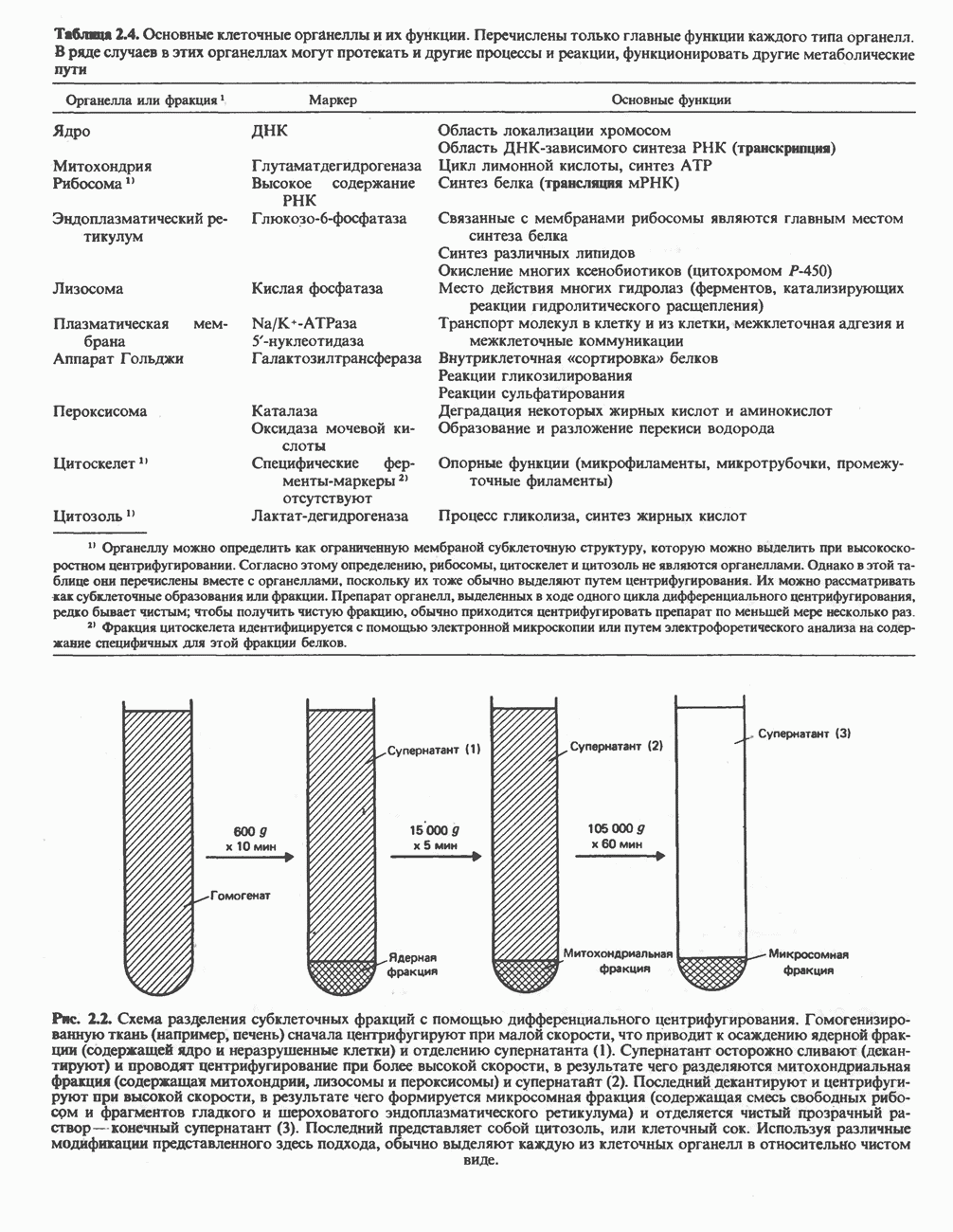
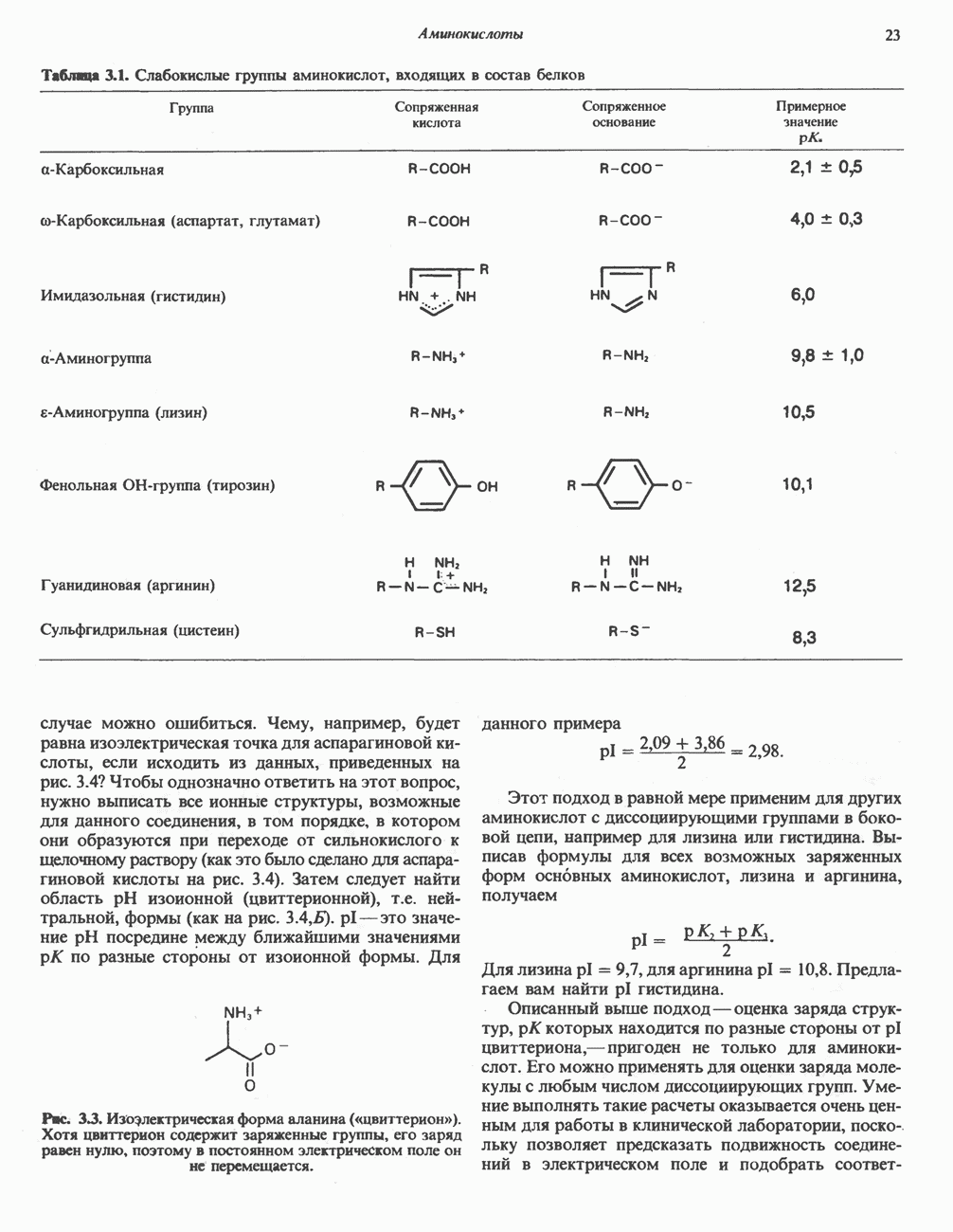
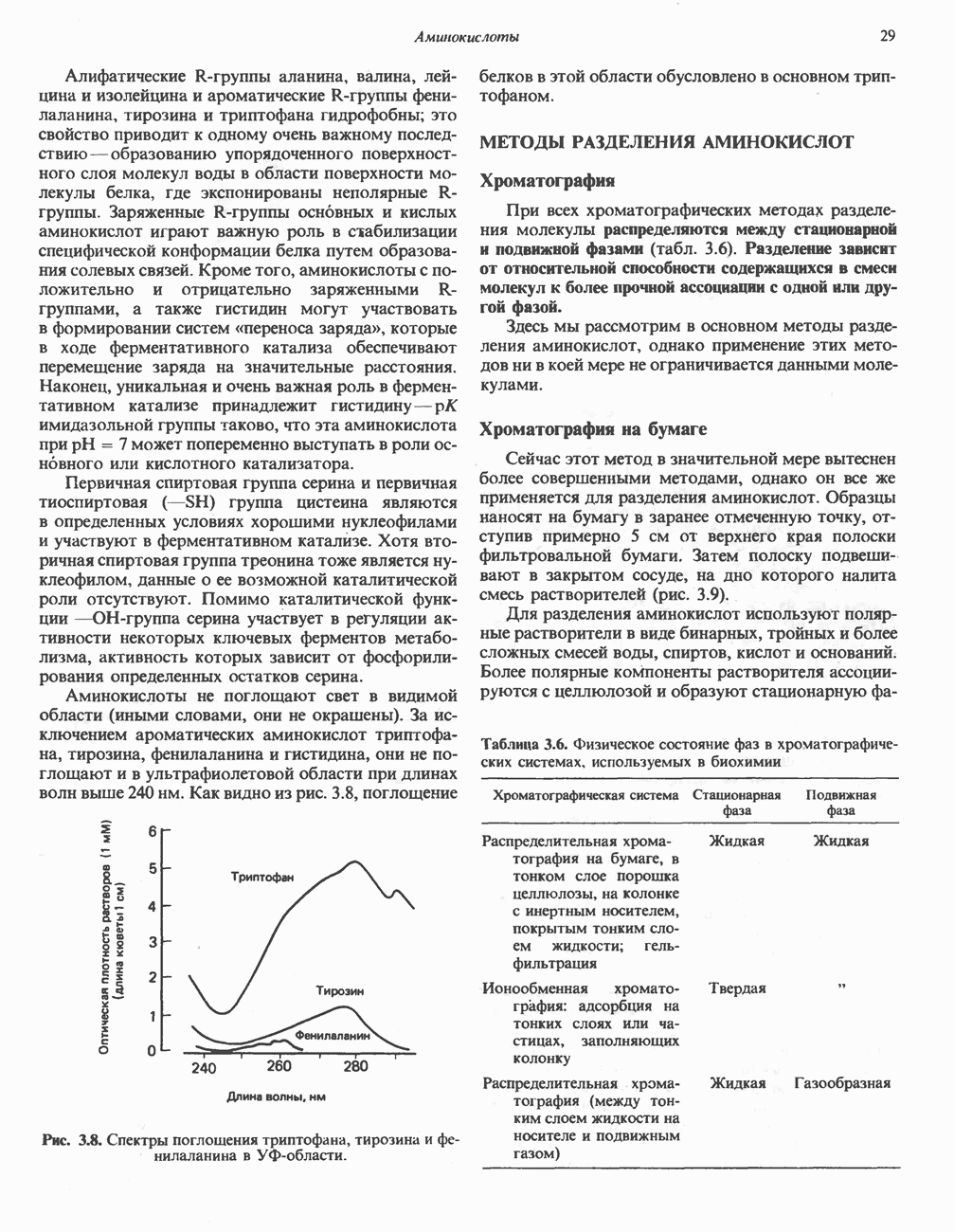
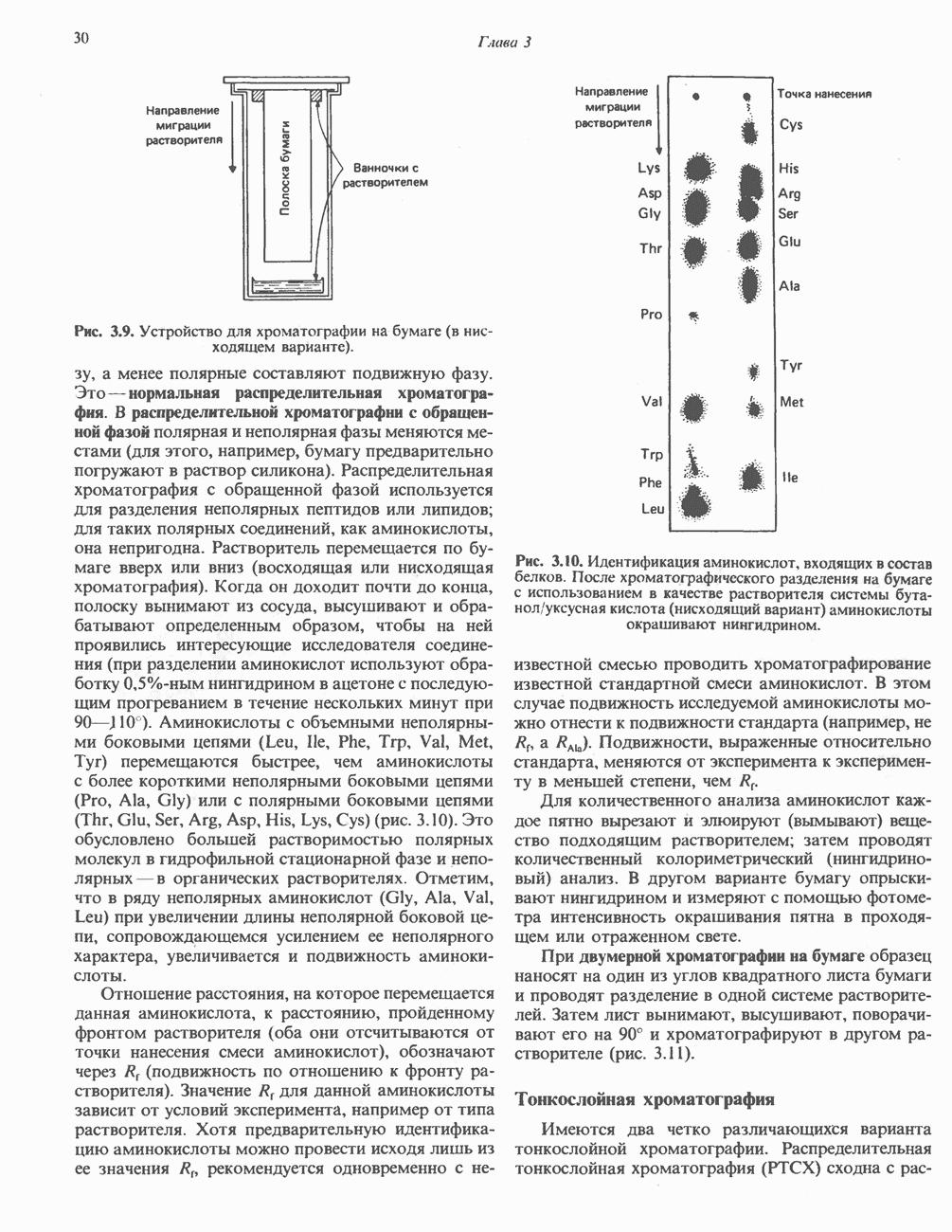
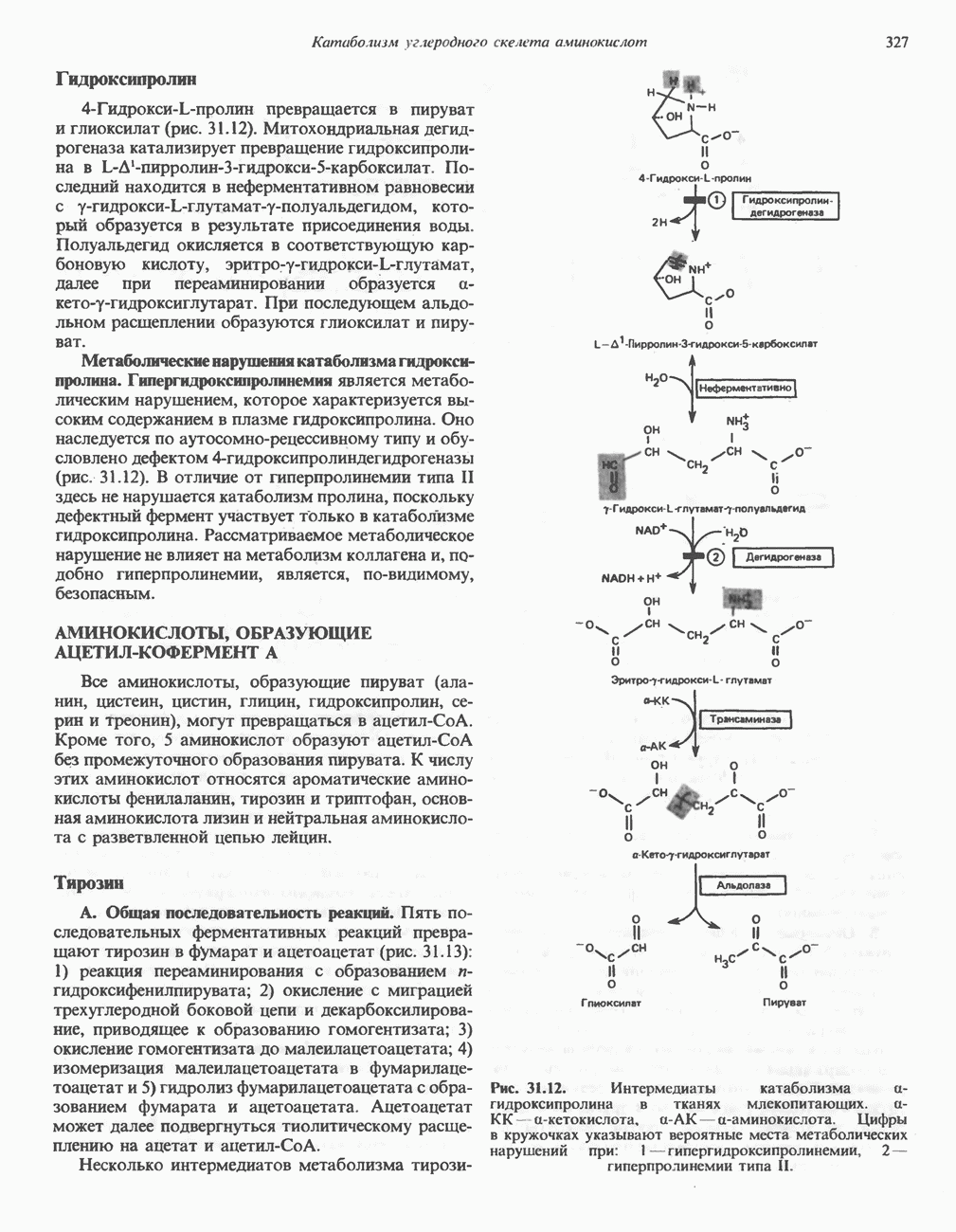
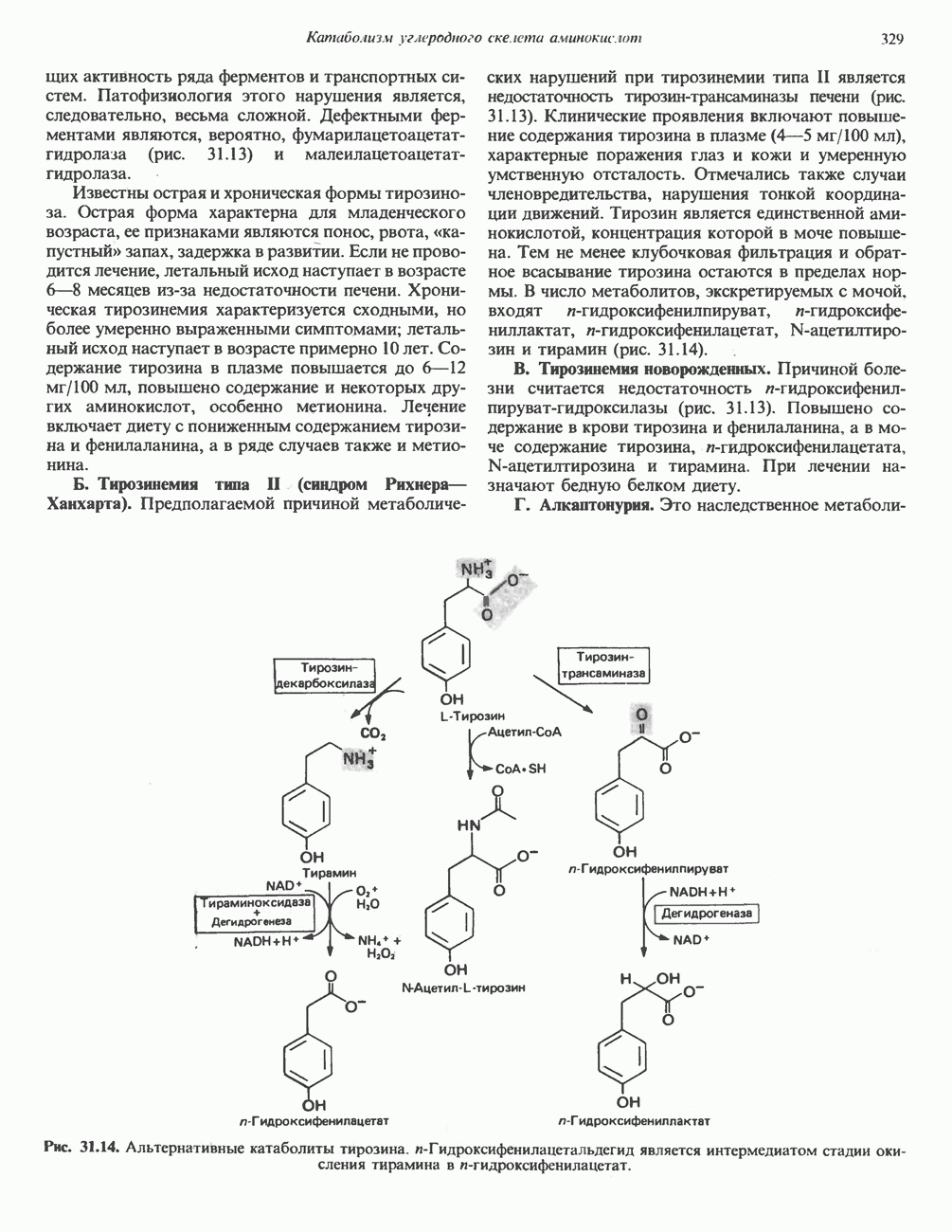
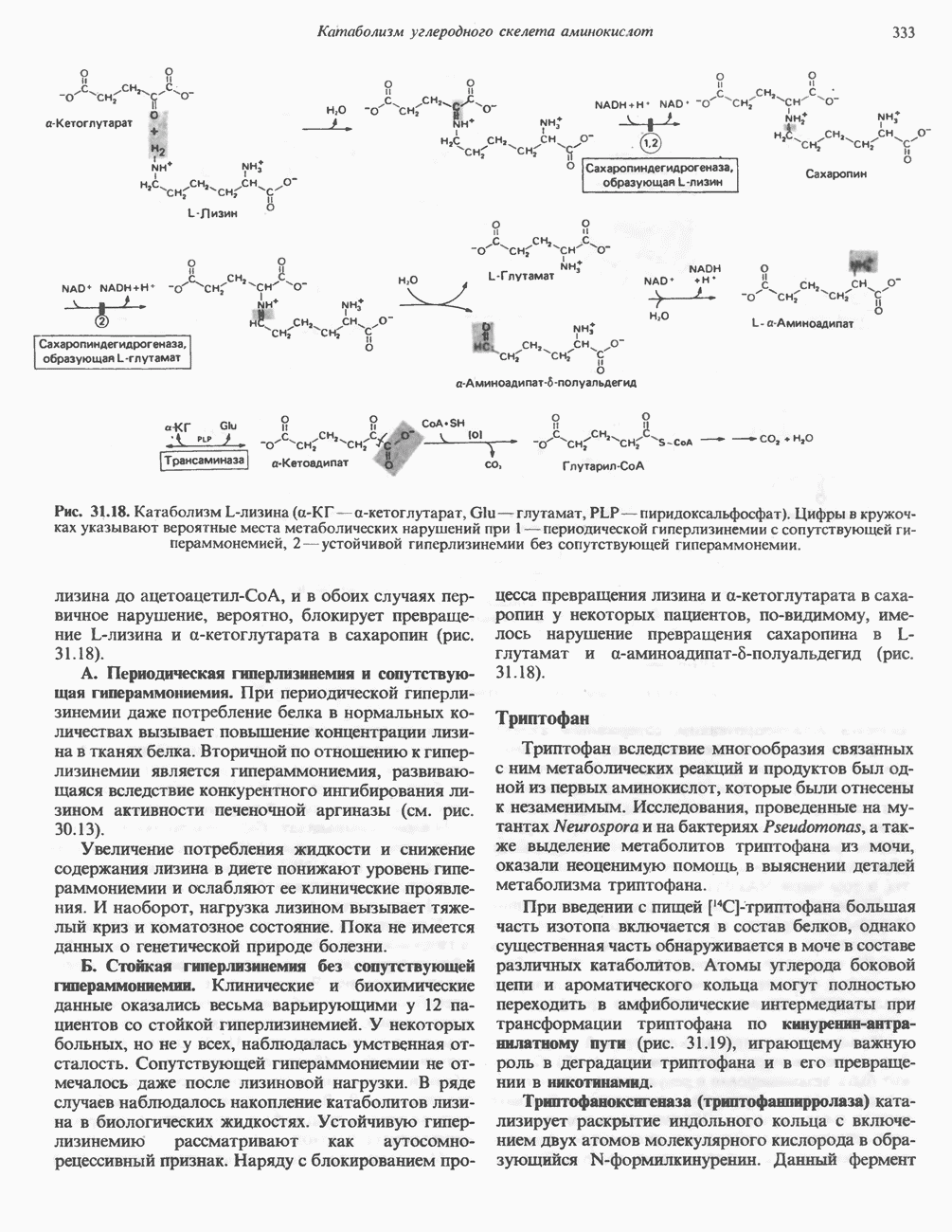
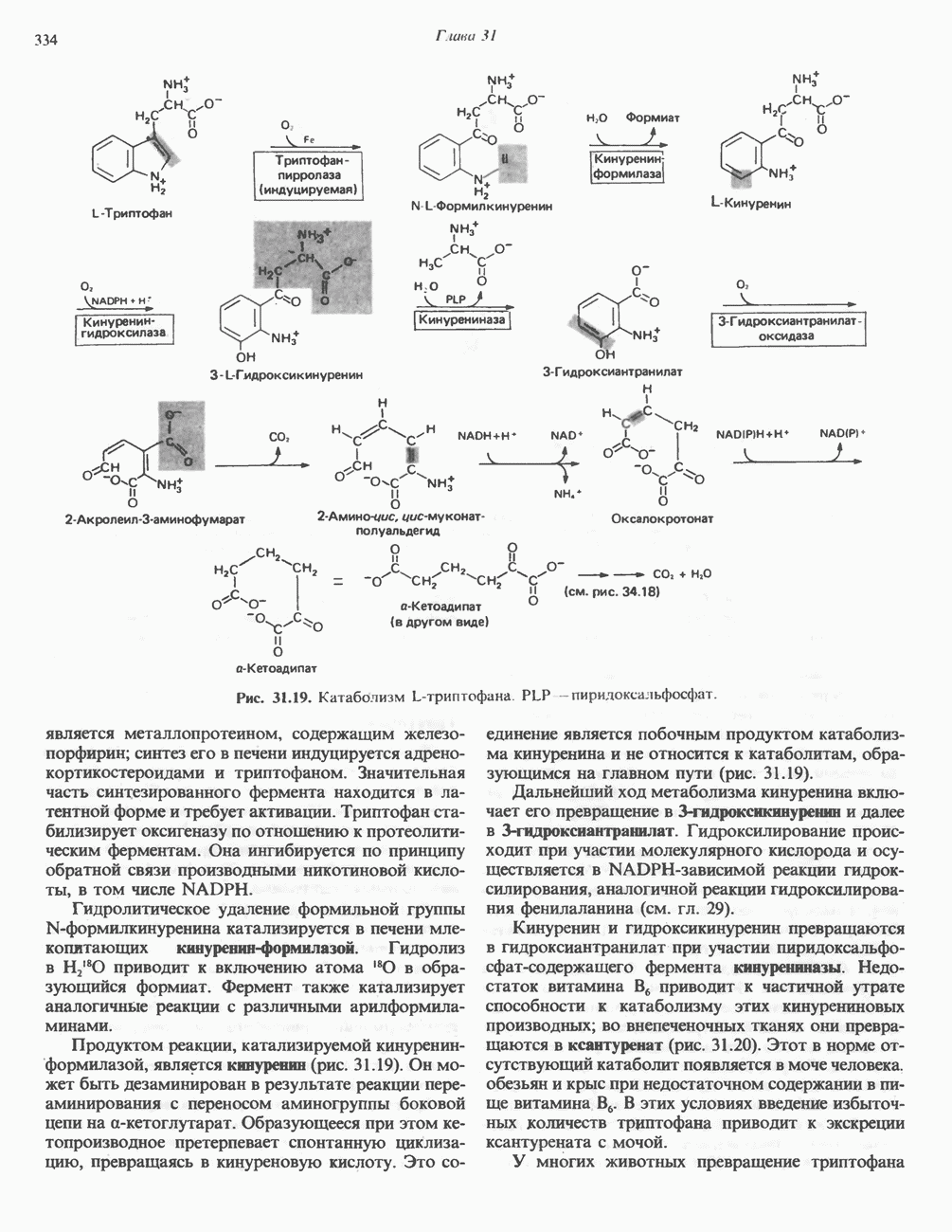
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
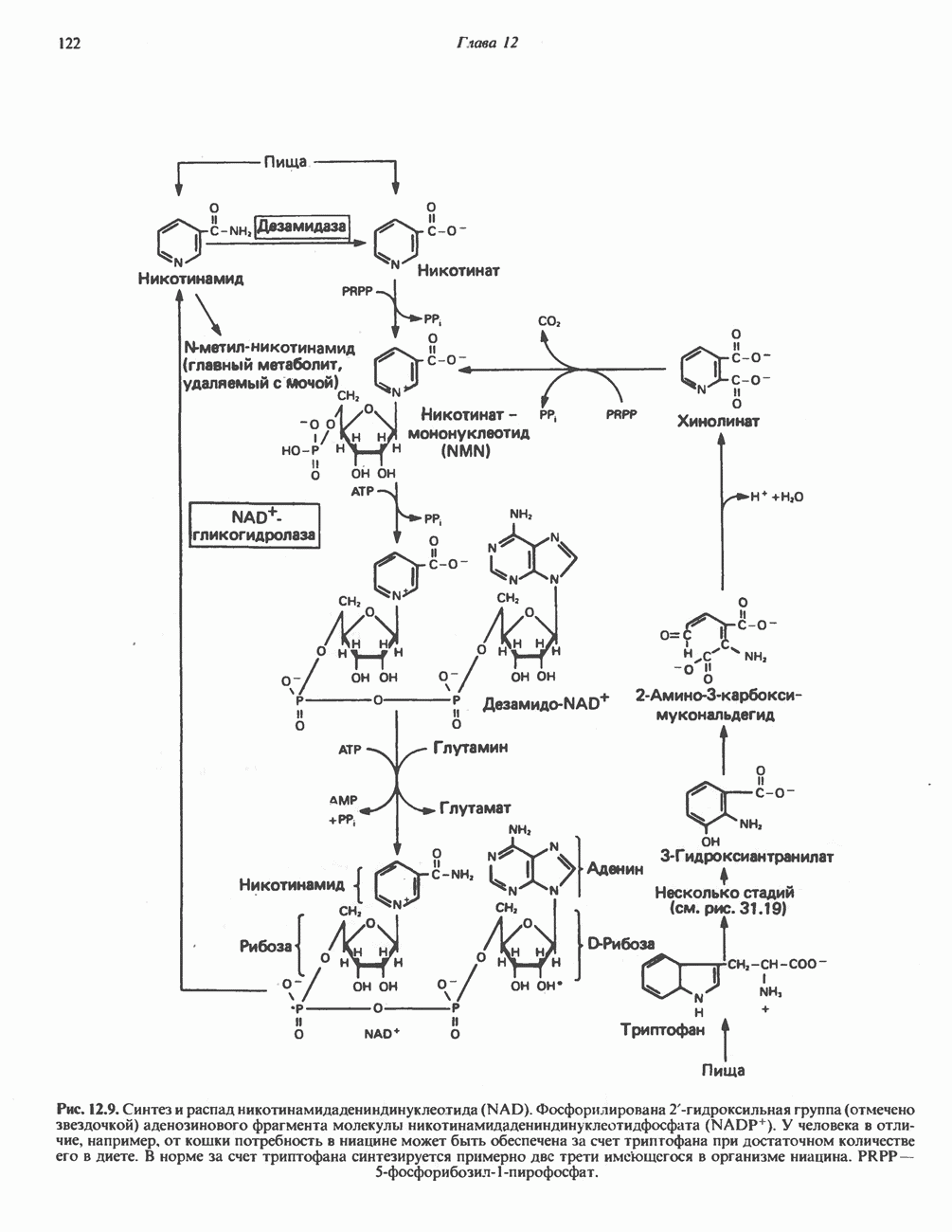
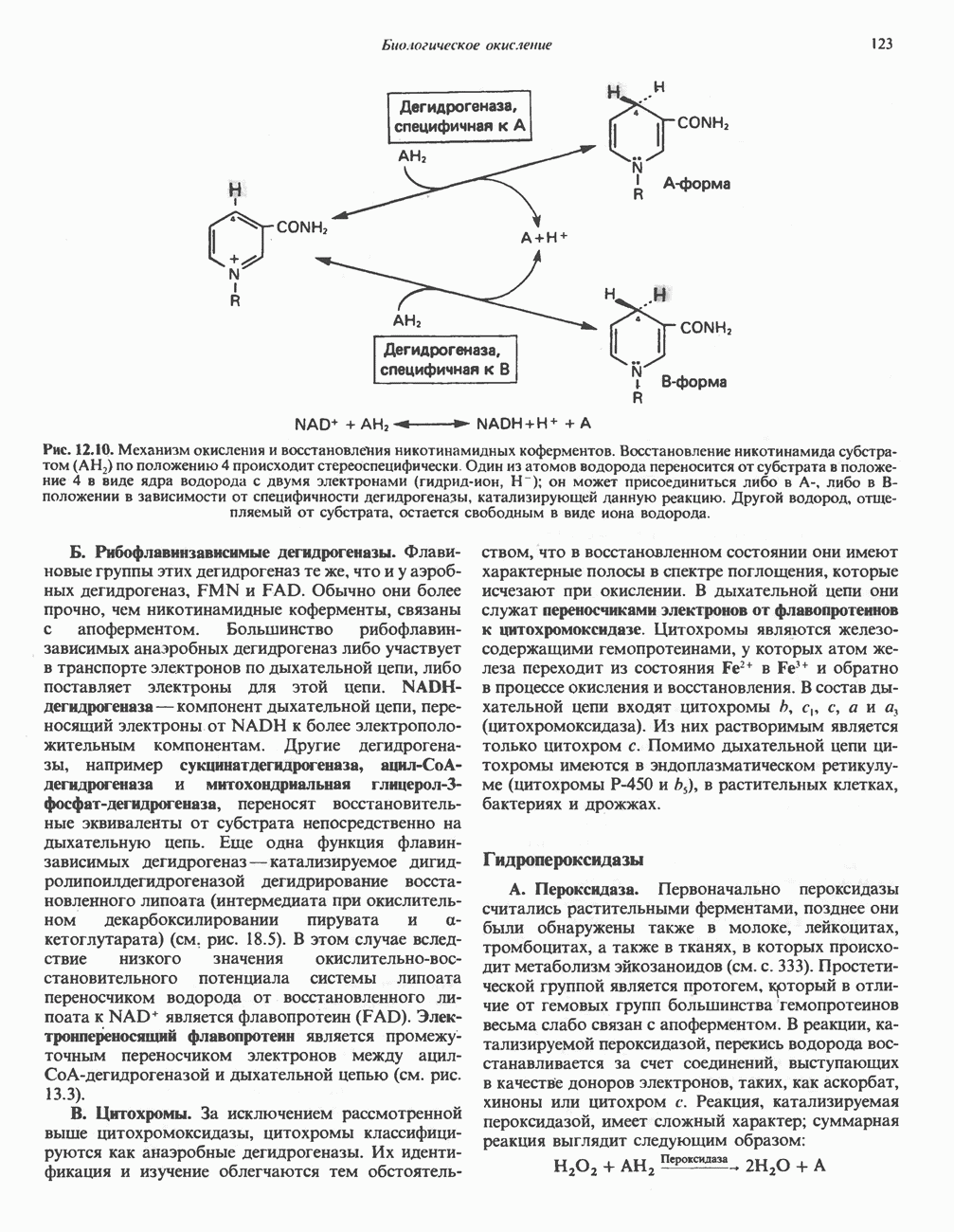
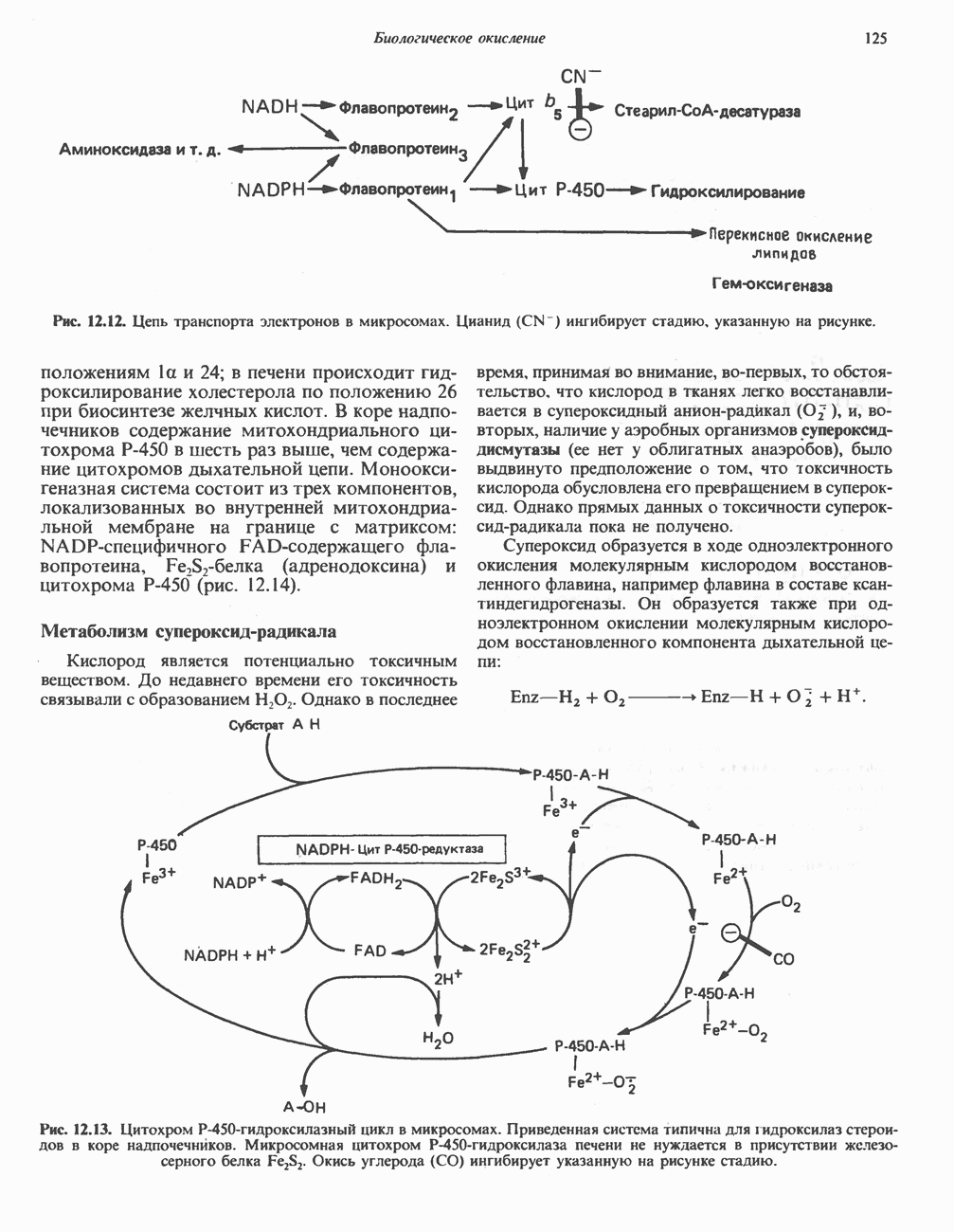
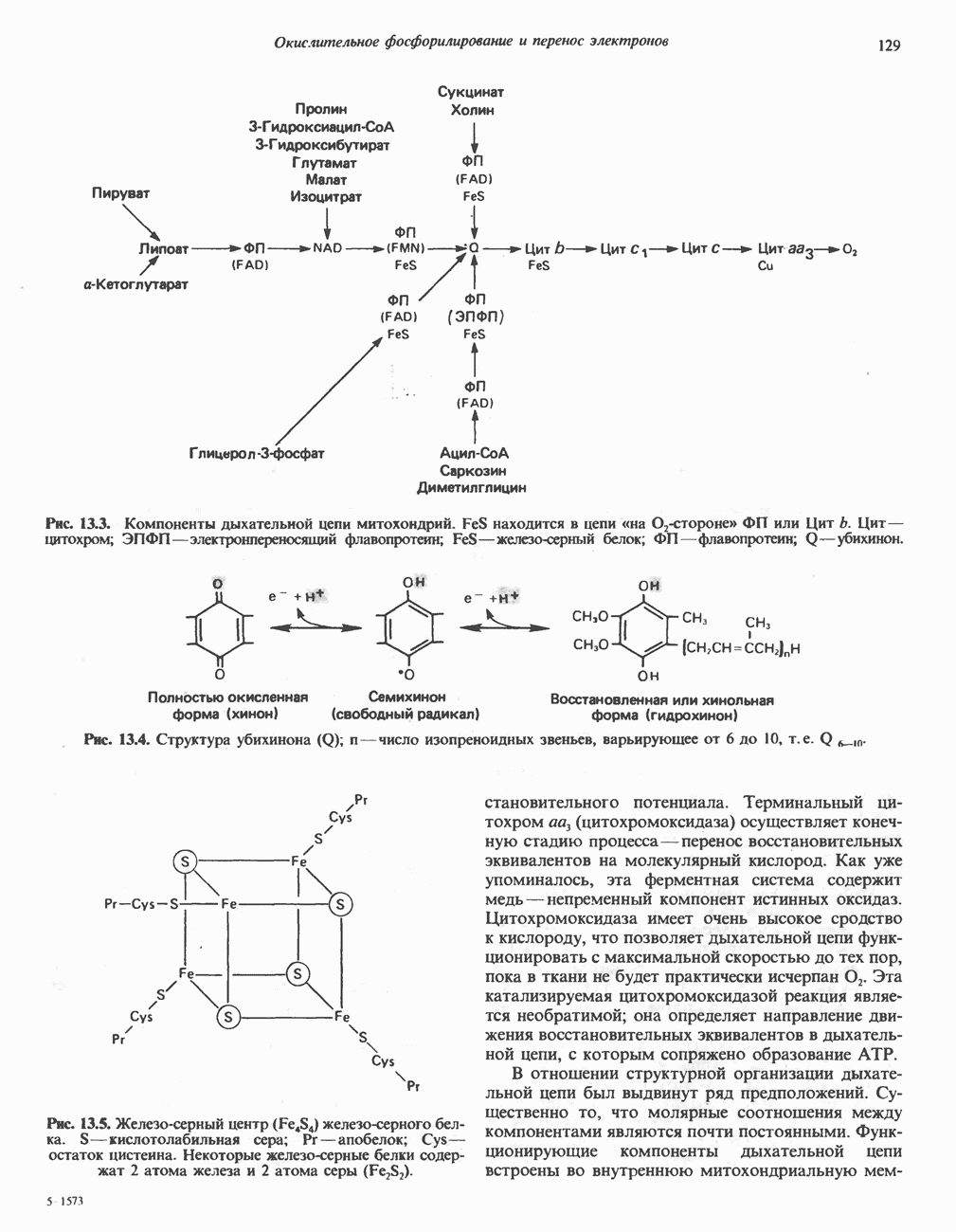
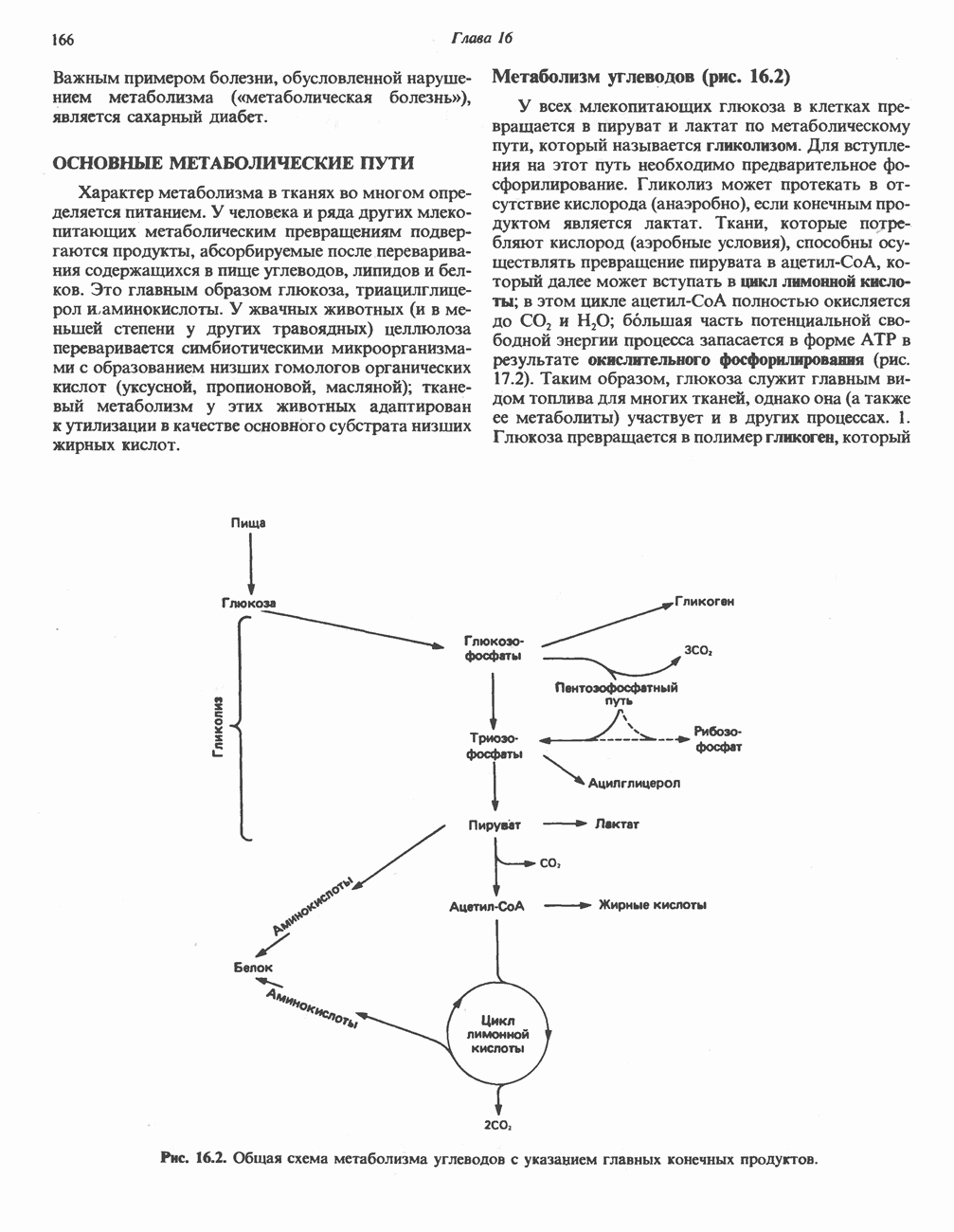
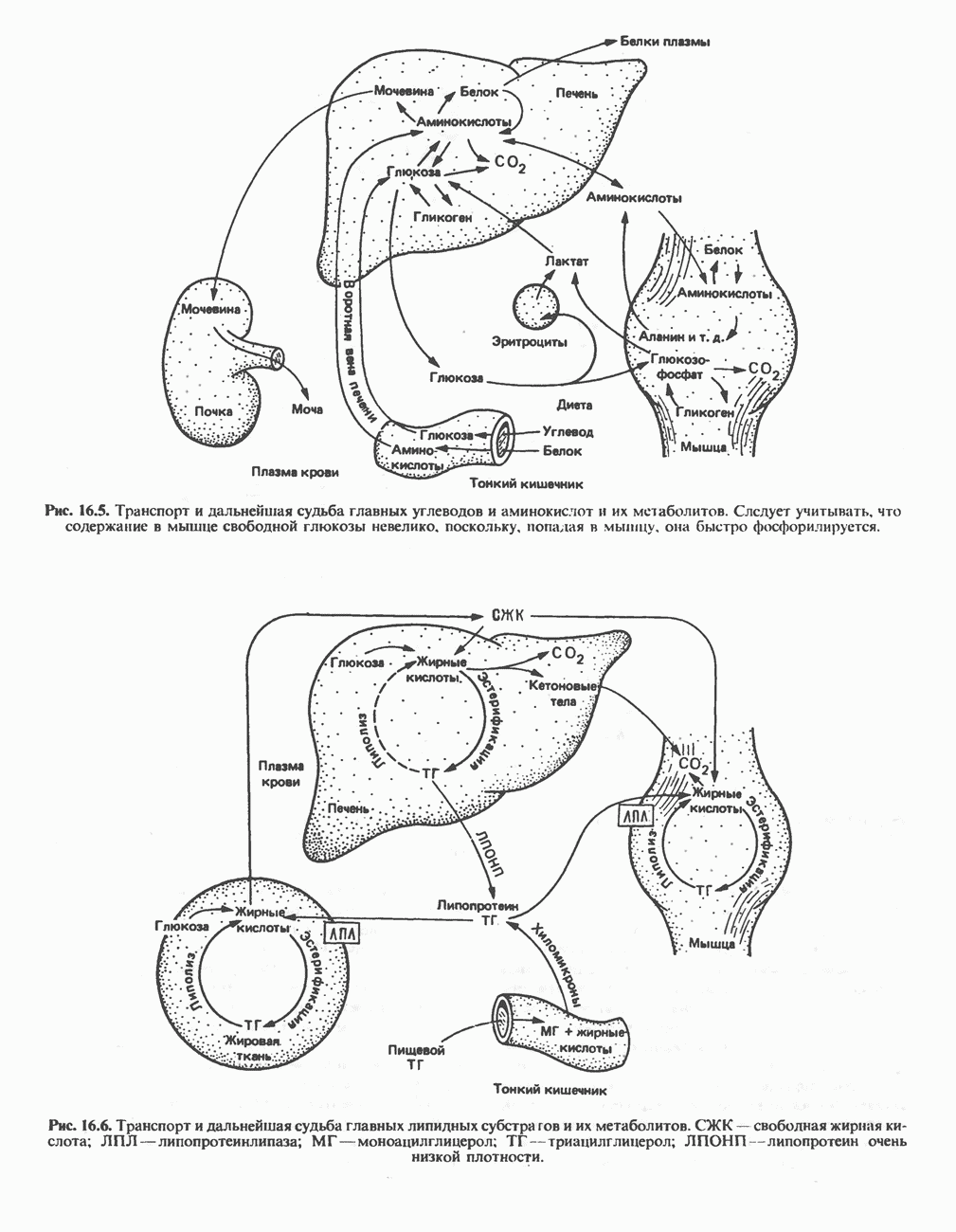
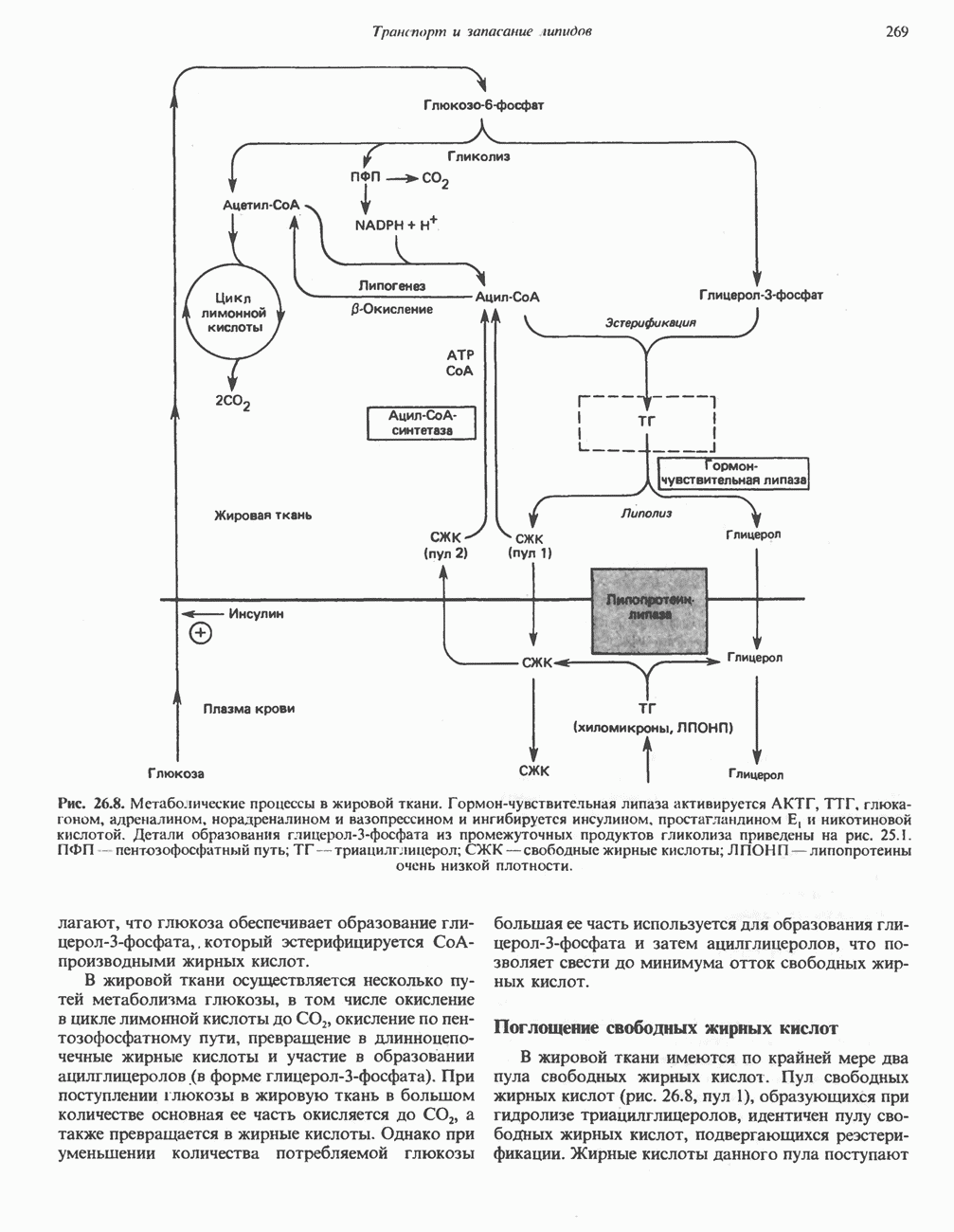
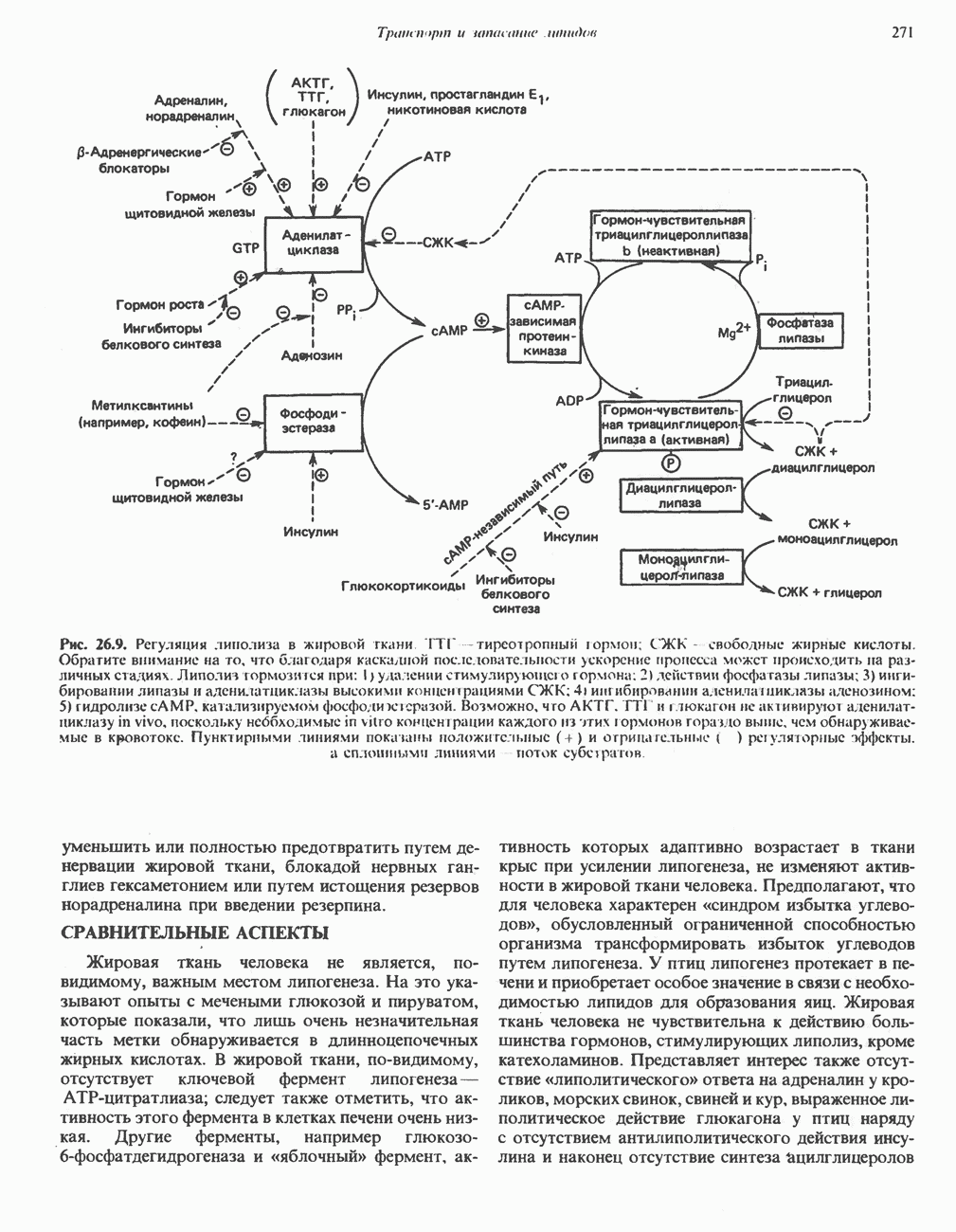
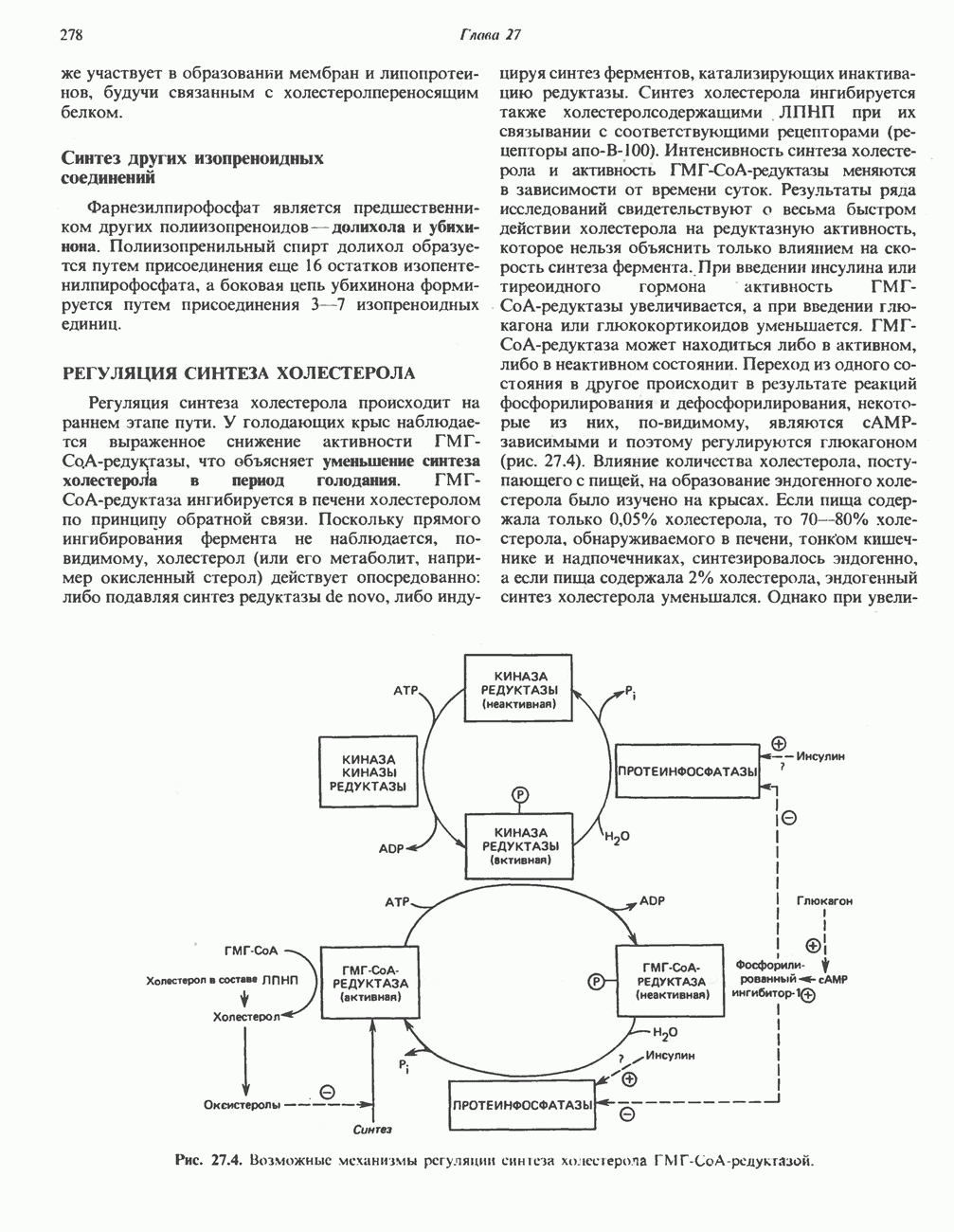
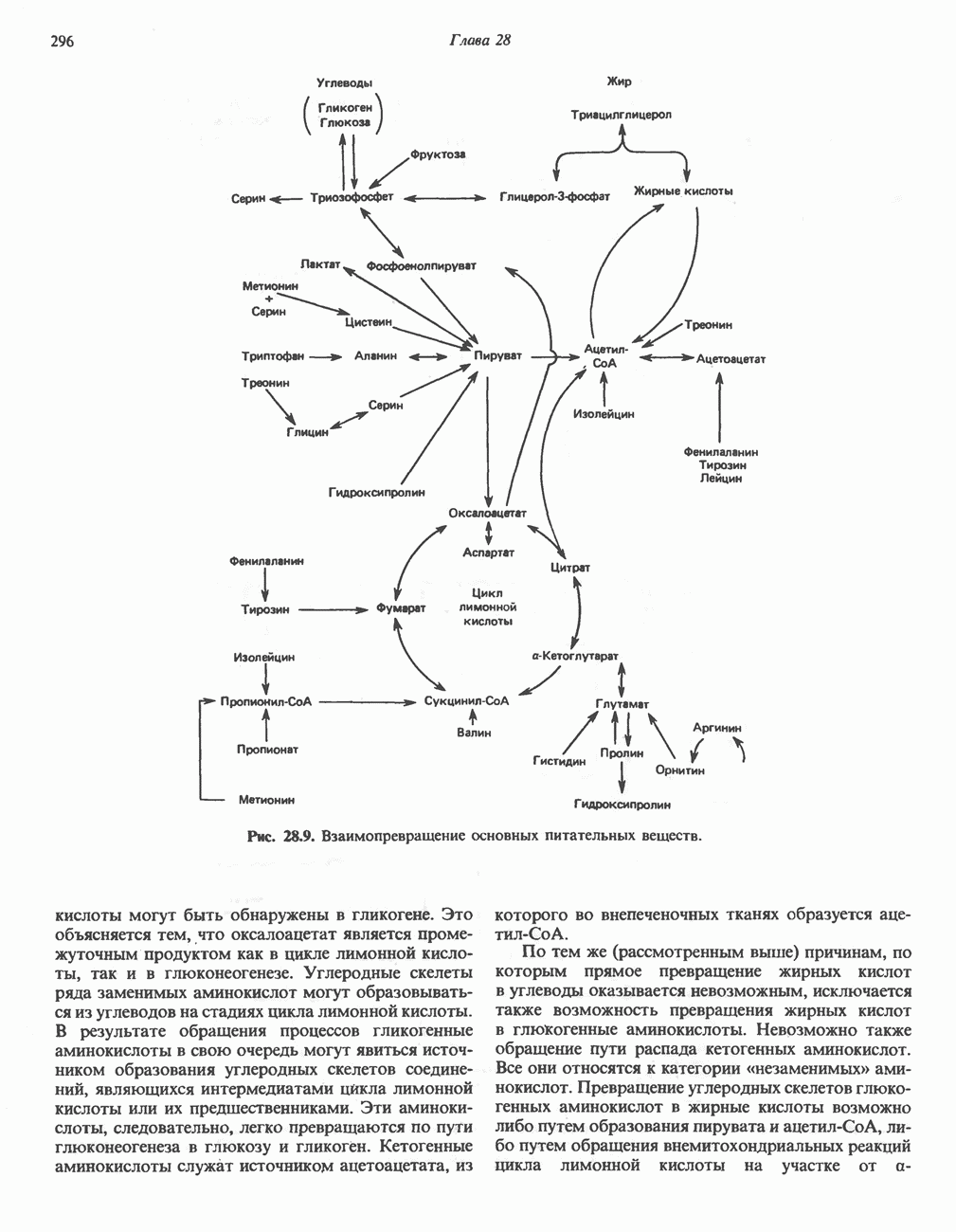
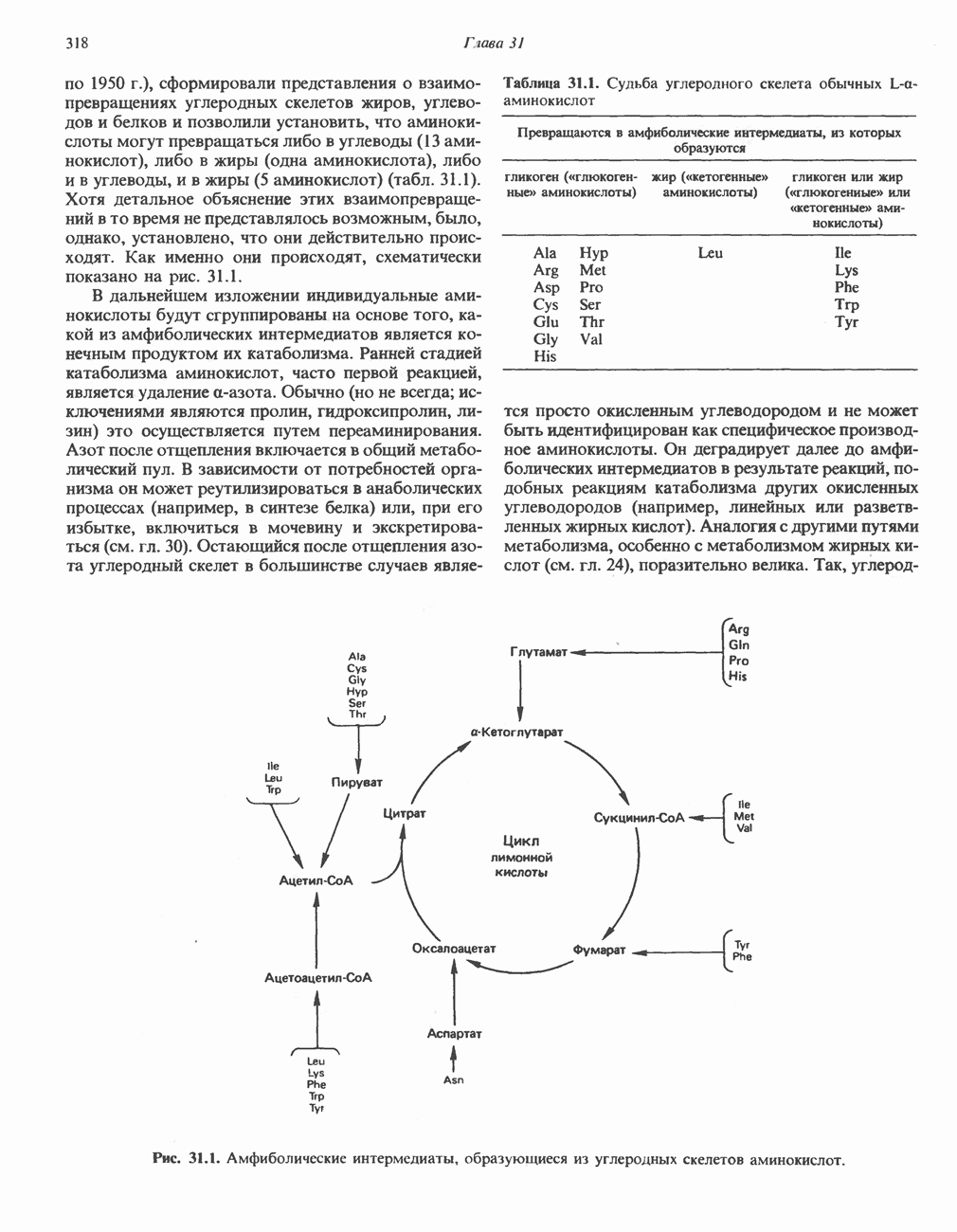
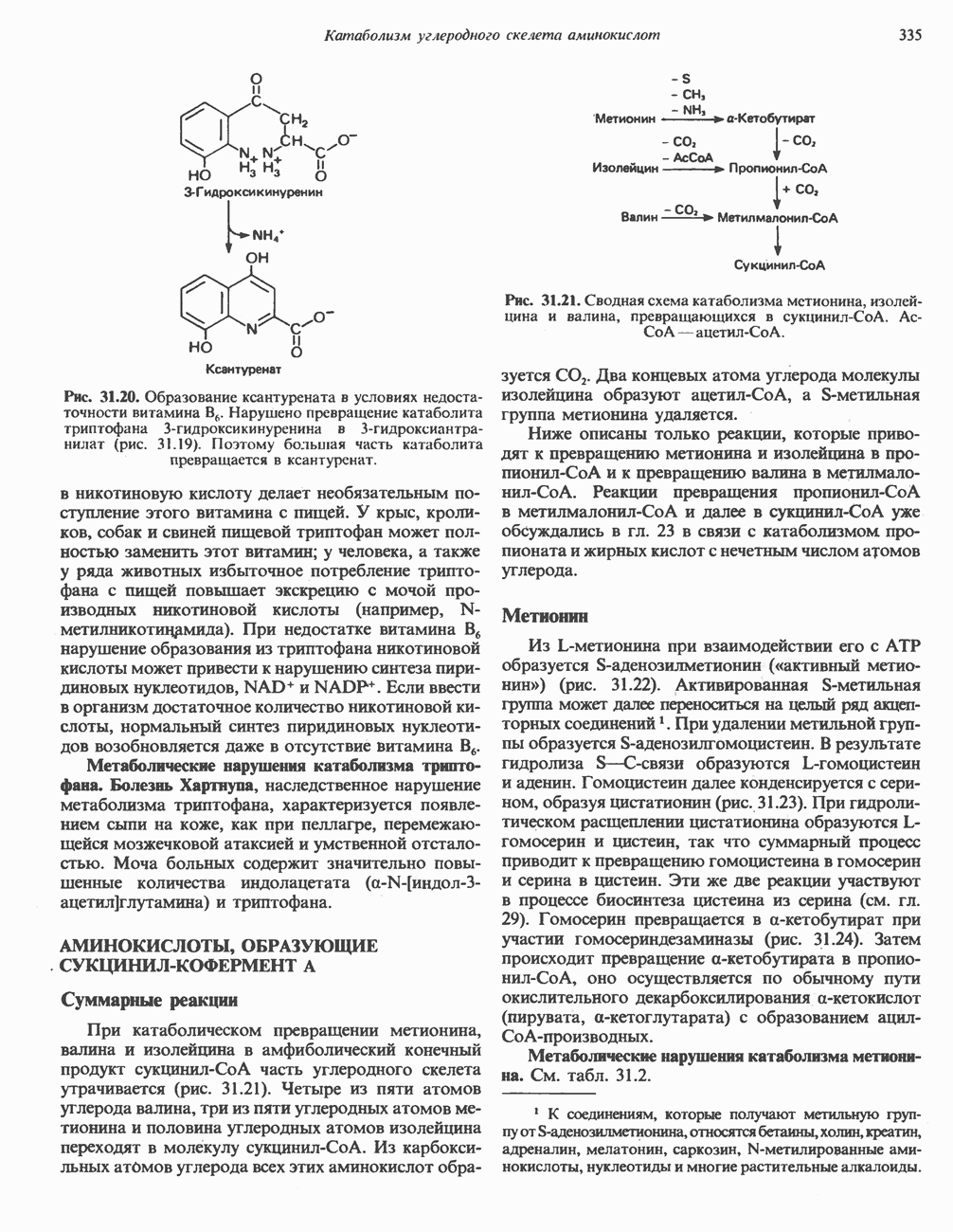
АМИНОКИСЛОТЫ, ОБРАЗУЮЩИЕ СУКЦИНИЛ-КОФЕРМЕНТ АСуммарные реакцииПри катаболическом превращении метионина, валина и изолейцина в амфиболический конечный продукт сукцинил-СоА часть углеродного скелета утрачивается (рис. 31.21). Четыре из пяти атомов углерода валина, три из пяти углеродных атомов метионина и половина углеродных атомов изолейцина переходят в молекулу сукцинил-СоА.
Рис. 31.21. Сводная схема катаболизма метионина, изолейцина и валина, превращающихся в укцинил-СоА. Ас-СоА - ацетил-СоА. Из карбоксильных атомов углерода всех этих аминокислот образуется Ниже описаны только реакции, которые приводят к превращению метионина и изолейцина в пропионил-СоА и к превращению валина в метилмалонил-СоА. Реакции превращения пропионил-СоА в метилмалонил-СоА и далее в сукцинил-СоА уже обсуждались в гл. 23 в связи с катаболизмом пропионата и жирных кислот с нечетным числом атомов углерода. МетионинИз L-метионина при взаимодействии его с АТР образуется S-аденозилметионин («активный метионин») (рис. 31.22). Активированная S-метильная группа может далее переноситься на целый ряд акцепторных соединений Метаболические нарушения катаболизма метионина. См. табл. 31.2.
Рис. 31.22. Образование (см. скан) Рис. 31.24. Превращение (см. скан) Рис. 31.23. Превращение метионина в пропионил-СоА. Лейцин, валин и изолейцинКак и можно было предполагать, учитывая структурное сходство L-лейцина, L-валина и L-изо-лейцина, их катаболизм на первых этапах идет по общему пути. Затем этот путь разветвляется и скелет каждой аминокислоты трансформируется по собственному пути с образованием амфиболических интермедиатов (рис. 31.25 и 31.26). В зависимости от природы этих амфиболических конечных продуктов аминокислоты относят к типу гликогенных (валин), кетогенных (лейцин) или к обоим типам (изолейцин). Многие из рассматриваемых реакций аналогичны реакциям катаболизма жирных кислот с линейными и разветвленными цепями. В силу сходства начальных реакций катаболизма всех трех аминокислот, которое видно на рис. 31.26, их удобно рассматривать вместе. В последующем тексте номера реакций будут соответствовать тем, которые приведены на рис. 31.26-31.29. А. Переаминирование. Обратимое переаминирование (реакция 1) всех трех разветвленных L-а-аминокислот в тканях млекопитающих осуществляется, вероятно, одной и той же трансаминазой. Обратимостью этой реакции объясняется возможность замены в диете L-a-аминокислот соответствующими Б. Окислительное декарбокснлирование с образованием ацил-СоА-тиоэфиров. Эта реакция (реакция 2) аналогична окислению пирувата до ацетил-СоА и Субъединицы дегидрогеназного комплекса а-кетокислот аналогичны соответствующим субъединицам пируватдегидрогеназы. Комплекс включает субъединицы с декарбоксилазной (по отношению к
Рис. 31.25. Катаболизм аминокислот с разветвленной цепью у млекопитающих. Реакции 1—3 — общие для всех трех аминокислот; далее пути их катаболизма расходятся. Перечеркнутые стрелки показывают стадии, на которых метаболизм блокирован в случае двух редко встречающихся заболеваний: 2-болезни «кленового сиропа» (нарушен катаболизм всех трех аминокислот), 3 - изовалериановой ацидемии (нарушен катаболизм лейцина). Из разветвленных a-кетокислот самым сильным ингибитором является а-кетоизока-проат (а-кетолейцин). В. Дегидрогенирование с образованием а, Реакции, специфичные для катаболизма лейцина (рис. 31.27)Реакция
Рис. 31.26. Сходство трех первых реакций катаболизма лейцина, валина и изолеицина. Обратите внимание, что реакции 2 и 3 сходны также с соответствующими реакциями катаболизма жирных кислот. Подобная аналогия обнаруживается и на ряде последующих стадий (см. последующие рисунки). того факта, что при превращении концевой изопропильной группы лейцина в ацетоацетат происходит «фиксация» (т.е. ковалентное связывание) 1 моля Реакция Реакция Реакции, специфичные для катаболизма валина (рис. 31.28) Реакция 4V: гидратация метилакрилил-СоА. Эта реакция идущая достаточно быстро и в отсутствие фермента, катализируется кротоназой, представляющей собой гидролазу с широкой специфичностью, действующую на Реакция 5V: деацилированне Реакция 6V: окисление Р-гидроксиизобутирата. В тканях млекопитающих содержится фермент,
Рис. 31.27. Катаболизм Р-метилкротонил-СоА, образующегося из L-лейцина (см. рис. 31.26). Звездочкой отмечены атомы углерода, поступившие из катализирующий NAD+-зависимое окисление первичной спиртовой группы Р-гидроксиизобутирата в альдегидную (реакция 6V, рис. 31.28) с образованием метилмалонового альдегида. Реакция легко обратима. Реакция 7V: дальнейшие превращения метилмалонового полуальдегида. Дальнейший метаболизм метилмалонового полуальдегида в тканях млекопитающих может идти по двум путям: в результате переаминировання из него образуется Р-аминоизобутират (реакция 7V, рис. 31.28), а при превращении по пути, изображенному на рис. 31.28 (реакции 8V-10V), - в сукцинил-СоА. Переаминирование с образованием а-аминоизо-бутирата—аминокислоты, в норме присутствующей в моче,— осуществляется в различных тканях млекопитающих, в том числе в почках. Второй основной путь — окисление в метилмалонат с последующим образованием метилмалонил-СоА и изомеризацией последнего в сукцинил-СоА (реакции 8V-10V, рис. 31.28). Изомеризация (реакция 10V, рис. 31.28) протекает при участии аденозилко-баламинового кофермента, она катализируется метилмалонил-СоА-мутазой. Эта реакция важна для катаболизма не только валина, но и пропионил-СоА-катаболита изолейцина (рис. 31.29). В условиях дефицита кобаламина (витамина Реакции, специфичные для катаболизма изолейцина (рис. 31.29)Подобно тому как это было в случае валина и лейцина, первые сведения о катаболизме изолейцина были получены в ходе наблюдений над животными, которые содержались на различных рационах; в результате были выявлены гликогенные и слабокетогенные свойства изолейцина. Возможность синтеза гликогена из изолейцина была подтверждена при использовании соединений показано, что углеродный скелет изолейцина расщепляется с образованием ацетил-СоА и пропионил-СоА (рис. 31.29). Реакция 41: гидратация тиглил-СоА. Эта реакция протекает аналогично реакции катаболизма валина (реакция 4V, рис. 31.28). Она катализируется кротоназой тканей млекопитающих.
Рис. 31.28. Катаболизм метакрилил-СоА, образующегося из L-валииа (см. рис. 31.26). а-КК - а-кетокислота, а-АК - а-аминокислота. Реакция 51: дегидрогенирование а-метил-р-гидроксибутирил-СоА. Реакция аналогична реакции 5V катаболизма валина (рис. 31.28). Напомним что в ходе катаболизма валина соответствующий ацил-СоА сначала гидролизуется и лишь затем окисляется. Реакция 61: тнолиз а-метилацетоацетил-СоА. Тиолитическии разрыв ковалентной связи между углеродными атомами 2 и 3 в молекуле а-метилацетоацетил-СоА катализируется Метаболические нарушения катаболизма разветвленных аминокислот (лейцина, валина и изолейцина) Известны четыре метаболических нарушения. Наиболее обстоятельно изучена болезнь «кленового сиропа» (названная по характерному запаху мочи больных). Описано более 50 случаев заболевания. Частота его оценивается в 5—10 случаев на миллион новорожденных. Остальные три болезни — гипервалинемия,
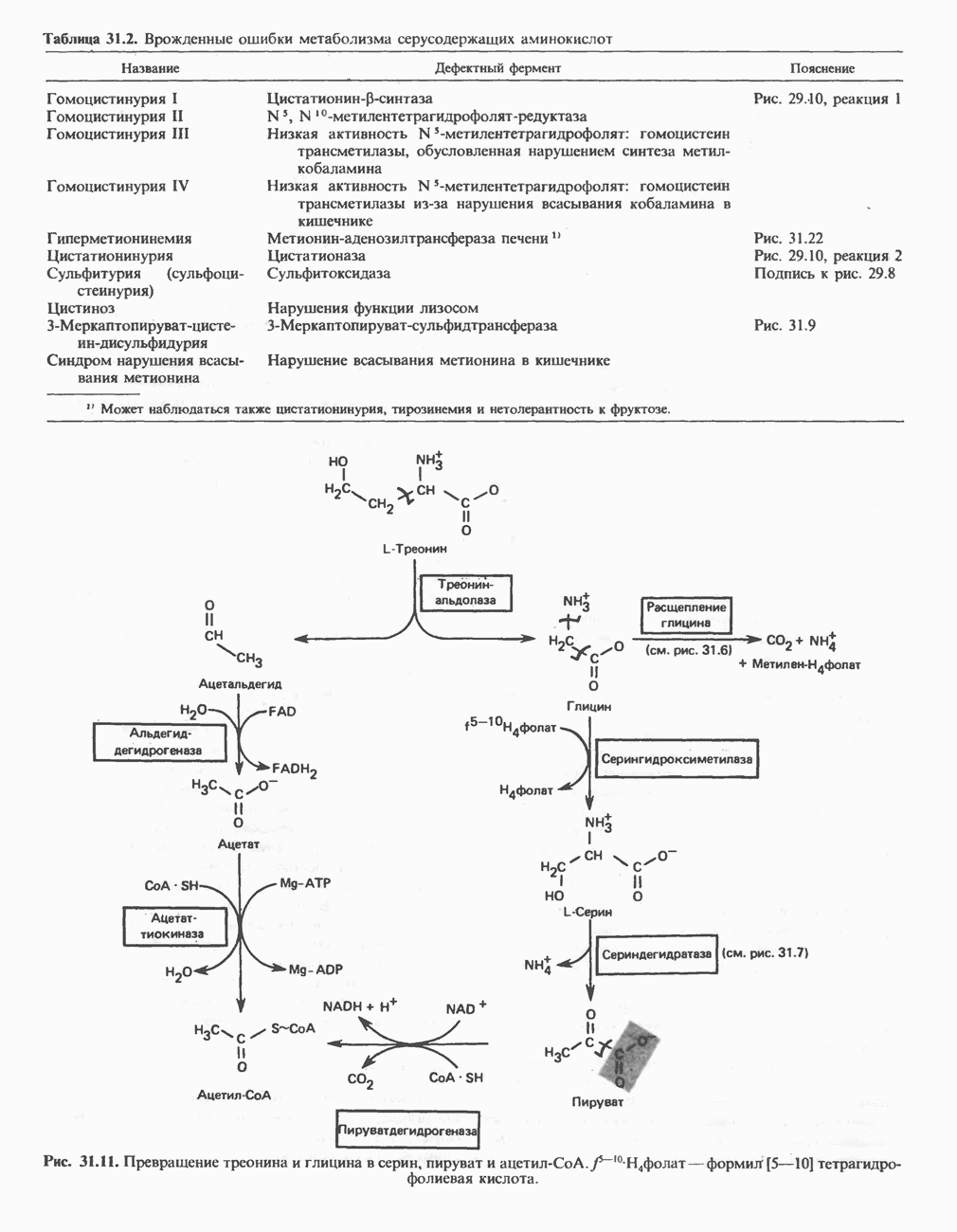
Рис. 31.29. Катаболизм тиглил-СоА, образующегося из L-изолейцина (см. рис. 31.26). скачкообразная кетонурия (повышение содержания в моче кетокислот с разветвленной цепью) и изовалериановая ацидемия — наблюдались менее чем у пяти детей каждая. A. Гипервалинемия. Это метаболическое заболевание характеризуется повышенным содержанием в плазме валина (но не лейцина или изолейцина), что обусловлено нарушением переаминировання валина с образованием а-кетоизовалерата (реакция 1, рис. 31.26). При этом переаминирование лейцина и изолейцина (реакция 1, рис. 31.26) не нарушается. Б. Болезнь «кленового сиропа». Наиболее ярким отличительным признаком этой наследственной болезни является характерный запах мочи больного, напоминающий запах кленового сиропа или жженого сахара. В плазме и в моче сильно повышается содержание аминокислот с разветвленной цепью — лейцина, изолейцина и валина, а также соответствующих им a-кетокислот. Поэтому болезнь «кленового. сиропа» иногда называют кетонурией разветвленных кетокислот. Отмечено также присутствие в моче разветвленных а-гидроксикислот, образующихся при восстановлении а-кетокислот. Характерные признаки болезни проявляются в конце первой недели после рождения. Наряду с описанными выше биохимическими нарушениями возникают трудности при кормлении ребенка, может наблюдаться рвота. Иногда наблюдается летаргия. Диагноз ранее конца первой недели возможен только с помощью ферментного анализа. У выживших детей отмечены выраженные нарушения мозговой деятельности. При отсутствии лечения летальный исход наступает к концу первого года жизни. Биохимической причиной болезни является отсутствие или сильное снижение активности декарбоксилазы a-кетокислот, катализирующей превращение всех трех разветвленных Больным назначают диету с заменой белков на смесь очищенных аминокислот, не содержащую лейцина, изолейцина и валина. После того как содержание этих трех аминокислот в плазме снизится до нормального уровня, можно вводить их в пищу, например, в составе молока и других продуктов, но лишь в таком количестве, чтобы обеспечить (но не превысить) потребности в аминокислотах с разветвленной цепью. Нет сведений о том, можно ли впоследствии ослабить ограничения в диете и когда именно. Если лечение было начато в первую неделю жизни ребенка, удается значительно смягчить тяжелые проявления болезни. B. Скачкообразная кетонурия (разветвленных кетокислот). Эта болезнь является одной из форм болезни «кленового сиропа» и, вероятно, связана со сравнительно меньшими структурными изменениями декарбоксилазы Болезнь «кленового сиропа» и скачкообразная кетонурия разветвленных кетокислот могут служить иллюстрацией положения, о котором говорилось во введении к этой главе, — о мутациях, вызывающих различные изменения в структуре одного и того же фермента. Вероятно, изменения активности фермента проявляются в спектре форм заболевания, включающем выраженную форму, скачкообразную кетонурию и практически нормальное состояние. Г. Изовалериановая ацидемия. Симптомами заболевания являются запах сыра у выдыхаемого воздуха и жидкостей организма, рвота, ацидоз и даже кома; последняя может возникнуть при избыточном потреблении белка или как следствие присоединившегося инфекционного заболевания. В трех известных случаях наблюдалась умеренная задержка умственного развития. Причиной болезни является недостаточность изовалерил-СоА-дегидрогеназы (реакция 3, рис. 31.26). Накапливающийся в результате Дополнительные метаболические нарушения, связанные с катаболизмом аминокислот (пропионат, метилмалонат и витамин В12)Пропионил-СоА (рис. 31.21) может образоваться из изолейцина (рис. 31.26 и 31.29), метионина (рис. 31.23), а также из боковой цепи холестерола и жирных кислот с нечетным числом атомов углерода. Превращение Вскоре после того, как было установлено участие изомеризации метилмалонил-СоА в сукцинил-СоА, было показано, что пациенты с недостаточностью витамина В12 экскретируют с мочой большие количества метилмалоната. Метилмалоновая ацидурия прекращалась после введения достаточных количеств витамина А. Пропионовая ацидемия. Недостаточная активность пропионил-СоА-карбоксилазы характеризуется высоким содержанием пропионата в сыворотке и нарушением катаболизма пропионата в лейкоцитах. Лечение состоит в назначении бедной диеты и в принятии мер по снижению метаболического ацидоза. Б. Метилмалоновая ацидурия. Известны две формы метилмалоновой ацидурии. Одна из них прекращается после парентерального введения физиологических доз витамин Приведенные выше примеры наследственных нарушений катаболизма аминокислот относятся к сравнительно хорошо изученным заболеваниям. По этой теме имеется более обстоятельный обзор (Wellner and Meister, 1981). ЛИТЕРАТУРАBremer H.J. et al. Amino Acid Metabolism: Clinical Chemistry and Diagnosis, Urban and Schwarzenberg. 1981. Cooper A.J.L. Biochemistry of the sulfur-containing amino acids, Annu. Rev. Biochem., 1983, 52, 187. Felig P. Amino acid metabolism in man, Annu. Rev. Biochem., 1975, 44, 933. Frimter G. W. Aminoacidurias due to disorders of metabolism. (2 parts), N. Engl. J. Med., 1973, 289, 835, 895. Paxton R„ Harris R. A. Isolation of rabbit liver branched chain a-ketoacid dehydrogenase and regulation by phosphorylation, J. Biol. Chem., 1982, 257, 14433. Paxton R., Harris R.A. Regulation of branched-chain a-ketoacid dehydrogenase kinase, Arch. Biochem Biophys, 1984, 231, 48. Rosenberg L. E., Scriver C. R. Disorders of amino acid metabolism, Chapter 11. In: Metabolic Control and Disease, Bon-dy P. K., Rosenberg L. E. (eds), Saunders, 1980. Schwarz V. A Clinical Companion to Biochemical Studies, Freeman, 1978. Stanbury J. B. et al. (eds), The Metabolic Basis of Inherited Disease, 5th ed., McGraw-Hill, 1983. Wellner D., Meister A. A survey of inborn errors of amino acid metabolism and transport in man, Annu. Rev. Biochem., 1981, 50, 911.
|
 |
Оглавление
|



































































































































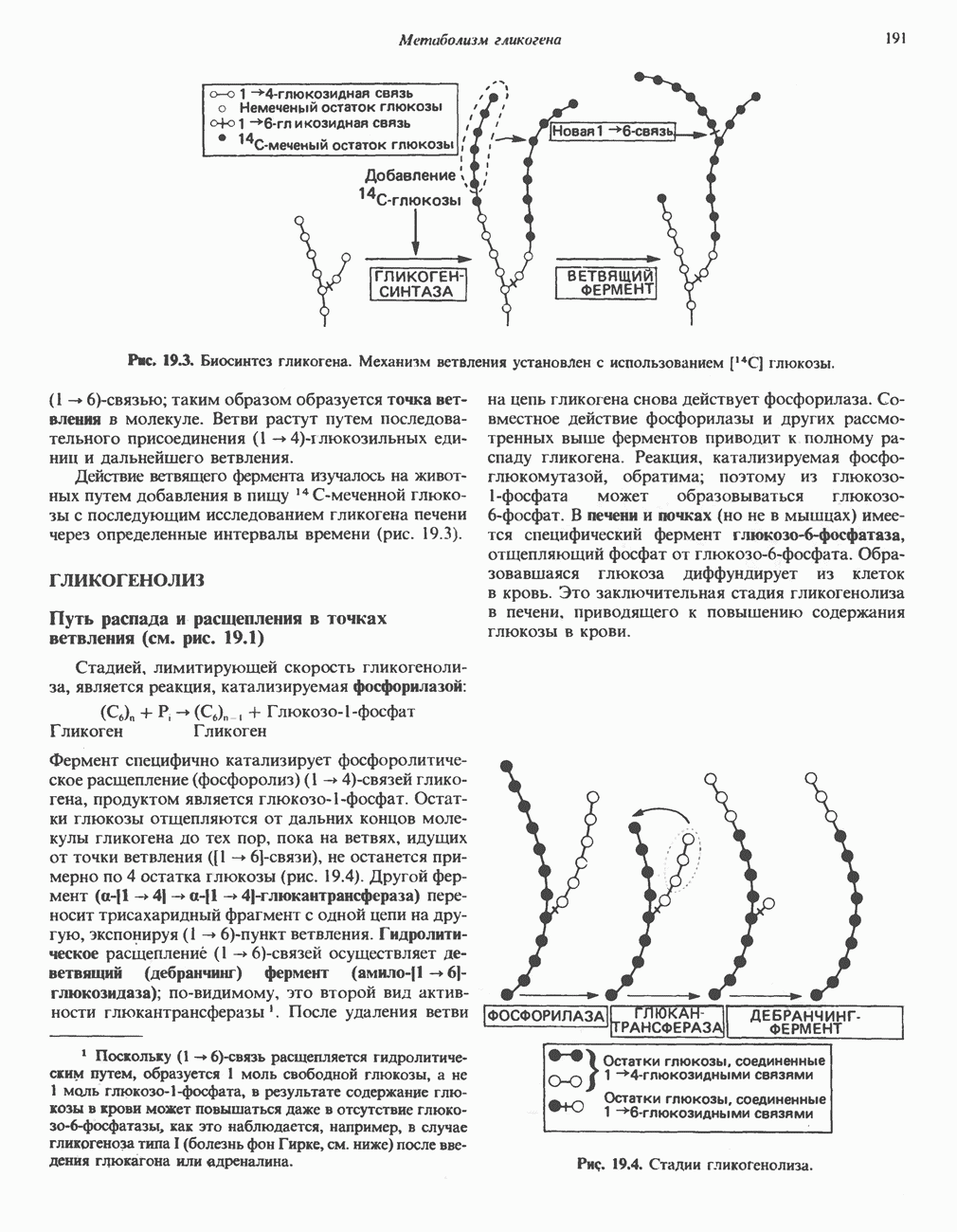
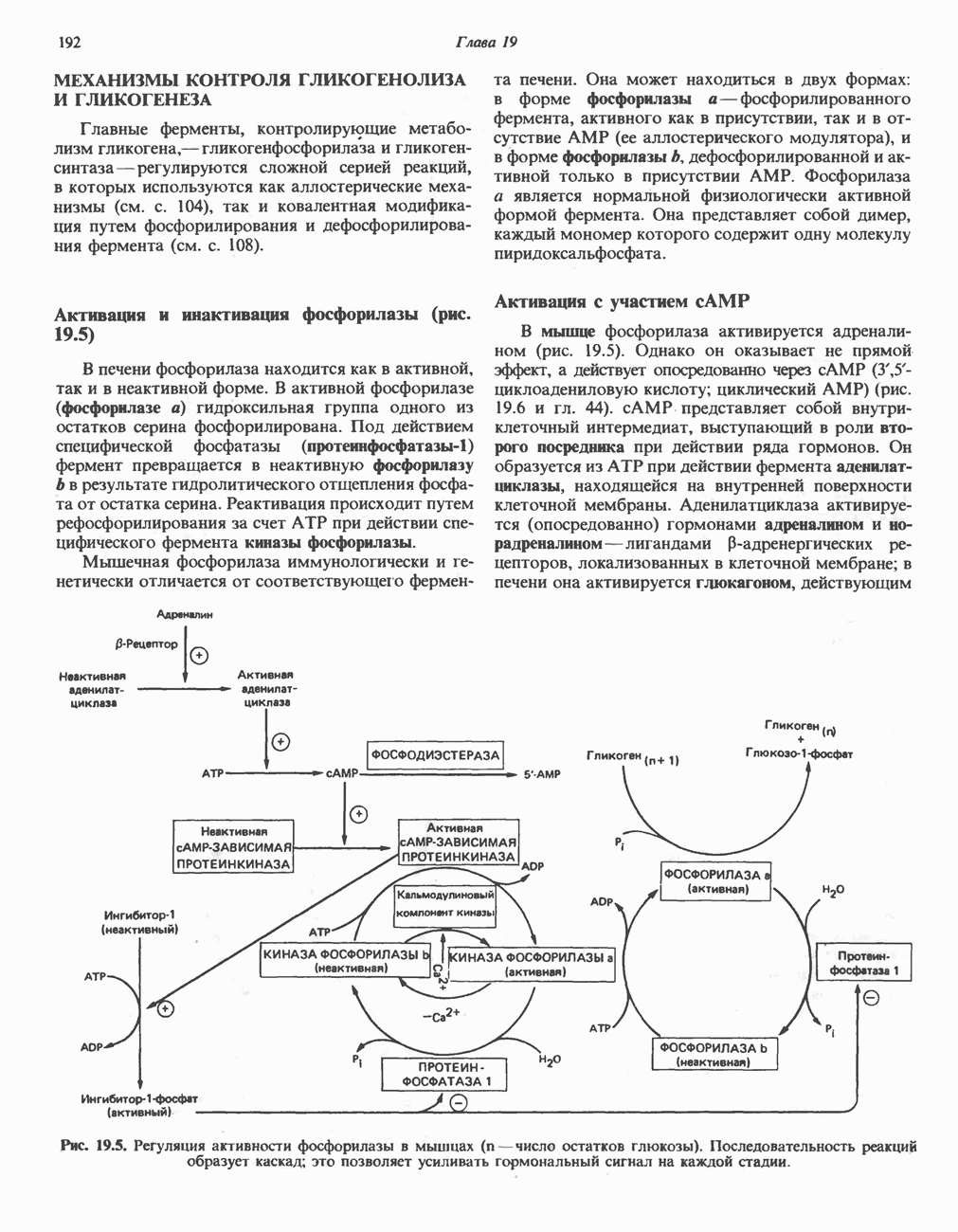

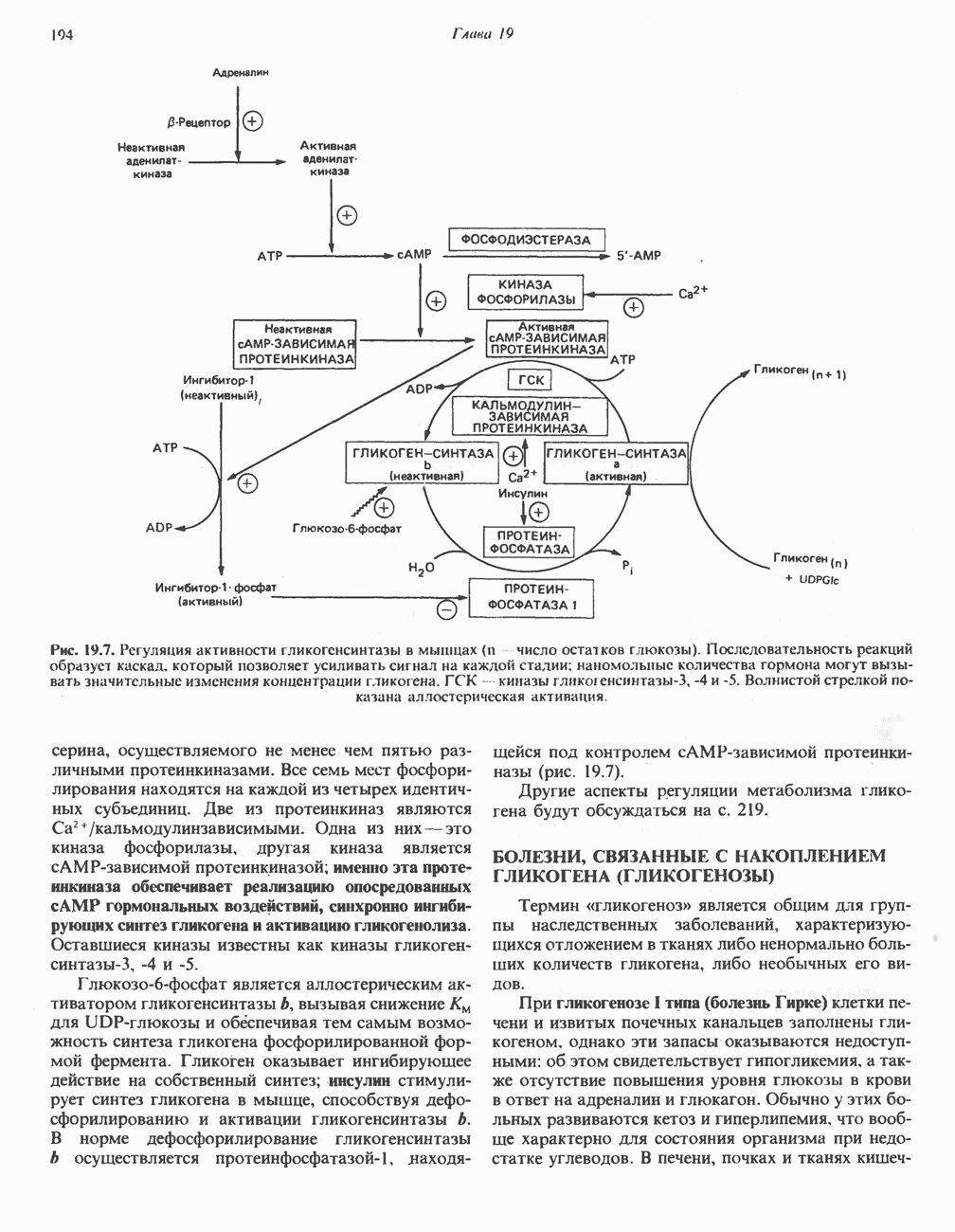


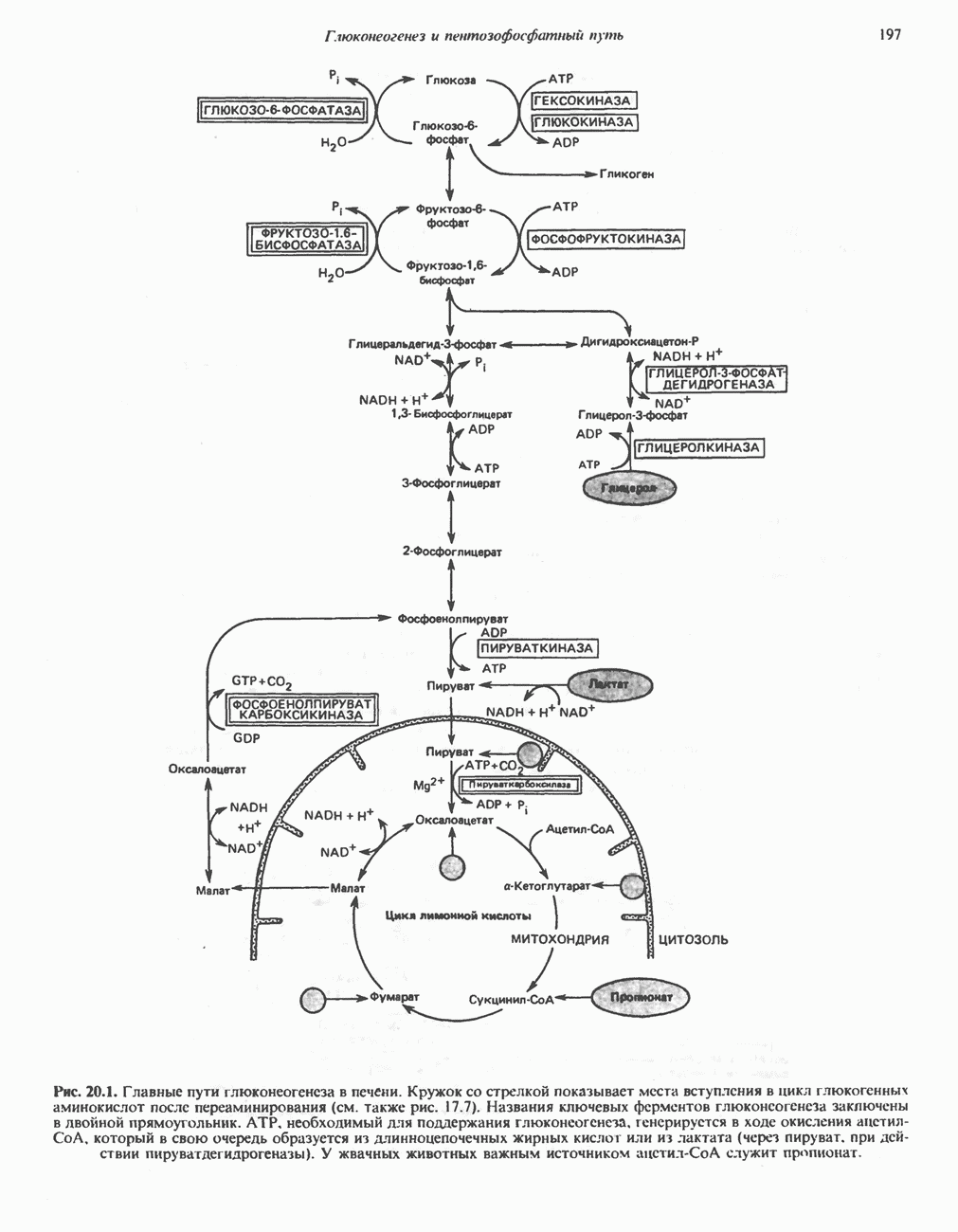











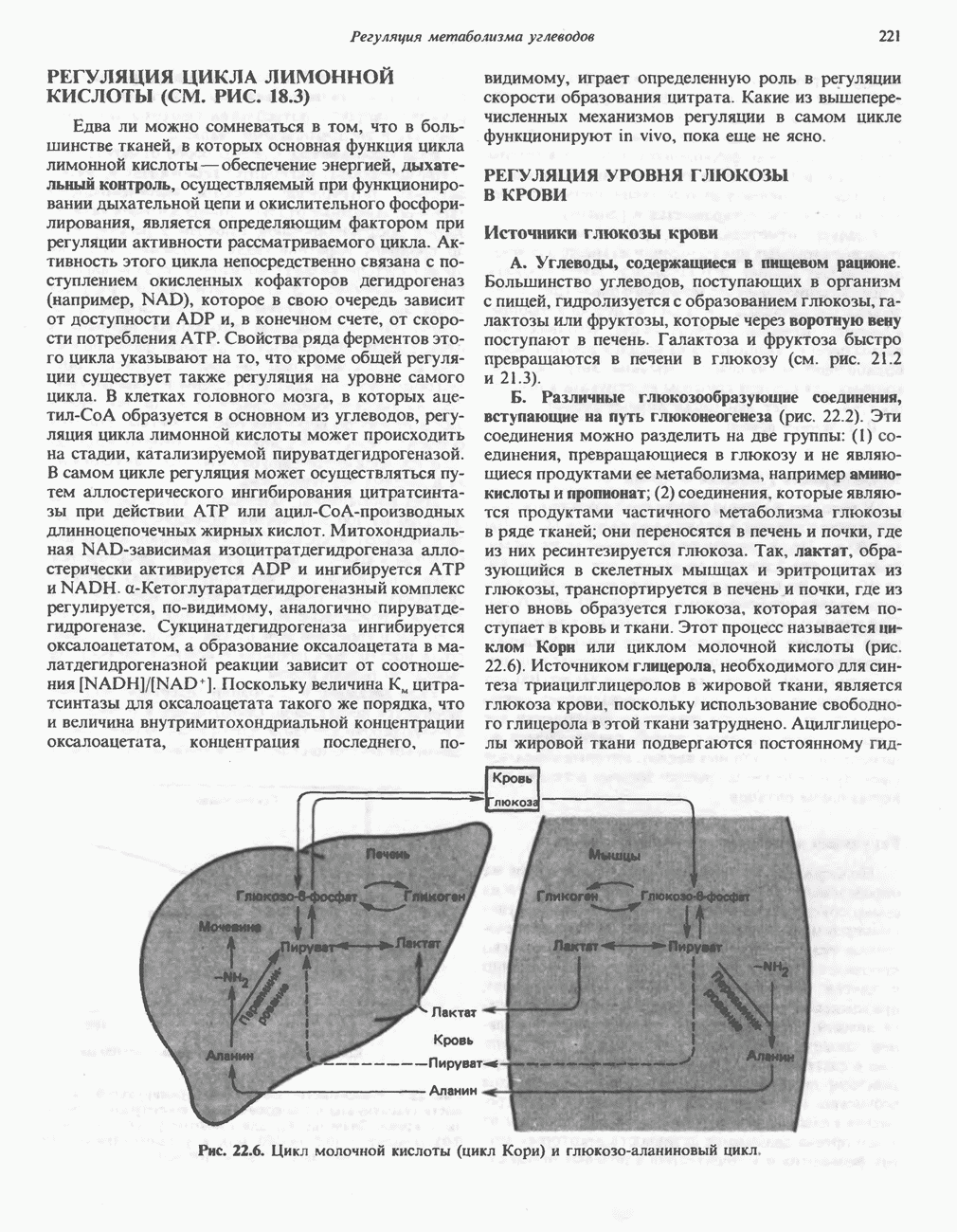





























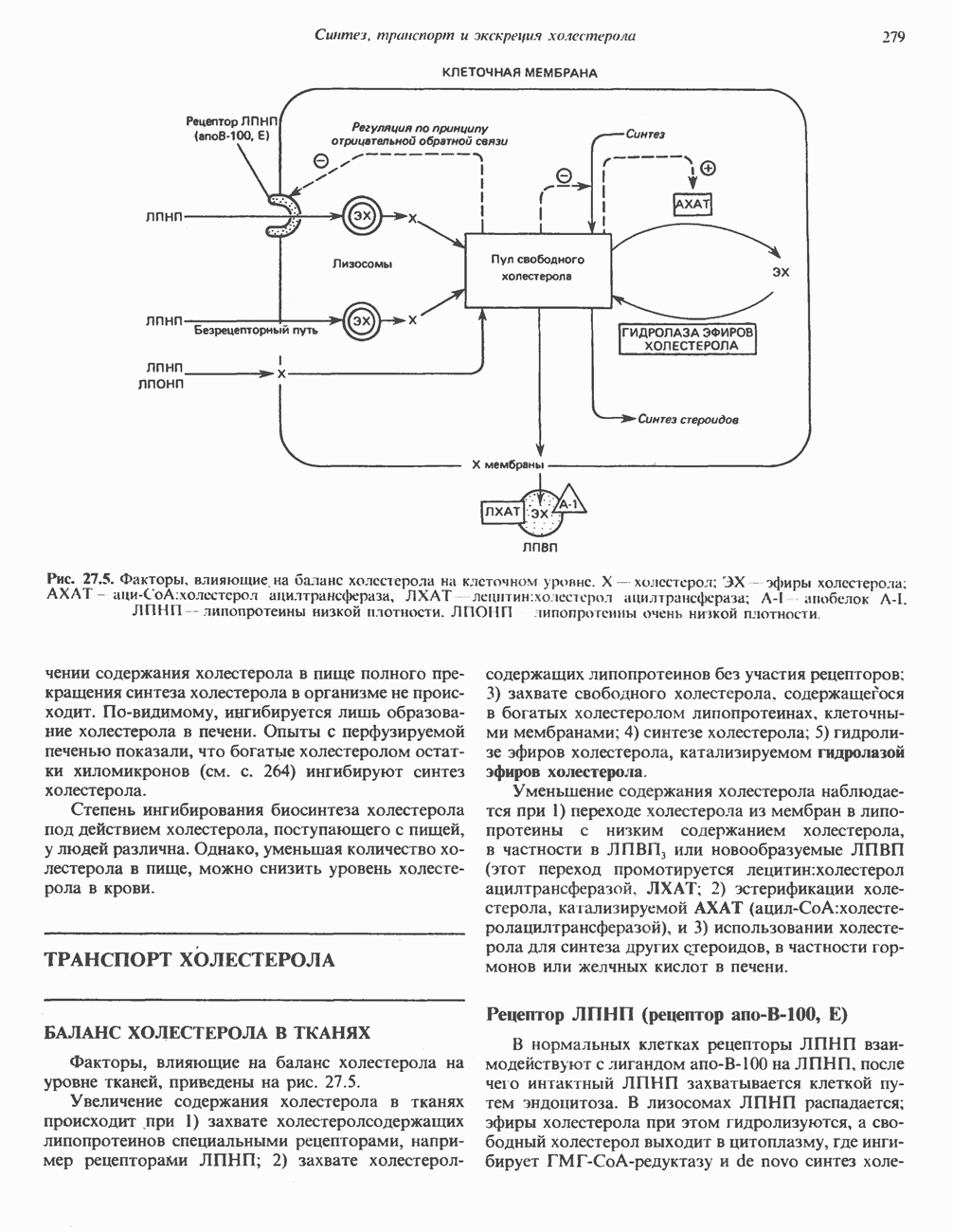
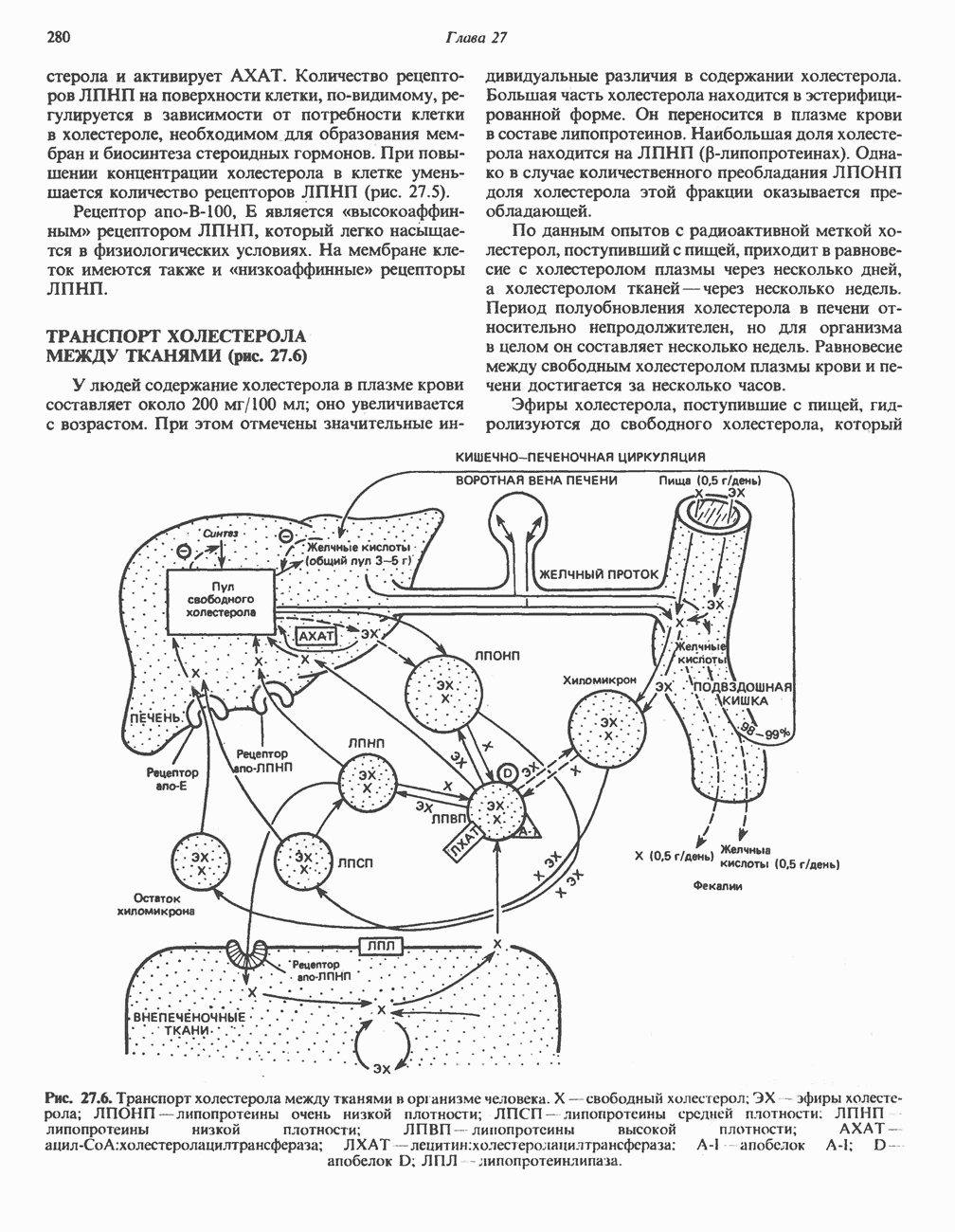

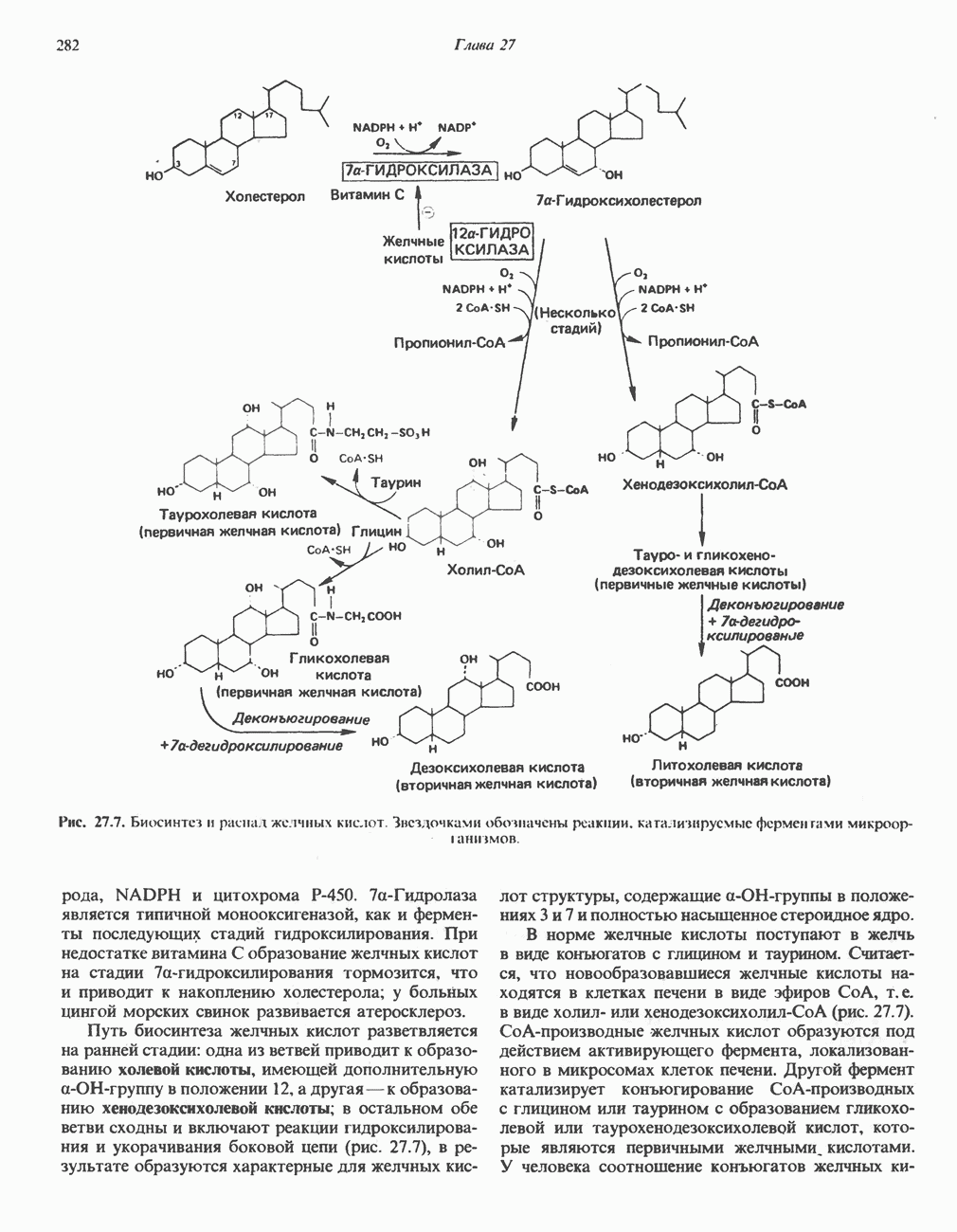















































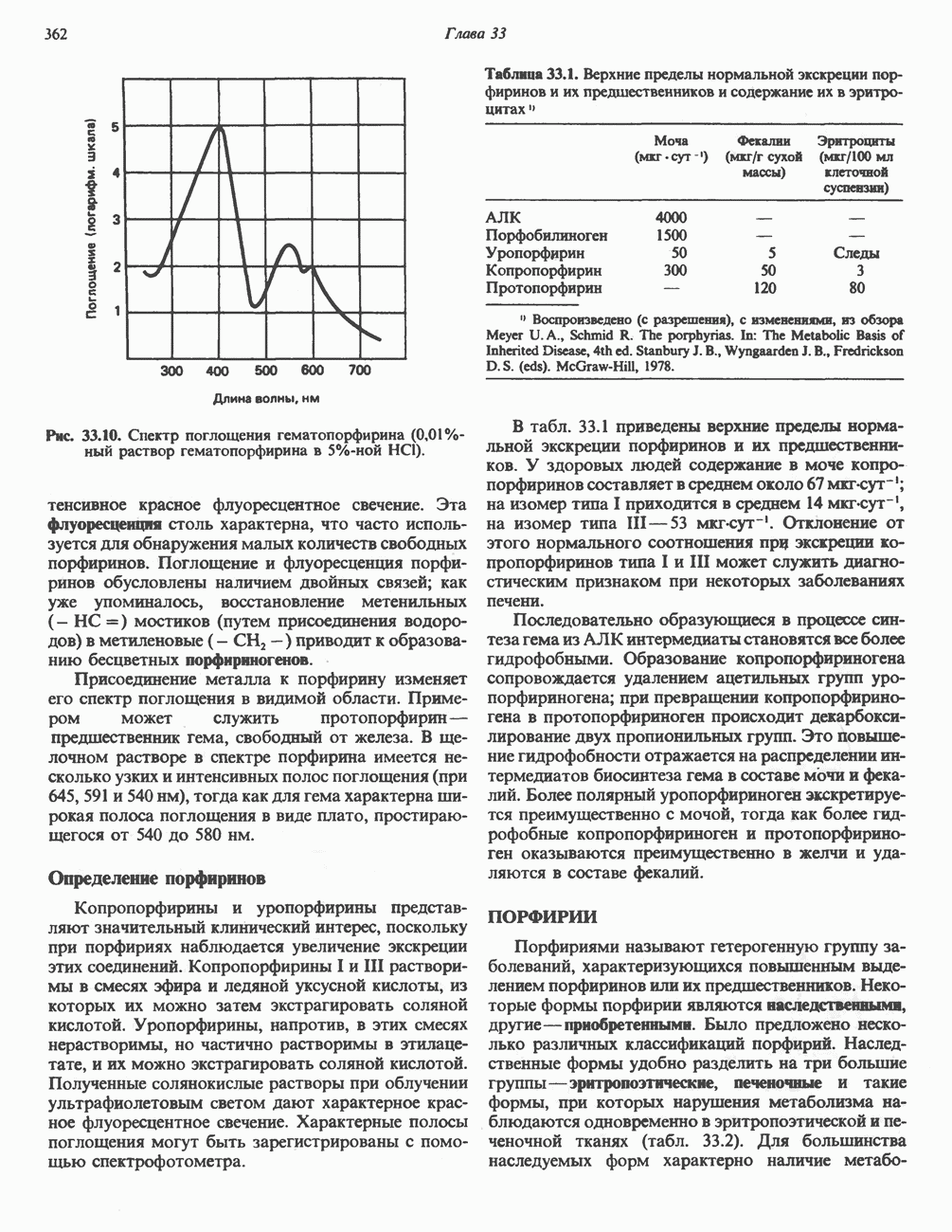
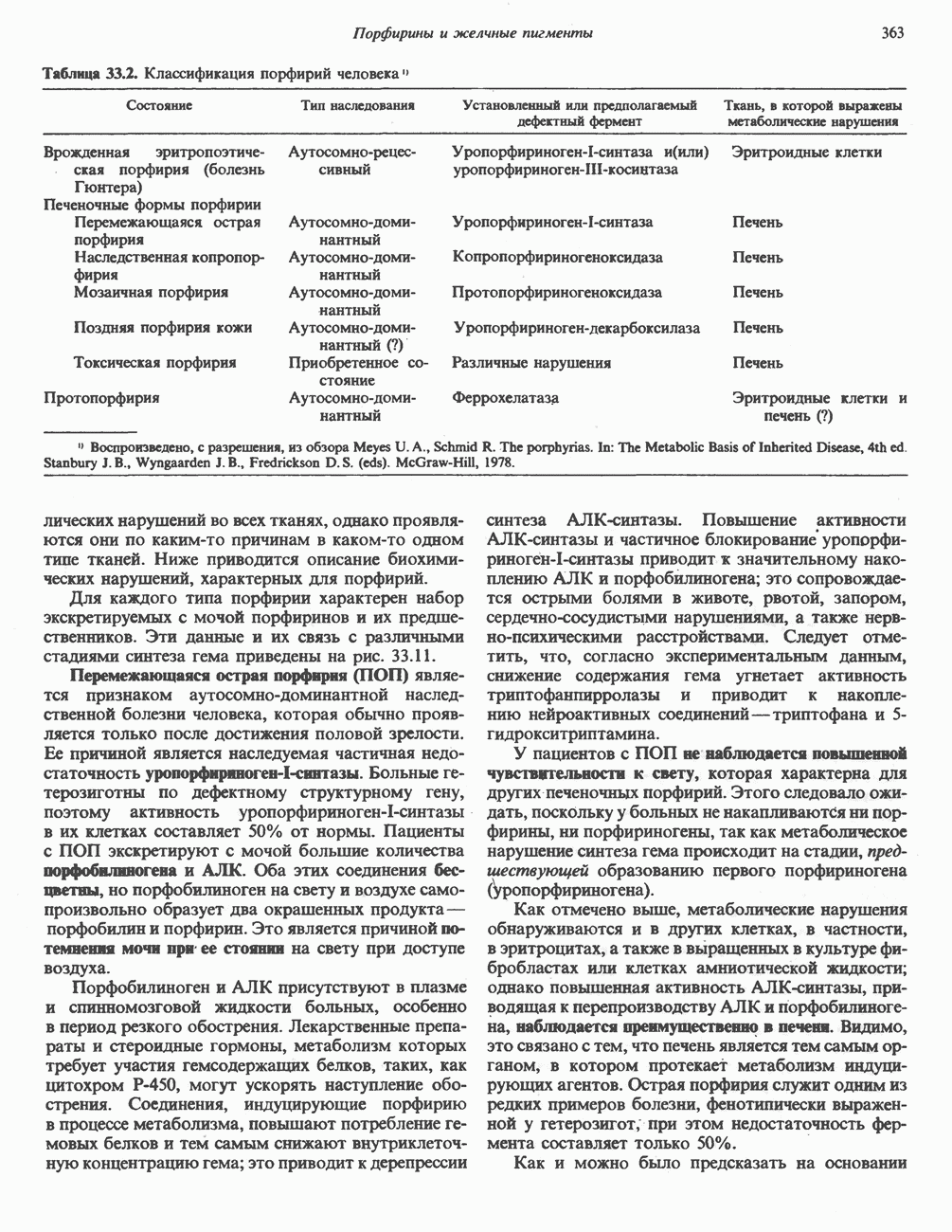


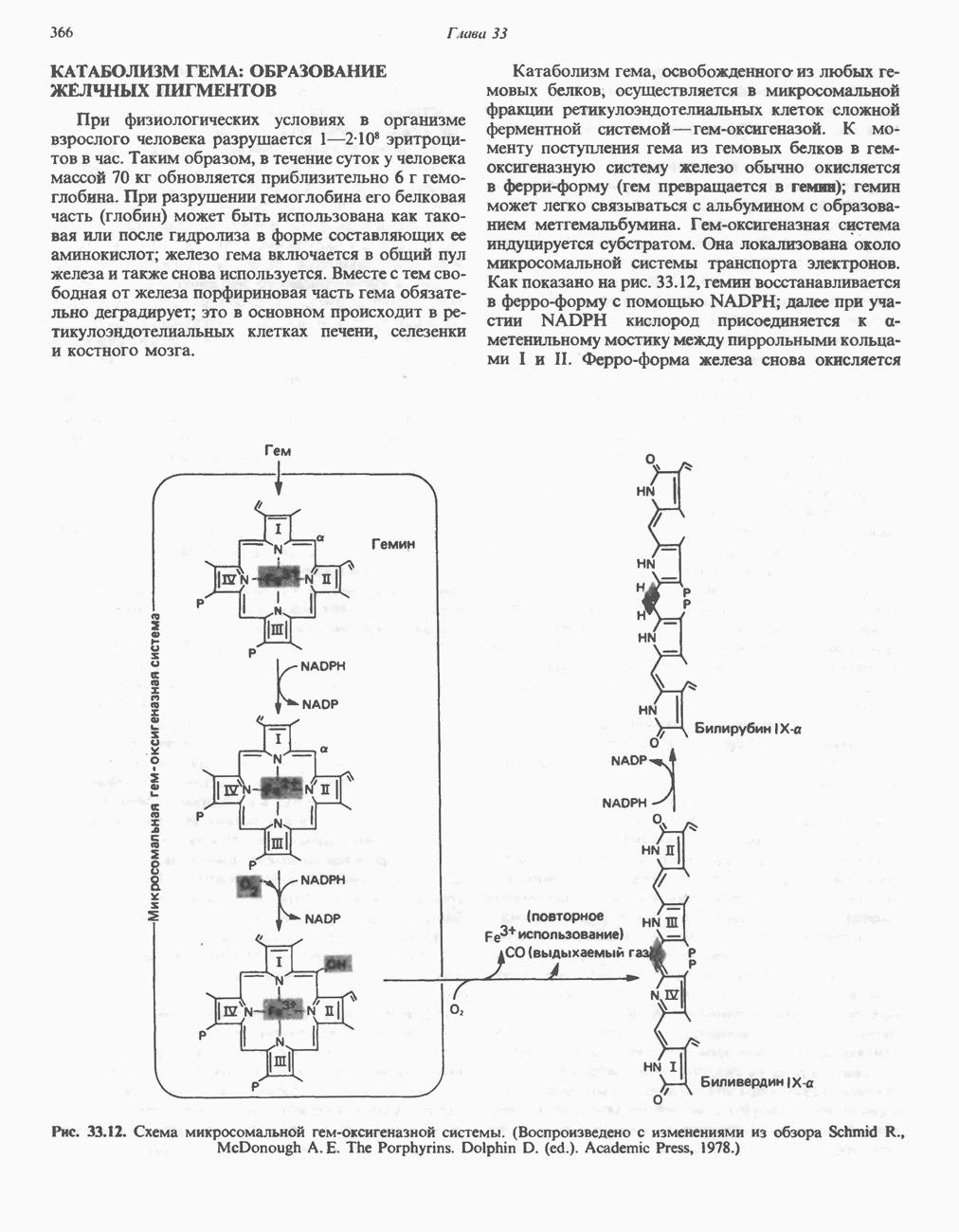




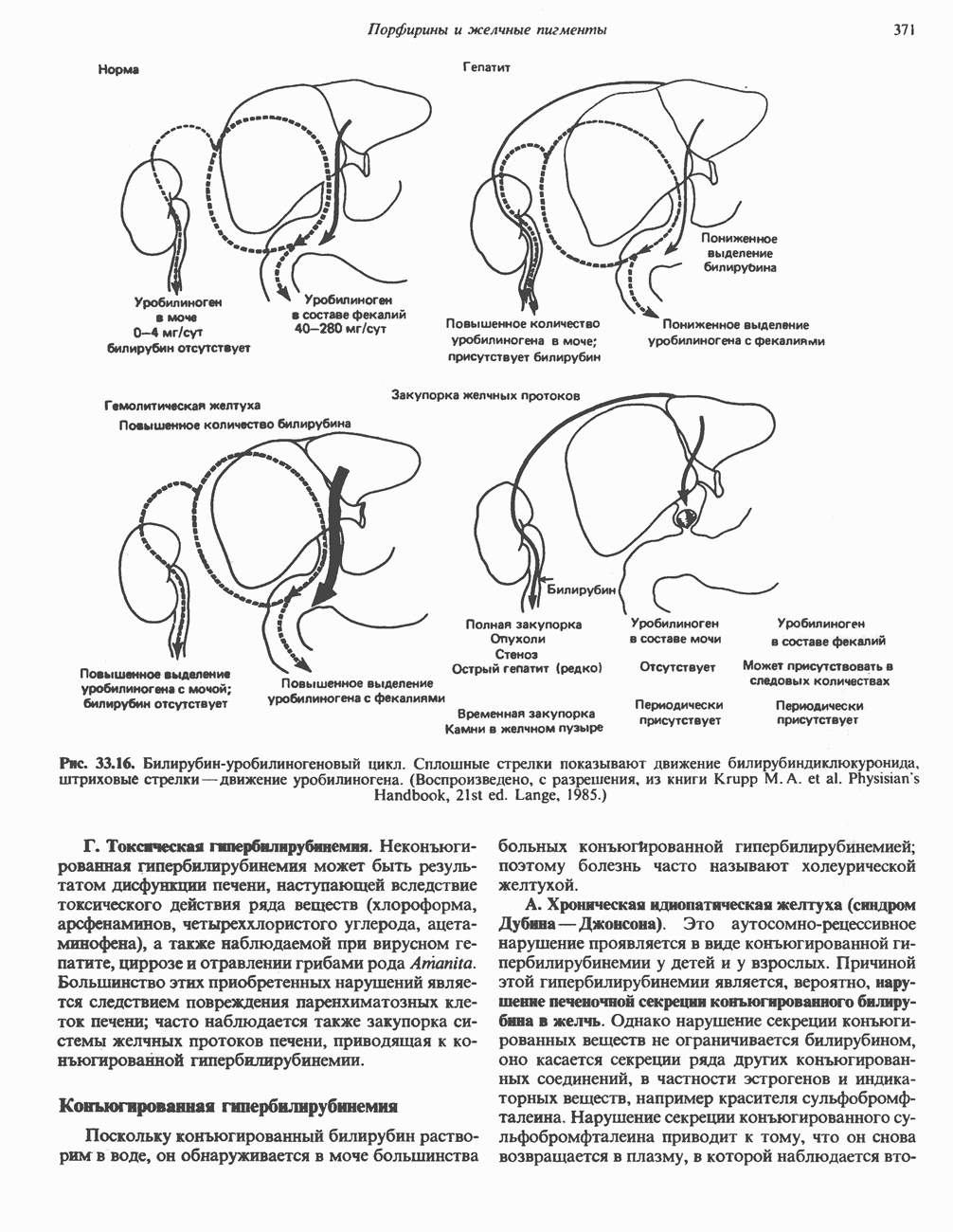



 . Два концевых атома углерода молекулы изолейцина образуют ацетил-СоА, а S-метильная группа метионина удаляется.
. Два концевых атома углерода молекулы изолейцина образуют ацетил-СоА, а S-метильная группа метионина удаляется.
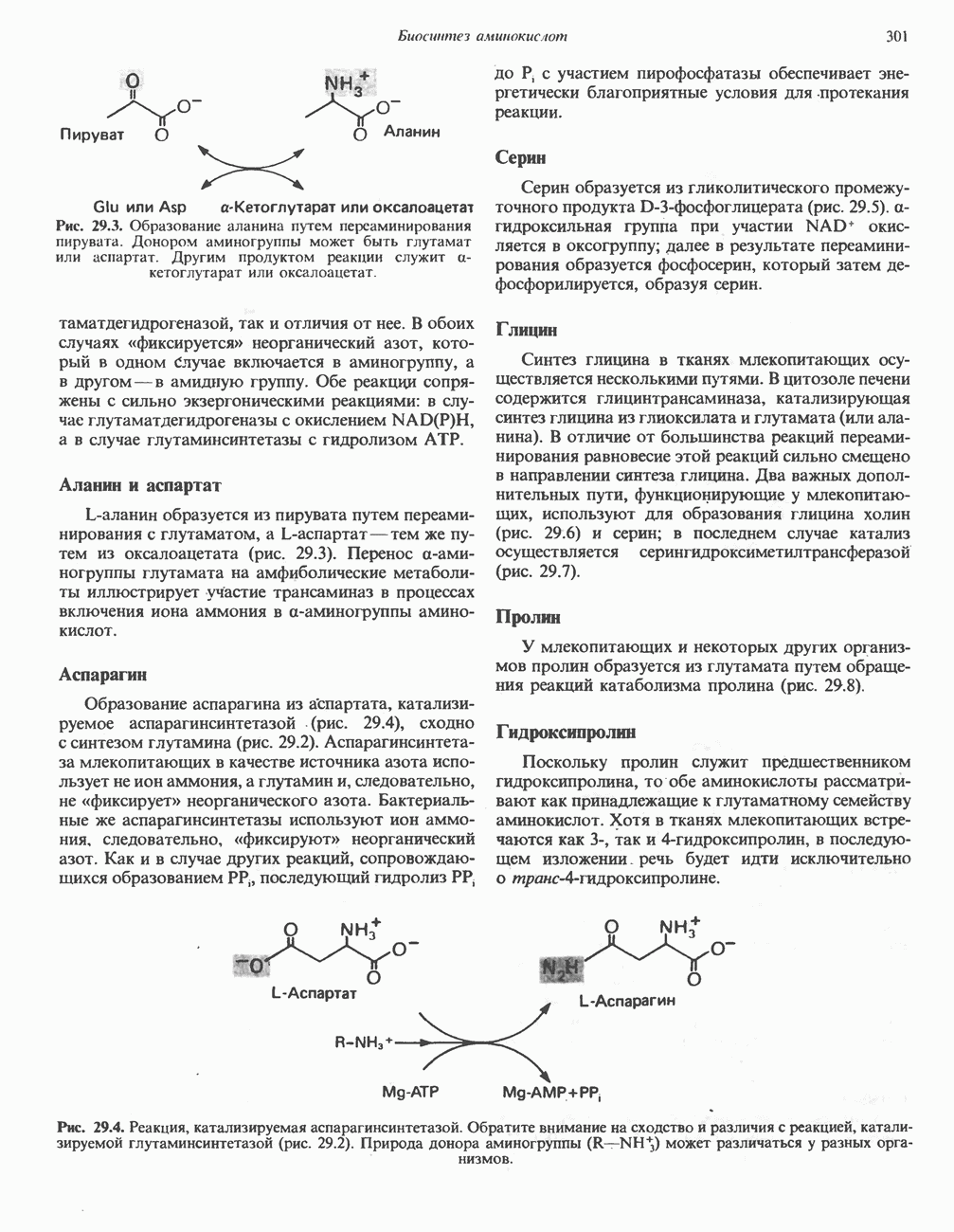
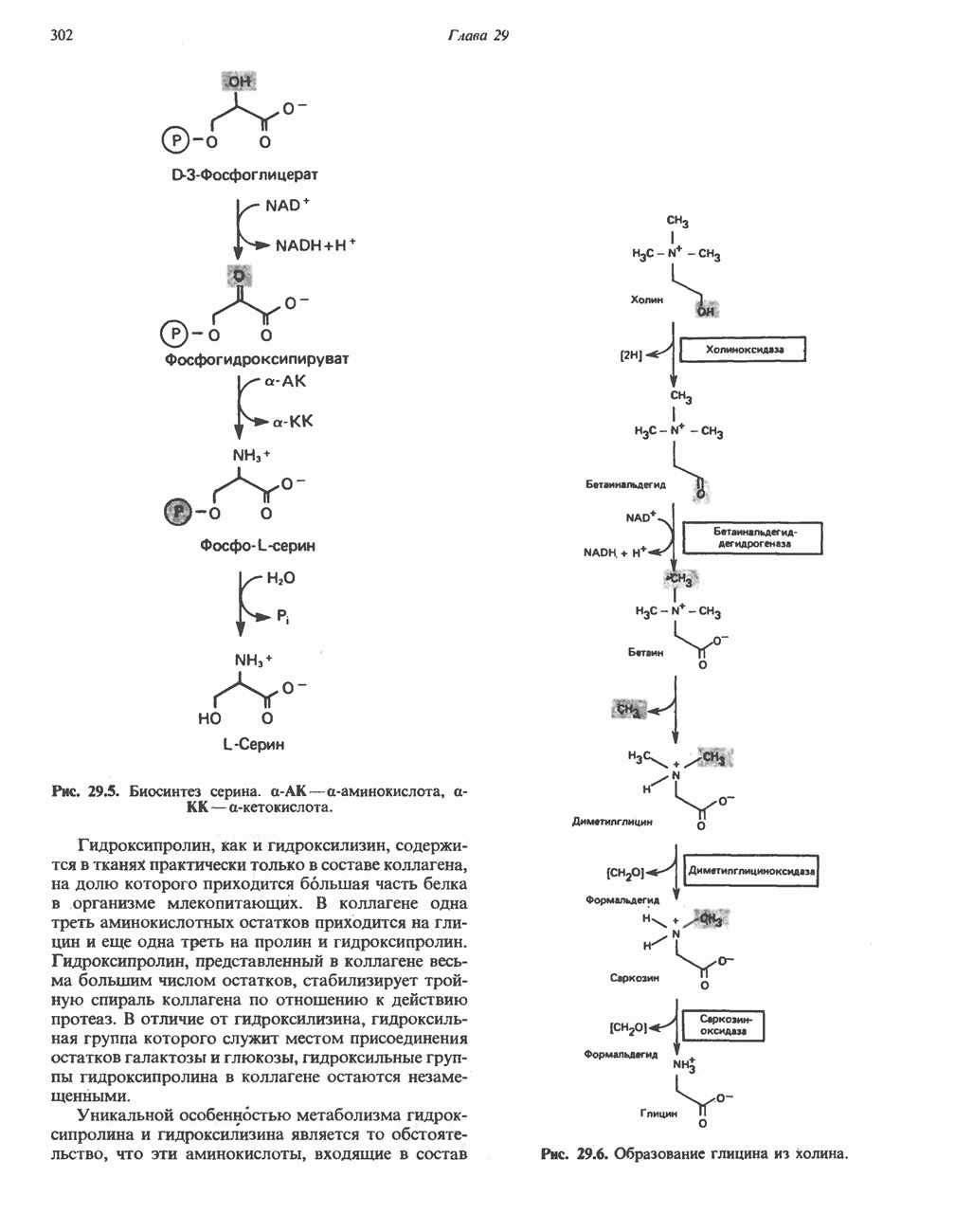
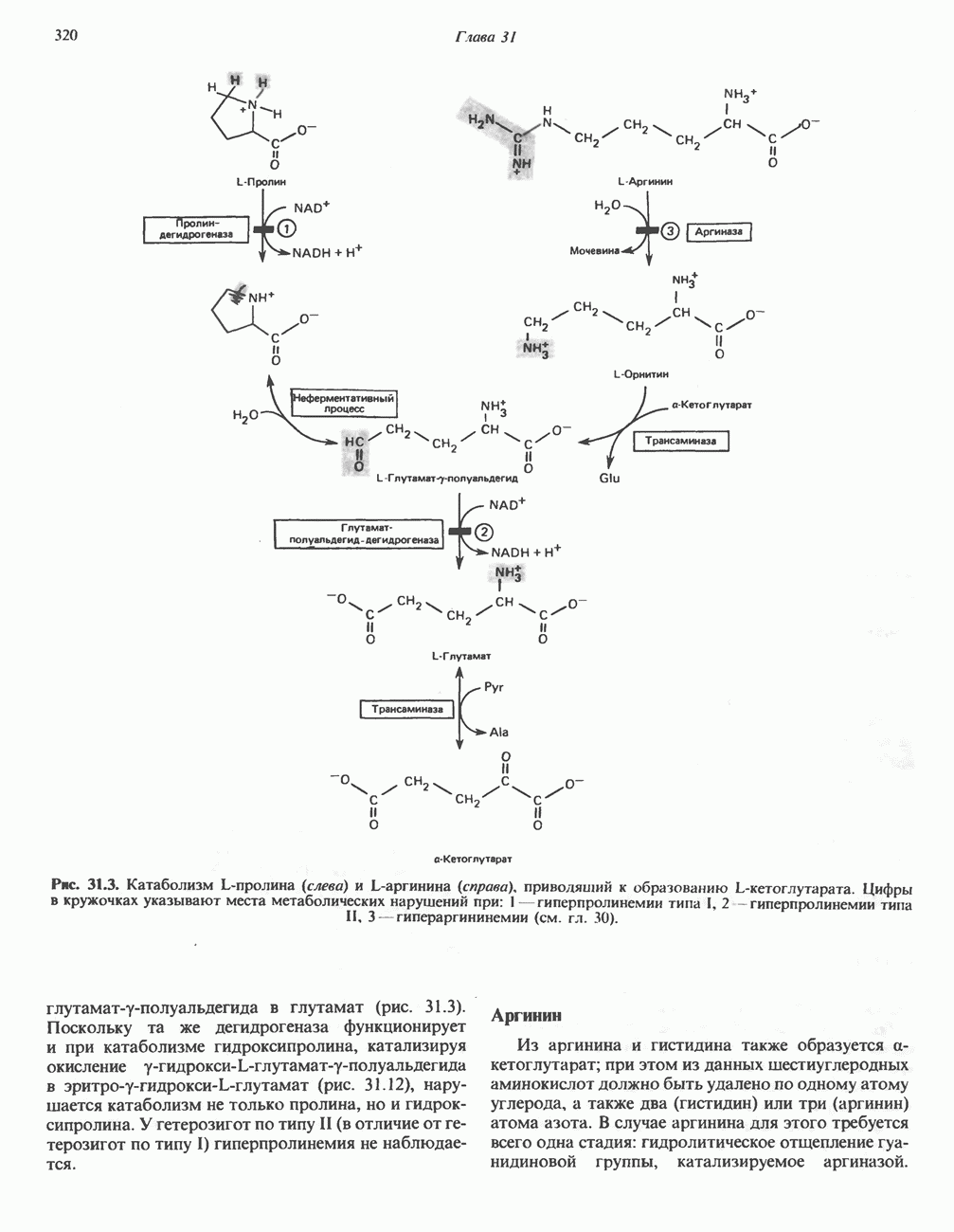
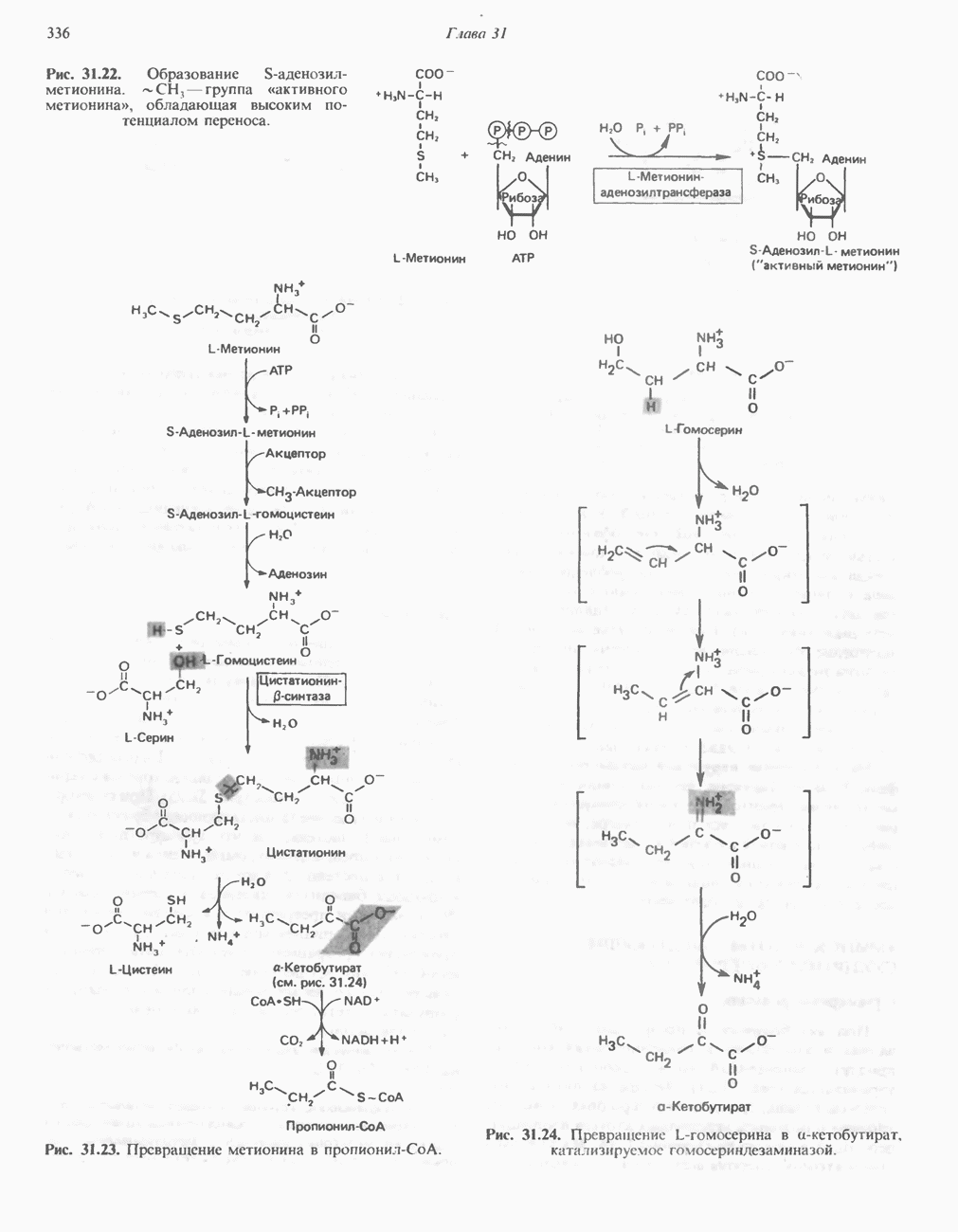
 При удалении метильной группы образуется S-аденозилгомоцистеин. В результате гидролиза S-С-связи образуются L-гомоцистеин и аденин. Гомоцистеин далее конденсируется с серином, образуя цистатионин (рис. 31.23). При гидролитическом расщеплении цистатионина образуются L-гомосерин и цистеин, так что суммарный процесс приводит к превращению гомоцистеина в гомосерин и серина в цистеин. Эти же две реакции участвуют в процессе биосинтеза цистеина из серина (см. гл. 29). Гомосерин превращается в а-кетобутират при участии гомосериндезаминазы (рис. 31.24). Затем происходит превращение а-кетобутирата в пропионил-СоА, оно осуществляется по обычному пути окислительного декарбоксилирования а-кетокислот (пирувата, а-кетоглутарата) с образованием ацил-СоА-производных.
При удалении метильной группы образуется S-аденозилгомоцистеин. В результате гидролиза S-С-связи образуются L-гомоцистеин и аденин. Гомоцистеин далее конденсируется с серином, образуя цистатионин (рис. 31.23). При гидролитическом расщеплении цистатионина образуются L-гомосерин и цистеин, так что суммарный процесс приводит к превращению гомоцистеина в гомосерин и серина в цистеин. Эти же две реакции участвуют в процессе биосинтеза цистеина из серина (см. гл. 29). Гомосерин превращается в а-кетобутират при участии гомосериндезаминазы (рис. 31.24). Затем происходит превращение а-кетобутирата в пропионил-СоА, оно осуществляется по обычному пути окислительного декарбоксилирования а-кетокислот (пирувата, а-кетоглутарата) с образованием ацил-СоА-производных.

 группа «активного метионина», обладающая высоким потенциалом переноса.
группа «активного метионина», обладающая высоким потенциалом переноса.
 -гомосерина в а-кетобутират, катализируемое гомосериндезаминазой.
-гомосерина в а-кетобутират, катализируемое гомосериндезаминазой.
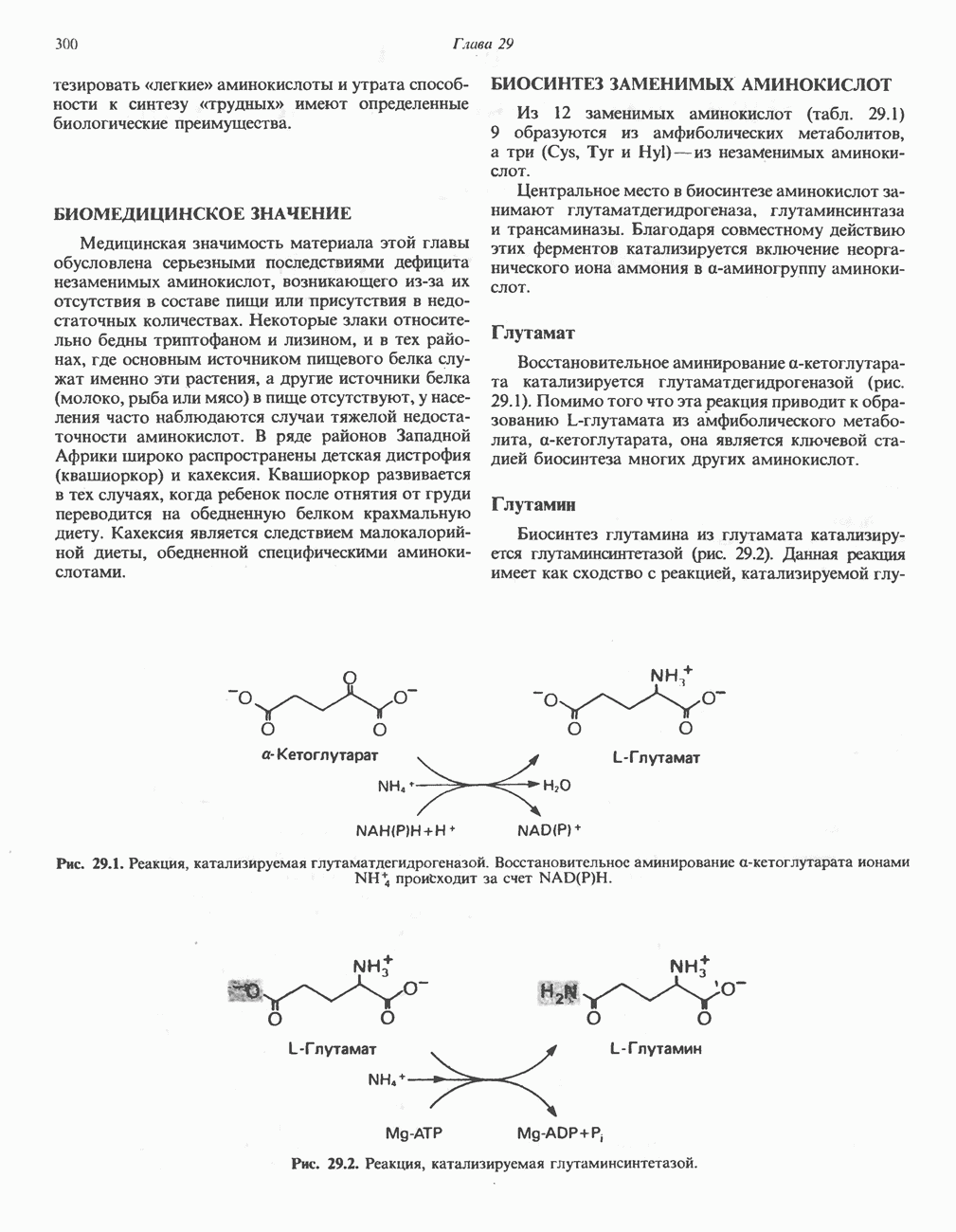
 -кетокислотами, если организму доступны адекватные источники азота.
-кетокислотами, если организму доступны адекватные источники азота.
 пируватдегидрогеназой и окислению а-кетоглутарата до
пируватдегидрогеназой и окислению а-кетоглутарата до  а-кетоглута-ратдегидрогеназой (см. гл. 18). У млекоптающих дегидрогеназа разветвленных a-кетокислот является митохондриальным мультиферментным комплексом, катализирующим окислительное декарбоксилирование
а-кетоглута-ратдегидрогеназой (см. гл. 18). У млекоптающих дегидрогеназа разветвленных a-кетокислот является митохондриальным мультиферментным комплексом, катализирующим окислительное декарбоксилирование  -кетоизокапроата (из лейцина),
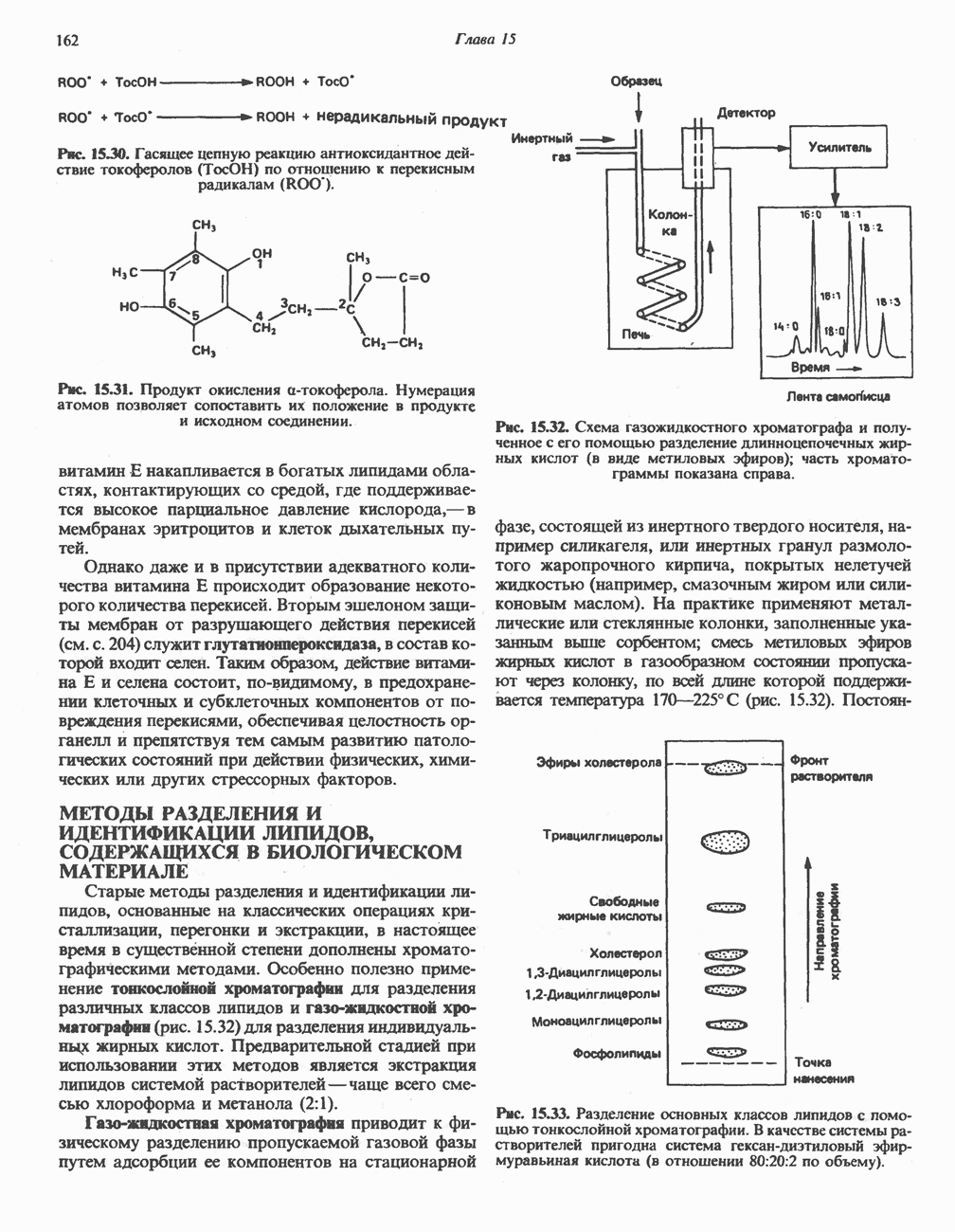
-кетоизокапроата (из лейцина),  -кето-Р-метилвалерата (из изолейцина) и а-кетоизовалерата (из валина).
-кето-Р-метилвалерата (из изолейцина) и а-кетоизовалерата (из валина).
 -кетокислотам), трансацилазной и дигидролипоилдегидрогеназной активностями. Как и в случае пируватдегидрогеназы, данный комплекс инактивируется при фосфорилировании за счет АТР в реакции, катализируемой протеинкиназой.
-кетокислотам), трансацилазной и дигидролипоилдегидрогеназной активностями. Как и в случае пируватдегидрогеназы, данный комплекс инактивируется при фосфорилировании за счет АТР в реакции, катализируемой протеинкиназой.  -Независимая фосфопротеинфосфатаза катализирует дефосфорилирование комплекса и, следовательно, реактивирует его. Таким образом, перевод фермента в фосфорилированное состояние может регулировать катаболизм аминокислот с разветвленной цепью. Протеинкиназа ингибируется ADP, а-кетокислотами с разветвленной цепью, гиполипидемическим агентом клофибратом и дихлорацетатом, а также тиоэфирами кофермента А (например, ацетоацетил-СоА).
-Независимая фосфопротеинфосфатаза катализирует дефосфорилирование комплекса и, следовательно, реактивирует его. Таким образом, перевод фермента в фосфорилированное состояние может регулировать катаболизм аминокислот с разветвленной цепью. Протеинкиназа ингибируется ADP, а-кетокислотами с разветвленной цепью, гиполипидемическим агентом клофибратом и дихлорацетатом, а также тиоэфирами кофермента А (например, ацетоацетил-СоА).

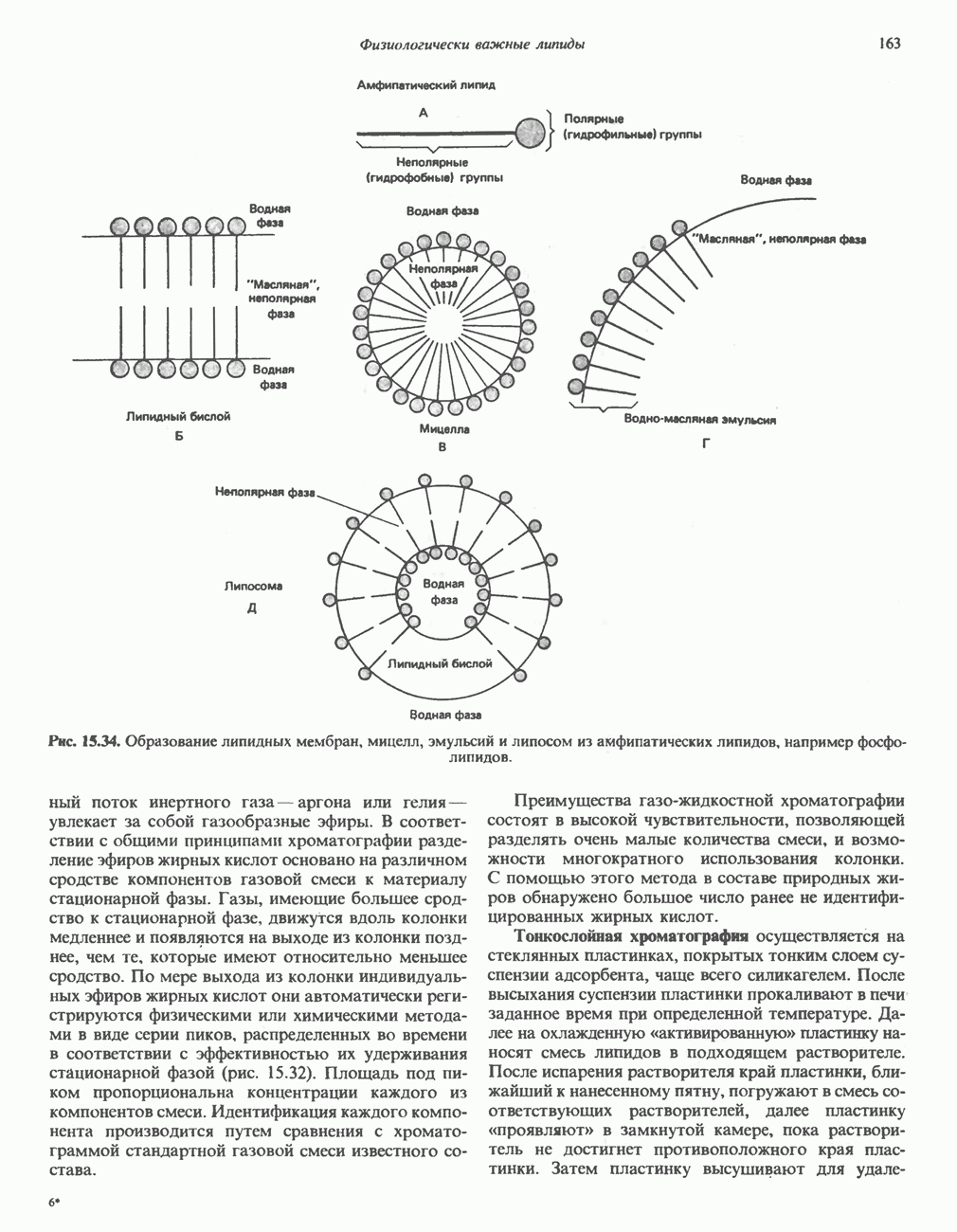
 -ненасьпценных тноэфнров ацил-СоА. Эта реакция (реакция 3) аналогична дегидрогенированию линейных ацил-СоА тиоэфиров в процессе катаболизма жирных кислот. Пока неизвестно, катализирует ли дегидрогенирование всех трех разветвленных ацил-СоА тиоэфиров одна и та же дегидрогеназа. Косвенные данные указывают на участие по крайней мере двух ферментов; они основаны на обследовании пациентов с нзовалернановой ацидемией, у которых после приема обогащенной белком пищи в крови накапливается изовалерат; при этом содержание других разветвленных a-кетокислот не повышается. Изовалерат образуется путем деацилирования
-ненасьпценных тноэфнров ацил-СоА. Эта реакция (реакция 3) аналогична дегидрогенированию линейных ацил-СоА тиоэфиров в процессе катаболизма жирных кислот. Пока неизвестно, катализирует ли дегидрогенирование всех трех разветвленных ацил-СоА тиоэфиров одна и та же дегидрогеназа. Косвенные данные указывают на участие по крайней мере двух ферментов; они основаны на обследовании пациентов с нзовалернановой ацидемией, у которых после приема обогащенной белком пищи в крови накапливается изовалерат; при этом содержание других разветвленных a-кетокислот не повышается. Изовалерат образуется путем деацилирования  являющегося субстратом упомянутой выше дегидрогеназы.
являющегося субстратом упомянутой выше дегидрогеназы.
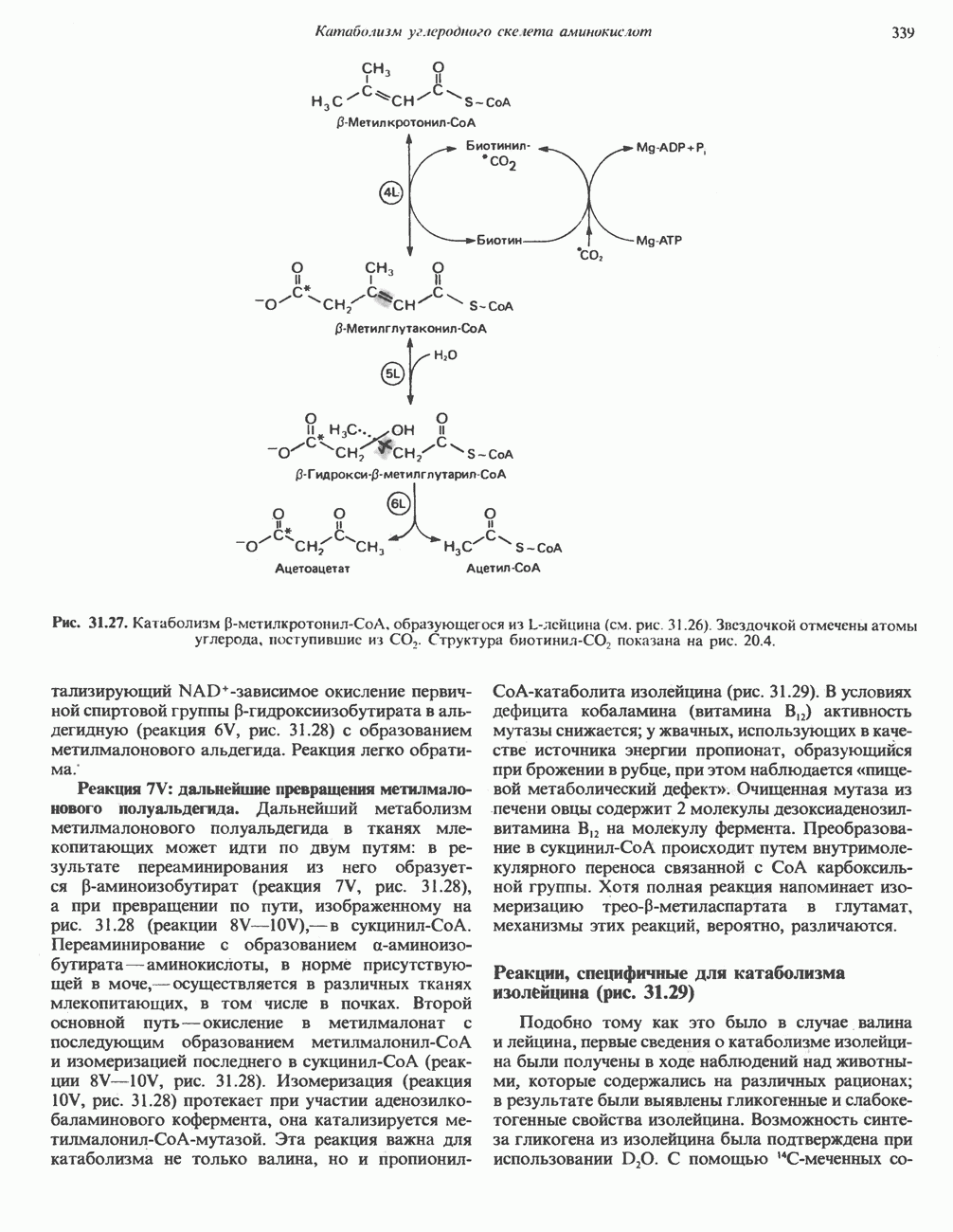
 карбоксилирование Р-метнлкротонил-СоА. Ключевым наблюдением, позволившим объяснить кетогенное действие лейцина, было обнаружении
карбоксилирование Р-метнлкротонил-СоА. Ключевым наблюдением, позволившим объяснить кетогенное действие лейцина, было обнаружении

 на моль изопропильных групп. Этой фиксации (реакция 4L, рис. 31.27) предшествует осуществляемое за счет энергии АТР присоединение
на моль изопропильных групп. Этой фиксации (реакция 4L, рис. 31.27) предшествует осуществляемое за счет энергии АТР присоединение  к связанному с ферментом биотину с образованием биотинил-
к связанному с ферментом биотину с образованием биотинил- . В результате фиксации
. В результате фиксации  образуется интермедиат
образуется интермедиат  -метилглутаконил-СоА.
-метилглутаконил-СоА.
 присоединение воды по двойной связи
присоединение воды по двойной связи  Продукт реакции
Продукт реакции  служит предшественником не только кетоновых тел (реакция 6L, рис. 31.27), но также и мевалоната, который далее может превращаться в холестерол и другие полиизопреноиды (см. гл. 28).
служит предшественником не только кетоновых тел (реакция 6L, рис. 31.27), но также и мевалоната, который далее может превращаться в холестерол и другие полиизопреноиды (см. гл. 28).
 расщепление
расщепление  Реакция, в ходе которой из
Реакция, в ходе которой из  образуются
образуются  и ацетоацетат, протекает у млекопитающих в митохондриях клеток печени, почек и сердца. Этим процессом объясняется сильное кетогенное действие лейцина, поскольку на 1 моль лейцина наряду с 1 молем ацетоацетата образуется опосредованно еще 1/2 моля кетоновых тел из ацетил-СоА (см. гл. 28).
и ацетоацетат, протекает у млекопитающих в митохондриях клеток печени, почек и сердца. Этим процессом объясняется сильное кетогенное действие лейцина, поскольку на 1 моль лейцина наряду с 1 молем ацетоацетата образуется опосредованно еще 1/2 моля кетоновых тел из ацетил-СоА (см. гл. 28).
 содержащие в оксиацильной части 4—9 атомов углерода.
содержащие в оксиацильной части 4—9 атомов углерода.
 Поскольку
Поскольку  -тиоэфир не является субстратом последующей реакции (реакция 6V, рис. 31.28), он должен быть сначала деацилирован в Р-гидроксиизобутират (реакция 5V, рис. 31.28). Эта реакция катализируется деацилазой, присутствующей во многих животных тканях; деацилаза может действовать также еще на один субстрат -
-тиоэфир не является субстратом последующей реакции (реакция 6V, рис. 31.28), он должен быть сначала деацилирован в Р-гидроксиизобутират (реакция 5V, рис. 31.28). Эта реакция катализируется деацилазой, присутствующей во многих животных тканях; деацилаза может действовать также еще на один субстрат -  -гидроксипропионил-СоА.
-гидроксипропионил-СоА.

 Структура биотинил-СO2 показана на рис. 20.4.
Структура биотинил-СO2 показана на рис. 20.4.
 ) активность мутазы снижается; у жвачных, использующих в качестве источника энергии пропионат, образующийся при брожении в рубце, при этом наблюдается «пищевой метаболический дефект». Очищенная мутаза из печени овцы содержит 2 молекулы дезоксиаденозип-витамина
) активность мутазы снижается; у жвачных, использующих в качестве источника энергии пропионат, образующийся при брожении в рубце, при этом наблюдается «пищевой метаболический дефект». Очищенная мутаза из печени овцы содержит 2 молекулы дезоксиаденозип-витамина  на молекулу фермента. Преобразование в сукцинил-СоА происходит путем внутримолекулярного переноса связанной с СоА карбоксильной группы. Хотя полная реакция напоминает изомеризацию трео-р-метиласпартата в глутамат, механизмы этих реакций, вероятно, различаются.
на молекулу фермента. Преобразование в сукцинил-СоА происходит путем внутримолекулярного переноса связанной с СоА карбоксильной группы. Хотя полная реакция напоминает изомеризацию трео-р-метиласпартата в глутамат, механизмы этих реакций, вероятно, различаются.
 . С помощью
. С помощью  -меченных
-меченных

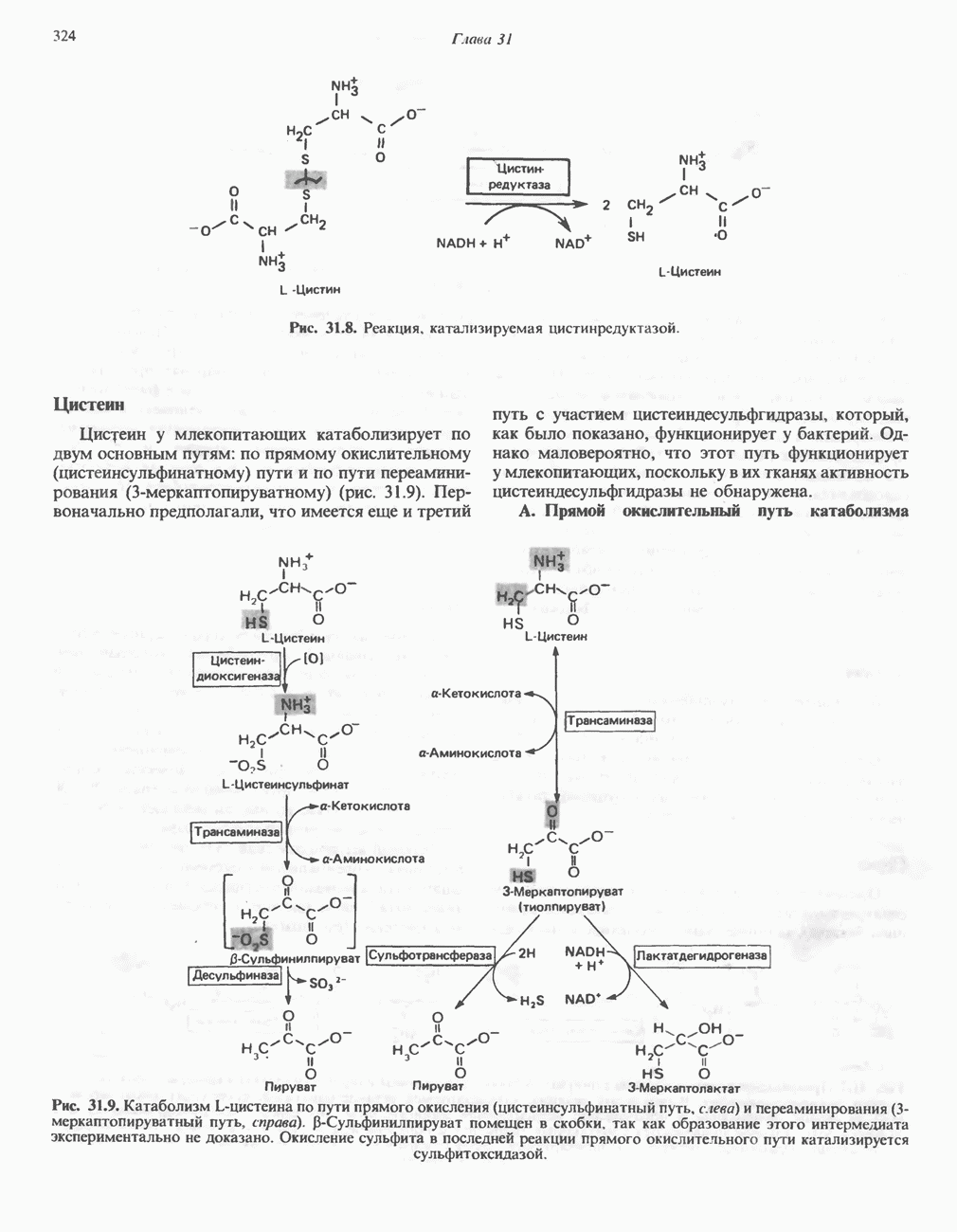
 -кетотиолазой подобно тиолизу ацетоацетил-СоА с образованием двух молекул ацетил-СоА. Природа образующихся продуктов, ацетил-СоА (кетогенный) и пропионил-СоА (гликогенный), позволяет понять кетоген-ные и гликогенные свойства изолейцина.
-кетотиолазой подобно тиолизу ацетоацетил-СоА с образованием двух молекул ацетил-СоА. Природа образующихся продуктов, ацетил-СоА (кетогенный) и пропионил-СоА (гликогенный), позволяет понять кетоген-ные и гликогенные свойства изолейцина.

 -кетокислот в ацил-СоА-тиоэфиры с выделением
-кетокислот в ацил-СоА-тиоэфиры с выделением  (реакция 2, рис. 31.26). Это было установлено путем ферментного анализа лейкоцитов и клеток культуры фибробластов кожи больных детей. Механизм токсического действия накапливающихся соединений неизвестен.
(реакция 2, рис. 31.26). Это было установлено путем ферментного анализа лейкоцитов и клеток культуры фибробластов кожи больных детей. Механизм токсического действия накапливающихся соединений неизвестен.
 -кетокислот. Активность декарбоксилазы в лейкоцитах и фибробластах значительно ниже нормы, но все же существенно выше, чем в классических случаях болезни «кленового сиропа». Поскольку в этом случае больные обладают хотя и сниженной, но все же определенной способностью к катаболизму лейцина, изолейцина и валина, типичные симптомы болезни «кленового сиропа» проявляются позднее и наблюдаются лишь эпизодически. Прогноз при назначении соответствующей диеты более благоприятный.
-кетокислот. Активность декарбоксилазы в лейкоцитах и фибробластах значительно ниже нормы, но все же существенно выше, чем в классических случаях болезни «кленового сиропа». Поскольку в этом случае больные обладают хотя и сниженной, но все же определенной способностью к катаболизму лейцина, изолейцина и валина, типичные симптомы болезни «кленового сиропа» проявляются позднее и наблюдаются лишь эпизодически. Прогноз при назначении соответствующей диеты более благоприятный.
 гидролизуется, образующийся изовалерат выделяется в составе мочи и пота.
гидролизуется, образующийся изовалерат выделяется в составе мочи и пота.
 в амфиболические интермедиаты включает биотинзависимое карбоксилирование с образованием
в амфиболические интермедиаты включает биотинзависимое карбоксилирование с образованием  последний может образоваться и непосредственно из валина (при этом не происходит предварительного образования
последний может образоваться и непосредственно из валина (при этом не происходит предварительного образования  ) (рис. 31.21 и 31.28, реакция
) (рис. 31.21 и 31.28, реакция  результате изомеризации, коферментом которой является производное витамина
результате изомеризации, коферментом которой является производное витамина  превращается в
превращается в  цикла лимонной кислоты, который окисляется до
цикла лимонной кислоты, который окисляется до  и воды.
и воды.
 -дезоксиаденозилкобаламина в качестве кофактора
-дезоксиаденозилкобаламина в качестве кофактора
 .
.
 , а другая — нет. В последнем случае улучшение наступает после введения больших (фармакологических) доз витамина
, а другая — нет. В последнем случае улучшение наступает после введения больших (фармакологических) доз витамина  (1 г в сутки). Фибробласты больных, культивируемые в среде, содержащей витамин
(1 г в сутки). Фибробласты больных, культивируемые в среде, содержащей витамин  (концентрация 25 пгмл-1), слабо окисляют [14С]-пропионат. Содержание 5-дезоксиаденозилкобаламина в клетках культуры составляло только около 10% от его содержания в контрольных клетках. После повышения концентрации витамина
(концентрация 25 пгмл-1), слабо окисляют [14С]-пропионат. Содержание 5-дезоксиаденозилкобаламина в клетках культуры составляло только около 10% от его содержания в контрольных клетках. После повышения концентрации витамина  в среде в 10000 раз скорость окисления пропионата и внутриклеточная концентрация 5-дезоксиаденозилкобаламина приблизились к норме. В то же время не наблюдалось отклонений в связывании кофермента с апоферментом мутазы. Таким образом, причиной второй формы метилмалоновой ацидурии, по-видимому, является неспособность к образованию 5-дезоксиаденозилкобаламина при нормальных концентрациях витамина.
в среде в 10000 раз скорость окисления пропионата и внутриклеточная концентрация 5-дезоксиаденозилкобаламина приблизились к норме. В то же время не наблюдалось отклонений в связывании кофермента с апоферментом мутазы. Таким образом, причиной второй формы метилмалоновой ацидурии, по-видимому, является неспособность к образованию 5-дезоксиаденозилкобаламина при нормальных концентрациях витамина.