Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
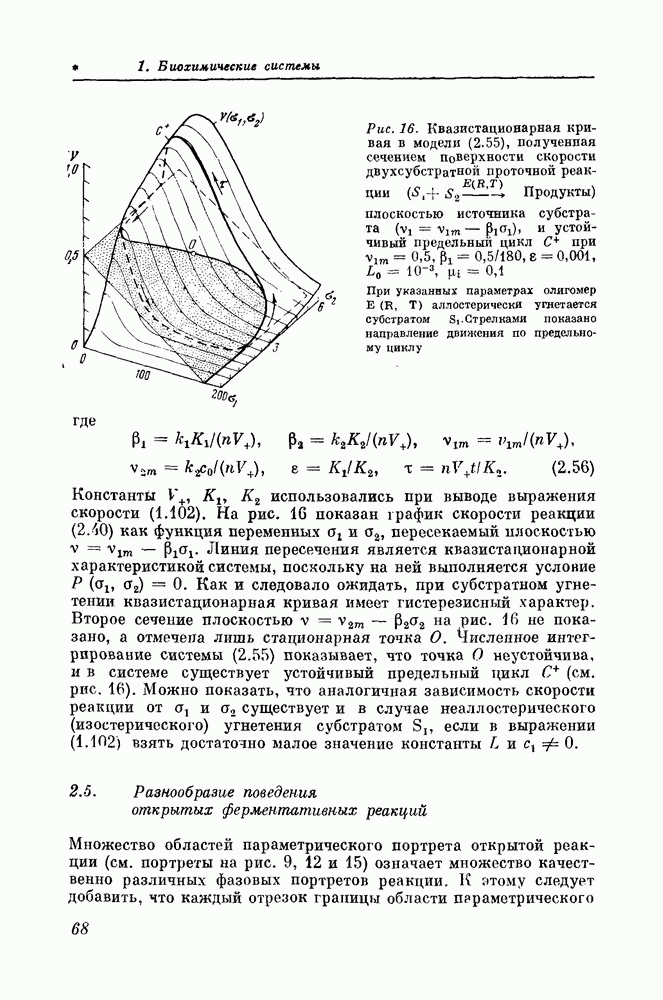
2.6. Эквивалентные математические и кинетические моделиМатематически сходство динамического поведения различных систем, имеющих гистерезис квазистационарной характеристики, можно интерпретировать как простое следствие эквивалентности их математических моделей. Например, если для описания кинетики действия фермента
или реакцию
использовать уравнение вида (2.42), то математические модели этих реакций будут иметь тот же вид, что математическая модель (2.36) реакции (2.32). Если же фермент имеет олигомерную структуру и кинетика его действия описывается более сложным уравнением (2.54) или еще более сложным уравнением (1.102), то математические модели этих реакций, хотя и отличаются от модели (2.36) реакции (2.32) видом своих правых частей и количественными характеристиками взаимоотношений между переменными, все же остаются топологически эквивалентными более простой модели (2.36). Под топологической эквивалентностью здесь подразумевается одинаковое строение параметрических и, следовательно, фазовых портретов моделей. Очевидно, что усилия, затрачиваемые на анализ новой, еще не изученной биохимической системы, можно резко сократить, если доказать, что ее математическая модель эквивалентна более детально изученной модели. В таком случае все свойства более полно изученной модели автоматически распространяются на новую модель. Таким образом, наиболее детально изученная модель некоторого множества эквивалентных моделей может служить эталоном, с которым сравниваются другие менее изученные модели, принадлежащие этому множеству. Приведение данной модели к эталонной может быть осуществлено с помощью соответствующей замены переменных [72]. Так, например, математическую модель открытой реакции с продуктной активацией
имеющей в безразмерных переменных вид [80—84]
где
с помощью простой замены переменных
можно привести к эквивалентной форме
где
Новую переменную
в которой
Это уравнение описывает гиперболическую зависимость скорости от концентрации Из-за малости Простейшие из эквивалентных моделей целесообразно использовать для замещения более сложных при моделировании полиферментных систем. При этом подгонкой параметров легко добиться хорошего количественного совпадения между моделью-объектом и моделью-заместителем. В работе [85], например, показано, что феноменологическая модель (2.60) с высокой точностью замещает существенно более сложную модель реакции (2.59), катализируемую олигомером [86]. В дальнейшем кинетические модели, описываемые топологически эквивалентными математическими моделями, будем называть эквивалентными. Эквивалентными будем называть и регуляторные связи, если они принадлежат таким кинетическим моделям. Так, например, только что было показано, что кинетические модели (2.59) и (2.65) описываются топологически эквивалентными моделями. Следовательно, эти кинетические модели и их регуляторные связи — субстратное угнетение в модели (2.65) и продуктная активация в модели (2.59) — также эквивалентны.
|
 |
Оглавление
|








































































































































































































































































































 катализирующего реакцию
катализирующего реакцию


 в реакциях (2.57) и (2.58)
в реакциях (2.57) и (2.58)
 порядка
порядка






 можно рассматривать как концентрацию субстрата
можно рассматривать как концентрацию субстрата  некоторой эквивалентной реакции
некоторой эквивалентной реакции

 ингибитор фермента
ингибитор фермента  связанный законом сохранения с продуктом
связанный законом сохранения с продуктом  полная концентрация
полная концентрация  Правда, выражение (2.64) для безразмерной скорости реакции (2.65) является «нехимическим» — оно предсказывает
Правда, выражение (2.64) для безразмерной скорости реакции (2.65) является «нехимическим» — оно предсказывает  при
при  Однако этот недостаток легко устраняется малым возмущением уравнения (2.64) с помощью малого параметра
Однако этот недостаток легко устраняется малым возмущением уравнения (2.64) с помощью малого параметра 

 и кривую с максимумом
и кривую с максимумом  характерную для субстратного угнетения.
характерную для субстратного угнетения.
 модели (2.63), (2.64) и (2.63), (2.66) имеют почти совпадающие фазовые портреты, различия существуют лишь в малой
модели (2.63), (2.64) и (2.63), (2.66) имеют почти совпадающие фазовые портреты, различия существуют лишь в малой  -полосе, примыкающей к оси
-полосе, примыкающей к оси  Поэтому параметрические портреты этих моделей, построенные в плоскости параметров
Поэтому параметрические портреты этих моделей, построенные в плоскости параметров  не только топологически эквивалентны, но даже количественно почти совпадают. А так как модель (2.63) с уравнением для скорости реакции (2.66) с точностью до обозначений совпадает с эталонной моделью (2.36), то ее можно считать топологически эквивалентной эталонной модели.
не только топологически эквивалентны, но даже количественно почти совпадают. А так как модель (2.63) с уравнением для скорости реакции (2.66) с точностью до обозначений совпадает с эталонной моделью (2.36), то ее можно считать топологически эквивалентной эталонной модели.