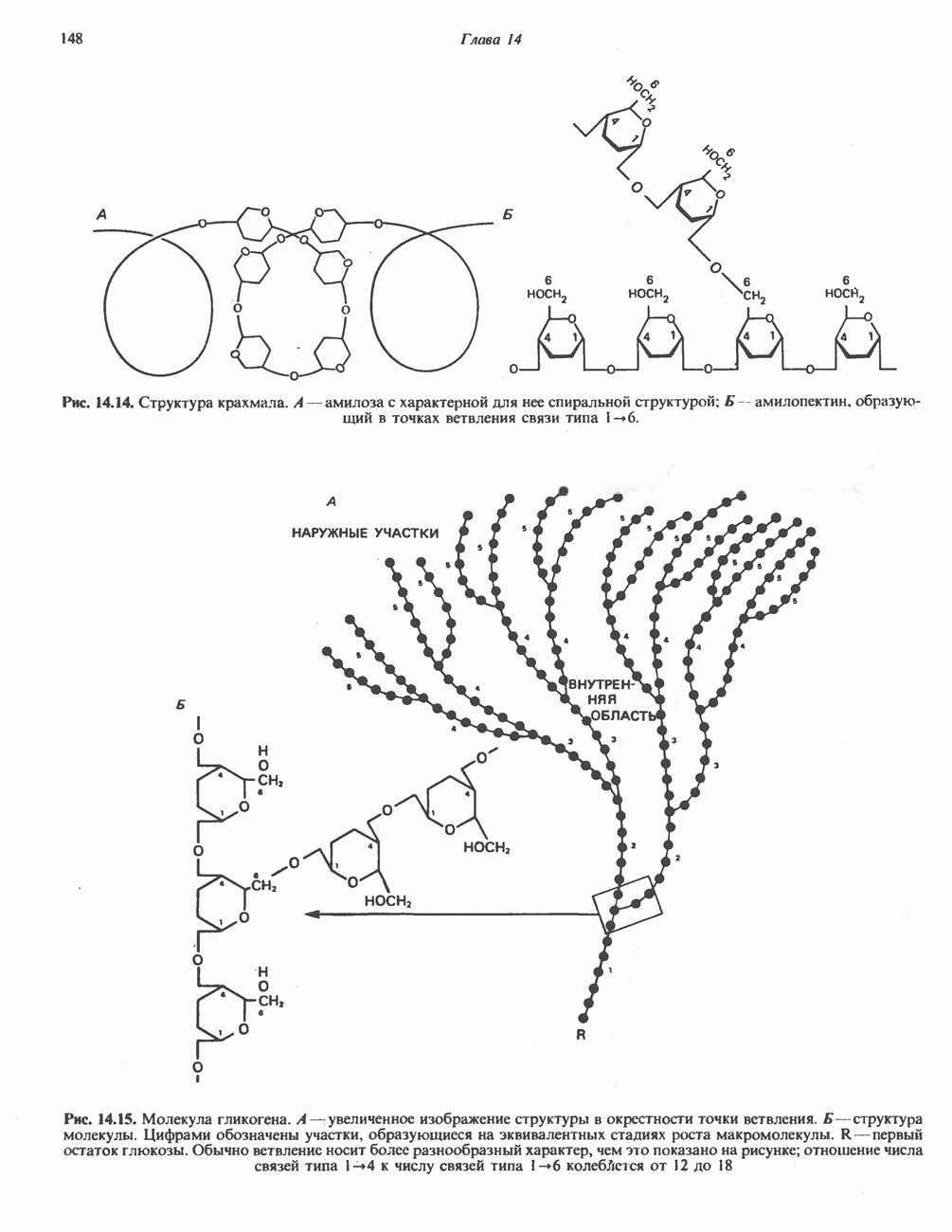
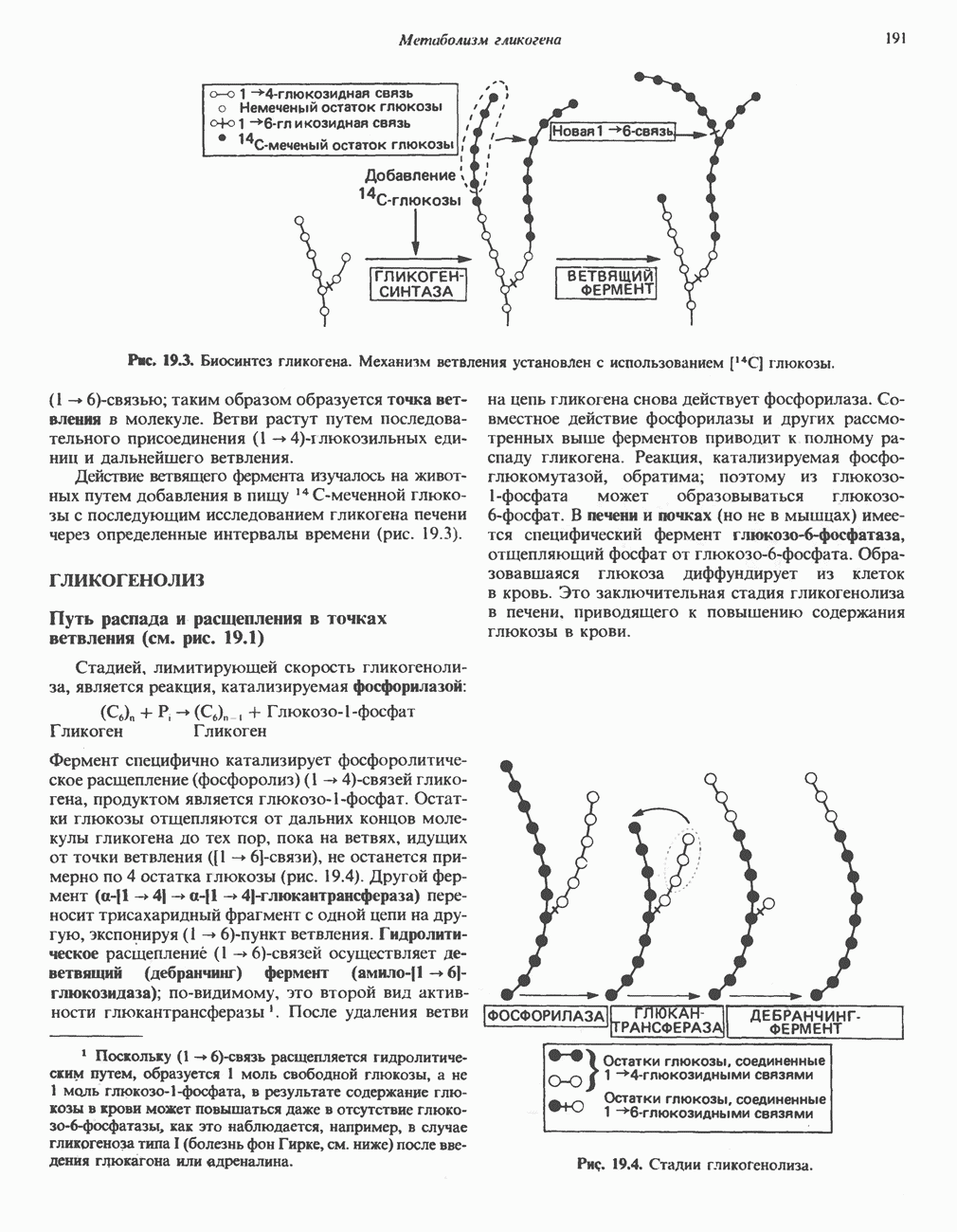
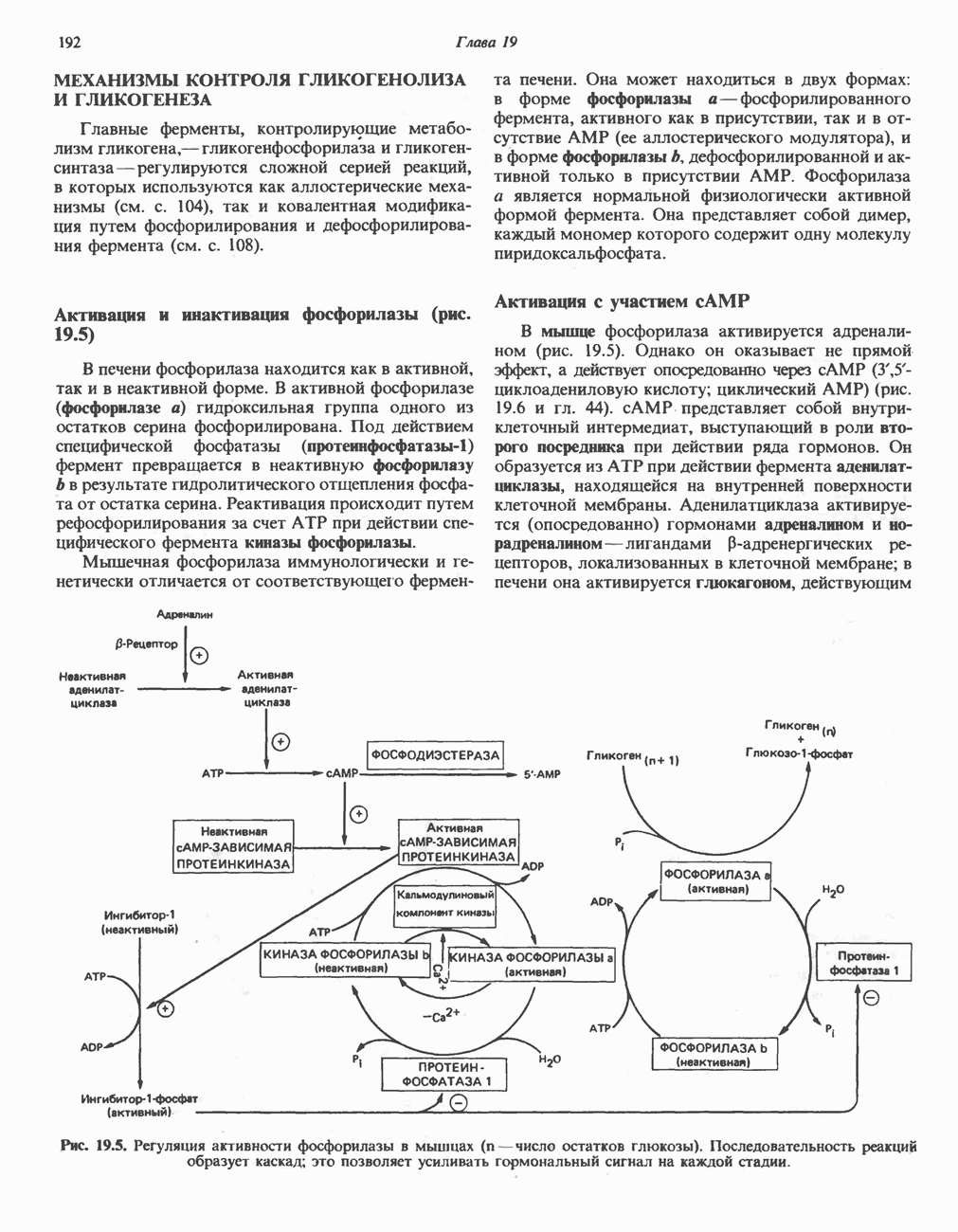
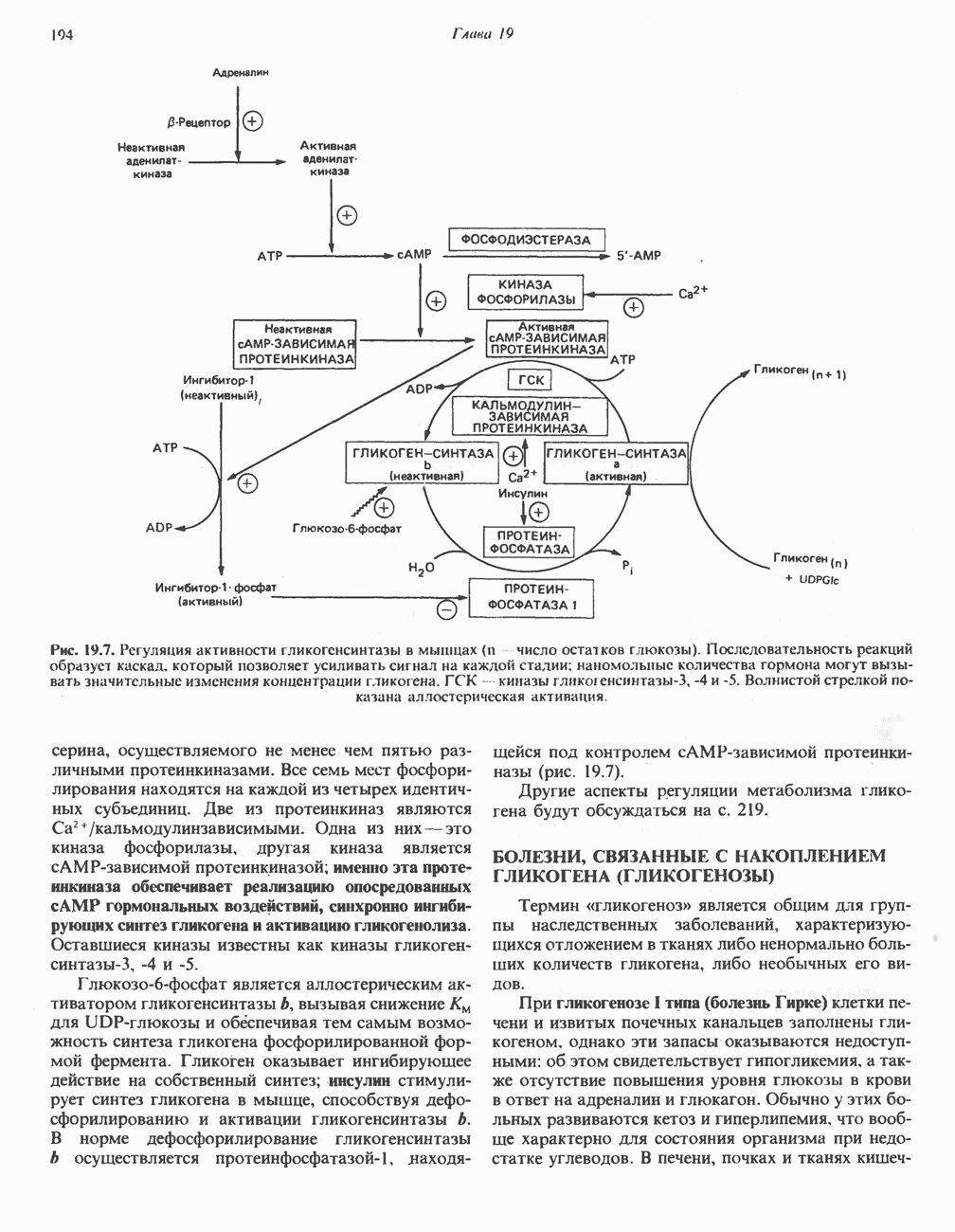
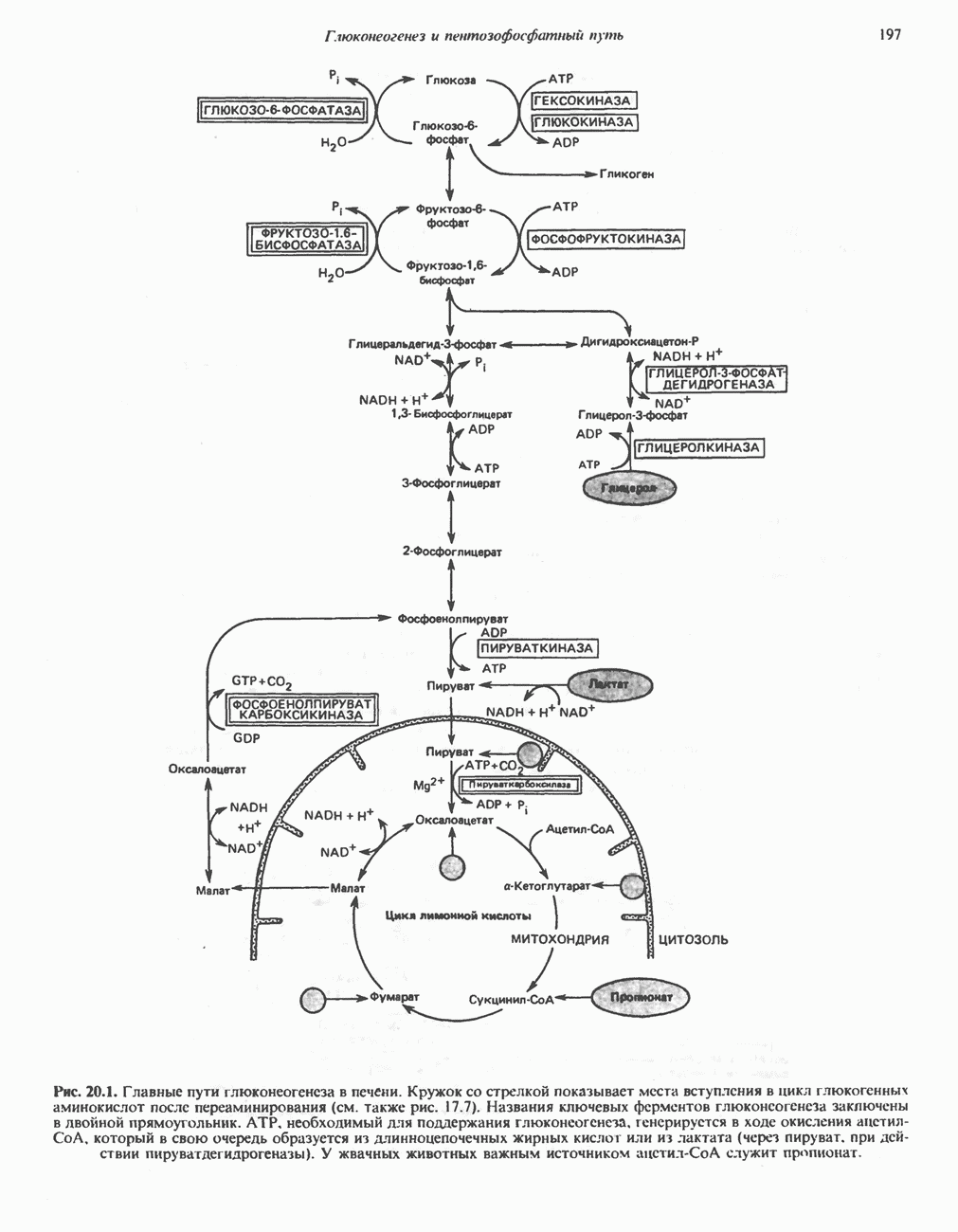
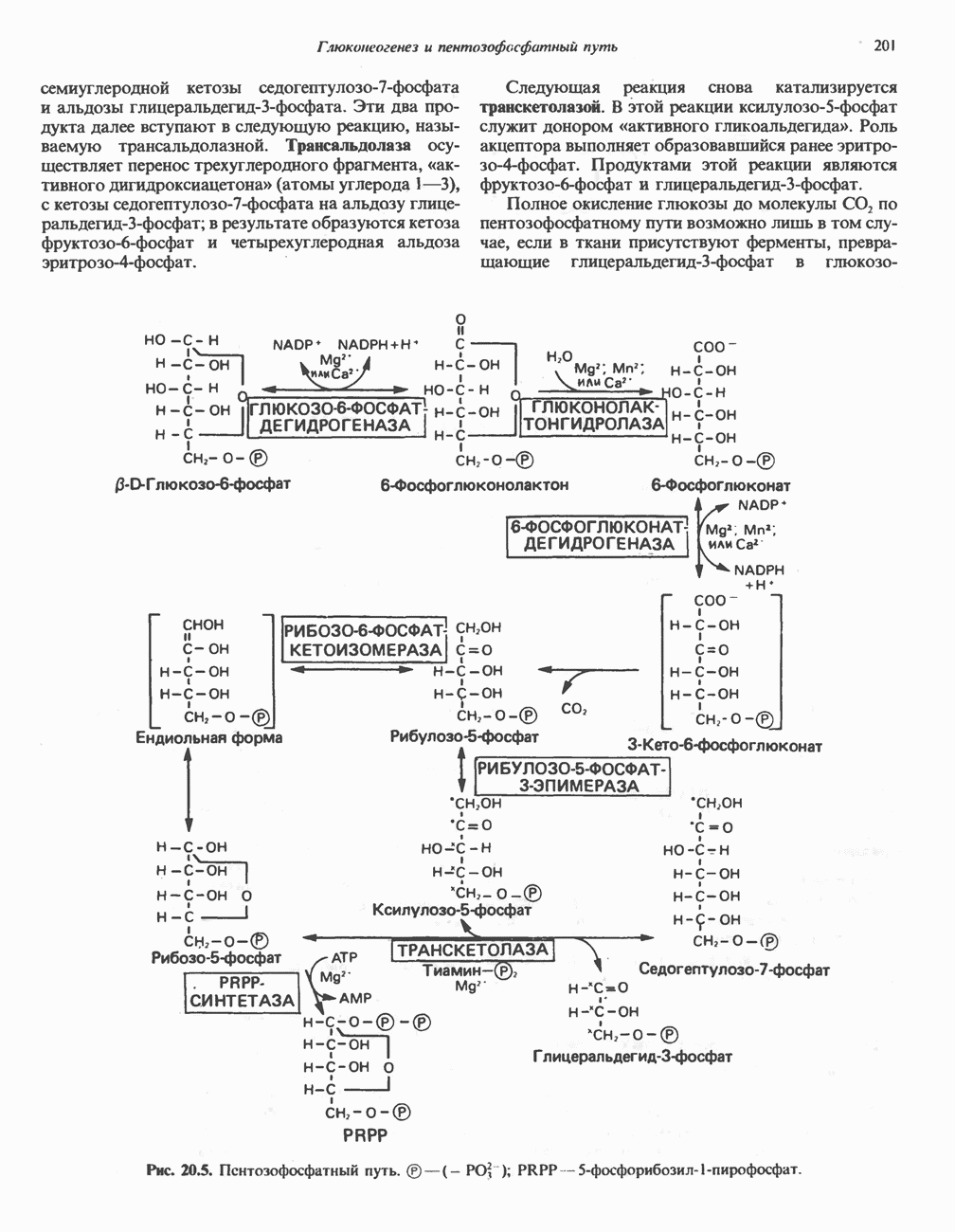
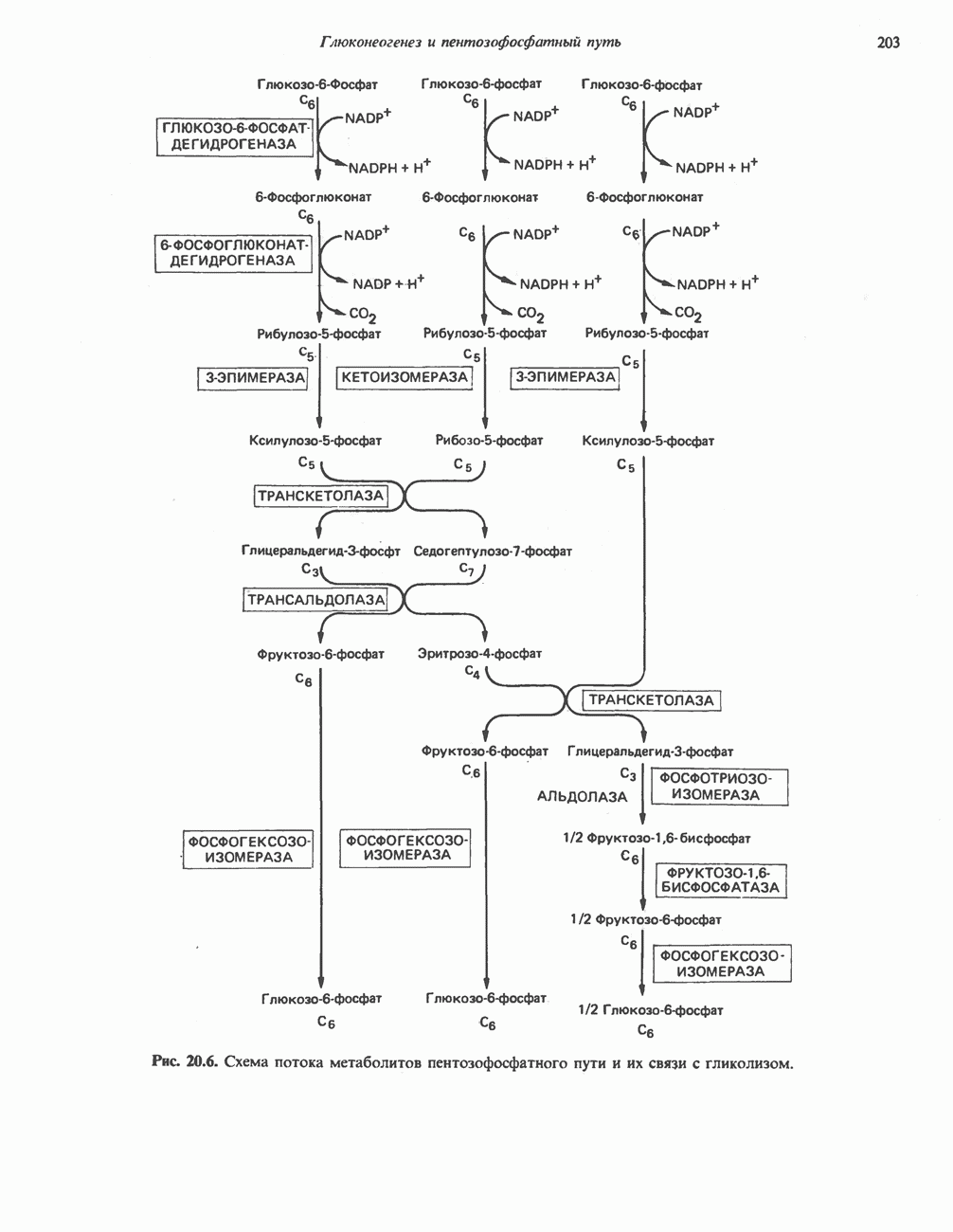
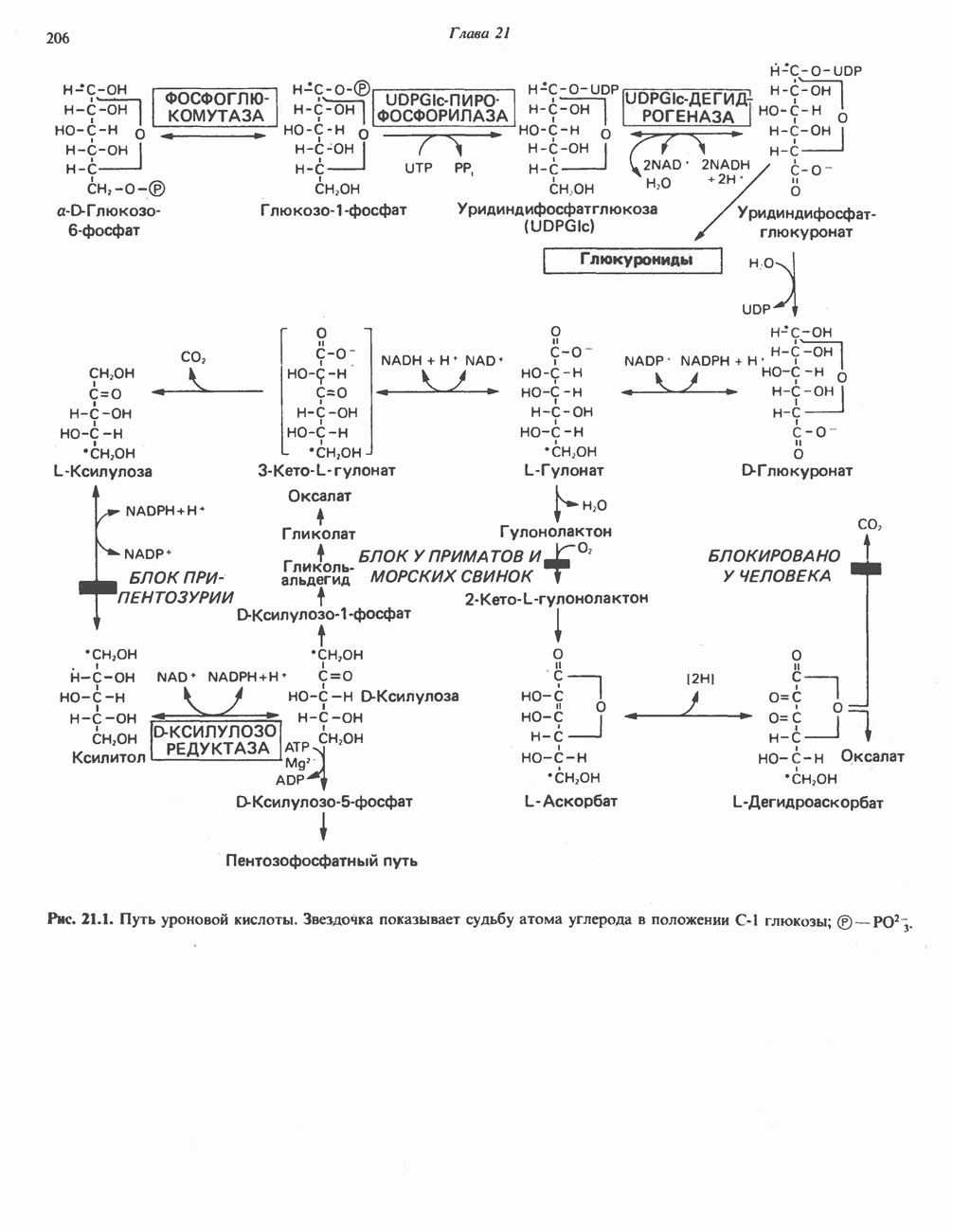
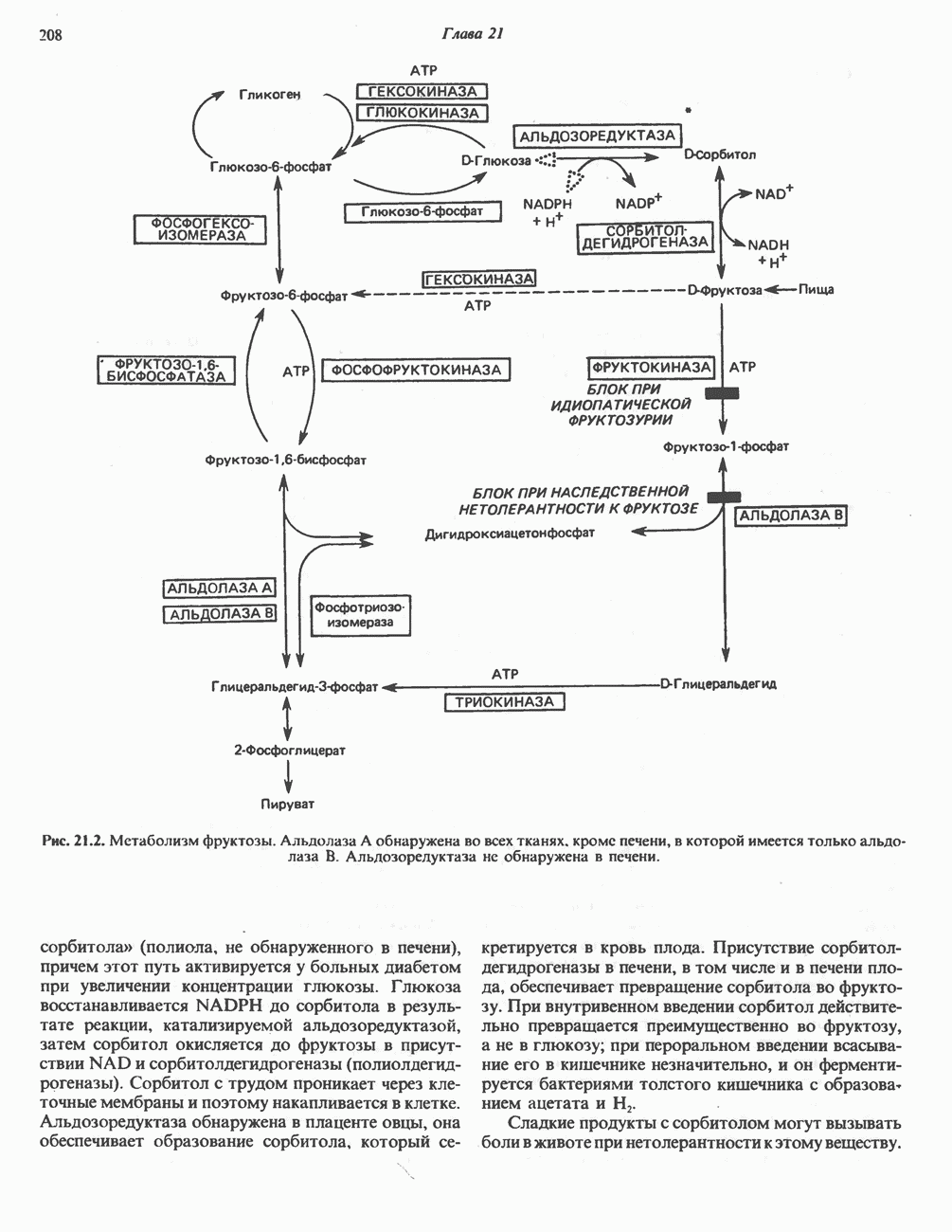
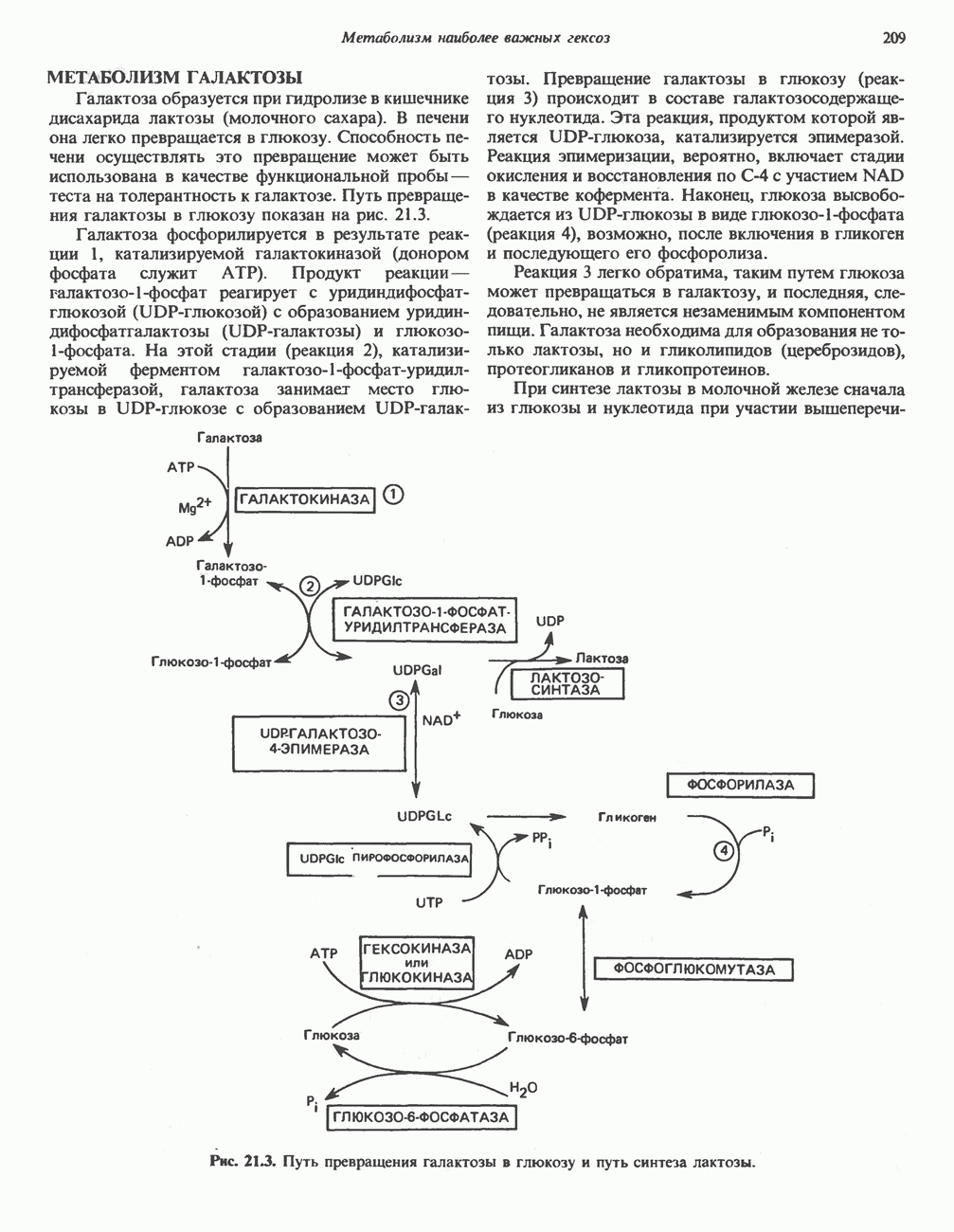
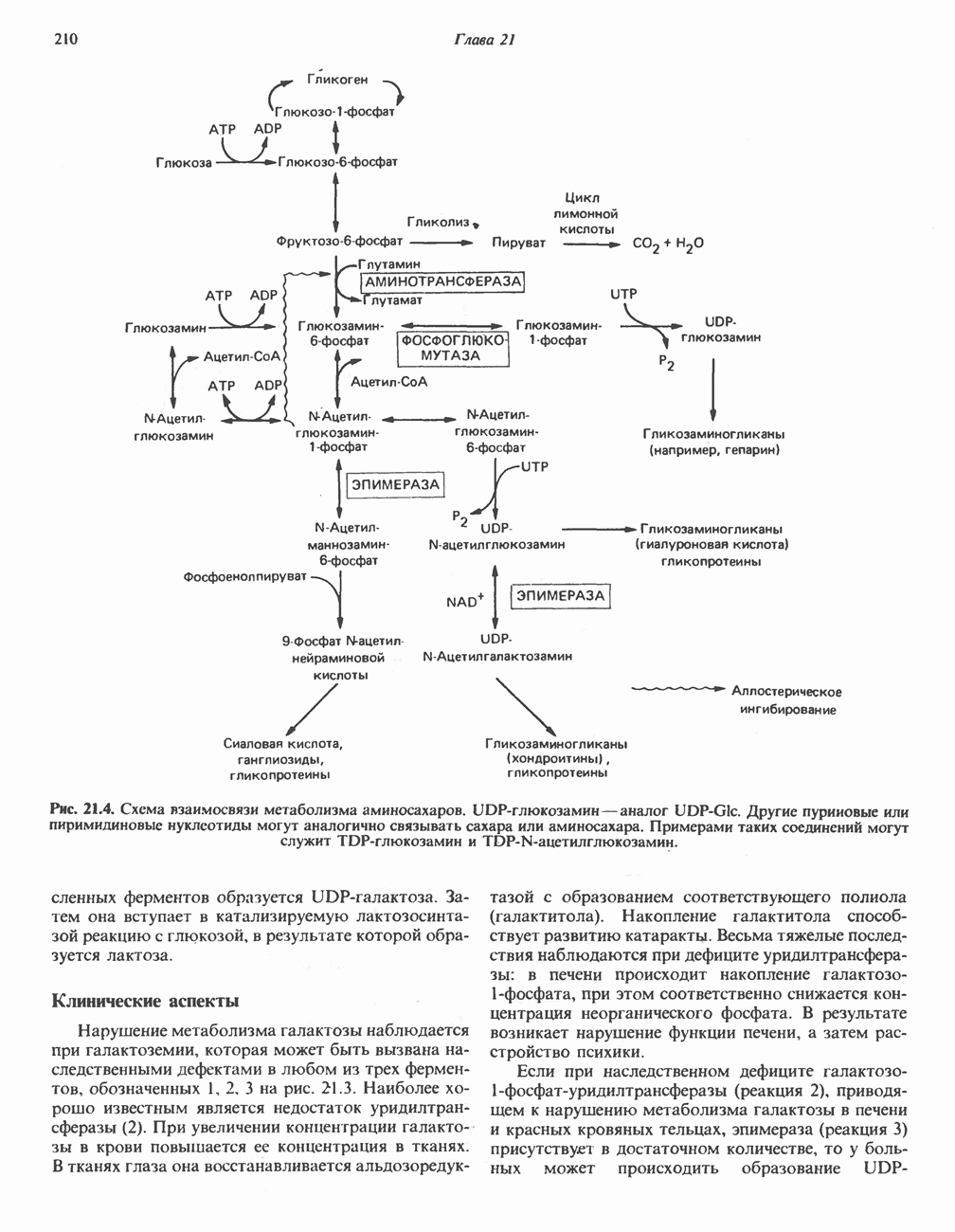
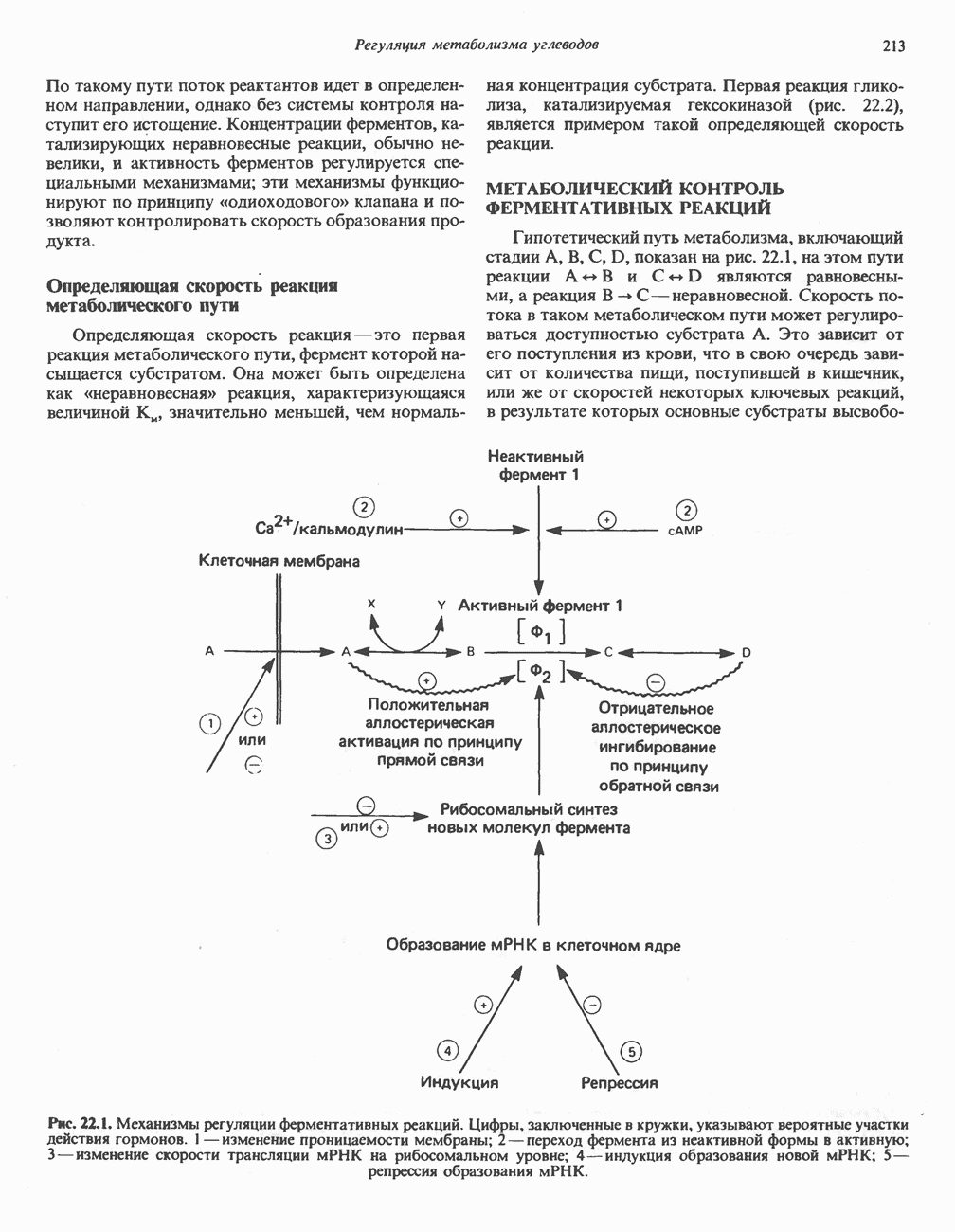
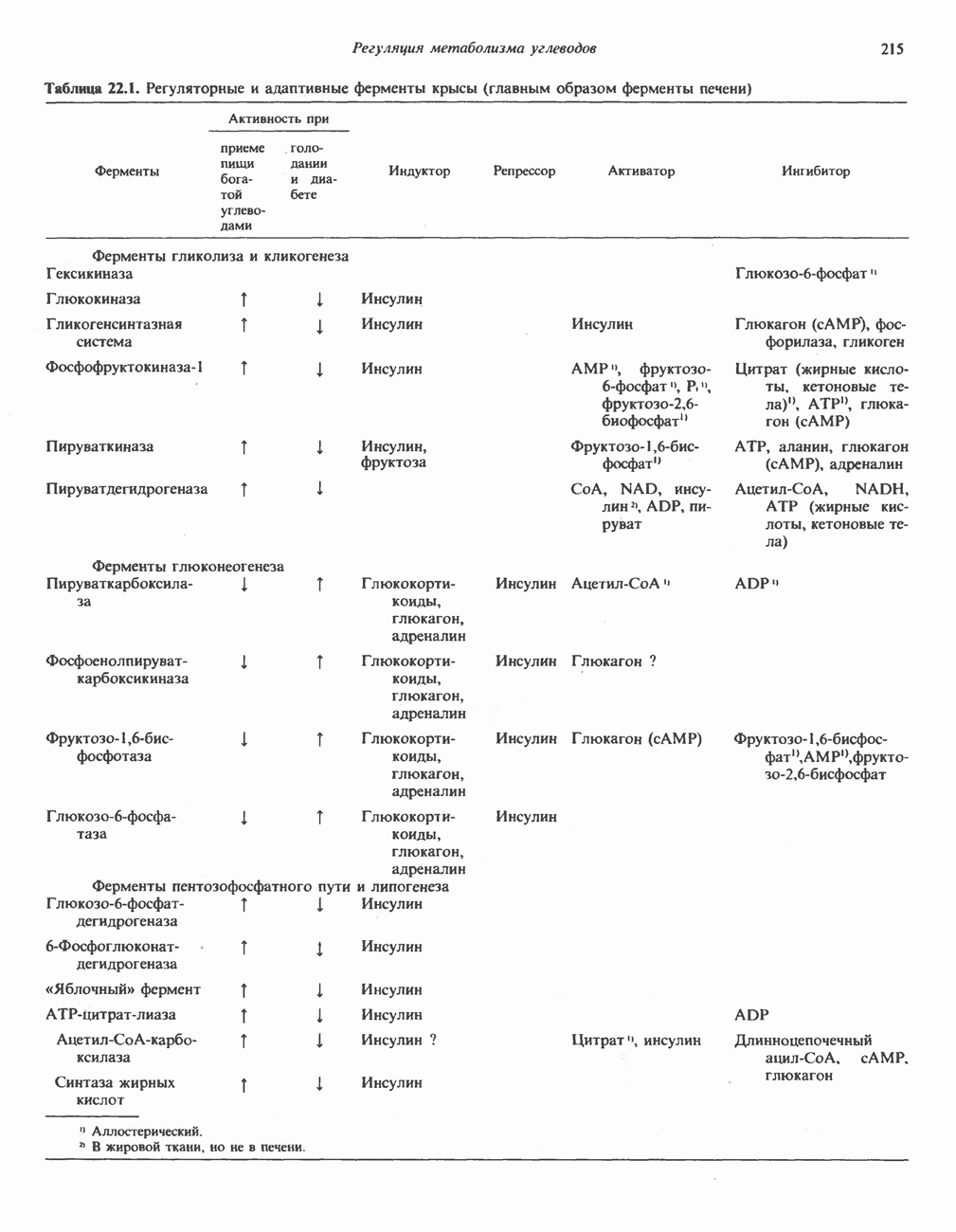
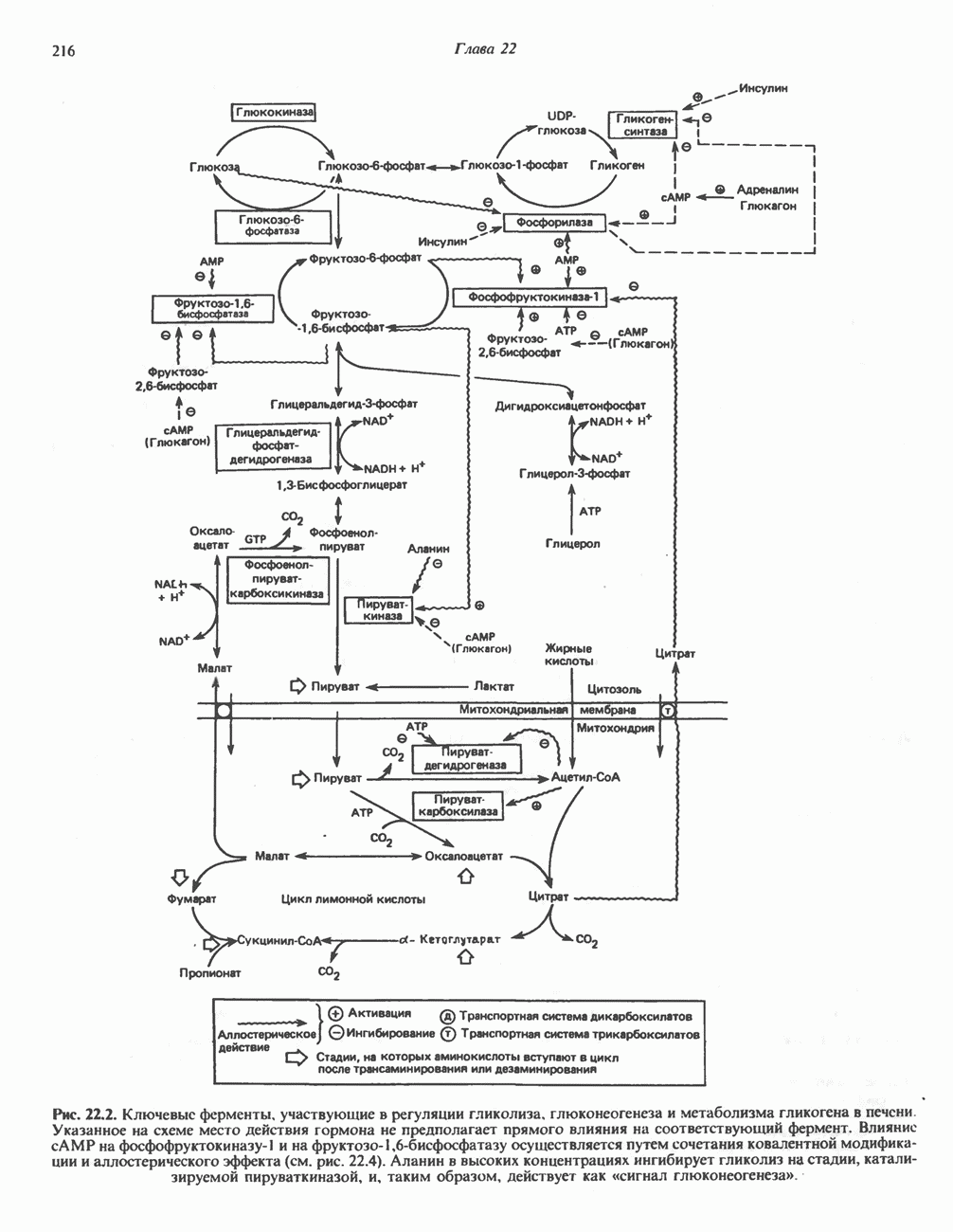
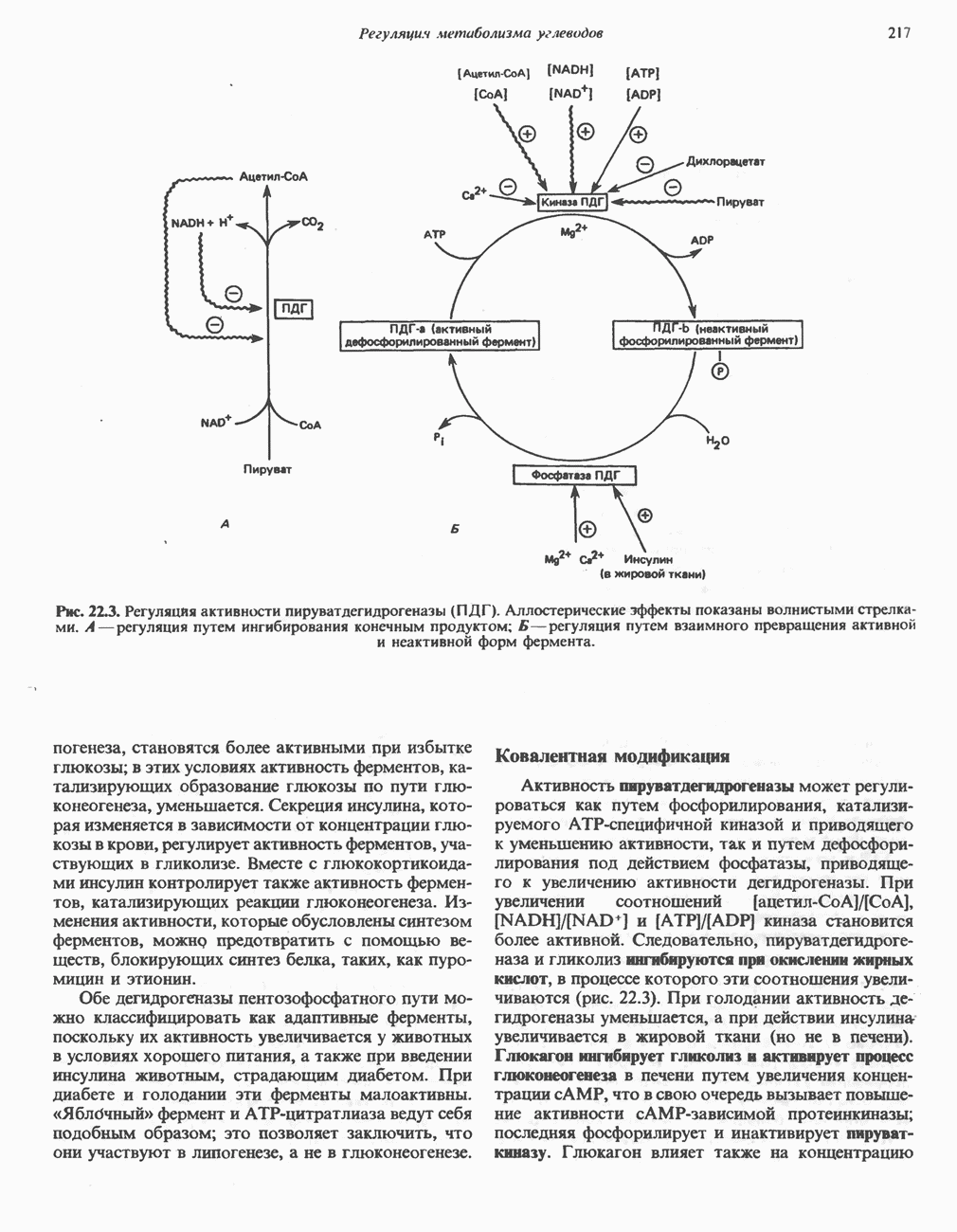
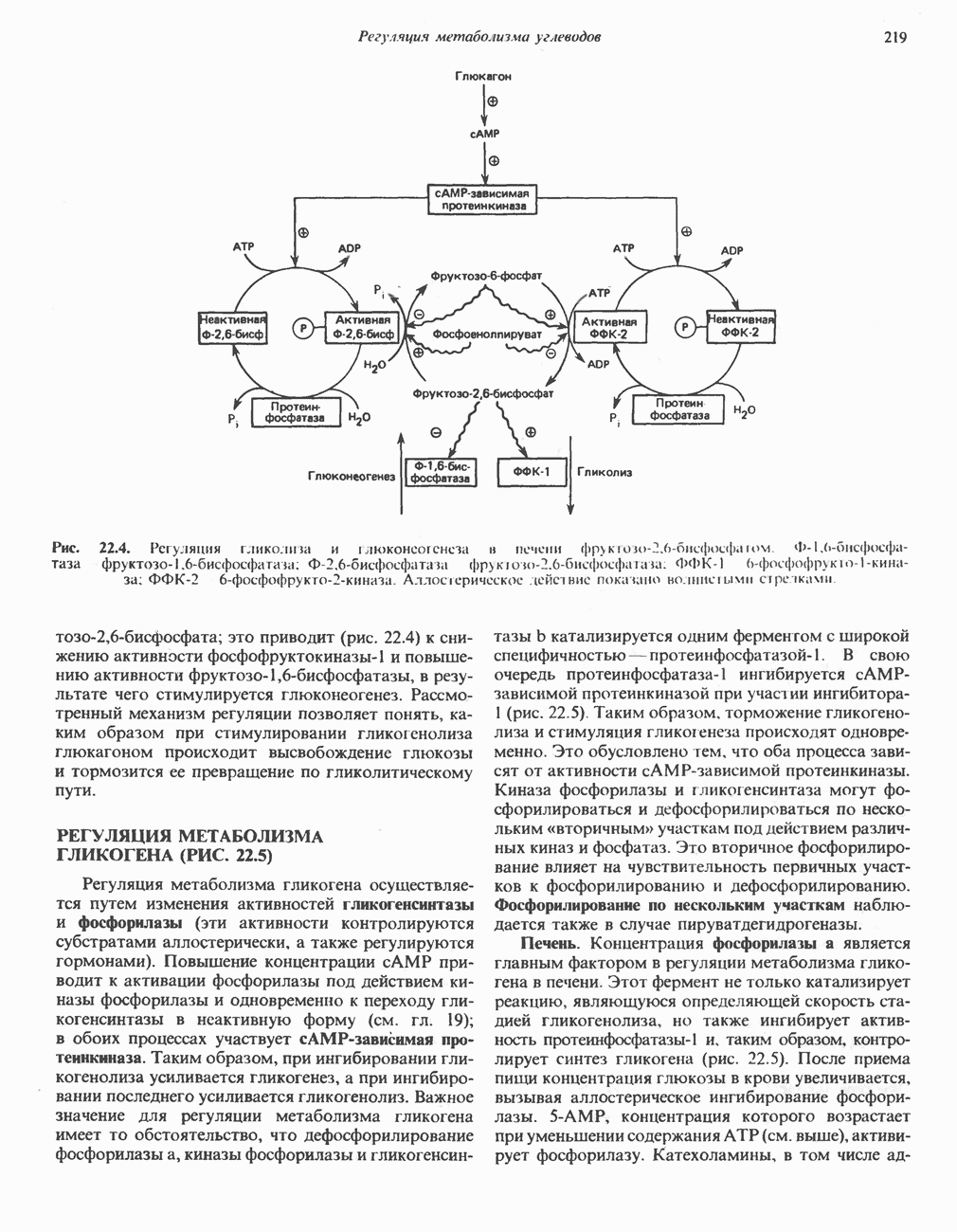
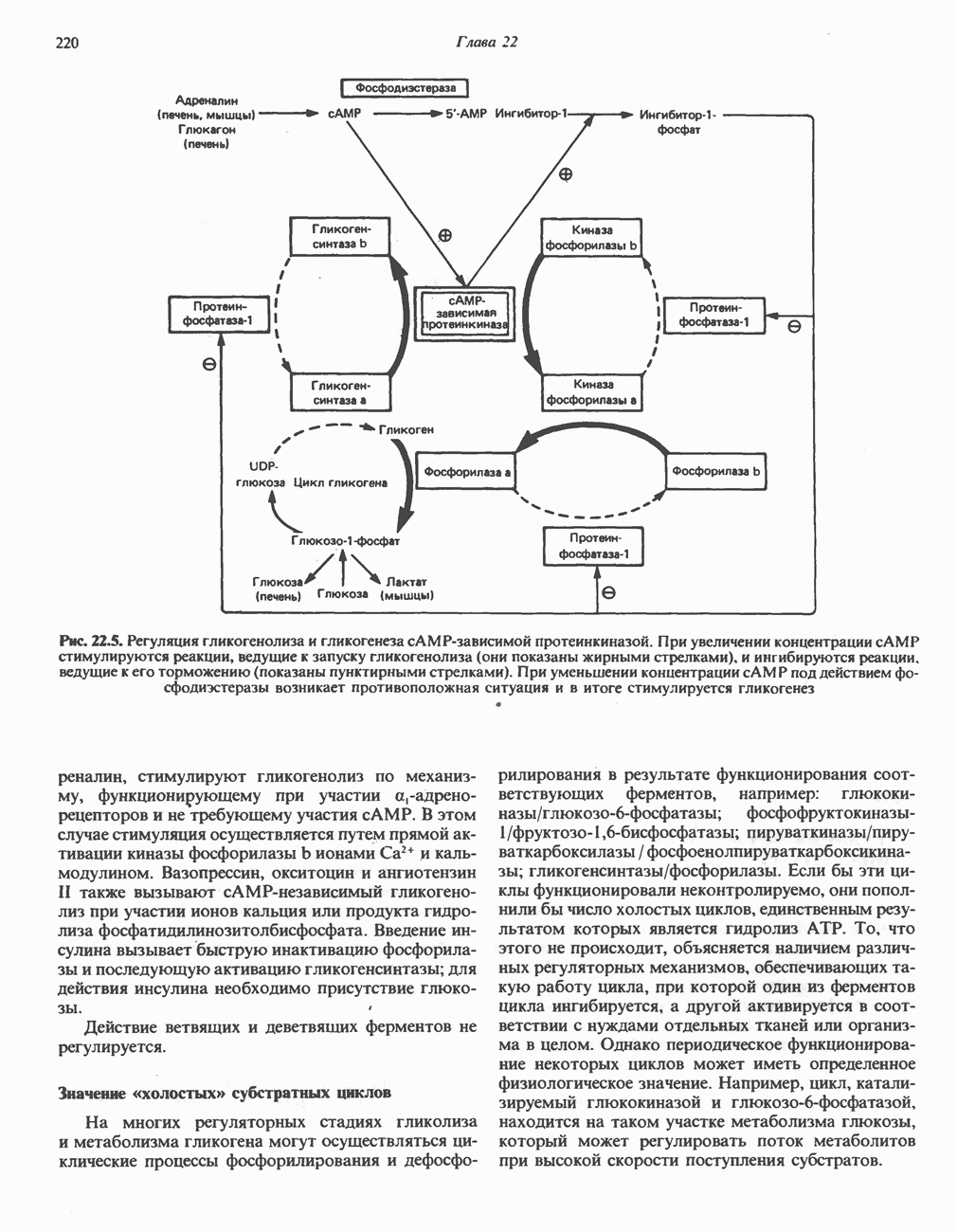
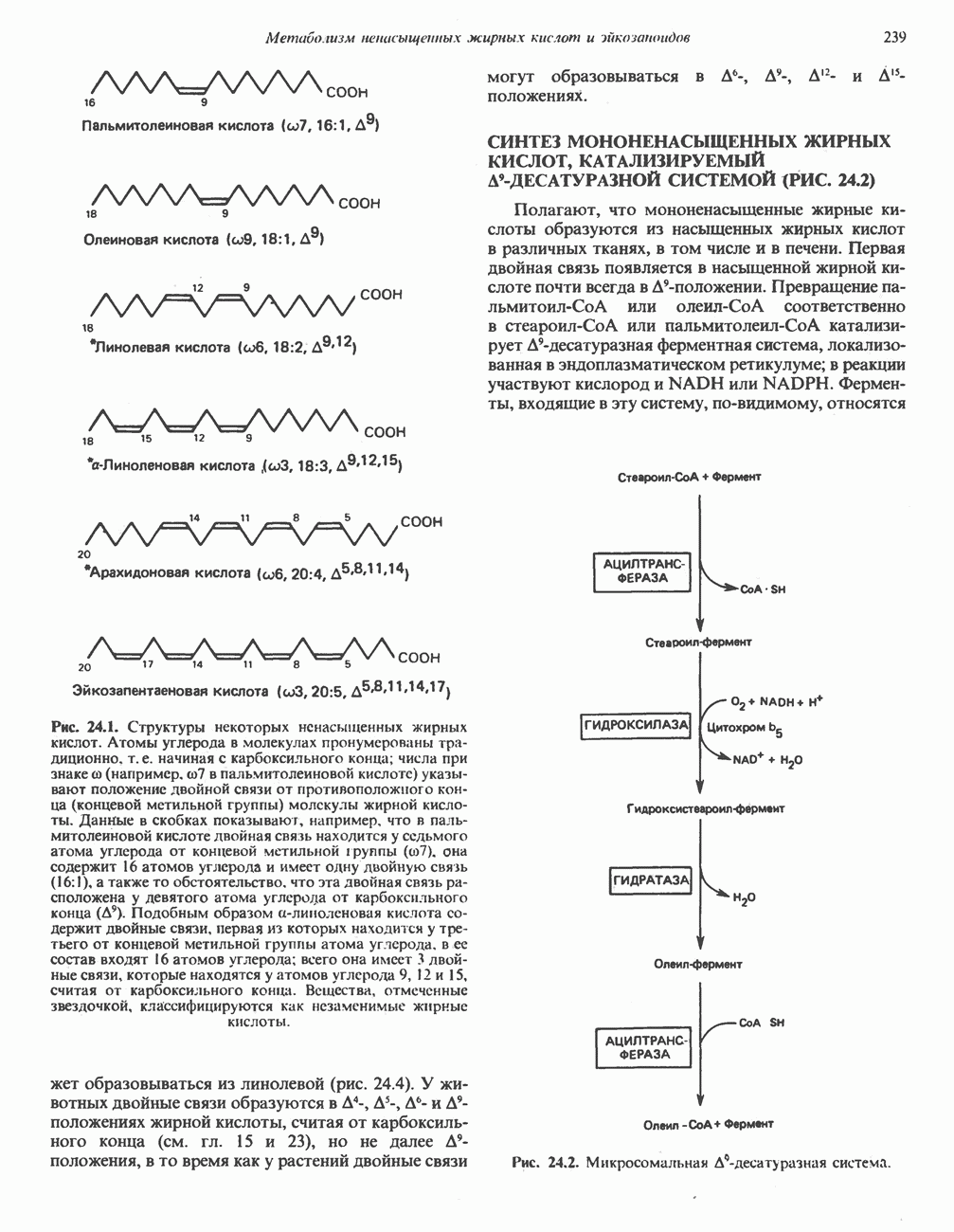
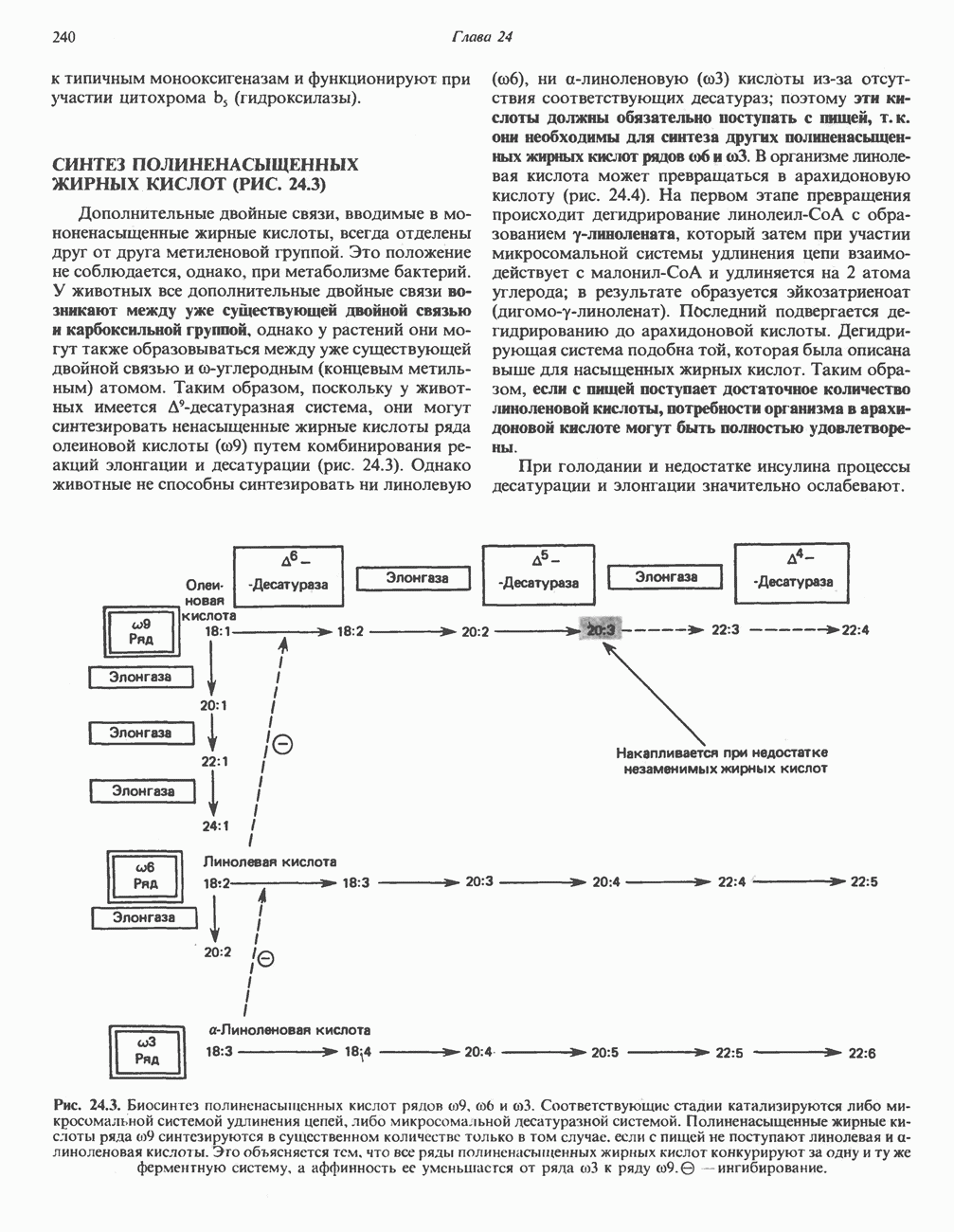
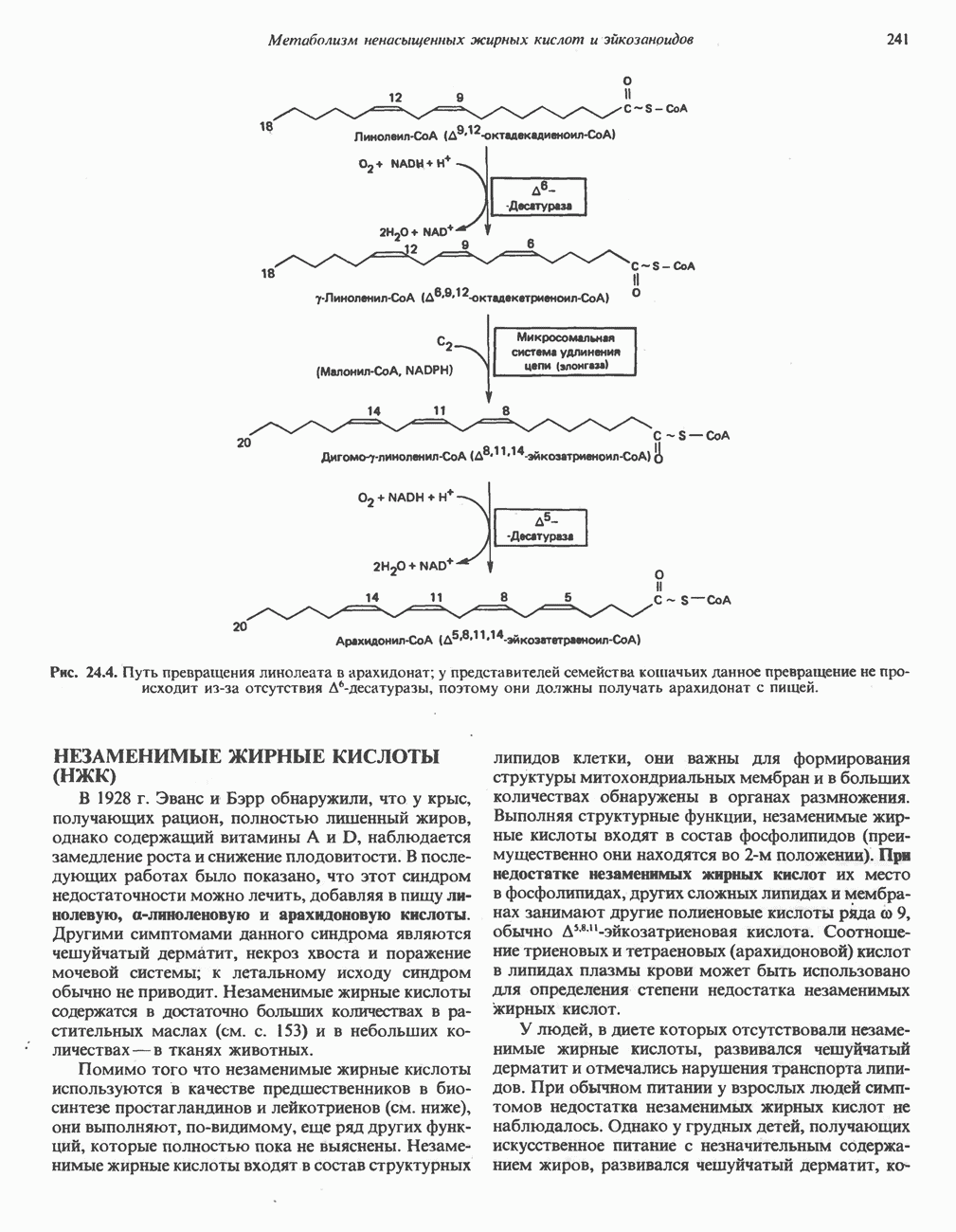
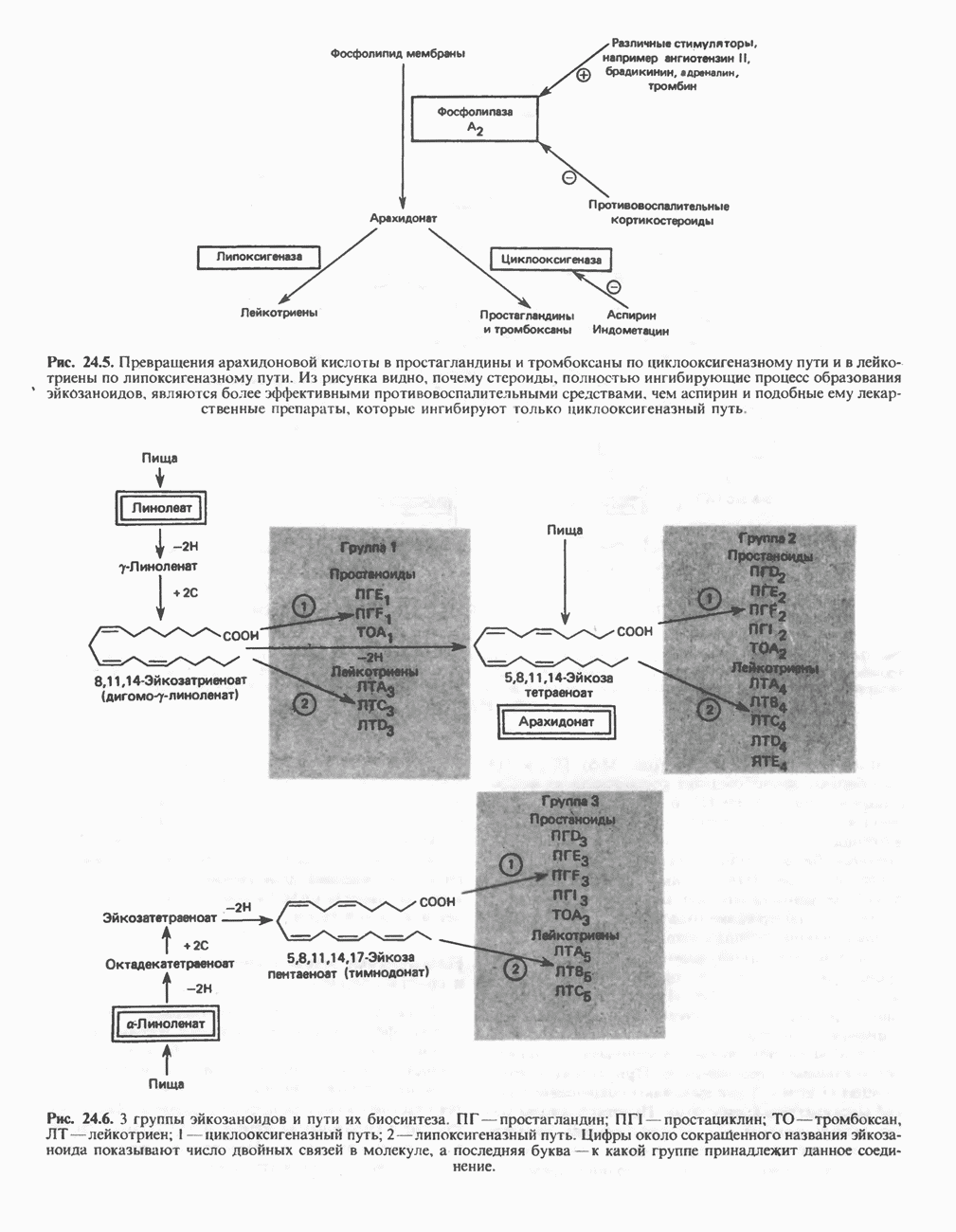
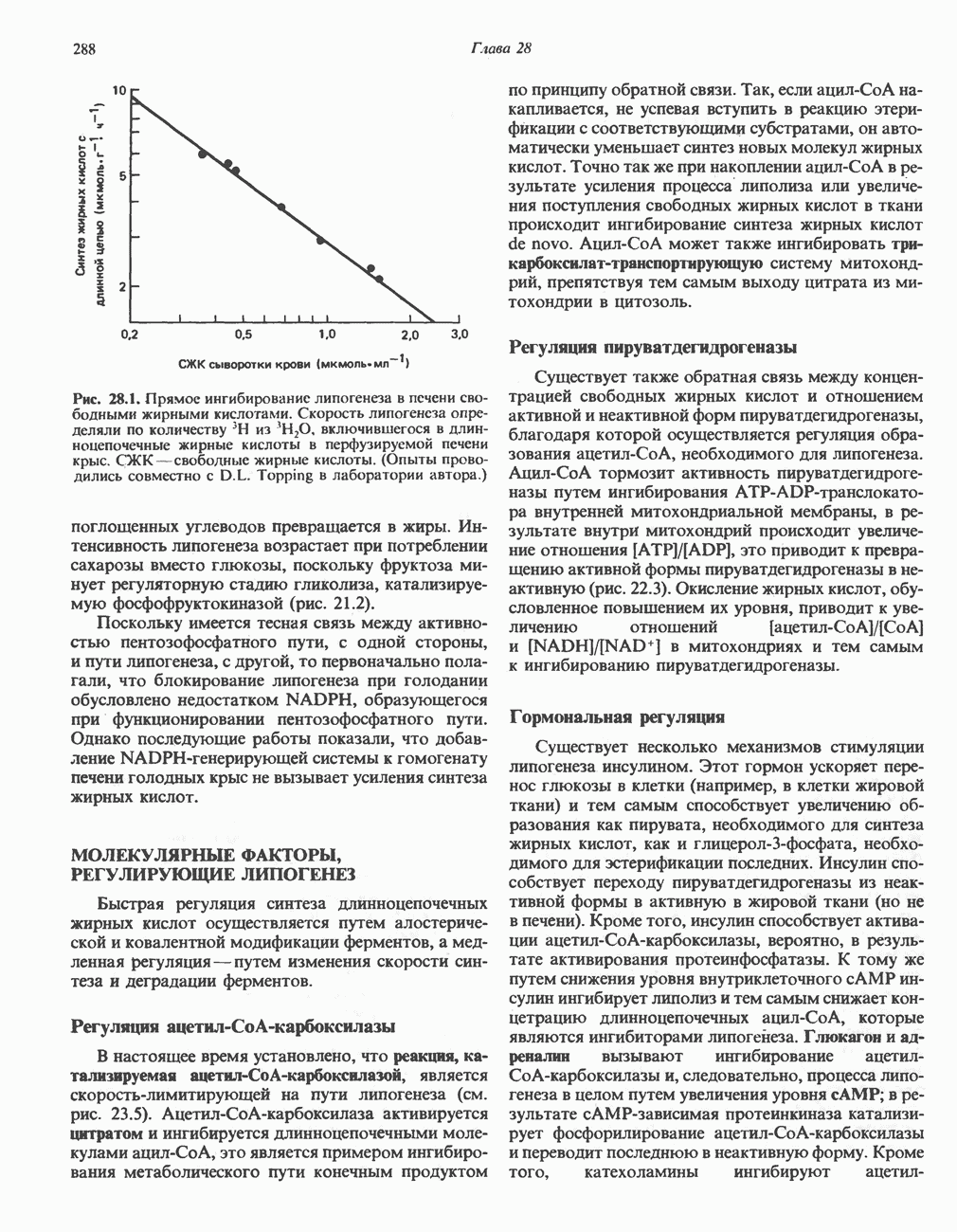
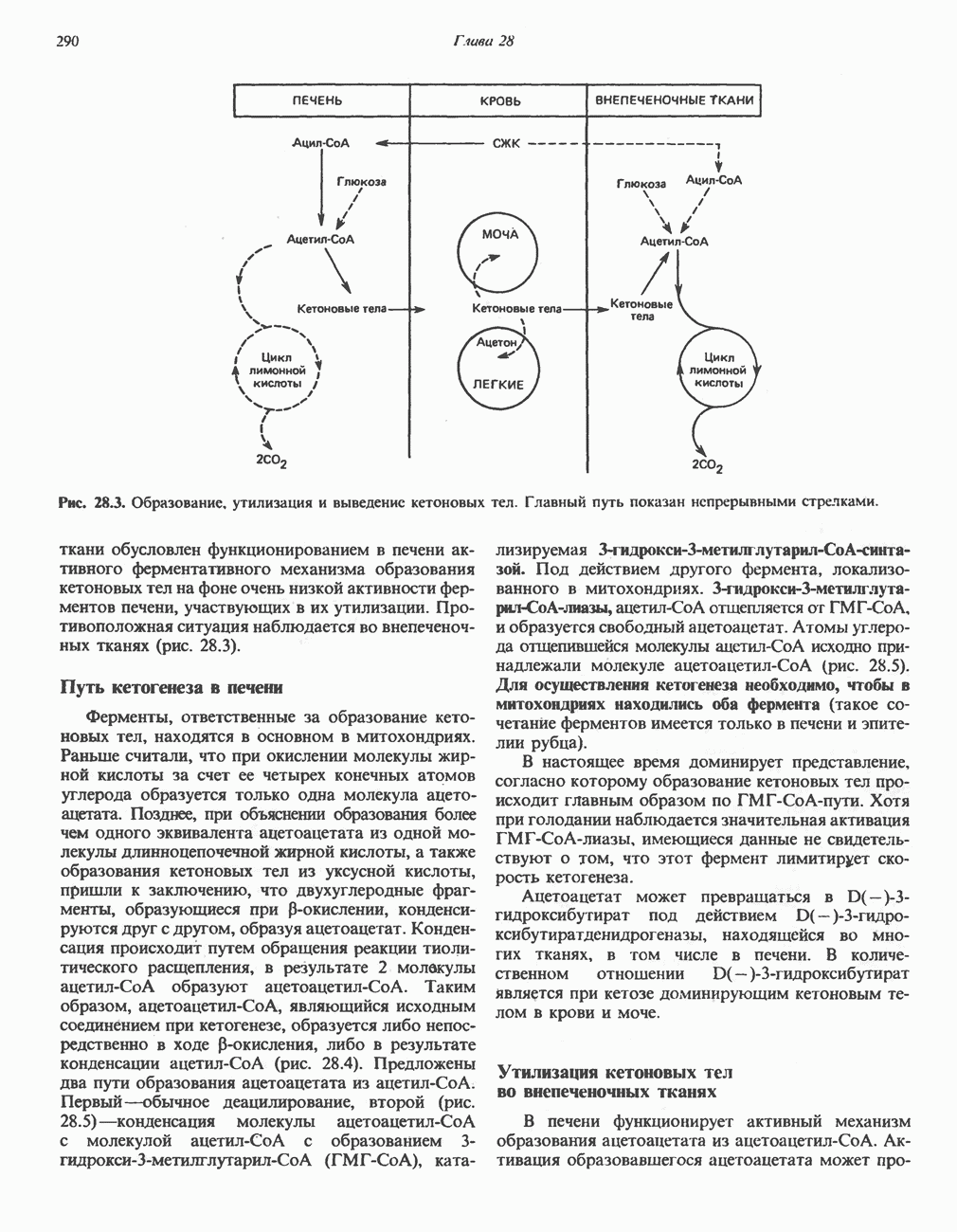
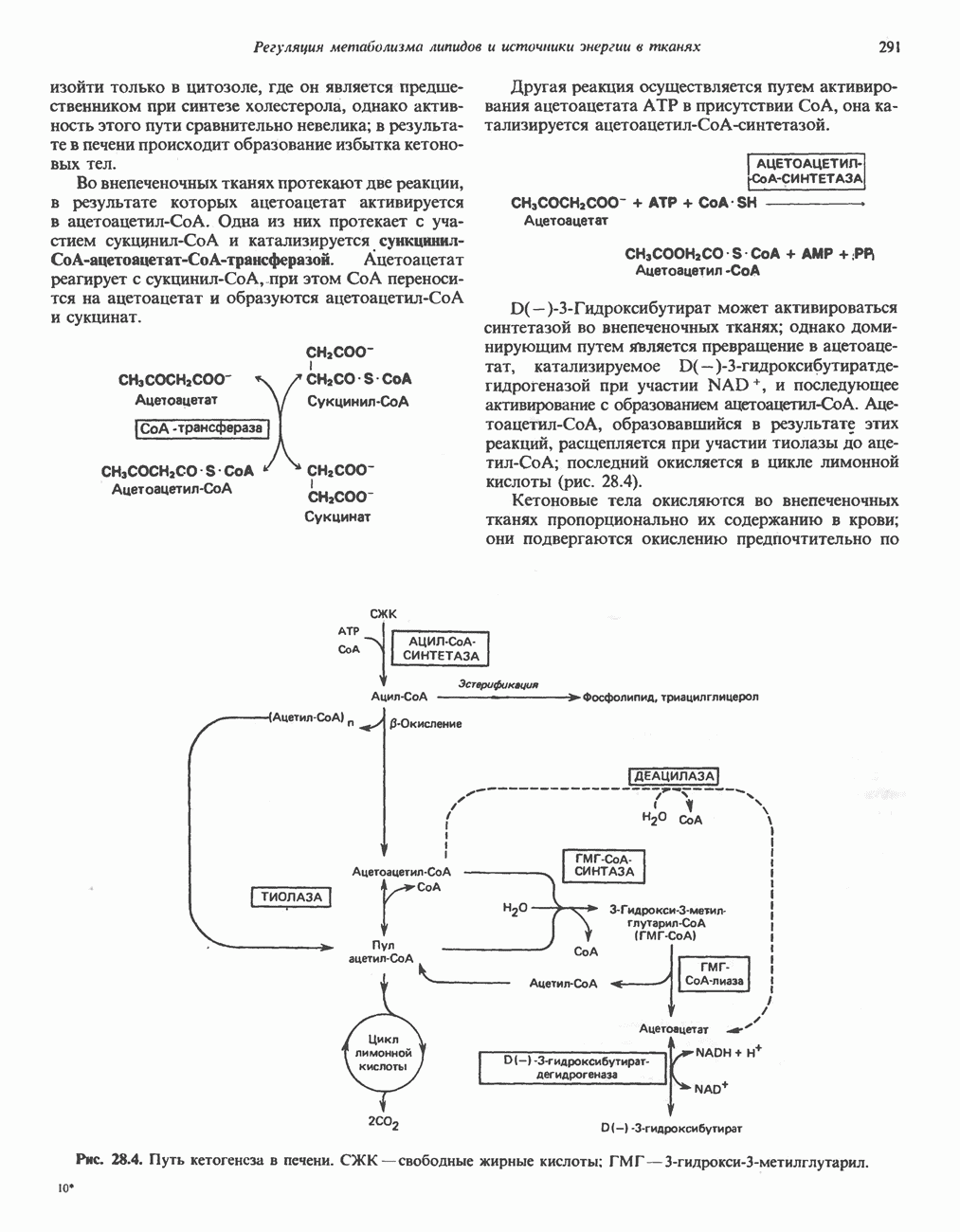
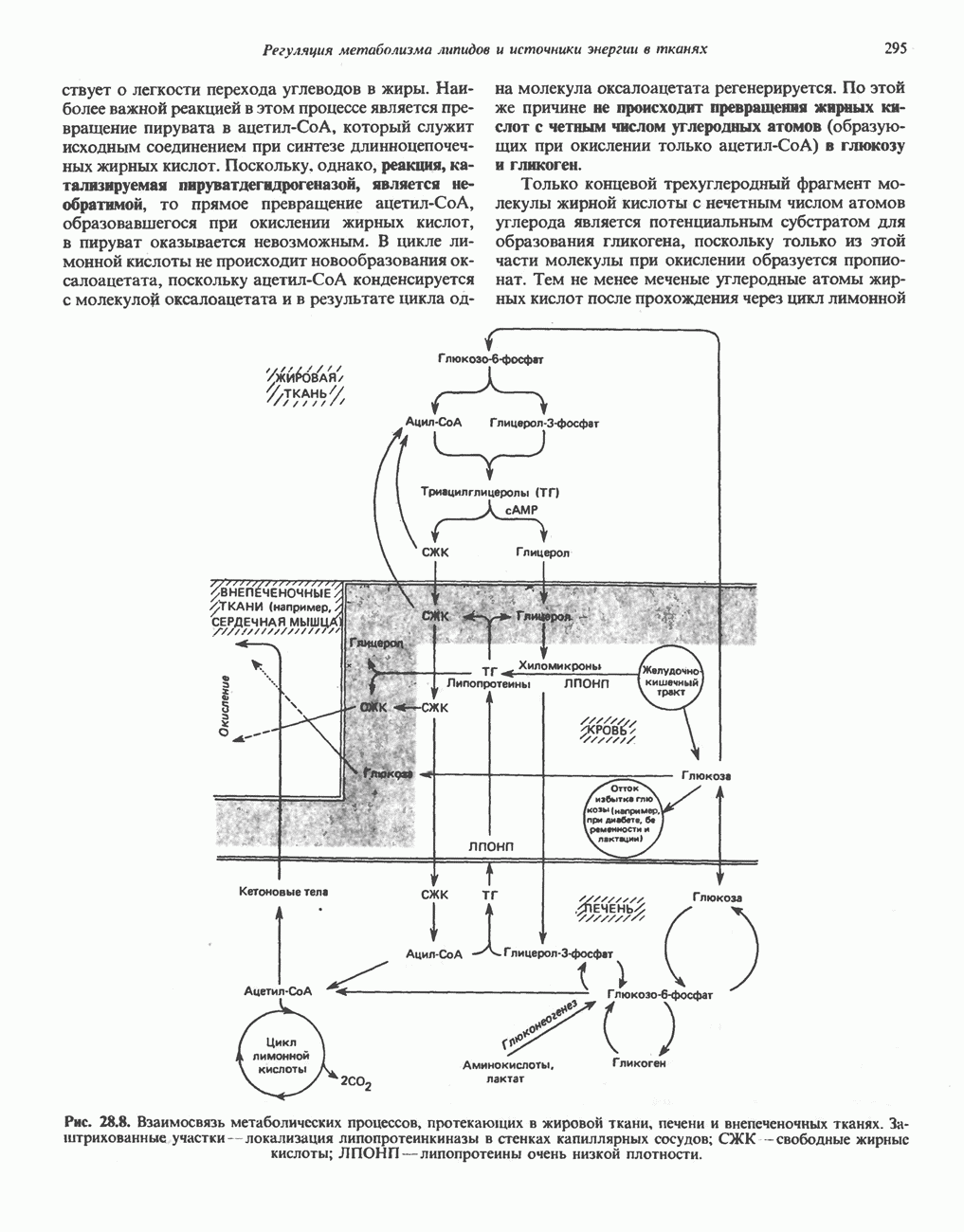
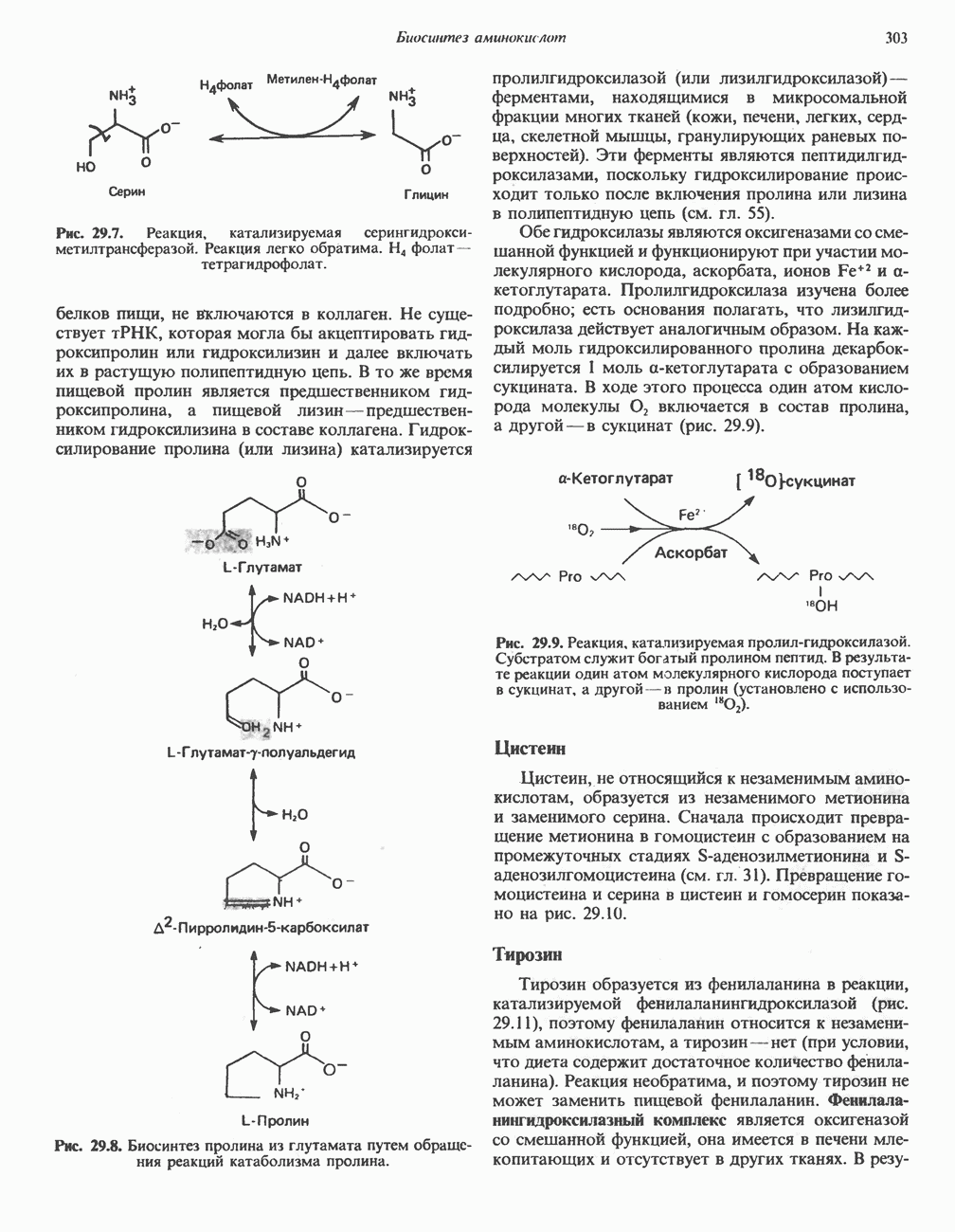
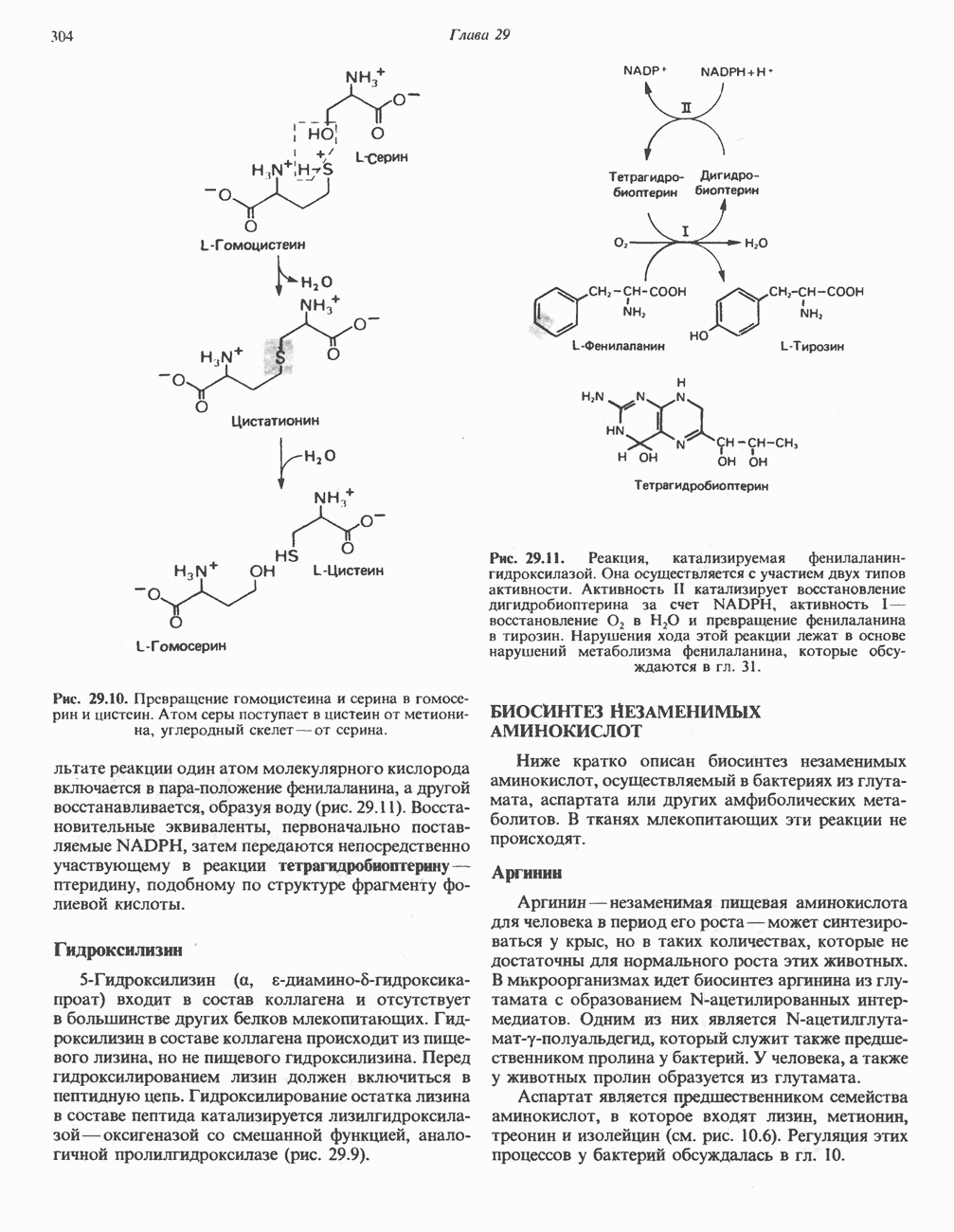
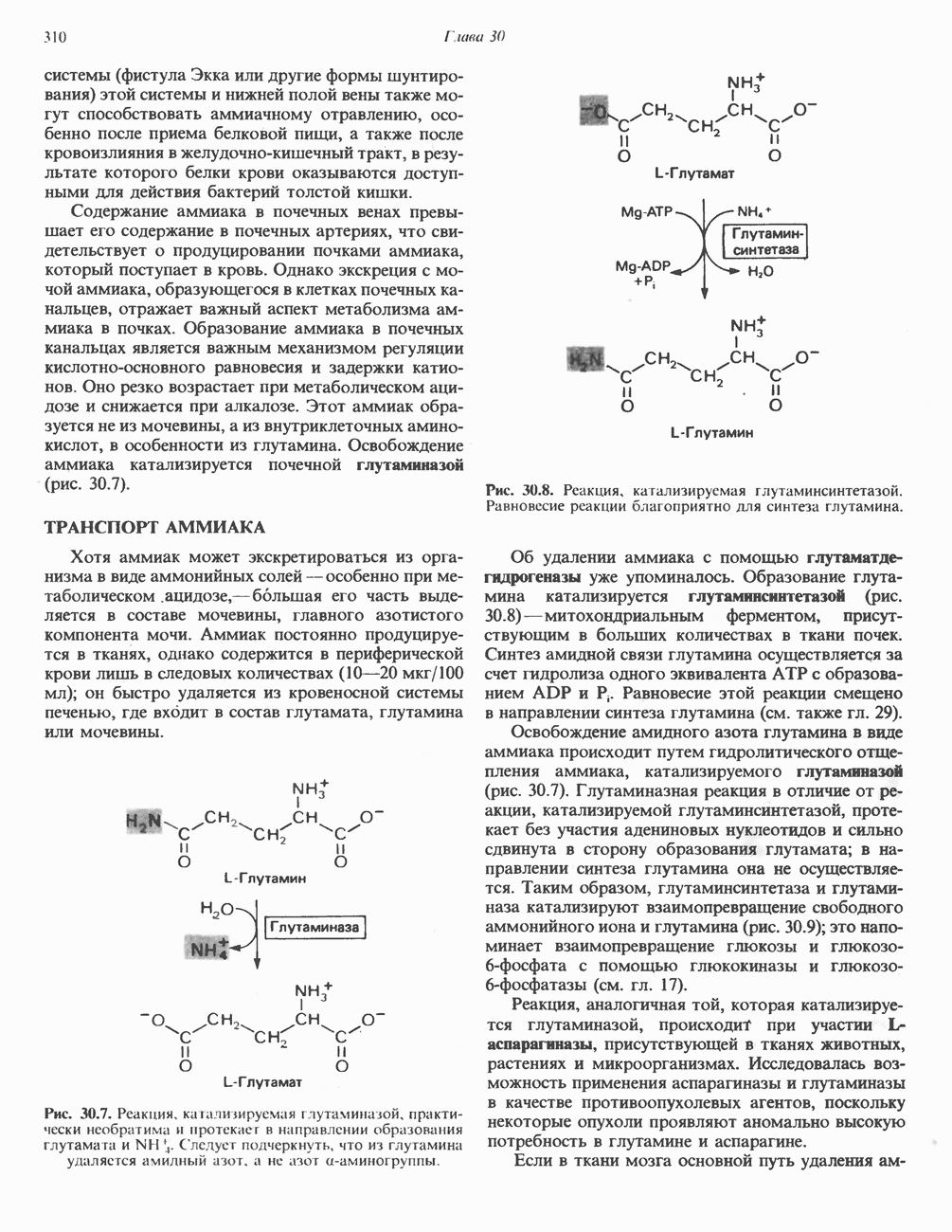
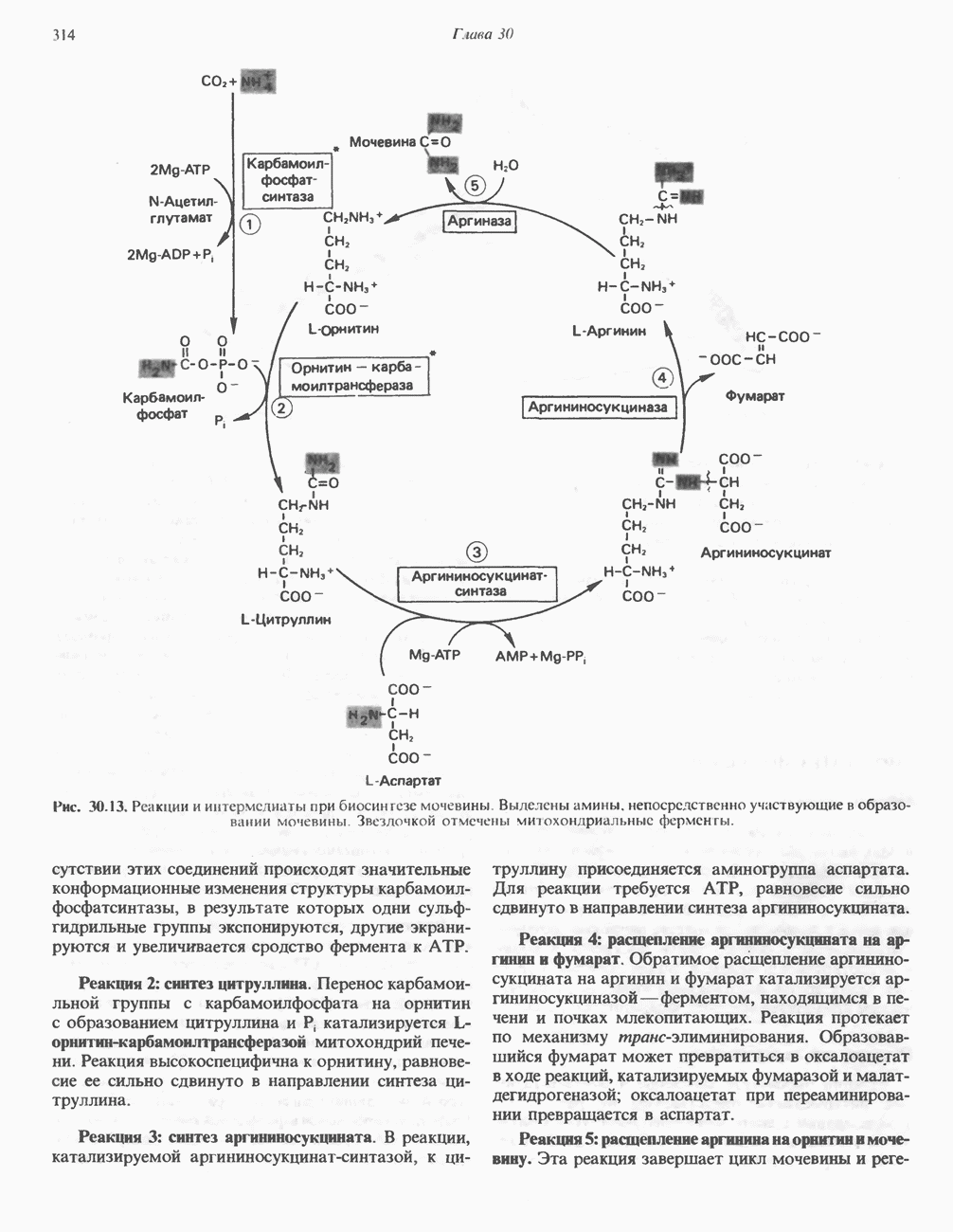
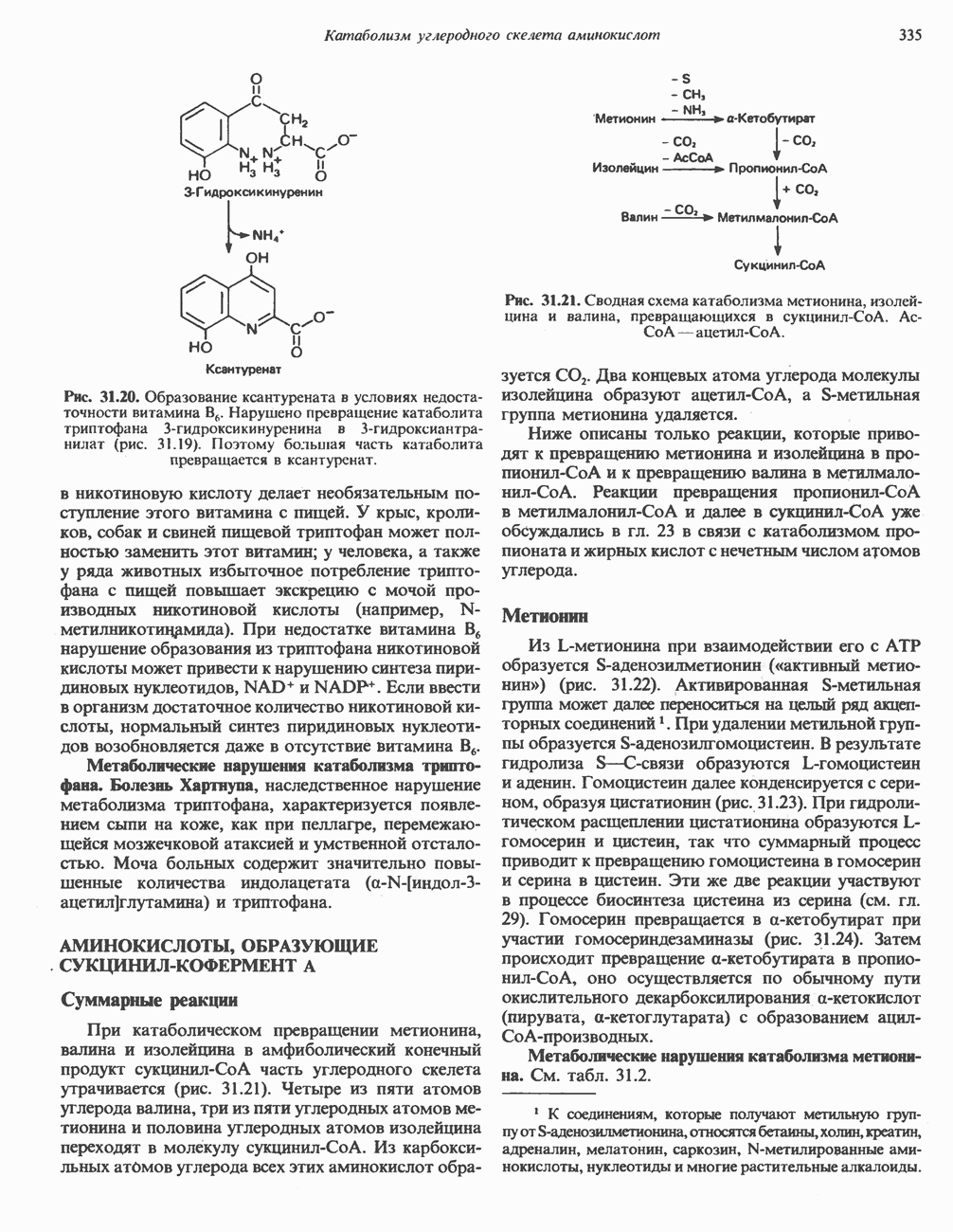
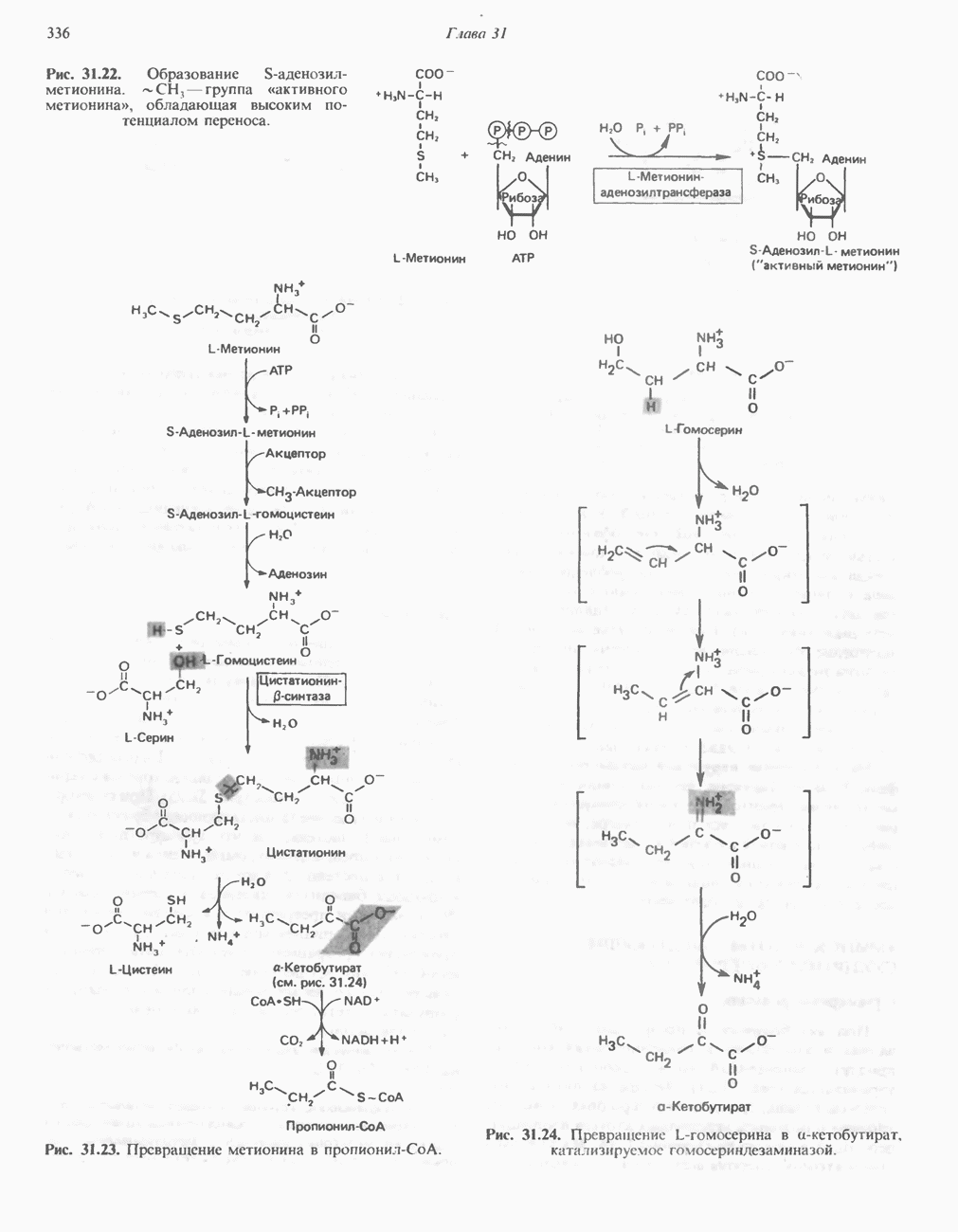
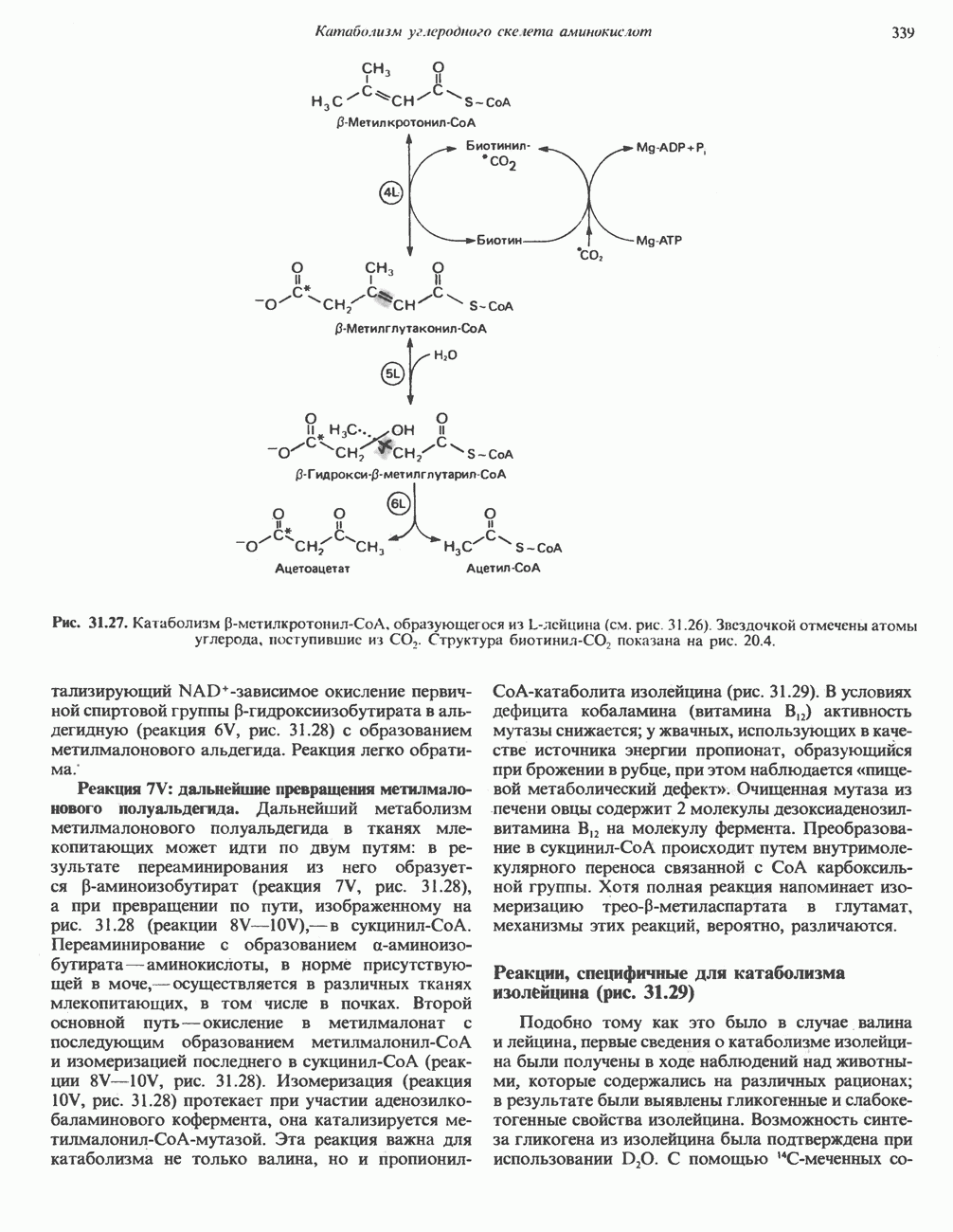
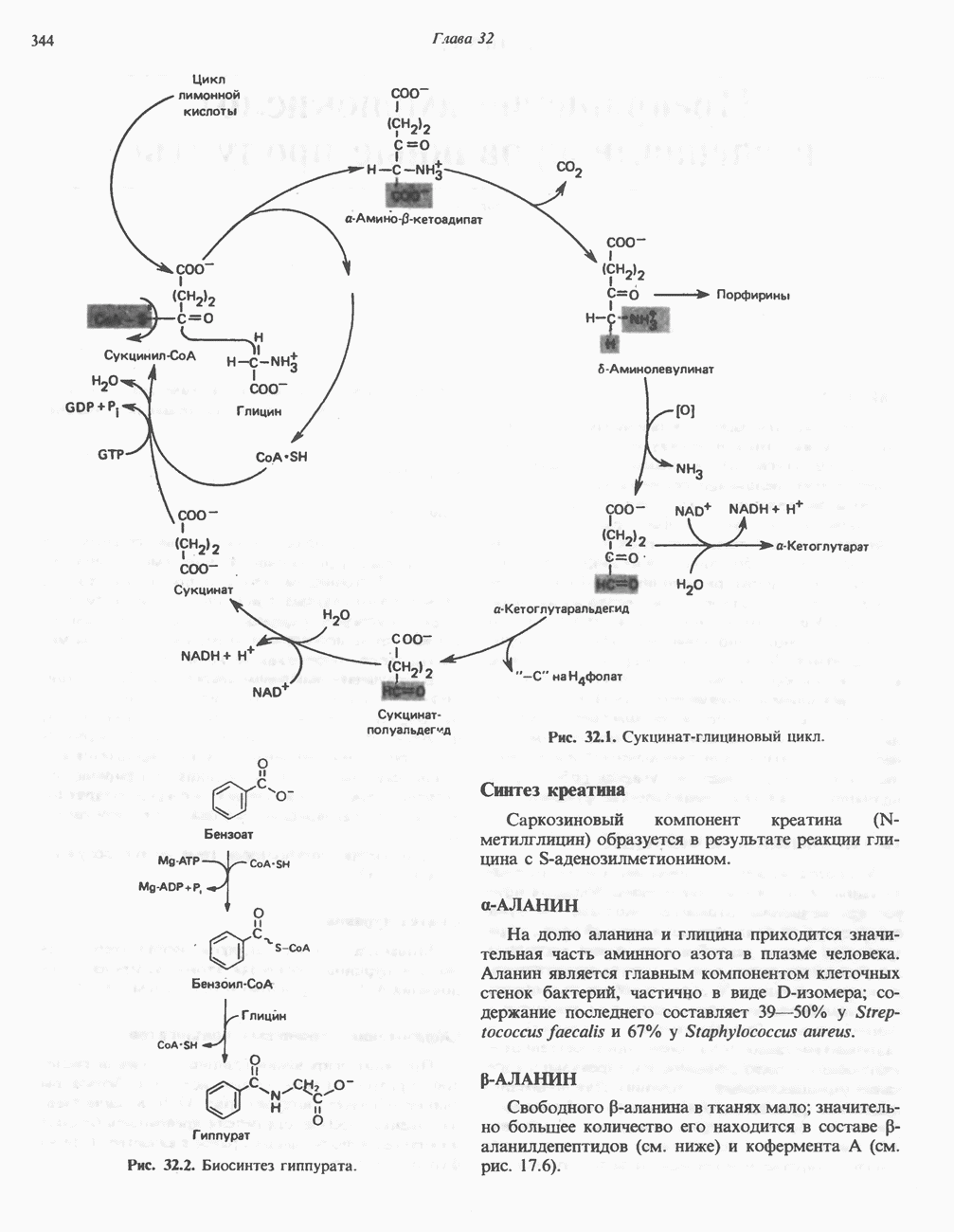
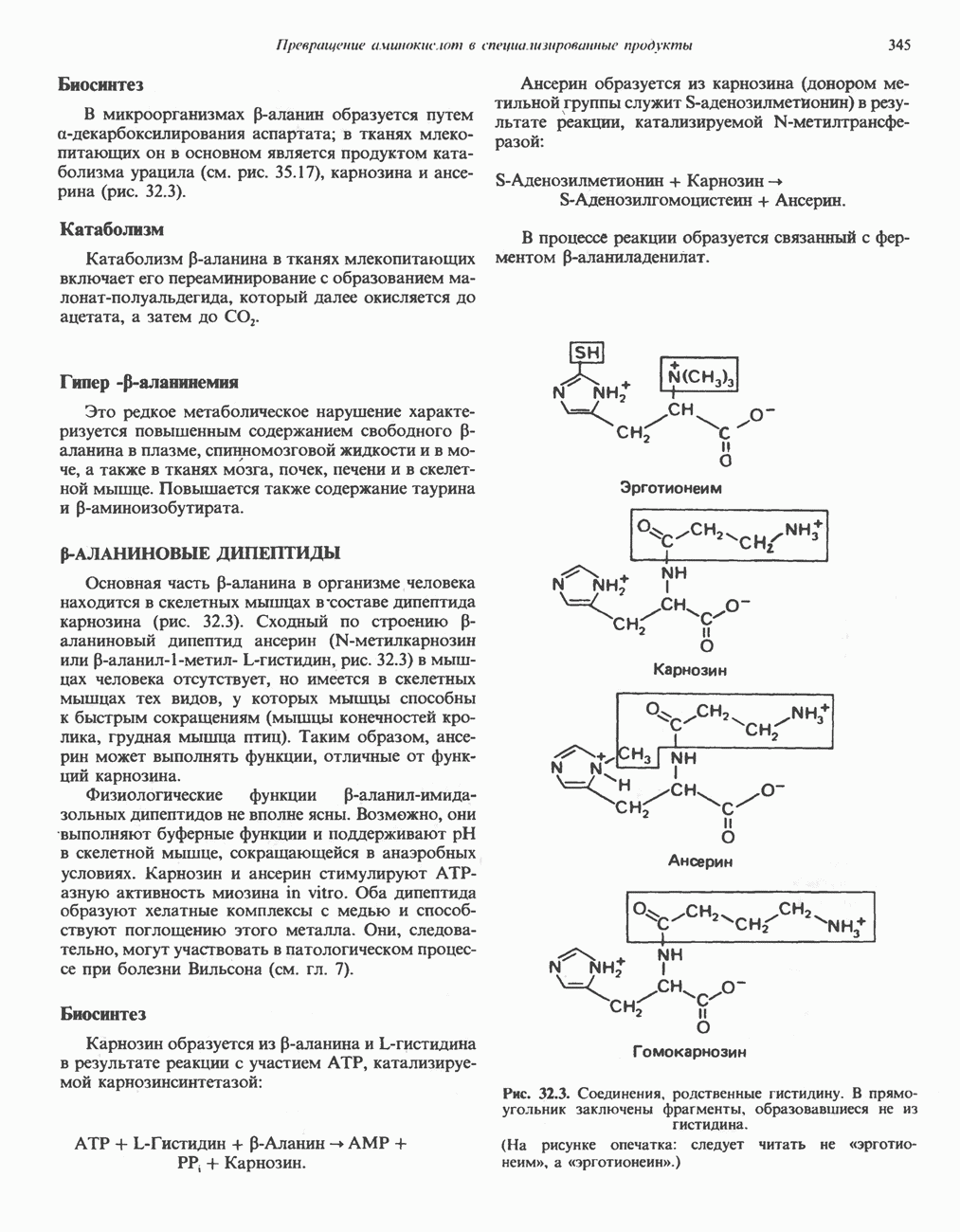
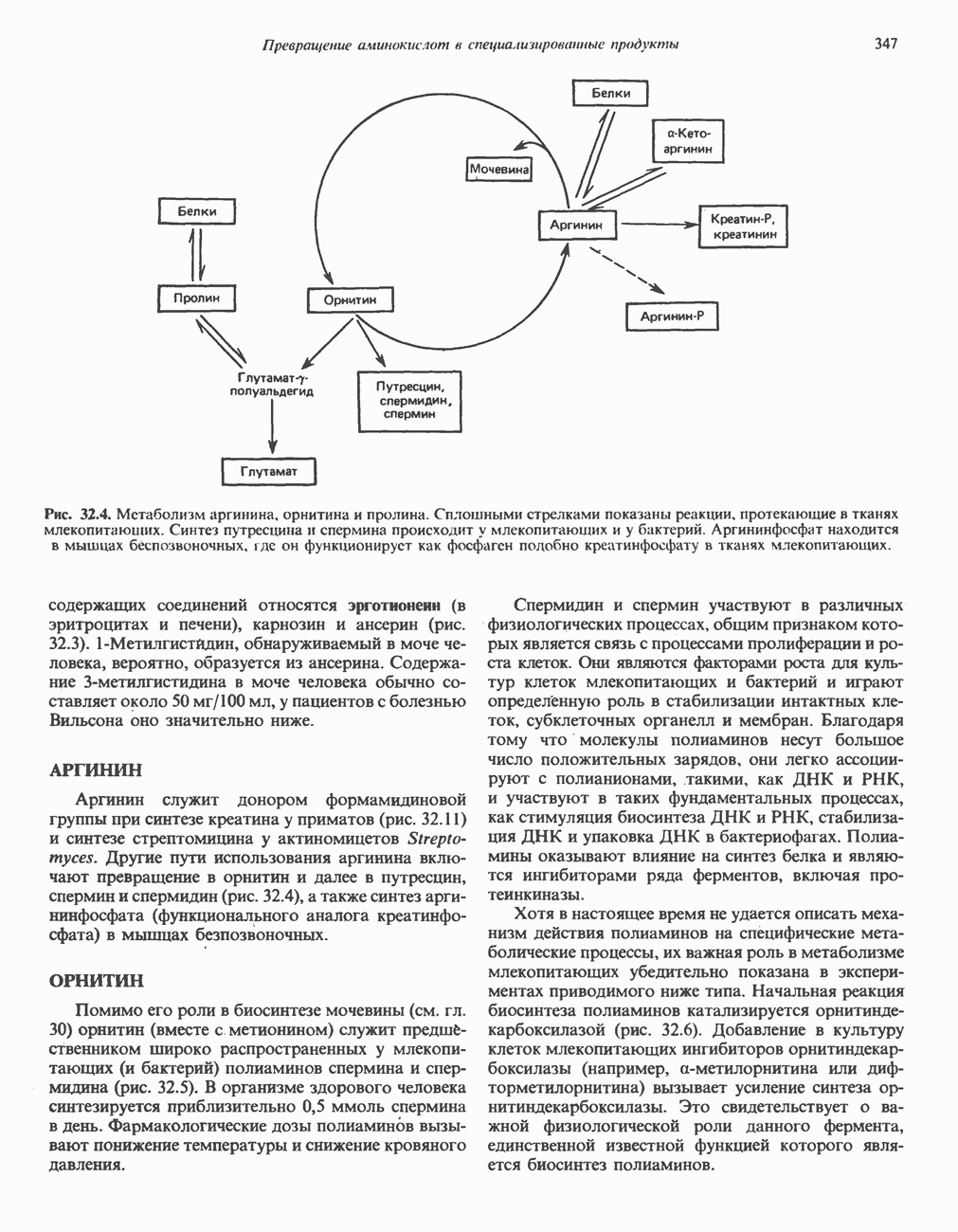
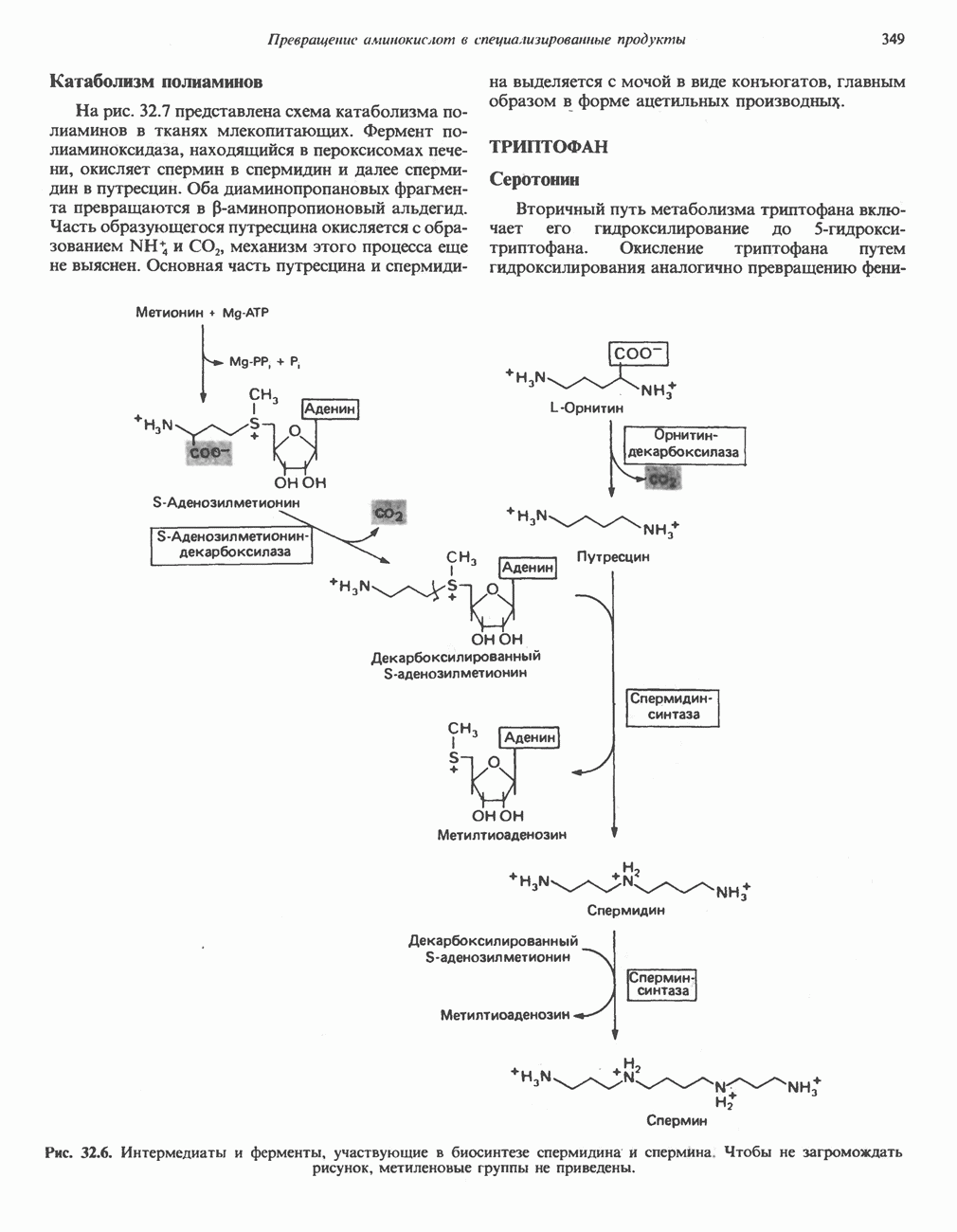
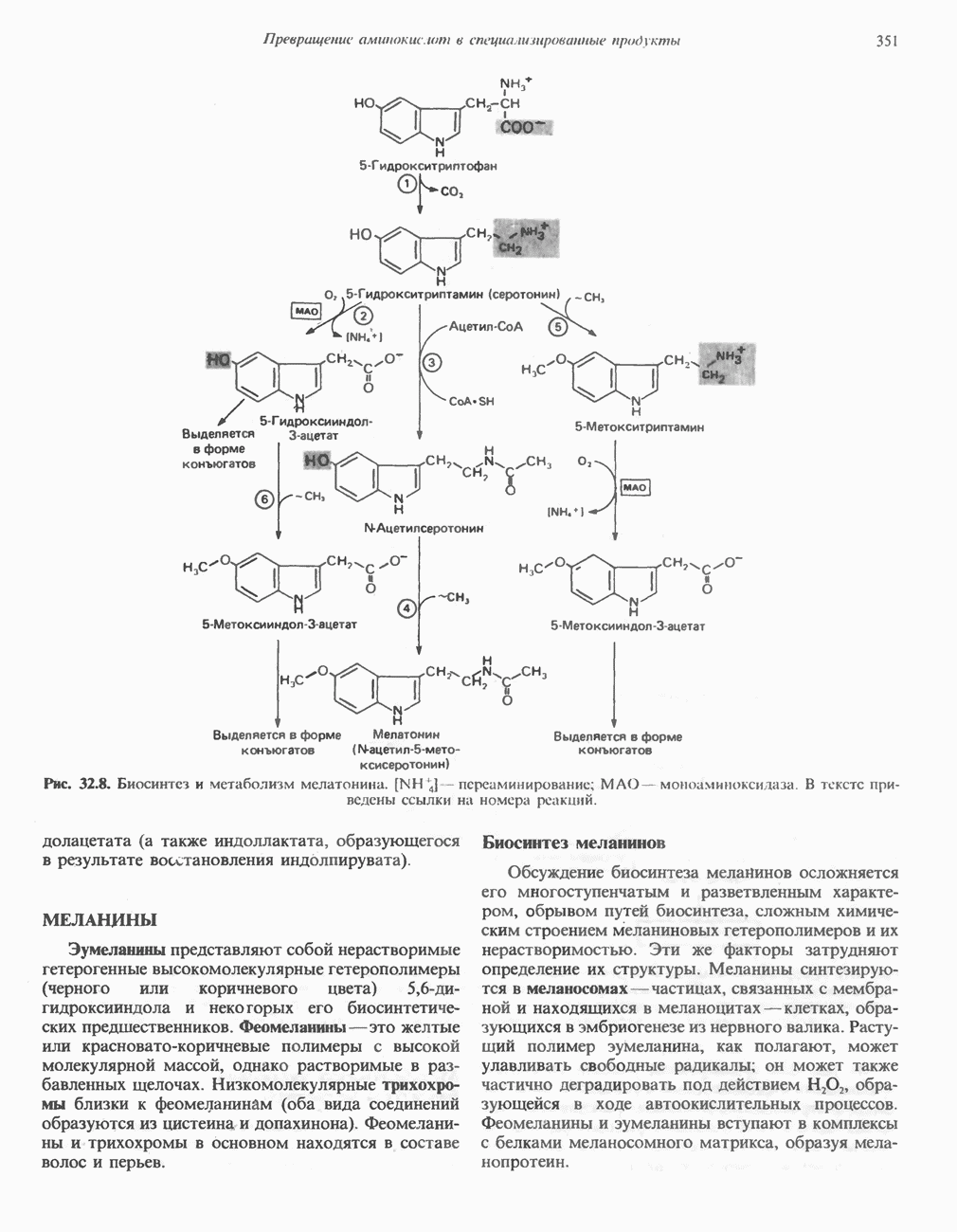
Пред.
След.
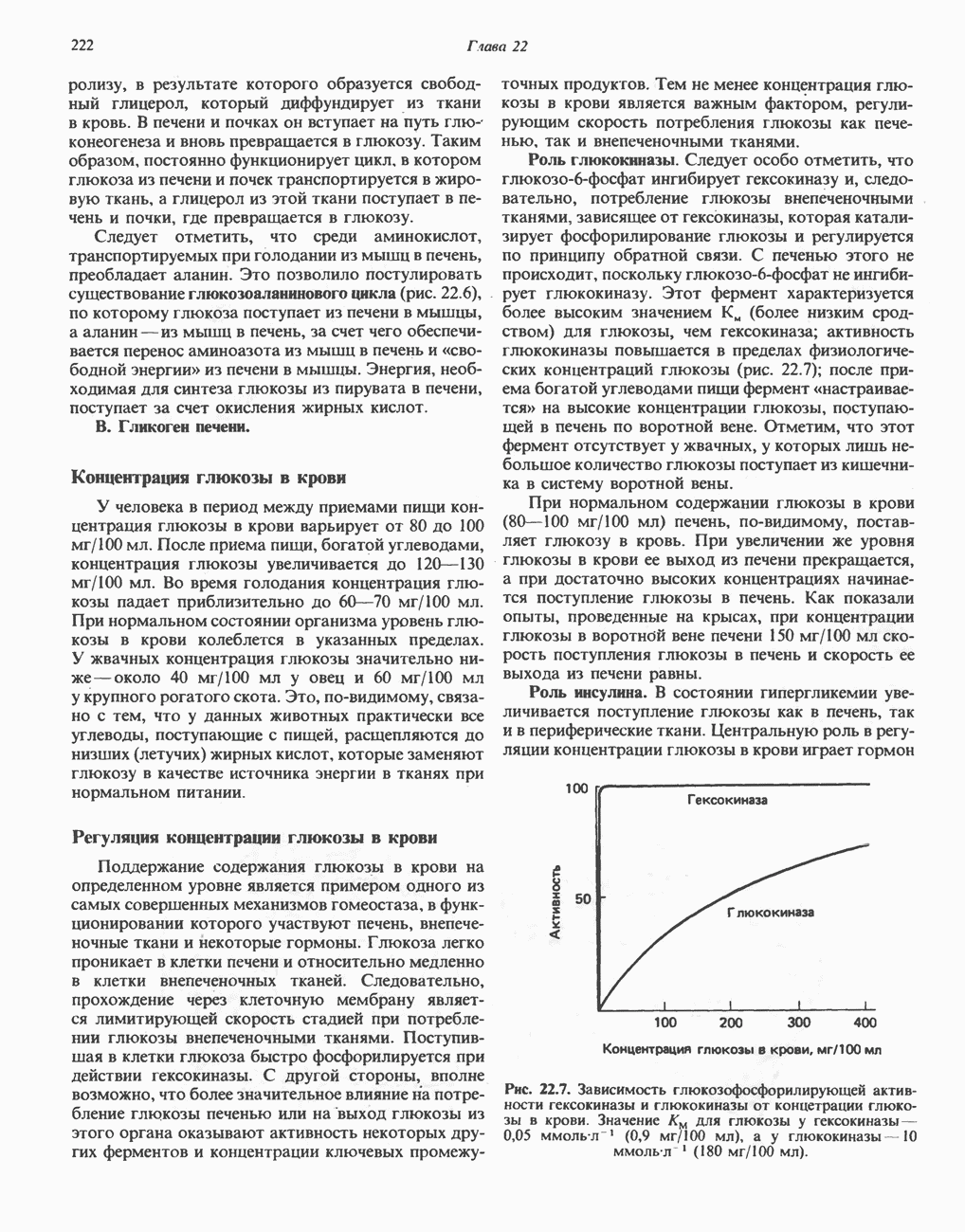
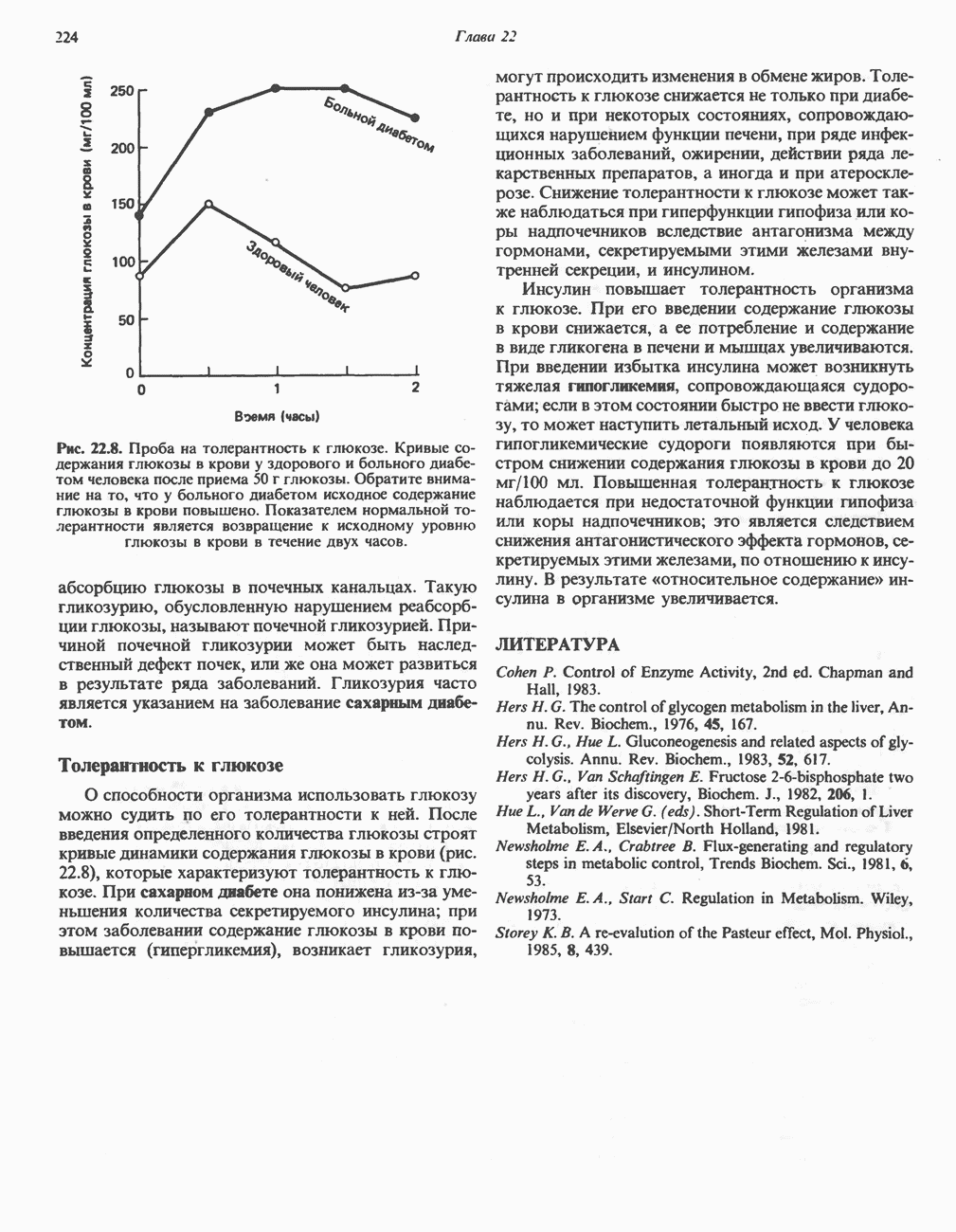
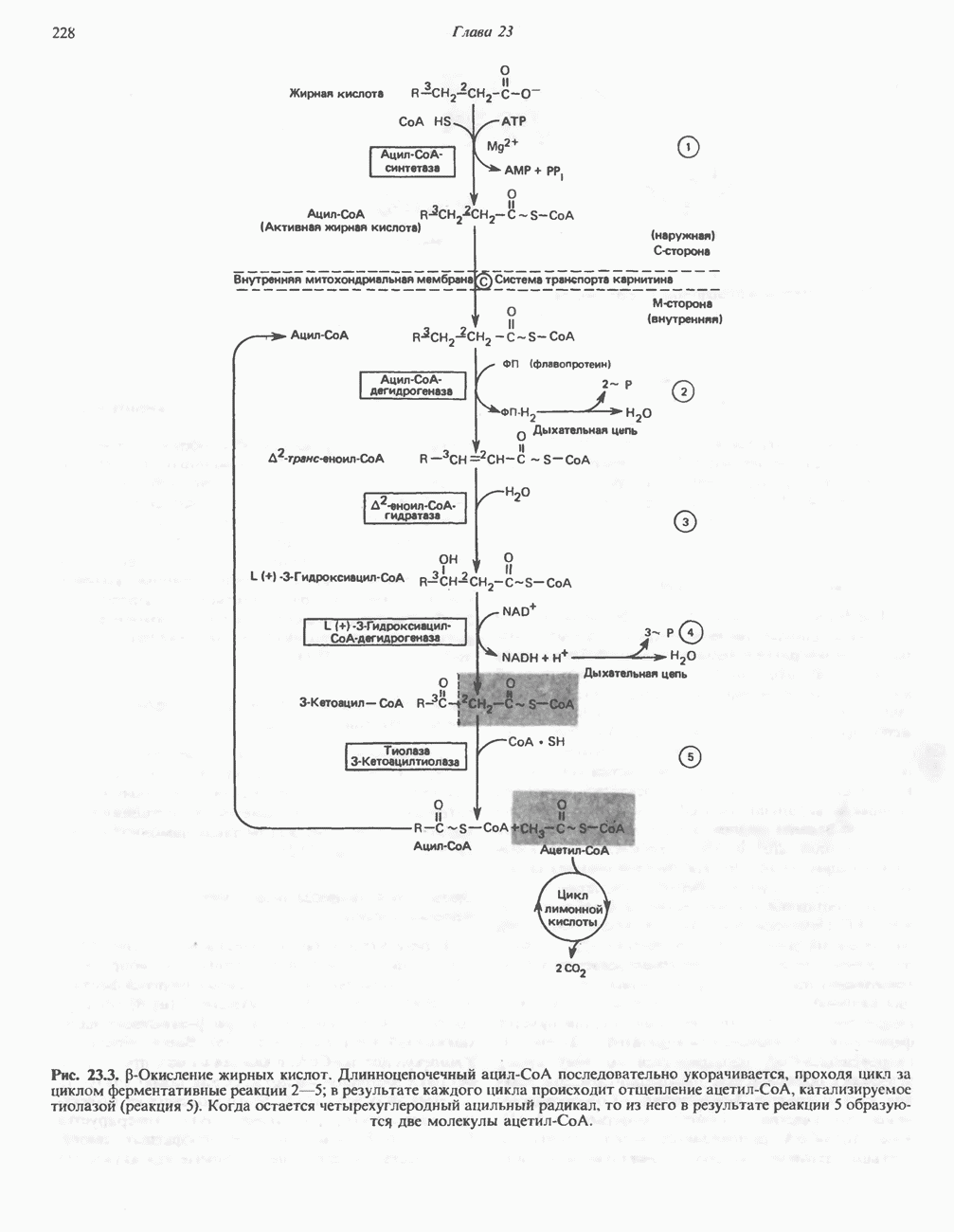
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
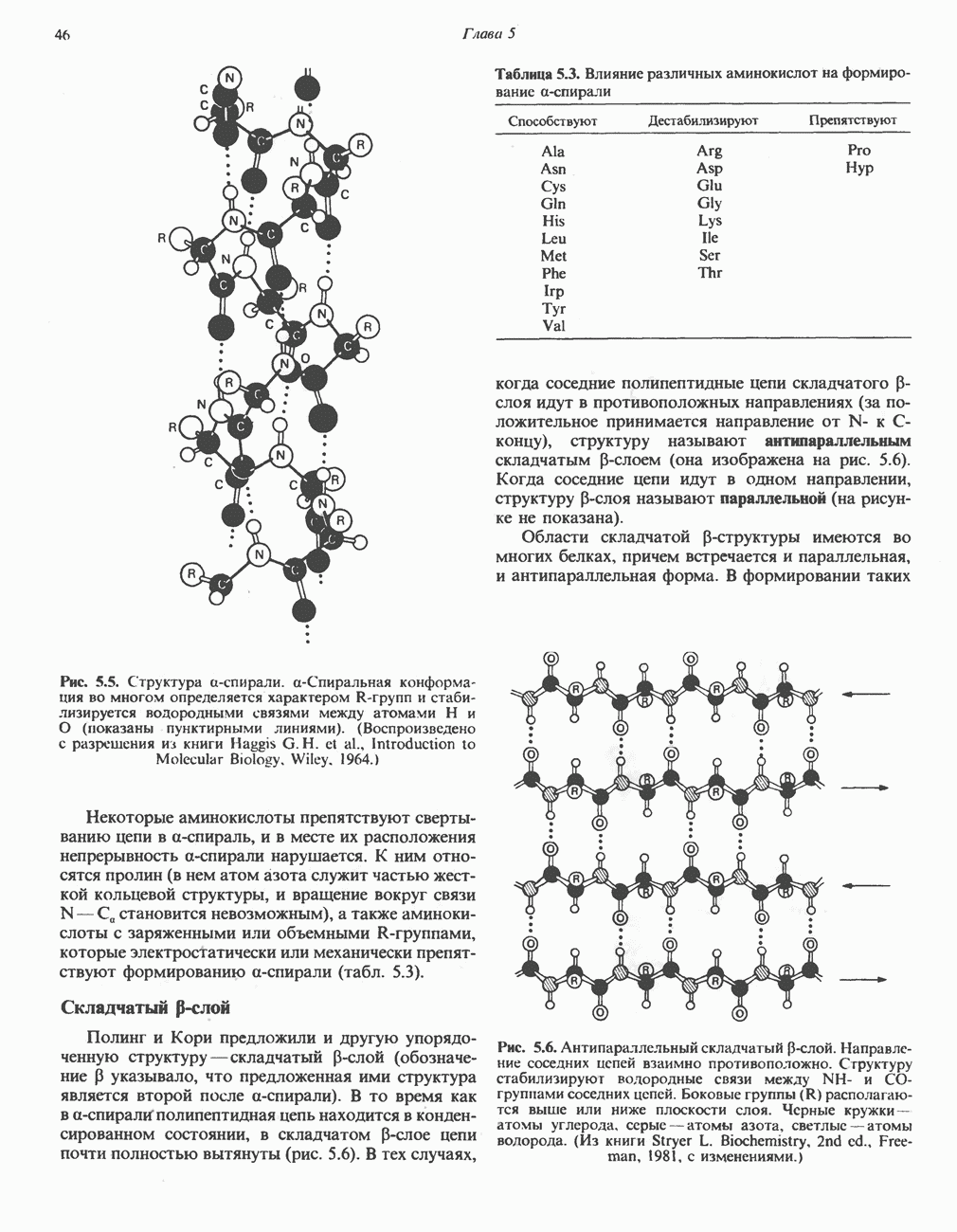
ZADANIA.TO
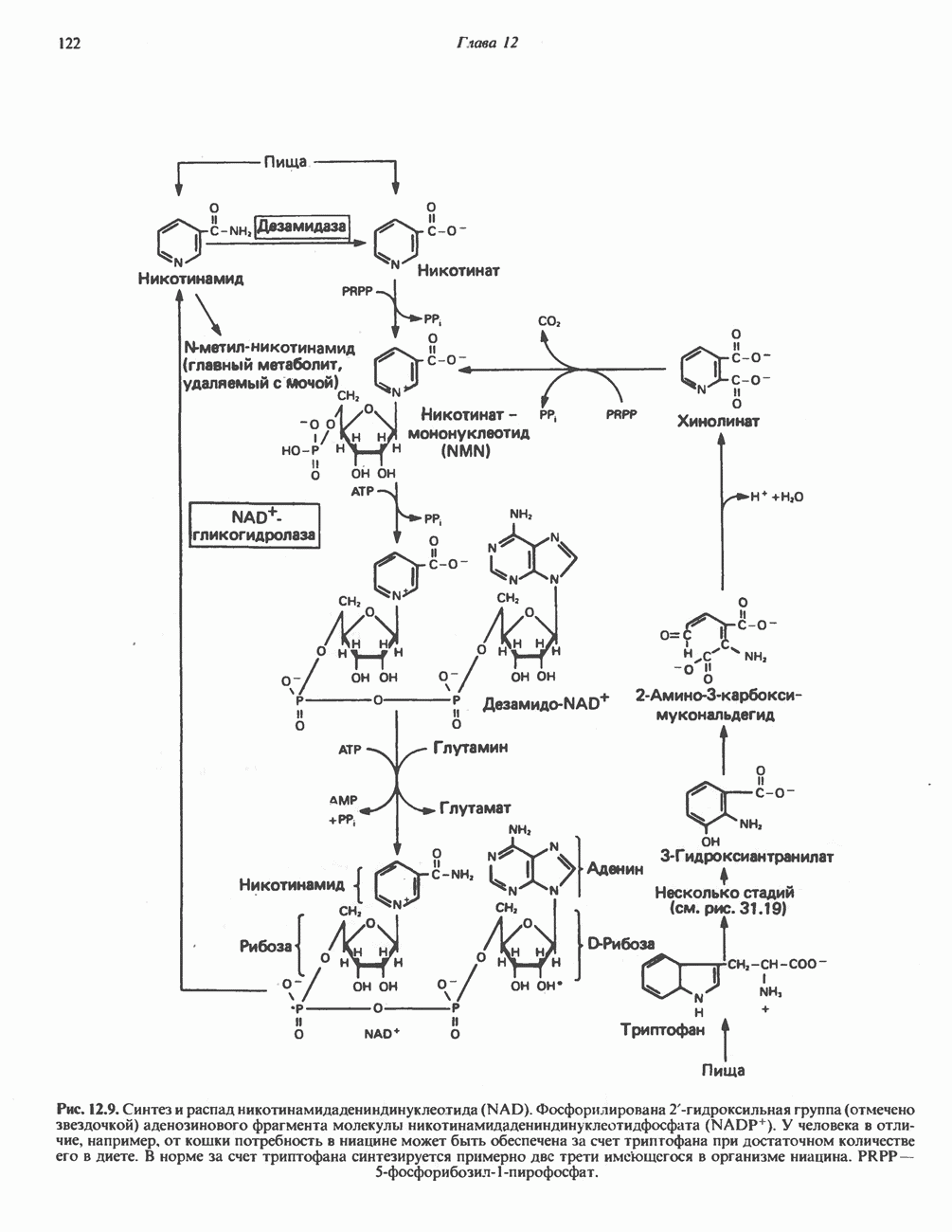
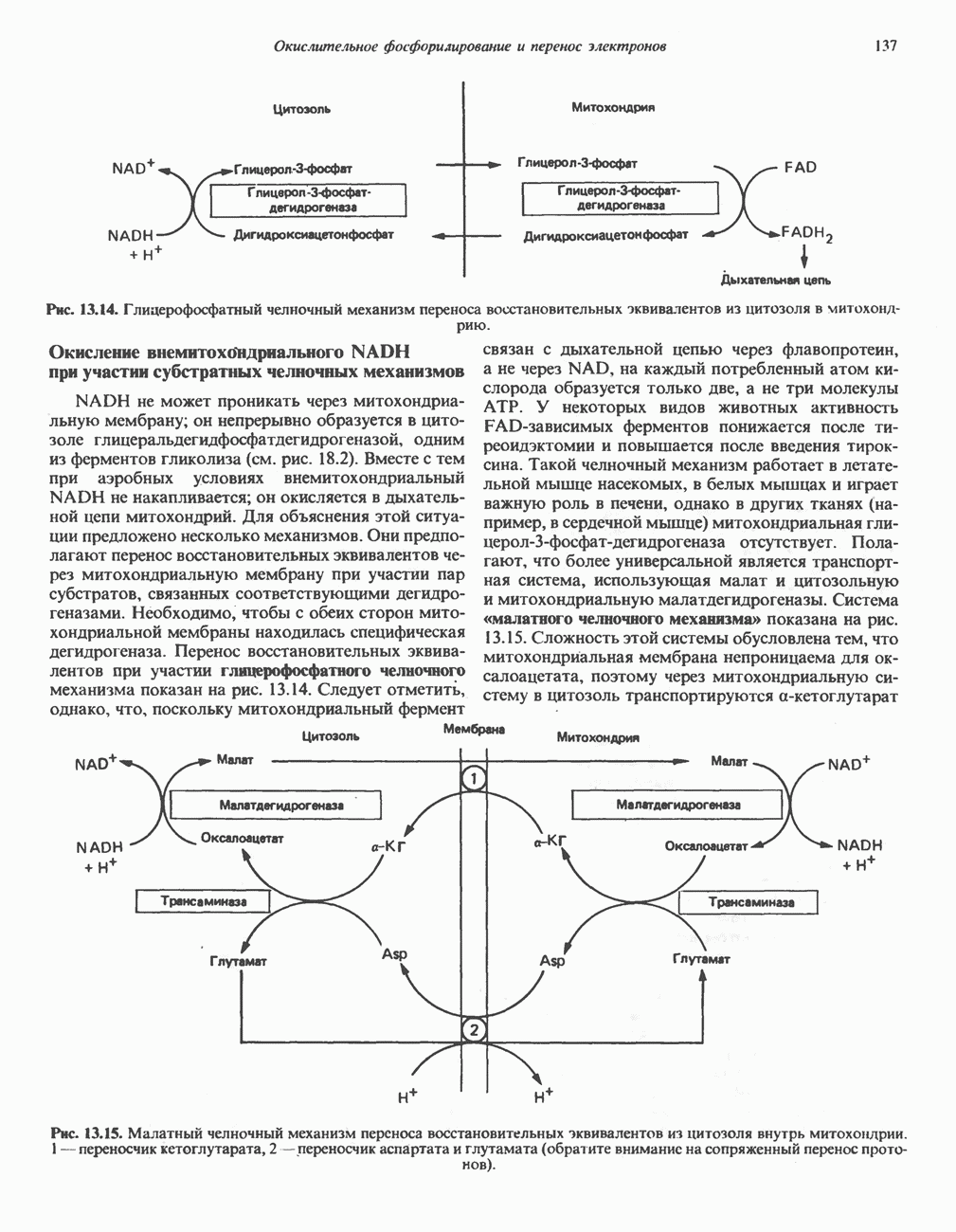
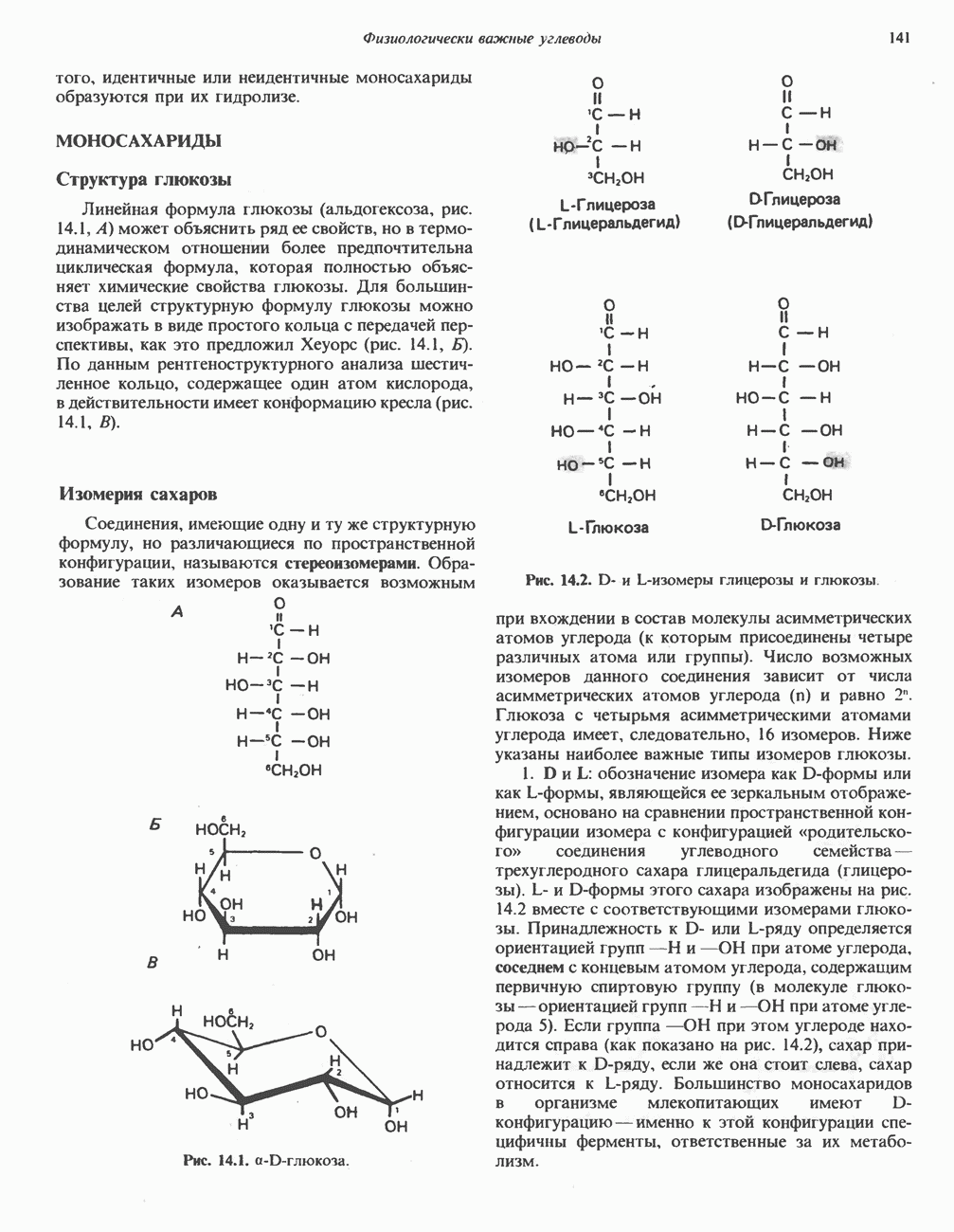
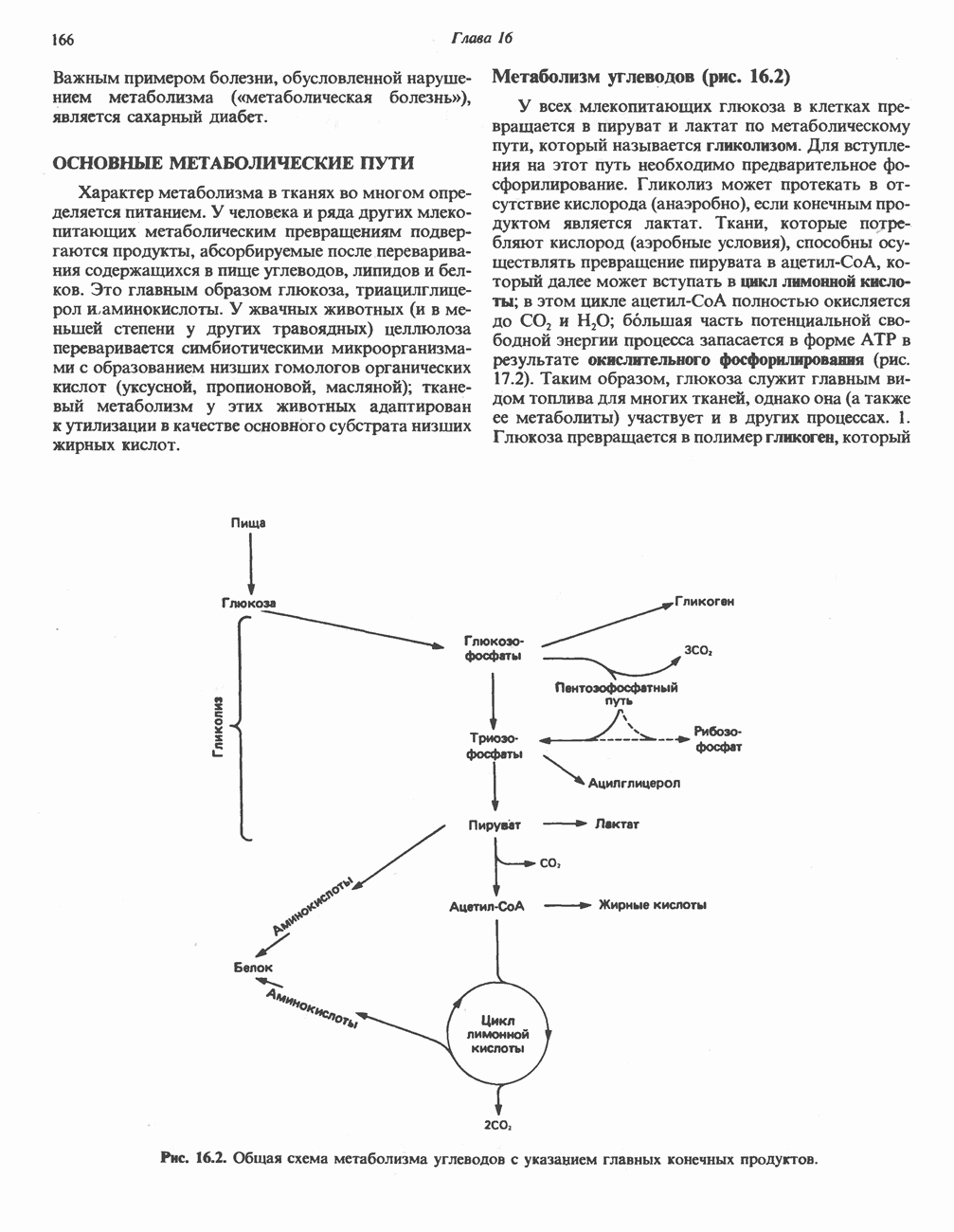
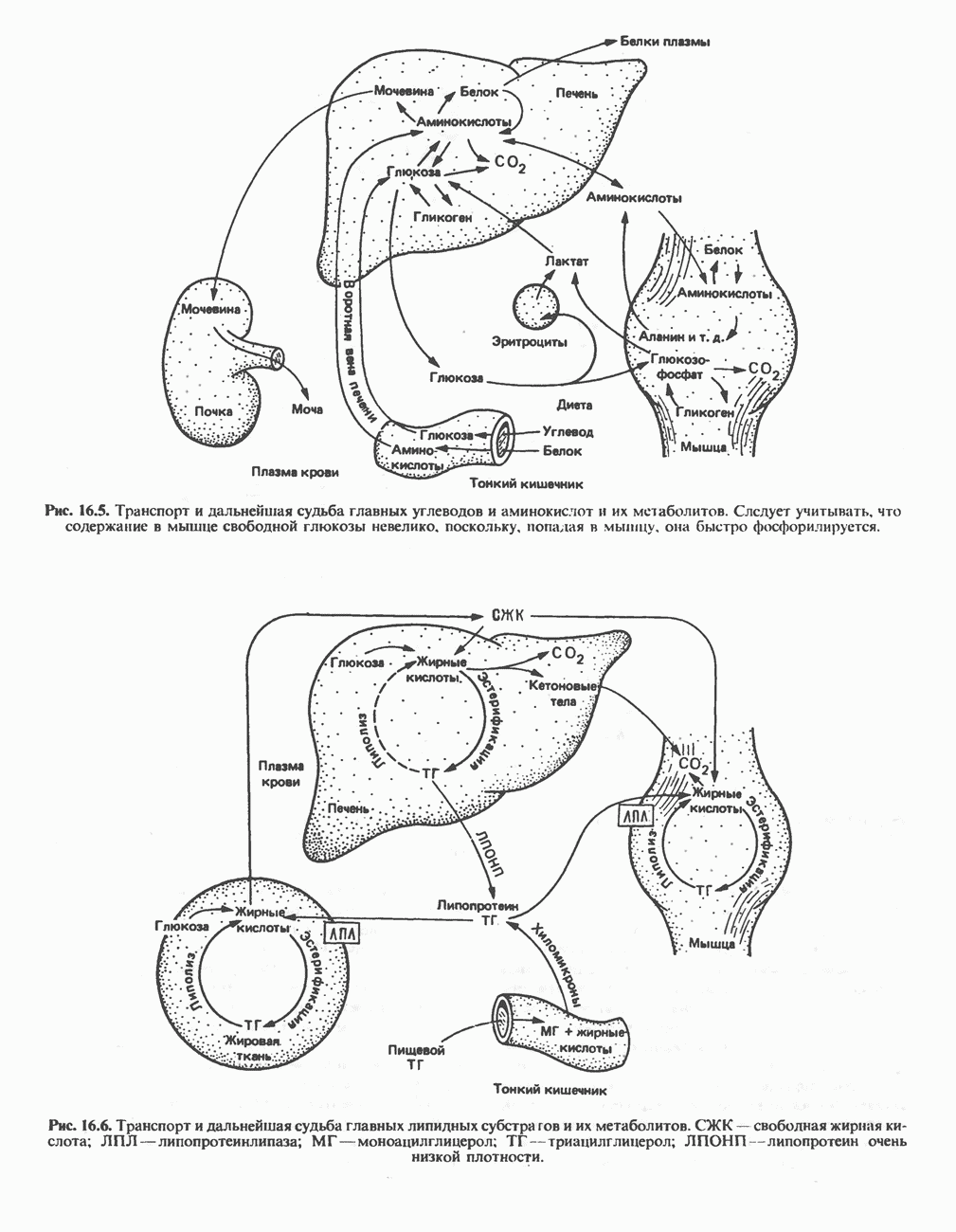
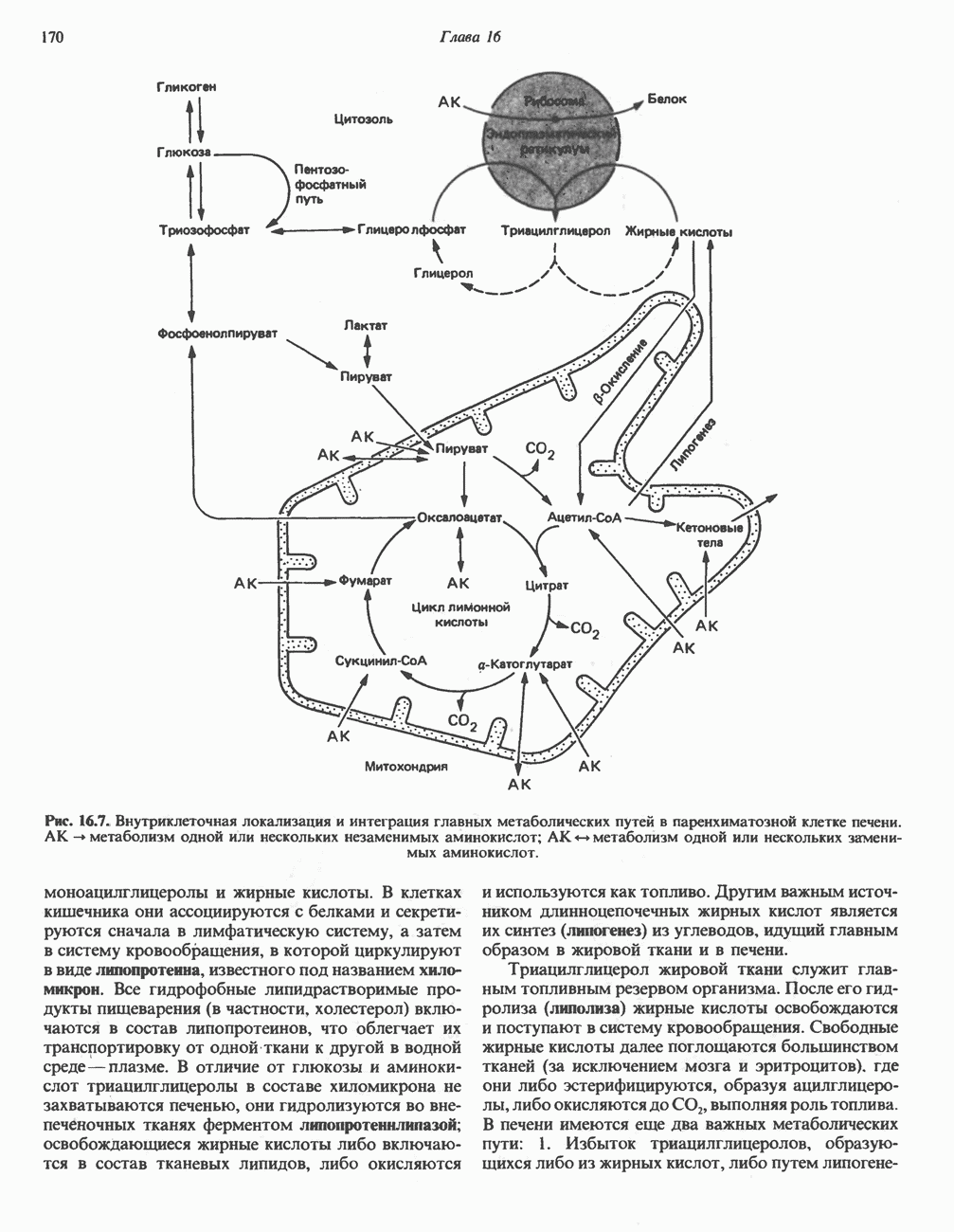
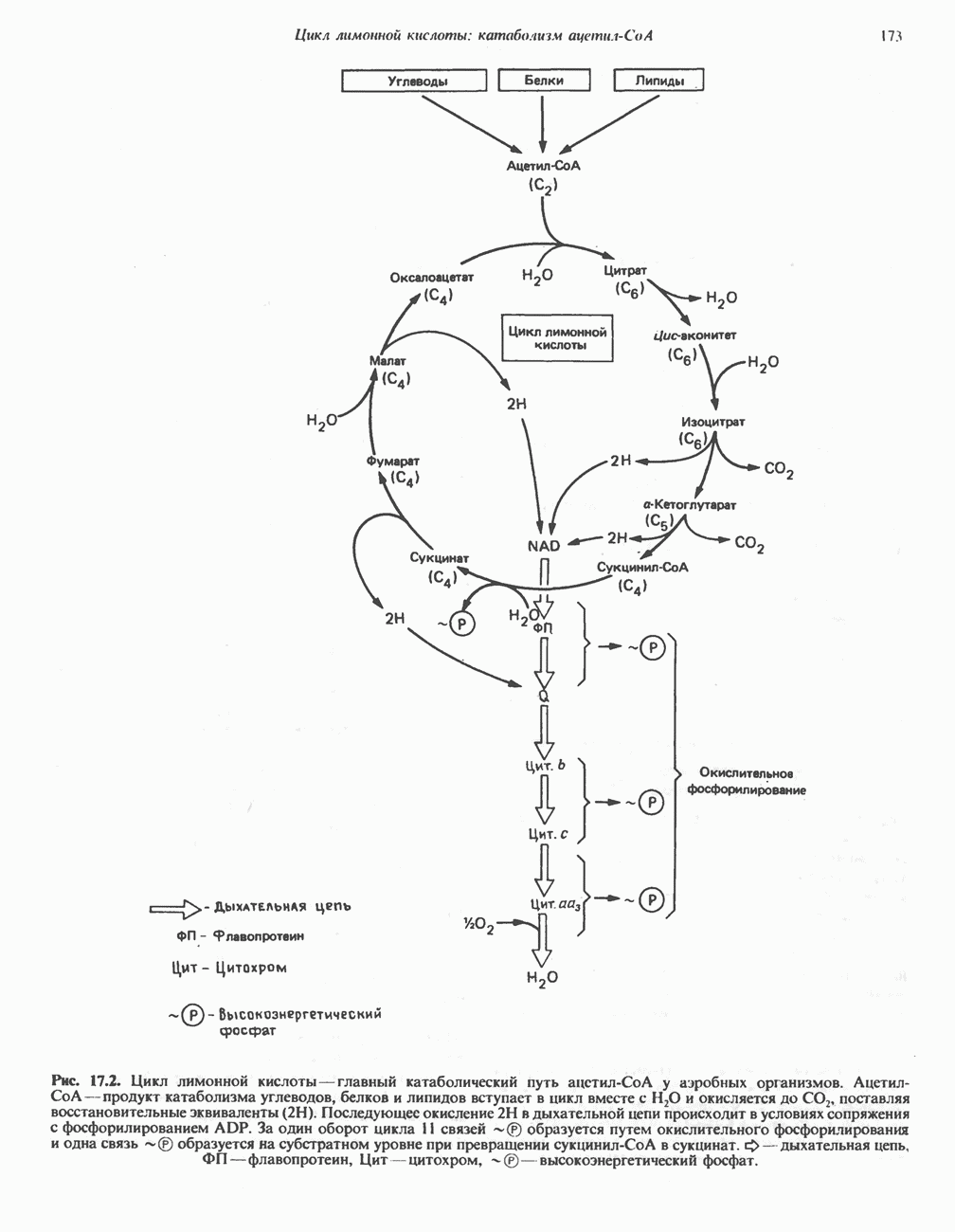
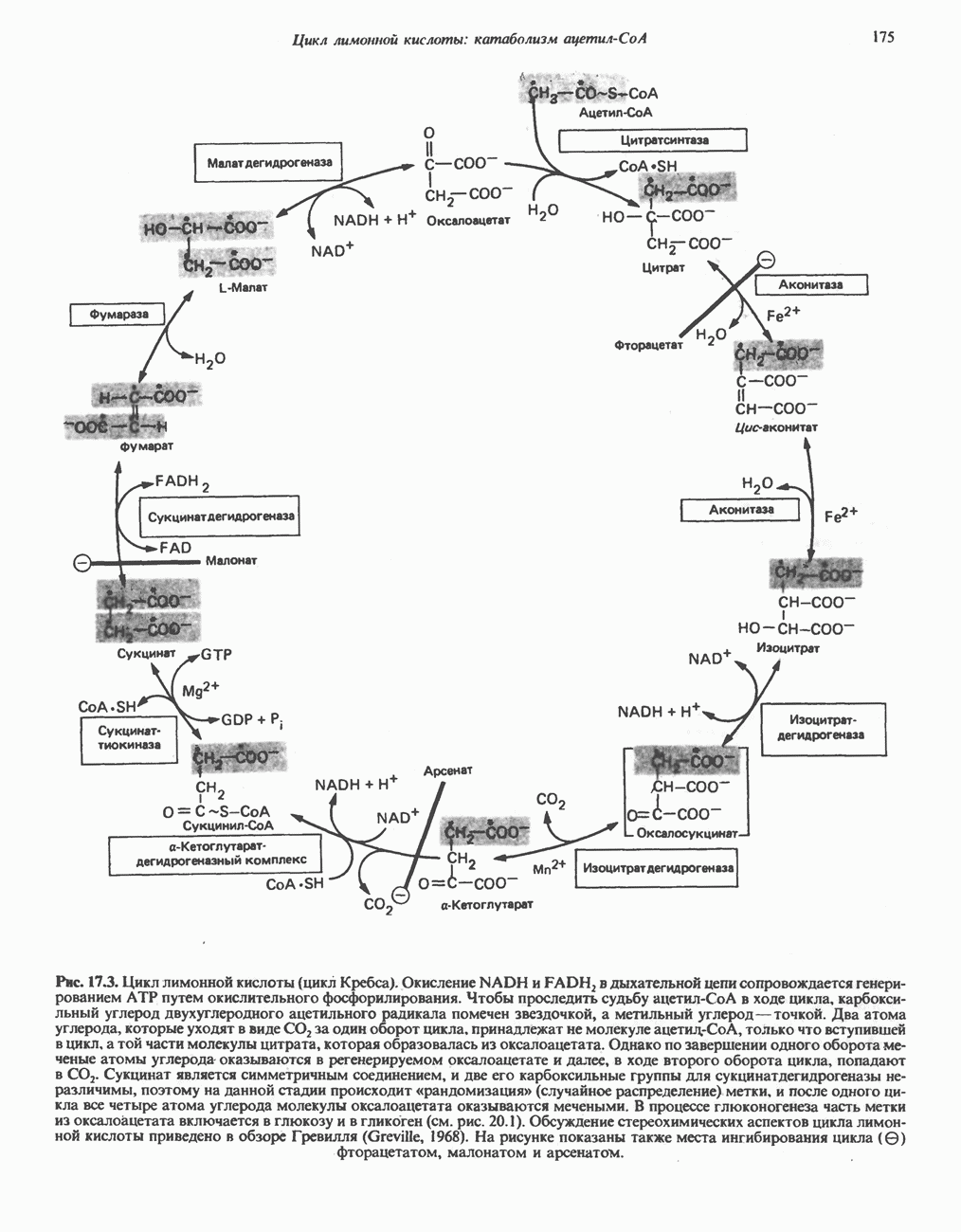
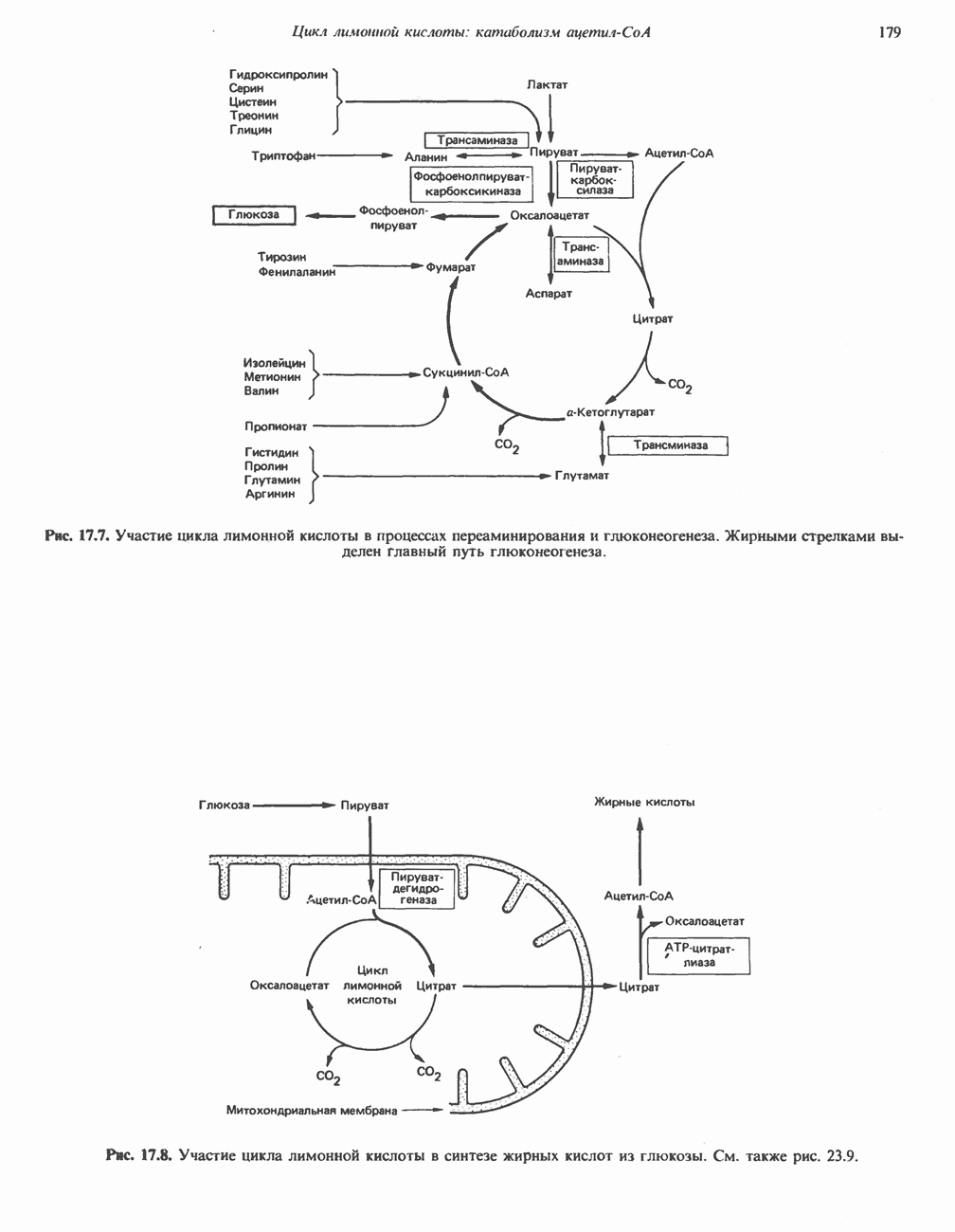
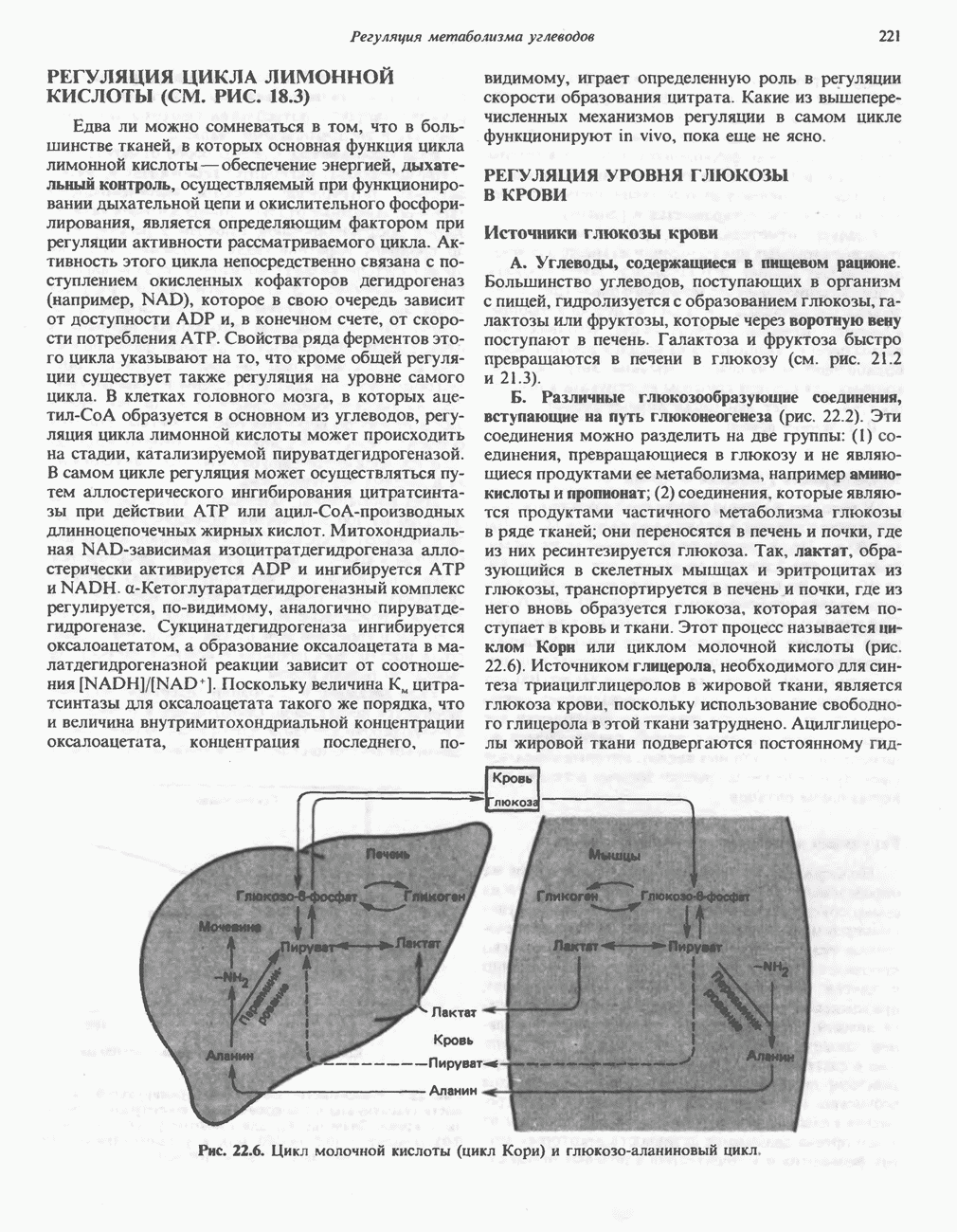
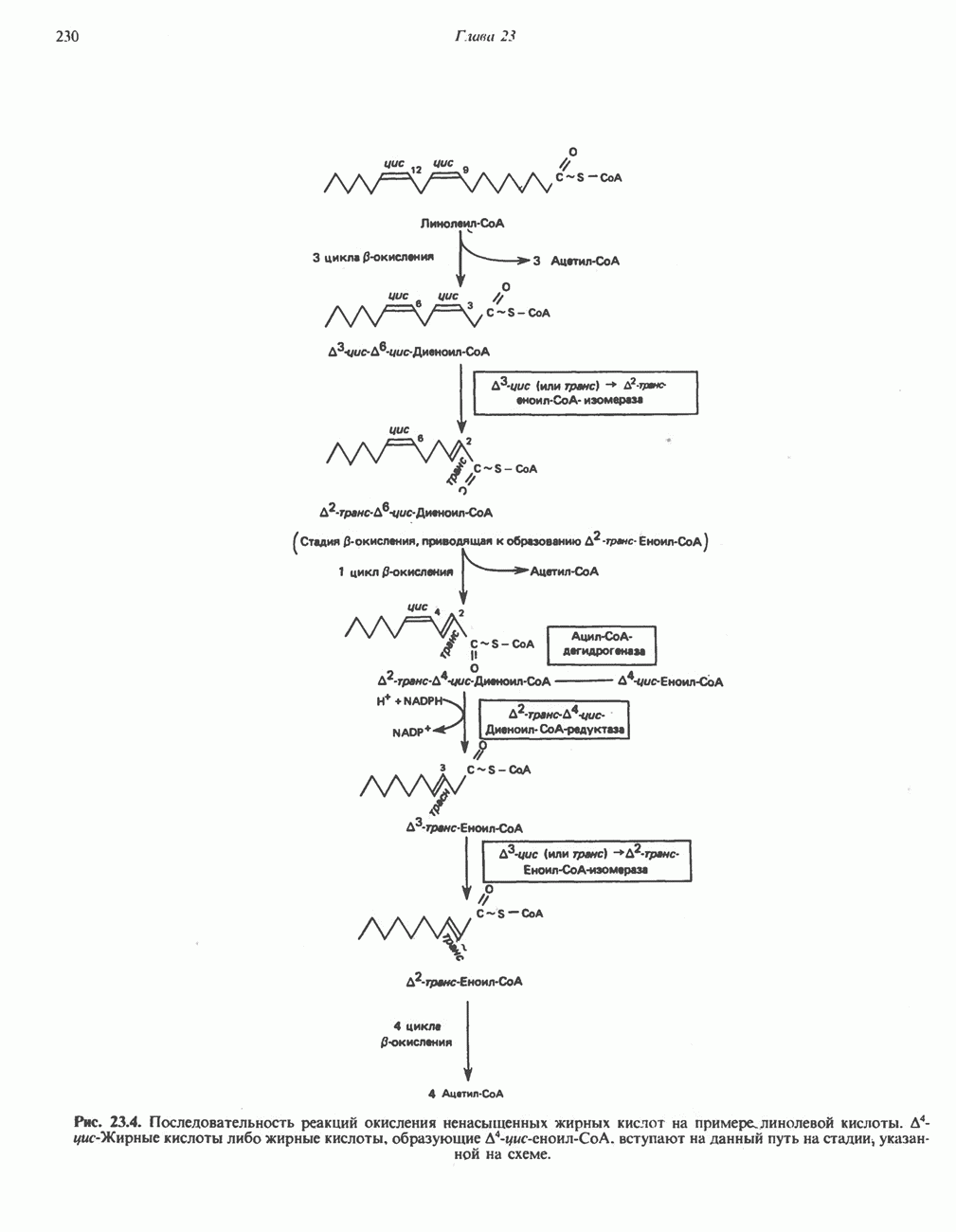
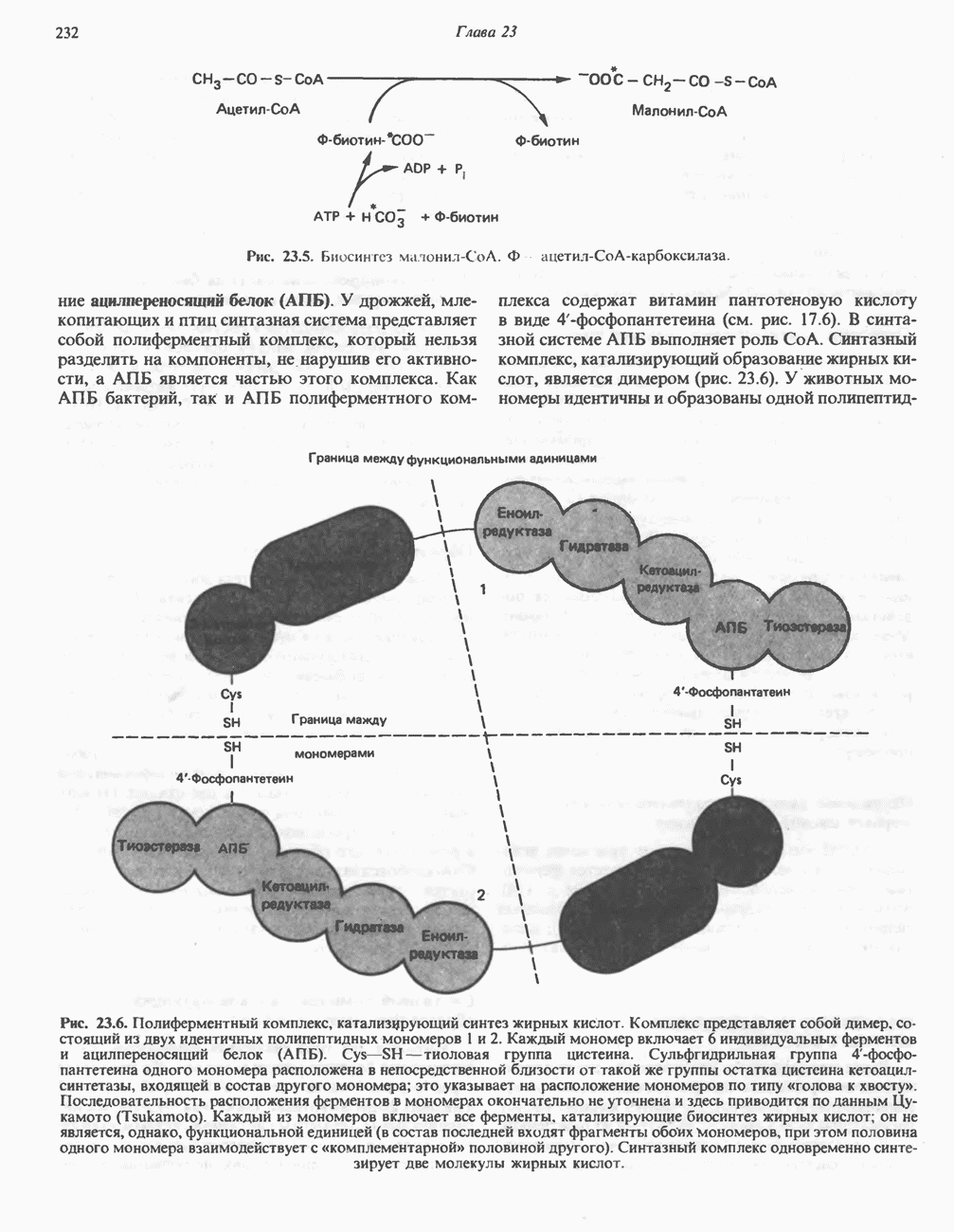
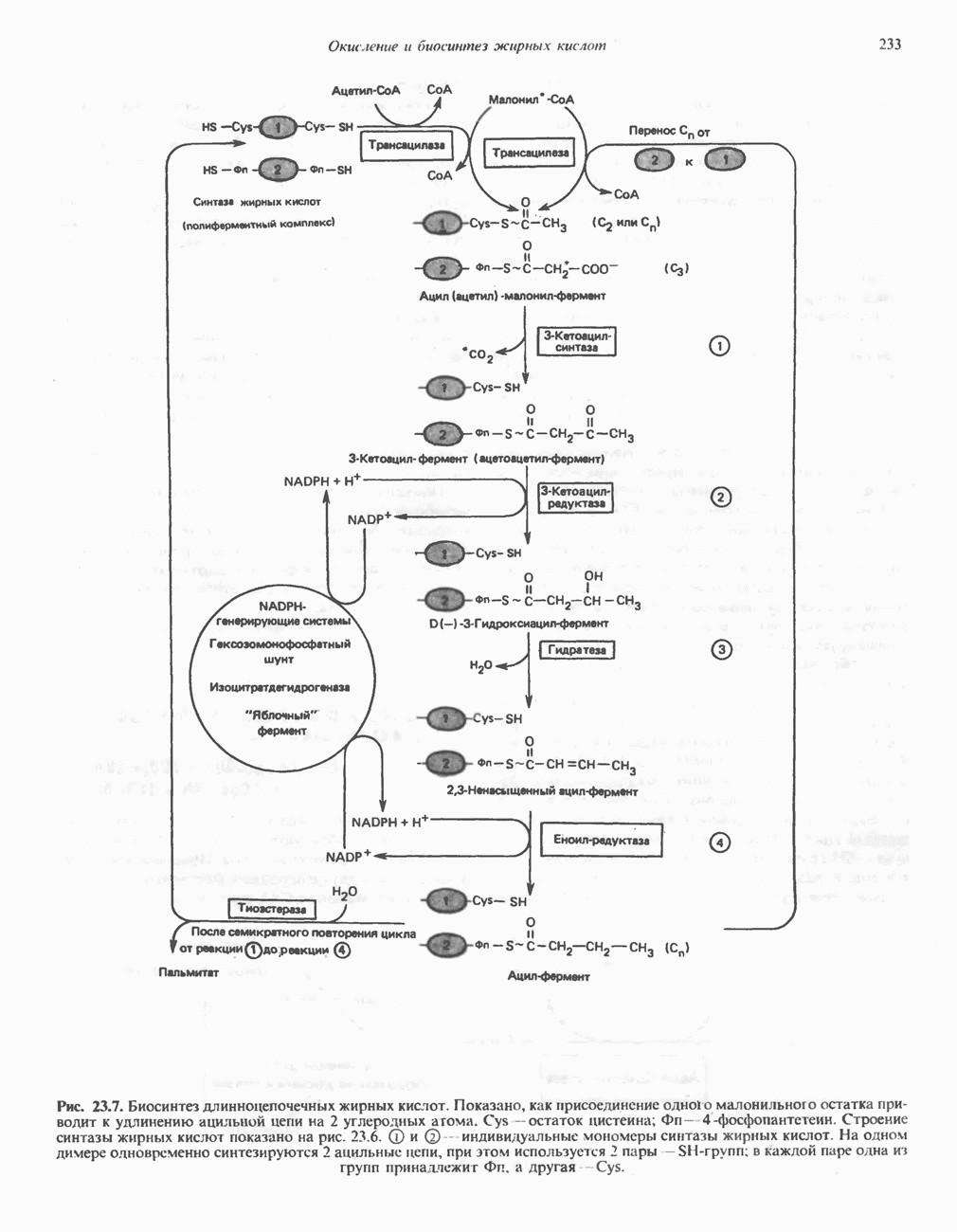
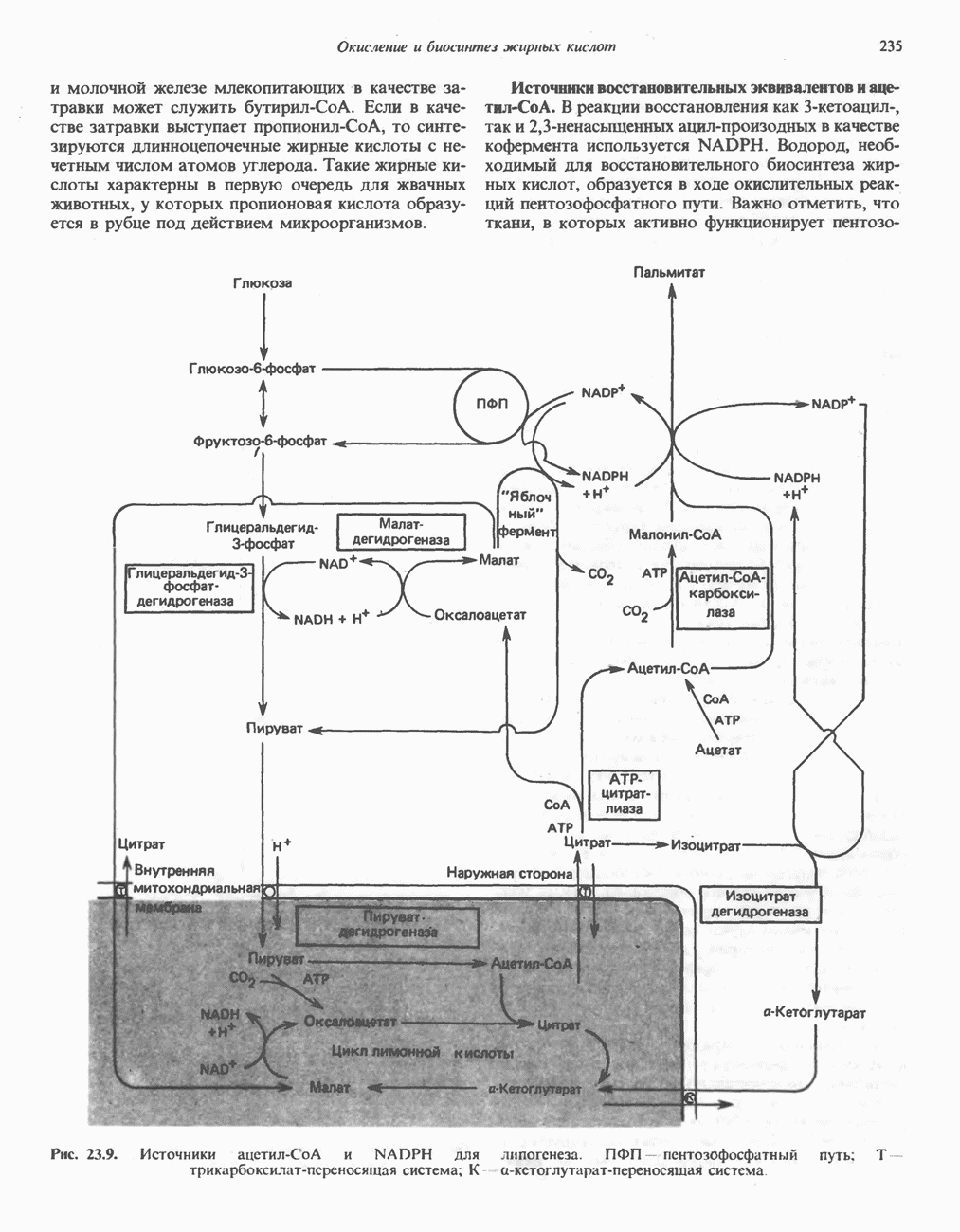
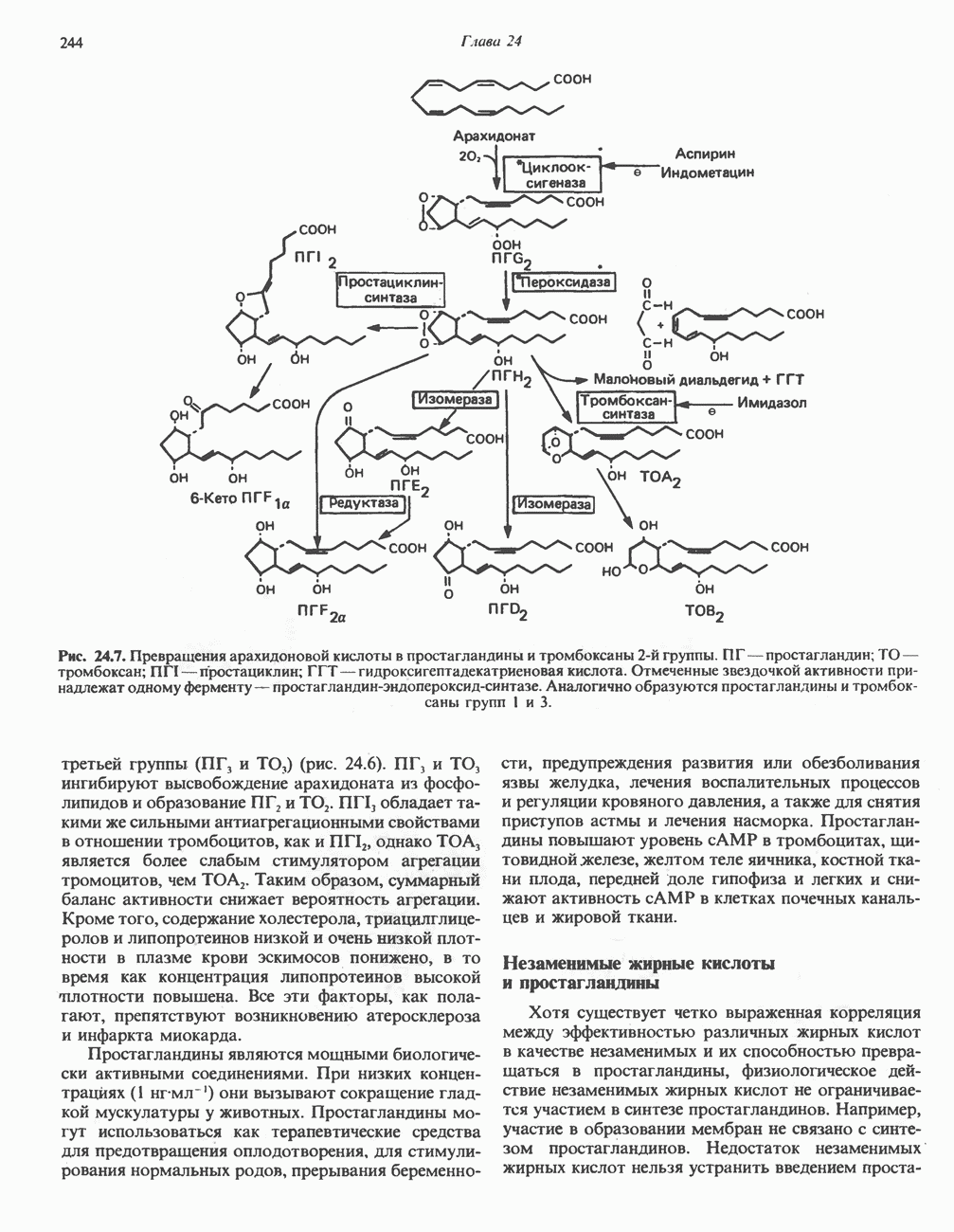
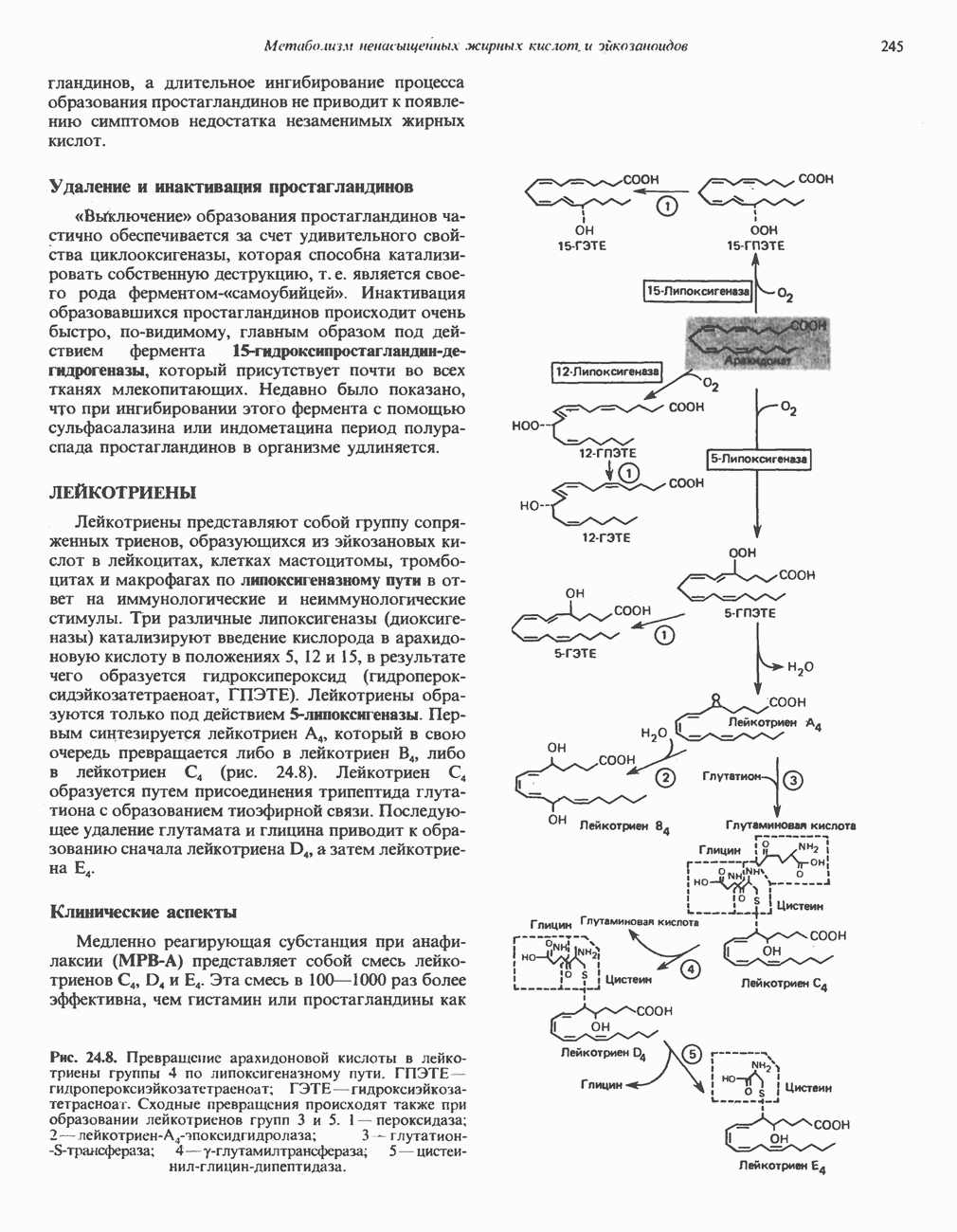
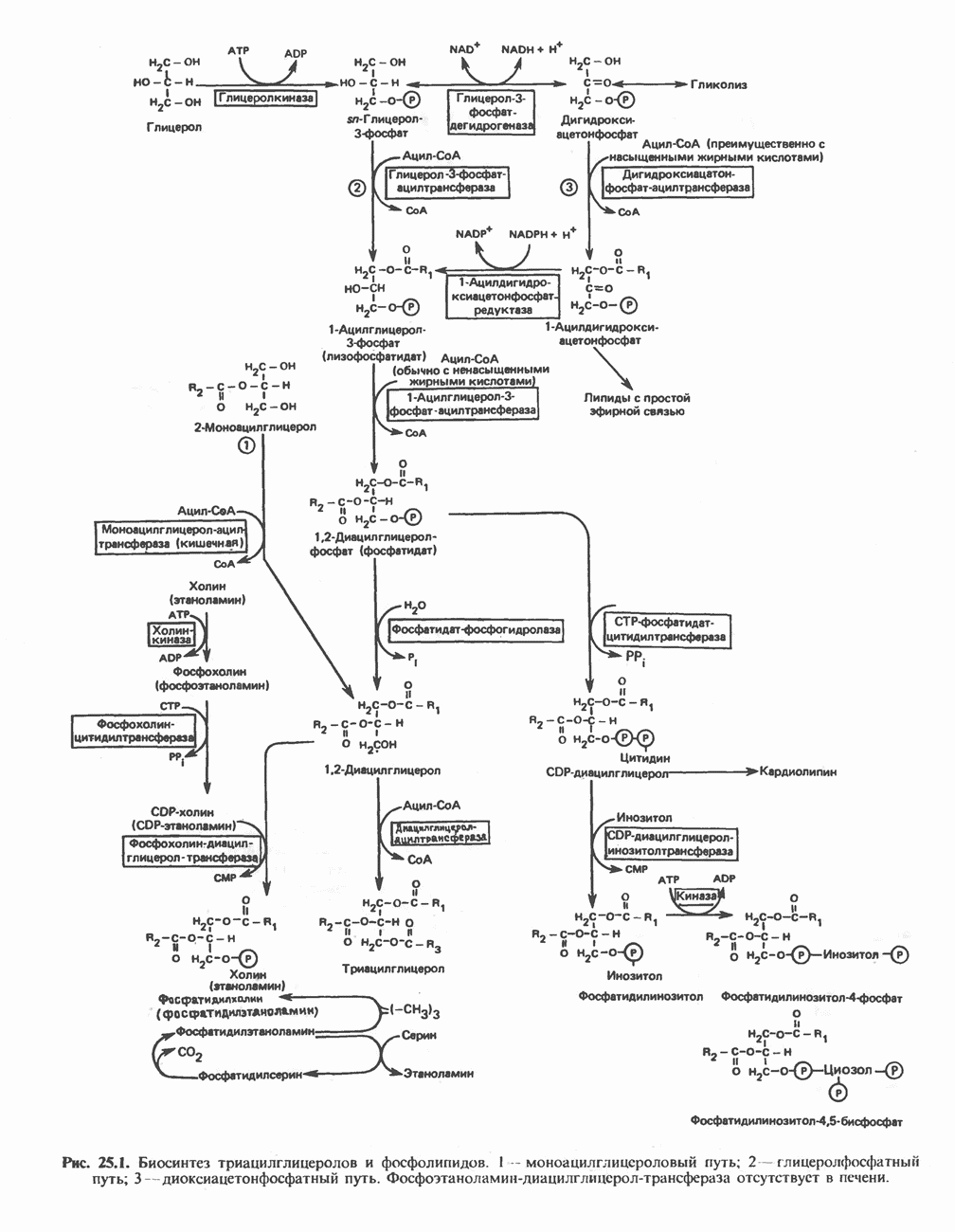
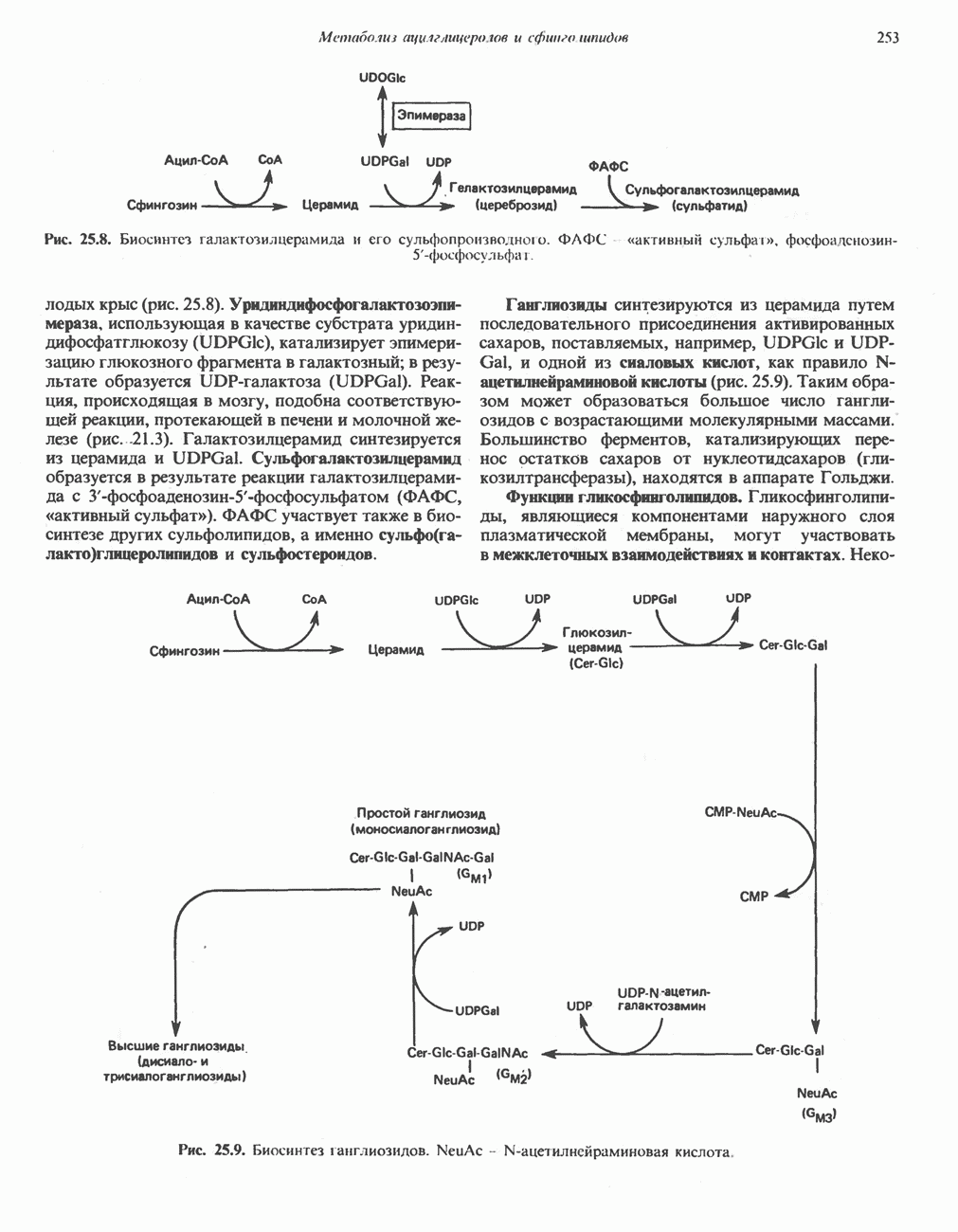
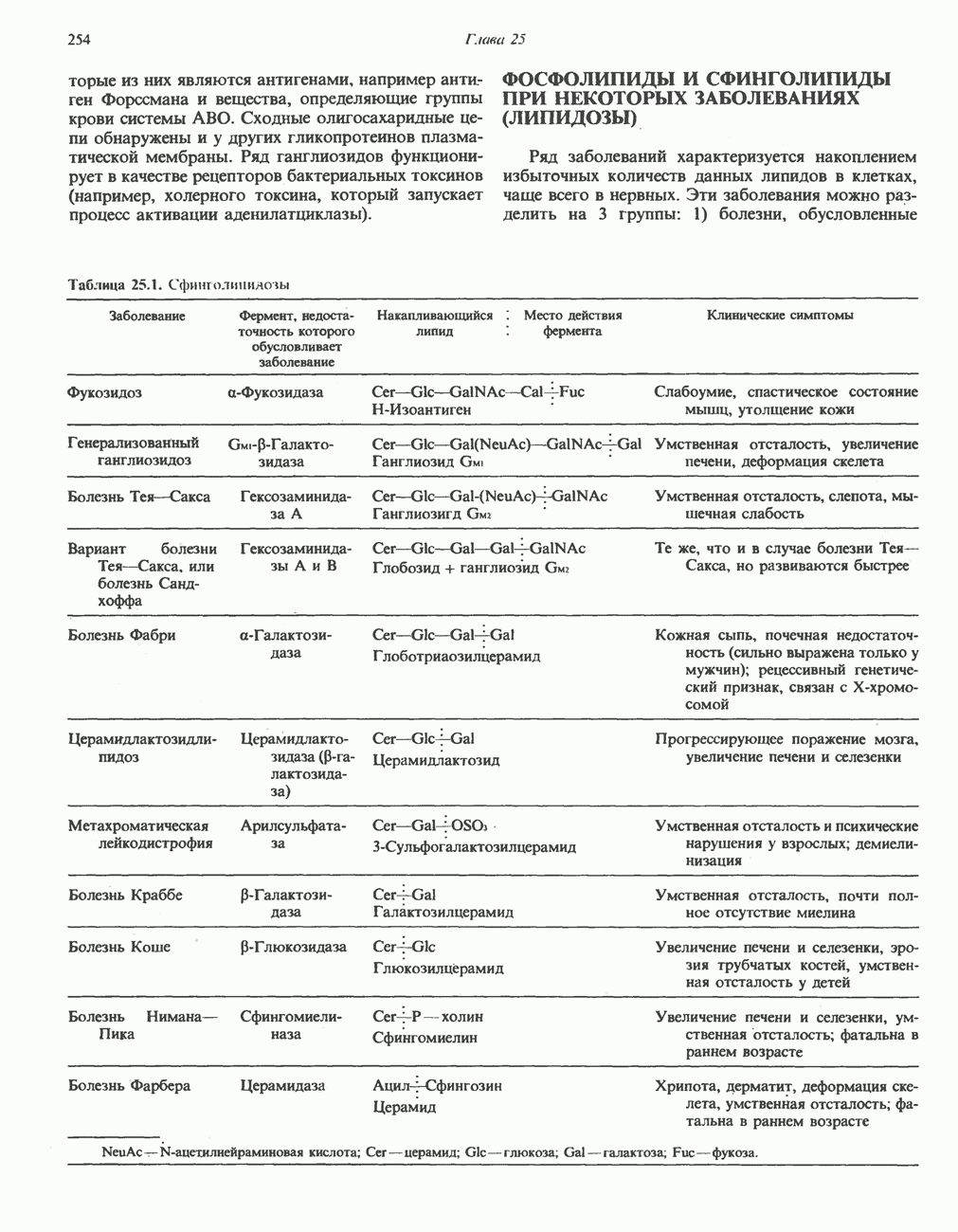
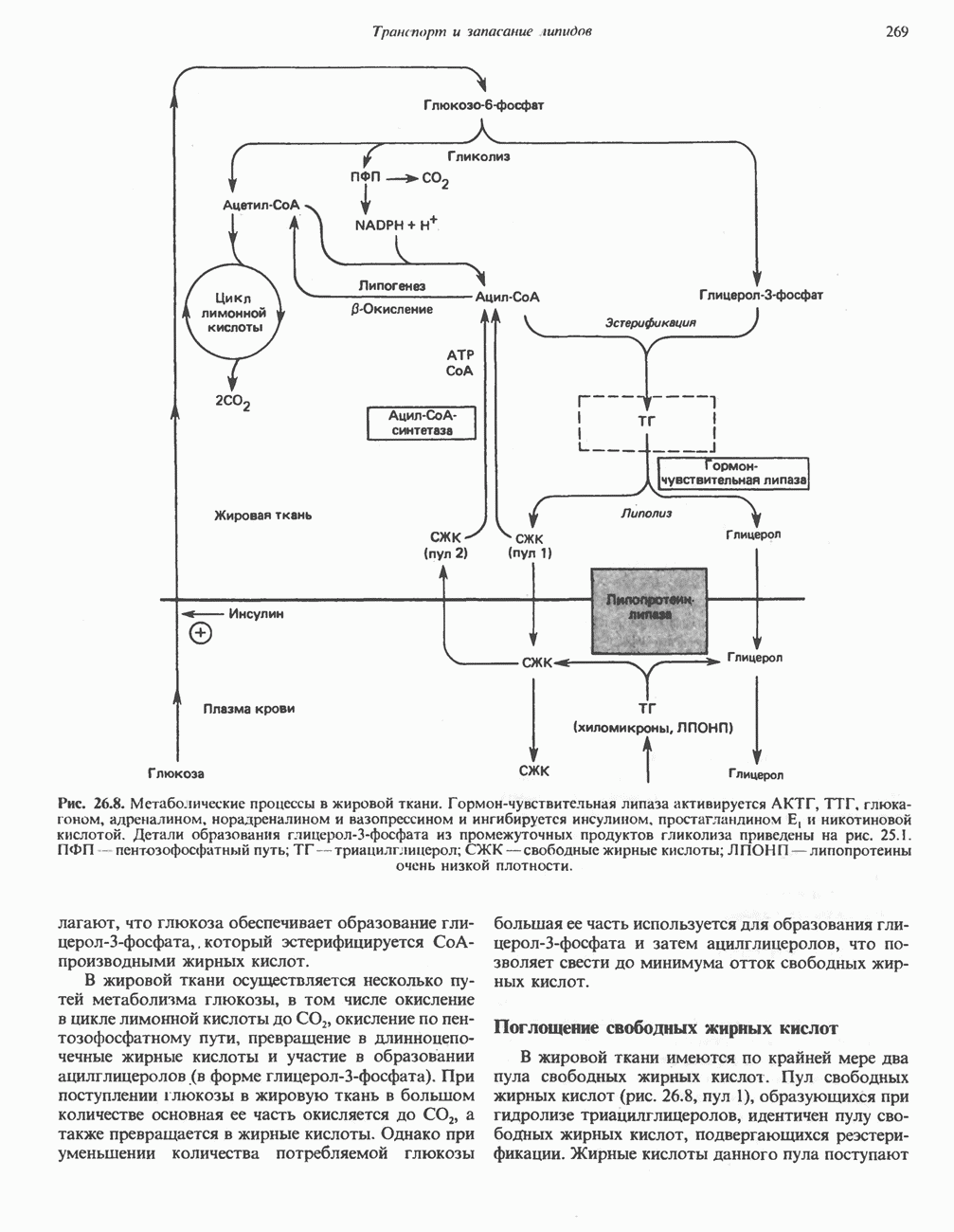
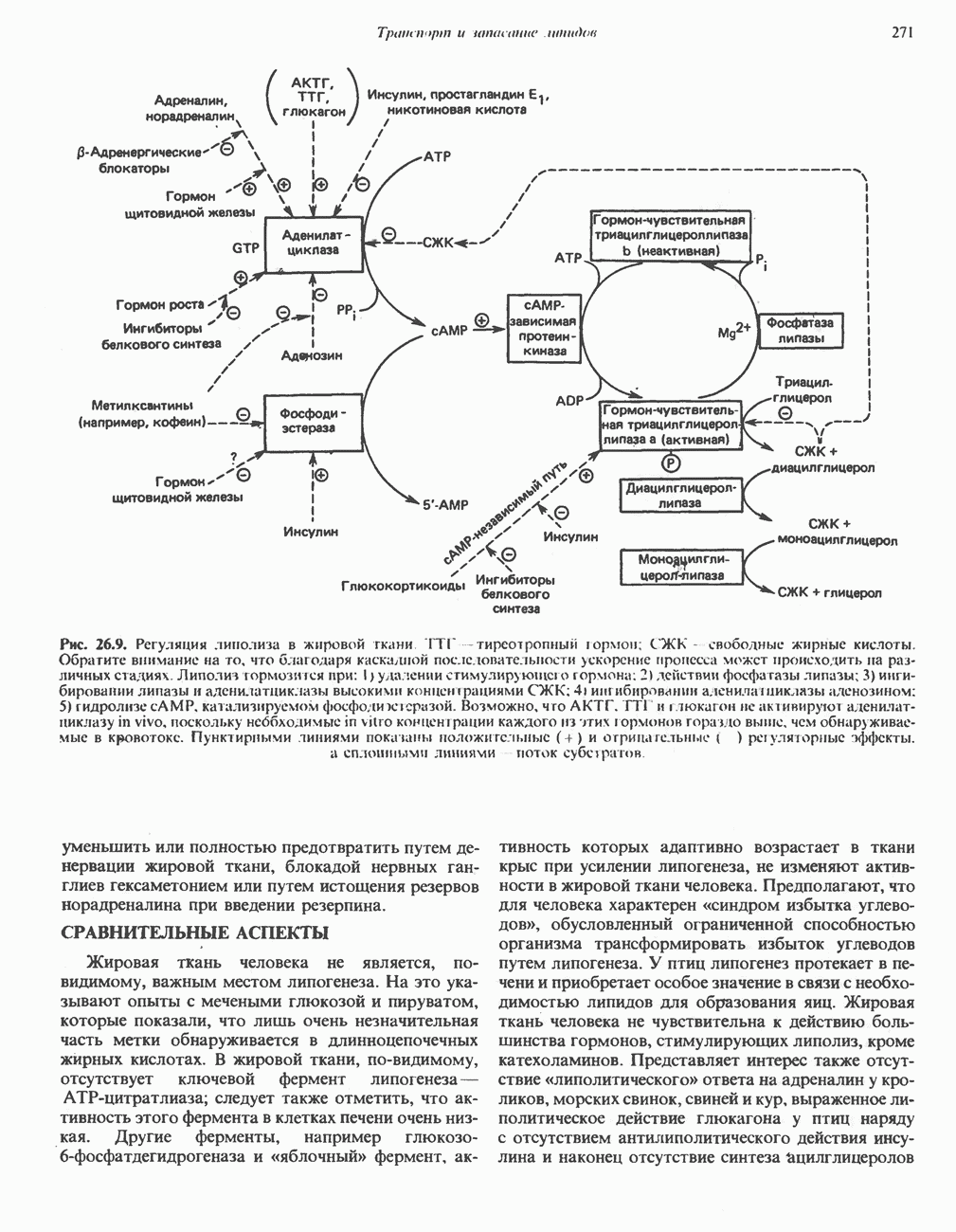
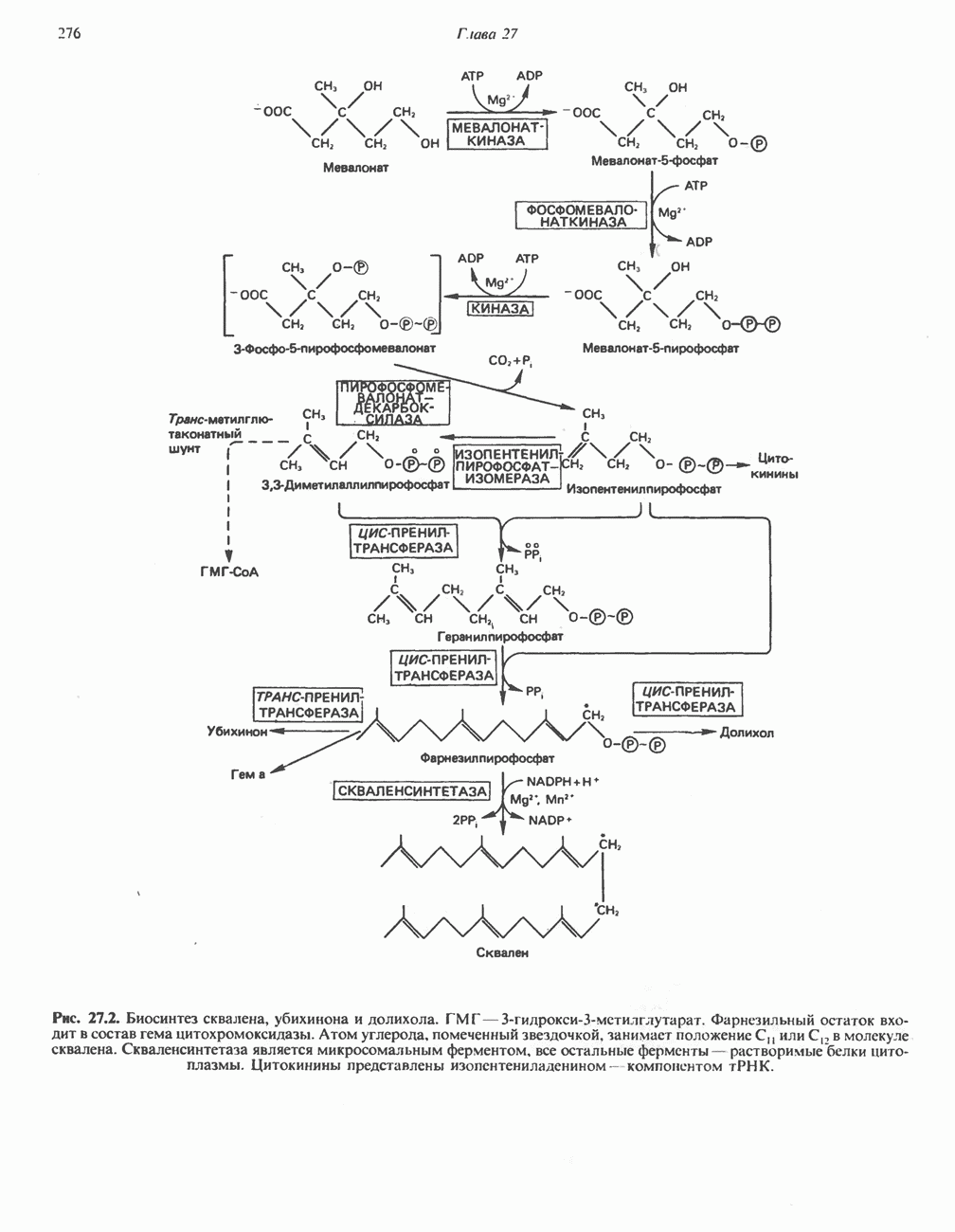
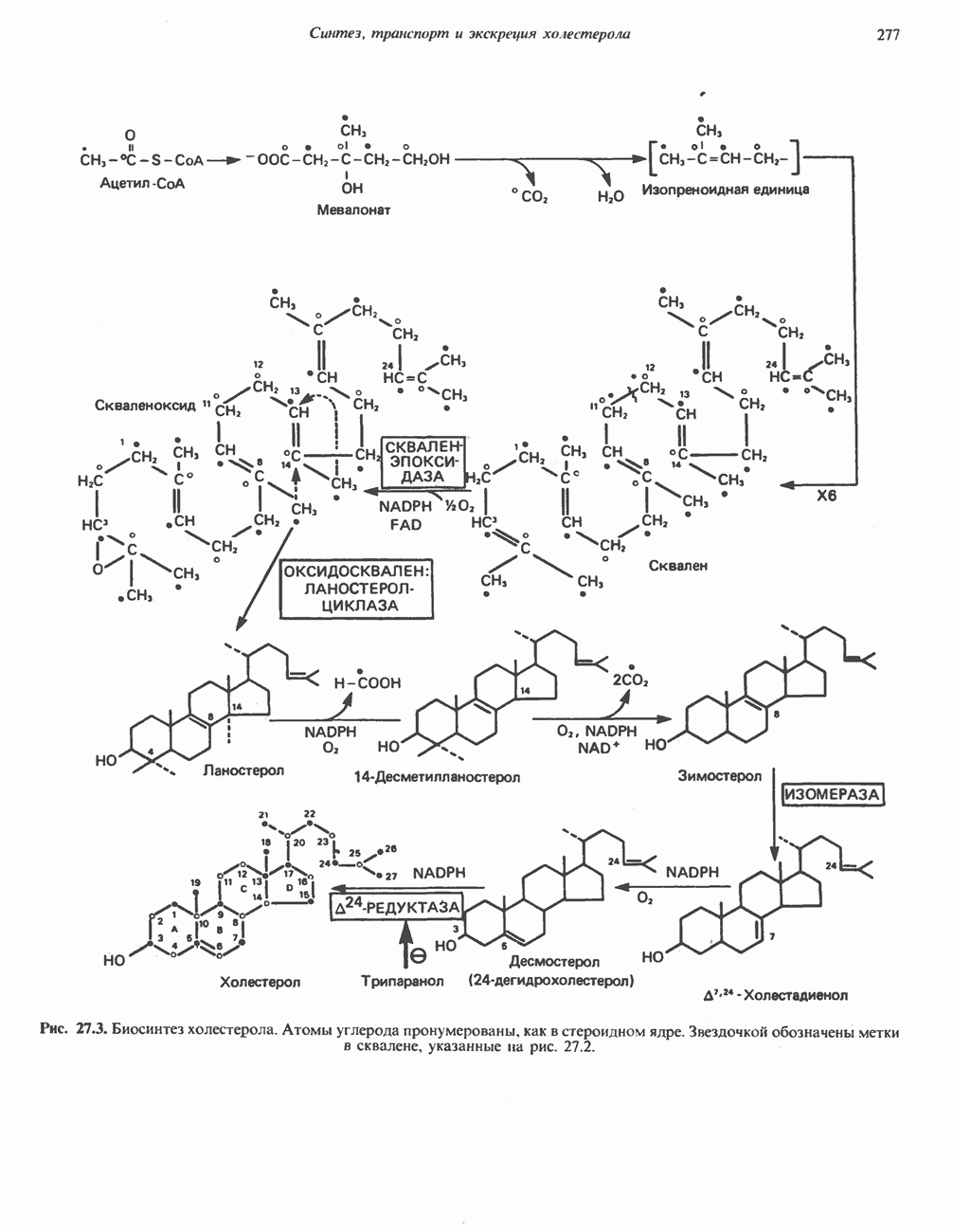
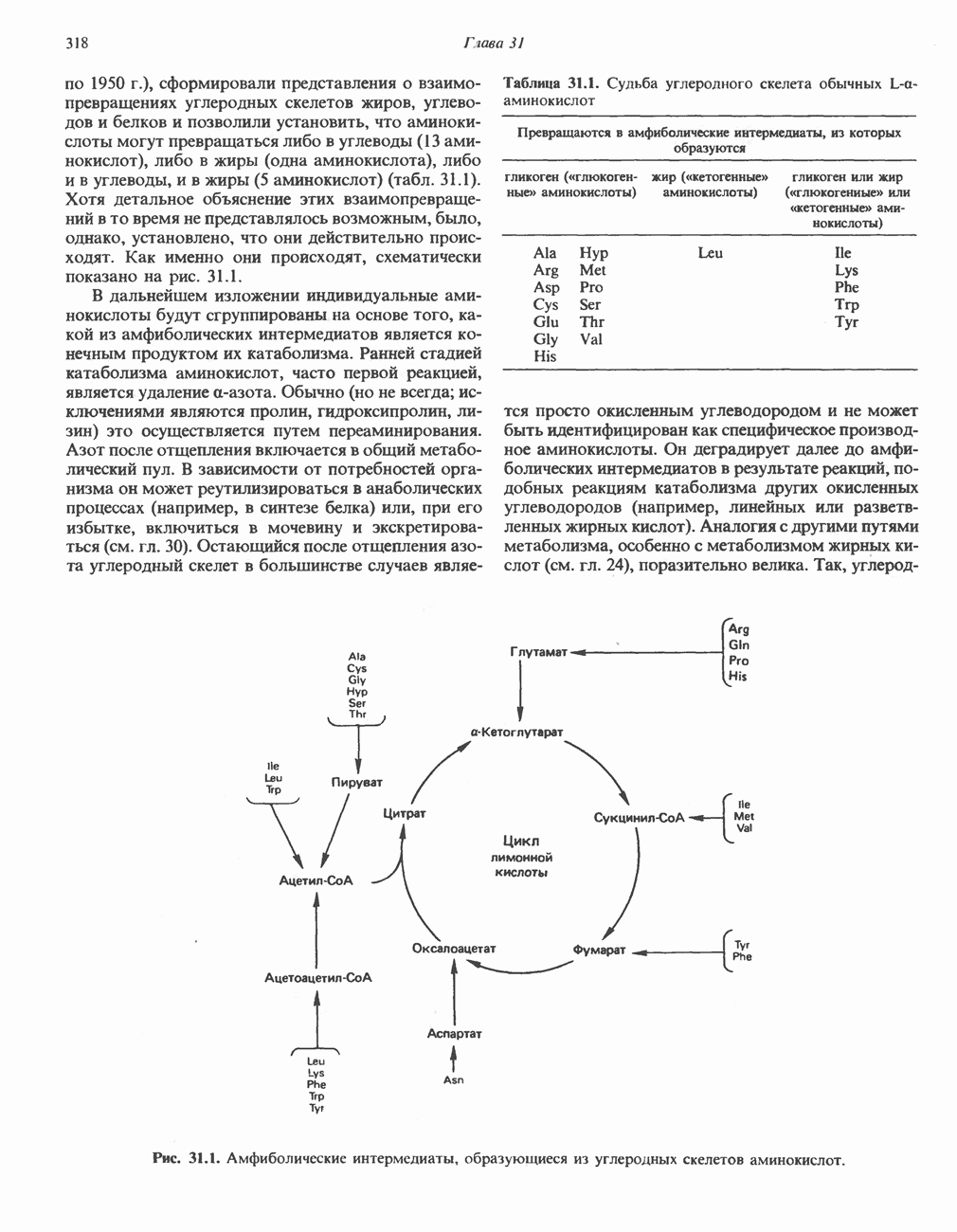
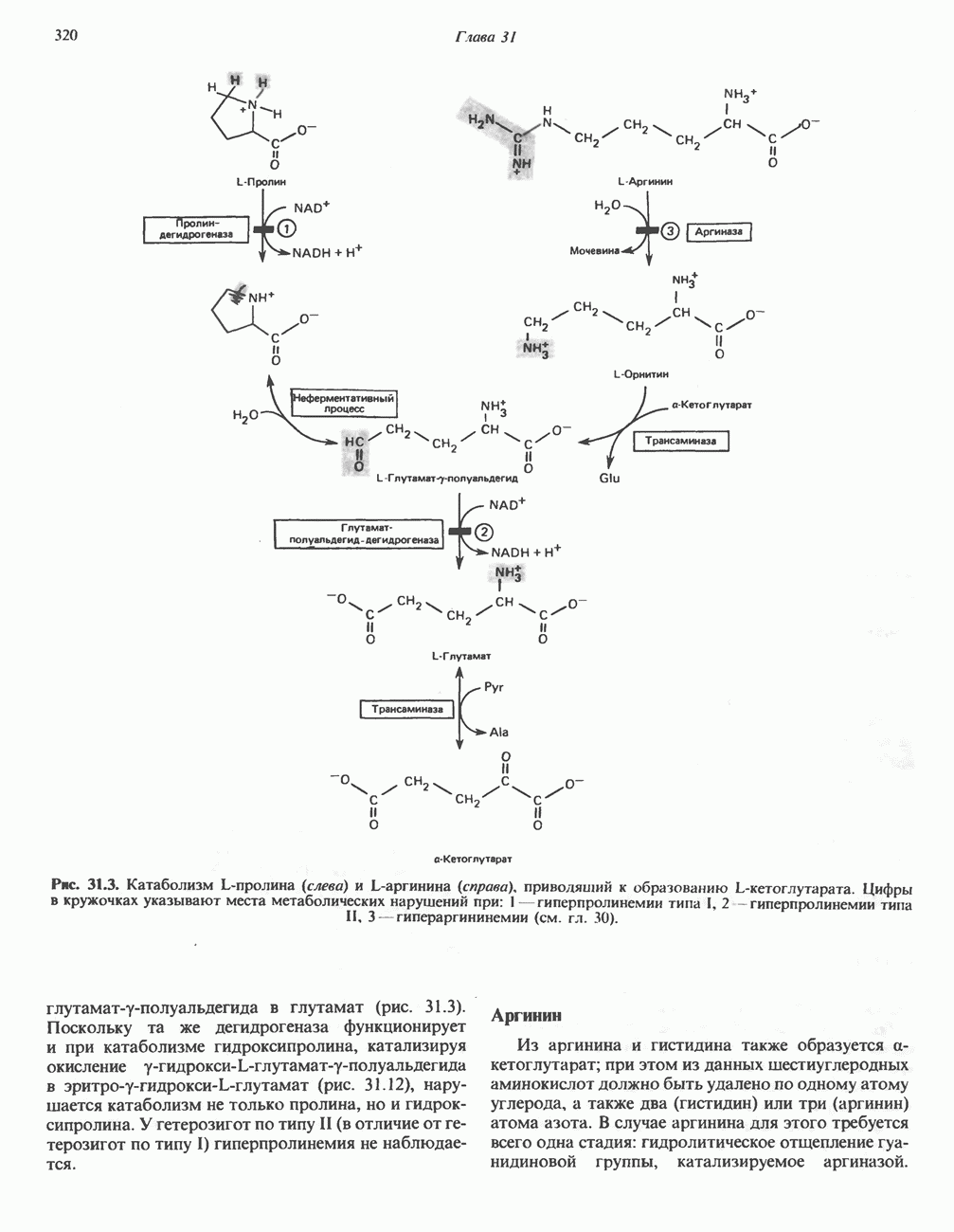
МЕТАБОЛИЗМ АЦИЛГЛИЦЕРОЛОВКатаболизм триацилглицероловПуть катаболизма триацилглицеролов начинается с их гидролиза до жирных кислот и глицерола под действием липазы; в основном этот процесс происходит в жировой ткани. Высвободившиеся жирные кислоты поступают в плазму крови, где связываются сывороточным альбумином. Затем свободные жирные кислоты переходят в ткани, где они либо окисляются, либо вновь подвергаются эстерификации. Ткани многих органов (печени, сердца, почек, мышц, легких, семенников, мозга), а также жировая ткань способны окислять длинноцепочечные жирные кислоты. Однако поступление этих кислот в клетки мозга затруднено. Что касается судьбы глицерола, то она зависит от того, присутствует ли в данной ткани необходимый активирующий фермент — глнцеролкиназа (рис. 25.1). Значительное количество этого фермента обнаружено в печени, почках, кишечнике, бурой жировой ткани и в молочных железах в период лактации. Биосинтез ацилглицероловХотя в лабораторных условиях можно осуществить реакции, обратные реакциям расщепления триацилглицеролов, в организме биосинтез ацилглицеролов протекает иным путем. Перед образованием ацилглицеролов глицерол и жирные кислоты должны быть активированы при участии АТР. Глнцеролкиназа катализирует фосфорилирование глицерола, в результате образуется (см. скан) Рис. 25.1. Биосинтез триацилглицеролов и фосфолипидов. 1 — моноацилглицсроловый путь; 2— глицеролфосфатный путь; 3 — диоксиацетонфосфатный путь. Фосфоэтаноламин-диацилглицсрол-трансфераза отсутствует в печени. или при его низкой активности, как это имеет место в мышцах и жировой ткани, большая часть глицерол-3-фосфата должна образовываться из промежуточного соединения гликолитического пути — дигидроксиацетонфосфата, восстановление которого за счет NADH до глицерол-3-фосфата катализируется глицерол-3-фосфатдегидрогена зон (рис. 25.1). А. Триацилглицерол. Жирные кислоты активируются в результате взаимодействия с СоА с образованием ацил-СоА; реакция катализируется ацил-СоА-синтетазой и протекает с участием АТР. Две молекулы ацил-СоА взаимодействуют с глицерол-3-фосфатом, в результате образуется 1,2-диацилглицеролфосфат (фосфатидат). Этот процесс протекает в две стадии. Сначала глицерол-3-фосфат-ацилтрансфераза катализирует образование лизофосфатидата, а затем 1 -ацилглицерол-3-фосфат-ацилтрансфераза (лизофосфатидат-ацилтрансфераза) катализирует образование фосфати-дата. Далее фосфатидат гидролизуется фосфатидат—фосфогндролазой до Дигидроксиацетонфосфат может ацилироваться и превращаться в лизофосфатидат путем востановления с участием NADPH. Относительно значения этого пути еще нет единого мнения. По-видимому, он играет важную роль в пероксисомах, где участвует в биосинтезе липидов с простой эфирной связью. Б. Фосфоглицеролы. Эти фосфолипиды образуются либо из фосфатидата (например, фосфатидилинозитол), либо из 1,2-диацилглицерола (например, фосфатидилхолин и фосфатидилэтаноламин). При синтезе фосфатидилинозитола цитидинтрифосфат (СТР) взаимодействует с фосфатидатом с образованием цитидиндифосфатдиацилглицерола (CDP-диацилглицерол), который при участии фермента CDP-днацилглицерол—инозитолтрансферазы реагирует с инозитолом, в результате чего образуется фо-сфатидилинозитол (рис. 25.1). Последовательное фосфорилирование фосфатидилинозитола приводит к образованию сначала фосфатидилинозито)1-4-фосфата, а затем фосфатидилинозитол-4,5-бисфосфата. Последний гидролизуется с образованием диацилглицерола и инозитолтрифосфата (процесс запускается гормонами, в частности вазопрессином, которые повышают концентрацию В процессе биосинтеза фосфатидилхолина и фосфатидилэтаноламина (лецитина и кефалина) холин и этаноламин должны сначала перейти в активную форму. На первой стадии процесса в результате реакции с АТР образуется соответствующий монофосфат, который затем реагирует с СТР, в результате чего образуется либо цитидиндифосфохолин ( Фосфатидилсерин синтезируется путем прямого взаимодействия фосфатидилэтаноламина и серина. Фосфатидилсерин может декарбоксилироваться, в результате образуется фосфатидилэтаноламин. В печени существует альтернативный путь, по которому фосфатидилхолин синтезируется из фосфатидилэтаноламина путем последовательного метилирования остатка этаноламина с участием S-аденозилметионина в качестве донора метильных групп. В митохондриях присутствует фосфолипид кардиолипин (дифосфатидилглицерол). Он образуется из фосфатидилглицерола, который в свою очередь синтезируется из CDP-диацилглицерола (рис. 25.1) и глицерол-3-фосфата, как показано на схеме, приведенной на рис. 25.2.
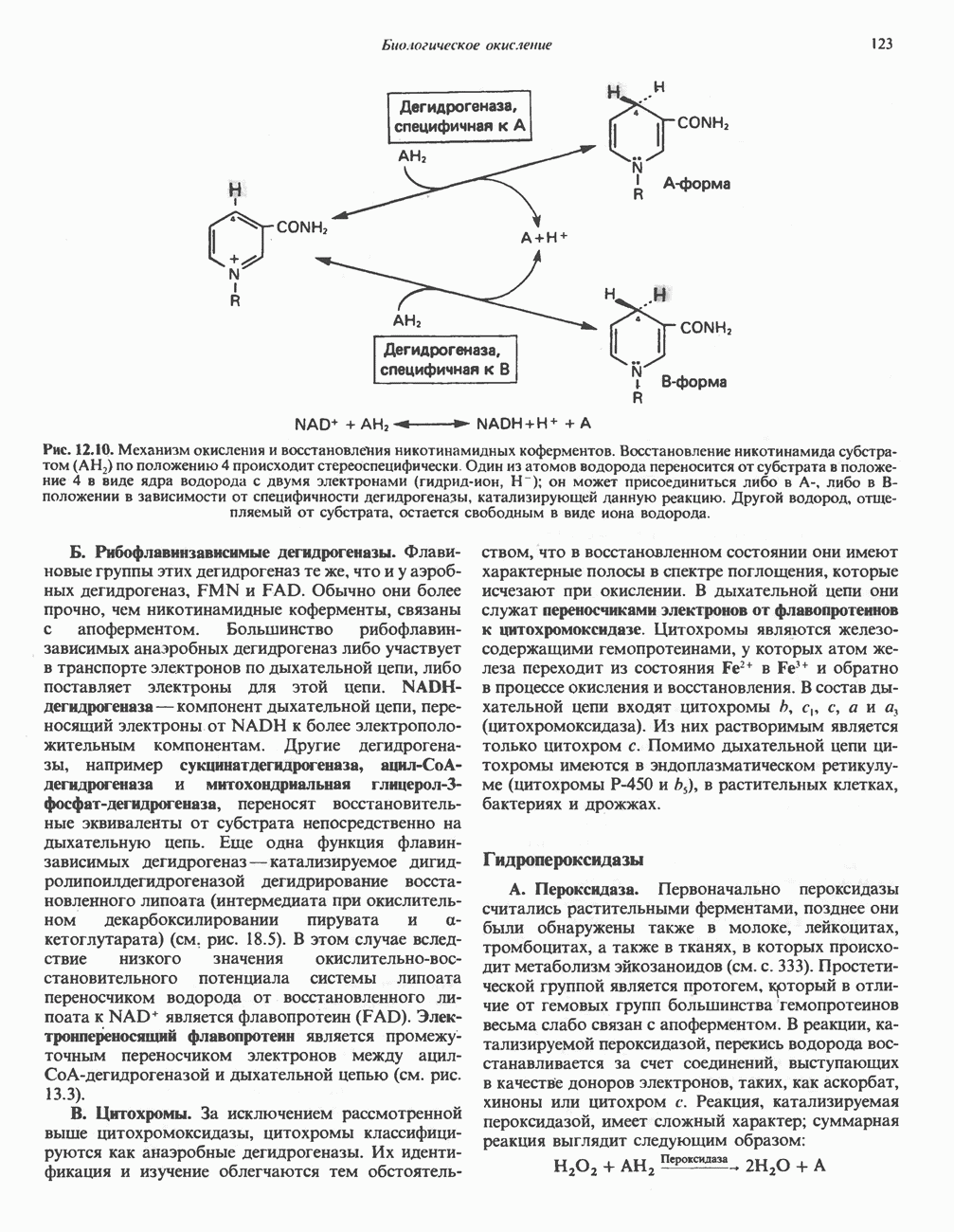
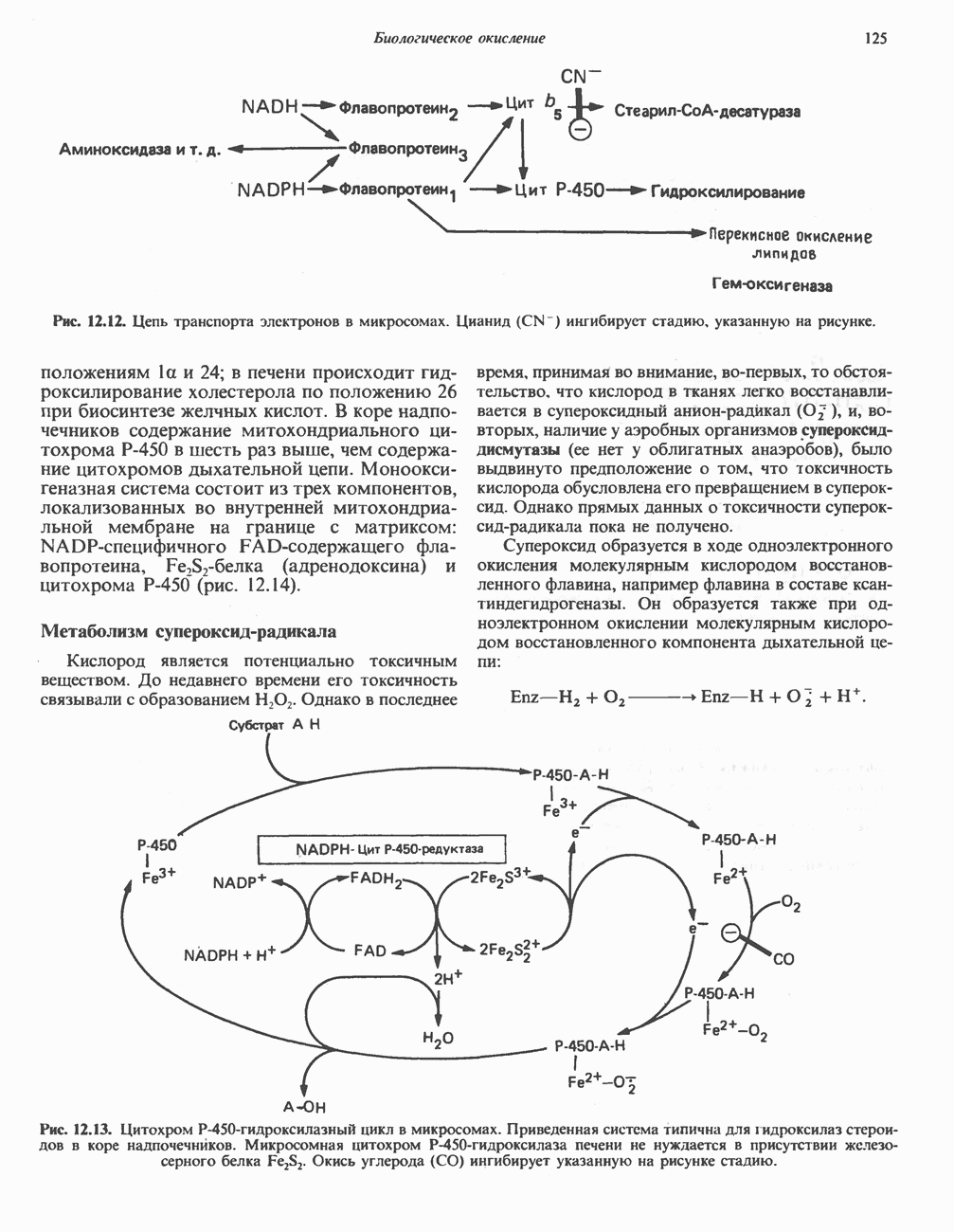
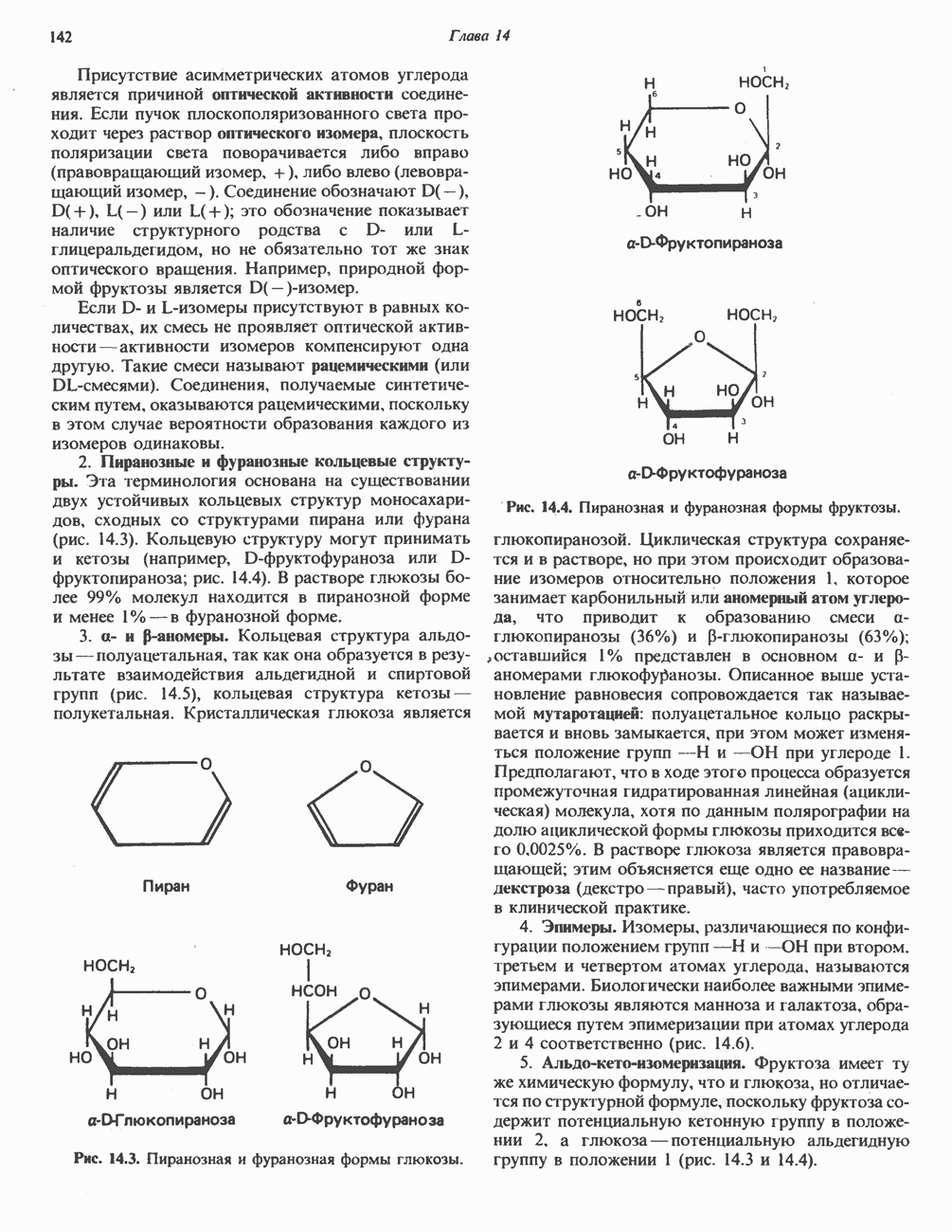
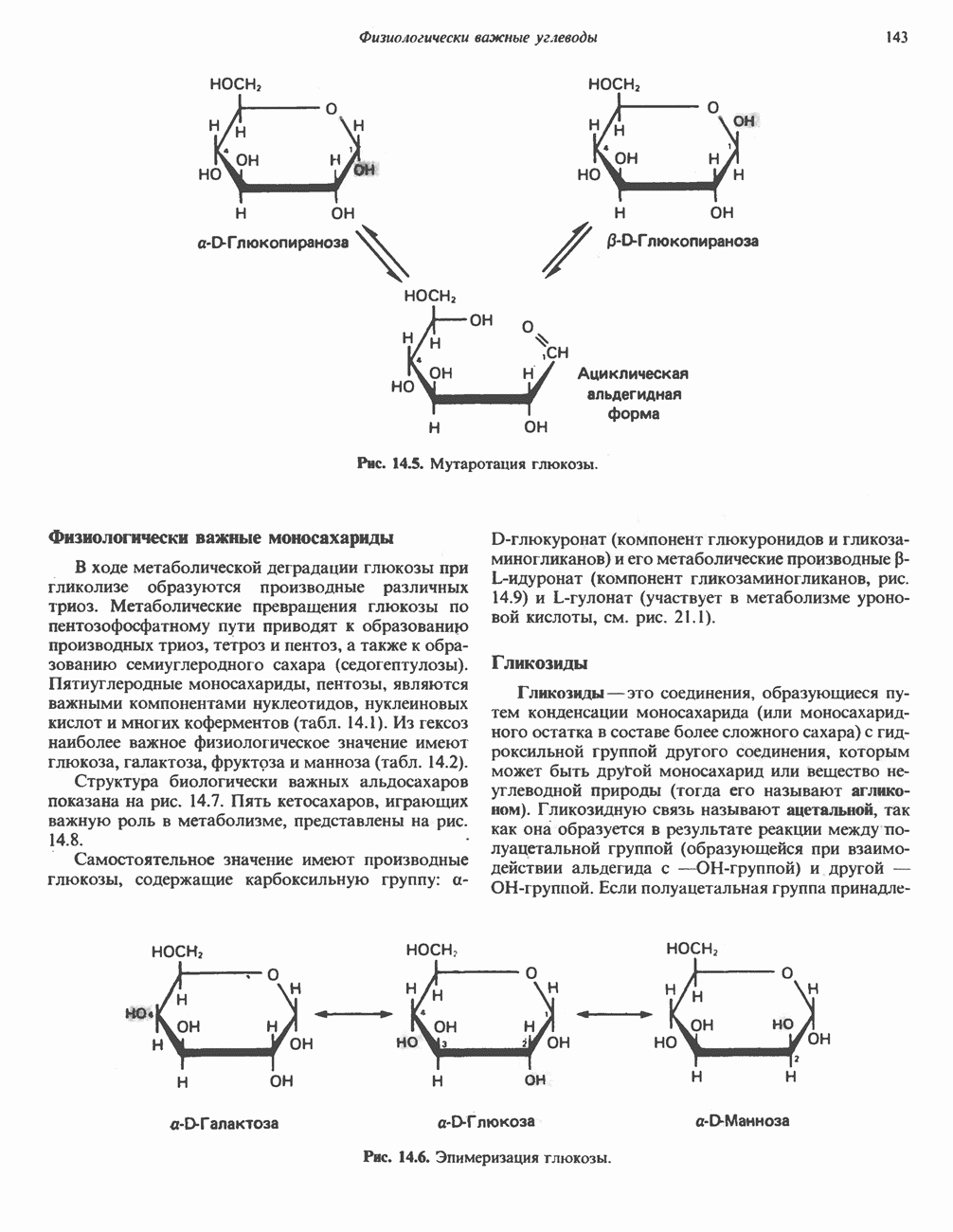
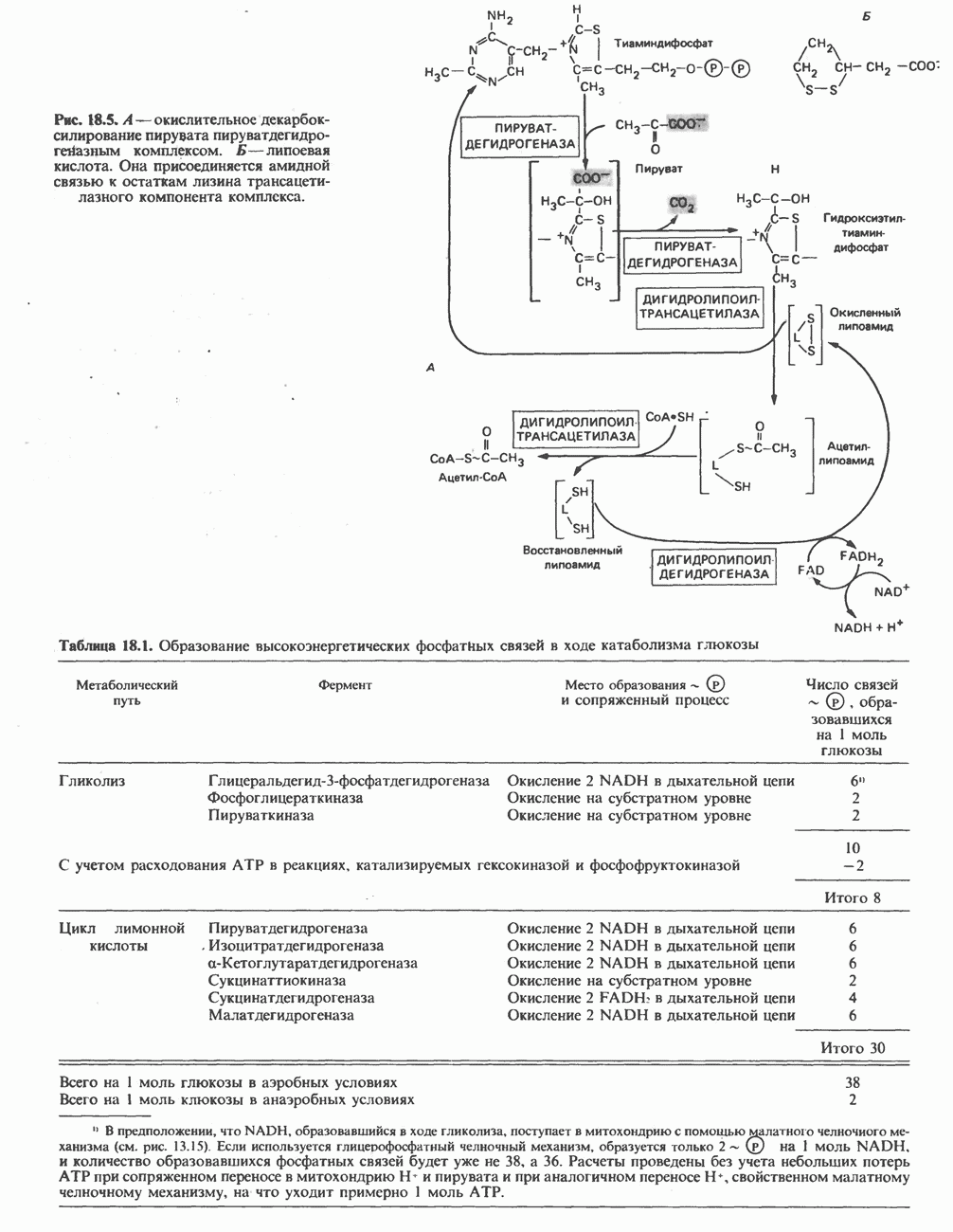
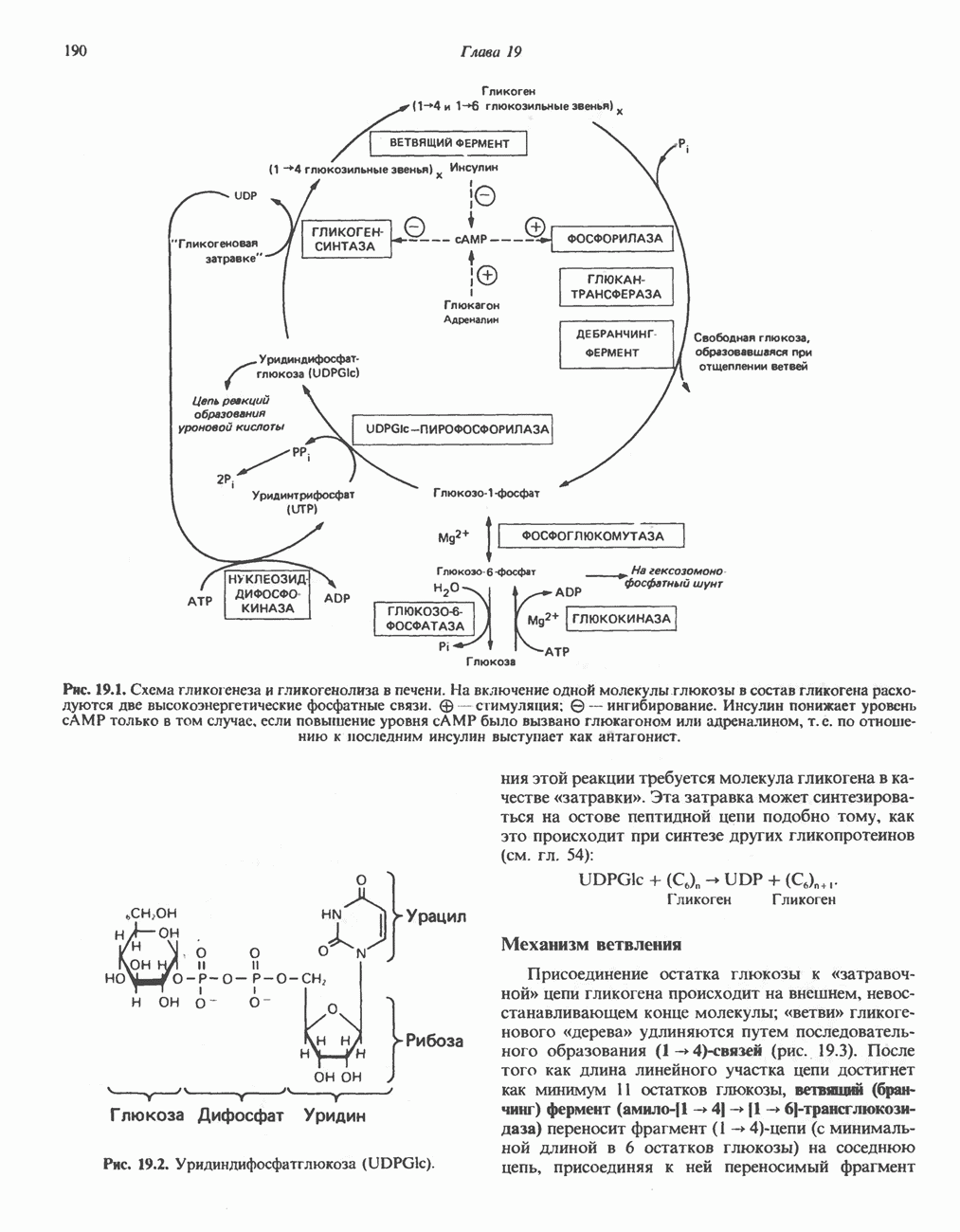
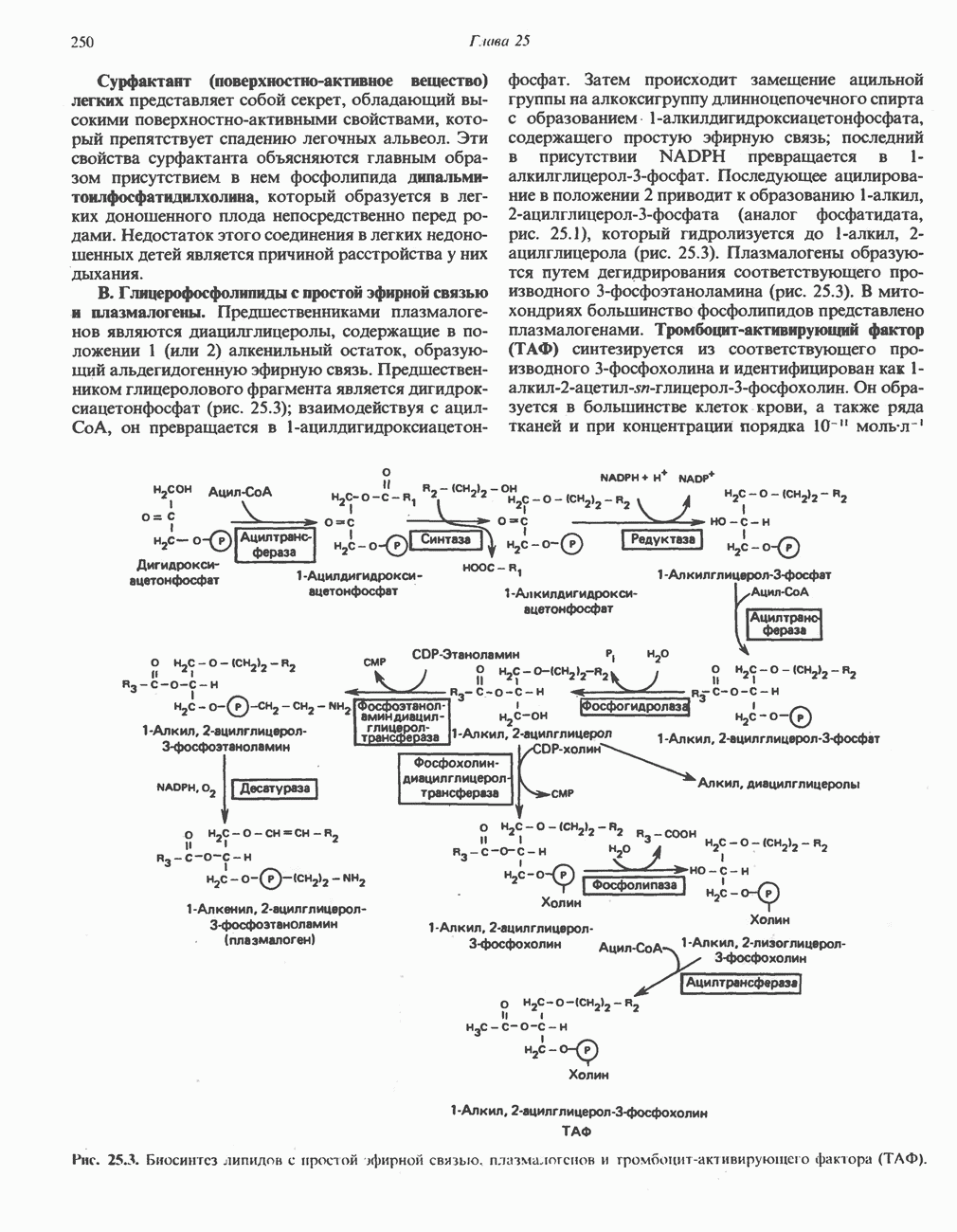
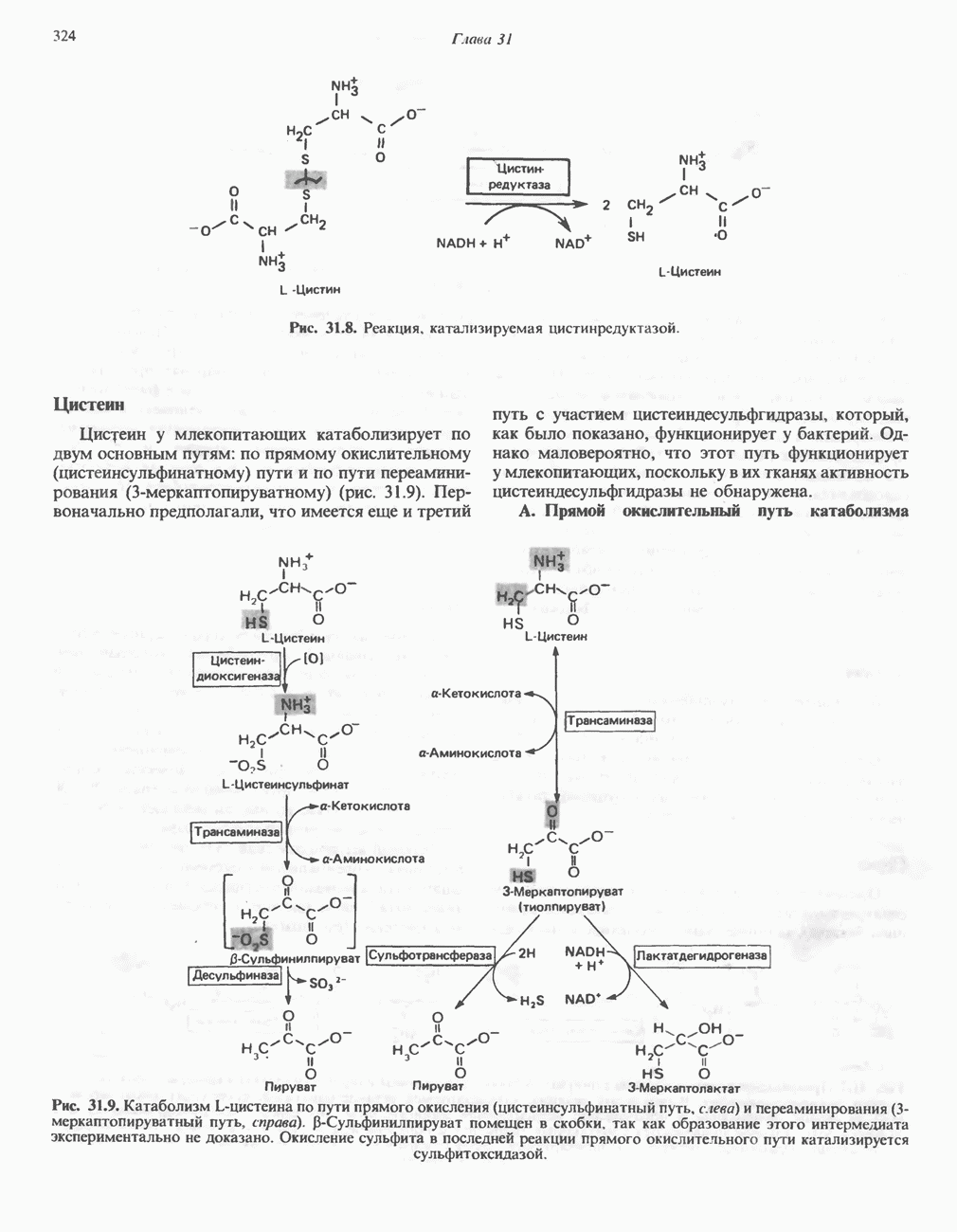
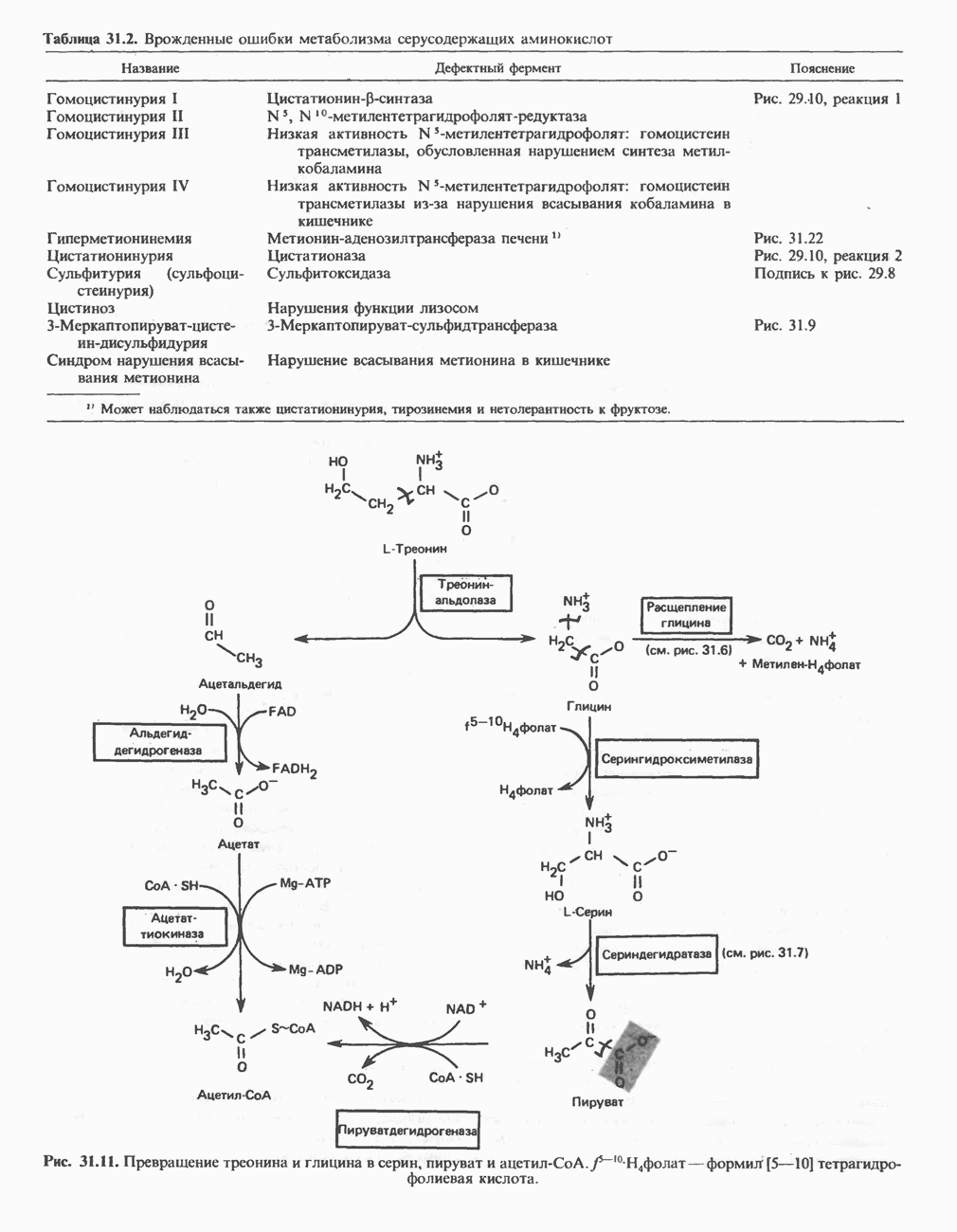
Рис. 25.2. Биосинтез кардиолинина. Сурфактант (поверхностно-активное вещество) легких представляет собой секрет, обладающий высокими поверхностно-активными свойствами, который препятствует спадению легочных альвеол. Эти свойства сурфактанта объясняются главным образом присутствием в нем фосфолипида дипальмитонлфосфатиднлхолнна, который образуется в легких доношенного плода непосредственно перед родами. Недостаток этого соединения в легких недоношенных детей является причиной расстройства у них дыхания. В. Глицерофосфолипиды с простой эфирной связью и плазмалогены. Предшественниками плазмалогенов являются диацилглицеролы, содержащие в положении 1 (или 2) алкенильный остаток, образующий альдегидогенную эфирную связь. Предшественником глицеролового фрагмента является дигидроксиацетонфосфат (рис. 25.3); взаимодействуя с ацил-СоА, он превращается в 1-ацилдигидроксиацетон-фосфат. Затем происходит замещение ацильной группы на алкоксигруппу длинноцепочечного спирта с образованием 1-алкилдигидроксиацетонфосфата, содержащего простую эфирную связь; последний в присутствии NADPH превращается в 1-алкилглицерол-3-фосфат. Последующее ацилирование в положении 2 приводит к образованию (см. скан) Рис. 25.3. Биосинтез липидов с простой эфирной связью, плазмалогснов и тромбоцит-активируюшего фактора (ТАФ). вызывает агрегацию тромбоцитов. ТАФ вызывает также снижение кровяного давления. Распад и обновление глицерофосфолипидов Многие сложные молекулы, например молекулы белков, расщепляются в тканях полностью. Поэтому для них можно определить время обновления. Фосфолипиды также активно распадаются, но в этом случае для каждой части молекулы время обновления различно. Например, время обновления фосфатной группы отличается от времени обновления 1-ацильной группы; это обусловлено наличием ферментов, вызывающих частичный гидролиз фосфолипидов, вслед за которым может снова происходить их синтез (рис. 25.4). Фосфолипаза
Рис. 25.4. Метаболизм фосфатидилхолина (лецитина).
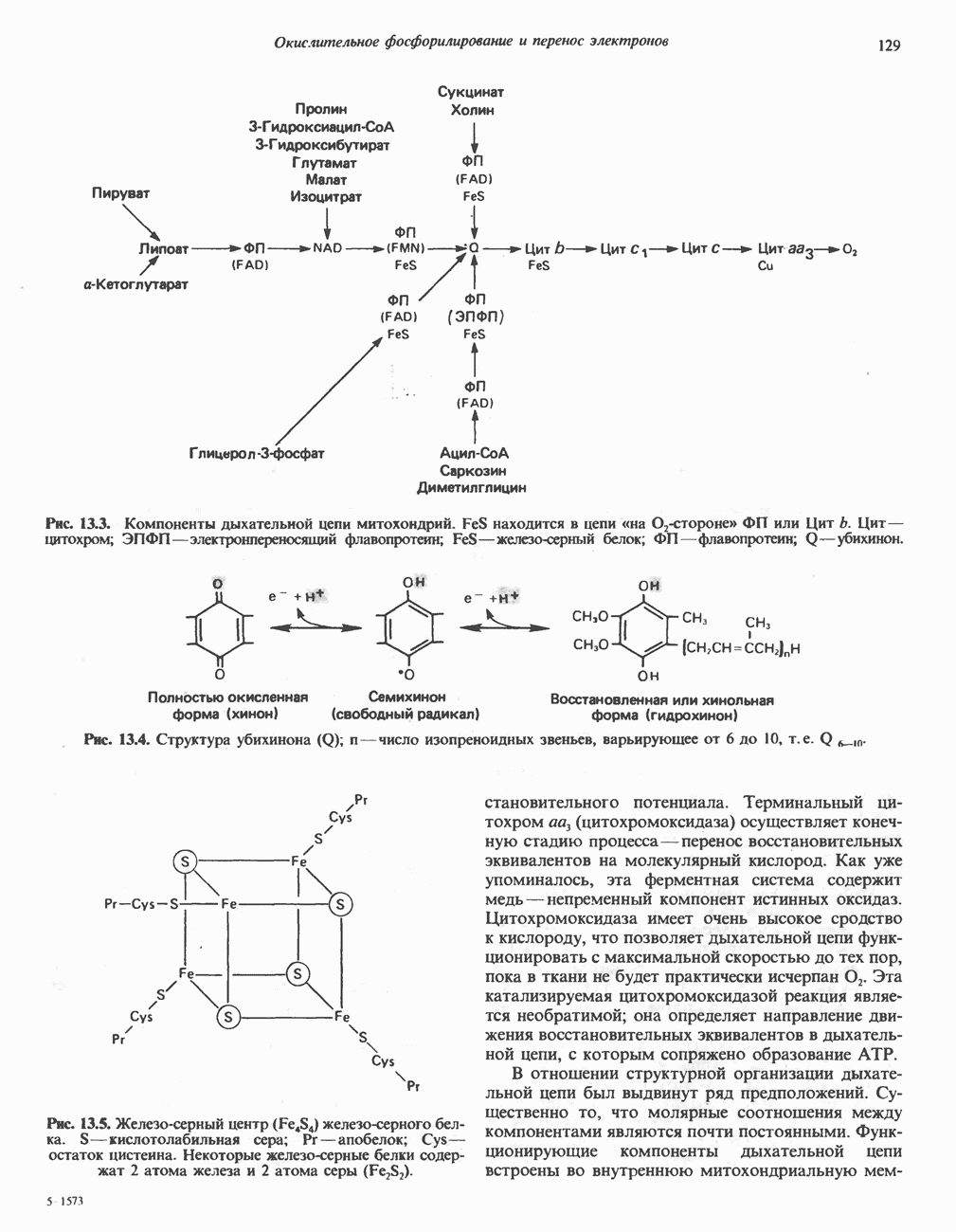
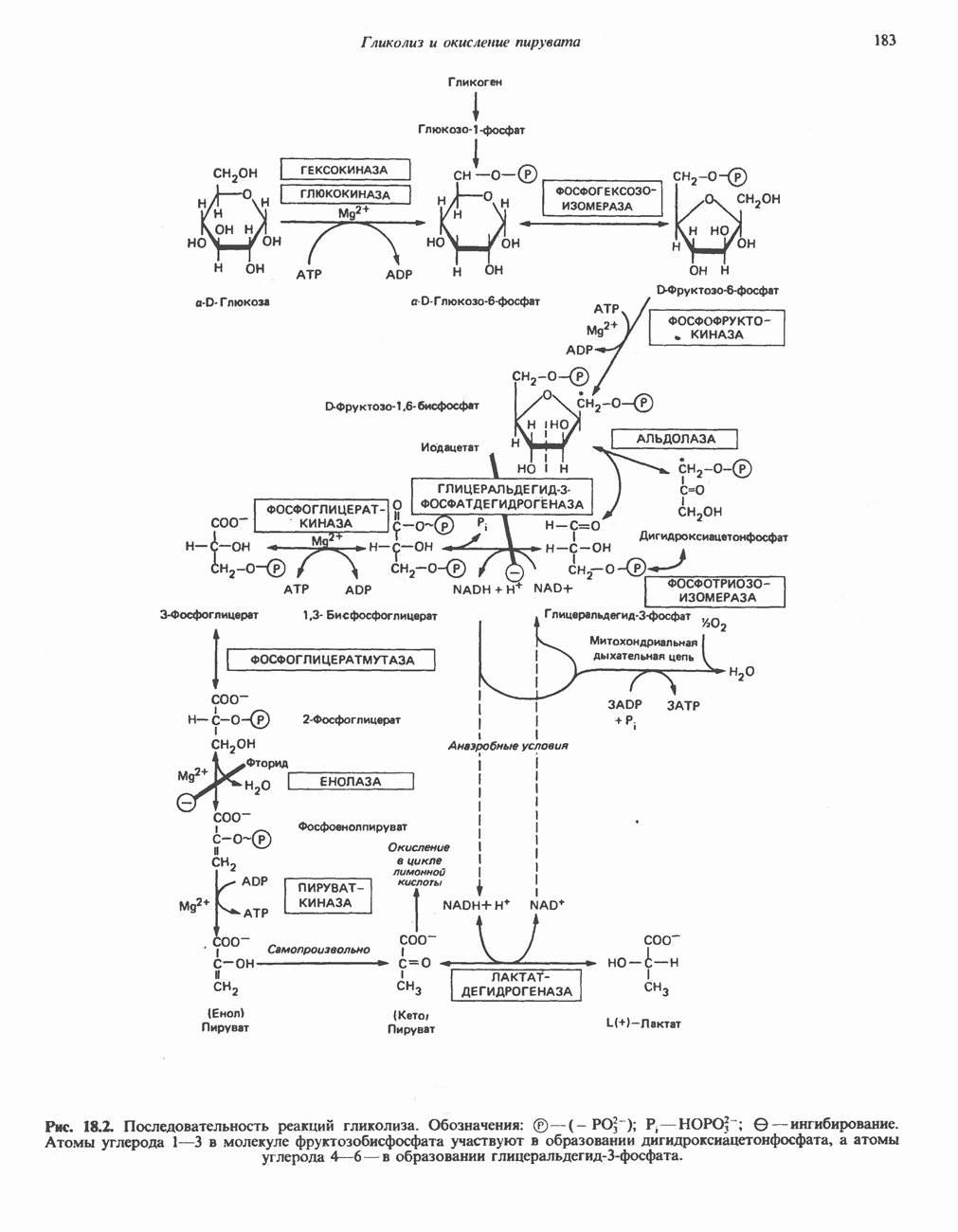
Рис. 25.5. Эфирные связи фосфолипидного субстрата, гидролизуемые фосфолипазами. В альтернативном варианте лизофосфолипид (например, лизолецитин) атакуется лизофосфолипазой (фосфолипазой В), при этом отщепляется оставшаяся 1-ацильная группа и образуется соответствующее глицеролфосфорильное основание. Последнее в свою очередь может расщепляться гидролазой до глицерол-3-фосфата и основания. Фосфолипаза А, атакует эфирную связь фосфолипидов в положении 1 (рис. 25.5), а фосфолипаза С - в положении 3 (в последнем случае образуется 1,2-диацилглицерол и фосфорильное основание). Фосфолипаза С является одним из главных бактериальных токсинов. Фосфолипаза D, встречающаяся главным образом у растений, катализирует отщепление от фосфолипида азотистого основания. Лизолецитин может синтезироваться по альтернативному пути с участием лецитин: холестерол ацилтрансферазы (ЛХТА). Этот фермент, находящийся в плазме крови и образующийся в печени, катализирует перенос остатка жирной кислоты из положения 2 молекулы лецитина на холестерол, в результате образуется сложный эфир холестерола. Считается, что именно под действием ЛХТА синтезируется большая часть сложных эфиров холестерола— компонентов липопротеинов плазмы крови. Последствия недостатка ЛХАТ обсуждаются на с. 281.
В фосфолипидах длинноцепочечные насыщенные жирные кислоты находятся главным образом в положении 1, в то время как полиненасыщенные жирные кислоты (например, предшественники простагландинов) - чаще всего в положении 2. Включение жирных кислот в молекулу лецитина происходит при полном синтезе фосфолипида, при трансацилировании между эфиром холестерола и лизолецитином, а также при прямом ацилировании лизолецитина ацил-СоА. Таким образом может происходить постоянное обновление жирных кислот, особенно важным является включение в молекулы фосфолипидов незаменимых жирных кислот.
|
 |
Оглавление
|


































































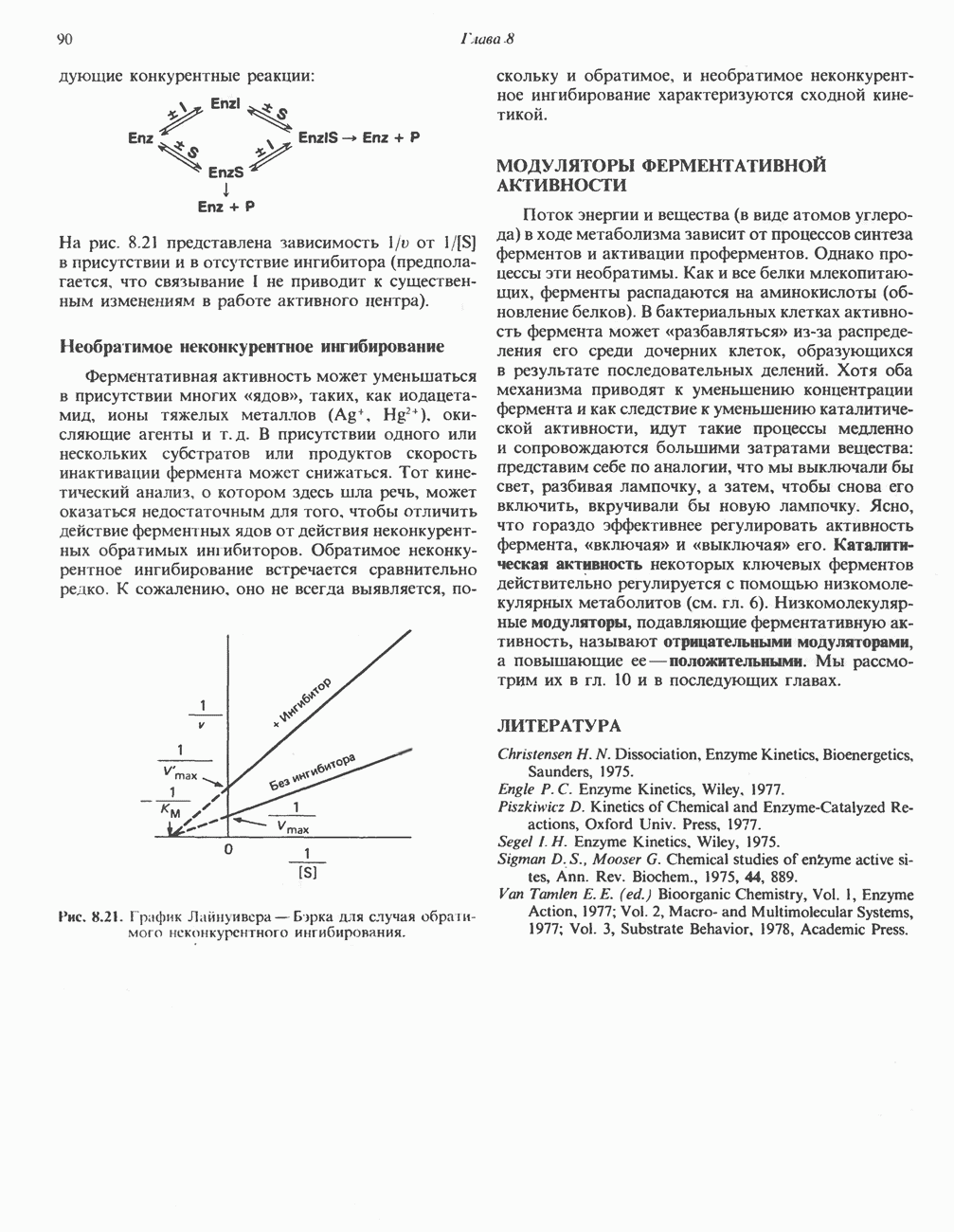

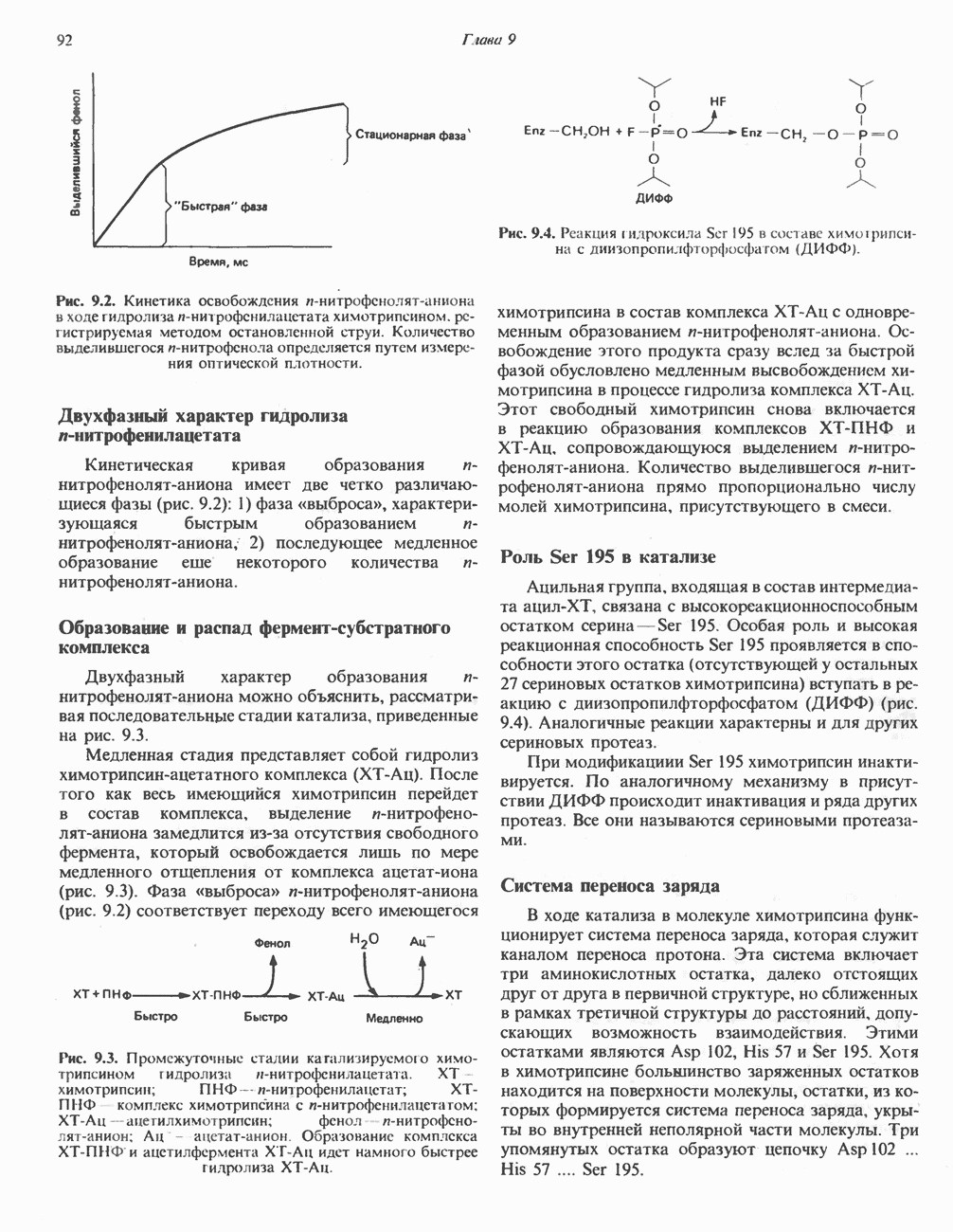










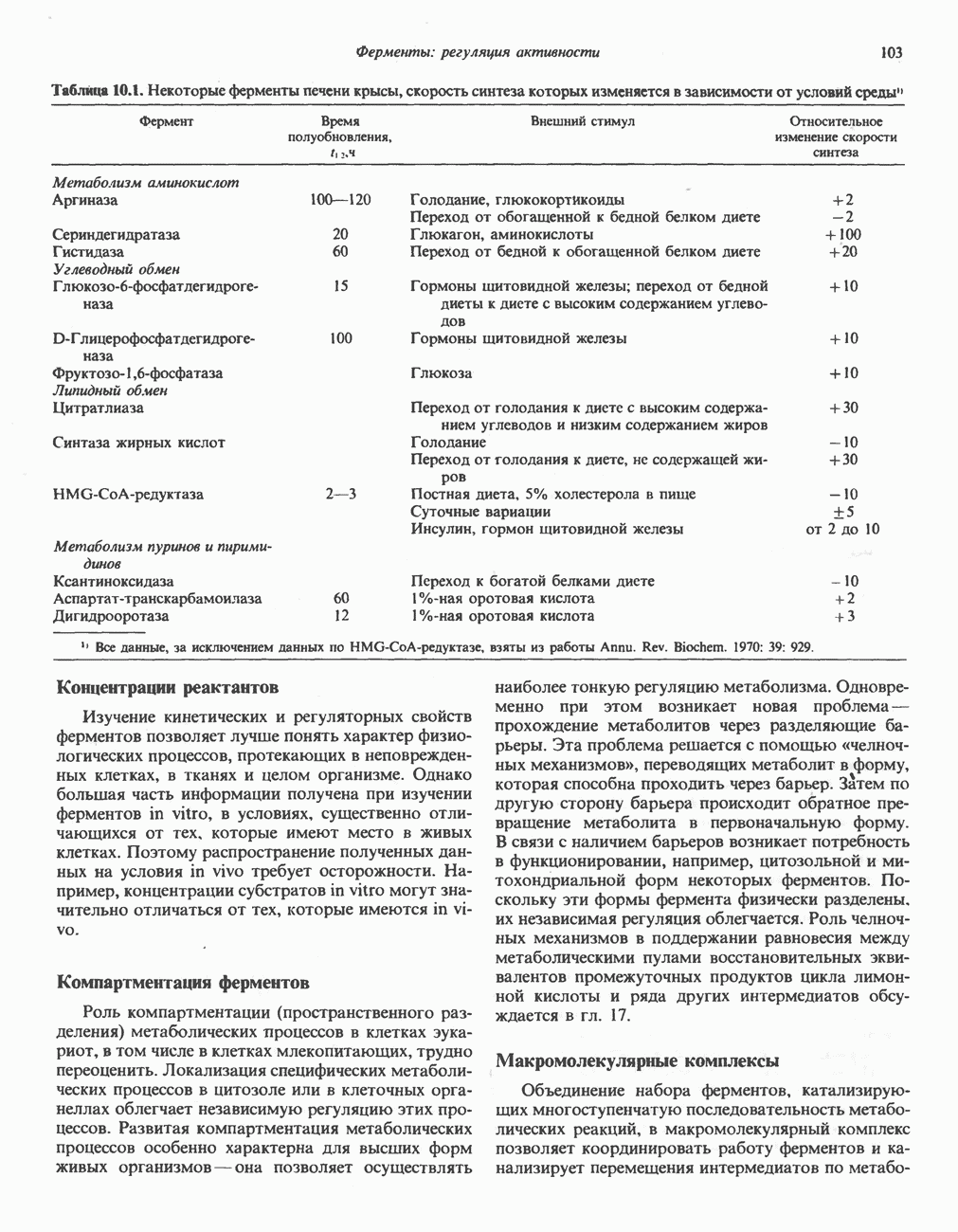


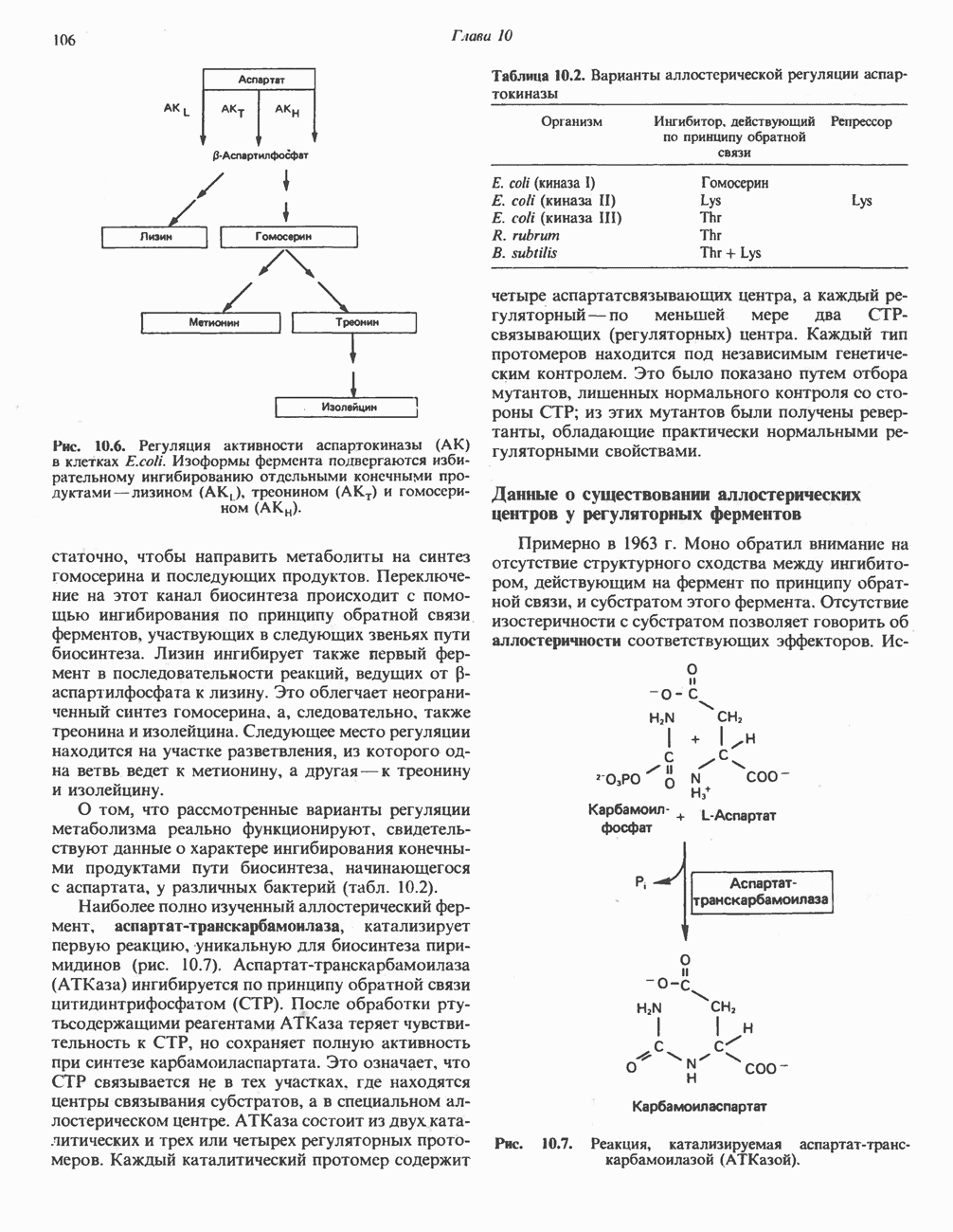



































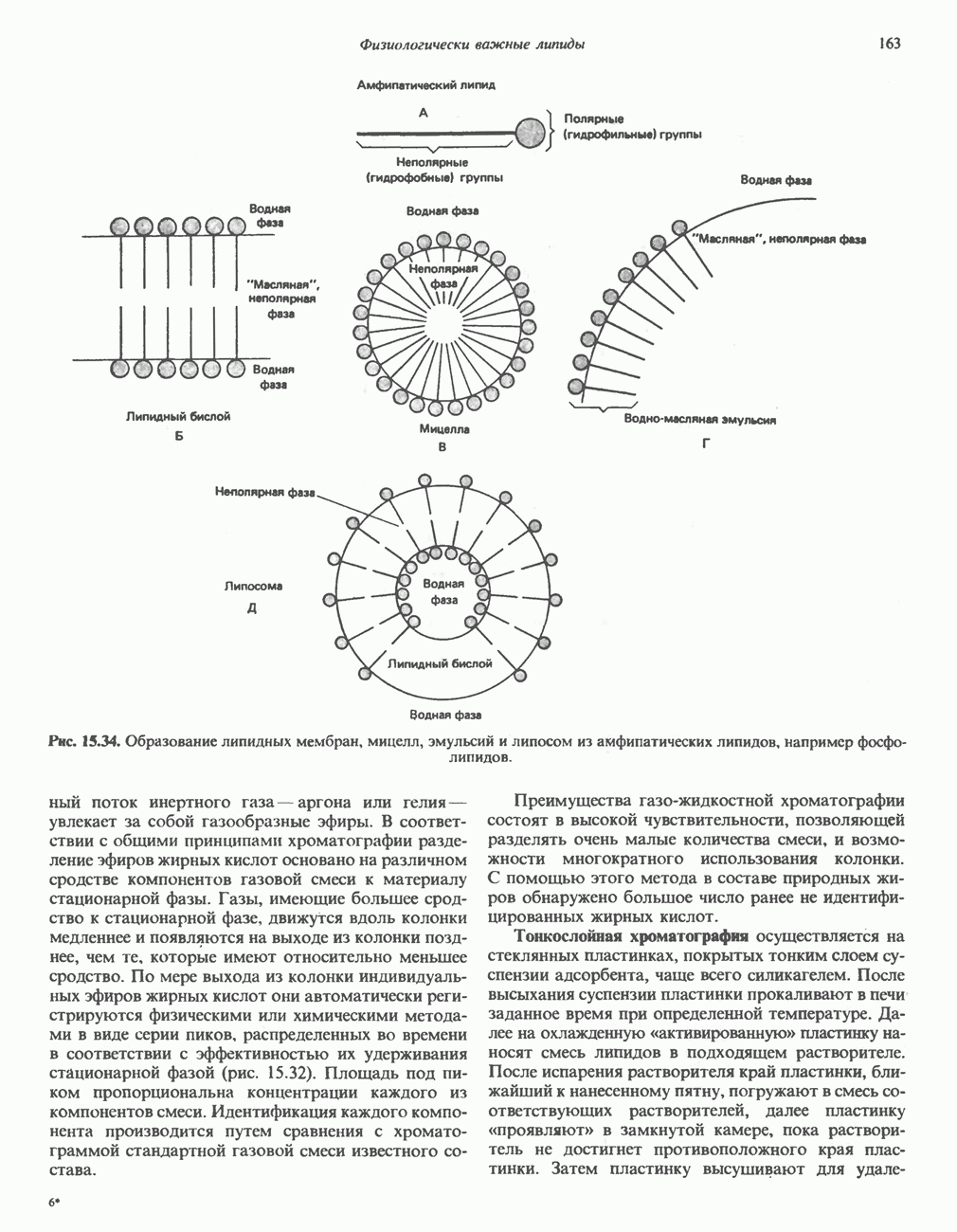




















































































































 При отсутствии данного фермента
При отсутствии данного фермента
 -диацилглицерола. В слизистой оболочке кишечника функционирует путь образования 1,2-диацилглицерола из моноа-цилглицерола, реакция катализируется моноацилгли-церол-ацилтрансферазой. Далее 1,2-диацилглицерол ацилируется третьей молекулой ацил-СоА и превращается в триацилглицерол (эта реакция катализируется диацилглицерол-ацилтрансферазой). Большинство ферментов данного пути находятся в эндоплазматическом ретикулуме, и только некоторые, например глицерол-3-фосфат-ацилтрансфераза, в митохондриях. Фосфатидатфосфогидролазная активность обнаруживается главным образом в супернатантной фракции, часть ее связана с мембранами.
-диацилглицерола. В слизистой оболочке кишечника функционирует путь образования 1,2-диацилглицерола из моноа-цилглицерола, реакция катализируется моноацилгли-церол-ацилтрансферазой. Далее 1,2-диацилглицерол ацилируется третьей молекулой ацил-СоА и превращается в триацилглицерол (эта реакция катализируется диацилглицерол-ацилтрансферазой). Большинство ферментов данного пути находятся в эндоплазматическом ретикулуме, и только некоторые, например глицерол-3-фосфат-ацилтрансфераза, в митохондриях. Фосфатидатфосфогидролазная активность обнаруживается главным образом в супернатантной фракции, часть ее связана с мембранами.
 ). Эти два продукта действуют как вторые посредники при действии гормонов (рис. 44.5).
). Эти два продукта действуют как вторые посредники при действии гормонов (рис. 44.5).
 -холин), либо цитидиндифосфоэтаноламин QCDP-этаноламин). В такой активной форме холин (или этаноламин) вступает в реакцию с 1,2-диацилгли-церолом; происходит перенос фосфорилированного основания (фосфохолина или фосфоэтаноламина) на диацилглицерол и образуется либо фосфатидилхолин, либо фосфатидилэтаноламин. Регуляторным ферментом на пути образования фосфатидилхолина является, по-видимому, цитидилтрансфераза.
-холин), либо цитидиндифосфоэтаноламин QCDP-этаноламин). В такой активной форме холин (или этаноламин) вступает в реакцию с 1,2-диацилгли-церолом; происходит перенос фосфорилированного основания (фосфохолина или фосфоэтаноламина) на диацилглицерол и образуется либо фосфатидилхолин, либо фосфатидилэтаноламин. Регуляторным ферментом на пути образования фосфатидилхолина является, по-видимому, цитидилтрансфераза.

 -алкил, 2-ацилглицерол-З-фосфата (аналог фосфатидата, рис. 25.1), который гидролизуется до 1-алкил, 2-ацилглицерола (рис. 25.3). Плазмалогены образуются путем дегидрирования соответствующего производного 3-фосфоэтаноламина (рис. 25.3). В митохондриях большинство фосфолипидов представлено плазмалогенами. Тромбоцит - активирующий фактор (ТАФ) синтезируется из соответствующего производного 3-фосфохолина и идентифицирован как
-алкил, 2-ацилглицерол-З-фосфата (аналог фосфатидата, рис. 25.1), который гидролизуется до 1-алкил, 2-ацилглицерола (рис. 25.3). Плазмалогены образуются путем дегидрирования соответствующего производного 3-фосфоэтаноламина (рис. 25.3). В митохондриях большинство фосфолипидов представлено плазмалогенами. Тромбоцит - активирующий фактор (ТАФ) синтезируется из соответствующего производного 3-фосфохолина и идентифицирован как  Он образуется в большинстве клеток крови, а также ряда тканей и при концентрации порядка
Он образуется в большинстве клеток крови, а также ряда тканей и при концентрации порядка 
 катализирует гидролиз эфирной связи в положении 2 глицерофосфолипидов, в результате чего образуются свободная жирная кислота и лизофосфолипид, который в свою очередь реацилируется ацил-СоА при участии ацилтрансферазы.
катализирует гидролиз эфирной связи в положении 2 глицерофосфолипидов, в результате чего образуются свободная жирная кислота и лизофосфолипид, который в свою очередь реацилируется ацил-СоА при участии ацилтрансферазы.


