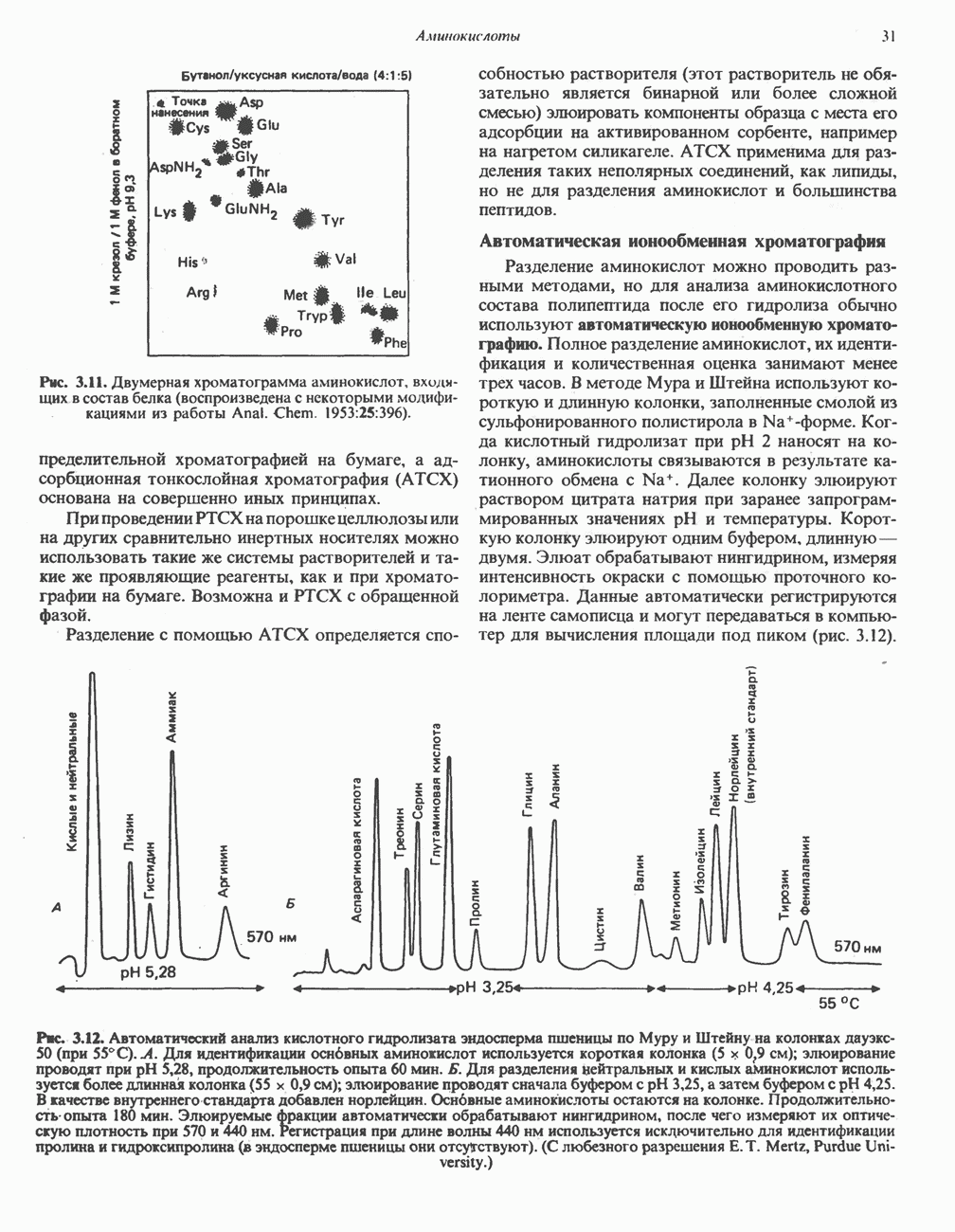
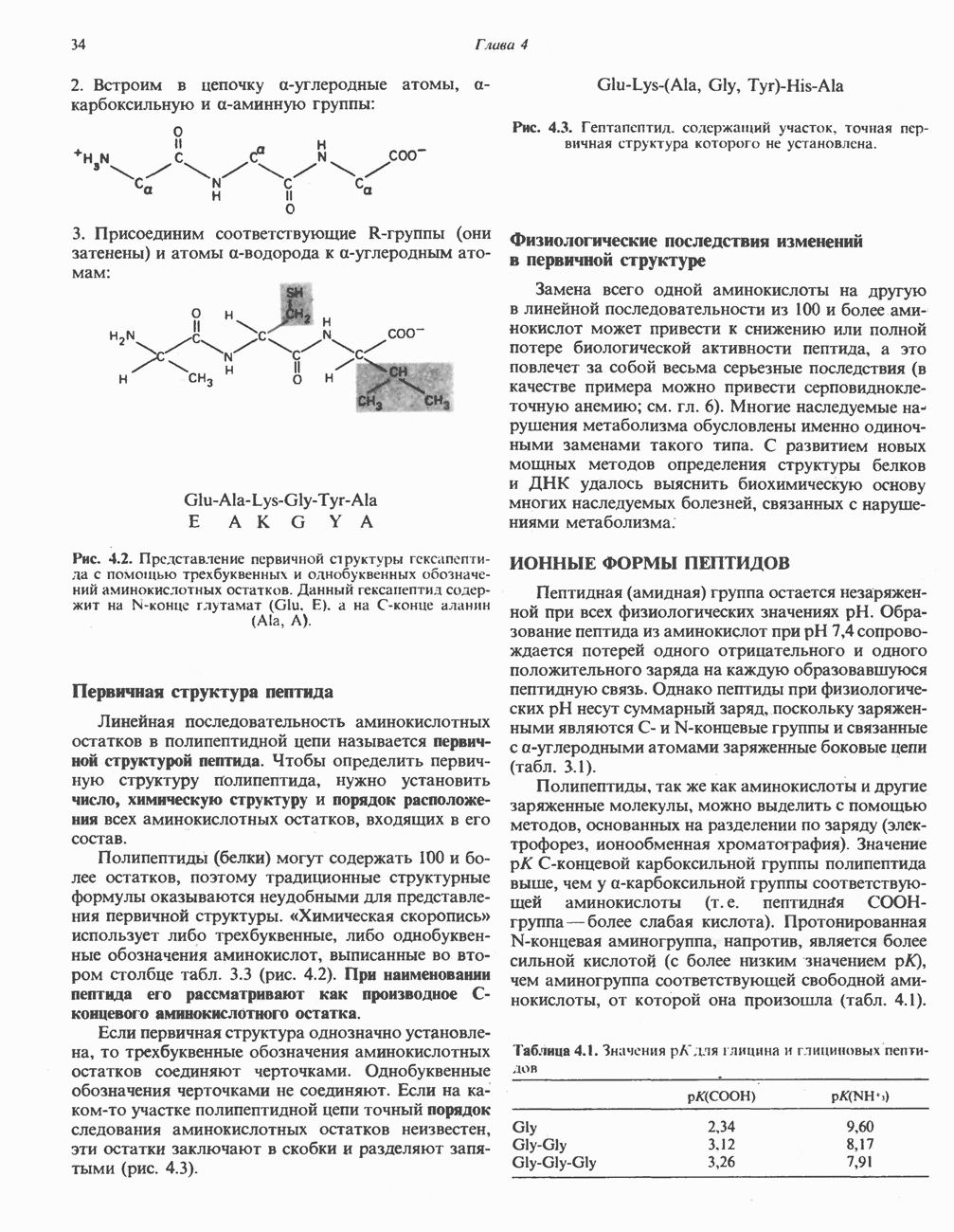
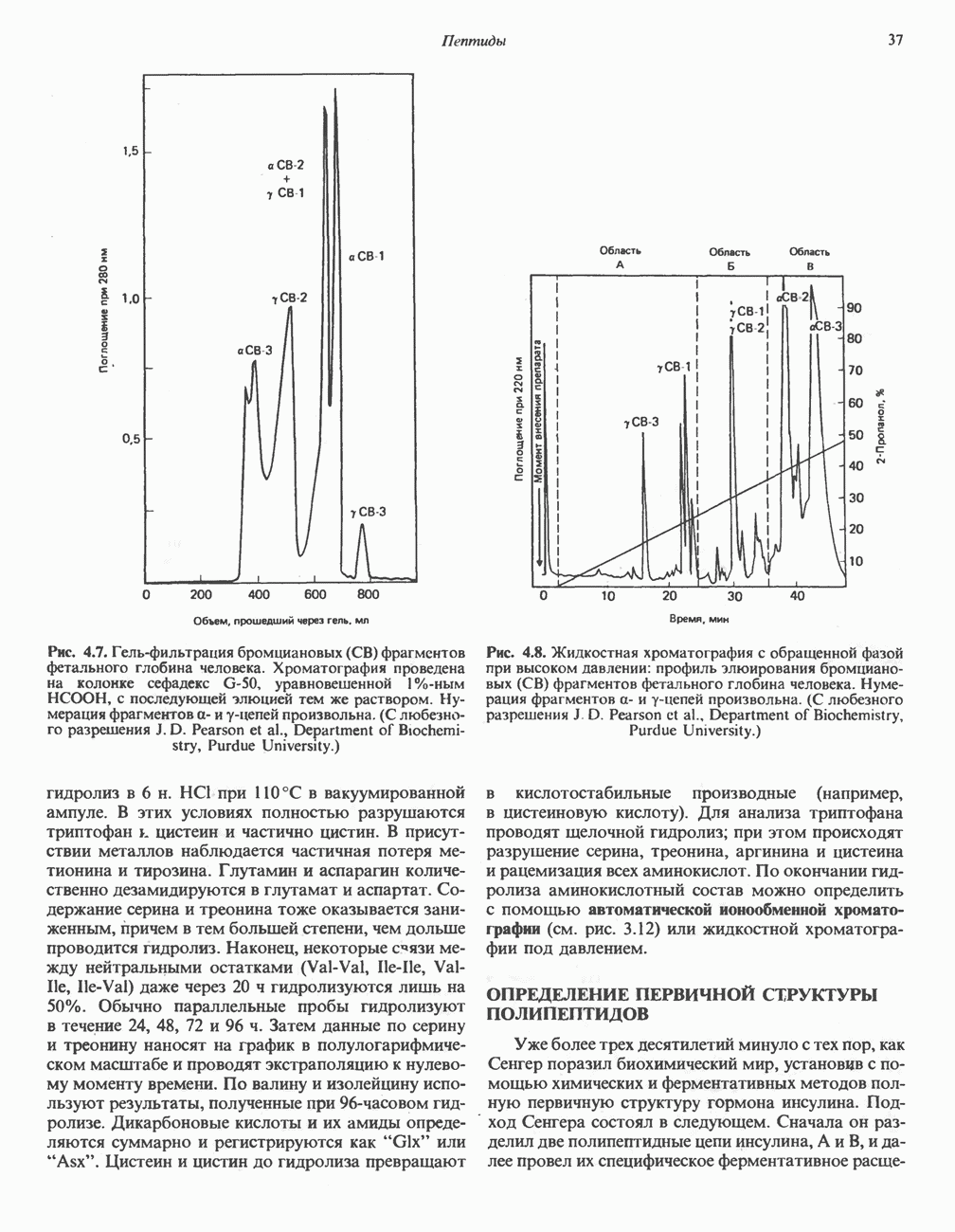
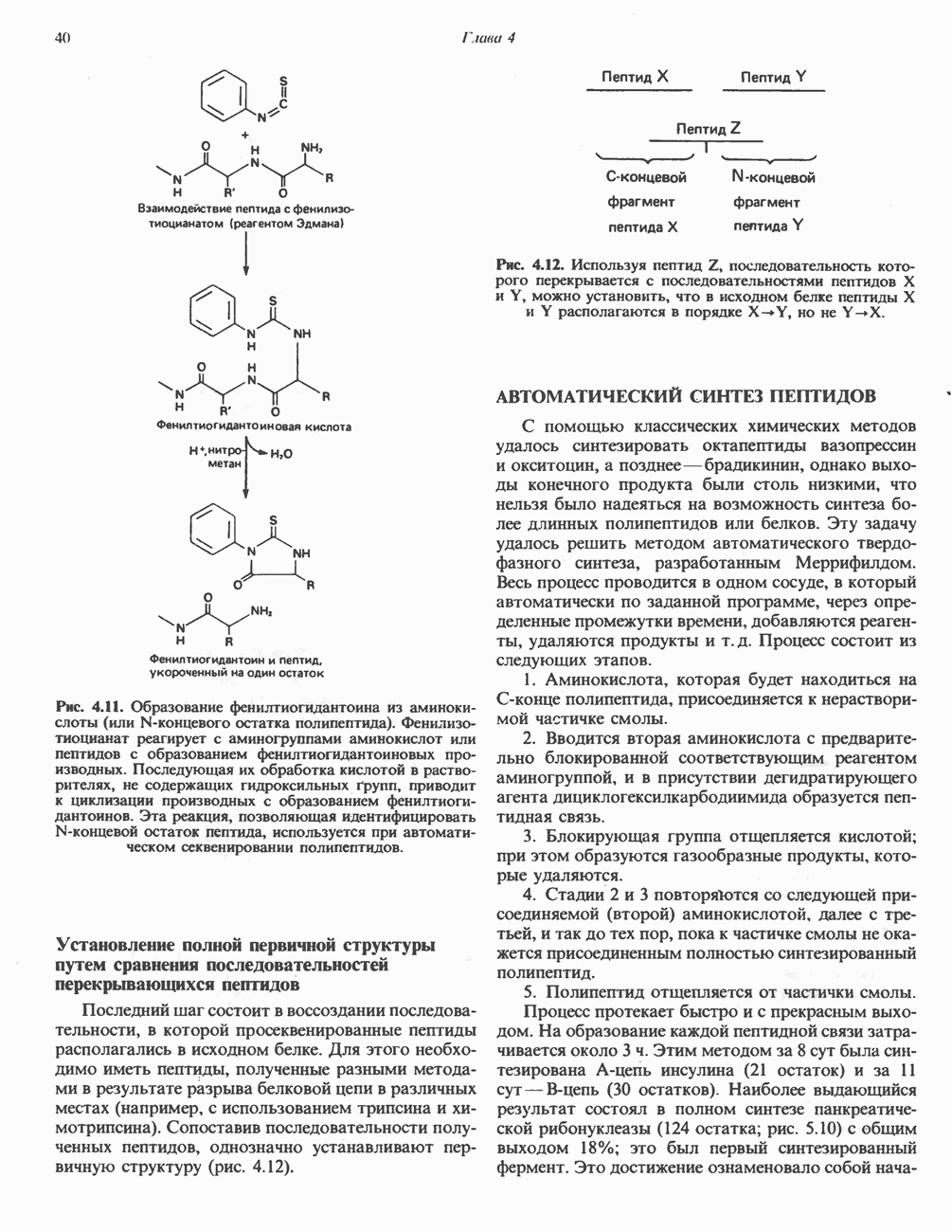
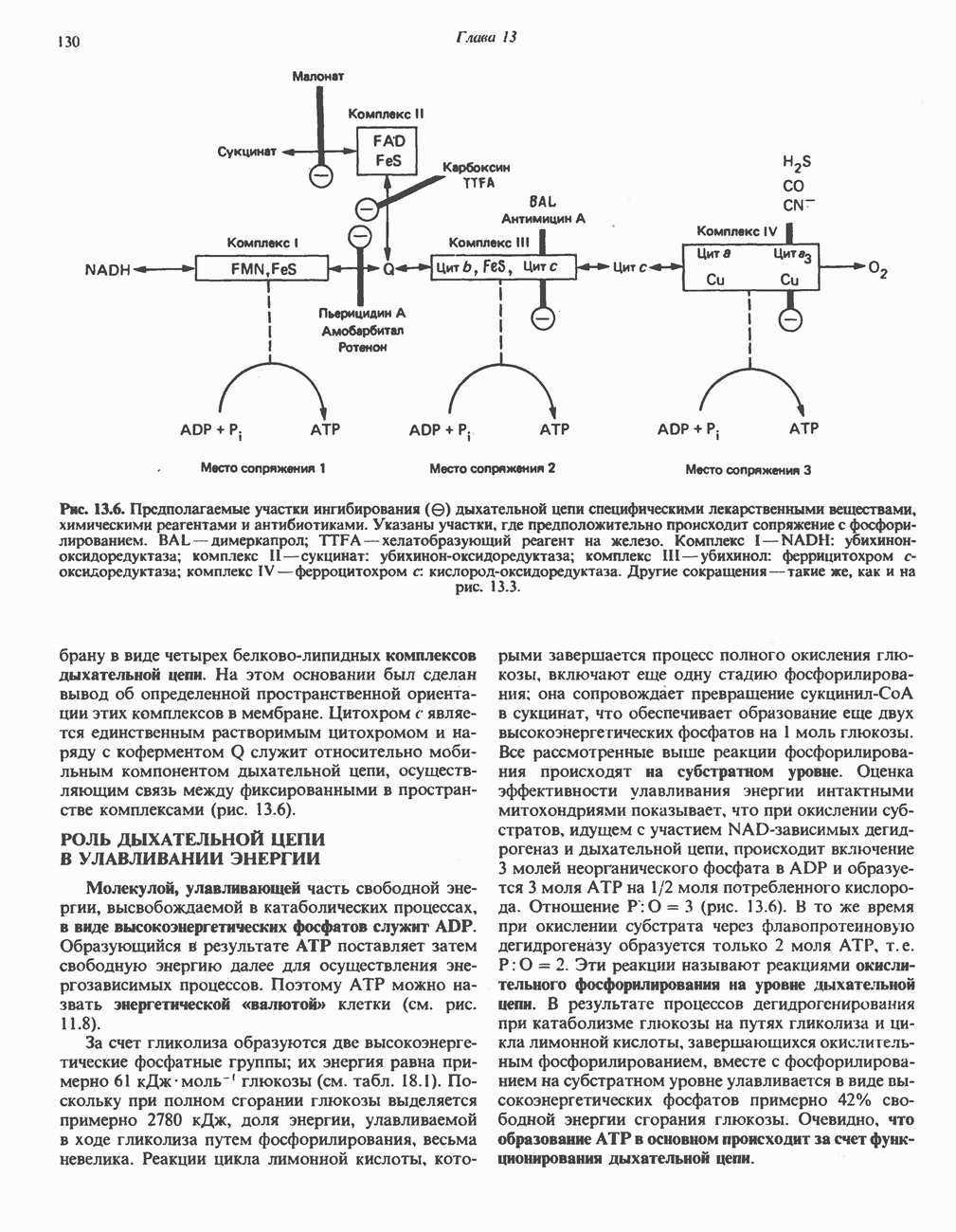
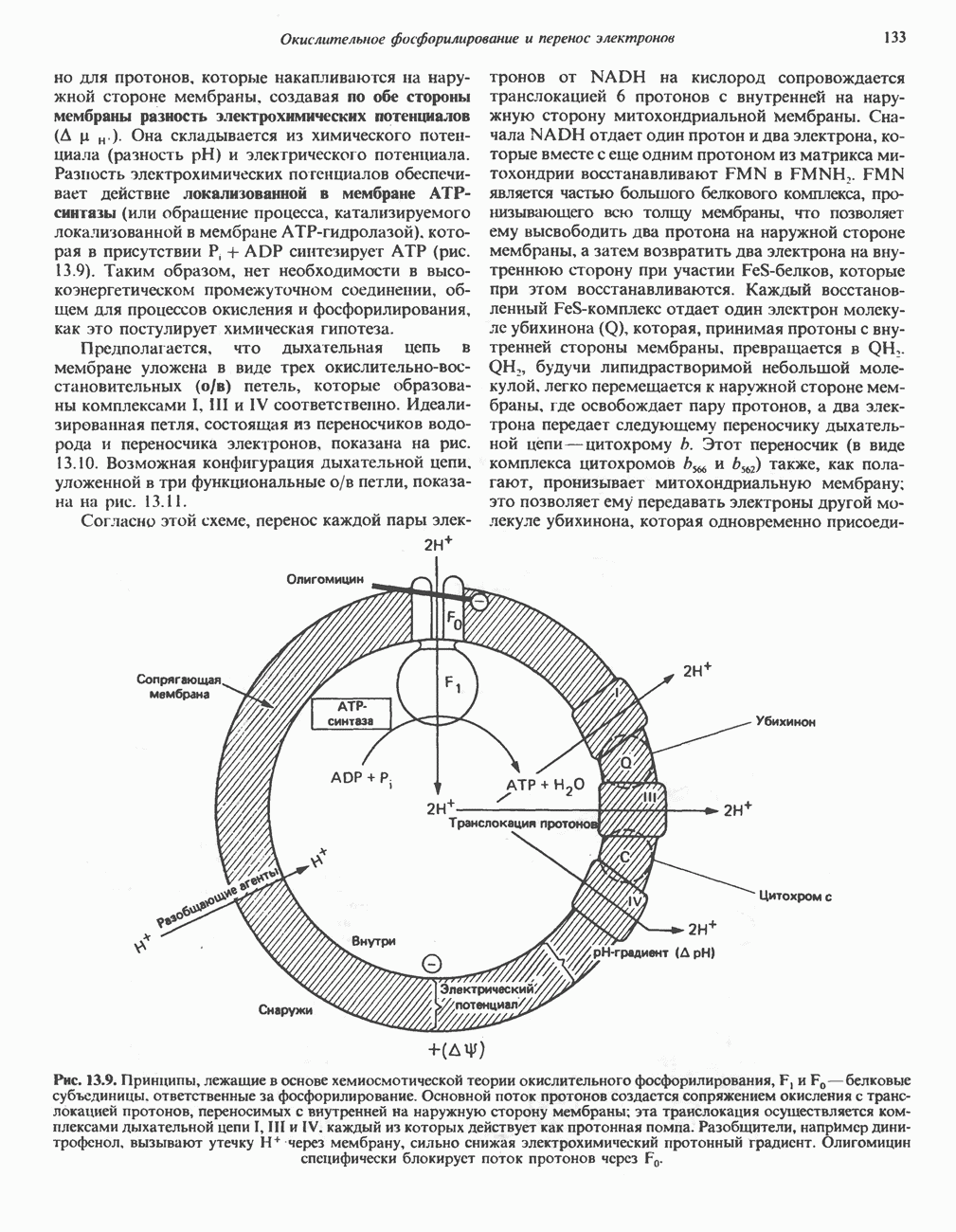
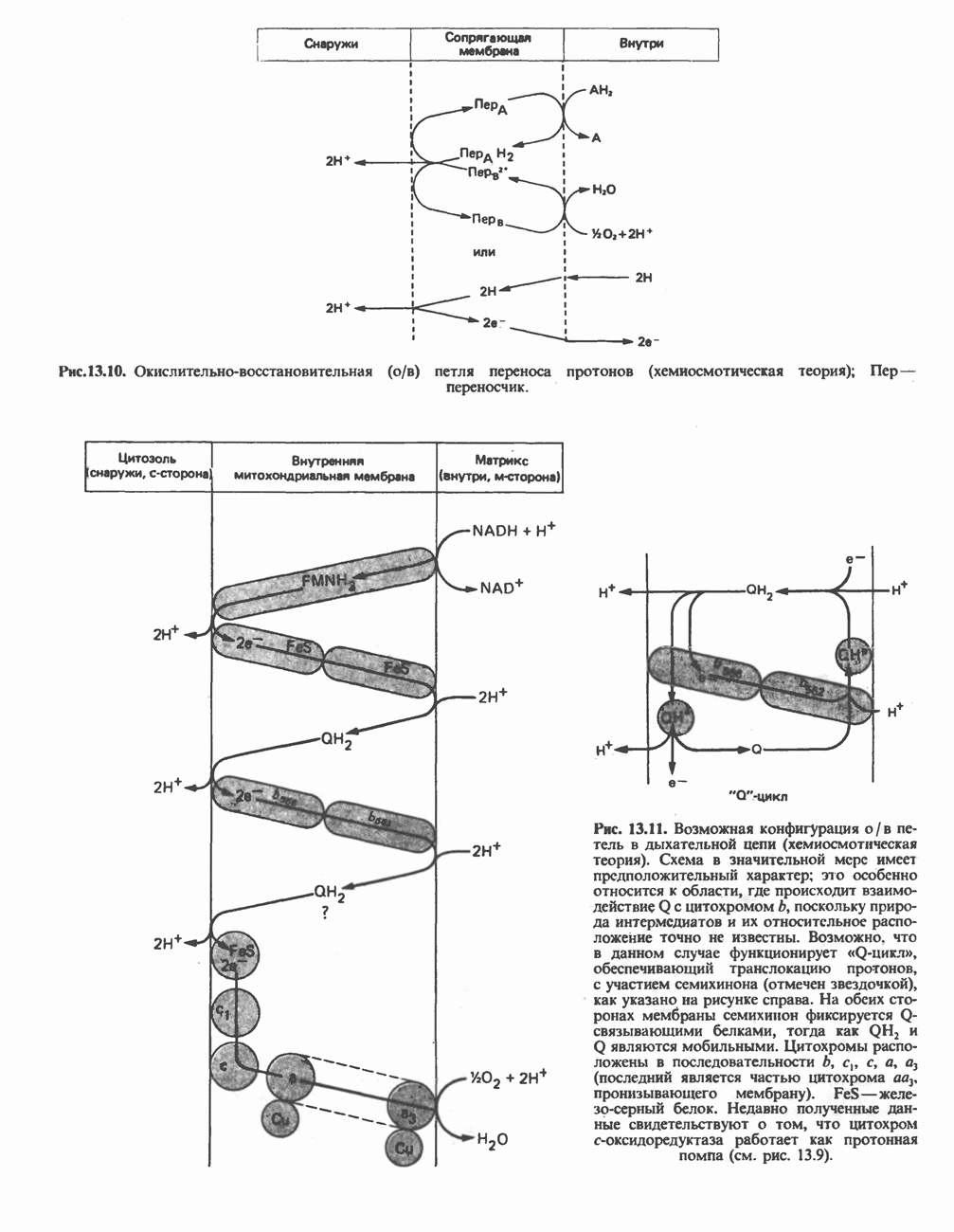
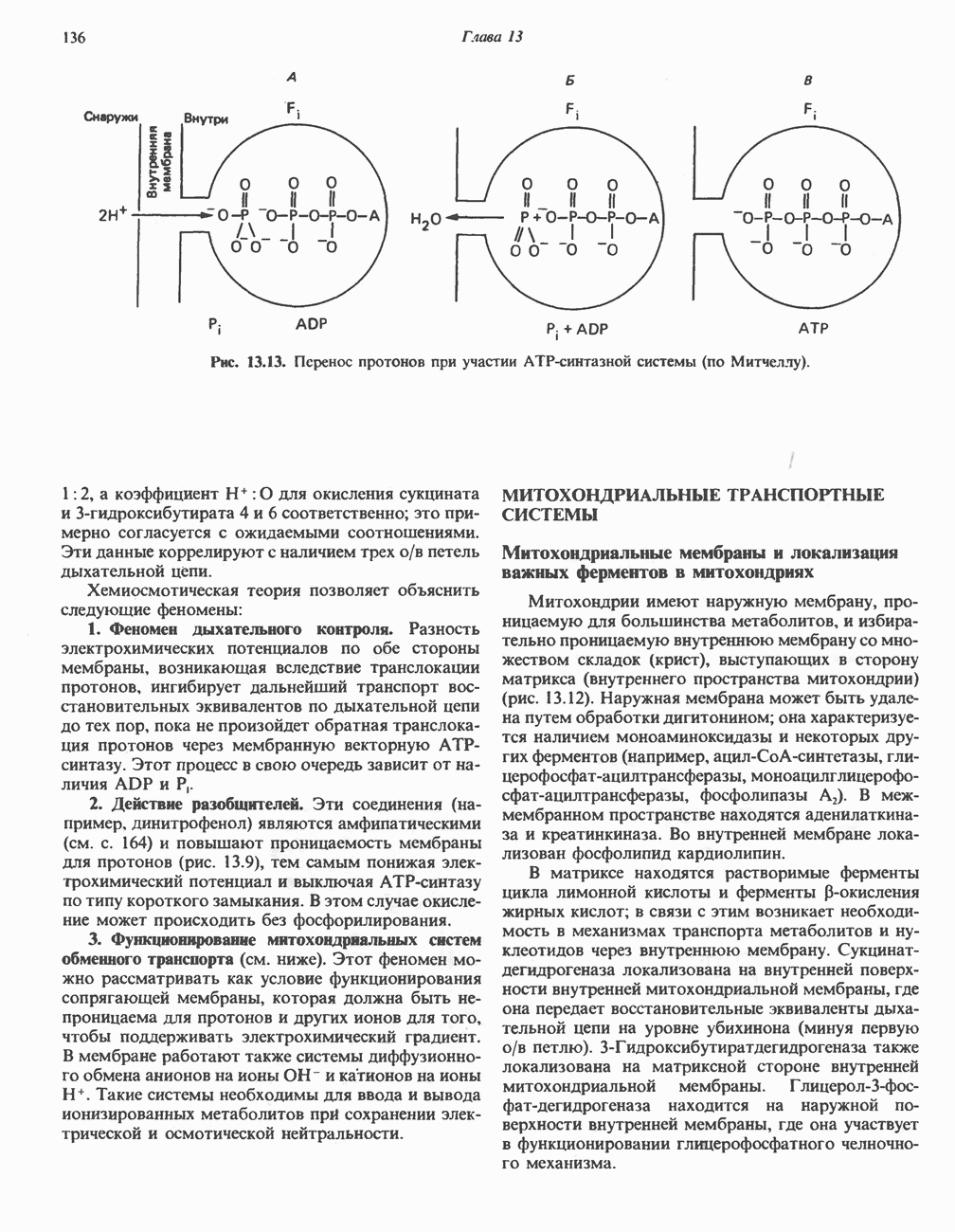
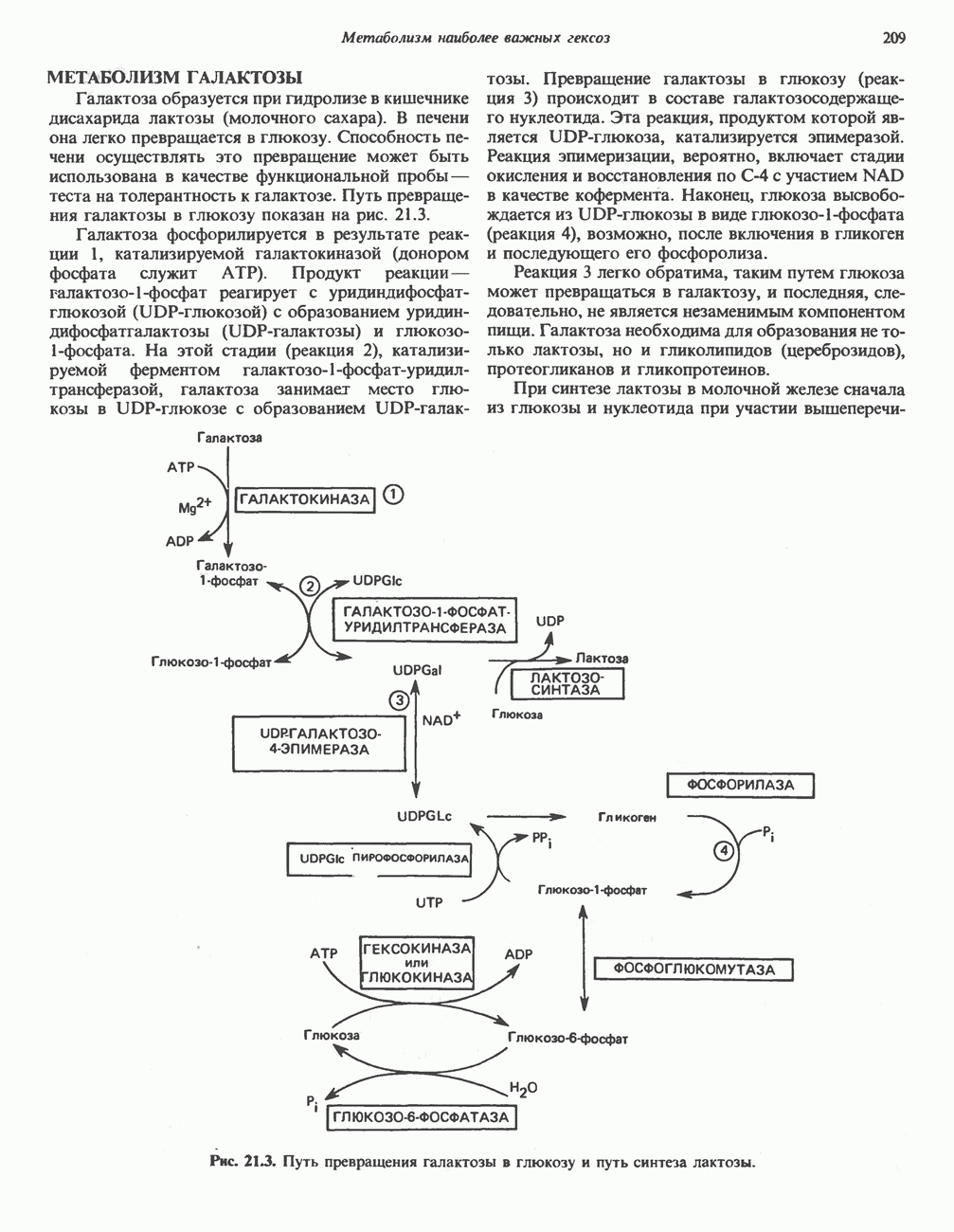
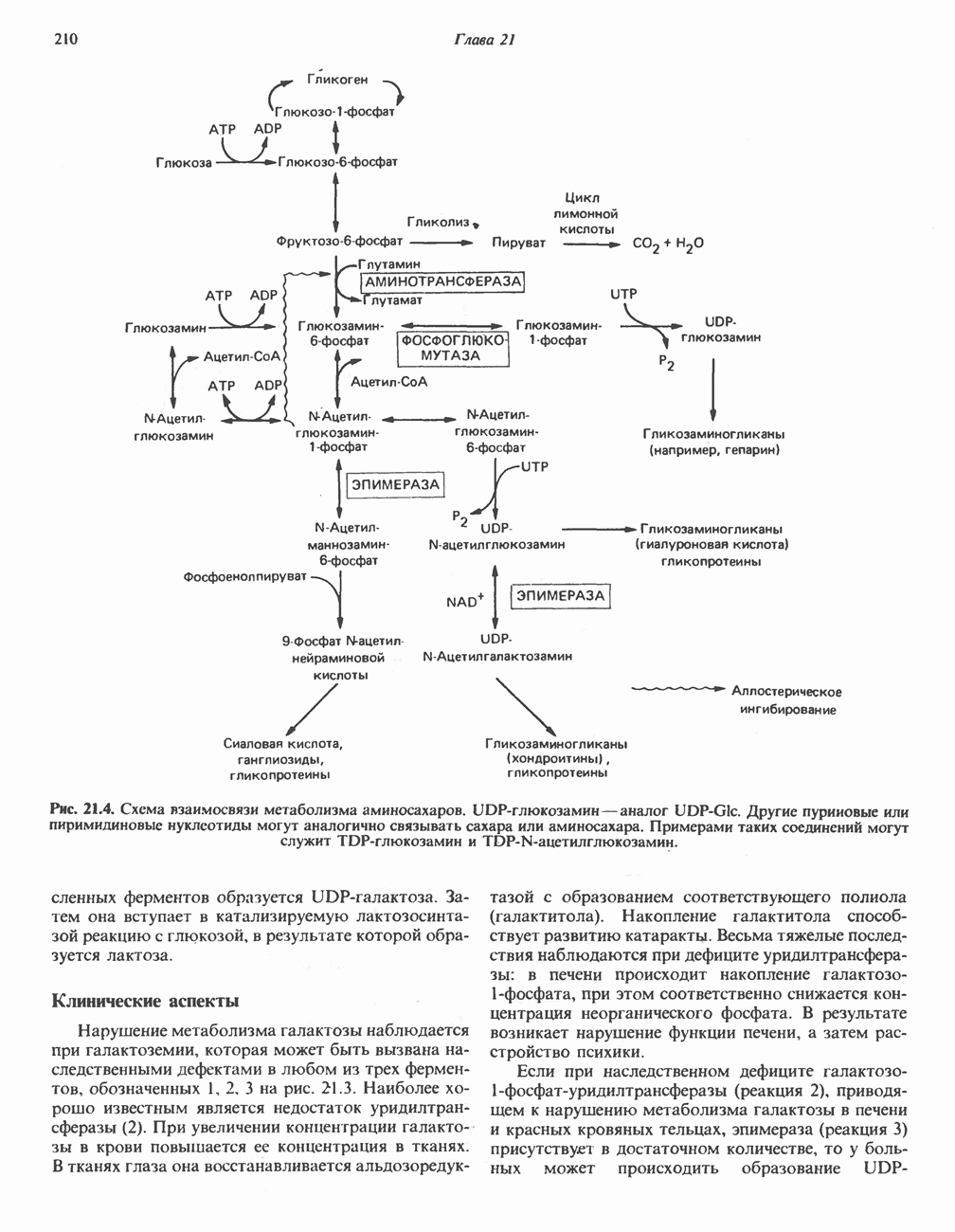
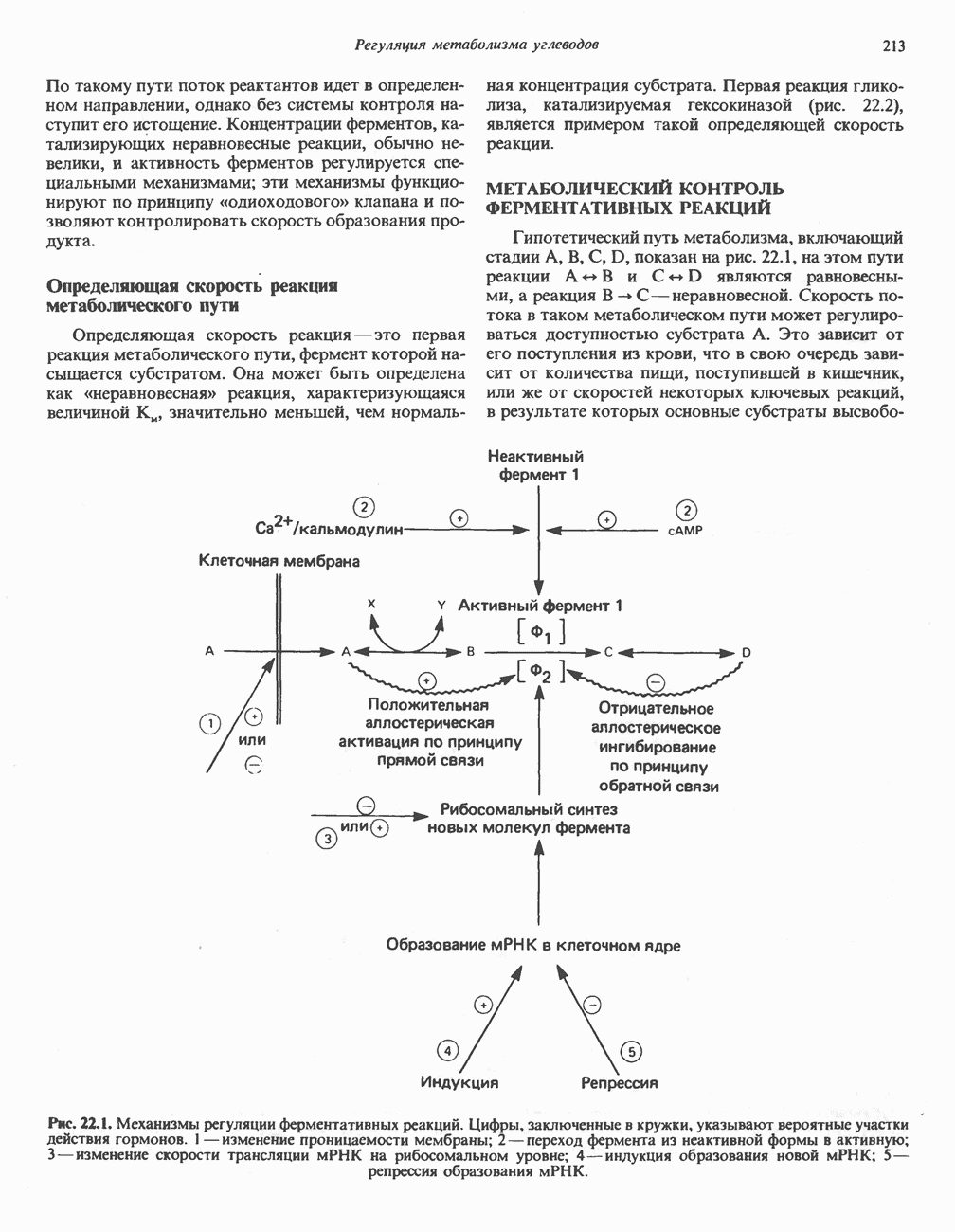
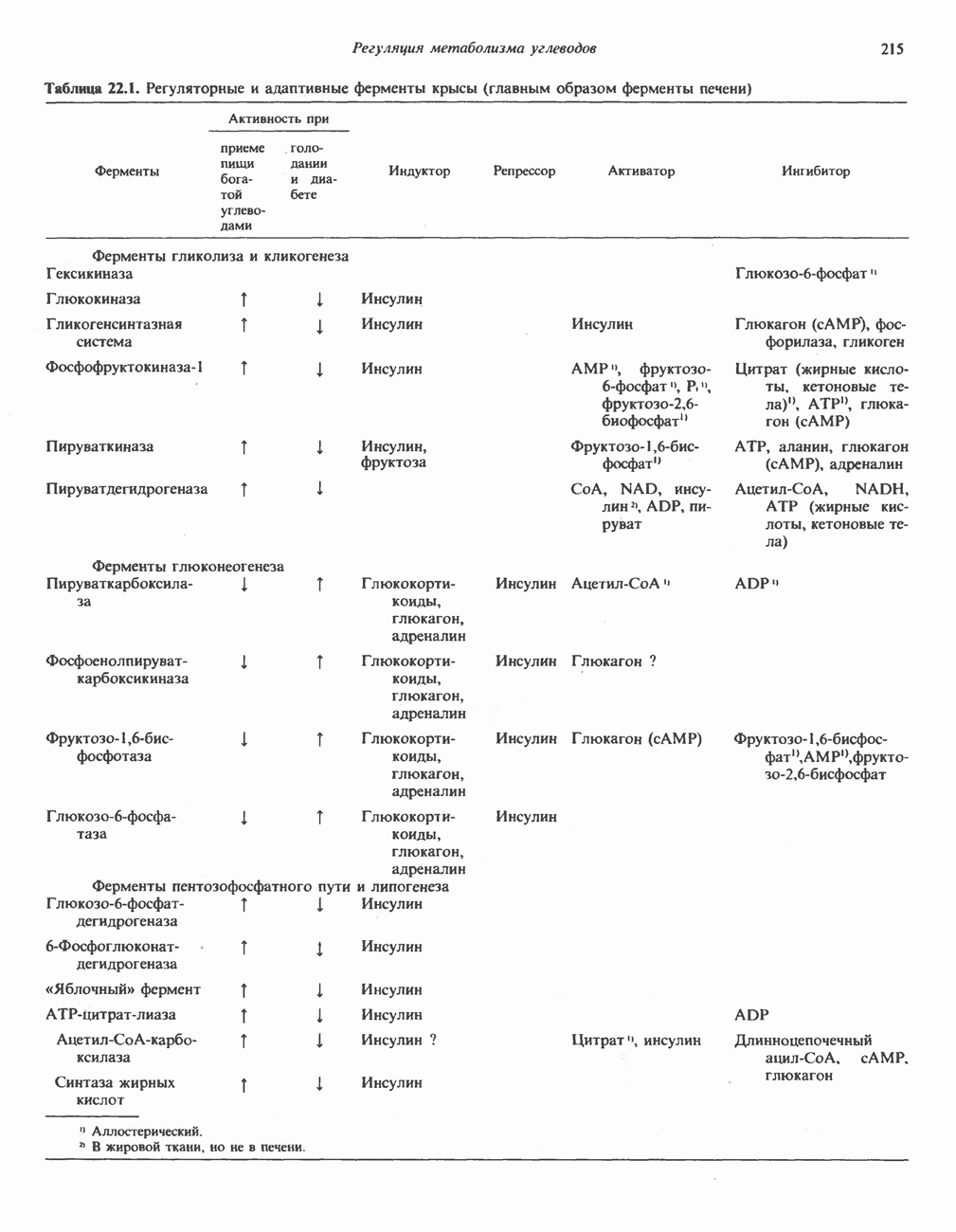
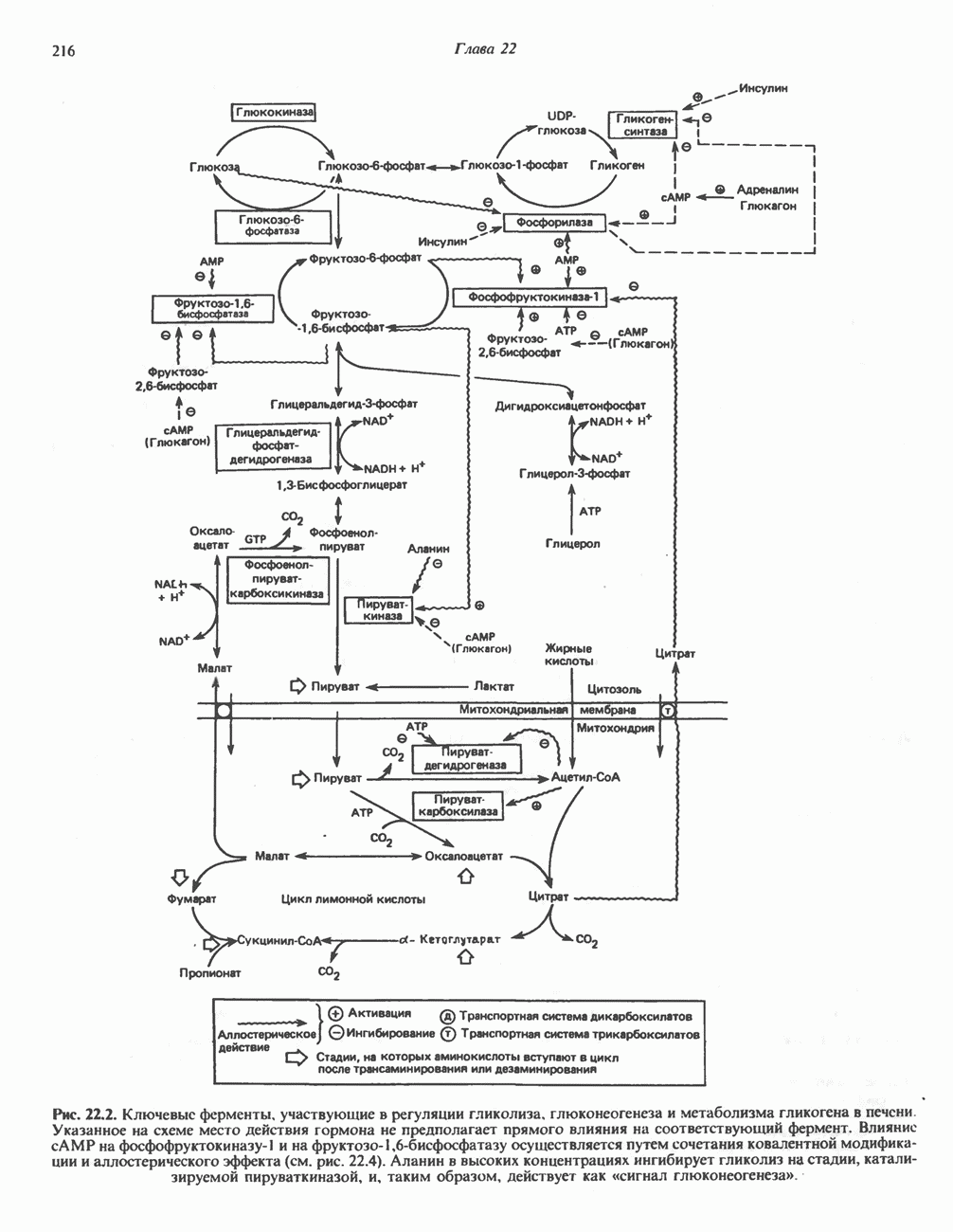
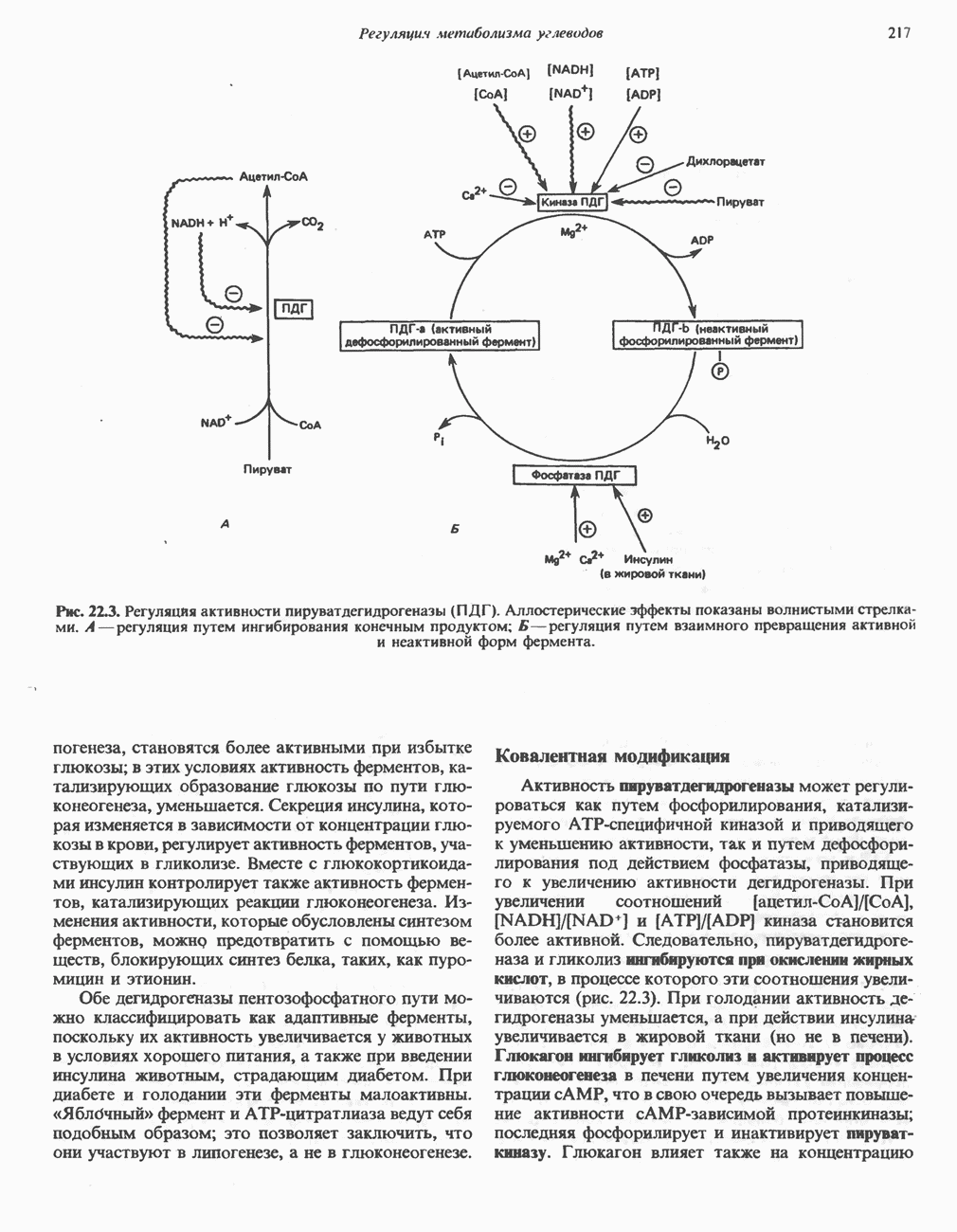
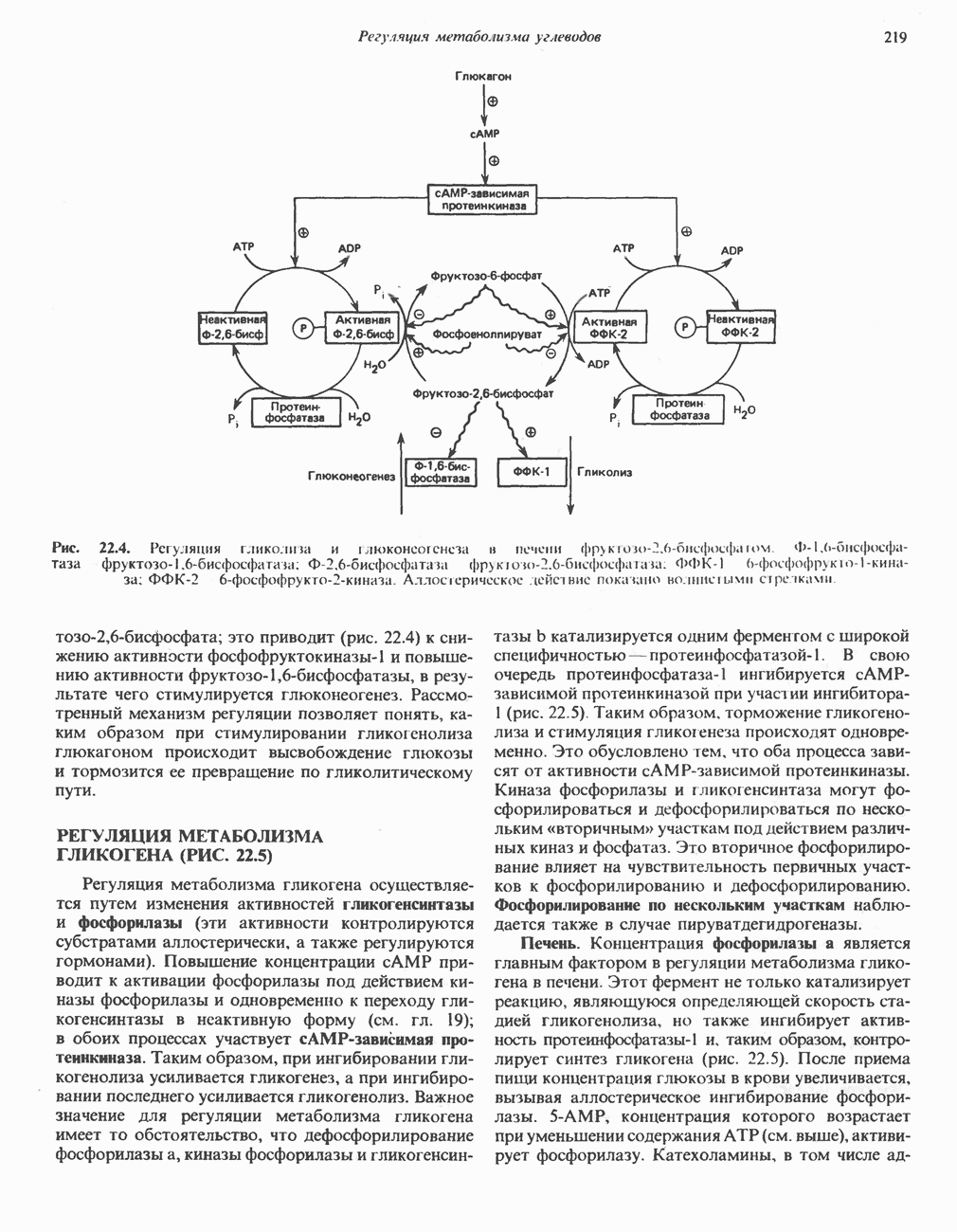
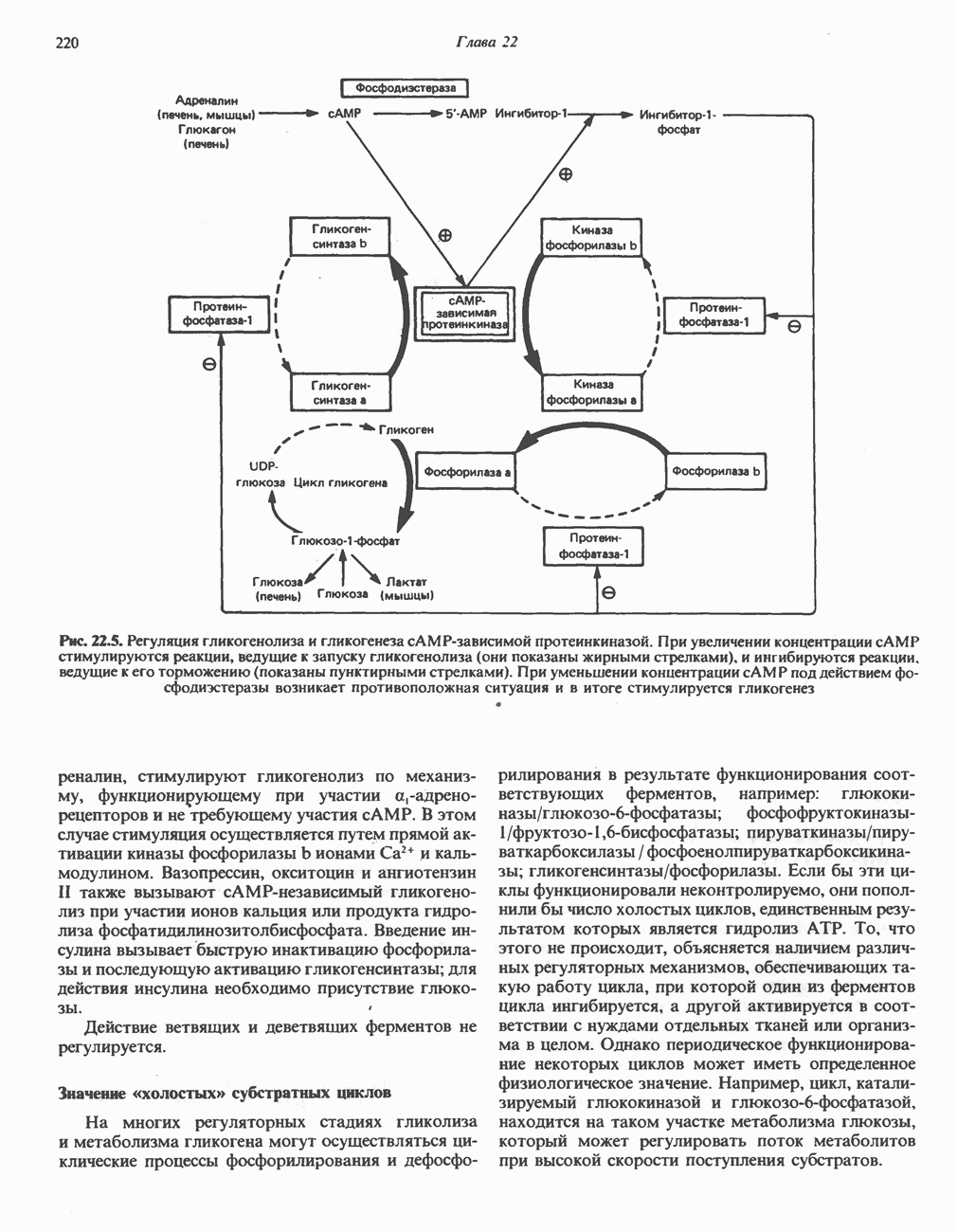
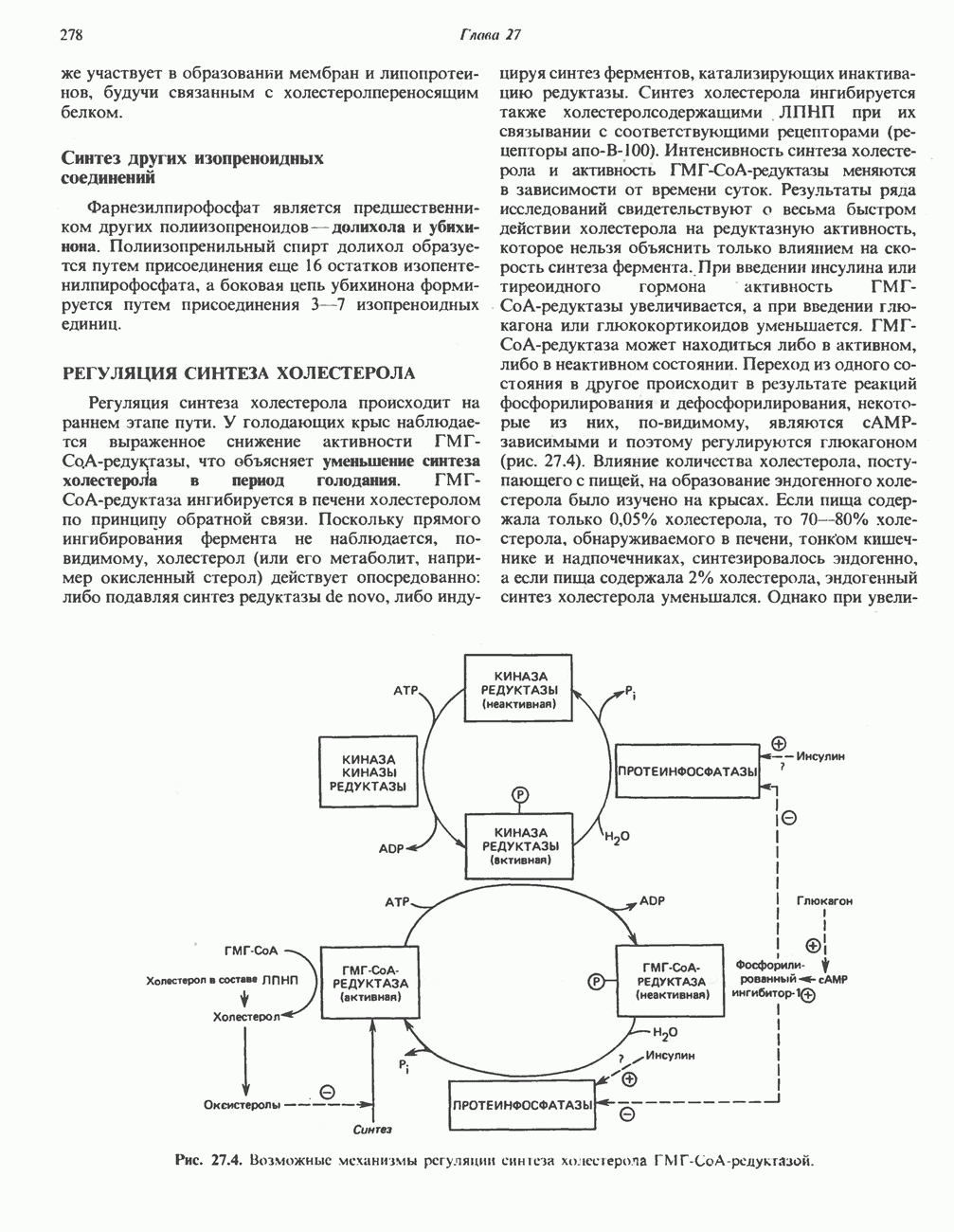
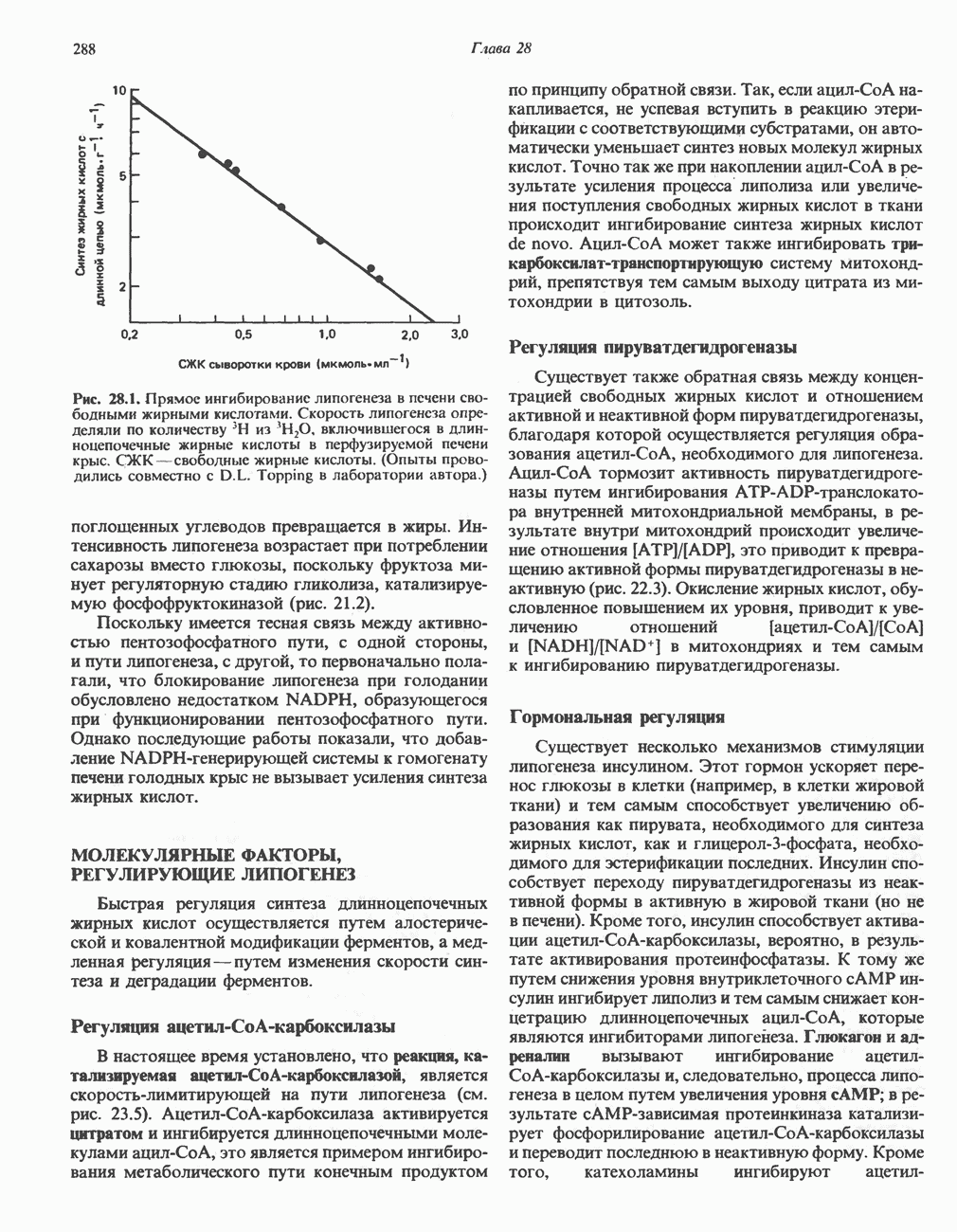
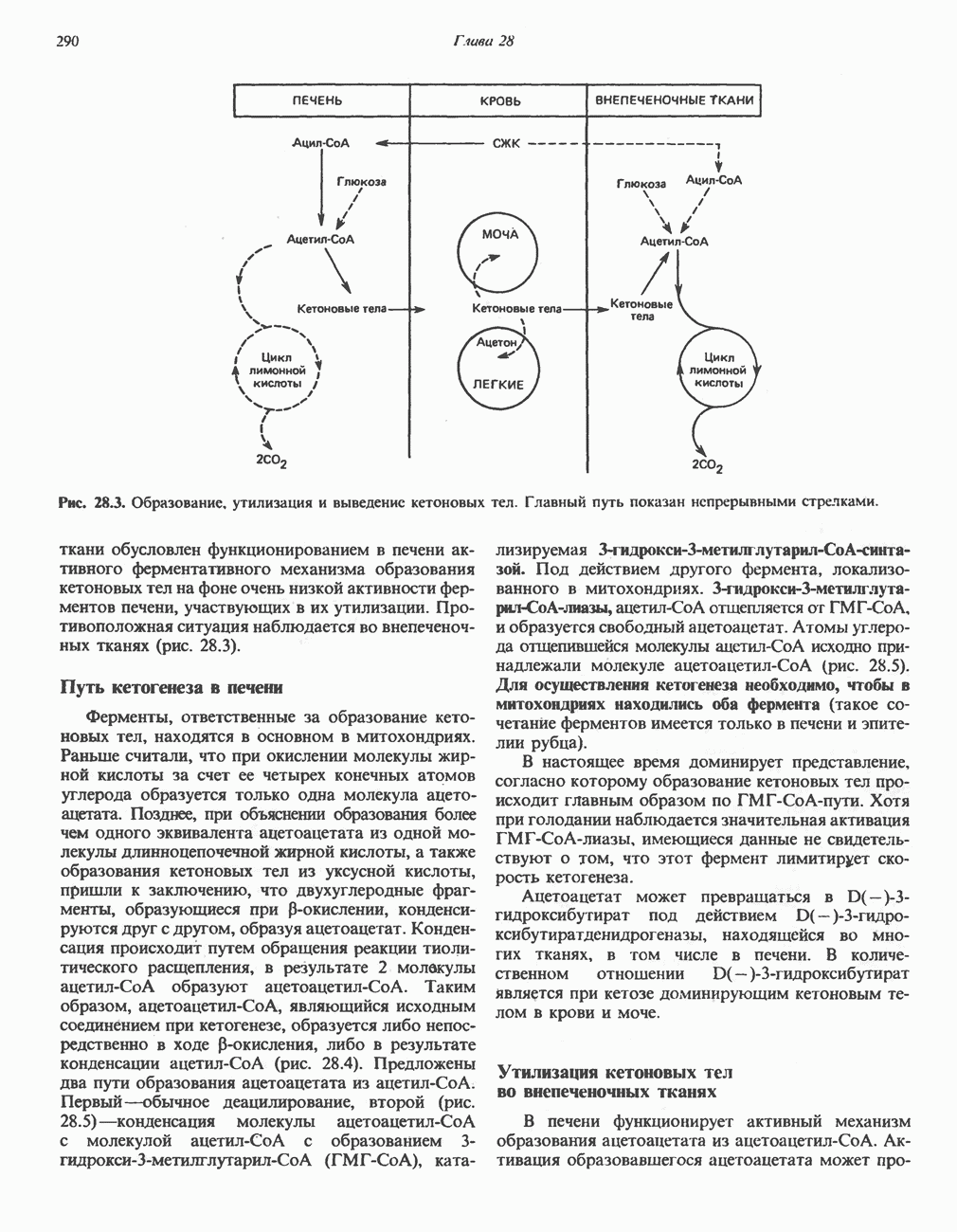
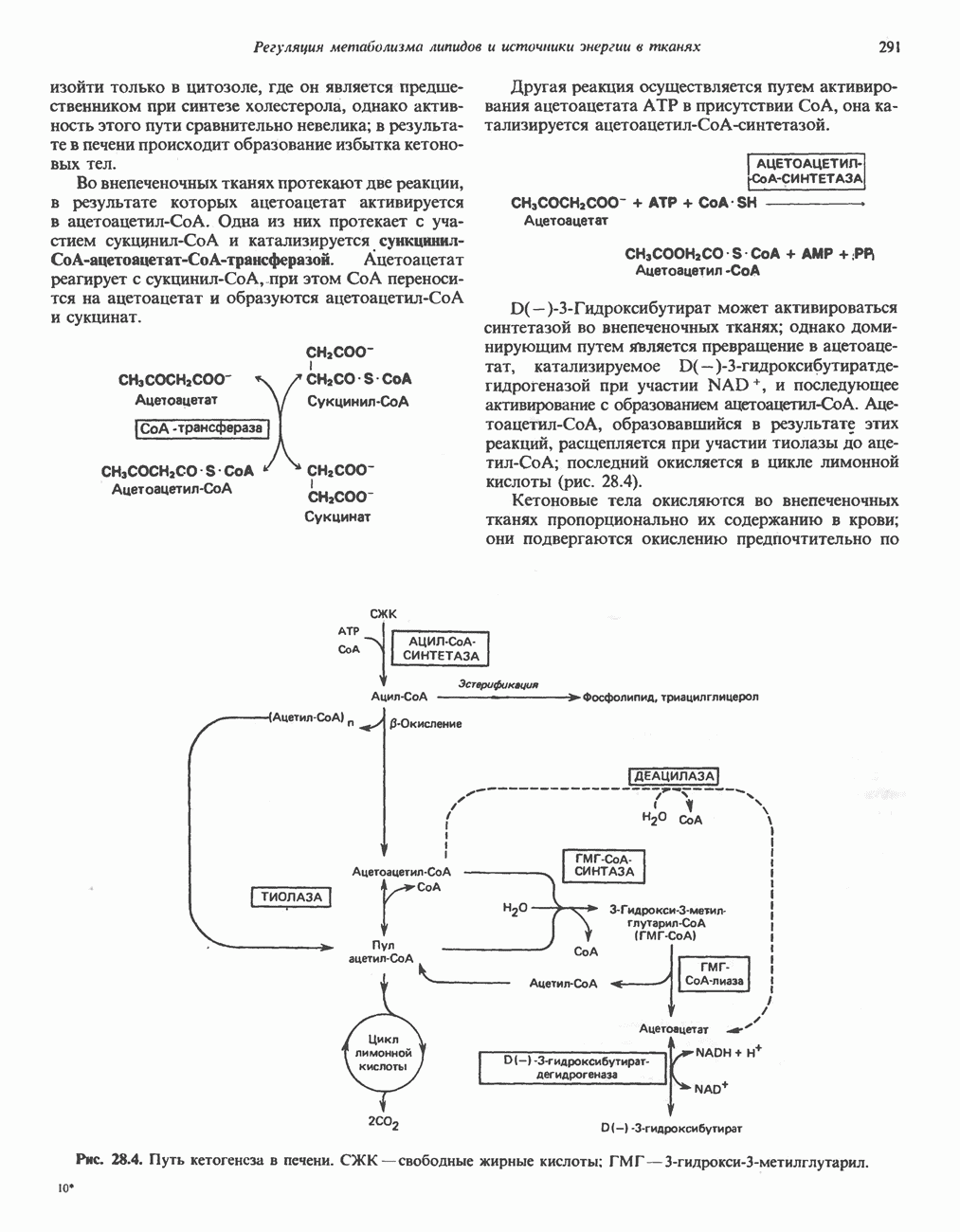
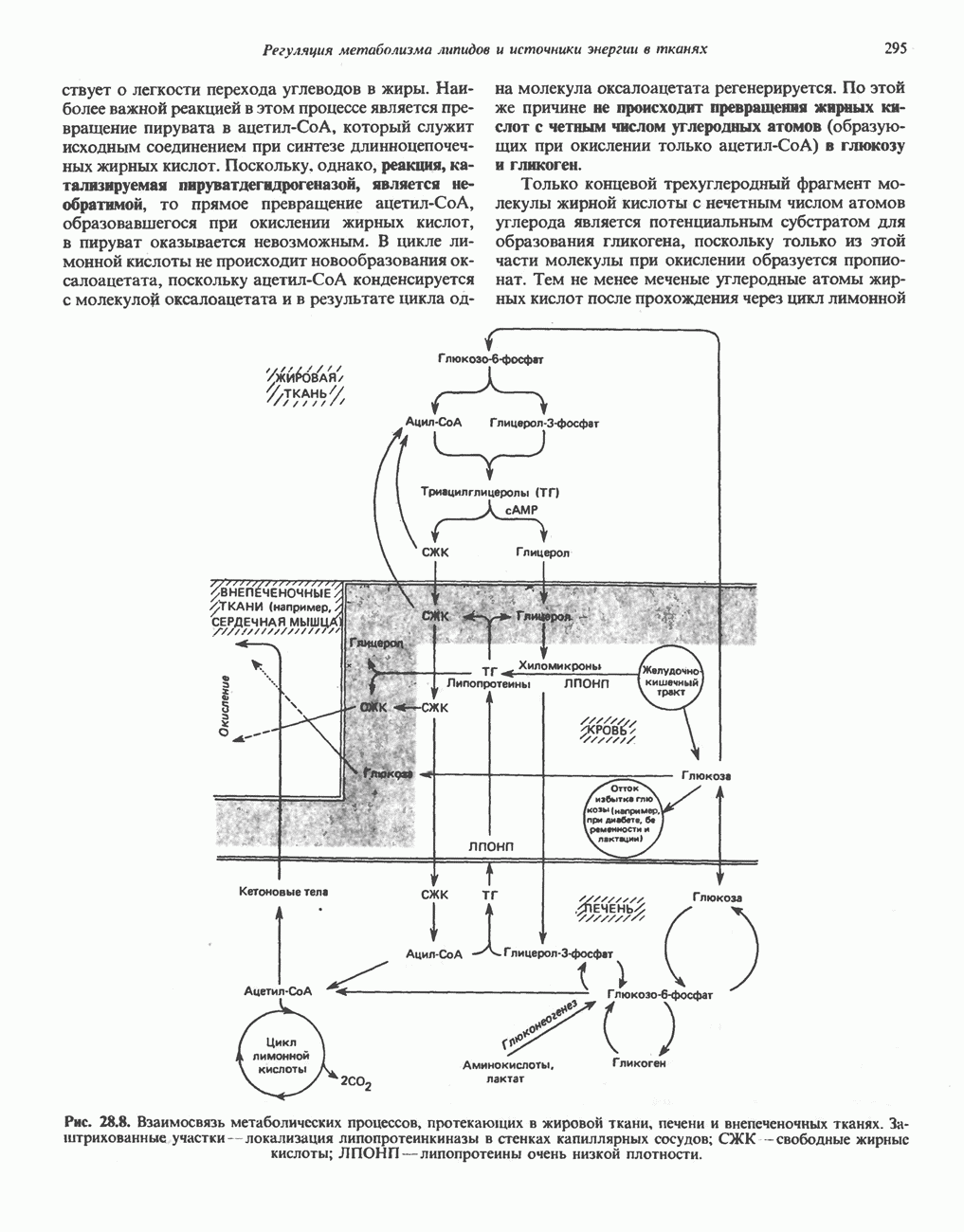
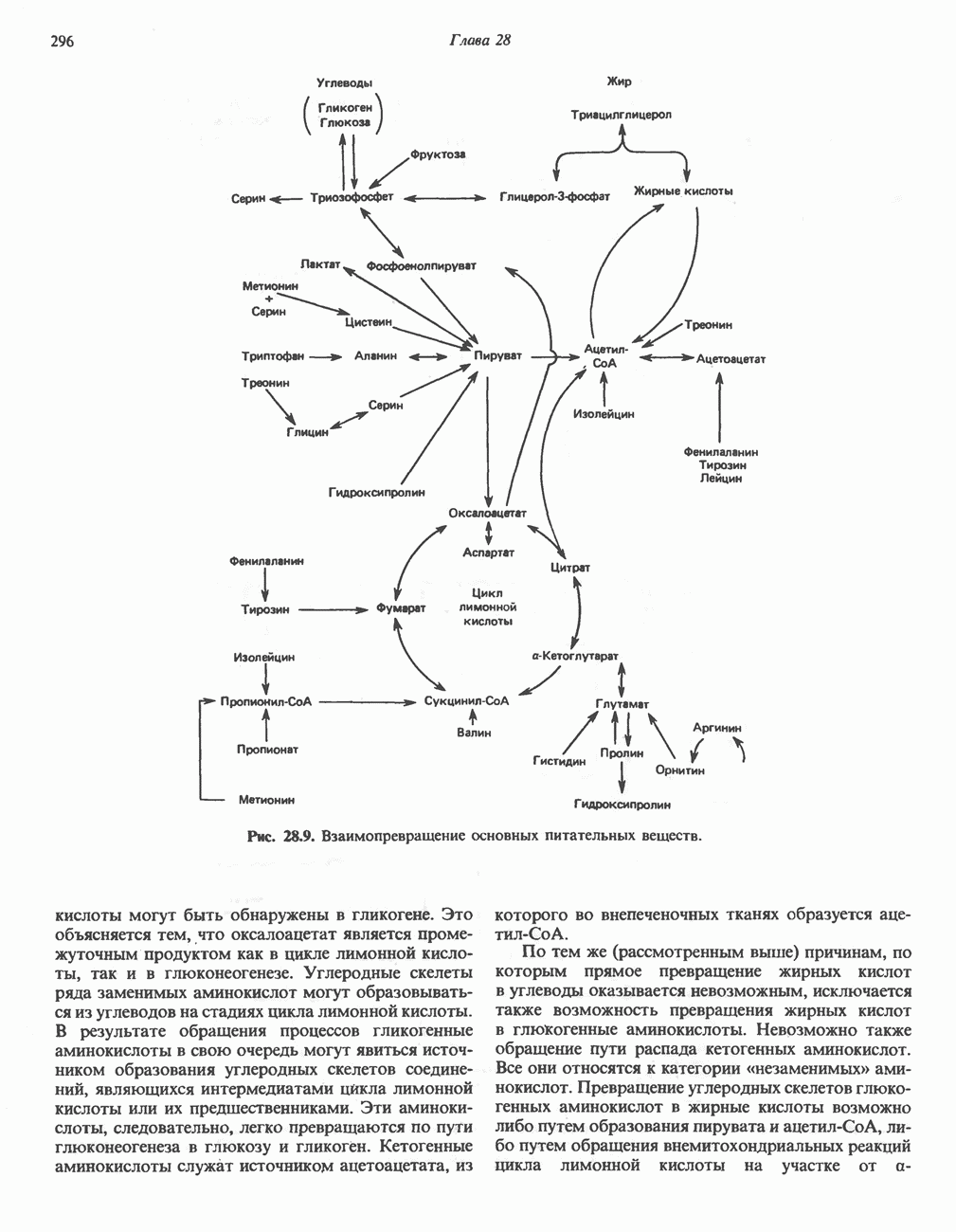
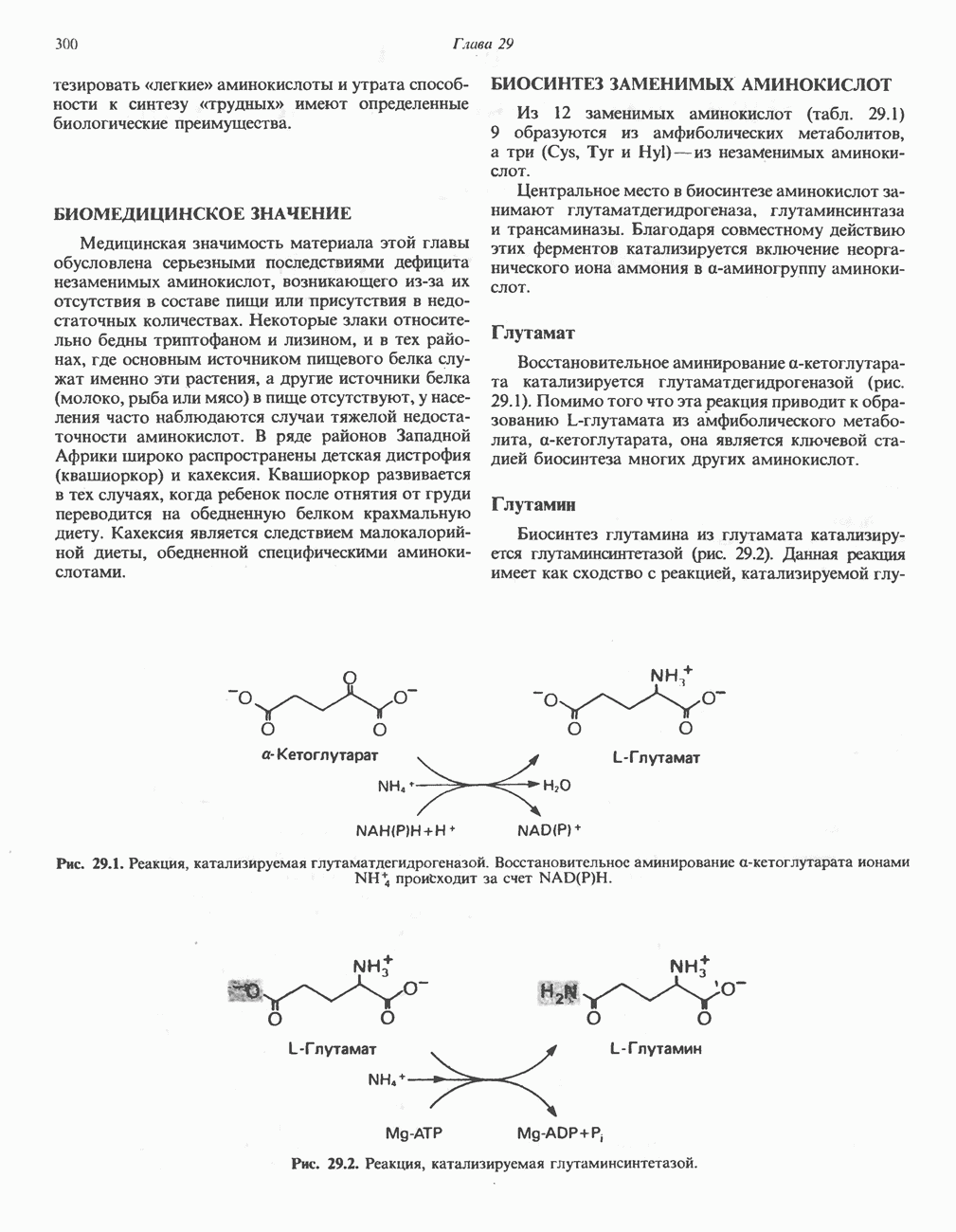
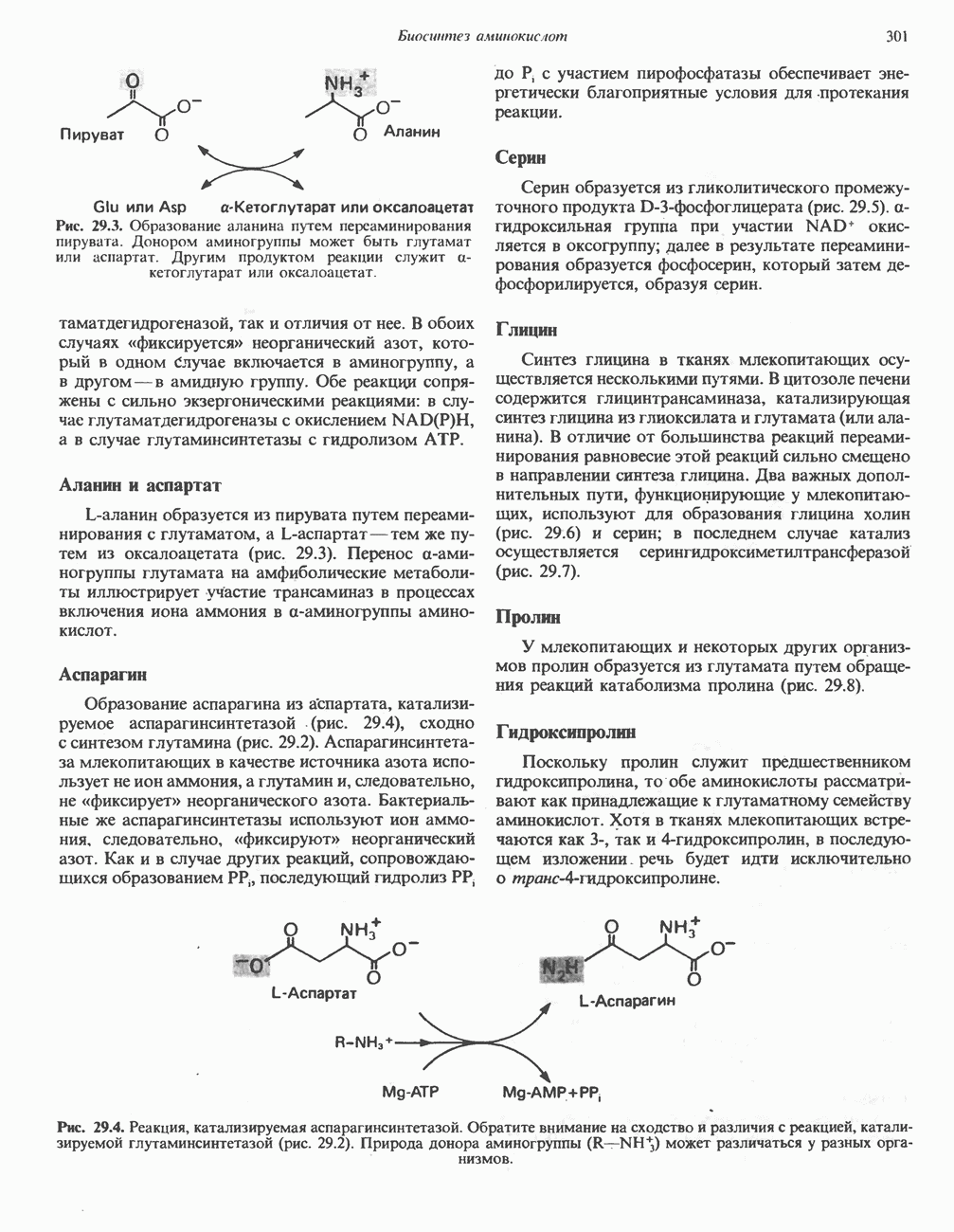
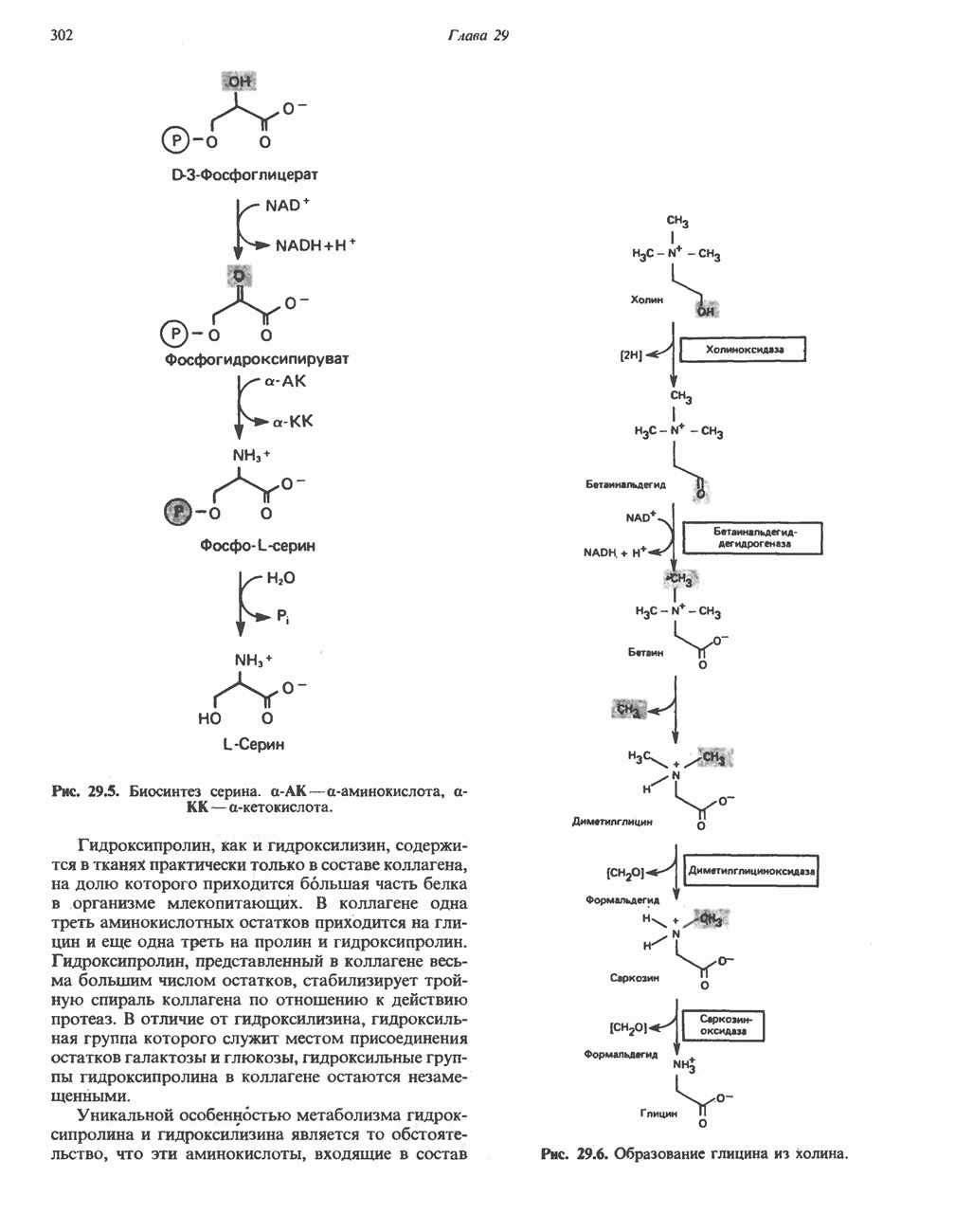
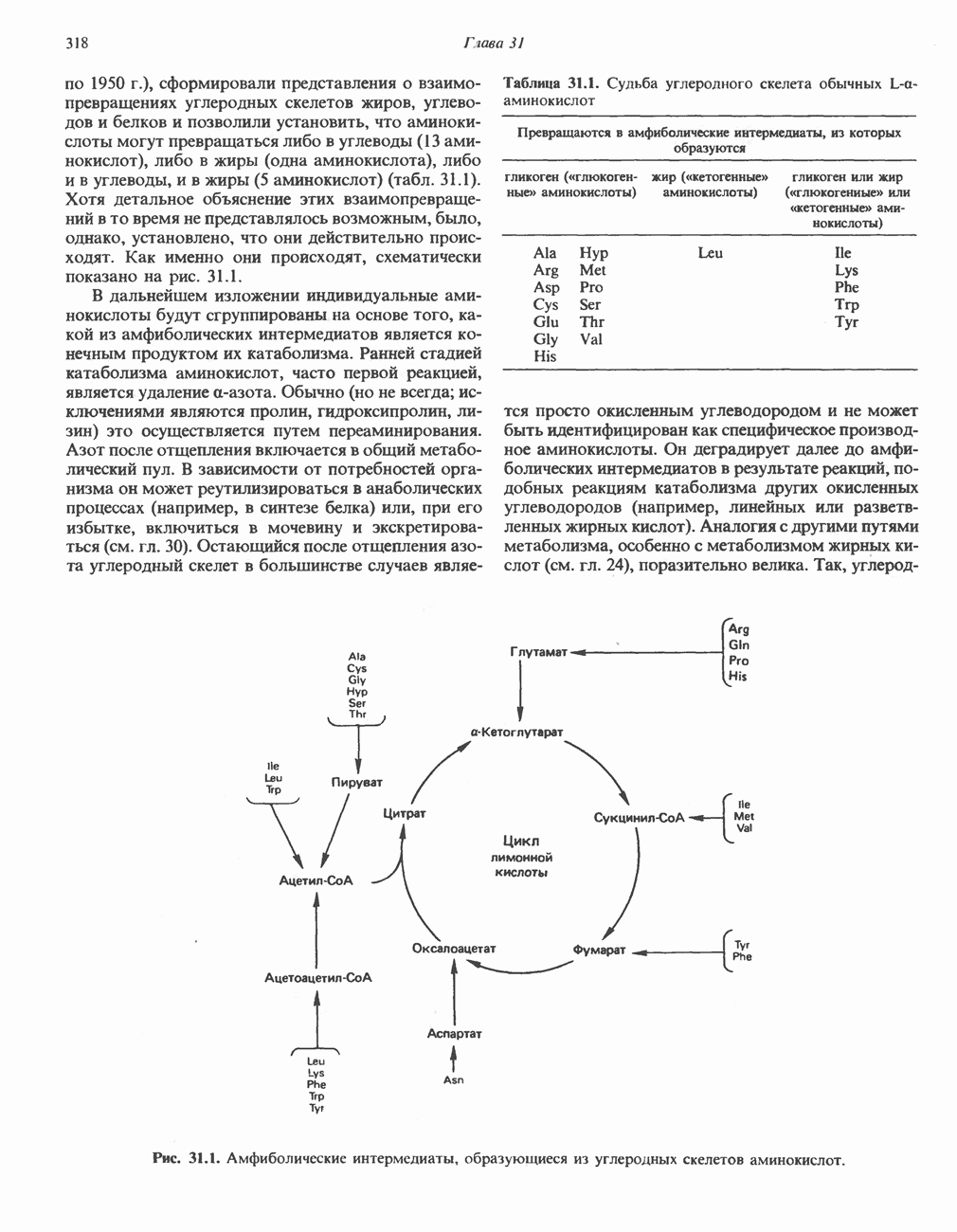
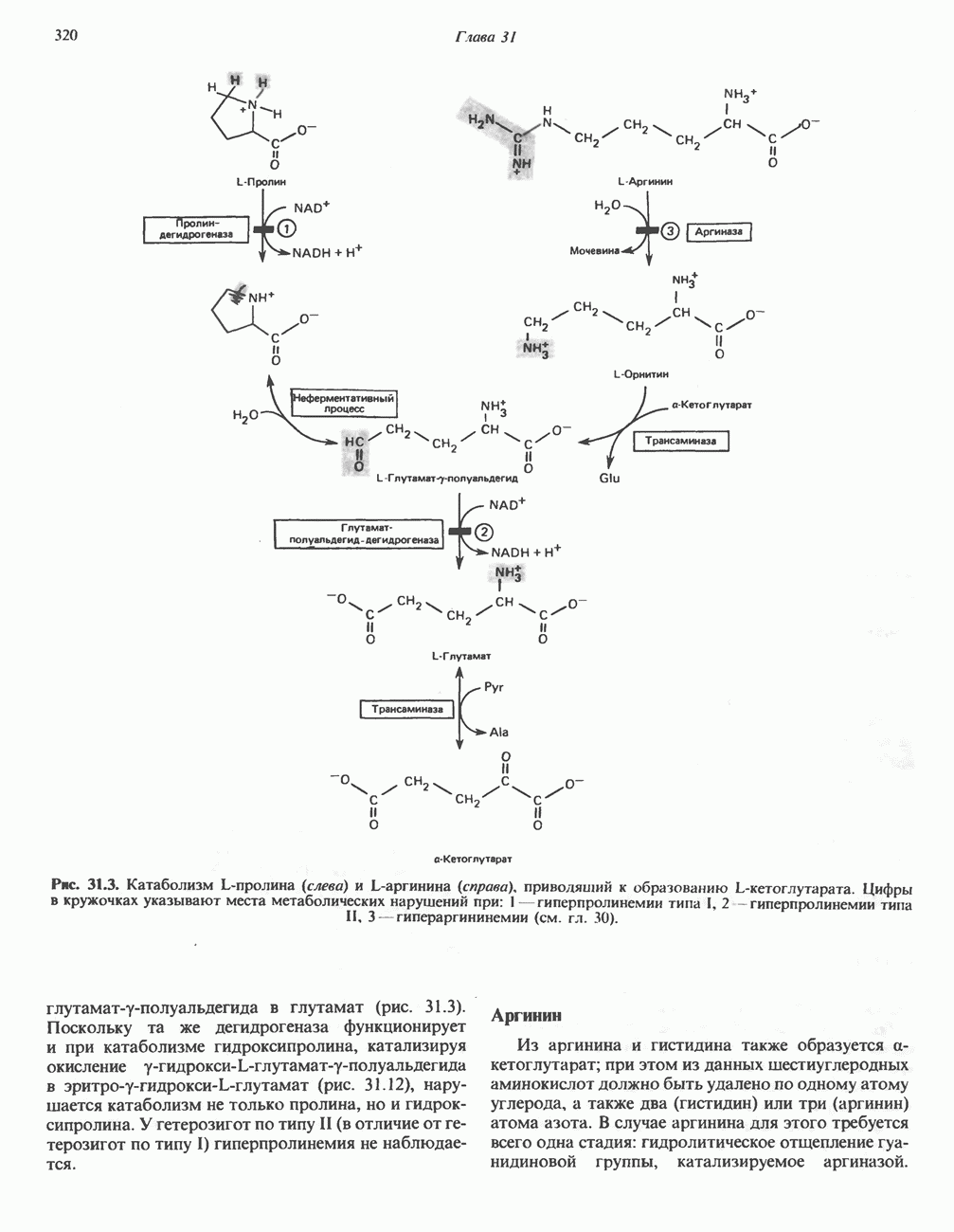
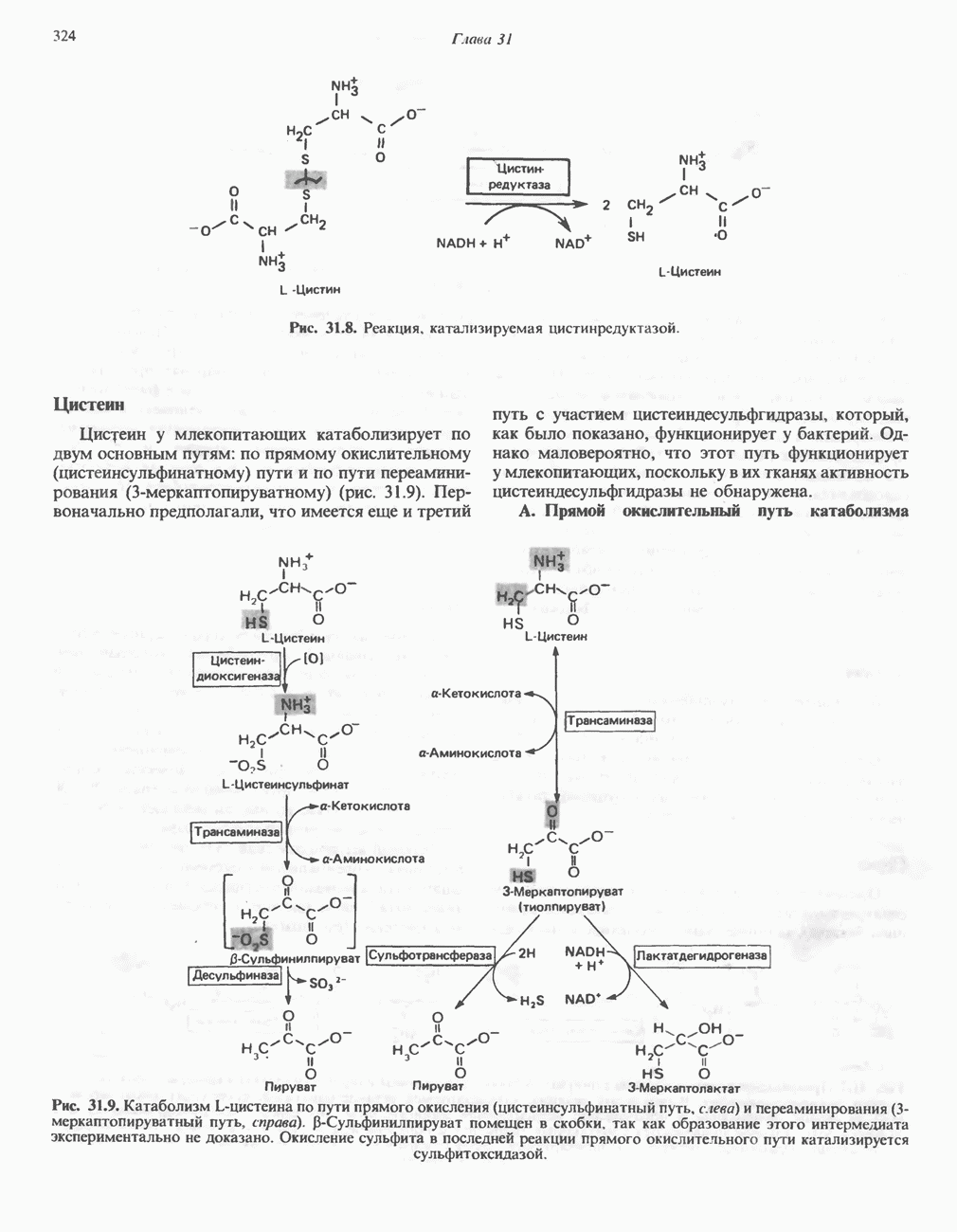
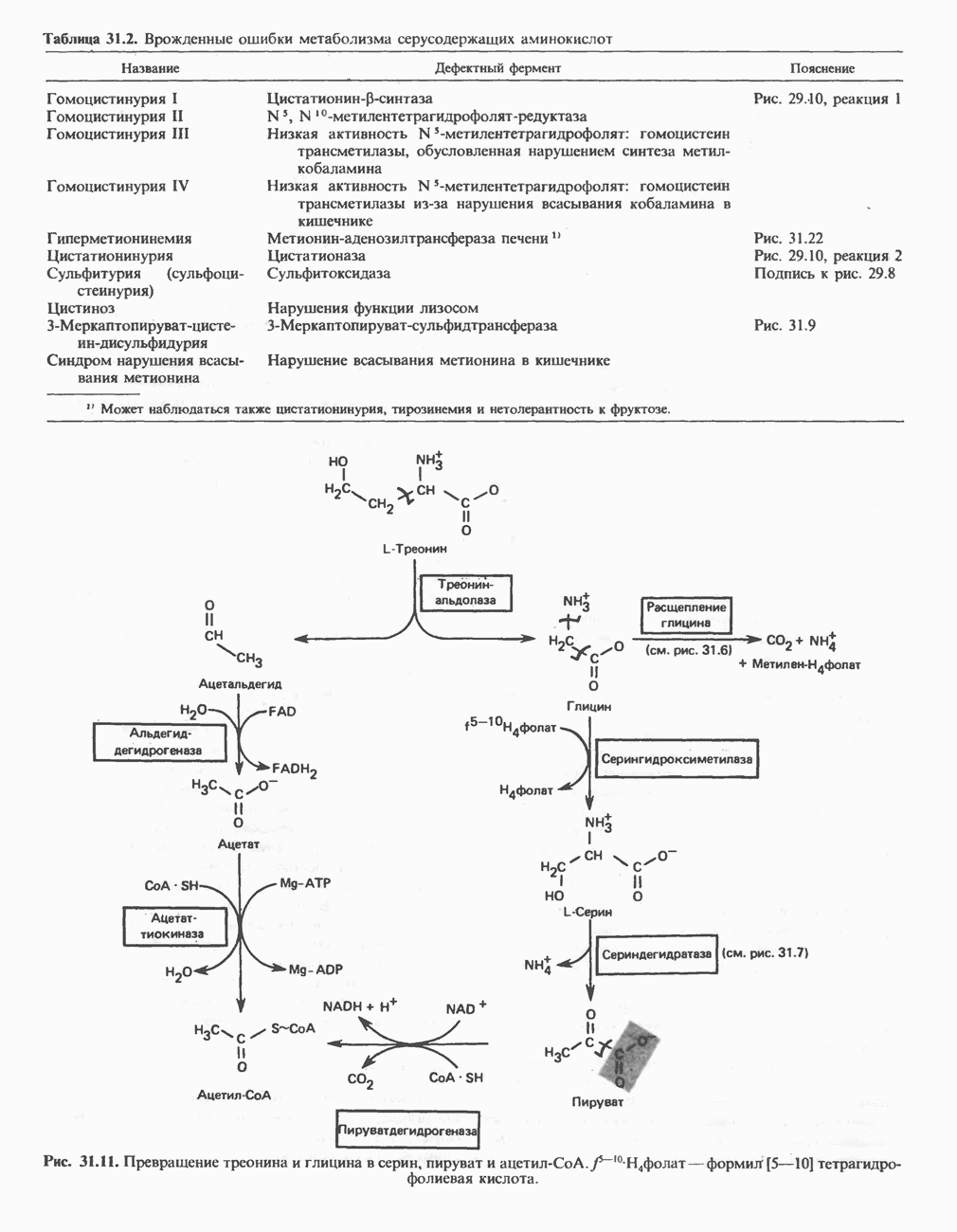
Пред.
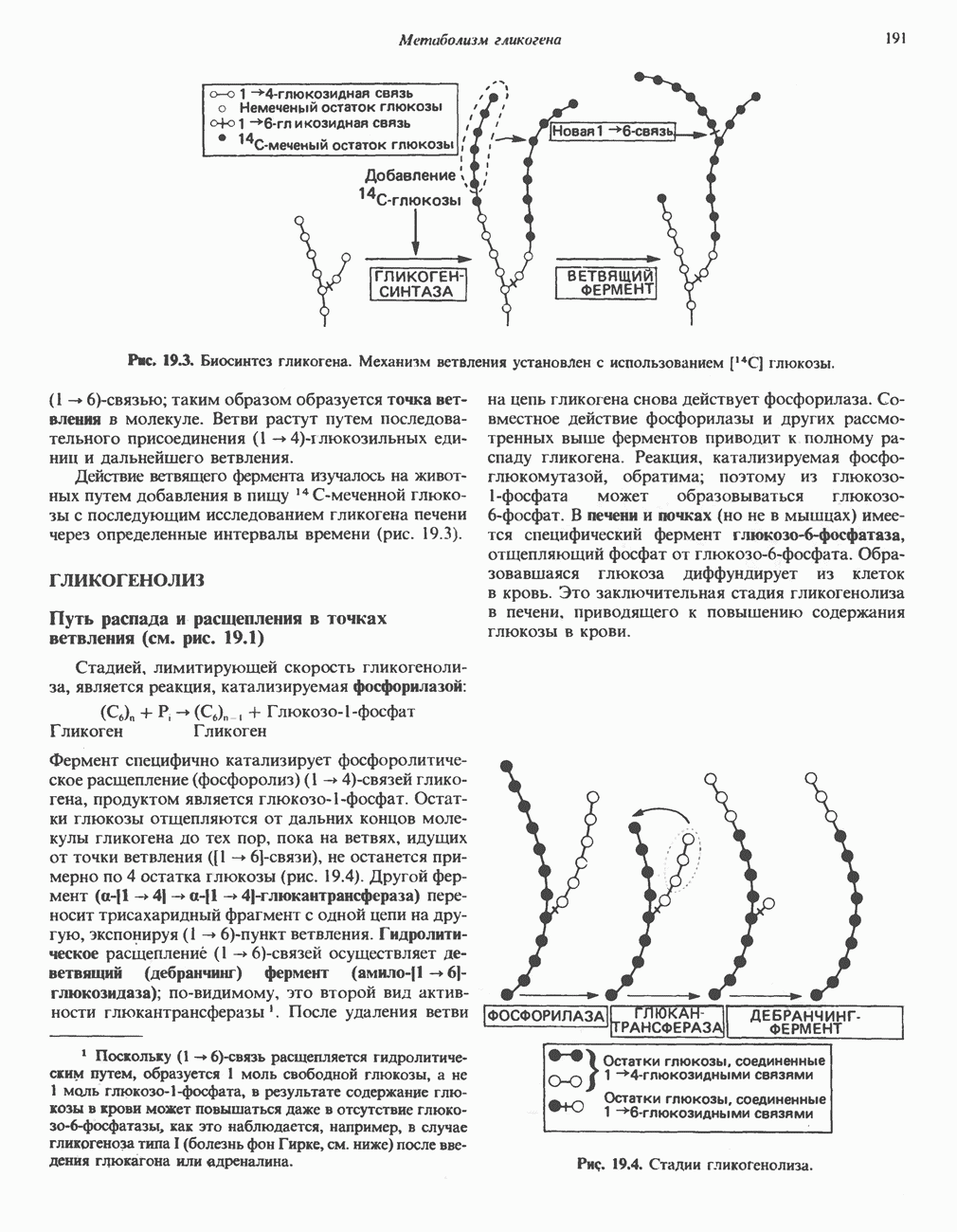
След.
Макеты страниц
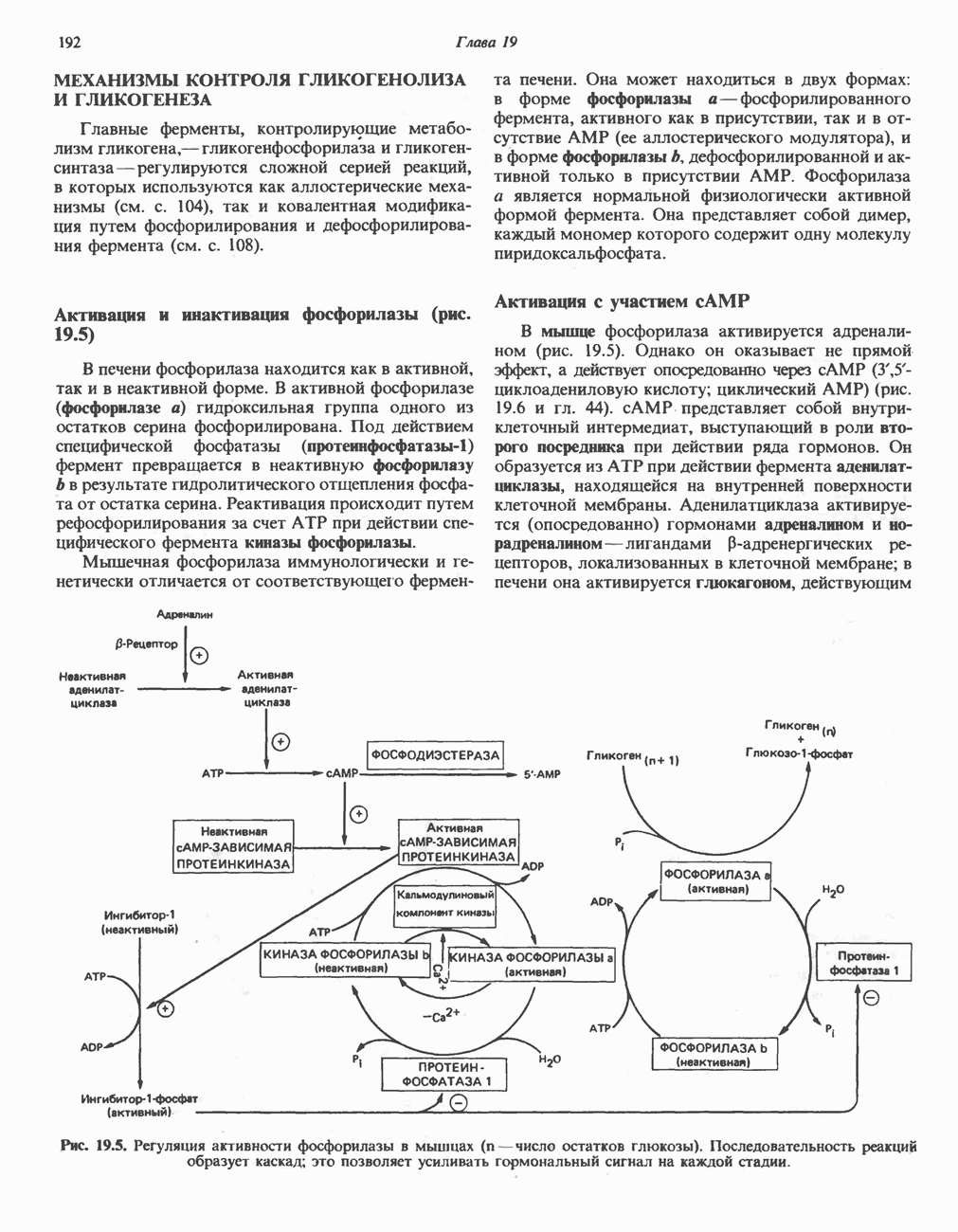
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
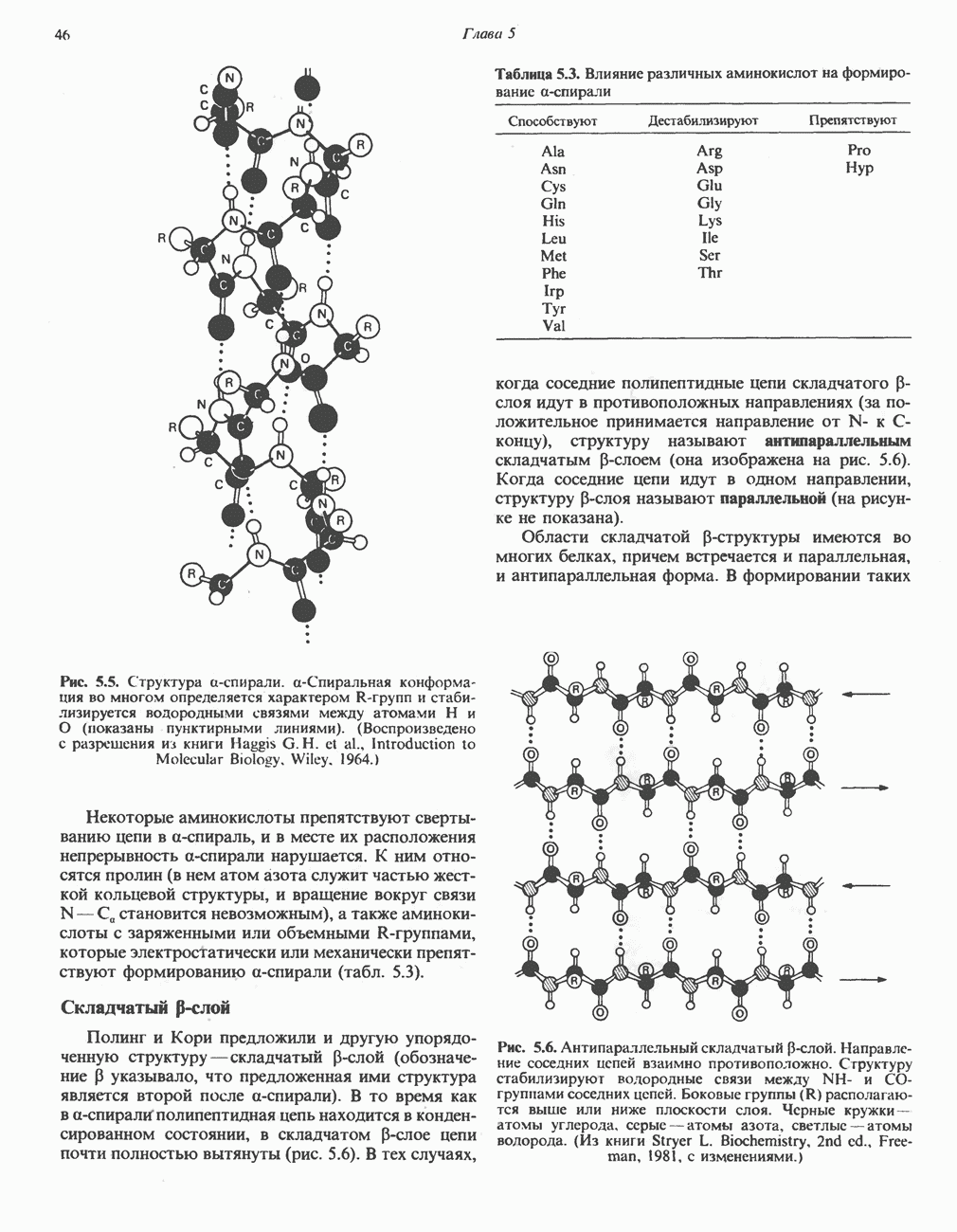
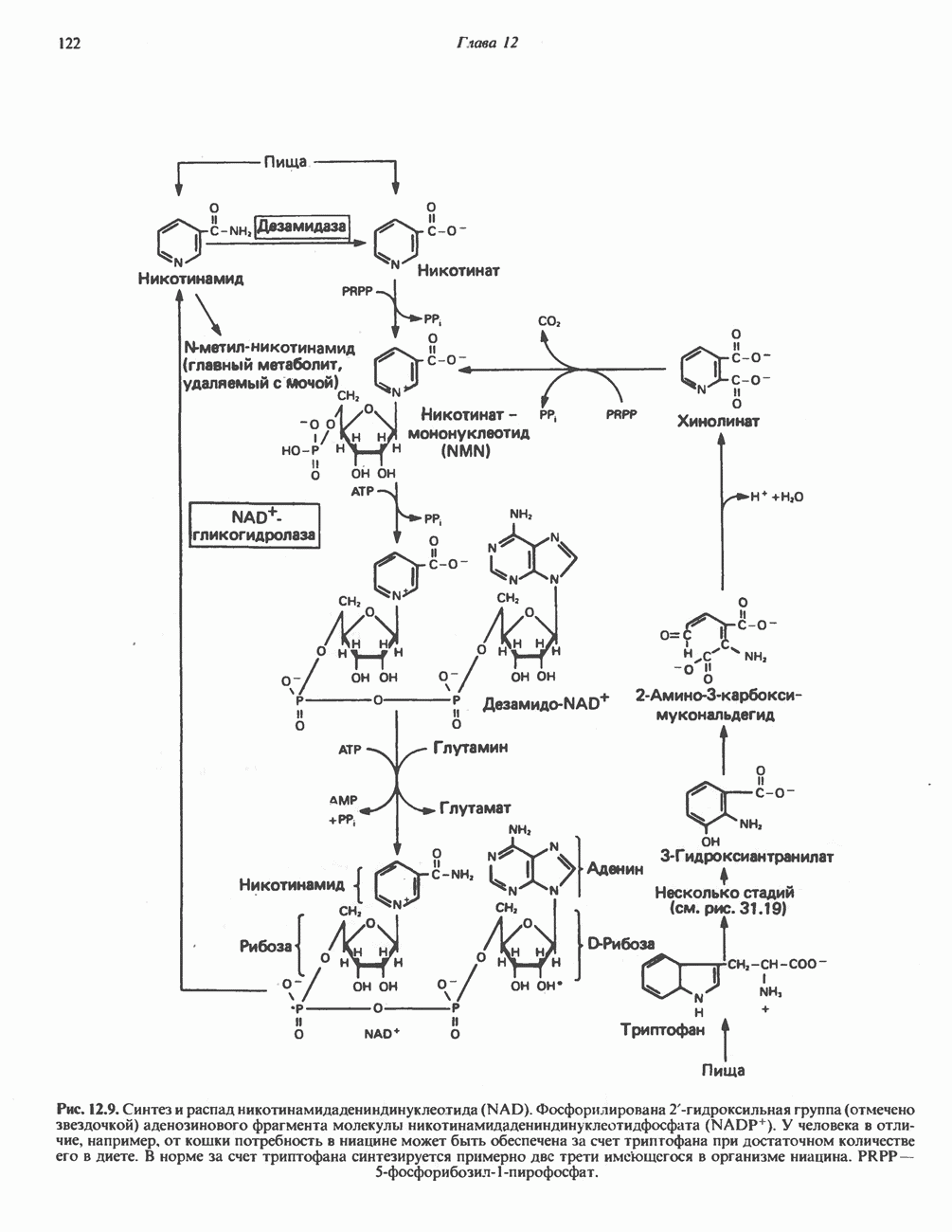
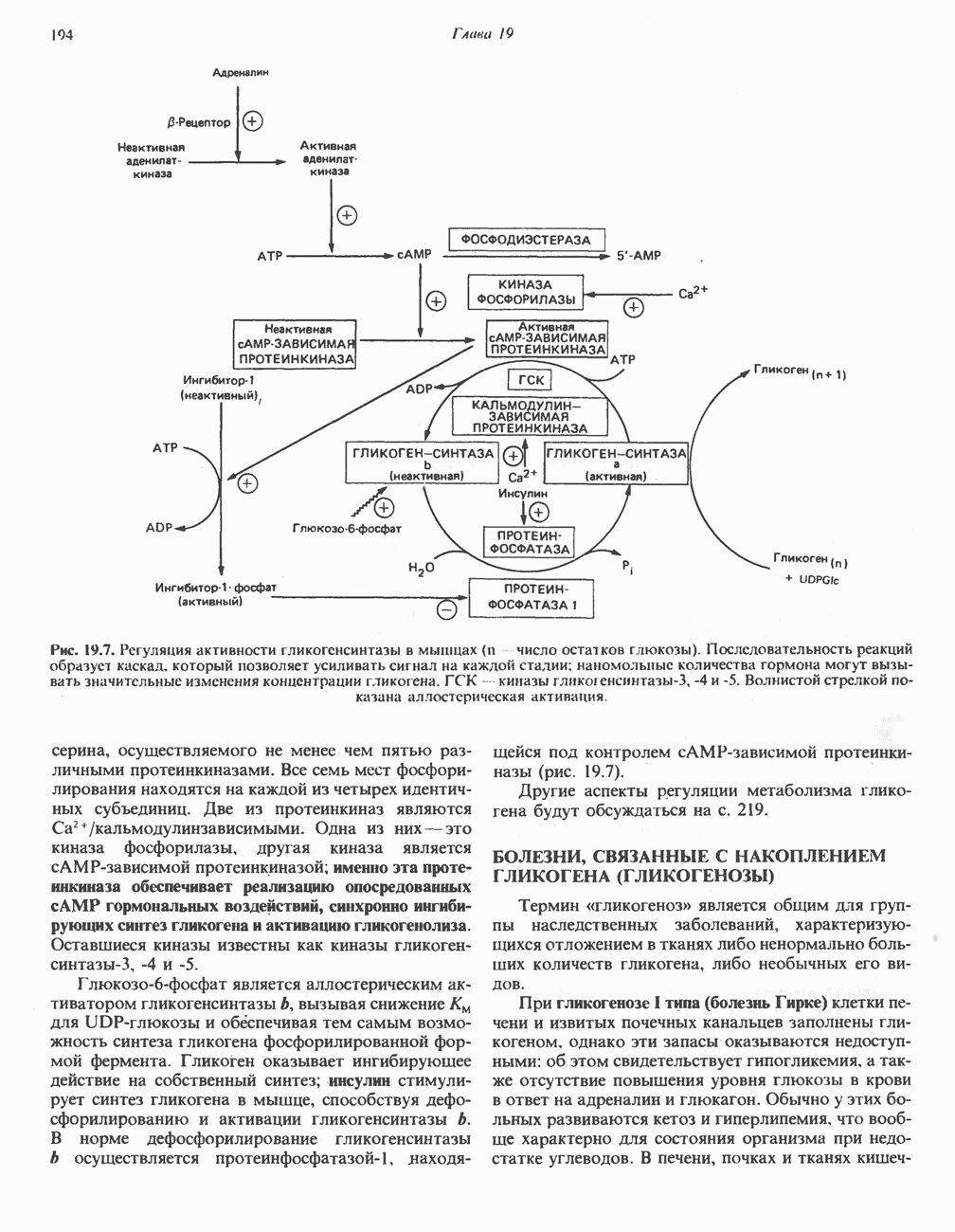
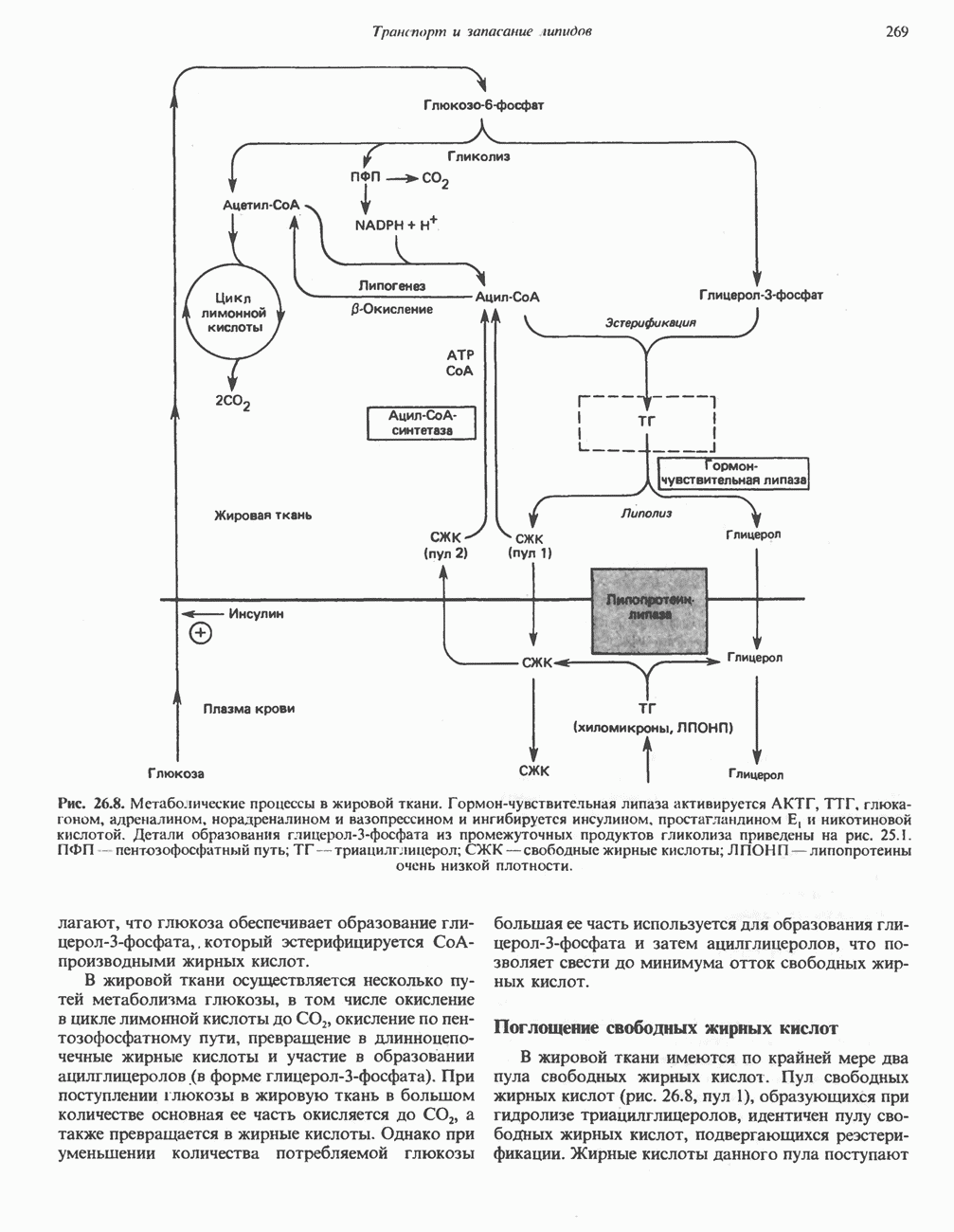
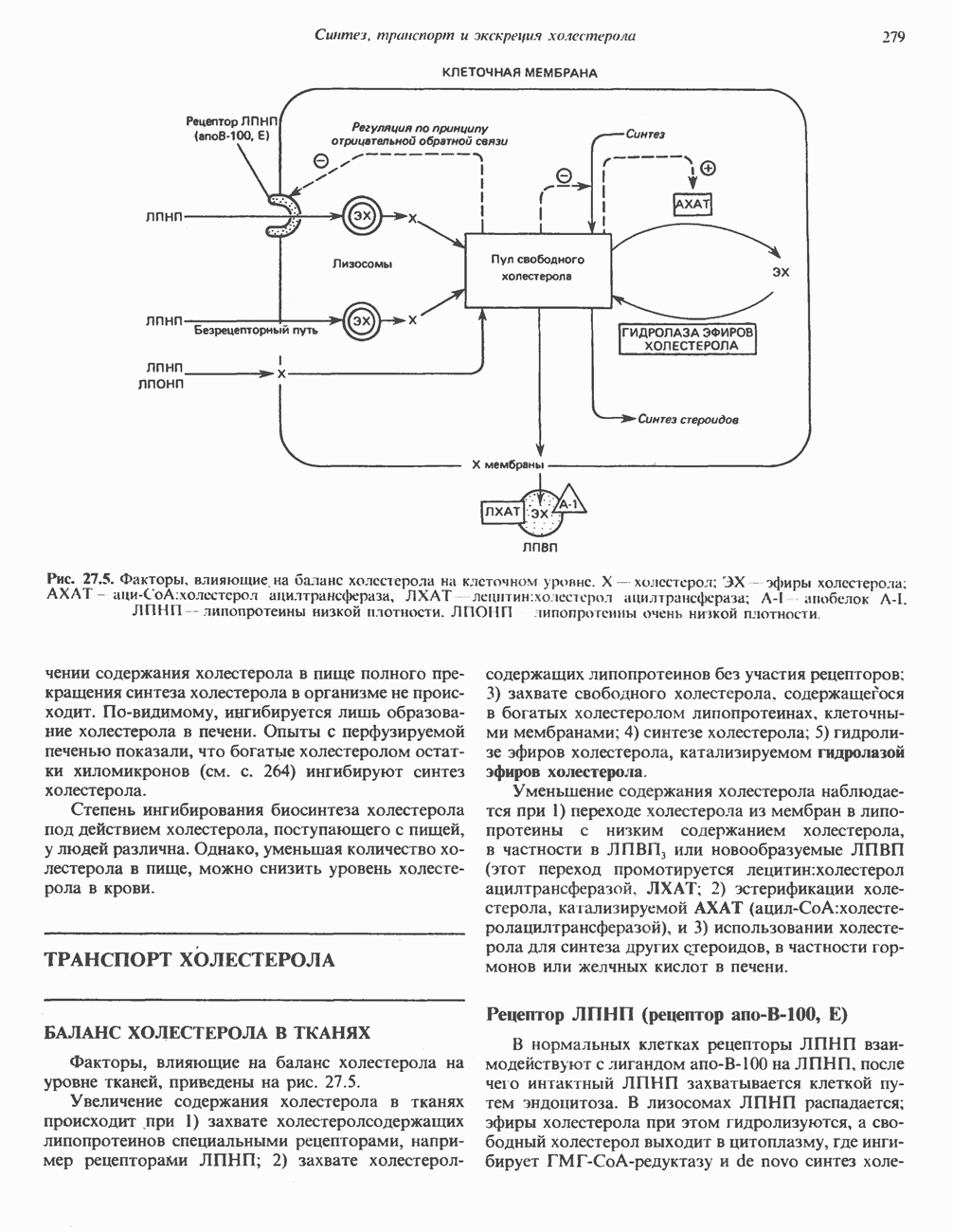
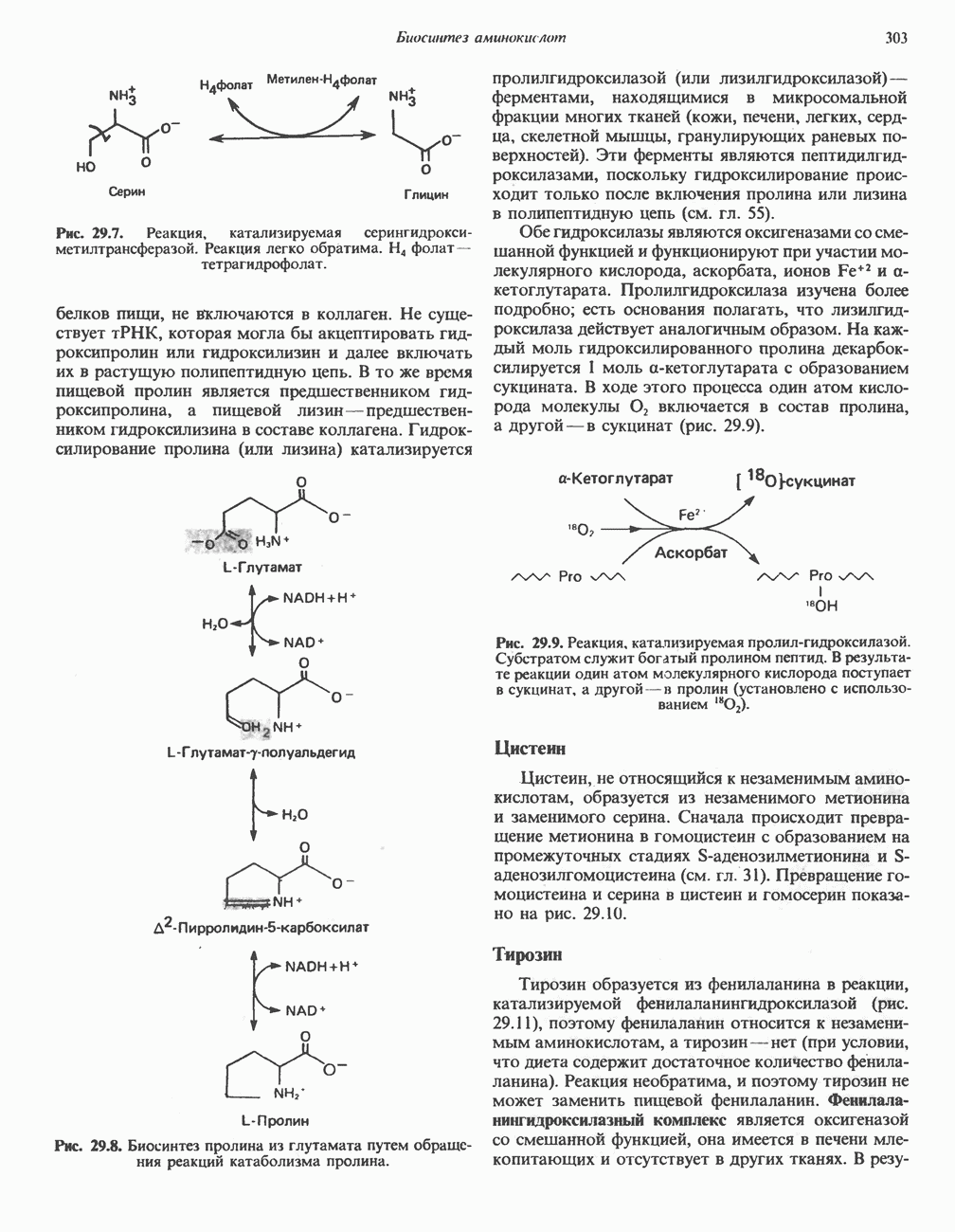
р-АЛАНИНОВЫЕ ДИПЕПТИДЫОсновная часть Р-аланина в организме человека находится в скелетных мышцах всоставе дипептида карнозина (рис. 32.3). Сходный по строению Физиологические функции БиосинтезКарнозин образуется из Р-аланина и
Ансерин образуется из карнозина (донором метильной группы служит
В процессе реакции образуется связанный с ферментом
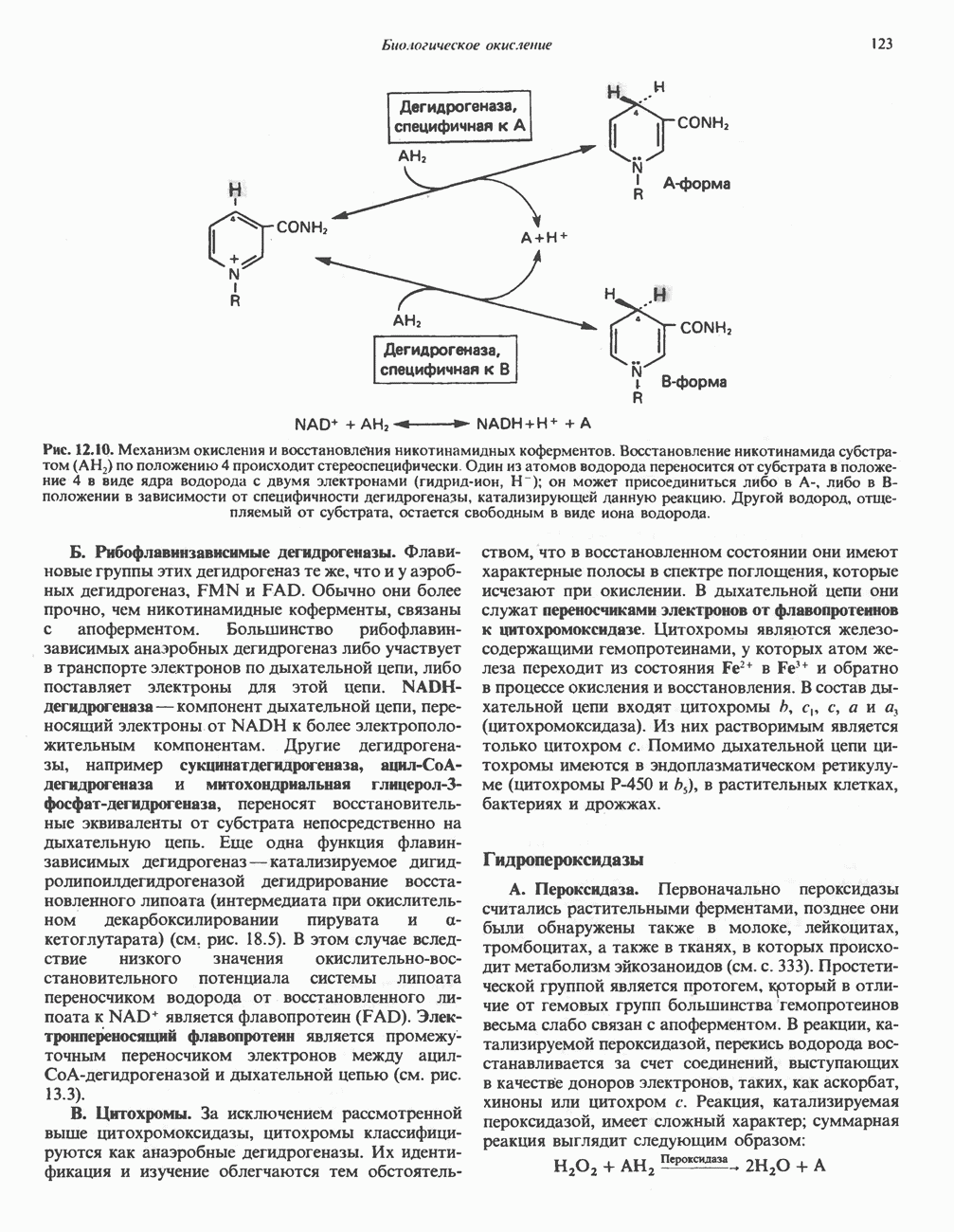
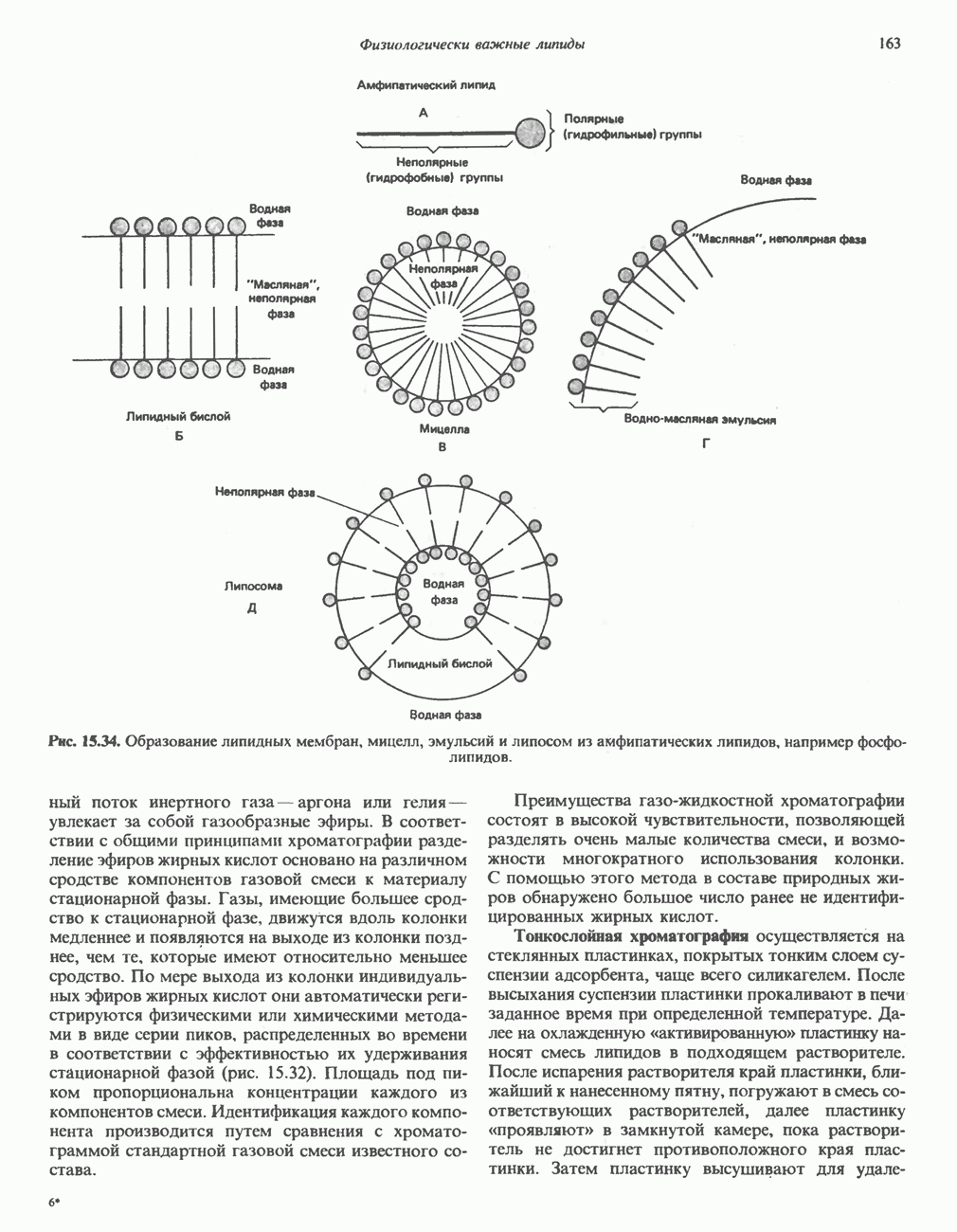
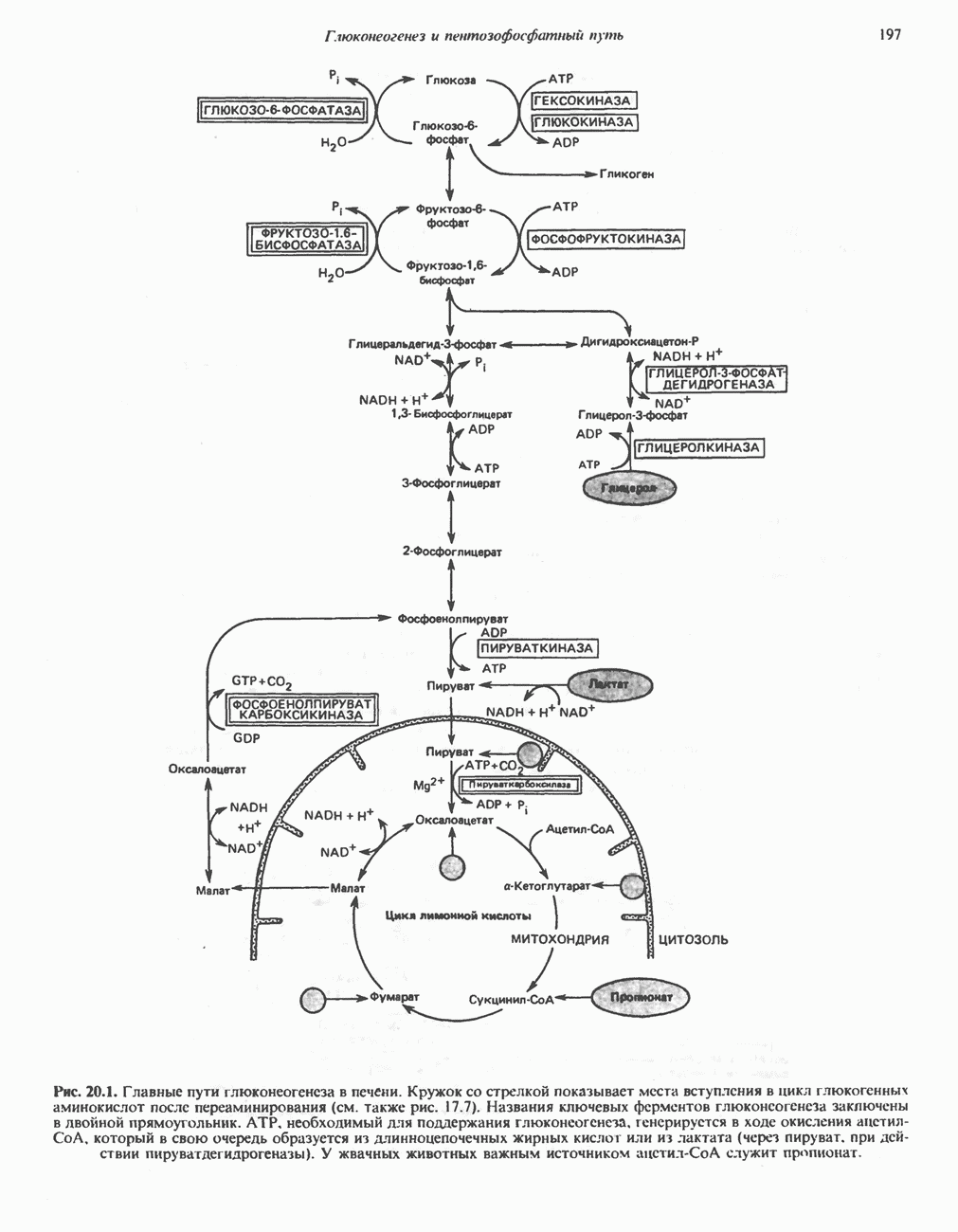
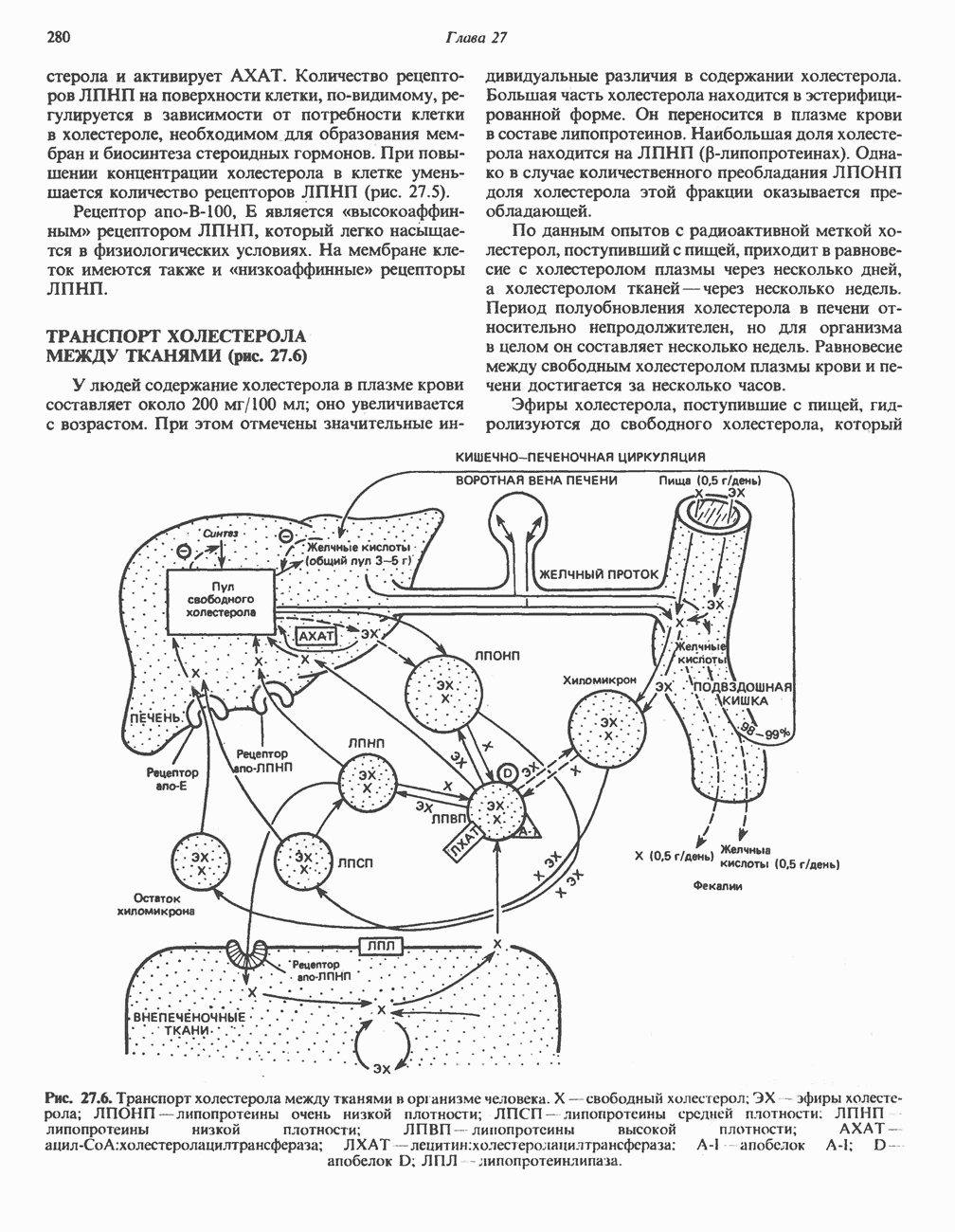
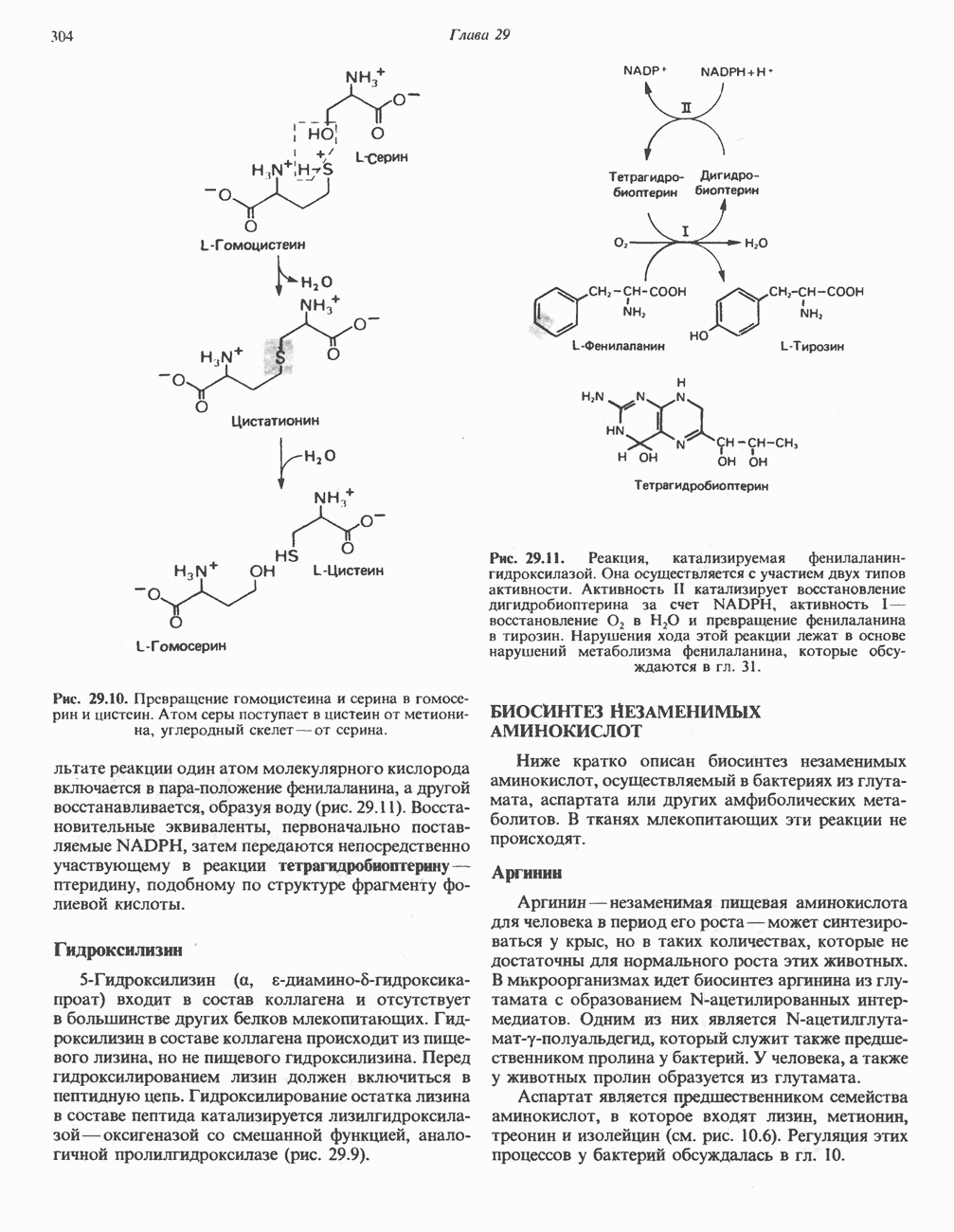
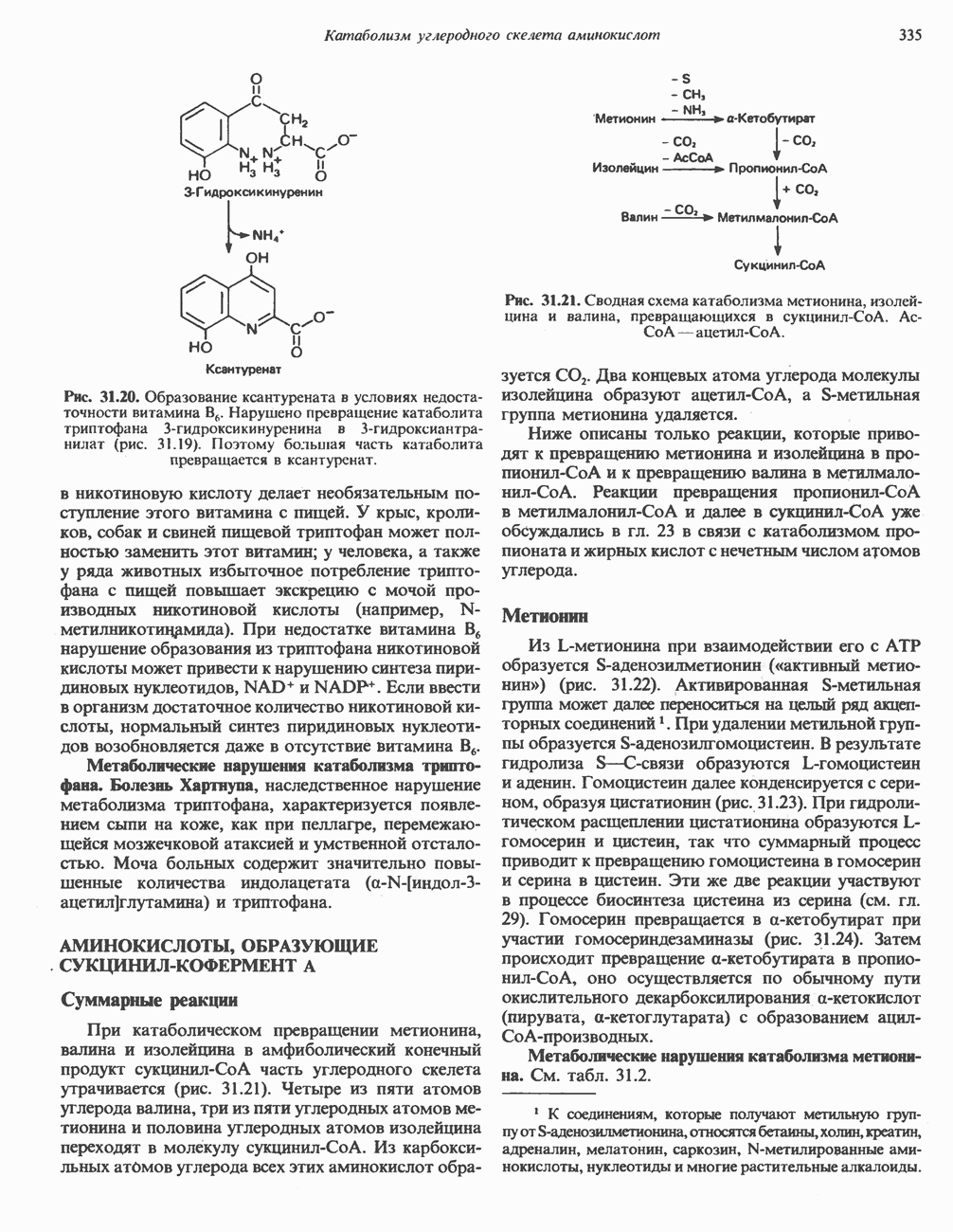
Рис. 32.3. Соединения, родственные гистидину. В прямоугольник заключены фрагменты, образовавшиеся не из гистидина. На рисунке опечатка: следует читать не «эрготионеим», а «эрготионеин».) Абсорбция b-аланиновых дипептидовВ ткань почек и энтероциты кишечника карнозин и (З-аланин поступают при участии мембранных переносчиков, которые отличают один субстрат от другого и оба субстрата от других дипептидов. Способность переносчиков отличать (З-аланин от карнозина используется при диагностике болезни Хартнуна — наследственного нарушения транспорта некоторых нейтральных а-аминокислот (см. гл. 31). У пациентов с болезнью Хартнупа содержание в крови гистидина соответствует норме после приема с пищей карнозина, но оказывается ниже нормы после приема КатаболизмКарнозин гидролизуется с образованием Р-аланина и ГомокарнозинГомокарнозин (у-аминобутирил-Ь-гистидин, рис. 32.3) - дипептид центральной нервной системы, структурно и метаболически родственный карнозину; его физиологическая функция пока не ясна. Этот дипептид, образованный Недостаточность сывороточной карнозиназыЭто наследуемое, предположительно аутосомнорецессивное заболевание характеризуется стойкой карнозинурией и периодической карнозинемией. Карнозинурия не прекращается даже при исключении карнозина из диеты. ГомокарнозинозПри этом состоянии содержание гомокарнозина повышается в ткани мозга и в спинномозговой жидкости, но не в плазме и моче. Описан один случай заболевания, СЕРИНБольшая часть серина в фосфопротеинах находится в форме О-фосфосерина. Серин участвует в синтезе сфингозина (см. гл. 25). Он участвует также в синтезе пуринов и пиримидинов. ТРЕОНИНТреонин в реакциях переаминировання не участвует; его МЕТИОНИНМетионин как донор метильной группы уже рассматривался в гл. 31. В форме ЦИСТЕИНСодержащийся в моче сульфат практически полностью образуется при окислении ГИСТИДИНГистамин образуется из гистидина путем декарбоксилирования; в тканях млекопитающих эта реакция катализируется декарбоксилазой ароматических К числу находящихся в организме гистидин
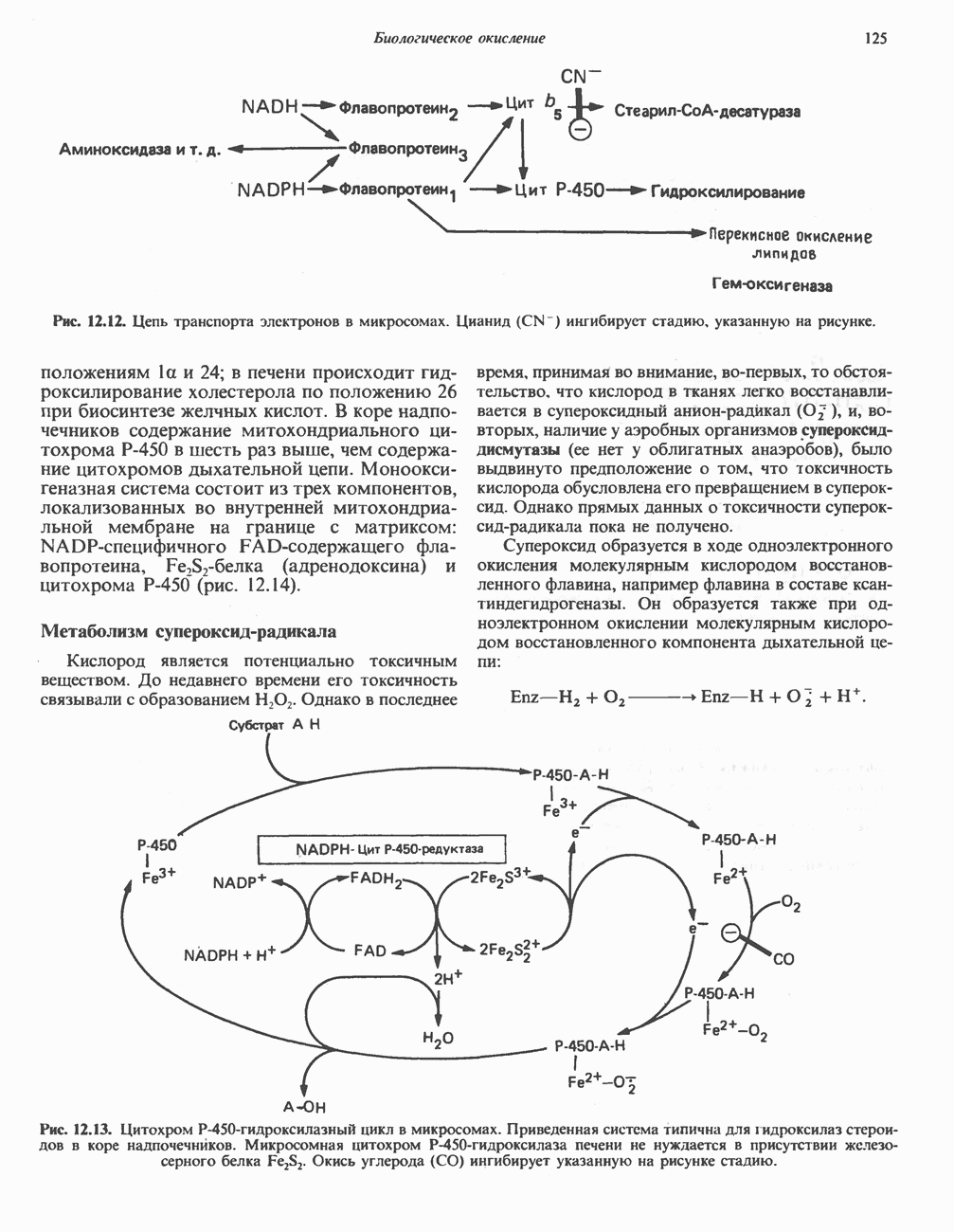
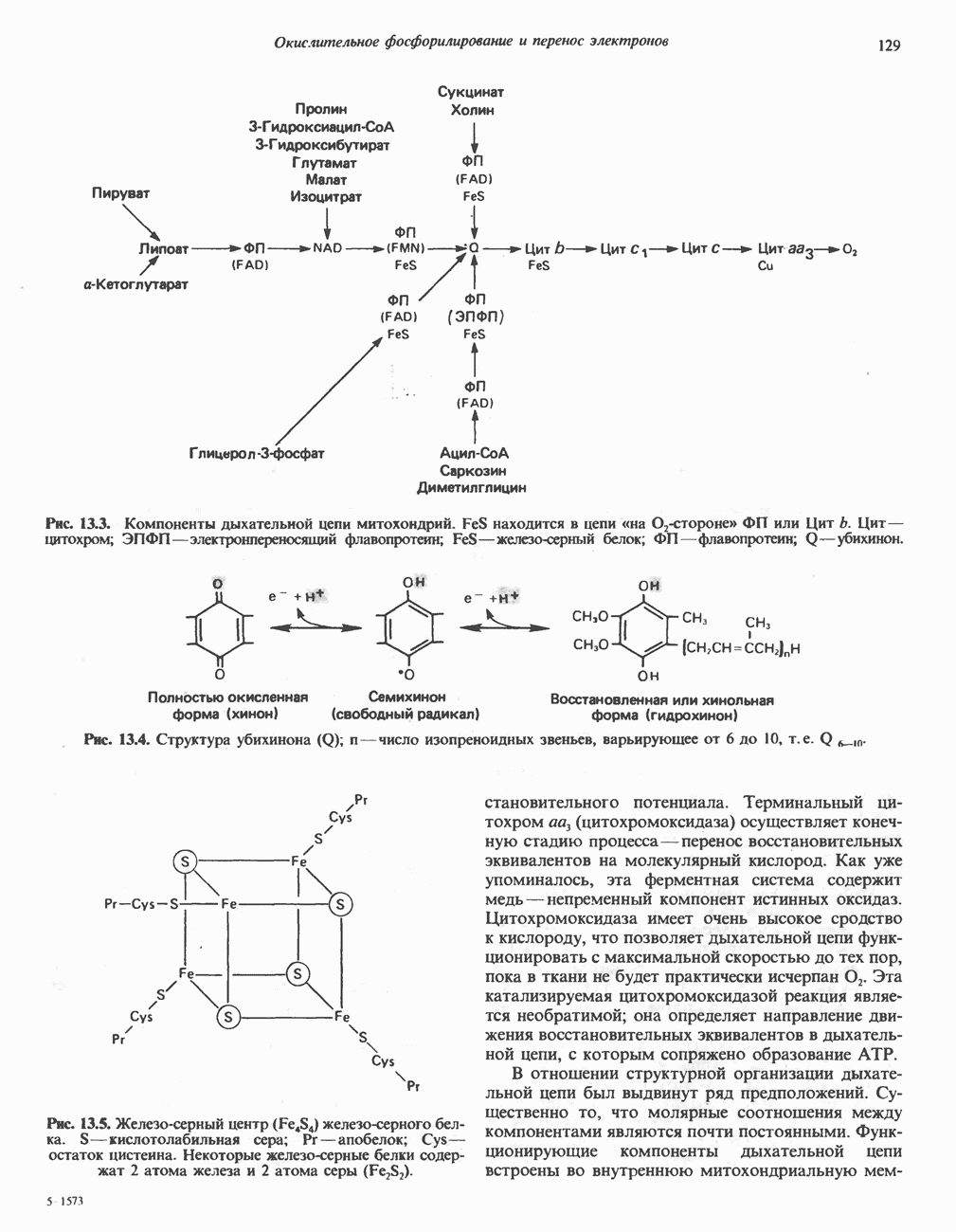
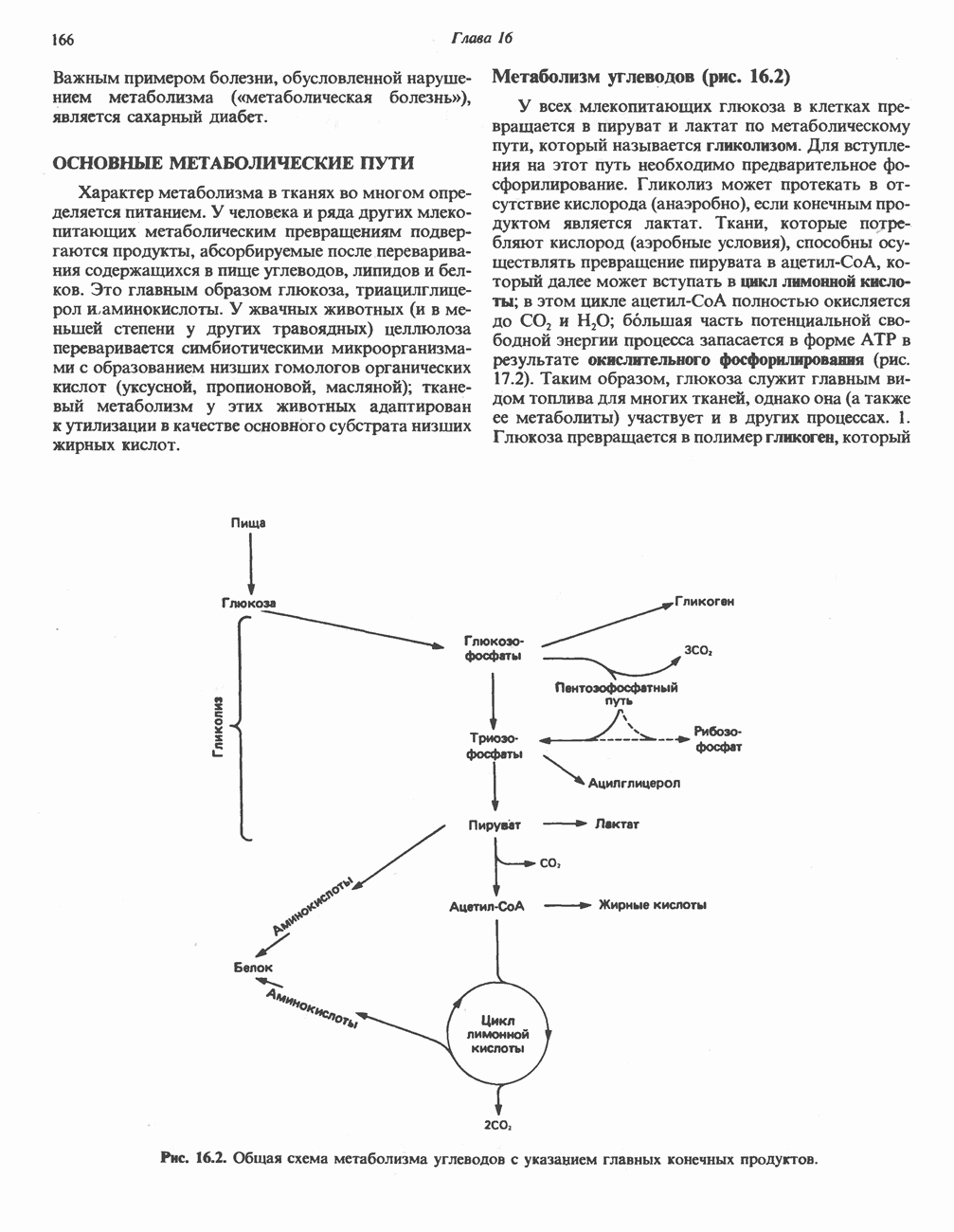
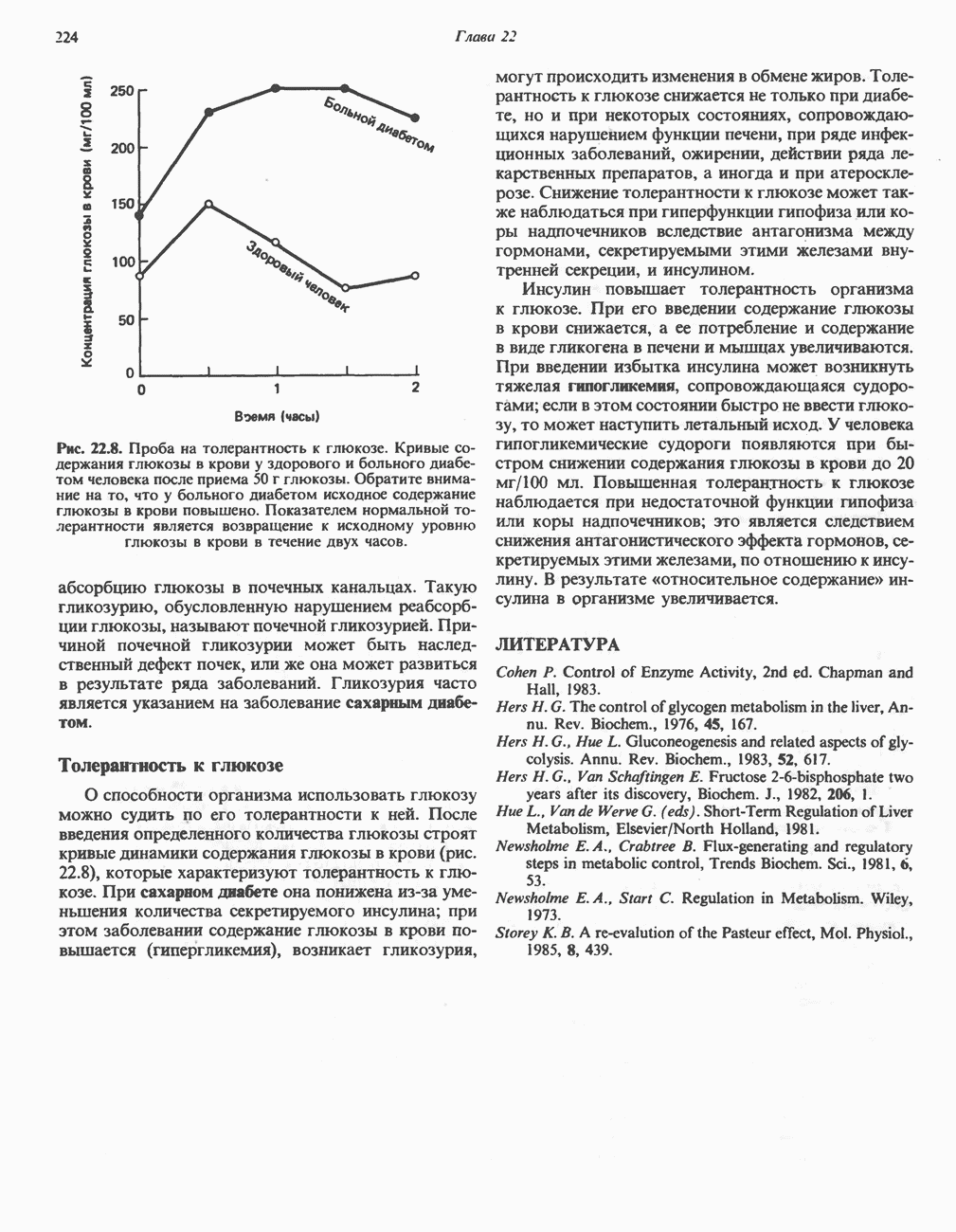
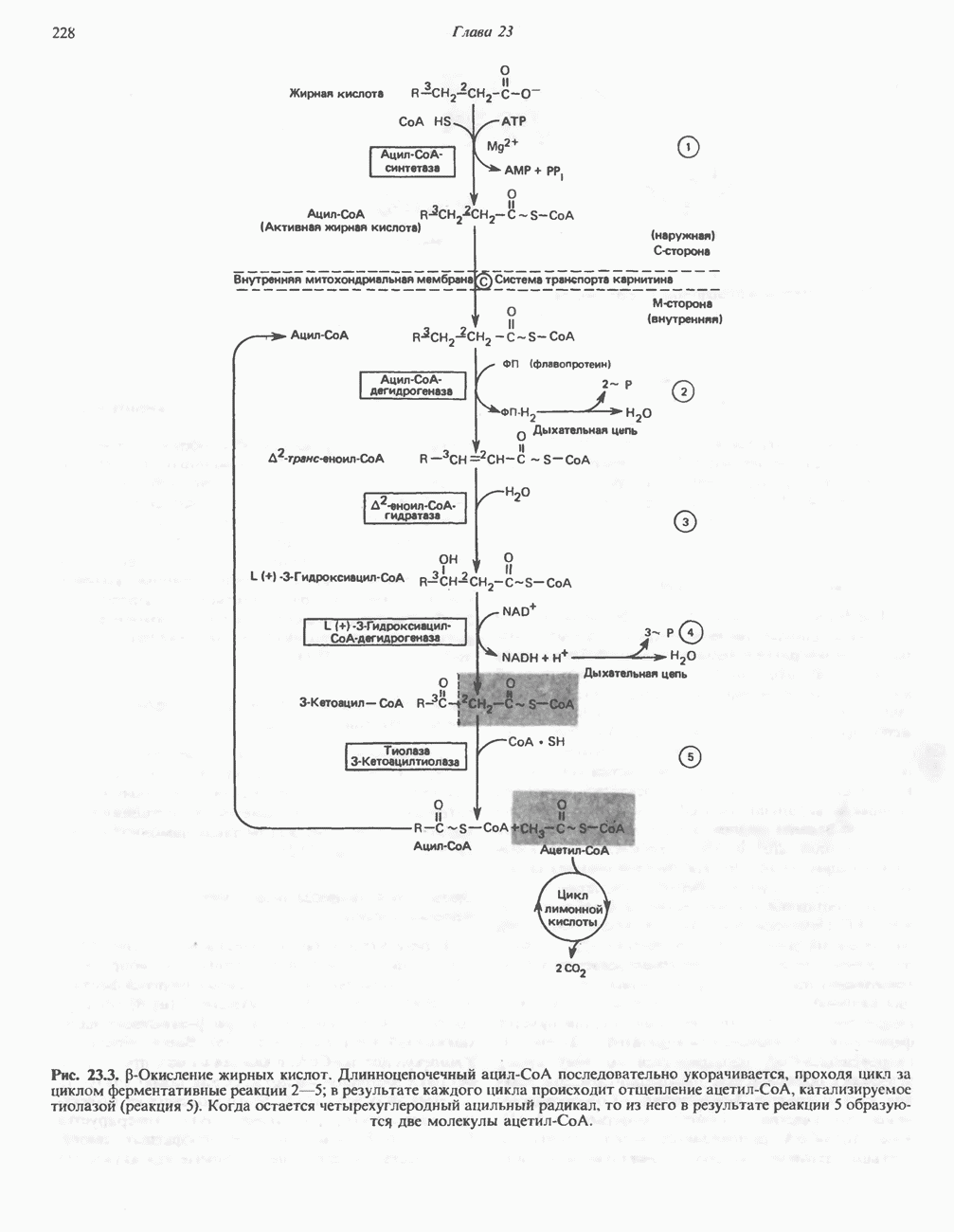
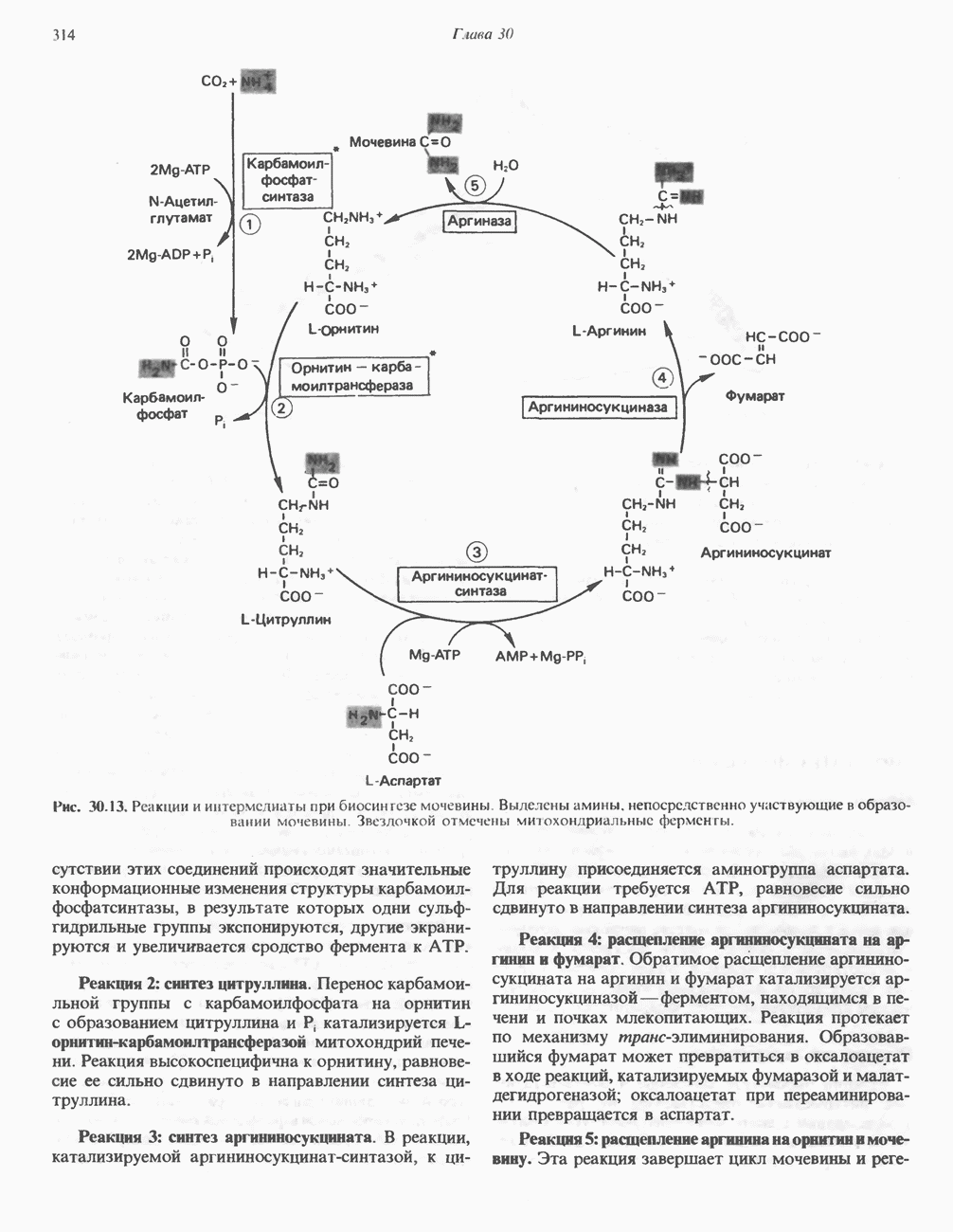
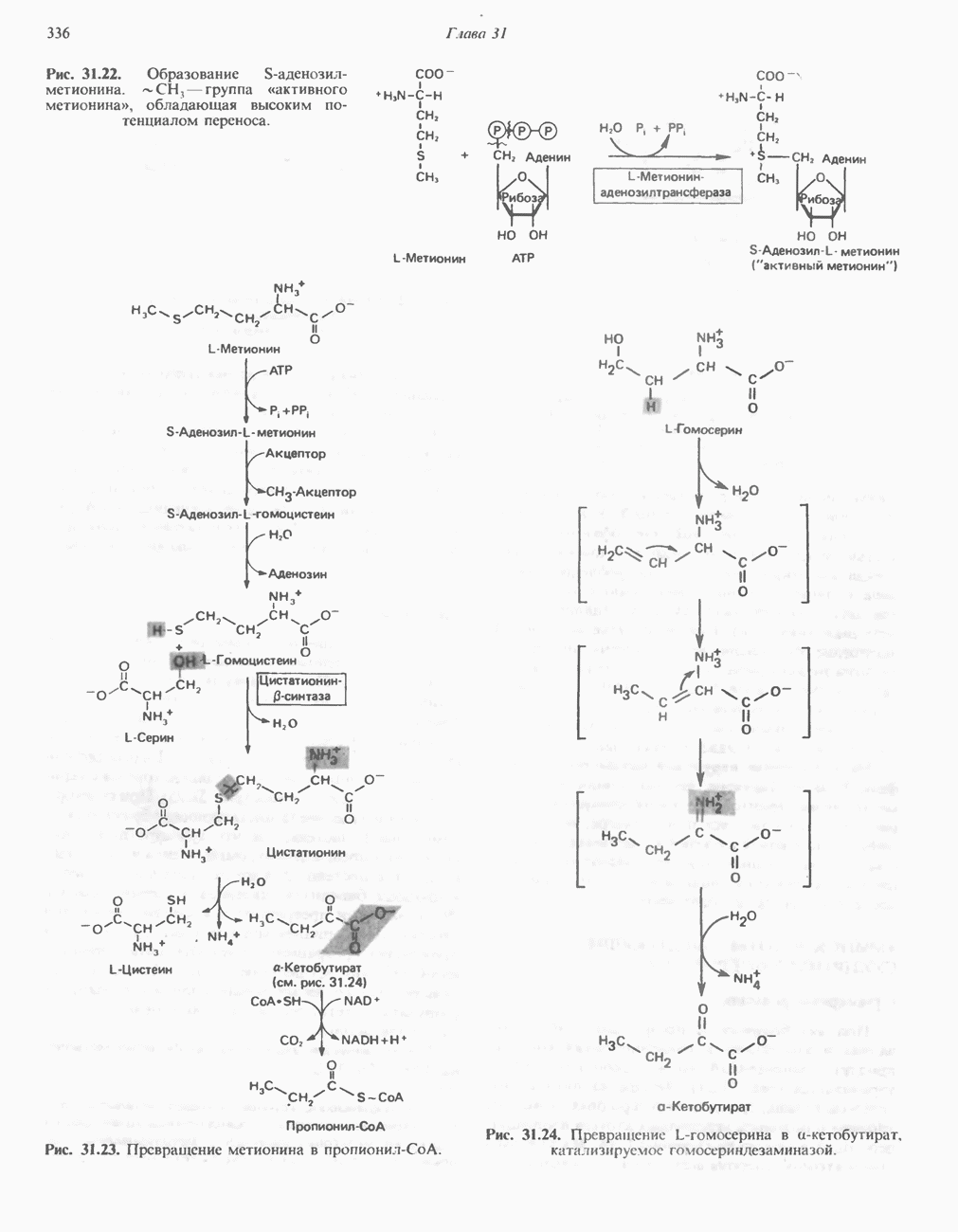
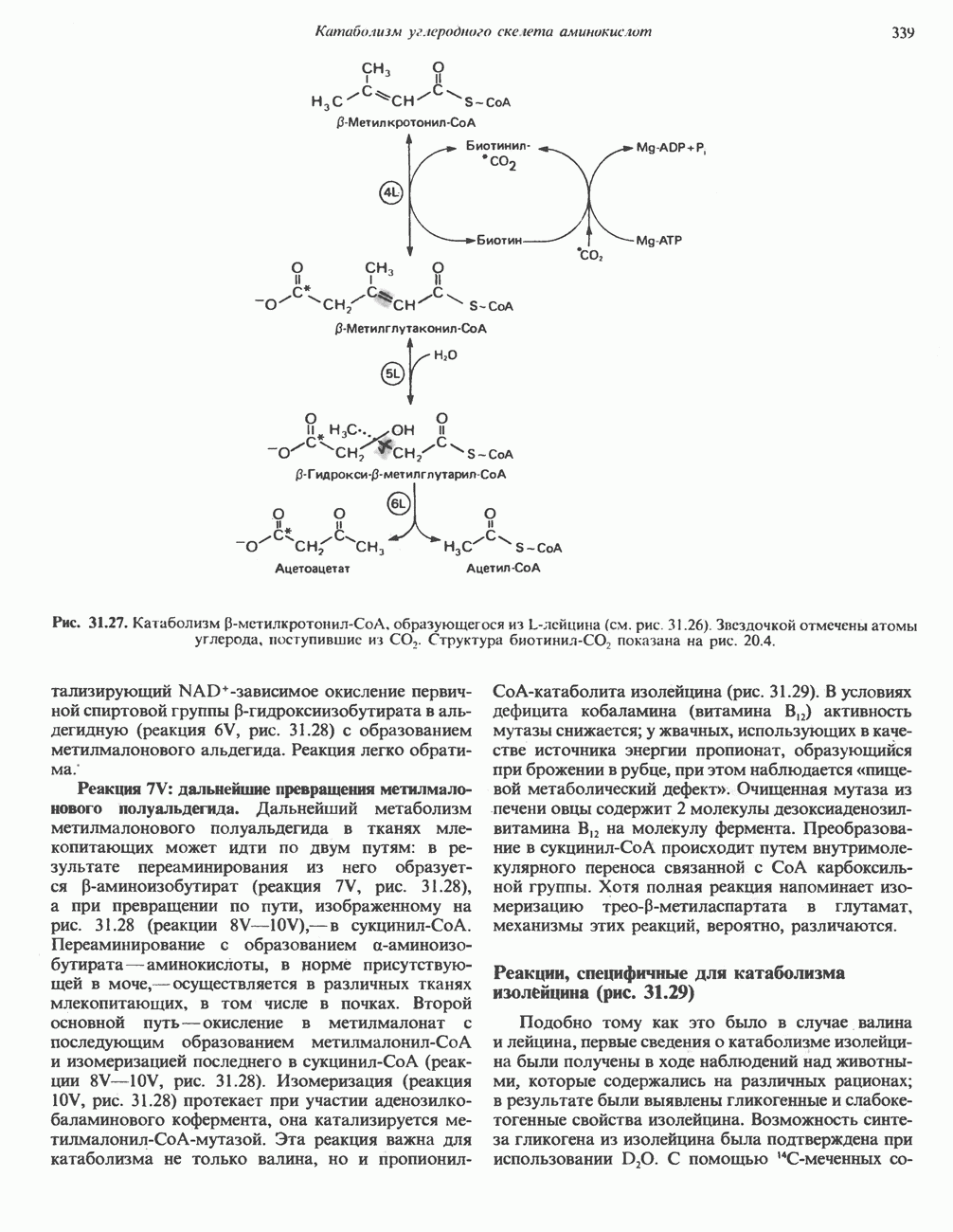
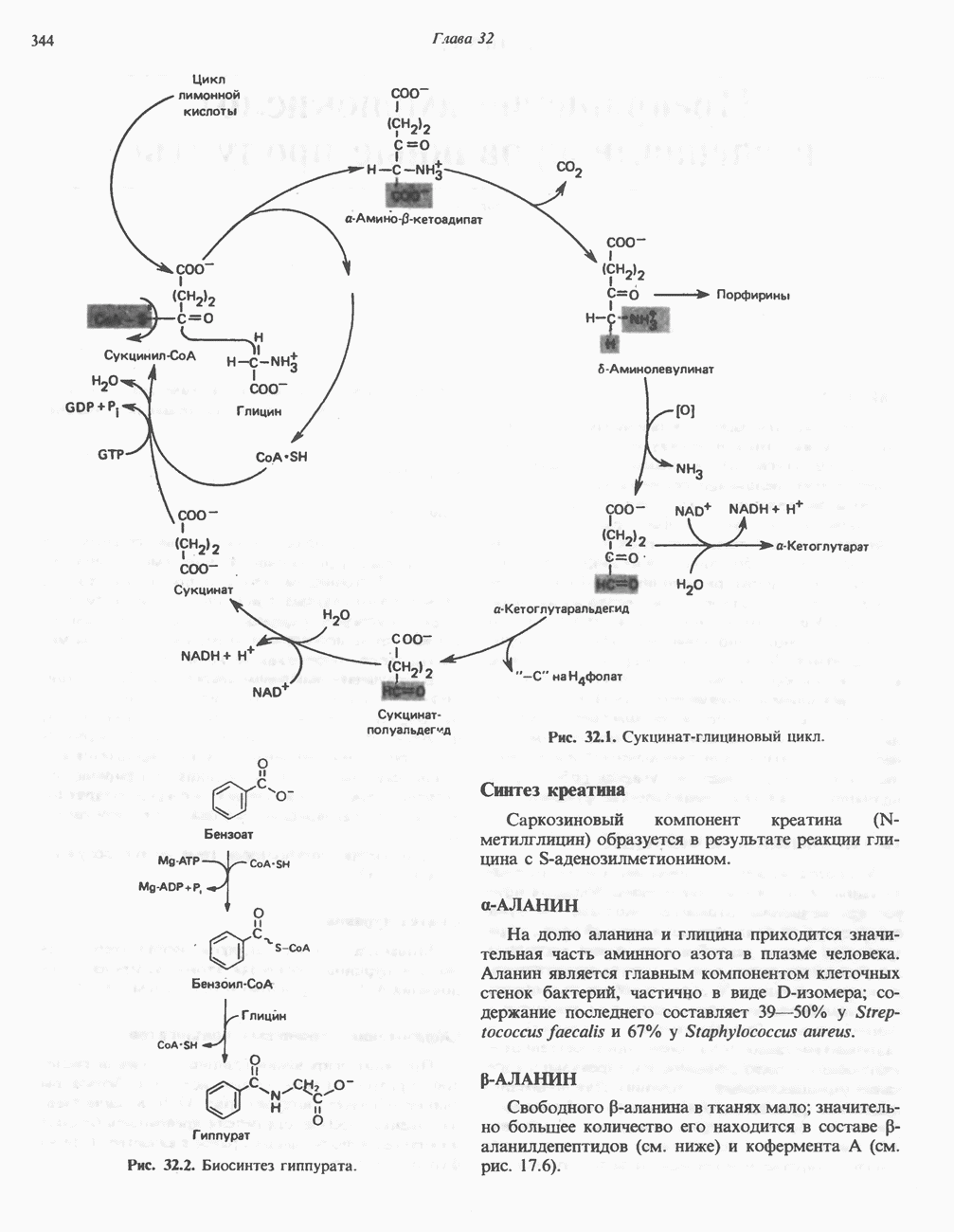
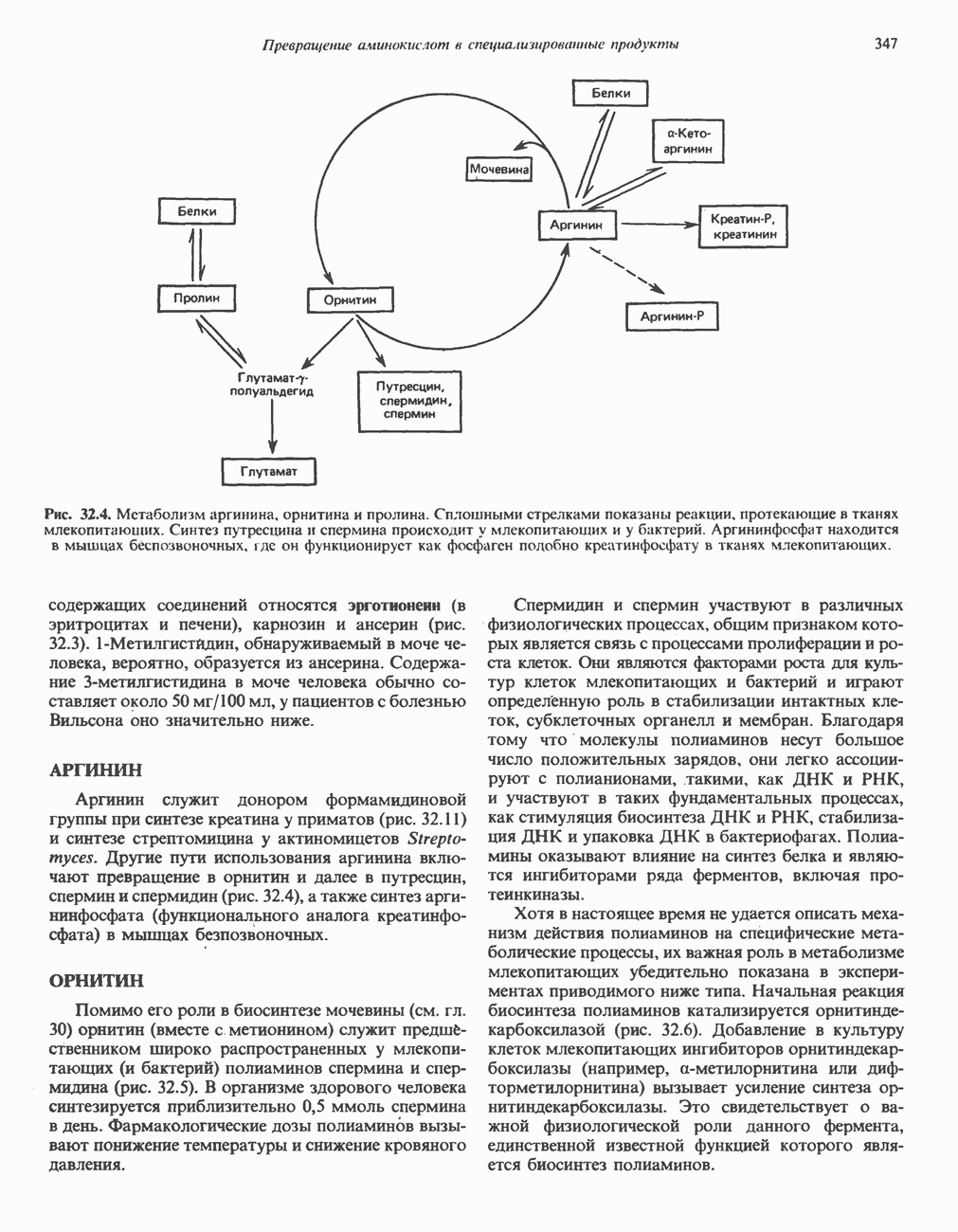
Рис. 32.4. Метаболизм аргинина, орнитина и пролина. Сплошными стрелками показаны реакции, протекающие в тканях млекопитающих. Синтез путресцина и спермина происходит у млекопитающих и у бактерий. Аргининфосфат находится в мышцах беспозвоночных, где он функционирует как фосфаген подобно креатинфосфату в тканях млекопитающих. содержащих соединений относятся эрготионеин (в эритроцитах и печени), карнозин и ансерин (рис. 32.3). 1-Метилгистидин, обнаруживаемый в моче человека, вероятно, образуется из ансерина. Содержание АРГИНИНАргинин служит донором формамидиновой группы при синтезе креатина у приматов (рис. 32.11) и синтезе стрептомицина у актиномицетов Strepto-myces. Другие пути использования аргинина включают превращение в орнитин и далее в путресцин, спермин и спермидин (рис. 32.4), а также синтез аргининфосфата (функционального аналога креатинфо-сфата) в мышцах безпозвоночных. ОРНИТИНПомимо его роли в биосинтезе мочевины (см. гл. 30) орнитин (вместе с метионином) служит предшественником широко распространенных у млекопитающих (и бактерий) полиаминов спермина и спермидина (рис. 32.5). В организме здорового человека синтезируется приблизительно 0,5 ммоль спермина в день. Фармакологические дозы полиаминов вызывают понижение температуры и снижение кровяного давления. Спермидин и спермин участвуют в различных физиологических процессах, общим признаком которых является связь с процессами пролиферации и роста клеток. Они являются факторами роста для культур клеток млекопитающих и бактерий и играют определенную роль в стабилизации интактных клеток, субклеточных органелл и мембран. Благодаря тому что молекулы полиаминов несут большое число положительных зарядов, они легко ассоциируют с полианионами, такими, как ДНК и РНК, и участвуют в таких фундаментальных процессах, как стимуляция биосинтеза ДНК и РНК, стабилизация ДНК и упаковка ДНК в бактериофагах. Полиамины оказывают влияние на синтез белка и являются ингибиторами ряда ферментов, включая протеинкиназы. Хотя в настоящее время не удается описать механизм действия полиаминов на специфические метаболические процессы, их важная роль в метаболизме млекопитающих убедительно показана в экспериментах приводимого ниже типа. Начальная реакция биосинтеза полиаминов катализируется орнитиндекарбоксилазой (рис. 32.6). Добавление в культуру клеток млекопитающих ингибиторов орнитиндекарбоксилазы (например,
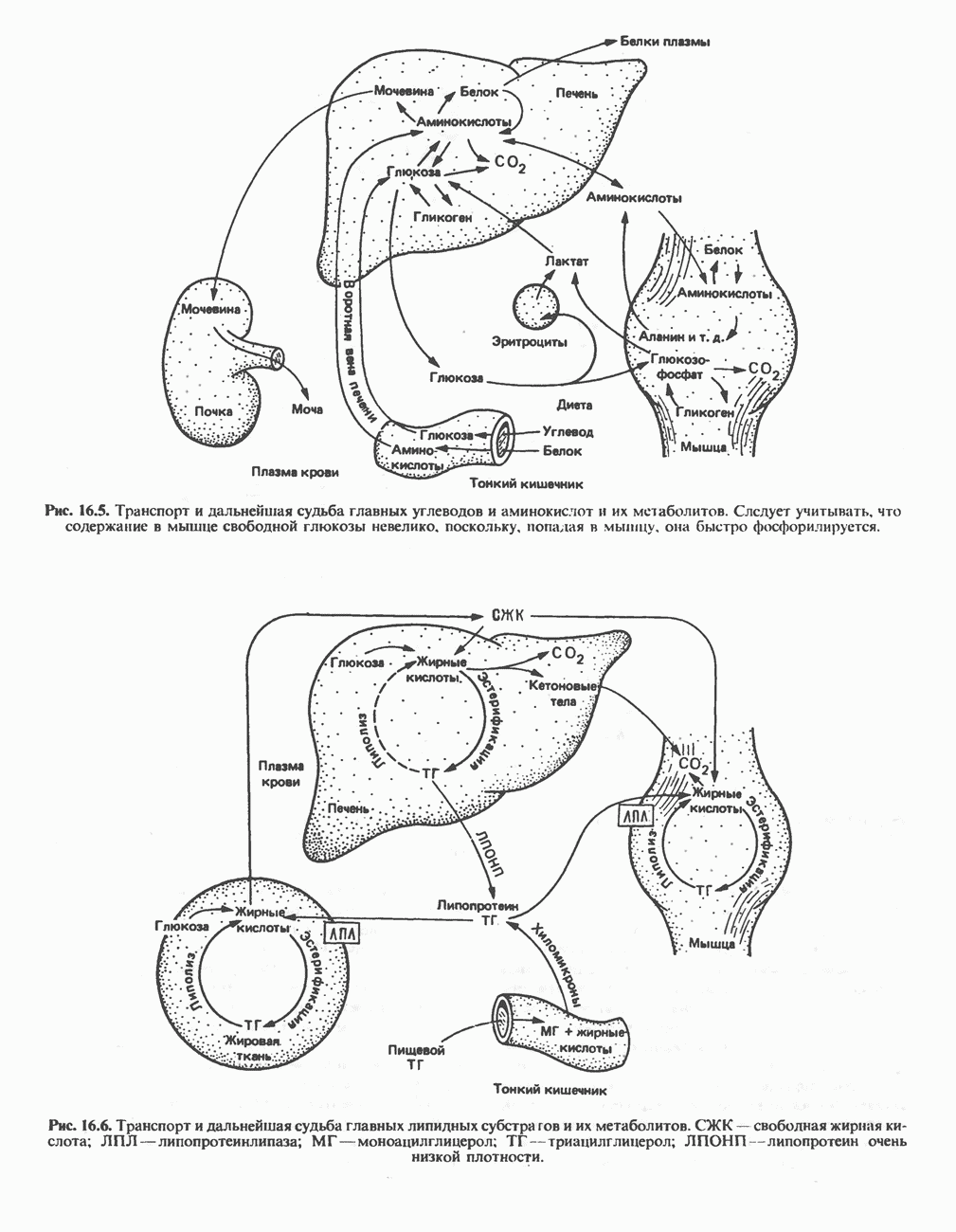
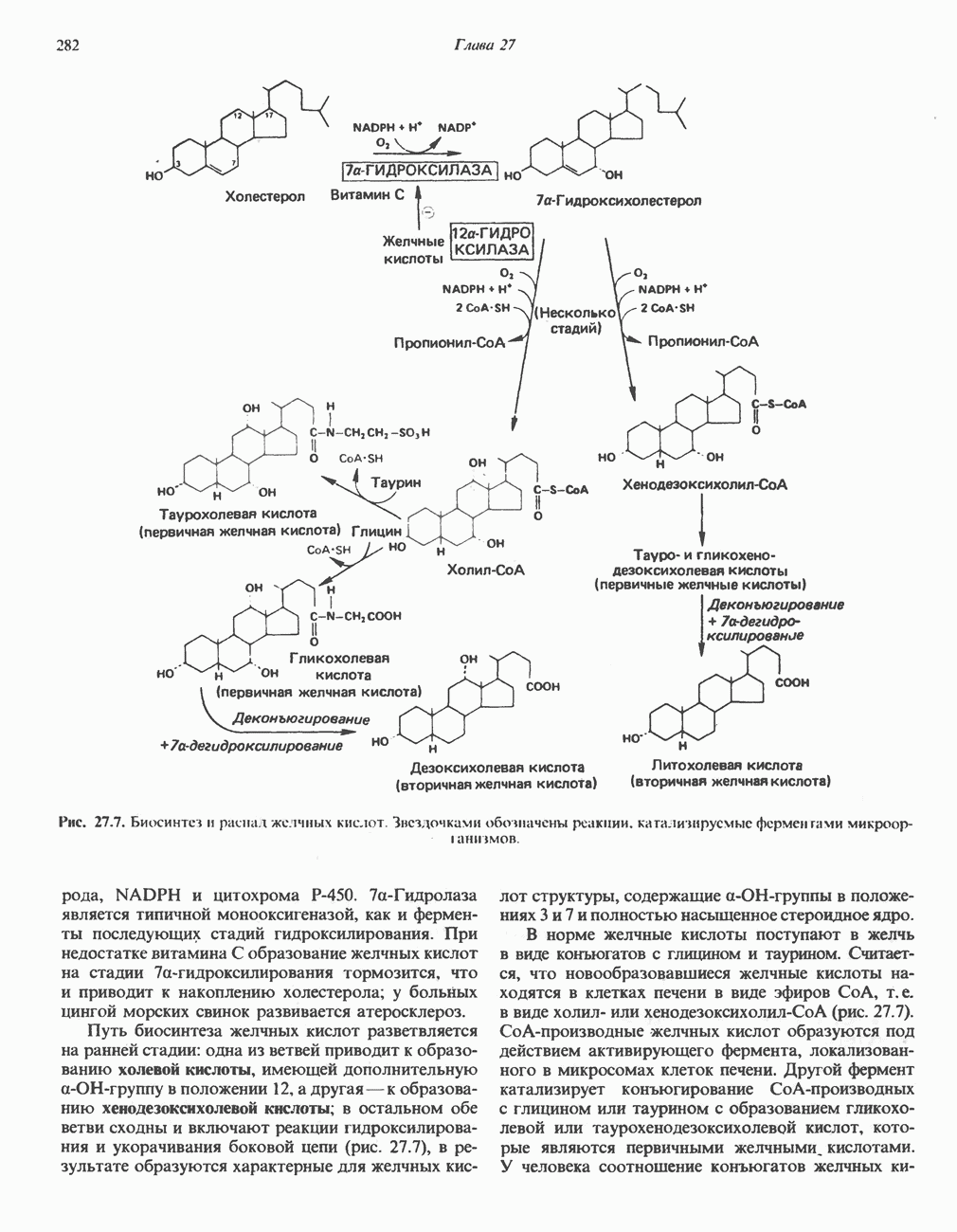
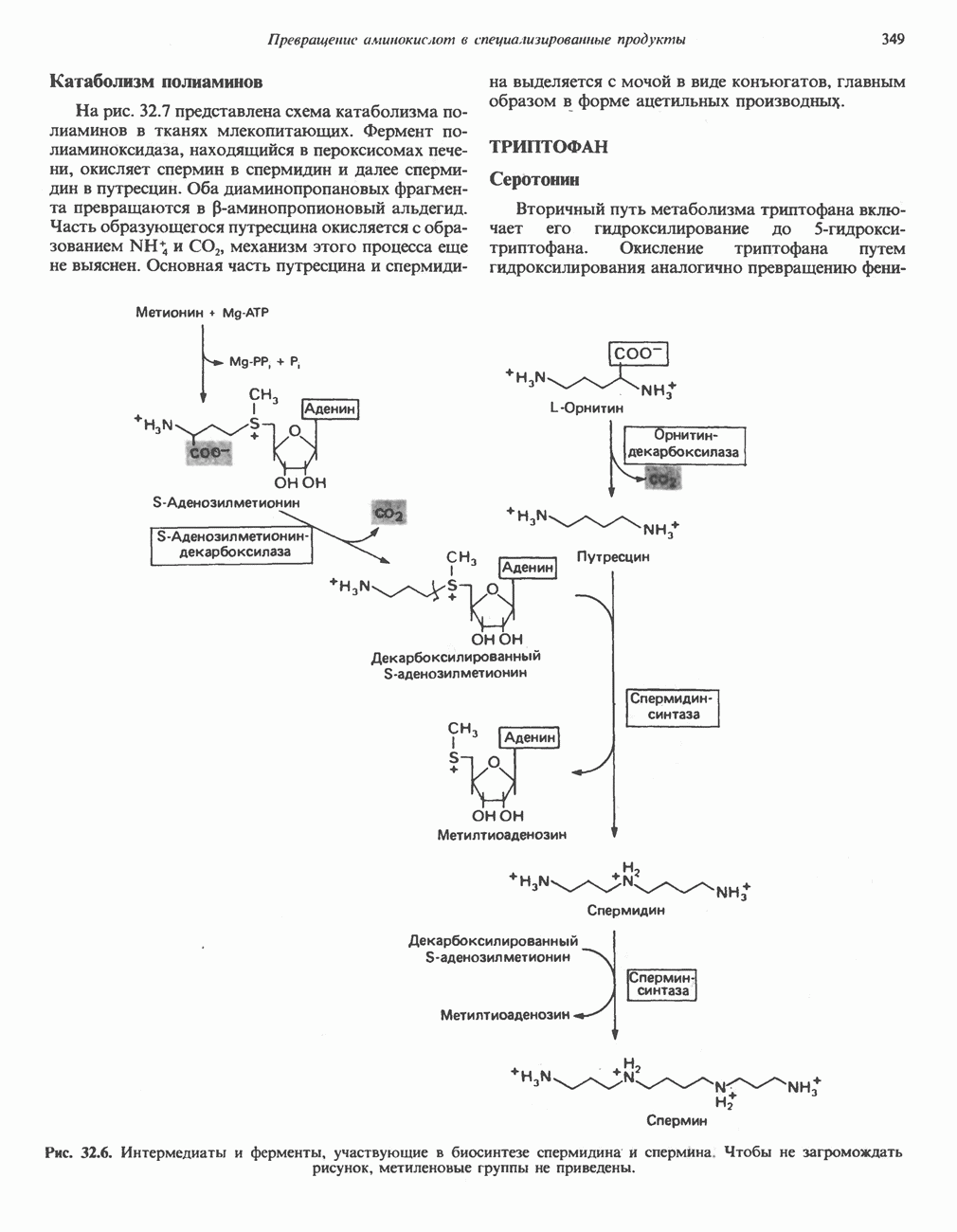
Рис. 32.5. Структура природных полиаминов. Обратите внимание, что спермидин и спермин являются полимерами диаминопропана (А) и диаминобутана (Б). В тканях млекопитающих присутствует также диаминопентан (кадаверин). Биосинтез полиаминовНа рис. 32.6 представлен метаболический путь биосинтеза полиаминов в тканях млекопитающих. Обратите внимание на то, что путресциновый фрагмент спермидина и спермина образуется из L-орнитина (интермедиата цикла мочевины, см. гл. 30), а диаминопропановый фрагмент — из L-метионина (с промежуточным образованием S-аденозилметионина). Орнитиндекарбоксилаза и S-аденозилметионин-декарбоксилаза являются индуцируемыми ферментами с коротким периодом полужизни. Спермин - и спермидинсинтаза, напротив, не являются индуцируемыми и относительно «стабильны». Из числа ферментов биосинтеза полиаминов у млекопитающих орнитиндекарбоксилаза и S-аденозилметионин-декарбоксилаза представляют интерес в плане их регуляции и возможности направленной химиотерапии. Время полужизни орнитиндекарбоксилазы (приблизительно 10 мин) меньше, Чем у любого другого известного фермента млекопитающих; активность этого фермента очень быстро и масштабно изменяется в ответ на различные стимулирующие воздействия. Добавление в культуру клеток млекопитающих гормона роста, кортикостероидов, тестостерона или фактора роста эпидермиса быстро повышает активность орнитиндекарбоксилазы в 10—200 раз. При добавлении в культуру клеток полиаминов индуцируется синтез белкового антифермента (антизима), который связывается с орнитиндекарбоксилазой и ингибирует ее активность. Таким образом, активность орнитиндекарбоксилазы контролируется белок-белковым взаимодействием подобно регуляции активности трипсина ингибиторами белковой природы. По отношению к орнитиндекарбоксилазе дифторметилорнитин можно рассматривать как «суицидальный ингибитор», т.е. соединение, превращающееся в ингибитор под действием самого фермента. Он используется для выделения штамма мутантных клеток, характеризующихся гиперпродукцией орнитиндекарбоксилазы, а также как ингибитор репликации клеток (он действует как химиотерапевтический фактор, мишенью которого является фермент). S-аденозилметионин-декарбоксилаза является единственным известным эукариотическим ферментом, содержащим пируват как ковалентно связанный кофактор (обычно декарбоксилазы содержат пиридоксальфосфат, но в S-аденозилметионин-декарбоксилазе он отсутствует). S-Аденозилметионин-декарбоксилаза имеет сравнительно небольшой период полужизни (1-2 ч) и реагирует на действие стимуляторов клеточного роста подобно орнитиндекарбоксилазе, однако несколько медленнее и менее масштабно. Активность S-аденозилметионин-декарбоксилазы (рис. 32.6) ингибируется декарбоксилированным S-аденозил-метионином и стимулируется путресцином. Катаболизм полиаминовНа рис. 32.7 представлена схема катаболизма полиаминов в тканях млекопитающих. Фермент полиаминоксидаза, находящийся в пероксисомах печени, окисляет спермин в спермидин и далее спермидин в путресцин. Оба диаминопропановых фрагмента превращаются в (
|
 |
Оглавление
|

































































































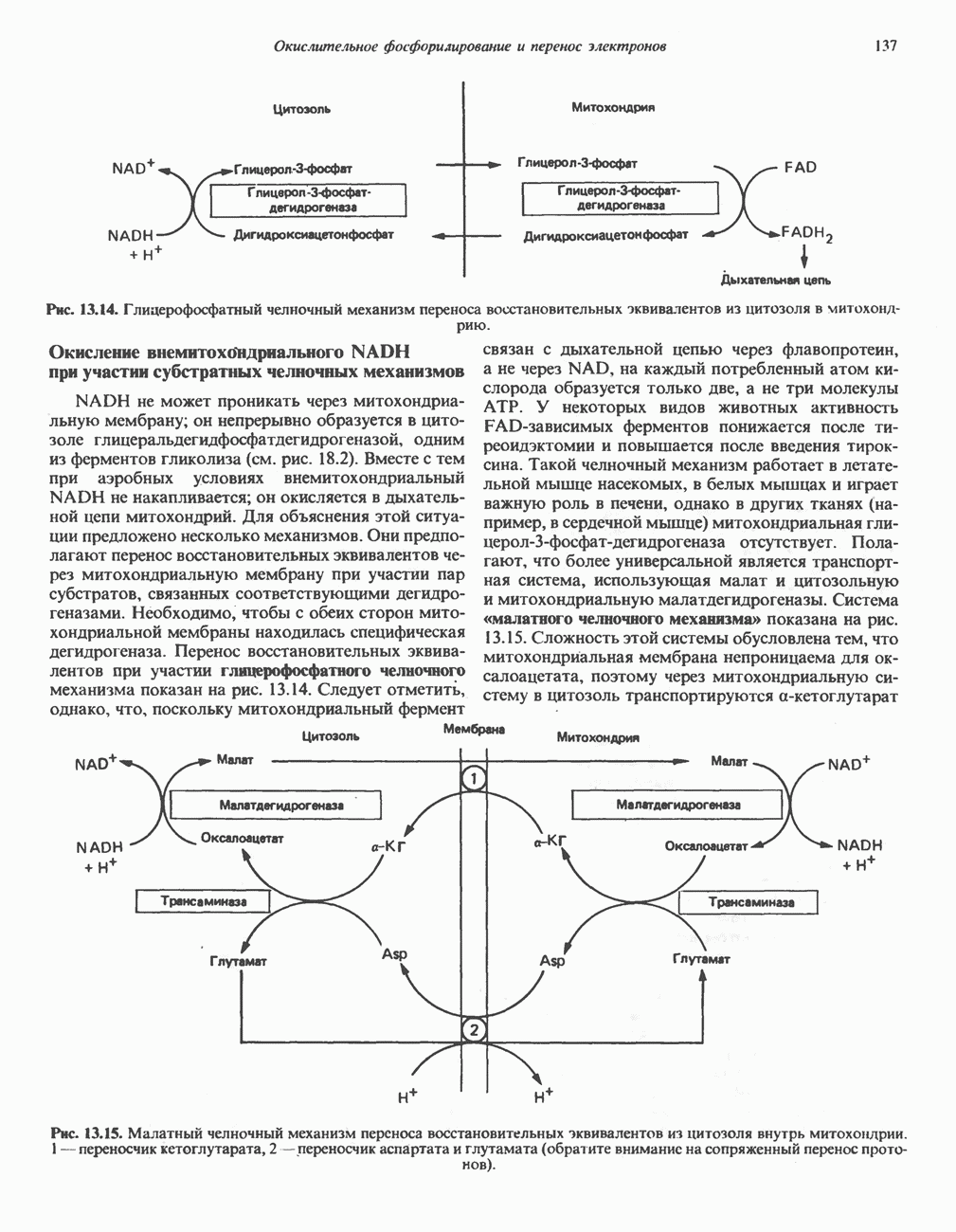



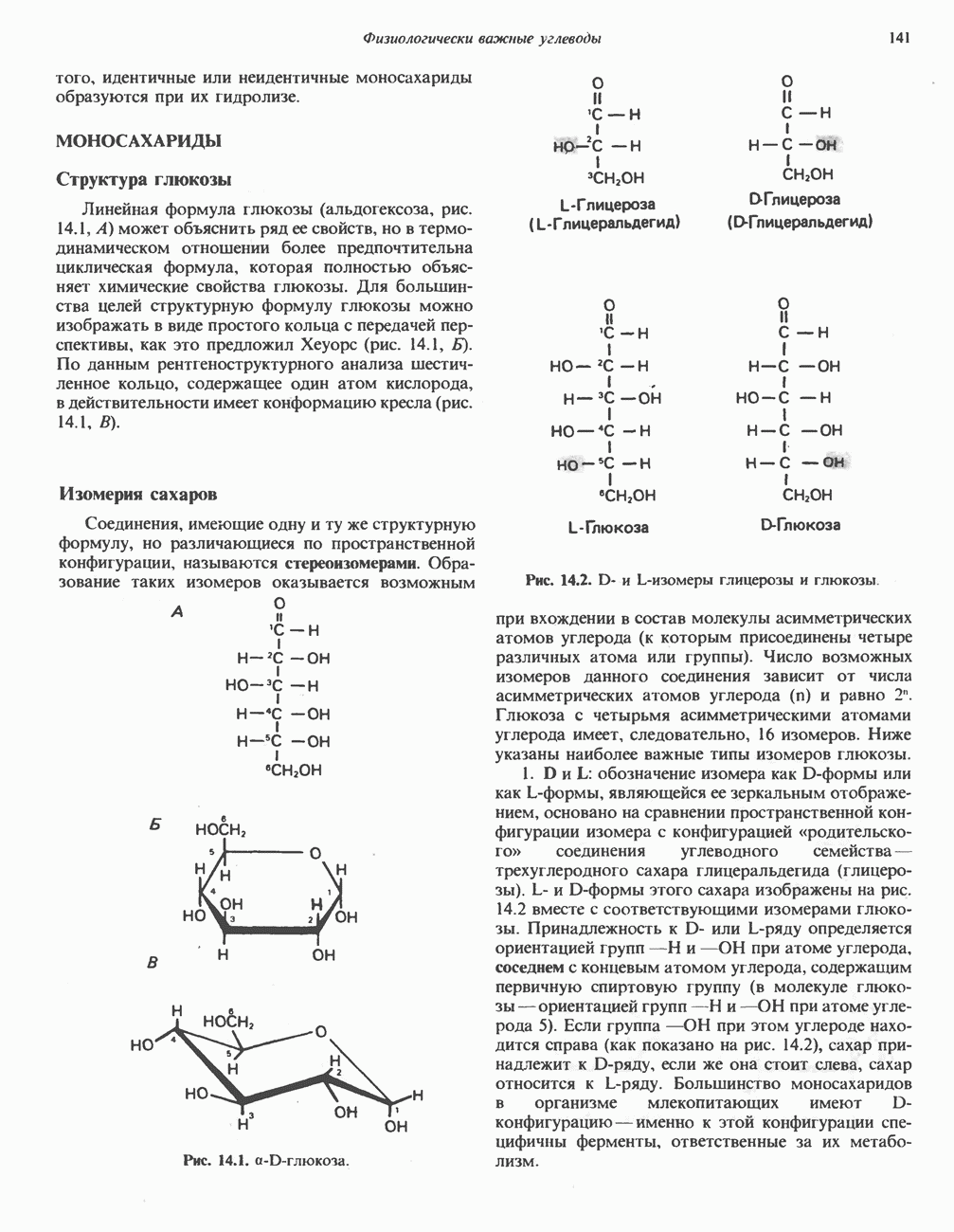
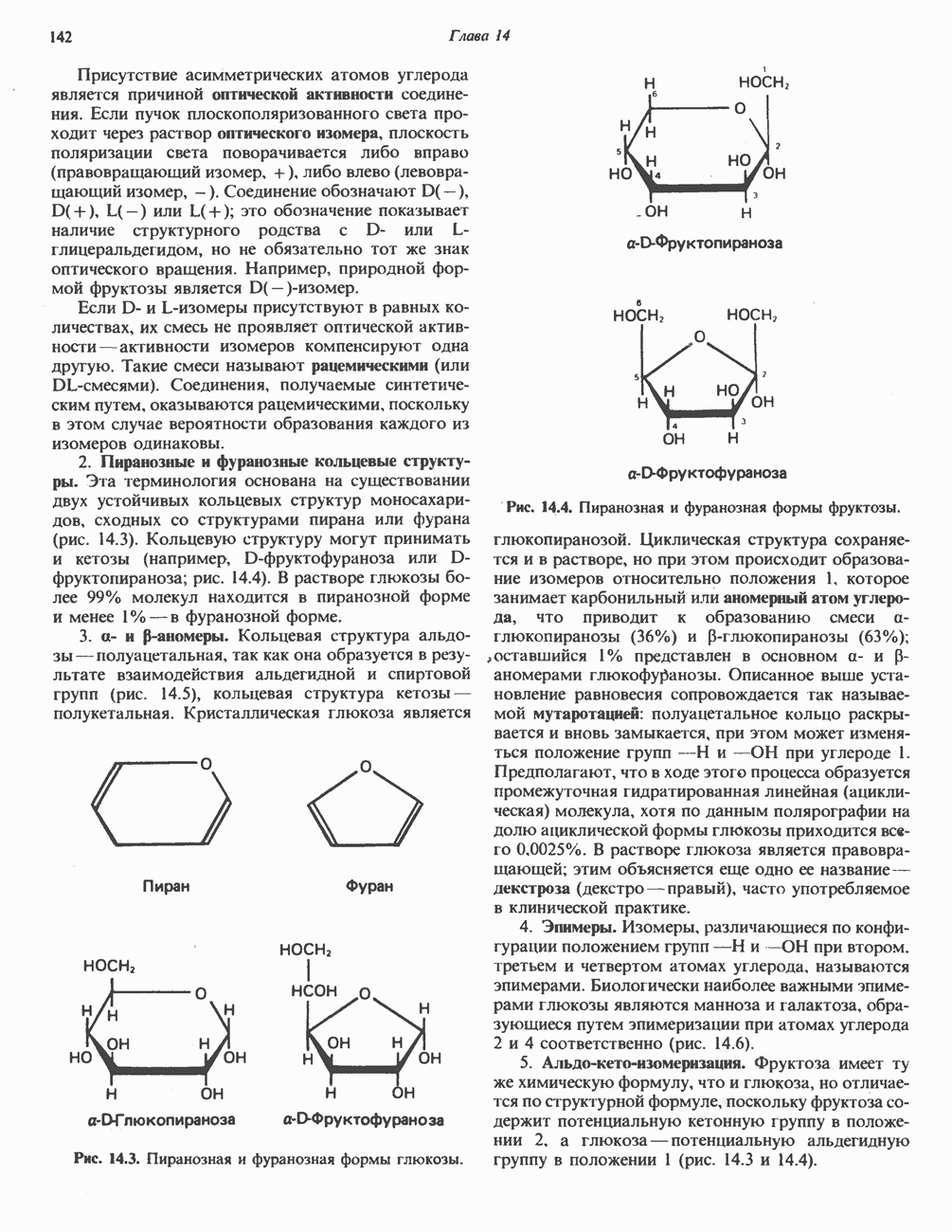
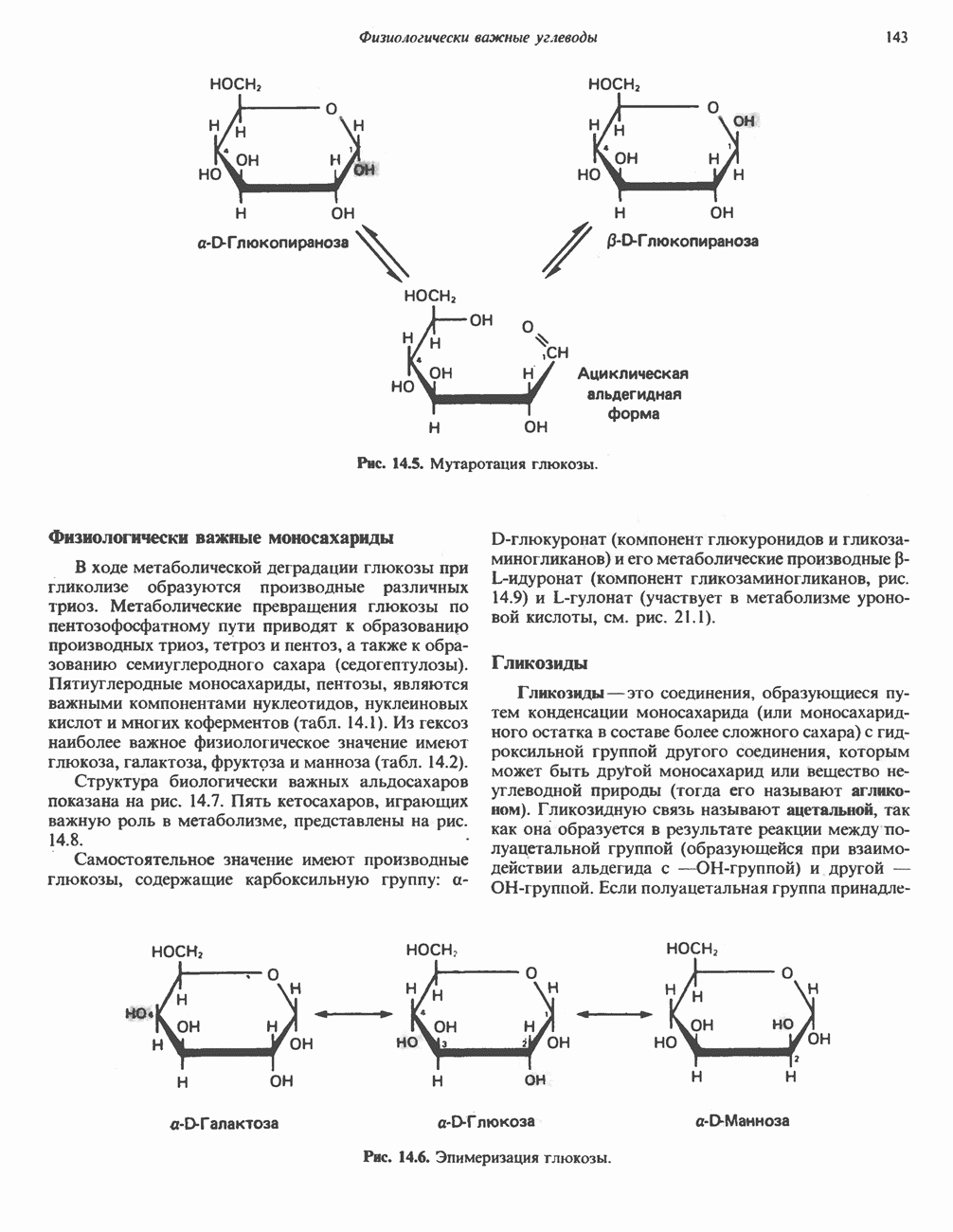

















































































































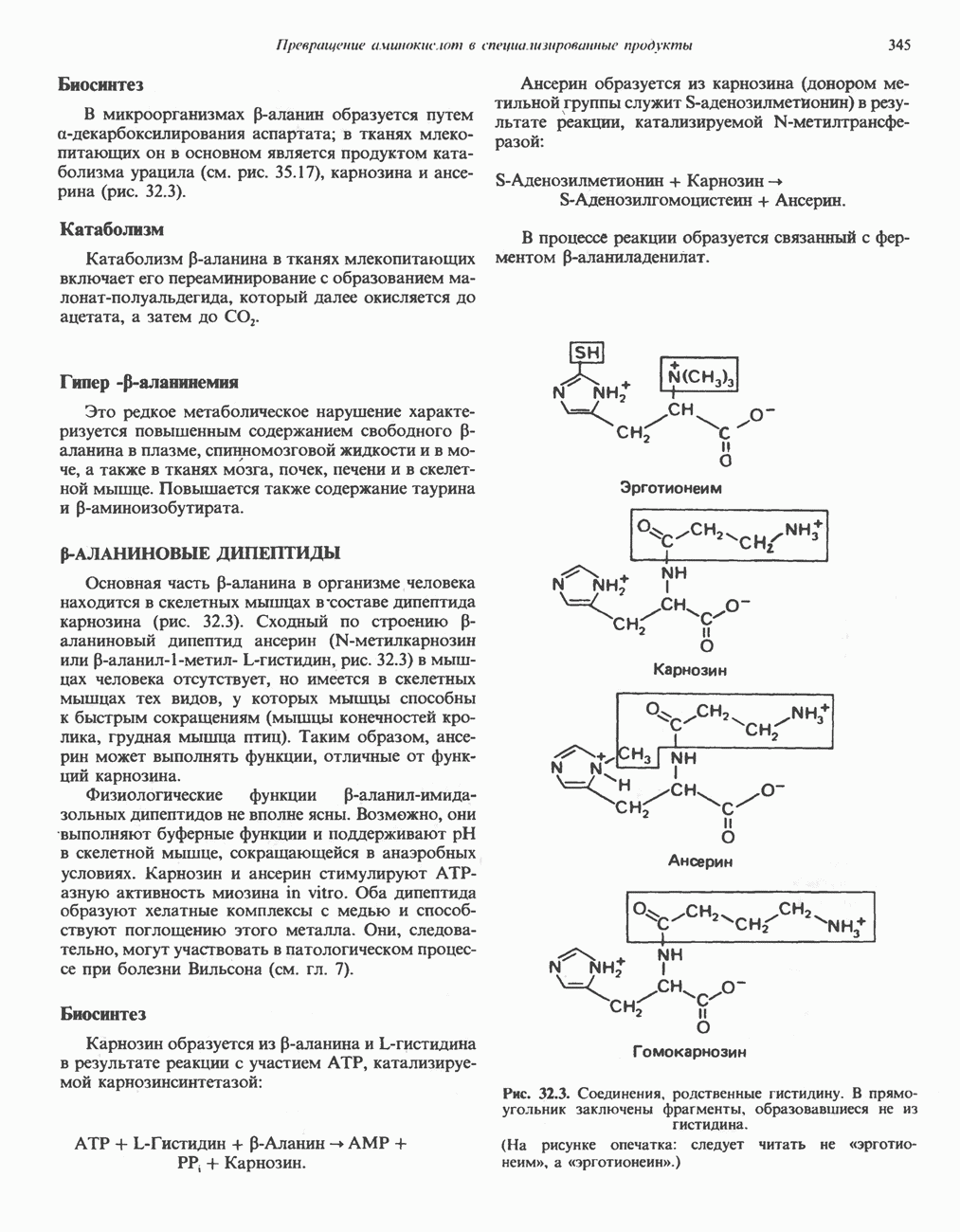

















 -аланиновый дипептид ансерин (
-аланиновый дипептид ансерин ( -метилкарнозин или
-метилкарнозин или  рис. 32.3) в мышцах человека отсутствует, но имеется в скелетных мышцах тех видов, у которых мышцы способны к быстрым сокращениям (мышцы конечностей кролика, грудная мышца птиц). Таким образом, ансерин может выполнять функции, отличные от функций карнозина.
рис. 32.3) в мышцах человека отсутствует, но имеется в скелетных мышцах тех видов, у которых мышцы способны к быстрым сокращениям (мышцы конечностей кролика, грудная мышца птиц). Таким образом, ансерин может выполнять функции, отличные от функций карнозина.
 -аланил-имида-зольных дипептидов не вполне ясны. Возможно, они выполняют буферные функции и поддерживают pH в скелетной мышце, сокращающейся в анаэробных условиях. Карнозин и ансерин стимулируют АТР-азную активность миозина in vitro. Оба дипептида образуют хелатные комплексы с медью и способствуют поглощению этого металла. Они, следовательно, могут участвовать в патологическом процессе при болезни Вильсона (см. гл. 7).
-аланил-имида-зольных дипептидов не вполне ясны. Возможно, они выполняют буферные функции и поддерживают pH в скелетной мышце, сокращающейся в анаэробных условиях. Карнозин и ансерин стимулируют АТР-азную активность миозина in vitro. Оба дипептида образуют хелатные комплексы с медью и способствуют поглощению этого металла. Они, следовательно, могут участвовать в патологическом процессе при болезни Вильсона (см. гл. 7).
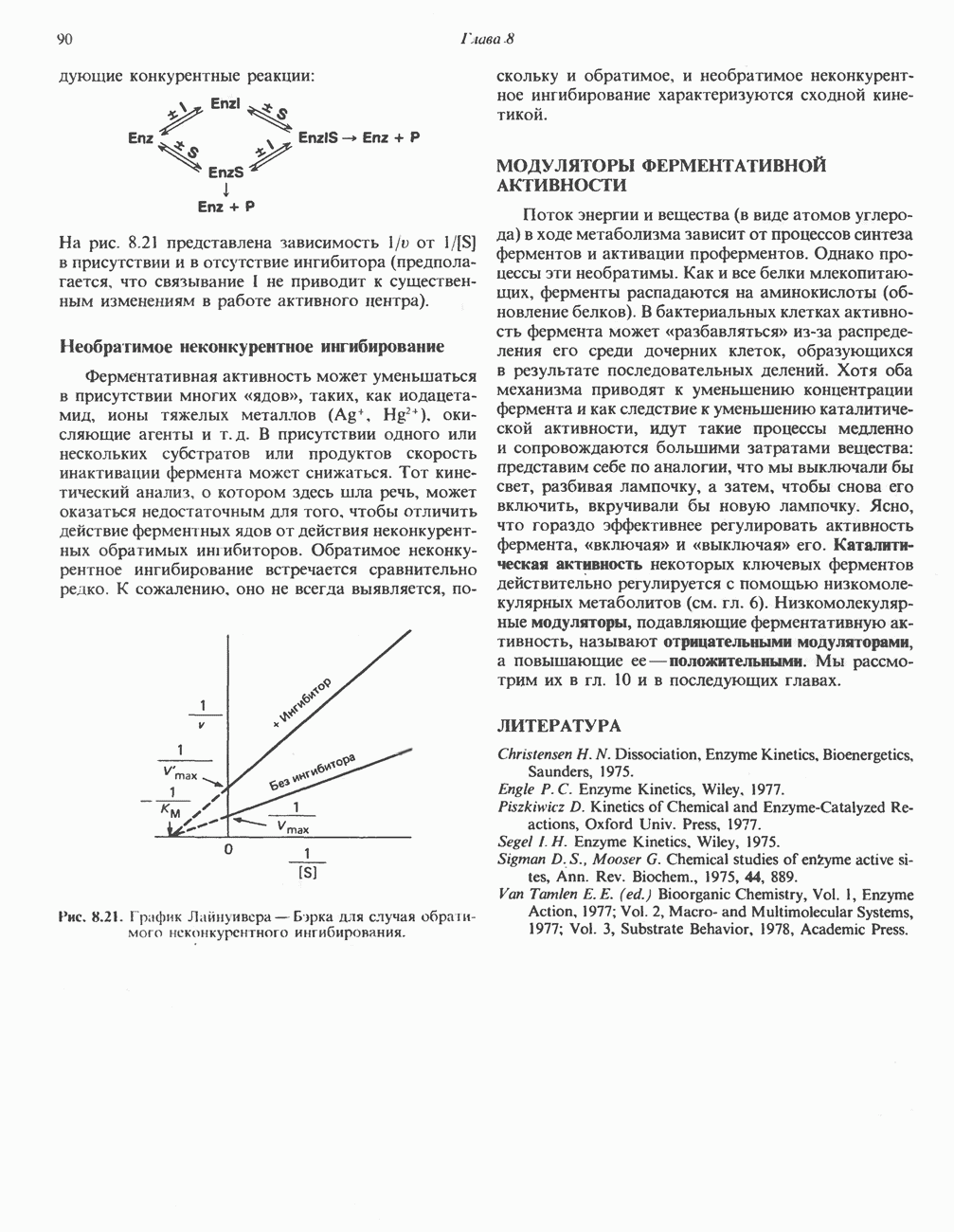
 -гистидина в результате реакции с участием АТР, катализируемой карнозинсинтетазой:
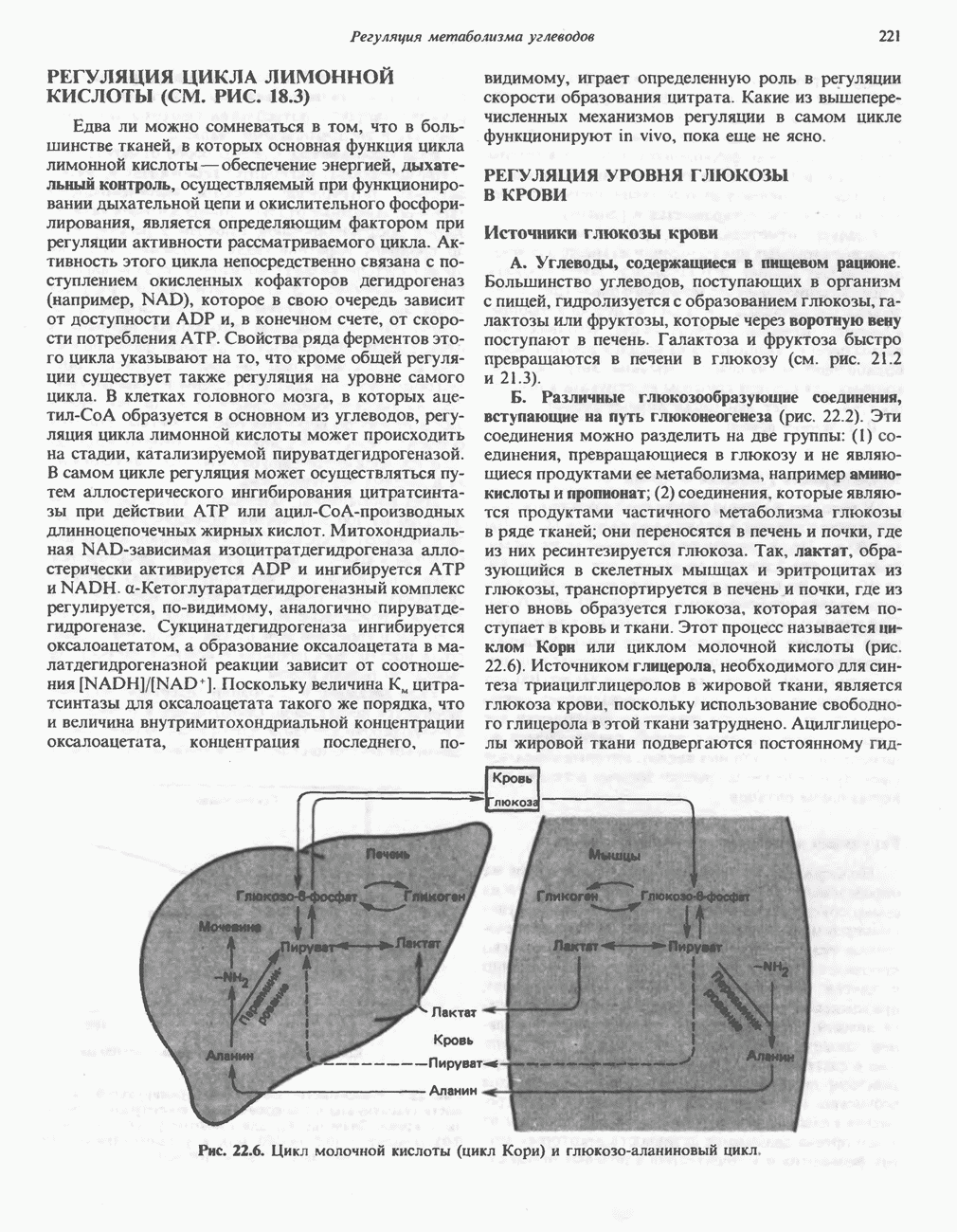
-гистидина в результате реакции с участием АТР, катализируемой карнозинсинтетазой:

 -аденозилметионин) в результате реакции, катализируемой
-аденозилметионин) в результате реакции, катализируемой  -метилтрансферазой:
-метилтрансферазой:

 -аланиладенилат.
-аланиладенилат.

 -гистидина
-гистидина
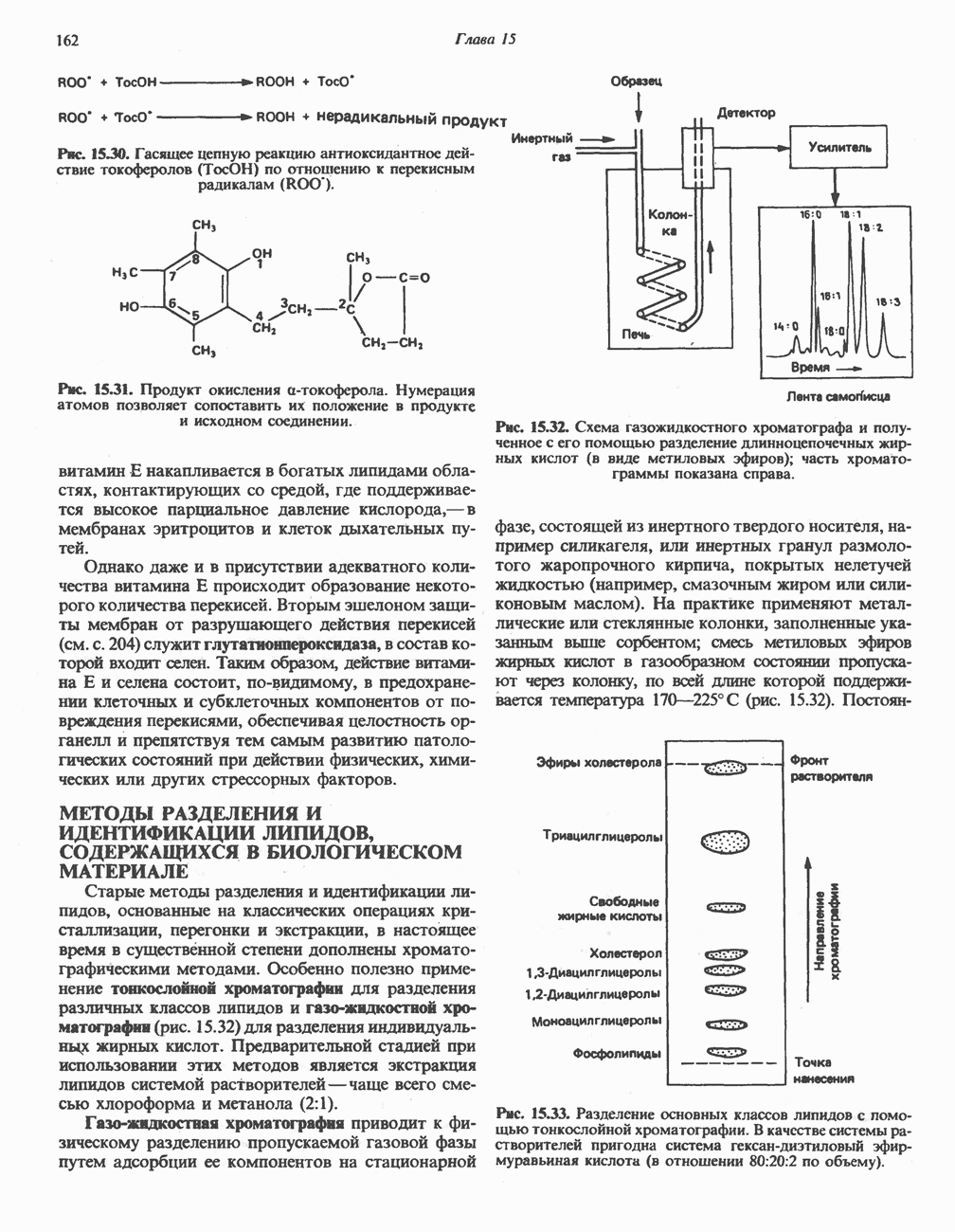
 -гистидина содержащимся в сыворотке цинксодержащим металлоферментом карнозиназой (карнозин-гидролазой). В сыворотке имеются две формы карнозиназы.
-гистидина содержащимся в сыворотке цинксодержащим металлоферментом карнозиназой (карнозин-гидролазой). В сыворотке имеются две формы карнозиназы.
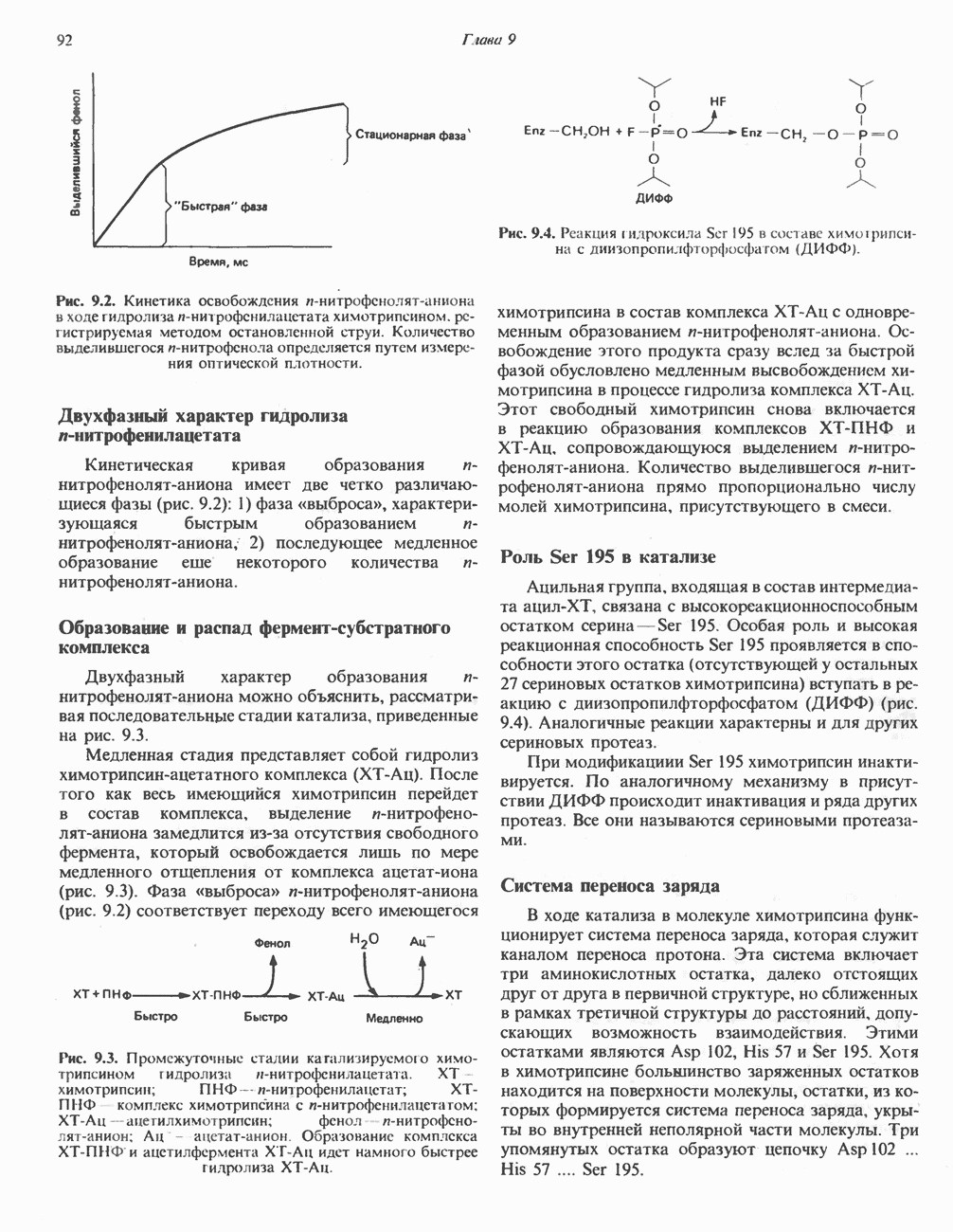
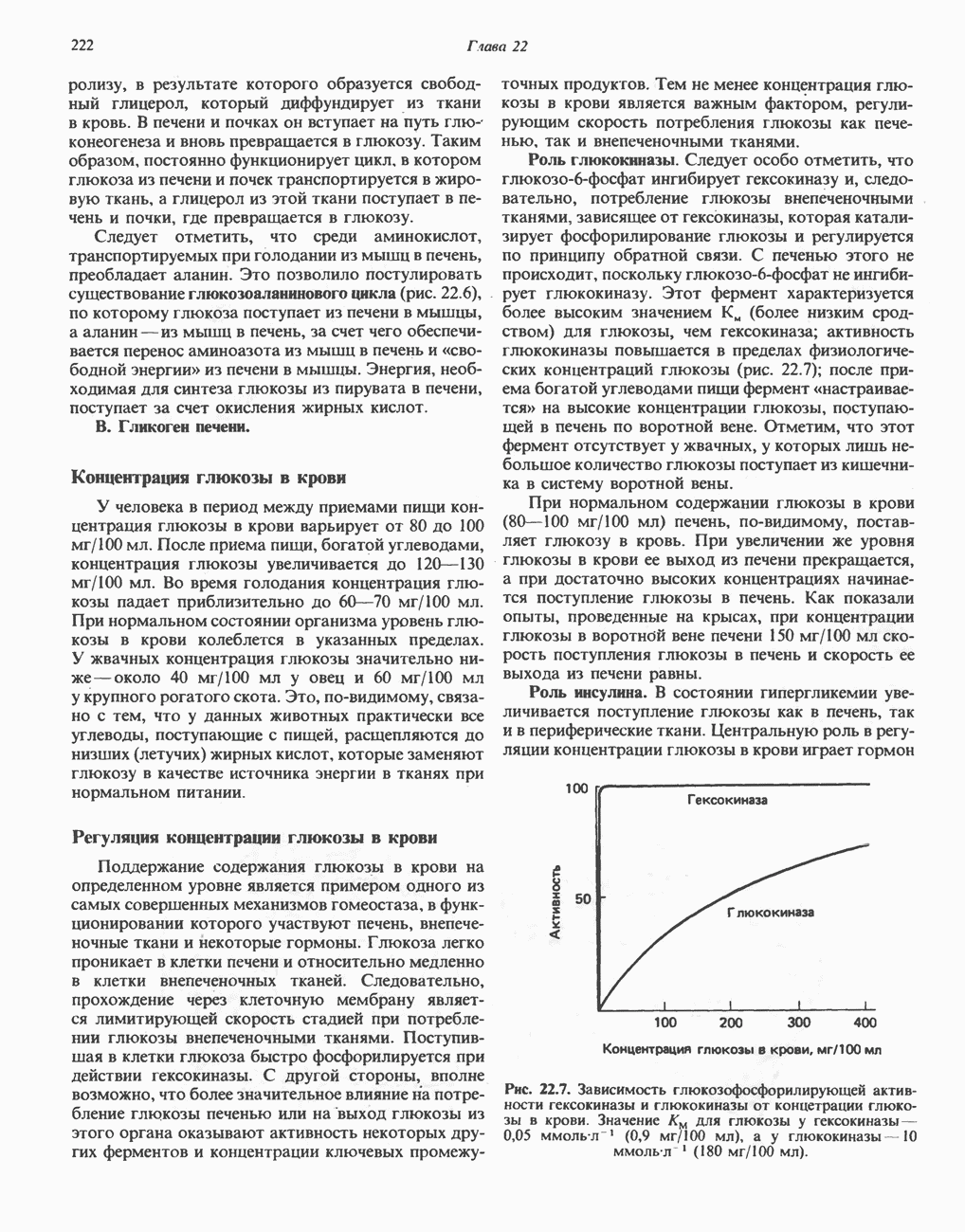
 -аминобутиратом и
-аминобутиратом и  -гистидином, находится в ткани мозга человека; концентрация его в разных отделах мозга различна. Биосинтез гомокарнозина в мозгу человека катализируется, по-видимому, карнозинсинтетазой. Сывороточная карнозиназа гомокарнозин не гидролизует.
-гистидином, находится в ткани мозга человека; концентрация его в разных отделах мозга различна. Биосинтез гомокарнозина в мозгу человека катализируется, по-видимому, карнозинсинтетазой. Сывороточная карнозиназа гомокарнозин не гидролизует.
 -Углерод является источником метильных групп тимина (и холина) и атомов углерода в положениях 2 и 8 пуринового ядра (см. гл. 35).
-Углерод является источником метильных групп тимина (и холина) и атомов углерода в положениях 2 и 8 пуринового ядра (см. гл. 35).
 -изомер и соответствующая а-кетокислота млекопитающими не усваиваются. В ряде белков треонин находится в форме О-фосфотреонина.
-изомер и соответствующая а-кетокислота млекопитающими не усваиваются. В ряде белков треонин находится в форме О-фосфотреонина.
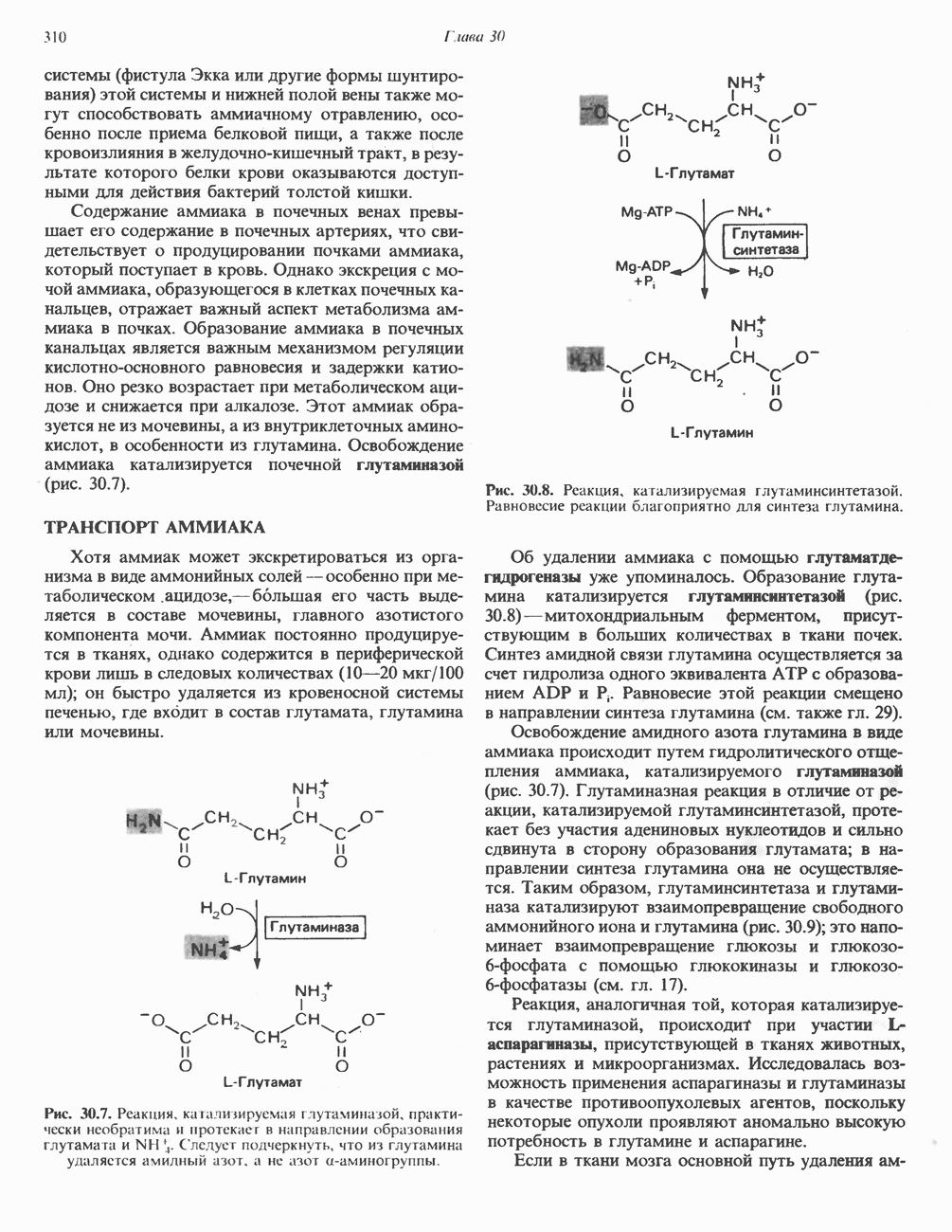
 -аденозилметионина он является главным источником метильных групп в организме. Метильные группы могут использоваться как непосредственно, так и в окисленном виде. Метальный углерод может переходить в состав одноуглеродного фрагмента, который конъюгируется с глицином при синтезе серина. В форме
-аденозилметионина он является главным источником метильных групп в организме. Метильные группы могут использоваться как непосредственно, так и в окисленном виде. Метальный углерод может переходить в состав одноуглеродного фрагмента, который конъюгируется с глицином при синтезе серина. В форме  -аденозилметионина метионин служит предшественником
-аденозилметионина метионин служит предшественником  -диаминопропанового фрагмента полиаминов, спермина и спермидина (см. ниже раздел «Орнитин» и рис. 32.6).
-диаминопропанового фрагмента полиаминов, спермина и спермидина (см. ниже раздел «Орнитин» и рис. 32.6).
 -цистеина. Сера метионина (после образования гомоцистеина) переносится на серин (см. рис. 29.10) и участвует в образовании сульфата мочи опосредованно (т.е. после образования цистеина).
-цистеина. Сера метионина (после образования гомоцистеина) переносится на серин (см. рис. 29.10) и участвует в образовании сульфата мочи опосредованно (т.е. после образования цистеина).  -Цистеин является предшественником тиоэтаноламинового фрагмента кофермента А. Он является также предшественником таурина, образующего конъюгаты с желчными кислотами с образованием таурохолевой кислоты и других продуктов.
-Цистеин является предшественником тиоэтаноламинового фрагмента кофермента А. Он является также предшественником таурина, образующего конъюгаты с желчными кислотами с образованием таурохолевой кислоты и других продуктов.
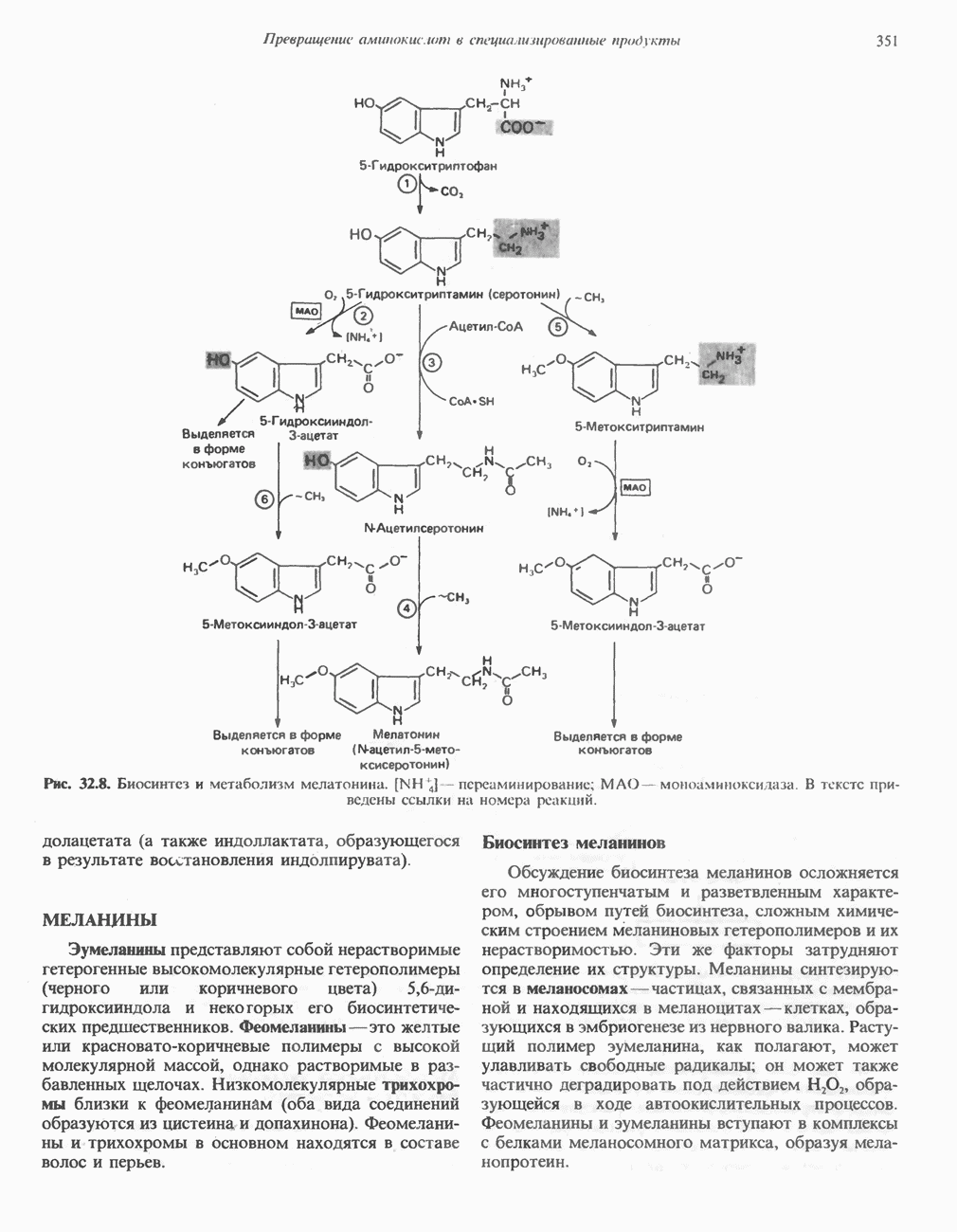
 -аминокислот. Этот фермент катализирует также декарбоксилирование ДОФА, 5-гидрокситриптофана, фенилаланина, тирозина и триптофана (см. ниже). Декарбоксилаза in vitro и in vivo ингибируется а-метиламинокислотами, которые применяются в клинике в качестве гипотензивных средств. Наряду с декарбоксилазой ароматических аминокислот в большинстве клеток имеется гистидиндекарбоксилаза, катализирующая декарбоксилирование гистидина.
-аминокислот. Этот фермент катализирует также декарбоксилирование ДОФА, 5-гидрокситриптофана, фенилаланина, тирозина и триптофана (см. ниже). Декарбоксилаза in vitro и in vivo ингибируется а-метиламинокислотами, которые применяются в клинике в качестве гипотензивных средств. Наряду с декарбоксилазой ароматических аминокислот в большинстве клеток имеется гистидиндекарбоксилаза, катализирующая декарбоксилирование гистидина.

 -метилгистидина в моче человека обычно составляет около
-метилгистидина в моче человека обычно составляет около  у пациентов с болезнью Вильсона оно значительно ниже.
у пациентов с болезнью Вильсона оно значительно ниже.
 -метилорнитина или дифторметилорнитина) вызывает усиление синтеза орнитиндекарбоксилазы. Это свидетельствует о важной физиологической роли данного фермента, единственной известной функцией которого является биосинтез полиаминов.
-метилорнитина или дифторметилорнитина) вызывает усиление синтеза орнитиндекарбоксилазы. Это свидетельствует о важной физиологической роли данного фермента, единственной известной функцией которого является биосинтез полиаминов.

 -аминопропионовый альдегид. Часть образующегося путресцина окисляется с образованием
-аминопропионовый альдегид. Часть образующегося путресцина окисляется с образованием  механизм этого процесса еще не выяснен. Основная часть путресцина и спермидина выделяется с мочой в виде конъюгатов, главным образом в форме ацетильных производные.
механизм этого процесса еще не выяснен. Основная часть путресцина и спермидина выделяется с мочой в виде конъюгатов, главным образом в форме ацетильных производные.