Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
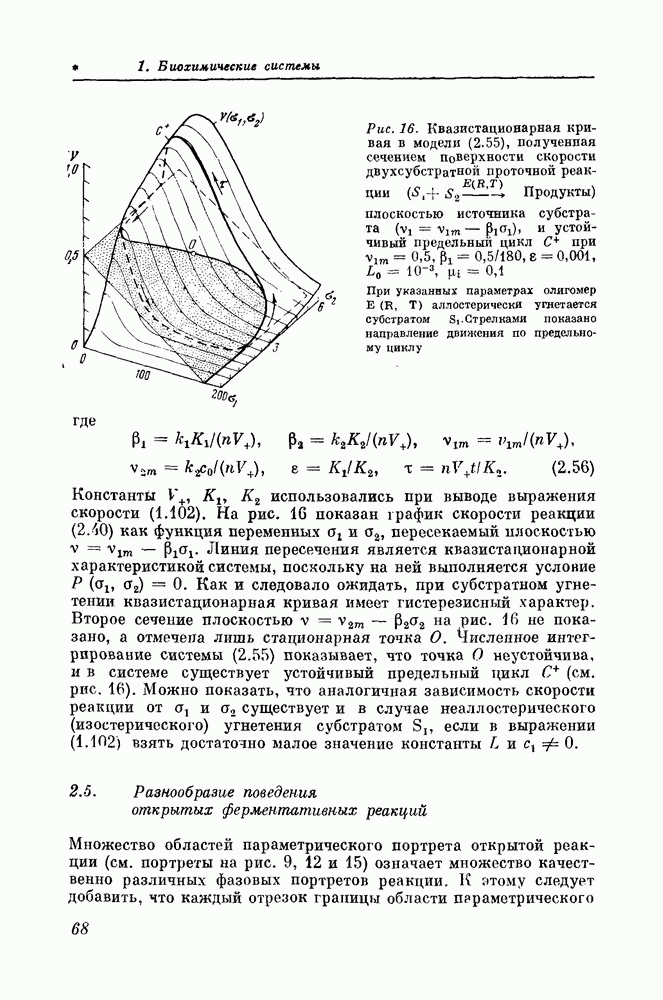
ВВЕДЕНИЕСовременные представления о процессах, протекающих в живой клетке, позволяют сравнивать ее с хорошо организованным и ритмично работающим заводом, поставляющим другим клеткам «изделия» своей специализации и постоянно обновляющим собственное, в основном белковое, оборудование. Жизнедеятельность клетки зависит как от регулярного поступления извне сырья, а с ним и запасенной в нем энергии, так и от безотказной работы всех звеньев химического производства. В настоящее время известны тысячи химических превращений, протекающих в клетке, причем многие из них несовместимы друг с другом. Такие несовместимые биохимические превращения должны быть изолированы соответствующей организацией их во времени и пространстве. Пространственная организация ферментов в клетке осуществляется многими способами: агрегацией функционально связанных ферментов в крупные макромолекулярные комплексы, присоединением ферментов в определенном порядке к внутриклеточным мембранам, разнесением несовместимых ферментов и полиферментных систем в различные отсеки (компартменты), разделенные мембранами, обладающими селективной проницаемостью, и т. д. В частности, в клетке многие чисто анаболические (биосинтетические) или чисто катаболические (разлагающие внутриклеточные вещества) процессы разнесены в различные клеточные органеллы. Однако в клетке имеются биохимические процессы, протекающие в зависимости от условий внутри или внеклеточной среды либо в анаболическом, либо в катаболическом направлении. Такие процессы, называемые амфиболическими, в принципе не могут быть эффективно организованы в пространстве, так как ферменты, катализирующие противоположно направленные процессы (например, ферменты, синтезирующие и расщепляющие гликоген), должны находиться в одном и том же компартменте. Для таких биохимических процессов единственно возможной формой или, по крайней мере, основной формой организации является временная организация, т. е. определенный (как правило, периодический) порядок работы несовместимых биохимических процессов во времени. Пытаясь ответить на вопрос, какова организация работы клетки, целесообразно поставить ряд более простых вопросов. 1. Каким образом регулируются сложные открытые полиферментные системы в бесструктурной гомогенной среде? 2. Какими свойствами обладают клеточные мембраны, разделяющие полиферментные системы и клетки друг от друга? 3. К каким следствиям приводит пространственная распределенность биохимических, в частности возбудимых мембранных систем? 4. Каким образом можно получить трехмерные реконструкции внутриклеточных структур по данным оптической или электронной микроскопии, дающим практически лишь двумерные сечения или проекции? Теоретическое решение этих вопросов и составляет основную цель нашей книги. Материал, посвященный математическому моделированию биохимических систем, содержится в части 1, состоящей из трех глав. Глава 1 посвящена выводу уравнений, описывающих зависимость квазистационарной (или так называемой начальной) скорости ферментативной реакции от концентрации реактантов (субстратов, продуктов, модификаторов и т. п.). Основное внимание здесь уделено выводу и анализу уравнений, описывающих регуляторные ферменты. Такие ферменты имеют олигомерную (субъединичную) структуру, включающую несколько активных и регуляторных (аллостерических) центров. До сих пор теоретическое рассмотрение кинетики действия таких ферментов не выходило за рамки простых моделей (модель Моно — Уаймена — Шанжё, модель Кошлэнда, модели Фридена — Курганова), описывающих моносубстратную необратимую реакцию. Между тем подавляющее большинство биохимических реакций, катализируемых регуляторными олигомерными ферментами, представляют собой многосубстратные, часто легкообратимые реакции. Поэтому в главе 1 приводится вывод уравнений, описывающих скорости реакций, имеющих произвольное число реактантов. Эти уравнения, являющиеся существенным обобщением модели Моно, Уаймена и Шанжё и модели Фридена — Курганова, могут быть использованы для описания очень широкого класса биохимических реакций. Глава 2 посвящена описанию динамических свойств открытых ферментативных реакций. Описанию предпослан вводный раздел 2.1, кратко знакомящий с основными методами качественного исследования математических моделей, используемых в этой главе и в некоторых главах части 2. Хотя в последние годы математические исследования динамики проточных химических и биохимических реакций довольно популярны, очень мало сделано для того, чтобы сформулировать достаточно общую теорию таких реакций. Поэтому часто публикуются работы, результаты которых можно было бы заранее предсказать на основании того, что многие кинетические модели открытых (био)химических и мембранных систем математически топологически эквивалентны. Целесообразно подробно изучить лишь одну из множества эквивалентных моделей, а затем ограничиваться установлением принадлежности новой, еще неизученной модели к некоторому классу эквивалентных моделей. Если удается доказать такую принадлежность, то все свойства наиболее детально изученной модели, которую можно считать эталонной моделью данного класса, автоматически переносятся на новую, еще неизученную модель. С учетом этих соображений в разделах 2.2 и 2.3 проведено весьма детальное исследование математических моделей двух типов открытых ферментативных реакций — обратимой реакции с субстратным угнетением и необратимой двухсубстратной реакции с субстратным угнетением. Обе эти реакции, несмотря на несходство кинетических схем, математически эквивалентны: они имеют одинаковое строение параметрических и фазовых портретов. Эти модели можно рассматривать как эталонные модели широкого класса (био)химических систем — систем, квазистационарные характеристики (входные или выходные характеристики, главные изоклины) которых имеют гистерезис. В частности, в разделах 2.4 и 2.6 показано, что различные варианты моделей открытых реакций эквивалентны моделям, разобранным в разделах 2.2 и 2.3. Суммированные в разделе 2.5 основные результаты показывают, каким богатством динамического поведения обладают даже очень простые ферментативные реакции. При подходящих условиях они способны генерировать устойчивые или неустойчивые автоколебания и однократные спайкообразные импульсы стандартной формы (после действия соответствующего сверхпорогового возмущения); способны переходить из одного устойчивого режима в другой, причем альтернативные устойчивые режимы могут иметь как колебательный, так и неколебательный характер, а период колебаний в открытых биохимических системах из-за эффекта депонирования (раздел 2.7) может быть очень большим — порядка суток. В главе 3 рассматривается математическое моделирование полиферментных систем. В литературе, посвященной этому типу моделирования, очень часто приводятся исключительно сложные математические модели, насчитывающие многие десятки переменных и параметров. Можно показать, что такие модели содержат много лишней информации, обременяющей их исследование, и много «шума», вызванного неточностью экспериментального определения параметров, неизбежной произвольностью в части допущений о механизмах реакций и условиях их протекания, а также некоторой неустойчивостью вычислительного алгоритма, используемого при анализе модели на компьютере. Поэтому подобные модели создают лишь иллюзию высокой точности описания полиферментной системы, подобную той, какая возникает при распечатке результатов вычислений, производившихся на компьютере, скажем, с десятью значащими цифрами: из-за малейшей неточности в исходных данных или программе сомнительными могут оказаться не только первые цифры, но и порядок величин результата. После обзора математических моделей полиферментных систем, проведенного в разделах 3.1 и 3.2, вводится представление о трех иерархических уровнях организации полиферментных систем, последовательно возникших в ходе биологической эволюции. Эти уровни названы первичной, вторичной (стехиометриче-ской) и третичной (нестехиометрической) структурами полиферментных систем. На основе этой классификации предлагается метод теоретического анализа механизма регуляции клеточного метаболизма, основанный на последовательном рассмотрении свойств различных уровней. В качестве примера использования этого метода в разделах 3.3 и 3.4 показано, что исследование вторичной структуры энергетического метаболизма позволяет выявить ряд существенных его свойств и предсказать множество регуляторных связей, принадлежащих более молодой (в эволюционном отношении) и существенно более сложной третичной структуре. Обосновывается утверждение, что третичная структура полиферментных систем, содержащая всевозможные аллостерические регуляторные взаимодействия, может быть теоретически реконструирована, исходя из более древней, потому существенно более простой, вторичной структуры. В разделе 3.3 показано, что кинетическая модель энергетического метаболизма при определенных условиях может быть редуцирована к модели одной эквивалентной реакции. Этот вывод означает, что математические модели полиферментных систем могут быть такими же простыми, как и модели отдельных ферментативных реакций, и что о свойствах большой полиферментной системы можно судить по свойству всего лишь одной эквивалентной реакции, вроде той, какие были рассмотрены в главе 2. Часть 2 книги посвящена возбудимым мембранам. Мембраны в клетке выполняют весьма разнообразные функции: ионные каналы мембраны избирательно пропускают одни ионы и задерживают другие, насосы мембраны накачивают в клетку необходимые вещества против градиента концентрации, на мембранах локализованы многие важные ферменты. Возбудимые мембраны используются клеткой для передачи информации. Нарушения работы мембран приводят к различным заболеваниям — от небольших нервных расстройств до тяжелых психических нарушений, параличей и нарушений работы сердечной мышцы. При исследовании возбудимых мембран сейчас много внимания уделяется их молекулярной организации. Но знание того, какие молекулы и в каком порядке стоят в каких каналах и рецепторах, вряд ли является окончателыш целью. Для практического приложения в будущем результатов мембранных исследований важно научиться сознательно воздействовать на феноменологические характеристики мембран, выработать принципы создания необходимых биологически активных и лекарственных веществ. Важно понять, на какие ионные токи и как необходимо воздействовать, чтобы изменять характеристики мембран в желательном направлении. Для уяснения принципиальных механизмов связи ионных, токов и электрофизиологических характеристик мембран, много дали работы Ходжкина и Хаксли. Но метод Ходжкина — Хаксли трудоемок, и вряд ли его целесообразно использовать в повседневной экспериментальной работе при изучении действия биологически активных веществ на мембраны. Важно иметь доступный для повседневной экспериментальной работы метод, позволяющий определять, какие из регистрируемых изменений ионных токов ответственны за определенные изменения характеристик и поведения мембран. В настоящей книге для решения этой задачи предлагается следующий подход. Не составлять полных уравнений мембраны и вообще не составлять никаких уравнений. А связи между ионными токами и электрофизиологическими характеристиками мембраны устанавливать непосредственно по записям ионных токов при фиксации потенциала, используя для их обработки качественные методы теории колебаний (см. раздел 2.1). Этот подход применяется и проверяется сначала на математических моделях мембран (глава 4), а затем переносится на анализ «живых» возбудимых мембран (глава 5). В главе 4 качественные методы анализа используются для исследования модели Ходжкина — Хаксли для мембраны аксона кальмара и модели Нобла для волокна сердца теплокровного. Уравнения типа Ходжкина — Хаксли с помощью методов малого параметра сводятся к уравнениям второго порядка (типа уравнения Ван дер Поля), после чего они легко исследуются методом фазового портрета и качественными методами по нуль-изоклинам. В результате применения качественных методов теоретический анализ моделей мембран стал менее трудоемким и более мощным. Особенно важно, что использование качественных методов позволяет значительно упростить экспериментальную работу при исследовании мембран. Этому вопросу посвящена глава 5, где излагается применение качественных методов для анализа записей ионных токов при фиксации потенциала. Качественные методы позволяют легко и быстро определить в экспериментах с фиксацией потенциала, за счет изменения каких компонент мембранного тока происходит изменение тех или иных электрофизиологических характеристик. Особенно большое значение эти методы могут иметь при поиске биологически активных веществ с заданными свойствами. В главе 5 также описываются разработанные методы построения уравнений типа Ван дер Поля для исследуемых мембран непосредственно по записям ионного тока при фиксации потенциала. Такие модели были построены для ряда объектов: волокна предсердия лягушки, волокна Пуркинье сердца теплокровного, волокна поперечнополосатой мышцы, гигантского аксона кальмара, перехвата Ранвье, нейрона улитки. Часть 3 книги посвящена волнам в возбудимых (активных) средах, в частности в возбудимых мембранах. Волны здесь распространяются без затухания, заново генерируясь каждой точкой мембраны. Волны возбуждения являются сильно нелинейными волнами; при столкновении они гаснут, отражения от границ среды отсутствуют. Эти свойства обеспечивают их важное отличие от аддитивных волн (электромагнитных, звуковых), хорошо изученных в математической физике. Наибольшее внимание уделяется процессам возникновения волн, так называемым автоволновым процессам, аналогичным автоколебательным процессам в сосредоточенных системах. Автоволновые процессы лежат в основе функционирования самых различных активных сред — распределенных экологических, электрохимических и химических систем, физических систем с инверсной населенностью (рабочие тела лазеров), различных технических устройств — цепочек мультивибраторов, линий из туннельных диодов и нейристоров, а также распределенных систем, образованных возбудимыми биологическими мембранами, нейронных структур, нервных и мышечных волокон, сердечной ткани. В Институте биологической физики АН СССР была предсказана возможность возникновения источников спиральных волн и источников периодических концентрических волн в неоднородных активных средах, состоящих из неавтоколебательиых элементов; теоретически изучены свойства этих источников волн; здесь же были выполнены первые эксперименты по наблюдению этих источников волн в распределенной химической активной среде. Результаты, полученные при анализе автоволновых процессов в возбудимых средах, нашли широкое применение для исследования концентрационных волн в химических системах, а также для анализа механизмов аритмий сердечной мышцы. Главы 6 и 7 посвящены исследованию новых типов источников волн, специфических для возбудимых сред — вращающейся спиральной волны, так называемого ревербератора (глава 6), и источ-. ника эха (глава 7). В главе 8 рассматриваются процессы, происходящие в возбудимых средах при взаимодействии источников волн, и их приложение к анализу сердечных аритмий. В главе 6 рассмотрены процессы, приводящие к возникновению источников волн в возбудимых средах. Покавано, что распространение волн в возбудимых средах с размерностью Детально изучены свойства и поведение источников спиральных волн (ревербераторов). Обнаружено, что в неоднородных средах эти источники волн имеют конечное время жизни; они исчезают в результате того же процесса образования разрывов волн, который привел к их возникновению. Время жизни спиральной волны оказывается тем больше, чем больше длительность возбужденного состояния элементов среды и чем меньше степень неоднородности среды; в однородных средах ревербератор устойчив и имеет неограниченное время жизни. Однако в однородных средах распространение волн не приводит к формированию ревербераторов. Одно из важнейших свойств источников волн — их способность размножаться в неоднородных средах. Размножение источников возможно в результате того, что волны, посылаемые ими, в неоднородной среде разрываются (так как эти источники посылают волны с максимальной частотой, которую способна пропустить данная среда); из разрывов возникают новые источники волн. Изучены условия, при которых возможно размножение. Отмечены аналогии с цепными или автокаталитическими реакциями: тип режима в среде зависит от соотношения скоростей двух противоположных процессов — размножения источников волн и их исчезновения. Если скорость размножения источников волн превышает скорость их исчезновения, в среде возникает нерегулярный режим, связанный с большим числом источников волн, работающих с разными частотами, исчезающими в одних местах и появляющимися в других. Ставится задача о нахождении критических значений параметров возбудимой среды, при превышении которых в среде становятся возможными нерегулярные режимы. Полученные результаты используются для создания нового подхода к поиску сердечных противоаритмических лекарственных средств. В части 4 представлены работы, которые иллюстрируют различные области применения новых методов исследования пространственной организации клеточных структур: трехмерную реконструкцию по электронно-микроскопическим снимкам биологических структур, обладающих определенными видами симметрии (на примере реконструкции хвоста фага); голографический синтез объемных биологических объектов по их ракурсным снимкам, полученным в режиме вторичных электронов (на примере восстановления поверхностного рельефа мембраны нейрона); моделирование гипотетической молекулярной структуры объекта путем расчета плоского изображения и сопоставления с истинным электронномикроскопическим изображением (на примере изучения структуры лейцинаминопептидазы); машинное исследование изображений клеточных структур, наблюдаемых в оптический микроскоп, путем их сканирования с помощью телевизионных автоматов (на примере определения геометрических параметров глиальных клеток и метафазных хромосом); восстановление первичной структуры биополимеров по набору их фрагментов (на примере анализа первичной последовательности РНК). В просвечивающем электронном микроскопе имеют дело с проекциями клеточных структур. В световой микроскопии в зависимости от глубины резкости объектива наблюдают либо сечение клетки, либо ее проекцию на плоскость. Проекции и сечения биологических структур описываются двумерными функциями оптической плотности. Дальнейший путь исследования может состоять либо в том, что ограничиваются плоскими представлениями о пространственной структуре, либо определяются некоторые количественные характеристики трехмерной структуры по имеющемуся в наличии набору плоских картин. Иной подход состоит в том, чтобы, набрав достаточное количество плоских представлений объемной клеточной структуры, построить ее пространственную модель (аналогично тому, как на основе трех ортогональных проекций детали конструктор воссоздает ее пространственную форму). Если объект симметричен, то для его моделирования в ряде случаев достаточно одной проекции. Например, при исследовании спиральных структур (таких, как волокна биополимеров или чехлы бактериофагов) можно избежать трудоемких операций съемки объекта в разных ориентациях в электронной микроскопии. Модель структуры чехла бактериофага, реконструированная таким методом, рассмотрена в разделе 9.3. Следует заметить, что организация отростка бактериофага, являющегося простейшей структурой, способной к сокращению, представляет исключительный интерес. Возможны и другие применения этих методов. Часто бывает достаточно найти внешнюю форму структуры, например поверхность клетки, рельеф этой поверхности. Изменение внешней мембраны клеток отражает как метаболические процессы, происходящие внутри клеток, так и характер контакта между клетками. Рельеф клетки сравнительно легко поддается наблюдению. Прогресс этих исследований стал заметен с распространением растровых электронных микроскопов, работающих на отражение во вторичных электронах. Однако, несмотря на заметный «эффект объемности», возникающий при рассмотрении изображений, полученных в растровом электронном микроскопе, количественная оценка размеров деталей рельефа затруднительна. Связано это с отсутствием параллакса в рассматриваемом изображении. Здесь оказались полезными методы голографии, которые позволили объединить на одной голограмме информацию о различных ракурсах объекта. Эта информация содержится в наборе снимков, сделанных в растровом электронном микроскопе под различными углами. Изображение, восстановленное с голограммы, отражает особенности поверхности клетки, так как объект выглядит объемным. Такой метод позволяет перенести изображение клетки размерного диапазона электронного микроскопа в размерный диапазон обычных оптических наблюдений, где возможны прямые измерения ее рельефа с помощью светящихся меток, сопряженных с измерительными механизмами. Этот метод рассмотрен в разделе 9.4. При исследовании трехмерных клеточных структур по их сечениям и проекциям исследователь обычно сталкивается с проблемой некорректности обратных задач, т. е. задач, которые решаются в обратном порядке причинно-следственных отношений (по следствию необходимо выяснить причину). Почти все научные исследования — это решение обратных задач; наблюдается эффект, производимый изучаемой системой (следствие), а восстанавливается ее строение (причина). Такие задачи часто не имеют решения. Иногда решение можно найти, но из-за неоднородности структуры возникает его неоднозначность, т. е. необходимая информация имеется, но ее недостаточно. Наконец, возможен третий случай: если удалось измерить интенсивность, поляризацию и фазу световых волн, рассеянных структурой клетки по отношению к падающему свету, как функцию углов рассеяния, то согласно формальной постановке обратной задачи этой информации, в принципе, достаточно для получения количественного описания микроструктуры объекта. Однако вычислительная схема подобных задач очень чувствительна к небольшим ошибкам регистрируемых данных. Если интегральные значения измеряются с ошибкой (а при измерениях погрешность всегда существует), то вычисляемая подынтегральная функция может сильно отличаться от истинного значения. Механизм такой слабой помехозащищенности очевиден. Параметры объемной клеточной структуры вычисляются как разности близких и больших величин, измеряемых на сечениях или проекциях. Чтобы уменьшить ошибку, необходимо увеличивать точность и статистику измерений при определении параметров клетки, описываемых интегральным выражением. Однако существует еще один путь борьбы с некорректностью обратных задач. Это моделирование путем использования априорной дополнительной информации о структуре объекта. Если что либо известно об организации структуры подынтегральной функции, то можно попытаться найти решение, перебирая варианты подынтегральной функции и вычисляя каждый раз интеграл, значение которого сравнивается с измеренным до их совпадения. Такая прямая схема вычислений путем моделирования клеточной структуры более помехоустойчива, но требует предварительной, хотя бы приближенной, информации о характере этой структуры. Одному из простейших вариантов такого моделирования клеточных структур посвящен раздел 9.5. Опыт показывает, что при опознавании объектов их контур — одна из наиболее информативных частей изображения. Основная математическая проблема, возникающая при машинном анализе изображений клеток и их частей, состоит в получении исходного словаря, в терминах которого можно моделировать строение объекта, и набора правил машинного конструирования из его слов трехмерной структуры клеток. В качестве примера рассмотрен словарь, в основе которого лежит контурное описание структурных клеточных элементов на срезах или проекциях. Целью анализа контура является вычисление геометрических параметров структурных клеточных элементов или нахождение характерных точек, позволяющих определить их форму. Эти вопросы освещены в главе 10. Глава 11 посвящена анализу биологических клеточных структур, описываемых одномерными функциями. Для расшифровывания цепи нуклеиновых кислот длиной в несколько сот нуклеотидов необходима разработка алгоритмов, позволяющих восстанавливать первичную структуру по отдельным перекрывающимся блокам разорванной цепи. «Интуитивного» перебора вариантов для определения длинных последовательностей оказывается недостаточно. Математическая постановка этой задачи может быть сформулирована в терминах восстановления неизвестного слова по известному набору его фрагментов, полученных определенным способом. В разделе 11.5 даны рекомендации для планирования биохимического эксперимента по расшифровке первичной последовательности нуклеотидов.
|
 |
Оглавление
|








































































































































































































































































































 качественно мало отличается от распространения импульса в волокне, если среда однородна, и рассматриваются «естественные» начальные условия (соответствующие состоянию покоя элементов среды). Качественно новые режимы возникают в неоднородных средах, где могут происходить разрывы волн. При эволюции разрыва его края скручиваются, что приводит к формированию вращающихся спиральных волн. Большое число спиральных волн создает картину, внешне похожую на турбулентность.
качественно мало отличается от распространения импульса в волокне, если среда однородна, и рассматриваются «естественные» начальные условия (соответствующие состоянию покоя элементов среды). Качественно новые режимы возникают в неоднородных средах, где могут происходить разрывы волн. При эволюции разрыва его края скручиваются, что приводит к формированию вращающихся спиральных волн. Большое число спиральных волн создает картину, внешне похожую на турбулентность.